
Morphological changes on nerves and histopathological effects on liver and kidney of rats by...
11
Camp. Biachem. F!ysiol, Vol. IOlC, No. 2, pp. 353-363, 1992 Printed in Great Britain 0306~92192 S.00 + 0.00 Q 1992 Pergamon Press pIc MORPHOLOGICAL CHANGES ON NERVES AND HISTOPATHOLOGICAL EFFECTS ON LIVER AND KIDNEY OF RATS BY PENTACHLOROPHENOL (PCP) F. VILLENA,* G. MoNroYA,*t R. KLAASEN,~ R. FLECKENSTEIN$ and M. SUWALSKY$ *Faculty of Biological Sciences and Natural Resources, $Faculty of Medicine and §Faculty of Sciences, University of Conception, Casilla 2407, Conception, Chile (Received 19 March 1991) Abstract-i. The chronic toxicity of ~nt~hlorophenol (PCP) was studied in rats after 90-120 days of oral administration ad fjbiru~ of 0.3-3 mM PCP aqueous solutions. 2. Morphological studies of their sciatic nerves were performed by optical and electron microscopy. 3. They showed degenerative changes in about 10% of the A and B type of nerve fibers. 4. The myelin sheath was discontinued by complete separation in several concentric rings while some other parts of the nerve exhibited a variable loss of neurotubules, neurofilaments and other axoplasmic components. 5. However, the C type of nerve fibers, the blood vessels and the perineurium did not show any morphologic alteration. 6. It was also found that in the liver PCP caused hemodynamic vein changes and injury in the hepatocytes such as cellular swelling and vacuolar degeneration. 7. The damage in the kidney occurred primarily in the glomeruli and secondarily in the proximal tubules causing turbid tumefaction and the formation of casts in the tubular lumen. Pentachlorophenol (PCP) and its salts are used as algicides, bactericides, fungicides, herbicides, insecti- cides and molluscides with a variety of applications in the industrial, agricultural and domestic fields (Cirelli, 1978; Hess, 1978). PCP is one of the contami- nants of ground water (Buhler et al., 1973; McCarthy et al., 1981), soils (Schellenberg et al., 1984; Tam and Trevors, 1981) and cattle (Schull et al., 1986). Conse- quently, this compound is routinely detected in human tissues, body fluids (Conklin and Fox, 1978) and in food (Wu et al., 1978). It has been shown that PCP is also a product of the metabolic conversion of other pesticides such as hexachloro~nzene and pentachloronitrobenzene (Engst et al., 1976; Renner, 1981). It is now generally accepted that the major mode of PCP toxicity is related to its liopopi ilicity. In fact, it is able to partition into the lipid biiayer of cell membranes disrupting several functions (Hayes, 1978). It uncouples the oxidative phosphorylation (Weinbach and Garbus, 1965), alters the microsomal electron transport system (Weinback and Garbus, 1965) and inhibits the amino acid transport across cell membranes (Hissin and Silf, 1978). PCP is readily absorbt ,l th, ough the skin as well as through the respiratory : nd gas rointestinal tract. In rats, which were given a single c ral dose of 10 mg of “C-PCP per kg of body weight, the peak plasma concentration (50 mg/l) was attained only 4-6 hr after administration (Brau,i et al., 1977). The body com- ponent containing the highest level of PCP appeared to be the liver, followed by the kidney and blood ~To whom correspc,ldence should be addressed. (Larsen et al., 1972). A feeding study described by McConnell et d. (1980) indicated that the chronic toxicity of PCP was primarily due to accompanying microcontaminants such as chlorodibenzodioxins and chlorodi~nzofurans rather than to PCP itself. The ubiquitous presence of PCP in the environ- ment, as well as in man, stimulates the need to understand the precise mechanism of its toxicity. The present work was performed primarily to examine the chronic effects of PCP on nerve fibers. Furthe~ore, the damage produced by PCP to liver and kidney was also studied. MATERIALS AND METHODS Male Wistar albino rats of 300-350 g of body weight were divided into five groups. The first group was given PCP (Sigma, Lot 127F3439) 0.3 mM in water ad Iibitum for 60 days. The second received 1 .OmM PCP for 60 days. The third drank the same solution but for 90 days. The fourth was given PCP 3.0 mM 120 davs and the fifth was a control group. The animals were killed by decapitation and their sciatic nerves, liver and kidneys were extracted. The nerves were tixed in 2.5% glutaraldehyde in 0.15 M sodium phos- phate buffer pH 7.4 for 30min (Sabatini et al., 1963), followed by a rinse in the buffer and post-fixed with 2% osmium tetroride in the same buffer for 1 hr (Milonig, 1961). After a short rinsing with buffer the specimens were dehydrated in graded et’<anoi series, embedded in Epon (Luft. 1961) and process i f~ ultra-thin section using a MT2 Porter Rlum ultran i:rotoml (Du Pont Instruments, Sorvall). Sectiofis for clectrlm microscopy were stained with 2% aqueous solution of uranyl acetate and lead cirtrate stain (Reynolds, 19C3). Thick sections for light microscopy were stained with Tduidine Blue (Trump et al,, 1961). The liver and kidneys were fixed in 10% formaline and included i? paraffin for processing by conventional histological tech- niques. Sections were stained with haematoxylin and eosin. 353
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Morphological changes on nerves and histopathological effects on liver and kidney of rats by...

Camp. Biachem. F!ysiol, Vol. IOlC, No. 2, pp. 353-363, 1992 Printed in Great Britain
0306~92192 S.00 + 0.00 Q 1992 Pergamon Press pIc
MORPHOLOGICAL CHANGES ON NERVES AND HISTOPATHOLOGICAL EFFECTS ON LIVER AND
KIDNEY OF RATS BY PENTACHLOROPHENOL (PCP)
F. VILLENA,* G. MoNroYA,*t R. KLAASEN,~ R. FLECKENSTEIN$ and M. SUWALSKY$
*Faculty of Biological Sciences and Natural Resources, $Faculty of Medicine and §Faculty of Sciences, University of Conception, Casilla 2407, Conception, Chile
(Received 19 March 1991)
Abstract-i. The chronic toxicity of ~nt~hlorophenol (PCP) was studied in rats after 90-120 days of oral administration ad fjbiru~ of 0.3-3 mM PCP aqueous solutions.
2. Morphological studies of their sciatic nerves were performed by optical and electron microscopy. 3. They showed degenerative changes in about 10% of the A and B type of nerve fibers. 4. The myelin sheath was discontinued by complete separation in several concentric rings while some
other parts of the nerve exhibited a variable loss of neurotubules, neurofilaments and other axoplasmic components.
5. However, the C type of nerve fibers, the blood vessels and the perineurium did not show any morphologic alteration.
6. It was also found that in the liver PCP caused hemodynamic vein changes and injury in the hepatocytes such as cellular swelling and vacuolar degeneration.
7. The damage in the kidney occurred primarily in the glomeruli and secondarily in the proximal tubules causing turbid tumefaction and the formation of casts in the tubular lumen.
Pentachlorophenol (PCP) and its salts are used as algicides, bactericides, fungicides, herbicides, insecti- cides and molluscides with a variety of applications in the industrial, agricultural and domestic fields (Cirelli, 1978; Hess, 1978). PCP is one of the contami- nants of ground water (Buhler et al., 1973; McCarthy et al., 1981), soils (Schellenberg et al., 1984; Tam and Trevors, 1981) and cattle (Schull et al., 1986). Conse- quently, this compound is routinely detected in human tissues, body fluids (Conklin and Fox, 1978) and in food (Wu et al., 1978). It has been shown that PCP is also a product of the metabolic conversion of other pesticides such as hexachloro~nzene and pentachloronitrobenzene (Engst et al., 1976; Renner, 1981). It is now generally accepted that the major mode of PCP toxicity is related to its liopopi ilicity. In fact, it is able to partition into the lipid biiayer of cell membranes disrupting several functions (Hayes, 1978). It uncouples the oxidative phosphorylation (Weinbach and Garbus, 1965), alters the microsomal electron transport system (Weinback and Garbus, 1965) and inhibits the amino acid transport across cell membranes (Hissin and Silf, 1978).
PCP is readily absorbt ,l th, ough the skin as well as through the respiratory : nd gas rointestinal tract. In rats, which were given a single c ral dose of 10 mg of “C-PCP per kg of body weight, the peak plasma concentration (50 mg/l) was attained only 4-6 hr after administration (Brau,i et al., 1977). The body com- ponent containing the highest level of PCP appeared to be the liver, followed by the kidney and blood
~To whom correspc,ldence should be addressed.
(Larsen et al., 1972). A feeding study described by McConnell et d. (1980) indicated that the chronic toxicity of PCP was primarily due to accompanying microcontaminants such as chlorodibenzodioxins and chlorodi~nzofurans rather than to PCP itself.
The ubiquitous presence of PCP in the environ- ment, as well as in man, stimulates the need to understand the precise mechanism of its toxicity. The present work was performed primarily to examine the chronic effects of PCP on nerve fibers. Furthe~ore, the damage produced by PCP to liver and kidney was also studied.
MATERIALS AND METHODS
Male Wistar albino rats of 300-350 g of body weight were divided into five groups. The first group was given PCP (Sigma, Lot 127F3439) 0.3 mM in water ad Iibitum for 60 days. The second received 1 .O mM PCP for 60 days. The third drank the same solution but for 90 days. The fourth was given PCP 3.0 mM 120 davs and the fifth was a control group. The animals were killed by decapitation and their sciatic nerves, liver and kidneys were extracted. The nerves were tixed in 2.5% glutaraldehyde in 0.15 M sodium phos- phate buffer pH 7.4 for 30min (Sabatini et al., 1963), followed by a rinse in the buffer and post-fixed with 2% osmium tetroride in the same buffer for 1 hr (Milonig, 1961). After a short rinsing with buffer the specimens were dehydrated in graded et’<anoi series, embedded in Epon (Luft. 1961) and process i f~ ultra-thin section using a MT2 Porter Rlum ultran i:rotoml (Du Pont Instruments, Sorvall). Sectiofis for clectrlm microscopy were stained with 2% aqueous solution of uranyl acetate and lead cirtrate stain (Reynolds, 19C3). Thick sections for light microscopy were stained with Tduidine Blue (Trump et al,, 1961). The liver and kidneys were fixed in 10% formaline and included i? paraffin for processing by conventional histological tech- niques. Sections were stained with haematoxylin and eosin.
353

354 F. VILLENA ei al
Trans mission electron microscopy observations were made in a P ‘hilips 200 EM, while a Leitz Orthoplan was used for optica .I microscopical studies.
Ele :ctron and optical observations on the sciatic nerve from rats fed with 0.3 mM PCP for 60 days (grou p 1) and 1.0 mM for 60 days (group 2) did not show any significant difference when compared with those of the control group. However, about IO% of
Fig. IA. Thick section: myelin dissociation (thin arrows) and myelin islots (thick arrow). Toluidine Bfue. optical microscopy. (All the micrographs correspond to samples obtained from the group 3 of rats.)
RESULTS
type A and B fibers from rats belonging to groups 3 (1 .O mM PCP for 90 days) and 4 (3.0 mM PCP for 120 days) showed visible changes in their myelin sheath. In fact, its structure was discontinued by complete separation in two or three concentric rings (Fig. 1A and 8) or zones of separation along the perimeter that looked like empty spaces between the phospholipid bilayers (Fig. 2A and B). A consider- able number of these nerve fibers showed variable degrees of dissociation of the layers that form the myelin sheath. Besides, some of these fibers had
Fig. IB. Myelin fibers with three concentric rings; electron microscopy.
Morphological and histopathological changes by PCP 355
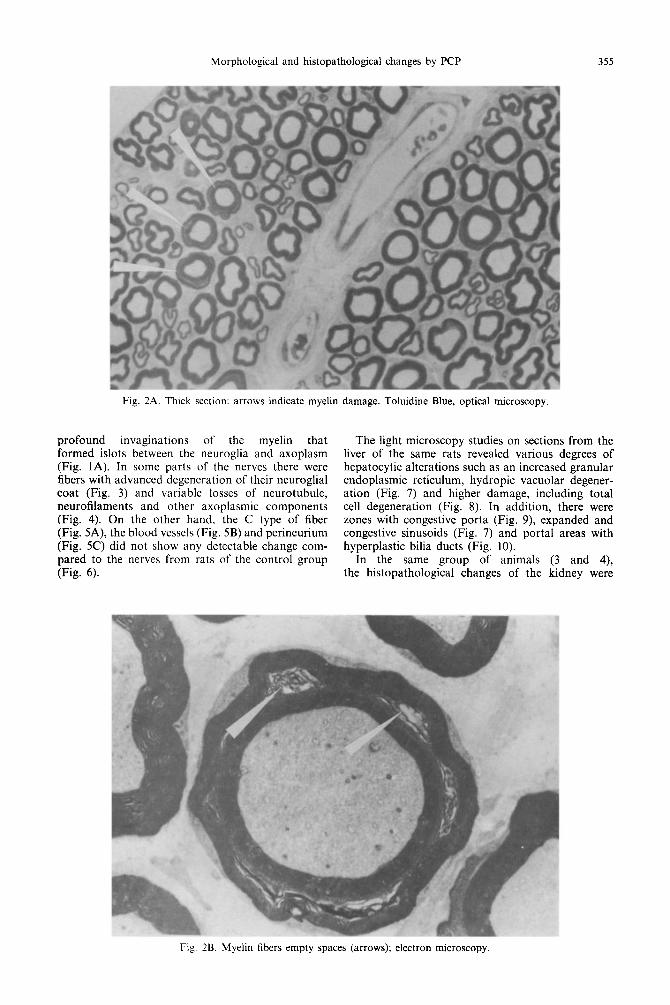
Fig. 2A. Thick section: arrows indicate myelin damage. Toluidine Blue, optical microscopy.
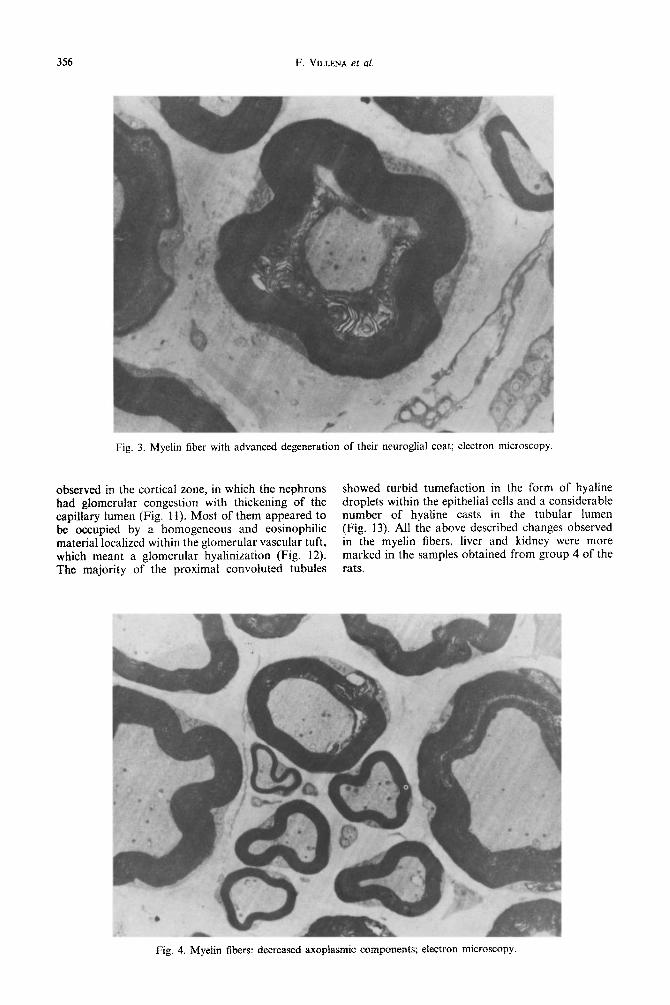
profound invaginations of the myelin that formed islots between the neuroglia and axoplasm (Fig. 1A). In some parts of the nerves there were fibers with advanced degeneration of their neuroglial coat (Fig. 3) and variable losses of neurotubule, neurofilaments and other axoplasmic components (Fig. 4). On the other hand, the C type of fiber (Fig. 5A), the blood vessels (Fig. 5B) and perineurium (Fig. 5C) did not show any detectable change com- pared to the nerves from rats of the control group (Fig. 6).
The light microscopy studies on sections from the liver of the same rats revealed various degrees of hepatocytic alterations such as an increased granular endoplasmic reticulum, hydropic vacuolar degener- ation (Fig. 7) and higher damage, including total cell degeneration (Fig. 8). In addition, there were zones with congestive porta (Fig. 9) expanded and congestive sinusoids (Fig. 7) and portal areas with hyperplastic bilia ducts (Fig. 10).
In the same group of animals (3 and 4), the histopathological changes of the kidney were
Fig. 2B. Myelin fibers empty spaces (arrows); electron microscopy.
356 F. VILLENA et al.
Fig. 3. Myelin fiber with advanced degeneration of their neuroglial coat; electron microscopy
observed in the cortical zone, in which the nephrons showed turbid tumefaction in the form of hyaline had glomerular congestion with thickening of the droplets within the epithelial cells and a considerable capillary lumen (Fig, 11). Most of them appeared to number of hyaline casts in the tubular lumen be occupied by a homogeneous and eosinophilic (Fig. 13). All the above described changes observed material localized within the glomerular vascular tuft, in the myelin fibers, liver and kidney were more which meant a glomerular hyalinization (Fig. 12). marked in the samples obtained from group 4 of the The majority of the proximal convoluted tubules rats.
Fig. 4. Myelin fibers: decreased axoplasmic components; electron microscopy

Morphological and histopathological changes by PCP 351
Fig. 5A. Normal amyelinic fibers (arrow); electron microscopy
DlSCUSSlON
The present study has shown that PCP is able to produce neurotoxicity and damage to the liver and kidney of rats by its chronic ingestion.
In general, PCP concentration and/or exposure time determined the degree of injury on the nerve structure, which may culminate in degeneration of the myelinated nerve fibers. This is supported by observations made with the neurotoxic phenol which, at low concentration and short exposure time, only
reversibly blocks the nerve conduction. However, at high concentrations it produces a long-lasting block with subsequent mo~hologi~al nerve damage (Burke and McPhee, 1970; Nathan et al., 1965). On the other hand, it is of interest to relate the above-reported damage that PCP produced on the myelin sheath with that produced by this compound on lipid bilayers. Structural studies performed in this laboratory have shown that PCP is able to perturb to a significant extent the bilayer structure of two membrane-related phospholipids (Suwalsky et al., 1990). At present, an
Fig. 5B. Thick section: blood vessel (arrow), Toluidine Blue; optical microscopy.

358 F. VILLENA et al.
Fig. 5C. Thick section: perineurium (arrow), Toluidine Blue; optical microscopy
X-ray study on the damage produced to the sciatic
nerve of PCP-fed rats is in progress. Neurotoxic effects of PCP had been previously reported in vitro (Walun and Peterson, 1984) in which cultured mice neuroblastoma cells showed an increase of cell detachment with exposure to PCP. It has also been found that myelin maturation is affected by PCP, although it is not clear whether this effect is due to the direct action of PCP on the myelin sheet or to an interference with either food intake or
with lipid metabolism. The latter is essential for the increase of the myelin sheath thickness during the late maturation period. In this respect it is of interest to note that prenatal and postnatal malnutrition may also cause impairment of myelin development (Hedley-White, 1973; Krigman and Hogan, 1976; Oldfors and Ulmann, 1980; Sima, 1974).
In a previous study in vitro carried out in this laboratory it was found that the axonal conduction
Fig. 6. Myelin fiber (thick arrow) of rat sciatic nerves without treatment (Control). Electron microscopy.

Morphological and histopathological changes by PCP 359
Fig. 7. Liver. Hidropic vacuolar degeneration of cells and expanded sinusoids. Optical microscopy.
in nerve fibers and synaptic transmission in toad sympathetic ganglia were blocked by PCP (Montoya et al., 1988). These effects were dose-time dependent and could not be reversed. These results indeed agree with those found in the present study.
The present study has shown that the organ most affected by PCP is the kidney, in which the glomeru- lar congestion that implies a structural manifestation of a type of nephritis, has the potential of progressing to a widespread destruction of the renal parenchymal and, therefore, a loss of function. In fact, since hyalinization of the glomeruli implies irreversible
damage to the glomerular structure and function, these observed in such conditions represent the latest and most serious stage of glomerular alteration. The hyaline droplets in the proximal convoluted tubular cells represent, as in other forms of renal diseases, the reabsorption of excessive amounts of protein from the glomerular filtrate. This indicates that the primary damage is localized in the filtrating barrier. This conclusion is consistent with the hyaline casts observed in the lumen of a great number of proximal tubules and the scant proportion of them present in the collector tubular system.
Fig. 8. Liver. Islot of cells with advanced degeneration. Optical microscopy.

360 F. VILLENA et ai.
Fig. 9. Liver. Congestive portal blood vessel. Optical microscopy.
Fig. 10. Liver. Hyperplastic bilia ducts. Optical microscopy.

Morphological and histopathological changes by PCP 361
Fig. Il. Kidney. Congestive renal corpuscle. Optical microscopy.
Fig. 12. Kidney. Glomerular hyalinization. Optical microscopy.

362 F. VILLENA er ul.
Fig. 13. Kidney. Tumefaction and hyaline casts in proximal convoluted tubules. Optical microscopy.
Acknowledgements-These studies were supported by grants from FONDECYT (0625/89 and 0783/88) and from the University of Conception (20.36.02 and 20.13.79).
REFERENCES
Buhler D. R., Rassmuson M. E. and Natrahue H. S. (1973) Ocurrence of hexachlorophene and penthachlorophenol in sewage and water. Enciron. Sci. Technol. I, 929-934.
Burke1 W. E. and McPhee M. (1970) Effect of phenol iniection into peripheral nerve of rat; electromicroscopy studies. Arch Physiol. Med. Rehab. 51, 391-397.
Braun W. H.. Young J. D.. Blau G. E. and Gehring P. J (1977) The’ pharmacokinetics and metabolism of penta- chlorophenol in rats. Toxical appl. Pharmacol. 41, 395406.
Cirelli D. P. (1978) In Pentachlorophenol, Chemistry, Phar- macology and Environmental Toxicity (Edited by Rao K. R.), pp. 3-11. Plenum Press, New York.
Conklin P. J. and Fox F. R. (1978) Environmental impact of pentachlorophenol and its products-a round-table discussion. In PentachloroDhenol (Edited by Rao K. R.), pp. 389-394. Plenum Press, New‘York. .
Debets F. M. H., Strik J. J. T. and Olie K. (1980) Effects of pentachlorophenol on rat liver changes induced by hexachlorobenzene with special reference to porphiria and alterations in mixed-function oxygenases. Toxicology 15, 181-195.
Engst T., Machloz R. M. and Kujawa M. (1976) The metabolism of hexachlorobenzene (HCB) in rat. Bull. Environ. Contam. Toxicol. 16, 248-252.
Hayes A. (1978) Transport processes as sites of action of toxic chemicals. Fedn. Proc. 38. 2218-2219.
Hedley-White E. T. (1973) Myelination of rat sciatic nerve: Comparison of undernutrition and cholestero biosyntesis inhibition. J. Neuropathol. exp. Neural. 32, 284-302.
Hissin P. J. and Hilf R. (1978) Characteristic of proline transport into R3230 AC mammary tumor cells. Biochim. biophys. Acta 508, 401-412.
Hoss R. A. W. (1978) In Pentachlorophenol, Chemistry, Pharmacology and Enviromental Toxicity (Edited by Rao K. R.), pp. 3-l 1. Plenum Press, New York.
Krigman M. R. and Hogan E. L. (1976) Undernutrition in the developing rat: effect upon myelination. Brain Res. 107, 239.-255.
Larsen R. V., Kirsh L. E., Shaw S. M., Christin J. E. and Born G. S. (1972) Excretion and tissue distribution of uniformly labelled ‘*C-pentachlorophenol in rats. J. Pharm. Sri. 61, 2004-2006.
Luft J. (1961) Improvements in epoxy resin embeddings method. J. biophJ’.s. biochrm Cyt;l. 4, 409414.
McCarthv P. L.. Rinhard M. and Rittman B. E. (1981) Trace organics’in ground water. Environ. Sci. Tech&. 15; 40-51.
McConnell E. E., Moore J. A., Grupta B. N., Rakes A. H., Luster M. I., Goldstein J. A., Haseman J. K. and Parker C. E. (1980) The chronic toxicity of technical and analytical pentachlorophenol in cattle. I. Clinico- pathology. To.uicol. appl. Pharmacol. 52, 468490.
Milloning G. (1961) Advantages of a phosphate buffer for 0~0, solutions in fixation. J. uppl. Physiol. 32, 1637.
Montoya G. A., Roa J., Crux F., Villena F. and Pezo P. (1988) The actions of phenol and pentachlorophenol (PCP) on axonal conduction, ganglionic synaptic trans- mission, and the effect of pH changes. Comp. Biochem. Physiol. 89C, 377-382.
Nathan P. W., Sears T. A. and Smith M. C. (1965) Effect of phenol solutions on the nerve roots of the cat: an electrophysiological and histological study. J. neural. Sci. 2, 7-29.
Oldfors A. and Ulmann M. (1980) Motor nerve conduction velocity and nerve fiber diameter in exper- imental protein deprivation. Acta Neuorpathol. Berl. 51, 215-221.
Ravanel P., Taillandier G., Tissut M. and Benoit-Guyod J. L. (1985) Effect of chlorophenols on isolated plant mitochondria activities: a quantitative structure relation- ship study. Ecotoxirol. Emiron. Safety 9, 300-320.
Renner G. (1981) Biotransformation of the fungicides hexa- chlorobenzene and pentachloronitrobenzene. Xenobiotica 11, 435446.
Reynolds E. S. (1963) The use of lead citrate at high pH as an electro-opaque stain in electromicroscopy. J. Cell Biol. 17, 208.

Morphological and histopathological changes by PCP 363
Sabatini D. D., Beensche K. and Bamet R. J. (1963) Cytochemistry and electronmicroscopy: the preservation of cellular ultrastructure and enzymatic activity by alde- hyde fixation. J. Cell. Biol. 17, 19-58.
Schellenberg K., Lewenberber C. and Schwarzenback R. P. (1984) Sorption of chlorinated phenols by natural sedi- ments and aquiter materials. Environ. Sci. Technol. 18, 652-657.
Shull L. R., Olson Ba. A., Hughes B. J., Kensie R. M. and Kinsell J. H. (1986) Effect of pentachlorophenol on microsomal mixed-function oxidases in cattle. Pesfic. Biochem. Toxicol. 25, 31-39.
Sima A. (1974) Relation between the number of myelin lamellae and axon circumference in fibers of ventral and dorsal roots and optic nerve in normal, undernourished and rehabilited rats, an ultrastructural morphometic study. Acta physiol stand. 410, l-38.
Suwalsky M., Espinoza M. A., Bagrara M. and Sotomayor C. P. (1990) X-ray and fluorescence studies on phospho-
lipid bilayers. IX. Interactions with pentachlorophenol. Z. Naturjorsch. 45, 265-272.
Tam T. Y. and Trevors J. T. (1981) Effect of pentachloro- phenol on asymbiotic nitrogen fixation in soil. Water Air Soil Pollut. 16, 40%414.
Trump B. H., Smuckler E. A. and Benditt E. P. (1961) A method for staining apoxy sections for light microscopy. J. Ultrastruct. Res. 5, 343-348.
Walun E. and Peterson A. (1984) On the application of culture neuroblastoma cells in chemical toxicity screening. J. Toxicol. Environ. Health 13, 511-520.
Weinbach E. C. and Garbus J. (1965) The interaction of uncoupling phenols with mitochondria and with mito- chondrial protein. J. biol. Chem. 240, 1811-1819.
Wu A., Lech J. J., Glickman A. and Pearson M. L. (1978) Mass fragmentographic determination of pentachloro- phenol in rainbow trout. J. Ass. Ofl Anal. Chem. 61, 1303-1306.




















