
Habitat preferences and fitness consequences for fauna ...
185
Habitat preferences and fitness consequences for fauna associated with novel marine environments Luke T Barrett orcid.org/0000-0002-2820-0421 Submitted in total fulfilment of the requirements of the degree of Doctor of Philosophy September 2018 School of BioSciences University of Melbourne
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Habitat preferences and fitness consequences for fauna ...

Habitat preferences and fitness consequences for
fauna associated with novel marine environments
Luke T Barrett
orcid.org/0000-0002-2820-0421
Submitted in total fulfilment of the requirements of the degree of Doctor of
Philosophy
September 2018
School of BioSciences
University of Melbourne

i
ABSTRACT
The rapidly expanding reach of anthropogenic environmental change means that animals must
now navigate landscapes comprised largely of modified and degraded habitats. Individuals that
correctly perceive habitat quality will be best placed to survive and reproduce in novel
environments, but where environmental change outpaces the evolution of behavioural
responses, mismatches can arise between cues and the underlying value of habitats. These
mismatches can lead individuals to select habitats that offer relatively poor fitness outcomes,
creating ecological traps. In environments where ecological traps are likely to occur, data on
habitat preferences and fitness consequences can fundamentally change predictions of
metapopulation models and increase our understanding of the role that novel habitats play in
population persistence, but such data are rarely collected. In this thesis, I first conduct a global
meta-analysis to assess the state of knowledge on habitat preference and fitness metrics in
animal populations, using wildlife populations associated with aquaculture as a case study. My
findings reveal that responses to aquaculture vary widely across taxa and farming systems,
ranging from large increases in abundance to near complete displacement. However, the
influence of aquaculture on wildlife populations remains poorly understood, as researchers
rarely obtain appropriate measures of habitat preference, survival or reproductive success.
Accordingly, in subsequent chapters I apply the ecological trap framework to assess marine
habitats modified by aquaculture or invasive species. In the first application, I collect wild
Atlantic cod (a species known to be attracted to salmon farms) from areas of high and low
salmon farming intensity, and compare reproductive fitness via a captive spawning trial with
hatchery-rearing of offspring. I found limited negative effects of high farming intensity on quality
of offspring. In the second application, I show that the threat of predation by a native keystone
predator may limit the ability of an invasive seastar to exploit a food-rich habitat at shellfish
farms. In the third application, I show that an invasive canopy-forming marine macroalga
provides viable habitat for native fishes and may help to maintain fish biodiversity in areas where
urban impacts have driven a decline in native macroalgal canopy cover. Together, this thesis
demonstrates the utility of individual-level data on habitat preference and fitness outcomes—
via the application of the ecological trap conceptual framework—in assessing the impacts of
novel habitats on animals, and recommends greater use of this approach in future investigations
into the impacts of human-induced rapid environmental change in coastal marine ecosystems.

ii
DECLARATION
This is to certify that:
The thesis comprises only my original work towards the PhD except where indicated in the
Preface.
Due acknowledgement has been made in the text to all other material used.
The thesis is fewer than 100 000 words in length, exclusive of tables, maps, bibliographies and
appendices.
Luke Barrett
September 2018
Cover image: Mesocosm reef stocked with invasive wakame kelp (Undaria pinnatifida)

iii
PREFACE
I am the primary author and principle contributor on all chapters presented in this thesis. My
supervisors, Stephen E Swearer and Tim Dempster, are co-authors on all chapters.
Article publication status and author contributions
Chapter Two: Published by Reviews in Aquaculture on 14 Aug 2018. Co-authored by Tim
Dempster and Stephen E Swearer. LTB, TD and SES conceived and designed the experiment; LTB
conducted the experiment and collected data with assistance from technical staff and
volunteers; LTB analysed the data and wrote the manuscript; TD and SES provided editorial
comments.
Contributions: LTB 80 %, TD 10 %, SES 10 %
Chapter Three: Published by Aquaculture Environment Interactions on 16 Aug 2018. Co-
authored by Tim Dempster, Stephen E Swearer, Ørjan Karlsen, Torstein Harboe and Sonnich
Meier. LTB, TD, SES, ØK and TH conceived and designed the experiment; LTB conducted the
experiment and collected data with assistance from ØK, TH and SM, as well as technical staff at
the Norwegian Institute of Marine Research; LTB analysed the data and wrote the manuscript;
TD, SES and SM provided editorial comments.
Contributions: LTB 75 %, TD 5 %, SES 5 %, ØK 5 %, TH 5 %, SM 5 %
Chapter Four: Unpublished material not submitted for publication. Co-authored by Tim
Dempster and Stephen E Swearer. LTB, TD and SES conceived and designed the experiment; LTB
conducted the experiment and collected data with assistance from technical staff and
volunteers; LTB analysed the data and wrote the manuscript; TD and SES provided editorial
comments.
Contributions: LTB 80 %, TD 10 %, SES 10 %
Chapter Five: Unpublished material not submitted for publication. Co-authored by Stephen E
Swearer, Tim Dempster. LTB, SES and TD conceived and designed the experiment; LTB
conducted the experiment and collected data with assistance from technical staff and
volunteers; LTB analysed the data and wrote the manuscript; SES and TD provided editorial
comments.
Contributions: LTB 80 %, SES 10 %, TD 10 %

iv
This research was funded by grants from the Holsworth Wildlife Research Endowment (Chapters
Four and Five), the PADI Foundation (Chapter Five), the Victorian Environmental Assessment
Council (Chapter Five), the Sustainable Aquaculture Lab – Temperate and Tropical (all chapters),
the Research on the Ecology and Evolution (REEF) Lab (all chapters), and the Norwegian Seafood
Research Fund (Chapter Three). All animal research was conducted in accordance with the
animal ethics requirements of the University of Melbourne (Chapter Five: approval numbers
1413133 and 1413193) and Norwegian legislation on animal experimentation (Chapter Three:
approval number 8264). Permits were obtained from the Victorian state government for
collection and translocation of marine animals and algae for (Chapters Four and Five: RP919,
RP1185, NP280, NP282).

v
ACKNOWLEDGMENTS
I would firstly like to thank my supervisors, Steve Swearer and Tim Dempster, for their
unwavering support over the duration of my PhD. They have been everything I could have hoped
for in a pair of supervisors.
My friends and colleagues in the REEF and SALTT labs, past and present, provided helpful
discussions and comments on my thesis chapters, and were universally great company to have
a coffee or beer with, as were all my officemates in 131 and others around BioSciences 4. Special
mentions go to Simon, Emily, Ben, Fran, Qike, Valeriya, Tyler, Matt, James, Jack, Ollie and Fletch
for making me feel welcome in my first couple of years in Melbourne.
Many people combined forces with me to get fieldwork done on the cold and murky waters of
Port Phillip Bay. In alphabetical order: Dean Chamberlain, Seann Chia, Ben Cleveland, Emily
Fobert, Molly Fredle, Akiva Gebler, Kevin Jensen, Valeriya Komyakova, Nina Kriegisch, Kevin
Menzies, Rebecca Morris, Jack O’Connor, Simon Reeves, Juan Manuel Valero Rodriguez, Kyler
Tan, Chris Taylor, João Teixiera, Oliver Thomas and Rod Watson (Victorian Marine Science
Consortium). Apologies if I forgot anyone! Lance Wiffen provided access to his aquaculture
leases at Clifton Springs and Grassy Point. John Ahern and Tania Long averted a couple of
aquarium-related catastrophes in my absence, thanks and sorry!
Thanks to my Norwegian collaborators and surrogate supervisors during my time there: Torstein
Harboe, Ørjan Karlsen and Sonnich Meier. The work was made possible by numerous technical
staff, researchers, expert cod fishermen and all-round nice people. I’d especially like to thank
Margareth Møgster, Stig Ove Utskot, Theresa Aase, Michal Rejmer, Inger Semb Johansen, Nele
Gunkel-Sauer, Kristine Hovland Holm, Yvonne Rong, Terje van der Meeren, Tord Skår, Velimir
Nola and Glenn Sandtorv. I’d also like to thank the staff and students at Austevoll High School
for taking us to Brandasund and back with a boatload of live cod.
My family and friends back home in WA tolerated my long absence and my
incommunicativeness during the busy times, and largely stayed away from the question “when
will you be finished?”. Well played!
Finally, but most importantly, I’d like to thank my partner Marina, who provided constant love
and support, and made quite a few sacrifices to ensure that I had a clear run at this thesis. I hope
it’s been worth it!

vi
CONTENTS
List of Tables ....................................................................................................................... ix
List of Figures ....................................................................................................................... x
Chapter One | General introduction
General introduction ........................................................................................................... 1
References .......................................................................................................................... 9
Chapter Two | Impacts of marine and freshwater aquaculture on wildlife: a global meta-
analysis
Abstract ............................................................................................................................. 14
Introduction ...................................................................................................................... 14
Methods ............................................................................................................................ 17
Results ............................................................................................................................... 21
Discussion.......................................................................................................................... 28
References ........................................................................................................................ 36
Chapter Three | Limited evidence for differential reproductive fitness of wild Atlantic cod in
areas of high and low salmon farming density
Abstract ............................................................................................................................. 50
Introduction ...................................................................................................................... 50
Methods ............................................................................................................................ 53
Results ............................................................................................................................... 62
Discussion.......................................................................................................................... 66
References ........................................................................................................................ 71

vii
Chapter Four | Native predator prevents an invader from exploiting food-rich habitat
Abstract ............................................................................................................................. 79
Introduction ...................................................................................................................... 79
Methods ............................................................................................................................ 82
Results ............................................................................................................................... 86
Discussion.......................................................................................................................... 89
References ........................................................................................................................ 97
Chapter Five | An invasive habitat-former mitigates impacts of native habitat loss for endemic
reef fishes
Abstract ........................................................................................................................... 102
Introduction .................................................................................................................... 103
Methods .......................................................................................................................... 105
Results ............................................................................................................................. 113
Discussion........................................................................................................................ 119
References ...................................................................................................................... 123
Chapter Six | General discussion and conclusions
General discussion and conclusions ............................................................................... 128
References ...................................................................................................................... 137
Appendices
Appendix 2.1 ................................................................................................................... 142
Appendix 2.2 ................................................................................................................... 152
Appendix 2.3 ................................................................................................................... 153
Appendix 2.4 ................................................................................................................... 154
Appendix 2.5 ................................................................................................................... 155

viii
Appendix 3.1 ................................................................................................................... 156
Appendix 3.2 ................................................................................................................... 157
Appendix 3.3 ................................................................................................................... 158
Appendix 4.1 ................................................................................................................... 161
Appendix 4.2 ................................................................................................................... 164
Appendix 5.1 ................................................................................................................... 165
Appendix 5.2 ................................................................................................................... 166
Appendix 5.3 ................................................................................................................... 167
Appendix 5.4 ................................................................................................................... 168
Appendix 5.5 ................................................................................................................... 170
Appendix 5.6 ................................................................................................................... 171
Appendix 5.7 ................................................................................................................... 172

ix
LIST OF TABLES
Table 2.1. Mean effects of aquaculture sites on wildlife populations ........................................ 33
Table 3.1. Body size and condition metrics for low and high farm density groups .................... 62
Table 3.2. Egg and larval quality metrics for cod from low and high farm density areas ........... 67
Table 4.1. Population metrics for native (Coscinasterias muricata) and invasive (Asterias
amurensis) seastars inside and outside farms at Grassy Point and Clifton Springs Aquaculture
Fisheries Reserves ....................................................................................................................... 96
Table 5.1. Summary of fish recruitment to artificial reefs stocked with Undaria pinnatifida,
Ecklonia radiata, or left barren. ................................................................................................ 114
Table 5.2. Comparison of reef fish relative abundance estimated by diver catch per unit effort in
Undaria and Ecklonia habitats. ................................................................................................ 117
Table 5.3. Comparison of body condition and reproductive condition metrics in reef fishes
collected from Undaria and Ecklonia habitats. ......................................................................... 118

x
LIST OF FIGURES
Figure 1.1. Conceptual representation of habitat selection responses to habitats of varying
quality ............................................................................................................................................ 2
Figure 2.1. Distribution of research effort for studies that met the criteria for inclusion ......... 20
Figure 2.2. Distribution of research effort on interactions between aquaculture sites and wild
fauna among countries and territories ....................................................................................... 22
Figure 2.3. Summary statistics for log response ratios for each variable in our meta-analysis. 25
Figure 3.1. Map of collection sites relative to active salmon farms in south-western Norway . 54
Figure 3.2. Weight-at-length relationship for female and male cod .......................................... 61
Figure 3.3. Multidimensional scaling (MDS) plot showing dissimilarly (Euclidean distance) of
multivariate fatty acid profiles in Atlantic cod ovaries according to salmon farm density ........ 63
Figure 3.4. Daily egg production per tank during the captive spawning period ......................... 64
Figure 4.1. Population density of Asterias amurensis and Coscinasterias muricata inside and
outside the Clifton Springs and Grassy Point Fisheries Aquaculture Reserves ........................... 87
Figure 4.2. Armspan and gutted weight of Asterias amurensis inside and outside the Clifton
Springs Fisheries Aquaculture Reserve ....................................................................................... 88
Figure 4.3. Condition metrics for Asterias amurensis inside and outside the Clifton Springs
Fisheries Aquaculture Reserve .................................................................................................... 90
Figure 4.4. Density plot of Asterias amurensis size distribution inside and outside the farm
boundary at Clifton Springs Aquaculture Fisheries Reserve ....................................................... 91
Figure 4.5. Effect of prey, conspecifics and predators on habitat selection decisions by the
invasive seastar Asterias amurensis in laboratory trials ............................................................. 92
Figure 5.1. Archetypal examples of three rocky reef habitats in northern Port Phillip Bay ..... 103
Figure 5.2. Map of study locations in Port Phillip Bay, Australia .............................................. 106
Figure 5.3. Weight-at-length relationships for Heteroclinus perspicillatus, H. heptaeolus and
Neoodax balteatus .................................................................................................................... 113
Figure 5.4. Habitat choice trial results for Heteroclinus perspicillatus and Neoodax balteatus
................................................................................................................................................... 114

xi
Figure 5.5. Canonical analysis of principle coordinates (CAP) showing variation in fish
communities across underwater visual census (UVC) plots with and without Undaria pinnatifida
canopy ...................................................................................................................................... 120
Figure 5.6. Fish community metrics from underwater visual census plots .............................. 122
Figure 6.1. Conceptual diagram showing speculative positioning of responses of focal species to
novel habitats ............................................................................................................................ 130
Figure. 6.2. Conceptual representation of the ecological trap framework applied to an
assessment of the role of artificial reefs for fish production .................................................... 132
Figure. 6.3. Current distribution of Undaria pinnatifida within Port Phillip Bay in 2017 ......... 133

1
CHAPTER ONE: GENERAL INTRODUCTION
Anthropogenic impacts on the natural environment are occurring at an unprecedented rate
and spatial scale, with nearly every part of the globe affected by one or more human impacts
(Vitousek et al. 1997, Sanderson et al. 2002, Halpern et al. 2008, Vörösmarty et al. 2010). The
effects of these drivers are referred to collectively as human-induced rapid environmental
change (HIREC), and include those arising from habitat loss or habitat change, pollution,
species introductions, human harvesting, and climate change (Sih et al. 2011). As a result,
animals must now navigate a landscape comprised of modified or degraded environments that
have altered the ecological playing field from that of their evolutionary past (often to the
advantage of non-native over native species: Byers 2002, Crooks et al. 2011).
Impacts of HIREC on animal populations will depend on how individuals respond to novel cues
and environmental conditions. Animals will be best placed to survive and reproduce in
modified or degraded habitats if they exhibit adaptive behaviours, such as selecting the best
available habitats, choosing suitable food items and recognising and evading novel predators
(Sih et al. 2011, Sih 2013, Wong & Candolin 2014). However, HIREC frequently outpaces the
evolution of the indirect decision-making cues used by animals to assess current and future
conditions, which can result in individuals incorrectly evaluating risks and resources
(Schlaepfer et al. 2002).
Novel habitats, ecological traps and metapopulation ecology
These scenarios, in which ecological novelty leads individuals to choose behaviours or habitats
that lead to poor fitness outcomes, are termed evolutionary traps (Schlaepfer et al. 2002,
Robertson et al. 2013). Incorrect evaluations may arise through one of two sensory
mechanisms: a reliance on ‘outdated’ cues that no longer reliably predict fitness outcomes
because conditions have changed, or the introduction of novel cues that mimic or overpower
pre-existing cues but lead to poor fitness outcomes (Schlaepfer et al. 2002, Robertson et al.
2013, Wong & Candolin 2014).
A specific case of the evolutionary trap, the ecological trap, occurs when individuals either
prefer or fail to avoid low quality habitats when higher quality alternatives are available
(Robertson & Hutto 2006, Hale, Treml, et al. 2015). In doing so, they also fall into the corollary,

2
a perceptual trap, whereby individuals avoid or fail to prefer a relatively high quality habitat
(Fig. 1.1, Kokko and Sutherland 2001; Gilroy and Sutherland 2007; Patten and Kelly 2010).
Although the ecological trap concept concerns individual-level phenomena, traps directly drive
population-level processes, and can exacerbate population-level effects of environmental
change in impacted landscapes. Attractive population sinks caused by ecological traps can
impact metapopulation persistence disproportionately by drawing in animals that would
otherwise settle in source habitats (Battin 2004, Hale, Treml, et al. 2015). Similarly, perceptual
traps cause potential source habitats to be underutilised (Fig. 1.1), increasing effective habitat
loss in fragmented landscapes and potentially driving Allee effects for any individuals that do
choose to reside in the perceptual trap habitat (Kokko & Sutherland 2001, Gilroy & Sutherland
2007, Patten & Kelly 2010).
Habitat quality
High (potential source)
Low (potential sink)
Habitat selection
response
Preferred Correct decision Ecological trap
Avoided Perceptual trap Correct decision
Figure 1.1. Conceptual representation of the ecological trap framework in terms of possible
habitat selection responses to habitats of varying quality, sensu Patten and Kelly (2010).
In early source-sink metapopulation models, population growth was considered to be
approximately proportional to the spatial extent of source and sink habitats, with populations
in sink habitats maintained by passive dispersal from source habitats (Holt 1985). Later,
researchers began to incorporate active dispersal, allowing individuals to preferentially settle
in the best available habitats (population sources), and then spill over into more marginal
habitats in a density-dependent fashion (Pulliam 1988). Such models have significantly better
predictive ability than their predecessors and drove an important shift in thinking about
metapopulation ecology (Dias 1996). However, by assuming that individuals will tend to make
adaptive habitat selections, these models are vulnerable to making overly optimistic
predictions about population persistence whenever this assumption is violated, with
potentially dire consequences for the management of threatened animal populations (Battin
2004). The ecological trap concept provides a framework for assessing habitats through the
lens of individual habitat preferences and habitat quality (Fig. 1.1). This framework can, by

3
linking habitat-specific indices of habitat preference and fitness, distinguish between attractive
population sources (‘traditional’ population sources), attractive population sinks (ecological
traps), unattractive potential population sources (perceptual traps), and unattractive
population sinks (‘traditional’ population sinks) (Fig. 1.1, Patten and Kelly 2010; Hale et al.
2015; Hale and Swearer 2016). Incorporating such an assessment can improve the predictive
ability of source-sink models, and in some cases may fundamentally alter expectations of
population persistence in degraded or modified environments (Kokko & Sutherland 2001,
Battin 2004, Hale, Treml, et al. 2015). Despite this, uptake of the ecological trap assessment
framework has been relatively slow (Hale & Swearer 2016).
Current evidence for ecological traps
In their influential review, Robertson and Hutto (2006) proposed a set of criteria for
demonstrating an ecological trap: (1) there must be a suitable measure of habitat preference
indicating preference for, or non-avoidance of, the putative trap habitat relative to alternative
habitats, and (2) there must be a reasonable surrogate measure of fitness that is significantly
lower in the putative trap habitat relative to alternative habitats. Put simply, the preferred
habitat should not confer the best fitness outcomes. Habitat preference can be particularly
difficult to assess in the field, especially in systems where observed population densities are
determined by a combination of passive recruit supply, active habitat preference and early
stage survival (Underwood & Fairweather 1989, Hixon & Beets 1993, Shima 2001, Stevens
2003, Railsback et al. 2003). Robertson and Hutto (2006) proposed that estimates of habitat
preference should be based on one or more of the following lines of evidence: settlement
patterns, distribution of dominant individuals, temporal variance in population size, and
controlled choice experiments. Robertson and Hutto (2006) considered that only 5/45 putative
demonstrations at that time had provided sufficient evidence of habitat preference.
Terrestrial systems
While ecological traps may arise naturally (where a broadly adaptive preference is maladaptive
in certain situations), most proposed and demonstrated cases have been driven by HIREC (Hale
& Swearer 2016). The bulk of evidence for ecological traps comes from terrestrial systems,

4
with birds and ovipositing insects providing especially convenient model systems as fitness can
be easily compared between habitats by tracking clutch sizes and survivorship of adults or
offspring.
Habitat changes associated with human land use are perhaps the most well-documented
drivers of ecological traps. The creation of artificial materials such as plastic, asphalt, oils, glass,
and paint can provide hyperattractive cues that lead insects to choose unsuitable ovipositing
sites (e.g. Horváth et al. 1998; Kriska et al. 1998), while built structures may mimic natural/
nesting sites for birds, but offer poor fitness outcomes (Sumasgutner et al. 2014).
Similarly, human disturbance of native vegetation may mimic natural grasslands or disturbed
forest, but present novel risks for animals that choose to reside in these areas. Balme et al.
(2010) found that the efficacy of a wildlife reserve was reduced by a failure of leopards to
avoid adjacent areas of farmland, where they come into conflict with humans. Negative edge
effects were also observed in birds nesting in vegetation patches fragmented by human land
use: indigo buntings preferentially nested in patches with a high edge length to area ratio—an
adaptation that allows them to exploit natural disturbance—but experienced high chick
mortality in anthropogenic patches (Weldon & Haddad 2005).
Habitat structure is also influenced by exotic or invasive habitat-forming plants, and such
species can drive the creation of ecological traps by altering the quality of the habitat without
affecting associated attractive cues. Lloyd and Martin (2005) found that the presence of an
exotic grass did not alter the attractiveness of a native prairie habitat but was associated with
lower nest survival for longspurs, while Rodewald et al. (2011) reported lower reproductive
success in cardinals that selected exotic honeysuckle as nesting substrate in urban areas. Some
traps may arise due to phenology of exotic habitat-formers: returning migrant blackcaps
preferentially settled in plantations of an exotic tree, perhaps because the associated shrubs
develop spring foliage earlier, but experienced reduced nesting success (Remeš 2003).
Aquatic systems
Marine and freshwater systems have not escaped the reach of HIREC (Halpern et al. 2008,
Vörösmarty et al. 2010), yet there has been comparatively little research effort on behavioural
responses of aquatic fauna to habitat changes (Hale, Coleman, et al. 2015, Hale & Swearer

5
2016). This lack of research reflects difficulties in obtaining habitat preference and fitness
metrics in many aquatic animals, especially broadcast spawning fishes, but it is nonetheless
important to broaden the research effort beyond North American and European birds, as
terrestrial and aquatic fauna are likely to differ in their vulnerability to the various causes of
ecological traps (Hale & Swearer 2016).
Aquatic fauna are likely to be particularly vulnerable to ecological traps caused by pollution,
given the tendency for aquatic environments to transport and accumulate pollutants from
surrounding watersheds, intensifying exposure to toxins (Hale, Coleman, et al. 2015). Perhaps
as a result, pollution-driven ecological traps were among the earliest documented in the
aquatic environment. Juvenile flatfish detect and avoid heavily-oiled sediments, but not lightly-
oiled sediments (Moles et al. 1994). Likewise, juvenile crabs select preferred sediment grain
size regardless of the presence of oil, despite avoiding the oil when it was present on less
preferred sediments (Moles & Stone 2002). Negative fitness effects of oil contamination mean
that affected sediments are likely to function as ecological traps (Moles & Norcross 1998, Khan
2003). Other forms of pollution, such as noise or seismic pollution tend to be avoided,
although exposure prior to avoidance, or sub-avoidance levels of exposure, may have
deleterious effects that could lead to ecological trap formation to the extent that they co-
occur with particular habitats or locations (Codarin et al. 2009, Miller et al. 2014).
Much of the subsequent work on marine ecological traps has assessed habitat selection in
relation to fishing pressure. Natural floating debris signals oceanic convergence zones and high
food availability for pelagic predators, but there is now broad-scale use of artificial fish
aggregation devices (FADs) deployed by fishing fleets in non-convergence zones. These FADs
are associated with lower food availability and high fishing mortality for fish that are attracted
to them (Marsac et al. 2000, Hallier & Gaertner 2008). Similarly, casitas—concrete shelters
used to attract lobsters for harvesting—act as ecological traps for lobsters that find the casitas
more attractive than natural reefs (Gutzler et al. 2015). Fishing pressure may drive ecological
traps even without the need for attractive structures: Abrams et al. (2012) predicted that
some animals may find harvested areas to be more attractive than adjacent marine protected
areas because: (a) potential predators and competitors are rare due to harvesting pressure, (b)
prey species are abundant due to the rarity of predators, and (c) the risk of harvesting
mortality is undetectable. Indirect fishing-driven traps also arise for predators that compete
with fisheries for food. Verhulst et al. (2004) found that oystercatcher populations did not
redistribute following the establishment of a shellfish reserve, despite individuals outside
reserves having fewer shellfish in the diet, poorer condition indices and higher mortality rates.

6
Similarly, African penguins rely on sea surface temperature and primary productivity cues to
find optimum feeding areas, but a combination of fishing pressure and climate change has
driven large scale changes in the size and distribution of forage fish populations such that
these cues no longer reliably predict prey abundance (Sherley et al. 2017).
High densities of wildlife at some marine and freshwater aquaculture systems have prompted
research into the effects of association with farms on wildlife populations. Šigutová et al.
(2015) found that pond management regimes caused high mortality rates for endangered
dragonfly larvae, and recommended measures to make the ponds less attractive to ovipositing
dragonflies. Stocking of fish ponds or sea cages for aquaculture can also alter the availability of
food for piscivorous birds, leading to ecological traps where food availability is lower than
expected (Kloskowski 2012), where high densities of breeding waterbirds attracted to fish
ponds in turn attract nest predators (Broyer et al. 2017), or where culling is carried out (Quick
et al. 2004, Callier et al. 2017). Wild fish attracted to sea cage fish farms had higher condition
indices and a lower incidence of internal parasites, but also higher incidences of external
parasites (Dempster et al. 2011), while captive feeding trials indicate that consuming farm
waste may have deleterious effects on reproduction (Lavens et al. 1999, Mazorra et al. 2003,
Salze et al. 2005, Bogevik et al. 2012). Other studies indicate that marine infrastructure
(whether for aquaculture or other purposes) is probably both attractive and productive for
many fish and benthic invertebrate populations (Reubens et al. 2013, Borgert 2015).
Exotic or invasive species can have disastrous effects on native animal populations (Clavero &
García-Berthou 2005), and are drivers of several demonstrated terrestrial ecological traps
(Robertson & Hutto 2006, Hale & Swearer 2016). Despite this, there have been few
assessments of potential ecological traps driven by non-native aquatic species (candidates may
include novel predators, prey, competitors or habitats). One such case concerns the invasive
macroalga Caulerpa taxifolia, a habitat-forming species that functions as an ecosystem
engineer in soft sediment habitats. In Australia, native bivalves do not avoid C. taxifolia as a
substrate, and recruit in large numbers (Gribben & Wright 2006, Gribben et al. 2009). Recruits
fare well, but there is evidence that mortality risk over an individual’s lifetime is higher in the
invasive habitat (Gribben & Wright 2006, Wright & Gribben 2008, Byers et al. 2010).

7
Thesis aims and structure
Habitat association studies are a cornerstone of the animal ecology field, yet only a fraction
provide sufficient evidence to place habitats within a source–sink metapopulation framework,
let alone determine whether individuals utilising or avoiding habitats are making adaptive
habitat selection decisions (Hale & Swearer 2016). This is especially true for marine systems,
where most investigations provide only basic population- or community-level data. The
examples cited in the previous section constitute nearly all published investigations of
potential ecological traps in the marine environment. This is despite the ecological trap
concept being first articulated more than four decades ago (Dwernychuk & Boag 1972, Gates &
Gysel 1978), and refined over subsequent decades (Battin 2004, Robertson & Hutto 2006,
Gilroy & Sutherland 2007, Patten & Kelly 2010, Hale & Swearer 2016). Related calls have been
made regarding the analogous attraction–production controversy around fish populations on
artificial reefs (Bohnsack 1989, Osenberg et al. 2002, Brickhill et al. 2005), but studies that
separate these key population processes remain rare.
Accordingly, this thesis examines habitat selection and fitness consequences for animals in
several human-impacted coastal marine environments using the ecological trap assessment
framework, and aims to link these individual-level traits to the potential for population
persistence in degraded environments. The aim was not necessarily to demonstrate the
existence of ecological traps in these environments, but rather to demonstrate the application
of the ecological trap assessment framework to questions around the value of novel habitats
for fauna. I worked across three study systems modified by aquaculture and invasive species to
demonstrate the broad applicability of this individual-level approach.
Chapters 2-5 were written as standalone manuscripts, with minor changes to fit the thesis
format. As a result, some repetition of key concepts—particularly HIREC and ecological trap
theory—has been unavoidable. I have minimised unnecessary repetition by confining
descriptions of my study systems to the relevant chapters.
In Chapter Two, I conduct a global systematic review and meta-analysis of research on
interactions between all forms of aquaculture and vertebrate wildlife, with an emphasis on the
impacts of aquaculture on the distribution, fitness and population persistence of wildlife. I
demonstrate that there are considerable knowledge gaps around very basic questions,
including individual responses to aquaculture (i.e. the degree of attraction or repulsion relative

8
to natural habitats), and whether proximity to aquaculture is likely to have positive or negative
effects on fitness and population persistence of vertebrate wildlife.
Chapter Three concerns one of the key knowledge gaps highlighted by Chapter 2: the effect of
proximity to aquaculture on the fitness of wild fish. We have multiple lines of evidence that
wild fish are attracted to aquaculture (see Chapter 2), where they experience high food
availability (Dempster et al. 2011), but the effects of proximity to aquaculture on reproductive
fitness are poorly known. In this chapter, I collect adult Atlantic cod (Gadus morhua) in
spawning condition from sites within two areas of high and low salmon farming density, and
conduct a hatchery spawning experiment to compare egg production, egg quality, and larval
fitness metrics.
In Chapter Four, I study a complex interaction between shellfish aquaculture, a native
keystone predator (the eleven-arm seastar Coscinasterias muricata) and an invasive keystone
predator (the northern Pacific seastar Asterias amurensis). Shellfish aquaculture provides a
large trophic subsidy for benthic fauna, and has the potential to act as an important population
source for both species. However, as the native seastar has been reported to prey upon the
invasive seastar, there may be some predation pressure occurring that reduces the value of
the shellfish farm habitat for the invader, potentially leading to an ecological trap. Accordingly,
I investigate the behavioural responses of the invasive seastar to the shellfish farm habitat and
the native predator, and assess fitness metrics for seastars inside and outside the farms.
In Chapter Five, I consider the attractiveness and fitness value of habitat created by an
invasive ecosystem engineer (the wakame kelp Undaria pinnatifida) for native fish on urban-
impacted temperate reefs. This invasive habitat-forming kelp provides a seasonal canopy on
reefs where urchin-grazing and urban impacts have driven the decline of native kelp cover. I
combine a laboratory habitat choice experiment, recruitment data on mesocosm reefs, fish
community surveys on natural reefs, and fitness metrics to provide a holistic assessment of the
relative value of this novel habitat.
Finally, Chapter Six provides a general discussion of this body of work, and reflects on the
utility of the ecological trap framework in assessing the role of novel habitats in population
persistence of marine fauna.

9
REFERENCES
Abrams PA, Ruokolainen L, Shuter BJ, McCann KS (2012) Harvesting creates ecological traps:
consequences of invisible mortality risks in predator-prey metacommunities. Ecology
93:281–293
Balme GA, Slotow R, Hunter LTB (2010) Edge effects and the impact of non-protected areas in
carnivore conservation: leopards in the Phinda–Mkhuze Complex, South Africa. Anim
Conserv 13:315–323
Battin J (2004) When good animals love bad habitats: ecological traps and the conservation of
animal populations. Conserv Biol 18:1482–1491
Bogevik AS, Natário S, Karlsen Ø, Thorsen A, Hamre K, Rosenlund G, Norberg B (2012) The
effect of dietary lipid content and stress on egg quality in farmed Atlantic cod Gadus
morhua. J Fish Biol 81:1391–405
Bohnsack JA (1989) Are high-densities of fishes at artificial reefs the result of habitat limitation
or behavioral preference. Bull Mar Sci 44:631–645
Borgert RJ (2015) Value of off-bottom oyster aquaculture baskets as habitat for juvenile blue
crab. MSc thesis. Auburn University, Alabama
Brickhill MJ, Lee SY, Connolly RM (2005) Fishes associated with artificial reefs: attributing
changes to attraction or production using novel approaches. J Fish Biol 67:53–71
Broyer J, Chavas G, Chazal R (2017) The effects of cessation of fish farming on duck breeding in
French fishpond systems. Hydrobiologia 788:47–53
Byers JE (2002) Impact of non-indigenous species on natives enhanced by anthropogenic
alteration of selection regimes. Oikos 97:449–458
Byers JE, Wright JT, Gribben PE (2010) Variable direct and indirect effects of a habitat-
modifying invasive species on mortality of native fauna. Ecology 91:1787–1798
Callier MD, Byron CJ, Bengtson DA, Cranford PJ, Cross SF, Focken U, Jansen HM, Kamermans P,
Kiessling A, Landry T, O’Beirn F, Petersson E, Rheault RB, Strand Ø, Sundell K, Svåsand T,
Wikfors GH, McKindsey CW (2017) Attraction and repulsion of mobile wild organisms to
finfish and shellfish aquaculture: a review. Rev Aquac. DOI: 10.1111/raq.12208
Clavero M, García-Berthou E (2005) Invasive species are a leading cause of animal extinctions.
Trends Ecol Evol 20:110
Codarin A, Wysocki LE, Ladich F, Picciulin M (2009) Effects of ambient and boat noise on
hearing and communication in three fish species living in a marine protected area
(Miramare, Italy). Mar Pollut Bull 58:1880–1887

10
Crooks JA, Chang AL, Ruiz GM (2011) Aquatic pollution increases the relative success of
invasive species. Biol Invasions 13:165–176
Dempster T, Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere JT, Nilsen R, Bjørn P-A, Uglem
I (2011) Proxy measures of fitness suggest coastal fish farms can act as population
sources and not ecological traps for wild gadoid fish. PLoS ONE 6:e15646
Dias PC (1996) Sources and sinks in population biology. Trends Ecol Evol 11:326–330
Dwernychuk LW, Boag DA (1972) Ducks nesting in association with gulls - an ecological trap?
Can J Zool 50:559–563
Gates JE, Gysel LW (1978) Avian nest dispersion and fledging success in field-forest ecotones.
Ecology 59:871–883
Gilroy JJ, Sutherland WJ (2007) Beyond ecological traps: perceptual errors and undervalued
resources. Trends Ecol Evol 22:351–356
Gribben PE, Wright JT (2006) Invasive seaweed enhances recruitment of a native bivalve: roles
of refuge from predation and the habitat choice of recruits. Mar Ecol Prog Ser 318:177–
185
Gribben PE, Wright JT, O’Connor WA, Steinberg P (2009) Larval settlement preference of a
native bivalve: the influence of an invasive alga versus native substrata. Aquat Biol 7:217–
227
Gutzler BC, Butler MJ, Behringer DC (2015) Casitas: a location-dependent ecological trap for
juvenile Caribbean spiny lobsters, Panulirus argus. ICES J Mar Sci 72:i177–i184
Hale R, Coleman R, Pettigrove V, Swearer SE (2015) Identifying, preventing and mitigating
ecological traps to improve the management of urban aquatic ecosystems. J Appl Ecol
52:928–939
Hale R, Swearer SE (2016) Ecological traps: current evidence and future directions. Proc R Soc B
Biol Sci 283:494–499
Hale R, Treml EA, Swearer SE (2015) Evaluating the metapopulation consequences of
ecological traps. Proc R Soc London B Biol Sci 282:20142930
Hallier J-P, Gaertner D (2008) Drifting fish aggregation devices could act as an ecological trap
for tropical tuna species. Mar Ecol Prog Ser 353:255–264
Halpern BS, Walbridge S, Selkoe KA, Kappel C V, Micheli F, D’Agrosa C, Bruno JF, Casey KS,
Ebert C, Fox HE, Fujita R, Heinemann D, Lenihan HS, Madin EMP, Perry MT, Selig ER,
Spalding M, Steneck R, Watson R (2008) A global map of human impact on marine
ecosystems. Science 319:948–952
Hixon MA, Beets JP (1993) Predation, prey refuges, and the structure of coral‐reef fish

11
assemblages. Ecol Monogr 63:77–101
Holt RD (1985) Population dynamics in two-patch environments: some anomalous
consequences of an optimal habitat distribution. Theor Popul Biol 28:181–208
Horváth G, Bernáth B, Molnár G (1998) Dragonflies find crude oil visually more attractive than
water: multiple-choice experiments on dragonfly polarotaxis. Naturwissenschaften
85:292–297
Khan RA (2003) Health of flatfish from localities in Placentia Bay, Newfoundland, contaminated
with petroleum and PCBs. Arch Environ Contam Toxicol 44:485–492
Kloskowski J (2012) Fish stocking creates an ecological trap for an avian predator via effects on
prey availability. Oikos 121:1567–1576
Kokko H, Sutherland WJ (2001) Ecological traps in changing environments: ecological and
evolutionary consequences of a behaviourally mediated Allee effect. Evol Ecol Res 3:537–
551
Kriska G, Horváth G, Andrikovics S (1998) Why do mayflies lay their eggs en masse on dry
asphalt roads? Water-imitating polarized light reflected from asphalt attracts
Ephemeroptera. J Exp Biol 201:2273–2286
Lavens P, Lebegue E, Jaunet H, Brunel A, Dhert P, Sorgeloos P (1999) Effect of dietary essential
fatty acids and vitamins on egg quality in turbot broodstocks. Aquac Int 7:225–240
Lloyd JD, Martin TE (2005) Reproductive success of chestnut-collared longspurs in native and
exotic grassland. Condor 107:363–374
Marsac F, Fonteneau A, Ménard F (2000) Drifting FADs used in tuna fisheries: an ecological
trap? In: Gall J-Y Le, Cayre P, Taquet M (eds) Pêche Thonière et Dispositifs de
Concentration de Poissons, 28th edn. Actes Colloq. - IFREMER, p 537–552
Mazorra C, Bruce M, Bell JG, Davie A, Alorend E, Jordan N, Rees J, Papanikos N, Porter M,
Bromage N (2003) Dietary lipid enhancement of broodstock reproductive performance
and egg and larval quality in Atlantic halibut (Hippoglossus hippoglossus). Aquaculture
227:21–33
Miller PJO, Antunes RN, Wensveen PJ, Samarra FIP, Catarina Alves A, Tyack PL, Kvadsheim PH,
Kleivane L, Lam F-PA, Ainslie MA, Thomas L (2014) Dose-response relationships for the
onset of avoidance of sonar by free-ranging killer whales. J Acoust Soc Am 135:975–993
Moles A, Norcross BL (1998) Effects of oil-laden sediments on growth and health of juvenile
flatfishes. Can J Fish Aquat Sci 55:605–610
Moles A, Rice S, Norcross BL (1994) Non-avoidance of hydrocarbon laden sediments by
juvenile flatfishes. Netherlands J Sea Res 32:361–367

12
Moles A, Stone RP (2002) Habitat preferences of juvenile tanner and red king crabs: substrate
and crude oil. In: Paul AJ, Dawe EG, Elner R, Jamieson GS, Kruse GH, Otto RS, Sainte-Marie
B, Shirley TC, Woodby D (eds) Crabs in Cold Water Regions: Biology, Management, and
Economics. University of Alaska Sea Grant, p 631–644
Osenberg C, Mary CM St., Wilson JA, Lindberg WJ (2002) A quantitative framework to evaluate
the attraction-production controversy. ICES J Mar Sci 59:S214–S221
Patten MA, Kelly JF (2010) Habitat selection and the perceptual trap. Ecol Appl 20:2148–2156
Pulliam HR (1988) Sources, sinks, and population regulation. Am Nat 132:652–661
Quick NJ, Middlemas SJ, Armstrong JD (2004) A survey of antipredator controls at marine
salmon farms in Scotland. Aquaculture 230:169–180
Railsback SF, Stauffer HB, Harvey BC (2003) What can habitat preference models tell us? Tests
using a virtual trout population. Ecol Appl 13:1580–1594
Remeš V (2003) Effects of exotic habitat on nesting success, territory density, and settlement
patterns in the blackcap (Sylvia atricapilla). Conserv Biol 17:1127–1133
Reubens JT, Vandendriessche S, Zenner AN, Degraer S, Vincx M (2013) Offshore wind farms as
productive sites or ecological traps for gadoid fishes? Impact on growth, condition index
and diet composition. Mar Environ Res 90:66–74
Robertson BA, Hutto RL (2006) A framework for understanding ecological traps and an
evaluation of existing evidence. Ecology 87:1075–1085
Robertson BA, Rehage JS, Sih A (2013) Ecological novelty and the emergence of evolutionary
traps. Trends Ecol Evol 28:552–560
Rodewald AD, Shustack DP, Jones TM (2011) Dynamic selective environments and evolutionary
traps in human‐dominated landscapes. Ecology 92:1781–1788
Salze G, Tocher DR, Roy WJ, Robertson DA (2005) Egg quality determinants in cod (Gadus
morhua L.): egg performance and lipids in eggs from farmed and wild broodstock. Aquac
Res 36:1488–1499
Sanderson EW, Jaiteh M, Levy MA, Redford KH, Wannebo A V, Woolmer G (2002) The human
footprint and the last of the wild. Bioscience 52:891–904
Schlaepfer MA, Runge MC, Sherman PW (2002) Ecological and evolutionary traps. Trends Ecol
Evol 17:474–480
Sherley RB, Ludynia K, Dyer BM, Lamont T, Makhado AB, Roux J-P, Scales KL, Underhill LG,
Votier SC (2017) Metapopulation tracking juvenile penguins reveals an ecosystem-wide
ecological trap.
Shima JS (2001) Recruitment of a coral reef fish: roles of settlement, habitat, and

13
postsettlement losses. Ecology 82:2190–2199
Šigutová H, Šigut M, Dolný A (2015) Intensive fish ponds as ecological traps for dragonflies: an
imminent threat to the endangered species Sympetrum depressiusculum (Odonata:
Libellulidae). J Insect Conserv 19:961–974
Sih A (2013) Understanding variation in behavioural responses to human-induced rapid
environmental change: a conceptual overview. Anim Behav 85:1077–1088
Sih A, Ferrari MCO, Harris DJ (2011) Evolution and behavioural responses to human-induced
rapid environmental change. Evol Appl 4:367–387
Stevens BG (2003) Settlement, substratum preference, and survival of red king crab
Paralithodes camtschaticus (Tilesius, 1815) glaucothoe on natural substrata in the
laboratory. J Exp Mar Biol Ecol 283:63–78
Sumasgutner P, Nemeth E, Tebb G, Krenn HW, Gamauf A (2014) Hard times in the city -
attractive nest sites but insufficient food supply lead to low reproduction rates in a bird
of prey. Front Zool 11:48
Underwood AJ, Fairweather PG (1989) Supply-side ecology and benthic marine assemblages.
Trends Ecol Evol 4:16–20
Verhulst S, Oosterbeek K, Rutten AL, Ens BJ (2004) Shellfish fishery severely reduces condition
and survival of oystercatchers despite creation of large marine protected areas. Ecol Soc
9:17
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of Earth’s
ecosystems. Science (80- ) 277:494–499
Vörösmarty CJ, McIntyre PB, Gessner MO, Dudgeon D, Prusevich A, Green P, Glidden S, Bunn
SE, Sullivan CA, Liermann CR, others (2010) Global threats to human water security and
river biodiversity. Nature 467:555–561
Weldon AJ, Haddad NM (2005) The effects of patch shape on indigo buntings: evidence for an
ecological trap. Ecology 86:1422–1431
Wong BBM, Candolin U (2014) Behavioral responses to changing environments. Behav Ecol
26:665–673
Wright JI, Gribben PE (2008) Predicting the impact of an invasive seaweed on the fitness of
native fauna. J Appl Ecol 45:1540–1549

14
CHAPTER TWO: IMPACTS OF MARINE AND FRESHWATER AQUACULTURE ON WILDLIFE: A
GLOBAL META-ANALYSIS
ABSTRACT
The global expansion of aquaculture has raised concerns about its environmental impacts,
including effects on wildlife. Aquaculture farms are thought to repel some species and function
as either attractive population sinks (‘ecological traps’) or population sources for others. We
conducted a systematic review and meta-analysis of empirical studies documenting
interactions between aquaculture operations and vertebrate wildlife. Farms were associated
with elevated local abundance and diversity of wildlife, although this overall effect was
strongly driven by aggregations of wild fish at sea cages and shellfish farms (abundance: 72x;
species richness: 2.0x). Birds were also more diverse at farms (1.1x), but other taxa showed
variable and comparatively small effects. Larger effects were reported when researchers
selected featureless or unstructured habitats as reference sites. Evidence for aggregation
‘hotspots’ is clear in some systems, but we cannot determine if farms act as ecological traps
for most taxa, as few studies assess either habitat preference or fitness in wildlife. Fish
collected near farms were larger and heavier with no change in body condition, but also faced
higher risk of disease and parasitism. Birds and mammals were frequently reported preying on
stock, but little data exists on the outcomes of such interactions for birds and mammals –
farms are likely to function as ecological traps for many species. We recommend researchers
measure survival and reproduction in farm-associated wildlife to make direct, causal links
between aquaculture and its effects on wildlife populations.
INTRODUCTION
Aquaculture infrastructure (farms hereafter) presents a novel environment for wild animal
populations. High stocking densities within farms aggregate biomass far beyond natural levels
(commonly 5-45 kg m-3 final biomass: FAO Fisheries and Aquaculture 2018), and in open
systems, provide considerable trophic subsidies for animals that take advantage of the
opportunity, potentially benefitting some wildlife. However, there are also deleterious effects
associated with proximity to farms, and the net impact of aquaculture on productivity and

15
persistence of wildlife populations will depend both on behavioural responses to farms and
the fitness consequences of those responses. Where individuals are attracted to a habitat that
confers poorer fitness outcomes than other available habitats, they have fallen into an
‘ecological trap’ (Robertson & Hutto 2006; Hale & Swearer 2016). While the concept is defined
at the individual level, trap habitats have population-level consequences by drawing
individuals from surrounding habitats into attractive population sinks (Hale et al. 2015). Even
in the absence of an ecological trap, changes in the abundance and spatial distribution of
influential species may indirectly affect other species and drive large-scale shifts in biodiversity
and ecosystem function (Gamfeldt et al. 2015).
A range of attractive and repulsive mechanisms for wildlife can occur simultaneously at farms
(Callier et al. 2017). The primary attractive mechanism in most systems is probably food
availability, either in the form of direct predation on stock, or an indirect trophic subsidy in the
form of farm waste (spilled feed, faeces and dead stock). Birds, pinnipeds and otters are well-
documented predators of stock at sea cage or pond fish farming systems (Carss 1993; Pitt &
Conover 1996; Adámek et al. 2003; Güçlüsoy & Savas 2003; Quick et al. 2004; Freitas et al.
2007; Dorr et al. 2012; Sepúlveda et al. 2015), while farm waste from sea cages also attracts
significant aggregations of opportunistic wild fish (Dempster et al. 2002, 2009; Tuya et al.
2006; Sanchez-Jerez et al. 2011). A high local abundance of fish is likely to lead to secondary
attraction of large predators, such as dolphins (Diaz López 2006; Piroddi et al. 2011). Shellfish
and algae farming do not require inputs of feed, but high densities of filter feeding shellfish in
farms do accumulate biomass, attracting wild fish and invertebrate species (Dealteris et al.
2004; Powers et al. 2007; McKindsey et al. 2011; Segvic-Bubic et al. 2011), while algae farming
attracts wild herbivores (Hehre & Meeuwig 2016). Farm structures themselves may also be
attractive, functioning in a similar manner to fish aggregation devices or artificial reefs
(Tallman & Forrester 2007; Sanchez-Jerez et al. 2011). Farm structures provide three-
dimensional habitat complexity, and associated light, noise and novel biofouling communities
may all be attractive to a range of wild taxa (Dumont et al. 2011; Callier et al. 2017).
Paradoxically, many of these environmental changes associated with farms, such as light,
noise, eutrophication and high densities of predators, may have repulsive effects on wary or
functionally specialised taxa (Markowitz et al. 2004; Becker et al. 2011).
Attraction to farms may increase or decrease the fitness of wildlife. One expectation is that
increased food availability will lead to faster growth, higher body condition and increased
reproductive output. Accordingly, there is some evidence that farm-associated wild fish have
higher body condition and reproductive investment indices than fish from reference sites

16
(Dempster et al. 2011), but little is known about potential benefits for other taxa. In broadcast
spawning taxa, high local population densities at farms are likely to confer greater mating
efficiency (Inglis & Gust 2003). Such benefits for farm-associated wildlife are likely to be at
least partially counteracted by potential deleterious fitness effects related to dietary shifts,
contamination, disease, parasitism, and elevated mortality rates. For example, a shift from fish
oils to terrestrially-derived ingredients in aquaculture feed may result in deficiencies of long-
chain polyunsaturated fatty acids in animals that feed regularly at farms (eg. Salze et al. 2005;
Fernandez-Jover et al. 2007a; Gonzalez-Silvera et al. 2017). Additionally, farm waste can create
an anoxic environment with significant effects on benthic and estuarine communities (Wu
1995; Yucel-Gier et al. 2007; Herbeck et al. 2013; Valdemarsen et al. 2015), while in some
areas, wildlife may also accumulate elevated tissue loadings of contaminants such as
antibiotics, pyrethroid parasiticides, metals and organohalogens (Samuelsen et al. 1992; Boyd
& Massaut 1999; Burridge et al. 2010; Bustnes et al. 2010) with potentially nontrivial effects
(e.g. Crump & Trudeau 2009; Berg et al. 2016). For fish, the primary concern may be the effect
of proximity to farms on disease and parasitism rates: high population densities within farms
create favourable conditions for outbreaks of diseases and parasites such as sea lice (Krkosek
et al. 2005, 2006; Costello 2009; Lafferty et al. 2015; Krkošek 2017). Wild fish populations may
also act as reservoirs for parasites and diseases, and as they move between cages to take
advantage of feeding opportunities, they act as potential transmission vectors that may
increase reinfection rates for farms, driving positive feedbacks (Uglem et al. 2009; Hayward et
al. 2011).
Despite this suite of environmental concerns, the aquaculture industry is the world’s fastest-
growing food production sector (FAO Fisheries and Aquaculture 2015). For this growth to be
sustainable in terms of environmental impacts and ‘social license’ to operate, the industry
must grapple with issues arising from interactions between aquaculture activities and the
natural environment and develop solutions to minimise negative effects on wildlife (and vice
versa). The first step should be to assess the state of knowledge on these issues and identify
the most severe effects. Recent reviews have outlined the range of interactions that occur
between aquaculture activities and wild fauna (e.g. Uglem et al. 2014; Taranger et al. 2015;
Glover et al. 2017; Callier et al. 2017), but there has been not yet been a quantitative global
synthesis of the impacts of aquaculture on wildlife. Here, we conduct a systematic review and
meta-analysis of studies documenting interactions between aquaculture activities and wildlife,
primarily to quantify the effects of these interactions on abundance, diversity, and fitness of
farm-associated wildlife, and secondarily to highlight potential drivers of conflict between

17
wildlife and aquaculture. Thereafter, we recommend directions for future research to address
key knowledge gaps in this area.
MATERIALS AND METHODS
Literature search and systematic review
Primary publications up to November 2017 were discovered by searching for the following
terms using the ISI Web of Science: (aquaculture OR mariculture OR "fish farm*" OR "shellfish
farm*" OR "mussel farm*" OR "oyster farm*" OR "sea cage*" OR "net pen*" OR "fish pond*"
OR "seaweed farm*" OR "macroalgal farm*" OR "algal farm*") AND (attract* OR avoid* OR
wild OR aggreg* OR impact* OR depredat* OR predat* OR disease) AND (wildlife OR animal*
OR fauna* OR fish* OR shark* OR mammal* OR dolphin* OR cetacean* OR otter* OR seal* OR
sea lion* OR bird* OR avian OR reptile* OR snake* OR amphibian* OR frog*). >9000 results
were manually screened on an individual basis, by title and abstract alone where the topic was
clearly irrelevant, or else after accessing the full text. Additional articles missed by our initial
search were discovered using informal exploratory searches using Google Scholar, and by
reading the reference lists of all relevant articles returned by our initial search. Our search
focused on interactions with vertebrate wildlife (defined here as fish, birds, mammals and
reptiles), as these animals are typically highly mobile and are therefore more able to make
decisions about whether to reside at and interact with farms.
For inclusion, publications were required to have provided empirical field data on at least one
of the following: (1) distribution, behaviour, condition, disease or mortality of wildlife in the
vicinity of aquaculture sites, or (2) direct interactions between wildlife and stock at
aquaculture sites (e.g. predation of stock). To minimise potential duplication of data, we only
included peer-reviewed English-language journal articles.
To document the distribution of research effort in the field, we recorded the year, country,
region, environment, culture system, culture taxa and the wild taxa for each study.

18
Meta-analysis
Studies were included in the subsequent meta-analysis if they provided quantitative data
sufficient to calculate effect sizes for variables at aquaculture sites relative to ‘natural’ or
‘reference’ sites. We extracted a range of quantitative variables that were representative of
the dominant types of interactions between aquaculture operations and wild vertebrates,
relating to spatial distribution (Abundance, Species Richness), size structure (Length, Weight),
food availability (Body Condition, Stomach Fullness), disease and parasite infection levels –
either infection loads on individuals or prevalence of infected individuals in the population
(Infection Level), as well as direct measures of Mortality and Fertility. Reproductive condition
metrics (e.g. relative gonad size) were considered a component of Body Condition.
Natural log response ratios were calculated for each variable: RR = ln(F/R), where F is the trait
mean at farm sites and R is the trait mean at reference sites. Taking the natural log of the
response ratio normalises the error distribution by reducing the influence of positive
responses (Hedges et al. 1999). Studies employed a variety of sampling designs, including
random or matched farm and reference sites, and stocked or unstocked farms. All were
treated as random for the purposes of this meta-analysis, with RR calculated from the mean
trait values across all farm and reference sites regardless of how sites were selected by the
authors. Where multiple complementary measures were available for a response variable, we
took the mean of those measures (for example, Fulton’s K, hepatosomatic index and
gonadosomatic index all contribute to the Body Condition variable). Where a study provided
data on a response variable from multiple species or sites, we combined data to provide a
single replicate, except where data spanned multiple culture systems (e.g. cages and ponds),
taxonomic classes (e.g. birds and mammals), environments (e.g. marine and freshwater), or
countries. No article contributed more than two studies to our database. This was done to
prevent studies that provided data on numerous species from having a disproportionate
influence on our findings, and to ensure spatial independence between replicates given the
high mobility of most species studied. Where data was provided for farms with and without
exclusion measures (e.g. fenced and unfenced sites), we used data from sites without
exclusion measures.
Some variables were not easily quantified for statistical analysis but were nonetheless
important in understanding interactions between farms and wild fauna. These included
changes in tissue fatty acid profiles, trace elements and stable isotopes, contamination from

19
antibiotics, heavy metals and other substances, and behavioural data such as residence time or
visitation rates. For these variables, we recorded the response ratio if possible, otherwise we
noted the direction or nature of the effect.
Statistical analyses
To test for a significant effect of farm-association on response variables, we checked normality
before conducting one sample t-tests on RR data (mean RR under null hypothesis of no farm
effect = 0) using R software (R Core Team 2017).
Exploratory model selection was used to determine which of the following factors best
predicted effects of farms on wildlife (abundance and species richness responses only, as
remaining responses had insufficient sample sizes for exploratory analysis): Year, Country,
Continent, Environment (Marine, Freshwater), Culture System (Cage, Pond, Longline, Rack,
Bed), Cultured Taxa (Fish, Shellfish, Crustacean, Alga), Wild Taxa (Fish, Bird, Mammal, Reptile,
Amphibian), and Reference Habitat (Structured, Unstructured). We fitted a global general
linear model using R, and employed the dredge() function in the MuMIn package (Barton 2016)
to compare the second-order Akaike’s Information Criterion (AICC) score of every possible
subset of the global model. AICC includes a correction for finite sample sizes and yields more
conservative models than AIC (Burnham and Anderson 2002). We selected the model with the
lowest AICC score, and then used the likelihood ratio to test whether the selected model
offered a significantly better fit than the null (intercept only) model, tested the significance of
model terms, and then conducted post-hoc tests with a Tukey correction to test pairwise
effects within significant model terms.
There was orders-of-magnitude variation in RRs for abundance and species richness among
studies and systems, and accordingly, the overall trends that we report may be strongly
influenced by a small number of studies with unusually large RRs. To test this possibility, we
conducted a sensitivity analysis by ranking studies (replicates) according to the absolute value
of the RR, removing the studies with the largest RR in a stepwise fashion, and rerunning the
model between each removal (Bancroft et al. 2007; Kroeker et al. 2010). We then report the
number of studies than can be removed from the dataset without altering the statistical
significance of the farm effect.

20
To test whether the geographic distribution of research effort on this topic corresponds to the
distribution of aquaculture production, we fitted a zero-inflated Poisson model (using the pscl
package for R: Zeileis et al. 2008) to compare the number of studies contributed by each
country with the reported aquaculture production (t) by that country (FAO Fisheries and
Aquaculture 2017). To account for the large disparity in peer-reviewed English language
research output between developed and developing nations, we also included the United
Nations Human Development Index as a model term (United Nations Development
Programme 2017).
Figure 2.1. Distribution of research effort for studies that met the criteria for inclusion in our
database, according to (A) Country, (B) Culture system, (C) Region, (D) Culture taxa, (E)
Environment, and (F) Wild taxa.

21
RESULTS
Our searches discovered 204 relevant studies across 191 articles published between 1978 and
2017 (Appendix 2.1). 91 studies provided comparative data on wildlife populations at farms
and reference sites suitable for inclusion in the meta-analysis of log response ratios (RR).
Distribution of research effort
There was a clear geographical bias in research effort within our database, with 114 peer-
reviewed English language studies conducted in Europe and 46 in North America (Fig. 2.1).
Among nations, Norway, the United States and Spain accounted for the most research (Fig.
2.1). Research effort across nations was significantly predicted by an interaction between the
size of the nation’s aquaculture industry and the developmental index of the nation (p = 0.03,
Appendix 2.2), wherein highly developed nations (especially those in Europe and North
America: Fig. 2.2) with large production contributed more studies than those with low
production (p <0.0001, Appendix 2.2). Several major aquaculture-producing nations were
either poorly represented or entirely absent from our database: most notably, mainland China
is by an order of magnitude the largest aquaculture producer in the world (FAO Fisheries and
Aquaculture 2017), yet was entirely absent from our database. Other leading producers,
namely Indonesia, India, Vietnam, Philippines and Bangladesh, were also either absent or
represented by only a single study.
Most studies in our database assessed interactions with wildlife in marine or estuarine
environments (Fig. 2.1), despite global animal aquaculture production being considerably
higher in freshwater environments (47 cf. 27 million t in 2014) (FAO Fisheries and Aquaculture
2015). 105/144 studies in the marine environment took place at sea cage farms, while 49/60
freshwater systems were pond-based (Fig. 2.1). Fish were the most common cultured taxa
studied (163 studies) – primarily salmonids (69 studies) in western Europe and the Americas,
and sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax) in southern Europe (43
studies). The research effort on environmental effects of salmon farming is in line with the
predominance of salmonids in the marine fish farming sector, although freshwater cyprinid
culture is the most productive pisciculture sector overall (FAO Fisheries and Aquaculture
2015). Sea bream, sea bass and marine shellfish systems are overrepresented in our dataset

22
relative to the size of these sectors, perhaps due to their importance for nations with high
marine research activity (particularly Spain). Algal and crustacean culture (5 and 3 studies,
respectively) were dramatically underrepresented here relative to the size of the sectors (FAO
Fisheries and Aquaculture 2015).
Most studies reported interactions with wild fish (108 studies), followed by birds (53 studies),
mammals (38 studies), reptiles (3 studies) and amphibians (2 studies) (Fig. 2.1).
Figure 2.2. Distribution of research effort on interactions between aquaculture sites and wild
fauna among countries and territories. Production data taken from the Fishstatj database (FAO
Fisheries and Aquaculture Department 2017).
Effects on wildlife
Abundance
We discovered 65 studies that quantified the abundance of wildlife at aquaculture farming
sites compared to reference sites, using various forms of Control-Impact (CI), Before-After (BA)
and Control-Impact-Before-After (BACI) designs. These studies used a variety of sampling
methods, including visual census, catch-per-unit-effort and tagging/tracking. 17 studies
reported a lower abundance near farms, two no difference, and 46 a higher abundance. The

23
mean effect was a 49x increase in abundance near farms (RR = 1.05, t64 = 4.3, p <0.0001), but
this value was strongly influenced by a few outlier studies reporting very large aggregations of
wild fish around sea cages (for example, a mean 1327x increase over three sampling dates at
one Australian offshore farm compared to featureless mid-water reference sites: Dempster et
al., 2004). Fish demonstrated the largest abundance changes, while changes in bird and
mammal abundance were highly variable in both effect size and direction and not significantly
different to zero (Fig. 2.3, Table 1). We were not able to calculate RR for an additional six
studies reporting differential abundance at farms (fish: 2/2 higher; mammals: 1/2 higher; birds:
2/2 higher).
A sensitivity analysis revealed that it was possible to conduct stepwise removal of 25/65
replicates with the largest effect sizes without losing statistical significance, indicating that the
overall trend was robust. However, when studies that assessed changes in wild fish abundance
at sea cage systems were omitted from the analysis, the remaining studies did not provide
support for an overall effect of aquaculture on wildlife abundance (t38 = 0.81, p = 0.42),
indicating that wild fish aggregations around sea cages were largely responsible for this overall
effect.
Model selection indicated that differential abundance was best predicted by a model
containing Environment, Cultured Taxa and Reference Habitat (R2 = 0.33, F = 7.4, p <0.0001;
Appendix 2.3). The Cultured Taxa term was significant (p = 0.0001), as was Reference Habitat
(p = 0.003), while Environment was not (p = 0.12). Post-hoc testing revealed that increases in
abundance of wild fauna tended to be higher at fish farms than at shellfish farms (p = 0.001)
and that studies comparing abundance at farm sites to unstructured or featureless reference
sites (e,g, sandy seabed or open ocean) generally found a larger response than those that
chose natural reef, unstocked farms or other structured habitats as reference sites (p = 0.006;
Appendix 2.4).
Species richness
Most studies only assessed a limited number of target species, but 37 studies provided useful
data on species richness at farms and reference sites. Of these, all but six reported higher
species richness at farm sites, with a mean 1.7x increase (RR = 0.30, t36 = 3.1, p = 0.004). This
effect was strongest in fish (RR = 0.43) but was also significant in birds (RR = 0.13) (Table 1).

24
Only one study in our database quantified differential species richness in mammals, and one in
amphibians (Roycroft et al. 2004; Kloskowski 2010, Table 1).
There was large variation in effect size and direction across studies, but a sensitivity analysis
found the overall trend to be remarkably robust (25/37 studies removed without losing
statistical significance). As with abundance, the effect was not significant when sea cage
systems were omitted from the analysis (t21 = 1.6, p = 0.11).
Species richness effects were best predicted by a model containing Reference Habitat and Wild
Taxa (R2 = 0.27, F = 4.3, p = 0.007; Appendix 2.3). Post-hoc testing revealed that the only
significant pairwise effect was between fish and amphibians, with fish species richness
positively affected and amphibian species richness negatively affected by the presence of
aquaculture sites in their respective environments (p = 0.03; Appendix 2.5).
Size structure, body condition and stomach fullness
Wild fish collected near aquaculture sites were on average 1.2x larger and 1.7x heavier than
their counterparts from reference sites (Table 1), but no size comparisons were available for
non-fish taxa.
Most studies (11) reported trends toward higher condition metrics in farm-associated wildlife,
while two found no difference and two lower condition metrics at farms, although there was
no significant effect overall (Table 1). Similarly, 8/11 studies found higher rates of stomach
fullness in farm-associated wildlife, but these effects tended to be small and were not
significant overall (Table 1). All but two comparisons of body condition or stomach fullness
data concerned wild marine fish, while Gregory and Nelson (1991) estimated a 1.9x higher rate
of stomach fullness in snakes at fish hatcheries, and Kloskowski et al. (2017) reported higher
physiological stress indicators (=lower condition for our purposes) in grebes nesting on fish
ponds.
Physiological changes
All 16/17 studies that reported looking for physiological or dietary changes in farm-associated
wild fish relative to those from reference sites found evidence of dietary shifts, while the

25
remainder found only minor differences in stable isotopes (Johnston et al. 2010). Evidence for
dietary shift included farm feed pellets in the stomachs of farm-associated wild fish (Skog et al.
2003; Fernandez-Jover et al. 2011; Arechavala-Lopez et al. 2011), taxonomic changes in
stomach contents (Demétrio et al. 2012; Fernandez-Jover & Sanchez-Jerez 2015), higher tissue
fat content and altered tissue fatty acid profiles that reflected the terrestrial-origin of lipids in
farm feed (Skog et al. 2003b, Fernandez-Jover et al. 2007, 2011, Arechavala-Lopez et al. 2011,
2015, Abaad et al. 2016). Arechavala-Lopez et al. (2015) also reported differing trace element
profiles in saithe near and far from salmon farms, while two studies reported altered taste and
other metrics of quality (Skog et al. 2003; Bogdanović et al. 2012).
Figure 2.3. Summary statistics for log response ratios (RR) for each variable in our meta-
analysis. All taxa are included. Boxes denote median, lower (25 %) and upper (75 %) quartiles,
whiskers denote 1.5x interquartile range. Data points are ‘jittered’ for clarity. Asterisk indicates
variables for which higher RR corresponds to poorer outcomes.
Contamination
Comparisons of contaminant levels in the tissues of farm-associated and non-associated fish
revealed mixed results. All three studies that tested for antimicrobial contamination in farm-
associated wild fish at farms where antimicrobials were in use found evidence of antimicrobial

26
residue in the majority of fish sampled, including oxytetracycline (0.2-1.3 µg g-1 muscle tissue:
Björklund et al. 1990), oxolinic acid (0.4-4.4 µg g-1 muscle tissue at two farms: Samuelsen et al.
1992) and flumequine (1.0-4.9 µg g-1 muscle tissue: Ervik et al. 1994). In each case, mean
concentrations for positive samples exceed the current European Union limits for these
substances in skin and muscle of finfish for human consumption: oxytetracycline: 0.1 µg g-1;
oxolinic acid: 0.1 µg g-1; flumequine: 0.6 µg g-1 (European Union 2010). It should be noted that
the development of new vaccines has allowed fish farmers in some areas (e.g. salmonid farms
in Norway and Scotland) to largely cease antimicrobial use despite rapid expansion of the
industry, but use remains high in other regions (Watts et al. 2017). It remains unclear whether
antimicrobial residue impacts fitness in farm-associated wild fish, whether through toxicity,
loss of gut microbiota or antimicrobial resistance in pathogens.
There have also been assessments of organohalogens and metals in the tissues of farm-
associated wild fish. One study reported significantly higher levels of organochlorines and
polybrominated diphenyl ethers in farm-associated fish relative to those from reference sites
(1.5x higher in cod, 1.2x higher in saithe: Bustnes et al. 2010). Another reported higher levels
(2.1x) of mercury in tissues of farm-associated rockfish (Sebastes spp.), potentially related to
an increase in trophic level near farms (DeBruyn et al. 2006). In the most comprehensive study
to date, Bustnes et al. (2011) measured concentrations of 30 elements in cod and saithe livers
from three regions in Norway. In saithe, Hg (2.0x), U (1.4x), Cr (1.9x) and Mn (1.6x)
concentrations were significantly higher in farm-associated fish, while Se, Zn, Cd, Cs and As
were higher at reference sites. In cod, U (1.4x), Al (1.5x) and Ba (1.9x) were higher in farm-
associated fish, while Se, Zn, Cd, Cs and As were higher at reference sites. While there is
evidence that some metals accumulate in sediments under fish farms, there is little evidence
so far that farm-associated wild fish are accumulating high concentrations in their tissues.
Infection rates
We discovered 22 studies that empirically investigated viral, bacterial or parasite transmission
between farmed and wild populations. In all cases, the authors concluded that the risk of
infection was either unchanged or elevated by interactions between farms and wild fish
populations. Of the 11 studies that quantified changes in infection levels with the presence of
active fish farms, all found higher levels of infection in farm-associated wild fish, with a mean
16x increase overall (RR = 2.1, t10 = 5.5, p = 0.0003). This large effect was primarily driven by

27
eight studies of sea louse infection loads on wild salmonids near salmon farms (3.5-73x
increase, RR = 2.5). One study reported higher infection densities of external parasites but
lower densities of internal parasites in farm-associated gadids, probably as a result of
consuming fewer infected wild fish and invertebrates in favour of commercial feed (Dempster
et al. 2011). Three studies provided molecular evidence for likely viral or bacterial transmission
between cultured and wild fish in the Mediterranean Sea (Zlotkin et al. 1998; Diamant et al.
2000; Colorni et al. 2002), and a molecular analysis of stomach contents revealed that wild cod
consumed escaped salmon stock infected with piscine reovirus (Glover et al. 2013). However,
molecular evidence did not always support the transmission hypothesis: Mladineo et al. (2009)
reported that monogenean and isopod parasites were not transmitted between wild and
farmed fish at one Mediterranean Sea farm.
Survivorship and fertility rates
Only two studies in our database estimated differential mortality rates in farm-associated
fauna. Kilambi et al. (1978) used age structure to infer a 21 % increase in survivorship of
largemouth bass following the establishment of cage culture in a freshwater lake, while in
contrast, Broyer et al. (2017) recorded 39 % higher mortality of ducklings at fish ponds. In sea
cage systems, elevated external parasitism rates (especially sea louse infections on salmonids)
may increase mortality in farm-associated fish, but to our knowledge, differential mortality
between farm and reference sites has not yet been empirically demonstrated. A further six
studies quantified culling of numerous birds at farms but did not compare mortality rates at
farms to those at reference sites. Two others reported dolphins being accidentally drowned in
anti-predator nets (Kemper & Gibbs 2001; Diaz López & Bernal-Shirai 2007), but again, did not
benchmark these against natural mortality rates. Several studies noted higher fishing effort
adjacent to sea cages, although we are only aware of two studies that quantified fishing effort
and catch rates (Akyol & Ertosluk 2010; Bacher & Gordoa 2016), and none assessed fishing
mortality rates among farm-associated fish.
Estimates of fertility (i.e. reproductive success) for wildlife at farms are similarly rare, but two
recent examples were returned by our search, both documenting probable ecological traps:
Kloskowski (2012) reported that fledging rates of grebes nesting on fish ponds stocked with +1
carp were only 37 % of those nesting on unstocked ponds, while Broyer et al. (2017) found

28
that high food availability was outweighed by high predation rates for ducks nesting on
stocked ponds (Table 1).
Conflict with aquaculture operations
Birds were usually predators of stock. 45/53 studies that documented interactions with birds
considered predation on stock to be the major habitat use, whether in cages, ponds, shellfish
beds or longlines. The most common avian predators were cormorants and herons. 24/38
studies of interactions with wild mammals considered predation to be the major habitat use,
in most cases by otters in ponds or sea cages (12 studies) or pinnipeds in sea cages (10
studies). Five studies reported herbivorous fishes inhabiting algae farms, but only one
presented clear evidence of fish consuming algal crops (Anyango et al. 2017). One study
reported predation of farmed mussels by wild fish (Segvic-Bubic et al. 2011), while three
reported snakes taking stock from hatchery ponds (Plummer & Goy 1984; Gregory & Nelson
1991; Nelson & Gregory 2000).
Of the 77 studies that reported predation on stock or damage to infrastructure, only 11
quantified stock losses as a proportion of potential production, with a mean loss of 15 %
(range 0-50 %). The lower end of that range was due to mammals taking only dead or
moribund fish from hatcheries (Pitt & Conover 1996), while the upper was due to predation by
cormorants in fish ponds (Barlow & Bock 1984). Other studies quantified consumption of stock
by individual predators without placing it in the context of potential production (e.g. Glahn et
al. 1999). In addition to predating stock, pinnipeds were reported to damage nets and cause
fish escapes (e.g. Güçlüsoy & Savas 2003; Sepúlveda & Oliva 2005).
DISCUSSION
Responses to aquaculture by wildlife vary greatly across taxonomic groups and culturing
systems, but our systematic review and meta-analysis reveals several key and well-supported
trends within taxonomic groups and culturing systems and identifies clear knowledge gaps to
inform future research.

29
Are wildlife attracted to aquaculture?
Fish
Multiple lines of evidence suggest that many fish species prefer aquaculture sites over natural
habitats, and on average, farms are associated with a much higher density and diversity of wild
fish. The few available tracking studies indicate that farm-associated wild fish tend to be either
residents or regular visitors (Otterå & Skilbrei 2014; Arechavala-Lopez et al. 2015a; Loiseau et
al. 2016; Tsuyuki & Umino 2017), with spilled feed and waste likely to be the major attractive
cues driving wild fish aggregations (Tuya et al. 2006; Ballester-Moltó et al. 2015; Bacher et al.
2015). Effects on fish abundance and diversity are also likely to depend on the functional group
being assessed, with most surveys of fish populations at farms and reference sites targeting
mobile generalist carnivores (either by design or through choice of sampling method).
Birds
Studies of bird abundance revealed highly variable responses to farms, but our meta-analysis
indicates that aquaculture sites are associated with higher bird species richness overall.
Numerous studies documented large bird populations at farms without comparing them to
natural waterways, making it difficult to draw conclusions about the influence of farms on the
spatial distribution of wildlife. Furthermore, little work has been done to assess responses at
the individual level (i.e. migration or site fidelity) that can assist in inferring habitat preferences
(Robertson & Hutto 2006), but it is likely that many bird species (especially herons, cormorants
and gulls) find the availability of prey at fish and crustacean farms highly attractive (Barlow &
Bock 1984; Stickley et al. 1992, 1995; Carss 1993; Glahn et al. 1999; Harrison 2009). Shellfish
farms also increase local abundance of generalist or molluscivorous bird functional groups
(Roycroft et al. 2004; Kirk et al. 2007), but others, such as invertivorous wading birds, may be
displaced by shellfish farm infrastructure or associated ecological changes (Kelly et al. 1996;
Godet et al. 2009; Broyer & Calenge 2010).
Mammals

30
Marine mammals (pinnipeds and dolphins) also showed highly variable responses to the
presence of aquaculture, ranging from resident nuisance animals (Pemberton & Shaughnessy
1993; Hume et al. 2002; Güçlüsoy & Savas 2003; Sepúlveda and Oliva 2005), to periodic visitors
(Diaz López 2012, 2017; Díaz López and Methion 2017), to active avoidance of farms
(Markowitz et al. 2004; Watson-Capps & Mann 2005; Pearson 2009; Becker et al. 2011). Otters
were common at freshwater fish ponds (Kloskowski 2005; Kortan et al. 2007) and estuarine
sea cages in Europe (Freitas et al. 2007; Sales-Luis et al. 2009), but our search did not reveal
any data on abundance or attraction to farms relative to natural waterways.
How does aquaculture affect fitness of wildlife?
Fish
Our meta-analysis indicated that farm-associated fish tend to be larger and heavier, a finding
that is consistent with either aggregation of adult fish or higher growth rates due to a trophic
subsidy. This larger average size, together with greater abundance overall, results in a very
high local biomass of farm-associated wild fish. Despite this, farm-associated fish had similar or
higher body condition metrics and rates of stomach fullness than fish from reference sites
(Fernandez-Jover et al. 2007a; Dempster et al. 2011), indicating that farm environments may
have a higher carrying capacity for wild fish than reference sites. However, any potential
positive effects—such as higher reproductive potential—may be opposed by orders of
magnitude higher infection loads near farms (especially sea lice on salmonids: Krkošek 2017)
and possible impacts of a dietary shift from marine-derived to terrestrially-derived fatty acids
in commercial aquaculture feed (Lavens et al. 1999; Mazorra et al. 2003; Salze et al. 2005;
Bogevik et al. 2012; Arechavala-Lopez et al. 2015b). Little is known about how the plurality of
environmental changes at farms combine to influence survival and reproduction in wild fish.
Mortality rates are difficult to measure directly, but Kilambi et al. (1978) compared age
structure and recapture rates in a lake before and after the commencement of cage farming
and inferred that survivorship had increased with farming.

31
In this study, we only assessed direct interactions between aquaculture activities and wildlife,
but indirect interactions also occur, and are likely to have a considerable bearing on outcomes
for fish populations in farming areas. Dietary shifts may occur indirectly via benthic nutrient
loading and subsequent ecological changes across multiple trophic levels (Brown et al. 1987;
Wu 1995; Yucel-Gier et al. 2007; White et al. 2017), and potential deleterious effects of direct
or indirect dietary shifts or other changes may be most apparent in eggs or offspring of farm-
associated fish (Salze et al. 2005; Barrett et al. 2018). Aggregations of large predators around
sea cages may also reduce survivorship of fishes that inhabit the same area (Güçlüsoy & Savas
2003). Fish that escape from farms can reduce fitness in native populations through disease
transmission (Arechavala-López et al. 2013; Glover et al. 2013), genetic mixing (Glover et al.
2017), and interference with spawning or competition with offspring and adults (Jensen et al.
2010; Sundt-Hansen et al. 2015).
Birds
In birds, the effects of farm proximity on fitness are even less clear; only in a few cases were
we able to extract usable data on direct or indirect fitness metrics. Numerous studies reported
birds taking stock from ponds and cages, but none in our database compared feeding rates to
those on natural waterways. Nonetheless, we expect food availability to be high provided that
birds are able to access suitable food items (e.g. feed, stock or wild prey co-occurring at
farms). However, predatory birds also experience high mortality from culling and anti-predator
net entanglements where such methods are employed, potentially causing fish farms to act as
ecological traps for birds if mortality rates outweigh any benefits of higher food availability
(Carss 1994; Belant et al. 2000; Blackwell et al. 2000; Bechard & Márquez-Reyes 2003; Quick et
al. 2004). Negative effects will be exacerbated if food availability is lower than advertised, for
example if piscivores are attracted to fish ponds but cannot access fish due to anti-predator
nets, or if stocking regimes lead to cohorts of fish that are too large to be consumed. This latter
scenario was observed by Kloskowski (2012), who reported that European carp farms were
acting as ecological traps for red-necked grebes, as farmed fish were too large for fledglings to
consume leading to starvation. Predation risk for clutches may also be elevated at farms:
Broyer et al. (2017) observed high densities of breeding pairs and high food availability, but
also high offspring mortality – a probable ecological trap. Conversely, tuna ranching in
Australia was associated with a population boom for silver gulls and appears to be a clear case

32
of fish farms functioning as a strong population source for wildlife – reproductive success for
the gulls was dramatically increased by the trophic subsidy obtained by exploiting farm feed
(Harrison 2009). Similarly, long term trends in wading bird populations closely tracked the
scale of crayfish aquaculture in the southern USA (Fleury & Sherry 1995).
Mammals
Effects of aquaculture on mortality and reproduction of aquatic mammals are little known, but
as with piscivorous birds, are probably determined by the trade-off between high food
availability and high risk from culling and entanglements. Cetaceans may benefit from easy
food when they visit farms (Diaz López 2017) and culling and entanglements are relatively rare
(Diaz López & Bernal-Shirai 2007; Callier et al. 2017). As a result, attraction to farms may be an
adaptive trait that results in increased fitness on balance, although we lack direct evidence for
this. In contrast to cetaceans, pinnipeds experience heavy mortality from culling (Güçlüsoy &
Savas 2003; Quick et al. 2004; Callier et al. 2017) and are more vulnerable to accidental
entanglement (Callier et al. 2017). High mortality rates are likely to outweigh any increase in
food availability for a long-lived, slow breeding animal, such that seals that are attracted to
farms may be vulnerable to ecological traps driven by culling at farms.
Conflict and potential mutualism between aquaculture and wildlife
Our meta-analysis revealed that the nature of interactions between wild fauna and
aquaculture were highly dependent on the taxon. Wild fish generally do not interact directly
with stock unless small enough to enter sea cages through the mesh (although in rare cases
wild fish may damage nets: Moe et al. 2007, Sanchez-Jerez et al. 2008). Of more concern is the
role that wild populations play as reservoirs for pathogens and parasites, facilitating
reinfection of farms (Uglem et al. 2014b). This is an inevitable risk of farming in open systems,
but research is underway to lower infection rates by minimising spatiotemporal overlap
between stock and zones of high infection risk (Samsing et al. 2016; Wright et al. 2017).
Together with post-infection treatments, such measures also minimise the role that farms play
as amplifiers of pathogen and parasite populations.

33
Table 2.1. Mean effects of aquaculture sites on wildlife populations. F:W = mean at farms /
mean at reference sites. RR = ln(F:W). Positive RR indicates metric is higher at aquaculture
sites. t-stat and p refer to one sample t-test comparing RR data to null expectation of RR = 0.
Taxa are omitted where no comparative data are available. Asterisk indicates variables for
which higher RR corresponds to poorer outcomes.
N F:W RR ± SE t-stat p
Abundance
Fish 44 72 1.65 ± 0.29 5.7 <0.0001
Birds 13 1.8 0.13 ± 0.31 0.40 0.70
Mammals 7 1.1 -0.68 ± 0.67 -1.0 0.35
Amphibians 1 0.31 -1.17 – –
All taxa 65 49 1.05 ± 0.24 4.3 <0.0001
Species richness
Fish 28 2.0 0.43 ± 0.11 3.9 0.0005
Birds 7 1.1 0.13 ± 0.04 3.3 0.02
Mammals 1 0.50 -0.69 – –
Amphibians 1 0.32 -1.15 – –
All taxa 37 1.7 0.30 ± 0.10 3.1 0.004
Size (length)
Fish 18 1.2 0.15 ± 0.03 4.6 0.0002
Size (weight)
Fish 12 1.7 0.40 ± 0.13 3.0 0.01
Condition metrics
Fish 14 1.3 0.17 ± 0.09 1.9 0.08
Birds 1 0.7 -0.31 – –
All taxa 15 1.2 0.14 ± 0.09 1.5 0.15
Stomach fullness
Fish 10 1.4 0.04 ± 0.30 0.13 0.90
Reptiles 1 1.9 0.66 – –
All taxa 11 1.5 0.10 ± 0.28 0.35 0.73
Infection level+
Fish 11 16 2.09 ± 0.38 5.5 0.0003
Fertility
Birds 2 0.60 -0.60 ± 0.40 -1.5 0.37
Mortality+
Fish 1 0.82 -0.20 – –
Bird 1 1.4 0.33 – –

34
Most studies returned by our search concluded that predation or damage by birds and
mammals is an ongoing problem for managers, but stock losses were rarely quantified (but see
some recent examples: Sun et al. 2004; Sepúlveda & Oliva 2005; Morrison & Vogel 2009; Dorr
et al. 2012) Where suitably habituated, pinnipeds have a propensity to become ‘nuisance
animals’, damaging nets (leading to fish escapes) and consuming or stressing stock (Kemper et
al. 2003; Quick et al. 2004; Sepúlveda & Oliva 2005). Such problems are difficult to solve.
Culling is undesirable as it increases environmental impacts and negatively affects public
perceptions of aquaculture. Relocation is expensive and often ineffective (Hume et al. 2002)
and scaring devices have a limited effective lifespan before animals are desensitised. Exclusion
using steel mesh appears to be the only viable option in some cases (Pemberton &
Shaughnessy 1993).
While there tends to be a focus on negative interactions between farms and wild fauna, wild
fish can provide ecosystem services to aquaculture operations by increasing animal welfare
and reducing local environmental impacts of farming. Invertivorous fish that are small enough
to gain access to sea cages (such as wrasse and lumpfish in Norwegian salmon farms) can act
as cleaner fish and significantly reduce parasite loads on stock. Cleaner fish are now being
deployed in large numbers for this purpose (Imsland et al. 2014; Skiftesvik et al. 2014). Wild
fish and invertebrates ameliorate and disperse benthic nutrient loads by consuming spilled
feed, faeces and dead stock (Vita et al. 2004; Felsing et al. 2005; Fernandez-Jover et al. 2007b).
However, resident populations of large predators at fish farms may impede this waste
amelioration service by scaring or consuming wild fish (Diaz López 2006), resulting in more
severe benthic impacts, but such predators also prey on escaped fish, potentially reducing the
risk of genetic introgression from farmed to wild fish populations (Glover et al. 2017).
For fish farming to continue to grow, farmers need to demonstrate environmental
sustainability and good animal welfare standards. Protecting wild fish aggregations to take
advantage of the ecosystem services they provide may be an important part of achieving these
goals. Continued development of non-lethal bird and pinniped exclusion methods will be a
necessary step.
Recommendations for future research on impacts of aquaculture on wildlife
Simply documenting behaviour of wildlife at farms or changes in wildlife abundance provides
little information on the effects of aquaculture on persistence of wildlife populations.

35
Aquaculture can have qualitatively distinct effects on wildlife that are superficially
indistinguishable in the absence of data on habitat selection decisions, movement or fitness.
For example, an elevated density of wildlife at a farm relative to a reference site may support
various contradictory hypotheses, including but not limited to: (a) high survivorship or fertility
causing the farm to function as a productive population source, with or without strong
attraction, and typically with density-dependent spillover to surrounding areas (Pulliam 1988),
or (b) strong attraction to the farm habitat but high mortality rates or low reproductive success
for residents. The latter scenario describes an attractive population sink (ecological trap) that
draws animals in from surrounding areas and causes deleterious population effects
disproportionate to its area (Hale et al. 2015). Our meta-analysis reveals that in most cases we
do not have sufficient data on fitness outcomes, either direct or indirect, and as a result cannot
distinguish between attractive or productive population effects, or their resultant positive or
negative effects on wild populations.
Conceptual frameworks have been developed to distinguish between these two (non-
mutually-exclusive) processes on artificial reefs and fish aggregation devices (Osenberg et al.
2002; Brickhill et al. 2005; Reubens et al. 2013), and may be applied to aquaculture sites.
Evidence for attraction without significant production of wild fauna at aquaculture sites may
include: (i) rarity of younger cohorts relative to older cohorts, (ii) population declines at
adjacent reference sites corresponding to increases at farms, (iii) high mortality or
reproductive failure rates at farms, or (iv) tracking, microchemistry, tissue fatty acid or stable
isotope analysis indicating recent immigration to farms. Conversely, evidence for high
individual fitness leading to productive wild populations at farms may include, depending on
the taxa: (i) successful breeding pairs residing at farms, (ii) high densities of larvae or juveniles,
(iii) increases in abundance at farms followed by increases at adjacent reference sites
consistent with density-dependent spillover, (iv) tracking, microchemistry, tissue fatty acid and
stable isotope analysis indicating that most individuals are not recent immigrants.
Importantly, the above criteria for separating attraction and production are most relevant
when compared to reference habitats (i.e. is residing at a farm a good decision for an
individual, or likely to lead to an ecological trap?). Only 91/204 studies included in our
database allowed us to infer changes in at least one variable in farm-associated wildlife by
making comparison to reference sites or timepoints. In many cases, changes in distribution or
health of wildlife were not central to the study, but in other cases, there was a lost opportunity
to understand more about these interactions. Where relevant, we recommend that studies of
abundance or fitness of wild fauna at farms should benchmark their findings against reference

36
sites or timepoints (Underwood 1994; Osenberg et al. 2002; Brickhill et al. 2005). Reference
sites should be appropriate for the hypotheses being tested. For example, our meta-analysis
revealed that inferred increases in population densities at sea cage fish farms vary by orders of
magnitude depending on whether the reference habitat is a nearby natural reef or featureless
open water. Accordingly, researchers should be clear in their reasons for selecting a given
reference habitat.
Most importantly, we have highlighted the paucity of data on mortality rates and reproductive
success in farm-associated fauna. Such data are central to our understanding of the
environmental impacts of aquaculture but can be difficult to obtain. Population-level metrics
can be effective in closed or semi-closed systems (Kilambi et al. 1978), and researchers have
long been capable of tracking mortality and breeding success in birds, including at aquaculture
sites (Kloskowski 2012; Broyer et al. 2017). Open systems with highly mobile taxa (such as wild
fish associated with sea cage aquaculture) present a greater challenge, but in coastal
environments, acoustic tags in conjunction with external tags can provide excellent data on
spatiotemporal movement and mortality rates in areas with differing levels of farm activity
(Olsen & Moland 2011; Olsen et al. 2012; Fernández-Chacón et al. 2015).
It is now well established that wild fish are typically more abundant at sea cage fish farms than
reference sites, and that such fish are likely to consume farm waste, experience nutritional
shifts and depending on the system, be exposed to elevated parasite loads. The challenge now
is to develop an equivalent state of knowledge for other wild taxa and aquaculture systems,
and to obtain more direct measures of the effects of farm-association on wildlife populations.
REFERENCES
Abaad M, Tuset VM, Montero D, Lombarte A, Otero-Ferrer JL and Haroun R (2016) Phenotypic
plasticity in wild marine fishes associated with fish-cage aquaculture. Hydrobiologia 765:
343–358.
Adámek Z, Kortan D, Lepič P and Andreji J (2003) Impacts of otter (Lutra lutra L) predation on
fishponds: A study of fish remains at ponds in the Czech Republic. Aquaculture
International 11: 389–396.
Akyol O and Ertosluk O (2010) Fishing near sea-cage farms along the coast of the Turkish
Aegean Sea. Journal of Applied Ichthyology 26: 11–15.
Anyango JO, Mlewa CM and Mwaluma J (2017) Abundance diversity and trophic status of wild

37
fish around seaweed farms in Kibuyuni, South Coast Kenya. International Journal of
Fisheries and Aquatic Studies 5: 440–446.
Arechavala-Lopez P, Izquierdo-Gomez D, Uglem I and Sanchez-Jerez P (2015a) Aggregations of
bluefish Pomatomus saltatrix (L) at Mediterranean coastal fish farms: seasonal presence
daily patterns and influence of farming activity. Environmental Biology of Fishes 98: 499–
510.
Arechavala-Lopez P, Sæther B-S, Marhuenda-Egea F, Sanchez-Jerez P and Uglem I (2015b)
Assessing the influence of salmon farming through total lipids fatty acids and trace
elements in the liver and muscle of wild saithe Pollachius virens. Marine and Coastal
Fisheries 7: 59–67.
Arechavala-Lopez P, Sanchez-Jerez P, Bayle-Sempere JT, Fernandez-Jover D, Martinez-Rubio L,
Lopez-Jimenez JA and Martinez-Lopez FJ (2011) Direct interaction between wild fish
aggregations at fish farms and fisheries activity at fishing grounds: a case study with
Boops boops. Aquaculture Research 42: 996–1010.
Arechavala-López P, Sánchez-Jerez P, Bayle-Sempere JT, Uglem I and Mladineo I (2013) Reared
fish farmed escapees and wild fish stocks—a triangle of pathogen transmission of
concern to Mediterranean aquaculture management. Aquaculture Environment
Interactions 3: 153–161.
Bacher K and Gordoa A (2016) Does marine fish farming affect local small-scale fishery
catches? A case study in the NW Mediterranean Sea. Aquaculture Research 47: 2444–
2454.
Bacher K, Gordoa A and Sagué O (2015) Feeding activity strongly affects the variability of wild
fish aggregations within fish farms: a sea bream farm as a case study. Aquaculture
Research 46: 552–564.
Ballester-Moltó M, Sanchez-Jerez P, Garcia-Garcia B and Aguado-Giménez F (2015) Husbandry
and environmental conditions explain temporal variability of wild fish assemblages
aggregated around a Mediterranean fish farm. Aquaculture Environment Interactions 7:
193–203.
Bancroft BA, Baker NJ and Blaustein AR (2007) Effects of UVB radiation on marine and
freshwater organisms: a synthesis through meta-analysis. Ecology Letters 10: 332–345.
Barlow CG and Bock K (1984) Predation of fish in farm dams by cormorants Phalacrocorax spp.
Australian Wildlife Research 11: 559–566
Barrett LT, Swearer SE, Harboe T, Karlsen Ø, Meier S and Dempster T (2018) Limited evidence
for differential reproductive fitness of wild Atlantic cod in areas of high and low salmon

38
farming density. Aquaculture Environment Interactions doi: 10.3354/aei00275
Barton K (2016) MuMIn: multi-model inference R package version 1156. URL: https://cran.r-
project.org/package=MuMIn
Bechard MJ and Márquez-Reyes C (2003) Mortality of wintering ospreys and other birds at
aquaculture facilities in Colombia. Journal of Raptor Research 37: 292–298.
Becker BH, Press DT and Allen SG (2011) Evidence for long-term spatial displacement of
breeding and pupping harbour seals by shellfish aquaculture over three decades. Aquatic
Conservation: Marine and Freshwater Ecosystems 21: 247–260.
Belant JL, Tyson LA and Mastrangelo PA (2000) Effects of lethal control at aquaculture facilities
on populations of piscivorous birds. Wildlife Society Bulletin 28: 379–384.
Berg V, Kraugerud M, Nourizadeh-Lillabadi R et al. (2016) Endocrine effects of real-life
mixtures of persistent organic pollutants (POP) in experimental models and wild fish.
Journal of Toxicology and Environmental Health Part A 79: 538–548.
Björklund H, Bondestam J and Bylund G (1990) Residues of oxytetracycline in wild fish and
sediments from fish farms. Aquaculture 86: 359–367.
Blackwell BF, Dolbeer RA and Tyson LA (2000) Lethal control of piscivorous birds at
aquaculture facilities in the northeast United States: effects on populations. North
American Journal of Aquaculture 62: 300–307.
Bogdanović T, Šimat V, Frka-Roić A and Marković K (2012) Development and application of
quality index method scheme in a shelf-life study of wild and fish farm affected bogue
(Boops boops L). Journal of Food Science 77: S99–S106.
Bogevik AS, Natário S, Karlsen Ø, Thorsen A, Hamre K, Rosenlund G and Norberg B (2012) The
effect of dietary lipid content and stress on egg quality in farmed Atlantic cod Gadus
morhua. Journal of Fish Biology 81: 1391–405.
Boyd CE and Massaut L (1999) Risks associated with the use of chemicals in pond aquaculture.
Aquacultural Engineering 20: 113–132.
Brickhill MJ, Lee SY and Connolly RM (2005) Fishes associated with artificial reefs: attributing
changes to attraction or production using novel approaches. Journal of Fish Biology 67:
53–71.
Brown JR, Gowen RJ and McLusky DS (1987) The effect of salmon farming on the benthos of a
Scottish sea loch. Journal of Experimental Marine Biology and Ecology 109: 39–51.
Broyer J and Calenge C (2010) Influence of fish-farming management on duck breeding in
French fish pond systems. Hydrobiologia 637: 173–185.
Broyer J, Chavas G and Chazal R (2017) The effects of cessation of fish farming on duck

39
breeding in French fishpond systems. Hydrobiologia 788: 47–53.
Burnham KP and Anderson DR (2002) Model selection and multimodel inference: a practical
information-theoretic approach, 2nd edn. Springer, New York, NY.
Burridge L, Weis JS, Cabello F, Pizarro J and Bostick K (2010) Chemical use in salmon
aquaculture: a review of current practices and possible environmental effects.
Aquaculture 306: 7–23.
Bustnes JO, Lie E, Herzke D, Dempster T, Bjørn PA, Nygård T and Uglem I (2010) Salmon farms
as a source of organohalogenated contaminants in wild fish. Environmental Science &
Technology 44: 8736–43.
Bustnes JO, Nygard T, Dempster T, Ciesielski T, Jenssen BM, Bjorn PA and Uglem I (2011) Do
salmon farms increase the concentrations of mercury and other elements in wild fish?
Journal of Environmental Monitoring 13: 1687–1694.
Callier MD, Byron CJ, Bengtson DA et al. (2017) Attraction and repulsion of mobile wild
organisms to finfish and shellfish aquaculture: a review. Reviews in Aquaculture doi:
10.1111/raq.12208
Carss DN (1993) Grey heron Ardea cinerea L predation at cage fish farms in Argyll western
Scotland. Aquaculture and Fisheries Management 24: 29–45.
Carss DN (1994) Killing of piscivorous birds at Scottish fin fish farms, 1984-87. Biological
Conservation 68: 181–188.
Colorni A, Diamant A, Eldar A, Kvitt H and Zlotkin A (2002) Streptococcus iniae infections in Red
Sea cage-cultured and wild fishes. Diseases of Aquatic Organisms 49: 165–170.
Costello MJ (2009) How sea lice from salmon farms may cause wild salmonid declines in
Europe and North America and be a threat to fishes elsewhere. Proceedings of the Royal
Society B 276: 3385–3394.
Crump KL and Trudeau VL (2009) Mercury-induced reproductive impairment in fish.
Environmental Toxicology and Chemistry 28: 895.
Dealteris JT, Kilpatrick BD and Rheault RB (2004) A comparative evaluation of the habitat value
of shellfish aquaculture gear, submerged aquatic vegetation and a non-vegetated seabed.
Journal of Shellfish Research 23: 867–874.
DeBruyn AMH, Trudel M, Eyding N et al. (2006) Ecosystemic effects of salmon farming increase
mercury contamination in wild fish. Environmental Science & Technology 40: 3489–3493.
Demétrio JA, Gomes LC, Latini JD and Agostinho AA (2012) Influence of net cage farming on
the diet of associated wild fish in a Neotropical reservoir. Aquaculture 330: 172–178.
Dempster T, Sanchez-Jerez P, Bayle-Sempere JT and Giménez-Casalduero F (2002) Attraction of

40
wild fish to sea-cage fish farms in the south-western Mediterranean Sea: spatial and
short-term temporal variability. Marine Ecology Progress Series 242: 237–252.
Dempster T, Sanchez-Jerez P, Bayle-Sempere JT and Kingsford M (2004) Extensive aggregations
of wild fish at coastal sea-cage fish farms. Hydrobiologia 525: 245–248.
Dempster T, Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere JT, Nilsen R, Bjørn PA and
Uglem I (2011) Proxy measures of fitness suggest coastal fish farms can act as population
sources and not ecological traps for wild gadoid fish. PLOS ONE 6: e15646.
Dempster T, Uglem I, Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere J and Nilsen R (2009)
Coastal salmon farms attract large and persistent aggregations of wild fish: an ecosystem
effect. Marine Ecology Progress Series 385: 1–14.
Diamant A, Banet A, Ucko M, Colorni A, Knibb W and Kvitt H (2000) Mycobacteriosis in wild
rabbitfish Siganus rivulatus associated with cage farming in the Gulf of Eilat, Red Sea.
Diseases of Aquatic Organisms 39: 211–219.
Diaz López B (2006) Bottlenose dolphin (Tursiops truncatus) predation on a marine fin fish
farm: some underwater observations. Aquatic Mammals 32: 305–310.
Diaz López B (2012) Bottlenose dolphins and aquaculture: interaction and site fidelity on the
north-eastern coast of Sardinia (Italy). Marine Biology 159: 2161–2172.
Diaz López B (2017) Temporal variability in predator presence around a fin fish farm in the
Northwestern Mediterranean Sea. Marine Ecology 38: e12378.
Diaz López B and Bernal-Shirai JA (2007) Bottlenose dolphin (Tursiops truncatus) presence and
incidental capture in a marine fish farm on the north-eastern coast of Sardinia (Italy).
Journal of the Marine Biological Association of the UK 87: 113–117.
Díaz López B and Methion S (2017) The impact of shellfish farming on common bottlenose
dolphins’ use of habitat. Marine Biology 164: 83.
Dorr BS, Burger LW, Barras SC and Godwin KC (2012) Economic impact of double-crested
cormorant Phalacrocorax auritus depredation on channel catfish Ictalurus punctatus
aquaculture in Mississippi, USA. Journal of the World Aquaculture Society 43: 502–513.
Dumont CP, Gaymer CF and Thiel M (2011) Predation contributes to invasion resistance of
benthic communities against the non-indigenous tunicate Ciona intestinalis. Biological
Invasions 13: 2023–2034
Ervik A, Thorsen B, Eriksen V, Lunestad BT and Samuelsen OB (1994) Impact of administering
antibacterial agents on wild fish and blue mussels Mytilus edulis in the vicinity of fish
farms. Diseases of Aquatic Organisms 18: 45–51.
European Union (2010) Commission Regulation (EU) No 37/2010 of 22 December 2009 on

41
pharmacologically active substances and their classification regarding maximum residue
limits in foodstuffs of animal origin Brussels European Union. URL:
https://eceuropaeu/food/plant/pesticides/max_residue_levels_en
FAO Fisheries and Aquaculture (2015) FAO Yearbook - Fishery and Aquaculture Statistics
(2014).
FAO Fisheries and Aquaculture (2017) Fishery and Aquaculture Statistics - Global aquaculture
production 1950-2015 (FishstatJ). URL:
http://www.fao.org/fishery/statistics/software/fishstatj/en
FAO Fisheries and Aquaculture (2018) Cultured aquatic species fact sheets. URL:
http://www.fao.org/fishery/culturedspecies/search/en
Felsing M, Glencross B and Telfer T (2005) Preliminary study on the effects of exclusion of wild
fauna from aquaculture cages in a shallow marine environment. Aquaculture 243: 159–
174.
Fernández-Chacón A, Moland E, Espeland SH and Olsen EM (2015) Demographic effects of full
vs partial protection from harvesting: inference from an empirical before-after control-
impact study on Atlantic cod. Journal of Applied Ecology 52: 1206–1215.
Fernandez-Jover D, Jimenez JA, Sanchez-Jerez P, Bayle-Sempere JT, Giménez-Casalduero F,
Lopez FJM and Dempster T (2007a) Changes in body condition and fatty acid composition
of wild Mediterranean horse mackerel (Trachurus mediterraneus Steindachner 1868)
associated to sea cage fish farms. Marine Environmental Research 63: 1–18.
Fernandez-Jover D, Martinez-Rubio L, Sanchez-Jerez P et al. (2011) Waste feed from coastal
fish farms: a trophic subsidy with compositional side-effects for wild gadoids. Estuarine
Coastal and Shelf Science 91: 559–568.
Fernandez-Jover D and Sanchez-Jerez P (2015) Comparison of diet and otolith growth of
juvenile wild fish communities at fish farms and natural habitats. ICES Journal of Marine
Science 72: 916–929.
Fernandez-Jover D, Sanchez-Jerez P, Bayle-Sempere J, Carratala A and Leon VM (2007b)
Addition of dissolved nitrogen and dissolved organic carbon from wild fish faeces and
food around Mediterranean fish farms: Implications for waste-dispersal models. Journal
of Experimental Marine Biology and Ecology 340: 160–168.
Fleury BE and Sherry TW (1995) Long-term population trends of colonial wading birds in the
southern United States: the impact of crayfish aquaculture on Louisiana populations. Auk
112: 613–632.
Freitas D, Gomes J, Luis TS et al. (2007) Otters and fish farms in the Sado estuary: ecological

42
and socio-economic basis of a conflict. Hydrobiologia 587: 51–62.
Gamfeldt L, Lefcheck JS, Byrnes JEK, Cardinale BJ, Duffy JE and Griffin JN (2015) Marine
biodiversity and ecosystem functioning: what’s known and what’s next? Oikos 124: 252–
265.
Glahn JF, Tomsa T and Preusser KJ (1999) Impact of great blue heron predation at trout-rearing
facilities in the northeastern United States. North American Journal of Aquaculture 61:
349–354.
Glover KA, Solberg MF, McGinnity P et al. (2017) Half a century of genetic interaction between
farmed and wild Atlantic salmon: status of knowledge and unanswered questions. Fish
and Fisheries doi: 10.1111/faf.12214
Glover KA, Sørvik AGE, Karlsbakk E, Zhang Z and Skaala Ø (2013) Molecular genetic analysis of
stomach contents reveals wild Atlantic cod feeding on piscine reovirus (PRV) infected
Atlantic salmon originating from a commercial fish farm. PLOS ONE 8: e60924.
Godet L, Toupoint N, Fournier J, Le Mao P, Retiere C and Olivier F (2009) Clam farmers and
oystercatchers: effects of the degradation of Lanice conchilega beds by shellfish farming
on the spatial distribution of shorebirds. Marine Pollution Bulletin 58: 589–595.
Gonzalez-Silvera D, Guardiola FA, Cordero H, Cuesta A, Esteban MA, Martínez-López FJ and
López-Jiménez JA (2017) The short-term effects of farmed fish food consumed by wild
fish congregating outside the farms. Marine Pollution Bulletin 114: 689–698.
Gregory PT and Nelson KJ (1991) Predation on fish and intersite variation in the diet of
common garter snakes Thamnophis sirtalis on Vancouver Island. Canadian Journal of
Zoology 69: 988–994.
Güçlüsoy H and Savas Y (2003) Interaction between monk seals Monachus monachus
(Hermann 1779) and marine fish farms in the Turkish Aegean and management of the
problem. Aquaculture Research 34: 777–783.
Hale R and Swearer SE (2016) Ecological traps: current evidence and future directions.
Proceedings of the Royal Society B 283: 494–499.
Hale R, Treml EA and Swearer SE (2015) Evaluating the metapopulation consequences of
ecological traps. Proceedings of the Royal Society B 282: 20142930
Harrison SJ (2009) Interactions between Silver Gulls (Larus novaehollandiae) and Southern
Bluefin Tuna (Thunnus maccoyii) aquaculture in the Port Lincoln area. PhD thesis. Flinders
University, Australia.
Hayward CJ, Svane I, Lachimpadi SK, Itoh N, Bott NJ and Nowak BF (2011) Sea lice infections of
wild fishes near ranched southern bluefin tuna (Thunnus maccoyii) in South Australia.

43
Aquaculture 320: 178–182.
Hedges LV, Gurevitch J and Curtis PS (1999) The meta-analysis of response ratios in
experimental ecology. Ecology 80: 1150–1156.
Hehre EJ and Meeuwig JJ (2016) A global analysis of the relationship between farmed seaweed
production and herbivorous fish catch. PLOS ONE 11: e0148250.
Herbeck LS, Unger D, Wu Y and Jennerjahn TC (2013) Effluent nutrient and organic matter
export from shrimp and fish ponds causing eutrophication in coastal and back-reef
waters of NE Hainan, tropical China. Continental Shelf Research 57: 92–104.
Hume F, Pemberton D, Gales R, Brothers N and Greenwood M (2002) Trapping and relocating
seals from salmonid fish farms in Tasmania 1990-2000: was it a success? Papers and
Proceedings of the Royal Society of Tasmania 136: 1–6.
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Foss A, Vikingstad E and Elvegård TA (2014)
The use of lumpfish (Cyclopterus lumpus L) to control sea lice (Lepeophtheirus salmonis
Krøyer) infestations in intensively farmed Atlantic salmon (Salmo salar L). Aquaculture
424–425: 18–23.
Inglis GJ and Gust N (2003) Potential indirect effects of shellfish culture on the reproductive
success of benthic predators. Journal of Applied Ecology 40: 1077–1089.
Jensen Ø, Dempster T, Thorstad EB, Uglem I and Fredheim A (2010) Escapes of fishes from
Norwegian sea-cage aquaculture: causes consequences and prevention. Aquaculture
Environment Interactions 1: 71–83.
Johnston TA, Keir M and Power M (2010) Response of native and naturalized fish to salmonid
cage culture farms in northern Lake Huron, Canada. Transactions of the American
Fisheries Society 139: 660–670.
Kelly JP, Evens JG, Stallcup RW and Wimpfheimer D (1996) Effects of aquaculture on habitat
use by wintering shorebirds in Tomales Bay, California. California Fish and Game 82: 160–
174.
Kemper C, Pemberton D, Cawthorn M et al. (2003) Aquaculture and marine mammals: co-
existence or conflict? In: Gales N, Hindell M and Kirkwood R (ed.) Marine Mammals:
Fisheries Tourism and Management Issues, pp. 208–225. CSIRO Publishing, Collingwood.
Kemper CM and Gibbs SE (2001) Dolphin interactions with tuna feedlots at Port Lincoln, South
Australia and recommendations for minimising entanglements. Journal of Cetacean
Research and Management 3: 283–292.
Kilambi RV, Adams JC and Wickizer WA (1978) Effects of cage culture on growth abundance
and survival of resident largemouth bass (Micropterus salmoides). Journal of the Fisheries

44
Research Board of Canada 35: 157–160.
Kirk M, Esler D and Boyd WS (2007) Morphology and density of mussels on natural and
aquaculture structure habitats: implications for sea duck predators. Marine Ecology
Progress Series 346: 179–187.
Kloskowski J (2010) Fish farms as amphibian habitats: factors affecting amphibian species
richness and community structure at carp ponds in Poland. Environmental Conservation
37: 187–194.
Kloskowski J (2012) Fish stocking creates an ecological trap for an avian predator via effects on
prey availability. Oikos 121: 1567–1576.
Kloskowski J (2005) Otter Lutra lutra damage at farmed fisheries in southeastern Poland II:
exploitation of common carp Cyprinus carpio. Wildlife Biology 11: 257–261.
Kloskowski J, Kaczanowska E, Krogulec J and Grela P (2017) Hematological indicators of habitat
quality: erythrocyte parameters reflect greater parental effort of red-necked grebes
under ecological trap conditions. The Condor 119: 239–250.
Kortan D, Adámek Z and Poláková S (2007) Winter predation by otter Lutra lutra on carp pond
systems in South Bohemia (Czech Republic). Folia Zoologica 56: 416.
Krkošek M (2017) Population biology of infectious diseases shared by wild and farmed fish.
Canadian Journal of Fisheries and Aquatic Sciences 74: 620–628.
Krkosek M, Lewis MA, Morton A, Frazer LN and Volpe JP (2006) Epizootics of wild fish induced
by farm fish. Proceedings of the National Academy of Sciences of the USA 103: 15506–
15510.
Krkosek M, Lewis MA and Volpe JP (2005) Transmission dynamics of parasitic sea lice from
farm to wild salmon. Proceedings of the Royal Society B 272: 689–96.
Kroeker KJ, Kordas RL, Crim RN and Singh GG (2010) Meta-analysis reveals negative yet
variable effects of ocean acidification on marine organisms. Ecology Letters 13: 1419–
1434.
Lafferty KD, Harvell CD, Conrad JM et al. (2015) Infectious diseases affect marine fisheries and
aquaculture economics. Annual Review of Marine Science 7: 471–496.
Lavens P, Lebegue E, Jaunet H, Brunel A, Dhert P and Sorgeloos P (1999) Effect of dietary
essential fatty acids and vitamins on egg quality in turbot broodstocks. Aquaculture
International 7: 225–240.
Loiseau N, Kiszka JJ, Bouveroux T, Heithaus MR, Soria M and Chabanet P (2016) Using an
unbaited stationary video system to investigate the behaviour and interactions of bull
sharks Carcharhinus leucas under an aquaculture farm. African Journal of Marine Science

45
38: 73–79.
Markowitz TM, Harlin AD, Wursig B and Mcfadden CJ (2004) Dusky dolphin foraging habitat:
overlap with aquaculture in New Zealand. Aquatic Conservation: Marine and Freshwater
Ecosystems 14: 133–149.
Mazorra C, Bruce M, Bell JG et al. (2003) Dietary lipid enhancement of broodstock
reproductive performance and egg and larval quality in Atlantic halibut (Hippoglossus
hippoglossus). Aquaculture 227: 21–33.
McKindsey CW, Archambault P, Callier MD and Olivier F (2011) Influence of suspended and off-
bottom mussel culture on the sea bottom and benthic habitats: a review. Canadian
Journal of Zoology 89: 622–646.
Mladineo I, Segvic T and Grubisic L (2009) Molecular evidence for the lack of transmission of
the monogenean Sparicotyle chrysophrii (Monogenea, Polyopisthocotylea) and isopod
Ceratothoa oestroides (Crustacea, Cymothoidae) between wild bogue (Boops boops) and
cage-reared sea bream (Sparus aurata). Aquaculture 295: 160–167.
Moe H, Dempster T, Sunde LM, Winther U and Fredheim A (2007) Technological solutions and
operational measures to prevent escapes of Atlantic cod (Gadus morhua) from sea cages.
Aquaculture Research 38: 91–99.
Morrison SS and Vogel P (2009) Aquaculture ponds a Jamaican study: The impact of birds on
fish production. African Journal of Agricultural Research 4: 1447–1454.
Nelson KJ and Gregory PT (2000) Activity patterns of garter snakes Thamnophis sirtalis in
relation to weather conditions at a fish hatchery on Vancouver Island, British Columbia.
Journal of Herpetology 34: 32.
Olsen EM, Heupel MR, Simpfendorfer CA and Moland E (2012) Harvest selection on Atlantic
cod behavioral traits: implications for spatial management. Ecology and Evolution 2:
1549–1562.
Olsen EM and Moland E (2011) Fitness landscape of Atlantic cod shaped by harvest selection
and natural selection. Evolutionary Ecology 25: 695–710.
Osenberg C, St Mary CM, Wilson JA and Lindberg WJ (2002) A quantitative framework to
evaluate the attraction-production controversy. ICES Journal of Marine Science 59: S214–
S221.
Otterå H and Skilbrei OT (2014) Possible influence of salmon farming on long-term resident
behaviour of wild saithe (Pollachius virens L). ICES Journal of Marine Science 71: 2484–
2493.
Pearson HC (2009) Influences on dusky dolphin (Lagenorhynchus obscurus) fission-fusion

46
dynamics in Admiralty Bay, New Zealand. Behavioral Ecology and Sociobiology 63: 1437–
1446.
Pemberton D and Shaughnessy PD (1993) Interaction between seals and marine fish-farms in
Tasmania and management of the problem. Aquatic Conservation: Marine and
Freshwater Ecosystems 3: 149–158.
Piroddi C, Bearzi G and Christensen V (2011) Marine open cage aquaculture in the eastern
Mediterranean Sea: a new trophic resource for bottlenose dolphins. Marine Ecology
Progress Series 440: 255–266.
Pitt WC and Conover MR (1996) Predation at Intermountain West fish hatcheries. The Journal
of Wildlife Management 60: 616–624.
Plummer M V and Goy JM (1984) Ontogenetic dietary shift of water snakes (Nerodia
rhombifera) in a fish hatchery. Copeia 1984: 550.
Powers M, Peterson C, Summerson H and Powers S (2007) Macroalgal growth on bivalve
aquaculture netting enhances nursery habitat for mobile invertebrates and juvenile
fishes. Marine Ecology Progress Series 339: 109–122.
Pulliam HR (1988) Sources sinks and population regulation. The American Naturalist 132: 652–
661.
Quick NJ, Middlemas SJ and Armstrong JD (2004) A survey of antipredator controls at marine
salmon farms in Scotland. Aquaculture 230: 169–180.
R Core Team (2017) R: A language and environment for statistical computing. URL:
http://www.r-project.org/
Reubens JT, Vandendriessche S, Zenner AN, Degraer S and Vincx M (2013) Offshore wind farms
as productive sites or ecological traps for gadoid fishes? Impact on growth condition
index and diet composition. Marine Environmental Research 90: 66-74.
Robertson BA and Hutto RL (2006) A framework for understanding ecological traps and an
evaluation of existing evidence. Ecology 87: 1075–1085.
Roycroft D, Kelly TC and Lewis LJ (2004) Birds seals and the suspension culture of mussels in
Bantry Bay a non-seaduck area in Southwest Ireland. Estuarine Coastal and Shelf Science
61: 703–712.
Sales-Luis T, Freitas D and Santos-Reis M (2009) Key landscape factors for Eurasian otter Lutra
lutra visiting rates and fish loss in estuarine fish farms. European Journal of Wildlife
Research 55: 345–355.
Salze G, Tocher DR, Roy WJ and Robertson DA (2005) Egg quality determinants in cod (Gadus
morhua L): egg performance and lipids in eggs from farmed and wild broodstock.

47
Aquaculture Research 36: 1488–1499.
Samsing F, Johnsen I, Stien LH, Oppedal F, Albretsen J, Asplin L and Dempster T (2016)
Predicting the effectiveness of depth-based technologies to prevent salmon lice infection
using a dispersal model. Preventive Veterinary Medicine 129: 48–57.
Samuelsen OB, Lunestad BT, Husevag B, Holleland T and Ervik A (1992) Residues of oxolinic
acid in wild fauna following medication in fish farms. Diseases of Aquatic Organisms 12:
111–119.
Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere J, Valle C, Dempster T, Tuya F and Juanes F
(2008) Interactions between bluefish Pomatomus saltatrix (L) and coastal sea-cage farms
in the Mediterranean Sea. Aquaculture 282: 61–67.
Sanchez-Jerez P, Fernandez-Jover D, Uglem I et al. (2011) Coastal fish farms as fish aggregation
devices (FADs). In: Bortone SA, Brandini FP, Fabi G and Otake S (ed.) Artificial Reefs in
Fishery Management, pp. 187–208. Taylor & Francis Group, Florida.
Segvic-Bubic T, Grubisic L, Karaman N, Ticina V, Jelavic KM and Katavic I (2011) Damages on
mussel farms potentially caused by fish predation – self service on the ropes?
Aquaculture 319: 497–504.
Sepúlveda M, Newsome SD, Pavez G, Oliva D, Costa DP and Hueckstaedt LA (2015) Using
satellite tracking and isotopic information to characterize the impact of South American
sea lions on salmonid aquaculture in southern Chile. PLOS ONE 10: e0134926–e0134926.
Sepúlveda M and Oliva D (2005) Interactions between South American sea lions Otaria
flavescens (Shaw) and salmon farms in southern Chile. Aquaculture Research 36: 1062–
1068.
Skiftesvik AB, Blom G, Agnalt A-L et al. (2014) Wrasse (Labridae) as cleaner fish in salmonid
aquaculture – the Hardangerfjord as a case study. Marine Biology Research 10: 289–300.
Skog T-E, Hylland K, Torstensen BE and Berntssen MHG (2003) Salmon farming affects the fatty
acid composition and taste of wild saithe Pollachius virens L. Aquaculture Research 34:
999–1007.
Stickley AR, Glahn JF, King JO and King DT (1995) Impact of great blue heron depredations on
channel catfish farms. Journal of the World Aquaculture Society 26: 194–199.
Stickley AR, Warrick GL and Glahn JF (1992) Impact of double‐crested cormorant depredations
on channel catfish farms. Journal of the World Aquaculture Society 23: 192–198.
Sun YH, Wu HJ and Wang Y (2004) Tawny fish-owl predation at fish farms in Taiwan. Journal of
Raptor Research 38: 326–333.
Sundt-Hansen L, Huisman J, Skoglund H and Hindar K (2015) Farmed Atlantic salmon Salmo

48
salar L parr may reduce early survival of wild fish. Journal of Fish Biology 86: 1699–1712.
Tallman JC and Forrester GE (2007) Oyster grow-out cages function as artificial reefs for
temperate fishes. Transactions of the American Fisheries Society 136: 790–799.
Taranger GL, Karlsen Ø, Bannister RJ et al. (2015) Risk assessment of the environmental impact
of Norwegian Atlantic salmon farming. ICES Journal of Marine Science 72: 997–1021.
Tsuyuki A and Umino T (2017) Spatial movement of black sea bream Acanthopagrus schlegelii
around the oyster farming area in Hiroshima Bay, Japan. Fisheries Science 83: 235–244.
Tuya F, Sanchez-Jerez P, Dempster T, Boyra A and Haroun RJ (2006) Changes in demersal wild
fish aggregations beneath a sea-cage fish farm after the cessation of farming. Journal of
Fish Biology 69: 682–697.
Uglem I, Dempster T, Bjørn PA, Sanchez-Jerez P and Økland F (2009) High connectivity of
salmon farms revealed by aggregation residence and repeated movements of wild fish
among farms. Marine Ecology Progress Series 384: 251–260.
Uglem I, Karlsen Ø, Sanchez-Jerez P and Sæther B (2014) Impacts of wild fishes attracted to
open-cage salmonid farms in Norway. Aquaculture Environment Interactions 6: 91–103
Underwood AJ (1994) On Beyond BACI: sampling designs that might reliably detect
environmental disturbances. Ecological Applications 4: 3–15.
United Nations Development Programme (2017) International Human Development Indicators.
Available at: http://hdrundporg/en/countries [Accessed October 30 2017].
Valdemarsen T, Hansen PK, Ervik A and Bannister RJ (2015) Impact of deep-water fish farms on
benthic macrofauna communities under different hydrodynamic conditions. Marine
Pollution Bulletin 101: 776–783.
Vita R, Marin A, Madrid JA, Jimenez-Brinquis B, Cesar A and Marin-Guirao L (2004) Effects of
wild fishes on waste exportation from a Mediterranean fish farm. Marine Ecology
Progress Series 277: 253–261.
Watson-Capps JJ and Mann J (2005) The effects of aquaculture on bottlenose dolphin (Tursiops
sp) ranging in Shark Bay, Western Australia. Biological Conservation 124: 519–526.
Watts JEM, Schreier HJ, Lanska L and Hale MS (2017) The rising tide of antimicrobial resistance
in aquaculture: sources sinks and solutions Marine Drugs 15: 158.
White CA, Bannister RJ, Dworjanyn SA, Husa V, Nichols PD, Kutti T and Dempster T (2017)
Consumption of aquaculture waste affects the fatty acid metabolism of a benthic
invertebrate. Science of The Total Environment 586: 1170–1181.
Wright DW, Stien LH, Dempster T, Vågseth T, Nola V, Fosseidengen J-E and Oppedal F (2017)
“Snorkel” lice barrier technology reduced two co-occurring parasites the salmon louse

49
(Lepeophtheirus salmonis) and the amoebic gill disease causing agent (Neoparamoeba
perurans) in commercial salmon sea-cages. Preventive Veterinary Medicine 140: 97–105.
Wu RSS (1995) The environmental impact of marine fish culture: towards a sustainable future.
Marine Pollution Bulletin 31: 159–166.
Yucel-Gier G, Kucuksezgin F and Kocak F (2007) Effects of fish farming on nutrients and benthic
community structure in the Eastern Aegean (Turkey). Aquaculture Research 38: 256–267.
Zeileis A, Kleiber C and Jackman S (2008) Regression models for count data in R. Journal of
Statistical Software 27. doi: 10.18637/jss.v027.i08
Zlotkin A, Hershko H and Eldar A (1998) Possible transmission of Streptococcus iniae from wild
fish to cultured marine fish. Applied and Environmental Microbiology 64: 4065–4067.

50
CHAPTER THREE: LIMITED EVIDENCE FOR DIFFERENTIAL REPRODUCTIVE FITNESS OF WILD
ATLANTIC COD IN AREAS OF HIGH AND LOW SALMON FARMING DENSITY
ABSTRACT
Sea cage fish aquaculture attracts large aggregations of wild fish that feed on farm waste. Fish
that associate closely with farms undergo physiological changes, and captive feeding trials
indicate possible negative effects on reproductive fitness. However, little is known about the
significance of this phenomenon for reproduction in wild fish over larger spatial scales. To
assess if coastal areas with intensive aquaculture impact the fitness of wild fish, we collected
Atlantic cod (Gadus morhua) from two areas of high and low salmon farming density (HFD and
LFD respectively) in south-western Norway, a region that hosts the world’s largest coastal fish
aquaculture industry. We conducted a captive spawning trial and compared a range of
reproductive fitness metrics. Two fatty acids that occur at high levels in commercial feed, oleic
and lineoleic acid, were strongly correlated in the ovaries of HFD fish, but a comparison of lipid
profiles between HFD and LFD fish showed no differences in total lipids or essential fatty acids.
Although HFD fish were slightly larger than LFD fish and had similar body condition, the volume
of eggs produced did not differ, indicating relatively lower reproductive investment by HFD
fish. HFD eggs were 5 % smaller than LFD eggs, which did not lead to differential hatching
success but may have contributed to HFD larvae being 8 % smaller than their LFD counterparts
at 40 days post spawning. The potential for cumulative effects of smaller eggs and larvae on
fitness at later life stages warrants further investigation.
INTRODUCTION
Intensive culture of fish within sea cages leads to considerable benthic nutrient loads via
spilled feed and waste, providing a trophic subsidy that attracts large and persistent
aggregations of ‘farm-associated’ wild fish (Dempster et al. 2002, 2009, 2010). Individuals may
reside in the vicinity of farms for months to years (Uglem et al. 2009, Otterå & Skilbrei 2014),
and during this time are exposed to a variety of environmental changes including elevated
infection risk from parasites and other pathogens (Dempster et al. 2011, Johansen et al. 2011,
Arechavala-López et al. 2013, Glover et al. 2013), artificial lighting regimes that may delay

51
maturation and alter behaviour (Porter et al. 1999, Hansen et al. 2001, McConnell et al. 2010,
Otterå and Skilbrei 2014, Skilbrei and Otterå 2016, reviewed in Trippel 2010), contamination
from chemicals and metals used in aquaculture (Burridge et al. 2010), elevated predation risk
due to the abundance of large predatory fish, and where permitted, fishing mortality (Akyol &
Ertosluk 2010, Bagdonas et al. 2012). However, perhaps the most obvious change is the high
availability of waste feed that typically results in higher somatic and gonadal condition indices
for farm-associated wild fish (e.g. Dempster et al. 2011). 67-90% of fish sampled near Spanish
sea cages had consumed pellets (Fernandez-Jover et al. 2008), while pellets made up 71 and
25%, respectively, of the diet for farm-associated saithe and cod in Norway (Dempster et al.
2011).
Superficially, this trophic subsidy appears to benefit wild fish, but it also results in a dietary
shift from marine-derived highly-unsaturated omega-3 fatty acids to terrestrially-derived
omega-6 fatty acids. This in turn translates to a compositional shift in tissues (Fernandez-Jover
et al. 2007, Fernandez-Jover et al. 2011, Olsen et al. 2014, Arechavala-López et al. 2015), and
given that dietary lipids are reflected in the egg stores (e.g. Lavens et al. 1999, Czesny et al.
2000, Salze et al. 2005), may result in deficiencies in several of the fatty acids required for
reproduction and development. For example, captive feeding trials have found that essential
fatty acids 20:5 n-3 (eicosapentaenoic acid, EPA) and 22:6 n-3 (docosahexaenoic acid, DHA), as
well as 20:4 n-6 (arachidonic acid, AA), are important for fecundity, egg and sperm quality,
hatching success and larval development in fish (Reitan et al. 1994, Rainuzzo et al. 1997,
Sargent et al. 1999, Sargent et al. 1999, Rahman et al. 2014). Dietary deficiencies in these
reproductive nutrients contribute to low fertilisation and hatching rates in Atlantic cod (Gadus
morhua) broodstock relative to their wild counterparts (Salze et al. 2005) and have been linked
to changes in egg quality metrics in turbot (Scophthalmus maximus) (Lavens et al. 1999),
Atlantic halibut (Hippoglossus hippoglossus) (Mazorra et al. 2003) and cod (Bogevik et al.
2012). Furthermore, while data are lacking for fish, evidence from other taxa indicate that
additional effects may become apparent only after fertilisation; sea urchins (Echinus acutus)
reared on commercial salmon feed had higher gonad indices but lower fertilization and larval
survival rates, leading to an overall reduction in reproductive fitness (White et al. 2016, White
et al. 2017a).
The net effect of farm proximity on fitness, whether positive or negative, has a considerable
bearing on wild fish populations in farming areas. If trophic subsidies and associated conditions
provide a net fitness benefit, farm-associated fish populations will experience higher
production than those in neighbouring areas, and farms will act as a population source.

52
However, because farms are highly attractive to many fish species, any decline in fitness in
farm-associated wild fish may cause farms to function as ‘ecological traps’ (Robertson & Hutto
2006, Hale & Swearer 2016). Ecological traps are attractive but low quality habitats that can
have significant metapopulation-level impacts by drawing in individuals from higher quality
adjacent habitats, thus acting as attractive population sinks (Hale et al. 2015, Hale & Swearer
2016). To demonstrate the existence of an ecological trap, we must show that (1) individuals
prefer or show equal preference for the putative trap habitat relative to other available
habitats, and (2) that fitness outcomes in the putative trap habitat are lower than they would
have been in the other available habitats (Robertson & Hutto 2006, Patten & Kelly 2010). We
have strong empirical evidence that ecological traps affect birds and mammals in modified
terrestrial and aquatic environments (e.g. Remeš 2003, Weldon & Haddad 2005, Balme et al.
2010, Kloskowski 2012), but to date there have been very few tests of the theory in the marine
environment (Hallier and Gaertner 2008, Dempster et al. 2011, Sherley et al. 2017), and none
that have directly assessed fitness.
Norway operates the largest coastal fish aquaculture industry in the world, with Atlantic
salmon production reaching ~1.4 million tons in 2016 (Norwegian Directorate of Fisheries
2016). Fjord cod stocks, which are distinct from the more mobile offshore cod stocks
(Robichaud & Rose 2004), coexist with salmon farming in southern Norway and are at
historically low levels, with recent recruitment rates also low (Knutsen et al. 2016). Given these
concerns, the population can ill afford a potential ecological trap scenario. The >300 active
salmon farms in south-western Norway have widespread effects on fish movement and
population distribution and represent hundreds of potential ecological traps for fjord cod.
Gadid populations are strongly attracted to salmon grow-out cages (Dempster et al. 2011) and
move from farm to farm (Uglem et al. 2009, Otterå & Skilbrei 2014). This satisfies the first
component of an ecological trap assessment by showing preference for the putative trap
habitat (Robertson & Hutto 2006). However, we have no data on direct fitness measures in any
farm-associated fish, including gadids.
Farms may impact fitness in wild fish by altering either the survival or reproductive success of
individuals. Both pathways are difficult to measure directly in marine fishes, particularly highly
mobile broadcast spawning species, and as a result most previous work has relied on proxies
such as body condition (Fernandez-Jover et al. 2007, Dempster et al. 2011). Here we employ a
more direct approach by conducting a captive spawning experiment with wild-caught Atlantic
cod, to investigate whether reproductive fitness differs between areas of high and low salmon
farming intensity in south-western Norway. We assess potential reproductive effects spanning

53
initial adult body condition, ovarian fatty acid profiles and volumetric egg production, through
to egg quality metrics, hatching rates, and larval quality metrics including growth rates,
deformity rates and behaviour.
MATERIALS AND METHODS
Experimental design and fish husbandry
The density of salmon farms throughout Norway means that there are no longer any true
reference sites for the impacts of salmon farming on highly mobile wild fish in most parts of
the country. Instead, we make a comparison between two areas with differential farm density,
reflecting the typical spectrum of farm exposure for wild fish populations. We collected two
groups of mature live wild Atlantic cod (Gadus morhua) from the outer fjords of Hordaland
county in south-western Norway during February 2016. The first group (High Farm Density;
HFD) was collected by commercial fishers and technical staff from six locations along an 8 km
stretch of coastline in the Austevoll archipelago and 4 km away at Reksteren (Fig. 3.1). HFD
collection locations were all in relatively close proximity (300–1200 m) to six active salmon
farms, in an area of generally dense farming activity. The collections took place over ten days
between 1-15 February. The second group (Low Farm Density; LFD) was collected by
commercial fishers from two locations within the Bømlo archipelago, a neighbouring area with
relatively little fish farming activity (Fig. 3.1), between 5–15 February. HFD fish were collected
from sites exposed to a mean of 4.0 ha (range 1.5–5.2 ha) of sea cage surface area within 4 km
of their collection location, while LFD collection sites were exposed to mean 0.8 ha (range 0–
3.2 ha) within 4 km. These estimates include all sea cages and holding pens, including
enclosures that were unstocked at times during 2015 (for locations of active salmon grow-out
cages relative to collection sites, see Fig. 3.1). 75 % of LFD fish were collected ~9 km from the
nearest salmon cages, although it should be noted that the remaining 25 % of LFD fish were
collected within 2 km of a small salmon farm (2340 t capacity). As all fish were pooled within
groups, this is likely to reduce overall effect sizes observed in this study. We judged the farm
density within 4 km of the collection site to be a suitable metric of farm exposure based on
telemetry data describing the movements of tagged wild coastal cod in Norway and Shetland
(Neat et al. 2006, Svåsand et al. 2008, Uglem et al. 2008), but even if some proportion of fish
move larger distances, a considerable difference in the average level of farm exposure

54
between HFD and LFD groups will be maintained (Fig. 3.1). These two areas are otherwise
comparable environments in terms of gross hydrology, geology and ecology. Several logistical
factors prevented us from including additional sampling areas. Specifically, our need to collect
both HFD and LFD fish from shallow water to minimise barotrauma reduced the number of
suitable locations, as farms are typically placed in much deeper water. We were also restricted
to areas with similar latitude and water temperature to minimise temporal effects on
spawning (Kjesbu et al. 1994). In consideration of this geographic restriction, we consider our
results to be representative of outer Hardangerfjord but with relevance for similar systems
worldwide.
Figure 3.1. Map of collection sites relative to active salmon farms in south-western Norway.
Active outgrowing locations are taken from Norwegian Directorate of Fisheries aquaculture
biomass geodata.
Fish were captured using gill nets and fyke nets over reef or mixed sand-reef substrates at 5 -
30 m depth, and held in marine net pens, unfed, until the commencement of the experiment.
The experiment was conducted at the Austevoll Research Station, Norwegian Institute of
Marine Research (IMR). On 24 February 2016, we sedated all fish using 20 g L-1 tricaine
methanesulfonate (MS-222: Finquel). Fish with unhealed injuries or other welfare concerns

55
were killed with a blow to the head while sedated. We recorded length, wet weight and sex of
the remaining fish, inserted a Passive Integrated Transponder (PIT) tag into the peritoneal
cavity of every fish (allowing us to track individual weight loss between the start and finish of
spawning), and took ovarian biopsies from all females for storage at -80 °C. Within LFD and
HFD groups, 54 females and 24 males were allocated randomly among six cylindrical 7 m3
tanks per group (nine females and four males per tank, total 12 tanks).
Tanks were supplied with 6-8 °C sand-filtered and aerated seawater from 168 m depth and
exposed to a natural photoperiod through light-reducing shades. Hatchery facilities at IMR
Austevoll are described in detail by Karlsen et al. (2015). Winter spawning typically occurs in
the dark, during the early hours of the morning every 2-3 days from February to April. Each
spawning tank was appended with a cylindrical 100 L egg collection tank that filtered the full
volume of the spawning tank outflow via outlets at the top and bottom of the water column. A
constant circular flow was maintained within the egg collector to prevent eggs from being
pressed against the filter and damaged. The egg collectors were emptied every morning for
the duration of spawning and the volume of floating and sinking eggs was recorded.
Fish in this experiment were not fed for the duration of captivity to better preserve any effects
of diet prior to capture and to prevent clogging of egg collectors with waste matter. Reduced
feeding and significant weight loss is typical for both wild and captive cod during spawning
(Lambert & Dutil 2000), but to improve animal welfare, we removed fish from the experiment
early if their body condition dropped below acceptable levels (indicated by loss of muscle
mass, cessation of normal swimming behaviour, or unhealed wounds). Eggs were present after
the first night of egg collection, indicating that spawning had already begun. Accordingly, some
caution must be applied to interpretations of egg production and quality. However, the two
groups were collected at similar latitudes and housed in almost identical temperatures (the
main determinant of spawning commencement time: Kjesbu 1994), while temporal trends in
egg production do not suggest differential start or end times between groups (Fig. 3.4). Egg
production declined gradually toward the end of the spawning season, with the season
considered complete when all tanks had failed to produce viable eggs for four consecutive
days. All fish were humanely killed with a blow to the head while sedated with 20 g L-1 MS-222
before recording final length and weight.
On two occasions during the spawning season (3-4 March and 24 March), we took eggs for
incubation and hatching. Up to 350 mL eggs per tank were disinfected in 400 mg L−1

56
glutaraldehyde for 8 min to limit harmful bacterial growth (Harboe et al. 1994), and
transferred to 70 L black polyethylene conical incubators (one per spawning tank) with 0.5 L
min-1 seawater flow at 6 °C. Each morning during incubation, dead eggs were drained from the
bottom of the incubator and measured volumetrically. On 22 March, when the majority of
eggs from the first collection had hatched, we took approximately 6000 larvae from each
incubator and divided them across duplicate 50 L larval feeding tanks (24 tanks in total) at 8 °C.
Larvae remaining in the incubators were killed with a lethal dose of MS-222. Larvae were fed
size-fractionated copepod nauplii collected from the IMR sea-pond facility at Svartatjern.
Techniques for harvesting and preparation of copepods for rearing of cod larvae are described
in detail by van der Meeren et al. (2014) and Karlsen et al. (2015). Twice daily, larval rearing
tanks received 150000 nauplii, with 1.5 mL of algal paste per tank added to improve feeding
performance (Naas et al. 1992). The experiment was concluded on 13 April, with early season
larvae at day 42 and late season larvae at day 21 post fertilisation.
Ovarian fatty acid composition
We randomly selected ten ovarian samples per group (LFD and HFD) from the biopsies stored at the
beginning of the experiment. The samples weighed 60-100 mg each. All samples were methylated
and the respective fatty acid methyl esters were analysed on a HP-7890A gas chromatograph
(Agilent, USA) with a flame ionization detector (GC-FID), according to the method described in
Meier et al. (2006). The fatty acid 19:0 was added as an internal standard and 2.5 M dry HCl in
methanol was used as a methylation reagent. The methyl esters were extracted using 22 mL of
hexane, and the solution diluted or concentrated to obtain a suitable chromatographic response. 1
μL was injected splitless (the split was opened after 2 min) with the injection temperature set to
270 °C. The column was a 25 m × 0.25 mm fused silica capillary, coated with polyethylene-glycol of
0.25 μm film thickness, CP-Wax 52 CB (Varian-Chrompack, Middelburg, The Netherlands). Helium
(99.9999 %) was used as mobile phase at 1 ml/min for 45 min, then 3 ml/min for 25 min. The
temperature of the flame ionization detector was set at 300 °C. The oven temperature was
programmed to hold at 90 °C for 2 min, then heated to 150 °C at 30 °C/min and then to 240 °C at
2.5 °C/min and held steady for 30 min. Total analysis time was 70 min. Seventy well-defined peaks
in the chromatogram were selected, and identified by comparing retention times with a fatty acid
methyl ester standard (GLC-463 from Nu-Chek Prep. Elysian, MN, USA) and retention index maps
and mass spectral libraries (GC-MS) (http://www.chrombox.org/index.html) performed under the

57
same chromatographic conditions as the GC-FID (Wasta & Mjøs 2013). Chromatographic peak areas
were corrected by empirical response factors calculated from the areas of the GLC-463 mixture.
The chromatograms were integrated using the EZChrom Elite software (Agilent Technologies). Only
the 39 fatty acids that contributed more than 0.1 % of the total fatty acid amount were included in
the calculation. The total amount of fatty acids and cholesterol was calculated using the internal
standard 19:0.
Reproductive fitness traits
Body condition and weight loss
The amount and quality of egg production is likely to depend on fish condition. To allow
quantification of any such relationship, we compared body condition between LFD and HFD
groups (within sexes) using the relative condition index Krel = 100*(W/ExpW), where W is the
measured wet weight of the individual and ExpW is the expected weight (LeCren 1951). The
expected weight was calculated using a power function of the form ExpW = aLb fitted to the
full dataset including both LFD and HFD fish (Fig. 3.2). In this case, a = 0.0272 and b = 2.76 (R2 =
0.89). We also calculated proportional weight loss between the start and end of the spawning
season. This provides a general index of relative reproductive investment that is typically
closely correlated with egg production (e.g. Kjesbu et al. 1996).
Egg production
We quantified volumetric egg production in terms of daily egg production per tank, both with
and without a correction for the size of the females within a given tank, as well as any loss of
females during the season. Corrected egg production (relative daily egg production; RDEP) was
calculated as follows: RDEP = eggV / CFL, where eggV is the volume (mL) of eggs collected
from the tank, and CFL is the combined length (cm) of all females in the tank at the time of egg
collection. Four females (all HFD) experienced ≤10% weight loss, indicating little or no egg
release.
Egg quality

58
Viability, fertilization and early development
Egg viability rates were estimated from the proportion of eggs that were floating when the egg
collectors were emptied each morning. Fertilisation rates were assessed in subsamples of 100
eggs per tank on four occasions (three occasions for one HFD tank that finished spawning
early) during the spawning season (early season: 3-4 and 8-9 March, late season: 17 and 24-25
March), as egg quality typically declines during the spawning season (e.g. Kjesbu et al. 1996).
Where egg production was low on a given occasion, we combined fertilisation estimates from
two successive days. Eggs were scored as fertilised and normal (symmetrical cell divisions),
fertilised and abnormal (asymmetry or other abnormalities), or unfertilised.
Egg size and variability
Mean egg size and variability was measured on two occasions (3-4 March and 11 April) during
the spawning season. Eggs were taken from the egg collectors, stored in 6 °C seawater for 2-3
hours, and placed on a petri dish and photographed using a digital camera mounted on a light
microscope. The images had a 19 mm field of view at a resolution of 1024x768 pixels. We
measured the diameter of up to 50 eggs per tank (range 18-50) using the image analysis
software package ImageJ (Schneider et al. 2012), calibrated against a micrometer slide.
Hatching success
To estimate the proportion of viable eggs that successfully hatch, duplicate egg subsamples
(>100 eggs) were taken from incubators on two occasions, early and late in the season (early
season: 10 March, Day 8 of incubation; late season: 1 April, Day 9 of incubation) for a hatching
trial. Eggs were rinsed with filtered seawater, placed in covered containers filled with 200 mL
of filtered seawater, and maintained at 6-7 °C until hatching was complete. Dead eggs and live
and dead hatchlings were scored and removed daily until no viable embryos remained. As
there is large variation in growth rates between individual larvae, cannibalism makes it

59
impossible to reliably estimate impacts of farm density on larval survival in this system beyond
first feeding.
Larval quality
Larval development and growth rates
Subsamples of at least 30 larvae per tank were collected 40 days after spawning (23-28 days
post hatching), killed by a lethal dose of MS-222 and stored at 6 °C until required for
photography (<4 hrs). Larvae were transferred to a petri dish with a thin layer of seawater and
photographed laterally under dark field illumination on a light microscope with mounted
digital camera. The images provided a 12 mm field of view at a resolution of 1024x768 pixels.
Larvae were measured for length using the ImageJ measuring tool calibrated against a
micrometer slide. The measurement was taken along a polyline running from the tip of the
snout to the cranial vertebra and along the spine to the end of the caudal peduncle.
Phototaxis
Development of visual and cognitive systems in larvae (at day 42, ca. 23 days since first
feeding) was compared by means of a phototaxis trial in which we tested the proportion of
larvae exhibiting behavioural responses to a light gradient. Phototactic responses in this
context correlate with larval fitness (e.g. Karlsen and Mangor-Jensen 2001, Forsgren et al.
2013). Approximately 70 larvae (± 16.9 SD) were placed in a 60 x 10 x 5 cm tank and allowed to
disperse. We then completely covered the tank with black plastic to block any light. After 8
mins, we removed the covering on a 10 x 10 x 5 cm end section of the tank (thus exposing it to
ambient light from a fluorescent bulb) and scored the number of larvae already present in the
end section. This was treated as the control score: the number of larvae present in the end
section due to random dispersal without stimulus. The tank was then left for a further 5 mins,
and the number of larvae in the end section scored again. We then removed the rest of the
covering and counted the larvae that had not showed preference for light. The trial was
conducted for each of the 12 spawning tanks. The change in spatial distribution (the difference

60
in the proportion of larvae in the end section before and after the light treatment) provided a
measure of responsiveness to this environmental stimulus.
Statistical analysis
We compared initial fish size, weight-at-length and condition (Krel) metrics across LFD and HFD
groups (Group), using linear analysis of variance models constructed using the lm function in R
(R Core Team 2016). Data were log transformed as necessary to improve normality (and to
linearise weight-at-length curves). Proportional weight loss was compared between LFD and
HFD groups using a beta regression generalised linear model fitted using the betareg package
for R (Cribari-Neto & Zeileis 2010).
We conducted several ovarian fatty acid analyses: initally, we made univariate comparisons of
total lipids, cholesterol, and aquafeed markers oleic and linoleic acid across LFD and HFD
groups using linear analysis of variance models. We then compared the entire suite of fatty
acids across groups using a multivariate permutational analysis of variance (Permanova) fitted
to a Euclidean dissimilarity matrix using Primer 6 software (Anderson et al. 2008, White et al.
2017c).
Egg production metrics were compared between treatments and over time using negative
binomial generalised linear mixed models fitted using the glmmTMB package (Brooks et al.
2017). Group (farm density group) and Day (sampling day) were fitted as fixed effects. As tanks
were sampled repeatedly, nonindependence between samples was addressed by including a
random intercept term with TankID (tank identity) nested within Group. We report significance
of individual model terms within glmmTMB models by comparing the fit of the full model and
a null model with the term removed (X2 test on 1 df).
Effects of farm density on egg quality metrics were tested by fitting linear mixed models, also
using the glmmTMB package. Egg size data were best fitted with a gaussian model family,
while proportion data were fitted using a beta regression family. We included three fixed
terms: Group, Time (early or late season collections) and MeanFL (mean female length at the
tank level, to account for possible effects of female age on egg quality). As with egg production
models, we included the nested random intercept term Group/TankID. Proportion or rate
responses were analysed at the tank level. Egg size data were analysed at the egg level, with
Group, Time and MeanFL as fixed terms and Group/TankID as a random intercept term.
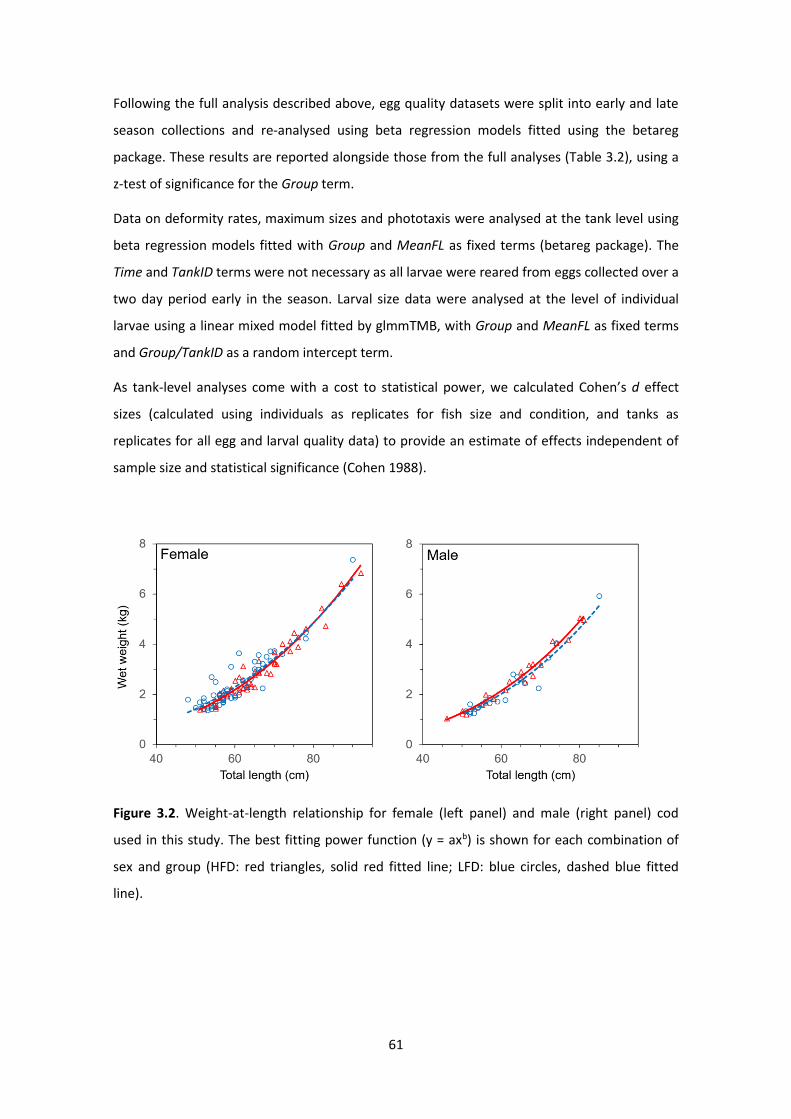
61
Following the full analysis described above, egg quality datasets were split into early and late
season collections and re-analysed using beta regression models fitted using the betareg
package. These results are reported alongside those from the full analyses (Table 3.2), using a
z-test of significance for the Group term.
Data on deformity rates, maximum sizes and phototaxis were analysed at the tank level using
beta regression models fitted with Group and MeanFL as fixed terms (betareg package). The
Time and TankID terms were not necessary as all larvae were reared from eggs collected over a
two day period early in the season. Larval size data were analysed at the level of individual
larvae using a linear mixed model fitted by glmmTMB, with Group and MeanFL as fixed terms
and Group/TankID as a random intercept term.
As tank-level analyses come with a cost to statistical power, we calculated Cohen’s d effect
sizes (calculated using individuals as replicates for fish size and condition, and tanks as
replicates for all egg and larval quality data) to provide an estimate of effects independent of
sample size and statistical significance (Cohen 1988).
Figure 3.2. Weight-at-length relationship for female (left panel) and male (right panel) cod
used in this study. The best fitting power function (y = axb) is shown for each combination of
sex and group (HFD: red triangles, solid red fitted line; LFD: blue circles, dashed blue fitted
line).
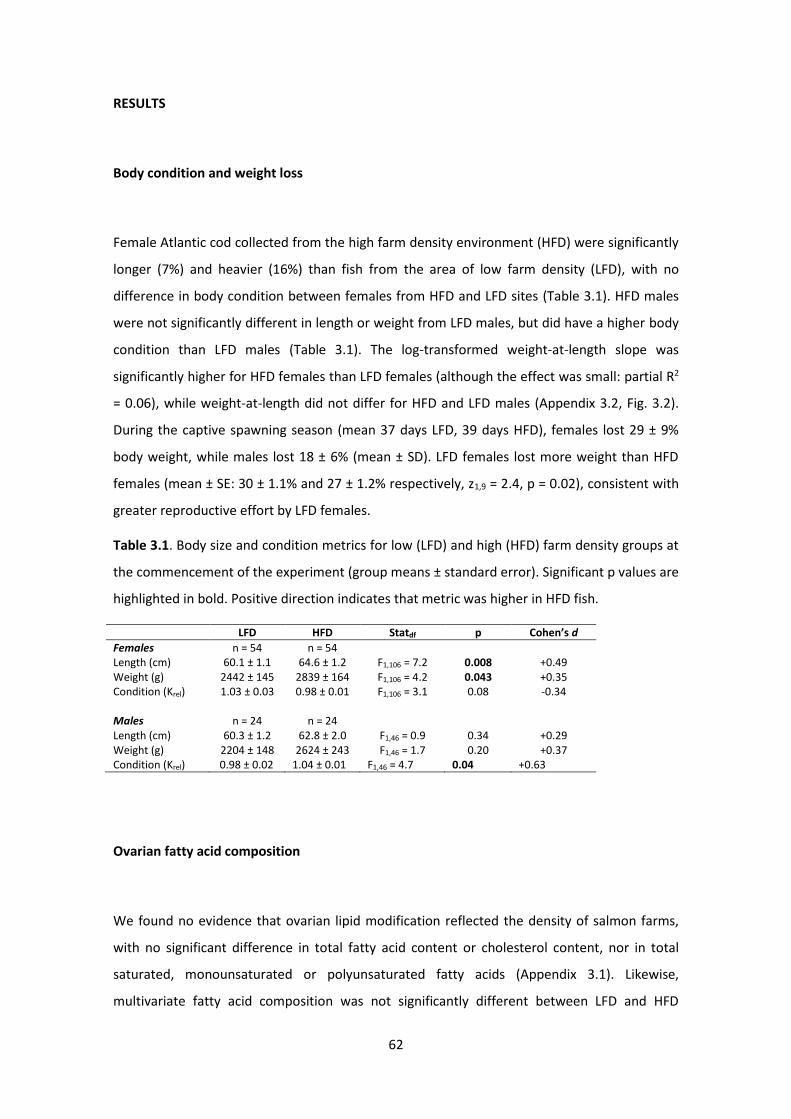
62
RESULTS
Body condition and weight loss
Female Atlantic cod collected from the high farm density environment (HFD) were significantly
longer (7%) and heavier (16%) than fish from the area of low farm density (LFD), with no
difference in body condition between females from HFD and LFD sites (Table 3.1). HFD males
were not significantly different in length or weight from LFD males, but did have a higher body
condition than LFD males (Table 3.1). The log-transformed weight-at-length slope was
significantly higher for HFD females than LFD females (although the effect was small: partial R2
= 0.06), while weight-at-length did not differ for HFD and LFD males (Appendix 3.2, Fig. 3.2).
During the captive spawning season (mean 37 days LFD, 39 days HFD), females lost 29 ± 9%
body weight, while males lost 18 ± 6% (mean ± SD). LFD females lost more weight than HFD
females (mean ± SE: 30 ± 1.1% and 27 ± 1.2% respectively, z1,9 = 2.4, p = 0.02), consistent with
greater reproductive effort by LFD females.
Table 3.1. Body size and condition metrics for low (LFD) and high (HFD) farm density groups at
the commencement of the experiment (group means ± standard error). Significant p values are
highlighted in bold. Positive direction indicates that metric was higher in HFD fish.
LFD HFD Statdf p Cohen’s d
Females n = 54 n = 54 Length (cm) 60.1 ± 1.1 64.6 ± 1.2 F1,106 = 7.2 0.008 +0.49 Weight (g) 2442 ± 145 2839 ± 164 F1,106 = 4.2 0.043 +0.35 Condition (Krel) 1.03 ± 0.03 0.98 ± 0.01 F1,106 = 3.1 0.08 -0.34 Males n = 24 n = 24 Length (cm) 60.3 ± 1.2 62.8 ± 2.0 F1,46 = 0.9 0.34 +0.29 Weight (g) 2204 ± 148 2624 ± 243 F1,46 = 1.7 0.20 +0.37 Condition (Krel) 0.98 ± 0.02 1.04 ± 0.01 F1,46 = 4.7 0.04 +0.63
Ovarian fatty acid composition
We found no evidence that ovarian lipid modification reflected the density of salmon farms,
with no significant difference in total fatty acid content or cholesterol content, nor in total
saturated, monounsaturated or polyunsaturated fatty acids (Appendix 3.1). Likewise,
multivariate fatty acid composition was not significantly different between LFD and HFD

63
groups (pseudo-F18 = 0.58, p = 0.71; Fig. 3.3). Multidimensional scaling (MDS) revealed that
three individuals—one LFD and two HFD—were separated from the main cluster. These three
fish were not remarkable in size or body condition (57-72 cm, 2022-3620 g, Krel 0.99-1.11), but
both HFD fish were high in oleic acid 18:1 (n-9) (15.2 and 17.5%) and one was also high in
linoleic acid 18:2 (n-6) (3.7%). The outlying LFD fish was also high in linoleic acid (2.5%). Both
oleic and linoleic acid can indicate consumption of commercial feed (White et al. 2017c), and
were marginally higher in the HFD group on average (6% and 18% increases, respectively), but
this effect was not significant for either oleic acid (F1,18 = 1.9, p = 0.2) or linoleic acid (F1,18 = 0.4,
p = 0.6). Levels of these fatty acids were strongly positively correlated with each other in the
HFD group (Pearson’s r = 0.90, t8 = 6.0, p <0.001), while this effect was nonsignificant in the
LFD group (r = 0.44, t8 = 1.4, p = 0.2). The highest levels of both oleic and linoleic acid (17.5%
and 3.7% respectively) occurred in one individual from the HFD group.
Figure 3.3. Multidimensional scaling (MDS) plot showing dissimilarly (Euclidean distance) of
multivariate fatty acid profiles in Atlantic cod ovaries according to salmon farm density.
Individuals are grouped by high (HFD; red triangles) and low (LFD; blue circles) salmon farm
density. Model stress is 0.08.
Egg production
Twelve spawning tanks produced a total of 137 L of eggs over the spawning season, with no
difference in the total egg volume from LFD and HFD tanks (11.3 ± 0.9 L and 11.3 ± 0.7 L,

64
respectively; Fig. 3.4). A model comparing raw daily egg production between HFD and LFD
tanks over time found no effect of the farm density factor (p = 0.08; Table S3). There was a
significant temporal decline in daily egg production (p <0.0001, Fig. 3.4), and a positive effect
of total female length in the tank on daily egg production (p <0.0001).
The relative daily egg production metric (daily egg production corrected for total female
length) also did not significantly differ across groups (LFD: 64.6 ± 6.2 cf. HFD: 57.2 ± 3.4 mL
female m-1 day-1 tank-1; p = 0.52), but again, there was a significant decline over time (p
<0.0001, Fig. 3.4).
Initial female body condition (tank mean values) did not significantly predict either raw daily
egg production or relative daily egg production metrics overall (p >0.10 in both cases), or
within farm density groups (p >0.05 in each case). However, mean female weight loss during
the season was strongly correlated with both egg production metrics (p <0.0004 in each case).
Figure 3.4. Daily egg production per tank during the captive spawning period (25 Feb 2016 – 11
April 2016). Tanks are grouped by high (HFD; red triangles, solid red fitted line) and low (LFD;
blue circles, dashed blue fitted line) farm density. Production is quantified by (left panel) raw
daily egg volume per tank and (right panel) with a correction for the total length of females in
the tank. The temporal trend within groups is fitted by third order polynomial functions.
Egg quality and survivorship
The proportion of viable (floating) eggs was almost identical across farm density groups (Table
3.2; Table S3), with the viable proportion significantly declining over the season (p <0.0001).

65
Neither mean female length (p = 0.18) nor body condition (p = 0.60) were correlated with egg
viability rates at the tank level, regardless of farm density group.
The LFD group produced larger eggs over the duration of the season (Table 3.2; Appendix 3.3).
LFD eggs were 4.8 % larger by diameter at the time of the early season collections, with no
significant difference later in the season (Table 3.2). We did not test for a temporal decline in
egg size, as late but not early season samples were fixed in formalin. Again, neither mean
female length (p = 0.065) nor body condition (p = 0.32) were significantly correlated with egg
size, regardless of farm density group.
Rates of asymmetrical cell division in pre-blastula eggs did not differ between LFD and HFD
groups in either early or late season collections (Table 3.2; Appendix 3.3). Neither mean female
size nor body condition were correlated with egg symmetry (p >0.06 in each case). Mean
asymmetry rates did not exceed 10% in either group on any collection date, although samples
ranged from 0-53% asymmetry.
Early season eggs hatched 12-17 days after collection, while late season eggs hatched 14-18
days after collection. Egg survival during incubation prior to the hatching trial did not differ
significantly between groups (Table 3.2; Appendix 3.3). Declines in survivorship between early
and late season collections did not differ (p >0.10), nor did the effect of mean female length or
body condition (p >0.10 in each case).
Hatching success rates were similar between groups (Table 3.2; Appendix 3.3), with no decline
between early and late season collections. One HFD tank ceased spawning before the late
season egg sampling, so late season HFD hatching data comes from only five tanks.
Larval quality
Larvae from LFD tanks were 8 % larger than those from HFD tanks (Table 3.2; Appendix 3.3).
Neither mean female size (p = 0.97) nor body condition (p = 0.34) were correlated with larval
size either overall or within farm density groups. The size of the largest larvae from each tank
did not differ across farm density groups, nor did the rate of deformities (Table 3.2; Appendix
3.3). Neither were significantly correlated with mean female length or body condition (p >0.10
in each case). Overall rates of larval deformity were 48% at day 40 from both LFD and HFD
groups (Table 3.2). Most deformities were of the spine (lordosis, kyphosis or vertebral
misalignment) (152/173 deformities), followed by deformities of the jaw (15/173) and skull

66
(6/173). Larvae from both groups exhibited phototaxis in response to a horizontal light
gradient (proportion in end section before exposure: 0.25 ± 0.03 cf. after exposure: 0.69 ±
0.04; t-test: t22 = -9.5, p <0.0001), but we found no evidence that the farm density groups
differed in the extent of the phototactic response (Table 3.2; Appendix 3.3), nor any strong
evidence for an effect of mean female length or body condition (p >0.13 in each case).
DISCUSSION
This study presents limited evidence for negative impacts of high salmon farm density on
reproductive fitness in the studied Atlantic cod (Gadus morhua) population. Female cod
collected from locations with a high density of salmon farms (HFD) were larger and heavier
than fish from the low farm density location (LFD), but with no consistent changes in female
body condition. Effects on ovarian lipid composition were also small and largely limited to two
HFD individuals. There was no significant difference in egg production, viability, fertilisation
rates, symmetry or hatching success, but eggs from HFD tanks were 5 % smaller and this likely
contributed to the observed -8 % disparity in mean larval length at 40 days post spawning for
the HFD group relative to the LFD group.
Among the viable proportion, fertilisation rates were similar for LFD and HFD tanks
respectively (Table 3.2; Appendix 3.3), but declined during the season (p = 0.009). Neither
mean female length nor body condition significantly predicted fertilisation rates overall (p
>0.06 in each case), although the effect of female length was significant within the LFD group
(p = 0.04). Given the very small difference in egg viability and fertilisation rates between
groups, and nonsignificant effects of male length and body condition on fertilisation rates
either overall or within groups (p >0.13 in each case), we did not test for differences in sperm
quality.
Gadid fishes accumulate significant energy reserves for reproduction, with lipids stored
primarily in the liver and proteins in muscle tissue (Kjesbu et al. 1991, Lambert & Dutil 1997).
Accordingly, body condition indices during vitellogenesis are typically good predictors of
fecundity in coastal cod from this region (Skjæraasen et al. 2006). In our case, mean female
size and percentage weight loss during the season positively tracked egg production at the
tank level; both were better predictors of egg production than initial female body condition.
Table 3.2. Egg and larval quality metrics for cod from low (LFD) and high (HFD) farm density
areas (group mean ± standard error). Positive effect sizes indicate that quality was higher in

67
the HFD group. Significance has not been corrected for false discovery rate of multiple
comparisons.
LFD HFD N (HFD, LFD) Stat p Cohen’s d
Viability rate
Early season 0.96 ± 0.01 0.94 ± 0.03 102, 102 z = 0.21 0.83 -0.25
Late season 0.78 ± 0.15 0.76 ± 0.11 70, 74 z = 0.56 0.56 -0.19
Overall 0.87 ± 0.04 0.85 ± 0.04 172, 176 X2 = 0.23 0.61 -0.14
Egg diameter (μm) (of viable eggs)
Early season 1183 ± 16 1129 ± 16 257, 289 X2 = 5.8 0.016 -1.36
Late season* 1261 ± 30 1210 ± 13 221, 271 X2 = 5.3 0.07 -0.92
Overall 1222 ± 20 1166 ± 16 478, 560 X2 = 6.5 0.011 -0.90
Fertilisation rate (of viable eggs)
Early season 0.75 ± 0.04 0.77 ± 0.03 19, 20 z = 0.68 0.49 +0.27
Late season 0.59 ± 0.09 0.68 ± 0.06 15, 18 z = 0.47 0.63 +0.46
Overall 0.67 ± 0.08 0.73 ± 0.04 34, 38 X2 = 1.0 0.60 +0.36
Egg symmetry rate (of fertilised viable eggs)
Early season 0.92 ± 0.01 0.90 ± 0.02 19, 20 z = 0.69 0.49 -0.29
Late season 0.88 ± 0.18 0.88 ± 0.08 15, 18 z = 0.48 0.63 -0.10
Overall 0.90 ± 0.03 0.89 ± 0.04 34, 38 X2 = 0.35 0.84 -0.08
Egg survival rate during incubation
Early season 0.85 ± 0.03 0.83 ± 0.05 6, 6 z = 0.66 0.49 -0.23
Late season 0.56 ± 0.11 0.68 ± 0.09 5, 6 z = 0.48 0.63 +0.41
Overall 0.70 ± 0.08 0.76 ± 0.06 11, 12 X2 = 0.01 0.95 +0.23
Hatching success rate
Early season 0.90 ± 0.03 0.85 ± 0.03 12, 12 z = 1.56 0.12 -0.64
Late season 0.92 ± 0.03 0.88 ± 0.08 11, 12 z = 0.15 0.88 -0.07
Overall 0.91 ± 0.02 0.86 ± 0.04 23, 24 X2 = 0.18 0.67 -0.48
Larval length (mm)
Early season 6.46 ± 0.05 5.98 ± 0.18 180, 176 X2 = 3.9 0.048 -1.46
Max larval length (mm)
Early season 8.05 ± 0.17 7.68 ± 0.21 6, 6 t1, 10 = 1.32 0.21 -0.77
Larval deformity rate
Early season 0.48 ± 0.06 0.48 ± 0.07 6, 6 z1,9 = 0.01 0.99 0
Larval phototaxis rate
Early season 0.71 ± 0.07 0.67 ± 0.04 6, 6 z1,9 = 0.79 0.43 -0.28
*Late season eggs were fixed in formalin prior to examination, which may have affected egg diameter.

68
However, the direction of the body condition trend was positive, and together, our findings are
consistent with greater reproductive investment relative to body length by LFD females.
Egg quality metrics revealed similar quality overall in LFD and HFD groups, but with
significantly smaller eggs from HFD fish. This was somewhat unexpected, as HFD females were
slightly larger; female size and age in cod is usually positively correlated with egg size (e.g.
Marteinsdottir & Steinarsson 1998, Vallin & Nissling 2000), although body condition can be
equally important (Chambers & Waiwood 1996, Marteinsdottir & Steinarsson 1998). The egg
size effect that we observed did not correspond to any significant decrease in other metrics of
egg quality, and the cumulative effect of the more direct metrics (viability, fertilisation,
developmental symmetry, survival during incubation and hatching success) was such that eggs
collected from HFD and LFD tanks did not differ substantially in their likelihood of successfully
hatching (HFD: 40 % cf. LFD: 37 %). However, previous studies have indicated that egg size can
predict larval quality, with larger size at hatching, faster growth rates and successful
development of the swim bladder leading to a survival advantage for larvae from larger eggs
(Knutsen & Tilseth 1985, Marteinsdottir & Steinarsson 1998). This prediction was consistent
with our larval size data; HFD larvae were 8 % smaller on average than their LFD counterparts
40 days after spawning.
Larger larvae often exhibit differing responses to stimuli such as light (e.g. Colton & Hurst
2010). In some fish, phototaxis along a horizontal light gradient correlates positively with other
metrics of development (e.g. first feeding: Karlsen & Mangor-Jensen 2001) and is affected by
the environment (Forsgren et al. 2013). In our case, LFD and HFD fish did not differ in their
phototactic response to a horizontal light gradient, although the effect was in the direction of
the greater phototaxis by LFD larvae (Cohen’s d = 0.28). Regardless, even small differences in
hatching size and larval growth rates can affect survival during the planktonic stage and even
influence post settlement fitness (e.g. Sclafani et al. 1993, Shima & Swearer 2010), so it is likely
that the HFD larvae in this study would have experienced nontrivial negative fitness effects
later in development.
Condition indices for fish in the present study were similar to those from wild caught fish in the
same region prior to the expansion of salmon farming (Botros 1962, cited in Kjesbu 1989). Egg
sizes in our study (mean 1.2 mm) were at the lower end of those reported by previous captive
spawning studies with Norwegian coastal cod (1.2-1.4 mm: Kjesbu et al. 1996, Otterå et al.
2006, van der Meeren & Ivannikov 2006, Bogevik et al. 2012, Karlsen et al. 2015), but this may
be related to the lack of feed during spawning rather than condition at the time of collection
(Kjesbu et al. 1990). Egg fertilisation, symmetry and hatching rates were all within the range of

69
those reported by previous studies (Morgan et al. 1999, van der Meeren & Ivannikov 2006,
Bogevik et al. 2012, Karlsen et al. 2015), while larval growth rates (6.2 mm cf. 7.4 mm at 25
days post hatching) were slightly below those reported by Karlsen et al. (2015).
While previous studies have found significant shifts in ovarian fatty acid composition in
captive-fed sea bream (Cejas et al. 2003) and farm-associated bogue (L. Martinez-Rubio
unpubl. data, cited in Fernandez-Jover et al. 2011b), the absence of such clear effects in this
study should not be taken as strong evidence that these fish have not fed at salmon farms.
Lipid profiles of the gonads are less affected by diet than those of the liver or muscle tissue, as
the gonads are composed almost entirely of phospholipids rather than dietary fatty acids.
Accordingly, our analysis of ovarian lipid composition may only have detected effects of a large
dietary shift, while the presence of a single active salmon farm near one LFD location (affecting
25 % of LFD fish) may have reduced overall differences in fatty acid profiles between HFD and
LFD groups. Indirect intake of farm waste via predation of farm-associated invertebrates and
fish, combined with dietary sparing and biosynthesis, may also weaken or mask fatty acid
signals in fish that are not strongly farm-associated (White et al. 2017c). The strong correlation
between levels of oleic and linoleic acids in the HFD group (less so in the LFD group) is
consistent with (but not necessarily strong evidence for) a spectrum of farm association, with
the most strongly farm-associated individuals in the HFD group having higher levels of oleic
and linoleic acids than their counterparts in the LFD group. Both oleic and linoleic acid are
present in the natural diet but are especially abundant in commercial salmon feed, and captive
feeding trials revealed a strong correlation between these two fatty acids in muscle, liver and
gonad tissue of saithe fed on salmon feed (Karlsen et al., unpublished data).
Our collection sites were selected to represent levels of farm exposure experienced by cod
populations in the south-western fjords (Fig. 3.1). Importantly, cod do not associate with farms
as closely or persistently as other species (e.g. saithe), and likely do not feed exclusively at
farms for extended periods (Dempster et al. 2011), so our experiment was designed to test for
effects of farm density on cod whose home ranges at the time of collection overlap with areas
of farming influence. Salmon farms have relatively localised nutrient footprints, with acute
deposition within 50-250 m and very diffuse deposition beyond 500 m (Kutti et al. 2007,
Bannister et al. 2016, White et al. 2017b). Accordingly, our collection sites are likely to reflect
the two dominant types of farm exposure for wild Norwegian fjord cod: temporally dynamic or
partial association versus little or no association. Tissue fatty acid profiles in cod are altered
within 3 weeks of a dietary shift (Kirsch et al. 1998), with vitellogenesis commencing 3-4
months prior to spawning (Skjæraasen et al. 2006). Individuals that reside within farm

70
footprints throughout vitellogenesis are likely to show the largest shifts in ovarian fatty acid
profiles and concomitant effects on reproductive physiology and output. We do not know how
long HFD and LFD cod have resided at their capture locations, but available telemetry and
mark-recapture data indicate that wild coastal cod have relatively restricted home ranges over
a scale of weeks and months. 87 % of tag returns for wild cod released in Heimarkspollen, a 2.9
km2 semi-enclosed fjord in the Austevoll archipelago, were recaptured within the fjord
(Svåsand 1990), while in Balsfjord in northern Norway, the majority of wild cod tagged and
released at a farm were still present at the same farm 9-12 weeks later (Uglem et al. 2008). In
the Shetland Isles, 133 wild cod were tagged and released, with 37/39 recaptures over a two
year period occurring within 15 km of the release site (Neat et al. 2006). While some
individuals move larger distances and may have spent time in areas that differ from their
capture location in terms of farm density, our study only assumes that fish collected from LFD
locations will, on average, be less affected by farms than fish from HFD locations.
Previous captive feeding experiments have shown that gadids and other fishes fed commercial
diets experience changes in reproductive fitness (e.g. Salze et al. 2005, Bogevik et al. 2012),
but very little work has been done to assess potential impacts in a real world ecological
context. Taken together, our findings indicate that salmon farming in this region has some
negative effects on the reproductive physiology of Atlantic cod on a fjord-wide scale, with
potential cumulative effects of egg and larval size on later developmental stages. More work is
needed to track fitness effects later in development, including potential effects of decreased
egg and larval size on later life stages. In addition, as our study was restricted in its geographic
extent and spatial replication, we are cautious of generalising these findings beyond our study
environment. We encourage others to replicate and extend this important line of research.
Future work may also consider the other potential pathway for fitness impacts: individual
mortality. Mortality may decline due to the provision of a trophic subsidy (Kilambi et al. 1978),
or increase due to elevated levels of contamination, infection, or predation at farms. Wild fish
aggregations are also an easy target for fishers, and fishing mortality will take on greater
importance if the current 100 m fishing restriction around Norwegian farms is lifted (Bagdonas
et al. 2012). Previous studies have employed acoustic tracking to good effect in comparing
spatiotemporal movement and mortality rates of Atlantic cod individuals across habitats
(Olsen et al. 2011, Olsen et al. 2012, Fernández-Chacón et al. 2015), and it would be entirely
feasible to apply the same approach to quantify differential mortality rates in wild fish across
multiple farm-affected and non-affected areas.

71
ACKNOWLEDGMENTS
We thank the staff at IMR Austevoll for their expert advice and assistance in the collection and
maintenance of the spawning fish, egg incubation and feeding of larvae: Margareth Møgster,
Stig Ove Utskot, Michal Rejmer, Inger Semb Johansen, Nele Gunkel-Sauer, Kristine Hovland
Holm, Yvonne Rong, Terje van der Meeren, Tord Skår, Velimir Nola and Glenn Sandtorv. We
also thank the staff and students at Austevoll High School for their assistance in transporting
live fish to the research station. Theresa Aase prepared the fatty acid analysis. Camille White
provided useful comments on lipid data. Francisca Samsing assisted with the creation of
figures. The manuscript was improved by comments from three anonymous reviewers. All
procedures were conducted in accordance with Norwegian animal welfare regulations by
experienced personnel.
This project was funded by the Sustainable Aquaculture Laboratory, University of Melbourne,
and the Norwegian Seafood Research Fund (Fiskeri og Havbruksnæringens Forskningsfond).
LITERATURE CITED
Akyol O, Ertosluk O (2010) Fishing near sea-cage farms along the coast of the Turkish Aegean
Sea. J Appl Ichthyol 26:11–15
Anderson M, Gorley R, Clarke R (2008) Permanova+ for Primer: guide to software and
statistical methods. Primer-E Limited, Plymouth.
Arechavala- López P, Sæther B-S, Marhuenda-Egea F, Sánchez-Jerez P, Uglem I (2015)
Assessing the influence of salmon farming through total lipids, fatty acids, and trace
elements in the liver and muscle of wild saithe Pollachius virens. Mar Coast Fish 7:59–
67
Arechavala-López P, Sánchez-Jerez P, Bayle-Sempere JT, Uglem I, Mladineo I (2013) Reared
fish, farmed escapees and wild fish stocks—a triangle of pathogen transmission of
concern to Mediterranean aquaculture management. Aquac Environ Interact 3:153–
161
Bagdonas K, Humborstad O-B, Løkkeborg S (2012) Capture of wild saithe (Pollachius virens)
and cod (Gadus morhua) in the vicinity of salmon farms: Three pot types compared.
Fish Res 134–136:1–5

72
Balme GA, Slotow R, Hunter LTB (2010) Edge effects and the impact of non-protected areas
in carnivore conservation: leopards in the Phinda–Mkhuze Complex, South Africa.
Anim Conserv 13:315–323
Bannister RJ, Johnsen IA, Hansen PK, Kutti T, Asplin L (2016) Near- and far-field dispersal
modelling of organic waste from Atlantic salmon aquaculture in fjord systems. ICES J
Mar Sci 73:2408–2419
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed effects models using
lme4. J Stat Softwar 67:1–48
Bogevik AS, Natário S, Karlsen Ø, Thorsen A, Hamre K, Rosenlund G, Norberg B (2012) The
effect of dietary lipid content and stress on egg quality in farmed Atlantic cod Gadus
morhua. J Fish Biol 81:1391–405
Botros G (1962) Die Fruchtbarkeit des Dorsches (Gadus morhua L.) in der westlichen Ostsee
und den westnorwegischen Gewassern. Kieler Meeresforsch 18:67–80
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ,
Maechler M, Bolker BM (2017) Modeling zero-inflated count data with glmmTMB.
bioRxiv preprint bioRxiv:132753.
URL: https://www.biorxiv.org/content/early/2017/05/01/132753
Burridge L, Weis JS, Cabello F, Pizarro J, Bostick K (2010) Chemical use in salmon aquaculture:
A review of current practices and possible environmental effects. Aquaculture 306:7–
23
Cejas JR, Almansa E, Villamandos JE, Badıa P, Bolaños A, Lorenzo A (2003) Lipid and fatty acid
composition of ovaries from wild fish and ovaries and eggs from captive fish of white
sea bream (Diplodus sargus). Aquaculture 216:299–313
Chambers RC, Waiwood KG (1996) Maternal and seasonal differences in egg sizes and
spawning characteristics of captive Atlantic cod, Gadus morhua. Can J Fish Aquat Sci
53:1986–2003
Cohen J (1988) Statistical power analysis for the behavioral sciences (2nd edn). Erlbaum,
Hillsdale
Colton AR, Hurst TP (2010) Behavioral responses to light gradients, olfactory cues, and prey
in larvae of two North Pacific gadids (Gadus macrocephalus and Theragra
chalcogramma). Environ Biol Fishes 88:39–49
Cribari-Neto F, Zeileis A (2010) Beta egression in R. J Stat Softw 34:1–24

73
Czesny S, Dabrowski K, Christensen JE, Eenennaam J Van, Doroshov S (2000) Discrimination
of wild and domestic origin of sturgeon ova based on lipids and fatty acid analysis.
Aquaculture 189:145–153
Dempster T, Sánchez-Jerez P, Bayle-Sempere JT, Giménez-Casalduero F (2002) Attraction of
wild fish to sea-cage fish farms in the south-western Mediterranean Sea: spatial and
short-term temporal variability. Mar Ecol Prog Ser 242:237–252
Dempster T, Sánchez-Jerez P, Fernandez-Jover D, Bayle-Sempere JT, Nilsen R, Bjørn P-A,
Uglem I (2011) Proxy measures of fitness suggest coastal fish farms can act as
population sources and not ecological traps for wild gadoid fish. PLOS ONE 6:e15646
Dempster T, Sánchez-Jerez P, Uglem I, Bjørn P-A (2010) Species-specific patterns of
aggregation of wild fish around fish farms. Estuar Coast Shelf Sci 86:271–275
Dempster T, Uglem I, Sánchez-Jerez P, Fernandez-Jover D, Bayle-Sempere J, Nilsen R (2009)
Coastal salmon farms attract large and persistent aggregations of wild fish: an
ecosystem effect. Mar Ecol Prog Ser 385:1–14
Fernández-Chacón A, Moland E, Espeland SH, Olsen EM (2015) Demographic effects of full vs.
partial protection from harvesting: inference from an empirical before-after control-
impact study on Atlantic cod. J Appl Ecol 52:1206–1215
Fernandez-Jover D, Arechavala-López P, Martinez-Rubio L, Tocher D, Bayle-Sempere J, López-
Jiménez J, Martinez-López F, Sánchez-Jerez P (2011) Monitoring the influence of
marine aquaculture on wild fish communities: benefits and limitations of fatty acid
profiles. Aquac Environ Interact 2:39–47
Fernandez-Jover D, Jimenez JAL, Sánchez-Jerez P, Bayle-Sempere JT, Giménez-Casalduero F,
López FJM, Dempster T (2007) Changes in body condition and fatty acid composition of
wild Mediterranean horse mackerel (Trachurus mediterraneus, Steindachner, 1868)
associated to sea cage fish farms. Mar Environ Res 63:1–18
Fernandez-Jover D, Martinez-Rubio L, Sánchez-Jerez P, Bayle-Sempere JT, Lopez Jimenez JA,
Martínez López FJ, Bjørn P-A, Uglem I, Dempster T (2011) Waste feed from coastal fish
farms: a trophic subsidy with compositional side-effects for wild gadoids. Estuar Coast
Shelf Sci 91:559–568
Fernandez-Jover D, Sánchez-Jerez P, Bayle-Sempere JT, Valle C, Dempster T (2008) Seasonal
patterns and diets of wild fish assemblages associated with Mediterranean coastal fish
farms. ICES J Mar Sci 65:1153–1160
Forsgren E, Dupont S, Jutfelt F, Amundsen, T (2013) Elevated CO2 affects embryonic
development and larval phototaxis in a temperate marine fish. Ecol Evol 3:3637–3646

74
Glover KA, Sørvik AGE, Karlsbakk E, Zhang Z, Skaala Ø (2013) Molecular genetic analysis of
stomach contents reveals wild Atlantic cod feeding on piscine reovirus (PRV) infected
Atlantic salmon originating from a commercial fish farm. PLOS ONE 8:e60924
Hale R, Swearer SE (2016) Ecological traps: current evidence and future directions. Proc R Soc
B Biol Sci 283:494–499
Hale R, Treml EA, Swearer SE (2015) Evaluating the metapopulation consequences of
ecological traps. Proc R Soc London B Biol Sci 282
Hansen T, Karlsen Ø, Taranger GL, Hemre G-I, Holm JC, Kjesbu OS (2001) Growth, gonadal
development and spawning time of Atlantic cod (Gadus morhua) reared under
different photoperiods. Aquaculture 203:51–67
Hallier J-P, Gaertner D (2008) Drifting fish aggregation devices could act as an ecological trap
for tropical tuna species. Mar Ecol Prog Ser 353:255–264
Harboe T, Huse I, Øie G (1994) Effects of egg disinfection on yolk sac and first feeding stages
of halibut (Hippoglossus hippoglossus L.) larvae. Aquaculture 119:157–165
Johansen L-H, Jensen I, Mikkelsen H, Bjørn P-A, Jansen PA, Bergh Ø (2011) Disease
interaction and pathogens exchange between wild and farmed fish populations with
special reference to Norway. Aquaculture 315:167–186
Karlsen Ø, Mangor-Jensen A (2001) A correlation between phototactic response and first-
feeding of Atlantic halibut (Hippoglossus hippoglossus L.) larvae. Aquacult Res 32: 907–
912
Karlsen Ø, Meeren T van der, Rønnestad I, Mangor-Jensen A, Galloway TF, Kjørsvik E, Hamre
K (2015) Copepods enhance nutritional status, growth and development in Atlantic
cod (Gadus morhua L.) larvae - can we identify the underlying factors? PeerJ 3:e902
Kilambi RV, Adams JC, Wickizer WA (1978) Effects of cage culture on growth, abundance, and
survival of resident largemouth bass (Micropterus salmoides). J Fish Res Board Canada
35:157–160
Kirsch PE, Iverson SJ, Bowen WD, Kerr SR, Ackman RG (1998) Dietary effects on the fatty acid
signature of whole Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 55:1378–1386
Kjesbu OS (1989) The spawning activity of cod, Gadus morhua L. J Fish Biol 34:195–206
Kjesbu OS (1994) Time of start of spawning in Atlantic cod (Gadus morhua) females in
relation to vitellogenic oocyte diameter, temperature, fish length and condition. J Fish
Biol 45:719–735

75
Kjesbu OS, Klungsøyr J, Kryvi H, Witthames PR, Walker MG (1991) Fecundity, atresia, and egg
size of captive Atlantic cod (Gadus morhua) in relation to proximate body composition.
Can J Fish Aquat Sci 48:2333–2343
Kjesbu OS, Solemdal P, Bratland P, Fonn M (1996) Variation in annual egg production in
individual captive Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 53:610–620
Kjesbu OS, Witthames PR, Solemdal P, Walker MG (1990) Ovulatory rhythm and a method to
determine the stage of spawning in Atlantic cod (Gadus morhua). Can J Fish Aquat Sci
47:1185–1193
Kloskowski J (2012) Fish stocking creates an ecological trap for an avian predator via effects
on prey availability. Oikos 121:1567–1576
Knutsen H, Olsen EM, Espeland SH (2016) Norwegian coastal cod south of 62° N (in
Norwegian). In: Institute for Marine Research Report (Havforskningsrapporten).
Norwegian Institute for Marine Research (Havforskningsinstituttet)
Knutsen GM, Tilseth S (1985) Growth, development, and feeding success of Atlantic cod
larvae Gadus morhua related to egg size. Trans Am Fish Soc 114:507–511
Kutti T, Ervik A, Hansen PK (2007) Effects of organic effluents from a salmon farm on a fjord
system. I. Vertical export and dispersal processes. Aquaculture 262:367–381
Lambert Y, Dutil J-D (1997) Can simple condition indices be used to monitor and quantify
seasonal changes in the energy reserves of cod (Gadus morhua)? Can J Fish Aquat Sci
54:104–112
Lambert Y, Dutil J-D (2000) Energetic consequences of reproduction in Atlantic cod (Gadus
morhua) in relation to spawning level of somatic energy reserves. Can J Fish Aquat Sci
57:815–825
Lavens P, Lebegue E, Jaunet H, Brunel A, Dhert P, Sorgeloos P (1999) Effect of dietary
essential fatty acids and vitamins on egg quality in turbot broodstocks. Aquac Int
7:225–240
LeCren ED (1951) The length-weight relationship and seasonal cycle in gonad weight and
condition in the perch (Perca fluviatilis). J Anim Ecol 20: 201–219
Marteinsdottir G, Steinarsson A (1998) Maternal influence on the size and viability of Iceland
cod Gadus morhua eggs and larvae. J Fish Biol 52:1241–1258
Mazorra C, Bruce M, Bell JG, Davie A, Alorend E, Jordan N, Rees J, Papanikos N, Porter M,
Bromage N (2003) Dietary lipid enhancement of broodstock reproductive performance
and egg and larval quality in Atlantic halibut (Hippoglossus hippoglossus). Aquaculture
227:21–33

76
McConnell A, Routledge R, Connors B (2010) Effect of artificial light on marine invertebrate
and fish abundance in an area of salmon farming. Mar Ecol Prog Ser 419: 147–156
Meeren T van der, Ivannikov VP (2006) Seasonal shift in spawning of Atlantic cod (Gadus
morhua L.) by photoperiod manipulation: egg quality in relation to temperature and
intensive larval rearing. Aquac Res 37:898–913
Meeren T van der, Karlsen Ø, Liebig PL, Mangor-Jensen A (2014) Copepod production in a
saltwater pond system: A reliable method for achievement of natural prey in start-
feeding of marine fish larvae. Aquac Eng 62:17–27
Meier S, Mjøs SA, Joensen H, Grahl-Nielsen O (2006) Validation of a one-step
extraction/methylation method for determination of fatty acids and cholesterol in
marine tissues. J Chromatogr A 1104:291–8
Morgan MJ, Wilson CE, Crim LW (1999) The effect of stress on reproduction in Atlantic cod. J
Fish Biol 54:477–488
Naas KE, Næss T, Harboe T (1992) Enhanced first feeding of halibut larvae (Hippoglossus
hippoglossus L.) in green water. Aquaculture 105:143–156
Neat FC, Wright PJ, Zuur AF, Gibb IM, Gibb FM, Tulett D, Righton DA, Turner RJ (2006)
Residency and depth movements of a coastal group of Atlantic cod (Gadus morhua L.).
Mar Biol 148:643–654
Olsen SA, Hansen PK, Givskud H, Ervik A, Samuelsen OB (2014) Changes in fatty acid
composition and stable isotope signature of Atlantic cod (Gadus morhua) in response
to laboratory dietary shifts. Aquaculture 435:277–285
Olsen EM, Heupel MR, Simpfendorfer CA, Moland E (2012) Harvest selection on Atlantic cod
behavioral traits: implications for spatial management. Ecol Evol 2:1549–1562
Olsen EM, Moland E (2011) Fitness landscape of Atlantic cod shaped by harvest selection and
natural selection. Evol Ecol 25:695–710
Otterå H, Agnalt A, Jorstad K (2006) Differences in spawning time of captive Atlantic cod from
four regions of Norway, kept under identical conditions. ICES J Mar Sci 63:216–223
Otterå H, Skilbrei OT (2014) Possible influence of salmon farming on long-term resident
behaviour of wild saithe (Pollachius virens L.). ICES J Mar Sci 71:2484–2493
Patten MA, Kelly JF (2010) Habitat selection and the perceptual trap. Ecol Appl 20:2148–2156
Porter MJ., Duncan N., Mitchell D, Bromagea N. (1999) The use of cage lighting to reduce
plasma melatonin in Atlantic salmon (Salmo salar) and its effects on the inhibition of
grilsing. Aquaculture 176:237–244
R Core Team (2016) R: A language and environment for statistical computing.

77
Rahman MM, Gasparini C, Turchini GM, Evans JP (2014) Experimental reduction in dietary
omega-3 polyunsaturated fatty acids depresses sperm competitiveness. Biol Lett 10
Rainuzzo JR, Reitan KI, Olsen Y (1997) The significance of lipids at early stages of marine fish:
a review. Aquaculture 155:103–115
Reitan KI, Rainuzzo JR, Olsen Y (1994) Influence of lipid composition of live feed on growth,
survival and pigmentation of turbot larvae. Aquac Int 2:33–48
Remeš V (2003) Effects of exotic habitat on nesting success, territory density, and settlement
patterns in the blackcap (Sylvia atricapilla). Conserv Biol 17:1127–1133
Robertson BA, Hutto RL (2006) A framework for understanding ecological traps and an
evaluation of existing evidence. Ecology 87:1075–1085
Robichaud D, Rose GA (2004) Migratory behaviour and range in Atlantic cod: inference from
a century of tagging. Fish Fish 5:185–214Salze G, Tocher DR, Roy WJ, Robertson DA
(2005) Egg quality determinants in cod (Gadus morhua L.): egg performance and lipids
in eggs from farmed and wild broodstock. Aquac Res 36:1488–1499
Sargent J, Bell G, McEvoy L, Tocher D, Estevez A (1999) Recent developments in the essential
fatty acid nutrition of fish. Aquaculture 177:191–199
Sargent J, McEvoy L, Estevez A, Bell G, Bell M, Henderson J, Tocher D (1999) Lipid nutrition of
marine fish during early development: current status and future directions.
Aquaculture 179:217–229
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image
analysis. Nat Methods 9:671–675
Sclafani M, Taggart CT, Thompson KR (1993) Condition, buoyancy and the distribution of
larval fish: implications for vertical migration and retention. J Plankton Res 15:413–435
Sherley RB, Ludynia K, Dyer BM, Lamont T, Makhado AB, Roux J-P, Scales KL, Underhill LG,
Votier SC (2017) Metapopulation tracking juvenile penguins reveals an ecosystem-
wide ecological trap. Curr Biol 27:563–568
Shima JS, Swearer SE (2010) The legacy of dispersal: larval experience shapes persistence
later in the life of a reef fish. J Anim Ecol 79:1308–1314
Skilbrei OT, Otterå H (2016) Vertical distribution of saithe (Pollachius virens) aggregating
around fish farms. ICES J Mar Sci 73:1186–1195
Skjæraasen JE, Nilsen T, Kjesbu OS (2006) Timing and determination of potential fecundity in
Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 63:310–320
Vallin L, Nissling A (2000) Maternal effects on egg size and egg buoyancy of Baltic cod, Gadus
morhua: implications for stock structure effects on recruitment. Fish Res 49:21–37

78
Svåsand T (1990) Comparisons of migration patterns of wild and recaptured reared coastal
cod, Gadus morhua L., released in a small fjord in western Norway. Aquac Res 21:491–
495
Uglem I, Bjørn PA, Dale T, Kerwath S, Økland F, Nilsen R, Aas K, Fleming I, McKinley RS (2008)
Movements and spatiotemporal distribution of escaped farmed and local wild Atlantic
cod (Gadus morhua L.). Aquac Res 39:158–170
Uglem I, Dempster T, Bjørn P-A, Sánchez-Jerez P, Økland F (2009) High connectivity of salmon
farms revealed by aggregation, residence and repeated movements of wild fish among
farms. Mar Ecol Prog Ser 384:251–260
Wasta Z, Mjøs SA (2013) A database of chromatographic properties and mass spectra of fatty
acid methyl esters from omega-3 products. J Chromatogr A 1299:94–102
Weldon AJ, Haddad NM (2005) The effects of patch shape on indigo buntings: evidence for
an ecological trap. Ecology 86:1422–1431
White CA, Bannister RJ, Dworjanyn SA, Husa V, Nichols PD, Kutti T, Dempster T (2017a)
Consumption of aquaculture waste affects the fatty acid metabolism of a benthic
invertebrate. Sci Total Environ 586:1170–1181
White CA, Dworjanyn SA, Nichols PD, Mos B, Dempster T (2016) Future aquafeeds may
compromise reproductive fitness in a marine invertebrate. Mar Environ Res 122:67–75
White CA, Nichols PD, Ross DJ, Dempster T (2017b) Dispersal and assimilation of an
aquaculture waste subsidy in a low productivity coastal environment. Mar Pollut Bull
120:309-321
White CA, Woodcock SH, Bannister RJ, Nichols PD (2017) Terrestrial fatty acids as tracers of
finfish aquaculture waste in the marine environment. Rev Aquac. Early view:
https://doi.org/10.1111/raq.12230

79
CHAPTER FOUR: NATIVE PREDATOR PREVENTS AN INVADER FROM EXPLOITING
FOOD-RICH HABITAT
ABSTRACT
Biodiverse ecosystems may be inherently resistant to invasion, but environmental change can
facilitate invasion by disturbing natural communities and providing resources that are
underutilised by native species. In such cases, abundant native predators may be crucial in
limiting invasive population growth. We studied invasive seastars feeding at shellfish farms
together with a larger native predatory seastar, to examine whether farms are reproductive
hotspots for the invader or whether the native predator causes farms to function as ecological
traps. Invaders were not more abundant at farms, despite individuals residing at farms having
higher body condition and reproductive investment than those on reference habitats. The
native predator was 25x more abundant at farms than surrounding habitats and was
negatively correlated with invader abundance. We also observed several native-on-invasive
predation events and a conspicuous absence of small invaders at farms despite high larval
recruitment to farms, consistent with biotic control by the native predator. In a choice
experiment, invaders were strongly attracted to mussels, but the presence of a single native
predator nullified the attractive effect, indicating that the invader recognises and avoids the
native predator. Attractive habitats add spatial structure and may leave invasive populations
more susceptible to top-down control. We provide evidence that the presence of a native
predator prevents an invader from exploiting an attractive rich food resource.
INTRODUCTION
Invasive species are economically costly and an important contributor to biodiversity loss
worldwide (Clavero and García-Berthou, 2005; Pimentel et al., 2001). Species introductions are
increasing in frequency as a result of human transport, with the likelihood of a newly
introduced species establishing and becoming invasive depending on multiple interacting
factors including environmental conditions, ecosystem characteristics and species traits
(Carlton, 1996; Keane and Crawley, 2002; Kennedy et al., 2002; Kimbro et al., 2013; Papacostas
et al., 2017).

80
Intact, biodiverse ecosystems sometimes have inherent biotic invasion resistance (Elton,
1958), conferred by low niche vacancy rates and strong competition for resources (Kennedy et
al., 2002), as well as a higher likelihood of the new arrival encountering predators or other
enemy species (Keane and Crawley, 2002; Kimbro et al., 2013). However, ecosystem-level
invasion resistance is being eroded by large scale habitat disturbance and loss of biodiversity
associated with human-induced rapid environmental change (HIREC: Vitousek et al. 1997;
Sanderson et al. 2002; Halpern et al. 2008; Vörösmarty et al. 2010; Sih et al. 2011). Non-native
species can benefit from the impacts of HIREC by taking advantage of man-made or degraded
habitats that are low in biodiversity and have resources that are underutilised by native
competitors or predators (Byers, 2002; Didham et al., 2005; MacDougall and Turkington, 2005;
Mack et al., 2000). In the early stages of invasion, or at range limits, these refuge habitats can
become invasion hubs: satellite populations that facilitate persistence and expansion (Carlton,
1996; Letnic et al., 2014; Russell et al., 2011; Suarez et al., 2001; With, 2002). Later, they may
function as reproductive hotspots for both native and invasive species, for example by
permitting high local population densities that overcome Allee effects (Inglis & Gust 2003; Ling
et al. 2012).
A single abundant predator species can exert considerable top-down control and play an
important role in limiting the eventual population size and distribution of an invader,
essentially providing invasion resilience when invasion resistance has failed (DeRivera et al.,
2005; Kimbro et al., 2013; Papacostas et al., 2017). Predator-mediated biotic control may be
particularly effective in cases where the invader does not recognise or respond appropriately
to the native predator; a scenario that is most likely to occur when the invader has no
evolutionary history with the native predator or its relatives (Cox and Lima, 2006; Pintor and
Byers, 2015; Sih et al., 2010). Such conditions favour the formation of an evolutionary trap:
where an individual chooses a habitat (ecological trap: Robertson & Hutto 2006; Hale &
Swearer 2016) or behaviour that leads to poor fitness outcomes (Robertson et al., 2013). Traps
usually arise as a result of environmental change outpacing the evolution of behaviour, and
account for some of the disastrous effects of invasive predators on naïve native prey (e.g.
juvenile fish avoid reefs with a resident native predator but not those with a resident invasive
predator: Benkwitt 2017). Most work has focused on mitigating traps that affect native
species, but evolutionary traps also have potential as tools in the management of non-native
species (Letnic et al., 2015; Robertson et al., 2017). One potential approach is to increase
mortality rates at attractive sites to create attractive sinks – Letnic et al. (2015) recently
demonstrated that excluding invasive toads from water sources converts invasion hubs into

81
ecological traps. Native predators may also contribute to the formation of ecological traps for
non-native species, but such traps have not yet been empirically demonstrated.
In this study, we employ the ecological trap framework to study interactions between native
and invasive seastars at shellfish farms in southern Australia. To demonstrate an ecological
trap, it is necessary that individuals either prefer or fail to avoid the putative trap habitat when
better options are available, and that fitness is reduced in the putative trap habitat (Robertson
and Hutto, 2006). Shellfish farms are an excellent model for a range of anthropogenic marine
habitats worldwide that are simultaneously highly disturbed and rich in food for benthic
predators (D’Amours et al., 2008; Inglis and Gust, 2003; McKindsey et al., 2011). This
combination of traits may cause such habitats to be highly vulnerable to exploitation by
invasive species (e.g. Ling et al. 2012). The invasive seastar Asterias amurensis (Asterias
hereafter) was introduced to Tasmania during the 1980s (Grannum et al., 1996), and mainland
Australia in 1995 (Parry and Cohen, 2001), with the mainland population reaching 75 million by
2000 (Parry and Cohen, 2001). The invader has decimated naïve bivalve populations (Hutson et
al., 2005; Ross et al., 2003), and has few predators in its new environment, but there have
been reports of predation by a native predator, the eleven-arm seastar Coscinasterias
muricata (Coscinasterias hereafter) (Byrne et al., 2013; Parry, 2017). The trophic subsidy
provided by shellfish farms attracts aggregations of both species, but the ability of the invader
to benefit from the high food availability will depend on competition and predation risk from
the native predator (Appendix 4.1). If the native predator does not impede the ability of the
invader to access the food resource, then such habitats are likely to act as population sources
(Letnic et al., 2014; Ling et al., 2012). Alternatively, if the invader is attracted to these sites but
then suffers high predation rates or competitive exclusion, there is potential for an ecological
trap to arise (Hale and Swearer, 2016). We combine data on population distribution, individual
body condition, reproductive investment and habitat preference to assess the likely role of
two coastal mussel farms for populations of the native and invasive seastars. In doing so, we
provide evidence indicating that an abundant native predatory seastar (a) prevents an
established invader from accessing a disturbed but food-rich habitat.

82
METHODS
Study sites and focal species
Port Phillip Bay is a 1930 km2 semi-enclosed marine water body in south-eastern Australia
subject to multiple impacts from invasive species and urban inputs (Hewitt et al., 2004;
Sampson et al., 2014). We studied native (Coscinasterias) and invasive (Asterias) seastar
populations at two shellfish aquaculture reserves 12 km apart in the south-western part of the
bay: Clifton Springs and Grassy Point. These leases primarily produce blue mussels (Mytilus
galloprovincialis) grown on suspended longlines. Clifton Springs and Grassy Point contain
approximately 81 and 42 ha of active bivalve aquaculture areas, respectively. Active areas
consist of parallel longlines approximately 20 m apart, which are stocked year-round with
short turnaround times between harvesting and restocking. Live mussels (typically 40-60 mm
shell length) were abundant on the seabed throughout the study (pers. obs.), with the natural
substrate entirely replaced by mussel shell debris within active areas. Both reserves are
located on soft sediment between 8-14 m depth, with constant mixing and exchange of water
by tidal cycling and wind-generated currents. Clifton Springs is the more sheltered of the two
locations and is dominated by silty substrate, while Grassy Point is characterised by sandy
substrate and lower turbidity. The natural benthos within farms and surrounding areas at both
locations is dominated by Halophila australis seagrass beds, with paper mussels (Electroma
georgiana) attached to available seagrass or macroalgae. Caulerpa spp. beds are common at
Grassy Point. Seastars are the predominant benthic predators within and around both
reserves.
Coscinasterias muricata (Asteroidea: Asteriidae) are a large (armspan commonly 30-45 cm)
subtidal reef dwelling invertivore. Bivalves are preferentially consumed, especially the mussels
that are abundant on reefs and marine infrastructure in southern Australia, but will
opportunistically consume a variety of invertebrate prey (Day et al., 1995; Parry, 2017).
Asterias amurensis (Asteroidea: Asteriidae) are smaller (armspan commonly 20-30 cm) but
highly mobile invertivores, and are typically more abundant on soft sediment habitats where
they consume primarily bivalve prey (e.g. clams, scallops and mussels) (Ross et al., 2003; Ross
and Johnson, 2002).

83
Experiment 1: Effect of shellfish farming on population density of native and invasive
seastars
We surveyed Asterias and Coscinasterias population densities along diver transects inside and
outside the two aquaculture reserves: Clifton Springs on six occasions between July 2014 and
June 2016, and Grassy Point on seven occasions between July 2014 and June 2015. On each
sampling occasion, we surveyed three replicate 2 x 25 m transects (extended to 2 x 50 where
<10 seastars were recorded) inside and outside the reserve boundaries (a total of six transects
per reserve per sampling date). ‘Inside’ transects were haphazardly placed within active
mussel longline areas where live mussels were present on the seabed. ‘Outside’ transects were
haphazardly placed on natural soft sediment habitat within a ~6 km2 area 1-2 km outside the
reserve boundary. Inside and Outside habitats were comparable in terms of depth (10-13 m),
water flow and sediment characteristics. Each transect consisted of the diver laying out a
transect centreline on a predetermined random heading and counting all seastars with their
central point within 1 m of the transect centreline. On rare occasions where a transect strayed
beyond the area of fallen mussel debris (typically >20 m from the outermost longline) we
altered the heading of the remainder of transect. We placed transects at least 50 m apart to
ensure that no seastars were sampled twice.
To test for effects of shellfish farms on seastar population density, we constructed generalised
linear mixed models (GLMMs) for each species using the glmmTMB package for R (Brooks et
al., 2017; R Core Team, 2017). A negative binomial model family was specified as there was a
higher frequency of low or zero counts than would be contained within a Poisson distribution.
We included Habitat (Inside or Outside) and Location (Clifton Springs or Grassy Point) as fixed
effects and Date as a random intercept term to account for temporal variation. As there was a
very strong effect of Location (largely driven by the rarity of Asterias at Grassy Point), we then
split the dataset by Location and re-ran the models without the Location term, reporting both
sets of results. In each case, we tested the significance of the Habitat effect by comparing the
fit of models with and without the Habitat term using a likelihood ratio test. To test for a
potential effect of Coscinasterias abundance on Asterias abundance, we fitted a negative
binomial GLMM in which Asterias density is regressed against Coscinasterias density, with
Habitat and Location also as fixed effects and Date as a random intercept term.

84
Experiment 2: Effect of shellfish farming on size and condition of native and invasive seastars
To track any differences in size between habitats, we measured the arm span of all seastars
encountered on each transect (or the first 30 of each species) and took the mean arm span for
each species on each transect.
We compared condition metrics between habitats by collecting up to 20 individuals per
species per farm per sampling occasion (collected on transects and supplemented with off-
transect collections when necessary). When fewer than 20 individuals were available in one
habitat on a given day, we limited our sample size in the other habitat to avoid temporal bias
caused by seasonal dynamics in condition metrics. Drained and gutted carcass weight provided
a measure of size that was largely independent of body condition, as seastars store nutrients
primarily in the coelomic fluid and digestive organs. The relative size of the pyloric caeca
(digestive organs that increase in size with feeding activity) provided a measure of body
condition and an indirect measure of food availability inside and outside the aquaculture
reserves (Oudejans et al., 1979; Xu and Barker, 1990). We calculated the pyloric caeca index
relative to somatic weight: PCI = PCW/CW, where PC = pyloric caeca wet weight and CW =
gutted carcass weight drained of coelomic fluid. We considered reproductive condition in
terms of two correlated metrics: relative reproductive investment (gonadosomatic index: GSI =
GW/CW, where GW = gonad weight) and potential fecundity (gonad size). A pilot study
demonstrated that PCI and GSI can be accurately estimated from a single limb (whole-body vs.
limb-only linear R2 with n = 20 for Asterias PCI = 0.79, GSI = 0.90; with n = 23 for Coscinasterias
PCI = 0.88, GSI = 0.78). This permitted the release of the native Coscinasterias after the
removal of a single limb. As Asterias is a declared noxious aquatic species, entire Asterias
individuals were taken, but only a single limb dissected. Asterias are sufficiently abundant and
mobile that the removal of up to 20 individuals per farm per sampling occasion likely had a
negligible effect on numbers in later surveys (Andrews et al., 1996). Limbs were cut open
longitudinally, drained of coelomic fluid and weighed. The pyloric caeca and gonads were then
removed and weighed separately. Individuals were sexed based on the colour of the gonads, a
method that was validated by microscopy during a pilot study (n = 20 per species, 100 %
accuracy).
To test for effects of shellfish farms on size and condition, we fitted linear mixed models with
Habitat as a fixed term and Date and Sex as a random intercept term. We tested the Habitat

85
term as per Experiment 1. Responses were transformed as necessary to improve linearity and
normality: log10 for Asterias armspan and 4th root for PCI and GSI.
Experiment 3: Interactions between native and invasive seastars
Any fitness benefits of high food availability may be offset if there is an elevated predation risk.
We recorded evidence of predation on transects as for population surveys in Experiment 1,
including predation events in progress and sublethal damage that is unlikely to be explained by
asexual reproduction (e.g. damage to arms consistent with crab or fish predation: Ling &
Johnson 2013). Predation may influence the size distribution of a prey species whenever
predation events and/or success rates depend on the relative size of the predator and prey.
Coscinasterias predation on Asterias is strongly size dependent, with a higher likelihood of prey
escaping when relative sizes are close (Parry 2017). Cannibalism of small Asterias by larger
conspecifics also occurs (pers. obs.). In light of this, we compared the size distributions of
Asterias inside and outside farms for evidence of size-dependent predation within and/or
avoidance of the farm environment.
Given evidence that the native seastar does prey on the invasive seastar, we devised a
laboratory-based choice experiment to test the strength of the invader’s attraction to fallen
mussels, and whether this attraction is moderated by the presence of either conspecifics or
native predators. We conducted the experiment in a 72 L flume tank (120 x 30 x 20 cm),
housed within a recirculating seawater facility containing ca. 12000 L of continuously filtered
and UV-treated seawater. A flow rate of 4 L min-1 was divided between the two chambers at
the head of the tank, with both chambers opening into an undivided downstream portion of
the tank containing the Asterias subject. We compared rates of attraction to four treatments:
a seawater control with no predatory nor prey cues (‘SW’), five thawed mussels (40-60 mm
shell length) placed in a perforated compartment at the upstream end of the chamber (‘M’),
five thawed mussels with three conspecifics in the chamber with the mussels (‘AM’), and five
thawed mussels with a large Coscinasterias individual also present in the chamber (‘CM’). All
seastars placed in the treatment chamber for AM and CM treatments remained by the end of
the trial. We applied treatments singly, with the cue randomly assigned to one of the two
chambers (all treatments were tested relative to a seawater control). We left the subjects
overnight (16 hrs), and considered them to be attracted to the habitat cue if they were present
inside the corresponding chamber at the completion of the trial, providing a binary outcome.

86
The flume tank was cleaned and flushed between trials. Subjects were randomly selected from
a large captive population (>200) collected from the wild within 2 months of the trial, fasted
for 2-4 days prior to the trial, and randomly assigned to a treatment. The size range of subjects
was 4-17 cm in every treatment group (mean 11 ± 0.2 cm).
We tested for differential attraction to treatments by fitting a binomial generalised linear
model using the glm function in R, before conducting post-hoc pair-wise X2 tests of proportions
using the prop.test function.
RESULTS
Experiment 1: Effect of shellfish farming on population density of native and invasive
seastars
The invasive seastar Asterias was common on silty substrate and mussel shell debris both
inside and outside the farm boundary at Clifton Springs, with very high abundance inside
throughout winter 2014 (Fig. 4.1). However, Asterias population density was highly variable,
with no significant effect of the farm habitat over the full study duration (2014-2016) (Table
4.1). Asterias was nearly absent from Grassy Point, being recorded on a single transect only
inside the Grassy Point boundary in winter 2014. (Fig. 4.1).
The native seastar Coscinasterias showed contrasting habitat preference (Fig. 4.1). At Grassy
Point, Coscinasterias was common inside and outside the Grassy Point farm boundary, with a
14x higher population density inside (Table 4.1). They were also common inside the boundary
at Clifton Springs, but absent on our transects outside the Clifton Springs boundary (Fig. 4.1).
Across the two locations, Coscinasterias occurred at a higher density (mean 25x) inside farms
on every sampling occasion between 2014-2016.
If both species were distributed according to availability of a common food source, we would
expect a positive correlation between the abundance of the two species. Instead, our
population density data reveal that Asterias density is negatively dependent on Coscinasterias
density (negative binomial mixed effects model with Coscinasterias density, Habitat and
Location as fixed effects and Date as random effect: X2 = 4.1, p = 0.04).

87
Experiment 2: Effect of shellfish farming on size and condition of native and invasive seastars
Both seastar species were larger on average inside farm boundaries. Due to the rarity of
Asterias outside Grassy Point and the rarity of Coscinasterias outside Clifton Springs, we
compared the size distribution of both species inside and outside a single farm (Asterias:
Clifton Springs, Coscinasterias: Grassy Point).
Figure 4.1. Population density of Asterias amurensis (plots A and B) and Coscinasterias
muricata (plots C and D) inside and outside the Clifton Springs and Grassy Point Fisheries
Aquaculture Reserves. Boxes denote median, lower (25 %) and upper (75 %) quartiles,
whiskers denote 1.5x interquartile range.
Asterias inside the farm had a 25 % wider arm span than their counterparts outside,
corresponding to a 92 % increase in gutted weight (Table 4.1). This effect was consistent across

88
all but one sampling date (Fig. 4.2). The difference in mean arm span of Asterias was driven by
a contraction at the lower end of the size distribution inside the Clifton Springs farm, rather
than the presence of larger individuals (Fig. 4.4). All body and reproductive condition metrics
that we assessed in Asterias were higher inside the farm than outside: drained weight was 77
% higher, gonad weight 57 % higher, PCI 26 % higher and GSI 57 % higher (Table 4.1). The PCI
effect was highly consistent over the study duration, while differences in GSI varied over time
(Fig. 4.3).
Figure 4.2. Armspan (A) and gutted weight (B) of Asterias amurensis inside and outside the
Clifton Springs Fisheries Aquaculture Reserve.
Likewise, Coscinasterias inside the farm had a 7 % wider arm span and 42 % heavier gutted
weight than those outside (Table 4.1). Individuals inside the farm also had higher body
condition (11 %) but there was no difference in either gonad size or GSI (Table 4.1).
Experiment 3: Interactions between native and invasive seastars
We observed three native-on-invasive seastar predation events on our transects inside the
Clifton Springs farm (total 3325 m2 surveyed, 160 Asterias scored), with none observed on the
same number of transects outside (3900 m2 surveyed, 126 Asterias scored). We also noted a
further ten predation events off-transect, as well as five skeletons of recently-consumed

89
Asterias, all inside the farm boundary. We observed one cannibalism event (a large Asterias
consuming a smaller conspecific), also on a transect within the Clifton Springs farm.
We found strong effects of prey and predator cues on habitat selection decisions by the
invasive Asterias in captive behavioural trials (n = 40 per treatment, p <0.0001) (Fig. 4.5). The
mussel cue (M) attracted Asterias subjects at a significantly higher rate than the seawater
control (SW) (93 vs. 28 % respectively, X2 = 32, p <0.0001). Mussels remained more attractive
than seawater when three Asterias conspecifics were present feeding on the mussels (AM) (88
%, X2 = 27, p <0.0001), and there was no difference between rates of attraction to M or AM
cues (93 % vs. 88 % respectively, X2 = 0.1, p = 0.7). In contrast, the presence of a single
Coscinasterias individual feeding on the mussels (CM) resulted in a dramatic decline in
attraction rates relative to both the M (28 vs. 93 % respectively, X2 = 32, p <0.0001) and AM
cues (28 vs. 88 % respectively, X2 = 27, p <0.0001). 2/11 individuals that selected the chamber
with the CM cue were predated by the Coscinasterias (one fully, one partially consumed
overnight). Asterias individuals that avoided the chamber with the CM cue exhibited a variety
of behaviours, such as moving immediately to the cue-free (clean seawater) chamber and
remaining there for the duration of the study, circling the downstream portion of the tank, or
attempting to reach the mussels before evading a predation attempt by Coscinasterias. We
found no evidence that the size of the subject influenced their response to the presence of
either conspecifics (z38 = 1.1, p = 0.3) or Coscinasterias (z38 = 1.0, p = 0.3). The size differential
between the subject and the three conspecifics in the AM treatment (natural log of subject
arm span / mean conspecific arm span) also did not influence the likelihood of attraction to
the mussels (z38 = 0.6, p = 0.5).
DISCUSSION
In disturbed ecosystems, abundant native predators may be an important line of defence
against invaders, limiting the probability of establishment and subsequent population growth.
In Experiment 1 we showed that a native predator occurs at 25x higher abundance inside two
shellfish farms relative to reference habitats, with the shellfish farm accounting for almost the
entire population in an area with non-preferred silty substrate. In contrast, the invasive species
was not significantly more abundant inside the farms and was negatively associated with the
native predator. Experiment 2 revealed that invasive seastars residing inside one farm
possessed considerably higher body condition and reproductive condition metrics than

90
conspecifics at reference sites, supporting the hypothesis that this shellfish farm may be a
reproductive hotspot for the invader. When contrasted with the lack of a clear effect on
abundance in Experiment 1, this suggests a failure or inability to take advantage of a high
quality food resource. Accordingly, in Experiment 3 we provided observational field evidence
of elevated predation risk for the invasive species inside shellfish farms, and found that in
captivity, the invader recognises and avoids the native predator even if it means forgoing a
favoured prey item. Taken together, these findings raise the possibility that the presence of
the native predator is preventing the invader from accessing a highly disturbed food-rich
habitat.
Figure 4.3. Condition metrics for Asterias amurensis inside and outside the Clifton Springs
Fisheries Aquaculture Reserve: drained carcass weight (A), gonad weight (B), gonadosomatic
index (C), and pyloric caeca index (D).
In the absence of abundant native predators, man-made habitats such as shellfish farms, fish
farms and mussel-fouled infrastructure can function as reproductive hotspots for invasive

91
Asterias amurensis populations, simultaneously supporting orders-of-magnitude higher
population density and elevated individual reproductive output (Ling et al. 2012). Within their
native range, Asterias species also have highly elevated population densities at shellfish farms,
perhaps indicating release from native predators such as crabs and sunstars (e.g. Olaso Toca
1979, 1982; Saranchova & Kulakovskii 1982, cited in McKindsey et al. 2011; D’Amours et al.
2008). The present study indicates that top-down control may alter the role played by these
food-rich habitats.
Figure 4.4. Density plot of Asterias amurensis size distribution inside and outside the farm
boundary at Clifton Springs Aquaculture Fisheries Reserve. N = 165 (inside) + 107 (outside).
Effective top-down control of non-native animals by native predators has been reported in
numerous marine, freshwater and terrestrial systems (e.g. DeRivera et al. 2005; Cheng & Hovel
2010; Tetzlaff et al. 2011; Freed & Leisnham 2014), but to our knowledge, this is the first clear
evidence of a native predator preventing invaders from accessing a high value food resource.
Asterias is thought to be primarily food limited in its new range: growth rates declined during
the first three years following its introduction to mainland Australia, coinciding with increasing
population densities and large declines in prey abundance (Parry and Cohen, 2001). Our body
condition and reproductive investment data show elevated condition inside farms, indicating
that food limitation still exists outside farm habitats. This means that regardless of the
frequency of predation events, if Coscinasterias discourage Asterias from accessing farms, the

92
native predator may exert indirect biotic control on the invasive population by maintaining
food limitation and thus a lower equilibrium population size.
Figure 4.5. Effect of prey, conspecifics and predators on habitat selection decisions by the
invasive seastar Asterias amurensis in laboratory trials. Treatments: SW = seawater control, M
= mussels, AM = mussels + Asterias conspecifics, CM = mussels + Coscinasterias. Bars show the
proportion of subjects that are attracted to the cue chamber (‘Y’) or not (‘N’) according to the
treatment. Individuals that were not attracted to the cue chamber either remained in the
downstream portion or moved into the control chamber. N = 160 (40 per treatment). Matching
letters indicate proportions that do not differ statistically.
Our data also raise the possibility that juveniles of the non-native species may be falling into a
predation-mediated ecological trap at shellfish farms. Asterias has a long pelagic larval
duration (79-112 days: Bruce et al., 1995), and in Australia, planktonic Asterias recruit in large
numbers directly to mussel lines and other suspended structures (e.g. Dommisse & Hough
2004). This creates the conditions for an ecological trap affecting juvenile Asterias, as juveniles
living on mussel lines are (a) unlikely to reach sexual maturity before the mussel lines are
harvested, and (b) likely to experience high predation risk if they fall to a benthic environment
populated by >2000 Coscinasterias ha-1 (this study). The truncated size distribution for Asterias
on the seabed under mussel farms is consistent the hypothesis that juvenile Asterias falling to
the seabed from mussel lines either leave the area or are consumed, although we cannot rule
out individuals leaving or avoiding the farm environment due to innate preferences (such as

93
for soft sediment rather than biofouled mussel shell substrate). The risk of predation and
potential for an ecological trap may be increased if juveniles do not have sufficiently strong
predator avoidance behaviour (Cox and Lima, 2006; Sih et al., 2010); in our habitat choice
experiment, small individuals were no more likely to avoid the native predator than large
individuals, despite being at higher risk of predation than larger conspecifics (size-dependent
predation success: Parry 2017).
As adults, the potential for an ecological trap at shellfish farms depends on whether individuals
are attracted to the farm (i.e. the strength of their preference for bivalve prey versus their
avoidance response to the native predator) and the net fitness effects of any preference for
the farm (i.e. high food availability versus high predation risk). Our choice experiment found
that Asterias were strongly attracted to bivalves, but only in the absence of the native
predators, and while actual predation rates are almost certainly higher inside the farms (this
study; Parry 2017), so is reproductive output (this study; Ling et al. 2012). Further work is
needed to calculate the net effect of these opposing factors, and to test whether Asterias
individuals are able to make informed habitat selection decisions at ecologically-relevant
distances. The high densities of mussels and predators at farms provide a relatively strong
scent trail, and all else being equal, even random movement of seastars will tend to result in
higher densities in preferred habitats if individuals move less after arrival (Benhamou 2011;
see also elevated density of Coscinasterias inside farms, this study).
Invading Asterias benefitted from the naivety of Australian bivalves (Hutson et al., 2005; Ross
et al., 2003), but to date there has been little evidence that Asterias responds naïvely to novel
predators, despite theoretical expectations arising from a lack of shared evolutionary history
(Cox and Lima, 2006; Sih et al., 2010). Behavioural data on predator-prey interactions in the
early days of invasion are not available, but the establishment and expansion of Asterias in
Australia may have been assisted by pre-existing predator cues evolved in response to
functionally similar predators in its native range (Sih et al., 2010). In particular, the sun star
Solaster paxillatus and king crab Paralithodes camtschaticus of the northern Pacific region are
notably similar in form and functional role to the major invertebrate predators in its new
range: the seastar Coscinasterias and the spider crab Leptomithrax gaimardii (Byrne et al.,
2013; Ling and Johnson, 2013), and may emit similar sensory cues.
Little is known about how the Asterias invasion affected native predators. A meta-analysis by
Pintor and Byers (2015) found that simultaneous access to native and non-native prey was
associated with increased native predator abundance, but exceptions are common where
native and non-native species rely on a shared food resource or where abundant non-native

94
species drive severe ecological change (e.g. Pothoven et al. 2001; Suarez & Case 2002). Here,
the invader competes with the native species for mussel prey, but localised high densities of
mussels, such as those on rocky reefs, shellfish farms and marine infrastructure may suit the
native species by improving their ability to exclude Asterias from the food resource, as well as
supplement the mussel diet with opportunistic intraguild predation of the invader.
Eradication of this invader is not feasible, but any top-down pressure may have ecological
benefits by partially releasing vulnerable prey populations from predation and reducing the
likelihood of new populations establishing (DeRivera et al., 2005; Letnic et al., 2009; Mack et
al., 2000). Conversely however, in cases where a non-native prey species provides a large
trophic subsidy for native predators, native predators can become overabundant and severely
impact their native prey species (Noonburg and Byers, 2005). It is unclear whether the
presence of Asterias at farms is sufficient to maintain locally elevated Coscinasterias
populations, but such predation events could conceivably be important during rare periods
when preferred prey (mussels) are scarce.
A range of harvesting methods have been employed to control seastars globally, but none are
cost-effective (Barkhouse et al., 2007). In particular, a lack of spatial population structure on
homogeneous landscapes limits the efficiency of control methods, but attractive habitats such
as shellfish farms and other marine infrastructure add spatial structure and may leave invasive
populations more vulnerable if mortality can be induced at such sites (Bascompte et al., 2002;
Letnic et al., 2015; Russell et al., 2011). In our study, biotic control by the native predator
apparently limits the ability of the invader to exploit a food-rich resource, and we provide
evidence that these habitats may function as ecological traps for the invader by combining
attractive cues from bivalve prey with a high predation risk from the native predator. In such a
case, the native predator provides a potentially valuable ecosystem service by suppressing
invasive populations. It may be feasible to improve the impact of this service by protecting and
augmenting predator populations in key areas such as localised sites of new introductions
(Parry, 2017); an approach has been used with some success in other invaded marine
environments (Atalah et al., 2016, 2013).
ACKNOWLEDGMENTS
This work was funded by a Holsworth Wildlife Research Endowment grant awarded to LB.
Lance Wiffen provided access to aquaculture leases, while Chris Taylor, Simon Reeves, Emily

95
Fobert, Dean Chamberlain, Jack O'Connor, Ben Cleveland, Kevin Jensen, Kevin Menzies, Oliver
Thomas and Rod Watson (Victorian Marine Science Consortium) assisted with fieldwork.
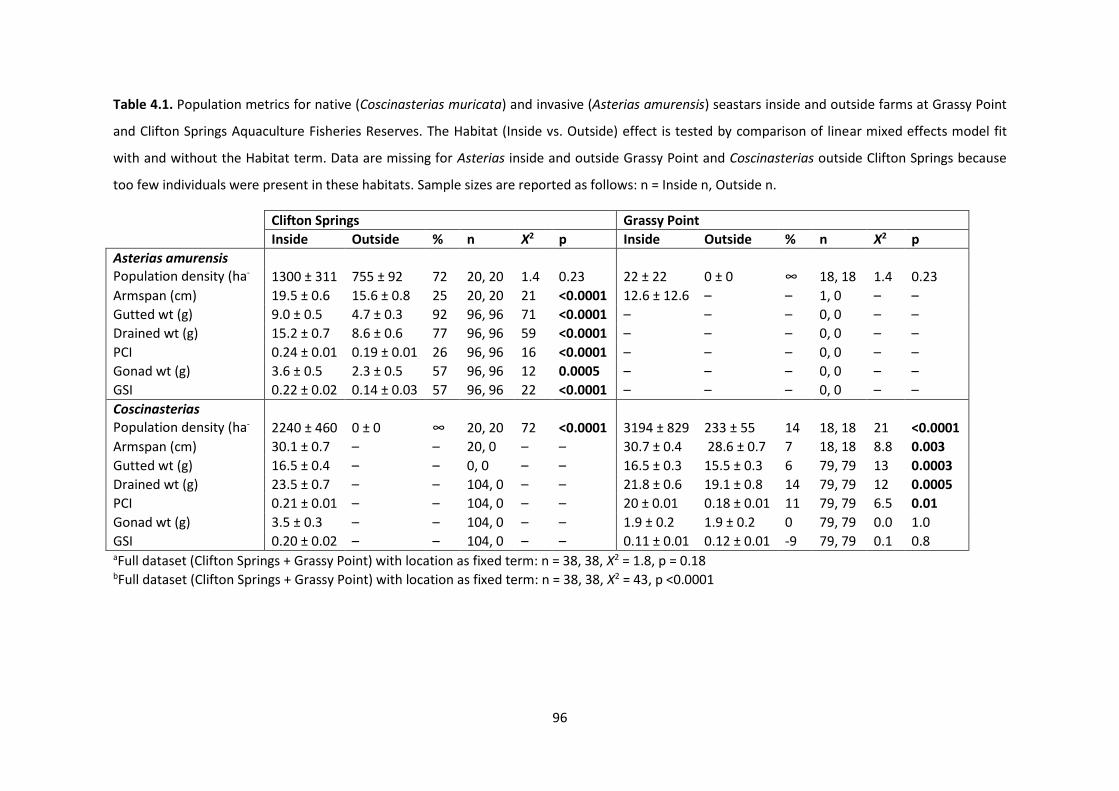
96
Table 4.1. Population metrics for native (Coscinasterias muricata) and invasive (Asterias amurensis) seastars inside and outside farms at Grassy Point
and Clifton Springs Aquaculture Fisheries Reserves. The Habitat (Inside vs. Outside) effect is tested by comparison of linear mixed effects model fit
with and without the Habitat term. Data are missing for Asterias inside and outside Grassy Point and Coscinasterias outside Clifton Springs because
too few individuals were present in these habitats. Sample sizes are reported as follows: n = Inside n, Outside n.
Clifton Springs Grassy Point
Inside Outside % n X2 p Inside Outside % n X2 p
Asterias amurensis Population density (ha-
1)a 1300 ± 311 755 ± 92 72 20, 20 1.4 0.23 22 ± 22 0 ± 0 ∞ 18, 18 1.4 0.23
Armspan (cm) 19.5 ± 0.6 15.6 ± 0.8 25 20, 20 21 <0.0001 12.6 ± 12.6 – – 1, 0 – –
Gutted wt (g) 9.0 ± 0.5 4.7 ± 0.3 92 96, 96 71 <0.0001 – – – 0, 0 – –
Drained wt (g) 15.2 ± 0.7 8.6 ± 0.6 77 96, 96 59 <0.0001 – – – 0, 0 – –
PCI 0.24 ± 0.01 0.19 ± 0.01 26 96, 96 16 <0.0001 – – – 0, 0 – –
Gonad wt (g) 3.6 ± 0.5 2.3 ± 0.5 57 96, 96 12 0.0005 – – – 0, 0 – –
GSI 0.22 ± 0.02 0.14 ± 0.03 57 96, 96 22 <0.0001 – – – 0, 0 – –
Coscinasterias muricata
Population density (ha-
1)b 2240 ± 460 0 ± 0 ∞ 20, 20 72 <0.0001 3194 ± 829 233 ± 55 14 18, 18 21 <0.0001
Armspan (cm) 30.1 ± 0.7 – – 20, 0 – – 30.7 ± 0.4 28.6 ± 0.7 7 18, 18 8.8 0.003
Gutted wt (g) 16.5 ± 0.4 – – 0, 0 – – 16.5 ± 0.3 15.5 ± 0.3 6 79, 79 13 0.0003
Drained wt (g) 23.5 ± 0.7 – – 104, 0 – – 21.8 ± 0.6 19.1 ± 0.8 14 79, 79 12 0.0005
PCI 0.21 ± 0.01 – – 104, 0 – – 20 ± 0.01 0.18 ± 0.01 11 79, 79 6.5 0.01
Gonad wt (g) 3.5 ± 0.3 – – 104, 0 – – 1.9 ± 0.2 1.9 ± 0.2 0 79, 79 0.0 1.0
GSI 0.20 ± 0.02 – – 104, 0 – – 0.11 ± 0.01 0.12 ± 0.01 -9 79, 79 0.1 0.8 aFull dataset (Clifton Springs + Grassy Point) with location as fixed term: n = 38, 38, X2 = 1.8, p = 0.18 bFull dataset (Clifton Springs + Grassy Point) with location as fixed term: n = 38, 38, X2 = 43, p <0.0001

97
REFERENCES
Andrews, D., Whayman, G., Edgar, G., 1996. Assessment of optimal trapping techniques to
control densities north Pacific seastars on marine farm leases. Tasmanian Department of
Primary Industries and Fisheries, Hobart.
Atalah, J., Hopkins, G.A., Forrest, B.M., 2013. Augmentative biocontrol in natural marine
habitats: persistence, spread and non-target effects of the sea urchin Evechinus
chloroticus. PLoS One 8, 1–10
Atalah, J., Newcombe, E.M., Zaiko, A., 2016. Biocontrol of fouling pests: effect of diversity,
identity and density of control agents. Mar. Environ. Res. 115, 20–27
Barkhouse, C., Niles, M., Davidson, L.-A., 2007. A literature review of sea star control methods
for bottom and off bottom shellfish cultures. Can. Ind. Rep. Fish. Aquat. Sci. 279, vii + 35p.
Bascompte, J., Possingham, H., Roughgarden, J., 2002. Patchy populations in stochastic
environments: critical number of patches for persistence. Am. Nat. 159, 128–137
Benhamou, S., 2011. Dynamic approach to space and habitat use based on biased random
bridges. PLoS One 6, e14592
Benkwitt, C.E., 2017. Predator effects on reef fish settlement depend on predator origin and
recruit density. Ecology 98, 896–902
Brooks, M.E., Kristensen, K., van Benthem, K.J., Magnusson, A., Berg, C.W., Nielsen, A., Skaug,
H.J., Maechler, M., Bolker, B.M., 2017. Modeling zero-inflated count data with glmmTMB.
bioRxiv 132753. doi:10.1101/132753
Bruce, B.D., Sutton, C.A., Lyne, V.D., 1995. Laboratory and field studies of the larval distribution
and duration of the introduced seastar Asterias amurensis with updated and improved
prediction of the species spread based on a larval dispersal model: final report to
Fisheries Research and Development Corporation, Canberra.
Byers, J.E., 2002. Impact of non-indigenous species on natives enhanced by anthropogenic
alteration of selection regimes. Oikos 97, 449–458
Byrne, M., O’Hara, T.D., Lawrence, J.M., 2013. Asterias amurensis, in: Lawrence, J.M. (Ed.),
Starfish: Biology and Ecology of the Asteroidea. The John Hopkins University Press,
Baltimore, MA, pp. 174–180.
Carlton, J.T., 1996. Pattern, process, and prediction in marine invasion ecology. Biol. Conserv.
78, 97–106.
Cheng, B.S., Hovel, K.A., 2010. Biotic resistance to invasion along an estuarine gradient.
Oecologia 164, 1049–1059

98
Clavero, M., García-Berthou, E., 2005. Invasive species are a leading cause of animal
extinctions. Trends Ecol. Evol. 20, 110
Cox, J.G., Lima, S.L., 2006. Naiveté and an aquatic-terrestrial dichotomy in the effects of
introduced predators. Trends Ecol. Evol. 21, 674–680
D’Amours, O., Archambault, P., McKindsey, C.W., Johnson, L.E., 2008. Local enhancement of
epibenthic macrofauna by aquaculture activities. Mar. Ecol. Prog. Ser. 371, 73–84.
Day, R., Dowell, A., Sant, G., Klemke, J., Shaw, C., 1995. Patchy predation: foraging behaviour
of Coscinasterias calamaria and escape responses of Haliotis rubra. Mar. Freshw. Behav.
Physiol. 26, 11–33
DeRivera, C.E., Ruiz, G.M., Hines, A.H., Jivoff, P., 2005. Biotic resistance to invasion: native
predator limits abundance and distribution of an introduced crab. Ecology 86, 3364–3376
Didham, R.K., Tylianakis, J.M., Hutchison, M.A., Ewers, R.M., Gemmell, N.J., 2005. Are invasive
species the drivers of ecological change? Trends Ecol. Evol. 20, 470–474
Dommisse, M., Hough, D., 2004. Controlling the northern Pacific seastar (Asterias amurensis)
in Australia. Final report for the Australian Government Department of the Environment
and Heritage, Canberra.
Elton, C.S., 1958. The ecology of invasions by animals and plants., The ecology of invasions by
animals and plants. Methuen & Co. Ltd., London.
Freed, T.Z., Leisnham, P.T., 2014. Roles of spatial partitioning, competition, and predation in
the North American invasion of an exotic mosquito. Oecologia 175, 601–611
Grannum, R.K., Murfet, N.B., Ritz, D.A., Turner, E., 1996. The distribution and impact of the
exotic seastar, Asterias amurensis (Lutken) in Tasmania.
Hale, R., Swearer, S.E., 2016. Ecological traps: current evidence and future directions. Proc. R.
Soc. B Biol. Sci. 283, 494–499
Halpern, B.S., Walbridge, S., Selkoe, K.A., Kappel, C. V, Micheli, F., D’Agrosa, C., Bruno, J.F.,
Casey, K.S., Ebert, C., Fox, H.E., Fujita, R., Heinemann, D., Lenihan, H.S., Madin, E.M.P.,
Perry, M.T., Selig, E.R., Spalding, M., Steneck, R., Watson, R., 2008. A global map of
human impact on marine ecosystems. Science 319, 948–952
Hewitt, C.L., Campbell, M.L., Thresher, R.E., Martin, R.B., Boyd, S., Cohen, B.F., Currie, D.R.,
Gomon, M.F., Keough, M.J., Lewis, J.A., Lockett, M.M., Mays, N., McArthur, M.A., O’Hara,
T.D., Poore, G.C.B., Ross, J.D., Storey, M.J., Watson, J.E., Wilson, R.S., 2004. Introduced
and cryptogenic species in Port Phillip Bay, Victoria, Australia. Mar. Biol. 144, 183–202
Hutson, K.S., Ross, D.J., Day, R.W., Ahern, J.J., 2005. Australian scallops do not recognise the
introduced predatory seastar Asterias amurensis. Mar. Ecol. Prog. Ser. 298, 305–309

99
Inglis, G.J., Gust, N., 2003. Potential indirect effects of shellfish culture on the reproductive
success of benthic predators. J. Appl. Ecol. 40, 1077–1089
Keane, R.M., Crawley, M.J., 2002. Exotic plant invasions and the enemy release hypothesis.
Trends Ecol. Evol. 17, 164–170
Kennedy, T.A., Naeem, S., Howe, K.M., Knops, J.M.H., Tilman, D., Reich, P., 2002. Biodiversity
as a barrier to ecological invasion. Nature 417, 636
Kimbro, D.L., Cheng, B.S., Grosholz, E.D., 2013. Biotic resistance in marine environments. Ecol.
Lett. 16, 821–833
Letnic, M., Crowther, M.S., Koch, F., 2009. Does a top-predator provide an endangered rodent
with refuge from an invasive mesopredator? Anim. Conserv. 12, 302–312
Letnic, M., Webb, J.K., Jessop, T.S., Dempster, T., 2015. Restricting access to invasion hubs
enables sustained control of an invasive vertebrate. J. Appl. Ecol. 52, 341–347
Letnic, M., Webb, J.K., Jessop, T.S., Florance, D., Dempster, T., 2014. Artificial water points
facilitate the spread of an invasive vertebrate in arid Australia. J. Appl. Ecol. 51, 795–803
Ling, S.D., Johnson, C.R., 2013. Native spider crab causes high incidence of sub-lethal injury to
the introduced seastar Asterias amurensis, in: Echinoderms in a Changing World:
Proceedings of the 13th International Echinoderm Conference, January 5-9 2009,
University of Tasmania, Hobart. p. 195.
Ling, S.D., Johnson, C.R., Mundy, C.N., Morris, A., Ross, D.J., 2012. Hotspots of exotic free-
spawning sex: man-made environment facilitates success of an invasive seastar. J. Appl.
Ecol. 49, 733–741
MacDougall, A.S., Turkington, R., 2005. Are invasive species the drivers of change or
passengers of change in degraded ecosystems? Ecology 86, 42–55
Mack, R.N., Simberloff, D., Mark Lonsdale, W., Evans, H., Clout, M., Bazzaz, F.A., 2000. Biotic
invasions: causes, epidemiology, global consequences, and control. Ecol. Appl. 10, 689–
710
McKindsey, C.W., Archambault, P., Callier, M.D., Olivier, F., 2011. Influence of suspended and
off-bottom mussel culture on the sea bottom and benthic habitats: a review. Can. J. Zool.
89, 622–646
Noonburg, E.G., Byers, J.E., 2005. More harm than good: when invader vulnerability to
predators enhances impact on native species. Ecology 86, 2555–2560
Olaso Toca, I., 1982. Ecologia de los equinodermos de la Ría de Arosa. Bol. Inst. Esp. Ocean. 7,
4–29
Olaso Toca, I., 1979. Biologia de los equinodermos de la Ría de Arosa. Bol. Inst. Esp. Ocean. 5,

100
81–127
Oudejans, R.C.H.M., van der Sluis, I., van der Plas, A.J., 1979. Changes in the biochemical
composition of the pyloric caeca of female seastars, Asterias rubens, during their annual
reproductive cycle. Mar. Biol. 53, 231–238
Papacostas, K.J., Rielly-Carroll, E.W., Georgian, S.E., Lomg, D.J., Princiotta, S.D., Quattrini, A.M.,
Reuter, K.E., Freestone, A.L., 2017. Biological mechanisms of marine invasions. Mar. Ecol.
Prog. Ser. 565, 251–268
Parry, G.D., 2017. Potential for biocontrol of the exotic starfish, Asterias amurensis, using a
native starfish. Biol. Invasions 19, 2185–2196
Parry, G.D., Cohen, B.F., 2001. The distribution, abundance and population dynamics of the
exotic seastar Asterias amurensis during the first three years of its invasion of Port Phillip
Bay, (incorporating a Report on the Bay Pest Day, 2 April 2000). Marine and Freshwater
Resources Institute, Melbourne.
Pimentel, D., McNair, S., Janecka, J., Wightman, J., Simmonds, C., O’Connell, C., Wong, E.,
Russel, L., Zern, J., Aquino, T., Tsomondo, T., 2001. Economic and environmental threats
of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 84, 1–20
Pintor, L.M., Byers, J.E., 2015. Do native predators benefit from non‐native prey? Ecol. Lett. 18,
1174–1180
Pothoven, S.A., Nalepa, T.F., Schneeberger, P.J., Brandt, S.B., 2001. Changes in diet and body
condition of lake whitefish in southern Lake Michigan associated with changes in
benthos. North Am. J. Fish. Manag. 21, 876–883
R Core Team, 2017. R: A language and environment for statistical computing. Vienna.
Robertson, B.A., Hutto, R.L., 2006. A framework for understanding ecological traps and an
evaluation of existing evidence. Ecology 87, 1075–1085
Robertson, B.A., Ostfeld, R.S., Keesing, F., 2017. Trojan females and Judas goats: evolutionary
traps as tools in wildlife management. Bioscience 67, 983–994
Robertson, B.A., Rehage, J.S., Sih, A., 2013. Ecological novelty and the emergence of
evolutionary traps. Trends Ecol. Evol. 28, 552–560
Ross, D.J., Johnson, C.R., 2002. Impact of introduced seastars Asterias amurensis on
survivorship of juvenile commercial bivalves Fulvia tenuicostata. Mar. Ecol. Prog. Ser.
241, 99–112
Ross, D.J., Johnson, C.R., Hewitt, C.L., 2003. Variability in the impact of an introduced predator
(Asterias amurensis: Asteroidea) on soft-sediment assemblages. J. Exp. Mar. Biol. Ecol.
288, 257–278

101
Russell, B.G., Letnic, M., Fleming, P.J.S., 2011. Managing feral goat impacts by manipulating
their access to water in the rangelands. Rangel. J. 33, 143–152
Sampson, J., Easton, A., Singh, M., 2014. Port Phillip Bay, in: Wolanski, E. (Ed.), Estuaries of
Australia in 2050 and Beyond. Springer, pp. 49–68
Sanderson, E.W., Jaiteh, M., Levy, M.A., Redford, K.H., Wannebo, A. V, Woolmer, G., 2002. The
human footprint and the last of the wild. Bioscience 52, 891–904
Saranchova, O.L., Kulakovskii, É.E., 1982. Effect of environmental salinity on different
developmental stages of sea stars Asterias rubens and mussels Mytilus edulis. Sov. J. Mar.
Biol. 8, 32–36
Sih, A., Bolnick, D.I., Luttbeg, B., Orrock, J.L., Peacor, S.D., Pintor, L.M., Preisser, E., Rehage, J.S.,
Vonesh, J.R., 2010. Predator-prey naïveté, antipredator behavior, and the ecology of
predator invasions. Oikos 119, 610–621
Sih, A., Ferrari, M.C.O., Harris, D.J., 2011. Evolution and behavioural responses to human-
induced rapid environmental change. Evol. Appl. 4, 367–387
Suarez, A. V, Case, T.J., 2002. Bottom-up effects on persistence of a specialist predator: ant
invasions and horned lizards. Ecol. Appl. 12, 291–298
Suarez, A. V, Holway, D.A., Case, T.J., 2001. Patterns of spread in biological invasions
dominated by long-distance jump dispersal: insights from Argentine ants. Proc. Natl.
Acad. Sci. USA 98, 1095–1100
Tetzlaff, J.C., Roth, B.M., Weidel, B.C., Kitchell, J.F., 2011. Predation by native sunfishes
(Centrarchidae) on the invasive crayfish Orconectes rusticus in four northern Wisconsin
lakes. Ecol. Freshw. Fish 20, 133–143
Vitousek, P.M., Mooney, H.A., Lubchenco, J., Melillo, J.M., 1997. Human domination of Earth’s
ecosystems. Science 277, 494–499
Vörösmarty, C.J., McIntyre, P.B., Gessner, M.O., Dudgeon, D., Prusevich, A., Green, P., Glidden,
S., Bunn, S.E., Sullivan, C.A., Liermann, C.R., others, 2010. Global threats to human water
security and river biodiversity. Nature 467, 555–561
With, K.A., 2002. The landscape ecology of invasive spread. Conserv. Biol. 16, 1192–1203
Xu, R.A., Barker, M.F., 1990. Laboratory experiments on the effects of diet on the gonad and
pyloric caeca indices and biochemical composition of tissues of the New Zealand starfish
Sclerasterias mollis (Hutton) (Echinodermata: Asteroidea). J. Exp. Mar. Biol. Ecol. 136, 23–
45

102
CHAPTER FIVE: AN INVASIVE HABITAT-FORMER MITIGATES IMPACTS OF NATIVE
HABITAT LOSS FOR ENDEMIC REEF FISHES
ABSTRACT
Impacts of environmental change on animal populations depend on how individuals respond
to novel or degraded habitats. Animals that make adaptive habitat selection choices can
maximise their fitness in altered landscapes, while maladaptive selection of low quality
habitats can lead to the formation of ‘ecological traps’ with increased mortality or lowered
reproductive success. Habitat-forming invasive species provide evolutionarily unfamiliar
habitat with the potential to create ecological traps. In southern Australia, urchin grazing has
driven a decline in native kelps enabling an invasive kelp (Undaria pinnatifida) to fill the vacant
niche and provide replacement canopy with unknown effects. We assessed the value of this
novel habitat for macroalgae-associated fishes by combining laboratory habitat choice
experiments, an artificial reef experiment, before-after-control-impact field surveys and
estimates of fitness. In captive choice experiments, macroalgae-associated fishes preferred
rock with macroalgal cover to rock without, but did not distinguish between invasive and
native macroalgae. Cryptobenthic reef fishes occurred in higher abundance and diversity on
urchin-grazed reefs with invasive kelp than those without, while the invader did not affect
communities of large or highly mobile fishes. More fish recruited to artificial reefs that were
stocked with kelp compared to barren reefs, with no preference between native and invasive
kelp canopy. Fitness metrics, including stomach contents, body condition, liver and gonad
indices and fecundity in fish collected from invasive kelp habitats were similar to those in fish
from adjacent native kelp patches, indicating that this invader can function as a partial
replacement for lost native habitat-forming species. These findings illustrate how invasive
habitat-formers can play a role in maintaining biodiversity in heavily impacted ecosystems, and
reinforces the need for managers to consider effects of habitat-forming invasive species within
the broader context of environmental change.

103
INTRODUCTION
Ecosystems will continue to undergo dramatic anthropogenic shifts in the coming century,
with impacts on fauna likely to depend on how individuals respond to modified or degraded
habitats. Animals that make adaptive decisions by responding appropriately to novel risks and
resources and that prefer habitats conferring the best fitness outcomes will be most able to
persist in impacted landscapes (Sih et al. 2011, Wong & Candolin 2014). However, a lack of
evolutionary history in altered environments can create a mismatch between cues and
outcomes, causing individuals to make maladaptive decisions (evolutionary trap: Robertson,
Rehage & Sih 2013). In the case of habitat selection decisions, this may result in individuals
avoiding high quality but unattractive habitats (perceptual traps) or selecting low quality but
attractive habitats (ecological traps) (Robertson & Hutto 2006; Patten & Kelly 2010; Hale &
Swearer 2016). In impacted landscapes, such individual-level responses to altered habitats
could exacerbate population-level effects of environmental change, as animals are either
drawn into attractive population sinks from surrounding higher quality habitats, or fail to take
advantage of remaining viable habitats in fragmented landscapes because habitat cues have
changed (Hale et al. 2015).
Traditional density-based habitat association studies may come to erroneous conclusions
about the ecological value of a habitat if individual-level processes are not considered (van
Horne 1983). Density-dependent source-sink population models account for differential fitness
between habitats (Pulliam 1988), but such models typically assume (often incorrectly) that
individuals will choose the best available habitat, and thus ignore the potential for attractive
sinks or underutilised sources (Kokko & Sutherland 2001, Battin 2004). By linking individual
habitat selection and fitness outcomes, the ecological trap framework provides a more
informative approach to assessing the role of habitats in population persistence (Hale et al.
2015, Hale & Swearer 2016).
Invasive habitat-forming ecosystem engineers, such as plants, algae and sessile marine fauna,
can alter ecosystems by competing for space with native habitat-formers and by changing the
availability of food and shelter for animals (Crooks 2002, MacDougall & Turkington 2005,
Gribben & Wright 2006, Pyšek et al. 2012). It is often assumed that invasive species reduce
biodiversity in invaded areas, but the body of empirical evidence is equivocal, particularly for
animals (Gribben & Wright 2006, Pyšek et al. 2012, Dijkstra et al. 2017). Effects on biodiversity
will depend on a range of conditions, including the state of the ecosystem prior to invasion,

104
the quality of the novel habitat for native fauna, and on behavioural responses by native fauna
to the novel habitat.
Figure 5.1. Archetypal examples of three rocky reef habitats in northern Port Phillip Bay,
Australia: (A) dense canopy of the native kelp Ecklonia radiata, (B) urchin-grazed barren reef,
and (C) spring growth of the invasive kelp Undaria pinnatifida on urchin-grazed reef.
Where anthropogenic stressors lead to declines in native habitat-forming species, invasive
habitat-formers may take advantage of vacant niches (MacDougall & Turkington 2005). A
marine macroalga, the Japanese kelp (Undaria pinnatifida), is one such invader. Native to the
north-west Pacific Ocean, and has now established in Europe, the Americas, New Zealand and
Australia. However, despite its high profile as a serial invader and a considerable body of
literature on its physiology and reproductive ecology (Schaffelke & Hewitt 2007, Davidson et
al. 2015), very little is known about its impacts on animal populations in invaded ecosystems
(Raffo et al. 2009, 2014, Thomsen et al. 2009, Irigoyen et al. 2011, Howland 2012). In south-
eastern Australia, Undaria fills a macroalgal niche on artificial substrates and degraded reefs
where poor water quality and urchin grazing have driven the decline of native kelps such as
Ecklonia radiata (Fig. 5.1). Undaria is a weak competitor in undisturbed macroalgal
communities, especially those with laminarian kelps (Valentine & Johnson 2003, Edgar et al.
2004, Farrell & Fletcher 2006, de Leij et al. 2017), but quickly takes advantage of bare
substrate and appears better able to persist on degraded reefs (Campbell & Burridge 1998,
Valentine & Johnson 2003, Edgar et al. 2004, South & Thomsen 2016).
Seasonal Undaria growth may mitigate loss of fish biodiversity on urchin-grazed rocky reefs if
native fish utilize the potential refugia provided by the invasive habitat, and the invasive
habitat is of higher quality than the urchin barren habitats that it replaces. Potential benefits

105
will be maximised if the invasive habitat does not act as an ecological trap by attracting fish
that might otherwise settle in higher quality native habitats (Robertson & Hutto 2006, Patten
& Kelly 2010, Hale & Swearer 2016). In this study, we employ the ecological trap framework to
test three predictions about the effects of the invasive habitat-former on native fauna, by
surveying fish populations on comparable natural rocky reefs and artificial boulder reefs with
differing canopy cover. The directions of our predictions are based on a common expectation
that invaded habitats will be both less attractive and of poorer quality than uninvaded
habitats, but that the invasive habitat-former may offer better habitat than urchin barrens:
native fish will prefer native over invasive kelps over urchin barrens (Prediction 1); native fish
communities will differ and be more abundant and diverse in invasive kelp than urchin barrens
(Prediction 2); and measures of fitness will be higher for fish in native than invasive kelp
habitat (Prediction 3). In testing these predictions, we inform management responses to this
established invader, and demonstrate how the ecological trap framework can be employed in
future assessments of the effects of invasive habitat-formers on native fauna.
METHODS
Study system
We conducted this study at five locations in Port Phillip Bay (Fig. 5.2; see Appendix 5.1 for
location descriptions), a 1930 km2 semi-enclosed marine embayment in south-eastern
Australia. The bay is affected by numerous anthropogenic impacts arising from population
centres and industrial inputs (Sampson et al. 2014). Soft sediment habitats are predominant,
with scattered rocky shorelines and reef patches in 0-8 m depth. Rocky reef habitats in the
northern half of the bay have been degraded by a combination of abiotic conditions and urchin
grazing, leading to large areas of urchin barrens and epilithic sediment matrix with turfing
algae (Ling et al. 2010, Filbee-Dexter & Wernberg 2018). Canopy-forming macroalgae such as
the native common kelp (Ecklonia radiata) have been in decline for decades (Jung et al. 2011).
Concurrently, the invasive kelp Undaria pinnatifida has been spreading since its introduction in
the late 1980s, and is now widespread on rocky reefs and marine infrastructure throughout
the bay, with high densities in the northern half (pers. obs.). Throughout its native range and
most of its introduced range (including Australia), Undaria is a winter annual, forming dense
stands during winter-spring and senescing in summer.

106
Figure 5.2. Map of study locations in Port Phillip Bay, Australia. Round markers denote
locations with established populations of Undaria pinnatifida. Governor Reef (square marker)
is a comparatively unimpacted reef with dense Ecklonia radiata canopy.
Prediction 1: Native fish will prefer native over invasive kelps and urchin barrens habitat
The first criterion for demonstrating an ecological trap is to show that animals either prefer or
do not avoid the novel habitat (Robertson & Hutto 2006).
Habitat choice experiment
To investigate whether native reef fish prefer native over invasive kelp, we collected common
weedfish (Clinidae: Heteroclinus perspicillatus) and little weed whiting (Odacidae: Neoodax
balteatus) individuals from rocky reef habitats and subjected them to a habitat choice trial.
These species are among the most common macroalgae-associated fishes in the Bay and are
easily housed in aquaria for the duration of the experiment. Both species use macroalgae and
seaweed structure as shelter. Clinids are predators of small fish and motile invertebrates and
use available cover to stalk and ambush prey. N. balteatus are more active in browsing
macroalgal surfaces for epifaunal invertebrate prey such as molluscs and crustaceans.

107
We placed individuals in the centre of a four-chambered crucifix-shaped tank (McDermott and
Shima 2006; Appendix 5.2) and offered them a simultaneous choice between one reference
(bare rock, simulating an urchin barren) and three macroalgal habitat cues (similarly-sized
rocks with a Undaria, Ecklonia or Sargassum linearifolium thallus attached via a black cable tie
on the holdfast). We provided equal volumes of each species of macroalgae (measured by
water displacement) and randomised their positions within the tank for each replicate fish.
Undaria and Ecklonia are very similar in height and gross morphology at a given volume, with
large fronds making up the bulk of the volume and spreading to cover a large area of
substrate. Sargassum tends to be more upright, with gas-filled bladders holding foliage above
the substrate. At least 10 thalli of each macroalgal species were used throughout the
experiment; for each replicate trial, a thallus was randomly selected and attached to a
randomly-selected rock. We held fish in a transparent plastic cylinder in the centre of the tank
for 5 min to visually survey the habitat options prior to the commencement of the trial. The
cylinder was then raised by a string-and-pulley system with the researcher out of sight (but
able to view the fish through a small hole in a screen), and the fish allowed to swim freely
around the choice tank. As measures of preference, we recorded both the initial choice of the
fish, which was likely to be a primarily vision-based decision (first chamber entered after
release from the cylinder) and the location of the fish after 20 mins, which is expected to be a
decision based on multiple senses. No fish were recorded in the centre of the tank at the 20
min mark.
Recruitment to artificial reefs
To complement the laboratory habitat choice experiment, we conducted an artificial reef
experiment at Half Moon Bay to test the effect of native and invasive kelp canopies on fish
recruitment under controlled field conditions. Half Moon Bay was selected as it offered a large
expanse of sandy substrate at a suitable depth and in an area of the Bay where Undaria has
been established since at least 2009 (Primo et al. 2010). During September 2014, we
constructed 20 replicate boulder reefs (1 m2) on sandy substrate, arrayed 20 m apart in a 4 x 5
grid pattern, and randomly assigned each reef to one of three treatments (n = 7 with Undaria
canopy, n = 7 with E. radiata canopy and n = 6 as unstocked controls) using kelp thalli collected
from an adjacent subtidal reef and attached to the artificial reef using rope and cable ties. We
stocked kelp treatments in late July and early August 2015 and recorded natural recruitment of

108
reef fishes on three occasions throughout spring (16 September, 26 October and 31 November
2015). Kelp canopies were replenished as necessary to maintain similar percent coverage
(Undaria treatments required regular replenishment as thalli regressed during October and
November). As artificial reefs were initially devoid of fish, relative recruitment was quantified
in terms of the abundance and diversity of fish present on the reef at each survey date.
Prediction 2: Native fish will be more abundant and diverse in invasive kelp than urchin
barrens
To test whether habitat preferences in experimental conditions correspond to patterns of fish
abundance in the field, we compared fish communities on heavily urchin-grazed rocky reefs
with and without seasonal Undaria canopy using the underwater visual census (UVC) survey
method for benthic fish species and baited remote underwater video (BRUV) for larger, more
mobile fish species. Survey locations were selected based on the presence of Undaria patches
on urchin-grazed subtidal reefs. We primarily focused on a comparison of fish populations in
invasive Undaria habitat relative to urchin barrens—rather than native Ecklonia habitat—
because (a) the small size and rarity of remaining Ecklonia habitats in northern Port Phillip Bay
limited the potential for spatial replication of comparable sites, with Williamstown currently
the only known location where urchin barrens, Undaria patches and remnant Ecklonia patches
co-occur on rocky reef, and (b) this invader fills vacant habitat on degraded reefs or artificial
substrates rather than directly outcompeting native kelps, making the comparison to barren
reefs more ecologically-relevant. Nonetheless, we did compare fish populations in Undaria and
Ecklonia beds using a diver catch-per-unit-effort metric at one location (Williamstown), as well
as BRUV deployments at two locations (Williamstown and St Leonards) (Fig. 5.2). All surveys
and collections were conducted by the same researcher (LB) under calm weather conditions
(<15 knots) between 1000-1600 hours. Survey and collection efforts alternated randomly
between the two habitat types to avoid temporal bias.
Underwater visual census survey
The UVC was conducted on paired circular plots of 12.6 m2 (2 m radius) with comparable
rugosity and depth but differing in their coverage of Undaria. The paired design only extended

109
to placement of plots to maximise comparability of plots with and without Undaria, rather
than a paired statistical analysis. We employed a before-after-control-impact (BACI) design by
surveying plots with and without seasonal Undaria canopy, during and after the Undaria
growing season (when the canopy was at its most dense and after it had completely
regressed). In this environment, adult Undaria undergo a complete die-off over summer, with
no holdfasts remaining by the time of the “after” surveys. We haphazardly placed paired plots
10-15 m apart, and >30 m from adjacent pairs at that location. Populations of small benthic
and cryptic fishes are strongly influenced by habitat characteristics at this scale (Willis &
Anderson 2003). We dropped a weight to mark the middle of the plot and surveyed larger
benthic and benthopelagic fish initially by swimming a circular path outside the plot. Small or
cryptic species were detected by a diver moving in concentric circular paths from the outer
edge to the centre of the plot, searching within the reef structure and among algal cover. If fish
of the same species were observed multiple times within the plot, they were counted as a
single fish unless they were clearly distinct individuals (size, sex or markings). Several members
of the Heteroclinus genus (H. perspicillatus, H. heptaeolus, H. adelaidae, H. wilsoni, H.
eckloniae, H. macrophthalmus) are difficult to reliably distinguish in situ, so were treated as a
single species. Accordingly, the species richness metric is likely underestimating the true
diversity of the Heteroclinus genus. Upon completing the fish census, we counted the number
of urchins present within the plot and estimated percent coverage of kelp, all macroalgae, and
algal turf.
Baited video survey
The BRUV system consisted of a GoPro Hero3+ camera mounted on a weighted milk crate with
a 1 m long bait arm made from 20mm PVC conduit. Two pilchards (Sardinops sagax) were cut
in half and placed in a 20 x 20 cm plastic mesh bag attached to the end of the bait arm. Larger
baits are typically used in BRUV systems, but we wished to minimise the spread of the bait
plume as elevated rocky reefs in the bay are often very small (10-100 m). BRUV deployments
were made at least 50 m apart on comparable reef substrate at depths of 2-4.5 m. Fish
observations were made for 20 min from the time when the BRUV unit settled on the sea
floor. To assess relative abundance of each species at each site while preventing potential
double counting of individual fish, we used a conservative metric termed maxN, where maxN is
the maximum number of individuals of a given species occurring simultaneously in the video

110
field of view (Willis & Babcock 2000). In the case of sexually dimorphic species or juvenile
fishes, we summed the maxN counts for males, females and juveniles of that species. Where
an individual could not be identified to species level, we identified it to the lowest taxonomic
rank possible (usually Family or Genus). We included the southern calamari squid (Sepioteuthis
australis) in the analysis as it is similar in habit to many inshore fishes.
Catch-per-unit-effort
Collection efforts for N. balteatus, H. perspicillatus and H. heptaeolus in Ecklonia and Undaria
habitat patches at Williamstown were timed to allow quantification of catch per unit effort
(CPUE). The Williamstown location contained a mosaic of seasonal Undaria growing on urchin-
grazed reefs between well-defined remnant patches of Ecklonia and Sargassum. Collections
were made by hand net on SCUBA. We calculated CPUE on a per species basis, with each
collection dive treated as a single statistical replicate where CPUE = N fish collected / duration
of dive. Collection dives alternated between habitats over the course of nine days between 24
October to 28 November 2016.
Prediction 3: Measures of fitness will be higher for fish in native than invasive kelp habitat
The second criterion for demonstrating an ecological trap is to show that a suitable measure of
fitness is lower in the putative trap habitat than other habitat options (Robertson & Hutto
2006). We expect that the native kelp habitat will be of higher quality and may lead to higher
fitness metrics, although individual fitness (Prediction 2) may not be independent of
population density (Prediction 1); population density tends to be higher in high quality
habitats, leading to equalisation of individual fitness between habitats of differing quality
(ideal free distribution: Fretwell & Lucas 1969).
Fish collected from natural reefs
We compared body condition metrics in N. balteatus, H. perspicillatus and Ogilby's weedfish
(Heteroclinus heptaeolus) collected at Williamstown from Undaria and Ecklonia habitats

111
(collection efforts described above). The habitat occupied by each individual was defined by
the dominant kelp species within 2 m of the collection site, with percentage coverage of
macroalgal species estimated visually. Fish were collected using handnets and killed using
clove oil, and placed in sealed plastic bags in an ice slurry. Within 24 hours of collection,
specimens were weighed (wet weight), measured (total length) and dissected to determine sex
and weight of the stomach contents, liver and gonads. Overall body condition was quantified
using the relative condition metric recommended by (Le Cren 1951): Krel = W/Wexp, where W is
the measured gutted weight and Wexp is the gutted weight predicted by the weight-at-length
power curve fitted to all available samples (Fig. 5.3). Individuals with Krel values >1 are heavier
than the population mean for their length. One particularly large H. perspicillatus individual
from Undaria habitat was removed from the Krel analysis (145 mm, 4.7 standard deviations
above the mean). Liver and gonad condition was quantified using hepatosomatic and
gonadosomatic indices respectively (HSI or GSI = OW/GW, where OW = wet organ weight and
GW = gutted weight). For gravid females, we also photographed and counted subsamples of
ova (N. balteatus) or embryos (Heteroclinus spp.) to assess fecundity (Neoodax are broadcast
spawners, while Heteroclinus are livebearers).
Fish collected from artificial reefs
We collected H. perspicillatus individuals from artificial reefs (see Prediction 1) from Jan-Nov
2015 and compared body condition and reproductive investment metrics in fish from reefs
stocked with Undaria or Ecklonia. Collection methods were identical to those on natural reefs,
with collection efforts alternating between Undaria and Ecklonia stocking treatments. There
were not enough H. perspicillatus recruits on control (unstocked) reefs for meaningful
comparisons of condition on reefs with and without kelp.
Statistical analysis
Choice data (Prediction 1) were tested for habitat preference using a X2 test of proportions
implemented in R (R Core Team 2017), with expected (null) proportions equally distributed
across the four habitat cues. Artificial reef recruit abundance and species richness data
(Prediction 1) were compared across kelp treatments using Poisson generalised linear mixed

112
effects models implemented in the lme4 package for R (Bates et al. 2015). We included
Treatment (stocked kelp species: Undaria, Ecklonia or barren) as a fixed term, KelpCover
(percentage coverage of canopy) as a covariate, and as reefs were surveyed repeatedly, a reef
identity random intercept term (ReefID) nested within Treatment. There was some data
overdispersion, so we included an observation-level random term to account for excess
variation and avoid overestimating the predictive ability of model terms (Harrison 2014). We
tested for a significant Treatment effect by comparing the fit of models with and without the
Treatment term, while the lsmeans package for R provided Tukey’s pairwise post-hoc
comparisons of the three treatments (Lenth 2016).
Fish community data from UVC and BRUV surveys (Prediction 2) were fitted to a permutational
multivariate ANOVA model implemented in PRIMER 6 with the PERMANOVA+ add-on
(Anderson et al. 2008). Data were log(x+1) transformed to reduce the influence of a few highly
abundant species and fitted to a Bray-Curtis similarity resemblance matrix with a dummy
variable of 1. The models contained two fixed factors, Habitat and Location, as well as an
interaction term (Habitat×Location). The Location factor contained two levels (North and
West), reflecting environmental differences between sites in the north (Point Cook, Altona
Bay, Williamstown, Half Moon Bay) and west (Kirk Point) of the Bay. The models were fitted
using Type III sums of squares, with unrestricted permutation of raw data and 9999
permutations. We also extracted diversity metrics using the Diversity function and identified
species that were associated most strongly with the observed differences between habitats
and locations using the similarity percentage (SIMPER) function. Finally, data were visualised
using multidimensional scaling (MDS) and canonical analysis of principal coordinates (CAP)
plots.
Catch per unit effort data were log(x+1) transformed to improve normality, and compared
across Ecklonia and Undaria habitats using a linear analysis of covariance (lm function in R)
with Habitat as a fixed factor and sampling Date as a temporal covariate.
We compared fitness metrics (Prediction 3) in native and invasive kelp habitats using a series
of univariate linear models implemented in R. Response variables were checked for normality
and equality of variance and transformed as necessary. We analysed both weedfish species (H.
perspicillatus and H. heptaeolus) with a single model and included terms for kelp habitat type
(‘Habitat’), species (‘Species’), day of the season (‘Day’), and sex (‘Sex’). Models for the
sequentially hermaphroditic N. balteatus included only Habitat and Day terms. Fitness metrics
for H. perspicillatus recruits collected from artificial reefs (Prediction 3) were compared across
kelp treatments using univariate linear mixed models implemented in lme4, and included

113
Treatment, KelpCover and Sex as fixed terms and Treatment/ReefID as a random intercept
term. In all cases, we tested the significance of the effect of interest by comparing the fit of
models with and without the relevant term.
Plots were produced using the ggplot2 package for R (Wickham 2009).
Figure 5.3. Weight-at-length relationships for (A) Heteroclinus perspicillatus and H. heptaeolus
and (B) Neoodax balteatus individuals collected from Undaria pinnatifida and Ecklonia radiata
habitats in Port Phillip Bay.
RESULTS
Prediction 1: Native fish will prefer native over invasive kelps and urchin barrens habitat
Habitat choice experiment
Common weedfish (Heteroclinus perspicillatus) were more likely to select macroalgal cover
than barren rock, both initially (1.25x more than expected; p <0.0001) and after 20 mins (1.22x
more than expected; p <0.0001) (Fig. 5.4A; Appendix 5.3). Of those that chose macroalgae
during their initial decision, Undaria was preferable to Ecklonia or Sargassum (2.0x and 2.7x
more likely, respectively; p = 0.015), but after 20 mins, individuals were evenly distributed
across the three macroalgal options (p = 0.9) (Fig. 5.4A; Appendix 5.3).

114
Figure 5.4. Habitat choice trial results for (A) common weedfish (Heteroclinus perspicillatus): N
= 48; (B) little weed whiting (Neoodax balteatus): N = 23.
Little weed whiting (N. balteatus) also preferred macroalgae to bare rock, both initially (1.16x
more than expected; p = 0.0004) and after 20 mins (1.22x more than expected; p <0.0001) (Fig.
5.4B; Appendix 5.3). However, we observed no clear preference among macroalgal options,
either initially (p = 0.12) or after 20 mins (p = 0.4) (Fig. 5.4B; Appendix 5.3).
Table 5.1. Summary of fish recruitment to artificial reefs stocked with Undaria pinnatifida,
Ecklonia radiata, or left barren.
Treatment Kelp canopy cover (%) Abundance reef-1 Species richness reef-1 N surveys
Undaria 42 ± 3 0.9 ± 0.1 0.8 ± 0.1 89
Ecklonia 73 ± 2 1.1 ± 0.1 1.1 ± 0.1 89
Barren 5 ± 5 0.4 ± 0.2 0.4 ± 0.2 18
Pairwise
U-E z = 1.7, p = 0.21 z = 1.6, p = 0.24
U-B z = 2.3, p = 0.05 z = 2.8, p = 0.02
E-B z = 3.0, p = 0.008 z = 3.3, p = 0.003
Recruitment to artificial reefs
A 2.5x higher abundance (p = 0.01) and 2.4x higher species richness (p = 0.007) of reef fish
recruited to artificial reefs stocked with kelp relative to those with turfing algae only (Table

115
5.1). There was no evidence for differential recruitment between reefs stocked with either
kelp (p = 0.21), despite the Undaria treatment having less canopy cover (42 vs. 73 %) (Table
5.1). The canopy cover covariate positively predicted recruit abundance (p = 0.05), indicating
that the amount of cover may be more important than the macrophyte species providing the
cover. Canopy cover did not affect species richness (p = 0.36).
Prediction 2: Native fish will be more abundant and diverse in invasive kelp than urchin
barrens
Visual census survey
We recorded 19 fish species across 25 Undaria and 26 barren plots during in-season surveys
(September-October 2016) (Appendix 5.4) and found a significant effect of Habitat (Undaria or
Barren), Location (west or north Port Phillip Bay), and Habitat×Location on dissimilarity
between plots (PERMANOVA: Appendix 5.5). Canonical analysis of principle coordinates (CAP)
plots revealed visual separation of sites with and without Undaria, driven primarily by a
greater abundance of Heteroclinus spp. and Diodon nicthemerus in Undaria plots (Fig. 5.5).
SIMPER analysis determined that weedfish (Heteroclinus spp.: 28 %), Clarke’s threefin
(Trinorfolkia clarkei: 23 %), Tasmanian blenny (Parablennius tasmanianus: 10 %) and globefish
(D. nichthemerus: 7 %) contributed most to the observed dissimilarity between habitats
(weedfish and globefish in Undaria plots, threefins and blennies in Barren plots). Gross
community metrics also differed; species richness and total abundance were both higher on
Undaria plots (2.0x and 1.9x higher, respectively: Fig. 5.6A, Appendix 5.6). Barren and Undaria
plots were placed at comparable depths (mean ± SD: 2.6 ± 0.5 m cf. 2.7 ± 0.4 m, respectively).
Barren plots contained 6 ± 2 % macroalgal cover, mostly sea lettuce (Ulva spp). Undaria plots
contained 72 ± 3 % macroalgal cover, dominated by Undaria (58 %) with some secondary cover
from other ephemeral macroalgae, including Ulva spp. and Gracilaria spp. Undaria plots did
not contain any other brown algal species such as Ecklonia radiata or Sargassum spp. Where
Ulva or Gracilaria were present on Undaria plots, clinids (Heteroclinus spp. and Cristiceps
australis) were generally found in Undaria microhabitat (12/16 fish).
We resurveyed 18 plots (9 Undaria, 9 Barren) after the Undaria had completely regressed
(April-May 2017). Sparse early stage Undaria recruits (<10 cm) were present in May, but

116
provided a negligible amount of macroalgal cover. The effect of the Habitat factor on overall
fish community structure was significant (PERMANOVA: Appendix 5.5), but neither species
richness nor abundance significantly differed (Appendix 5.6).
Analysing in-season and off-season data together revealed significant overall effects of
Habitat, Season and Location on fish community structure, as well as a Habitat×Location
interaction (PERMANOVA: Appendix 5.5). Off-season Undaria and Barren plots both contained
abundant non-canopy-forming ephemeral macroalgae, dominated by Ulva spp., Gracilaria spp.
and Caulerpa spp., with greater mean coverage (51 ± 12 %) on Undaria plots relative to Barren
plots (32 ± 12 %).
Overall, macroalgal cover negatively correlated with urchin density (r = -0.42, t67 = -3.7, p =
0.0004). We found no evidence that off-season macroalgal cover on resurveyed plots was
predicted by in-season cover of either Undaria (F16 = 0.5, p = 0.5) or all macroalgae (F16 = 1.5, p
= 0.24).
Baited video survey
We recorded 28 fish species across 14 Undaria and 15 barren deployments (Appendix 5.4). Fish
communities did not differ between habitats or locations (PERMANOVA: Appendix 5.6), and
nor did species richness or combined MaxN (Appendix 5.6). BRUV data included some non-
reef-associated species, but restricting the analysis to reef-associated species did not alter our
interpretation.
Diver catch per unit effort
Collection efforts provided 45 Neoodax balteatus, 24 Heteroclinus perspicillatus and 18
Heteroclinus heptaeolus from 12 dives (total 465 min) in Undaria and 13 dives in Ecklonia (total
690 min) habitats over 9 days between 24 October and 28 November 2016. Catch per unit
effort (CPUE) of N. balteatus was 2.6x higher in Ecklonia than Undaria habitat (p = 0.04), but
CPUE of Heteroclinus spp. did not differ (Table 5.2). CPUE of N. balteatus also increased
throughout spring, coinciding with warmer water and greater fish activity (Date covariate: R2 =
0.26, p = 0.007; Table 5.2). There was no evidence that CPUE of Heteroclinus spp. increased
over time (p = 0.6; Table 5.2).

117
Table 5.2. Comparison of reef fish relative abundance, estimated by diver catch per unit effort
(CPUE), in Undaria and Ecklonia habitats, and over time, at Williamstown. Number of
collection efforts (N dives) are given as ‘Undaria, Ecklonia’. N dives differ between species as
not all dives targeted all species. Terms are tested by maximum likelihood ratio comparison of
fitted and null models (X2). Positive Cohen’s d effect sizes indicate metrics were higher in
Undaria habitat.
Habitat effect Undaria Ecklonia N dives X2 p Cohen’s d
Heteroclinus perspicillatus 0.9 ± 0.3 2.0 ± 0.6 13, 10 1.4 0.12 -0.74
Heteroclinus heptaeolus 0.7 ± 0.2 1.0 ± 0.4 13, 12 0.04 0.91 -0.30
Heteroclinus spp. 1.5 ± 0.4 2.7 ± 0.8 13, 12 0.32 0.73 -0.48
Neoodax balteatus 1.6 ± 0.4 4.1 ± 1.0 9, 10 2.1 0.04 -1.08
Temporal effect R2 N dives X2 p
Heteroclinus perspicillatus -0.02 13, 10 0.74 0.33
Heteroclinus heptaeolus 0.10 13, 12 0.77 0.19
Heteroclinus spp. 0.03 13, 12 0.14 0.87
Neoodax balteatus 0.26 9, 10 3.3 0.007
CPUE was not formally assessed in barren habitats, as both Heteroclinus spp. and N. balteatus
are rare in areas without macroalgae. In the course of surveying 26 barren UVC plots during
the Undaria growing season in the northern part of the Bay, we only observed one clinid—a H.
perspicillatus individual inhabiting a macroalgal microhabitat on an otherwise barren plot—
and no N balteatus (Appendix 5.4). Accordingly, CPUE in the barrens would likely be close to
zero.
Prediction 3: Measures of fitness will be higher for fish in native than invasive kelp habitat
Fish collected from natural reefs
N. balteatus individuals collected from Ecklonia habitat were 1.3x larger (p = 0.007) and 2.4x
heavier (p = 0.01) than those in seasonal Undaria habitat (Table 5.3), and a higher proportion
were male (Ecklonia: 7/23; Undaria: 1/22). We found no evidence that body condition or
reproductive fitness metrics differed between Ecklonia and Undaria habitats, although the
direction of effect for reproductive metrics was generally positive in Undaria habitats (Table
5.3).
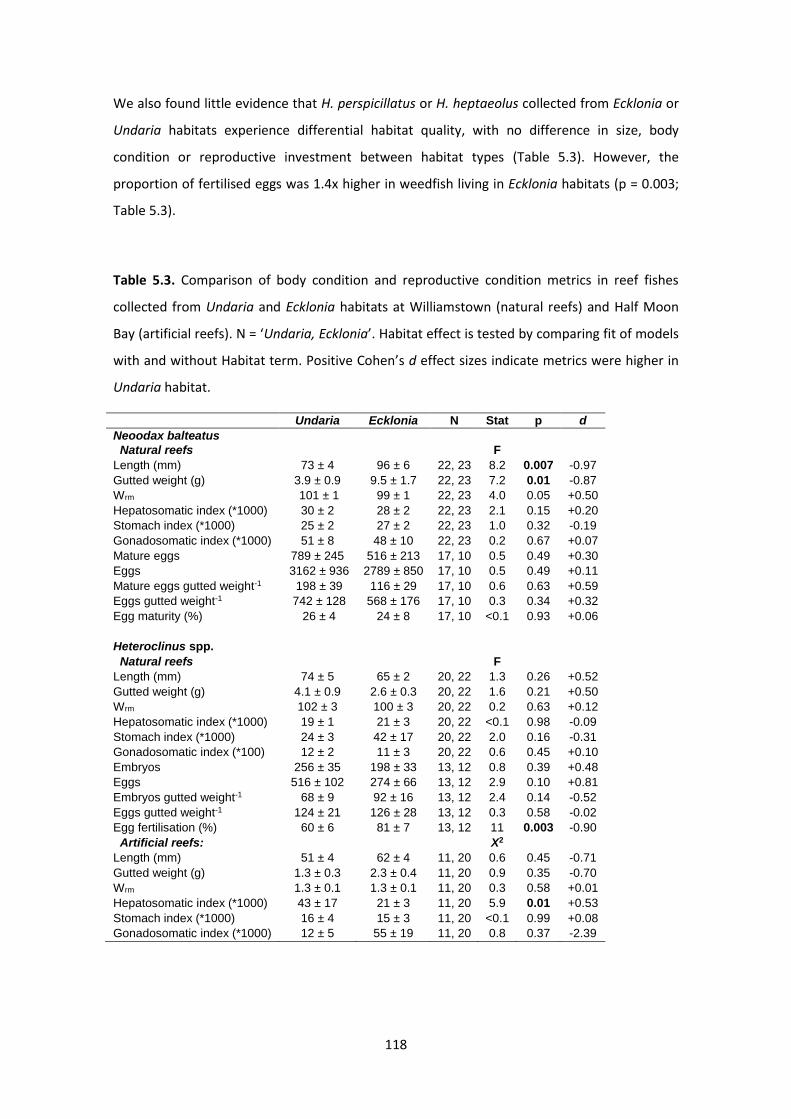
118
We also found little evidence that H. perspicillatus or H. heptaeolus collected from Ecklonia or
Undaria habitats experience differential habitat quality, with no difference in size, body
condition or reproductive investment between habitat types (Table 5.3). However, the
proportion of fertilised eggs was 1.4x higher in weedfish living in Ecklonia habitats (p = 0.003;
Table 5.3).
Table 5.3. Comparison of body condition and reproductive condition metrics in reef fishes
collected from Undaria and Ecklonia habitats at Williamstown (natural reefs) and Half Moon
Bay (artificial reefs). N = ‘Undaria, Ecklonia’. Habitat effect is tested by comparing fit of models
with and without Habitat term. Positive Cohen’s d effect sizes indicate metrics were higher in
Undaria habitat.
Undaria Ecklonia N Stat p d
Neoodax balteatus
Natural reefs F
Length (mm) 73 ± 4 96 ± 6 22, 23 8.2 0.007 -0.97
Gutted weight (g) 3.9 ± 0.9 9.5 ± 1.7 22, 23 7.2 0.01 -0.87
Wrm 101 ± 1 99 ± 1 22, 23 4.0 0.05 +0.50
Hepatosomatic index (*1000) 30 ± 2 28 ± 2 22, 23 2.1 0.15 +0.20
Stomach index (*1000) 25 ± 2 27 ± 2 22, 23 1.0 0.32 -0.19
Gonadosomatic index (*1000) 51 ± 8 48 ± 10 22, 23 0.2 0.67 +0.07
Mature eggs 789 ± 245 516 ± 213 17, 10 0.5 0.49 +0.30
Eggs 3162 ± 936 2789 ± 850 17, 10 0.5 0.49 +0.11
Mature eggs gutted weight-1 198 ± 39 116 ± 29 17, 10 0.6 0.63 +0.59
Eggs gutted weight-1 742 ± 128 568 ± 176 17, 10 0.3 0.34 +0.32
Egg maturity (%) 26 ± 4 24 ± 8 17, 10 <0.1 0.93 +0.06
Heteroclinus spp.
Natural reefs F
Length (mm) 74 ± 5 65 ± 2 20, 22 1.3 0.26 +0.52
Gutted weight (g) 4.1 ± 0.9 2.6 ± 0.3 20, 22 1.6 0.21 +0.50
Wrm 102 ± 3 100 ± 3 20, 22 0.2 0.63 +0.12
Hepatosomatic index (*1000) 19 ± 1 21 ± 3 20, 22 <0.1 0.98 -0.09
Stomach index (*1000) 24 ± 3 42 ± 17 20, 22 2.0 0.16 -0.31
Gonadosomatic index (*100) 12 ± 2 11 ± 3 20, 22 0.6 0.45 +0.10
Embryos 256 ± 35 198 ± 33 13, 12 0.8 0.39 +0.48
Eggs 516 ± 102 274 ± 66 13, 12 2.9 0.10 +0.81
Embryos gutted weight-1 68 ± 9 92 ± 16 13, 12 2.4 0.14 -0.52
Eggs gutted weight-1 124 ± 21 126 ± 28 13, 12 0.3 0.58 -0.02
Egg fertilisation (%) 60 ± 6 81 ± 7 13, 12 11 0.003 -0.90
Artificial reefs: X2
Length (mm) 51 ± 4 62 ± 4 11, 20 0.6 0.45 -0.71
Gutted weight (g) 1.3 ± 0.3 2.3 ± 0.4 11, 20 0.9 0.35 -0.70
Wrm 1.3 ± 0.1 1.3 ± 0.1 11, 20 0.3 0.58 +0.01
Hepatosomatic index (*1000) 43 ± 17 21 ± 3 11, 20 5.9 0.01 +0.53
Stomach index (*1000) 16 ± 4 15 ± 3 11, 20 <0.1 0.99 +0.08
Gonadosomatic index (*1000) 12 ± 5 55 ± 19 11, 20 0.8 0.37 -2.39

119
Dietary assessment revealed that almost all clinids had consumed mysid shrimp, both in
Undaria and Ecklonia habitats. Some individuals had also consumed amphipods (Undaria and
Ecklonia), isopods (Undaria only) or decapods (Ecklonia only). Amphipods were the most
frequently identifiable prey items for N. balteatus (Ecklonia only). Animal prey items from N.
balteatus in Undaria habitats were not identifiable, although there was no significant
difference in the weight of stomach contents (Table 5.3).
Fish collected from artificial reefs
Hepatosomatic index was 2.0x higher in H. perspicillatus individuals collected from reefs
stocked with Undaria canopy versus Ecklonia canopy (p = 0.01), while other fitness metrics
were unaffected by kelp canopy treatment (Table 5.3).
DISCUSSION
Some results indicate that the presence of the invasive habitat-forming ecosystem engineer
does not create an ecological trap for native fish. Contrary to Prediction 1, that native fish will
prefer native to invasive kelp, the laboratory habitat choice experiment revealed that two
macroalgal-associated reef fishes were equally willing to utilise shelter provided by native and
invasive kelp. Similarly, fish recruits did not distinguish between artificial reefs stocked with
invasive or native kelp, but preferred both to barren reefs. Our population survey data broadly
supported Prediction 2, that fish communities will be more abundant and diverse in native
kelp than invasive kelp than urchin barrens, with some evidence for higher population
densities in native kelp beds, but invasive kelp patches greatly improved fish abundance and
diversity on heavily urchin-grazed reefs where native canopy-forming macroalgae are absent.
Body condition and reproductive investment metrics indicate that fish inhabiting these
invasive kelp patches have similar or better body condition to those in adjacent native kelp
beds, contradicting Prediction 3 (that fish in native kelp habitats will have higher fitness
metrics than those in invasive kelp habitats), although we did not make any direct comparisons
of mortality rates between habitats. Overall, this invasive habitat-forming kelp appears to
provide valuable habitat for native fishes on urchin-grazed urban-impacted rocky reefs.
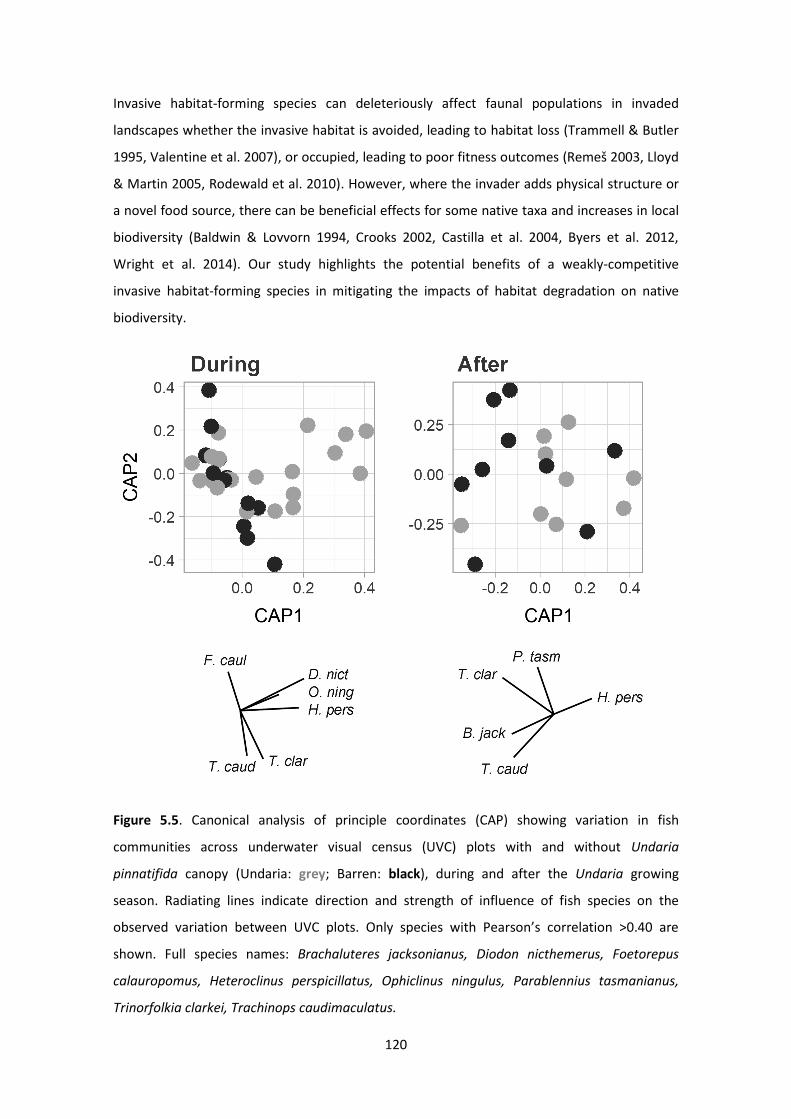
120
Invasive habitat-forming species can deleteriously affect faunal populations in invaded
landscapes whether the invasive habitat is avoided, leading to habitat loss (Trammell & Butler
1995, Valentine et al. 2007), or occupied, leading to poor fitness outcomes (Remeš 2003, Lloyd
& Martin 2005, Rodewald et al. 2010). However, where the invader adds physical structure or
a novel food source, there can be beneficial effects for some native taxa and increases in local
biodiversity (Baldwin & Lovvorn 1994, Crooks 2002, Castilla et al. 2004, Byers et al. 2012,
Wright et al. 2014). Our study highlights the potential benefits of a weakly-competitive
invasive habitat-forming species in mitigating the impacts of habitat degradation on native
biodiversity.
Figure 5.5. Canonical analysis of principle coordinates (CAP) showing variation in fish
communities across underwater visual census (UVC) plots with and without Undaria
pinnatifida canopy (Undaria: grey; Barren: black), during and after the Undaria growing
season. Radiating lines indicate direction and strength of influence of fish species on the
observed variation between UVC plots. Only species with Pearson’s correlation >0.40 are
shown. Full species names: Brachaluteres jacksonianus, Diodon nicthemerus, Foetorepus
calauropomus, Heteroclinus perspicillatus, Ophiclinus ningulus, Parablennius tasmanianus,
Trinorfolkia clarkei, Trachinops caudimaculatus.

121
In this study system, the invasive habitat-former is highly seasonal, and may influence the
reproductive success of local reef fishes to the extent that it coincides with reproductive
provisioning and larval settlement phases (conceptual model: Appendix 5.7). Common
macroalgae-associated reef fishes such as Heteroclinus spp. and N. balteatus inhabit Undaria
patches throughout vitellogenesis and into the spawning period that occurs from late winter to
late spring (Gunn & Thresher 1991; Neira & Sporcic 2002; this study). Our data on stomach
contents, body condition and reproductive investment indicate that adults residing in Undaria
habitat experience comparable food availability to those in native Ecklonia habitats – perhaps
as a result of lower population density and therefore less competition in Undaria patches. This
would be consistent with the ideal free distribution theory (Fretwell & Lucas 1969). Regardless,
it is unlikely that individuals that choose to reside in Undaria habitats during vitellogenesis are
falling into a condition-driven ecological trap. We did find evidence that fertilisation rates were
lower in the ovaries of H. perspicillatus and H. heptaeolus collected from Undaria patches. This
may reflect mate-finding difficulty in small habitat patches that contain few if any conspecifics
and are separated from adjacent patches by urchin barrens. Undaria patches on these
degraded reefs are increasing in size with successive seasons (pers. obs.), and we expect that
this possible Allee effect will ameliorate over time.
We focus on juvenile and adult fish, but larval settlement is also important in assessing the role
of the invasive habitat-former. At present, no literature exists on the settlement preferences
of larval Heteroclinus spp. or Neoodax balteatus. Clinid larvae are at peak densities in the
water column in spring, with settlement occurring during spring-summer (Gunn & Thresher
1991, Neira & Sporcic 2002). This coincides with peak densities of Undaria canopy, and it is
likely that if larvae have similar habitat preferences to adults, large numbers may recruit to
these habitats shortly before the summer die-off of Undaria canopy (Appendix 5.7). Such a loss
of cover on otherwise barren reefs warrants investigation as a potential temporal ecological
trap for clinid recruits. Such a trap may occur if the benefits of inhabiting the invasive kelp are
outweighed by the eventual loss of cover. However, the rapid appearance of late stage
juvenile and adult fish on our plots and artificial reefs in summer—together with anecdotal
reports of clinids ‘rafting’ in unattached macroalgae—suggests that migration is common and
perhaps does not greatly increase mortality rates. As quantifying movement and mortality of
cryptobenthic fishes settling in invaded and non-invaded habitats remains a considerable
challenge, future studies would benefit from developing novel approaches to discriminate
between these two key demographic processes. N. balteatus settlement peaks later in summer
(Neira & Sporcic 2002), by which time Undaria thalli have regressed to sporophyll and
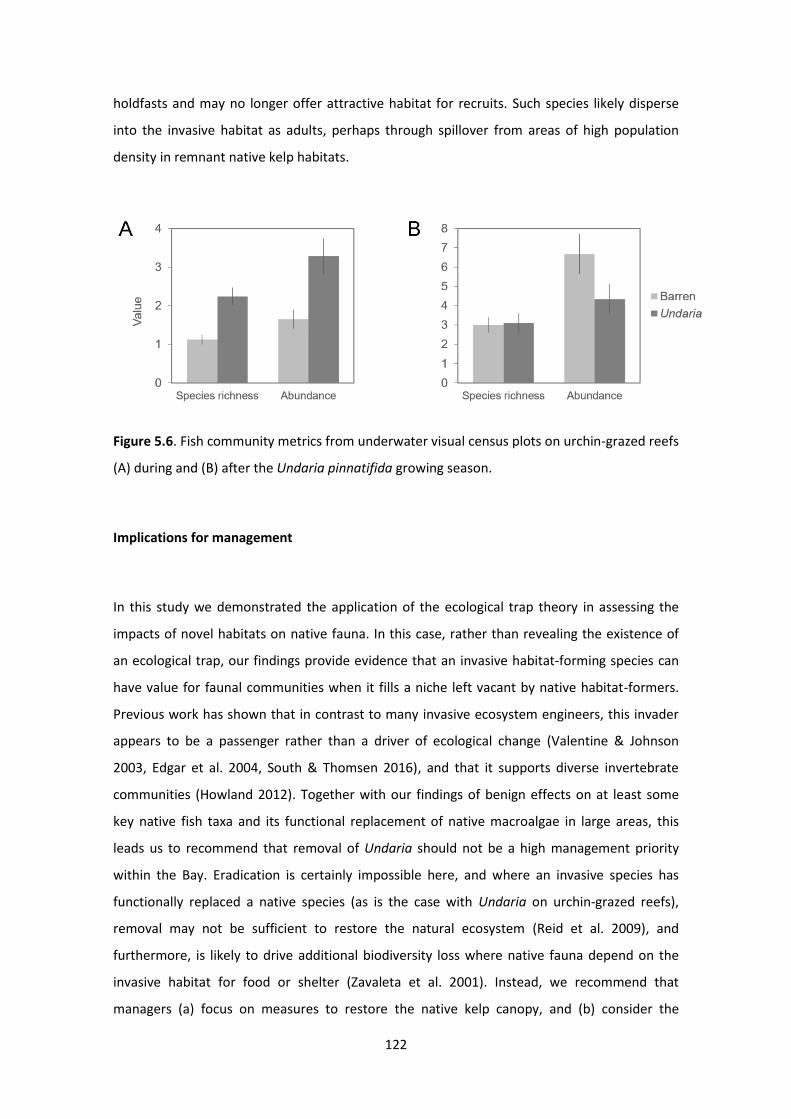
122
holdfasts and may no longer offer attractive habitat for recruits. Such species likely disperse
into the invasive habitat as adults, perhaps through spillover from areas of high population
density in remnant native kelp habitats.
Figure 5.6. Fish community metrics from underwater visual census plots on urchin-grazed reefs
(A) during and (B) after the Undaria pinnatifida growing season.
Implications for management
In this study we demonstrated the application of the ecological trap theory in assessing the
impacts of novel habitats on native fauna. In this case, rather than revealing the existence of
an ecological trap, our findings provide evidence that an invasive habitat-forming species can
have value for faunal communities when it fills a niche left vacant by native habitat-formers.
Previous work has shown that in contrast to many invasive ecosystem engineers, this invader
appears to be a passenger rather than a driver of ecological change (Valentine & Johnson
2003, Edgar et al. 2004, South & Thomsen 2016), and that it supports diverse invertebrate
communities (Howland 2012). Together with our findings of benign effects on at least some
key native fish taxa and its functional replacement of native macroalgae in large areas, this
leads us to recommend that removal of Undaria should not be a high management priority
within the Bay. Eradication is certainly impossible here, and where an invasive species has
functionally replaced a native species (as is the case with Undaria on urchin-grazed reefs),
removal may not be sufficient to restore the natural ecosystem (Reid et al. 2009), and
furthermore, is likely to drive additional biodiversity loss where native fauna depend on the
invasive habitat for food or shelter (Zavaleta et al. 2001). Instead, we recommend that
managers (a) focus on measures to restore the native kelp canopy, and (b) consider the

123
potential for beneficial effects of introduced habitat-formers, particularly in degraded
environments, and when possible, focus control or eradication efforts on invaders that are
highly competitive in the invaded environment and/or drive declines in biodiversity of native
species.
ACKNOWLEDGEMENTS
We wish to thank Dean Chamberlain, Seann Chia, Ben Cleveland, Emily Fobert, Molly Fredle,
Akiva Gebler, Kevin Jensen, Valeriya Komyakova, Nina Kriegisch, Kevin Menzies, Jack O’Connor,
Simon Reeves, Juan Manuel Valero Rodriguez, Kyler Tan, Chris Taylor and João Teixiera for
their assistance with fieldwork. Members of the SALTT and REEF labs provided useful
comments on the manuscript. All work was conducted in accordance with permits from the
Victorian Fisheries Authority (RP919) and the University of Melbourne’s Faculty of Science
Animal Ethics Committee (1413193 and 1413133). This research was funded by grants to LB
from the Holsworth Wildlife Research Endowment, the PADI Foundation, and the Victorian
Environmental Assessment Council. The authors have no competing interests to declare.
REFERENCES
Anderson M, Gorley R, Clarke R (2008) Permanova+ for Primer: guide to software and
statistical methods. Plymouth.
Baldwin JR, Lovvorn JR (1994) Expansion of seagrass habitat by the exotic Zostera japonica, and
its use by dabbling ducks and brant in Boundary Bay, British Columbia. Mar Ecol Prog Ser
103:119–127
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed effects models using lme4.
J Stat Softwar 67:1–48
Battin J (2004) When good animals love bad habitats: ecological traps and the conservation of
animal populations. Conserv Biol 18:1482–1491
Byers JE, Gribben PE, Yeager C, Sotka EE (2012) Impacts of an abundant introduced ecosystem
engineer within mudflats of the southeastern US coast. Biol Invasions 14:2587–2600
Campbell SJ, Burridge TR (1998) Occurrence of Undaria pinnatifida (Phaeophyta: Laminariales)
in Port Phillip Bay, Victoria, Australia. Mar Freshw Res 49:379–381

124
Castilla JC, Lagos NA, Cerda M (2004) Marine ecosystem engineering by the alien ascidian
Pyura praeputialis on a mid-intertidal rocky shore. Mar Ecol Prog Ser 268:119–130
Cren ED Le (1951) The length-weight relationship and seasonal cycle in gonad weight and
condition in the perch (Perca fluviatilis). J Anim Ecol 20:201–219
Crooks JA (2002) Characterizing ecosystem-level consequences of biological invasions: the role
of ecosystem engineers. Oikos 97:153–166
Davidson AD, Campbell ML, Hewitt CL, Schaffelke B (2015) Assessing the impacts of
nonindigenous marine macroalgae: an update of current knowledge. Bot Mar 58:55–79
Dijkstra JA, Harris LG, Mello K, Litterer A, Wells C, Ware C (2017) Invasive seaweeds transform
habitat structure and increase biodiversity of associated species. J Ecol 105:1668–1678
Edgar GJ, Barrett NS, Morton AJ, Samson CR (2004) Effects of algal canopy clearance on plant,
fish and macroinvertebrate communities on eastern Tasmanian reefs. J Exp Mar Biol Ecol
312:67–87
Farrell P, Fletcher RL (2006) An investigation of dispersal of the introduced brown alga Undaria
pinnatifida (Harvey) Suringar and its competition with some species on the man-made
structures of Torquay Marina (Devon, UK). J Exp Mar Biol Ecol 334:236–243
Filbee-Dexter K, Wernberg T (2018) Rise of turfs: a new battlefront for globally declining kelp
forests. Bioscience 68:64–76
Fretwell SD, Lucas HL (1969) On territorial behavior and other factors influencing habitat
distribution in birds. Acta Biotheor 19:16–36
Gribben PE, Wright JT (2006) Invasive seaweed enhances recruitment of a native bivalve: roles
of refuge from predation and the habitat choice of recruits. Mar Ecol Prog Ser 318:177–
185
Gunn JS, Thresher RE (1991) Viviparity and the reproductive ecology of clinid fishes (Clinidae)
from temperate Australian waters. Environ Biol Fishes 31:323–344
Hale R, Swearer SE (2016) Ecological traps: current evidence and future directions. Proc R Soc B
Biol Sci 283:494–499
Hale R, Treml EA, Swearer SE (2015) Evaluating the metapopulation consequences of
ecological traps. Proc R Soc London B Biol Sci 282:20142930
Harrison XA (2014) Using observation-level random effects to model overdispersion in count
data in ecology and evolution. PeerJ 2:e616
Horne B van (1983) Density as a misleading indicator of habitat quality. J Wildl Manage
47:893–901
Howland S (2012) Variation and impacts on biodiversity in kelp holdfast assemblages: a

125
comparison of the introduced Undaria pinnatifida and the indigenous Ecklonia radiata.
University of Melbourne
Irigoyen AJ, Eyras C, Parma AM (2011) Alien algae Undaria pinnatifida causes habitat loss for
rocky reef fishes in north Patagonia. Biol Invasions 13:17–24
Jung CA, Dwyer PD, Minnegal M, Swearer SE (2011) Perceptions of environmental change over
more than six decades in two groups of people interacting with the environment of Port
Phillip Bay, Australia. Ocean Coast Manag 54:93–99
Kokko H, Sutherland WJ (2001) Ecological traps in changing environments: ecological and
evolutionary consequences of a behaviourally mediated Allee effect. Evol Ecol Res 3:537–
551
Leij R de, Epstein G, Brown MP, Smale DA (2017) The influence of native macroalgal canopies
on the distribution and abundance of the non-native kelp Undaria pinnatifida in natural
reef habitats. Mar Biol 164:156
Lenth R V (2016) Least-squares means: the R package lsmeans. J Stat Softw 69:1–33
Ling SD, Ibbott S, Sanderson JC (2010) Recovery of canopy-forming macroalgae following
removal of the enigmatic grazing sea urchin Heliocidaris erythrogramma. J Exp Mar Biol
Ecol 395:135–146
Lloyd JD, Martin TE (2005) Reproductive success of chestnut-collared longspurs in native and
exotic grassland. Condor 107:363–374
MacDougall AS, Turkington R (2005) Are invasive species the drivers of change or passengers
of change in degraded ecosystems? Ecology 86:42–55
McDermott CJ, Shima JS (2006) Ontogenetic shifts in microhabitat preference of the temperate
reef fish Forsterygion lapillum: implications for population limitation. Mar Ecol Prog Ser
320:259–266
Neira FJ, Sporcic MI (2002) Use of ichthyoplankton ecology to evaluate ecosystem changes: a
case study in a large, semi-enclosed Australian bay. Mar Freshw Res 53:339
Patten MA, Kelly JF (2010) Habitat selection and the perceptual trap. Ecol Appl 20:2148–2156
Primo C, Hewitt CL, Campbell ML (2010) Reproductive phenology of the introduced kelp
Undaria pinnatifida (Phaeophyceae, Laminariales) in Port Phillip Bay (Victoria, Australia).
Biol Invasions 12:3081–3092
Pulliam HR (1988) Sources, sinks, and population regulation. Am Nat 132:652–661
Pyšek P, Jarošík V, Hulme PE, Pergl J, Hejda M, Schaffner U, Vilà M (2012) A global assessment
of invasive plant impacts on resident species, communities and ecosystems: the
interaction of impact measures, invading species’ traits and environment. Glob Chang

126
Biol 18:1725–1737
R Core Team (2017) R: A language and environment for statistical computing.
Raffo MP, Eyras MC, Iribarne OO (2009) The invasion of Undaria pinnatifida to a Macrocystis
pyrifera kelp in Patagonia (Argentina, south-west Atlantic). J Mar Biol Assoc UK 89:1571–
1580
Raffo MP, Russo V Lo, Schwindt E (2014) Introduced and native species on rocky shore
macroalgal assemblages: Zonation patterns, composition and diversity. Aquat Bot
112:57–65
Reid AM, Morin L, Downey PO, French K, Virtue JG (2009) Does invasive plant management aid
the restoration of natural ecosystems? Biol Conserv 142:2342–2349
Remeš V (2003) Effects of exotic habitat on nesting success, territory density, and settlement
patterns in the blackcap (Sylvia atricapilla). Conserv Biol 17:1127–1133
Robertson BA, Hutto RL (2006) A framework for understanding ecological traps and an
evaluation of existing evidence. Ecology 87:1075–1085
Robertson BA, Rehage JS, Sih A (2013) Ecological novelty and the emergence of evolutionary
traps. Trends Ecol Evol 28:552–560
Rodewald AD, Shustack DP, Hitchcock LE (2010) Exotic shrubs as ephemeral ecological traps for
nesting birds. Biol Invasions 12:33–39
Sampson J, Easton A, Singh M (2014) Port Phillip Bay. In: Wolanski E (ed) Estuaries of Australia
in 2050 and Beyond. Springer, p 49–68
Schaffelke B, Hewitt CL (2007) Impacts of introduced seaweeds. Bot Mar 50:397–417
Sih A, Ferrari MCO, Harris DJ (2011) Evolution and behavioural responses to human-induced
rapid environmental change. Evol Appl 4:367–387
South PM, Thomsen MS (2016) The ecological role of invading Undaria pinnatifida: an
experimental test of the driver–passenger models. Mar Biol 163:175
Thomsen MS, Wernberg T, Tuya F, Silliman BR (2009) Evidence for impacts of nonindigenous
macroalgae: a meta-analysis of experimental field studies. J Phycol 45:812–819
Trammell MA, Butler JL (1995) Effects of exotic plants on native ungulate use of habitat. J Wildl
Manage 59:808–816
Valentine JP, Johnson CR (2003) Establishment of the introduced kelp Undaria pinnatifida in
Tasmania depends on disturbance to native algal assemblages. J Exp Mar Biol Ecol
295:63–90
Valentine LE, Roberts B, Schwarzkopf LIN (2007) Mechanisms driving avoidance of non-native
plants by lizards. J Appl Ecol 44:228–237

127
Wickham H (2009) ggplot2: elegant graphics for data analysis. Springer-Verlag New York
Willis T, Anderson M (2003) Structure of cryptic reef fish assemblages: relationships with
habitat characteristics and predator density. Mar Ecol Prog Ser 257:209–221
Willis TJ, Babcock RC (2000) A baited underwater video system for the determination of
relative density of carnivorous reef fish. Mar Freshw Res 51:755–763
Wong BBM, Candolin U (2014) Behavioral responses to changing environments. Behav Ecol
26:665–673
Wright JT, Byers JE, DeVore JL, Sotka EE (2014) Engineering or food? mechanisms of facilitation
by a habitat‐forming invasive seaweed. Ecology 95:2699–2706
Zavaleta ES, Hobbs RJ, Mooney HA (2001) Viewing invasive species removal in a whole-
ecosystem context. Trends Ecol Evol 16:454–459

128
CHAPTER SIX: GENERAL DISCUSSION
Incorporating data on individual-level responses to environmental change can fundamentally
improve our understanding of associations between novel habitats and wild animal
populations, and especially the role of novel habitats for population persistence in a source–
sink metapopulation framework. The ecological trap literature highlights the importance of
combining individual- and population-level data to distinguish between attractive population
sources, unattractive population sources, attractive population sinks and unattractive
population sinks.
HOW PREVALENT ARE ECOLOGICAL TRAPS IN THE MARINE ENVIRONMENT?
Ecological traps are probably more common than the current evidence indicates. Clear
demonstrations of traps are rare, but it would be surprising if this reflects consistently
adaptive habitat selection by animals in novel and degraded environments (Sih et al. 2011,
Robertson et al. 2013, Sih 2013). More likely it reflects a lack of research effort on the topic,
with the vast majority of habitat association studies focusing on community-level survey data.
As a result, most researchers are not obtaining suitable estimates of habitat preference, nor
studying the fitness consequences of those habitat selection decisions. Furthermore, Battin
(2004) noted that researchers in terrestrial systems frequently presented findings that were
consistent with the existence of an ecological trap but did not interpret the findings within the
ecological trap framework. Similar cases exist within the marine literature: several of the
examples I cite in this thesis as evidence for marine ecological traps were not interpreted as
such by the authors, including flatfish and crabs selecting oil-contaminated sediment (Moles et
al. 1994, Moles & Norcross 1998, Moles & Stone 2002), bivalves selecting sediment modified
by Caulerpa taxifolia (Wright & Gribben 2008, Gribben et al. 2009a, 2009b, Byers et al. 2010),
and fish recruits avoiding reefs with resident native predators but not those with invasive
lionfish (Benkwitt 2017). In some cases, authors referred to probable ecological trap habitats
as ‘population sinks’ (e.g. Gribben et al. 2009b), consistent with greater awareness of the
source-sink framework, or otherwise noted likely deleterious population effects. However, as
ecological trap models demonstrate, an attractive population sink (ecological trap) has more
severe implications for population persistence than a ‘traditional’ non-preferred population

129
sink as it actively draws in individuals from surrounding higher quality areas (Battin 2004,
Abrams et al. 2012, Hale et al. 2015). In other words, non-preferred population sinks are likely
to conform to the ideal free distribution (with each individual optimising its fitness within
available habitats: Fretwell & Lucas 1969), while ecological traps do not, causing fauna
populations to underperform relative to expectations.
Additional evidence for the likely prevalence of ecological traps comes from the rate at which
researchers looking for traps find them. While it may be impossible to account for bias in
research effort (i.e. researchers selecting systems that are likely to be vulnerable to ecological
traps, or setting aside the ecological trap hypothesis when no evidence is found), there are
notably few cases of researchers looking for traps and not finding them (but see Dempster et
al. 2011; Reubens et al. 2013). In Chapters 3-5, I investigated three novel marine systems that I
considered to be vulnerable to the formation of ecological traps. In Chapter 3 I found some
limited evidence that offspring of Atlantic cod from an area of high salmon farming density
produced smaller eggs and larvae, potentially leading to reduced fitness. In Chapter 4, an
absence of small Asterias seastars within mussel farms was consistent with either high
mortality or emigration in this habitat, while adult Asterias may be avoiding a habitat rich in
food but also potential predators. In Chapter 5, I concluded that juvenile and adult weedfish
and weed whiting are probably making broadly adaptive habitat-selection decisions in Undaria
habitats (consistent with ideal free distribution), but that there are possible Allee effects in
small patches and that loss of canopy during summer senescence might be costly for weedfish
larvae that recruit to the Undaria canopy in spring if the value of the Undaria canopy is
outweighed by its eventual disappearance.
Intriguingly, in all cases, my findings indicated that fauna associated with novel marine habitats
were most likely either making adaptive habitat selection decisions or falling into ecological
traps. I found no evidence that any of these novel habitats were likely to function as
perceptual traps (Fig. 6.1). This is consistent with the current body of literature, in which novel
habitats are more commonly found to be ecological traps than perceptual traps (but see
Patten and Kelly 2010). I see three possible reasons for this trend: (1) novel habitats may, on
average, offer poorer fitness outcomes than natural habitats for native species due to a lack of
evolutionary adaptation to the novel habitat and likely disadvantages relative to non-native
competitors or predators (Byers 2002, Crooks et al. 2011); (2) if a novel habitat appears similar
to an evolutionarily-familiar habitat, individuals will likely attempt to utilise it; and (3) an
abundance of a native species in a novel habitat is more likely to arouse the interest of

130
ecologists than an absence, as we are accustomed to native taxa avoiding novel habitats and
implicitly assume this to be a broadly adaptive response.
Figure 6.1. Conceptual diagram showing speculative positioning of responses of focal species
to novel habitats, indicating potential differences in responses and fitness outcomes across life
stages (Asterias and Coscinasterias at shellfish farms; Heteroclinus and Neoodax in Undaria
canopy; Atlantic cod at salmon farms) placed within the ecological trap framework. Species
placed within the green area exhibit adaptive habitat selection responses. Juvenile
Coscinasterias were not observed inside the shellfish farm, and it is unclear whether this
reflects habitat preferences or high mortality.
HOW DOES THE ECOLOGICAL TRAP CONCEPT RELATE TO THE ATTRACTION–PRODUCTION
DEBATE?
Ultimately, the ecological trap and attraction–production concepts are directed toward the
same objective: understanding the role of a given habitat for population growth, with a focus
on ecologically-novel habitats. The ecological trap concept pre-dates both source–sink models
and the attraction–production debate (Dwernychuk & Boag 1972, Gates & Gysel 1978), but the
source–sink model experienced more rapid uptake among ecologists, while the attraction–
production debate arose in response to questions about the role of artificial reefs for
production of targeted fish species during the 1980s (Lindberg 1997).

131
Uptake of the ecological trap concept has been slow in the fisheries management literature,
but in my view, it offers something that the attraction–production distinction does not. The
typical application of the attraction–production models looks at two questions: are fish
attracted to the given habitat, and does the existence of this habitat lead to more fish being
produced? Under the attraction hypothesis, fish move from surrounding areas to the new
habitat, with no net increase in production. Under the production hypothesis, the new habitat
increases the carrying capacity of a habitat-limited environment and leads to increased fish
production. These two hypothesis are not mutually exclusive and likely co-occur in many cases,
but the limitation of the attraction–production framework is that researchers are liable to miss
a third possibility – that poor individual fitness outcomes in an attractive novel habitat will
create an ecological trap that leads to serious negative effects on fish production, such that it
would be better for the habitat not to exist at all (Reubens et al. 2014). The ecological trap
assessment framework (see Fig. 1.1) accounts for this possibility, and its corollary, an
unattractive but high quality habitat that is underutilised (termed a perceptual trap by Patten
and Kelly 2010). Gutzler et al. (2015) considered both frameworks in their assessment of the
role of casitas for targeted lobsters as individuals and as a population, while Reubens et al.
(2014) proposed a hybrid ‘attraction–ecological trap–production’ framework for assessing the
role of artificial reefs for fish populations, within which researchers ask the following
questions:
(1) Does attraction towards the artificial reef occur?
(2) If there is attraction, is it age-group specific?
(3) Which mechanisms or processes influence production in the ecosystem investigated,
and are these mechanisms/processes affected by the artificial reef?
(4) What is the species-specific behavioural ecology in this ecosystem?
(5) If there is production, is it sufficient to offset associated fishing mortality?
This integration of two assessment frameworks, with two largely independent bodies of
research focusing on terrestrial birds (ecological traps) and artificial reefs (attraction–
production), can also be expressed as a conceptual diagram (Fig. 6.2). The four scenarios
outlined in the diagram are extreme outcomes, and intermediate outcomes are possible. For
example, newly-created, unattractive, low quality habitats may have a small positive effect on
production in habitat-limited systems if they offer a sufficient fitness value for individuals
spilling over from densely-populated preferred habitats (adaptively following the ideal free
distribution).

132
Relative fitness outcomes for individuals
Good Poor
Relative habitat attractiveness
Attractive Source ↑ Ecological trap ↓
Unattractive Perceptual trap — Sink —
Figure 6.2. Conceptual representation of the ecological trap framework applied to an
assessment of the role of artificial reefs for fish production. “↑” indicates an increase in fish
production (e.g. fish utilise a viable new habitat in a habitat-limited system), “↓” indicates a
decrease in fish production (e.g. fish are attracted to a habitat that offers low availability of
food or mates, or confers high mortality), and “—" indicates a neutral effect on fish production
(as the new habitat is not utilised).
BETTER LEFT ALONE? LESSONS FOR THE MANAGEMENT OF HABITAT-FORMING INVASIVE
SPECIES
Most human-mediated species introductions do not establish, but of those that do, a few will
become invasive (Richardson & Pyšek 2012). Such invasions may be an important factor in
native species extinctions worldwide (Clavero & García-Berthou 2005), and some particularly
influential invasive ecosystem engineers have driven phase shifts in natural ecosystems
(Crooks 2002). Management of non-native arrivals often begins with immediate eradication
attempts, but once the window for effective eradication has passed, management strategies
must be reassessed. In cases where the invader is an opportunistic passenger rather than a
driver of ecological change, control measures may be neither effective nor desirable (Zavaleta
et al. 2001, Didham et al. 2005, MacDougall & Turkington 2005). This may be especially true
where invasive habitat-forming species replace native habitat-formers and native animals
come to rely on the invasive habitat (Zavaleta et al. 2001). However, a major limitation in our
understanding of the impacts of invasive habitat-formers is a scarcity of data on habitat
preferences and fitness outcomes for native fauna inhabiting invasive habitats.

133
Figure 6.3. Current distribution of Undaria pinnatifida within Port Phillip Bay in 2017. Red
markers denote documented established populations on rocky reefs or marine infrastructure;
blue markers denote recent infestations. A satellite population has also established at Apollo
Bay (97 km SW) and another was discovered and eradicated at Flinders (45 km SE). Both
expansions were likely the result of boat transport.
The invasive kelp Undaria pinnatifida, a habitat-forming ecosystem engineer, provides an
illustrative case study. Despite being one of only two macroalgal taxa included on the IUCN’s
list of 100 of the world’s worst invasive alien species (Lowe et al. 2000), the subsequent
literature on invasive Undaria populations in Australia, New Zealand and Europe indicates that
this species is a passenger of general ecological degradation, taking advantage of rocky reef
substrate left vacant by declining native kelp canopy but only persisting at low population
densities in intact native macroalgal habitats (Valentine & Johnson 2003, Edgar et al. 2004,
South & Thomsen 2016, Kriegisch et al. 2016, de Leij et al. 2017, South et al. 2017). Effects of
the invasive canopy on native faunal biodiversity are less well known, but in South America,
invasive Undaria cover was associated with elevated macroinvertebrate species richness,
diversity and abundance compared to plots where early stage Undaria recruits were removed
to prevent canopy formation (Irigoyen et al. 2011). Similarly, work in Australia found that
invasive Undaria and native Ecklonia radiata holdfasts contain comparable abundance and
species richness of epibenthic invertebrates (Howland 2012), indicating that Undaria is likely to
have neutral or positive effects on biodiversity in degraded areas. In Chapter Five, I

134
demonstrated that the presence of Undaria on degraded reefs is associated with elevated fish
biodiversity and provides viable habitat for a variety of endemic fish species in the months
leading up to spring spawning. This work is a rare assessment of outcomes for native species
utilising invasive habitats.
Despite this growing body of evidence that Undaria canopy is utilised by native fish and
invertebrates, coupled with broadly benign effects on native macroalgal communities,
authorities are undertaking ongoing eradication efforts at local sites. Such efforts are unlikely
to be effective in the long term (e.g. Hewitt et al. 2005), and as they necessarily occur during
the Undaria growing season in spring and summer, are likely to displace resident macroalgae-
associated fish in spawning condition. Accordingly, I recommend that eradication efforts cease
in locations where Undaria is established (see Fig. 6.3 for distribution in Port Phillip Bay), and
instead focus on eradicating satellite populations using approaches modelled on successful
removals of localised infestations at Flinders (Parry & Cohen 2001) and the Chatham Islands
(Wotton et al. 2004). Where the invader is established, management efforts may be better
directed toward promoting the recovery of Ecklonia and other native macrophytes that
compete with Undaria (Ling et al. 2009, Carnell & Keough 2016, Kriegisch et al. 2016), for
example by investigating methods to reduce sea urchin grazing pressure (e.g. Pert et al. 2018)
and other stressors.
Farming of Undaria may also be considered in areas where the invader is already abundant
and where farming is unlikely to increase the severity of any ecological impacts. Accordingly,
the New Zealand government allowed farming of Undaria in several heavily infested areas in
2012, with interest along similar lines in Europe (Peteiro et al. 2016). Farming would greatly
increase propagule pressure locally if the reproductive plants reach maturity before
harvesting, so the potential for increased local impacts as well as the risk of accelerated range
expansion should be considered carefully. Undaria spores do not naturally disperse large
distances (Forrest et al. 2000), but caution is required given the likelihood of detached mature
plants drifting large distances (Russell et al. 2008), and a history of long distance dispersal
mediated by maritime transport.
Undaria may not be alone in being more benign than initially feared. A recent global review of
non-native seaweeds found that introduced seaweeds generally had negative effects on
macroalgal diversity, but neutral or positive effects on faunal communities (Thomsen et al.
2016). In many cases, there is evidence that the spread of non-native taxa is facilitated by prior
declines in native cover (Ceccherelli et al. 2014, Gennaro & Piazzi 2014, Luigi & Giulia 2017).

135
More research is needed to understand the value of non-native habitats for native fauna, and
to avoid conflating effects of non-native habitat-formers with negative effects of other forms
of HIREC.
WHERE TO NEXT FOR THIS WORK?
The meta-analysis in Chapter Two revealed substantial gaps in our understanding of the
impacts of marine and freshwater aquaculture on wildlife. In particular, studies that rigorously
demonstrate attractive or repulsive effects of farms are rare, as are studies quantifying direct
or indirect fitness effects on wild taxa. Chapter Three addressed one of these key knowledge
gaps—effects of farm-association on reproductive fitness of wild fish—and revealed some
evidence for negative effects, but the study was nonetheless limited in spatial replication,
experimental power and duration of larval rearing. Ideally, the geographical extent of this work
should be expanded, with additional resources to support larger sample sizes, and/or raise
larvae to juvenile or adult stages to test for delayed effects of farm-association on offspring.
Adult mortality is also important in assessing the impacts of aquaculture on wild fish
populations, especially where farms bring elevated infection, predation and fishing mortality
rates. Long term acoustic tracking of adult fish within farm-affected or non-affected areas,
replicated across multiple locations, may be the most viable approach for quantifying mortality
and emigration in semi-enclosed systems such as fjords (Olsen & Moland 2011, Olsen et al.
2012, Fernández-Chacón et al. 2015).
In Chapter Four, I showed that despite an underlying attraction to shellfish prey and benefits
of high food availability for individuals residing inside shellfish farms, the invasive seastar
Asterias amurensis was not more abundant inside farms. Predator avoidance experiments
together with a truncated size distribution indicated that the threat of predation by the native
seastar Coscinasterias muricata may be preventing the invader from exploiting the food-rich
resource. However, I was unable to clearly partition the effects of behavioural predator
avoidance and mortality. A follow-up study quantifying relative predation risk for tethered
seastars, as well as mass mark-recapture of seastars at locations inside and outside farms
(Loosanoff 1937, Lamare et al. 2009, Barahona & Navarrete 2010) may shed light on the
relative importance of these processes.

136
In Chapter Five, I provided estimates of habitat preference (via a controlled habitat choice
experiment and recruitment patterns to mesocosm reefs) and relative fitness (via condition
and fecundity metrics) for native reef fish inhabiting native and invasive kelp canopy, but
further work is required to quantify patterns of movement, site fidelity and mortality,
especially during the summer senescence of the Undaria canopy. Acoustic tags can be
appropriate for tracking movement of large-bodied taxa (Parsons et al. 2003), while site
fidelity of smaller site-attached taxa may be estimated by a mark-recapture study using visible
implant elastomer tags (Willis et al. 2001). However, a pilot mark-recapture study of the
common weedfish Heteroclinus perspicillatus translocated to mesocosm reefs in Chapter Five
indicated low retention rates, with simultaneous high immigration of new juveniles and adults
indicating that this species is highly transient, consistent with previous work tracking temporal
trends in fish communities on artificial seaweed beds (Jenkins & Sutherland 1997). Novel
techniques may be required to track movement and mortality of small cryptobenthic fishes
during and after the Undaria growing season. The little weed whiting Neoodax balteatus may
be a better focal species for a study of survivorship rates in native and invasive kelp habitats,
as dominant terminal phase males maintain harems and appear to be more strongly site-
attached (pers. obs.), such that it may be reasonable to infer mortality rates from the loss of
dominant individuals, although a full assessment of the ecological role of Undaria patches
should consider a wider suite of associated vertebrate and invertebrate fauna.
CONCLUSION
A reliance on population- and community-level metrics can result in a failure to detect
important individual-level processes that influence population growth and persistence, such as
habitat selection, survivorship and reproductive output. Wherever possible, researchers
assessing the role of novel or degraded habitats as population sources or sinks should
complement population- and community-level metrics with direct or indirect measures of
habitat preference and individual fitness. Applying the ecological trap assessment framework
can also improve our understanding of source–sink dynamics by accounting for maladaptive
habitat selection decisions that can lead to attractive population sinks or underutilised
population sources. Correctly characterising the value of specific habitats for fauna will assist
in targeting of management measures, for example by: (1) protecting attractive, highly
productive habitats, (2) disarming ecological traps by removing attractive cues or drivers of

137
poor fitness outcomes, (3) investigating ecological engineering approaches to increase
utilisation of perceptual traps by native fauna, or (4) leveraging ecological traps to assist in the
control of non-native fauna.
REFERENCES
Abrams PA, Ruokolainen L, Shuter BJ, McCann KS (2012) Harvesting creates ecological traps:
consequences of invisible mortality risks in predator-prey metacommunities. Ecology
93:281–293
Barahona M, Navarrete SA (2010) Movement patterns of the seastar Heliaster helianthus in
central Chile: relationship with environmental conditions and prey availability. Mar Biol
157:647–661
Battin J (2004) When good animals love bad habitats: ecological traps and the conservation of
animal populations. Conserv Biol 18:1482–1491
Benkwitt CE (2017) Predator effects on reef fish settlement depend on predator origin and
recruit density. Ecology 98:896–902
Byers JE (2002) Impact of non-indigenous species on natives enhanced by anthropogenic
alteration of selection regimes. Oikos 97:449–458
Byers JE, Wright JT, Gribben PE (2010) Variable direct and indirect effects of a habitat-
modifying invasive species on mortality of native fauna. Ecology 91:1787–1798
Carnell PE, Keough MJ (2016) The influence of herbivores on primary producers can vary
spatially and interact with disturbance. Oikos 125:1273–1283
Ceccherelli G, Pinna S, Cusseddu V, Bulleri F (2014) The role of disturbance in promoting the
spread of the invasive seaweed Caulerpa racemosa in seagrass meadows. Biol Invasions
16:2737–2745
Clavero M, García-Berthou E (2005) Invasive species are a leading cause of animal extinctions.
Trends Ecol Evol 20:110
Crooks JA (2002) Characterizing ecosystem-level consequences of biological invasions: the role
of ecosystem engineers. Oikos 97:153–166
Crooks JA, Chang AL, Ruiz GM (2011) Aquatic pollution increases the relative success of
invasive species. Biol Invasions 13:165–176
Dempster T, Sanchez-Jerez P, Fernandez-Jover D, Bayle-Sempere JT, Nilsen R, Bjørn P-A, Uglem
I (2011) Proxy measures of fitness suggest coastal fish farms can act as population

138
sources and not ecological traps for wild gadoid fish. PLoS One 6:e15646
Didham RK, Tylianakis JM, Hutchison MA, Ewers RM, Gemmell NJ (2005) Are invasive species
the drivers of ecological change? Trends Ecol Evol 20:470–474
Dwernychuk LW, Boag DA (1972) Ducks nesting in association with gulls - an ecological trap?
Can J Zool 50:559–563
Edgar GJ, Barrett NS, Morton AJ, Samson CR (2004) Effects of algal canopy clearance on plant,
fish and macroinvertebrate communities on eastern Tasmanian reefs. J Exp Mar Bio Ecol
312:67–87
Fernández-Chacón A, Moland E, Espeland SH, Olsen EM (2015) Demographic effects of full vs.
partial protection from harvesting: inference from an empirical before-after control-
impact study on Atlantic cod. J Appl Ecol 52:1206–1215
Forrest BM, Brown SN, Taylor MD, Hurd CL, Hay CH (2000) The role of natural dispersal
mechanisms in the spread of Undaria pinnatifida (Laminariales, Phaeophyceae).
Phycologia 39:547–553
Fretwell SD, Lucas HL (1969) On territorial behavior and other factors influencing habitat
distribution in birds. Acta Biotheor 19:16–36
Gates JE, Gysel LW (1978) Avian nest dispersion and fledging success in field-forest ecotones.
Ecology 59:871–883
Gennaro P, Piazzi L (2014) The indirect role of nutrients in enhancing the invasion of Caulerpa
racemosa var cylindracea. Biol Invasions 16:1709–1717
Gribben PE, Wright JT, O’Connor WA, Doblin MA, Eyre B, Steinberg PD (2009a) Reduced
performance of native infauna following recruitment to a habitat-forming invasive
marine alga. Oecologia 158:733–745
Gribben PE, Wright JT, O’Connor WA, Steinberg P (2009b) Larval settlement preference of a
native bivalve: the influence of an invasive alga versus native substrata. Aquat Biol 7:217–
227
Gutzler BC, Butler MJ IV, Behringer DC (2015) Casitas: a location-dependent ecological trap for
juvenile Caribbean spiny lobsters, Panulirus argus. ICES J Mar Sci 72:i177–i184.
Hale R, Treml EA, Swearer SE (2015) Evaluating the metapopulation consequences of
ecological traps. Proc R Soc London B Biol Sci 282:20142930
Hewitt CL, Campbell ML, McEnnulty F, Moore KM, Murfet NB, Robertson B, Schaffelke B (2005)
Efficacy of physical removal of a marine pest: the introduced kelp Undaria pinnatifida in a
Tasmanian Marine Reserve. Biol Invasions 7:251–263
Howland S (2012) Variation and impacts on biodiversity in kelp holdfast assemblages: a

139
comparison of the introduced Undaria pinnatifida and the indigenous Ecklonia radiata.
University of Melbourne
Irigoyen AJ, Trobbiani G, Sgarlatta MP, Raffo MP (2011) Effects of the alien algae Undaria
pinnatifida (Phaeophyceae, Laminariales) on the diversity and abundance of benthic
macrofauna in Golfo Nuevo (Patagonia, Argentina): potential implications for local food
webs. Biol Invasions 13:1521–1532
Jenkins G., Sutherland C. (1997) The influence of habitat structure on nearshore fish
assemblages in a southern Australian embayment: colonisation and turnover rate of
fishes associated with artificial macrophyte beds of varying physical structure. J Exp Mar
Bio Ecol 218:103–125
Kriegisch N, Reeves S, Johnson CR, Ling SD, Jernakoff P (2016) Phase-shift dynamics of sea
urchin overgrazing on nutrified reefs (BD Russell, Ed.). PLoS One 11:e0168333
Lamare MD, Channon T, Cornelisen C, Clarke M (2009) Archival electronic tagging of a
predatory sea star - testing a new technique to study movement at the individual level. J
Exp Mar Bio Ecol 373:1–10
Leij R de, Epstein G, Brown MP, Smale DA (2017) The influence of native macroalgal canopies
on the distribution and abundance of the non-native kelp Undaria pinnatifida in natural
reef habitats. Mar Biol 164:156
Lindberg WJ (1997) Can science resolve the attraction-production issue? Fisheries 22:10–13
Ling SD, Johnson CR, Frusher SD, Ridgway KR (2009) Overfishing reduces resilience of kelp beds
to climate-driven catastrophic phase shift. Proc Natl Acad Sci USA 106:22341–22345
Loosanoff VL (1937) Use of nile blue sulfate in marking starfish. Science 85:412
Lowe S, Browne M, Boudjelas S, Poorter M De (2000) 100 of the world’s worst invasive alien
species: a selection from the global invasive species database. Auckland
Luigi P, Giulia C (2017) Eutrophication affects the resistance of fucoids to an introduced alga
spread. Mar Environ Res 129:189–194
MacDougall AS, Turkington R (2005) Are invasive species the drivers of change or passengers
of change in degraded ecosystems? Ecology 86:42–55
Moles A, Norcross BL (1998) Effects of oil-laden sediments on growth and health of juvenile
flatfishes. Can J Fish Aquat Sci 55:605–610
Moles A, Rice S, Norcross BL (1994) Non-avoidance of hydrocarbon laden sediments by
juvenile flatfishes. Netherlands J Sea Res 32:361–367
Moles A, Stone RP (2002) Habitat preferences of juvenile tanner and red king crabs: substrate
and crude oil. In: Paul AJ, Dawe EG, Elner R, Jamieson GS, Kruse GH, Otto RS, Sainte-Marie

140
B, Shirley TC, Woodby D (eds) Crabs in Cold Water Regions: Biology, Management, and
Economics. University of Alaska Sea Grant, p 631–644
Olsen EM, Heupel MR, Simpfendorfer CA, Moland E (2012) Harvest selection on Atlantic cod
behavioral traits: implications for spatial management. Ecol Evol 2:1549–1562
Olsen EM, Moland E (2011) Fitness landscape of Atlantic cod shaped by harvest selection and
natural selection. Evol Ecol 25:695–710
Parry GD, Cohen BF (2001) Exotic species established in Western Port, including an assessment
of the status of the exotic species Corbula gibba, Alexandrium spp, Gymnodinium spp and
Undaria pinnatifida. Queenscliff
Parsons DM, Babcock RC, Hankin RKS, Willis TJ, Aitken JP, O’dor RK, Jackson GD (2003) Snapper
Pagrus auratus (Sparidae) home range dynamics: acoustic tagging studies in a marine
reserve. Mar Ecol Prog Ser 262:253–265
Patten MA, Kelly JF (2010) Habitat selection and the perceptual trap. Ecol Appl 20:2148–2156
Pert CG, Swearer SE, Dworjanyn SA, Turchini GM, Francis DS and Dempster T (2018) Barrens of
gold: gonad conditioning of an overabundant sea urchin. Aquac Environ Interact 10:345–
361
Peteiro C, Sánchez N, Martínez B (2016) Mariculture of the Asian kelp Undaria pinnatifida and
the native kelp Saccharina latissima along the Atlantic coast of southern Europe: an
overview. Algal Res 15:9–23
Reubens JT, Degraer S, Vincx M (2014) The ecology of benthopelagic fishes at offshore wind
farms: a synthesis of 4 years of research. Hydrobiologia 727:121–136
Reubens JT, Vandendriessche S, Zenner AN, Degraer S, Vincx M (2013) Offshore wind farms as
productive sites or ecological traps for gadoid fishes? Impact on growth, condition index
and diet composition. Mar Environ Res 90:66–74
Richardson DM, Pyšek P (2012) Naturalization of introduced plants: ecological drivers of
biogeographical patterns. New Phytol 196:383–396
Robertson BA, Rehage JS, Sih A (2013) Ecological novelty and the emergence of evolutionary
traps. Trends Ecol Evol 28:552–560
Russell LK, Hepburn CD, Hurd CL, Stuart MD (2008) The expanding range of Undaria pinnatifida
in southern New Zealand: distribution, dispersal mechanisms and the invasion of wave-
exposed environments. Biol Invasions 10:103–115
Sih A (2013) Understanding variation in behavioural responses to human-induced rapid
environmental change: a conceptual overview. Anim Behav 85:1077–1088
Sih A, Ferrari MCO, Harris DJ (2011) Evolution and behavioural responses to human-induced

141
rapid environmental change. Evol Appl 4:367–387
South PM, Floerl O, Forrest BM, Thomsen MS (2017) A review of three decades of research on
the invasive kelp Undaria pinnatifida in Australasia: an assessment of its success, impacts
and status as one of the world’s worst invaders. Mar Environ Res 131: 243–257
South PM, Thomsen MS (2016) The ecological role of invading Undaria pinnatifida: an
experimental test of the driver–passenger models. Mar Biol 163:175
Thomsen MS, Wernberg T, South PM, Schiel DR (2016) Non-native seaweeds drive changes in
marine coastal communities around the world. In: Hu Z-M, Fraser C (eds) Seaweed
Phylogeography: Adaptation and Evolution of Seaweeds under Environmental Change.
Springer Netherlands, Dordrecht, p 147–185
Valentine JP, Johnson CR (2003) Establishment of the introduced kelp Undaria pinnatifida in
Tasmania depends on disturbance to native algal assemblages. J Exp Mar Bio Ecol
295:63–90
Willis TJ, Parsons DM, Babcock RC (2001) Evidence for long‐term site fidelity of snapper
(Pagrus auratus) within a marine reserve. New Zeal J Mar Freshw Res 35:581–590
Wotton DM, O’Brien C, Stuart MD, Fergus DJ (2004) Eradication success down under: heat
treatment of a sunken trawler to kill the invasive seaweed Undaria pinnatifida. Mar
Pollut Bull 49:844–9
Wright JI, Gribben PE (2008) Predicting the impact of an invasive seaweed on the fitness of
native fauna. J Appl Ecol 45:1540–1549
Zavaleta ES, Hobbs RJ, Mooney HA (2001) Viewing invasive species removal in a whole-
ecosystem context. Trends Ecol Evol 16:454–459

142
APPENDICES
Appendix 2.1. List of 191 articles included in systematic review and meta-analysis.
Abaad, M., Tuset, V.M., Montero, D., Lombarte, A., Otero-Ferrer, J.L. and Haroun, R. (2016) Phenotypic
plasticity in wild marine fishes associated with fish-cage aquaculture. Hydrobiologia 765, 343–358.
Adámek, Z., Kortan, D., Lepič, P. and Andreji, J. (2003) Impacts of otter (Lutra lutra L.) predation on
fishponds: A study of fish remains at ponds in the Czech Republic. Aquaculture International 11,
389–396.
Anyango, J.O., Mlewa, C.M. and Mwaluma, J. (2017) Abundance, diversity and trophic status of wild fish
around seaweed farms in Kibuyuni, South Coast Kenya. International Journal of Fisheries and
Aquatic Studies 5, 440–446.
Arechavala-Lopez, P., Izquierdo-Gomez, D., Uglem, I. and Sanchez-Jerez, P. (2015a) Aggregations of
bluefish Pomatomus saltatrix (L.) at Mediterranean coastal fish farms: seasonal presence, daily
patterns and influence of farming activity. Environmental Biology of Fishes 98, 499–510.
Arechavala-Lopez, P., Sæther, B.-S., Marhuenda-Egea, F., Sanchez-Jerez, P. and Uglem, I. (2015b)
Assessing the influence of salmon farming through total lipids, fatty acids, and trace elements in
the liver and muscle of wild saithe Pollachius virens. Marine and Coastal Fisheries 7, 59–67.
Arechavala-Lopez, P., Sanchez-Jerez, P., Bayle-Sempere, J., Fernandez-Jover, D., Martinez-Rubio, L., Lopez-
Jimenez, J.A. and Martinez-Lopez, F.J. (2011) Direct interaction between wild fish aggregations at
fish farms and fisheries activity at fishing grounds: a case study with Boops boops. Aquaculture
Research 42, 996–1010.
Arechavala-Lopez, P., Uglem, I., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J.T. and Nilsen, R.
(2010) Movements of grey mullet Liza aurata and Chelon labrosus associated with coastal fish
farms in the western Mediterranean Sea. Aquaculture Environment Interactions 1, 127–136.
Avery, M.L., Eiselman, D.S., Young, M.K., Humphrey, J.S. and Decker, D.G. (1999) Wading bird predation
at tropical aquaculture facilities in central Florida. North American Journal of Aquaculture 61, 64–
69.
Bacher, K. and Gordoa, A. (2016) Does marine fish farming affect local small-scale fishery catches? A case
study in the NW Mediterranean Sea. Aquaculture Research 47, 2444–2454.
Bacher, K., Gordoa, A. and Sague, O. (2012) Spatial and temporal extension of wild fish aggregations at
Sparus aurata and Thunnus thynnus farms in the north-western Mediterranean. Aquaculture
Environment Interactions 2, 239–252.
Bacher, K., Gordoa, A. and Sagué, O. (2015) Feeding activity strongly affects the variability of wild fish
aggregations within fish farms: a sea bream farm as a case study. Aquaculture Research 46, 552–
564.
Bagdonas, K., Humborstad, O.-B. and Løkkeborg, S. (2012) Capture of wild saithe (Pollachius virens) and
cod (Gadus morhua) in the vicinity of salmon farms: three pot types compared. Fisheries Research
134–136, 1–5.
Ballester-Moltó, M., Sanchez-Jerez, P. and Aguado-Giménez, F. (2017) Consumption of particulate wastes
derived from cage fish farming by aggregated wild fish. An experimental approach. Marine
Environmental Research 130, 166–173.
Ballester-Moltó, M., Sanchez-Jerez, P., Garcia-Garcia, B. and Aguado-Giménez, F. (2015) Husbandry and
environmental conditions explain temporal variability of wild fish assemblages aggregated around
a Mediterranean fish farm. Aquaculture Environment Interactions 7, 193–203.
Baltrūnaitė, L. (2009) Diet of otters in fish farms in Lithuania. Acta Zoologica Lituanica 19, 182–187.
Barlow, C.G. and Bock, K. (1984) Predation of fish in farm dams by cormorants, Phalacrocorax spp.
Australian Wildlife Research 11, 559–566.
Bechard, M.J. and Márquez-Reyes, C. (2003) Mortality of wintering ospreys and other birds at aquaculture
facilities in Colombia. Journal of Raptor Research 37, 292–298.

143
Becker, B.H., Press, D.T. and Allen, S.G. (2011) Evidence for long-term spatial displacement of breeding
and pupping harbour seals by shellfish aquaculture over three decades. Aquatic Conservation-
Marine and Freshwater Ecosystems 21, 247–260.
Belant, J.L., Tyson, L.A. and Mastrangelo, P.A. (2000) Effects of lethal control at aquaculture facilities on
populations of piscivorous birds. Wildlife Society Bulletin 28, 379–384.
Bergman, K.C., Svensson, S. and Ohman, M.C. (2001) Influence of algal farming on fish assemblages.
Marine Pollution Bulletin 42, 1379–1389.
Beynon, J.L., Hutchins, D.L., Rubino, A.J., Lawrence, A.L. and Chapman, B.R. (1981) Nocturnal activity of
birds on shrimp mariculture ponds. Journal of the World Mariculture Society 12, 63–70.
Bjordal, Å. and Johnstone, A.D.F. (1993) Local movements of saithe (Pollachius virens L.) in the vicinity of
fish farm cages. In: ICES Mar Sci Symp, Vol. 196. pp 143–146.
Björklund, H., Bondestam, J. and Bylund, G. (1990) Residues of oxytetracycline in wild fish and sediments
from fish farms. Aquaculture 86, 359–367.
Bjørn, P.-A., Uglem, I., Kerwath, S., Sæther, B.-S. and Nilsen, R. (2009) Spatiotemporal distribution of
Atlantic cod (Gadus morhua L.) with intact and blocked olfactory sense during the spawning season
in a Norwegian fjord with intensive salmon farming. Aquaculture 286, 36–44.
Bjørn, P.A., Finstad, B. and Kristoffersen, R. (2001) Salmon lice infection of wild sea trout and Arctic char
in marine and freshwaters: the effects of salmon farms. Aquaculture Research 32, 947–962.
Blackwell, B.F., Dolbeer, R.A. and Tyson, L.A. (2000) Lethal control of piscivorous birds at aquaculture
facilities in the northeast United States: effects on populations. North American Journal of
Aquaculture 62, 300–307.
Bogdanović, T., Šimat, V., Frka-Roić, A. and Marković, K. (2012) Development and application of quality
index method scheme in a shelf-life study of wild and fish farm affected bogue (Boops boops, L.).
Journal of Food Science 77, S99–S106.
Bonizzoni, S., Furey, N.B., Pirotta, E., Valavanis, V.D., Wuersig, B. and Bearzi, G. (2014) Fish farming and its
appeal to common bottlenose dolphins: modelling habitat use in a Mediterranean embayment.
Aquatic Conservation-Marine and Freshwater Ecosystems 24, 696–711.
Boyra, A., Sanchez-Jerez, P., Tuya, F., Espino, F. and Haroun, R. (2004) Attraction of wild coastal fishes to
an Atlantic subtropical cage fish farms, Gran Canaria, Canary Islands. Environmental Biology of
Fishes 70, 393–401.
Brandt, M.J., Höschle, C., Diederichs, A., Betke, K., Matuschek, R., Witte, S. and Nehls, G. (2013) Far‐
reaching effects of a seal scarer on harbour porpoises, Phocoena phocoena. Aquatic Conservation:
Marine and Freshwater Ecosystems 23, 222–232.
Brehmer, P., Gerlotto, F., Guillard, J., Sanguinède, F., Guénnegan, Y. and Buestel, D. (2003) New
applications of hydroacoustic methods for monitoring shallow water aquatic ecosystems: the case
of mussel culture grounds. Aquatic Living Resources 16, 333–338.
Bridgman, G.K., Rave, E.H. and Rafferty, J.M. (2000) Piscivorous bird depredation at northern Minnesota
aquaculture facilities. Prairie Naturalist 32, 17–28.
Broyer, J. and Calenge, C. (2010) Influence of fish-farming management on duck breeding in French fish
pond systems. Hydrobiologia 637, 173–185.
Broyer, J., Chavas, G. and Chazal, R. (2017) The effects of cessation of fish farming on duck breeding in
French fishpond systems. Hydrobiologia 788, 47–53.
Bustnes, J.O., Lie, E., Herzke, D., Dempster, T., Bjørn, P.A., Nygård, T. and Uglem, I. (2010) Salmon farms
as a source of organohalogenated contaminants in wild fish. Environmental Science & Technology
44, 8736–43.
Bustnes, J.O., Nygard, T., Dempster, T., Ciesielski, T., Jenssen, B.M., Bjorn, P.A. and Uglem, I. (2011) Do
salmon farms increase the concentrations of mercury and other elements in wild fish? Journal of
Environmental Monitoring 13, 1687–1694.
Caldow, R., Beadman, H., McGrorty, S., Kaiser, M., Goss-Custard, J., Mould, K. and Wilson, A. (2003) Effects
of intertidal mussel cultivation on bird assemblages. Marine Ecology Progress Series 259, 173–183.
Carss, D.N. (1990) Concentrations of wild and escaped fishes immediately adjacent to fish farm cages.
Aquaculture 90, 29–40.

144
Carss, D.N. (1993a) Cormorants Phalacrocorax carbo at cage fish farms in Argyll, western Scotland. Seabird
15, 38–44.
Carss, D.N. (1993b) Grey heron, Ardea cinerea L., predation at cage fish farms in Argyll, western Scotland.
Aquaculture and Fisheries Management 24, 29–45.
Carss, D.N. (1994) Killing of piscivorous birds at Scottish fin fish farms, 1984-87. Biological Conservation
68, 181–188.
Cartier, L.E. and Carpenter, K.E. (2014) The influence of pearl oyster farming on reef fish abundance and
diversity in Ahe, French Polynesia. Marine Pollution Bulletin 78, 43–50.
de Carvalho, L.L., de Souza, E.G.A., da Mata Júnior, M.R. and Villaça, R.C. (2017) Assessment of rocky reef
fish assemblages close to seaweed farming. Aquaculture Research 48, 481–493.
Cervencl, A., Troost, K., Dijkman, E., de Jong, M., Smit, C.J., Leopold, M.F. and Ens, B.J. (2015) Distribution
of wintering common eider Somateria mollissima in the Dutch Wadden Sea in relation to available
food stocks. Marine Biology 162, 153–168.
Chesney, E.J. and Iglesias, J. (1979) Seasonal distribution, abundance and diversity of demersal fishes in
the inner Ria de Arosa, Northwest Spain. Estuarine and Coastal Marine Science 8, 227–239.
Clynick, B.G., McKindsey, C.W. and Archambault, P. (2008) Distribution and productivity of fish and
macroinvertebrates in mussel aquaculture sites in the Magdalen islands (Québec, Canada).
Colorni, A., Diamant, A., Eldar, A., Kvitt, H. and Zlotkin, A. (2002) Streptococcus iniae infections in Red Sea
cage-cultured and wild fishes. Diseases of Aquatic Organisms 49, 165–170.
Connolly, L.M. and Colwell, M.A. (2005) Comparative use of longline oysterbeds and adjacent tidal flats
by waterbirds. Bird Conservation International 15, 237–255.
D’Amours, O., Archambault, P., McKindsey, C.W. and Johnson, L.E. (2008) Local enhancement of
epibenthic macrofauna by aquaculture activities. Marine Ecology Progress Series 371, 73–84.
Dealteris, J.T., Kilpatrick, B.D. and Rheault, R.B. (2004) A comparative evaluation of the habitat value of
shellfish aquaculture gear, submerged aquatic vegetation and a non-vegetated seabed. Journal of
Shellfish Research 23, 867–874.
DeBruyn, A.M.H., Trudel, M., Eyding, N., et al. (2006) Ecosystemic effects of salmon farming increase
mercury contamination in wild fish. Environmental Science & Technology 40, 3489–3493.
Demétrio, J.A., Gomes, L.C., Latini, J.D. and Agostinho, A.A. (2012) Influence of net cage farming on the
diet of associated wild fish in a Neotropical reservoir. Aquaculture 330, 172–178.
Dempster, T., Fernandez-Jover, D., Sanchez-Jerez, P., Tuya, F., Bayle-Sempere, J., Boyra, A. and Haroun,
R.J. (2005) Vertical variability of wild fish assemblages around sea-cage fish farms: implications for
management. Marine Ecology Progress Series 304, 15–29.
Dempster, T., Sanchez-Jerez, P., Bayle-Sempere, J.T. and Kingsford, M. (2004) Extensive aggregations of
wild fish at coastal sea-cage fish farms. Hydrobiologia 525, 245–248.
Dempster, T., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J.T., Nilsen, R., Bjørn, P.-A. and
Uglem, I. (2011) Proxy measures of fitness suggest coastal fish farms can act as population sources
and not ecological traps for wild gadoid fish. PLoS ONE 6, e15646.
Dempster, T., Sanchez-Jerez, P., Uglem, I. and Bjørn, P.-A. (2010) Species-specific patterns of aggregation
of wild fish around fish farms. Estuarine, Coastal and Shelf Science 86, 271–275.
Dempster, T., Uglem, I., Sanchez-Jerez, P., Fernandez-Jover, D., Bayle-Sempere, J. and Nilsen, R. (2009)
Coastal salmon farms attract large and persistent aggregations of wild fish: an ecosystem effect.
Marine Ecology Progress Series 385, 1–14.
Diamant, A., Banet, A., Ucko, M., Colorni, A., Knibb, W. and Kvitt, H. (2000) Mycobacteriosis in wild
rabbitfish Siganus rivulatus associated with cage farming in the Gulf of Eilat, Red Sea. Diseases of
Aquatic Organisms 39, 211–219.
Diaz López, B. (2006) Bottlenose dolphin (Tursiops truncatus) predation on a marine fin fish farm: some
underwater observations. Aquatic Mammals 32, 305–310.
Diaz López, B. (2012) Bottlenose dolphins and aquaculture: interaction and site fidelity on the north-
eastern coast of Sardinia (Italy). Marine Biology 159, 2161–2172.
Diaz López, B. (2017) Temporal variability in predator presence around a fin fish farm in the Northwestern
Mediterranean Sea. Marine Ecology 38, e12378.

145
Diaz López, B. (2009) The bottlenose dolphin Tursiops truncatus foraging around a fish farm: effects of
prey abundance on dolphins’ behavior. Current Zoology 55, e11221–e11221.
Diaz López, B. and Bernal-Shirai, J.A. (2007) Bottlenose dolphin (Tursiops truncatus) presence and
incidental capture in a marine fish farm on the north-eastern coast of Sardinia (Italy). Journal of
the Marine Biological Association of the United Kingdom 87, 113–117.
Diaz López, B. and Bernal-Shirai, J.A. (2008) Marine aquaculture and bottlenose dolphins’ (Tursiops
truncatus) social structure. Behavioral Ecology and Sociobiology 62, 887–894.
Dorr, B.S., Burger, L.W., Barras, S.C. and Godwin, K.C. (2012) Economic impact of double-crested
cormorant, Phalacrocorax auritus, depredation on channel catfish, Ictalurus punctatus,
aquaculture in Mississippi, USA. Journal of the World Aquaculture Society 43, 502–513.
Draulans, D. and van Vessem, J. (1985) The effect of disturbance on nocturnal abundance and behaviour
of grey herons (Ardea cinerea) at a fish-farm in winter. Journal of applied ecology, 19–27.
Dunthorn, A.A. (1971) The predation of cultivated mussels by eiders. Bird Study 18, 107–11.
Eklöf, J.S., de la Torre-Castro, M., Nilsson, C. and Rönnbäck, P. (2006) How do seaweed farms influence
local fishery catches in a seagrass-dominated setting in Chwaka Bay, Zanzibar? Aquatic Living
Resources 19, 137–147.
Erbland, P.J. and Ozbay, G. (2008) A comparison of the macrofaunal communities inhabiting a Crassostrea
virginica oyster reef and oyster aquaculture gear in Indian River Bay, Delaware. Journal of Shellfish
Research 27, 757–768.
Ervik, A., Thorsen, B., Eriksen, V., Lunestad, B.T. and Samuelsen, O.B. (1994) Impact of administering
antibacterial agents on wild fish and blue mussels Mytilus edulis in the vicinity of fish farms.
Diseases of Aquatic Organisms 18, 45–51.
Fernandez-Jover, D., Faliex, E., Sanchez-Jerez, P., Sasal, P. and Bayle-Sempere, J.T. (2010) Coastal fish
farming does not affect the total parasite communities of wild fish in SW Mediterranean.
Aquaculture 300, 10–16.
Fernandez-Jover, D., Jimenez, J.A.L., Sanchez-Jerez, P., Bayle-Sempere, J.T., Giménez-Casalduero, F.,
Lopez, F.J.M. and Dempster, T. (2007a) Changes in body condition and fatty acid composition of
wild Mediterranean horse mackerel (Trachurus mediterraneus, Steindachner, 1868) associated to
sea cage fish farms. Marine Environmental Research 63, 1–18.
Fernandez-Jover, D., Martinez-Rubio, L., Sanchez-Jerez, P., et al. (2011) Waste feed from coastal fish
farms: a trophic subsidy with compositional side-effects for wild gadoids. Estuarine, Coastal and
Shelf Science 91, 559–568.
Fernandez-Jover, D. and Sanchez-Jerez, P. (2015) Comparison of diet and otolith growth of juvenile wild
fish communities at fish farms and natural habitats. ICES Journal of Marine Science 72, 916–929.
Fernandez-Jover, D., Sanchez-Jerez, P., Bayle-Sempere, J., Carratala, A. and Leon, V.M. (2007b) Addition
of dissolved nitrogen and dissolved organic carbon from wild fish faeces and food around
Mediterranean fish farms: Implications for waste-dispersal models. Journal of Experimental Marine
Biology and Ecology 340, 160–168.
Fernandez-Jover, D., Sanchez-Jerez, P., Bayle-Sempere, J.T., Arechavala-Lopez, P., Martinez-Rubio, L.,
Lopez Jimenez, J.A. and Martinez Lopez, F.J. (2009) Coastal fish farms are settlement sites for
juvenile fish. Marine environmental research 68, 89–96.
Fernandez-Jover, D., Sanchez-Jerez, P., Bayle-Sempere, J.T., Valle, C. and Dempster, T. (2008) Seasonal
patterns and diets of wild fish assemblages associated with Mediterranean coastal fish farms. ICES
Journal of Marine Science 65, 1153–1160.
Fleury, B.E. and Sherry, T.W. (1995) Long-term population trends of colonial wading birds in the southern
United States: The impact of crayfish aquaculture on Louisiana populations. Auk 112, 613–632.
Freitas, D., Gomes, J., Luis, T.S., et al. (2007) Otters and fish farms in the Sado estuary: ecological and
socio-economic basis of a conflict. Hydrobiologia 587, 51–62.
Gabrielsen, S.-E. (1999) Effects of fish-farm activity on the limnetic community structure of brown trout,
Salmo trutta, and Arctic charr, Salvelinus alpinus. Environmental biology of fishes 55, 321–332.
Gaitán‐Espitia, J.D., Gómez, D., Hobday, A.J., Daley, R., Lamilla, J. and Cárdenas, L. (2017) Spatial overlap
of shark nursery areas and the salmon farming industry influences the trophic ecology of Squalus
acanthias on the southern coast of Chile. Ecology and evolution 7, 3773–3783.

146
Giannoulaki, M., Machias, A., Somarakis, S. and Karakassis, I. (2005) Wild fish spatial structure in response
to presence of fish farms. Journal of the Marine Biological Association of the United Kingdom 85,
1271–1277.
Glahn, J.F., Rasmussen, E.S., Tomsa, T. and Preusser, K.J. (1999a) Distribution and relative impact of avian
predators at aquaculture facilities in the northeastern United States. North American Journal of
Aquaculture 61, 340–348.
Glahn, J.F., Tomsa, T. and Preusser, K.J. (1999b) Impact of great blue heron predation at trout-rearing
facilities in the northeastern United States. North American Journal of Aquaculture 61, 349–354.
Glover, K.A., Sørvik, A.G.E., Karlsbakk, E., Zhang, Z. and Skaala, Ø. (2013) Molecular genetic analysis of
stomach contents reveals wild Atlantic cod feeding on piscine reovirus (PRV) infected Atlantic
salmon originating from a commercial fish farm. PLoS ONE 8, e60924.
Godet, L., Toupoint, N., Fournier, J., Le Mao, P., Retiere, C. and Olivier, F. (2009) Clam farmers and
oystercatchers: effects of the degradation of Lanice conchilega beds by shellfish farming on the
spatial distribution of shorebirds. Marine Pollution Bulletin 58, 589–595.
Goodbrand, L., Abrahams, M. V and Rose, G.A. (2013) Sea cage aquaculture affects distribution of wild
fish at large spatial scales. Canadian Journal of Fisheries and Aquatic Sciences 70, 1289–1295.
Gregory, P.T. and Nelson, K.J. (1991) Predation on fish and intersite variation in the diet of common garter
snakes, Thamnophis sirtalis, on Vancouver Island. Canadian Journal of Zoology 69, 988–994.
Güçlüsoy, H. and Savas, Y. (2003) Interaction between monk seals Monachus monachus (Hermann, 1779)
and marine fish farms in the Turkish Aegean and management of the problem. Aquaculture
Research 34, 777–783.
Hayward, C.J., Svane, I., Lachimpadi, S.K., Itoh, N., Bott, N.J. and Nowak, B.F. (2011) Sea lice infections of
wild fishes near ranched southern bluefin tuna (Thunnus maccoyii) in South Australia. Aquaculture
320, 178–182.
Hehre, E.J. and Meeuwig, J.J. (2016) A global analysis of the relationship between farmed seaweed
production and herbivorous fish catch. PLoS ONE 11, e0148250.
Hume, F., Pemberton, D., Gales, R., Brothers, N. and Greenwood, M. (2002) Trapping and relocating seals
from salmonid fish farms in Tasmania, 1990-2000: was it a success? Papers and Proceedings of the
Royal Society of Tasmania 136, 1–6.
Iglesias, J. (1981) Spatial and temporal changes in the demersal fish community of the Ria de Arosa (NW
Spain). Marine Biology 65, 199–208.
Hilgerloh, G., Halloran, J.O., Kelly, T.C. and Burnell, G.M. (2001) A preliminary study on the effects of oyster
culturing structures on birds in a sheltered Irish estuary. Hydrobiologia 465, 175–180.
Jacobs, S.R. and Terhune, J.M. (2000) Harbor seal (Phoca vitulina) numbers along the New Brunswick coast
of the Bay of Fundy in autumn in relation to aquaculture. Northeastern Naturalist 7, 289–296.
Jimenez, J.E., Arriagada, A.M., Fonturbel, F.E., Camus, P.A. and Avila-Thieme, M.I. (2013) Effects of exotic
fish farms on bird communities in lake and marine ecosystems. Naturwissenschaften 100, 779–
787.
Johansson, T., Hakanson, L., Borum, K. and Persson, J. (1998) Direct flows of phosphorus and suspended
matter from a fish farm to wild fish in lake southern Bullaren, Sweden. Aquacultural Engineering
17, 111–137.
Johnston, T.A., Keir, M. and Power, M. (2010) Response of native and naturalized fish to salmonid cage
culture farms in northern Lake Huron, Canada. Transactions of the American Fisheries Society 139,
660–670.
Kelly, J.P., Evens, J.G. and Stallcup, R.W. (1996) Effects of aquaculture on habitat use by wintering
shorebirds in Tomales Bay, California. California Fish and Game 82, 160–174.
Kemper, C.M. and Gibbs, S.E. (2001) Dolphin interactions with tuna feedlots at Port Lincoln, South
Australia and recommendations for minimising entanglements. Journal of Cetacean Research and
Management 3, 283–292.
Kilambi, R. V., Adams, J.C. and Wickizer, W.A. (1978) Effects of cage culture on growth, abundance, and
survival of resident largemouth bass (Micropterus salmoides). Journal of the Fisheries Research
Board of Canada 35, 157–160.

147
Kloskowski, J. (2010) Fish farms as amphibian habitats: factors affecting amphibian species richness and
community structure at carp ponds in Poland. Environmental Conservation 37, 187–194.
Kloskowski, J. (2012) Fish stocking creates an ecological trap for an avian predator via effects on prey
availability. Oikos 121, 1567–1576.
Kloskowski, J. (2005) Otter Lutra lutra damage at farmed fisheries in southeastern Poland, II: exploitation
of common carp Cyprinus carpio. Wildlife Biology 11, 257–261.
Kloskowski, J. (2009) Size-structured effects of common carp on reproduction of pond-breeding
amphibians. Hydrobiologia 635, 205–213.
Kloskowski, J., Grendel, A. and Wronka, M. (2000) The use of fish bones of three farm fish species in diet
analysis of the Eurasian otter, Lutra lutra. Folia Zoologica 49, 183–190.
Kloskowski, J., Kaczanowska, E., Krogulec, J. and Grela, P. (2017) Hematological indicators of habitat
quality: erythrocyte parameters reflect greater parental effort of red-necked grebes under
ecological trap conditions. The Condor 119, 239–250.
Kloskowski, J., Nieoczym, M., Polak, M. and Pitucha, P. (2010) Habitat selection by breeding waterbirds at
ponds with size-structured fish populations. Naturwissenschaften 97, 673–682.
Kortan, D., Adámek, Z. and Poláková, S. (2007) Winter predation by otter, Lutra lutra on carp pond systems
in South Bohemia (Czech Republic). Folia Zoologica 56, 416.
Krkosek, M., Lewis, M.A., Morton, A., Frazer, L.N. and Volpe, J.P. (2006) Epizootics of wild fish induced by
farm fish. Proceedings of the National Academy of Sciences of the United States of America 103,
15506–15510.
Krkosek, M., Lewis, M.A. and Volpe, J.P. (2005) Transmission dynamics of parasitic sea lice from farm to
wild salmon. Proceedings. Biological sciences 272, 689–96.
Laffargue, P., Begout, M. and Lagardere, F. (2006) Testing the potential effects of shellfish farming on
swimming activity and spatial distribution of sole (Solea solea) in a mesocosm. ICES Journal of
Marine Science 63, 1014–1028.
Lanszki, J., Pallos, Z.S., Nagy, D. and Yoxon, G. (2007) Diet and fish choice of Eurasian otters (Lutra lutra L.)
in fish wintering ponds in Hungary. Aquaculture international 15, 393–402.
Lin, H.-J., Shao, K.-T., Hsieh, H.-L., Lo, W.-T. and Dai, X.-X. (2009) The effects of system-scale removal of
oyster-culture racks from Tapong Bay, southwestern Taiwan: model exploration and comparison
with field observations. ICES Journal of Marine Science 66, 797–810.
Loiseau, N., Kiszka, J.J., Bouveroux, T., Heithaus, M.R., Soria, M. and Chabanet, P. (2016) Using an unbaited
stationary video system to investigate the behaviour and interactions of bull sharks Carcharhinus
leucas under an aquaculture farm. African Journal of Marine Science 38, 73–79.
Ludwig, G.X., Hokka, V., Sulkava, R. and Ylönen, H. (2002) Otter Lutra lutra predation on farmed and free-
living salmonids in. Wildlife Biology 8.
Machias, A., Karakassis, I., Giannoulaki, M., Papadopoulou, K.N., Smith, C.J. and Somarakis, S. (2005)
Response of demersal fish communities to the presence of fish farms. Marine Ecology Progress
Series 288, 241–250.
Machias, A., Karakassis, I., Labropoulou, M., Somarakis, S., Papadopoulou, K.N. and Papaconstantinou, C.
(2004) Changes in wild fish assemblages after the establishment of a fish farming zone in an
oligotrophic marine ecosystem. Estuarine Coastal and Shelf Science 60, 771–779.
MacKenzie, K., Longshaw, M., Begg, G.S. and McVicar, A.H. (1998) Sea lice (Copepoda: Caligidae) on wild
sea trout (Salmo trutta L.) in Scotland. ICES Journal of Marine Science 55, 151–162.
Manikowska-Ślepowrońska, B., Szydzik, B. and Jakubas, D. (2016) Determinants of the presence of conflict
bird and mammal species at pond fisheries in western Poland. Aquatic Ecology 50, 87–95.
Marenghi, F.P. and Ozbay, G. (2010) Floating oyster, Crassostrea virginica Gmelin 1791, aquaculture as
habitat for fishes and macroinvertebrates in Delaware Inland Bays: the comparative value of oyster
clusters and loose shell. Journal of Shellfish Research 29, 889–904.
Markowitz, T.M., Harlin, A.D., Wursig, B. and Mcfadden, C.J. (2004) Dusky dolphin foraging habitat:
overlap with aquaculture in New Zealand. Aquatic Conservation-marine and Freshwater
Ecosystems 14, 133–149.
Marques, C., Rosalino, L.M. and Santos-Reis, M. (2007) Otter predation in a trout fish farm of central-east
Portugal: preference for “fast-food”? River Research and Applications 23, 1147–1153.

148
McAllister, P.E. and Owens, W.J. (1992) Recovery of infectious pancreatic necrosis virus from the faeces
of wild piscivorous birds. Aquaculture 106, 227–232.
McConnell, A., Routledge, R. and Connors, B. (2010) Effect of artificial light on marine invertebrate and
fish abundance in an area of salmon farming. Marine Ecology Progress Series 419, 147–156.
McPeek, K.C., McDonald, P.S. and VanBlaricom, G.R. (2015) Aquaculture disturbance impacts the diet but
not ecological linkages of a ubiquitous predatory fish. Estuaries and Coasts 38, 1520–1534.
Middlemas, S.J., Fryer, R.J., Tulett, D. and Armstrong, J.D. (2013) Relationship between sea lice levels on
sea trout and fish farm activity in western Scotland. Fisheries Management and Ecology 20, 68–74.
Mladineo, I., Segvic, T. and Grubisic, L. (2009) Molecular evidence for the lack of transmission of the
monogenean Sparicotyle chrysophrii (Monogenea, Polyopisthocotylea) and isopod Ceratothoa
oestroides (Crustacea, Cymothoidae) between wild bogue (Boops boops) and cage-reared sea
bream (Sparus aurata) . Aquaculture 295, 160–167.
Morrisey, D.J., Cole, R.G., Davey, N.K., Handley, S.J., Bradley, A., Brown, S.N. and Madarasz, A.L. (2006)
Abundance and diversity of fish on mussel farms in New Zealand. Aquaculture 252, 277–288.
Morrison, S.S. and Vogel, P. (2009) Aquaculture ponds, a Jamaican study: The impact of birds on fish
production. African Journal of Agricultural Research 4, 1447–1454.
Morton, A., Routledge, R. and Krkosek, M. (2008) Sea louse infestation in wild juvenile salmon and Pacific
herring associated with fish farms off the east-central coast of Vancouver Island, British Columbia.
North American Journal of Fisheries Management 28, 523–532.
Morton, A., Routledge, R., McConnell, A. and Krkosek, M. (2011) Sea lice dispersion and salmon survival
in relation to salmon farm activity in the Broughton Archipelago. ICES Journal of Marine Science
68, 144–156.
Morton, A., Routledge, R., Peet, C. and Ladwig, A. (2004) Sea lice (Lepeophtheirus salmonis) infection rates
on juvenile pink (Oncorhynchus gorbuscha) and chum (Oncorhynchus keta) salmon in the
nearshore marine environment of British Columbia, Canada. Canadian Journal of Fisheries and
Aquatic Sciences 61, 147–157.
Morton, A., Routledge, R.D. and Williams, R. (2005) Temporal patterns of sea louse infestation on wild
Pacific salmon in relation to the fallowing of Atlantic salmon farms. North American Journal of
Fisheries Management 25, 811–821.
Mott, D.E. and Flynt, R.D. (1995) Evaluation of an electric fence system for excluding wading birds at
catfish ponds. The Progressive Fish-Culturist 57, 88–90.
Nelson, K.J. and Gregory, P.T. (2000) Activity patterns of garter snakes, Thamnophis sirtalis, in relation to
weather conditions at a fish hatchery on Vancouver Island, British Columbia. Journal of
Herpetology 34, 32.
Neofitou, N. (2016) Waste feed from fish farms of the eastern Mediterranean and attraction of wild fish.
Universal Journal of Geoscience 4, 112–115.
Oakes, C.T. and Pondella, D.J. (2009) The value of a net‐cage as a fish aggregating device in southern
California. Journal of the World Aquaculture Society 40, 1–21.
Otterå, H. and Skilbrei, O.T. (2014) Possible influence of salmon farming on long-term resident behaviour
of wild saithe (Pollachius virens L.). ICES Journal of Marine Science 71, 2484–2493.
Özgül, A. and Angel, D. (2013) Wild fish aggregations around fish farms in the Gulf of Aqaba, Red Sea:
implications for fisheries management and conservation. Aquaculture Environment Interactions 4,
135–145.
Parkhurst, J.A., Brooks, R.P. and Arnold, D.E. (1992) Assessment of predation at trout hatcheries in central
Pennsylvania. Wildlife Society Bulletin (1973-2006) 20, 411–419.
Pemberton, D., Brothers, N. and Copson, G. (1991) Predators on marine fish farms in Tasmania. Papers
and Proceedings of the Royal Society of Tasmania 125, 33–35.
Pemberton, D. and Shaughnessy, P.D. (1993) Interaction between seals and marine fish-farms in
Tasmania, and management of the problem. Aquatic Conservation-marine and Freshwater
Ecosystems 3, 149–158.
Pitt, W.C. and Conover, M.R. (1996) Predation at intermountain west fish hatcheries. The Journal of
Wildlife Management 60, 616–624.

149
Plummer, M. V. and Goy, J.M. (1984) Ontogenetic dietary shift of water snakes (Nerodia rhombifera) in a
fish hatchery. Copeia 1984, 550.
Powers, M., Peterson, C., Summerson, H. and Powers, S. (2007) Macroalgal growth on bivalve aquaculture
netting enhances nursery habitat for mobile invertebrates and juvenile fishes. Marine Ecology
Progress Series 339, 109–122.
Quick, N.J., Middlemas, S.J. and Armstrong, J.D. (2004) A survey of antipredator controls at marine salmon
farms in Scotland. Aquaculture 230, 169–180.
Radomski, A.A. and Zimba, P. V (2010) Does pond water reflectance influence double‐crested cormorant
selection of aquaculture ponds? Journal of the World Aquaculture Society 41, 430–437.
Ramos, I.P., Brandão, H., Zanatta, A.S., Zica, E. de O.P., da Silva, R.J., de Rezende-Ayroza, D.M.M. and
Carvalho, E.D. (2013) Interference of cage fish farm on diet, condition factor and numeric
abundance on wild fish in a Neotropical reservoir. Aquaculture 414, 56–62.
Ribeiro, S., Viddi, F.A., Cordeiro, J.K. and Freitas, T.R.O. (2007) Fine-scale habitat selection of Chilean
dolphins (Cephalorhynchus eutropia): interactions with aquaculture activities in southern Chiloe
Island, Chile. Journal of the Marine Biological Association of the United Kingdom 87, 119–128.
Riera, R., Tuset, V.M., Rodríguez, M., Monterroso, Ó. and Lombarte, A. (2017) Analyzing functional
diversity to determine the effects of fish cages in insular coastal wild fish assemblages. Aquaculture
479, 384–395.
Robinson, S., Gales, R., Terauds, A. and Greenwood, M. (2008a) Movements of fur seals following
relocation from fish farms. Aquatic Conservation-marine and Freshwater Ecosystems 18, 1189–
1199.
Robinson, S., Terauds, A., Gales, R. and Greenwood, M. (2008b) Mitigating fur seal interactions: relocation
from Tasmanian aquaculture farms. Aquatic Conservation-marine and Freshwater Ecosystems 18,
1180–1188.
Ross, B., Lien, J. and Furness, R.W. (2001) Use of underwater playback to reduce the impact of eiders on
mussel farms. ICES Journal of Marine Science 58, 517–524.
Roycroft, D., Kelly, T.C. and Lewis, L.J. (2004) Birds, seals and the suspension culture of mussels in Bantry
Bay, a non-seaduck area in Southwest Ireland. Estuarine, Coastal and Shelf Science 61, 703–712.
Roycroft, D., Kelly, T.C. and Lewis, L.J. (2007) Behavioural interactions of seabirds with suspended mussel
longlines. Aquaculture International 15, 25–36.
Sales-Luis, T., Freitas, D. and Santos-Reis, M. (2009) Key landscape factors for Eurasian otter Lutra lutra
visiting rates and fish loss in estuarine fish farms. European Journal of Wildlife Research 55, 345–
355.
Samuelsen, O.B., Lunestad, B.T., Husevag, B., Holleland, T. and Ervik, A. (1992) Residues of oxolinic acid in
wild fauna following medication in fish farms. Diseases of Aquatic Organisms 12, 111–119.
Sandilyan, S. (2017) A preliminary assessment on the role of abandoned shrimp farms on supporting
waterbirds in Pichavaram mangrove, Tamilnadu, Southern India. Journal of Coastal Conservation
21, 255–263.
Sanz-Lázaro, C., Belando, M.D., Navarrete-Mier, F. and Marín, A. (2011) Effects of wild fish and motile
epibenthic invertebrates on the benthos below an open water fish farm. Estuarine, Coastal and
Shelf Science 91, 216–223.
Schramm JR, H.L., French, B. and Ednoff, M. (1984) Depredation of channel catfish by Florida double-
crested cormorants. The Progressive Fish-Culturist 46, 41–43.
Segvic-Bubic, T., Grubisic, L., Karaman, N., Ticina, V., Jelavic, K.M. and Katavic, I. (2011) Damages on mussel
farms potentially caused by fish predation - self service on the ropes? Aquaculture 319, 497–504.
Segvić Bubić, T., Grubišić, L., Tičina, V. and Katavić, I. (2011) Temporal and spatial variability of pelagic wild
fish assemblages around Atlantic bluefin tuna Thunnus thynnus farms in the eastern Adriatic Sea.
Journal of Fish Biology 78, 78–97.
Sepúlveda, F., Marin, S.L. and Carvajal, J. (2004) Metazoan parasites in wild fish and farmed salmon from
aquaculture sites in southern Chile. Aquaculture 235, 89–100.
Sepúlveda, M., Newsome, S.D., Pavez, G., Oliva, D., Costa, D.P. and Hueckstaedt, L.A. (2015) Using satellite
tracking and isotopic information to characterize the impact of South American sea lions on
salmonid aquaculture in southern Chile. PLoS ONE 10, e0134926–e0134926.

150
Sepúlveda, M. and Oliva, D. (2005) Interactions between South American sea lions Otaria flavescens
(Shaw) and salmon farms in southern Chile. Aquaculture Research 36, 1062–1068.
Sepúlveda, M., Pavez, G., Santos‐Carvallo, M., Balbontín, C., Pequeño, G. and Newsome, S.D. (2017)
Spatial, temporal, age, and sex related variation in the diet of South American sea lions in southern
Chile. Marine Mammal Science 33, 480–495.
Skilbrei, O.T. and Otterå, H. (2016) Vertical distribution of saithe (Pollachius virens) aggregating around
fish farms. ICES Journal of Marine Science: Journal du Conseil 73, 1186–1195.
Skog, T.-E., Hylland, K., Torstensen, B.E. and Berntssen, M.H.G. (2003) Salmon farming affects the fatty
acid composition and taste of wild saithe Pollachius virens L. Aquaculture Research 34, 999–1007.
Snow, M., Black, J., Matejusova, I., McIntosh, R., Baretto, E., Wallace, I.S. and Bruno, D.W. (2010)
Detection of salmonid alphavirus RNA in wild marine fish: implications for the origins of salmon
pancreas disease in aquaculture. Diseases of Aquatic Organisms 91, 177–188.
Stickley, A.R., Glahn, J.F., King, J.O. and King, D.T. (1995) Impact of great blue heron depredations on
channel catfish farms. Journal of the World Aquaculture Society 26, 194–199.
Stickley, A.R., Warrick, G.L. and Glahn, J.F. (1992) Impact of double‐crested cormorant depredations on
channel catfish farms. Journal of the World Aquaculture Society 23, 192–198.
Strictar-Pereira, L., Agostinho, A.A. and Gomes, L.C. (2010) Cage culture with tilapia induces alteration in
the diet of natural fish populations: the case of Auchenipterus osteomystax. Brazilian Journal of
Biology 70, 1021–1030.
Sudirman, H.H., Jompa, J., Zulfikar, Iswahyudin and McKinnon, A.D. (2009) Wild fish associated with
tropical sea cage aquaculture in South Sulawesi, Indonesia. Aquaculture 286, 233–239.
Sun, Y.H., Wu, H.J. and Wang, Y. (2004) Tawny fish-owl predation at fish farms in Taiwan. Journal of Raptor
Research 38, 326–333.
Tallman, J.C. and Forrester, G.E. (2007) Oyster grow-out cages function as artificial reefs for temperate
fishes. Transactions of the American Fisheries Society 136, 790–799.
Tsuyuki, A. and Umino, T. (2017) Spatial movement of black sea bream Acanthopagrus schlegelii around
the oyster farming area in Hiroshima Bay, Japan. Fisheries Science 83, 235–244.
Tuckett, Q.M., Ritch, J.L., Lawson, K.M. and Hill, J.E. (2017) Landscape-scale survey of non-native fishes
near ornamental aquaculture facilities in Florida, USA. Biological Invasions 19, 223–237.
Tully, O., Gargan, P., Poole, W.R. and Whelan, K.F. (1999) Spatial and temporal variation in the infestation
of sea trout (Salmo trutta L.) by the caligid copepod Lepeophtheirus salmonis (Krøyer) in relation
to sources of infection in Ireland. Parasitology 119, 41–51.
Tuya, F., Sanchez-Jerez, P., Dempster, T., Boyra, A. and Haroun, R.J. (2006) Changes in demersal wild fish
aggregations beneath a sea-cage fish farm after the cessation of farming. Journal of Fish Biology
69, 682–697.
Uglem, I., Dempster, T., Bjørn, P.-A., Sanchez-Jerez, P. and Økland, F. (2009) High connectivity of salmon
farms revealed by aggregation, residence and repeated movements of wild fish among farms.
Marine Ecology Progress Series 384, 251–260.
Ulenaers, P. and Dhondt, A.A. (1994) Great crested grebe Podiceps cristatus chick mortality in relation to
parental fishing. Bird Study 41, 211–220.
Ulenaers, P. and van Vessem, J. (1994) Impact of great crested grebes (Podiceps cristatus L.) on fish ponds.
Hydrobiologia 279, 353–366.
Valle, C., Bayle-Sempere, J.T., Dempster, T., Sanchez-Jerez, P. and Gimenez-Casalduero, F. (2007)
Temporal variability of wild fish assemblages associated with a sea-cage fish farm in the south-
western Mediterranean Sea. Estuarine Coastal and Shelf Science 72, 299–307.
Valtonen, E.T. and Koskivaara, M. (1994) Relationships between the parasites of some wild and cultured
fishes in two lakes and a fish farm in central Finland. International Journal for Parasitology 24, 109–
118.
Vita, R., Marin, A., Madrid, J.A., Jimenez-Brinquis, B., Cesar, A. and Marin-Guirao, L. (2004) Effects of wild
fishes on waste exportation from a Mediterranean fish farm. Marine Ecology Progress Series 277,
253–261.

151
Wallace, I.S., Donald, K., Munro, L.A., et al. (2015) A survey of wild marine fish identifies a potential origin
of an outbreak of viral haemorrhagic septicaemia in wrasse, Labridae, used as cleaner fish on
marine Atlantic salmon, Salmo salar L., farms. Journal of Fish Diseases 38, 515–521.
Wallace, I.S., Gregory, A., Murray, A.G., Munro, E.S. and Raynard, R.S. (2008) Distribution of infectious
pancreatic necrosis virus (IPNV) in wild marine fish from Scottish waters with respect to clinically
infected aquaculture sites producing Atlantic salmon, Salmo salar L. Journal of Fish Diseases 31,
177–186.
Watson-Capps, J.J. and Mann, J. (2005) The effects of aquaculture on bottlenose dolphin (Tursiops sp.)
ranging in Shark Bay, Western Australia. Biological Conservation 124, 519–526.
Wisniowska, L. (2007) Otter (Lutra lutra L.) damage in commercial carp ponds of southern Poland. Hystrix,
the Italian Journal of Mammalogy 17.
Yasue, M. and Dearden, P. (2009) The importance of supratidal habitats for wintering shorebirds and the
potential impacts of shrimp aquaculture. Environmental Management 43, 1108–1121.
Zlotkin, A., Hershko, H. and Eldar, A. (1998) Possible transmission of Streptococcus iniae from wild fish to
cultured marine fish. Applied and Environmental Microbiology 64, 4065–4067.
Zydelis, R., Esler, D., Kirk, M. and Sean Boyd, W. (2009) Effects of off-bottom shellfish aquaculture on
winter habitat use by molluscivorous sea ducks. Aquatic Conservation: Marine and Freshwater
Ecosystems 19, 34–42.

152
Appendix 2.2. Summary of results for zero-inflated Poisson model comparing research effort on
interactions between wildlife (number of articles included in our systematic review) and
aquaculture between nations according to both domestic aquaculture production (t) in 2015
and Human Development Index (HDI) in 2015. N = 188 nations.
Model term Estimate SE z p
HDI 9.4 2.3 4.1 <0.0001 Production -0.01 0.006 -2.2 0.02 HDI × Production 0.02 0.01 2.2 0.03 HDI > 0.8, N = 55 Production 0.008 0.002 5.6 <0.0001

153
Appendix 2.3. List of the six most parsimonious linear models predicting log response ratios for
wildlife abundance (a) and species richness (b) at aquaculture sites relative to reference sites. In
each case, the full model was specified using the following terms: Response ~ Year + Country +
Region + Environment + Culture System + Culture Organism + Wild Taxa + Reference Habitat. A
model selection process using the MuMIn package for R (Barton 2016) fitted every possible
combination of these terms using maximum likelihood estimation and ranked them by AICC
score. ‘+’ indicates the corresponding term was included in the given model.
Model ID
Cultured Taxa
Env Ref Hab Wild Taxa
Year logLik AICC ΔAICC Wt
27 + + + -115.6 247.2 0 0.33 91 + + + + -111.7 247.6 0.3 0.28
155 + + + + -115.3 249.2 2.0 0.12 75 + + + -113.9 249.2 2.0 0.12 83 + + + -114.3 250.0 2.8 0.08
219 + + + + + -111.6 250.3 3.1 0.07
Model ID
Cultured Taxa
Env Ref Hab Wild Taxa
Year logLik AICC ΔAICC Wt
81 + + -24.8 64.4 0 0.34 65 + -26.5 65.0 0.6 0.26
209 + + + -24.1 66.0 1.7 0.15 89 + + + -24.4 66.7 2.4 0.11 73 + + -26.2 67.2 2.9 0.08
193 + + -26.5 67.8 3.4 0.06

154
Appendix 2.4. ANOVA table and Tukey’s post-hoc test results (multiple comparisons of means
with 95% family-wise confidence level) for best fitting linear model for factors predicting log
response ratios (RR) for wildlife abundance at aquaculture sites. The best fitting model was
selected from a full model containing the following factors: Year, Country, Continent,
Environment (Marine, Freshwater), Culture System (Cage, Pond, Longline, Rack, Bed), Cultured
Taxa (Fish, Shellfish, Crustacean, Alga), Wild Taxa (Fish, Bird, Mammal, Reptile, Amphibian), and
Reference Habitat (Structured, Unstructured). Post-hoc testing compared all pairwise
combinations of levels of significant model terms (Cultured Taxa and Reference Habitat).
Positive values for ‘Difference’ indicate that the former level has a higher response to farms than
the latter level in the pairwise comparison.
ANOVA Model term df SS MS F p
Environment 1 6.87 6.87 2.7 0.11 Cultured Taxa 3 62.0 20.7 8.1 0.0001 Reference Habitat 1 24.9 24.9 9.8 0.003 Residuals 57 144 2.54 POST HOC Significant model term Difference Lower Upper Adjusted p
Cultured Taxa Crustacean-Alga -2.33 -6.95 2.29 0.54 Fish-Alga 1.77 -0.24 3.78 0.10 Shellfish-Alga 0.09 -2.01 2.19 0.99 Fish-Crustacean 4.11 -0.17 8.38 0.06 Shellfish-Crustacean 2.42 -1.89 6.74 0.45 Shellfish-Fish -1.68 -2.84 -0.52 0.002 Reference Habitat Unstructured-Structured 1.17 0.37 1.97 0.005

155
Appendix 2.5. ANOVA table and Tukey’s post-hoc test results for best fitting linear model for
factors predicting log response ratios (RR) for wildlife species richness at aquaculture sites. The
best fitting model was selected from a full model containing the following factors: Year, Country,
Continent, Environment (Marine, Freshwater), Culture System (Cage, Pond, Longline, Rack, Bed),
Cultured Taxa (Fish, Shellfish, Crustacean, Alga), Wild Taxa (Fish, Bird, Mammal, Reptile,
Amphibian), and Reference Habitat (Structured, Unstructured). Post-hoc testing compared all
pairwise combinations of wild taxonomic groups represented in the dataset. Positive values for
‘Difference’ indicate that the former level has a higher response to farms than the latter level in
the pairwise comparison.
ANOVA Model term df SS MS F p
Wild Taxa 3 3.63 1.21 4.4 0.01 Reference Habitat 1 0.81 0.81 3.1 0.09 Residuals 32 8.27 0.26 POST HOC Significant model term Difference Lower Upper Adjusted p
Wild Taxa Bird-Amphibian 1.22 -0.25 2.70 0.13 Fish-Amphibian 1.53 0.13 2.93 0.03 Mammal-Amphibian 0.41 -1.54 2.35 0.94 Fish-Bird 0.30 -0.28 0.89 0.50 Mammal-Bird -0.82 -2.29 0.65 0.44 Mammal-Fish -1.12 -2.53 0.28 0.15

156
Appendix 3.1. Ovarian fatty acid profiles in cod (Gadus morhua) collected from areas of high and
low salmon farming density. Fatty acids that represent less than 0.1 % of the total fatty acids are
omitted. Data are presented as percentage of total fatty acids (mean ± SD, n = 10 per group).
Statistical comparisons are from univariate linear analyses of variance with 1 on 18 df.
Fatty acids Low farm density High farm density F p
14:0 1.63 ± 0.36 1.53 ± 0.39 Iso 15:0 0.25 ± 0.12 0.20 ± 0.04 Iso 16:0 0.17 ± 0.06 0.15 ± 0.04 Iso 17:0 0.46 ± 0.09 0.46 ± 0.07
Antiso 17:0 0.23 ± 0.09 0.21 ± 0.05 17:0 0.37 ± 0.06 0.41 ± 0.08
iso 18:0 0.18 ± 0.05 0.19 ± 0.04 18:0 2.44 ± 0.40 2.55 ± 0.54
∑SFA 23.87 ± 0.71 24.00 ± 1.05 0.10 0.76
16:1 (n-11) 0.14 ± 0.04 0.13 ± 0.05 16:1 (n-9) 1.27 ± 0.13 1.30 ± 0.28 18:1 (n-9) 12.77 ± 0.71 13.57 ± 1.67 1.9 0.18 18:1 (n-7) 4.25 ± 0.89 3.98 ± 0.47 18:1 (n-5) 0.28 ± 0.04 0.31 ± 0.07
20:1 (n-11) 0.33 ± 0.23 0.20 ± 0.08 20:1 (n-9) 1.02 ± 0.56 0.61 ± 0.23
22:1 (n-11) 0.38 ± 0.32 0.21 ± 0.10 22:1 (n-9) 0.08 ± 0.04 0.07 ± 0.01 24:1 (n-9) 0.94 ± 0.20 1.02 ± 0.34 24:1 (n-7) 0.24 ± 0.05 0.32 ± 0.12
∑MUFA 26.16 ± 3.10 26.12 ± 1.51 0.00 0.97
18:2 (n-6) 1.06 ± 0.53 1.26 ± 0.89 0.34 0.57 18:3 (n-6) 0.18 ± 0.05 0.17 ± 0.04 20:2 (n-6) 0.32 ± 0.15 0.25 ± 0.07 20:3 (n-6) 0.10 ± 0.01 0.09 ± 0.03 20:4 (n-6) 3.90 ± 0.69 4.11 ± 1.72 22:4 (n-6) 0.68 ± 0.20 0.50 ± 0.20 22:5 (n-6) 0.36 ± 0.04 0.40 ± 0.05 18:3 (n-3) 0.39 ± 0.07 0.52 ± 0.25 18:4 (n-3) 0.44 ± 0.13 0.68 ± 0.59 20:3 (n-3) 0.14 ± 0.02 0.14 ± 0.03 20:4 (n-3) 0.45 ± 0.11 0.50 ± 0.19 20:5 (n-3) 11.14 ± 1.00 10.34 ± 0.94 21:5 (n-3) 0.19 ± 0.04 0.17 ± 0.03 22:5 (n-3) 1.54 ± 0.47 1.35 ± 0.45 22:6 (n-3) 26.64 ± 3.43 26.87 ± 2.08
∑PUFA 47.53 ± 2.78 47.33 ± 1.03 0.04 0.84
∑PUFA (n-3) 40.92 ± 3.26 40.57 ± 2.34 0.08 0.78
∑PUFA (n-6) 6.61 ± 0.84 6.77 ± 1.78 0.07 0.80
(n-3)/(n-6) 6.32 ± 1.20 6.47 ± 2.12 0.03 0.86
Fatty acids relative to sample wet weight (%)
2.12 ± 0.47 2.27 ± 0.73 0.32 0.58
Cholesterol relative to sample wet weight (%)
0.21 ± 0.06 0.20 ± 0.03 0.17 0.68

157
Appendix 3.2. Model summaries for linear regression of log-transformed weight-at-length
relationships in female and male Atlantic cod from areas of high and low salmon farming density.
FEMALES
Model term Estimate SE t p
Intercept 1.63 0.10 17 <0.0001
log(W) 0.32 0.01 26 <0.0001
Treatment(LFD) -0.02 0.01 -2.5 0.014
R2 = 0.88 Residual df = 105
MALES
Model term Estimate SE t p
Intercept 1.56 0.96 19 <0.0001
log(W) 0.33 0.00 31 <0.0001
Treatment(LFD) 0.02 0.00 2.0 0.06
R2 = 0.95 Residual df = 45

158
Appendix 3.3. Model summaries for effects of farm density on egg production, egg quality and
larval quality of wild Atlantic cod. Model terms are Farm density group (LFD and HFD), Day, Time
(early or late season sampling), total female length (TotalFL) and mean female length (MeanFL).
Model summaries for X2 likelihood ratio tests of model terms are omitted, as are model
summaries for analyses within farm density groups.
Daily egg production (DEP) Model specification: DEP ~ Group + Day + TotalFL + (1|Group/Tank) Model term Estimate SE z p
Intercept 1.55 0.48 3.2 0.001 Group (LFD) 0.53 0.28 1.9 0.056 Day -0.017 0.008 -2.0 0.042 TotalFL 0.008 0.006 14 <0.0001 Observations 420 Residual df 413
Relative daily egg production (RDEP) Model specification: RDEP ~ Group + Day + (1|Group/Tank) Model term Estimate SE z p
Intercept 5.32 0.19 27 <0.0001 Group (LFD) 0.12 0.18 0.7 0.52 Day -0.07 0.007 -10 <0.0001 Observations 420 Residual df 414
Egg viability rate (ViablePr) Model specification: ViablePr ~ Group + Time + meanFL + (1|Group/Tank) Model term Estimate SE z p
Intercept 3.7 1.48 2.5 0.012 Group (LFD) -0.09 0.18 -0.5 0.61 Time (late season) -0.81 0.03 -6.3 <0.0001 MeanFL -0.03 0.02 -1.4 0.18 Observations 348 Residual df 341
Egg diameter (EggSize) Model specification: EggSize ~ Group + Time + meanFL + (1|Group/Tank) Model term Estimate SE z p
Intercept 0.82 0.16 5.6 <0.0001 Group (LFD) 0.07 0.02 3.9 0.0001 Time (late season) 0.08 0.004 20 <0.0001 MeanFL 0.005 0.002 2.1 0.045 Observations 1038 Residual df 1031
Egg fertilisation rate (FertPr) Model specification*: FertPr ~ Group + Time + (1|Group/Tank) *MeanFL omitted due to poor model fit Model term Estimate SE z p

159
Intercept 0.96 0.22 4.4 <0.0001 Group (LFD) -0.16 0.25 -0.6 0.52 Time (late season) -0.64 0.25 -2.6 0.009 Observations 72 Residual df 66
Egg symmetry rate (SymPr) Model specification: SymPr ~ Group + Time + (1|Group/Tank) Model term Estimate SE z p
Intercept -2.0 2.7 -0.7 0.50 Group (LFD) 0.15 0.35 0.4 0.67 Time (late season) 0.36 0.26 1.4 0.18 MeanFL 0.05 0.04 1.3 0.20 Observations 72 Residual df 65
Egg survival rate during incubation (SurvPr) Model specification: SurvPr ~ Group + Time + MeanFL + (1|Group/Tank) Model term Estimate SE z p
Intercept -2.2 3.6 -0.6 0.54 Group (LFD) 0.03 0.41 0.07 0.95 Time (late season) -1.0 0.39 -2.7 0.007 MeanFL 0.06 0.06 1.0 0.31 Observations 23 Residual df 16
Egg hatching rate (HatchPr) Model specification: HatchPr ~ Group + Time + MeanFL + (1|Group/Tank) Model term Estimate SE z p
Intercept 3.1 2.6 1.2 0.24 Group (LFD) 0.13 0.30 0.43 0.67 Time (late season) 0.57 0.26 2.2 0.030 MeanFL -0.02 0.04 -0.54 0.59 Observations 45 Residual df 38
Larval length (LarvL) Model specification: LarvL ~ Group + MeanFL + (1|Group/Tank) Model term Estimate SE z p
Intercept 6.4 1.7 3.6 0.0002 Group 0.46 0.21 2.2 0.029 MeanFL -0.006 0.03 -0.2 0.83 Observations 356 Residual df 350
Maximum larval length (MaxLarvL) Model specification: MaxLarvL ~ Group + MeanFL Model term df SS F p
Group 1 0.39 1.6 0.24 MeanFL 1 0.01 0.04 0.84 Residuals 9 2.2

160
Observations 12
Larval deformity rate (DeformPr) Model specification: DeformPr ~ Group + MeanFL Model term Estimate SE z p
Intercept 0.88 3.1 0.28 0.77 Group -0.07 0.38 -0.18 0.85 MeanFL -0.02 0.05 -0.31 0.75 Observations 12 Residual df 8
Larval phototaxis rate (PhotoPr) Model specification: PhotoPr ~ Group + MeanFL Model term Estimate SE z p
Intercept 0.07 3.3 0.02 0.98 Group 0.30 0.41 0.74 0.46 MeanFL 0.01 0.05 0.19 0.85 Observations 12 Residual df 8

161
Appendix 4.1. (Left) Large Asterias amurensis individuals (>20 cm) feeding on fallen mussel
clumps with Coscinasterias muricata. (Right) Attempted predation of Asterias amurensis by
Coscinasterias muricata. Image credit: Emily Fobert.

162
Appendix 4.2. Model summaries for effects of shellfish farms on seastar population metrics.
Model summaries for X2 likelihood ratio tests of model terms are omitted, as are model
summaries for analyses within locations.
Asterias population density (Density) Model specification: Density ~ Habitat + Location + (1|Date) Model term Estimate SE z p
Intercept 7.6 0.96 8.1 <0.0001 Habitat (Out) -0.81 0.60 -1.4 0.18 Location (GP) -10.5 1.5 -6.8 <0.0001 Observations 76 Residual df 71
Asterias mean transect arm span at Clifton Springs (Size) Model specification: Size ~ Habitat + (1|Date) Model term Estimate SE z p
Intercept 3.0 0.05 55 <0.0001 Habitat (Out) -0.2 0.04 -5.5 <0.0001 Observations 38 Residual df 34
Asterias gutted weight at Clifton Springs (GuttedW) Model specification: GuttedW ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 9.2 1.1 8.4 <0.0001 Habitat (Out) -4.3 0.46 -9.4 <0.0001 Observations 192 Residual df 187
Asterias drained weight at Clifton Springs (DrainedW) Model specification: DrainedW ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 16 2.0 8.1 <0.0001 Habitat (Out) -6.5 0.78 -8.3 <0.0001 Observations 192 Residual df 187
Asterias gonad weight at Clifton Springs (GonadW) Model specification: GonadW ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 4.2 1.8 2.3 0.022 Habitat (Out) -1.1 0.33 -3.5 0.0004 Observations 192 Residual df 187
Asterias gonadosomatic index at Clifton Springs (GSI) Model specification: GSI ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 0.42 0.15 2.8 0.005 Habitat (Out) -0.08 0.02 -4.9 <0.0001

163
Observations 192 Residual df 187
Asterias pyloric caeca index at Clifton Springs (PCI) Model specification: PCI ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 0.69 0.02 33 <0.0001 Habitat (Out) -0.04 0.01 -4.1 <0.0001 Observations 192 Residual df 187
Coscinasterias population density (Density) Model specification: Density ~ Habitat + Location + (1|Date) Model term Estimate SE z p
Intercept 7.1 0.31 22.7 <0.0001 Habitat (Out) -6.5 1.1 -5.8 <0.0001 Location (GP) 4.2 1.1 3.7 0.0002 Observations 76 Residual df 71
Coscinasterias mean transect arm span at Grassy Point (Size) Model specification: Size ~ Habitat + (1|Date) Model term Estimate SE z p
Intercept 31 0.56 55 <0.0001 Habitat (Out) -2.1 0.66 -3.2 0.001 Observations 32 Residual df 28
Coscinasterias gutted weight at Clifton Springs (GuttedW) Model specification: GuttedW ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 16.4 1.5 11 <0.0001 Habitat (Out) -2.0 0.53 -3.7 0.0002 Observations 158 Residual df 153
Coscinasterias drained weight at Clifton Springs (DrainedW) Model specification: DrainedW ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 21 2.3 9.1 <0.0001 Habitat (Out) -2.7 0.76 -3.6 0.0004 Observations 158 Residual df 153
Coscinasterias gonad weight at Clifton Springs (GonadW) Model specification: GonadW ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 1.5 0.56 2.6 0.009 Habitat (Out) -0.01 0.23 -0.04 0.96

164
Observations 158 Residual df 153
Coscinasterias gonadosomatic index at Clifton Springs (GSI) Model specification: GSI ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 0.41 0.12 3.4 0.0008 Habitat (Out) -0.005 0.02 -2.8 0.78 Observations 158 Residual df 153
Coscinasterias pyloric caeca index at Clifton Springs (PCI) Model specification: PCI ~ Habitat + (1|Date) + (1|Sex) Model term Estimate SE z p
Intercept 0.66 0.01 50 <0.0001 Habitat (Out) -0.02 0.009 -2.6 0.010 Observations 158 Residual df 153

165
Appendix 5.1. Study site characteristics for Chapter Five.
We conducted surveys of reef fish communities at Kirk Point, Point Cook, Altona Reef,
Williamstown and Half Moon Bay (Fig. 5.2). These locations had dense Undaria patches adjacent
to reference habitats of turfing algae or urchin barrens. Kirk Point is primarily urchin barrens
with areas of ephemeral macroalgae, including Undaria, Ulva and Gracilaria. Point Cook is
adjacent to a no-take marine reserve and contains a variety of rocky reef habitats, including
large areas of urchin barrens adjacent to mixed Ecklonia and Sargassum patches and areas of
Undaria, Ulva, Gracilaria and Caulerpa. Altona Reef is largely barren, with patchy, ephemeral
growth of Undaria, Ulva and Gracilaria. Williamstown contain large remnant beds of Ecklonia,
alongside habitats such as urchin barrens, turf, mixed Sargassum/Cystophora forests and diverse
ephemeral macroalgae including dense Undaria patches. Half Moon Bay sites were
characterised by extensive urchin barrens, mussel beds, and diverse macroalgae including
Sargassum and seasonal growth of Undaria and Ulva.

166
Appendix 5.2. Diagram of the tank used for the habitat choice experiment.

167
Appendix 5.3. Results of habitat choice experiment. Preference was tested using X2 test of
proportions, using equal preference for habitat options as the null hypothesis. Sample sizes are
lower for comparisons between macroalgae species as fish that selected barren rock were
omitted from this comparison.
N df X2 p
Heteroclinus perspicillatus
Initial choice
Overall 48 3 20 0.0002
Macroalgae vs. barren 48 1 36 <0.0001
Between macroalgae 45 2 8.4 0.02
20 min choice
Overall 48 3 7.3 0.06
Macroalgae vs. barren 48 1
121
33 <0.0001
Between macroalgae 44 2 0.2 0.9
Neoodax balteatus
Initial choice
Overall 23 3 6.7 0.08
Macroalgae vs. barren 23 1 12 0.0004
Between macroalgae 20 2 4.3 0.12
20 min choice
Overall 23 3 16 0.13
Macroalgae vs. barren 23 1
121
16 <0.0001
Between macroalgae 21 2 2.0 0.4

168
Appendix 5.4. Species counts from underwater visual census (UVC) and baited remote underwater video (BRUV) surveys. Number of UVC plots or BRUV
deployments given in parentheses. Focal species for this study are highlighted in bold type.
Family Scientific name Common name
In-season UVC Off-season UVC In-season BRUV
Undaria Barren Undaria Barren Undaria Barren Ecklonia
(25) (26) (9) (9) (14) (15) (5)
Apogonidae Vincentia conspersa Southern cardinalfish 2 2 1 1 2
Apogonidae Siphamia cephalotes Wood's siphonfish 1
Atherinidae Atherinosoma microstoma Smallmouth hardyhead 1
Blenniidae Parablennius tasmanianus Tasmanian blenny 3 8 2 8 2
Callionymidae Foetorepus calauropomus Common stinkfish 1 4 1 4 1
Centrolophidae Seriolella brama Blue warehou 16 3
Clinidae Cristiceps australis Crested weedfish 2 1 1
Clinidae Heteroclinus tristis Longnose weedfish 2
Clinidae Heteroclinus spp. Common weedfish and allies 38 1 8 3
Clinidae Ophiclinus ningulus Variable snakeblenny 1
Dinolestidae Dinolestes lewini Longfin pike 1
Diodontidae Diodon nicthemerus Globefish 5 1 1 4 3
Engraulidae Engraulis australis Australian anchovy 65
Gobiidae Nesogobius pulchellus Sailfin goby 2 1 4
Gobiidae Unidentified Goby 1
Hemiramphidae Hyporhamphus melanochir Southern garfish 11 65
Heterodontidae Heterodontus portusjacksoni Port Jackson shark 1 1
Kyphosidae Girella zebra Zebrafish 5
Kyphosidae Tilodon sexfasciatus Moonlighter 1
Labridae Notolabrus tetricus Bluethroat wrasse 1 3
Labridae Neoodax balteatus Little weed whiting 3 9 4 7
Labridae Pictilabrus laticlavius Senator wrasse 1
Loliginidae Sepioteuthis australis Southern calamari squid 1 1
Monacanthidae Brachaluteres jacksonianus Southern pygmy leatherjacket 1 2 4 1
Monacanthidae Meuschenia freycineti Sixspine leatherjacket 1 1 2 1 1
Monacanthidae Scobinichthys granulatus Rough leatherjacket 1 1 2
Monacanthidae Acanthaluteres spilomelanurus Bridled leatherjacket 1
Monacanthidae Eubalichthys mosaicus Mosaic leatherjacket 1
Monacanthidae Meuschenia hippocrepis Horseshoe leatherjacket 4 5
Mullidae Upeneichthys vlamingii Bluespotted goatfish 1 1
Platycephalidae Platycephalus laevigatus Rock flathead 1 1
Plesiopidae Trachinops caudimaculatus Southern hulafish 5 3 9 34 6
169
Pomacentridae Parma victoriae Scalyfin 1
Rhinobatidae Trygonorrhina dumerilii Southern fiddler ray 3 1
Sillaginidae Sillaginodes punctata King George whiting 1
Sphyraenidae Sphyraena novaehollandiae Snook 5
Tetraodontidae Tetractenos glaber Smooth toadfish 5 4
Antennariidae Trichophryne mitchellii Spinycoat anglerfish 1
Tripterygiidae Trinorfolkia clarkei Clark's threefin 20 19 15 25 1
Urolophidae Urolophus paucimaculatus Sparsely-spotted stingaree 2 1
Unidentified Unidentified Unidentified 31
Species richness 14 9 13 11 19 18 11
Combined MaxN 81 43 39 61 169 100 54

170
Appendix 5.5. Results of permutational ANOVA comparing fish communities surveyed by underwater
visual census (UVC) and baited remote underwater video (BRUV). Habitats are Undaria and Barren;
Locations are pooled into western or northern Port Phillip Bay.
df pseudo-F p
UVC IN-SEASON
Habitat 1 10 0.0002
Location 1 4.4 0.002
Habitat x Location 1 4.9 0.002
Residual 47
UVC OFF-SEASON
Habitat 1 2.5 0.04
Location 1 1.9 0.10
Habitat x Location 1 1.1 0.41
Residual 14
UVC OVERALL
Habitat 1 7.6 <0.0001
00 Season 1 6.9 <0.0001
Location 1 4.0 0.002
Habitat x Season 1 1.3 0.26
Habitat x Location 1 3.0 0.01
Season x Location 1 1.7 0.14
Habitat x Season x Location 1 1.0 0.43
Residual 61
BRUV IN-SEASON
Habitat 1 1.2 0.28
Location 1 1.9 0.06
Habitat x Location 1 1.6 0.11
Residual 25
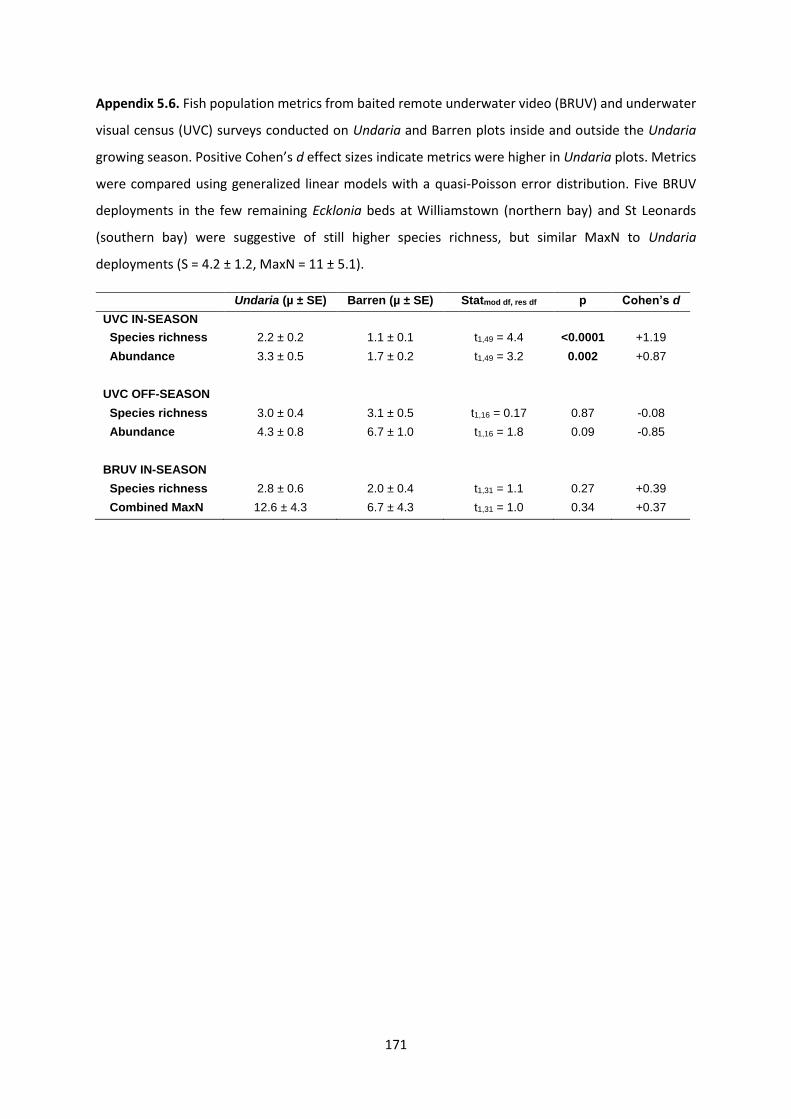
171
Appendix 5.6. Fish population metrics from baited remote underwater video (BRUV) and underwater
visual census (UVC) surveys conducted on Undaria and Barren plots inside and outside the Undaria
growing season. Positive Cohen’s d effect sizes indicate metrics were higher in Undaria plots. Metrics
were compared using generalized linear models with a quasi-Poisson error distribution. Five BRUV
deployments in the few remaining Ecklonia beds at Williamstown (northern bay) and St Leonards
(southern bay) were suggestive of still higher species richness, but similar MaxN to Undaria
deployments (S = 4.2 ± 1.2, MaxN = 11 ± 5.1).
Undaria (µ ± SE) Barren (µ ± SE) Statmod df, res df p Cohen’s d
UVC IN-SEASON
Species richness 2.2 ± 0.2 1.1 ± 0.1 t1,49 = 4.4 <0.0001 +1.19
Abundance 3.3 ± 0.5 1.7 ± 0.2 t1,49 = 3.2 0.002 +0.87
UVC OFF-SEASON
Species richness 3.0 ± 0.4 3.1 ± 0.5 t1,16 = 0.17 0.87 -0.08
Abundance 4.3 ± 0.8 6.7 ± 1.0 t1,16 = 1.8 0.09 -0.85
BRUV IN-SEASON
Species richness 2.8 ± 0.6 2.0 ± 0.4 t1,31 = 1.1 0.27 +0.39
Combined MaxN 12.6 ± 4.3 6.7 ± 4.3 t1,31 = 1.0 0.34 +0.37

172
Appendix 5.7. Conceptual model outlining the temporal relationship between Undaria pinnatifida and
reproductive phases of macroalgae-associated reef fishes. Undaria habitat quality is most likely to
influence reproductive success when seasonal Undaria canopy coincides with periods of reproductive
provisioning, larval settlement, or both. The kernel area where Undaria density overlaps with a
reproductive phase is proportional to the likelihood of an individual being influenced by Undaria
habitat during that phase. Plots represent (A) minimal temporal overlap between Undaria canopy and
reproductive provisioning and no overlap with settlement, (B) moderate coincidence during
reproductive provisioning but settlement largely decoupled (e.g. Neoodax balteatus), and (C) strong
coincidence with both reproductive provisioning and settlement (e.g. Heteroclinus spp.).

Minerva Access is the Institutional Repository of The University of Melbourne
Author/s:
Barrett, Luke
Title:
Habitat preferences and fitness consequences for fauna associated with novel marine
environments
Date:
2017
Persistent Link:
http://hdl.handle.net/11343/216400
File Description:
Complete thesis
Terms and Conditions:
Terms and Conditions: Copyright in works deposited in Minerva Access is retained by the
copyright owner. The work may not be altered without permission from the copyright owner.
Readers may only download, print and save electronic copies of whole works for their own
personal non-commercial use. Any use that exceeds these limits requires permission from
the copyright owner. Attribution is essential when quoting or paraphrasing from these works.




















