
Description of Lactobacillus backi sp. nov., an Obligate Beer-Spoiling Bacterium
Upload
independentCategory
view
3download
0
-1
Obligate groundwater fauna of France: diversity
patterns and conservation implications
DAVID FERREIRA*, FLORIAN MALARD, MARIE-JOSEDOLE-OLIVIER and JANINE GIBERTUMR CNRS 5023, Ecologie des Hydrosystemes Fluviaux, Equipe Hydrobiologie et Ecologie
Souterraines, Universite Claude Bernard Lyon 1, Bat. Forel, 43 Boulevard du 11 Novembre 1918,
F-69622 Cedex, France; *Author for correspondence (e-mail: [email protected]; phone: +33-4-
72432945; fax:+33-4-72431523)
Received 31 January 2005; accepted in revised form 6 June 2005
Key words: Biodiversity, Conservation, Endemism, France, Ground water, Stygobiotic fauna
Abstract. We examined taxonomic and geographic patterns of the obligate groundwater fauna (i.e.
stygobiotic fauna) by assembling in a distributional data base all species occurrences reported from
France since 1805. A simulated annealing algorithm was used to identify conservation targets. Until
the 60s, biological surveys were restricted to caves but the proportion of sampling sites in
unconsolidated sediments increased from 1 to 16% over the last 40 years. A total of 380 species and
subspecies in 40 families were collected, 70% of which being restricted to France. As observed in
other temperate regions, the stygobiotic fauna was dominated by crustaceans (65% of species) and
molluscs (22%). The cumulative number of species did not level off over time, clearly showing that
biodiversity was underestimated. Temporal trends in the cumulative number of obligate ground-
water and surface water species suggested that groundwater comprised more crustaceans than
surface freshwater. Endemism was high although the geographic range size of species increased as
distributional data accumulated. Of 380 species, 156 were known from a single 400-km2 cell, among
which 73% were located in the southern third of France. The distribution map of species richness
changed dramatically over time, indicating that the location of richness hotspots was sensitive to
sampling effort. Less than 2% of the French landscape was needed to capture 60% of known
species. Thus, a large proportion of species could be protected by focusing habitat conservation
efforts on a few complementary species-rich aquifers located in distinct regions.
Introduction
In the last few decades, groundwater ecology has developed rapidly forming afertile discipline of aquatic ecology (Gibert et al. 1994). Whereas the subter-ranean domain has long been considered as a species-poor environment,worldwide syntheses revealed an unexpectedly high diversity of living forms ingroundwater (Botosaneanu 1986; Juberthie and Decu 1994, 1998, 2001).Botosaneanu (1986) listed about 7000 obligate groundwater species worldwide.Groundwater ecosystems harbor different kinds of animal organisms fromtypically accidental obligate-surface water species (i.e. stygoxenes) to a highlyspecialized obligate groundwater fauna (i.e. stygobionts), the members ofwhich developed adaptive strategies for life in a dark and energy-limited
Biodiversity and Conservation (2007) 16:567–596 � Springer 2007
DOI 10.1007/s10531-005-0305-7

environment (Marmonier et al. 1993; Langecker 2000). Because moststygobionts have a narrow distribution range, the risk of species extinction isexpectedly high in face of the increase in multiple anthropogenic pressures(Malard et al. 1996; Gibert and Deharveng 2002; Danielopol et al. 2003). Thehigh level of endemicity in groundwater systems requires specific protectionmeasures for maintaining their ecological integrity and biological diversity(Notenboom et al. 1994). However, the current incomplete state of knowledgeon groundwater biodiversity and the lack of sound conservation strategiesseverely constrain the implementation of protection policies (Bouchet 1990;Holsinger 1993; Gibert 2001).In order to reveal the hidden biodiversity of groundwater, ecologists have
recently begun to synthesize and map existing data on the diversity anddistribution of stygobiotic fauna. Culver et al. (2000) mapped the distribu-tion of 973 aquatic and terrestrial obligate cave species in U.S.A. andshowed that 61% of the species were found only in a single county. TheItalian Ministry of Environment maintains a distributional database ofstygobionts as a background for developing adequate conservation strategies(Stoch 2000, 2001). In France, groundwater biodiversity patterns are stillpoorly known despite the existence of syntheses on the distribution of sev-eral taxonomic groups (Lescher-Moutoue 1967; Rouch 1968; Coineau 1971;Henry 1976; Magniez 1976). Works by Ginet and Juberthie (1987) andJuberthie and Ginet (1994) were the first attempts to document globaldiversity patterns based on distribution data for a restricted number ofspecies.The objectives of this paper were (1) to summarize our present-day knowl-
edge of groundwater biodiversity in France based on distributional data col-lected since 1805; (2) to examine the taxonomic and geographic patterns ofspecies diversity; and (3) to discuss the conservation implications of thesepatterns.
Material and methods
Data sources and validation procedure
Over the last three years, we assembled in a database the occurrences of allstygobionts reported from France since 1805. With the exception of Nematoda,Tardigrada and Hydrachnidia, all invertebrate groups known to have repre-sentatives within the stygobiotic fauna were included in the database. Infor-mation available in the literature, existing databases, and personal collectionswere used to gather as many records as possible and to produce lists anddistribution maps of species for each taxonomic group. In order to minimizethe risk to include spurious records, lists of stygobiotic species, records, anddistribution maps were submitted to taxonomists for correction and validation(Ferreira et al. 2003).
568

Database and mapping
In order to be exported in the European database on groundwater biodiversitydeveloped under the 4D software (Ferreira et al. 2003), all data were entered in4 Excel spreadsheets containing for each record (i.e. line) a total of 28 fields (i.e.columns). The following fields were used in the present study: (1) class, order,family, genus and species names; (2) species authorities and description year;(3) spatial coordinates of the site; (4) data sources (e.g. bibliographic reference,personal collection); (5) collection year; and (6) habitat type (e.g. karst orporous aquifers). The spatial coordinates of the centroid of the civil parish(gazetteer GEOFLA� Commune, National Geographic Institute, Paris) wasused for most records because we lacked the precise coordinates of the sam-pling site. France is divided into 36,582 civil parishes, the average area of whichis 15±15.2 km2. Data were exported into a geographical information system(ArcView 3.2 software) to map the distribution of species and species richness.
Data analysis
Because the collection data were presence-only, they could not be used toevaluate spatial heterogeneity in sampling effort. Following the proceduredeveloped by Culver et al. (2004) for the analysis of species richness in Dinariccaves, Slovenia, we utilized time snapshots of the groundwater fauna toexamine stability of diversity patterns. Based on an intensive literature search,we used the number of sampling sites at which groundwater invertebratecommunity studies were carried out to assess spatial variation in samplingeffort during three overlapping periods: prior 1907, 1960 and 2003. These datescorresponded to stepping-stones in the historical development of research ongroundwater biodiversity: the creation of the association named ‘Biospeologica’(Racovitza 1907), the emergence of several groundwater research groups in theearly 60s (Juberthie and Ginet 1994), and the launching of the present databasewithin the framework of the European program PASCALIS (i.e. Protocols forthe Assessment and Conservation of Aquatic Life In the Subsurface). Siteswhich were sampled within the framework of taxon-oriented studies (e.g. re-gional inventory of selected species) were discarded in order to restrict taxo-nomic biases in our estimate of sampling effort. The point coverage of samplingsites was intersected with a geological polygon coverage (i.e. geological map ofFrance, scale 1/1,000,000, BRGM 1997) to determine differences in the numberof sampling sites between areas comprising karst aquifers and porous aquifers.According to Geze (1973), karst areas were further divided into highly kars-tified (thick and compact limestone), well karstified (thin limestone or alter-nating with shales), and poorly karstified (chalk) areas.The year at which a species was first described or reported from French
groundwater was used to examine changes in the cumulative number of speciessince 1805. We also compared temporal changes in the cumulative numbers of
569

stygoxen and stygobiotic crustacean species. Although only the descriptionyear of species was available for stygoxen crustaceans (i.e. a species could bereported from France several years after its description), the comparisonproved to be valuable for assessing the phase difference between knowledge ofbiodiversity in groundwater and surface water.The frequency distribution of range size was examined at three different
periods to determine how increasing sampling of groundwater could modifyour perception of rarity among stygobiotic species. According to Gaston(1991), we distinguished between two measures of range size: the area ofoccupancy of a species and its extent of occurrence. The number of 400-km2
cells (see below for the definition of the grid system) in which a species wascollected was used as a measure of its area of occupancy. The latitudinal extentof a species, i.e., the straight-line distance (km) between latitudinally mostwidely separated occupied sites, was used as a measure of the extent ofoccurrence.A grid-based distribution map of species richness was produced for each
period by counting the number of species present in the 400-km2 cells of thegrid coverage. The choice of the cell area was dictated by the spatial resolutionof the data; one cell contained in average 15 civil parishes. We examinedtemporal changes in the frequency distribution of species richness as well as therelationships between species richness of cells and the number of sampling sites.Marxan 1.8.6, an optimization package designed for marine protected area
site selection (Possingham et al. 2000; Ball and Possingham 2001), was used todetermine the minimum number and potential location of cells for representingall species at least once. We performed adaptive simulated annealing followedby the summed irreplaceability heuristic algorithm to select cells. Then, theswap iterative improvement algorithm was used to ensure that no selected cellswere superfluous. We ran the simulated annealing 1000 times and measured theselection frequency of each cell by recording the number of times it was in-cluded in the ‘reserve’ network. Selection frequency is a measure of theimportance of a cell for meeting the conservation goal (i.e. representing allspecies at least once).
Results
Sampling effort
The number of sampling sites at which groundwater invertebrate communitystudies were carried out increased from 53 in 1907 to 1567 in 2003. Themajority of the sampling sites was concentrated in southern and eastern Francein areas comprising highly karstified limestone (Figure 1). Almost no inverte-brate community studies were conducted in alluvial aquifers of several majorrivers (e.g. the Seine and the Loire Rivers) and aquifers in fissured rocks andunconsolidated sediments of the Paris Basin, Armoricain Massif, and Central
570
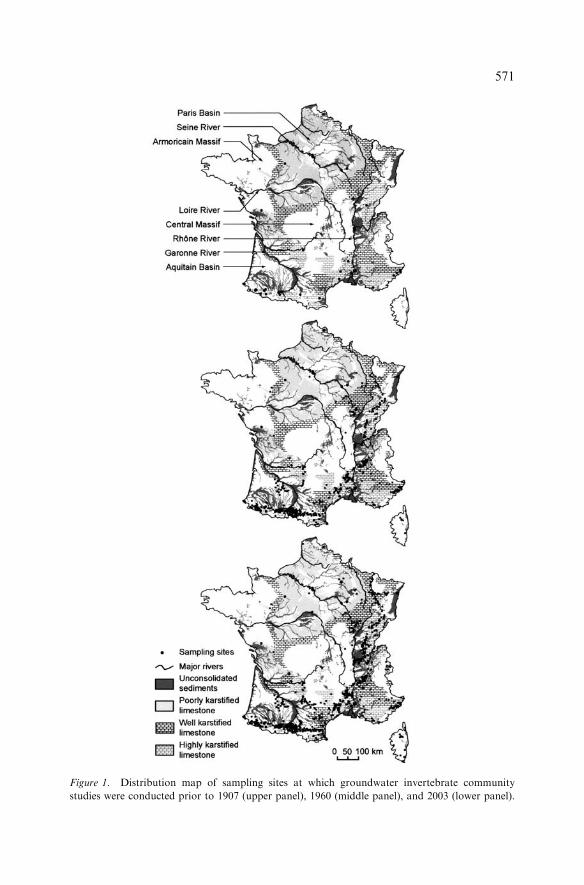
Figure 1. Distribution map of sampling sites at which groundwater invertebrate community
studies were conducted prior to 1907 (upper panel), 1960 (middle panel), and 2003 (lower panel).
571

Massif. Until the 60s, sampling was almost totally restricted to karst areas(99%) and more precisely to caves (95% of sampled sites). Over the last40 years, the proportion of sampling sites in areas comprising extensivedeposits of unconsolidated sediments increased from 1.2 to 15.9%.
Taxonomic patterns
The database contained 380 species and subspecies corresponding to more than5700 records of which 83.5%, 12% and 2.5% corresponded to occurrences ofcrustaceans, molluscs and annelids, respectively. The list of species is providedin Appendix 1. Species were distributed among 40 families and 100 genera(Table 1). Crustaceans and molluscs accounted for 65 and 22% of stygobioticspecies richness, respectively, whereas they represented only 6 and 15%,respectively, of all stygoxen species collected in France. Crustaceans accountedfor 60% of the generic and family richness. About 43% of crustaceans werecopepods, among which 33% belonged to the Cyclopidae family (Cyclopoida)and 63% to the Ameiridae, Canthocamptidae and Parastenocarididae families
Table 1. Number of families, genera, and species within the stygobiotic fauna of France.
Groundwater Surface water
Family Genus Species Species
Taxa
Nemertina 1 1 1 1
Planaria 2 6 24 11
Annelida
Aphanoneura 2 2 2 10
Oligochaeta 4 12 21 144
Polychaeta 1 1 1 na
Hirudinea 1 1 1 22
Molluscs 3 17 82 150
Crustacea
Branchiopoda 1 1 2 88
Decapoda 1 1 1 10
Ostracoda 3 10 23 68
Syncarida 2 11 21 0
Isopoda 5 7 49 1
Amphipoda 6 10 45 12
Copepoda Cyclopoida 1 7 35 31
Copepods Harpacticoida 4 10 68 35
Copepods Gelyelloida 1 1 1 0
Copepods Calanoida 1 1 1 24
Insects 1 1 2 3200
All taxa 40 100 380 4305
The last column provides estimates of the number of surface water obligate species in France
(na: not available). Data from different sources.
572

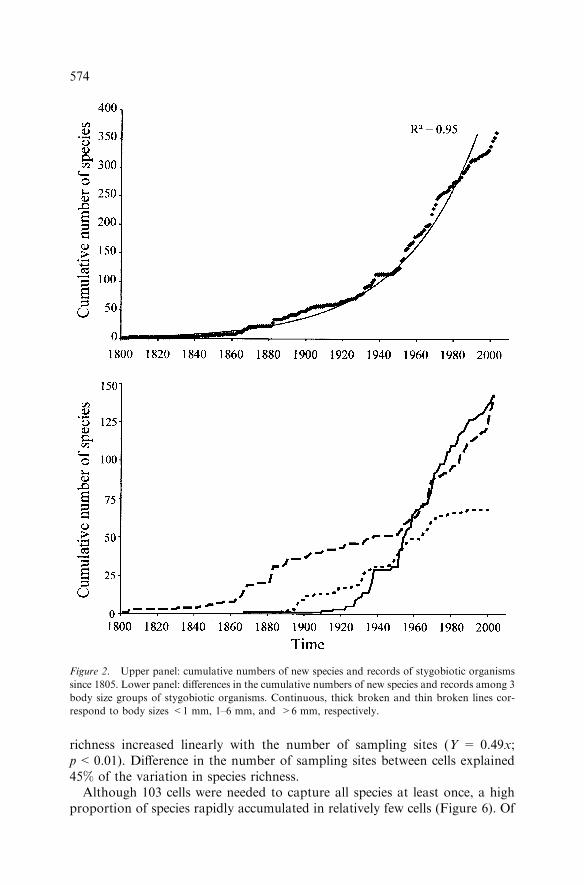
(Harpacticoida). Out of ten crustacean groups, six had more species withinstygobiotic fauna than in stygoxen fauna. Whereas insects represented about74% of all invertebrate species known from surface water, the stygobiotic faunacomprised only two insect species belonging to the genus Siettitia (ColeopteraDytiscidae).The cumulative number of species increased exponentially with time
(Figure 2). During the last 50 years, the number of species increased three-fold(i.e. from 124 in 1951 to 380 in 2003). However, the rate of reporting of newspecies and records has decreased since the 80s. From 1978 to 2003, only 82species were reported whereas 136 species had been reported during the sameduration from 1948 to 1973. The rate of reporting of species belonging to themeiofauna (i.e. body size <1 mm) suddenly increased in the 30s, and sincethen, has remained markedly higher than the reporting rate of speciesbelonging to the macrofauna (body size >1 mm) (Figure 2). Consequently, theproportion of meiofaunal species (i.e. almost all Copepoda, Ostracoda,Syncarida and Branchiopoda) increased from 4% in 1909 to 40% in 2003.There was almost a 100-year phase difference between the curves of the
cumulative number of stygoxen and stygobiotic species of crustaceans(Figure 3). About 80% of stygoxen species were described before 1900, whereasonly 4% of stygobiotic species had been reported. Since the 70s, only fivestygoxen species have been described against 70 species for the stygobioticfauna. Despite this pronounced time lag between the curves, the present-daynumber of stygobiotic crustaceans (246 species among which 18 undescribedspecies) almost equals that of stygoxenes (266 species).
Geographic patterns
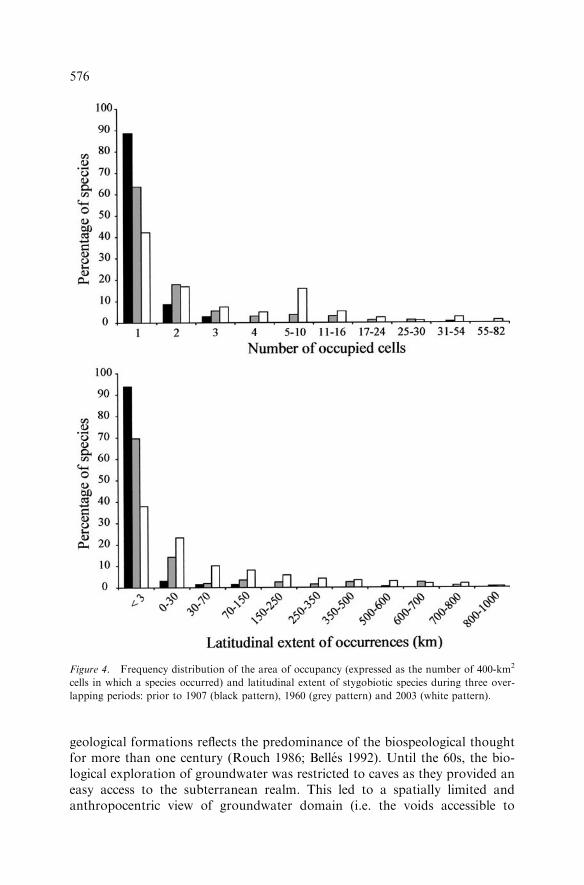
The distributions of range size were strongly skewed towards small areas ofoccupancy and latitudinal extents (Figure 4). In 2003, 41% of species werecollected from a single cell and 38% had a latitudinal extent less than 3 km.Only 5 and 2.5% of species had an area of occupancy higher than 25 cells and alatitudinal extent higher than 600 km, respectively.However, the average and maximum range sizes increased over time. From
1960 to 2003, the average area of occupancy and average latitudinal extentincreased from 2.2±3.9 to 5.6±10.4 cells and from 58±159 to102±184 km, respectively. The maximum area of occupancy increased fromthree cells in 1907 to 54 and 82 cells in 1960 and 2003, respectively. Themaximum latitudinal extent was only 82 km in 1907 but 900 and 940 km,respectively, in 1960 and 2003.Over the last 100 years, the distribution patterns of species richness changed
dramatically (Figure 5). From 1907 to 2003, the proportion of cells containingno species decreased by 35%. Meantime, the number of species in the richestcell increased from 4 to 52. However, in 2003, only 2.7% of the cells containedmore than 11 species, whereas 33% had between 1 and 10 species. Species
573
richness increased linearly with the number of sampling sites (Y = 0.49x;p<0.01). Difference in the number of sampling sites between cells explained45% of the variation in species richness.Although 103 cells were needed to capture all species at least once, a high
proportion of species rapidly accumulated in relatively few cells (Figure 6). Of
Figure 2. Upper panel: cumulative numbers of new species and records of stygobiotic organisms
since 1805. Lower panel: differences in the cumulative numbers of new species and records among 3
body size groups of stygobiotic organisms. Continuous, thick broken and thin broken lines cor-
respond to body sizes <1 mm, 1–6 mm, and >6 mm, respectively.
574

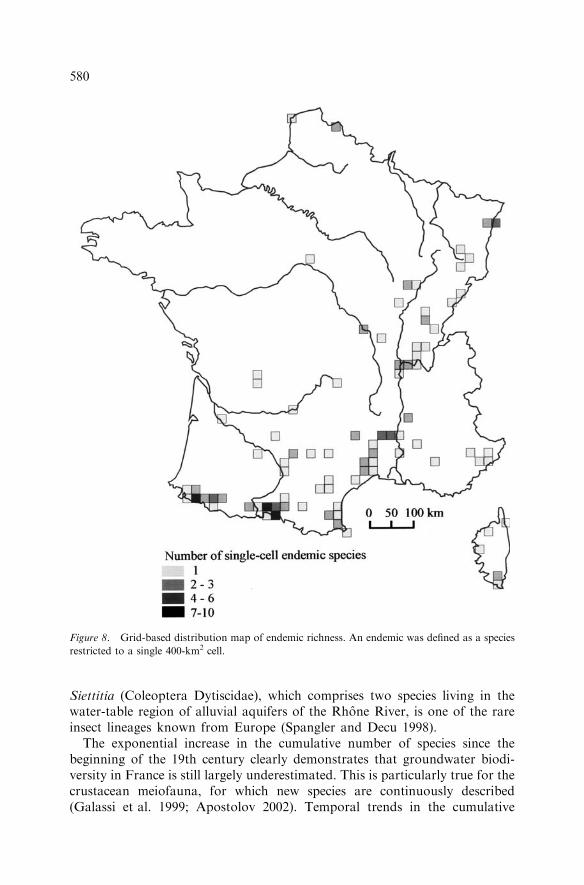
a total of 380 species, 227 (60%) were represented in only 17 cells and 75% ofspecies were represented in 34 cells. Beyond 34 cells, adding an additional cellresulted in an increase of only two or one species. Figure 7 indicates theselection frequency of cells during 1000 runs of the optimization algorithm.Only 145 cells out of 445 appeared in at least one solution, among which 90appeared in all solutions, representing 362 species. Of the 90 cells appearing inall solutions, 86 contained the entire distribution of one or more species andwere therefore irreplaceable. The location in southern France of two thirds ofever-selected cells reflected the concentration of endemic species. Of the 156species restricted to a single cell (i.e. single-cell endemics), 114 occurred in thesouthern third of France (Figure 8).
Discussion
Sampling effort
Groundwater fauna has certainly been more investigated in France than inmost other countries (except Slovenia). A total of 1800 caves were sampled inEurope within the framework of the Biospeologica program, among which60% were located in France (Juberthie and Ginet 1994). The French databaseincludes about the same number of records for the aquatic cave fauna (i.e. 3000records) as the U.S. database (i.e. 2774 records, but see Culver et al. 1999,2000). However, sampling is still highly incomplete and spatially concentratedin limestone regions. The disparity in sampling effort between karsts and other
Figure 3. Cumulative numbers of new species and records of stygobiotic (open squares) and
stygoxen (full diamonds) crustaceans since 1758.
575
geological formations reflects the predominance of the biospeological thoughtfor more than one century (Rouch 1986; Belles 1992). Until the 60s, the bio-logical exploration of groundwater was restricted to caves as they provided aneasy access to the subterranean realm. This led to a spatially limited andanthropocentric view of groundwater domain (i.e. the voids accessible to
Figure 4. Frequency distribution of the area of occupancy (expressed as the number of 400-km2
cells in which a species occurred) and latitudinal extent of stygobiotic species during three over-
lapping periods: prior to 1907 (black pattern), 1960 (grey pattern) and 2003 (white pattern).
576

Figure 5. Grid-based distribution map of species richness during three overlapping periods: prior
to 1907 (upper panel), 1960 (middle panel) and 2003 (lower panel).
577

humans), which retarded the comprehension of their structure and functioning.However, research conducted during the last 40 years in a variety of uncon-solidated sediments (e.g. marine, alluvial, colluvial and morainic deposits)revealed that stygobiotic animals were likely to be found wherever there wasgroundwater. The disproportionate sampling effort allocated to karst aquifersin many regions probably biases our estimate of groundwater biodiversitybecause nearby karst and alluvial aquifers usually harbor dissimilar inverte-brate assemblages. Rouch (1988) showed that the Baget karst system, Pyrenees,and its ensuing alluvial aquifer contained 22 and 21 species, respectively, withonly 12 species in common. Similarly, the Dorvan karst system, Jura, and thecontiguous alluvial aquifer of the Albarine River harbored 22 and 21 species,respectively, but shared only 8 species Gibert et al. 2000). About 47 and 29% ofspecies collected in France are exclusive to karst and unconsolidated habitats,respectively, and 20% occur in both habitats,
Taxonomic patterns
Groundwater fauna of France is more diversified (i.e. 380 species) than thatof other countries including U.S.A. (300 species; Culver et al. 2000), Italy(265 species; Stoch 2001), Slovenia (210 species; Sket 1999a) and Croatia(about 200 species; Gottstein-Matocec et al. 2002). This elevated number ofspecies is most likely the result of a higher sampling effort in various aquifers.Although all invertebrate groups are represented in groundwater, the stygo-biotic fauna is dominated by crustaceans at different systematic levels. Thisoverrepresentation of crustaceans (i.e. 65% of species) has been observed ingroundwater of all temperate regions (Sket 1999a, b). Danielopol et al. (2000)estimated that the stygobiotic fauna accounted for 40% of the total numberof aquatic crustacean species in Europe. The lack of competitors such as
Figure 6. Species accumulation curve for 103 cells representing all species at least one.
578

aquatic insects has been advocated by several authors for explaining the highdiversity of crustaceans in groundwater (Stoch 1995; Sket 1999b). Beingdependent upon air for breathing or reproducing, aquatic insects areextremely rare in groundwater whereas they represent 50% of all animalspecies living in surface water of Europe (Danielopol et al. 2000). The genus
Figure 7. Grid-based distribution map of species richness showing the location and selection
frequency of cells for representing all species at least one (out of 1000 runs of the selection algo-
rithms). Cells numbered from 1 to 17 correspond to the 17 first most-complementary cells con-
taining 60% of known species.
579
Siettitia (Coleoptera Dytiscidae), which comprises two species living in thewater-table region of alluvial aquifers of the Rhone River, is one of the rareinsect lineages known from Europe (Spangler and Decu 1998).The exponential increase in the cumulative number of species since the
beginning of the 19th century clearly demonstrates that groundwater biodi-versity in France is still largely underestimated. This is particularly true for thecrustacean meiofauna, for which new species are continuously described(Galassi et al. 1999; Apostolov 2002). Temporal trends in the cumulative
Figure 8. Grid-based distribution map of endemic richness. An endemic was defined as a species
restricted to a single 400-km2 cell.
580

number of obligate groundwater and surface water species suggest thatgroundwater comprises more crustaceans than surface freshwater. This lendssupport to the idea that the contribution of stygobiotic fauna to continentalfreshwater biodiversity is probably far more important than previously sus-pected (Gibert and Deharveng 2002). Decrease in the report rate of species forthe last 20 years essentially reflects the deteriorating capacity of an everdecreasing number of active taxonomists to identify biological material andpublish updated lists of species (Reid 1992; Galassi 2001; Valdecasas andCamacho 2003). As observed for two decades in all branches of biology (Oliver1988; Gaston and May 1992), this crisis in biosystematics impoverishes con-siderably the quality of taxonomic information which is, however, the mostfundamental requirement for any assessment and study of groundwater bio-diversity. A high proportion of records (i.e. from 12% for Copepoda up to 52%for Syncarida) that are not identified to species do not figure on distributionmaps and there is an increasing number of potentially new species (i.e. about 19species) still awaiting description. On the other hand, new information forunderrepresented taxonomic groups such as Oligochaeta progressively becomeavailable (Martınez-Ansemil et al. 1997; Giani et al. 2001).
Geographic patterns
Although the average latitudinal extent and area of occupancy of groundwaterspecies increased over time as distributional data accumulated, the geographicrange size of most stygobionts remained extremely small compared to those ofepigean species. Nearly 70% of stygobiotic species collected in France are re-stricted to the French landscape (including Corsica). For comparison, theFrench fauna of Ephemeroptera and Odonata (i.e. 252 species) have 80 and100% species in common with the Italian and Swiss fauna, respectively. Sket(1999a) reported that the Dinaric Mountains, probably the richest ground-water region in the world in relation to its area, also shared very fewstygobiotic species with neighboring regions (i.e. 22% of species). Recentgenetic studies reinforced the significance of endemism within groundwaterfauna by demonstrating that a number of species identified on the basis ofmorphological criteria corresponded in fact to clusters of distinct species(Cobolli-Sbordoni et al. 1990; Ketmaier et al. 2000; Lefebure et al. 2006).The exceptional richness of endemics is directly linked to the highly fragmentednature of the groundwater domain that fosters evolution processes (i.e. speci-ation) leading to the isolation of populations (Holsinger 2000; Humphreys2000). Because endemism is typically higher in groundwater than in surfacewater fauna, the contribution of groundwater fauna to the richness of conti-nental water increases with increasing spatial scale (Culver and Sket 2000;Gibert and Deharveng 2002).The high proportion of single-cell endemics in the southern third of France
(i.e. 73%) was unlikely the result of incomplete sampling. The Pyrenees
581

contained five time more endemic species (52 species) than the Jura (10 species),although these two mountain areas received comparable sampling efforts.Distribution maps of stygobiotic species recently produced within the frame-work of the European project PASCALIS also revealed that more than 80% ofsingle-cell endemic species (0.2 · 0.2 decimal degree cell) occurred in SouthernEurope (J. Gibert, unpublished data). The southern concentration of endemic-rich areas supports the hypothesis that the distribution of stygobiotic species,most of which colonized groundwater several millions years ago, was stronglymodified by cold Pleistocene events (Ginet 1971; Holsinger et al. 1983; Strayeret al. 1995; Magniez 1997). Many endemic species that were particularly vul-nerable due to their small population size might have gone extinct in northernareas during the last ice age.The distribution map of species richness changed dramatically over time as
more extensive surveys provided additional records, clearly indicating that thelocation of biodiversity hotspots varied as a function of sampling effort. Sincethe 60s, sampling in alluvial aquifers of the Rhone River resulted in several newhotspots. In contrast, Culver et al. (2000) argued that the concentration ofbiodiversity within a few cave regions of the U.S.A. was not sensitive to anincrease in sampling effort. In France, most species-rich cells (>20 species)corresponded to areas that received a disproportionate amount of samplingeffort. These included areas that were located within a short distance to re-search facilities such as the subterranean laboratory in Moulis, Pyrenees (52species), the groundwater ecology laboratory in Lyon (43 species), and thelaboratory of animal biology in Dijon, Cote-d’Or (24 species). Several studiesdemonstrated that sampling was positively biased towards areas designated ashotspots of biodiversity (Nelson et al. 1990; Reddy and Davalos 2003). Con-versely, this concentration of species in highly sampled areas strongly suggestedthat species richness was underestimated in most groundwater areas of France.For example, we would expect all cells located along the southern part of theRhone River to contain at least 20 species because most interstitial stygobiontscollected in the vicinity of the Lyon city are also known to occur in alluvialgroundwater in southern France.
Conservation implications
Although several studies documented the adverse effects of groundwater con-tamination and extraction on invertebrate assemblages (Rouch et al. 1993;Malard et al. 1996; Mosslacher and Notenboom 2000), the diverse and vul-nerable groundwater fauna of France has received almost no conservationattention to date (Bouchet 1990; Juberthie 1995). Despite the extremely narrowdistribution range of most obligate groundwater species known from France,only 20 species (18 molluscs, 1 copepod and 1 coleopteran) were included in thethreatened species list of the International Union for Conservation of Natureand Natural Resources (IUCN 2004). However, listing of threatened species
582

under the IUCN act or European Habitat Directive is only a minor step forconserving groundwater biodiversity. As stated by Culver et al. (2000, p. 395),‘‘there are simply too many subterranean species at risk to deal with them oneat time’’. Indeed, almost all groundwater species of France could potentially belisted in the European habitat directive based on legislative criteria used fordefining priority species (i.e. endemism, rarity, species characteristic of a singlebiogeographic region). A more efficient conservation solution is to protect aminimum network of complementary aquifers that maximizes the representa-tion of as many species as possible while minimizing the risk of biodiversity lossdue to human activities (Gaston et al. 2002; Margules et al. 2002). This strategywhich pays explicit attention to patterns of between-aquifer complementarity at anational scale enables the maintenance of regionally distinctive species-richassemblages while minimizing the duplication of conservation efforts acrossregional agencies (Mace 2000). Whereas heuristic algorithms are commonlyused for designing effective reserve networks in terrestrial and marine systems(Csuti et al. 1997; Stewart et al. 2003), the designation of priority groundwatersites is still essentially based on traditional selection methods (selection ofhotspots of species richness and of endemism). Culver et al. (2000) applied forthe first time a greedy algorithm to delineate potential subterranean prioritysites in the U.S.A and showed that 50% of obligate cave fauna occurred in lessthan 1% of the landscape. We also demonstrated using an adaptive simulatedannealing that only 1.2% (i.e. 17 cells) of the French territory was needed tocapture 60% of known stygobiotic species. Thus, it should be possible toconserve a large proportion of species by focusing habitat conservation effortsin a few complementary species-rich aquifers (i.e. conservation targets) locatedin distinct regions including the Pyrenean Mountains (cells 1, 5, 11, and 12 inFigure 7), the Roussillon Plain (9, 15), the North-Montpellieran karsts (3, 7,17), the Ardeche calcareous plateau (4, 10), the lower Rhone valley (2, 16), theOccidental Jura (13, 14), the Burgundy karsts (8), and the Rhine valley (6).Although many of these species-rich aquifers were previously identified byother scientists (Juberthie and Juberthie-Jupeau 1975; Malard et al. 1997;Gibert et al. 2000), their present-day location within each region reflects tosome extent the distribution of data-rich areas.We identified two major research avenues for implementing an effective
groundwater reserve network in France. A first important step is to shift from agrid cell selection approach to an aquifer selection approach that incorporatesnot only representation targets of species and minimum space requirement butalso socio-economic costs related to the vulnerability of aquifers and strengthof human activity in the catchment. Since the objective function of Marxanallows assigning different costs for each aquifer (Ball and Possingham 2001),the resultant reserve network would not only depend on the ecological valuesof aquifers but also on their conservation costs linked to socio-economicconstrains. The use of aquifers as planning units is all the more importantbecause aquifer classifications based on relevant ecological criteria (i.e. per-meability, pore size, hydrological connection with surface environment) can be
583

used as abiotic surrogates of groundwater biodiversity in data-poor regions. Asecond critical step consists in developing appropriate sampling strategies inresponse to well-defined conservation goals. Because beta diversity amongregions (i.e. dissimilarity in species composition between regions) makes by farthe highest contribution to total richness of stygobionts in groundwater ofFrance (Ferreira et al. unpublished), the inclusion of new conservation targetsnecessitates sampling aquifers in supposedly rich regions that have not yet beeninvestigated. In contrast, increasing the flexibility of the network of conser-vation targets requires sampling additional aquifers in regions that have al-ready been sampled. Although the high level of endemism within the obligategroundwater fauna would necessarily reduce the latitude in placement of pri-ority areas for conservation, the present lack of flexibility of the reserve net-work is primarily attributable to the weakness of sampling effort in mostregions. The use of aquifers as planning units, the integration of conservationcosts, and the urgent need for updating biological data are interdisciplinarygoals that require hydrogeologists, socio-economists, and biologists to worktogether for implementing an effective groundwater reserve network.
Acknowledgements
This work was supported by the European program PASCALIS (Protocols forthe ASsessment and Conservation of Aquatic Life In the Subsurface) (EVK2-CT-2001-00121). We thank the numerous scientists who provided data andcorrections: C. Bou, N. Coineau, A. Brancelj, L. Deharveng, M. andG. Falkner, D. Galassi, N. Giani, R. Ginet, C. Jouin-Toulmond, J. Juget,F. Lescher-Moutoue, G. Magniez, J.-P. Henry, P. Marmonier, J. Mathieu,P. Richoux, and M.-J. Turquin.
Appendix 1. List of groundwater obligate species of France.
NEMERTINA
Tetrastemmatidae
Prostoma puteale de Beauchamp 1932
PLANARIA
Dendrocoelidae
Dendrocoelopsis beauchampi (Gourbault 1969)
Dendrocoelopsis bessoni Gourbault, Benazzi & Helleouet 1976
Dendrocoelopsis brementi (de Beauchamp 1919)
Dendrocoelopsis chattoni (de Beauchamp 1949)
Dendrocoelopsis garmieri (de Beauchamp 1950)
Dendrocoelopsis vandeli (de Beauchamp 1931)
Dendrocoelum (Bolbodendrocoelum) agile de Beauchamp 1932
Dendrocoelum (Dendrocoelides) barbei de Beauchamp 1956
Dendrocoelum (Dendrocoelides) coiffaiti de Beauchamp 1956
Dendrocoelum (Dendrocoelides) col1ini (de Beauchamp 1919)
584

Appendix 1. (continued)
Dendrocoelum (Dendrocoelides) lescherae Gourbault 1971
Dendrocoelum (Dendrocoelides) regnardi (de Beauchamp 1919)
Dendrocoelum (Dendrocoelides) tuzetae Gourbault 1965
Dendrocoelum (Dendrocoelum) infernale (Steinmann 1907)
Dendrocoelum (Eudendrocoelum) gineti de Beauchamp 1954
Dendrocoelum (Eudendrocoelum) remyi de Beauchamp 1926
Dendrocoelum (Polycladodes) album (Steinmann 1910)
Dendrocoelum sollaudi de Beauchamp 1931
Miodendrocoelum parisi de Beauchamp 1929
Planariidae
Atrioplanaria delamarei Gourbault 1969
Atrioplanaria notadena de Beauchamp 1937
Phagocata albissima (Vejdovsky 1883)
Phagocata vitta (Duges 1830)
Plagnolia vandeli de Beauchamp & Gourbault 1964
ANNELIDA, HIRUDINEA
Erpobdellidae
Trocheta bykowskii Gedroyc 1913
ANNELIDA, APHANONEURA
Aeolosomatidae
Aeolosoma gineti Juget 1959
Potamodrilidae
Potamodrilus fluviatilis Lastochkin 1935
ANNELIDA, POLYCHAETA
Nerillidae
Troglochaetus beranecki Delachaux 1921
ANNELIDA, OLIGOCHAETA
Enchytraeidae
Enchytraeus flavus Moszynski 1938
Pachydrilus fossor Vejdovsky 1877
Haplotaxidae
Delaya corbarensis (Delay 1972)
Haplotaxis leruethi (Hrabe 1958)
Lumbriculidae
Cookidrilus speluncaeus Rodriguez & Giani 1987
Trichodrilus capilliformis Rodriguez & Giani 1994
Trichodrilus cernosvitovi Hrabe 1937
Trichodrilus intermedius (Fauvel 1903)
Trichodrilus 1eruthi Hrabe 1937
Trichodrilus pragensis (Vejdovsky 1875)
Trichodrilus tenuis Hrabe 1960
Tubificidae
Abyssidrilus cuspis (Erseus & Dumnicka 1988)
Gianius labouichensis (Rodriguez & Giani 1989)
Gianius cavealis Juget & Des Chatelliers 2001
Haber turquini (Juget & Lafont 1979)
Krenedrilus sergei Giani, Erseus & Martinez-Ansemil 1990
Rhyacodrilus amphigenus Juget 1987
Rhyacodrilus balmensis Juget 1959
Rhyacodrilus lindbergi Hrabe 1963
Rhyacodrilus subterraneus Hrabe 1963
Spiridion phreaticola (Juget 1987)
585

Appendix 1. (continued)
MOLLUSCA
Amnicolidae
Bythinella bouloti Girardi, Bichain & Wienin 2002
Bythinella cylindracea (Paladilhe 1869)
Bythinella eutrepha (Paladilhe 1867)
Bythinella galerae Girardi, Bichain & Wienin 2002
Bythinella geisserti Boeters & Falkner 2003
Bythinella padiraci Locard 1903
Bythinella pupoides phreaticola Bernasconi 1989
Hydrobiidae
Alzoniella (Alzoniella) haicabia Boeters 2000
Alzoniella (Alzoniella) navarrensis Boeters 1999
Alzoniella (Alzoniella) pyrenaica (Boeters 1983)
Alzoniella (Alzoniella) ,junqua Boeters 2000
Alzoniella (Navariella) elliptica (Paladilhe 1874)
Avenionia bourguignati (Locard 1883)
Avenionia brevis (Draparnaud 1805)
Avenionia berenguieri (Bourguignat 1882)
Belgrandiella saxatilis (Reynies 1844)
Belgrandiella ? dunalina (Moquin-Tandon 1856)
Fissuria boui Boeters 1981
Graziana provincialis (Boeters 2000)
Graziana rayensis (Caziot 1910)
Graziana ? cezairensis Boeters 2000
Heraultiella exilis (Paladilhe 1867)
Islamia bomangiana Boeters & Falkner 2003
Islamia bourguignati (T. Letourneux 1869)
Islamia consolationis (Bernasconi 1985)
Islamia germaini Boeters et Falkner 2003
Islamia minuta (Draparnaud 1805)
Islamia moquiniana (Dupuy 1851)
Islamia spirata (Bernasconi 1985)
Istriana falkneri Boeters 2000
Moitessieriidae
Bythiospeum anglesianum (Westerlund 1890)
Bythiospeum articense Bernasconi 1985
Bythiospeum bourguignati (Paladilhe 1866)
Bythiospeum bressanum Bernasconi 1985
Bythiospeum charpyi charpyi (Paladilhe 1867)
Bythiospeum charpyi giganteum Bernasconi 1969
Bythiospeum diaphanoides Bernasconi 1985
Bythiospeum diaphanum diaphanum (Michaud 1831)
Bythiospeum diaphanum michaellense Girardi 2002
Bythiospeum dorvani Bernasconi 1985
Bythiospeum drouetianum (Cless in 1882)
Bythiospeum francomontanum Bernasconi 1973
Bythiospeum garnieri (Sayn 1889)
Bythiospeum klemmi (Boeters 1969)
Bythiospeum michaudi (Locard 1882)
Bythiospeum moussonianum (Paladilhe 1869)
Bythiospeum racovitzai (Germain 1911)
586

Appendix 1. (continued)
Bythiospeum rhenanum rhenanum (Lais 1935)
Bythiospeum terveri (Locard 1882)
Henrigirardia Wienini Girardi 2001
Moitessieria bourguignati Coutagne 1883
Moitessieria cocheti Boeters & Falkner 2003
Moitessieria heideae Boeters & Falkner 2003
Moitessieria fontsaintei Bertrand 2001
Moitessieria,juvenisanguis Boeters & E. Gittenberger 1980
Moitessieria lescherae Boeters 1981
Moitessieria locardi Coutagne 1883
Moitessieria massoti Bourguignat 1863
Moitessieria nezi Boeters & Bertrand 2001
Moitessieria olleri Altimira 1960
Moitessieria rhodani Coutagne 1883
Moitessieria rolandiana Bourguignat 1863
Moitessieria simoniana (Saint-Simon 1848)
Palacanthilhiopsis vervierii Bernasconi 1988
Palacanthilhiopsis ? margritae Boeters & Falkner 2003
Paladilhia conica Paladilhe 1867
Paladilhia gloeeri Boeters & Falkner 2003
Paladilhia pleurotoma Bourguignat 1865
Paladilhia pontmartiniana (Nicolas 1891)
Paladilhia sp.
Paladilhia umbilicata (Locard 1902)
Palaospeum bessoni bessoni (Bernasconi 1999)
Palaospeum bessoni rebenacqense Boeters & Bertrand 2001
Palaospeum? nanum Boeters & Bertrand 2001
Plagigeyeria deformata (Nicolas 1891)
Spiralix burgundina (Locard 1882)
Spiralix collieri (Nicolas 1891)
Spiralix corsica (Bernasconi 1994)
Spiralix hofmanni Boeters & Falkner 2003
Spiralix puteana (Coutagne 1883)
Spiralix rayi (Locard 1882)
Spiralix vitrea (Draparnaud 1801)
CRUSTACEA
CLADOCERA
Chydoridae
Alona bessei Dumont 1983
Alona phreatica Dumont 1983
DECAPODA
Atyidae
Troglocaris inernris Fage 1937
ISOPODA
Stenasellidae
Stenasellus buili Remy 1949
Stenasellus racovitzai Razzauti 1925
Stenasellus virei angelieri Magniez 1968
Stenasellus virei boui Magniez 1968
Stenasellus virei hussoni Magniez 1968
Stenasellus virei virei Dollfus 1897
587

Appendix 1. (continued)
Asellidae
Gallasellus heilyi (Legrand 1956)
Proasellus albigensis (Magniez 1965)
Proasellus aquaecalidae (Racovitza 1922)
Proasellus beroni/ Henry & Magniez 1968
Proasellus boui Henry & Magniez 1969
Proasellus burgundus Henry & Magniez 1969
Proasellus cavaticus (Leydig 1871)
Proasellus chauvini Henry & Magniez 1978
Proasellus claudei Henry & Magniez 1996
Proasellus coiffaiti Henry & Magniez 1972
Proasellus coxalis (Dollfus 1892)
Proasellus meridianus (Racovitza 1919)
Proasellus nolli (Karaman 1952)
Proasellus racovitzai Henry & Magniez 1972
Proasellus rouchi Henry 1980
Proasellus spelaeus (Racovitza 1922)
Proasellus strouhali puteanus (Henry 1966)
Proasellus synaselloides (Henry 1963)
Proasellus valdensis (Chappuis 1948)
Proasellus vandeli Henry & Magniez 1969
Proasellus walteri (Chappuis 1948)
Proasellus sp. 1
Microparasellidae
Microcharon angelieri Coineau 1963
Microcharon boui Coineau 1968
Microcharon doueti Coineau 1968
Microcharon juberthiei juberthiei Coineau 1968
Microcharon juberthiei ramosus Coineau 1968
Microcharon reginae Dole & Coineau 1987
Microcharon rouchi Coineau 1968
Microcharon sisiphus Chappuis & Delamare 1954
Microcharon sp. 1
Microcharon sp. 2
Microcharon sp. 3
Microcharon sp. 4
Microcharon sp, 5
Microcharon sp. 6
Microcharon sp. 7
Sphaeromatidae
Caecosphaeroma burgundum burgundum Dollfus 1898
Caecosphaerorma burgundum rupisfucaldi Hubault 1934
Caecosphaeroma virei Dollfus 1896
Cirolanidae
Sphaeromides raymondi Dollfus 1897
Faucheria faucheri Dollfus & Vire 1900
Cirolanidae n. sp.
AMPHIPODA
Karstogiella lautieri
Bogidiellidae
Bogidiella albertimagni Hertzog 1933
588

Appendix 1. (continued)
Bogidiella nicolae Karaman 1988
Medigidiella chappuisi Ruffo & Delaimare 1952
Niphargidae
Niphargus angelieri Ruffo 1953
Niphargus aquilex Schiodte 1855
Niphargus balazuci Schellenberg 1951
Niphargus boulangei Wichers 1964
Niphargus burgundus Graf & Straskraba 1967
Niphargus ciliates Chevreux 1906
Niphargus corsicanus Schellenberg 1950
Niphargus delamarei Ruffo 1953
Niphargus fontanus Bate 1859
Niphargus foreli Humbert 1877
Niphargus gallicus Schellenberg 1935
Niphargus gineti Bou 1965
Niphargus gr.jovanovici S, Karaman 1931
Niphargus kieferi Schellenberg 1936
Niphargus kochianus kochianus Bate 1859
Niphargus ladmiraulti Chevreux 1901
Niphargus laisi Schellenberg 1936
Niphargus nicaensis Isnard 1916
Niphargus pachypus Schellenberg 1933
Niphargus plateaui Chevreux 1901
Niphargus renei G, Karaman 1986
Niphargus rhenorhodanensis Schellenberg 1937
Niphargus robustus Chevreux 1901
Niphargus schellenbergi S, Karaman 1932
Niphargus setiferus Schellenberg 1937
Niphargus thienemanni Schellenberg 1934
Niphargus vandeli Barbe 1961
Niphargus virei Chevreux 1896
Niphargopsis casparyi (Pratz 1866)
Crangonyctidae
Crangonyx subterraneaus Bate 1859
Salentinellidae
Parasalentinella rouchi Bou 1971
Salentinella angelieri Ruffo & Delamare 1952
Salentinella delamarei delamarei Coineau 1962
Salentinella delamarei macrocheles Coineau 1968
Salentinella gineti Balazuc 1957
Salentinella major Barbe 1965
Salentinella petiti Coineau 1968
Salentinella sp. 1
Pseudoniphargidae
Pseudoniphargus adriaticus S. Karaman 1955
Ingolfiellidae
Ingolfiella (Tyrrhenidiella) catalanensis Coineau 1963
Ingolfiella (Tyrrhenidiella) thibaudi Coineau 1968
OSTRACODA
Ostracoda n. sp.
Candonidae
589

Appendix 1. (continued)
Candonopsis boui Danielopol 1978
Cryptocandona kieferi (Klie 1938)
Fabaeformiscandona breuili (Paris 1920)
Fabaeformiscandona cf. breuili sp.1
Fabaeformiscandona cf breuili sp. 2
Fabaeformiscandona wegelini (Petkovski 1962)
Mixtocandona juberthieae Danielopol 1978
Mixtocandona laisi (Klie 1938)
Pseudocandona delamarei Danielopol 1978
Pseudocandona rouchi Danielopol 1973
Pseudocandona simililampadis Danielopol 1978
Pseudocandona zschokkei (Wolf 1920)
Schellencandona belgica (Klie 1937)
Schellencandona cf. schellenbergi sp. 1
Schellencandona cf. schellenbergi sp. 2
Schellencandona triquetra (Klie 1936)
Cyprididae
Cavernocypris subterranea (Wolf 1920)
Psychrodromus betharrarmi Baltanas, Danielopol, Roca & Marmonier 1993
Entocytheridae
Sphaeromicola cebennica cebennica Remy 1948
Sphaeromicola cebennica juberthiei Danielopol 1977
Sphaeromicola hamigera Remy 1948
Sphaeromicola topsenti Paris 1916
COPEPODA, CYCLOPOIDA
Cylopidae
Acanthocyclops hispanicus Kiefer 1937
Acanthocyclops rhenanus Kiefer 1936
Acanhocyclops sensitivus (Graeter & Chappuis 1914)
Diacyclops cf. paolae Pesce & Galassi 1987
Diacyclops clandestinus Kiefer 1933
Diacyclops zschokkei (Graeter 1910)
Eucyclops graeteri (Chappuis 1927)
Graeteriella (Paragraeteriella) bertrandi Lescher-Moutoue 1974
Graeteriella (Paragraeteriella) gelyensis Lescher-Moutoue1978
Graeteriella (Paragraeteriella) laisi (Kiefer 1936)
Graeteriella (Paragraeteriella) vandeli Lescher-Moutoue 1969
Graeteriella boui Lescher-Moutoue 1974
Graeteriella brehmi Lescher-Moutoue 1968
Graeteriella rouchi Lescher-Moutoue 1968
Graeteriella unisetigera (Graeter 1910)
Kieferiella delamarei (Lescher-Moutoue 1971)
Megacyclops brachypus Kiefer 1954
Speocyclops arregladensis Chappuis & Kiefer 1952
Speocyclops anomalus Chappuis & Kiefer 1952
Speocyclops castereti Lindberg 1954
Speocyclops demetiensis (Scourfield 1932)
Speocyclops gallicus Chappuis & Kiefer 1952
Speocyclops kieferi Lescher-Moutoue 1968
Speocyclops orcinus Kiefer 1937
Speocyclops proserpinae Kiefer 1937
590

Appendix 1. (continued)
Speocyclops racovitzai (Chappuis 1923)
Speocyclops racovitzai boscensis Kiefer 1954
Speocyclops racovitzai gouillounensis Kiefer 1954
Speocyclops racovitzai incerta Chappuis & Kiefer 1952
Speocyclops racovitzai liguensis Chappuis & Kiefer 1952
Speocyclops racovitzai noustensis Chappuis & Kiefer 1952
Speocyclops racovitzai peyortensis Chappuis & Kiefer 1952
Speocyclops racovitzai,sabartensis Kiefer 1954
Speocyclops racovitzai sandetsi Chappuis & Kiefer 1952
Speocyclops sisyphus Kiefer 1937
COPEPODA, HARPACTICOIDA
Ectinasomatidae
Pseudectinosoma vandeli (Rouch 1969)
Pseudectinosoma janineae Galassi, Dole-Olivier & De Laurentiis 1999
Ameiridae
Parapseudoleptomesochra subterranea subterranea (Chappuis 1928)
Parapseudoleptomesochra subterranea deminuta (Chappuis 1928)
Nitocrellopsis elegans (Chappuis & Rouch 1959)
Nitocrellopsis rouchi Galassi, De Laurentiis & Dole-Olivier 1999
Nitocrella beatricis Cottarelli & Bruno 1994
Nitocrella delayi Rouch 1970
Nitocrella gracilis Chappuis & Rouch 1959
Nitocrella dussarti Chappuis 1955
Nitocrella omega Hertzog 1936
Nitocrella sp.1 groupe hirta
Nitocrella sp, 2 groupe hitra
Canthocamptidae
Elaphoidella boui Rouch 1988
Elaphoidella bouilloni Rouch 1964
Elaphoidella brehieri Apostolov 2001
Elaphoidella calypsonis Chappuis & Rouch 1959
Elaphoidella cavatica Chappuis 1957
Elaphoidella coiffaiti Chappuis & Kiefer 1952
Elaphoidella elaphoides elaphoides (Chappuis 1923/124)
Elaphoidella federicae Pesce & Galassi 1988
Elapheidella garbeteneis Rouch 1980
Elaphoidella infernalis Rouch 1970
Elaphoidella leruthi leruthi/ Chappuis 1937
Elaphordella leruthi meridionalis Chappuis 1953
Elaphoidella longifurcata Chappuis & Kiefer 1952
Elaphoidella madiracensis Apostolov 1998
Elaphoidella mauro Chappuis 1956
Elaphoidella pyrenaica Rouch 1970
Elaphoidella reducta Rouch 1964
Elaphoidella vandeli Chappuis & Rouch 1958
Elaphoidella vasconica Rouch 1970
Ceuthonectes boui Apostolov 2002
Ceuthonectes bulbiseta Apostolov 2002
Ceuthonectes chappuisi Rouch 1980
Ceuthonectes gallicus Chappuis 1928
Ceuthonectes serbicus Chappuis 1923/24
591

Appendix 1. (continued)
Ceuthonectes vievilleae Rouch 1980
Moraria catalana Chappuis & Kiefer 1952
Moraria varica (Graeter 1911)
Bryocamptus (Articocamptus) vandowei (Kessler 1914)
Bryocamptus (Articocamptus) zschokkei triarticulata Kiefer 1929
Bryocamptus (Rheocamptus) alosensis Apostolov 1998
Bryocamptus (Rheocamptus) dentatus Chappuis 1937
Bryocamptus (Rheocamptus) pyrenaicus (Chappuis 1923)
Bryocamptus (Rheocamptus) unisaetosus Kiefer 1930
Antrocamptus catherinae Chappuis & Rouch 1960
Antrocamptus chappuisi Rouch 1970
Antrocamptus coffaiti Chappuis 1956
Antrocamptus longifurcatus Rouch 1970
Anlrocamptus stygius Rouch 1970
Parastenocarididae
Parastenocaris aedes Hertzog 1938
Parastenocaris corsica Cottarelli, Bruno & Berera 2000
Parastenocaris dentulatus Chappuis & Rouch 1959
Parastenocaris dianae Chappuis 1955
Parastenocaris fontinalis borea Kiefer 1960
Parastenocaris fontinalis fontinalis Schnitter & Chappuis 1915
Parastenocaris fontinalis meridionalis Hertzog 1936
Parastenocaris glareola Hertzog 1936
Parastenocaris hippuris Hertzog 1938
Parastenocaris mangini Rouch 1990
Parastenocaris micheli Chappuis & Rouch 1959
Parastenocar is nertensis Rouch 1990
Parastenocaris nicolasi Rouch 1996
Parastenocaris psammica Songeur 1961
Parastenocaris silvana Cottarelli, Bruno & Berera 2000
Parastenocaris stammeri gallicus Chappuis & Rouch 1959
Parastenocaris vandeli Rouch 1987
COPEPODA, GELYELLOIDA
Gelyellidae
Gelyella droguei Rouch & Lescher-Moutoue 1977
COPEPODA, CALANOIDA
Diaptomidae
Spelaeodiaptomus rouchi Dussart 1970
SYNCARIDA
Bathynellidae
Bathynella gallica Delamare & Chappuis 1954
Bathynella natans natans Vejdovsky 1882
Bathynella natans picardi Delamare 1961
Bathynella pyrenaica Delamare & Chappuis 1954
Delamareibathynella debouttevillei Serban 1989
Delamareibathynella motasi Serban 1992
Gallobathynella (Clamousella) delayi (Serban, Coineau & Delamare 1971)
Gallobathynella (Gallobathynella) boui Serban, Coineau & Delamare 1971
Gallobathynella (Gallobathynella) coiffaiti (Delamare 1963)
Gallobathynella (Gallobaihynella) juberthiae Serban, Coineau & Delamare 1971
Gallobathynella (Gallobathynella) tarissei Serban, Coineau & Delarnare 1971
592

References
Apostolov A. 2002. Copepodes Harpacticoıdes stygobies de France. 1. Le genre Ceuthonectes
Chappuis, 1924 avec la description de deux especes nouvelles. Crustaceana 75: 777–790.
Ball J.R. and Possingham H.P. 2001. The design of marine protected areas: adapting terrestrial
techniques. In: Ghassemi F., Whetton P., Little R. and Littleboy M. (eds), Proceedings of the
International Congress on Modelling and Simulation, Natural Systems (vol 2). The Modelling
and Simulation Society of Australia and New Zealand Inc, Canberra, Australia, pp. 769–774.
Belles X. 1992. From dragons to allozymes. A brief account on the history of biospeleology. In:
Camacho A.I. (ed.), The Natural History of Biospeleology. Monografias del Museo National de
Ciencias Naturales, vol. 7, Madrid, pp. 3–24.
Botosaneanu L. 1986. Stygofauna Mundi, Brill, E.J. and Dr W. Backhuys Leiden, The
Netherlands.
Bouchet P. 1990. La malacofaune francaise: endemisme, patrimoine naturel et protection.
Rev. d’Ecol. 45: 259–288.
Cobolli-Sbordoni M., Mattocia M., La Rosa G., de Matthaeis E. and Sbordoni V. 1990. Secondary
sympatric occurrence of sibling species of subterranean shrimps in the karst. Int. J. Speleol. 19:
9–27.
Coineau N. 1971. Les Isopodes interstitiels. Documents sur leurecologie et leur biologie. Memoires
du Museum National d’Histoire Naturelle, Paris.
Csuti B., Polasky S., Williams P.H., Pressey R.L., Camm J.D., Kershaw M., Kiester A.R., Downs
B., Hamilton R., Huso M. and Sahr K. 1997. A comparison of reserve selection algorithms using
data on terrestrial vertebrates in Oregon. Biol. Conserv. 80: 83–97.
Culver D.C., Hobbs H.H.III, Christman M.C. and Master L.L. 1999. Distribution map of caves
and cave animals in the United States. J. Cave Karst Stud. 61: 139–140.
Culver D.C., Master L.L., Christman M.C. and Hobbs H.H.III 2000. Obligate cave fauna of the 48
contiguous United States. Conserv. Biol. 14: 386–401.
Culver D.C. and Sket B. 2000. Hotspots of subterranean biodiversity in caves and wells. J. Cave
Karst Stud. 62: 11–17.
Culver D.C., Christman M.C., Sket B. and Trontelj P. 2004. Sampling adequacy in an extreme
environment; species richness patterns in Slovenian caves. Biodivers. Conserv. 13: 1209–1229.
Appendix 1. (continued)
Meridiobathynella (Paraiberobathynella) rouchi Serban, Coineau & Delamare 1971
Parameridiobathynella gardensis Serban 1989
Pseudobathynella magniezi Serban, Coineau & Delaware 1971
Vandelibathrynella vandeli (Delamare & Chappuis 1954)
Vejdovskybathynella balazuci Serban & Leclerc 1984
Vejdovskybathynella espattyensis Serban & Leclerc 1984
Vejdovskybathynella leclerci/ Serban 1989
Parabathynellidae
Hexabathynella knoepffleri Coineau 1965
Parabathynella stygia Chappuis 1926
Paraiberobathynella (Paraiberobathynella) fagei (Delamare & Angelier 1950)
INSECTA
COLEOPTERA
Dytiscidae
Siettitia avenionensis Guignot 1925
Siettitia balsetensis Abeille de Perrin 1904
593

Danielopol D.L., Pospisil P. and Rouch R. 2000. Biodiversity in groundwater: a large-scale view.
Trends Ecol. Evol. 15: 223–224.
Danielopol D.L., Griebler C., Gunatilaka A. and Notenboom J. 2003. Present state and future
prospects for groundwater ecosystems. Environ. Conserv. 30: 104–130.
Ferreira D., Dole-Olivier M.-J., Malard F., Deharveng L. and Gibert J. 2003. Faune aquatique
souterraine de France: base de donnees et elements de biogeographie. Karstologia 42: 15–22.
Galassi D.M.P., De Laurentiis P. and Dole-Olivier M.-J. 1999. Nitocrellopsis rouchi sp. n., a new
ameirid harpacticoid from phreatic waters in France (Copepoda: Harpacticoida: Ameiridae).
Hydrobiologia 412: 177–189.
Galassi D.M.P. 2001. Groundwater Copepods: diversity patterns over ecological and evolutionary
scales. Hydrobiologia 453/454: 227–253.
Gaston K.J. 1991. How large is a species geographic range? . Oikos 61: 434–438.
Gaston K.J. and May R.M. 1992. Taxonomy of taxonomists. Nature 356: 281–282.
Gaston K.J., Pressey R.L. and Margules C.R. 2002. Persistence and vulnerability: retaining bio-
diversity in the landscape and in protected areas. J. BioSci. Suppl. 27: 361–384.
Geze B. 1973. Lexique des termes francais de speleologie physique et de karstologie. Ann. Speleol.
28: 1–20.
Giani N., Sambugar B., Rodriguez P. and Martınez-Ansemil E. 2001. Oligochaetes in southern
European groundwater: new records and an overview. Hydrobiologia 463: 64–74.
Gibert J., Danielopol D.L. and Stanford J.A 1994. Groundwater Ecology. Academic Press,
New York.
Gibert J., Malard F., Turquin M.-J. and Laurent R. 2000. Karst ecosystems in the Rhone River
basin. In: Wilkens H., Culver D.C. and Humphreys W.F. (eds), Subterranean Ecosystems.
Ecosystems of the World 30. Elsevier, Amsterdam, pp. 533–558.
Gibert J. 2001. Protocols for the assessment and conservation of aquatic life in the subsurface
(PASCALIS): a European Project. In: Culver D.C., Deharveng L., Gibert J. and Sasowsky I.D.
(eds), Mapping Subterrenean Biodiversity/Cartographie de la biodiversite souterraine, Pro-
ceedings of the International Workshop held, 18–20 March 2001. Laboratoire Souterrain du
CNRS, Moulis, Ariege, France. Karst Waters Institute, Special Publication 6, Charles Town,
West Virginia, pp. 19–21.
Gibert J. and Deharveng L. 2002. Subterranean ecosystems: a truncated functional biodiversity.
BioScience 52: 473–481.
Ginet R. 1971. Biogeographic de Niphargus et Caecosphaerom (Crustaces troglobies) dans les
departements francais du Jura et de l’Ain Origine; influence des glaciations. Proceedings of the
4th Swiss Congress of Speleology, Neuchatel, Switzerland, 26–27 September 1970, pp. 186–198.
Ginet R. and Juberthie C. 1987. Le peuplement animal des karsts de France. I. La faune aquatique.
Karstologia 10: 43–51.
Gottstein-Matocec S., Bakran-Petricioli T., Bedek J., Bukovec D., Buzjak S., Franicevic M., Jalzic
B., Kerovec M., Kletecki E., Kralj J., Kruzic P., Kucinic M., Kuhta M., Matocec N., Ozimec, R.,
Tonci R., Stamol V., Ternjej I. and Tvrtkovic N. 2002. An overview of the cave and interstitial
biota of Croatia. Nat. Croatica 11: 1–112.
Henry J.-P. 1976.Recherches sur les Asellidae hypoges de la lignee cavaticus (Crustacea Isopoda
Asellota). Ph.D. thesis, University of Dijon, France.
Holsinger J.R., Mort J.W. and Recklies A.D. 1983. The subterranean crustacean fauna of Castle
guard Cave, Columbia Icefields, Alberta, Canada, and its zoogeographic significance. Arctic
Antarctic Alpine Res. 15: 543–549.
Holsinger J.R. 1993. Biodiversity of subterranean amphipod crustaceans: global patterns and
zoogeographic implications. J. Nat. Hist. 27: 821–835.
Holsinger J.R. 2000. Ecological derivation, colonization and speciation. In: Wilkens H., Culver
D.C. and Humphreys W.F. (eds), Subterranean Ecosystems. Ecosystems of the World 30,
Elsevier, Amsterdam, pp. 399–415.
594

Humphreys W.F. 2000. Relict faunas and their derivation. In: Wilkens H. Culver D.C. and
Humphreys W.F. (eds), Subterranean Ecosystems. Ecosystems of the World 30, Elsevier,
Amsterdam, pp. 417–432.
Juberthie C. and Juberthie-Jupeau L. 1975. La reserve biologique du laboratoire souterrain du
CNRS a Sauve (Gard). Aiulales Speleol. 30: 539–551.
Juberthie C. and Decu V. 1994. Encyclopaedia Biospeleogica I, Societe de Biospeleologie. Moulis –
Bucarest.
Juberthie C. and Ginet R. 1994. France. In: Juberthie C. and Decu V. (eds), Encyclopoedia
Biospeleogica I, Societe de Biospeologie. Moulis, Bucarest, pp. 665–692.
Juberthie C. 1995. Underground Habitats and their Protection. Nature and Environment, 72.
Council of Europe, Strasbourg.
Juberthie C. and Decu V. 1998. Encyclopaedia Biospeleogica II, Societe de Biospeleologie. Moulis
– Bucarest.
Juberthie C. and Decu V. 2001. Encyclopaedia Biospeleogica III, Societe de Biospeleologie. Moulis
– Bucarest.
Ketmaier V., Messana G., Cobolli M., de Matthaeis E. and Argano R. 2000. Biochemical bioge-
ography and evolutionary relationships among the six known populations of Stenasellus rac-
ovitzai (Crustacea, Isopoda) from Tuscany, Corsica and Sardinia. Arch. Hydrobiol. 147:
297–309.
Langecker T.G. 2000. The effects of continuous darkness on cave ecology and cavernicolous
evolution. In: Wilkens H., Culver D.C. and Humphreys W.F. (eds), Subterranean Ecosystems.
Ecosystems of the World 30, Elsevier, Amsterdam, pp. 135–157.
Lescher-Moutoue F. 1967. Notes sur la biogeographic et les biotopes du genre Speocyclops dans la
region pyreneenne. Spelunca Memoires 5: 277–284.
Mace G.M. 2000. It’s time to work together and stop duplicating conservation efforts. Nature 405:
393.
Magniez G. 1976.Contribution a la connaissance de la biologie des Stenasellidae (Crustacea
Isopoda Asellota des eaux souterraines). PhD. thesis, University of Dijon, France.
Malard F., Gibert J. and Laurent R. 1997. L’aquifere de la source du Lez: un reservoir d’eau et de
biodiversite. Karstologia 30: 49–54.
Magniez G. 1997. Facteurs intrinseques et extrinseques de la distribution actuelle des Crustaces
Isopodes des eaux souterraines d’Europe. Proceedings of the 12th International Congress of
Speleology, La Chaux-de-Fonds, Switzerland, 10–17 August 1997, pp. 341–344
Malard F., Plenet S. and Gibert J. 1996. The use of invertebrates in groundwater monitoring: a
rising research field. Ground Water Monitor. Remediat. 16: 103–116.
Margules C.R., Pressey R.L. and Williams P.H. 2002. Representing biodiversity: data and pro-
cedures for identifying priority areas for conservation. J. BioSci. Suppl. 27: 309–326.
Marmonier P., Vervier P., Gibert J. and Dole-Olivier M.-J. 1993. Biodiversity in ground waters.
Trends Ecol. Evol. 8: 392–395.
Martınez-Ansemil E., Sambugar B. and Giani N. 1997. Groundwater Oligochaetes from Southern-
Europe. I. A new genus and three new species of Rhyacodrilinae (Tubificidae) with a rede-
scription of Tubifex pescei (Dumnicka) comb. n. Ann. Limnol. 33: 33–44.
Mosslacher F. and Notenboom J. 2000. Groundwater biomonitoring. In: Gerhardt A. (ed.), Bio-
monitoring of Polluted Water. Trans Tech Publications Ltd, Zurich, Switzerland, pp. 119–140.
Nelson B.W., Ferreira C.A.C., da Silva M.F. and Kawasaki M.L. 1990. Endemism centres, refugia
and botanical collection density in Brazilian Amazonia. Nature 345: 714–716.
Notenboom J., Plenet S. and Turquin M.-J. 1994. Groundwater contamination and its impact on
groundwater animals and ecosystems. In: Gibert J., Danielopol D.L. and Stanford J.A. (eds),
Groundwater Ecology. Academic Press, New York, pp. 477–504.
Oliver J.H.J. 1988. Crisis in biosystematics of Arthropods. Science 240: 967.
Possingham H., Ball I. and Andelman S. 2000. Mathematical methods for identifying represen-
tative reserve networks. In: Ferson S. and Burgman M. (eds), Quantitative Methods for Con-
servation Biology. Springer-Verlag, New York, pp. 291–305.
595

Racovitza E.G. 1907. Essai sur les problemes biospeologiques. Arch. Zool. Exp. Generale 6: 371–
488.
Reddy S. and Davalos L.M. 2003. Geographical sampling bias and its implications for conserva-
tion priorities in Africa. J. Biogeogr. 30: 1719–1727.
Reid J.W. 1992. Taxonomic problems: a serious impediment to groundwater ecological research in
North America. Proceedings of the First International Conference on Ground Water Ecology,
U.S. Environmental Protection Agency, Tampa, Florida, 26–29 April 1992, pp. 133–142
Rouch R. 1968. Contribution a la connaissance des Harpacticides hypoges (Crustaces -
Copepodes). Ann. Speleol. 23: 5–167.
Rouch R. 1986. Sur l’ecologie des eaux souterraines dans le karst. Stygologia 2: 352–398.
Rouch R. 1988. Sur la repartition spatiale des Crustaces dans le sous-ecoulement d’un ruisseau des
Pyrenees. Ann. Limnol. 24: 213–234.
Rouch R., Pitzalis A. and Descouens A. 1993. Effets de pompage a gros debit sur le peuplement des
Crustaces d’un aquifere karstique. Ann. Limnol. 29: 15–29.
Sket B. 1999a. High biodiversity in hypogean waters and its endangerment. The situation in
Slovenia, the Dinaric karsts. and Europe. Crustaceana 72: 767–780.
Sket B. 1999b. The nature of biodiversity in hypogean waters and how it is endangered. Biodiver.
Conserv. 8: 1319–1338.
Spangler P.J. and Decu V. 1998. Coleoptera Aquatica. In: Juberthie C. and Decu V. (eds),
Encyclopoedia Biospeleogica II. Societe de Biospeologie, Moulis, Bucarest, pp. 1031–1046.
Stewart R.R., Noyce T. and Possingham H.P. 2003. Opportunity cost of ad hoc marine reserve
design decisions: an example from South Australia. Mar. Ecol. Prog. Ser. 253: 25–38.
Stoch F. 1995. The ecological and historical determinants of Crustacean diversity in groundwaters,
or: why are there so many species? Memoires Biospeleol. 22: 139–160.
Stoch F. 2000. How many endemic species? Species richness assessment and conservation priorities
in Italy Belg. J. Entomol. 2: 125–133.
Stoch F. 2001. Mapping subterranean biodiversity : structure of the database, mapping software
(CKMAP), and a report of status for Italy. In: Culver D.C., Deharveng L., Gibert J. and
SasowskyI.D. (eds), Mapping Subterrenean Biodiversity/Cartographie de la biodiversite sout-
erraine. Proceedings of the International Workshop held, 18–20 March 2001, Laboratoire
Souterrain du CNRS, Moulis, Ariege, France, Karst Waters Institute, Special Publication 6,
Charles Town, West Virginia, pp. 29–35
Strayer D.L., May S.E., Nielsen P., Wollheim W. and Hausam S. 1995. An endemic groundwater
fauna in unglaciated eastern North America. Can. J. Zool. 73: 502–508.
IUCN 2004. 2004 IUCN Red List of Threatened Species. http://www.iucnredlist.org. Downloaded
on 10 November 2004.
Valdecasas A.G. and Camacho A.I. 2003. Conservation to the rescue of taxonomy. Biodivers.
Conserv. 12: 1113–1117.
596
Copyright © 2022 FDOKUMEN




















