
(O lECOu. /ji) 1 uiI'l(A
89
(O lECOu. /ji) 1 uiI'l(A
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of (O lECOu. /ji) 1 uiI'l(A

(O lECOu. /ji) 1 uiI'l(A

ImY BEAN! F1íV5HlU16Y OF VlEW f'OmmAl A!II) SHlE55 T!LERAM:E
,l~lfr., W. I4hite, Centro Internaoonal .le Agrímltura Tropical,
(CIAH
ana
Juan l!quíerd", FAO Regi~lal Dflice lar
latin ~ie. ana toe Caribbean.
Sanhago, Chile FAO Regional Gllíee lar latin Amerita and lhe Carlbbean
198'1

ThE' dl'sígnatlons l'iIl!lloyed ilnd ti*! presenhtíon 01 material in this publícatíon do nat ímply the e:<pression 01 anl' apinion whitsoever Of! the part 01 the Food and Agrículture Organízation 01 the Urnted rlatlons concerning the lega! status 01 ani country, terdtory, cí ty or area or 01 its iluthorities, or cOfleprning the delimitalion 01 its frontíers or boundaríes.

This manuscript was preíured origina!!", ny t!essrs. JeHrey W. White, Ph.D., f'hnt Physiologist, !lean F'rC-gra.iIl, Centro InternacIOnal de Agricultura Tropical (ClAn, Apartado Aéreo biD, Cali Coloobia, and Juan Izquierdo, Ph.D., Croll f'hysiologist, Regional Plant f'roouctioo OHicer, FA!) Regional affiee fOf latin America ano tne Caribbean, P.O. Box l~j15, Santiago Chile, as one chapter (lf tne OOOt Sean F'roouction illlá I!IijlfoveiIBf1t in the Tropiés which was ¡ntended to be pu!J!ished by CIAT. Very long delays in preparation of otner chapter manuscripts are predudiog tne produdion 01 the boo~, so in order to fii'lf:e toe information ilHilable to beil.11 researchers and e¡Üenslonist that are participating at toe FAD/RLAC Fooa Crep; Teconical Cooperatioo Netwod: and ta CIAr !lean Progralll cooperators, it Has decided to publish this document through a joint effort of [IAI ilnd FAD.
As of luIy, 1989, it is expected that the boo~ now entitIed Common Bean: Research for Crop Improvement will be availabIe in 1990.


l. Introduction 11. Basie Processes
A. Srowth Processes l. Fhotosynthesis 2. Respiratioo }. Ni trogen Hetabol i sm
3. lli!\'eIilflll!ef1tat f'rocessEs l. Germinatían 2. Vegetatíve Srowth 3. Reproductiva Srowth 4. S~eSi:ence and Ahseí s5i on
C. Integraban of f'rOCe5Se5
LONTENI
l. CarDon and Ni trogen Partí tianíng 2. The 5ouree-Sink l10del
Ill. Yield PotentíaI A. Introduetion
l. Estimates of rieId PotentiaI B. (ield f'otential 's Agronomic Hanaqeíll€i1t C. Physiologíeal Strategies lar lncreasing \'ield
l. Saure. Limitatians - Photosynthesis - Respiration - Light utilizatian
2. Sinl Limitabans - Flowar, seed and pod abortion - Pod growth rates - Ilarvest inde~
3. Assimilate partítianing 4. VieId Components S. Integrated Strategies 6. A New Strategy 7. 5pecial F'fobl elllS
- Nediuil! to large s.edad cultivars - Early maturity
8. Potefitial of Biotechnology or otnar Strategies in the Fuiure
7
7
11
lb !~
l7 19 19 l S' 21 21 21 21
25 25 29
" .JJ
34 7< .,;;..1
36 36 -39 41

page
IV. F'raductivity Under Stress ~3 A. Ilrought 43
1. EHects of drought 43 - Ra¡¡íd respOO5e!i 4S - Responses liithin hoors 4b - Respoose over looger perlads 46 - Campad son of drought respüflse of ather IegUJll!!5 4í
2. Ilrooght Tolerance MechanislI5 48 - Ilrought escape 48 - Ilrought tolerante wit" high plant water potential 5IJ
Maintenance of water uptal:e Sil Redudiüfl 01 water IOS5 5IJ
- Ilrought tolerance Hith low plant water potential 53 Maintenance of turgor 53 Desiccation tolerante 55 lntegrated mechanisMs: Assi.ilate partitiüfling 55
and remobilization , Ilrought tolerance thrcugh adaptatino to conditions 56 .,. assQciated Hith drought
4. [ooc!usion 59 B. Excess Soi! Water 59 C. 5alíne Soils 59 D. General¡zed Stress Tolerance: "rustid tr" bl
v. Conclusioo 65

1. INTI\'OIlUCTHlN
Tne COImOfl bean (fnaseolus vulgaris l has a paor reputation both in teras of yíeld potential and toleranee to physiological stresses such as drought, flooding, aOO saline soils. lt is iaperativ! and necessiry then to ¡ocrease be.n productivity in view al íti agricultural ¡aportanee and essentíal participation in the diet of .¡llíons of inhabitant. of Latin Aierica and ths Caribbean as He!l "s úf other regions in ths world. While agricultural ressarch progresses without being ibis to aake a breach in tbs barriers th.t restraín th!! yield of this crop, particularly under stress eonditions, regional bean produdivíty has decreased over the last 40 years. In llraziJ, the lain producing country, bean productivity in the paorer regions is 405 l:g ha-l. (illves, 191m. 1
Various researcn prograls have reported experilental yields over 5 tonlha in bush beans, and genetic variability lor yield potential and stress toleranee llave besn di!ilOStrated.
AJtn¡¡~gh tllese findings have not lead to drailatic increases in bean productívity as has occurr"d with other crop species, ph~'siological studies 01 beans, and otller grain leguIII!5 sutil as soytean (6hcine iIIax) and cOllpea (Vigna unguiculata), have generated useful inforutíon lor guiding agronOilic manageillent and suggesting strategies lor varietal iaproveillent of beaos, aod as our knowledge ¡ncreases, further practica! benefits lIill certain!y be forthco~ing.
Traditiona! vadeties of beaos respond paorly, in general I to the two agronoilic lIIanagl!íllent e!elents recOHended by tlle green revolution, aOO which have lIIade it püssíble to obtain lIigh yidds in otller cr¡¡ps: Iligh plaot density and fertilization. lo spite 01 thi!se constraints. traditiotial varieties contaio a valuable genetic variabilí ty which is use/ul lar plant breeders Ul the search lor resistance ta pest. and diseases or atoer coaracteristics as.ociated to l1igh yield. New varietie. of beaos and appropriate crap fianagl!leot tecnnologies suitible for the e~lditions of the smill farlars in developing countries, are eSSi!l1tidl io the next )'sars. lo this conte~t, biotechnology can provide, in suppart to genetic iaproveeent, new alternativ!!s to acnieve tois obiective. Howe~'er, adaptatioo of these future innavatíons to the traditionaJ farlling pradices 01 the Sliall farll!fs and ta tne ecologieal conte~t in whieh tlley will be introduced, IllUst be specifically considered. Senetic impravement and pl~lt physiology IllUSt converge in results expressed as new varieties oi oean., more "rustie' and adapted to marginal conditions. Tnese Cin initiate the expected "neH revolutioo' in this crop. A dear understanding of the processes and physiological phenalena tnat deterline anó regu!ate toe expression 01 yield, is basie far toi. new stage.
Due to the diversity and comple:dty al' physiological eVi!I1t. deterllining yield potential, the autllors naVE! presented both a descriptive aod integrative lOOK at tlle pnysiolagy af the bean plaot. rile chapter first discusses basic physiological processes which deteraine bean yields, and than applies this informatioo to yield potentía! and stress tolerance. Han~' unpublished results obtained al research work conducted in latín America were i"e!uded to illustrate physiological phenomena not considered in previous studies.
I IUves, E. 1'i'87. Os desafios de e~teosdo rural brasileira. Mimeograph. (lJnpublishedl.

- 2 -
Texts and journals an plant pnysiology cantain .any studies an the role of grOHth regulators in bean plant;, but tfil shall 001)' refer to sucll wor!: whea evidente edst; for eHl!Cts 00 5pl!Cífic physialogical process!!s. Although tllere are lIIaoy unknOHns coocerning plant grOHtll regulators, tlleir overal! role appears to be ooe of translllissioo of regulatory inforlUtioo frOl ane tissue to another. lo relatían to crup grOHth probleA\s, studíes of growth regulators are generally of ooIr indirl!Ct use, as e~pressed by lIit aod Penníng de Vrie; (198.3): "/\ horlllOn!l systeA\ i s a c_ic.lUoo systeil. SUch syst!!11lS lIay be anal ysed eí ther in terlllS of means of the hardware used or in terllll of purpose of tlle lessages transferred. In d)namic models of crop growth, interest should be focused on lile latter, whereas tlle existence of tlle hardware, the hormonal systellls, lIay be taken for granted".
5pace lilllitations preclude a cüíIljllete literature review 00 the physíology of this crup but interested readers can fínd pertinent and complelllentary inforlUtion in Grahaa (197B), Laing, Jone5 aod Ilavi 5 (1984) I Tanaka and Fuji ta (l979! I and Wallace (l9B5).
1

------- ~ --
- 3
n. 8I\SIC f'ROCESSES
Physiological processes in a plant lay be classified ioto aetabolic events sucn as photosyothesis and rempiratiOl1 Hhich nave a direct eHect 011 grOHth, and IIOfphogenic OI1es Hhích aHect the change in far. or development of the planto The tIlO groups of procesSi!s interad strongly, but are best discussed separately.
A. Growth ProcesSi!5
Growth llliy be lllE!asured in eany ways (slee elongatíon, accumulatiOl1 of nades, leal e~pansion, etc.l, but emphasis is given here to the processes Hhich alfect dry weight accumulation and its partitioning sinee yield is the ultie.te e.pressíon of these processes. Accumulatíon of dry weight in bean plants is a dírect consequence 01 the balance ilIIIOng photosynthesis, respiraUon, and lossos due to senescence iOd abscission. Partitíoning, on the other hand, establíshes an equilíbriua between vegetative and repraductive growth integrated over tne development of beans, resulting in an end praduct of yield.
1. Photosynthesi SI
1\5 in other grain I egUffi!!S , beans have a C~ photosynthetic pathway. EsU.ates of lftiI.iilUfi le.f photasynthetit rates (!\P) at albíeot Co. concentrations have ranged tra. 12 fig CO. dm-2 h-' to 35 fig
Co. d.-' h-' O.b1e U. Recent estiMatos have been relatively hígh, presumably reflecting i.proved measureaent techniques, but are still well below rates ot soybean., which CAn reaeh 45 to 6ú mg CO. d.-' tShibles et ~, 1975).
AP varies greatly Hitn leal age and eoviranmental cooditions. Leaves íncrease in photosi~thetic tapilcity as they e~pand, maiotain a lllaximum far a highly variable periad depending un cultivar and leal positian, and then decline as toe leal senesces (Fraser and Bidwell, 1974). TOis decline in AP appears to result primarlly frOlll 1055 ot photosynthetic capacity, but also involve. iocreased stOlllatal resistance (Dav!s and McCree, 19781. Tanaka and Fuiita 119791 detected a s¡gnilieant positíve carrelation between nítrogen cantent and AP in leal'25 at ages past the tilfte of M"illlUl rate, suggesting that 1055 01 photosynthetic cepacity is related to nitrogen remobilization. Si.ilar conclusions were reacned in soybeans ILugg and Sinclair, 19911, with toe clarific.tíon to.t toe linear relation only holds below a critical leal prote!n concentration levels 112 9 protein .• -· of leal in their studyJ.
Table 1. Reported values 01 1I\a.~ifiU. leal photosynthetic rah at near-ambient CO, concentrations.
____ ,Photosynthetic Rate __ _
Austin ~ Hiclean, 1972. Fraser ~ Bidwell, 1974 Sustril, 1962 He,leth ~ tlass, 1961 lzhar & Mallate, 1967 louwerse ~ lweerde, 1977 ~uene.an et. ~, 1979 Tanal.a & Fuiita, 1979 laíng et. al., 1'194
Ig eo, da-' h- I 9 CH,o .-2 h- I
24 1.6 19 1.2 22 1.4 12 1).9 15 1.0 22 1.4 JI 2 .. 1 35 2. '3 34 ~ ~
L ••

- 4 -
Ma~imul pnotosynthetic rates in beans are reached at irradiances 01 30Q N •• -2, the same level reparted lar soybeans IShibles et al., 1975). Burga (19781 found evidence in beans that rI' varies with the position of the leal "ithin the canopy, and reaches ¡ts !a~imus at the time of initial pod filling. Laing et fu (1'184) s!te unpoblished 1!Or~ 01 EI-SI1arl:a"ay detIonstratíng differenees in respoose of AP io two beao cultivars, with aptílld varying from 21"1: to 31"1:. No effect of water vapour pressure deficit (UFO) 00 AP Has detected by Hesse and Leaz (1982) usiog ~ tl~ illueioatioo at 22-C, but EI-SI1arkaway et fu (1984) fouod that beans had eo intermediate level 01 sensítívity as cOlpared to other craps. A possible explanatíon for the different conelusioos lies in ioteracting effects 01 irradiance, telperature and relative hUlidity on stomata opening, as reported in other species ILarcher, 19151.
Louwerse and Zweerde 119m lound tIlat maxi_ AP Has obtained frllll plants under high radiatíon regimes, and that AP Has pasitively correlated Hith leaf thickness, specific leaf weight and nueber of chlaroplasts, but fiot lIith chloraphyll contento Di Herent sources of ni trogen aHect AP too, apparently through eHeds 00 photoreactioos aHectiog oitrogen tetabolislII (Marques et fu. 1983'. Even siaple differences io container .ize can aHect leaf phatos\,nthetic rates as io toe study of ear.i et fu (1'183) where plants which had been stunted by beiog grown io small pots showed higher AF'.
Further complicatioos result froa the possibility that other parts of toe bean plant lay regulate photosynthesis independent of elfects of water or outrients. Carllli and Koller 11978! detected root eHeds 00 photosynthesis in priill3l"y leaves 01 beaos. Research in sOrtean suggests th!t il delilod from growing tissues is insuflicient le.g. as induced by artificial depodding), phot05yOthesis .ay be inhibited. Starch aceumulatian in chlarap1asts lIay playa I:ey role (Chattertoo and Silviu., 1979', although Carmi aod Sh~ (1979) found that bean lIIesaphyll cell. appeared able to .ccumulale large amounts 01 stareh without disruptiog the chloroplasls. Leal starcn io sOrbeaos has beeo found to be negatively eorrelaled with tne activity 01 the sucrose-P synthetase lHuber and Israel, (992). These findings support the postulate that sucrose"P sin!heta;e is I"ey in leguees in regulating photosynthetie lormatioo of suerase, and nence, stareh. The role of thi s enzyme, strongl y related to nitrogen content, appears to be critical in legume metabolic pracesses.
Breen bean pods fi~ ro. at 10_ rates lhan adjacent lea,es, and the rate decrease5 Hith pod age <Tanal,a aod Fuiita, 1979). Bean pods recyc1e substantial .mounts of internally-released COz, but are not ao i¡portant photosynthetie 50Urte of dry latter to developing seeds ICrooKstoo, O'Toole and Ozbun, 1974a).
Under eonditioos 01 high mesophy!l 62 concentratioos, such as oecurs when stOlale close uoder high light inlensities bul limited water availabi!ity, Oz competes Hith CO2 in reacting Hith F~Dr tarboxylase, with lhe illlflorlant dílference that chellical energy is expended and COz evolved. It Has lir5t thought that photorespiratíon, as it is terled, Nas either a Naste preces. or per~itted production of eOlllpounds essential for plant growth. No 5ueh fundion has been d2tlO!1strated, and !IOst researchers nOH favor the idea that photorespiration serVEs to dissipate light energy" I"l,en pnotOSyntheSi5 is blocked due to low COz cOl1centratioos (Pm;!es and OsillOlld, 1979; Tolbert, 1'I8Q) or to maintain COz levels relatívely constant Nithio 1!!5ophyll tissties (Leopold and r.íed!!ll!l1, 1975). Nevertheles5, Ogren U'I841 coosidered that oceurrence 01 photorespiratioo in anaerobic bacteria dlsproves the protectíoo funetion, and concluded that lower photorespiration rates still may be attaioable. Rites in beans have been shONn to vary w¡th environlental conditions lMartío el al., 19711. Hallace et ilc (1976) felt that this variabílity would .ake detection af gene!!e dilferenees in beaos diffitult.
Efficiency of the photo-assieilative preces5 i5 severely affected by .¡croelimate at the crap level. The distrlbutioo of radialion Híthio a be.n plant cammunitv i5 one 01 the most iaportant elements 01 .i(raclilate 01 the crop. Interceptioo 01 light cao be approxieated by the e.ponential decay funetion,
1

- 5 -
I = lo • ~p(-k t LI,
where lo is ne! radi.tíon aboye the canopy, 1 Is radi.tíon at a level correspondlng to L unit. 01 leal area index, and t Is the eKtinction coefficient, a characterlstic depending on leal orientation and di.tributíon U1ilthorpE! and IIoorby, 19791. !he review of the valuE!s of extinction coefficients lar various craps by tIonteith ((%91 Indicates th.t 1: faoges ¡rOll ¡.OS (horizontal leavesl to 1).24 (gras.!!. Nith vertical leaves). Data ¡roa Gardlner et ~(19791 lar two varietles plao!ed at 46 and 91 cm row spacings (Figure 11 gave va!ue5 of k o, 0.49 at 4b el and 0.56 at 91 el, suggesting that b~in leaves are ari~1ted In a relatively horizontal aanner, and confir.ing the expeetatlon th.t le .. "e. are l1IOre ereet at narrower spaclngs. !he SalE! tendeney was found by Burga 119781, Nho .lso reparted a highly signillcan! negative correlation hetween k and leaf angle, thís refleeting the abilityo, the hean plant to orient their le_ves with the position 01 the sun in the sky to increase canapy Iight interceptíon. Wien and Wallace !l973) and Sato and Gotoh II'1!lJ) shOHed that lea! orientation responds to light, that sensors are contained bolh in pulvínules and leaflets, and that varietal dHferences in patterns Di arienhtíon existo
2. Respiratíon:
Althougn tt i5 biochesic.lly not a realistic separation, respiration 15 IOst easily considered as cüllposed of two coq:¡onents. One i. termed 'm.intenance respiration', and represent; respiratioo attributable to activitles associated Hith aaintenance 01 tissue lunctioo. Growth respiration is the other cüllponent, ano represents respiration associated with transport and biosynlhesls in the construdion al new tíssues during growth.
Hainlenance respiration i5 si.ilar to hasal eetabolislI\ in animals, and i5 thought to vary Hith tis.ue cüllposi!ion and teeperature. Austin and HeLean (19720) found that roots 01 bean plants in the prilWy leal stage had rales approxieately twice thoSl! 01 leaves and stems, and that rates doubled HIth a W"C increase in t~erature. Sale (1975) detected a si.ilar level of sensitivity lar dark respiration ef bean leaves.
Assuming varietal diflerences in plant strurture and teeperature adaptation, there i5 every reason to e~(pect as IllUCh variabílíty in eainlenance respiration rates as is found far photosynthesis. Genotypie varíation in several graln legUJleS Has detected by f'andey et al. 11976). Thls poiot
appear. unexplored in beans except Nhere dlHerences in dark resplration of leaves have been deteeted in .tudies 01 photosynthesis !Izhar and Hallare, 19b7l.
Estimates o, growth respiration rates are obtained from costs oi biosynthesis (Penning de \ries et. ~, 19741 de Wit et. ~ 19791. Using data on ti.sue coaposition 01 beans frOl various SOUlces ITable 21, approximate respiration costs lar roo!s, leaves, stefiS and pods have been calculated tWhite, 19911. Pods are the mast costlr due to their high protein content, while roats and stees require mucn less d~ to their relallvely high carbohydrate conteot (Table 2).
Since growth respíration is direetly related to energy requirE!fl!ents lor specifíc chl!fllical readions, ít is believed relatlvely teeperature insE!nsitive. 8íosyothetic pathHays are si.Har amng many specíes, so genetic varíability Is supposed ION beyond expeeted dilferences due to variability in plant cOIIPosition. f1oNever, estima tes 01 efficiency of carbohydrate use in nitrogen fhation (Nhich IS norllally accounted lor in aS5UlllJltions about (Osts 01 proteio synthesi.l indicate eajar varietal difference. (Kipe-Nolt, personal coffilllUl1icatíonl, aod enviroomental eHects and genetic varlability in beans need additional researcn.
Penning de Vríes and van Laar (t977) used sequentia! tissue analy5es to estiMate respiration costs of beans during gerlination and early seedling growth under dilferent temperature regimes.

- 6 -
.oo
o· _-0--.-.-75
50
/ /
/ /
/ í'
~
./ ./
./ . ./ eo
/
• /,,4"
/" o O
.,.....-p..--
0-- "'cm .- .....
o ~------r-------r-------.-------.-------. o • •
Hg. I Relatíoo betHeell lea! area !ndex and light ioterceptioo at 4ó ca and '11 ca row "!dths. Estimated IrOll data ¡¡f Sardiner et al •• 1'n7.
Table 2. Coapositioo of bean plant tissues and costs 01 bi05ynthesis. Data lar coap05itioos coopiled frOll Piper and I1!Jrse (mm. Jardan and Chíbnall 11933'. I'1cKee et al. 11'155) and Sindair and de Hit (1976).
Carboh)ilrate Protel" lipid ligoin Ash
leal Root stem Ped
-····----------··-1 01 dry weight--··-----------
61.7 20.11 7.7 4.0 5.8
79.9 9.5 0.9 7.0 3.7
77.4 11.9 0.6 7.0 3.2
60.9 30.2 1.5 4.0 3.4
------.---------.g CH.D g" tissue-----····-----
Cost ¡¡f bi05ynthesis' 1.50 1.29 1.32 1.52
• Source: Hhite. 1991.
I

3. Nitrogeo Metabalí!.:
The high proteín conteot of leguee seerls explaios the particular iapartance af nitrogen &et.bolís. in grain legule physiology. lo beao seeds,. protein content 01 20% to 241 iaplies a oitrogen content 01 approximately 41, "hieh in tUfO fieans that every 1,000 kg 01 yield ¡apiles a neerl lar 4~ ¡~ 01 N, not including amounts neerled to replace la.,e; due to leaehing or residual N in ntber tissues. It was found that up to 807, 01 tn. foliar N is reallocated to other plant parts belore th. onset 01 leal absci,ion (Izquierdo, 1981). And it Nill be reMeMbered that N content af lea,es is high, tnat cootent varíes with ontogeny Hable 3), and tnat it can show a direct eorrelation Hitb Ar.
Althaugh N aúy be introduced through the foliage, the Aor.al raute 01 uptake is through the root;, either as nitrate ar N:., whieh i s eooverted in root nadules to atllllOfliu. ion;. For our purpases, tt will suffice ta observe that once fixed as alMlOOiu., N is In a lorll readily convertible to aminü acids. In the case al N absorbed as nitrate, the nitrate Is first eooverted IrOl nitrate to nitrite by nitrate reducta.e, and tñen lrOl nitrite lo ammoniua by nitrite reductase. Th~
dífferent ~taboli[ pathways required with different N scurces appear to eflect efl¡cieney of NlIartitíoning U/!!stermann et al., 1995).
In beans, nitrate reductase activity is greatest in leal tis.ue. This per.it. tha reductioo reaction to be coupled to energy directly available from photos)nthesis, and this should reduce the apparent energi' cost as coapared to trops whith have greatest redactase activíty in roots (Hunter et ~, 1982). Nitrate reductase activity varies greatly with leal age and crop growth stage (Franco el ~,1919), and so~ level af actívity' i5 maintained even when nitrate is e~c1uded as a nítrogen source (Tiopo and Neyra, 1983). In other trops, considerable effort has gone to determining whether nitrate reductase activity is yield límiting, but oa clear positiv! results hav!! been obtaíoed (e.y. fal:orede anó Mock. 19191. The great phsticíty 01 nitrog!!flase aclivity in bean plants suggests the saoe wryJld be true far beans.
B. Deve!opmental Processes
As a bean plant grows. it undergoes not ooly a change in siz!!. hut changes in for. which are associated with lormation of flawers afid pods, anó eventually, yield. These ehanges are the result 01 developmental processes such as ste. elongation, ¡lower buó differentiation, and pod growtn. The processes are c10sely Iinl:ed to growth, but are here considered independently lor clarity. Rates of developoent are sensitiv!! to photoperiod and teoperature.
l. Germinationl
During the drying phase .ssociated with pod maturation, a bean seed enters a state af redueed /letabolic activity characterized by ehang!'S in dehydration of eellular membran!'S and asevere reduction in resplration rateo This condition is eaintained as long as the seed aoisture conteot reffiain. low. Upon contaet with a ~iu. 01 sulficiently high water availability, the seed illbib!!s water and respiration rates inereases óramatically. Cultivar. seem to vary in toe prOPortion af water taken up thrcugh toe micropyle or the raphe and hilufll, while upta!e thrcugh tbe seerl coat is negligible IKorban et ~,I~Bl). Some bean seeds present a coodition of 'hard seededness' such that water upta!!! is extr!!/ll!ly restricted. TM. appears to be due to a combination 01 lactors including restrictioos 01 the micropyle 1K)'le and Randall, 19631. Austin and Mac Lean (1972b) "oled tnat under tool 1l2"C) tl?ll!jJeratures, large seerled culthars germinat!! lIIOre rapidly than small seerled ooes.
2. Vegetativ!! Ilrowth:
Although growth analv.is (the Uge uf sequential samples 01 erop dry weight and leal area to describe crop growth) requires approximatioos innerent in coosidering the dry weight accuffllilation and

- 8 -
distribution as the SUI of the various growth and develapmental processes, suco studies have proven useful in describing trap plastic response to environment and effects of differences amoog cultivars.
Table 3. Mean organ values fO/' nitragen far nine dry bean genotypes at foor reproducti ve stages", Eas! lansing.
_____ ....!Reproducti ve stage-:--_-:::--:--:--:--c Hid-pod Hid-seed Physialagical
Plant argan mthesis elongation f¡¡llng laturity
-------------------------g .-'------------------------
Roo! .23 ab" .29 a .27 ab .24 ab Stetl I.SO ab 3.21 a 3 .. 31) a I.n b I'etiole .bS a .7b a .81 a .26 b leaf blade 6.36 b 7.91 a 5.4b b .S7 e I'od Hall 5.60 a 2.M b Seed S.16 b 13.10 a
• Means folloHed by the Sale letter in rows do not differ significatnly Duncan aultiple range test at 51.
• Source: Izquierdo, 1991.
Alter seedling establishaent, bean (raps under non-stress conditions generally follow a near expooential growth phas! until onsat 01 rapid pod growth. Growth then sl_, 5_ craps reaching zera crap growth and toen declining in weight, and others Obtaíning their .a~ieu. weight very near eaturity (figure 2).
HaKillUll crap growth rates (C6Rl reported for bush beans range Iroe 14 to lB 9 .-2.day-· (Table 41. Clilbing beans on trellises have rea(hed rates 01 ~) 9 .-2, the greater rates presuaably reflecting better canapy structure and less investment in support tissues. These values are less !han balf 01 vataes reparted lar ather C. craps (Manteith, 197B).
Met assililation rate (NAR, c¡¡¡cul.led as CGRILAI) usually declines wilh (rap aye. Typical valaes during Ilowering are lroIt 4 to 7 9 .-2.dar'. "igh vatues af l/AA ear reflect ca.pensation 10/' restricted leal area le.y. due to a determinate growth hahit) rather than hlyh photosynthetic efficienty sinee NAR during flowering and lAO froe emergenee to ooset 01 flowerlng eay be negatively carrelated aeong different cultivars (figure 3).
LAI increases with crap growth, and as Hith C6R, alten reaches a peak around tiee 01 onsat of rapid pod growth, the haH in leal production presUlilbly reflecting incrused dl!lland fO/' nitrogen In growing pads IFigure 21. LAI in deter.inate beans .ay stap increasing at a relatively early stage due lo lack 01 nodes ta produce additional luves. Nost bun craps Ul1derga nearly cOIIPlete leal senescente belDre aaturlty, but soee 'non-senescing' traps do occur. At CIAT-Pamira, non-senescing craps usually ínvolve cultívars with poor pod ar seed set, thus the lad 01 senescence prObah1V reflect. weak sink detland. In telllperate regíon., such a pattern eay ínvolve prObl1!lS 01 phatoperiod-teeperature adaptation, partlcularly .lowed growth with (ooler teeperatures (Hallace,

~ g ! r • ~ o
Al
!lOO
200
O
Bl
o
10
I i
I ,/
ICA eh ... 11
30
lAl ,
/ /
/ ',
/ .1
.1
10 '0
- 9 -
• •
•
O
50
• 3
,
50 'O
Fig. 2 Change in reproductive and total trap dry weights in relation te LAI in two be!" cultivar.: A) Porrillo Sintetice: 9) ICA Guali.
Table 4. Reported maKimum trap growth rates ICGRl and leal area inde~ leal area indeK IlAD) at time 01 maJiilllUll! tSR.
6rowth lladlllUl Culti var habit « CSR LAI Sourte
9 m-' day-"
HawkeSOury' iIooder 18 5.0 Sale, 1974 Micnoacan 12-A-3 1 17 5.0 Aguílar et al., 1977 Porrillo Sintetico , 14 3.2 Laing et !le, 1994 <
Porrillo Sintetico , 11 3.3-4.4 CIAr, unpubl i 5h~d <
JU 84-7 , 16 Rodr í guez, I 'iBb L
Flor de Mayo 4 21 7. O-Il. O Fi!I1jul et !le, 1982 X-'b441
• CIAr classification.

- 10 -
• • • • y. ".95 -O.0391C
;;. 'E g ~ ~
! 3
~
~ : • ; ~
2
•
•• 60 1.
lo' Ik .. dwat!Ort
fiq. 3 Relation between leaf area duratian IrOl emergence to flowering and Mean net assi.il.lian rate Irom Ilowering to Ilowering plus 10 days for lB bush bean lines. C!AT-Paleira, 197B.
/t;xillllll LAI'. are often in the range 01 1 to 4 unit.. These are Irequently below cpU.al lA!'. estieated as LAI at .a~ilU. LGR ¡Table SI.
Table 5. Comparision 01 maxiMUI LAI and opti.al LA! calcutated as l~1 estimated .a,imum CGR lor 1I sets 01 data lor ev. Porrillo SinteticQ al CIAT.
3.t 3.3 3.1 3.4 2.0 3.0 3.4 3.5 4.! 2.7 4.6
/lean 3.2
Optílial LAI
3.0 3.1 2.9 4.8 2.2 3.7 2.6 5.1 3.3 2.6 5.4
3.5
Yield
g .-2
219 227 277 277 2'15 29B 312 347 J5S lbS 412

- 11 -
Specific leal area (SLA = leal area/leal dry weightl usually start. low, iocreases to a ••• imum arOllnd flowering, and then declines toward .aturity (Figure 41. Iletereinate I'arieti.s oftm have cOflIparati,ely high SLA, again presullably re!l&ting cOflIpmsation lor restricted LA!.
UndEr non-limiting conditions, main stem leal initiation and expansion procede at near linear ¡ates (Figure 5). leal initiation and e~pansion rates diller greallyamong cultivar; (Table bl. Flo.a- ínitlation has he!!{l detected as early as 10 days after plantíng (WallacE, 1'i851.
Al
• • • • • • • ••
• • • "'" •
. ~ e (11'< Rdktud g • • ~ '00 Ob .... veddat • 11 • " ., •• - RfrQl'''si4::Jn .l! • """ & • ~ •
• •• e. e • • • •
• CY. E"Attco 23
200 o 00 D • .,. en .. p1en1int
Fíg. 4 Change in specifíc leal area aver tille in two!lean cultivars: A!RedUoud; Si Ex-Rico 23. Source I Nhite, 1981
3. Reproductive Growth:
Seans usually self-pollinate. Patterns 01 flowering opening are well known. Anther dehiscence accur just h2lor2 bud opming, usually at night. In P. acutHolius dehiscence i5 pr&eded by appearance al stigutic secretions, cOflIposed prÍllarily of carbohydrates and lipids, and the 5ecretions rupture the stigma cuticle (Lord and Kohorn, 199bl. Pollen tubes grows down the hollc~ ,tyle dnO fert¡l¡sation occurs within 12 hOllrs, ovules nearest the sttle being fertilised first.

I ! 8
J lo
11
i
- 12 -
tu' pfimot'die
o+---______ .-________ .-________ ~-o 2.
Hy. 5 l:utlparison 01 lIaifi stem leal initiatioo and leal expansion rates 01 ev. lCA Pijao grown under lield conditions. CIAr-Pal.ira, 1977.
rabie 6. Main ,tea leal ¡niti.Uan and expansian ratl!S lar cultivar; 01 differiog yrowth habito Palmira, 1977.
100 Leal Leal seed initihtion expansion
Cultivar ".bit weight rate rate
mg primordia day-l leaves day-l
63WJ7 la 0.45 I'J.YJ G 4498 1 16 0.42 O.3i G3816 1 60 O.:ro 0.21 G 1540 27 0.25 'Y. 18
G 1995 11 18 0.54 ü.32 G 3645 II 17 0.56 0.13 ¡¡ 4495 11 15 0.48 0.27 G 4525 11 19 0.48 0.26
S 1046 III 22 0.50 0.3J G 2\.~i6 III 23 0.43 0.29 63353 !ll 27 0.50 0.32 G 4122 III 22 0.45 0.26
!l 1091 IV ·30 0.52 0.31 6 2327 IV 23 0.60 1).29 G 2540 IV 19 0.65 0.33 G 2525 IV 20 0.60 0.37
Mean 0.48 0.29

- 13 -
Overall pod growth patterns in Il!9WReS are repartedly 50 siailar that they uy al! be Su.lrized as involving three distínct phaSB5: pod Hall growth, seed grOHth, and desiccation (Dure, 19751. In be.o pod. reaching aaturity 48 day. ¡¡lter flower opening, lIia~i.ue pod length HaS reached in 24 days, and pod freah Height dropped only in the last ¡, to 9 days, although seed Heíght continued to íncrease durlng the last phase (Oliker et al., 1979). Figure b ¡Ilustrates these events far black beans. lhe growth pattern 01 individual pods appears relativ~ly ínsensitive to source-sink manipulations (Figure ]). As would be expected in a trop Hith such diversity in phenology and seeds size, pod development rate varies greatly alllong cultivars lTable 7), but is not strongly correlated Hith ather variables Hallle S). TEiIIjloral changes af pod tomposition vary signifitantly illliOOQ 50ap bean cultivar; (Stork and Mahoney, 1942; Guyer and ~rallier, 19521.
t20 • 450
1>-.-.- tOO • "d 350
".
P i' l ,- 8. 4 :E 1 I r i 1 • 250 I 1"'" • - j • "'" .' w.~ht ""11 3 •
L''''Qln I .. ~
~
d 1 • I " I .., 2 tSO I
I
~ I 2. ,
I 50
Ó _.l.-_~ o o
•• o • 10 " 20 25 30 3' .., 4' • Or,s .her SO% flú_rif'l9
PM
l'íg. 6 f'oo and seed growth 01 cv. "Blaü Turtle Soup' bean. 5oorce: Izquierdo 1981.
Genetic dilferences exist within cultivars and breeding lines lor both rate and duration 01 seed fillíng in rice (Jones et al., 1979), soybeans (raplan and Kaller, 1974), aiize (Daynard et al., 1971; Iluncao, 1900; and Johnson and Tiinner, 19721, and tOHpea IHien and Adah, 19781. Several studies have shOHn that the duration 01 seed fillíng wa; lIiore dosel y related to yield than Kas the rate 01 lill109. Other warks (Egli and leggett, 1976; Sofield et al., 19771 agreed that the rate 01 seed lil1109 were rel.ted ta cultivar diflerences in final grain weight. In beans, negativa oon-sigoilitant correlations between the linear filling rate and yield Has reparted by Izquierdo and Hosfield (19831. HOl-lEver, toe same work, significaot positive corr.latíons were found between the lioear filling duration and eJlective lilli09 periad to yiEld, nu~ber 01 seed per pod, seeds.m-2 , and 100 seed weight among cultivar; and lines Hith differing architectural lores. Some correlatioo WiS
10000d among linear lilling duration and bean yields (f'aredes 19861.

- 14 -
2.' - Conm>I ....... Pod femov.l
D--<J Gefol¡.tMm
1.5
~
J t 1,0 > ~
'& .. 0,5
o .., .0 .0
o.ys from pl."hNQ
Hg. 7 Growth pattern af individual pods: Al as affected by defoliation and pod relaval in line ICA linea 24; Bl under norlllill conditions far foor bean Unes·.
rabIe 7. Vield, tla~i_ pod growth rate, and other par_ters of 20 lines. Palaira, 19I15a.
100 Mean rod Days to Dars to Oaily seed pod growth
line Yield flower. laturity yield weight dry wt. rate
9 .-2 9 day-' 9 9 pod-1 9 .-2 day-l
G 44'15 2400 40 78 10.8 19.& I.J3 0.125 RAIl 24 2370 42 82 29.1 21.0 1.04 .088 llAi 477 23(,1) 45 77 30.6 20.0 1.16 .II'? A 149 2350 48 B4 27.8 36.3 1.30 .083 llAr 271 Z29Q 42 SI 27.9 19.1 1.21 .092 G 400Q 2200 44 111 26.6 17.0 1.05 .097 65059 2190 40 so 27.4 19.2 1.13 .105 llAi 85 2020 42 79 25.6 20.0 1.26 .130 G 4446 1810 44 94 22.1 'l2 .. 7 0.93 • Q'i'I A54 1830 43 83 22.0 21. 2 1.27 .106 snm 1820 45 82 22.2 38.9 1.44 .093 IlAT 304 1910 37 70 25.7 14.3 l./)4 .1l0 6 4499 17(,1) 40 14 23.B 18.1 0.93 .078 IlAT 41 1740 33 6B 25.6 15.2 0.89 .094 G 3807 1650 31 71 23.5 14.4 0.72 .077 XAA 146 1570 38 71 21.6 22.5 1.04 .107 S 76 1510 38 7Q 21.6 39.4 1.39 .117 64494 1410 4Q 74 19.1 41.9 1.30 .102 61017 1110 28 58 19.0 14.9 0,79 .113 G 1540 8SO 36 68 12.2 17.1 0.7B .064 Mean lB5l! 76 24.2 22.6 1.10 .101 S.E. ISO I 2.0 3.3 0.12 .015

- 15 -
Considerable research has been dooe 00 iSjleds 01 bean !leed growth related Lo seed biochellistry (loewenberg, 1955; Carr and 5kene, 1961; Sussex and Dale, 19m, but such ~Is haye shed little light 00 factor; affectíng overall pod growth and eaturatioo.
rabIe 8. Correlatioos .u\ong rield, pod growth rate, aoa other par""",ter. measured io 20 lines. Palmira, 1?S5a.
Days to maturity 'íield/day /1)0 seed wt. Pod weight red growth r ate
Pod grcwth rate (excludiog 6 1541))
VieJd
.74U
.951$ -.03 .45~
.26
-.02
Vays to maturiti'
.501 ~2b ~51t
-.04
-.20
Daily rield
-.15
" .·)1
.00
gi(' S!!Ed
weight
.nu Jil,
-.02
Pod weight
.50
-----------,---------------t, U P ( .1)5 and p <: .01 respecti\'ely.
4. Seo.seence and Abscissioo:
~ter photesynthesis aod respiratiOO, the processes of senescence ana abscission play the lOS!
i"Portan! roles io determiniog plaot growth. Any tissue may uoderga seneseence, and in stem borne tis,ues, especially 1 eaflets, petioles, flower and pods, this process is usually acc""Piníea hy an apparently progr~med pattern 01 abscissioo. However, the tissues where SEnescence appears to be af greatest illportance are in leaves, flOHers and pods. Hypotheses coocerning the role of seneSCEnte and abscissioo usually follow three lines.
The mast frequeotly citad hypothesis far legumes ís that tissues which are at a dísadvantage io cOSjletition «lOng SDUrees aod sin~:s are eliminated, thu5 IIdintaining the overal! ellícieoey 01 tha planto Leaves are dropped which no langer fix sufficíent e02 (are too near the e02 cOSjlensatioo pointl. This aight result frcm loss of N due to remobilizatian ta growing pods, ar leaves losing efliciency due to shadíng by leaves higher in the canopy.
Abnormd! flower buds (Ya'ez et ~, 1984), and failures af pallinatioo or fertili,ation can resul t in abortion of flowers and young pods. However, it has been rep1lrted that 001 y ooe ovule needs to be fertili¡ed to prevent pod abscissioo, and stress enhaneed beao pad abscíssioo was 5t.ted nat to be caused by fai!ure in fertilization (Harterleio et ~,19801. Tnis dala ís supported by fiodings in soybeao reproducti ve shedding lllbernathy et ~, 1977; Wiedbold et ~, 19B1l. Studies of timing 01 bean flower and pad absclssioo nave shawo that pod ahscíssiao cao continue up to 4S days alter anthesis (Izquierdo and HOsfield, ¡98tbl. lt has been stated that older pads may drop if adequate carbon assimilate is nol available (fanal a ana Fuiíta, 19791. Sinca legu~es have a particular high oilrogen demand during podlill, sené5cence resuJting Irom competitioo lor N usually results in legume craps 'sell-destructing' (Sinclalr and de Nit, 1976).
A secood hypothesis i5 tnat senescence, delined bl' leapold, O?8l)) as detErior.Uve process", conducive to plant death, is a programmed process nf that occurs irregardless of the soorce-sink balance 01 the planto This Mechanís. is most easily imagioed as fuoctíooing in perennial plant. in

- 16 -
the prOCHS .¡gM persist as a relictua! charaderistic in armua! craps, having been reparted in pea (locthart and Gotlschall, IV61!, bean Ulareing and Seth, 19671 and 50ybean (Uru!oo and Nood!n, 1977/. Such a process HOIlld still sh!IH relations with pod set if grotrlng pods prOOuced a senescence
signal INhich cruld be either an inhibitor or stillUlator, depending en the pOO gr!IHth shgel.
The last groop of hypotheses censider 5enescence as a phy;iological disorder triggered by illbalances of endogenru5 growth regulatars. The reparted rise in abscisic acid (ABAI le,'eh during senescence lIa~es ABA an attractive candidate as a general inducer of senescence. but not all studies suppart this. In tbe case of beans, Nidely varying levels of ABA were frund during and before leaf sen!!scence IColquhoon and "iIlun, 19751. 1111 the other band, íf leaf senescence at sturity is triggered by ethylene prOOuctíon in uturing pOOs, rapid leaf fall .ight reflect nothing aDre tban an excessive sensitivity of leaves to ethylene.
The strongest argulle!lts far source-sink relations ccae froa studiH of eHech of depOOding and defoliation. Continual depadding, whether through artificial eeans ar physiological stresses causing fl!lHer and pod drap, generally result in delayed leaf senescence (Tanaka and Fujita, 19791. The fad that senescence uItieately does occur (an observation used to suppart the second hypothesis; NOod~n, 19901 eay be ascribed to eventual depletion of nutrient rHerves (particularly Nith artificial depoddingl, accumulation of diseases and pests, changes in agronOilic conditions such as cooler tellperatures or failures to Minlain irrigatioo treat~ents, or deleterious effects Di large accumulations 01 starch in leaves.
A secood Iíne Di evidence e_s froa studies 01 carbobydrate and nitrogen reaobilizatioo and leal photO!lynthetic rates, where patterns 01 senescence and relati005 betNeen nitrogen content and llF are in lull agreement with lhe idea that SOUfce sink balances detereine patterns Di senescente. Although the balance 01 evidence appears to favor a relation between sOUfcl!-sink balance and senescence, th!! question is lar frOil reso¡"ed. Tbimann (19901 and NOodén 119001 have written lengthy rev!ew! defending other lI!!Chanisas.
C. lntegration Di f'roce55es
l. CarbObydrate and Nitrogen Partitioning:
Sludies 011 ,oC distribution io beans [Seth and lIareing, 191.7; lukas et fu. 1916; Tanal:a and FUjita, 1979; Water el fu, 19001 have provided a fairly complete picture of qualitaUve patterns of carbohydrate distribution during crop growth. YOUllg leaves are th!!ir 0HI1 sink, incorporating all fi.ed carbon iota leaf tissues, buí as the leal expands, up to OO! of lixed carbon ea~' be expar!ed fro. a leal. As a leaf approache§ senescence, e.ports fal! Nith declining photosynthetic capability and intreased respiratary costs. A source-sink unit has been defined, consisting of a leal, any branch or rateme in the .xil of the leaf, and the internade below. The degree to which this unlt exparts assiMi!ate to other parts of tne plant varíes Nith growth habit, leal position, and growth slage. Stephenson and Nilson !19m, studyiog soybean Hlth I·C analysi., showed that this unH Nas able to account far up to 701 01 the ¡ssi,tllle distribution. !bis unit, initially tereed a 'nutritional unit' [Adall5, 19671, Nas subsequently naeed the 'phytoaeric unit' (Adall5, 19811 due to Its great physlological significante in carbohydrate sourcl!-siok balance.
Although the phytoaeric unit is a useful appro~i.ation, it sillplifies diHerences atIDfI!I leaves. l!lHer leaves eKpart a larg! prapartien to roots, while leaves at the highest nades of indeler.inate plan!s I!l!part relaUvely large prapartions to 1!IHer nades. RelIOving raceees at one nad!! results in greater eKportiltioo to adjacent nades both above and bel!IH. Vegetative tissues can serve as 5ites of actumulation of assi.ilate reserves. ReaDving pods Nill augaent reserves, while defoliation and Nater stress triggers assiailat. and N reMObilization (Izquierdo and Hernandez, 1984, unpublise4¡ lable 9).
J

- 17 -
rabIe 9. Water str!!'iS and ID deloliatien' .ean eHeds en dry.atter N sta retl!lb!ll2atien of shteen blad bean genotype'i, Chillan. (Hernandez, 1984 ¡ .
__ StH retl!lbilization (- )"_ Treaben! Ilry IlaUer Ni trogen
.13l: defoliation Ilrought Control
Sa y!!'i lSIl5X
21.63 20.32 19.&
2.55 I.B3 0.42
• The Hater holding (droughtl and n;: defoliation Nere applied at 501 flawering.
• Change in dry MUer and N content at st_ frOll 501 Howering to physiological aaturity.
5tudies on nitrogen distribution have not provided as ca.plete a picture due to the lack af techniques as convenient as those used "ith I·C. IIowever patteros of nitrogen ilwliIIlIlation and retl!lbilízation deterained through analysl!s ol tissue N suggest patterns siailar to carbon Hith necessary diflerences due to N entering tn!! plant through the roots, and to tne relatively high N requireoaents of leaves and pods (ilesteraan!1 al. , 19B5¡.
Basic patterns both of carbon and nitrogen distribution are prohably identical to those found in otner grain leguaes (Shibles et !le, 1975; Pate, 19751, and are consistent Hith the idea that muen uf bean plant grOHth and develapaent is regulated by eOlllpetition amng sources and sinl:9. Tbe si~e, eetabolic ¡ctivity, and proKiaity 01 a sinl: to sources S!!etl to be priMary deterMinants 01 the ability of a gi ven sinl: to ca.pete aaong other sinl:s.
Wñile this qualitative andel is of great use in understanding crap growth, lOre quantitative data are needed to deteraine Hhether allocation processes lílit bean yield. Ad_ el al. 11979) shoNed diflerences in accuaulation uf .tarcn-like sustances aenng diflerent cultivars, hut did not establish Hhether the differen~es Here due to variability specifically related to retlobilizatíon, or to otner lactors such as overall plant vigour ""íeh could indirecUy control !he allOUnt of assi.ilate available lar remobilizatíon. However the technique they used is apprapriate only far very gross genotypic separation, and Has fOUM to hay! a IOH correlation Hith quantitative staren analyses (Izquierdo, et. al., I~BSI.
2. Tb! Source-Sink Hadell
Having revil!Hed the various cOflpenent processes 01 bean crap growtn and develop_t, there rellains the tasI: of integrating th!! ea.ponents iota an overall andel of nOH a bean plant functions. The IOst satisfactaryandel available assWlleS that basie processes are integrated by eOlllpetition !I!OIlQ sinks lar Ii.ited resources, and that the relative proportions of resourc!!'i allocated to di I/erent organs depends on Hhat BrIJUller (963) deseribed as "fundional balance". Reduced availability of water or nutrients reduces partitioning to leaf grOHth, but increases partitioning to roots {Figure el. Defoliation results in eKcess root eapacity in relation tu leal area (too high a root tu $hoot ratio), and partitioning HiII favor leal grOHth until the functional balance is

- 18 -
restared. Central to the lIOdel 15 the aSSUlIptiOll that resources, usually carbohydnte or nitrog!!!1, are li.iting. lbóever, thls prHIse Is not universally accepted, and the doobts _it dlscU5sion.
'.0
i i (Ul
J
o ~----------- _______________ ~.
Fíg. e Illustratioo of 'functiooal balance" (Broower, 1%1' betwW! relative partitíooíng oi grawth in lea,es Of roots as .Heded by soi! fertil!ty Of water avaílabiblity.
Inhibitlan al photos)~thesis due to ínsulllcient sínt strength 15 OIIe argu.ent. II this condítiOll occurs, then it loUows that resources are not U.íUng. Evídence lar the eflect C0fl(!5
prilarily IrOl eKperílental lanipulatiOlls creatíng abrupt changes io source-sint relations le.g. throogh depoddingl, 50 Nhile sint lilitatian al photosynthetic mar occur, OIIe fiight argue that it is not a llajor fadar under IIOSt ¡¡eld conditions. lo appostion, up to 35 9 .-2 al unrelObilized total oonstructural carbohydrates were lound io stees 01 Type 11 beaos in ~ulture with a norlal plant stand at physlological eaturity IIzquierdo, 1991). This could indicate lack ¡¡f use ¡¡f available reserve resources due to 51all sint. size and/ar to delayed senescence .,Uh relatively high fE'.
Another lioe 01 criticis. COleS ¡rOl wart. primarily ef laIbers (1979, 1982, and 19941 on cyanide resistan! respiration. This respiration utilizes pathways independent al toose _tioned previously, and SeetlS to produce no products considered essential ta growing craps. Indeed, its principal e/lect. are a depletion 01 carbohydrate and g!!!1eratian 01 hea!. The process appears Midespread in plaot species, and occurs in beans IAzean-Bieta and laobers, 1993). As Lalbers 119S3) has argued, such apparently 'Haste" respiratory activity is incOIIsistent with any theory asslI5iog that resources are 11.ltiog. At the eoaent, perhaps the best argUlents agaiost this ctiticisa are that lUdel. assU/lIing Ii.ited resources are raart.ably good a! predicting trap growth le.g. SOYIiRO by IiIHerson et ~, 1983), and tha! research an cyanide reslstant respiration ís sufficiently neH and the techniques ¡nvolved are difficult enough ihat Its lunction lay relaio to be discovered.
A third líoe 01 critie!SI stefiS ¡rOl fiodings that roots can release large aaounts of carbohydrates and .. ino acids into the soil where they sti.ulate .¡crobial activity fRussell, 1977). under conditions al nutrient stress or in the presence al soil pathogens, 5uch activity light
benefit !he plant via sti.ulation 01 benelicial arganis.! such as .ycorrhizal fungi or bacteria which inhibit palhogens. Hawever I candi tíans ligllt occur Nhere such secretions IIOUld serve no lunction.
J

- 19 -
m. Vla.o POTOOIAl
A. Inlroduction
/Ia.ilU. yields for bush beans are on the arder of 5 lo" ton ha-' (Table 101. !kcasional report. of higher yields are found, but are usually attributable to slall plot sizes with slrong border eflects. Clillbíng beaos on trellíses have yielded B tons ha-' (Fanju! et al., 1'Ill2I, but cO!1lParisoos Hito bu.h beans are af li.iled aeaníng since trellíses eli.inate lodging while reducing the noed for allocalion of assi.il.te to tissues lar ~hanical suppart. Allhough yields 01 5 to b ton ha-' are !eprass!ve 1000er 10 tlees the lean yield in latin Aeerlcal, they are 50 lnlrequently oblaieed lhat considerable research has gane iolo fioding Hays to maxieize yields io beans.
rabIe 10. MaxilllM bush be_o experimental yields reported Ira. the Bean Inleroational Yield and Adaptalion Nurseries, 1976-1984.
Crap Year locatian Variely Vield duratian
kg h.-' days-'
1976 Santiago, Chile Puebla 158 3705 113 1977 liraneros, Chi le Pinto Dorado 5500 94 1978 liraneros, Chile ICA PUao 4400 95 1979 Pal.ira, Colombia OOTB5 4bOO 75 1980 liraneros, Chile Jamapa 4400 19BI liraneros, Chile OOT58 5100 101 1982 liraneros, Chile Jamilpa 4900 90 1983 liraneros, Chile 1AN 109 5OOf) 9'3 1984 Popayan, ColOlbia ZAV 83% 5100 108
Nork DO 'yleld potential', as it lo terled, generally asou.es th.t far a given clilatic regl.e, 50il fertility and water availability are managed to provide good grawing condítions, aAd th.t eHect. of pests and di5eases are _initllal, if not cO!1lPlelely absent. Theoretícally, agronOlilc praclices should be ·opti.al", but sioc!! this .ighl inelude use of CO. fertilization, suppleeentary radialioo, or arti Udal supparts, east researchers have studied yield potefltial in lhe conteKt of e~isting cropping Systel5, either lar bush or climbing beans. He follaw thi. precedent, eephasizing bush beans in ~Q(ullure, but not assu.iog li.itations DO practices such as plant deflsity, raw spacing, irrigation, and fertilizer applications.
l. Estilates of Yield Potential!
As eentioned above, IaKi_ rields lar bean5 \Tibie (1») provide ORe estiNte of yield potential in beans. Yields of over 5 ton ha-' fra. lllVAN's in Chile iltustr.le the ieportance of considering production enviranment. The liran..-os site has fertile soil, high radiatian t1ittle cloud cover, and daylight of up to 15 hGurs at its '34"$ latitude), cool nights (109C to 139C linilUll, and little disease or pest pressure. Photoperiod elfeets .ay provide additional benefits, sinee long day. delay flowering and .aturity. While high yields can be obtained in beans, one should not e.peet tnat a tropical .ite Hith langer, Hareer nights, lower radiatian, and a arriad of other problees Hill yield OI'er 6 ton ha-'.

- 20 -
Coosidering eixillltlll yields of atller legume traps Hable !1), bean yields are si.ilar on a per day basís. Data lar noo-Iegume erops suggest a cansiderabli! 'yield gap" Hable 11).
Various indirect estiaates of bean yield potential are avaílable. AsSUling that greater Icdging resistanc!! and delayed aaturity are obtainable, yíelds af clilbiog beans Might approYi.ate th~ patential of buso beans. FIOHefing and aaturíty have been delayed in I'llrrillo Sintetico throogh use of photapericd ¡¡Hech provided by a gradient af artificial light giving a lB hour photapedcd. \'ields inerened 4S kg ha-' lar each day of delay in .aturity nable 121.
studies of heterosis effects aight provide another sauree of estiaates. Gutierrez and Sin~1 (1985) obtained Fl yields 2a7. to 47% over percent the .id-parent valae, but no eross yielded signilicantly better toan the best parent.
Table !l. Estimates of yield potential 01 grain legUBe5 and other craps.
Crop Crap Yield duratioo Reference
(kg/ha) (days)
legWll!l5 Chiclepea 401:00 Sa"ena, 1'1S4 Cowpea 42~>Q Mien and Su;mer{ield, 1984 6.-oundnut 59f.lQ Ashley, 1'1S4 Pigeonpea 4500 1lf.> Sheldr.ke, 1994 SOl'bean S6i>Q Shibles et !le, 1975
Noo-leguaes Caro 12í)¡)(} 132 Hiltharpe and Hoorby, 1979 Rice 12í)¡)(} 191) Miltharpe and Hoorby, 1979 SorghulI 7300 Miller and Kebede, 1984 lIheal l4loo Evaos et !le, 1915 Patato 19fJoo Miltharpe and Hoorby, 1919 (luber dry wU Sugar beat rolO ,)00 Miltharpe and Hoorby, 1979
Cruder esti •• tes 01 yield potential come frae sludies al Ilawer and pnd abscíssien. For a crap producing 251) Pod jI-', eaeh pod cootalnlng 1.2 9 of seed (b seeds weighting 20') II!J ea.[h). a :rol increase in pcd sel with no change in yield pcd-' ftlUst, by simple algebra, give a yield increase of 207., IrOll! 3 to 3.b ten ha-'. Coosidering that f10wer and pcd abartian rahs are frequently 00 the arder of Yl7. to ~)7., yield potential would be around 5.4 tan ha-'. Further detail. and data 00
estimate. 01 yield patenlial coosidering reprnductive abcissioo are discussed under the tapie al increasiog slnl: strength as a strategy to iocrease seed rield.
Befare reviewing strategles lar yield pütential, the role of agronoaic lanagemeot ~its aHention. Althoogh proposals usual! y asSUIH! 'near aptiaal' agroooaic coodi tiOl1S, ooe lIay questioo wIlether present agroooaic pradices truly approach tnase needed to obhin i1ay.illltlll yield. If f~lJr indapendent factars le.g. planting density, Ifilter lIIanageaent, lertilizer applicatiOl1s and soíl coapactionl are at 95! af their optllUB, this resul!s in a 191 yield reduction (100(1 - .~')),
enough ta reduce a hypothetical patential 01 b ton ha-' to 4.6 ton ha-'.

rabie 12. Yield 01 ev. Porrillo Sintetico under lonr photoperiod regi.es provided by a gradient of lighl intensity given an lB hour photoperiod. CIAT-Palaíra.
Ilislancf! Days to Seed Total ffOil laaps flower .atority yield bíoaass
11 ------tg ha-·------
3 51 'i5 41Qi) 7Qi)() 23 9 43 94 3500 5600 lb
15 3b 71 3Qi)() 4700 12 21 3b 69 2!lOO 4100 11
e. Yield f'otential Vi AgronOllic llanageeenl
8efore revil!lling stralegies for yield potential, toe role 01 ,¡gronDllic .anayeeent ~i ts altention. Although propasa!s u!iUally aS5_ "near opUlal' agrooDllic cendi tiOO5, one .ay question Hhelher present agrooOlic practices trul V approach thase needed to oblain lilXi_ yield. If fonr independent factor! le.y. planting density. water aanageeent, lertilizer app!icaUons and $Oil compactíon) are at 951 01 their optieu_, this results in a 19% yield reduction (10011 .954 )),
enough to reduce a hypothetical potential of lo too 0'-' to 4.t. ton ha-l.
r. Physíological 5trategies lor Increasing Yield
In the d!scussioos that 1011011, specific strategies lar lnereasing yield potenlial are reviewed, starting with strategies lor lnereasing sonrce and sink capacities. As a cooclusion, toe authors propase an ideotype lar _illlll yield. The problellS 01 earliness and seed _ize, and lar SOl!!
uneonventional strategles are also discussed.
Many strategies are based on the assumptioo that increasiny yield potential requires an ¡ncrease in SODree size. Support coees Iroe the observation that in cOlparisons of varieties, greater yield ii clasely associated Hilh greater growth, Hhether aeasored as erop dry weight, leal area duration, or other size par_ters Hable 131. Reductions of yield under shading IEscalante, !'IIlO) aOO yield iocreases with COz fertilization llaing, el al., 19841 also suppart tbe assulPtion that source size li.ils yield.
Tne silPlest strategy lor increasing source strength is siaply to increase lhe length of the growth eVele. In varietal trlals Managed lar high yields in tropical areas, yield Is usually carrelated with tlae to .atority Hable 13).

- 22 -
Table 13. Correlations of yield with harveit indeK, biOliSS, days to uturity and l!!af ar~a duration frllil variws studies.
leal HarVl!st Days to area
Trial indeK Billila5s utur. durat. Solrce
10 lines Habits I and 2 .51 .9Ut .75U .!IIlU l«1ite, 19S1
lB Unes Habíts 1,2, and 3 .46U .1I7U .4!U • 42ft l«1ite, 19111
Porrillo Sintetico 12 ekperilll!f1ts • 211 .9bU • lIS .. .B7U laing et fu. 19114
9 species of graln leguES .50 .91U .B6U .9JU laing et fu. 19114
length of growtb cycle
Table 14. Seed yields of 11 Unes 9rDWn Ill1der 3 daylengths. with and Hithout trellises. Pallira, 19B5b.
Üil tivar
JU 71t"'2 6 17648 BAr 450 JU 80-'3 llIJ! 49 XPll 112 ¡¡ 29S9 6 3807
JU 7ft"2 6 17MB BAr 450 JU SO-'3 llIJ! 49 XPll 112 G 2059 G 3807
o a y len 9 t h Ihrl 12.~ 13.5 14.5
-----------------kg ha-'-----------------
r«l TI1ElUSES 21l1S 19lIS 1692 1550 1236 873 2338 2439 2413 Z04Ó 2225 2154 2m 23'1B 2049 20lB 1bB6 2041 2333 2301 217tl 1590 11lIS 1m
2052 200B 1m
111m TRELllSES 2694 3200 3168 1992 2913 15Sb 2324 25911 2190 2073 2121 2444 23Q4 2lBI 21141 2043 2106 2446 1912 2900 253B 1731 1562 2144
21!5 2400 2416

- 23 -
lbe previously .entioned wort Hith photeperiod treataents to delay eaturíty deeonstrated this as did a yíeld tríal af 6 lines grllll1 under 3 philteperiod regille5 (Jable 141. ¡¡allac!! !I'1!I5I, reparted that at higher latitudes, yield is negatively carrelated Hitn ¡atenes., apparently becan!! late .aterials .ature poarly as a result af grOllth U.itaUan. Hlth cooler tl!llflleratures. Although there has been considerable work done on genetic cootrol af tillli! te f10wering and .aturity Ufallace, 1'1!I5I, and there have been effarts tD select far late .aturing Iines, expected yield increases have not been obtained. A ujar problH has been that selectioo far lateness resulted in selectioo af .aterials which are late because of poar photeperiod-tl!llfllerature adaptation, not greater vigor ar soorce capacity.
Photosynthesis
Another strategy far increasing soorce capacity i. to inerease the photosynthetic capadty af the trep. As in II05t treps, consistent positive carrelations between supposed laxi_ photosynthetíc rates and yleld have not been found {KuenHan et al., 19791. lbis failure probably results fro. interactions of factars detereing the efficiency of photosynthesis at th!! crap level. Ca.parisons af phot!l5ynthetic rates in 20 Iines of IIUflg bean {Chandra, et fu, 1'1!I5' supported thi. assUllption. ilhile .axieua rates were not eorrelaled "ith yield, rates during podfilling {which were ooly 50 to /1)% af aaxílUa rates) liad a corralatian af 0.66 Hitl! yield (signifieant at tne 0.01 probabilíty level'. Siven that photosynlhesis rates during pod filling usually decline rapidly due to nitrogen re.ohilization frae leaves to peds, tne IOSt critical proble. relating photosynthesis to yield potential .ay be oítrogen partitioning during reproduetive growth, as diseussed latero
Respiration
Reducing respíratian ilplies batl! a direct ioerease in avaílabilíty of assiMilate, and a possible indirect inerease througn higher aptilMl LAI. AssU/llÍng that growth respiraticn depends an plant cOlIpositioo, selection far decreased growth respiratioo _Id prooably prove difficult, and .ight resul t in lower proteio content in seeds. 5election far IOH iIIaintenance respiration appears lare proeising. Nork in perennial ryegras5 and feseU!! (Nilson and Jones, 1'IIl2; Volenec, et al., 1'184) have resulted in suece.stul identifieation of lioes Hith reduced respiration in Hara teeperatures, and these differences were issociated "ith increased yields. Gifford and Evans (1'181), in a review al relations a.ong phatosynthesis and partitioning, toncluded that such results represent the only known cases where selection lar a photosynthetic ar respiratary characteristic were successlully related to yield. However, to date, researen on respiratioo C!l5ts has received .iniul attention as cOtipared to phot05yntnesis.
In beaos, as in other treps, considerable elfor! has been e~pended io identifying lines coebining redueed photarespiration and greater yield, ¡Wallace, et !l., 1976). Results have been negativa, supportíog th!! tonclusion tl!at photorespíration is an unavoídable consequence 01 increased o. concentration. in the presence oi RBl'-carboxylase (I'OIIles and IIYond, 1979; Tolbert, 1'1801.
light utíli,ation
Sine!! crap photosynthetic rates depend on light interception patterns, yield potential should vary "ith canapy structure. lbe relatively horizontal leal orientation detected in studies of light interception in bean canapies ilply losi 01 elliciency due to e~cessive íllullination 01 upper leaves and shading 01 lower ones. As in aany other (reps, ideotypes lar beans aften suggest selecting lar lare erect lea! arientatlon. Nien and Wallace (1973) and Sato and Ilütoh (\979 and 1'183) detected varietal differences in leaflet llOVelM!l1ts, but the resalt. were nnt e~tended ta effects on canapy strncture.
5i~le ge_trlc argu_ts suggest th.t far a 91 ven LAI, both sllaller leaves and greater vertical separation of leaves Nill result in baltet penetration of light in tne canopy (lOOMis and Nillia.s,

19611. Reducing leaf s¡ze should not prove proble&itic, but the Principie of Sieililude (rno.pson, 19421 dictates that the taller the plant, an increasingly great aaoont of rescurees 8!.Ist be invesled in tissues Hith support functions (e.g. in slees and petioles). Une coneludes that far a given set 01 groHth conditions, there is an optilllal bean plant heighl. /lUantitatíve esti.ates 01 opU .. are lacting, but in cereal. redueed plant height has been associated "ilh higher yíe!ds IChandler, 1967).
Lodgillll i5 cOllllllonly seen in bush beaos, <lOO often reduces plant grONth severly ¡Fig. 9). 50me of the d~ge i5 usually attributable to mechanical da.age and subsequent disease ¡nfections, but los; of plant height reduces efficiency of light interception. Thus, lodging resistance i5 anather Mechani .. lar pholosynlhetic efficiency. Allhougo, use of trellises can reduce plant partitiül1iog far 5uppart tissu!!s, freeing additional assi.ilat!! for pod grOHtn, it Íi instruet!v!! to cül1sider data far yields with 3.d without artificial supports in rel.tian to ath~ ERperiMefltal .. nipulations. Delayad maturíty through photoperiod e~tensioo resulted in incre.sed yields woen trellises Here
pro'iided, but the yield elfect HU not detected in plets withoot support. Stofella et fu 11979) reported uphelogieal variatioo in roots stroogly aS50ciated wito lodging resistance and seed yield in black beao lines, indicating that a larger root sil!! may reduce lodgi.g aOO increasE seed yield.
210 Check with .,dfkt.t IUf:lpoft -- ---- -~-
250
230
Fig. 9 Effect 01 artificial lodging at dillerent dates frQM flowering on seed yield of c •• Porrillo Sintetico. lodging produced by rolli09 a bamboo pole OYer the crop. CIAr-Pal.ira, 1975.
Higher .. xi~ LAl is nat necessarily beneficial sinee greater leal area alone result! in greater respiratory casts and a reductioo in light intensi ti es in the lower eanopy. HOHever, if respiration is reduced ar elficiency 01 ligot interception iaproved, greater aaxi~ LAl's would be benelicial.
Increasing LAD is desirahle íf acnieved without eKceding optilal LAI, but achieving higher LAD's seeas diflíeult. AgronOlíc practices lavoríng rapid canopy establishMent (narrower rON spacings and higher plant populations) migot help. Anather possíbility is to reduce the rate of leaf senescence during pod fillíog. Supposedly non-senescing bean línes are known, but their yields are ION enougb that this charaeterislic was probibly achieved through redueed reproductive growth.

- 25 -
2. Sink li.itatioos:
Although rost stOOie> on the relative illportanee 01 500fce and sink strl!!lgtbs io deteflioing yield potential conclude tbat source li.itations are eore Íeportant, possible sink li.itatioos, IOsl ootably 'excess' Hower abartioo, are uf ten reported. Furlhermre, siok inhibiUon 01 photosyntbesis would aake source slrenglb appear lililing Nhlle actually resulling frOl iosu/fleient siok strengtb. Possible causes uf ION sink strength aay be elassed as either insuf~icient numbers uf sinks ar ION
strellgth uf indi ,idual sioks.
n ower, 5eed and poo abarti on
Fruit setting and pod retentioo procedes acropelally in legues. Abscissioo of Howers or SIlIall poos oceurs 00 pradically !!very raceae 01 the plant. There is evidellee tbat the basal and older pOOs regulat!! lhe absdssioo uf "ew flOH!!!"s and sllilll pOOs in several species af grain leguse. Tbis has beeo subslantiated in cowpea {Ojehoaoo, 19721, lupín (Van Stevenincl:, 1~57!, liaa bean (Cardner, 19JJI, faba bean (Jaquiery aod ~.eller, 1'1001, soybeao IMiebold et !le, 1981), pigeon pea (Ariyanayagan, 1'1001, and dry bean (Subhadrabandhu, 1976 and Ta •• s, 19761.
The abscíssioo uf reprOOuctive organs during fI_ring and,fruiting periOO in tb!! graio l!!gule> is often greater than 501. Binkley (19321 showed shedding 01 flowers uf .iM garden bean up to 76% while a range ol 42Z to 54! ot reproductive abscission on oine genotypes coroprisiog four differing plant architectures were found (Izquierdo, 1981_). roo drop accounted far up to B2Z 01 lb!! total reprOOuctive structures ab;cised, with tne shedding ot seall pOOs less ihan 10 lA in Jenglh accounting lar IOst of lhi. (Izquierda, 1981b).
Several lioes uf evidence suggest ihat if the shedding of reproductive parts io beans could be prevented or decreased, the yie!ds eight be increased {Subhadrabanhu et al., 1978 and Tuder el al., [9751. A be_o riela potent!al up to 6200 tg ha- t has been estieated iE the yield 1055 due to abscission was aaded to the actual yield (Table 151. Falterns and rates of flower and pOO shedding significantly differed alOng genotypes Hilh contrasting plant architecture and growth habil (Figure 10). Since pOOs absched al sucn an early stage ot developJll!!l11 ooe lIighl suggest tha! the lack ot fertilization is aainly responsible lar abscissian bul evidence is contradictory. Va;ez and coworl:ers (19841 fcund tbat flower buds !lith a bigh probabilíty (lf absdssing shOHed a lIid!! range of abooraalities iocluding large nlltlbers 01 ovules with necrosis and 151 uf buds lading the eabr¡'O saCo
NOs! researchers have assUlled that self pollinatioo procedes at a high enough rate in beans that pollinatian is not a limlting factor, but the fatt th.t pollination does occur in the absenee uf pollinatar! does ool establish that it is 01 suffieient eHiciency to IlaXÍ.ize yiela potential. Artificial tripping ot bean ¡Iowers, which ,¡.ies the effect of bee visits, nave ioereased seed set per pod and nuooer 01 pOOs per inflorescence, but eflect varies wilb genotype and seasoo (Table Ibl. Tbis variation prohibir exploios Nhy artUida! ... nipulatioos resulted io i'ield iocreases in SOIIe,
but not al! Jines lIhen flcwers opening io the ¡irst 7 days alter flOlll!fing were artificially tripped (Table 171. Unlortunately, these results stíll do not e5tablish that pollioation li.its yield polentia!. Inefficiency 01 pollination .ay be a character 01 lioes Nlth ION yield pütential, while the highest yielding .. terials already acnieve sulficient pollinatían.
Hdterlein et !le lI'IOOl stated that ooly ene lertilized ovule Nas needed to preven! pOO drop. Pechan and 1leb5ter (19!l61 also louod that abscissioo occurred pri_ily in fertllísed flONI!rs uf Red Kidney beans. And si.ilar results hay!'! been found in soybean (Aberna!hy et al., I'm; lIiedbold et ~. 19I11l.

- 26 -
Table 15. Estiaated losses in yield due to abscission, actual rields yield potential al nine dry bean entries coeprising four architectural laras, East Lansing, 1981.
IIrchitectur al {arll and entry
Estiaated yield loss due to absci ssi 00" Actual yield Yield potential V
---------------------t.g ha-·----------------------
ArcMtYlle m!!O 613Sb b1619
Clas,ic ¡¡ BTS Ne¡l-2
Stlall bush Seafarer Sanilac Tuscola
Erad sulI bush (-14
2661 3Ul5 3189
29114 2798
3496 3443 3922
3027
3577 a' 3076 ab 2'149 b
2SIl'I bcd 2!!45 b
2569 bcd ¡S23 e 2162 de
2366 cd
" {reproductive abstision/a21 • {seed/podl • {seed weightl.
6239 6262 6!37
54'f4 5644
5iJbb 5266 4984
53'14
y Yield potential = Estiaated rield los5 due to abscissloo ~ actual rield • • Mean separation in colulll/1s by Durn:an's range test, 5%,
mee: Izquierdo, 1991.
Perhaps lhe best way to raeneile these different reports is to assulte that causes of abscission vary greatly with growing conditions and cultivar. Abscission eay result froe water stress or fratl CIlIIPetition aeong de~-eloping pods far nitragen, atber nutrients and carbooydrates. I'Ilother cause of abscissioo is probably related to horllOOal regulatioo of abscissioo of younger d!!Veloping struelures in a raceE, lalnIy by the oIder frult. Regulatory control also coold be exerted throogh horllO!1l! availability, horeone balance or har_e inhibition of a¡¡si.ilate lIObiliz,¡tion to developing ¡eeds. DiHerences between bean cultivars far ethylene production by reproductive strudures has been reported (Izquierdo and Hosfield, 19!!O1 and related to leveIs al reproductive abscissioo. EI-geltagy and Hall (1975) suggested ethylene as the cause of abseisslon in bread beans, and spací/ie response to ethylene by target eells located at pod peduncles has been descríbed ¡Usborne, 1~77 and Nebster, 19751.

! • r e
1 i
1 .. 1 ~
o
Il
•
•
A
1
h......l·
7 " •
1
O Sm.n bush
• Clas'lrClI
.... Arehilyptl
IlSMS
• PhyslologicaJ mlltlKity
, '7 22 2' 32 37
F\ I \ I \ I \ I \ .11/1\ I b--fV\ \
I / \ \ . \ I ¡;;::
"..._.d Ii .\
~. <::¡. ~--'---'~. --'---'---<---'.'--'---'
11 H 21 11 32 31 42 41 62
Fig. 11) Patteras and lean rates 01 reproductive abscission IFlOHerS:i a.nd pods:bl 01 dry bean genotypes "íth differing plant architecture a.nd growth habito Seall bush: cvs. 'Seafarer', 'Sanílac' and 'Tuscola' (navíes, growth habit 1)1 classic 11: cvs. 'Black rurtle Soup' and 'Hep-I' (grOlith habit llIi a.nd architypes (erect, reduced branching ana narrOli profil!! canopy!: breeding lines 61380, 61lSb, and 61618 (grOHth habit 11). SourC!!: Izquierdo, 1981.
Table 16. [¡fect 01 artificial trípping of bean flOHers on pod and seed set in ten bean cultivar!. f'al.ira, 1984.
___ Pods/lnflorescence Seeds/Pod -Control Tripped Control Tripped
A 120 1.30 0.75 4.2 4.4 A 200 1.31 1.22 4.9 4.b 11 227 0.84 0.93 3.1 4.1 A 429 0.89 1.68 4.5 Ul A 463 0.32 0.55 2.S 6.4 A 488 0.22 0.41 5.0 5.9 BAl 202 0.14 0.37 2.9 3.8 BAr lOól 1.42 1.70 5.9 5.9 BAr 1291 0.96 D.n 5.0 5.0 BAr 1670 0.98 0.87 2.9 2.5 l1ean 0.84 0.94 4.1 4.7

- 28 -
Table 17. EHect of artUidal tríppíng of bean fl!lWl!f5 00 !íeed yiela four cultlvars. Falllra, 1985.
Cul tivar
8I\T 477 8I\T 1491 Caliu ¡¡ 1540
F Treatllll1.'!\t
__ Vield __ Control Tripped
-----ton na-'-----
2.43 2.45 2.51 2.99 1.36 1.57 1.4Q 1.39
7.Bt
Cultivar K Trt. 2 .. 9
• Significant at p = 0.5 level
_!ieeds Pod-'_ Fods .-. Control Tripped Control Tripped
5.4 5.4 210 2SO 5.3 6.2 200 2bO 2.0 2.0 95 JQJ 4 .. 2 4.1 Ib4 191
0.9
1 .. 2 Q.S
Ilnother explanatioo lar poor poli set <lS5UleS that the other organS ar older pods are competing sueces.lullr Hith the IlCHef5 and s.all pods lar assi.ilate. Wiebold (19Il1) speculated that iflere.sed abscission in canopy regions receíving IOH irradiance .Igh! be a consequence 01 non-availability of p1wtm;ynthates in soybean rae_s. localized cOiljletition allong older ana yaunger developing pods, at eaeh bean 'phytO&efic unit' cauld caUSE a locallzed abscissian. Carbohydrate availabllity and leve! Here studied .5 regulatary ayents in abscisslon. Subhradrabandhu (1<176) reported that carbon dioKide enrichment and long day. during tne reproductive phas!! decreased poli abscis.ion due to a high accuou!atian 01 earbohydrates in the stemi 01 tne ev. 'Blac~ Turtl!! Soup'. Higher [K[-starch scare values have been associated wlth hlgh!!r poli retentían and yleld 01 (v •• Nep-2 and Black Turtle Saup tnan the navy ev. 'Tusco!a' Uzqulerdo et al., 19801.
lo the sale líoe, a eodel based on vascular liaitatians has been postulated lor pigeon peas (Rawson and Canstable, 1981). Thís eKplanation assuaes that the carbon-assi.ilates can ool be IOVed
to the flOHers and slall pods rapldly enough becaus! the vascular systee 01 the pedicel is not fully operatianal. A!tilary 1l0000S llith short pedicles and a large eros. secUan Hith large leaves doS!! to the sinl could Increase the potential yield by reduclng the resistanc!! to the IOVE'f!ent 01 assi.llates. In wheat, phloel capacity appeared to be quite plastic, different species varying IQ-Iold in cross-sectional area 01 phloel. The variatíon Nas associated Hitn sink strength, thus leading to the conclusion !hat translocation HaS prohably not ¡i.iting (Evans et !le, 1970). JIOHever. leasureaents of translocatían rat!!s are difficul t to lIi~e, and toe problee oas not been resol ved IEvans, 1975).
Elphasis is usually glven to losses In sink strength due to 1l0Her ¡nd poli abstission, but abartion of individual ovules or developing embryoes HOUld ¡Iso reduce sink strength. Unfartunately, data en elbryo abortion is rare since deterainatíon of ovule nullbers requires tedious dissectiens of individual ovaries. Table 18 clllpares nWlber of seeds per poli at harvest, <lnd the IliKi_ oUllber al seeds per pod launa in the harvest saaple. Assu.ing ovule nullber indicates potential seed nullber, a 20% or lOfe los5 in yield potentíal (ould be ascribed to eebryo abortien. Dut agaio, it reaaios tohe dl'fllOllstrated Hhetner surh abortioo reflects defech in developeent, ar is sieply due to insuHicient assleilate.

- 29 -
Table lB. Ellbrtll aborUoo rute, lean and wi_ rnBIIber of S!!!!ds p!!r
lean rnBIIb!!r of ovules p!!r pod and sll!!d lII!ight in 5 !ines IfrOl Nhít~, 19811.
Úlbryol ab"rtioo I'ti!an No. IladlllUl No. I'ti!an Sel!d
Une rate sIl!!ds/pod seeds/pOO ovules!pod Keight
II!I
G 122 441 3.5 6 6.3 2'10 6 1540 421 J.l 5.S 5.3 270 G 6729 4tl 2.3 4.8 3.9 440 64495 m 5.9 9.5 8.1 ISO Il 4459 464 4.0 7.3 7.4 120
• Calculated u 100. (1 - sll!!ds per pOO/ovules per podl
POO growth rates
.!IOO
y"" ·400 ..... 57"X • • Al -0.43
• 1000 •
~ • • • ..
'" I '500 ,.
• '000
• 30 .2
fig. 11 Relatioo betlll!en sll!!d yield and duration of pOO filling in 20 bean lines. rlAT-Palaira, 19!14.

- 30 -
In addítioo to elfect; 00 nwnber 01 ;ínks, loo strength o/ índividu.! s¡n~s ligot límit yield. ti thís _e tfUE, pod gr!lWth fate migot be correlated wíth yíeld. For 19 bean culthars, m~>itl'}~
pod growth rate was only slíghtly correlated witn yíeld (Hg. !l!. As díscussed in the introduction to the sectien en rieló putenti_l the results ooly demoo5trate a reliltioo, nat caUSE and eflect. Ffrj
or seed growth rates and duratíon mal' rellect availabíliti' 01 assímilate as mueh as sin .. strength. However, íf tní; "ere the case , treatments whíeh alter the ;ource-sín!: balance, such as depodding and de/olíation, ;h!lUld alter pod gro"tb rates. Pod grol/th patterns under such treatments are ver)' símilar as mmpared to dUferencEs amang cultivars (figure li, supporting lhe possíhility 01 sinl: strength e/lects. When the pod elongatian fate o/ 25 línes Híthin an internatic~al replicited trial lar dr~Jght tolerance (BIVYT-erAT) was ~asured, both onder post-floweríng unífrigated ¡stress; and irrigated canditioos, a reducUoo up to 177. in the linear elongaUoo cate ... observed m.9b irrígated and 7.47 ffiJlI day-' under stress) Ilzquierdo and Huepe, 19Bb, unpublished).
lf siok strength ís límítíng, one wDuld expect a relatíve!y large pnjjlortíoo 01 crap W€ight at .,turHy as vegetative gro"th insteaá al seed yield. To Evaluate this possibilíty, a "harvest inde"' ls usually calculated as:
HI = Seed rie1d!Biological rieló,
where biolngical yield is símply total CfOp drl' ¡¡eightit maturity. l4hiIe í'ieId incre,se; in .,n)' uops have be_n linl.ed to incrpaseo h.rvest inde, iWallace et fu, 1972), diti sh¡¡¡;íng cilffe!atíoos affic"'l1g \'ield and hanest inde, must be iudged with raution if nat complete sleptícism. Sinee harl'Est inde, ís calculated in p.rt Iroo yíeld data, dOy cüfrelatioo .. ¡ll ¡ndude this MUfaC! and be artifícially ínflated (Charles-Eswards, 1982). Simply using paírs (jI nndc;¡¡ numb.rs to calcul.te .,tíhe;a! values (JI harvest inde", one can generate high!y sigoíliont cDrre13tions. Ca!cu!ating ,alves representatí;e lar beans, a mean harvest inde, 01 0.55 and mean "rielo;" 01 2.0 ton; h.-' general,..] Hith r.ndam numbers, 20 paífs DI data gal'e a correhtion 01 r 2 = 0.77 (Figure 12).
01 • • •
• • • • • 0' . • ; •••• ¡
• • • ~ • , .. o. • • y",. o 323 ... O.(X)()t29:X • •
M +-----------.,-----------,-'000 2000
Hg. 12 Artlllcial correlatioo DI yield and harvest índex calcuJated 1t0lft pairs 01 r.ndOOl ""mlJers.
Adding te the problem al inlerpreting harvest índex value. in be."s is that varietal differences in deloliatioo dnó .tero senescence during maturatíon 01 be •• s !lale it dilficult to decide which tissues to ¡nelude in a saople. Oi//ícullíes are a!so enc!lUOtered in decíding when to s_opIe, ana nOll to manage the saople subsequently. fldrvesting at a single date will ínflate values 01 eMIy

- 31 -
uturing Unes. I'Enitting salflles to air dry after lIarves! .igllt f!!Slllt in reduced diHerences since respiration can continue to cause Neigllt IOS5.
Jo cope witll soas li.ltatlon! due to cultivar differences in leaf abscission at laturity a aodified harvest inde~ (Kobasfli-Shibala and da Losta, 1'190) nas been proposed as: IIflI = seed yield/total cumulative biological yield at physiological eaturity + dry Neight of abscis.ed ergans IflONers, pods and leavesl. The collection receptable for field abscission studies described by Izquierdo and Hosfield (1982) is Nell suited for obtaining the dry Neigllt of absc!ssed cOIponents. Fi9. 13 shows a highly significan! correlation betNeen the eHI and final yield, altbough this resalt is ¡Iso an artifaet.
I1asaya {personal c_i(aUonl cOIfIared harvest index to other par_ters for individual plan! selection in F2 and ts populations, and found that it was essentially useless (Table 191. More research j5 needed to eVilaate IIflI as a tonl to overcOlle the li.Hatians found with the cooventiond harvest ind~.
,a
0.3 o. o .•
Fig. 13 ncdified harvest index relatíonship to yield for 9 dry bean cu!tívars Híth díllering plant architecture m: architype; o: da.sic 11; o: seall bushl and o: tal! ered bush). SourC!!: Izquierdo, 19B1a.

32 -
Tilble 19. Average seed yield. of f. progenies deri.ed frOll individual plan! .elections in the f. usíng variaus cOlbinations of selection criteria: HI = harvest indexl UN = plant dry weight DY = daily yield. NUmbers in parenthe~ indicate F. plant. in a given selection category. Masava, persooal cllfIIIUnicatíon.
Cross
11 429 x XAN 112
A 429 x f'ata de lOfle
A 429 ~
Pecho Altarill o
3. As.iailate Partitioning:
___ ....:Selectíon_critería __ _
Parental Vields 1 fe~ale Mal e HI
2 UN
Criterion 3 4 _4 plu._
YD ~~ 1+2 1+3 2+3
-------------------------g plot-·----------------------
268 256 266 312 283 270 287 (20) (lO) í3l (62) (n)
269 259 258 286 269 25B JI7 (lb) (9) mI (2) l4l
269 2B5 253 253 263 282 (21) (91 0281 (9)
Rather than ass~ing that specific characteri stics of the SDUrce or sink li.i t yield, an alternate hypothesis is that it is the partitioníng proces5 whích is defectíve. Adaes 11973, 1975, and 19911 has proposed a series of archítectural aodels eophasízing the need for cultivars .oich are efficient in mobilizing both dry .atter and H to the seed. If a cultivar arcUMUlates assi.ilate aud nitragen during .egetative growth, and is ilble lo refllObilíze it during podHlling, this shoold del ay leaf senescence, and lead lo higher yields. Ca.bined Hith a more erect plant type to reduce lodgíng, such a plant-type snoold perftit much greater efficiency during the podfílling periodo
Hodifieation in plant architecture does not necessaríly ha.e to Jead to increased efficiency in translocation althoogh yield advantages DI the i~roved varieties over old standard cultivars has been associated with ehanges in physíologícal perfor.ante (Gay et th. 19901. The upholagical aodeJ characterized by th. Adam's architype has produced apparently not onJy a llOre desirable bean plant type frOl the agronomit standpoint Ilodging resistancl!, and eas!! of .echanical harv!!st!, but has lead to higher yields due to íncreased physiological efficíency in.olving lilling duration, partitioning ind refllObilization oi assi.ilates (Izquierdo, 19911.
The hypothesís 01 stress tolerance due to rellObílízaUon of resenl!5 i; attracti"l! but has Ji.itatian;. The abílitv to withstand a stress by a.sieiJate remobilization is not necessarily lint.ed to storage capacity. Partítioning and refllObilization capacíty are both needed and IllUst be considered in a breeding prograa. Th!s suggestlan Is conslstent with the lart that bush type entries ha"e been characterized by a .hort bu! rapid filling period, high partition!ng, and hlgh r9Obilhation of dry Mtter. lIrn<ever, the.e entries, as ca.pared to the other types, are susceptible in different degrees to leal area reduction du!! to physical and biological stresses. Once the photosynthetic aechanis! beca.es impaired in bush type plant., seed fllling depends OH retebilization. Evidence that Hater stress enhantes assiellate retebllization (Salper and AdalS,

- 33 -
UI1flublishedl indicate the need for further researdt. Aspects of this problellfill be discussed in a subsequent sectian.
4. Yleld CoIpanents:
A special group of hypotlleses eancerning assi.ilate partitianing are tllose related to charaeteristics sutil as seed weight, seeds per pad ar inflore.cenee, and pod nuaber, which .ay be defined in such a way tllat when aultipled together, tlleir produd is equal to trap yield. Such 'yield tOllpanents', as tlley are terll!!d, lfete fin! used by EilgledllN (1'1251 in .ludies of wheat yields, but analyses of yield tempanents are frequently dane in leguainous traps. For beans, coapanents .ay be defioed as:
Yield = 5W K SP x 1m, HIlere 5W = weight of single seeds SP • seeds per pod 1'11 • pods per node !ti • nades per .2
!hese coaponents also serve to calculate othersl
SP K PN x 111 • SeedS per ... 5W K SP • rield per pod PN K 111 • pods per .. 5W K SP • PN • rield per nadE<
Cempanents .ay be subdlvided to distinguisll between coaponents borne en branches or .alO steas (e.g. Senoett et al., 19771.
The si!!plest strategies for iocreasing yleld throogll selection far yield cOllpanents observe that if tertain cempanen!s are increased wlthoot affectiog tlle others, then yield IUst increase. This lollows ¡rOl silple algebra, and as theory is unilpeachable. lo practice though, .ludies of varlation io tOlpOflents Easured in di fferent genotypes general! y show that far pairs of cempooent5 whose product equals leg. Seed Nelght ~ t.Ullber 01 Seed. per .21, lile correlatioo belween the palr 15 oegative !Table 2Q1. This effect Has teraed 'yleld coaponent cOIIpensatioo' by /Idus 09/,71, and j; attributed to cOII1jlensatory growth as distrlbution 01 SoIe yield lillting resource is affected by genetic and environtental factors. The principal ilp!ication 01 tllis process is that selectíon lor hígh leve!. of ooe caaponent will prohably not result in yield increases.
Table 2Q. Correlatioos between pairs al yield cllIpOOents Iwhose prodtd equal. rield) lor tIfO trials 01 bush beao.. Suurce: Nhite, 1981.
Seed weight vs. Seeds .-z Yie!d pod-' V5. rod5 .-z Yleld node-' vs. Nades .-0
tria! with: 10 culti'ars38 cultivar.
-.BlU -.!lbU -.65 -.SOU -.21 -.77U
',U Significaot at the p • 0.05 and p • 0.01 le,els, respectívely.

- 34 -
fkmever accepting that yield COIIPooent cOIIPensatioo occurs does net "Ii.inate the possibility of identifying an optieal cOlllbinatioo of COllPOI1l!f1tS. Graplls of varieta! dífferences in tIlO cOIIPooents ¡(¡ose prcdud !!IJuals yield (Hg. 14) i!lustrates the overall trend of cOllPl!f1satioo, hut deviatioos ¡rOl th!! curves lar coostant yie!ds soaw that there ís IlUCO variatioo besides that explained by cOllPensatioo. lhus oo!! .igot expect higher yielding lines to represent .eme optimal cOlllbinatioo 01 compooents.
5. Integrated Strategies:
As IIefltioned belare, plant breeders have IIilde no decisive breaHhrough io yields 01 dry heans Nith the exception of developing disease resishnt varieties Hhich aliOlI genetic potential to be !!:<pressed.
lotegrated approaches coosidering plant .orphology as Hell as physiological elliciency have been attelllpted by defining crop ideotypes adapted to specifk enviroolleflts. llooald (!9ó!l1 e.phasized that a plant ideotype must be a pocr competitor. lhis brea~ froe trad!tional thint.ing comes from the understanding that the individual plant with!n a coemunity Hill e'pre;s íts potentíal lar yíeld IOSt {ully if it suHers a .inimum of ínterference or competítion Iroe its neighbars. lhe crap ideotype is e,pected to aake a .inilUl deoand on reSQ!lfCe5 per unit 01 dry matter produced, but the cOlftUnity as a whole must e.tract a ••• imul aeount 01 tbe reSQ!lfces.
Hg. 14
~ 1400
~ = \200 ;; j E 11)00 , z
800
• • 00
.00 010 0.20 0.30 040 .50
Yíeld c01l!jlooent compensatíoo among 38 bean !ines. Curves represent levels of constant seed yield. Saurte: White, 1981.
In beans, characteristics such as deterainate habit, early lIaturity and slall leaves have heen cDnsidered to províde wide genotypíc adaptation (Sllvera, 19141. Ada.s (IV731 suggested a bean ideotype lar monoculture, and further hypothesízed that 0111 y lew coanges needed to be ude to adapt thís ideotype to be a successlul cllillPl'titar Hhen gr_ in lIi.ed culture Hita .aize. He suggested that the leal size be EdiWl to s ... l1, and tha! the sink capacity bl' increased by raí;¡ing the nUlber of f10wers per rac_ and nuaber 01 seeds per podo ~ the physiological side, he suggested it I«JUld he desirable to raíse toe net rate of Co" exchange, to ¡"crease the rate of translocatioo of pnatosynthate to sínk, and to sl'led lar a high narvest índeK.

- 33 -
Tanah 11"'74) suggested ao ¡ndetermínate beao type which _Id produce llaoy pods 00 toe .aio .te. for líxed culture Nith eaíle.
Adaes {l973l eentíooed the isogenic líne, eodel building and factor analysis as three eethods that had been used in coostructioo 01 plaot identtlles, but very IimUed flUIIber of studies have been cOllduded in beaos. Ilenis (1971) utilized a factor aoalysi. 01 bean plaot-type variables. lIith the sale approach, lare than bOt of total variaoce Nas attributable to the factor naeed ·partitiooing" under stress cooditioo. (Saalper and Adallls, 19S5, UIlpublíshed).
These theoretical, IIorpho-physiological plant Iodels have been apprOltieated in breOOing eHorts (Adaes, I98H, aOO illlllrovOO lines hay'!! appeared to shaN greater yíeld in _oculture.
6. A NeH Strategy:
Having revíewed varioos strategies tor increasing y-ield potential, a logical cooclusioo ís to provide a neN strategy. Basic cOIpooents 01 our strategy are nat radically different froa those described far cereal s (e.g. Donald, 1968; Chaodler, 1969), and al50 5hare cOIIIpooents 01 prevíous propasals lar graio leguaes.
0Ur fir.t preeíse is that the strategy IUst be developOO in the context 01 apprapriate agronoeic lanagement. We suggest narrON rOH spacings le.g. 30 ca) at densities 01 40 plants". The lact toat tbese pradices do not give maxi_ yields Nith I!Kisting IIateriáls is irrelevant sinee the ínterest is not in eurrent cultivars, but ratber a new t~~e of bean crop. An abundant nitrogen ~lPply Míll be r~luired. Far a 1 ton ha-' yield inerease appr~il3tely SO to loa tg ha-' 01 additional nitrogen are needed, lSslliting 507. losses ta non-seed tiSSUI!S aod leaehing ar volatilization. Achieviog such levels .ay requíre ;;plit applieations of oitrogen or ;;petial aanageaent to laxilize N-fiKatíon. Recen! work on flood culture of so}beans has produced 4O! ¡nereases in grOHth Qver conventiooal irrigation, suggesting that water eanagi!tllel1t May requíre lurther illjlrovement (Nathansoo et fu, 19841.
A specific morpbology (e.g. deterMinate .tees and na lateral branches) is less illjlartant than crop response to high fertility. As in changes introdueed in to ríee cultivar. (Chandler, l~b7),
plaots HIlith .haN liaited N response of ste. eloogatioo aod leaf expaosioo are needed. This response will prohibly be carrelatoo Nitb rOOucOO braoch grOHth aod sllaller Mxi_ leal .he. Iloot [Jf
.tee strength Nill have to be increased to i8pro\~ lodging resistanee.
lhe demonstration that photoperiod response is olten closely relatOO to yield perfor.ar.~e establishes the fleed lar assuring proper cliaatic adaptation. ROOuction 01 aaifltenance resPiration at high teaperatures may prove uselul. Nitrogen partitioning should permit starage in nan-Ieaf tisElles far remobilization during podfill. This eay also require a rOOuction of pod sink strength in early Etages of podfilling.
Specilic combinatian of yield component. should not be sought, besides aaintaining s.aller seed siles USO to 2SO lIg per seedl. It is possible toat number 01 seeds per pod shoold be lOIlitored in relalían to ovules per pod ta assure that seed fore.tian is efficient.
In the fir.t 15 to 20 days after emergence, toe growth curve of tbe crop may actuallr be slower than Hith eKisting cultivars, reflecting less allocation to leaf growth. Early reproductive grOHth .ay also be reducOO, thi. beiog associat.d with longer leaf life during pod-filling stage. Sreater leal life will assure eaintenance 01 high seed-fílling rates toward maturíty.
lIhile tM. crap will have a hígher yi.ld potential, eaillPetítive ability will have been sacrificed (Jennings Afld Je5us, 1968). The crap should perfor. pODrIr in .idures Hith traditiooal cultivars

- 36 -
!lf oth!!r crcps, and in the presente 01 heavy wead cOIIjIetition. I!ariability in individual ple"t ¡'ields will be ION. The crop Hill have a pOOl'" tolerante to defoliation, whatev!!r tne cause.
lhe 60st ellieienl Hay to develcp cultivar. satislying thes!! requísites is first to yield test a wide range of cli.atically adaptad .at!!ríal. under the agronOffiic cooditions speciliad above. Maturity date .hould be recordad to correet yields lar length 01 growth cycle. 5creening far specifie character; should begin with lodging resistance, nitrngen response, and reducad lad_ leal size. Reduead .... ifitenance respiration is desirable, but becau;!! 01 the equipllJ€f1t needad lor screening it is not suggested as a primary seleetion triterion.
For sHes where late lIIaturity may be e~ploited, selecUon lar lateness sflould prove uselul, but ooly if combinad with striet control to eliminate materials Hith 1011 reproductive sink strength ('poor adaptatioo' or low pod setl. Selection strategies might inelude selertian lar late flowering and high node p05ition of the first eainste inflarescence (not node o/ Hrst f1owerl. &c1usíve relianee DO parents known to be well adaptad Ilght nelp.
Dther desirable cnaracterístics described lar the ideot¡~e should l!Il!fge through direct selectíen lar yíeld. Resources should not be investad en seeking them individually.
7. Special Probl_=
lhe preceding sections have discussed yield potent!al ~ith Iittl@ r!!ference to specifíc cropping systi!lllS ar li.itatíons 01 seed type. !lowever, .a~imilation of yield potential is oHen sought Hith spedHe Ii.Hation •• ProblelltS assoclatad with .hed crcpping and mixtures 01 beao cultivars are particularly diffícult. lncre.sing yield patentíal io ledium to Iarge seeded and in early maturing línes has .Iso proven trouoleso.e. Ihese two toplcs arl! discussed here.
l1ediUJI lo large seeded cultivars
rrñ cooditions per.itting ylelds 01 2.5 to 3 ton ha- t , ~dium to large seedad lioes (hereafter referred to as 'large seaded' lar simplicityl typically yield 0.5 ton ha- t less than sI311 seeded ones. Compariog!lean yields over three trials al 57 lines varying IrOll! !7Q Ig to 470 lIlQ seed weight, seed size e~plained approxieately 491 01 variatioo in yield Irig. 151. This would 5Uggest that lar •• ~imum yielos, ,mall seeded lines are needed, but this has to be balanced against consume; preferentes lar largee oeans. Ihus, there 15 considerable interest in identilying causes 01 this diHerenee in yield potential.
Tne Mt persu.,i ve set 01 hypotheses deri ve support fratl the recent lindlng th.t 51all and large seeded cultivars represen! two genetically distioct groups, reflecting diflerent origios. The small seeded líoes are predominantly !lesoamerican in origin, while large seedad ones have ao Andean origlo (Septs et fu, 1984). !he groups are suHiciently diHerent that progenies IrOl inter-group crOSSES
usually produce dwarf rl plants ISingh ano Sutierrez, 1984).
Accepting that the groups have dillerent origins aod are diffieult enough to intercros; that there has been little introgression among the groups, the dlflerence in yleld potential simply reflect differences in adaptation 01 the cultivars. The s.all seeded, Mesoamerican types are adapled to pests, diseas~s, soU and clilatic conditions tl'pical 01 .iddle altitude sites trough!y !lOO m to 1600 ., typieal 01 MesolMerica, while the larger seeded types are adaptad to eooditions found at higher eleYatioos 0000 m lo :rooo m) along the Andes. Although Iafly cultl vars are nON grown oohioe the reglans they were ariginally adapted to, the difliculty o/ crossing aIOng groups has li.ited the possibili Hes of transferriog cOlljlonents 01 adaptation which would improve yield perforaance.
Evidence supparting differences in adaptation Js fllWld in cOlljlarisons of photoperiod response io dilferent seed size groups (Iable 21).

Hg. 15
• • •
- 37 -
Y.-Id "" 2640. i.SOO.SW
RI-OA9""·
ÚlIIIjlarisoo 01 lIIean 01 'leed yields over three trials al twa locatioo'! and individull seM weight lar 57 bean cultivar'l. CIAr, 1984 and 1985.
Table 21. Photoperiod response 01 flowering lar dlflerent germplaslII accessíons at CIAr grouped according to 'leed sile.
!leed 'lile
Slaall (Iess than 28 ¡¡¡gl
Hediuo (28 to 3S ¡¡¡gl
large (greater than15 ¡¡¡gi
Nuober 01 accessions wi th gi ven ::-_-,photoperiod Jesponse _ol_floweríngt __ Day neutral Interoediate Sensitive
53 27 ']})
24 36 52
16 24 59
ChF = 47, P «).I)o!
t Based en delay in flowering in lB hr artificial photoperiod as compared to natural 12 daylength at F'all.ira, Colollllia.
large seeded /laterials are predominantl\' photoperiod-sen5itive and 5mall seMed ooe5, day-neutral. In yield trials perforl!led at different elevations within Cololllli., large seeded materials yielded poorly at the warmer 'lites, but were superior at the coolest ,ite tFig. lb). Austin and ""[lean t1912b) alsQ noted th.t large seeded /laterial. germinated laster at cooler temperatures tE"'!:! th'n SIlIall seeded one., suggesting thi •• ight provide a uselul screening characteristic.
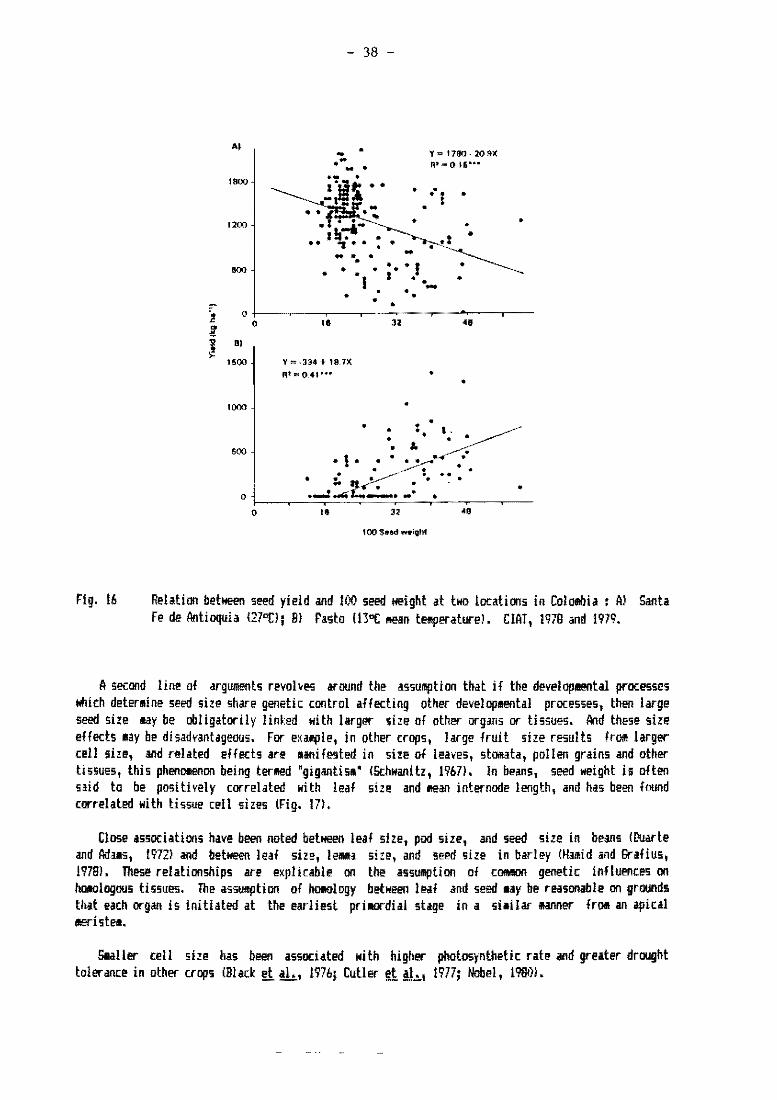
Fig. 16
A' ,_ 'lOO
600
l! O ~
O
" :!! O, • ~
1600
'000
500
O
O
- 38 ~
y ... 1190· :w 9)( R''''''OH ... •
• .* ¡ •
.... .. • • • ••• f
• *1 I • *_ • .a •• lO
y=, .334 + 19,1X
RI "" 0.41'"
•
•
•
•
• •
• •
•
•
• : :~·I ... .... . . 1. .* • ••• .A. . ~ ... • :. tt ... ~. ••. • •
• ...: ..... L..;_ ... * • ,. '.
Relatioo be!ween seed yield and 100 seed Height at two locatioos in Coloebia : Al Santa Fe de Antioquia 127"tl; Bl Pasto !13"t lean teMperature). CIAT, 1975 and 1979.
A second line 01 .rguroents revolve5 .round tne assumption that if the develap~tal processes which determine seed sile share genetic cootrol allecting otner develapaental processes, then large seed sile may be obligatorily linl:ed with larger size 01 other organs or tissues. And theS!! siz!! effeds lay be disadvantageous. For example, in other craps, large Irult sile results IrOlft larger cell she, and related effects are uni fested in she 01 lea,es, stOlftata, pollen grains and otner tissu!!s, this phenoeenon being termed 'gigantisl' ISchwanitz, 19b71. In beans, seed weight ls o/ten said to be positively eorrelated with leal sile and mean internade length, and has been found correlated with tissue cell siles (Fig. 171.
Clase IS5ociations have been noted between leal size, pod size, and seed size in beans lIluarte and Ada .. , 19121 and between leal size, Iem.a siz!, lOd seed size in barley (Halid and Grafius, 1979). These relationships are explicable on the assamption 01 eOlOOO genetic influenees on hMologous tisSUe5. !he assulIjItion ol hMology between leal and seed .ay be reasonable 00 grounds thilt eileh organ is initiated at the earli!!lit prillOf'dial shge in a siililar NIlfl!!r IrOll lO apical lI!!ri .tel.
liIlaller cel! size has been a5sociated Kith higher photos~nthetic rate and greater drought tolerance in other craps !Black et al., 1976; Cutler et al., 1977; Nobel, 1980).

Hg. 17
- 39 -
• Y-154J; +665.;(
•
•
• 200
o 0.2 04
Seed ~'qht 101
Relatlon between endodermls cell sile and seed weight in B bean cultivar.. ~ell .¡le Is a@asured as cross sectional are~ In arbltrary unlts.
Po.sible relations between.eed size and pod growth rate also .íght ¡nfluence yield potential. If larger seed; illjlly larger individual pod Hl!lghh, and lhis in tum requires hlgher pod growth rates ar longer filllng perlod., large seeds sight result in excessive siok strength Hithin a leaf-race.e unit. White tll'Bl) foond highly .igoiHcant positive carrelations 01 tl.!!b and 1).85 dIlOflg seed slze and pod dry weight in cOMparlsons 01 10 and 38 lines, respectlvely. Hawever, in studies of individual pod growth patteros, seed size and pod growth rate were not earrelated (CIAT, 1~85I, dillereoce. In pod slze prlmarily resulted IrOlll variatlon in duration al pod filling, nat rate al 111110g. Thís 15 In agree.ent with abserved díflerences In the linear seed lillíng periad found belween dllferíng plant afehiteclures and strongly re!ated ta final yleld (Izquierdo and Hosfield, 191m.
Clearly, the causes 01 the differenee in yíeld potential aeong seed size groups are not Hell established, although hypotheses abaund. Adaptatíon dilferences are prabably af tajar llIjIartance, but accepting this, one is still lelt "ith the passibility that characteristirs abligatarily a5sociated Hith large seeds are adaptive in eooler (limates.
Ear I Y Maturity
Far eonditions where grawth se_sons are lili ted , there is a strong IIOtivation lar Ia.'illlizing yields. Althaugh lIany arguments lar yield pateotial can be e,tanded directly to early maturing lioes, researchers have suggested that certain characteristic5 shauld be advantageous in early IUturlng Iine5.
Befare cansidering soee strategies lar inereasing yield potent!al In early laturing lines, He shauld consider the definitian 01 earliness. Given the stroog eflects 01 climate and phataper!ad 00
.atUfalían rate, no rigid definltlon can be given, nar Nill specific lioes be early under all tonditions. However, as a rough guideline, He will assume in these discussions that iE "normal" Ilnes at CIAr, Pal.ira mature in 75 lo BtJ days, then e.rly lines HOuld ¡ature in sa to 69 days.

- 40 -
The illjlartance 01 specialized agroooeit lanageaent lar eMly lines also _its note. Accepting that source li.itations Nill be critica! in early aaturinq aaterials, rapid estahlisheent 01 leal area should be critical. Sesides seetiog varieties with this Eharacteristic, plantiog patter" and density.ay he eodified lar ADre rapid establishment 01 crop Eover. Far loor early lines, ltllina (l9!lbl lound that a 20 CI by 20 el planting pattern resulted in greater canopy rover at 30 days, Erop dry Height at 50 day., and seed yield. than row spaciogs 01 30, 4S or bO c. with lhe sale plant population per uoit area !Table 221.
Table 22. Yield. of lOilr early eaturing lines sown at lour row spacings, but conserving a populatioo 01 20 ptants •• ~2. Pal.ira, 1985.
-----------------------kg ha-·-----------------------
RAIl bQ 2520 2100 229Q 2700 XAN 145 2J.W 2060 2240 2b1Q IlAT 41 2110 2UlO 2090 2300 G J8iJ7 1b40 1990 1870 1S'ji)
Mean 2150 2ObO 2120 2390
5.E. lar difference of treatment lII!!ans = 102
lMge seed sile is also Irequently aentioned as a eechanis. lar achieving rapid establishment !Rodríguez, t9SbI, but in cOfipMison 0140 eMly laturing Unes, seed Height HaS not carrelated with yield or yield day-' nable 23).
Table 23. Correlations iIOOg yield and other par_ters lar W early eaturing lines plus G 4525. Palaíra, t984b.
Yield!day Dar; lo aaturity Cro¡¡ dry weight Harvest índex 100 seed weight Seeds/pod Pods!.'
Yield
.9'1"
. son
.8m
.18
.14
.28
.bBU
tI Signilicant at the p = 0.01 level
Yíeld/day
.72U
.85U
.23
.14
.26
.bBU
Day; to liturity
.71U -.08 .18 .23 .mt

- 41 -
¡I a shart growth eyrle results in a ¡i.ite!! ouaber 01 sites lar pad lilliog, rel.tively I.rge pads or nulllber 01 pods per nade woold increase sinl: size. In eontr«st, lar a mljl with a shart periad lar pad ¡illing, very ellieient distrihutioo 01 siok demand ftight be crucial. Thi5, io turo, sugg!!sts an advantage lar ea,ly aaterials with smaller pads distribute!! over a wide range 01 .ites.
Argu_ts lar either a relatively long ar short duration 01 pad filling tan also be given. Sine!! duration of podfilling is positively carrelate!! Hith yield, one might seek tultivar. maNimiziog tbis periad withio the growth (Vde 01 early lioes. Alternately, if total vegetative grOHth deterMines crop yield, a Iíoe whicb f10wered late, but filie!! pods rapidl y woold be preferred.
Many other strategles have been sugggeste!! lar increasing yield potential io early m.turing lines. The reader 15 re/erre!! to Rodriguez (19Sb1 far details.
B. Potenti.1 of 8iotechoology or other Strategles In the Fulure:
f'revious dlscusslons have adhered to what Jlight b~ eonsidered conventíonal stralegies lar increasing yleld potential. However, with the .dvances in t;:chnlques lar maoipulatíog genes and propagating plant" there e,ists the possibility that advanCEs will resillt frcm strategles considerably outside the scope 01 those oullines above.
~lthough e~ploitatíon 01 heteros!s through use 01 h~trid sE.d has shown little prooisE io comeoo bean; (Guti!rrez and Siogh, 19841, other strategíes lar e'ploitirig heterosis e,lst. In scme crops, seed 15 produced without tbe genetí, recOl1lbínatioo occurring in meiosls (Bashaw, 19BOi, the process being terme!! "apoaüls' ar eore specifica!ly "aga_permy' (seed larl1li!tion without praductíon 01 gametesl. ApOOli~is occurs in very IOH Irequencíes in grain legumes (Sobolev et. al., 1976), ;··,t with a eore cOI1Iplete understanding 01 control 01 eeiosis it light be possible to coostruet a apolictic cultivars mueh in the way as has been proposerl lar potatoes lHer.seo, (980). Alternatí,~ly, iprnllíxis .¡ght be sought through mutation bree!!ing. ApOftlhis cnmplicates breeding stretegies, but colllbioing a genetic prapeosíty for apomi,is with a requír!!l1lent lar chemical ínductioo woold mini.ize thi. drawback Uwanaga, 198,i.
Another a,antia lar exploitíng apOl1li,is i5 through vegetative prop.gation. A few areas 01 bean praduction already ¡nvalve transplant 01 seedlings (5ingb, p!'fsonal co_nication), and introouting vegetatively propagated material should prava straight lor",.rd. PresentIr, it is dífficult to propagate beaos through tissua culture 1l1roginsH and ~artha, tS'!I4), but i I barriers are Q,ercol!\!!, FI hybríds .ight be propagated through tissue culture, aod distributed as "somitic e!IIbryoids", that is, e!IIbryo-like propagules.
lllthough coosid!'fable wark has been devate!! to tissue culture witb P •• ulgaris, to date, there is no publishe!! and confirme!! procedure available lar de novo plant g!'l1eration Iroa either callus, ceH suspension, ar protoplast cultures. Reg!'l1eration 01 plants by in vitra culture has sU!:ceeded Hitll apical meristems 01 zygotic embrvos only.
SIloot apex, callus and cel! suspeosion cultures ar@ relatively easy to establisll and were empla~ed in sevenl studies but results are very poar. lo a report tha! has oever been conl . ,1, I;;;¡
plaots were regenerated ¡rom about 400 c¡¡llus tultures when e'plan!s originated ¡rOl leaves 01 P. vulqari s were cul ture!! on a mediUl~ supplemented with bean seed extrad (CrocOfllo et &,., 19ni. An average o/ 15 regenerate!! buds per eath explanted ,haot apex was obtained ¡roo the cultivar "Palmital F'retoce" with fuI! plants recovery aHer lhe third suheulture wito gíhb!'felie fGA.l _ud naftalen 3cetic acids iNAAl growth regulators (~lartins and Sondhal, 19841. V!'ry fecent r.5ufts 01 fese3rch in tM, area at CIAl have shoHf1 tlle induction 01 ,om.tic embryo-Iike structures on beao c.lIus cultures lRoca, 1986, personal tommuníntioo). But, to re¡¡IUe ooy DI lhese possibilities a comple(e breaHhrough in regeoerating bean plants IrDm tissue cultures is needed.

- 42 -
Altllrugh current tedmiques lar h)'bridiüng I~ have peraitted Ií.íted crOS5eS between leguae gl!f1era ar species IAlvarez et al., 19I1l1, investigation on the US! oi protlljllast lusioo to prooocl! sOiutic hybrids suggest the possibility 01 prooucing grain legui!e5 conhining desirable ¡eatures 01 dishntly related QI!f1era ISchieder, 1'182). Thus, bean cultivan Hith the late .aturityand architocture of soj'bean plants, light result frOll sottatic hybrids 01 COllil!OO bean and soybean. Considerable infar.ation eKists on ¡usion 01 so)'bean protlljllast "ith non-legUles, but in no case have plants bl!l!f1 regenerated (lIroginsH and ~artha, 1'i'Il4).
Assuling that beans Nill be grown in envirOOleOts Hhere a C. photosynthetic pathway are advantageous, one .igot reconsider the possibilityof convertlng beans lo a C. pathway. Screening 01 rl'ier 60000 planls of so)'bean, !!ah and Hheat falled to detect a single individual Hith a IOH C~ compensation point IEvans, 1975), and identical lailure rates in other crlljls suggest that suco a change HilI not be achieved using current breeding techniques. However, Nith improved abtl i tr to identífy and _ipulate spedfic genes, the complete set of genes required lar the C. pathway light be transferred.
6enetic IodHícation of plants by genetic Itanipulatíon 01 the!lNA level are still lar in the luture. IIost 01 this research currently uses Iodel plants and genes. However, recOlbinant!lNA technology .. V, on a II!!diu. tere basis, be used to devellljlllOlecular genetic ...-kers Hhich in turo can be use to buílt genetic linkage laps lar ocooOlic impartant traits (Vallejos et al., 1'1S61. lhe availabillty of tMs procedure HOUld help bean breeding by Kcel!!ratiog the introgression of lareing genes into cOllercíal cultivar and is under study at CIAl (Roca, 1'1S6, p.c.l.
Other strategies are !!a,¡lr envisioned, but the aboye e.amples should serve as Marnlng that Hhile 'standard' strategies hold the greatest prOlise lar the ¡_iate future, on!! shoold not be blind to other strategies. l'erhaps the best rocOill!ll!f1dation e_s frOl Ralph N. [lIerson in the 19th century 'A fOQlish eoosisteney is the hobgoblin 01 little .ind,'.

, '1M. ' N'
- 43 -
IV. PRODUCTlViTY UN/lER STRESS
For the vast majority al bean production regions, yield potentials are never realized because af cOfflbined ellects 01 edaphic, elimatie, disease and pest problems. In most (ases, there is genetic variability lar responses lo strEsses, and considerable eflort has gone to the sludy of talerance or resistance to diflerent stresses. The discU5Sions that lallow efiPhasize abiotíc strEsses sueols drought and eKCESS water. Toe last section di5cusses general stress tolerante ar 'rusticity' in beans.
A. Drought
The import~lce 01 water delicits in liliting crap yields tan ultimately be traced to the faet that lor most field conditians, water vapar dilfuses out of a plant .are easily than tOo, and that saO water is oot sufficient to permit the plant to eontinu.lIy replace lost water. Conservation of water through st_tal closure or other liiecnanisms reduces pholosynthesis, taus erap growth is limited. This results in a clase rel.lían bel"een Hitar avaílabilily and yield (Fig. !ll).
A high relatíanshíp belween lea! area inde, ard eViljlotranspiration has been observed in Sflap heans by Bonanno and Milel (19931 in agreement wi tn the linear fundian described in Fig. 19 b~tW
Fig, lB
Hg. 19
- 44 -
o o o
A 170 o o"
" " o " o ." Ohu;!ol • C .. "" • •
• • '. 2000 • ~
" • D Da " • ,. g • . ! ~ • • " O
1000 " • 11 'a • 04 • " . • • • • O I
O 100 200 300
W.ter Ipplisd !mml
Yields oi A 170 and ev. Uiacol ealila al dif/erent levels nI applied water using A line soorce sprinkler inshll.tíon. CIAT-I'allin, 1'185 •
4000
'000
" "
1000
250
Y-17 42X 3591.66 'Al", .1133 F "'30.0
. ' .. .. '
"
" .' o'
300
"
, ' .. " '
. . ,,'
.. -~ .. ••.••• O
"
O
.' . .' .... . .. '
.' '
350
.' . " '
. ' .. , ' " .' " . , .
O XAN·'.
'000
, ' "
.. ~ ... ,o'
linear relationship ni evapolranspiratioo on seed yield af two block bean genotypes, ('I5l confídence li.itl are indicated by dotted lines). !iO!.Ircel Jara, Izquierdo. y llatta 1'lllS.

- 45 -
Rapid oo.et al drooght CilUses inlerruptions bolh in photasynthesis and in tissue eKpansioo.
EHeds on photosynthesis may be ascribed to two c011Iponents (O'Toole et al., 1977b). Onl! 15 reduced gas eKchange due to sloaatal t!o.ure. In beans, fu!! clasure .ay be reached in a feH .¡outes after ao app!icatioo oí a .tress (Hg. 20). With sloaatal dosure, C!b uptal:e is .ioieal and O" ¡(cumulates, causing decreased Cu. fi~atioo, but iocreased pnotorespifation (POKles and OSlOOd, 1779). There i5 .till considerable argument as to Khat actually triggers sta.at.1 clasure. Abscissic acid (ABA) levels increase under droughl and can trigger (Iasure (AspiRall, 1980). but clasure in beaos Has detected lfell helore iru:reases in ABA (NaHon et ~, 19m. StOltatal clasure has also been eorrelated Hith decreasing leal Hater polential, PI (Moldau, 1973) and iocreased leal tesljlerature !Fig. 211. Recenl warl: suggests thal in beans as lfell as sany other mljls, rapid responses under field conditions aay reflect sensitivity to decreasing atsospheric hueidity, eeasured as increased 01 vapor pressure de/icit (EI-Sharkaway et ~,1985). Seans appear inlerlediate in stoeatal sensitivity to hwlidity !EI-Sharkaway, et ~, 1985), suggesting that they lollow a strategy interm!diate water eonservation (drought avoidance).
The other C011lponent reducing photosynthesls i5 los5 01 efliciency in the physiological processes 01 photasynthesis, reflected a5 iIIl iocrease in eesophyll resistante (r.). Under dessication regiees siallar to lield conditions suco ellects occur only at IOH water patentlals. Holdau (1973) fouAd no increase in rM resistance at leal water potentials dOHn to -1.3 MPa.
Atteopts to determine which photasynthetic processes are resposible lar locreased r. are inconclusÍI'e. O'Tonle et al. 11977b) lound that tne decline in RuIlftase was actively greater than the iocrease io f.. Dessieation 01 chloroplasts and associated disruption 01 the chlorophyll-thylakoid membrane co.plex .ay also be iovolved. Osmond et. al. (1980) proposed that, in beans, internal cycling 01 tn. through photorespiration plays a majar role in protecting against stress associated daaage, and tn.t muen of reparted d.aage under stress i5 due to rapid stresses applied in laboratory e~periments.
Fig. 'lf)
'00
•
• 0
o ,
JO ,
.0 I
00
Til!le needed to reaeh stOftlatal c10surE in a detached leal 01 MI 477 as indicated by onset of linear phase of water 10s5. CIAT-Pal.ira, 1986.

- 46 -
Vrought stress lay affect tissue expansion both through redured cell expansion and dívi.íon. Usíng sadiu¡ ehlaride concentratians to generate diflerent water potentíals in culture solutioos, BrOUlfer (1963) eonduded that interruption ¡¡f eel! expansíon Nas eore íllflortant than eHeds on cel! divisíoo in lilitíng leal growth. Bunce (19m lound a linear relaHan bet_ soybean leal elongation rate and turgor pressure, and further noled thal plan!s grown in drier conditíoos required less turgar to _chieve a gi ven i!!IlOUnt 01 eloogatíon. However, eare recent _~ has lead to a tonsen.U5 that reduced turgor pressure í s not the pri.ary cause 01 cessation 01 tí SStie expansioo lIIlder dr ough t.
C.nopy tempetafUHI difffi&Jl!'.¡ f'Ct
n J • • 12
I ,.
-.2 '~
• l .4 ;!.
1 " •. $ ~
! • • -: ·8 ~
·'.0
R'."."'O.993 ...
• 1.1:
fig. 21 Relation between leal water potential and canopy te;operature diflerential lleasured between stressed and irrigated plot. frcm 4 bean lines.
Responses Hith!n hours
As a stress continues to be applied, plant water potential declines, rellecting coobined eftects 00 turgor pressure and oSllOtiC patential as water is last. Extreme redurtiOfls in turgor pressure result in wilting includíng loss af ability ta orlent leaves. Estilates al peroanent Hiltíng points in field 9rowo beans .re variable. 5ponchíada (1995) obtained sufficient stress to induce 'burning" of leaf largin., but PI wás never 10Hef than -1.1 tIPa. Jara (I'1!lS) obtained values ¡¡f -1.2 It'a as the leaf potentíal of stressed plants at 82 day •• Her planting.
80th ABA and prolin! accu~latiOfl have been detected in drought stressed beans (Stewart, 19721, ABA appear. involved in stOl!at.1 funcUon lIIlder drought, but sinee it acculllUlates in other tissues, additiOflal functiOfls lay be i"volved. AccUlflulatioo of proline, ana nitrate ana _ium i005.ay coolribute to asmatic adjust~t and re/lect inhibition of proteio synthesís IStewart and Hansoo, 1'I8f)1 frota afid fucker, 1978l.
Response ¡¡ver longer periad.
The previously discussed ellects ulti~ately result in .ajar changes in overall plant growlh. RedUl:ed growth is ";ua!ly detectable soon aHer onset 01 stress, "ith reductioo in leaf area being particularly earked. For a stress applied before flowering and sustained to maturity, Spoocñiado

- 47 -
(1985) laund that lina! bi~5s and lAD were reduced to SOl 01 levels in contral plots, althaugh yield reductions were appro~iaately 40Z and 801 lar drought tolerant and susceptible lines, respectively (Table 24).
Studies in other craps have shown that stress during early reproductiva growth usually has particularly severa effects (5latyer, 1969), and the sa/líe holds in baans. 5toker (19741, using lysi.eters to provide equal stresses at dilferent stages, obtained 20Z yield reductions with stress during early or late vagetative growth, 507. redudioo in early podfill, and no reduction in l.te podHII. In soybeans, eflects on yield cíli!Ijlooentes HerE easily interpreted in terlis 01 tiling al drought stress (Shaw and laing, (966). In bea"., the same would appear to be true although the relations have "ot been tested as carelully. ror a stress sustained to maturity, all yield components sbowed reductions, while a stress ter~ínated by rains during podfilling caused seed W9ight to ¡ncrease relative to the irrigated control (Fig. 221.
Drought stress in beans is usually associated with accelerated eaturíty (Fig. 23), although late 5eason release ¡roa stress may trigger recuperati.e growth resultíng in delayed maturity.
Cíli!Ijlari 5011 01 drought response of otnar legu,.es
Drought of response 01 other legumes vary greatly. Nithi" the genus fflaseolus, P. acutifolius is aften cited as being lOfe drougtht tolerant than r. vulgaris (~ary and Webster, 1984), but its small seed size has liaited its cOllercial accephbility.
Table 24. Comparisoo of growth parameters 01 two draught toleraat and drought susceptible materials grow dfaught ID) and irrigated (el plet; at Pallira, Colombia (Sponchiado, 1985).
__ Tolerant -- ____ SUsceptible __ Variable Treatment BAT 85 lIAr 471 BAT 1224 A 70
Yield (kg ha-!! o 14/,0 ¡' 1460 d 550 b 550 b e 2520 a ;:'54') .lb 2140 .¡ 2771) b
BiO;¡¡ilss ikg ha-') D 2bOO 25(1) 2200 1900 .lb e 5200 5300 5700 5400 a
LAO D na 83 b 74 ah BOa C 171) b Ibl b 171) b 186 a
HI (Xl D 55 a 51! a 25 b t!I b e 4Sa 4!l ¡¡ 41¡¡ 51 ¡¡
! Val ues lellowed bl' the same letter wi thin a row are not di fferent at p = .05 level 01 Duncan'; test.
Some traditiooa! cowpea cultivars responde ta drought thraugh extreme 5totlatal closure and restriction 01 water 1055 (Shackel and Hall, 1979), In a comparison of .ungbean, soybean, cowpea and peanuts ¡Pendy, et ~, 1984), the first tbree species showed patterns of response s¡_¡lar to beans,

- 48 -
including reacning eini.ue Pp al -1.2 to -1.4 HPa, but peanuts were muen less alfected, reaching only -0.4 1'1"a.
"" .00 •
i ~ r • ~ 1 • • 1 '" ~ 3 I ! '00 '" t 2 .. • "
o o
~""IH<g:.IH, O -Oroughl
Fig. 22 EHed al drought frae preflowering to end of pad fi 11 at Pal.ira and 1M lichao, Colombia an yield components of susceptible line A 70. Source: Sponchiada, 1995.
2. llr!lUght Tolerante /!echan!s.s:
A great eany drought tolerante _hani5fl!s nave been prapased, and deaonstrated to fundian in very specifíc cootexts. flowever, the warning frae the previ!lUs sectian that drooght effects varV greatly depending an spetiiic dr!lUgh! canditions is equally valid lor cansideratian al toleranti? tleChan i ses.
He define 'drougnt tolerante" in an agrooaeic sense. ror a group al laterials which give si_ílar yields ander noo-stress canditions, a m_tedal is tolerant if it yields welI coepared to the others under cooditions involving water delícits. A.1Ioog mechanisms which aay can/er tolerante are tOMe which ecologists usually dilssily as drought escape and drought avoidance. AH rutline 01 possibll! IleChanises is províded in Table 25. it lollows classifications used by Turner (1994) and others, with {he notable additioo that for cooditians whefe beans are grOl<l1, tolerante to factors related ta drooght such as high temperatures or saline soil. lay be essential. The discussion that fallos ampoasize the physiology 01 the ba.ie IleChani'ls.
Ilrooght escape
Bean traps tliay "escape" ur at least reduce drougM stress ei ther by ILlturing earlr or delaying eaturity until dro'lght stresses have abated, this laUer strategy terlled recuperation. lhe usefulness 01 the two strategies varies with timing and iotensity af the stress. Severe stresses occuring later in toe grlJllth cycle favor earliness, while .ild stresses relatively eMIr Kili favor later eaturing línes with capability lar recuperativa grOHth.

2500
2000
N
.:. .r.: O> é lSOO ." -¡¡ ¡;:
1000 •
• •
• •
- 49 -
• • • •
•• o •
• ••
•
•
sJ,----O 'r-------T'-----'-' ~, Hg. 23
S 9 12 15
Reductlon in time to maturity. (day.)
Acceleratioo 01 $aturatioo UI1der droug/lt stress as iodícated by reductioo in days to eaturity oi stress V5 irrigated plots lor 77 líoes. CIAT-Palaira, 1984.
lable 25. Classificatioo ol possíble drought tolerance aechanislI$ in beill1s.
l. Ilrought escape A. Earliness B. Recuperatlon
11. Ilrought tolerill1ce Hith high plant water potenBal A. ilaintenance 01 water aptal:e
l. Sreater root growth 2. Irnnased hydraulic coodudance
B. Redudioo oi water IOS5 l. Reduced area of evaporatioo 2. Ilreater resístance aloog pathway 01 water 1055 .3. Reduced gradíent fros leal lo atlosphere
lll. Ilrought tolerance Nith loo; water patenBal A. Hai ntenance 01 turgor
l. OSllOtiC adjushent 2. Increased cell elasticíty
B. Oessicatioo tolerance l. lI •• !Ir d.,., sf;¡bl I ity 2. Protein functlon
IV. lntegrated .echanis., asslmilate partitíooing and remobilizatioo V. Droug/lt tolerance through adaptation to factor. related to droug/lt
A. Heat tolerance B. loo; soil ~ertllity C. San tolerante D. Soil diseases and pests

- 50 -
Drought tolerance with high plant water potential
VMious tolerance _hanis~s thooght to fundioo in bean craps result in .aiotenaoce of relatively high plant water potentials. ThEse include those Hhich per.it greater water uptake, aod those Hhich redUCE water 1055.
/!alntenance 01 water uptal:e:
As in .. ny other craps, drcught tolerance in beaos has been associated Nith greater root grcwth. Coapariog tolerant and susceptible materials in a fertile soil per.itting deep root penetratioo,
Sponchiado (1985) showed that roots tolerant il\atEfials penetrated aboot 51) c. deeper than susceptible ooes (Hg. 241. Difference; in root growth and yield _e also associated .. ith greater l!l(ploitatioo of available soil moisture by the tolerant línes. However, this Mechanis. was not ef/ective in a shallcwer soH where a pH 01 S at depths below 3<) CM prevented I!J(tensi ve root growth (Hg. :15i, and in this environllel1t, yield diHerences refledíng suppased tolerance were oot fcuna (Table 26).
Inereased eflicieney 01 roots in extracting soil MOisture IUst also result in greater water uptal:e. Doe alternative í5 lor a erap to be able lo tolerale lcwer root water potentials and lhus extr1Ct soi! water held al 1000r potential5. HOwever, sinee this is hasically a eechanis.s al tolerating dessication, it is diseussed latero Anolher alternative is lor roots to have greater hydraulie cooductance, elther axially Hor transport along the *y191 or radially (for uptake frQfll soí! lo the ~yI9¡. Taylor 119801 concluded tha! axial resishnce in sorbean roots lIiIy be iaportan! at roo! depths ayer 1 1, but Passíoura 0'1ll31 suggested that becauSl! of capadty lor secoodary growth of Kyles, a.~ial resistance woold not be ¡¡ei ting in dirotyledooeol.l5 plants.
Reductioo 01 water IOS51
8esides inereasing water uplake, a crap lay have adaptatioos which reduce water loss. litfortunately, reduced f10w 01 water vapor usuall y iaplies reduced diffusioo of COz into the leaf, and thus lieit.tioos in photosyothesis.
As IIef1tiooed prel'iously, drwght causes Marl'ed reducti0l15 in un, but there is no evidente that tolerant be!n lines have gre.ter reductian than susceptible ~aterials. Tolerant .. terials shON si.ilar LAD as susceptibles Isee Table 241.
Reduced leal area aay be cOl1lflensated lor throogh a greater Mesophyll sudace area per uní! leal area INobel, 1'1I!i)), this occurring with increased leal thiclnes> and/or reduced cel! she, Hhieh in turn .Hects stA. ChangEs in leal thicl:ness was coosídered in an e~perilllEl1t with tIlO bean lines dillering on their response to nater stress {fig. 26). lino A 211 Nas able ta lai"tai" higher LAI without increasing leal thiel:n.s; as mueh as the Une XI1/HO did. Drooght yield of :15 genotypes have been correlated wíth the leal thidness iodex Ir = .7QUl, but the correlatioo under irrigatioo HU
even larger Ir o .91U, Fig. 271. This index !lay be related to photosynthetic eHiciency in tenIs &1 carb~ylating area, and in absence 01 water stress, .. y be used lar assessíng genottVic photoassimilatiOl1 ability.
Meehanis.. involvíng incre.sed resistance primarily involv~ barríers to water loss at the leaf-atllOSphere interface. lh" gre.test amount 01 water los; 15 usually via stoeata, 50 coosiderable attentioo has been given to studying diflerenees in sta.atal reslstance aMOng bean lines. SiMple relatlons a.ong stOMatal resistance and level of tolerance are not expected because of the relalian ~etween transpiratioo and COo uptake. Rather, stlliUtal respoose to enviroo_tal coodi tioos Is prollably crltlcal 1ft detllfmioing water use effidency. Ideally a plant shoold regulate transpiratian so that it nccurs when IOlslure qradients are ION (ION vapor pressure deficit In atlOspherel but

- 51 -
radiatim levels are aboye the cOIlflensatim poínt. 111e aain _hanis. whích would previda this response in C3 plants is stOiatal sensítivity to humidity.
I .<: o. ~
" ~
Fig. 24
Fig. 25
Root Length (cm)
o 20 40 60 Or-----~----~----~-
,."
o" "S~$ceptibles
40
60 : Tol~ranu
120
o Available Soil Moisture (%)
20
Toleranu:
40
" ", .•.•
60
COfIIIariSOll Ot lI!?an root growth and available seU lIIIlisture in drOOjjht túlerant mAr B5 and BAr 4771 and susceptible lBAr 1224 and A 701 line - Pal.ira.
Roor Length (cm)
o 60 120 Ot----~----~-
30
60
90
..... --,." 0'- __
Tolerants
8AT 1224
A 70
Available Soil MoIstl.H'I! (%)
20 60 100
COIlflarison 01 eean root growth and availaMe soil roisturl! in drought tolerant lBAT B5 and BAT 477) and susceptible (BAT 1224 and A 70) lines - Quilichao.

Hg.2b
r E
" r i ~ • < ~ E
3
- 52 -
s
o ,.....-----~--~----,--_.,.'~--T-~--__r·~--_,.._
•
1
•
•
4·
., 70 16 .el 90 98
0 ______ 0
....
b b
• •
.o~ ,,,,,,,'" b --O
/' b
• •
-+--~.~~-,.......--~~.~,-----,-~-
5& 11 8'591
Mean leaf Mea index and leaf thidness index of tIfO bean genotypes UAN-401 susceptible and A 2111 tolerantl with diflering response to water strells under loor ~tress conditions. ArrOH ShOHS 50l flOHl!ring. Mean separation by MIlT Duncan 51. Chillan, 1995. Sourcel Jara, Izquierdo and Matta, 19BB.

- 53 -
Amount of cuticular wa~ Has related tD rate 01 water loss in sarghum ([bercan, el al., 1977), and large varietal dilferences in cuticular resistanc!! 01 rice leaves were loana (Yoshida and de los Reyes, 1976). Woodlin (persona! cOfimunication) reparted considerable variati~1 in cuticular W3~ larMatian ~1g bean cultivars.
Bourulary layer resistance wi!! increase with greater leal pubescence, {Ehlednger, 19801 although this mechanisms has not been studied in bedos.
Assuming resistance along pathways 01 water loss are equall, cultivars may diHer in water use through e¡lect. on the water vapar pressure gradient IrOi leal tissue to the at~sphere. The e,ternal pressure will be equall, but internal vapar pressure Hi!l vary with lei! temperature. Thus lactar allecting leal temperature Hill reduce water loss. Leal arientatian can reduce radiation ioterception, but at the cost 01 same light energy intercepted lar photosynthesis. Leal pubescence and cuticular H.xes someti~5 appear to re!lect iofrared !ight Hitn relativ~ly little ellec! 01 PAR. Vari.tion in leal colar both .Il1009 ~arieties and between darsal and ventral sides of the Sdi\!! leal
area considerable in beans, but do not have not been ex~ined in relatíon to leaf teoperature eHeds.
Redueed leal size Nill can also reduce telljlerature sinee lar a given total lea! area convecti,e cooliog 15 greater far a surface divided into s.aller sertians (Gates, 1965). lbis effect has lead breeders in other traps lo breed lar saaller leal si¡es. Hiebseh et !197b) did not detect illjlroYe\lIeI'lts Hith i50-lín!!s 01 soybea"s diHering in leal size. LeaHet si.!! W~5 signifícantly reduced by an inlense post-Iloweriog water stress, aut the relationship between drought leaflet area and stress degree-day measured by inlrared therlllOmetry was not consistent amang 25 bean genotypes (Izquierdo, 1985, unpublishedi.
Drought tolerance Nitó low plant water potential
Although the precedíng IlieCnanis.s Hill reduce water 1055, most stress cooditioos are suHiciently severe that water potentials drop to lev~ls which affect plant lunctíon. Thi. leads to the possíb!lity 01 adaptations permitting lhe plant to function at 10H Hater potent!als.
J'faintenance 01 turgor
Sinee water enters roots tnrough flow along a gradient, lower root water potentials !F'rl wiH result in greater e~ploitatíQn 01 soil voltimE. Thus a crap whieh ha. a greater reduction in Fp or F~ lar a given aJIIOilllt 01 water 1055, should show more drought tolerance. However, Níth 1055 of turgar, evidenced by Nilting, !he plant loses the shility to eontinue ti.sue eKpaosion or to utili,e IeChanislS such as leal arientation which utili,e diflerencEs in turgOf pressure. This indicates lhe need far IlleChani.ls which per .. lt low Pp ar Pr wlthoot los. of turgor.
When water delieit is severe, plant eell. acc~late salutes, whicn lowers their internal os.otic potential, in arder to at least partially •• intain turgor pressure, and thus permit continued cell expansion and growth. Osmotic potent!al may be regulated through soift. in concentrations of potassiulI, sugars, dllino arias, and arganic adds Hurner and Janes, 1900; I'Iorgan, 1984). H&rl.harl (1985) cOIpared oslOtic adjustment of Phaseolus vulgar!. and Phaseolus acuti/olius grown io containers, ann did nnt delect d!fferenees. Jara (1985) found significant ailferences of about 0.0,5 If'a at tiee of flowering allOO9 ha hean cultivars under field conditions, but nol al other growlo stages.
Assuaing ~hanislls of osmatic adjusteent are the .ame in dilferent growth stages, groHth 01 seedling root. lo media with lOH potentials lIight proviae a simple index 01 capacity lar turgor lIIaintenance through osllotic adjustMent. In a comparison 01 25 cultivars, percent inhibition 01

- 54 -
radlcal e11l"l~ahon ;,:as ni~:,!y correlated (r ;: 0.72) Wltt1 ~rc,lj~'1t 3U5Cf!ptl~i~ity ~leaS1Jred with Fisher index (Hg. 2S1.
file mechaoieal characteristies af tissues al50 affect turgor maiotenanee. Reduced tissue elasticity, indieated by a high a buH: tissue elastic lIIOdulus 18l!1, will resalt in a small decrease in Hater cootent per uni t reductioo in Hater potential. Tne pratoplasl wi!l retaio lIIOfe Hater under stress, but at toe cost of 1055 oi turgor. Kim and Lee-Stadelmann 119841 (uuld nat detect difference5 in El of beaos in stress and control eonditions, and .hile Jara (19851 did find sueh differences lar four water regillie5 under field conditions, varietal diflerenee. were nat signHicant.
Table 26. Vield, root growth and other par.meters al two drought tolerant lines (BAT as and 477l and two susceptibles iB!\T 1224 and A ](11
far plantings at CIAT-Pal.ira and CIAr-UUilichao. D = Ilrought treatment! e = Irrigated (ontrol.
Treat- &AT BAT BAT A Variable ment as 477 1224 71)
Palmira
Yie!a Ilg ha-') O 145B.· 1457. 548 b 57Q b e 2517. 25.l!l.1! 234~a 2769b
~ü! root depth Icm) D 100 le.) bu 60 e 50 51) en 4. 5f.l
Root dry weight (ag) D 541 S'i'1 534 494 e 660 715 O"" 598 :1.J
Canopy temper atUfe [< 34.0 14.1 }b.a )7~S
IDC)' C "" " 29.6 2'1.6 29.9 ':'7.':'
leal water potentíal D 8to) B40 !tOÚ 11')1) U:Pa)' e 451) 46') 4/ll) 470
Quilíchao
Vield (I'g ha-') D S99a 724a 7233 192 b C 28%a 2V52ab 2661 b 2124
90. root depth (CM) D :ro .') }O 3(1
e 30 20 2~) 2Q
Root dry weight lag) D 924 899 792 bba e 47ó bll bse 492
Cdnopy temperature D ,-8.4 39.8 39.8 41.4 (·Cl' e 211.6 28.0 28.5 211.5 Leaf water pütential 1) 84lJ 96(1 '!50 W:ro (~Pa) • e 4(10 45f.l 45f.l 5Qí)
• Yields lollowed by the .ame letter do nat difler ac(oráing to Donean's p : .05.
• Measured duriflg perioos al .axi_uro stress.

Fig. 27
- 55 -
• 2.900 y = 10628 Q7-1354.39X A =0.9'·· • ... ,95
1550 2.800 • • , •
1450 • 2.700 • ~
1350 • • 2.600 • • • • •
:- • 1250
• 2.500 • ~ • ~
" 1150 ~ 2.400 :!
y = 4285 8-491.3X ~ 1050 • 2.300 R =0.70"
950 2.200 • BATses;
• Irngll@d
850 Post·llo"",errng w.ter stress 2.100
750 • 2.000 'C.,.: , , <, , ,
5.8 5.8 8.0 62 8 .. S., 5.6 5.8 6.0 8.2 5' 85
le.1 thl~knflSS mdfllllmq cm"')
linear regression 01 leal thid:ness index on yield 01 25 hean genotypes (CIAT-BIDYT, 1985) under irrigated and post-Ilowering water stress. The index measured during pod lilling. Source: Huepe, 1986.
Dessication tolerance:
At very IOH plant water potential physiological processes may be disrupted. This subiect is poorl¡· understood, hut it appears that effects 01 dessication may he divided among those on ce!! membrane lunction and those on protein lundion. Under dessication ce!! ~ranes appear to lose integrity, and the elliciency of processes requiring membranes are reduced (Leopold, et ~, 1981). Si.ilar!y, although it is easy to hypothesize effects 01 dessication on enzymatic proteins, such effeds, IlUch less varietal differences, are apparently unresearched in heans.
lntegrated mechanis.: assieilate partitioning and remobilization.
The previous .echanis.s have heen discussed as results 01 independent physiological processes, hut it should he reali!ed that they interad and ~a\' share COllfl1Of1 hasis. For exa"llle rembilization 01 N and stored carhohydrates (starch) may he enhanced under drought stress, and this would per.it greater root grOHth or capacity lor osmotic adjustment.
Studies 01 carbohydrate relObilization under drought conditions (Sa"ller, (984) suggest that tolerant cultivars are more efficient at remobilizing carhohydrate during podli!!ing. Induced or accelerated re.1IObilization 01 stor~d dry matter due to water stress has heen observed in harley hy Ga!!aher et ~,1975. Remobilization could he triggered hy the stress and thEm support the seed filling instead 01 the i"llaired photosynthetic leal area. Figure 29 shOHs a yield redudion due to a post flOHering Hater stress affecting lb bean genotypes. A tendency 01 genot¡·pes that accumulated stem dry .... tter ei ther under stress 01 check condi tions to produce signi fi cantl y reduced )'ields Has ooserved. On the other hand, genot¡]les which rembilized stem reserves produced superior yeild, but there are exceptions, and the response is not clear under the post Ilowering stress.

Vig. ?E
13.
". I I U6
I 1.06 • ! ~
! •• I ~ e.
,.
• ,o
- 56 -
o
• o
'"" 04366 + O O141 .• 'coleno ~
• R .... 72n
• &Al·es
15 20
R_dfcl. O'owt" intüb!UOI1 t%)
pfG~·8b •• j
lO 3.
linear regression 01 the radicle growth inhibition caused by rES (-8 bar) 00 the fisher drought susceptibility iode~ 01 25 bean genotypes, Chillan. Saurre: Izquierdo and Huepe, 1986, unpublished.
3. Drought Tolerance Through Adaptatian to Canditians Associated Hith Drought:
Mast discus,ioos 01 drought tolerance do not coosider the illlportance 01 charaderhtics Ilhich, Ilhile perhaps not directly related to drought, are potentially essential cOlllpanents 01 drought tolerant bean varieties. Drought in bean groHing areas is oHen <lssoci.ted wUh higher telilperature cooditioos, and because 01 I¡mit.tians on transpir.tion, plants under stress mar ~perience unusually high leal telllper,¡tures ¡Figure 3Q). Thus high tesper,¡ture tolerance _hanisllS sy be needed in drought areas.
lúny regians in tropical areas combine poor soil fertility with drought, and lill,¡t,¡tioo. 00
iIIOve!il?nt al water wi!! also reauce nutriEilt availability. Genotvpes Ilhich Me especially efficient undet poor fertílitr may be advantageous. lt has oIten been speculated that lII,¡terials tnlerant to paor soíl fertility shoold have greater root 9rOl<th, and thus ooe .igbt ~pect uteríals tolerant to inlertile saOs to perlor. well under drought coodítioos in si.i1ar snes. However, in a cOIIIparisoo 01 2 drought tolerant lines and B materials selected lor tolerance to acid so!!s, the drought tolerant •• terial. were superior fT.ible 27).
Nitrogen liKatioo i. reduced under drought stress ILaing el al., 1984!. In .oybeans, fiKatioo a.ppeMed to be IIOfe sensitive to drought stress than Has photosynthesis llIeisz l:!.lli, 1995), suggesting to,¡t some drought yield reductians fiay reflect nitrogen eflec!s.
Ilrought environments where lillited irrigatioos are practiced .ay c.!.Use proble!ls 01 soil salinitt·, Although salinity C'¡U5eS what is physiologically a drought sítuatioo ISBe sectian 00 salt
tolerance!, additiooal probless are c,¡used by todc Of deliciency sHeet. 01 cer!ain ioos. Thus droug/lt toler_e under saline tonditioos uy require other ada.ptatioos.

- 57 -
Finally, certaln disease5 and pests are particularly probleaatic In drought situations. Ferhaps tlle II!05t notable is Ilacr!Jl!hoaina sol.mi whích only becO«oes problell!atie under drought conditíoos rScnwartz, et al., 1978). 5creening lor drought toleraRee and llacrophoalna tolerance at CIAT have produced reNrkably siailar resulh (Abawi and Pastor-Corrales, persooal clltMllUlicationl,.md it is speculated that this reHects toMOO to!eranee lllechanlsls lor tlle twa probleas.
Fig. 29
A OS
H 7
-------- lO·:;-3.' X ,
21 \J ..
" l.. l' ,....,ItAr •• 1
2'.23'5 r .10 '. o • 10 .. 2O 2 • :lO ,. !MP·'. ! .- OOR IU
~ 158Af-»t 8- KAJUO
" • 7 __
~ , , • Orf .. ·INtA. ... , 1: EMP ea I
I tOc "·2'31 I , "tlAf·5f
3.' L.J .. U:XAN.11J
13:~. Slnt ...
... · ... ·l" 2,1 tl5' Oomkwlo
~i 11. $w.., v.u.y , • u ,
... eH ...... ~....,.., o •. .....,.,itIW
.. O ~ HIohKC'~iofo 1M 10w y;.1d
20 ,. 30
e. !h.mdrywt fQ/pl1l"'U
Quadrant analysis 01 yield and ste. dry weight changes during tlle reproductive grawth perlod 01 lb beao genotypes under: Al irrígated and SI postfloweríng water stress. Chillan, 1984. Source: flernandel, 1'184.

Fig. 30
- 58 -
3.
,.
,. -- -- IUag.l~ pIO'" atllin ..
_ ... - OfOu0hl9AT 411 •• "U'AT 85
- - Otought, A10 lH\d SAl 1 %2.
Air and loliage temperatures far loor bun lines grOH!! under drought and írrigated condítions. Sourre: Sponchíado, I~B5.
Table 27. Coeparison 01 yields unda- drought stress in acid soils lar 9 línes tolerant to acid soils, and 2 lines loleranl to drought. Santander de Quilichao, ColOlbía.
Une Yield
Tolerant to acid soíls: A 257 A 254 Carioca YA 67 NAlibO S 4830 BAr 1297 A 440
llrought toleranh BAl 477 S 5059
l1ean S.E.
862 843 825 BIS 815 812 1110 713
1096 870
846 11

- 59 -
4. Cooclusioo:
Tbe reader eay con~lude froe this review that drought toleranee is too complieated to lerit study. He would argue that while the complexity 01 effects of drought has ell.inated siaple 50lutioos, the prospects far íaproving yield undar drought are 9000. Illlf3rol'ements in bean crap productivity can be m.de ooly through two basie approaches: 1) by modífying plant genotypes to fit their envirOOlllef1t better ¡ and 2) by ~hangyng the environment to ameliarale the eHects of enviroomental stresses. To over~oee the cOIlstraints in both approaehes, agrOllOillists, breeder, physiologysts and genetic engineers require a dear undershnding of the physiological basis underlying rield and stress toleranee.
Studies 011 invidual eechanisMs ~ay be integrated by simulatiOll models which can predict elfscts of changes in climate and soil t)'pe. Far exalllflle, Janes aod Zur m84l used aynamie sieulatioos 01 sorbeao growth to test the relatiye importance 01 three mechanisms, 05motie adiustment, íncriased root growth, ana increased stOlatal resistanee at full turgor, under elay ana sandr soils concluded that greater root growth Has the IlIOst eHecti ve meehaní .m, wI1i le increased stO!!atal resi Stance always resulted in reduced photosynthesis.
B. Exes.s Soi I Water
Heavy rains oHen lead to 50il lloOOing ¡water in e~cess of {jeld capacit!'¡, especially in poorly drained soi I s. Be'll15 are sensiti ve to such cOIlai tiOlls, ana yield redudiOlls are frequentl y greater than OIle might e~pect. Thi5 can be attributed to the "earlr i_diate adverse eHeds on plant growth. Wadlllan-l'aIl Schravendiil: ana van Andel m8S) faund that leal ana rrot growth halted within 2 days 01 HoOOing, tnis being associated with asevere drap in transpiration, but a riie in leal AB~ concentration.
Tbe cause of damage ¡rOl inundatiOll is directly related to redueed gas diflusiOll in floOOed soils, resulting in depletion 01 available O. and accu.ulation 01 CO. ano ethylene in the rout zene. Aceompao~ing this is an inhibition 01 respiration, partial loss DI rout function, and possibly,
accumulation 01 to~ins due to blodage 01 respiratioo or activation 01 alternate, an,erobie pro-:esses. Higher soíl tl!lllf3eratures, frequently lound in tropical regiOlls, Ciln eKilCerbate probl!!1ltS of gas di Husien due to reduced solubílity 01 gases and greater respifiltary acti vi ty (Forsythe et al., 1979). Probl~5 with root rols are olten ilssociated with Ilouding.
True toleraoee mecnanisms have oot been ideotified in beans, but as in other eraps, recovery Irom inundatiOlli is re1ated to ability to pruduee oeH ad,entitíous roots (K,hn et al., 1995), lo trials at CIAT, floOOing by maintenanee of the water table to within 7 c. al the tap 01 beds resulted in an average of approxill\atel y 5IlX yieló redudi 011 , but coosiderable variet.1 diflerences were founiJ (Figure 311.
C. Salina Soil5
!lean. ara coo~idered saa sensiti ve, although F'h.seolus eocdneus has beeo elassi lied as intereedi.te between lOderately tolerant and moderately sensitiye legu.e spe~ies (Lauchli, 19H4). Problems in this crop a •• oci.ted with salinity are generally graate.t in arid reglans under irrlgatíoo, but localized areas 01 satine soils are found in many l!iesic environments.
!m~iate ellects of excess salinity inelude reduced plant growth, especially lea! area, and burning 01 leal marglns. These mal' relled three undE'flying proe".se.:
l. Physiological drought due to low os..atlc potenial; (high solute coocentrations).

- 60 -
2. Nutrient imb.lances due to relatívely high concentrations 01 specilic elements, especially sodiua, ínhibiting uptake 01 others.
3. TQl(ic eHect. of specific ions, particularly chloride and sodiu ••
E.idence lar an osmotic eflec! cotes frcm deeonstrations that high relative hulídity reduces stress ellec!s, and that leal relatív!! water content is reduced ~arkedly under stress (Ayoub and lshag, 1974). Aroub and lshag (1974) also detected reductions 01 calciua and potassium cation5 under Eooditions of high sodiua, but noted tbat tne redudions did not re.eh levels thought to produce deficiency syeptoms. High leveh of cMoride in bean leave; _e associated wHlt reduced photosynthesis, stomatal cooductance, and RuBP carboxylase efficiency iSeEfflan and Critchley, (985). These and si.ilar findings sU9ge.t tbat eflects of ;alinity are prohably multiple,and it would be a aistake to eaphasize a single mechanisl.
Fig. 31
• •
• • ~ l $ ~
~ "''''' ~ • •• l! • ~
. " • • •
.000 2000 "00
Control yieJd ttt. ha")
Yields 01 13 oean cultivars gr_ under flooded cootrol field conditioos. CIAT-f'aJ¡¡¡ira, 1976.
lit eoderate leve!; of .;llt tolerante oeans do show have been astríbed to their abilíty to accumulate sodiu. ions in root parenchyaa vacuoles. 5Dme soyheans appeaf ta coehine this IeChanjs! with an ability to eMclude E!KteSS chlaride from .hoots, and this eight be a promising avenue for increa.ing salt tolerance in l'Itaseolus vulgari. (Lauchli, 1m). General recOfiilil!fldatioos iar screening for salt tolerante in all craps were given by Greenway and Hunns 09001. S.íIlie and Nott (1982) have descrlbed a chloraphyll flourescence as a rapid test for salt toleranee, they usiog bean as a susceptible check.
Studies of genetic variati~l far salt tolerance among l'Itaseolus vulgari. cultivar. are rareo HoHler (personal eommuníeatioo! detected variet.l differentes under field conditions, but noted that variabilíty in salinity under field conditions lIIa~_es large scale screening dilficult. l!ebouck Ip!!rsooal cOilHIlI1nication) collected }and raees of oeans rrom epparently saline soU. in llei<ico.
¡¡;¡ !!!!!!! -= = -¡¡;¡ -¡¡;¡
=

- 61
Several investigators have reported the selection of salt-tolerant cell lines in SHeet pepper, 5ugar can;¡. callee, rice and taro. However, regef1eratioo of plants frOl!! salt toleran! ce!!s have beeo liaíted io these crops, ami has apparef1tly never beeo attea;pted w!th heans.
D. 6eiteraU~ed Stress Toleranc,,: "Rusticity'
It ís frequeotly c~ted tbat Nhile tradi!íonal beao cultivars lay lack specil¡c díseas" or other stress resistanc:es, they alteo outperlor. supposedly 'ílIlproved' ateríals under far. eondítions.
Alfara 11981) concluded froe 12~ bean cultivar trials at SO locations and 20 countries that lhe yields of 'Jaaapa' and 'Porrillo Sintetico·. used as standard checks of tnown adaptability, were nol frequeotly surpassed by di verse _teria!;. However I lhe 1011 values of tbe stability index lb! for ev. Porrillo Siotetico in tbe analyses 01 VO¡'sest and Sarcia (1984), Stiggest that this cultiviI/" perfores relatively better in poor sites as compared to sieilarly yieIding cvs. Jaaapa and ICA rijao (Table 28). lOH b valties are indícative of relatívely goOO yield in poor eoviron_ts and lael; of response in den sites. el'. Porrillo Sintetico stanas out coosistantly lor lower b values for si.ilar eean yielas in a given IBYAN. Sinee stresses varied greatly among .ites in the IUYAN's. the better perforaance af cv. Forrillo Sintetico in poor siles presw.ably reflected S~ generalized stress tolerance ar 'rusticity' rather than resistance to a few specific'stresses andlar disea;!!s.
Other examples al rusticity COfteS ¡rOl studies of cultivar response to artificial defoliatioo. In a comparison 01 17 tultivars receiving a 60Z deloliation at 10 days frOl planling el'. Carinca adually increased yield under toe defoliation stress Nhile !ine EIi' B4 shOHed a earl:ed yield reduction IFig. 321. This would suggest that el'. Cariaca would .. intain its yield under a wide range uf stress!!s alfecting leaf growth, a conelusion coosi;tent with cv. Carioca'; popularily in BraziL
rabIe 28. Coeparison of regression response lb) and Mean yields across sites 01 lhree cultivars used as checks in International Sean Vield and Adaptation tlurseries ffOll 197b to 1981. Data ¡fOlll Voysest and Garcia 119841.
lt:J.of Porrillo Sintetico _Jaapa_ JeA Pij"o_ Year tríals b Yield b Yield b Yield
I:g ha-' kg ha-' ~g h-'
1976 S4 1),77 1539 1.02 1572 0.79 15b6 m7 31 0.00 1623 1.09 1627 1978 33 0.94 1486 I.Q6 1498 1.17 152'1 1979 32 Q.SO 1501 1.06 1579 1.06 1554 1980 49 0.95 1584 1.06 Ib5b 1.06 Ib37 1991 26 0.99 15bS t. 01 1723
/lean 37 O.Sé 1550 1.05 lblO 1.02 1570

- 62 -
Calculatíng a generalized stress yield as the geometric lean 01 yields under 331 defoliation and under post n_ríng drought, data ¡rOil Hernandez (I'1!l4) show a 1011 correlation between stress and control yíelds (Fig. 331. Ihe contrast between A 231 and Orleo INIA ii partícularly striting with both línes yielding about the saae in control plots, but di Hering by !lOO I:g ha-' under stress. The cOMparison 01 (v •• Porrillo Sintetico and Jaaapa is also instructiva, Hith JaAapa outyielding Porrillo Sintetico by !lOO kg ha-' in the control, but being slightly inferior under stress.
1:1 2200
".
2 o
C#lRIOCA • " g • • JI • i .800 • t • •
" • . -• '" • •
1400 lBOO 2200
Hg. 32 Relativa yield perlorml/lce 01 17 bean Iínes under 6Q1 deloliation atJl.l days aHer planting. CIAT-Palair., 1993.
Possible characteristics which .ight conler rusticity inelude:
l. Abilíty to produce recuperative growth, presulably resulting (rOl relObilízation 01 carbohydrate Of nitrogen reserves and an indetermínate growth habito
2. 600d competitive ability. 3. High tissue roncentrations 01 compounds sueh as phenolics with ínhibitory eflect. DO a broad
range of pathogens ar pests arganises. 4. Greater partitioniog to root growth. 5. !!uf/er ability lar adequate ped retention and seed lílling.
Earlíness also might be considered a MeChaní5a since it per.its escape 01 Iany,tresies. Vata (roa the 197b ISYAN supported lhis. For ,ítes Hilhin JOo of the Equator (lhus eliminating trials Hitn severe photoperíod e((ectsl, early maturing cultivar. had signilícantly lower regressiDO response DO site mean yield lb parameter), índieating that they yielded relatively belter as compared to late cultivar. al the poorer sites IFig. 34J. Thal the regression response HaS not signifieantly correlated Hith yield (r = Q.Z21 deaonstrated that low values 01 b in early cultivar! did nat sieply rellect poor yields across sites IWhite, 19941.

Fig. 33
Fig. 34
a o .. i ¡ ¡ ::!
2700 y-sa3 +O .• '6.X R .... 0.11:-
2«lO • • • Pau,lta S""ttfieo
21""
• • 1800 • • >100
- 63 -
• A231
• •
•
• Qtf." INtA
3000 3300
Comparison 01 stres! and irrigated control yield on lb bean lines. stress yield Is geoeetric ~!ll 01 deloliation and drought treatments. Sased on data ffOll Hernandez I
1'I!l4.
• y ft ·1.43'" O.0307.X •
••
1.0
•
08
•
" 78 .0 ••
Relationship beb;een lII!an days to aaturity and regression response 01 yield lbl lar 20 cultivars 01 the 1976 leVAN. Source, White, 1'1!l4.

- 64 -
Nhile there is evidence suggestive DI diflerences in 'rustlclty' DI bean cultivars, it is diHicult lo decide Hhether such a characleristic is best sought by selecting under a .oolnte level DI various stresses, or identifying specilic characteristics related lo rusticity. EHects DI local adaptation could conf_d results unless such _k Is restricted ta speciHc sites.

- 6S -
V. C[KlU5I~
Crop physiology is oft!!fl criticized far the lad of tangible benefit. it has pnl'Yided to larllen, thi. crititis. certainly extending to research on the COlllon bean. Hhile in SoIe cases critieis. Has
invited by ayer entousiastic prOlises about the utility af certain lines af work, we would argue th!t lhe coaplexity 01 interactions 01 physiological processes preclude. rapid solulions. It is seldOl possible to reaeh fir. conclusions about a giv!!fl set al physialogical processes, hecause eath proces. interatt. HitO others. The evolution of our lnowledge of physiological processes is perhaps best W:ened to the aanufacture of a cOt1lplex ~hine. Although great strides llay have heen lIade in constructing our physiology .achine, and it lIay eY!!fl have soo-units which Seelt to functioo well, 001 Y as the lIachine nears cOlljlleUan wilI it approaeh its int!!flded level of produdivity. EV!!fl tllen, if 501!! parts were poorly aanufactured, it IlaY oot .eet expectations until tne Ilaws are identified and [arrected. lIe would argu!! that patience and care are needed to assure that the ph)'siology 1I.!Chím, is well built. As in any eonstruction project, priarities can and should be seto Nitrogen reQ¡ilizatian in relatían to leaf s!'neseence and seed growth is amoog the area where oor understanding is mast deficient. In cOIparisan to photosynthesis, respiratian is .llOst untouched, yet the few studies in other craps indicate great potential far increasing yields. And in all research, greater paios should be taken to assure lhat candi Hans used are relevant to HIlat My accor in bean fields, nol laboratary beoches or greenhooses. .


67 -
REFERENCES
- Abernathy R.a. Palier, R. Shibles and I.C. Anderson. 1977. Histological observatioos on abscissing and retained soybean flowers. Canadian J. Plant Sei. 57:713-7Ib.
- Adaas, M.N. 19b7. Basis 01 yield component compensatioo in crap plants with special references to toe lield bean, Ftaseolus vulgaris. Crap Sei. 71505-510.
- ---- 1973. Plant architecture and physiologital ef/iciency in toe field bean. p. 266-278. In Potentials DI lield beans and otoer load leguoes in latin Amerita. CIAT, Cali, Colofibia.
- ---- 1975. PI.nt architecture and yield in toe grain leguffles. p. I-Ib. In Report 01 the TAe working groop on the biology 01 yield 01 grain legulIl!!. Pub. No. DORRIllill175/2, FIlO, Rome.
- ---- 1981. updated new bein architype. Hichigan Dry Bean Dig. 5(2):12-13.
- ----, J.V. Hiers.a and Julio Salazar. 1978. Di/lerences in stareh accumulation among dry bean cultivars. (rop 5ei. 19:155-157.
- Aguilar M, 1, R.A. Fisher and J. Kooashi. 1977. Elfeüs 01 plant density and thinniog on high-yielding dry beans 1 Phaseolus 'lUlgaris ) in Mexica. Experimental Agriculture 11:325-335.
- Alfara, R. 1'191. Adaptabilidade e est.sbilidade de cOlllportamenta de liohagens e cultivares de fiejao preto l Phaseolus vulgaris L.l el! 124 ensaias internacionales. M.Se. thesis. Vi con, Minu Gerais, Brasil.
- Aharez, tI.tl., P.D. Ascher aod D.II. Vavis. 1981. Interspecific h)'tJrídízation in Euphaseolus through !!ftIhryo culture. fIort. Sci. 161541-543.
- Ariyanayagae, R.P. 198(J. rigeon pea breeding in the Caribbean F~ional Progra~e. Vol. 2. 415-426. In Praceedings DI the ioternatiroal worl:shop on pigeon peas. ICRISAT \ rahncheru, Htderabad, India.
- Ashley, J.II. 1'194. Groundnut. p. 453-494. In "./l. Goldsworthy and !I.H. Fisner (eds.). The physiology al tropical field (rap.. Joon Hile;' ~ Sons; New York.
- Aspinall, D. stress. p. 155-172. te~erature stress.
1990. Role of abscissic acid and other hormones in adaptatíon to water In N.C. Turner and r.J. ~n_ (eds. l. Ad!ptatioo 01 plants to water ill1d higo Joon Wiley & Sons; New Yarl.
- Austin, G.B. and M.S.N. Macle!n. 1972.. 5oIIl!! ellects 01 te!perature on the rates of photosynthesis and respiration of Philseolus vulgaris L •• Fhoto5ynthetica 6:41-50.
- ---- and ---- 1972b. A methnd far sereening Phasealus genatypes lor tolerance to low temperature5. J. Hart. 5ci. 47:279-290.
- Ayoub, A. T. and H.M. 15hag. 1974. (Fhaseolus vulgaris L.) J. Agr. 5ci., Cambo
Sodiu. toxicity and cation iMbalance 111 in dry beaos S2:339-342.
- Alcon-9ieto, J., H. laMbers ilnd V.A. Day. 1983. Re5piratory praperties 01 develaping be3n and pea leaves. Australian J. Plan! Phrsiol. 19:237-245.

- 68 -
- Bashaw, Le, 1'1SQ, Apomhis <lIld its applieation in crap illljlrovement, p, 45-1>3, In fI,R, Fehr <lIld H,H, Hadley leds.). Hyúridizat!on of crop plan!s. Amer. Soc. Agron.; Madison.
- Benfll'tt, J.P., M.N. AddlllS, and C. Ilurga. 19n. rüd yield cOlllpooent variatioo dnd intercorrelations in Phaseolus .ulqaris l. as affected by planting density. trap Se!. 17¡13-75.
- Binkley, A.M. 1932. fhe a~t of blossom and pod drop in si, varieties of graden heans. Pro-:. Amer. Soco Hart. Sci. 29:489-492.
- Blad, C.[" l.D. Soldstein, LB. Ilay, V.f'. Kestler and a.c. /1ayne. 1976. The relatíonship 01 pl_nt metabolism to internal leal and cell morphology and to the effieiency 01 COz assimilatioo. p. 113-139. In R.H. BUfrís ~ e.c. Blad (eds.!. ca. metabolís. and plant productrviti'
Uni versity Park f'ress; Balti_e.
- Bonanno, A.R. ar.d H.J. Mad. 19E3. Water relations illlO growth 01 snap oeans as in influen[ed by dífferentiel irrigation. J. Amer. 50e, Hort. Sei. 108:837-844.
- Brouwer, R. !9b2a. Distributions 01 dry matter in the planto 10:361-376.
Netherhnds J. Agr. 5cí.
1962b. Nutritive influences on the distribution 01 dry matter in the planto Netherlands J. Agr. Seí. !O:399-4ll!l.
- ---- 1963. Tne influence 01 the su[tian tensioo 01 the nutrient solutians on growtn, transpiration ano diffusion pressure defiei t 01 bean leaves ( Fllaseolus ~l1lgaris ). Ada Bot. tleer. 12: 24B-260.
- Ilunee, J.A. 1979. EHeds 01 water stress on leaf e"pa.nsion, net photosynthesis, <lIld vegetati ve growth 01 sOI'beans and eottan. (anadian J. Bot. Sb:1492-14'1!!.
- llurga, C.A. 1978. Canapy arehitecture, llght dístribution, and photosynthesis 01 dífferent dry bean (f'ha;¡eoll§ vulgari. l.) plant types. Ph.D. Tnesi.. Miehigan State University, East lansing.
- (armi, A. ana D. Koller. 1978. Ellects of the roots DO the rate of photosynthesis in prioary leaves 01 hean ( F'haseolus vulgarti l.!. F'hotosynthetica 12:17B-184.
- ----, J.D. H!;sleth, ti. T. Enos, and D.e. Petees. 1'1!!3. Interrel<ltionships between .hooe growth and photosynthesis, as .Heded by root growth restriction. F'hatosrllthetica 17:240-245.
- ---- ano l. Shomer. 1979. 5tarch accUflulabon and phatosynthetie activity in prillary leaves of hean I Phaseolus vulgar!s l,). Ann. Bot. 44:479-484.
- earr, V.J. and K.G./!. Skene, 19M. Diau~ie growto curves of óeeos HitO specíal reference to lrench beans ( Phaseolus vulgaris l.). Australian J. Biol. Sei. 14:1-12.
- [handl sr , R. F. 1969. PI ant morphology and stand geometry in relation lo ni tro!Jl!ll. p. 265-299. In J.D. Eastin, et al. (eds.). rhysiological aspects 01 crap yield. ASA, CSSAJ Madison, Miseon.in.
- Chanar. Babu, R., P.S. Brinivasan, N. Relationship Det_sen lea! photo5ynthetic rate and genotrpes. Photosynthetiea 19: 159-lb~.
Natarajaratnam and S.R. Bree Rangasamy. ¡g¡j5. yield in blae~gram (Vigo. 1Uflg9 (l) Hepper)

- 69 -
- Charles-Edwards, O.A. 19B2. Physíolagícal deter.in'fits of Erop growth. Ikademic Fr.?:;;; Sydney. lb! pp.
- [hattertan, N.J. ana J.E. Silvius. 1979. Fnotüsyoth.te partitianing into st.reh io sOrhedn leaves. Plaot F'hysiol. 64:749-753.
Coheo, D. 1971. Haximizing final yield .nen growth is lhlited by tiae or by liliting resources. J. Theor. Bia!' 33:299-1')7.
- ColQuhoun, A.J. and J.R. Hillman. 1975. Endagenous ahseisie aeid and the seneseence ef ¡eaves 01 poaseolu. vulgaris l. Z. PIlanzehpysio!. 76'326-332.
- Cordner, H.B. 1931. Externa! and internal factors aflecting blossom drop dnd set 01 paos in lima bean. F'roc. Ilffler. Soc. Hart. Sci. 30:571-571:.
- Cory, C., and B.D. I/roster. 1984. Ass€sslft€!1t Df drought tolerance in cultivars al po.seolu; vulgaris aná !'... acutifolius. Hart. Sd. 19 (3, Sed. W:b8.
- Crocomo, O.J., \I/.R. Sharp and J.E. Peters. 1976. Plant morphogenesis and the cantrol 01 calltis growth and root inductian 01 Fh.seolus vulgaris with the .ddition of be.n seea e"tr.ct. Z. Pfl anzenphysi 01. 7B: 451:-4bol.
- CrookstDn, R.K. and J. O'Toole, and J.L. Q,hun. 1974.. Characterizatian 01 the hean pod as • photo.ynthetic Ofgan. Crop Scí. 14:708,,712.
- Cutler, 3.M., D.W. Rains and R.S. lonmis. 1977. Tne importance 01 cell size in the water rel.Uans 01 plants.l'I1ysiol. Plant 40:255-260.
- D3vis, S.O. and J.K. MeCree. 1978. Ff1otosynthetic rate and diHusioo condudance as • funetion 01 age in leaves 01 bean pl.nts. Crop Sei.IB:~)-282.
- D.ynard, LS., J.14. Tanner and W.G. !)unean. 1971. !)uratioo 01 toe graín filling periad and its re.etian to grain yíeld in corn Zea mays L. Crop Sei. 11:45-4B.
- De Maura, R.L. and K.W. Faster. 1986. Spatial distrihulion 01 seed yield within plants of hean. Crap 5ei. 2b,337-341.
- Denis, J.C. and M.W. Adam.. 1978. A factor analysis of plant vilfiables rel.ted to yield in dry besos. l. Morphological traits. Crap Sei. 18:74-78.
- Don_Id, C.I1. 1%9. The breeding 01 crop ideotl'pes. Euphytica 17:385-403.
- Duarte, R.A. and M.W. Adams. 1972. A path coeffieient analy.i. 01 some yield component interrelalions in field beans { fflaseolus vul~arís ¡ trap Sei. 12,57~-582.
- !)oJflLan, W.G. 1980. Maize, p. 23-50. In L. T. E,ans ledo i. Erúp pnysiúlagy: SOfliE case histories. Cambridge Universíty F~ess; England.
- ----, n.E. MeCloud, fU. MeGraw and K.J. Boate. 1979. F'I1ysiological aspeds al peanut yíeld improv!!lI!!!nt. Crup Sei. 18: WIS-W20.
- !)ur., loS. 1975. 5eed For"atioo. Ano. Re,. F'lant Phl'sial. 2b:Z!W-278.

- 70 -
- Ebercan, A., A. Blum and N.R. Jordan. 1977. A rapid colorimetric melhod lor epicuticular wax conlent 01 sorghum ¡eaves. Crop 5ci. 17:179-180.
- Egli, D.B. ana J.t. Leggett. 1973. Dry matter accumulation patterns in determínate and indeterlinale 5lJybeans. Crop Sei. 13:220-222.
- ---- and --- 1976. Rate 01 dr\' mattEr ¡¡(cumulaban in soybeao SEedS source-sink ratios. Agron. J. b8:171-374.
Hi th \arying
- Ehleringher, J.R. 1980. leal morphology and reHertanc. in relation to r.ater ana teill!lerature stress. p. 295-309. In N.C. Turner and r.J. Wraffier (eas.l. Aaaptation 01 plants to water and high temperature stress. John Hiley ~ Son.; New York.
- ---- and O. Bjorl:iun. 1978. A cOll!parison 01 photosynthetic characteristics of Encelia species possessing glabrous aoó pubescent leaves. Plant Fhysíol. 12:185-191).
- El-Belhgy, A.S. dnd M.S. Hall. 1975. Studies 011 endog!mous lel'els 01 etnrlne and auxin in Vicia faba lo during grQ"tn and development. !le. fhyto!. 75:215-224.
- EI-Shar'."ay, M., J.H. Cad, afió A.A. Held K. 1984. Water use efficienq; al Cass3"3. lI. Oifleriog sensitívity 01 stomata lo air humidíty in cassava.n other waf!-climate spe~ies. Crop 5cí. 24:S0Hi07.
- EngledOli, F.L. 1925. Investigations DO yieló in (he cefeal S. 1I. A spacíng e,periment Hitn wneat. J. Agríe. Sei., Cambo 15:125-14b.
- Escalante, J.A.S. 198(1. Efedo de sOtllbreado artificial sobre el rendimiento y sus cOll!ponet1tes en Iríjol (ff¡~setllus vulgaris varo MichoacAn 12-11-3i. M.Se. thesis. Chapingo, rM¡'¡co, Colegio de Post-graduados. 34 pp.
- Evans, lo T. 1175. 8e)'ond photosynthesis---the mle 01 respiratíoo, transloeatian aod grOlith potential in deteraíning productivity. p. 501-507. In fhotosynthesís and produetivity in different enviraneents. Ed. by J.P. Coopero Cambridge University Fress; Cambridge.
- ----, R.l. Dunstone, H.H. Rawsoo and R.F. Williams. 1?70. The ph¡~m 01 toe wneat stem in relatian to requirements lar assimilate by the ear. Australian J. Biol. 5eí. 23:743-752.
- Fakorede, M.A.B. and j.J. Mock. 1978. Nitrate reauctase activity and graio yield 01 aai<E cultivar hibrids. Crop !leí. 18:bOO-b94.
- Fa.njul, L., J. Kohashí S., ano E. Hernánoez C. 1982. Yield potentia! and stratílied growth anal ¡si s 01 dO indetermínate climbing pole bean. E.pl. Ilj¡r. 19:1b7-175.
- Fisher, R.A. aod R. Mauer. 1978. Drought resistance in spring wneat cultivars. l. Grain yield respOfisEs. Australia" J. Agr. Res. 29:897-
- Farsythe, W.Il., A. \lictor a"d M. 6óme,. 1979. requirements 01 Fhaseolus vulgaris l. p. 205-214. Phy·sic31 properties and trop produdion in the tropics.
F!ooding toleranee and surfaee drainage In R. lal and O.J. Greenland leds.). Soil
- Franco, A.A., J.C. F'ereir. ~nd C.A. Neyra. 1979. Seasonal patterns of nitrate reductase and nitrogenase adi viti es in Phaseolus vul gar i s l. Plant Fir¡siol. 63:421 -424.

- 71 -
- FraSff, O.E. ad" R.S.S. Bidwell. 1974. PhDtDsynthesis and photarespiratiOll during the ontogeny ni lhe bean planL Canadian J. BoL 52:2561-2570.
- Frota, J./U. dIlrl T.C. Tucker. 1978. Salt and l/~ter stress influences nitrogen fl!'tabo!isffl in Red !<idn~y Beans. Soil Sd. AliIff. J. 42:743-746.
- Saastra, P. 1959. Photosynthesis 01 crup plants as inlluencOO by light, carboo dinxide, temperature, ana stomatal dillusiOll resist_nce. Medelingen landbouw, Wageningen 59:1-6B.
- ---- 1%2. Photosyllthesis of leaves and Held crops. Netherl.nds J. Agr. Sci. 10:311-324.
- Gallaher, J., 1'. Biscoe and R. Sentt. 1975. Barley and its !!!1virOllment. \l. Stability DI graio weight. The Journal 01 I\ppliOO tcology 12:119-336.
- Sardiner, T.R., D.M. Vietor and L.E. Cra¡er. 1979. Growth habit and row width elleds Q/1
leaf ar2. developffient and líght interceptíon of líeld beans. Canadian J. f'lant ,cí. 59: 1~1-199.
- Gales, r. 1. 1969. Water deficits and growth of hefo.tecHs plants. p. 135-190. In 1. T. KozloHsU (ed.). Water deficits and plant growth. 11. Plant water consu.1ljltion and respoosE. AcadEd!íc Pfe"j NetI Yo"';:.
- Gates, D.M. 1768. Transpíratioo and leal temperature. p.211-23S. In L. Hacoll" W. Sríggs dIld R. F'ark IOOs.). Ann. Rev. Fhnt Physiol.
- G3y, S., D.B. Egli and D.A. ReicQs~y. 1900. Physíologieal aspeds 01 i'ield i"'prOl'aeot in sorbeans. Agron. J. 72:387-391.
- Gepts, P., LC. G5oorn, r. Rashka and r.A. Dli;s. 1'194. Phasoolin variability in the eDilltT.oQfl bean (F'haseolus vulgaris l. 11 E~idence lar independent domesticatioo cEflters in I1esoamerin and the Andes. Ano. Rept. Bean llllprov. Coo¡¡. 27:62-64.
- Gifford, P .H. and lo T Evan;. 1'191. F11Otosynthesi 5, tarDOil parU Uaning and yield. Ann. Re" PI ant F'hy·siol.. 32: 4B5-50!j'.
- Graham, P.N. 1978. Some problems and potentials of ¡ield beans IPhaseolus yulgaris l.) in Latín A,~eriea. Field Crops Res. 1:295-317.
- Green way, H. and R. Hunns. 1980. Mechani5MS DI salt toleranee in nonhalophytes. Ann. P.e •• f'lant Physiol.31: 149-190.
- Gutiérre" J.A. and S.P. Singh. 1985. Heteros!s and inbreeding depression in dry bush bean., Phasealus vulgaris lo Canadian J. F'lant 5d, 65:243-249.
- Gu,'er, R.S. and A. t'ramer. 1952. Studies of factor s affediny the qualit)" ol grEEn ano wax beans. Meryland. Agr. E,p. Sta. Bull. Abe: H2.
- Ha.id, l.A. 3m! J.E. Gralius. 1978. Developmental allomelr)' and ils í¡¡¡¡¡licatioos to grain yield in barley. Crop SeL 18:83-86.
ft¡,-terlein, A.J., C.V. Clayberg, anó I.D. Teave. palien grain viabilityand palien tube growth in lhe !!ort. Se!. 11)5:12-14.
1980. ¡nfluence 01 high telllperature ..... style ol F'haseolus vulgar!s L. J. Amer. Soc.

- 72 -
- Herssen, J.S. In. 1 98Q. Breeding lar apomixis in patato: Fursuing a utopían scheme. Euph\,tkl 2?:595-b07.
- Hern~ndez M., Maria H. 1984. Estudio de la relación entre re~ílización de asi.ilados y rendillliento de Ireiol negro Whaseolus vulgaris L.), bajo coodiciooi!s sillluladas de "stre!,;" ambiental. lng.;\gr. thl!Sis. Universidad de Concepción, Chill!n, Chile. b2 pp.
- Hes~eth, J.U., n.N. Hoss. 1%3. Variatioo in the respoose 01 photos)'nthesis to líght. trop Scí. 3: 11)7-110.
Hesse, N. and F. LeAz. 1982. lnfluence 01 waler supply 0!1 transpiratioo afió fiel phatosynthetic ratl!S 01 clilllbing french bean ("haseolus yulqaris varo ,u!g3fís). Gartenbauwissenschalt 47:145-152.
- Hiebsch, C.K., E. T. Kan!!l1lasu and C.D. Nid:ell. 1976. EHects 01 soybean leaflet type 0!1 nel carban dioxide ef.change, water use, and water use-elliciency. Canadian J. PI_nt Sei. 56:455-458.
- fllher, S.C. and O.M. 15rael. 1982. Biacnelllital hasis lar partitioning 01 photosynthetically Ihed carbon between starch and sucrose in sOrbean (6lydne !\!2!l Merr.) leaves. Plant PhysiaI. 69:691-696.
- Huepe, S. 1986. Indices de tolerancia a sequia en freíol IFnaseolus vulgaris l.) lng. Ágr. Ine.is, Universidad de Concepción, ChillAn, 6S pp.
- Hunter, W.J., e.J. Fahring, S.R. m.en and L-K. Porter. 1982. loutíon 01 ni trate reductase in diflerent sOrbean cultivars. trop Sei. 22:944-948.
- Iwanag., M. 1~83. Cn!!l1lical indudioo 01 aposporOU$ opo.!lictic seed produdioo. p. 11)4-11)5. In W.J. Hooker (ed.!. Researcn lar tbe patato in tne year 21~)Q. CIP; Litu, Perú.
- Izhar, S. and D.H. Nallace. 1967. Studies 01 the physiolagical basis far yield di/lerence •• 111. 6enetic variation in pnatosynthetic elllciency 01 f'haseolus vulgaris l. Crop Sci. 7:457-460.
- Izquierdo, J. 1981a. Tne ellett of arcumulation and r~ilization 01 carban .ssi.ilate and nitrogen on abscission, seed development, and yield 01 common beans (f'haseolus vulgaris L.) with differing architedural larms. Ph.n. Thl!Sis, lIichigan 5tate University, East Lansing.
- ---- and J.M. HernAndez. 1984. Estudio de la relación entre r~ilízación de .. ilililados y rendi.iento de Iríjol negro (Phaseolus vulgaris L.) bajo condiciones simuladas de stress alllbiental. Universidad de ConcepcUn, Proyecto D. l. 202606, (unpublíshed), ChillAn, Chile.
- ---- and G.l. Has/ield. 1980a. Ethylene production ¡rom bean Ilowers and pods. Ann. Rept. Bean llllllrov. Coop. 21:96-99.
- ---- and ---- ¡?Blh. A collection receptacle lar field abscission studies in c~ beans (Fnaseolu5 ,ulgaris l.l. Crop Sei. 21:622-625.
- ---- and ---- 1983. The relationship 01 seed lilling to yield aiOng dry beans with dillering architectural lorms. J. Amer. Soc. Hort. Sei. ln.a:l0b-lll.
- ----, ----, M.N. AdalllS and M.A. Ueversa~. 198Ob. C-assisilate partitioning relalionship to reprodudive abscission and yield 01 dry bean (P. VUlgMÍ> L.). Siennial LooI. Sean l!!prov. Coop. and Nat. ~f)' Be.o Councíl Prac., Madison, Wisc., Nov.7-9, p. 81-89.

, ----, N.A. Uehersa, and W.M. Ad3ffiS.
soluble sol id. test 40r evaluating carbohydrate 1(2):145-152 (ISSN 0716-1bS9).
13 -
1985. fhe utility 01 the ,t¡¡reh iodine and partitioning in dry be"ns. Agro-Ciencia
---- and 6. Huepe. 1986. Valor de un nuevo indiee de tolerancia a sequía"" frijol. !j¡lversidad de Coocepción, Proyecto V. r. 2026~.f:, no publicado, Chill1n, ChO".
- Jaquiery, R. and E.~. Keller. 1 \'SO. lnfltienee of the distribution 01 .ssmiliate on pad set in faha be~n ¡Vicia faba l./. Fabis 2:34. ¡CMDA.
- Jara R., J.c. 1985. Evaluación de la le.!.peratura de la cubierta vegetal y otros parámetros bio!íslcos en frejol negro lFhaseolus vulgaris L.). M.5e. thesl;. !j¡iversidad de Concepción, ChillAn, Chile. 133 pp.
- ----, J. Izquierdo, and R. !latta. 1988. Respuesta fisiomorfológiciI y produdi va del frijol negro ¡Pnaseolus vulgarÍs) Irente al déficit hídrito. Agro-Ciencia 4W:2l-2b.
- Jennings, P.R. ana J. de Jesus Jr. 19b5. Studies an tOl/!¡letitian in rice. l. C01'Iljletitioo in mi.tures 01 varieties. E,'ülutían 22:119-124.
- JohnsOIl, H.W., H.A. Sorthwid, iIl1d R.O. Lellel. 1961)., E¡¡ids 01 phatoperiod and time 01 planting an rates 01 development 01 the soybean in variDUs states 01 the 1 if" c¡d". Sot. 6,2.
and J.W. Tanoler. 1972. Caltul.tian 01 the fate and duration 01 graiol filling in (w-n (lea mays l. l. Crap Sei. 12:4!l5-48b.
- Jones, D.B., M.L. Peterson ana S. 6enq. 1979. Asso[iati~l between grain filliog rate and duration and rield t01'lljlonents in rice. erap Sci. 19:MI-M4.
- Jooes, J.W. and 8. Zuro 1984. 5iffi'Jlation 01 po.siole adaptive ll!e':hanisffis in traps subjeded to water stress. !rrigation Sei. 5:251-264.
- Jordán, R. and A. ChillnaU. 19,33. Ilbservations on the fat iletalloloism 01 leaves. 11. Fah and phosphatides in toe 'Runner Sean' (Fhasealus IllUltillorus). Ann. Ilot. 47:163-¡lló.
- Kahn, B.A., P.J. Sto/ella, R.I'. 5andsted and R.W. Zabel. 1985. lnfluence 01 lloOOing on root morphological cOI/!¡lonents 01 young Dlack beans. J. Amer. 50c. Hort. Sei. 110(5):623-627.
- ¡(.plan, S.L and H.R. KoUer. lW4. VariatiOl1 i!iOOng sOrbean cultivars in s~d growth nte during the linear poase of seed growth. Crap Sei. 14:bI3-614.
- Kim, J.H. and D. Y. lee-St.delmann. 1984. W.ter relatiOl1s and [el! wall ela,stieity quantities in fhaseolus vulgaris leaves. J. t~per. Ilot. 35:941-B58.
- r.ohdShi-Shibata, J. and C. da Costa. 1980. Harves! inde, in Phasealus vulgaris l. !lean ¡.prov. Coop. 23:97-89.
- Knrban, S.S., D.P. [oyne, afid J.L. W!?ihing. 1991. ¡¡ate 01 water uptake and sites 01 water entry in seed. o/ diflerent cultivars 01 dry be.ns. Hort. Sei. 16:545-546.
- tramer, !'.J. 1983. Water relatians o/ pl.nt •• Academit F'ress; New York. 4B? pp.

- 74 -
- Kueneman, E.A., V.H. lIallace and P.M. ludIar. 1979. Photosynthesls measurements of fleld-gr_ dry be.n, and their relatiOl1 to selection lar yie!d. J. Amer. 5oc. !lort. Sci. 104: 49:)-482.
- KVle, J.H. and T.E. RandalJ. 1163. 1\ new concept 01 the hard seed charactor io Fl1aseo!us vulgarís L. aod lts use in breeding aod lnhoritance studies. Proc. Amer. Soco Agr. Sei. 81:461-475.
-laíog, D.R.l" P.6. Jones and J.H.C. na~is. 1984. Cammnn bean (Phasl!Olus vulqaris L.). p. 305-351. lo P.R. Goldsworthy and N.M. Fisher leds.). The physio!og)' 01 tropical field crop5. John Wi ley & SOOS¡ !lew York.
- LImer., H. 1979. Eoerg)' ""tabolism in higher planls io dilleren! environ~.l!nls. Thesis, State University Groningen, Ne>therlands. p. 1-179.
- ---- 1?82. Cyanide-resistant respiratiolH A non-phnsphorylaUng elEdron transport pato.ay ilCting as 3n energy overflow. Physiol. Plant 55:478-485.
---- 1984. Ihe ,unctiunal equilibrium: Nibbling on the edgE5 01 a paradigm. Neth~rlaods J. Agr. Sei. 31: 3'15-312.
- lilfcher, W. 1975. FhysiolOflical Plant EcoIOflY. 5pringer-Verlag¡ Serlín. 252 pp.
- Lauchli, A. 1984. Salt e,dusion: An adaptatiOl1 01 legUlles fe-r crop; saline conditions. p. 171-IB7. lo R.e. Staples aod B.H. Toenniessen (eds.). lar ioplants strategies lar crap improvement. John Wiley & Sons; New York.
and pa;tures under Sa!inity toler_oce
- Leopold, A.C. 19/J1). Aging aad senescenCE in plant developil!t'nt. lo fhima.no, ~:. V., R.L Adalmao and 6.5. Roth (ed.). 5enescente in plants. CRe Pr,,;s, loe.; Soca Ratan, Florid •.
- ---- .nd P.L Kriedemaon. 1975. Plant grOlith and development. Mc!h~-Hill¡ Ne>H York. 545 pp.
- Undoo, S.J. aod L.D. Noodén. 1977. Studies on the beh.,iour oi tne senestente signal in II;¡'Jf:a soybe.os. Plant F"hysiol. 59: 1136-1140.
- Lockhart, J.A. and V. GoHschall. sati ,u~. Plant Fhysi al. 3b: 189-39B.
196!. Fruit-ioduced ano apical senescence io Pisull
- LoEtieoberg, J.R. 1955. The development al bean seeds Whaseolus vulgaris L.). Plant Physiol. 10: 244-251).
- Loomis, R.S. and N.A. ~i!!iams.
patteros with leaves. p. 27-27. In J.D. CSSA¡ Nadisoo, Wisconsio.
1969. Prooudi vily aod the morpholo-JY al crop stands.1 Eastio [ed.l. Physiological aspects 01 crop yield. ASA,
- Lord, E.H. ano L.U. ~ohorn. 19Bb. G¡noecial development, pollinatiOl1, ano the path of palien tube grilWth in the tepary beao, F"haseul us acutí loliti~ Ilim". J. BoL 730 ):70-7B.
- l.nuwerse, H. ano W. v.d. IHeerde. 1977. Photosynthesis, transpiratíOl1 ano leaf IIOrpholOflY of Phaseolus wlgaris dad lea map; grOlifl dt difieren! irradíaO!:es in artificial i!!1d 5uolight. Photasynthetica Il: 11-21.
- lueas, E.a, S.M. Hilbourn and R.N. ;Jhitford. 1~,'6. The translú~atiOf1 01 "co, phatosynthate ¡roa leaves and pods in .t.asenlus vulgaris. Ann. Appl. Biol. B3:28S-2?ú.

- 75 -
- lugg, D.G. and T.R. 5inclair. 1981. 5eaSOflal changes in photosynlhesis 01 lield-grown soybeilll leaflets. 2. Relation to nítrogan conten. I'holosynthetica 15: 139-144.
- Marthart, A. IlI. 19!!5. COtIparative water relations 01 Phaseolus vulgaris L. and I'haseolus acutifolius Gray. I'lant I'hysiol. 77:I1HI7.
- Marques, I.A., M.J. llJerhoher and K.H. [dsmann. 1983. EHects 01 different inarganic nítrogen sourC!!S on pholosynlhetíc carbon lIelabolisill in primary leaves on non;¡ooulaled Phaseolus vulgaris lo Plant Fllysiol. 7l:555-5b1.
- Martin, r.A., J.l. Dzbun and D.H. Nallace. 1971. In .easurements al photorespiralion. Plant F'hysiol. 4~:7b4-U.!l.
- McKee, H.5., R.N. ROberlson aod J.B. Lee. 195j. I'hysi 01 ogy 01 pea ¡ruits. l. The develo¡¡ing IruiE. Australian J. Biol. 5d. B:137-163.
- Hiller, F.R. and Y. Kebede. 1984. Genetic contríbutions to yield gains in sarghul, 1950 to 19!1(t. p. 1-14. lo N.R. Fehr (ed.l. Genetíc eontribution to yield gaios ¡¡f five lajar ero¡¡ plants. ASA, CESA; Hadi son , Wisconsin.
- Miltharpe, F.l. and J.Hoorhy. 1979. An Iotrooudion to Crap I'hysiology. Cambridge Univer5it~' Frsss; CaMbridge. 244 pp.
- roOldau, H. 197.). Hfects al vMiau. water regill!!s on stOllatal and IEsaph)'1l eondudances 01 heao leaves. Pholosyothetica 711-7.
- roOlina C, A. 1996. Efecto de la distancia entre surco sobre el rendimiento en grano y otros par411!!lras fisiol6gicos de cuatro cultivares de fríjol precoz IFllaseolus vulqarls l.). 109. Agr. Thesis, Universidad Nacional, Pal.ira, Colombia.
- Honteith, J.l. 1%7. Ught interception and r1ldiativ2 mithange in ero¡¡ stand •• p. 89-111. 1'! EasUn, J.D., t.A. Hasking, C.V. Sullivan aod e.H.tI. Van Sa.el (eds.l. F'fiysiological aspeds 01 Crap Vield. Amer. Soc. Agron.; Madi son.
1978. Reassessmeot 01 la~i~u. growth rates lar C~ and C. traps. Expl. Agr. 14, p.I-S.
- Morgan, J.N. 1984. OsmoregulaUon and water stress io higher plant.. Ano. llevo f'lant fllysiol. 35:299-319.
- Mroginski, l.A. and ~.K. Kartha. 19B4. Tissue culture 01 legu..e lar [ro¡¡ illljifOvEilO1!lt, Plant Sreeding Review 2215-264.
- Nathanson, K., R.J. lawn, P.LM. De Jabruo and O.E. ayth. 19B4. Grc+lth, oooulation and nitrogen aecumulation by soybe.o io saturated soil culture. Field traps Res. 8:71-92.
- Nieman, R.H. and loL. Aoulsen. l~b7. Interactive elfeets 01 salinity and ahospheric husíalty on the growlh of bean aod col ton plants. Soto Gaz. 128,69-73.
Nobel, P.S. 1980. leal aoatooy and water use eHiciency. p. n-5S. Kramer (eds.l. Adaptation 01 plants to water and high temperature stress. Yart.
In N.C. Turner and r.J. John Wiley ~ Soos; Ney

- 76 -
- lIood~n, l.D. 1'18<). Senescente in tlle whole planto p. 219-25B. In LV. Thí_o (!!d.l. Senesc!!!lce in Plants. CllC Prass, lne.; Boca Ratan, florida.
- {]gren, N.L. 1'1ll4. Photorespiratian: r.thways, regulation and lIOdificatian. Ann. Rev. Plant Physiol. 35:415-42.
- OjehDmOn, 0.0. 1972. Fruit abseissian in [owpe., (Vigoa unguiculata l. Walp.l. J. Exper. Bol. 23:75!-7/'1.
- OW:er, N., A. l'oljakofl-Hayher, and A./f. Hayar. 1978. Changas in weight, nítrogen aeeUIIUlatian, respiraban and photosynthesis during growth and develO¡¡liIl!nt al seeds and p¡¡.js 01 Fhaseolus vulgari.. Amer. J. Bot. 65:366-371.
- Osborne, D.J. 1977. Ehtylene and targat ce!!. in the growth of plants. 5ei. Pr¡¡g. b4:51-ó3.
- 05llOOd, I:.B., K. llinter and 5.B. Powles. 1'18<). Adaptive signifítance 01 carhan dio:dde c¡cling during pllotosrntllesis in water-stress!!d plants. p. 139-154. In N.e. Tumer and P.J. ~:ra!fter (eds.l. Adi!j1tatioo of plant. to water aad high temperature stress. JaIln Niley ~ 5ans; New York.
- O'Toole, J.c., P.M. Ludlord, and J.L. Ozbun. 1977a. Gas e,ellange and enzymg adivity during lea! expansion in Phasrolus vulgaris l. !len Phytol. 78:51,5-572.
- ----, J.l. Ozbun and D.H. Wall.ce. ¡Q77b. Photasynth!?tic respOl1se to water stress in Pha5!!!Jlu5 vulgari 5. Phy5iol. Plant. 4¡): 111-114.
Pandey, R.~:, N.A. T. Herrera .nd J.W. Pendletoo. 1'l84. lJrooght response af grlin leguffii's unoer irrigation gradient: 11. Plant water st.tus and canopy temperature. Agron. J. 76:553-557 •
• ----, K.C. S •• en., tI.H. ~:.lubarffie, V.B. Sing" and V.V.S.S. Pfasad. 1971,. GE!lotypie variatioos in photosl'l'thetic fate and fespiratory losses in some grain legumes. Pl3l1t ~iochem. J. }(!):72-!lé'.
- Paredes, O. N. 192b. A study 01 se!!d lilling and dry ffi;tter p;rtitiooing (h.r.deristi[s .na their eombining abilityeHects and reJatíonsllips to yield amang drl' beaos with dWeriog grOl;th habits, architectures and maturHies. M. Se. Thesis. lIichigan Shte th1iversíty.
- Passioura, J.B. 1983. 7:265-2g:)~
Root, and drought resislance. Agricultural Water Han,¡gement
- Pate, J.S. 1975. Pea. p. 191-224. In l.r. Evans (ed.l. ero¡¡ ph)'siolog)'1 some ene histories. Cambridge l'niversity Pres5; Caobridge.
- Pechan, P.M. and B.O. Webster. 19Bb. Se!!d i!I1d poo se! of r!!d Hdney beaos. J. Amer. Soc. Hort. 5ci. II1 (!):87~B9.
- Penning de \lríes, F.W. T. 1975. Ihe cost 01 maintenanee processes in pla.nl (ell;;. Anna/s. Bol. 39: 77-?2,
- ----, A.H.N. Brunstíny, and H.H. Van laar. 1974. Produets, requiremenls and eHicienq Di bios¡~!hE;i5: a quantitative approaeh. J. Theor. Bio!. 45:339-,77.
- ---- and H.H. van La.r. 1977. Substr.te utilízatian in germinating seeas. p. 2!7-22B. In Landsbery, J.J. and C.V. CuHing (eds.l. Enviran_t.l eHects on crop physíol¡¡gy. Acldel!ic Press; loodoo.

- 77 -
- Piper, C.V. and W.J. Norse. 1923. The Soybean. McGraw-Hill Bool: Co.; NeH I'ork. 329 pp.
- Powles, S.S. and C.B. O;mond. 1979. Photoinhibition 01 intaet attached leaves 01 C. plan}.s illuminated in the absence 01 both carbon dio~ide and 01 photOfEspiration. Plant Physiol. 64: 982-988.
- RaH50n, H.M. and G.A. Con;table. 1980. Gas exchange 01 pigeon pea: a cornparison Hith othEf craps and a roodel 01 carbon production and its distribution within the plant. Vol.2. 175-189. In Proceedings 01 the internalional Hor!:shop on pigeon peas. ICRIsAT, Patencheru, Hybera.bad, India.
- Rodriguez C., R.R. 1986. Caracterización morlo-lisiológica e identilicación de caracteres para mayor rendimiento en genotipos precoces de Iríjol comÚn !F'haseolus vulgaris L.). N.Se. Thesis, Colegio de Postgraduados, Chapingo, México. 178 pp.
- Russell, R.S. 1977. Plant root systems. Me GraH-Hi 11 Boof Comp.ny (IJO Limi ted; londan, 798 pp.
- Sale, r.J.M. 1975. Produclivity 01 vegetable crops ina region 01 high solar input. IV. Field chaffiber measurements on French beans !Phaseolus vulgari; L.) and cabb'!ges ( Bras ; ,! oleracea). Australian J. Plant Physiol. 2:461-470.
- Samper, C. 1984. Eflects 01 Hater stress imposed at mid-pOd filling upon yield and dry matter partitioning in dry beans (r.,. vulgaris). M.S. Thesis, mchigan State UnivEfsity.
- Sato, H. and K. Gotoh. 1979. The phototropic movement and it's varietal diHerence in th.e leal 01 common bean !F'haseolus vulgaris L. l. Jaurn. 01 HoHaido Un\'. 01 Educ. (Sedion 11 8) Vol.30, No.I,pp.45-51.
- ---- and ---- 1983. Studies on leal orientalion movements in kidney beans. l. The response to Iight intensity and location 01 photo-receptor. Japan J. Crüp Sci. 52(4):515-520.
- Sa.~ena, N.P. 1984. Chidpea. p. 419-452. In P.R. Goldsworthy and N.M. Fisher (eds.l. The physio10gy 01 trüpical field crüps. John Wi ley· ~ Sons; tle", York.
- Schieder, O. 1982. Somatic hybridization: a ne,l .. ethod le; plant improvement. p. 239-251. In l.K. Vasil, N.R. Snowcraft, and K.J. Frey (eds.l. Plant improvement and samatie cel! genetics. Arademic Press; Me", I'off:.
- SchHanitz, Hassachusetts.
1967. The origin 01 cuHivated plants. Harvard University Press; Cambridge,
- Sehwartz, H.F. 1978. Miscel!aneous lungal pathogens. p. 12]-152. In H.F. SchHartz and G.E. Galvez (eds.l. Bean production problems. CIAT, Cali, Colombia.
- Seemann, J.R. and C. Critch1ey. 1985. Elleds 01 saH stress an the growth, ion content, stomatal behaviaus and photosynthetic capadty 01 a salt-sensitive, spedes, Phaseolus ,ulgaris lo Planta 164: 151-162.
- Seth, A.K. and F'.F. Wareing. 1967. Hormane directed transport 01 Bot. 18: 65-77.
"€taboli tes and i ts possible role in plant senescence. J. Exper.
- Shackel, K.A. and A.E. Hall. 1979. Reversible leal movements in relatian to draught adaptation in cowpeas Vigna unguiculata (L.) Walp. Aust. J. Plant Physiol. 6:265-276.

- 78 -
- EhaH, R.H. and D.R. Laiog. 19bb. Hoislure stress aod plant response. p. 73-94. In L.T. Evan. (ed.). Crop Phy.iology: 501ft!! Case Histories. ASA, C5SA; Madi.oo, Wiscoosio.
- Sheldral:e, A.R. 1'lG4. Figeoo pM. p. 385-417. In P.R. Gold.worthy and tloM. Fisher (eds.). The pnysiology 01 tropical field erops. Jono Wiley" Son'; tlew York.
- Ehibles, R., 1.C. Aoderson and A.f!. Gibson. 197:i. Soybean. p. ISHS9. In loT. Evan. (ed.l. Crop physiology: S!lile case histories. G.m!Jridge University Pres.; Cambridge.
- 5iddique, M.O.A. and P.!). 6oodwin. 1900. 5eed vigor in bean ¡Phas!!ülus vulgaris L. CIC
Apollo! as influEfleed by temperature 'lod water rEglm~ during d~,i?lopmeflt and luturatioo. J. Exper. BoL 31:11:1-323.
- Silvera, G.A.C. 1974. lneorporatioo 01 germplas. 01 [he "Sealarer" nay» bean into several Central and South AiIIerican beao cultivars in relatíon to breeding lar im¡Jrove..a.lt in adaptation. f'It.D. Thesi;, Nichigan St_te Uni v. ~8 pp.
- Sine!aír, r.R. and c.r. de Wít. !~7ó. Analysís of the carUon and nitrogen limit.tioos to ¡¡oybe.n yield. Agroo. J. 69:319-324.
- Bingo, s.r., and J.A. &Jtí~rrel. 1984. Geographítal distribution al the DlI and nCt2 genes cdusing hybrid dN;rfís~ in Phaseo1us vulgaris L., their associ.tion Nith sead size, ind tneir significante lo breeding. Euphytica 13:137-345.
- Slatyer, R.O. 1967. P!ant-water re1ationships. Arademie Press; landan.
- Sfflillíe, R.M. ana R. Nott. 19!12. 511t tolerante in erap planEs monitored by chlora¡;hyll f1oure¡¡eence in vi va. Plant Physiol. 70: 1049-1054.
- S;¡¡irnofl, ti. and G.R. 5te;;art. 19B5. lIítrate assímilation and translocation by higher p13nts: Campar.Uve physiology and ecologica! eonsequenees. Physiol. Planto M:l33-14ü.
- Snedecor, S.H. and II.S. Caenran. 1967. Shtistical I1ethods. lowa State University¡ A!>es. 593 pp.
- Sobolev, N.A., E.K. AdalOChu!: and LV. lhila. 197b. Apomixis in pea. p. 221-277. In 5.5. Kho!hloo (ed.). Apomixis in breeding. Aillerind Publishing LO. r"t, Ud.; New Delhi.
- Sofield, l., L. T. Evans, M.G. Coo~ and I.F. Warlaw. 1977. Fadors influet1cing tlle rate and duration al grain filling in i'heat. Australian J. Plan! Physiol. 4:7B5-797.
- Spooehiado, B.N. 1985. Avalíacao de sistema ndieular no leijoeíro Wl1asealus vulgaris L.)
eDiO un aecanisffiD de tolerancia ao deljeít hidríco. M.Se. Thesís. univ. Pelotas, Rio Grande do Sul; Brasil. 11} pp.
- Shrk, LC. and e.H. tlahooey. IN2. A .tudr 01 the tillie 01 developll!!nt 01 tlle fibroos sheath in lhe sidewall 01 edib1e snap bean püds with fespect to quality. PrO(. Amer. ~. Hort. Sei. ~1:15H59.
- Stephenson, R.A. and G.L. Hilson. 1977. Fatterns 01 •• simil.te dístribution at ~aturity. !l. lhe hile (OlieSe 01 changes io 14C dístributíoo in pods and st!!lll sections. Australia« J. Agr. Res. 2B!395-4(~.

- 79 -
- Stewart. e.R. 1972. Ellects 01 proline and carbonydrates on toe metabolism 01 exogenous praline by excised hean leaves in toe darf:. f'lant Physio!. 5D:S51-5S5.
- ---- and A.D. Kanson. 19B!). Proline accuiIi'Jlation as a metabolic respOll'. to water stress. p. 173-189. In N.C. Turner and P.J. Vramer (eds.) Adaptatioo al pLmts tü IIdter and hi'jh temperature stress. Jonn Wiley & 5on5; New York.
- Stolella, P.J.,Il.F. Sandsled, R.N. lobal, ana ¡¡.lo Ilymes. 19BO. Root eharaeteristics 01 Mat!. beans. l. RelatiOllships of root size to lodging and seed (ield. ero¡¡ Sci. 19:823-830.
- 5toler, R. 1974. Effeet en dI/3rt heans 01 water stress at different phases of gfowth. N. l. J. [xper. Agr. 2:13-15.
- 5ubhadrabandhu, S. 1976. Control 01 abscission frf flm,ers anó Iruits of fl1a.SEOlus I'ulgaris lo f11.U. Thesis. /!ichigan 5tate University, East Lansing.
- ----, f'I.lI. M_ and D.II. Reicosky. 1~7B. ~b5cis5ion 01 f1DHff5 and ¡rutls in f'haseolus wlgaris lo 11. Cultivar dilferences in floweriog patterns and abscission. ero¡¡ !id. 1!l:B?1-89b.
- 5usse, , I.N. and R.M... Dale. 1979. HorlllOn3l tootrol 01 storage proteio ;yothe.is in Fhaseolus vulgaris. p. 129-141. lo The plant seect. At:ademis F-ress; New YorL
- ra"as, LA., D.H. Wallaee, P.M. Ludloro and J.L. Ozbun. 1979. EHect 01 older Ifuits en ah9ftion and abscisic aeid concentratien 01 \'ouoger Ifui ts in Phaseolus vulgaris lo Plaot Ft1ysiol. 64:620-622.
- Tanal:a, A. 19H. Studies en physiology 01 f11aseolus vulgaris. t1flfival Ilep. lañ. f'lant tMrition, Faeulty 01 Agrie. fIolhido Univ. Sapporo, Japao, 38 pp.
and K. fujita. 1979. browth photosynthesís and yield components in relalion lo grain yield 01 tbe lield bean. J. Fae. Agr. fIoHaido Uní,. 5~(2):145-2'l!l.
- Ta¡lor, 11.1'1. I'IB<J. tfodifying root srsteills of eoUon ana sorne.ns to increa5~ water absorption. p. 75-84. In N.C. Turner and P.H. Kr.mer leds.). Maptation of plants to water ¡;,¡d high temperature stress. Wiley Interscience; New Vor!.
- Thimann, K. V. I'IB<). The senescence 01 leaves. p. 85-115. lo V. V. Thi~ano led.l. Senescence in plttnis. [Re Press; Soca Raton, Florida.
- Thomas, R.J., U. Feller and V.H. Erismann. 1?79. The ellect 01 diflerent inorganic nitrogen sGurces and plant age af the composition 01 ble~ding sap 01 Phaseolus vulgaris. New Phytol. 82:657-669.
- Thomps!!f1, D.N. 1942. un growth and for.. O?6? edition, J. T. ~onner, eds. l. Cambridge Ilniversity rres,; Cambridge.
- TiIljlO, LE. and C.A. /leyra. 1983. Expressioo 01 nitnte and nitrite fEdudase adivities under various lorms 01 nitcogen nutrition in Pbaseolus yulqaris l. Plant Phisiol. 72:71-75.
- Tolnert, N.E. 1981). Fhotü,espintion. p. 400-523. In D.n. Dades led.). Tne biocnemi stry 01 plants. Vol. 2. ""tabolis~ and respiration. A-:acJemic Press; New Yorl:.

- 80 -
- TroedsOrt, R.J., R.J. lailil and D.E. flyth. ¡'1St Water ~anag""ent 101' peal sOrtean prooudioo Ofl clay $OHs. p. 257-265. In M. A. Huirhead and E. fla~hreys !eds.). Root ~Orte Iiaitatioo- r.) crap production an clay sails.
- Tuder \ e.l., M.O. Miller and B.D. Ileh5ter. 1975. Effects 01 ethephan an seed yield 01 Phaseolus vulgari 5 lo Hart. 5d. 10: 15b-157.
- Turner, N.C. and M.M. Janes. ¡981). Turgor mainten.nce by oSilIOtic adjust~nt. A review and evaluatian. p. 81-103. In Il.e. Turner and P.J. ~ramer !eds.!. Adaptatian 01 plan!s ta water and high t~erature stress. Wi ley ~ Soos; NeH York.
- TrTee, tU. and H. T Hammel. 1972. lhe measum1li?nt 01 the turgor pressure a04 the w.ater relatioos 01 plants by the pres5ure-b01'h teehnique. J. Exper. Bot. Vol.23,No.H, p.267-B2.
- Vallejos, LE., 5.0. Tanl:sley, R. Bernatzl.y. 1986. localtzatioo in th~ tD<llatü genül1lE 01 DllI\ restrictioo fragille1lts cootaining sequences 01 homologous the rRr<A !~S 51, the ~djor chloraphyll ./b btnding polypeptide and vibulos. bíphosphate nrbo~ylese genes. Genetí !in press).
- Van Steveniocl, R.F.M. !~S7.
yellow lupios (Lupinus luteus U. E~peí. BoL 8: 373-3BI.
Factors aHecting lhe a!lsLÍssion 01 íeproductive organs in l. The ellect 01 dilferent pattero5 01 Ilower remov.l. J.
- Volenec, J.J., H.T. Nguyen, C.J. llelsan aad U.A. Sleper. 1984. Potential lar genetically IlIfldifying dad respiraUoo 01 tal! leseue leaves. trap Sci. 24:938-943.
- Voysest, O. and J. Earcia. 1984. Anal )sis 01 yield variatioo in the Internatianal Bean Yield and Adaplation Nursery: Focus 00 enviranmental represen tal ion. p.328-in In lo lernatior, il beao trials wOftshap. CIAl, eaH, ColOfllhia.
- Wadman~van 5chravendUl:, H. and D.N. vaii Andel. 1985. Interdepenrlence of groHth, Hlt!r relatioos and abscisie _cid level in f'haseolus vulqaris duriog wahr1ogging. f'hysiol. Planto b3:21S-220.
- Hallare, D.H. 1985. fhysiological genetics 01 plant ~aturity, adaptaban and yield. f'lant Ilreeding Reviews 3:21-lb7.
- ---- and I1.M. l1uoger. 116/'. Studies Ilfl the physiological basis lar rieló differences. 11. Variatians in dry matter distribution amog ¡¡erial argans lar several dry beao varieties. Crap Sel.
6:503-507.
- ----, J.l. Ozbuo, and I1.M. Munger. 1972. f'hysiological gentics al crap yieló. Advance. Agran. 14:97-14b.
- ----, M.H. F'eet i!l1d J.lo Ozbun. 1976. Studies 01 W, a.hbalism in F'hasoolus vulqaris l. anó applícatioos to breeding. In CO, Metabol i 5. 3.nd Plaot Proouctt vi ty. [d. by R.M. Suros aod C.e. Black. llniversity rark Press; Baltimore. p. 4.3-58.
- Haltan, D.C., E. Galsan and M.A. Harrison. 1977. The relationship bet~en .tomatal resistance and absdsic-acid levels in leaves 01 water-stress'?<l heao plant •. Planta 133:145-148.
- W.reing, f'.F. and A.K. 5eth. 1967. Agetng and senese.nce in the Hhole planto Sy~. Soc. Exp. BiaL 2It5·13-55B.

- 81 -
- Waters Jr., l., P.J. Breen and H.J. rlad. 1980. Tran.location 01 14C-phalosynthate, carbOhydrate content, and nitrogen lix.lían in Phaseolus vulgari. l. during repraductiYE developl!ll!l1l. J. Amer. Soc. Hart. Se!. 105(1):424-427.
- Webster B.n. ind H.W. Chiu. 1975. Ultrastructural .tudies 01 abscission in Fnaseolus: Characterislics 01 the floral abscíssion zane. J. Amer. 50c. Hart. Sei. 100(6):613-618.
- Wsisz, P.R., R.F. Venisoo, and T.R. Sinelair. 1985. Response to drought stress 01 nitrogen fí".Han (acetylene redudíonl rates by fíeld 9rO+ll\ soyheans. Plant Physíal. 78:525-530.
Westerman, l,K, Porter and M.A. o'ne;;¡¡. patterns in bean plants. Crap Sei. 25: 225-2?9.
1985. tUtrogen partHioning and ft1{1/)ilization
"'hite, J.M. 1981. A quantitatíve analysis 01 the growth and develapment 01 bean plants Ifhaseolus vulgaris L.l. Ph.D. thesis. lJniversity' 01 California; Berkeley.
- ---- 1984. Associ.tions aMng yield, yíeld slability and duraban 01 grawth qele in lBY!1N trials. p. 3!lO-41)f). In !nternational bean trials warlshop. ClAT; e.ti, ColOillbia.
- I/iehold, N.J., O.A. Ashley, <loe H.R. Boerma. liS!. P.eprodudíve abseissian levels and patterns lor eleven determinate sOrbean culti vars. Agran. J. 73: q}-4b.
- Míen, /l.e. ane E.E. Adab. 1979. I'ad develapllle!1t perlad in cowpeasl varietal diflerences as related to seed characters and environmenlal ellects. Crap Sel. IB:791-794.
- uu and R.J. SUlM!erfíeld, 1984. Cowpea (Vigo. unguiculata (L.) lIalp.). p. 353-3!l3. In P.R. lioldsworthy and 110M. Fisher (eds.l. The physiology of trapical lield craps. John Niley & Soos; New York .
• ---- and D.H. Nallaee. 1973. light-induced leaflet oríen!atian in F'hasealus vulgari. L Crop Sci. 13:721-724.
- Wilkerson, b.G., J.II. Janes, K.J. Boate, K.T. lngram, J.H. Hoshoe. 1983. Modeling sayhean gr~th lor crap ~aoageffient. Transactlons of the ASAE 26:b3-73.
- Nilsoo, D. and J.6. Janes. 1982. [Hect 01 selection lar d • .rl: respiratici1 rate 01 lIature leaves on crap yields of Lolium perenne ev. 521. Ann. Bol. 49:113-320.
- Hit, C.T de et al. 1979. Simulation al assímílation, r!!spiratíon and transpiratian 01 traps. Pudoc; Wageningen. 140 pp.
- ---- and F.N.T. Penning de Vries. 19B3. Crap growth models without hormones. Netherlands J. Agr. !id. 31:313-123.
- Yañez r., [. Pimienta, t. liad and J. I'ohashi. 19B4. ["".¡Jarati,e anatoffl)' 01 flower buds wíth and lIíthout pn!ential for abscissían in Phaseolus vulgaris. Turrialba VoL 34, No.4, p.431.435.
- Yoshida, 5. and E. de los Reyes. 1976. leal cuticular resislanee 01 rice i'arieties. 50il 5ei. Plant Nutr. 22(1¡:95-9S.





















