
Ontogenetic distribution of genetic lethality in Drosophila willistoni
24
ONTOGENETIC DISTRIBUTION OF GENETIC LETHALITY IN DROSOPHILA WILLISTONI M. T. M. RIZKI Deportment of Zoology, Colzirnbia University, New Pork, N. P. SEVEN FIGURES INTRODUCTION A Mendelian unit which causes the death of a zygote before the reproductive age may be designated as a lethal. The analysis of concealed variability of natural populations of Drosophila has revealed that a considerable proportion of recessive mutations have a lethal effect (Dobzhansky et al., '42; Dubinin, '46; Pavan et al., 51). These recessive muta- tions comprise a range of viability disturbances, from sub- vital factors, which kill only a few per cent of the individuals carrying the mutated unit, to lethals which result in the death of all organisms with the lethal factor. The various types of lethals cannot be sharply demarcated; moreover, the expression of a lethal factor may be conditioned by the residual genotype as well as by environmental changes. Changes in Mendelian factors are expressed through the developmental patterns of the organism. The effects of a lethal mutation, which alters the normal developmental pat- terns, can be assumed to depend upon an interaction between the lethal factor and the ontogenetic system of which it is a part. The kinds of lethals found in a species may therefore be expected to reveal some of the characteristics of the reactive system which the species has evolved as part of its adaptive requirement. Submitted in partial fulfillment of the requirement for the degree of Doctor of Philosophy in the Faculty of Pure Science, Columbia University, New York City. 327
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Ontogenetic distribution of genetic lethality in Drosophila willistoni

ONTOGENETIC DISTRIBUTION O F GENETIC LETHALITY IN DROSOPHILA
WILLISTONI
M. T. M. RIZKI Deportment of Zoology, Colzirnbia University, New Pork, N . P.
SEVEN FIGURES
INTRODUCTION
A Mendelian unit which causes the death of a zygote before the reproductive age may be designated as a lethal. The analysis of concealed variability of natural populations of Drosophila has revealed that a considerable proportion of recessive mutations have a lethal effect (Dobzhansky et al., '42; Dubinin, '46; Pavan et al., 51). These recessive muta- tions comprise a range of viability disturbances, from sub- vital factors, which kill only a few per cent of the individuals carrying the mutated unit, to lethals which result in the death of all organisms with the lethal factor. The various types of lethals cannot be sharply demarcated; moreover, the expression of a lethal factor may be conditioned by the residual genotype as well as by environmental changes.
Changes in Mendelian factors are expressed through the developmental patterns of the organism. The effects of a lethal mutation, which alters the normal developmental pat- terns, can be assumed to depend upon an interaction between the lethal factor and the ontogenetic system of which it is a part. The kinds of lethals found in a species may therefore be expected to reveal some of the characteristics of the reactive system which the species has evolved as part of its adaptive requirement.
Submitted in partial fulfillment of the requirement for the degree of Doctor of Philosophy in the Faculty of Pure Science, Columbia University, New York City.
327

328 M. T. M. RIZKI
Developmental analyses of lethal mutants in Drosophila have shown that death due to lethal factors may occur at different stages of development (Li, '27 ; Sivertzev-Dob- zhansky, '27 ; Medvedev, '39 ; Poulson, '40 ; Hadorn, '48, '51) ; but there is little information concerning the frequencies with which lethal mutations affect the various developmental stages of the organism. Does death due to lethal factors occur equally often throughout all stages of development, or are some developmental stages affected more often than others 7 Furthermore, do newly arisen lethals, which are a random sample of all lethal mutants, differ from lethals carried in a concealed state in natural populations and which therefore should be nearly neutral in heterozygous condition? The purpose of the present investigation is to determine the relative frequencies of lethals which cause death at different stages of development. Three samples of lethals were studied, namely, lethals extracted from natural popu- lations, lethals which arose spontaneously in laboratory strains, and lethals obtained in the offspring of flies treated with x-rays.
ACKNOWLEDGMENTS
The author wishes to express his gratitude to Professors Th. Dobzhansky, L. C. Dunn, and Howard Levene for their guidance and encouragement throughout the course of this investigation. He also wishes to thank Dr. C. Pavan, Mr. A. R. Cordiero, and Mr. B. Spassky for providing the spon- taneous lethal stocks.
MATERIALS AND METHODS
Since the main purpose of this investigation is to study the behavior of the "reactive system" in relation to the changed norms of reaction brought about by mutation in nature, the use of isogenic stocks has deliberately been avoided. Isogenic material would not only be unnecessary, but could even be misleading since the existence of an iso- genic stage in nature is unknown.

GENETIC LETHALITY IN DROSOPHILA 329
The three samples of DrosophiZa willistorti recessive lethals used in this study are: (1) Twenty-four spontaneous lethals extracted from various natural populations of Brazil. Twelve of these lethals were tested and shown to be non-allelic by Dr. C. Pavan, and the others by the author. (2) Twenty-nine spontaneous lethals derived from wild stocks which once had normal viability under laboratory conditions. The spontane- ous laboratory lethals were found during a determination of spontaneous mutation rates of D. wiZZistorti by Professor Th. Dobzhansky and B. Spassky, and were shown to be non-allelic by the author. (3) Twenty-two x-ray induced recessive lethals (6000 r). The two samples of spontaneous lethals consisted of lethal factors associated with the second and third chro- mosomes. Induced lethal factors were located on the second chromosome with the exception of one stock which carried a lethal factor on the third chromosome. All of the lethals used had been maintained in balanced lethal stocks.
The method of extracting autosomal lethals, both spon- taneous and induced, is essentially the same as that de- scribed by Dobzhansky et al. ('42) for D. pseudoobscura and Pavan et al. ('51) for D. wiZEisfoi& The general method is illustrated schematically in figure 1.
For step A in figure 1 one of two stocks may be used: a stock homozygous for the second chromosome with two reces- sive markers, abbreviated and brown; or a stock homozygous for the third chromosome with the recessive marker, pink. The males which were radiated came from a special stock made homozygous for a lethal-free second chromosome. There are two balancer chromosomes which may be used for step B, one for the second chromosome and the other for the third chromosome. (These chromosomes were synthesized in Pro- fessor Dobzhansky 's laboratory.) The second chromosome balancer has two dominant markers, Star and Hook, two recessive markers, abbreviated and brown, and inversion no. 207. The balancer stock for the third chromosome carries a dominant marker, Delta, and a recessive marker, pink, in addition to inversion no. 133. Both of these balancers are

330 M. T. M. RIZKI
good suppressors of crossing-over and both are lethal in homozygous condition.
Before a developmental analysis of a lethal stock can be made, the chromosome bearing the lethal factor to be tested must be separated from the balancer chromosome. This procedure is necessary because a lethal factor is associated
wild or irradiated cf 9 step A
P - X I 1
step B sinqle FI d ,
1
P 0 1 A -
I F “LGHAG
- I.
DIES PHENOTYPE-B WILD TYPE
F3 I: 2 :
DIES Fig. 1 General scheme of the crosses used to detect a lethal on the second
or third chromosome. The wild or irradiated chromosomes .are shown in black, the recessive marker chromosomes in white, and the balancer chromosomes in white with a curvature representing an inversion, and a dominant marker (B) .
with each balancer chromosome. A wild strain of D. willistoni, Standard, which has normal viability (-;*; x 100 = 85- 95%) has been used in this work for the purpose of outcrossing the lethal stocks. The method of outcrossing is represented in figure 2. When males from the lethal stock are crossed to Standard (normal) females two phenotypes are obtained in the F,, wild-type flies and flies with the dominant marker,

GENETIC LETHALITY I N DROSOPHILA 331
B. The phenotypically normal males and females are mated, and eggs from this cross are used for determining the stage of death of the lethal homozygotes. Some of the females heterozygous for the lethal factor (F,) are also crossed to wild males, and eggs obtained from this mating serve as a parallel control of the lethal cross. This control mating is used to estimate the mortality due to environmental and other unknown variables. Ideally each lethal under investi-
lethal stock normal d - x Q n
5 0 I a 0 I F f r om lefhol cross ( 1 ) control cross(2) 2 1 1 1 I I I 1 1
Fig. 2 Scheme of lethal and control crosses used t o obtain the flies for egg collection. The chromosomes with the lethal under test are represented a8 black bars, the balancer chromosomes are shown with curvatures and dots, and the wild chromosomes are white.
gation should have its specific control, but this condition requires an enormous amount of work and time. Therefore, the procedure adopted is to run a single control for a group of 5 or 6 lethals tested at one time. This combined control is obtained by crossing a mixture of Fl lethal heterozygous females representing each lethal under observation to the wild males.
TechNicaZ procedure. The lethal heterozygous parents (F,) are released in a fly chamber which is a glass tube 5cm in diameter and 10cm high. The bottom of this tube is closed

332 M. T. M. IlIZKI
with a piece of linen held in place by a rubber band. The upper end is covered by a lid made of nylon and a plastic rim. This chamber is placed on a wet filter paper which is constantly moistened by water from inverted bottles placed on the filter paper near the fly chambers. Paper spoons with food are placed on the floor of the chamber as illustrated in figure 3. This method of egg collection which is similar to the one described by Hodson and Chiang ( '48) was found necessary in order to maintain a high humidity. The survival of D. willistomi males and the per cent of egg hatching were
G.
Fig. 3 Arrangement f o r egg collection. F.CH., fly chamber; FD., food; F.P., filter paper; G.P., glass plate; INV.B., inverted bottle with water; SP., paper spoon.
greatly improved by this technique (90-95% hatching as compared to 3040% by usual methods of egg collection).
For the first two days the flies are given honey and yeast in a 1 : 1 mixture on the food (cream of wheat and molasses ; Spassky, '43). Thereafter plain yeasted food and honey yeasted food are given on alternate days. Eggs are collected either until the required number is obtained or until there is a drop in the per cent of egg hatching. Spoons were changed every 12 hours.
The eggs on the spoons are counted when the spoons are removed from the fly chambers. Spoons with the counted eggs are placed in stender dishes and left in a constant tem- perature room at 25 -+ 1°C.. After 16 hours the larvae which hatch are removed from the spoons and placed in creamers

GEKETIC LETHALITY IN DROSOPHILA 333
containing 8-10 em3 of yeasted cream of wheat-molasses food. Preliminary experiments showed that optimum hatching of adults is obtained with 20 larvae per creamer. The spoons are rechecked twice within the next 24 hours for any further hatching of larvae. After the final examination of the spoons the unecloded eggs are counted and the per cent egg hatching is determined.
The creamers are examined after 7 or 8 days. The pupal count is made the day the first flies begin to emerge. Adults are removed and counted every day until all of the pupae are left open by emerging adults. The pupae which do not give rise to adults even a week after the time that the first adults emerged are considered dead. Such dead pupae as well as dead larvae found on the food are fixed in chloroform- acetic-alcohol for further examination and comparisons.
Determination of the stages at which death occurs During the egg stage embryogenesis of the larval and
imaginal structures occurs. The larval organs are completely differentiated and prepared to function by the end of this stage. Mendelian factors causing death of the organism at any time before hatching are classified as egg lethals (E).
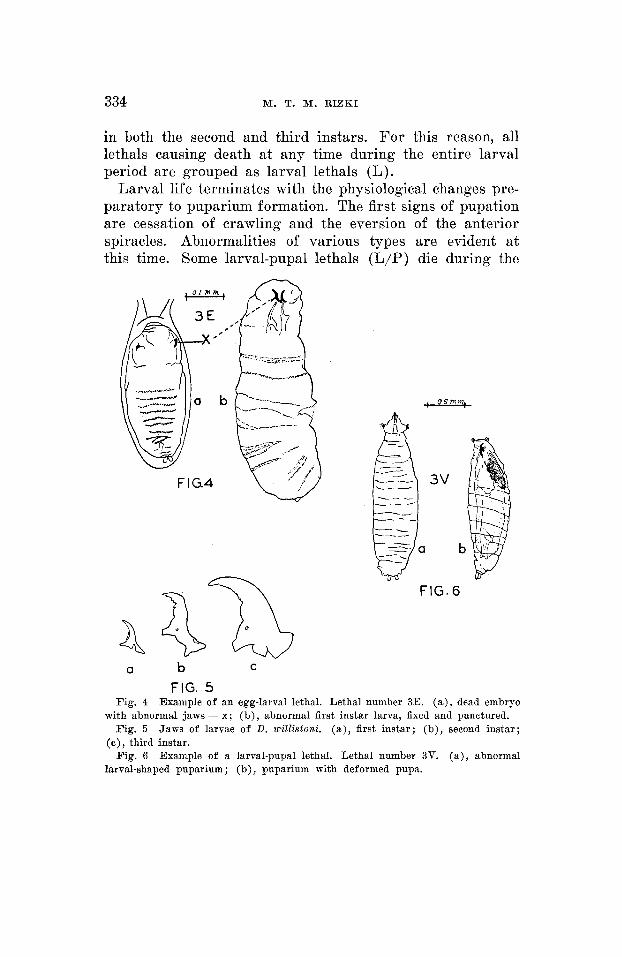
The beginning of the post-embryonic phase is characterized by the hatching movement of the completed embryo within the egg case. An egg-larval lethal (E/L) is the result of a lethal factor which allows the organism to complete embry- onic development, but arrests further developmental activity by causing death of some individuals at the stage of hatching movement and of other zygotes shortly after eclosion from the egg. An example of an egg-larval lethal is presented in figure 4.
The three larval instars of D. willistomi can be distinguished, like those of D. nzeZa,wogasier, by the dentition of the jaws (fig. 5 ) . Larval death caused by lethal factors can occur in any of the three instars or in a combination of two or three instars. For example, some lethals cause death in the first and second instars, while lethality in other cases may occur
334 M. T. M. RIZKI
in both the second and third instars. For this reason, all lethals causing death at any time during the entire larval period are grouped as larval lethals (L).
Larval life terminates with the physiological changes pre- paratory to puparium formation. The first signs of pupation are cessation of crawling and the eversion of the anterior spiracles. Abnormalities of various types are evident at this time. Some larval-pupal lethals (L/P) die during the
. 06mm.
3v
b
FIG. 6
a b C
FIG. 5 Fig. 4 Example of an egg-larval lethal. Lethal number 3E. (a) , dead embryo
Fig. 5 Jaws of larvae of D. wiZ2istoni. (a) , first instar; (b) , second instar;
Fig. 6 Example of a larval-pupal lethal. Lethal number 3V. (a) , abnormal
with abnormal jaws = x; (b), abnormal first instar larva, fixed and punctured.
(c ) , third instar.
larval-shaped puparium ; (b), puparium with deformed pupa.

G E N E T I C L E T H A L I T Y IX DROSOPHILA 335
early processes of puparium formation, with the result that they appear as larvae with a tanned puparium-like exoskele- ton. Other individuals possessing the same lethal factor form normal puparia, but fail to complete pupal development. Figure 6 is an example of a larval-pupal lethal.
A pupal lethal (P) factor causes death of the individual sometime after the pupal moult but before emergence of the imago.
Morphological determination of the stage of death
The presence of a large number of dead individuals among the offspring of parents heterozygous for a lethal factor and their absence in the corresponding control is one criterion for determining the stage at which death occurs. Lethals which show morphological abnormalities can be easily classi- fied as to the stage of death.
Xtatistical analysis
Even though egg hatchability was improved by using special techniques f o r collection, the problem of unfertilized eggs could not be avoided. This factor is especially difficult when dealing with D. willistoni. As pointed out by Hadorn ('51) unfertilized eggs cannot be distinguished from those whose death is due to genetic factors in the early embryonic stages. Cytological examination of every unhatched egg is imprac- ticable. Therefore, a statistical comparison of total mor- talities and stage mortalities in the progeny of the lethal cross and the control cross has been utilized in addition to morpho- logical observations.
Mortalities are calculated on the basis of the number of zygotes obtained in the lethal and control crosses. Assuming that the viability of lethal/normal is the same as normal/nor- mal, let M, be the total mortality among the offspring of the lethal cross and 11, the total mortality of the zygotes in the control cross. Theoretically every lethal homozygote

336 M. T. M. RIZKI
should die before attaining the reproductive age. The ex- pected mortality due to the class of lethal/lethal is 25% of the number of offspring. But ML is a composite mortality of the lethal homozygotes (4) plus mortality which occurs in the other 9 of the zygotes (refer to fig. 2). The latter mortality is also a component of M,. Therefore,
ML=4 + ~ M c
If a is the probability that a lethal homozygote will die, then
M, = a/4 + 4 M,
a = 4ML - 3Mc = Mc + 4 (ML - M,)
(3)'Mo (1 - Mc) NC
,,% - (4)' ML (1 - ML) + N L
ii-
where B is the sample estimate of a obtained by substituting observed values Mc and M, for the true values, and where N, and N, are the numbers of eggs from the lethal and control cross respectively.
Hypothetically at the 5% level,2 (2- 2 u 8 ) L a & (2 + 2u8)
I n order to determine mortality in the egg stage two values of B are computed:
where ML and M, are based on the value, 1 - ( totalno. eggs )
'(E) eggs -C ~ B ( E )
total no. adults
'(Lv) larvae -C ua(Lv)
total no. adults where the values M', and M', are based on, 1 - ( total no. larvae ) - 'The theoretical standard deviations are probably too small since they do not
take account of heterogeniety. Fo r example, 3WC (table 2) gives a value aE = 1.36 -C 0.10, which seems to be significantly greater than 100% mortality. Such results may be due to the presence of unfertilized eggs either in the lethal or control cross, or may be caused by differences between the heterozygous males and the normal standard males. The present work was not designed to study these diff erences.

GENETIC LETHALITY IN DROSOPHILA 337
If the &(Ly) value is significantly lower than 1, and there is no significant difference in the lethal and control larval and pupal stage mortalities as tested by,
M’L - M c V aZM’L + U’M‘O t1.96 =
then the lethal factor is concluded to cause death in the egg stage.
If the &,,, is significantly lower than 1 and there is also a significant difference for the t value, then the lethal factor is an egg-larval one.
Statistical determination of larval and pupal lethals is a simple comparison of per cent mortalities of these stages in the lethal and control.
RESULTS
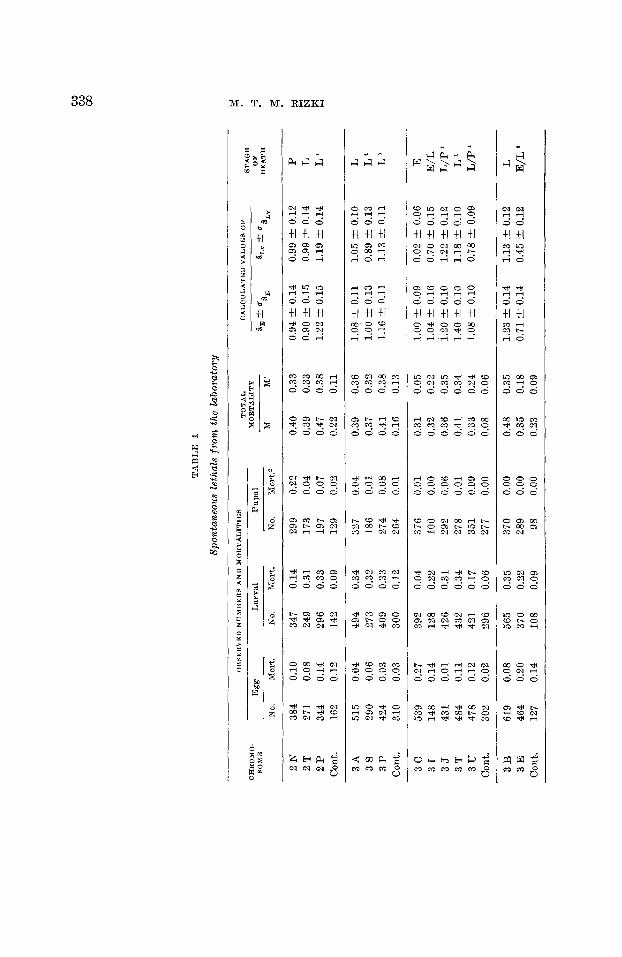
The data from the analysis of the two samples of spontane- ous recessive lethals are summarized in tables 1 and 2, and from the x-ray induced lethals in table 3.
Table 1 consists of 29 spontaneous lethals which originated in laboratory cultures. There is one egg lethal, 4 egg-larval lethals, 19 larval lethals, 4 larval-pupal lethals and one pupal lethal. The sample of 24 spontaneous lethals from wild popu- lations (table 2) is distributed as follows : one egg lethal, two egg-larval, 16 larval, two larval-pupal, and three pupal lethals. There is no significant difference between the frequency dis- tribution of lethals in the two samples of spontaneous lethal factors (table 4). Therefore these two samples are pooled together, the distribution of the stages of death of spontane- ous lethals being: two egg, 6 egg-larval, 35 larval, 6 larval- pupal, and 4 pupal lethals.
Among the 22 x-ray induced lethal chromosomes there are three lethals which cannot be treated in the statistical manner outlined above. These three lethal chromosomes (S,,, S35, SIl5) gave high mortalities (.55-.72 total mortality after correction for control mortality) in all developmental stages. No such lethals were found among the spontaneous
TA
BL
E 1
Spon
tane
ous
leth
als
fro
m t
he l
abor
ator
y
STA
GE
0L" -t c-
DE
AT
H
TO
TA
L
CA
LC
UL
AT
ED
VA
LU
ES
OF
O
HSE
RV
ED
NU
MB
ER
S A
ND
MO
RT
AL
ITIE
S
OF
H
MO
RT
AL
ITY
0,
+- u
M
M
8,
&L
V
Egg
L
arva
l P
upal
No.
M
ort.
No.
M
ort.
N
o.
hlor
t.2
CH
RO
MO
- SO
ME
K 2
N
384
0.10
34
7 0.
14
299
0.22
0.
40
0.33
0.
94 t 0.
14
0.99
2 0
.12
P
2P
34
4 0.
14
296
0.33
19
7 0.
07
0.47
0.
38
1.22
2 0
.15
1.19
2 0
.14
L'
2T
27
1 0.
08
249
0.31
17
3 0.
04
0.39
0.
33
0.90
2 0
.15
0.99
2 0
.14
1
z C
ont.
162
0.12
14
2 0.
09
129
0.02
0.
22
0.11
3A
51
5 0.
04
494
0.34
32
7 0.
04
0.39
0.
36
1.08
t 0
.11
1.05
t 0.
10
1
3s
29
0 0.
06
273
0.32
18
6 0.
01
0.37
0.
32
1.00
2 0
.13
0.89
C 0
.13
L'
3P
42
4 0.
03
409
0.33
27
4 0.
08
0.41
0.
38
1.16
2 0
.11
1.13
t 0
.11
L'
Con
t. 31
0 0.
03
300
0.12
26
4 0.
01
0.16
0.
13
3c
53
9 0.
27
392
0.04
37
6 0.
01
0.31
0.
05
1.00
& 0
.09
0.02
t 0
.06
E
31
14
8 0.
14
128
0.22
10
0 0.
00
0.32
0.
22
1.04
t 0
.16
0.70
t 0.
15
E/L
3
5
431
0.01
42
6 0.
31
292
0.06
0.
36
0.35
1.
20 t 0
.10
1.22
'-t
0.12
L
/P
3T
48
4 0.
11
432
0.34
27
8 0.
01
0.41
0.
34
1.40
2 0
.10
1.18
C 0
.10
L'
3u
47
8 0.
12
421
0.17
35
1 0.
09
0.33
0.
24
1.08
2 0
.10
0.78
t 0
.09
L/P
C
ont.
302
0.02
29
6 0.
06
277
0.00
0.
08
0.06
3B
61
9 0.
08
565
0.35
37
0 0.
00
0.48
0.
35
1.23
C 0
.14
1.13
'-t
0.12
1
3E
46
4 0.
20
370
0.22
28
9 0.
00
0.35
0.
18
0.71
k 0
.14
0.45
2 0
.12
E/L
* C
ont.
127
0.14
10
8 0.
09
98
0.00
0.
23
0.09

3v
35
8 0.
04
344
0.16
28
8 0.
24
0.38
0.
34
1.07
i 0.
12
1.03
+- 0
.11
L/P
' 3
W
397
0.05
37
8 0.
35
244
0.00
0.
39
0.35
1.
11 rt
0.15
1.
07 -C
0.1
1 1
3L
34
7 0.
04
333
0.38
25
0 0.
00
0.39
0.
39
1.11
4 0
.12
1.23
4 0
.12
L'
Con
t. 31
6 0.
04
300
0.10
27
0 0.
01
0.15
0.
11
3K
30
6 0.
25
228
0.13
19
9 0.
03
0.37
0.
15
1.06
i 0.
15
0.45
k 0
.10
E/L
3
M
384
0.09
35
1 0.
44
232
0.03
0.
41
0.36
1.
22 4
0.1
2 1.
29 k
0.1
1 1
3N
22
1 0.
20
178
0.38
11
0 0.
00
0.50
0.
38
1.58
rt
0.15
1.
47 &
0.1
5 1
1
3R
19
1 0.
10
171
0.30
12
0 0.
06
0.41
0.
34
1.22
4 0
.16
1.21
i 0
.15
1
Con
t. 22
9 0.
09
271
0.04
25
9 0.
01
0.14
0.
05
2 F
48
7 0.
07
454
0.34
30
1 2
D
465
0.04
44
8 0.
28
324
Con
t. 28
1 0.
06
266
0.09
24
1
3F
59
1 0.
14
505
0.27
36
9 3
6
463
0.09
41
9 0.
26
310
3H
50
4 0.
09
457
0.31
31
4 C
ont.
441
0.04
42
3 0.
04
407
0.10
0.
00
0.00
0.00
0.
00
0.00
0.
00
0.45
0.
41
1.38
4 0
.11
1.33
4 0
.11
L/P
c,
0.30
0.
28
0.78
4 0
.11
0.85
i 0.
10
1
0.14
0.
09
M 0.
38
0.27
1.
28 i 0.
09
0.96
i 0.
08
1
H
0.38
0.
32
1.28
i 0.
10
1.16
2 0
.09
1
0.08
0.
04
H ;
0.33
0.
26
1.08
4 0
.09
0.92
i 0.
09
L'
z 6 2
A
410
0.16
34
4 0.
35
224
0.00
0.
46
0.35
1.
06 4
0.1
3 1.
13 -t
- 0.
12
1
2E
48
2 0.
13
417
0.38
25
7 0.
04
0.48
0.
41
1.14
4 0
.12
1.37
4 0
.11
1
2G
18
8 0.
18
156
0.32
10
6 0.
02
0.46
0.
35
1.06
? 0
.17
1.13
i 0.
16
1
2H
41
3 0.
31
286
0.19
23
2 0.
06
0.48
0.
24
1.14
-t-
0.13
0.
69 i 0.
12
E/L
0.
26
0.09
C
ont.
241
0.19
18
8 0.
08
173
0.01
' Mor
phol
ogic
al a
bnor
mal
ity.
0.
00 m
eans
mor
tali
ty l
ess
than
0.0
1.
w
w
W

TA
BL
E 2
Spon
tane
ous
leth
als from n
atur
al p
opul
atio
ns
3W
A
258
0 13
22
4 0.
29
147
0.00
0
43
0.34
1.
18 t 0
.14
1.30
4 0
.13
L'
Con
t.
296
0.16
24
8 0
03
243
00
0
0.18
0.
02
3 W
B
469
0.06
44
2 0.
37
279
0.01
0
41
03
7 1.
25 -C
0.1
0 1.
27 ?
0.1
0 1
3W
P
307
00
3
299
00
8
274
0.25
0
33
0.31
0.
93 ?
0.1
2 1.
03 &
0.1
1 P
' 3
°F
32
3 00
7 29
9 0.
33
199
0.00
0.
39
0.33
1.
17 t 0
.12
1.11
31
0.12
1
C
ont.
389
0.07
34
7 0.
07
323
0.00
0.
13
0.07
3 W
E
495
0.25
38
2 0.
28
262
0.01
0
4i
0.32
1.
19 +
- 0.
14
1.01
t 0
.12
E/L
C
ont.
127
0.14
10
8 0.
09
98
0.00
0
23
0.09
3w
c
Cou
t. 52
2 30
2 0.
10
0.02
47
2 29
6 0.
32
0.06
32
0 0.
08
277
0.00
0.
40
0.35
0.
08
0.06
1.
36 t 0
.10
2W
D
2W
H
2W
I
2W
J
Con
t.
2W
K
2W
L
2W
M
2W
N
2 w
P
Con
t.
513
806
336
102
306
0.18
0.
21
0.17
0.
03
0.13
419
636
280 99
26
1
0.27
0.
29
0.36
0.
26
0.03
306
0.01
45
4 0.
01
178
0.02
73
0.
08
254
0.00
0.41
0.
27
0.44
0.
29
0.48
0.
37
0.44
0.
32
0.17
0.
03
1.13
C 0
.08
1.25
C 0
.10
1.41
t 0
.13
1.25
4 0
.20
380
604
738
480
374
421
0 08
0.
17
0.04
0.
12
0.16
0.
03
350
504
708
421.
313
408
0.29
0.
28
0.38
0.
19
0.09
0.
12
247
0.01
36
2 0.
06
442
0.02
34
0 0.
15
285
0.23
35
8 0.
01
0.36
0.
31
0.44
0.
32
0.41
0
35
0.40
0.
32
0.41
0.
30
0.16
0.
13
0.96
t 0
.11
1.36
& 0
.10
1.16
? 0
.09
1.12
? 0
.10
1.16
t 0.
12
1.22
t 0.
10
L'
z 0.
99 &
0.0
9 L
/P
H
1.07
C 0
.08
1
1.39
4 0
.12
1
z ,.
L'
1.19
4 0
.19
0.85
& 0
.09
L'
0.89
& 0
.10
E/L
' 1.
01 * 0.0
8 1
E E L
/P
0.89
I
0.11
0.
81 ?
0.1
1 P
9P
34
6 0.
06
12 P
36
1 0.
16
20 P
25
9 0.
23
21 P
45
4 0.
07
29 P
39
3 0.
06
35 P
31
3 0.
07
42 P
33
9 0.
04
Con
t. 29
4 0.
11
326
0.35
21
2 0.
02
0.40
0.
37
1.09
4 0
.12
1.30
? 0
.09
1
303
0.31
21
0 0.
00
0.42
0.
31
1.17
& 0
.12
1.06
t 0
.11
1
200
0.08
18
4 0.
04
0.32
0.
11
0.77
& 0
.14
0.26
t 0
.10
E
423
0.28
30
4 0.
11
0.40
0
36
1.09
t 0
.11
1.26
& 0
.10
L'
368
0.15
31
3 0.
25
0.40
0.
36
1.09
? 0
.12
1.26
? 0
.11
P'
291
0.31
20
0 0.
03
0.38
0.
34
1.01
C 0
.13
1.18
t 0.
12
L'
320
0.33
21
3 0.
01
0.38
0.
34
1.01
t 0
.12
1.18
-C 0
.11
IJ '
261
0.06
24
5 0.
00
0.17
0.
06
40 P
32
8 0.
06
307
0.28
22
1 0.
03
0.36
0.
30
0.90
C 0
.13
0.87
4 0
.12
1
41 P
44
2 0.
08
406
0.33
27
2 0.
02
0.40
0.
34
1.06
& 0
.12
1.03
-C 0
.11
L'
Con
t. 27
6 0.
08
254
0.09
23
0 0.
02
0.18
0.
11
Mor
phol
ogic
al a
bnor
mal
ity.
a
0.00
mea
ns m
orta
lity
les
s th
an 0
.01.

TA
BL
E 3
X-r
ay in
duce
d le
thal
s
STA
GE
O
F
&,+
a,
&I,
v fa
DE
AT
H
TO
TA
L
CA
LC
UL
AT
ED
VA
LU
ES
OB
SER
VE
D N
UM
BE
RS
AN
D M
OR
TA
LIT
IES
MO
RT
AL
ITY
L
arva
l P
upal
E
gg
CR
RO
MO
- SO
ME
N
o.
Mor
t. No.
Mor
t.'
M
M'
a,
81."
N
o.
Mor
t.
x7
38
2 0.
07
355
0.06
33
5 0.
23
0.32
0.
28
0.83
-1-
0.11
0.
94 k
0.0
7 P
0.
15
0.06
C
ont.
328
0.09
29
7 0.
05
281
0.01
x4
47
7 0.
25
355
01
2
311
0.08
0.
40
0.19
1.
18 k
0.1
0 0.
56 -I
0.10
E
/L
Con
t. 39
2 0.
05
376
0.10
33
9 0.
00
0.14
0.
10
S 18
47
2 0.
03
457
0.24
34
8 0.
08
0 32
0.
30
0 80
& 0
.11
1.02
t 0.
10
L'
8 S
48
378
00
8
348
0.28
25
1 0.
02
0.35
0.
29
0.92
C 0
.11
0.98
& 0
.10
L
~
S 5
6 37
6 0
30
336
0.20
27
0 0.
10
03
5
0.27
0.
92 2
0.1
2 0.
90 k
0.1
0 L
' M
z E s
2
452
0.39
27
6 0.
04
264
0.03
0.
44
0.06
1.
19 k
0.1
1 0
11 2
0.0
7 E
8
37
338
00
6
319
0.31
21
7 0.
01
0 36
0.
32
0.87
? 0
.11
1.07
t 0
.11
L
Con
t. 47
2 0.
13
411
0.06
38
5 0.
01
0.19
0.
08
s 10
42
2 0.
06
399
0.26
29
6 0.
03
0.33
0.
28
0 99
k 0
.10
0.97
-1-
0 09
S
84
299
0.08
27
8 0.
12
242
0.21
S
105
335
0.11
29
8 0.
07
278
0.26
0
39
0.31
1.
23 -1
- 0.
11
1.09
-1-
0.11
P
6
S 12
6 30
5 0.
19
247
0.04
23
8 0.
03
0.25
0
07
0.67
? 0
.11
0.13
k 0
.07
Con
t. 50
1 0.
07
468
0.04
44
7 0.
01
0.11
0
05
Con
t. 37
3 0.
03
362
0.09
32
8 0.
03
0.15
0.
13
S 5
8 35
0 0.
10
315
0.28
23
1 0.
02
s 120
50
0 0.
16
418
0.20
33
5 0.
00
0.33
0.
20
0.93
k 0
.10
0.47
t 0
.09
Con
t. 45
7 0.
03
444
0.10
40
1 0.
00
0 13
0.
11
Q
Con
t. 32
1 0.
08
289
0.05
27
4 0.
01
0.16
0.
06
H
L
P 0
36
0
30
1.
11 2 0
.11
1.05
k 0
.11
I?' 5
E
D
494
0.28
35
4 0.
10
319
0.01
0.
36
0.10
0.
99 I+
0.1
0 0.
01 t 0
.08
E
G U 3! 0
u,
0
0.35
0.
28
1.01
k 0
.11
0.79
& 0
.11
L
+d
E/L
x, F
S
281
0.57
11
9 0.
13
104
0.16
0
69
0.28
0.
93 C 0
.14
0.40
2 0
.19
E'
1.03
-1-
0.13
0.
76 2
0.1
3 E
/L
Con
t. 32
2 0.
49
164
0.13
14
2 0.
12
0.61
0.
24
358
0.13
31
3 0.
29
222
0.03
0.
40
0.31
s
121
S 14
6 39
7 0.
12
349
0.30
24
3 0.
02
0.40
0.
32
1.03
C 0
.13
0.80
-1-
0.13
L
C
ont.
206
0.04
19
8 0.
14
171
0.02
0.
19
0.16
~
~
S 24
57
0 0.
71
168
0.20
13
5 0.
02
0.77
0.
20
1.31
-1-
0.09
0.
17 &
0.1
4 E
' w
S 1
59
756
0.87
99
0.
13
86
0.02
0.
89
0.16
1.
79 ?
0.1
3 0.
01 -1
- 0.
17
E'
k
Con
t. 54
6 0.
45
300
0.18
24
6 0.
02
0.59
0.
21
Con
t. 15
3 0.
48
80
0.19
65
0.
03
0.59
0.
21
Mor
phol
ogic
al a
bnor
mal
ity.
a
0.00
mea
ns m
orta
lity
les
s th
an 0
.01.

342 M. T. M. RIZKI
lethal samples. A possible explanation for these results would be the presence of more than one lethal on the tested chromosome. When a chromosome is tested for its concealed genetic variability by the series of crosses discussed above, there is only a rare chance that crossing-over can occur in the chromosome under test. Since single males are mated in step B (fig. l), and the inversion in the balancer chromo- some suppresses crossing-over in the successive matings, the chromosome bearing the investigated lethal factor be-
TABLE 4
Comparison of Eethals from laboratory and natural populations
BTAQE
E E/L L p P Total
Laboratory No. observed 1 + 4 19 4 1 29
No. expected 4.38 19.15 3.28 2.19 - - 5
Wild population No. observed 1 + 2 16 2 3 24
No. expected 3.62 15.85 2.72 1.81 = 3
Total 8 35 6 4 53
Comparison o f laboratory with wild population xa 3 d.f. = 1.97 P 1 0.60
haves as a single Mendelian unit. However, this chromosome may be carrying two o r more lethal factors separable by crossing-over, a condition undetectable by the method of extraction of the lethal chromosomes. For determining the stage of death due to autosomal lethals in the present study it is necessary to outcross the lethal stock to the Standard strain. In so doing, the lethal-bearing chromosome will have a chance of exchanging its genetic variability once it has been removed from association with the crossing-over sup- pressor. Slizynski ( '42) suggested that some x-ray and ultra-

GEBETIC LETHALITY 111’ DROSOPHILA 343
violet-induced lethals may contain more than one lethal change, whereas multiple changes do not seem to occur in spontaneous lethals. Such high mortality may also be due to the presence of translocations. Since this type of effect is not comparable to the spontaneous changes in the hereditary norm, these three lethals are excluded from the sample of x-ray material used for comparison with spontaneous lethals.
The remaining 19 induced lethal chromosomes caused death at the following stages: 6 egg, three egg-larval, 7 larval, no larval-pupal, three pupal lethals.
TABLE 5
Comparison of spontaneous and induced lethals
Spontaneous No. observed 2 6 35 6 4 53 No. expected 5.89 6.62 30.91 4.42 5.15
Induced No. observed 6 3 7 0 3 19 No. expected 2.11 2.38 11.09 1.58 1.85
Total 8 9 42 6 7 72
Comparison of spontaneous with induced xa 4 d.f. = 15.14 P - 0.005
The ontogenetic distribution of death is different in spon- taneous and x-ray induced lethal Mendelian factors. The x2 test in table 5 shows that this difference is significant,
DISCUSSION
In 1939 Medvedev reported on a developmental analysis of 30 spontaneous sex-linked lethals in D. rnelavzogaster. He found that two lethals caused death in the egg stage, 10 lethals caused death during the larval period (9 in the first instar and one in the second), and 15 caused death in the prepupal and pupal stages. Medvedev concluded that there

344 31. T. M. RIZHI
are definite sensitive periods during development: the first occurs during embryonic development, the second during the first larval instar and the third at the time of pupation.
Hadorn ( ’48) tabulated the data collected by several authors concerning the time of death for lethals of D. melunoguster. No distinction was made among lethals of different origin; both spontaneous and x-ray induced lethals were included. According to Hadorn, the distribution of time of death is not a uniform one, “there being a clustering at four develop- mental periods. ” These periods are designated as sensitive phases, and the intervals during which no lethals have been found to die are the insensitive interphases. The first sensi- tive period is the early phase of embryonic development. A re-examination of the data shows that two of the lethals which die at this time, Nullo-x and N-8, are spontaneous chromosomal aberrations, while the remainder are induced by x-rays. The second sensitive period occurs at the com- pletion of embryonic development and in the early first larval instar. A group of 10 spontaneous lethals and two x-ray induced lethals were found to die at this stage. The third sensitive period occurs at the end of larval life and the begin- ning of the pupal period; 7 spontaneous lethals are listed for this period. Death of the last group of 9 lethals occurs during the late pupal stage or after emergence. All but two of these are spontaneous lethals.
The two samples of spontaneous autosomal lethals of D. willistomi analyzed in the present study show a low frequency of egg lethals as compared with a high frequency of larval lethals. The results on the whole agree with Hadorn’s scheme of sensitive periods. There are many egg-larval and first larval instar lethals among the spontaneous lethals analyzed. Another period of high mortality occurs during the late third larval instar and at the time of pupation. Finally, there are lethals which cause death during the late pupal period. However, only two egg lethals were found in the sample of 53 spontaneous lethals; both of these lethals die in the late embryonic period, apparently as completely de-

GENETIC LETHALITY I N DROSOPHILA 345
veloped larvae, and would be classified, according to Hadorn, under the second sensitive period of development. These observations on spontaneous lethals of D. willistonni do not support the idea of a sensitive period during early embryo- genesis.
Oster (’51) analyzed 43 ultraviolet-induced lethals of D. rnelauzogaster. Of these 4 died during the late embryonic period, 21 during the first larval instar, 7 during the third larval instar, 8 during the early pupal stage, and three during the late pupal period. Thus relatively few egg lethals are found among ultraviolet-induced lethals while there are many larval lethals. The distribution of the stage of death of ultra- violet-induced lethals appears to be similar to that found in spontaneous lethals of D. willistouzni.
Hadorn ( ’48) considers the second larval instar an example of an insensitive period, since almost no genetic lethality has been recorded for that period. Death in one spontaneous and one x-ray induced lethal in the present study occurred definitely during the second larval instar. Sivertzev-Dob- zhansky ( ’27), Medvedev ( ’39), and Brody ( ’40) also re- corded second instar larval lethals in D. melanogaster. Hadorn (’48) states that discrepancies with regard to phase-speci- ficity may be due to the presence of modifying factors, and “failure of phase specificity can therefore only be considered as proved if the residual heredity is made isogenic.” The material used in the present study was not isogenic, as already mentioned, since the intention was to examine the influence of lethal factors on ontogeny as it occurs in natural popu- lations. Referring to figure 7 , none of the larval instars of D. wiEEistovzi can be considered completely insensitive periods. Death due to lethal factors may occur mainly during one larval instar, or lethal factors may cause death during two or more larval instars.
The developmental analysis of the x-ray induced lethal factors shows an increase in the proportion of egg lethals as compared to the spontaneous lethal factors. All of the egg lethals found in this sample of x-ray material cause

346 M. T. M. RIZKI
death during early embryonic development. These 1etha.ls may be representative of the first sensitive period found by Hadorn.
No cytological information about the spontaneous and induced lethal factors analyzed in the present study is availa- ble. In view of the differences in the distribution of time of death of x-ray induced and spontaneous lethal factors, knowl-
I I I I
I
I I
I
I LI l L 2 I L 3 [ P I I I
I I
I I
I I I
I I I I I I ,
I t
I
I
ILl-2-3{ I I I
I I L 2 - 3 I I
I E G G I IST INSTAR12ND INSTAR 13RD INSTAR1 PUPAL
Fig. 7 Diagrammatic representation of the stage of death of different spon- taneous lethals. Each black rectangle which does not cross the stage boundaries dotted lines) represents a stage-specific lethal. Lethals dying in more than one stage are shown by bars which cross the stage boundaries.
edge of the salivary chromosome structure of these lethals would have been helpful in evaluating the results. However, the correlation of chromosomal changes with genetic lethal effects is complex. There are known cytological deficiencies which act as lethal factors (Slizynska, '38) ; lethal genetic deficiencies occur without any visible cytological changes (for example, N 264-8 in D. melarzogaster, described by Slizynska, '38) ; and finally there are cytological deficiencies which are not lethal (Demerec and Hoover, '36).

GENETIC LETHALITY I N DROSOPHILA 347
Altenburg ( '30, '31) convincingly demonstrated that ultra- violet produces gene mutations in Drosophila. This obser- vation was confirmed by Muller and Mackenzie ('39) and Mackenzie and Muller ( '40), who, in addition, showed that x-rays produce gross structural changes in Drosophila whereas ultraviolet is ineffective in causing aberrations. Stadler ( '39, '41) and Stadler and Sprague ('36) have also concluded that there are differences in genetic responses to x-ray and ultraviolet treatment in maize, Sacharov ( '35, '36) reported on cytological studies of 27 sex-linked lethals induced by x-ray treatment and 25 spontaneous lethals, He found 1 7 chromosomal aberrations and 4 deficiencies among the x-ray induced material while the spontaneous lethals were not correlated with any deficiencies or aberrations. Slizynski ( '38), on the other hand, found no great difference between spontaneous and x-ray induced lethals: 47.3% of the spon- taneous and 30% of the induced lethals were deficiencies. I n 1942, Slizynski analyzed a group of induced and spontaneous lethals in D. meZawogaster. Among 11 x-ray induced lethals, two were deficiencies, three translocations, and two inversions. Out of 21 cases of ultraviolet-induced lethals 5 were defi- ciencies, but no gross structural changes were found. There was one deficiency and one probable inversion among 8 spontaneous lethals. Dobzhansky ('51) found only two newly arisen spontaneous structural changes in an examination of at least 100,000 larvae of D. pseudoobscura.
The general conclusion derived from studies on induced and spontaneous lethals is that the x-ray material contains a larger per cent of gross structural changes than spontaneous and ultraviolet-induced lethal material.
The work of Poulson ('40, 45) on the various Notch de- ficiencies, white, Half-s and Nullo-x, all of which die in the early embryonic period, shows that the larger the deficiency the greater the chance of death taking place in early develop- ment. The increase in the number of egg lethals among the x-ray induced lethals as compared to the spontaneous lethals may be due to the presence of deficiencies and gross struc-

348 M. T. M. RIZKI
tural chromosomal changes in the x-ray material. I n addition to high mortality in the egg stage found in the induced lethals, S1,,, S,, and S, there is also a postembryonic effect in the offspring of these lethal and control crosses. Li ( '27) has noted such a maternal effect associated with some chromo- somal deficiencies.
Spontaneous lethals occurring in nature and in the labora- tory possess similar ontogenetic distributions of time of death. It has been shown that some lethals die in a strictly stage-specific manner while other lethals extend their effect over several stages of development. Few lethal factors (two out of 53) cause death during the embryonic period, and these are found in the late stages of embryonic development. This distribution pattern of lethality due to spontaneous lethals is probably a property of the species genotype. During the course of evolution, the reactive system of D. willistoni has been conditioned in such a manner that early embryo- genesis has a maximum amount of protection against the drastic effects (death) of lethal mutations. Early embryo- genesis is a very complex process (Demerec, '50) ; hence, a pattern of gene interaction which tends to protect the em- bryo against developmental upset would have a high adaptive value since the species seems to be exposed to the continual occurrence of lethal mutations.
SUMMARY
1. The time of death was studied in 75 recessive auto- soma1 lethals in Drosophila willistoni. Of these, 24 were found in natural populations of certain localities in Brazil, 29 were obtained in untreated laboratory cultures, and 22 were ob- served in the progeny of x-ray treated flies.
2. The lethals found in nature and those arising spon- taneously in laboratory cultures appear to be similar with respect to the developmental stages at which death OCCUFS.
Most frequently such lethals cause death of the larvae, although some egg lethals and pupal lethals also occur.

GENETIC LETHALITY IN DROSOPHILA 349
3. The x-ray induced lethals cause death in the egg stage relatively more frequently than do spontaneous lethals, al- though some of the former also kill the lsarvae an'd pupae. 4. The problem of sensitive periods of ontogeny and the
evolutionary aspects of the nature of the reactive system as revealed by the distribution of time of death of lethal factors have been discussed.
LITERATURE CITED
ALTENBURG, E. 1930 The effect of ultraviolet radiation on mutation. (Abstr.)
1931 Genetic effects of ultraviolet radiation. (Abstr.) Aaat. Rec.,
BRODY, G. 1944 The time of effect of some second chromosome lethals in DrosophiEa metanogaster. Genetics, 25 : 645-650.
DEMEREC, M., Editor 1950 Biology of Drosophila. John Wiley and Sons, Inc., New York.
DEMEREC, M., AND M. E. HOOVER 1936 Three related X-chromosome deficiencies in Drosophila. J. Heredity, 27: 207-212.
DOBZHANSKP, TH. 1951 Genetics and the Origin of Species. Third Ed. Colum- bia University Press, New York.
DOBZHANSKY, TH., A. M. HOLZ AND B. SPASSKY 1942 Genetics of natural populations. VIII. Concealed variability on the second and the fourth chromosomes of Drosophila pseudoobscura and its bearing on the prob- lem of heterosis. Genetics, 2 7 : 463-490.
DUBININ, N. P. 1946 On lethal mutations in natural populations. Genetics,
HAWRN, E. 1948 Gene action in growth and differentiation of lethal mutants
1951 Developmental action of lethal factors in Drosophila. I n M. Demerec, Ed. Advances in Genetics IV. Academic Press, New York.
HODSON, A. C., AND H. C. CHIANG A new method for rearing Drosophila. Science 107, No. 2772: 176-177.
LI, Ju-CHI 1927 The effect of chromosome aberrations on development in Drosophilu melunogaster. Genetics, 12 : 1-58.
MACKENZIE, K., AND H. J. MULLER 1940 Mutation effects of ultraviolet light in Drosophila. Proc. Roy. SOC. Lond., Series B, 129: 491-517.
MEDVEDEV, N. N. 1939 Studies in developmental genetics. C. R. Acad. Sci. U. R. 8. S., 22: 344-347.
MULLER, H. J., AND K. MACKENZIE 1939 Discriminatory effect of ultraviolet rays on mutation in Drosophila. Nature, 143: 83.
OSTER, I. I. An analysis of ultraviolet-induced lethals due to gene muta- tion. D. I. S., 25: 124-126.
€3. SPASSKY AND M. WEDEL 1951 Concealed genic variability in Brazilian populations of Drosophilu willistoni. Genetics, 36 : 13-30.
Anat. Ree., 4 7 : 383.
51, Suppl.: 108-109.
31: 21-38.
of Drosophila. Symp. SOC. Exp. Biol., 2 : 177-195.
1948
1951
PAVAN, C., A. R. CORDIERO, N. DORZHANSKP, TH. DOBZHANSKP, c. MALWOLOWKIN,

M. T. M. RIZKI
POULSON, D. F. 1940 The effects of certain X-chromsome deficiencies on the embryonic development of Drosophila melanogaster. J. Exp. Zoo].,
1945 Chromosomal control of embryogenesis in Drosophila. Am.
A cytological study of the lethals of the chromosomes
1936 On the specificity of the action of factors causing mutations.
SIVH;~TZEV-DOBZHANSKY, N. P. Uber den letalen Effekt einiger Gene bei
SLIZYNSKA, H. 1938 Salivary chromosome analysis of the white-facet region
SLIZYNSKI, €3. M. 1938 Salivary chromosome studies of lethals in Drosophila
1942 Deficiency effects of ultraviolet light in Drosophila melano-
83: 271-325.
Nat., 79: 340-363.
in Drosophila melanogaster. Doklady Akad. Nauk., 4 : 70-71.
Bull. Biol. Med. Exp., 1: 202-203.
Drosophila melanogaster. Arch. Entwmech. Org., 109 : 535-548.
of Drosophila melanogmter. Genetics, 83 : 291-299.
melanogaster. Genetics, 23: 283-290.
gaster. Proc. Roy. SOC. Edinb., 61 : 297-315.
SACHAROV, V. V. 1935
1927
SPASSKY, B. 1943 STADLEE, L. J. 1939 Genetic studies with ultraviolet radiation. Proc. Seventh
The comparison of ultraviolet and X-ray effects on mutation.
Genetic effects of ultraviolet radiation
Cream of wheat-molasses fly medium. D. I. S., 17: 67-68.
Intern. Genetics Congress, p. 269-276.
C. S. H. Symp. Quant. Biol., 9: 168-177. 1936
in maize. Proc. Nat. Acad. Sci., 2 2 : 572-591.
1941
STADLER, L. J., AND G. F. SPRAGUE




















