
Papers and reports upon forestry, forest schools, forest ...

JOFO jofo184 Dispatch: 8-19-2008 CE: AFL
Journal MSP No. No. of pages: 16 PE: Sonia
jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
J. Field Ornithol. 79(3):229–244, 2008 DOI: 10.1111/j.1557-9263.2008.00184.x
Identifying forest-obligate birds in tropical moist cloudforest of Andean Ecuador
C. Dustin Becker,1,4 Thomas M. Loughin,2 and Tatiana Santander3
1Life Net, 6423 South Bascom Trail, Willcox, Arizona 85643, USA2Department of Statistics, Simon Fraser University, Surrey, British Columbia, Canada
3Fundacion Ornitologica del Ecuador (CECIA), Quito, Ecuador
Received 15 August 2007; accepted 25 February 2008
ABSTRACT. Large-scale transformation of forested landscapes is a major factor in loss of biological diversityin the American tropics. Investigators examining the responses of species to deforestation rarely control for variationin the amount of forest relative to other habitats at the landscape-level. Bellavista Reserve on the western slope of theAndes in Ecuador is located between similar-sized areas of pristine, protected forest, and deforested landscapes. Weused strip-transect counts and mist netting to evaluate habitat use by passerine birds in a habitat mosaic consistingof abandoned pastures, forest edges, forest fragments, and large blocks of interior tropical montane cloud forest(TMCF). During 3600 net hours, we had 1476 captures, including 346 recaptures. Of 78 species captured in mistnets, 30 had sufficient counts for PRR modeling (a statistical method for comparing counts). Twelve species (40%)had capture patterns indicative of an affinity for mature TMCF, and six species (20%) had significantly higher countsin degraded areas (forest edge, forest fragment, and regenerating pastures) than in interior TMCF. The remaining40% showed no significant bias in detection among habitats. Combined with strip-count data, our results suggestthat about 38% of the 119 species sampled at the Bellavista Reserve occur primarily in mature TMCF, avoidingedges and early second-growth forest. Populations of these species may be vulnerable to further loss, fragmentation,and degradation of TMCF and, as such, deserve additional study and a place on lists of species of conservationconcern.
SINOPSIS. Identificando aves obligadas a bosque en un bosque montano tropical nubosoen los Andes de Ecuador
Las transformaciones a larga escala del paisaje de bosques es un factor principal en la perdida de biodiversidadbiologica en los tropicos de america. Los investigadores que examinan la respuesta de especies a la deforestacion,raras veces controlan las variaciones en la cantidad de bosque, relativa a otros habitats a nivel del paisaje. La ReservaBellavista, que se encuentra en la pendiente oeste de los Andes ecuatorianos, se encuentra localizada entre areas (desimilar tamano), pristinas de bosques protegidos y areas deforestadas. Utilizamos transectos lineares y capturas conredes de niebla para evaluar el uso de habitat, por parte de paserinos, en un mosaico de habitats que consistio detierras de pastoreo abandonadas, bordes de bosques, fragmentos de bosques y grandes bloques de bosque montanotropical nuboso (BMTN). En 3600 horas de trabajo, capturamos 1476 aves, incluyendo 346 recapturas. De 78especies capturadas con redes de niebla, en 30 tuvimos el numero suficiente para aplicarle el metodo estadistico PRR.Doce especies (40%) tuvieron un patron de captura asociado al BMTN, y seis especies (20%) tuvieron un conteosignificativamente mayor en areas degradadas (bordes, fragmentos y areas de pastizales en estado de regeneracion)que en el BMTN. El restante 40% no mostro sesgo significativo de detectabilidad entre los diferentes tipos dehabitats. Al combinar las capturas con los transectos, nuestros resultados sugieren que un 38% de las 119 especiesmuestreadas ocurren particularmente en el BMTN maduro, evitando bordes y etapas tempranas de sucesion. Laspoblaciones de estas especies pudieran ser vulnerables a mayor perdida de bosque, a la fragmentacion o degradacionde los BMTN. Las mismas ameritan mas estudios y deben colocarse en la lista de aves cuya conservacion ameritepreocupacion.
Key words: biodiversity conservation, forest fragmentation, habitat choice, tropical montane cloud forest
Rapid clearing and fragmentation of tropicalforest threatens many bird species (Stotz et al.1996). Tropical montane cloud forests (TMCF)in the northern Andes have declined in areaby as much as 90% (Hamilton et al. 1993).
4Corresponding author. Email: [email protected]
With 32 of 62 Choco regional endemics listedas near-threatened, vulnerable, or endangered(Long 1993, BirdLife International 2003), de-forestation appears to be taking its toll at theregional level.
Little is known about habitat use by birds incomplex tropical mosaics and, specifically, the ef-fects of habitat alteration on birds of the TMCF(Wiens 1994, Luck and Daily 2002). Although
C©2008 The Author(s). Journal compilation C©2008 Association of Field Ornithologists
229

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
230 C. D. Becker et al. J. Field Ornithol.
fragmentation and disturbance are known tocause declines in bird species richness andpersistence in tropical forests (Canaday 1999,Wijesinghe and Brooke 2005, Shahabuddin andKumar 2006), effects vary among species. Forexample, in Neotropical montane forest, frugi-vores, and nectar feeders appear to be attractedto edges, whereas some insectivores avoid them(Restrepo and Gomez 1998).
We studied a heterogeneous TMCF landscapemosaic and compared the relative abundance ofspecies in interior forest to those in regeneratingpastures, forest edge, and a forest fragment.Because anthropogenic clearing of forests farexceeds natural landslides in both intensity andarea (Dodson and Gentry 1991), we predictedthat many midelevation Andean bird specieswould show a preference for intact, interiorTMCF and would avoid forest fragments andedges. However, because landslides in TMCFalso create natural openings, we predicted thatlandslide specialists would show a preference fordegraded TMCF habitats.
METHODS
Study area. Located in Ecuador’s Choco en-demic bird area (Stattersfield et al. 1998), Bellav-ista reserve is a 900-ha, privately owned naturereserve located 62 km west of Quito (1400–2600 m above sea level at 0◦ 0.67′S, 78◦ 41′W).Local vegetation is tropical premontane andmontane cloud forest (Doumenge et al. 1993)characterized by abundant tree ferns (FamilyCyatheaceae), epiphytic ferns (Pteridophytes),mosses, bromeliads (Bromeliaceae), and orchids.
Bellavista is a buffer-zone property located atthe interface of heavily transformed TMCF andlarge tracts of pristine forest associated with theMindo-Nambillo Protective Forest. Forest clear-ing and regeneration at Bellavista have created amosaic of habitats that include a regionally repre-sentative gradient of forest degradation. Habitattypes range from clearings for pastures to large(>500 ha) intact blocks of TMCF connectedto the Mindo-Nambillo Protective Forest. Inour study, two fallow pastures abandoned forabout 6 yr represented the most degraded TMCFhabitat type. A 100-ha forest fragment (a patchof mature TMCF separated from mature forestby clearings, roads, and second growth) and aforest edge were considered intermediate levelsof degradation in the TMCF habitat mosaic.
The forest edge site was along a third abandonedpasture, with transitional vegetation betweenthe forest and the grass-dominated clearing. Weconsidered mature TMCF in large forest blocksconnected to the Mindo-Nambillo ProtectiveForest to be the least degraded habitat type ofthe gradient.
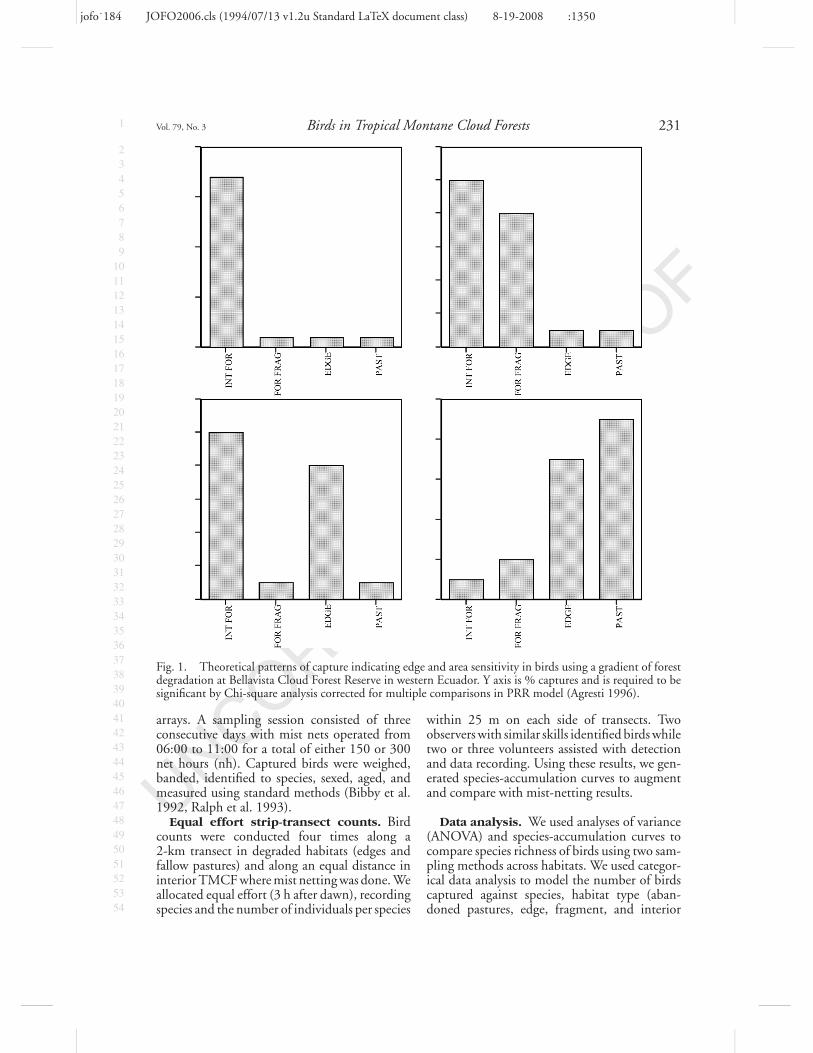
Hypotheses. We predicted that cloud forestspecies would vary in abundance as indicated bymist netting and observation (a proxy for habitatuse, habitat preference, and habitat avoidance)along the gradient. Specifically, we predicted that(1) counts of those species sensitive to area andedge effects in TMCF would be low in all butforest interior habitat, (2) counts of species sensi-tive to forest area, but not to edge effects (Parkeret al. 2005), would be low in a forest fragmentand higher counts in forest edge and interior,(3) counts of edge-sensitive species would berelatively high in fragments, but low along theedge of TMCF, and (4) counts of edge-lovingspecies would be higher in abandoned pasturesand forest edge and lower in the forest fragmentand the interior forest (Fig. 1).
Mist netting. From May to August 1998,1999, and 2000, we used mist nets to samplebirds in the understories of the four habitats,including pasture (N = 2), edge (N = 1),fragment (N = 1), and interior forest (N = 2).Due to availability, only one forest edge and oneforest fragment were sampled. Forest sites were>500 m from edge habitat (clearings for roads orpastures) and 2 km apart. Abandoned pastures(about 10 ha each) were at least 2 km apart andseparated by second-growth forest. The forestfragment was approximately 100 ha in size andsquare-shaped. On three sides of the fragment, a10-m-wide road, 300 m of scrubby edge, and ariver valley separated the fragment from second-growth forest. On the fourth side, the fragmentwas separated from forest by 200 m of scrubbyvegetation and bamboo created by a landslideand an abandoned pasture.
Either 10 or 20 mist nets (12 m × 2.8 m,36-mm mesh) were used in each habitat. Netswere positioned with random starting points onor adjacent to foot trails and were 10–50 m apart.In the fragment, nets were at least 200 m fromany edge. In pastures, nets were randomized ona grid and varied in distance from the “hard”edge of the forest, with some nets located asclose as 10 m from the edge. Elevation, terrain,orientation, and slope were similar for all net
jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 231
Fig. 1. Theoretical patterns of capture indicating edge and area sensitivity in birds using a gradient of forestdegradation at Bellavista Cloud Forest Reserve in western Ecuador. Y axis is % captures and is required to besignificant by Chi-square analysis corrected for multiple comparisons in PRR model (Agresti 1996).
arrays. A sampling session consisted of threeconsecutive days with mist nets operated from06:00 to 11:00 for a total of either 150 or 300net hours (nh). Captured birds were weighed,banded, identified to species, sexed, aged, andmeasured using standard methods (Bibby et al.1992, Ralph et al. 1993).
Equal effort strip-transect counts. Birdcounts were conducted four times along a2-km transect in degraded habitats (edges andfallow pastures) and along an equal distance ininterior TMCF where mist netting was done. Weallocated equal effort (3 h after dawn), recordingspecies and the number of individuals per species
within 25 m on each side of transects. Twoobservers with similar skills identified birds whiletwo or three volunteers assisted with detectionand data recording. Using these results, we gen-erated species-accumulation curves to augmentand compare with mist-netting results.
Data analysis. We used analyses of variance(ANOVA) and species-accumulation curves tocompare species richness of birds using two sam-pling methods across habitats. We used categor-ical data analysis to model the number of birdscaptured against species, habitat type (aban-doned pastures, edge, fragment, and interior
jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
232 C. D. Becker et al. J. Field Ornithol.
Table 1. Number of mist-netting sessions (150 nh)in each habitat during each year of the study.
Habitat type 1998 1999 2000
Forest 3 3 2Fragment 3 2 1Edge 1 2 1Pasture 2 2 2
forest), and their interaction. Capture rates (cap-tures per net hour [nh]) were analyzed using aPoisson rate regression (PRR) model (also calledPoisson Regression with Offsets) with a log link.PRR is a method of modeling counts whensampling efforts are not constant (Agresti 1996).The log of the number of netting sessions (150nh units) in a given habitat was used as the offset(Table 1).
Goodness of fit of the model was assessedusing the deviance/DF criterion. Effects of ex-planatory factors and interactions on capturerates were tested using Type 3 Likelihood Ratiotests. Pair-wise comparisons among levels ofsignificant factors were carried out using Waldchi-square tests for contrasts. Tests of simpleeffects of one factor within levels of anotherwere carried out for significant interactions inthe same manner as the pair-wise comparisons.
All fitting and testing of models were per-formed using PROC GENMOD (SAS Institute
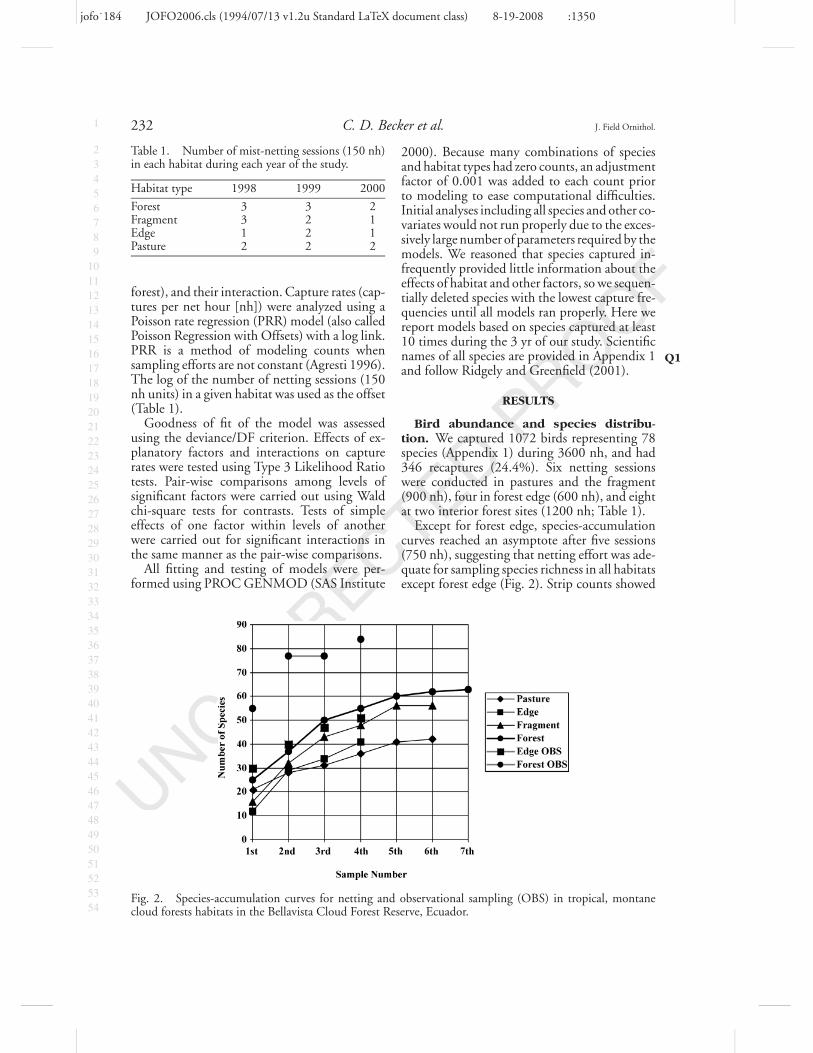
Fig. 2. Species-accumulation curves for netting and observational sampling (OBS) in tropical, montanecloud forests habitats in the Bellavista Cloud Forest Reserve, Ecuador.
2000). Because many combinations of speciesand habitat types had zero counts, an adjustmentfactor of 0.001 was added to each count priorto modeling to ease computational difficulties.Initial analyses including all species and other co-variates would not run properly due to the exces-sively large number of parameters required by themodels. We reasoned that species captured in-frequently provided little information about theeffects of habitat and other factors, so we sequen-tially deleted species with the lowest capture fre-quencies until all models ran properly. Here wereport models based on species captured at least10 times during the 3 yr of our study. Scientificnames of all species are provided in Appendix 1and follow Ridgely and Greenfield (2001).
Q1
RESULTS
Bird abundance and species distribu-tion. We captured 1072 birds representing 78species (Appendix 1) during 3600 nh, and had346 recaptures (24.4%). Six netting sessionswere conducted in pastures and the fragment(900 nh), four in forest edge (600 nh), and eightat two interior forest sites (1200 nh; Table 1).
Except for forest edge, species-accumulationcurves reached an asymptote after five sessions(750 nh), suggesting that netting effort was ade-quate for sampling species richness in all habitatsexcept forest edge (Fig. 2). Strip counts showed

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 233
a decline in species accumulation by the thirdcount. Accumulation curves for both nettingand strip counts indicated that species richnesswas higher in interior forest than in degradedhabitats. Abandoned pastures had the lowestrate of accumulation, reaching an asymptote at42 species, whereas forest fragment and interiorforest slowed to a similar rate at 60 species(Fig. 2).
Fewer birds and more, small understoryspecies, especially passerines, were captured inmist nets than were observed during strip counts.Large birds, such as Plate-billed Mountain-Toucans, Red-billed Parrots, Toucan Barbets,and Band-tailed Pigeons, made up nearly 20%of the observations during strip counts, butonly 0.2% of birds captured in mist nets(Appendix 1). Hummingbirds (Trochilidae),Flowerpiercers (Thraupidae), Wrens (Troglody-tidae), Warblers (Parulidae), and Brush-Finches(Emberizidae) were the most abundant fami-lies recorded by both methods (Appendix 1).Of 119 species sampled using nets and stripcounts, 60 were sampled by both methods.Four species (Dusky Bush-Tanager, Speckled
Table 2. The 10 most commonly netted bird species and their average captures per 150 nh (cap), and theirabundance rank (rank), in each habitat at Bellavista cloud forest reserve, Ecuador. Interior forest is used as areference. Blanks indicate no captures in that habitat.
Int. forest Fragment Edge Pasture
Species Cap Rank Cap Rank Cap Rank Cap Rank
Speckled Hummingbird 5.5 1 3.3 4 3.3 3 3.2 4Tawny-bellied Hermit 5.3 2 3.0 5.5 2.3 7 0.3 26Gray-breasted Wood-Wren 5.0 3 1.2 14.5 3.0 4 0.3 26Russet-crowned Warbler 4.8 4 4.7 3 2.0 10.5 1.3 7Dusky Bush-Tanager 4.6 5 5.8 2 2.0 10.5 6.7 1Chestnut-capped Brushfinch 4.3 6 0.8 18 0.7 18 0 –Gorgetted Sunangel 4.0 7 6.3 1 – – 1.2 8Yellow-bellied Chat-Tyrant 3.8 8 – – – – – –Sepia-brown Wren 3.0 9 1.5 11 4.5 1 – –Collared Inca 2.3 10 2.2 8 2.8 5 3.8 2Slaty Finch 2.4 11 3.0 5.5 – – – –Masked Flowerpiercer 0.8 23 2.3 7 – – 3.7 3Mionectes sp. – – 1.7 9 – – – –White-sided Flowerpiercer – – 1.7 10 – – 2.5 5Rufous-headed Pygmy-Tyrant – – – – 3.5 2 – –Black-eared Hemispingus – – – – 2.3 6 – –Three-striped Warbler – – – – 2.3 8 – –Beryl-spangled Tanager – – – – 2.0 9 – –Azara’s Spinetail – – – – – – 2.3 6Spectacled Whitestart – – – – – – 1.7 9Black-crested Warbler – – – – – – 1.0 10
Hummingbird, Russet-crowned Warbler, andGray-breasted Wood-Wren) were in the top 10most common species independent of samplingmethod (Tables 2 and 3).
PRR. For species captured at least 10 times(N = 30), we examined factors influencingcapture rates using a PRR model. The firstmodel included (1) level of forest degradation(low for interior forest, medium for edge andfragment, and high for pastures), (2) six net sites(two pastures, two interior forests, one edge,and one fragment) nested within degradationlevel, (3) 3 yr (1998, 1999, and 2000), and(4) 30 species. Results indicated that interac-tions between species and degradation level andbetween species and net sites were significant.For degradation levels analyzed separately, forestand pasture did not differ in species profiles,but edge and fragment did (� 2
29 = 55.4, P <0.002). Thus, we combined the two forestsand the two pastures, but not edge and frag-ment, resulting in a four-level factor represent-ing habitat types along a degradation gradient.The model was repeated, controlling for nettingeffort, and the species by habitat interaction

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
234 C. D. Becker et al. J. Field Ornithol.
Table 3. Mean number of observations of most abundant species observed during five strip counts conductedin 1999 and 2000 in forested (reference) versus edge and pasture habitats at Bellavista.
Forest Edge
Species Mean obs. Rank Mean obs. Rank
Dusky-bellied Bush-tanager 18.6 1 25.8 1Gray-breasted Wood-Wren 14.2 2 5.4 9Masked Flowerpiercer 9.8 3 12.4 2Russet-crowned Warbler 9.2 4 5.2 8Spillman’s Tapaculo 9.0 5 10.4 4Beryl-spangled Tanager 8.9 6 4.6 11Toucan Barbet 5.0 7 1.8 21Brown-capped Vireo 4.6 8.5 0.8 34Red-billed Parrot 4.6 8.5 2.4 17Band-tailed Pigeon 4.4 10 5.2 10Plate-billed Mountain-Toucan 4.0 12.5 2.6 15Slate-throated Whitestart 4.0 12.5 1.8 23Blue-winged Mountain-Tanager 4.0 12.5 7.2 5Speckled Hummingbird 4.0 12.5 6.2 6.5Buff-tailed Coronet 3.0 19 6.2 6.5Azara’s Spinetail 1.0 47 11.2 3Rufous-collared Sparrow 0 none 2.8 12
was again significant (� 287 = 414, P < 0.0001;
Table 4).We assessed habitat use probabilities for each
species by matching their capture profiles withpredicted distributions (Fig. 1). Chi-square anal-yses were used to determine where species werecaptured more or less often, and these resultswere used to plot capture profiles (Figs. 3–6).
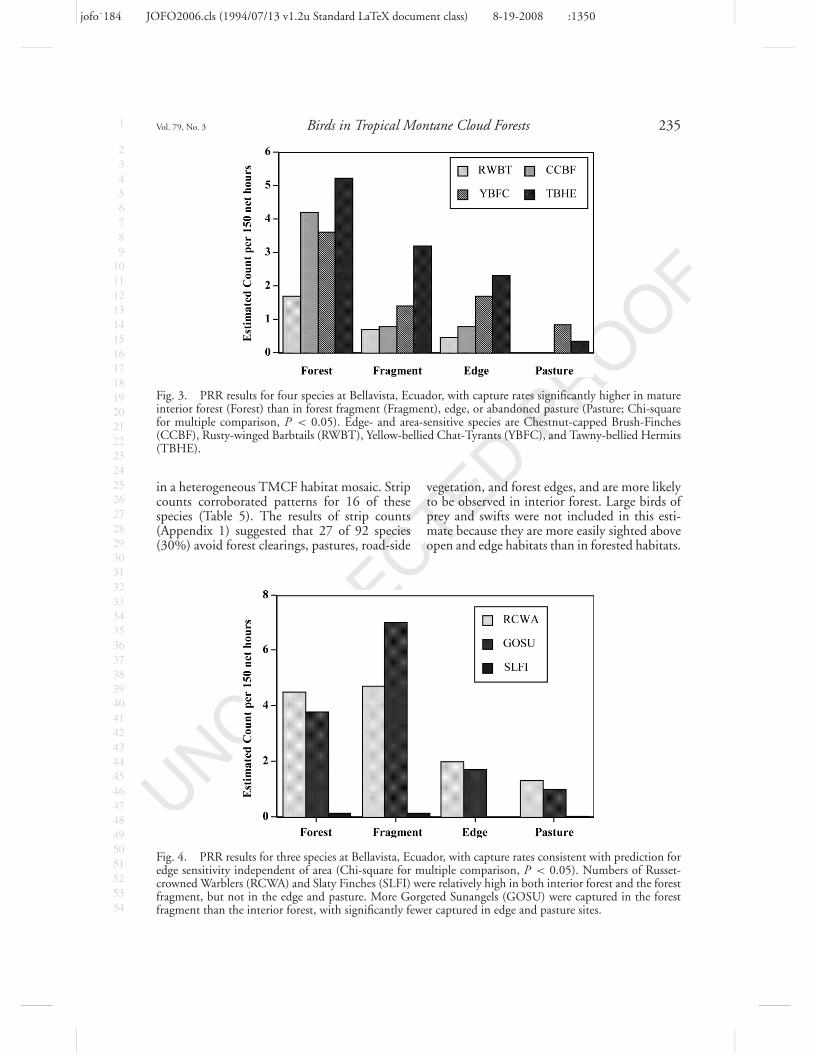
Sixteen of the 30 PRR species had signifi-cantly lower capture probabilities in abandonedpastures and edge than in forested habitats(Table 5). Four had capture probabilities match-ing the profile for edge and area sensitivity(Fig. 3). Two species exhibited edge sensitivityindependent of area sensitivity (Table 5, Fig. 4)
Table 4. Analysis of variance for PRR indicatingsignificance of variables explaining variation in countsof netted birds. Of particular interest to our centralhypothesis is species × habitat (degradation level).Q2
P ≥Source df Chi-square chi-square
Habitat 3 0.8 0.84Year 2 3.6 0.16Habitat × year 4 24.8 0.0001Species 29 415.1 0.0001Species × habitat 87 216.5 0.0001Species × year 58 160.8 0.0001
and five species exhibited forest area sensitivity(Table 5, Fig. 5). Eight species had trend-level(chi-square; 0.05 < P < 0.20) capture profilesindicative of a higher probability of finding themin forested habitat than in edge habitat (Table 5).For 14 species, strip-count data corroborated thePRR results and indicated a strong affinity forTMCF forest (Table 5).
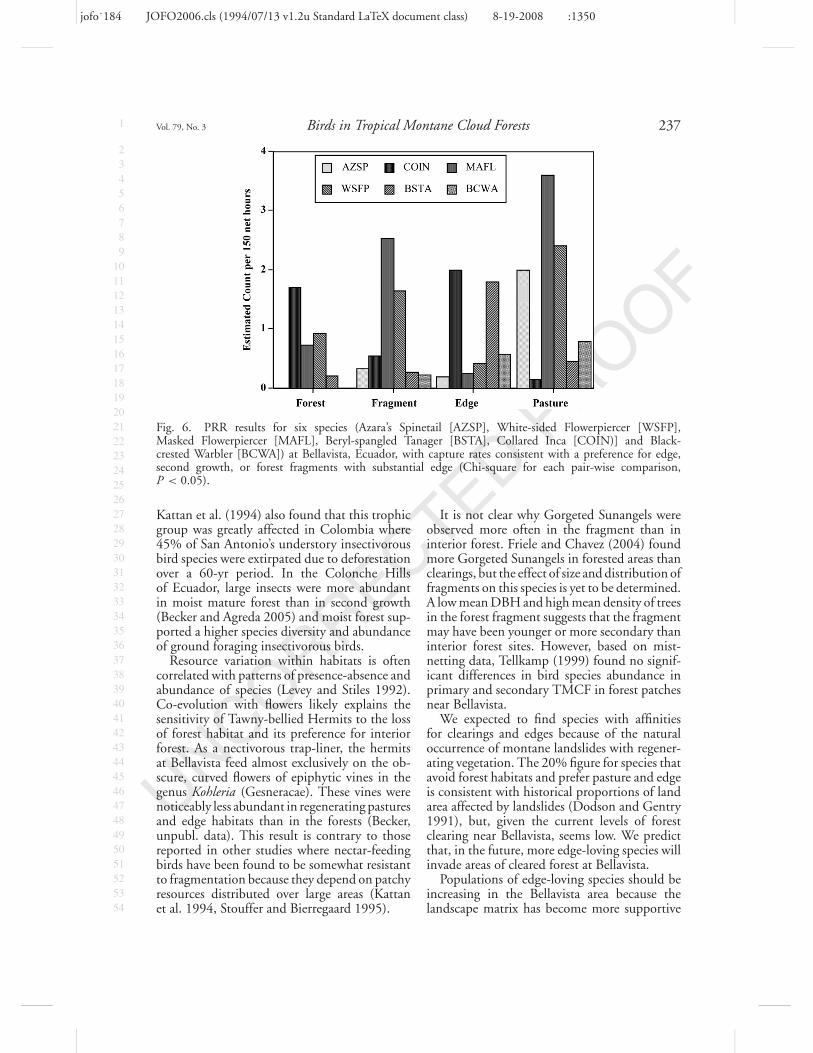
Two species, Beryl-spangled Tanager (Fig. 5)and Buff-tailed Coronet (Table 5), were equiv-ocal. Based on mist-netting results, Beryl-spangled Tanagers were more likely to be cap-tured in pasture and edge habitat, but stripcounts revealed higher numbers in forest habi-tats (Appendix 1). Buff-tailed Coronets werecaptured more often in forest habitats (Fig. 5),but were observed more frequently along edges(Appendix 1, Table 5).
Six species (20%) had capture profiles consis-tent with a preference for regenerating pastureand edge, or they failed to match any predictedpattern indicative of sensitivity to loss of forest(Table 5, Fig. 6). Azara’s Spinetails were nettedmost often in pastures, and were the third mostcommon species observed on transects in edgehabitat (Table 3). Dusky Bush-Tanagers hadsimilar numbers of captures in all habitats exceptedge (Tables 2 and 3).
In summary, based on PRR, 22 of 30 birdspecies (73%) had nonrandom capture profiles
jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 235
Fig. 3. PRR results for four species at Bellavista, Ecuador, with capture rates significantly higher in matureinterior forest (Forest) than in forest fragment (Fragment), edge, or abandoned pasture (Pasture; Chi-squarefor multiple comparison, P < 0.05). Edge- and area-sensitive species are Chestnut-capped Brush-Finches(CCBF), Rusty-winged Barbtails (RWBT), Yellow-bellied Chat-Tyrants (YBFC), and Tawny-bellied Hermits(TBHE).
in a heterogeneous TMCF habitat mosaic. Stripcounts corroborated patterns for 16 of thesespecies (Table 5). The results of strip counts(Appendix 1) suggested that 27 of 92 species(30%) avoid forest clearings, pastures, road-side
Fig. 4. PRR results for three species at Bellavista, Ecuador, with capture rates consistent with prediction foredge sensitivity independent of area (Chi-square for multiple comparison, P < 0.05). Numbers of Russet-crowned Warblers (RCWA) and Slaty Finches (SLFI) were relatively high in both interior forest and the forestfragment, but not in the edge and pasture. More Gorgeted Sunangels (GOSU) were captured in the forestfragment than the interior forest, with significantly fewer captured in edge and pasture sites.
vegetation, and forest edges, and are more likelyto be observed in interior forest. Large birds ofprey and swifts were not included in this esti-mate because they are more easily sighted aboveopen and edge habitats than in forested habitats.

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
236 C. D. Becker et al. J. Field Ornithol.
Fig. 5. PRR results for five species at Bellavista, Ecuador, with capture rates consistent with predictionfor sensitivity to area independent of edge (Chi-square for multiple comparison, P < 0.05). Numbers ofBuff-tailed Coronets (BTCO), Gray-breasted Wood-Wrens (GBWW), Slaty-backed Nightingale Thrushes(SBNT), Sepia-brown Wrens (SBWR), and Black-eared Hemispingus’s (BEHE) were higher in the interiorforest and forest edge, and significantly lower in the forest fragment.
Despite low counts, guans, forest-falcons, andparrots were included as forest-dependent basedon Ridgely and Greenfield (2001). Based on thetwo methods, on average of 38.5% of the speciessampled at Bellavista had a higher probabilityof being found in forest habitat than edges andregenerating pasture.
DISCUSSION
Because detection of a bird species in a partic-ular habitat may be confounded by the methodused (Remsen and Good 1996), defining habitatpreferences can be difficult. Also, the abundanceof individuals of species in particular habitatsdepends on levels of habitat availability, habitatuse, and animal movements (Kattan and Murcia2003). By using two methods to assess proba-bilities of bird species in the different habitats atBellavista, we feel confident about using termslike preference or affinity when both methodsprovided similar evidence.
Few investigators distinguish between edgeand area sensitivity, but any species sensitive toboth should be more vulnerable to extinctiondue to fragmentation and disturbance in forest
ecosystems than those sensitive to just edgeeffects or just area effects (Parker et al. 2005).Brooks et al. (1999) estimated that 41% of 124forest-dependent species in the Atlantic forestsof Brazil were area sensitive and vulnerable toextinction due to deforestation, a percentagesimilar to our estimates.
Edge- and area-sensitive Chestnut-cappedBrush-Finches (listed as uncommon by Ridgelyet al. 1998), Rusty-winged Barbtails (rare),Yellow-bellied Chat-Tyrants (uncommon), andTawny-bellied Hermits (uncommon) shouldbe considered species of conservation concern.With the exception of the brush-finch, thesespecies have been recognized as indicators ofhigh quality tropical midmontane (1600–2600m) evergreen forest (Stotz et al. 1996), so thecondition of TMCF forest understories in rela-tionship to the distribution and abundance ofthese species deserves further study.
Again, three species with both edge- and area-sensitivity were understory insectivores. Whentrend-level PRR results corroborated by strip-transect counts are also considered, 12 of 14bird species with an affinity for interior for-est are small understory insectivores (Table 5).
jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 237
Fig. 6. PRR results for six species (Azara’s Spinetail [AZSP], White-sided Flowerpiercer [WSFP],Masked Flowerpiercer [MAFL], Beryl-spangled Tanager [BSTA], Collared Inca [COIN)] and Black-crested Warbler [BCWA]) at Bellavista, Ecuador, with capture rates consistent with a preference for edge,second growth, or forest fragments with substantial edge (Chi-square for each pair-wise comparison,P < 0.05).
Kattan et al. (1994) also found that this trophicgroup was greatly affected in Colombia where45% of San Antonio’s understory insectivorousbird species were extirpated due to deforestationover a 60-yr period. In the Colonche Hillsof Ecuador, large insects were more abundantin moist mature forest than in second growth(Becker and Agreda 2005) and moist forest sup-ported a higher species diversity and abundanceof ground foraging insectivorous birds.
Resource variation within habitats is oftencorrelated with patterns of presence-absence andabundance of species (Levey and Stiles 1992).Co-evolution with flowers likely explains thesensitivity of Tawny-bellied Hermits to the lossof forest habitat and its preference for interiorforest. As a nectivorous trap-liner, the hermitsat Bellavista feed almost exclusively on the ob-scure, curved flowers of epiphytic vines in thegenus Kohleria (Gesneracae). These vines werenoticeably less abundant in regenerating pasturesand edge habitats than in the forests (Becker,unpubl. data). This result is contrary to thosereported in other studies where nectar-feedingbirds have been found to be somewhat resistantto fragmentation because they depend on patchyresources distributed over large areas (Kattanet al. 1994, Stouffer and Bierregaard 1995).
It is not clear why Gorgeted Sunangels wereobserved more often in the fragment than ininterior forest. Friele and Chavez (2004) foundmore Gorgeted Sunangels in forested areas thanclearings, but the effect of size and distribution offragments on this species is yet to be determined.A low mean DBH and high mean density of treesin the forest fragment suggests that the fragmentmay have been younger or more secondary thaninterior forest sites. However, based on mist-netting data, Tellkamp (1999) found no signif-icant differences in bird species abundance inprimary and secondary TMCF in forest patchesnear Bellavista.
We expected to find species with affinitiesfor clearings and edges because of the naturaloccurrence of montane landslides with regener-ating vegetation. The 20% figure for species thatavoid forest habitats and prefer pasture and edgeis consistent with historical proportions of landarea affected by landslides (Dodson and Gentry1991), but, given the current levels of forestclearing near Bellavista, seems low. We predictthat, in the future, more edge-loving species willinvade areas of cleared forest at Bellavista.
Populations of edge-loving species should beincreasing in the Bellavista area because thelandscape matrix has become more supportive

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
238 C. D. Becker et al. J. Field Ornithol.
Table 5. Bird species sensitivities to deforestation based on mist net and strip counts in a habitat mosaic atBellavista Reserve, Ecuador.
Detection profile Species Evidencea
Edge and area sensitive — Tawny-bellied Hermit BothMore likely in interior forest
than any other habitat typeRusty-winged BarbtailYellow-bellied Chat-tyrantChestnut-capped Brush-finch
Edge sensitive — Gorgeted Sunangel
Russet-crowned Warbler
Both, but strip counts higher infragment
BothAvoids edge habitats, but shows
no higher probability ininterior forest than forestfragment
Area sensitive — Black-eared Hemisphingus BothShows a higher probability in
interior forest than fragmentSlaty-backed Nightingale ThrushSepia-brown WrenGray-breasted Woodwren
Forest loss sensitive — Montane Woodcreeper Trend-level chi-square (0.05 < P <0.20) for capture profilescorroborated by strip-countresults
Low probability in pasture andedge compared with forest
Andean SolitaireStreak-necked FlycatcherFlavescent FlycatcherSlaty Finch Capture profile (P < 0.05); absent
from strip-count dataAffinity for edges — Azara’s Spinetail Both
Uses open areas and pasture;avoids forested habitatindependent of fragment size
Black-crested WarblerCollared Inca Capture profile significantly biased
toward pasture and edge;strip-count neutral
Green-and-Black FruiteaterThree-striped WarblerWhite-sided Flowerpiercer Strip counts support affinity for
edge, insufficient netting resultsMasked FlowerpiercerEquivocal habitat use Buff-tailed Coronet Netted more in forest than edge
(P < 0.05), strip counts oppositeBeryl-spangled Tanager Netted in edge and pasture more
than forest (P < 0.05); stripcounts opposite
aBoth refers to evidence based on both a significant capture profile and strip-count results.
of generalist species (Dunning et al. 1992,Andren 1994). Still, populations of forest-obligate species should also be sustainable atBellavista because extensive stands of matureTMCF are adjacent to the reserve in the 19,000-ha Mindo-Nambillo Protective Forest.
Conducting transect counts, along with mistnetting, helped clarify habitat preferences, biasesin methods, and added 41 species to our totalsample of 119 species. At least 190 species havebeen recorded at Bellavista (Krabbe and Becker,unpubl. data). Thus, in combination, the twomethods detected 60% of the bird species knownfor Bellavista, suggesting that more extensivesampling could improve the study that manyspecies are seasonally absent, or that some speciesare so rare that only long-term repetitive sam-pling would detect them. One bias that became
obvious involved captures of canopy-feedingbirds. For example, Beryl-spangled Tanagers andMasked Flowerpiercers fed on net-level fruitingand flowering shrubs in the pasture and edge,but foraged above the nets in mature forest.This likely explains the opposite results obtainedusing the two methods to detect birds in thosehabitats. Numbers of canopy frugivores andnectar feeders were probably underestimated ininterior forest by both methods because seeingand netting birds in high mature forest is moredifficult than detecting them in the shorter veg-etation associated with edges and regeneratingpasture.
Of the species that preferred interior forest atBellavista, eight are considered indicators of highquality tropical midmontane evergreen forestby Stotz et al. (1996), including three species

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 239
we found to be edge and area sensitive. Ourfindings reinforce the validity of such indicatorlists, and identified other species that should bemonitored. We suggest that Gorgeted Sunan-gels, Andean Solitaires, Chestnut-capped Brush-Finches, Streak-necked Flycatchers, and Mon-tane Woodcreepers be added to indicator lists formidmontane evergreen forest (sensu Stotz et al.1996) and also be considered for addition to listsof vulnerable species.
To conserve biodiversity, we must be ableto predict individual and population-level re-sponses of different species to changes in landuse. Studying habitat use in landscape mosaics isone way to assess habitat preferences of speciesand help predict that species will decline inresponse to losses of particular habitat types.
ACKNOWLEDGMENTS
Research by CDB was supported by Life Net(www.lifenetnature.org), Earthwatch Institute, andKansas State University. R. Parsons, owner of Bellavista,helped with logistics and accommodations. CECIAprovided institutional affiliation in Ecuador and theMinistry of the Environment provided research permits.We thank A. Agreda, F. Cupueran, M. Gardiner, O.Rodriguez, and P. Torres for assistance with data collectionand help with leading teams of Earthwatch volunteers.We thank G. Ritchison, D. Stotz, T. Povilitis, and twoanonymous reviewers for improving our manuscript. Weespecially thank the many volunteers who contributedboth labor and financial assistance to our project.
LITERATURE CITED
AGRESTI, A. 1996. An introduction to categorical dataanalysis. Wiley, New York, NY.
ANDREN, H. 1994. Effects of habitat fragmentation onbirds and mammals in landscapes with differentproportions of suitable habitat: a review. Oikos 71:355–366.
BECKER, C. D., AND A. AGREDA. 2005. Bird communitydifferences in mature and second growth garua forestin Machalilla National Park, Ecuador. NeotropicalOrnithology 16: 289–306.
BIBBY, C. J., N. D. BURGESS, AND D. A. HILL. 1992. Birdcensus techniques. Academic Press, San Diego, CA.
BIRDLIFE INTERNATIONAL. 2003. BirdLife’s online worldbird database: the site for bird conservation, ver-sion 2.0. BirdLife International, Cambridge, UK.Available at: <http://www.birdlife.org> (accessed6/11/2007).
BROOKS, T., J. TOBIAS, AND A. BALMFORD. 1999. Defor-estation and extinctions in the Atlantic forest. AnimalConservation 2: 211–222.
CANADAY, C. 1999. Loss of insectivorous birds along agradient of human impact in Amazonia. BiologicalConservation 77: 63–77.
DODSON, C. H., AND A. H. GENTRY. 1991. Biologicalextinction in western Ecuador. Annals of the MissouriBotanical Gardens 78: 273–295.
DOUMENGE, C., D. GILMOUR, M. R. PEREZ, AND J.BLOCKHUS. 1993. Tropical montane cloud forests:conservation status and management issues. In: Trop-ical montane cloud forests (L. S. Hamilton, J. O.Juvik, and F. N. Scatena, eds.), pp. 17–24. East-WestCenter. Honolulu, HI.
DUNNING, J. B., B. J. DANIELSON, AND H. R. PULLIAM.1992. Ecological processes that affect populations incomplex landscapes. Oikos 65: 169–175.
FRIELE, J. F., AND J. A. CHAVEZ. 2004. Interesting distribu-tional records and notes on the biology of bird speciesfrom a cloud forest reserve in north-west Ecuador.Bulletin of the British Ornithological Club 124: 6–16.
HAMILTON, L. S., J. O. JUVIK, AND F. N. SCATENA (eds.).1993. Tropical montane cloud forests: introductionand workshop synthesis. East-West Center, Hon-olulu, HI.
KATTAN, G. H. AND C. MURCIA. 2003. A review andsynthesis of conceptual frameworks for the studyof forest fragmentation. In: Ecological studies, vol.162: how landscapes change (G. A. Bradshaw, andP. A. Marquet, eds.), pp. 183–200. Springer-Verlag,Berlin, Germany.
———, H. ALVEREZ-LOPEZ, AND M. GIRALDO. 1994.Forest fragmentation and bird extinctions: San Anto-nio eighty years later. Conservation Biology 8: 138–146.
LEVEY, D. J., AND F. G. STILES. 1992. Evolutionary precur-sors of long-distance migration: resource availabilityand movement patterns in Neotropical landbirds.American Naturalist 140: 447–476.
LONG, A. J. 1993. Restricted-range and threatened birdspecies in tropical montane cloud forests. In: Tropicalmontane cloud forests (L. S. Hamilton, J. O. Juvik,and F. N. Scatena, eds.), pp. 47–65. East-West Cen-ter, Honolulu, HI.
LUCK, G.W., AND G. C. DAILY. 2002. Tropical countrysidebird assemblages: richness, composition, and foragingdiffer by landscape context. Ecological Applications13: 235–247.
PARKER, T. H., B. M. STANSBERRY, C. D. BECKER, ANDP. S. GIPSON. 2005. Edge and area effects on theoccurrence of migrant forest song birds. ConservationBiology 19: 1157–1167.
RALPH, J. C., G. R. GEUPEL, P. PYLE, T. E. MARTIN, ANDD. F. DESANTE. 1993. Handbook of field methodsfor monitoring landbirds. USDA General TechnicalReport PSW-GTR-144, Berkeley, CA.
REMSEN, J. V., AND D. A. GOOD. 1996. Misuse of datafrom mist-net captures to assess relative abundancein bird populations. Auk 113: 381–398.
RESTREPO, C., AND N. GOMEZ. 1998. Responses of un-derstory birds to anthropogenic edges in a Neotrop-ical montane forest. Ecological Applications 8:170–183.
RIDGELY, R. S., AND P. J. GREENFIELD. 2001. The birds ofEcuador, volumes I and II. Cornell University Press,Ithaca, NY.
———, P. J. GREENFIELD, AND M. GUERRERO. 1998.An annotated list of the birds of mainland Ecuador.

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
240 C. D. Becker et al. J. Field Ornithol.
Ornithological Foundation of Ecuador (CECIA),Quito, Ecuador.
SAS INSTITUTE. 2000. SAS/STAT user’s guide, Version8.1. SAS Institute, Inc., Cary, NC.
SHAHABUDDIN, G., AND R. KUMAR. 2006. Influence ofanthropogenic disturbance on bird communities ina tropical dry forest: role of vegetation structure.Animal Conservation 9: 404–413.
STATTERSFIELD, A. J., M. J. CROSBY, A. J. LONG, AND D.C. WEGE. 1998. Endemic bird areas of the world:priorities for conservation. Conservation Series No.7, Birdlife International, Cambridge, UK.
STOTZ, D. F., J. W. FITZPATRICK, T. A. PARKER, AND D.K MOSKOVITS. 1996. Neotropical birds: ecology andconservation. University of Chicago Press, Chicago,IL.
STOUFFER, P. C., AND R. O. BIERREGAARD. 1995. Effectsof forest fragmentation on understory hummingbirdsin Amazonian Brazil. Conservation Biology 9: 1085–1094.
TELLKAMP, M. P. 1999. The understory bird communityof a fragmented premontane forest in the westernAndes of Ecuador. M.S. thesis, University of Florida,Gainesville, FL.
WIENS, J. 1994. Habitat fragmentation: island vs. land-scape perspectives on bird conservation. Ibis 137:S97–104.
WIJESINGHE, M.R., AND M. L. BROOKE. 2005. Impact ofhabitat disturbance on the distribution of endemicspecies of small mammals and birds in a tropical rainforest in Sri Lanka. Journal of Tropical Ecology 21:661–668.

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 241
APP
EN
DIX
1.D
etec
tion
rate
s(%
ofca
ptur
esor
obse
rvat
ions
)of
bird
spec
ies
inm
atur
efo
rest
(int
erio
ran
dfr
agm
ent)
vers
usde
grad
edT
MC
Fha
bita
t(e
dge
and
seco
ndgr
owth
).Sp
ecie
sdes
igna
ted
wit
ha
sing
leas
teri
sk(N
=27
)are
thos
eex
hibi
ting
eith
ered
geor
area
sens
itiv
ity
and
expe
cted
tode
clin
ein
resp
onse
toco
ntin
ued
defo
rest
atio
n.Sp
ecie
sde
sign
ated
wit
ha
two
aste
risk
s(N
=16
)ar
eth
ose
likel
yto
bese
nsit
ive
tode
fore
stat
ion
base
don
obse
rvat
iona
lpat
tern
san
dm
istn
etti
ng,b
utw
ith
nost
atis
tica
lval
idat
ion
due
tolo
w-s
ampl
esi
zes.
Obs
erva
tion
Net
ting
Edg
eFo
rest
mea
nFo
rest
mea
nPa
stur
eE
dge
Frag
men
tFo
rest
Spec
ies
Com
mon
nam
eE
dge
%co
unt
%co
unt
%%
%%
Cat
hart
esau
raTu
rkey
Vul
ture
0.3
0.4
Cor
ygyp
satr
atus
Bla
ckV
ultu
re0.
61.
2C
hond
rohi
erax
unci
natu
sH
ook-
bille
dK
ite
0.1
0.2
0.1
0.2
Leuc
opte
rnis
prin
ceps
Bar
red
Haw
k0.
10.
2B
uteo
mag
niro
stris
Roa
dsid
eH
awk
0.1
0.2
B.l
euco
rrho
usW
hite
-rum
ped
Haw
k0.
10.
20.
10.
2O
roae
tusi
sidor
iB
lack
-and
-che
stnu
tEag
le0.
40.
60.
30.
6M
icra
stur
rufic
ollis
Bar
red
Fore
st-F
alco
n∗0.
10.
2Pe
nelo
pem
onta
gnii
And
ean
Gua
n∗0.
20.
4C
ham
aepe
tesg
oudo
tiiSi
ckle
-win
ged
Gua
n∗0.
10.
20.
81.
6O
dono
toph
orus
mel
anot
usD
ark-
back
edW
ood-
Qua
il∗0.
51.
0Pa
tagi
oenu
sfas
ciat
aB
and-
taile
dPi
geon
2.3
3.4
2.2
4.4
P.pl
umbe
aPl
umbe
ous
Pige
on1.
72.
60.
61.
2G
eotr
ygon
fren
ata
Whi
te-t
hroa
ted
Qua
il-D
ove∗
0.3
0.4
0.6
1.2
0.2
Pion
usso
rdid
usR
ed-b
illed
Parr
ot∗
1.6
2.4
2.3
4.6
Gla
ucid
ium
sp.
Pygm
yO
wlS
peci
es0.
2St
rept
opro
cne
zona
ris
Whi
te-c
olla
red
Swift
7.4
11.2
5.5
11.2
S.ru
tilus
Che
stnu
t-co
llare
dSw
ift0.
10.
2C
haet
ura
cine
reiv
entr
isG
ray-
rum
ped
Swift
6.9
10.4
5.1
10.4
Phae
thor
niss
yrm
atop
horu
sT
awn
y-b
elli
edH
erm
it0.
10.
21.
12.
20.
94.
75.
57.
4D
oryf
era
ludo
vica
eG
reen
-fro
nted
Lanc
ebill
∗0.
10.
20.
50.
30.
7C
olib
rith
alas
sinus
Gre
enV
iole
tear
0.8
1.2
.71.
40.
40.
90.
4A
delo
myi
am
elan
ogen
ysSp
eckl
edH
umm
ingb
ird
3.6
5.4
2.0
4.0
8.2
6.7
6.1
7.7
Am
azili
atz
acat
lR
ufou
s-ta
iled
Hum
min
gbir
d0.
2H
elio
doxa
rubi
noid
esFa
wn-
brea
sted
Bri
llian
t0.
4C
oelig
ena
wils
oni
Bro
wn
Inca
1.7
0.3
0.4
C.t
orqu
ata
Col
lare
dIn
ca1.
21.
8.9
1.8
15.
74.
03.
9B
oisso
nnea
uafla
vesc
ens
Buf
f-ta
iled
Cor
onet
3.9
5.8
1.5
3.0
1.7
2.6
0.9
2.6
Con
tinue
d

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
242 C. D. Becker et al. J. Field Ornithol.A
PPE
ND
IX1.
Con
tinu
ed.
Obs
erva
tion
Net
ting
Edg
eFo
rest
mea
nFo
rest
mea
nPa
stur
eE
dge
Frag
men
tFo
rest
Spec
ies
Com
mon
nam
eE
dge
%co
unt
%co
unt
%%
%%
Hel
iang
elus
strop
hian
usG
org
eted
Sun
ange
l0.
50.
81.
32.
63.
03.
611
.75.
6O
crea
tusu
nder
woo
dii
Boo
ted
Rac
ket-
tail
0.1
0.2
0.4
0.2
Agl
aioc
ercu
skin
giLo
ng-t
aile
dSy
lph
0.2
Philo
dice
mitc
helli
iPu
rple
-thr
oate
dW
oods
tar
0.2
Lafr
esna
yala
fres
naya
Mou
ntai
nV
elve
tbre
ast
0.9
0.6
0.2
Hyl
opha
edia
luge
nsH
oary
Puffl
eg0.
40.
50.
90.
4Ph
arom
achr
usan
tisia
nus
Cre
sted
Que
tzal
0.1
0.2
0.2
0.4
P.au
rice
psG
olde
n-he
aded
Que
tzal
0.1
0.2
0.2
0.4
0.3
Trog
onpe
rson
atus
Mas
ked
Trog
on∗
1.1
2.2
0.4
0.3
0.2
Sem
norn
isra
mph
astin
usTo
ucan
Bar
bet∗
1.2
1.8
2.5
5.0
0.2
And
igen
ala
min
iros
tris
Plat
e-bi
lled
Mou
ntai
n-To
ucan
∗1.
72.
62.
04.
00.
30.
2Pi
culu
sriv
olii
Cri
mso
n-m
antle
dW
oodp
ecke
r∗0.
91.
8Ve
nilio
rnis
fum
igat
usSm
oky-
brow
nW
oodp
ecke
r0.
2C
ampe
philu
spol
lens
Pow
erfu
lWoo
dpec
ker∗
0.1
0.2
Syna
llaxi
saza
rae
Aza
ra’s
Spin
etai
l4.
87.
20.
51.
06.
10.
50.
6S.
unir
ufa
Ruf
ous
Spin
etai
l∗0.
10.
20.
91.
80.
51.
50.
5M
arga
rorn
issq
uam
iger
Pear
led
Tree
runn
er∗
0.4
0.8
0.6
0.4
Prem
nopl
exbr
unne
scen
sSp
otte
dB
arbt
ail
3.1
1.5
2.5
Prem
norn
isgu
ttulig
era
Ru
sty
-win
ged
Bar
bta
il1.
01.
52.
8Ps
eudo
cola
ptes
boiss
onne
autii
Stre
aked
Tuft
edch
eek∗
0.7
1.4
0.3
0.2
Thr
ipad
ecte
shol
ostic
tus
Stri
ped
Tree
hunt
er0.
10.
20.
40.
50.
2T.
virg
atic
eps
Stre
ak-c
appe
dTr
eehu
nter
0.1
0.2
T.fla
mul
atus
Flam
ulat
edTr
eehu
nter
∗0.
10.
20.
7Sc
leru
rusm
exic
anus
Taw
ny-t
hroa
ted
Leaf
toss
er0.
4H
yloc
tiste
ssub
ulat
usSt
ripe
dFo
liage
-gle
aner
0.5
Synd
acty
lasu
bala
ris
Line
ated
Folia
ge-g
lean
er∗
0.3
0.6
0.3
Den
droc
incl
aty
rann
ina
Tyra
nnin
eW
oodc
reep
er∗
0.1
0.2
0.6
Xip
hoco
lapt
espr
omer
opir
hync
hus
Stro
ng-b
illed
Woo
dcre
eper
∗0.
10.
20.
81.
60.
30.
4Le
pido
cola
ptes
lacr
ymig
erM
on
tan
eW
oo
dcr
eep
er1.
83.
60.
91.
80.
7C
ampy
lorh
amph
uspu
sillu
sB
row
n-bi
lled
Scyt
hebi
ll0.
10.
2D
rym
ophi
laca
udat
aLo
ng-t
aile
dA
ntbi
rd0.
71.
00.
10.
2G
ralla
ria
rufic
apill
aC
hest
nut-
crow
ned
Ant
pitt
a∗0.
50.
80.
81.
60.
3C
ontin
ued

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
Vol. 79, No. 3 Birds in Tropical Montane Cloud Forests 243
APP
EN
DIX
1.C
onti
nued
.
Obs
erva
tion
Net
ting
Edg
eFo
rest
mea
nFo
rest
mea
nPa
stur
eE
dge
Frag
men
tFo
rest
Spec
ies
Com
mon
nam
eE
dge
%co
unt
%co
unt
%%
%%
Scyt
alop
ussp
illm
ani
Spill
man
’sTa
pacu
lo4.
16.
24.
49.
01.
00.
31.
2S.
vici
nior
Nar
ino
Tapa
culo
1.0
0.3
0.7
Acr
opte
rnis
orth
onyx
Oce
llate
dTa
pacu
lo0.
10.
2Ph
yllo
myi
asni
groc
apill
usB
lack
-cap
ped
Tyra
nnul
et0.
10.
2P.
cine
reic
eps
Ash
y-he
aded
Tyra
nnul
et0.
10.
2M
ecoc
ercu
lusp
oeci
loce
rcus
Whi
te-t
aile
dTy
rann
ulet
1.7
3.4
0.4
Mio
nect
esstr
iatic
ollis
Stre
ak-n
eck
edF
lyca
tch
er0.
61.
21.
31.
03.
11.
4Ps
eudo
tric
cusr
ufice
psR
ufo
us-
hea
ded
Py
gmy
-Ty
ran
t0.
40.
61.
32.
67.
30.
61.
1Ps
eudo
tric
cusp
elze
lni
Bro
nze-
oliv
ePy
gmy-
Tyra
nt0.
4Py
rrho
myi
asci
nnam
omea
Cin
nam
onFl
ycat
cher
0.3
0.4
0.2
0.4
Myi
opho
busfl
avic
ans
Fla
vesc
ent
Fly
catc
her
0.3
0.6
0.9
10.
91.
2C
onto
pusf
umig
atus
Smok
e-co
lore
dPe
wee
∗0.
30.
6O
chth
oeca
cinn
amom
eive
ntri
sSl
aty-
back
edC
hat-
Tyra
nt∗
0.3
0.6
1.6
0.5
S.di
adem
aY
ello
w-b
elli
edC
hat
-Ty
ran
t0.
30.
40.
51.
02.
23.
62.
55.
3M
yiod
ynas
tesc
hrys
ocep
halu
sG
olde
n-cr
owne
dFl
ycat
cher
0.8
1.2
0.3
0.6
Pipr
eola
rief
feri
iG
reen
-and
-bla
ckFr
uite
ater
0.3
0.4
0.3
0.6
1.7
0.5
1.2
0.2
Cya
noly
catu
rcos
aTu
rquo
ise
Jay
1.3
2.0
1.5
3.0
0.5
0.3
C.p
ulch
raB
eaut
ifulJ
ay0.
20.
4V
ireo
leuc
ophr
ysB
row
n-ca
pped
Vir
eo∗
0.5
0.8
2.3
4.6
Mya
deste
sral
loid
esA
nd
ean
Soli
tair
e0.
40.
61.
12.
20.
90.
51.
51.
4C
atha
rusf
usca
ter
Slat
y-b
ack
edN
igh
tin
gale
-Th
rush
0.5
1.0
0.4
4.1
0.3
2.5
Turd
usfu
scat
erG
reat
Thr
ush
0.5
0.8
0.1
0.2
0.4
0.5
0.2
T.se
rran
usG
loss
y-bl
ack
Thr
ush
0.9
1.4
0.5
1.0
0.9
0.5
0.6
0.9
Cin
clus
leuc
ocep
halu
sW
hite
-cap
ped
Dip
per
0.1
0.2
Not
ioch
elid
oncy
anol
euca
Blu
e-an
d-w
hite
Swal
low
0.3
0.4
0.4
0.8
Cin
nyce
rthi
aol
ivas
cens
Sep
ia-b
row
nW
ren
0.3
0.4
0.6
1.2
1.7
9.3
2.8
4.2
Thr
yoth
orus
euop
hrys
Plai
n-ta
iled
Wre
n1.
52.
21.
22.
4C
ontin
ued

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
1
23456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
244 C. D. Becker et al. J. Field Ornithol.A
PPE
ND
IX1.
Con
tinu
ed.
Obs
erva
tion
Net
ting
Edg
eFo
rest
mea
nFo
rest
mea
nPa
stur
eE
dge
Frag
men
tFo
rest
Spec
ies
Com
mon
nam
eE
dge
%co
unt
%co
unt
%%
%%
T.so
lstiti
alis
Mou
ntai
nW
ren∗
0.1
0.2
0.2
Hen
icor
hina
leuc
ophr
ysG
ray
-bre
aste
dW
oo
d-W
ren
3.1
4.6
7.0
14.2
0.9
6.2
1.8
7.0
Myi
obor
usm
inia
tus
Slat
e-th
roat
edW
hite
star
t1.
21.
82.
04.
01.
30.
50.
30.
4M
.mel
anoc
epha
lus
Spec
tacl
edW
hite
star
t0.
71.
00.
20.
43.
00
Bas
ileut
erus
trist
riat
usT
hree
-str
iped
War
bler
0.1
0.2
0.1
0.2
4.7
1.2
1.8
B.n
igro
crist
atus
Bla
ck-c
rest
edW
arbl
er1.
11.
62.
61.
60.
60
B.c
oron
atus
Ru
sset
-cro
wn
edW
arb
ler
3.5
5.2
4.5
9.2
3.5
4.1
8.6
6.7
Euph
onia
xant
hoga
ster
Ora
nge-
belli
edE
upho
nia
0.4
0.6
0.5
1.0
0.5
0.5
E.la
niir
ostr
isT
hick
-bill
edE
upho
nia
0.3
0.6
Con
iros
trum
albi
fron
sC
appe
dC
oneb
ill0.
10.
2D
iglo
ssacy
anea
Mas
ked
Flow
erpi
erce
r8.
212
.44.
89.
89.
50.
54.
31.
1D
.alb
ilate
raW
hite
-sid
edFl
ower
pier
cer
0.7
1.0
0.3
0.6
6.5
1.0
3.1
1.6
Pipr
aeid
eam
elan
onot
aFa
wn-
brea
sted
Tana
ger
0.2
0.4
Tang
ara
arth
usG
olde
nTa
nage
r0.
81.
20.
91.
80.
4T.
nigr
ovir
idis
Ber
yl-s
pang
led
Tana
ger∗
2.0
3.0
4.0
8.0
1.3
4.1
0.6
0.4
T.va
ssori
iB
lue-
and-
blac
kTa
nage
r∗1.
32.
60.
40.
52.
1T.
rufic
ervi
xG
olde
n-na
ped
Tana
ger
0.4
0.6
0.1
0.2
Ani
sogn
athu
sign
iven
tris
Scar
let-
belli
edM
ount
ain-
Tana
ger
0.5
0.3
A.s
ompt
uosu
sB
lue-
win
ged
Mou
ntai
n-Ta
nage
r4.
16.
22.
04.
01.
70.
51.
2A
.not
abili
sB
lack
-chi
nned
Mou
ntai
n-Ta
nage
r0.
10.
2B
uthr
aupi
smon
tana
Hoo
ded
Mou
ntai
n-Ta
nage
r0.
20.
4T
hrau
pisc
yano
ceph
ala
Blu
e-ca
pped
Tana
ger
1.6
2.4
0.3
0.6
0.5
Chl
oros
ping
usse
mifu
scus
Dus
kyB
ush-
Tana
ger
17.1
25.8
9.2
18.6
17.3
4.1
10.7
6.5
Hem
ispin
gusa
trop
ileus
Bla
ck-c
appe
dH
emis
ping
us0.
20.
4H
emisp
ingu
smel
anot
isB
lack
-ea
red
Hem
isp
ingu
s0.
71.
40.
40.
50.
92.
6C
hlor
orni
srie
fferi
iG
rass
-gre
enTa
nage
r1.
11.
61.
32.
61.
30.
2C
atam
blyr
hync
husd
iade
ma
Plus
hcap
∗0.
10.
20.
50.
5H
aplo
spiz
aru
stica
Slat
yF
inch
0.9
5.5
3.3
Atla
pete
slat
inuc
hus
Ruf
ous-
nape
dB
rush
-Fin
ch0.
5B
uarr
emon
brun
nein
ucha
Ch
estn
ut-
cap
ped
Bru
sh-F
inch
0.5
1.0
1.6
1.5
6.0
Ore
othr
aupi
sarr
emon
ops
Tana
ger
Finc
h1.
30.
3Z
onot
rich
iaca
pens
isR
ufou
s-co
llare
dSp
arro
w1.
92.
82.
20.
6

jofo˙184 JOFO2006.cls (1994/07/13 v1.2u Standard LaTeX document class) 8-19-2008 :1350
Queries
Q1 Author: Please specify what do the bold common name signify in the appendix table.
Q2 Author: The symbol “∗” has been replaced by the multi symbol “×” in Table 4. Please confirm.
Copyright © 2022 FDOKUMEN




















