
Ongoing activity in peripheral nerves: The physiology and pharmacology of impulses originating from...
14
EXPERIMENTAL NEUROLOGY 43, 580-593 (1974) Ongoing Activity in Peripheral Nerves: The Physiology and Pharmacology of Impulses Originating from a Neuroma ’ PATRICK D. WALL AND MICHAEL GUTNICK Department of Anatomy, University College, London, England, Department of Zoology, Hebrew University, Jerusalem, and Department of Neurology, Rotschild Hadassah University Hospital, Jerusalem, Israel Received February 1, 1974 In rats, the sciatic nerve was cut, drawn into a polythene tube with one end sealed, and a neuroma allowed to develop in the chamber. Activity in the dorsal and ventral roots terminating in the neuroma was examined be- tween 9 days and 4 months after the section. A fraction of the small myelinated a&rent fibers originating in the neuroma were carrying a steady ongoing barrage of nerve impulses. Many of the fine terminals were excited by slight mechanical distortion. The fine sprouts in the neuroma were elec- trically excitable. The ongoing afferent barrage was highly dependent on blood flow. It was abolished for long periods of time after a brief antidromic tetanus had invaded the active fibers. This suggests that some of the pain relief obtained from peripheral nerve stimulation may have a peripheral rather than a central mechanism. No signs of excitatory on inhibitory in- teraction were detected between volleys in one group of nerve fibers and the activity in other groups of fibers in the neuroma. Alpha active sympathetic amines, noradrenaline, excited the ongoing activity while beta agents, iso- prenaline, had no excitatory effect. This suggests that alpha blocking agents might be useful to test if the sympathetic system is involved in particular pains. INTRODUCTION When a peripheral nerve is cut, the sequence of degeneration followed by regeneration begins. The axons sprout and, if they fail to penetrate along the course of the original curve, they form a neuroma (4, 18). Human subjects report four sensory phenomena after a nerve has been cut. First, the part supplied by the nerve will be anesthetic but there may be a power- ful impression that the part still exists as a phantom. This phenomenon is due to an absence of nerve impulsessince it can be immediately imitated by local anesthetic block of normal nerve (13). Second, the cut nerve end and 1 This work was supported by the Medical Research Council and the Sloan Foundation. 580 Co yri ht 0 1974 by Academic Press, Inc. Alfrig%ts of nprcduction ia my form reserved.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Ongoing activity in peripheral nerves: The physiology and pharmacology of impulses originating from...

EXPERIMENTAL NEUROLOGY 43, 580-593 (1974)
Ongoing Activity in Peripheral Nerves: The Physiology and Pharmacology of Impulses Originating from a Neuroma ’
PATRICK D. WALL AND MICHAEL GUTNICK
Department of Anatomy, University College, London, England, Department of Zoology, Hebrew University, Jerusalem, and Department of Neurology, Rotschild Hadassah University Hospital, Jerusalem, Israel
Received February 1, 1974
In rats, the sciatic nerve was cut, drawn into a polythene tube with one end sealed, and a neuroma allowed to develop in the chamber. Activity in the dorsal and ventral roots terminating in the neuroma was examined be- tween 9 days and 4 months after the section. A fraction of the small myelinated a&rent fibers originating in the neuroma were carrying a steady ongoing barrage of nerve impulses. Many of the fine terminals were excited by slight mechanical distortion. The fine sprouts in the neuroma were elec- trically excitable. The ongoing afferent barrage was highly dependent on blood flow. It was abolished for long periods of time after a brief antidromic tetanus had invaded the active fibers. This suggests that some of the pain relief obtained from peripheral nerve stimulation may have a peripheral rather than a central mechanism. No signs of excitatory on inhibitory in- teraction were detected between volleys in one group of nerve fibers and the activity in other groups of fibers in the neuroma. Alpha active sympathetic amines, noradrenaline, excited the ongoing activity while beta agents, iso- prenaline, had no excitatory effect. This suggests that alpha blocking agents might be useful to test if the sympathetic system is involved in particular pains.
INTRODUCTION
When a peripheral nerve is cut, the sequence of degeneration followed by regeneration begins. The axons sprout and, if they fail to penetrate along the course of the original curve, they form a neuroma (4, 18). Human subjects report four sensory phenomena after a nerve has been cut. First, the part supplied by the nerve will be anesthetic but there may be a power- ful impression that the part still exists as a phantom. This phenomenon is due to an absence of nerve impulses since it can be immediately imitated by local anesthetic block of normal nerve (13). Second, the cut nerve end and
1 This work was supported by the Medical Research Council and the Sloan Foundation.
580
Co yri ht 0 1974 by Academic Press, Inc. Alfrig%ts of nprcduction ia my form reserved.

PERIPHERAL NERVES 581
the regenerating part becomes sensitive to mechanical distortion, giving rise to the Tine1 sign which may vary from a minor shooting sensation to frank pain. Third, the subject may experience a continuous sensation varying from mild paresthesias to serious pain. The Tine1 sign and these pains and paresthesias must originate as nerve impulses from the damaged nerve since they are temporarily abolished by local anesthesia (16, 26). Lastly these afferent nerve impulses may be influenced by activity of the autonomic system since the intensity of the sensations may be markedly influenced by increases or decreases of sympathetic activity (26).
The anatomy of degeneration, regeneration and neuroma formation has been extensively studied by light microscopy (4) and more recently by electron microscopy (1, 15, 17). Within l-2 days, multiple unmyelinated sprouts with diameters of less than 0.5 F grow out from the cut axons. If these fail to enter the distal Schwann cell tubes, a mass forms contain- ing Schwann cells fibrous tissue, blood vessels and skeins of fine un- myelinated axons which may extend 1-2 mm from the parent axon. Even two years after the section, some of these sprouts have growth cones at their ends while others do not. This has been taken to suggest that active probing and withdrawal of the sprouts continues even when no adequate receiving end stations are located by the sprouts (17). This may be so, but we now know that certain growing fibers do not have a terminal growth cone (12). Therefore the presence and absence of growth cones does not necessarily imply growth and withdrawal.
In spite of the clinical and basic interest of the activity of the fine fibers and the terminals in the usual environment of a neuroma, no physiological or pharmacological studies have been carried out. Immediately on crushing or cutting axons, there is a vigorous injury discharge which dies down in seconds and is followed by silence which lasts for at least some hours (23, 2.5). Kirk (personal communication) (10) reports that if a peripheral nerve is sectioned close to the dorsal root ganglion, spontaneous activity is generated in afferent fibers originating in the dorsal root ganglion itself some hours after the nerve section. The only published paper on the physiol- ogy of regeneraitng nerve of which we are aware (7) does not report spon- taneous ongoing activity but shows that the growing fibers are unduly sensi- tive to both mechanical distortion and to acetylcholine. We have therefore investigated the activity in fibers originating from a neuroma.
METHODS
Neuromas were prepared in 30 adult rats, Hebrew University strain, weighing 250-450 g. Under anesthesia, the sciatic nerve of one leg was exposed, dissected free and sectioned distal to the head of the femur. A suture was tied around the central cut end of the nerve and S-8 mm of the

582 WALL AND GUTNICK
sciatic was drawn into a snug fitting medical polyethylene tube (internal diameter 1.4 mm, outside diameter 2.0 mm). The distal end of the tube was heat-sealed with the suture running through the seal. This procedure formed a chamber containing proximally the normal vascularized nerve and a sealed distal end into which sprouts and neuroma tissue could grow without being able to escape. The chamber containing the cut end of the nerve was replaced in the animal, the incision was closed in layers and the neuroma was allowed to develop. The nerve fibers originating from the neuroma were examined in acute experiments at times varying between 9 days and 4 months after the nerve section. The animals were anesthetized with urethane (750 mg/kilo) paralyzed with gallamine and maintained on artifi- cial respiration. The neuroma in its chamber was exposed as was the intact sciatic nerve on the other side and stimulating electrodes installed, Fig. 1.
R2
FIG. 1. Diagram showing the experimental neuroma of the sciatic nerve in its chamber and the stimulus points Sl-6 and recording points R on a dorsal root filament and R2 on a ventral root filament.
The exposure of the cauda equina is difficult in the rat and is best achieved with miniature sharp triangular rongeurs. Stimulating and recording elec- trodes were placed on dorsal and ventral roots in an oil pool on the normal and neuroma side as shown in Fig. 1. The stimulation and recording equip- ment was conventional.
At the end of the experiment the sciatic nerve and its neuroma were fixed in formalin and stained by Bodian’s protargol method. The appearance was that classically described, with bundles of fine unmyelinated fibers meander- ing through disorganized connective tissue. Some bundles curved back into the connective tissue sheath of the nerve. In a few cases, the nerve had
PERIPHERAL NERVES 583
separated from the suture and withdrawn from the chamber so that the neuroma formed outside the chamber with penetrating strands invading nearby muscle. In some cases tension on the nerve produced a long (2.5 mm) tapering neuroma and here the bundles of fine fibers ran in mainly parallel streams from normal nerve to the tip of the neuroma.
RESULTS
Clinical Examination. The distal musculature of the leg on the side of the sciatic section was paralyzed and wasted. The lateral two-thirds of the foot appeared anesthetic since there was no response to pinch or pin prick. In 9 of the 30 animals, the animal had mutilated the anesthetic part of its foot. This gnawing of the anesthetic zone began as early as 9 days after the nerve section. This phenomenon has been reported (2) and it may imply that the animal is suffering from some form of anesthesia dolorosa or phantom pain. When it was observed, the animals were immediately anes-
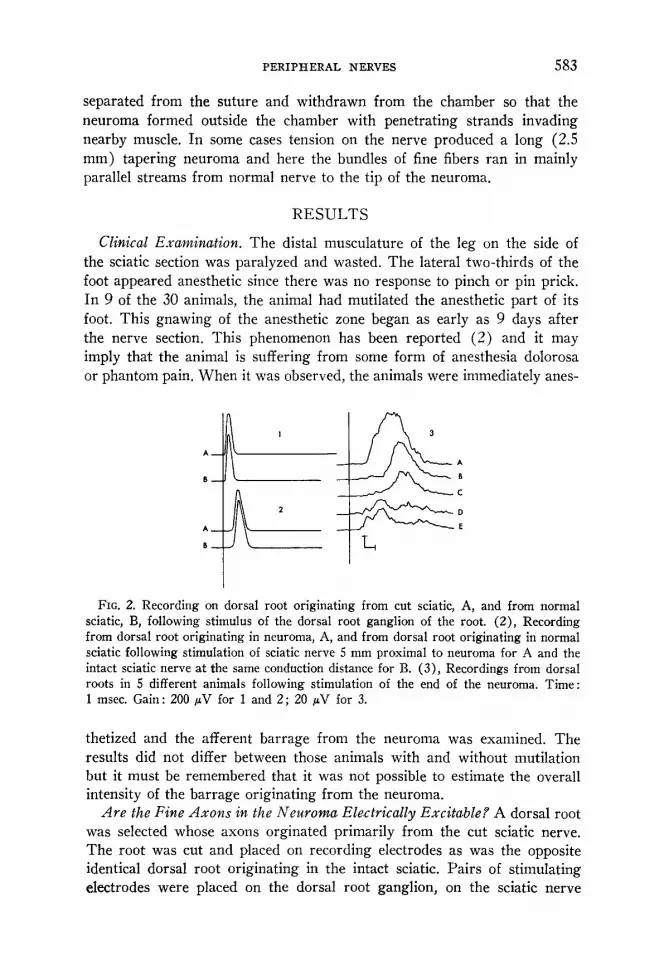
FIG. 2. Recording on dorsal root originating from cut sciatic, A, and from normal sciatic, B, following stimulus of the dorsal root ganglion of the root. (Z), Recording from dorsal root originating in neuroma, A, and from dorsal root originating in normal sciatic following stimulation of sciatic nerve 5 mm proximal to neuroma for A and the intact sciatic nerve at the same conduction distance for B. (3), Recordings from dorsal roots in 5 different animals following stimulation of the end of the neuroma. Time: 1 msec. Gain: 200 pV for 1 and 2; 20 pV for 3.
thetized and the afferent barrage from the neuroma was examined. The results did not differ between those animals with and without mutilation but it must be remembered that it was not possible to estimate the overall intensity of the barrage originating from the neuroma.
Are the Fine Axons in the Neuroma Electrically Excitable? A dorsal root was selected whose axons orginated primarily from the cut sciatic nerve. The root was cut and placed on recording electrodes as was the opposite identical dorsal root originating in the intact sciatic. Pairs of stimulating electrodes were placed on the dorsal root ganglion, on the sciatic nerve

584 WALL AND GUTNICK
just proximal to the chamber containing the neuroma and on the end of the neuroma Fig. 1: Sl, 2, and 3. The tip of the neuroma was stimulated by pushing two 26 gauge hypodermic needles through the wall of the chamber. Tracings of the results (Fig. 2) show that there was no significant differ- ence between the normal and abnormal sides of the compound action poten- tials generated by stimulating either on the dorsal root ganglion, Fig. 2, lA, B, or on the sciatic nerve proximal to the neuroma chamber, Fig. 2, 2A, B. The conduction distance from dorsal root ganglion to recording site was 13 mm and the fastest fibers conducted at 59 m per sec. In Fig. 2, 2A, B, the conduction distance was 4.5 mm and the fastest conduction velocity was 54 m per set both from the intact nerve and from the nerve with the neuroma. However, if the stimulus point was moved to the tip of the neuroma a grossly slowed and scattered compound action potential was always recorded. This stimulus point was only 5 mm distal to the stimulus point producing the fast compound action potential shown in Fig. 2, 2. Examples of the slow action potentials from 5 different animals are shown in Fig. 2, 3 A-E. Obviously the question is whether this delayed wave represents a group of slowly conducting fibers which send sprouts into the neuroma or whether it is produced by .action potentials in large diameter afferents which terminate in very fine extensions. That the latter possibility is the case was proved by first stimulating 5 mm proximal to produce this high velocity wave shown in Fig. 2, 2A, and then 1 msec later stimulating
at the tip of the neuroma. The result is complete abolition of the wave from
the neuroma stimulation. Evidently the fibers which carry the slow wave were made refractory by the first stimulus of the intact central myelinated
fibers and therefore must be continuations of the same fibers. The maximum
overall conduction velocity of the impulses over the distance between the tip
of the neuroma and the proximal end of the chamber is 6.5 m per set in the case of 3A and 2 m per set for 3B. However this estimate has little meaning
for three reasons. In the proximal end of the chamber, the fibers are of relatively normal diameter while at the distal end they are fine tapering
and branching. Therefore an impulse will accelerate as it moves proximally. Second, it is not possible to know the real conduction distance since the fibers meander markedly in the neuroma. Third, it is not possible to give
a maximal stimulus since as the stimulus current spreads from the tip, it is likely to stimulate more and more proximal therefore larger, therefore lower
threshold fibers. The nerve fibers immediately to the section are shrunken and changed (1.5). However, this change is not sufficient to explain the striking slowing of impulses in the neuroma region. In spite of these diffi-
culties it is clear that the fine fibers are electrically excitable and that the impulses are very slowly conducted.

PERIPHERAL NERVES 585
Is There Ongoing Activity Comnipzg from the NW-OWMX? Recordings of single units were made by dissecting fine filaments from dorsal and ventral roots. No spontaneous activity was observed in ventral root filaments originating in the neuromas. However dorsal root filaments consistently showed ongoing activity in the absence of any intentional stimulus. It was obviously of considerable importance to show that these afferent impulses originated from the neuroma and did not come from normal afferents. This was done in three ways. Individual units were isolated from multiunit re- cordings by a spike height discriminator. Units with ongoing activity could be fired by electrical stimuli applied directly to the tip of the neuroma in its chamber. Stimuli applied to surrounding tissue did not modify the firing of these spontaneous active units. This test by itself was not adequate because electrical stimulation of the neuroma also produced action potentials in many other fibers which did not have ongoing activity. A more delicate stimulation ‘test was to show that the ongoing activity was increased by gentle pressure on the neuroma chamber but not by more vigorous probing of surrounding tissue. Finally it was shown that ongoing activity was abolished if the neuroma chamber was perfused with 2% lidocaine. This was accomplished by placing two 26 gauge hypodermic needles at either end of the chamber and slowly injecting lidocaine into one and allowing it to escape from the other end. The frequency of ongoing activity for each unit was in the range of 0.5-5Hz. The spike heights of the spontaneously active units was small, less than one-third the height of the spindle afferent
spikes which are always recorded in normal root filaments. This implies that
large myelinated afferents were not spontaneously active. The technique
used did not allow the recording of unmyelinated single units. We can con- clude that a minority of the small myelinated afferents were spontaneously
active.
The Effect of Pressure on Ongoing Activity. Pressure was applied to the neuroma by distortion of the chamber either with externally applied
forceps or by perfusion with Ringer-Locke solution. As the pressure was
applied, a large number of fibers in birth dorsal and ventral rootlets became
active. However if the pressure was maintained, all of the dorsal root fibers
with large spike heights, most of the units with small spike heights, and all of the ventral root fibers became silent within seconds. These were not
injury discharges, since gentle repeated distortions produced repeated brief bursts of activity.
The Effect of Electrical Stiwmhtion on Ongoing Activity. It was noticed
that a silent period of ongoing activity followed a single electrical stimulus applied to the neuroma. Since direct stimulation of a region of impulse
generation may have a number of direct and indirect effects on the mem-

586 WALL AND GUTNICK
brane, this study was refined by investigating the effect of antidromic impulse invasion into the neuroma. The conditioning stimulus was applied to the cut end of the dorsal root filament (S4, Fig. 1). Bipolar recordings of ongoing activity were made on a more distal part of the filament (R, Fig. 1). The reason for this arrangement was to prevent any possibility that the fibers were blocked at the stimulus point itself. A single supra- maximal shock applied to the filament produced the antidromic wave passing the recording electrodes and was then followed by a period of Z-300 msec during which no ongoing afferent activity arriving from the neuroma was recorded. Far more dramatic decreases of activity were observed if the recording rootlet was stimulated tetanically. The standard conditioning stimulus was a maximal stimulus given as 0.1 msec square waves at 100 Hz for 6 sec. Figure 3 shows four examples of the effect of such repetitive
FIG. 3. Rate of ongoging activity in dorsal root filaments. 1 from normal intact nerve and Z-5 from neuromas. Combined rate of 4-6 single units. At the vertical line, the rootlet, from which recordings were taken, was stimulated for 6 sets with a maximal stimulus square wave 0.1 msec duration at 100 Hz. Time mark: 12 sec. Rate mark: 30 impulses per sec.
antidromic invasion of neuromas and one example of the effect on normal endings. The ongoing activity of normal afferents returns to normal within a few seconds of the end of such a tetanus. However, in the case of impulses originating in neuromas there was always a prolonged depression always lasting over 1 min and in some cases for over 1 hr. The records shown were intentionally multiunit recordings in which the summed rate of firing of 4-6 fibers was recorded so that the response of a population could be observed. In record ‘2, the firing rate of the population returned to normal in 76 set, in record 3, 100 set, in record 4, 120 set, and in record 5, the

PERIPHERAL NERVES 587
firing did not return to normal until 70 min later. When single units were observed it was found that if the tetanic stimulus was sufficiently strong to produce antidromic impulses in the observed fiber, then, after the tetanus, the orthodromic firing ceased or had a greatly decreased frequency for long periods as seen in the multiunit records.
Tetanic invasion of central terminals is known to produce a prolonged hyperpolarization (21, 24). Therefore it is possible that the reason for the cessation and decrease of firing observed here was due to an increased threshold of the fine fibers in the neuroma. To test this the electrical excit- ability of the neuroma was measured by a modification of the technique used to detect primary afferent membrane potentials (19). A submaximal shock was given once per 2 set at the neuroma (Sl, Fig. 1) and the com- pound orthodromic action potential was recorded on the rootlet (R) . Then the standard antidromic tetanus was given on the cut end of the recording rootlet. Ongoing activity was recorded as was the height of the compound action potential. It was found that the compound action potential was de- creased, the threshold was raised for some time after the tetanus. However, this decrease of excitability of the neuroma did not last as long as the
FIG. 4. Rate of ongoing activity in dorsal root filament originating in neuroma. Combined rate of 3-6 units. At the arrow, 5 JLG adrenaline injected iv. Time mark: 12 sec. Rate: 30 impulses per sec.
decrease of the ongoing activity. In the six cases tested, the excitability returned ‘to normal in 50-110 set while the spontaneous activity did not return to its normal rate for 1201360 sec.
The Effects of Blood Flow on Ongoing Activity. At the end of the experiments, when the animal was killed, it was noticed that ongoing activity from the neuroma ceased within seconds of the loss of blood flow whereas normal endings continue to fire for minutes.
The Efect of Sywspathetic Awines on Ongoing Activity. It is known that
in certain cases of nerve damage, the sympathetic nervous system plays some role in pain mechanisms, since anesthetic blockade or surgical removal of the sympathetic ganglia decreases the pain. In two animals, we perfused the neuroma chamber with adrenaline and noticed prolonged changes of firing. However, we decided not to pursue the development of the technique and returned instead to the more conventional intravenous injection of compounds, in this case through a cannula in the jugular vein. It should be remembered that these animals were under urethane and gallamine and

588 WALL AND GUTNICK
therefore the ongoing activity of their sympathetic system was likely to be low. Figure 4 shows the delayed and prolonged effect of intravenous adren- aline 5 pg. In this case, the firing returned to its control level after 5 min. In this and in other drug-produced excitations, the increased population firing rate was produced in part by an acceleration of ongoing activity in active units and also by the appearance of firing in previously silent units. Again, all the activated units had relatively small spike heights. To dissect which action of adrenaline was responsible for the acceleration and excita- tion we proceeded to investigate noradrenaline and isoprenaline. 1 pg/kg noradrenaline had no noticeable effect but 5 pg/kg led to a marked pro- longed excitation (traces 1, 2, Fig. 5). When this dose was given repeatedly at 15 min intervals, the latency of excitation became shorter decreasing from 32-10 set and there was a prolongation of firing which increased from 3-10 min. If the alpha blocking agent, phentolamine, 0.5 mg/kg was given by itself there was no significant effect (3, Fig. 5). When a combination of phentolamine 0.5 mg/kg with 5 pg/kg noradrenaline was given, the nor- adrenaline excitation was blocked (4, Fig. 5). Isoprenaline, a beta agent, 10 pg/kg produced no effect (5, Fig. 5). In one preparation, this dose
produced a slight rise followed by a prolonged decrease of firing. In a single
test, during excitation produced by 5 lug/kg noradrenaline, 10 pg/kg
isoprenaline was injected and there was a period of decreased firing lasting
about 1 min followed by a return of the excitation produced by the nor-
adrenaline. No other compounds were tested. One may conclude that an
alpha agent, noradrenaline, produced excitation which is blocked by
phentolamine and that a beta agent, isoprenaline, does not excite and may even decrease firing.
Were There Signs of Interaction Between Fibers in the Neurouna? The possibility was checked in a number of ways and all results were negative. A single supermaximal shock was delivered to the cut end of a dorsal root (S5, Fig. 1) and recordings were made from a filament dissected from the same root but not stimulated. No signs were observed of any orthodromic volley in the passive filament following the massive antidromic volley in all of its neighboring fibers. Equally negative results were obtained if a ventral root was stimulated and recording taken from a ventral root filament (S6, R2, Fig. 1) . S imilarly no interactions were observed if dorsal root was stimulated and ventral root recorded (S5, R2) or if the ventral root was stimulated and the dorsal root recorded (S6, R). Since there was such a marked depression of orthodromic ongoing firing following antidromic tetanus of the active fibers, the effect of tetanization of the complete root on activity in a component filament was observed. Again there was never signs of interruption of ongoing activity in fibers when their neighbors were tetanized. Evidently there is no obvious interaction between the fibers. In

PERIPHERAL NERVES 589
spite of these negative experiments, one must mention that the rate of on- going activity of multiunit recordings often showed small slow variations with a frequency of 5-20 sec. These small surges of activity can be seen in Fig. 3. The frequency of firing of individual units making up these multi- unit records showed a phase locked variation of their average frequency.
DISCUSSION
A technique is described here for growing a neuroma so that it is con- tained in a chamber. A similar technique had been used (1) for anatomical purposes but here we were able to use ‘the chamber for physiological and pharmacological purposes. Evidence is presented that some of the fibers in the neuroma are active in the absence of any intentional stimulation. There was no doubt that these impulses were originating in the neuroma since pressure and adrenaline applied locally to the neuroma increased their fre-
FIG. 5. The effect of various compounds given intravenously on the ongoing rate of activity, originating from a neuroma and recorded on a dorsal root filament. Drugs injected at the arrow: 1. 5 pg/kg noradrenaline; 2. 5 ag/kg noradrenaline; 3. 0.4 mg/kg phentolamine; 4. 0.4 mg/kg phentdamine with 5 pg/kg noradrenaline; 5. 10
@g/kg isoprenaline. Time mark: 12 sec. Rate mark: 30 impulses per sec.
quency and local lidocaine abolished it. The evidence suggests that the on- going activity was limited to small rather than large myelinated fibers. We can say nothing about unmyelinated fibers since the techniques used did not allow us to record from such axons. We could not always identify by elec- trical stimulation the conduction velocity of the axon carrying the ongoing activity. The reason for this is that the spikes were small in amplitude, usually below 150 pV, and electrical stimulation produced a large com- pound action potential in the small root filament. Sometimes the active unit could be identified by its size and shape within the larger wave of the com- pound action potential but frequently so many silent units were brought into

590 WALL AND GUTNICK
action by the peripheral electrical stimulus that it was difficult to be quite certain of the unit’s identity. However, it was clear that all spontaneously active units had a low amplitude. The relationship between spike height and conduction velocity has been investigated (6) and is frequently used in ventral root recordings to differentiate between alpha and gamma axons. Using these criteria and the few clearly identified units by electrical stimula- tion, we can say that activity in myelinated fibers appeared limited to the delta fibers. It is of interest that the larger diameter fibers whose endings are normally silent in the absence of stimulation (9) should also have silent terminals within a neuroma. It is not known if large diameter fibers have sprouts within a neuroma of different morphology from those originating in smaller fibers but it is apparent that these sprouts have a different pharmacology and physiology. It is also of interest that the smaller fibers include those normally transmitting nociceptive information and therefore this ongoing afferent barrage might be the source of pain.
The physiological and pharmacological properties of the impulse genera- tors in the neuroma evidently differ substantially from those reported for normal sensory endings; specifically they are sensitive to blood flow, en- forced activity, and to sympathetic amines. Since these impulses may be the source of pain, the unusual nature of their generation is of more than academic interest because it may offer therapeutic opportunities. High fre- quency stimulation of peripheral nerve was shown to produce analgesia (22) particularly in certain cases of peripheral nerve injury and this has now been extended into therapy by a number of groups (3). When this phenomenon was first observed, it was attributed to central inhibition pro- duced in the spinal cord and was one of the pieces of evidence to support the gate control theory of pain. However, it was puzzling that certain cases showed a very prolonged relief of pain following the end of the tetanic peripheral stimulation whereas the physiological studies of spinal cord excitability showed a relatively rapid return of central excitability follow- ing peripheral stimulation. One paper (5) suggests that the analgesic effect of peripheral stimulation is local block of the peripheral nerve under the stimulating electrodes but this work is unsatisfactory because it used very high stimulaiton currents, failed to demonstrate the block and failed to explain why the stimulus itself was not painful. However, the results re-
ported here suggest that in certain cases with peripheral nerve injury and sprouting, the relief of pain may have, in part, a peripheral explanation. It
is possible that as the stimulating current is raised, central inhibition occurs as previously suggested (20,22). This would allow the stimulus to be raised further to involve the smaller myelinated afferents which would then
transmit impulses antidromically into the damaged region. We have shown
here that such tetanic invasion will produce a prolonged turn off of the

PERIPHERAL NERVES 591
impulse generators. This peripheral mechanism would not only explain the prolonged effect but also the extreme localization of the effect. We noticed even in the initial cases that it was essential to stimulate the damaged nerve central to the damage and that stimulation of neighboring intact nerves failed to alleviate the pain (21, 22) ; others have reported the same phenomenon (3, 14). If the prolonged turn-off is peripheral and if, as we have shown here, no peripheral interaction occurs then one would not expect any pain relief on stimulation of unaffected nerves.
The reason for the turn-off following tetanic antidromic invasion is not known but we may speculate. The fibers involved must be fme, less than 1 pm diameter (1, 4) ; they are often closely packed (17) ; for some of their length they are not surrounded by Schwann cells (1, 17) ; they may be in the process of regeneration or degeneration (17) ; the vascularity of the neuroma is poor (14, 17). All of these factors might combine to make the impulse generator site slow to recover following high frequency driving. It has been shown that both in the kitten and during the early stages of degeneration, the central terminal arborizations are unable to sustain the normal range of high frequency response and recover more slowly than the mature endings (11). We have shown here that the electrical threshold of the neuroma is raised following ‘tetanic invasion but not for so long a time as the decrease of ongoing activity. A similar finding has been reported for central terminals during post-tetanic potentiation (24). The poor cor- relation of the two measures does not mean that they are unrelated because the electrical stimulation may well generate nerve impulses in the larger more proximal parts of the fine fibers which may recover more rapidly than the finer more distal terminals where the ongoing activity may be generated. Therefore it remains a possibility that the tetanus hyperpolarizes the ter- minals and raises their threshold thereby decreasing their tendency to generate impulses spontaneously.
The excitation of the terminals by the alpha type of sympathetic amine is striking and a surprising finding. The mechanism of this action is not known. Since local application has an effect, the result cannot be due to general systemic actions such as a rise of blood pressure. The latter is also ruled out by the long latency and prolonged effect of the drugs. If the action is within the neuroma we must ask if it is on any of the three components,
the vasculature, the surrounding cells, or the nerve terminals themselves. The local vasculature seems unlikely for the reasons that this would react
rapidly and that blood flow would tend to decrease and we know that this decreases firing. We cannot at this stage differentiate the possible effects
on surrounding cells or on the nerve fibers themselves. Whatever the mechanism may be, the results obviously suggest that clinical trials should be made to observe the effect of alpha blocking agents, such as phenoxy-

592 WALL AND GUTNICK
benzamine and guanethidine, on pain in those patients who are candidates for sympathectomy.
Our inability to detect interactions between the fibers in the neuroma was quite clear. It had been suggested that ephaptic connections developed in the region of damage and some experimental evidence for this was advanced (8). In experiments to be reported (23) we were unable to repeat these acute experiments and here in a region of chronic damage, we also saw no signs of interaction. This observation is probably the expected one for the following reasons ; no tight junctions have been seen between fibers in a neuroma (1, 17) ; current flow between fibers will be decreased by the high internal resistance of fine fibers; current flow will also be decreased by the high external resistance produced by close packing. Therefore the suggestion that pain from damaged nerve was produced by the ephaptic transmission of efferent impulses from the sympathetic and motor systems into afferent nerve fibers now seems unlikely. It seems more likely that the fine sprouts in damaged nerves possess nerve impulse trigger zones with unusual fragile properties which include sensitivity to circulating sym- pathetic amines and to high frequency nerve impulses.
REFERENCES
1. AGUAYO, A. J., J. M. PEYRONNARD and G. M. BRAY. 1973. A quantitative ultra- structural study of regeneration from isolated proximal stumps of transected unmyelinated nerves. J. Newopath. and Exp. Newel. 32: 256-270.
2. BASBAUM, A. I. 1974. Effects of central lesions on disorders produced by multiple dorsal rhizotomy in rats. Exfi. Neurol. 42: 490-501.
3. BONICA, J. 1974. Editor. Pain : Proceedings of the Seattle International Symposium, Raven Press, New York.
4. CAJAL, S. R. J. 1928. Degeneration and regeneration of the Nervous System. Ox- ford University Press, London.
5. CAMPBELL, J. N. and A. TAUB, 1973. Local analgesia from percutaneous electrical stimulation. Arch. Neural. 288 : 339-346.
6. CATON, W. T. 1958. Some properties of frog skin mechanoreceptors. J. Physiol. 141: 305-322.
7. DIAMOND, J. 1959. The effects of injecting acetyl choline into normal and re- generating nerves. J. Physiol. 145: 611-629.
8. GRANIT, R. and C. R. SKOGLUND. 1945. Facilitation and depression at the ‘artificial synapse’ formed by the cut end of a mammalian nerve. J. Physiol. 103 : 432-448.
9. Iooo, A. 1973. Handbook of Sensory Physiology, Vol. II Somatosensory System. Springer-Verlag, Berlin.
10. KIRK, E. J. 1973. Personal Communication. 11. KOSTYUK, P. G. 1963. Functional changes during degeneratidn of central synapses.
pp. 291-304. Zr, “The Effect of Use and Disease on Neuromuscular Junctions.” E. Gutmann and P. Hnik. [Eds.]. Elsevier, Amsterdam.
12. LOPRESTI, V., E. R. MACAGNO and C. LEVINTHAL. 1973. Structure and development of neuronal connections in isogenic organisms. Proc. Nat. Acad. Sci. U.S.A.
70 : 433-437.

PERIPHERAL NERVES 593
13. MELZACK, R. and P. R. BROMAGE. 1973. Experimental phantom limbs. Exp. Nezcrol. 39 : 261-269.
14. MEYER, G. A. and H. L. FIELDS. 1972. Causalgia treated by selective large fiber stimulation of peripheral nerve. Broiw 95: 163-168.
15. MORRIS, J. A., A. F. HUDSON and G. WEDDELL. 1972. A study of degeneration and regeneration in the divided rat sciatic nerve based on electron microscopy. 2. Fell. Mikrosk. Anat. 124: 76-103.
16. NOORDENBOS, W. 1959. Pain. Elsevier, Amsterdam. 17. SPENCER, P. S. 1971. Light and electron microscopic observations on localized
peripheral nerve injuries. Ph.D. Thesis, London1 University. 18. SUNDERLAND, A. 1968. Nerves and nerve injuries. Livingston, Edinburgh. 19. WALL, P. D. 1958. Excitability changes in afferent fiber terminations and their
relation to slow potentials. J. Physiol. 142: l-21. 20. WALL, P. D. 1973. Dorsal horn electrophysiology. pp. 253-270. In “Handbook of
Sensory Physiology,” Vol. II, A. Iggo [Ed.]. Springer-Verlag, Berlin. 21. WALL, P. D. and A. R. JOHNSON. 1958. Changes associated with post-tetanic poten-
tiation of a monosynaptic reflex. 1. Nezcrophysio~. 21 : 148-158. 22. WALL, P. D. and W. H. SWEET. 1967. Temporary abolition of pain in man.
Science 155: 108109. 23. WALL, P. D., S. G. WAXMAN and A. I. BASBAUM. 1974. Ongoing activity in
peripheral nerve: 3: the effect of acute injury. In preparation. 24. WALL, P. D. and B. WICKELGREN. 1968. Afferent hyperpolarization and post-tetanic
potentiation of a monosynaptic reflex. J. Plzysiol. 196: 135. 25. WAXMAN, S. G. and P. D. WALL. 1973. Acute neurophysiological effects of injury
on cat sural nerve. Nczlrology 23: 395. 26. WHITE, J. C. and W. H. SWEET. 1969. Pain and, the neurosurgeon. Thomas, Spring-
field.




















