
Nuevos Restos de Alligatoridae Cuvier, 1907 (Eusuchia, Crocodylia) del Neógeno del Sur de la...
168
UNIVERSIDAD NACIONAL DE SALTA FACULTAD DE CIENCIAS NATURALES ESCUELA DE BIOLOGÍA Tesis de Licenciatura en Ciencias Biológicas Nuevos Restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias Sistemáticas. FRANCISCO BARRIOS Director: Dr. Jaime Powell Co-Director: Dr. Fernando Lobo 2011
Transcript of Nuevos Restos de Alligatoridae Cuvier, 1907 (Eusuchia, Crocodylia) del Neógeno del Sur de la...

UNIVERSIDAD NACIONAL DE SALTA
FACULTAD DE CIENCIAS NATURALES
ESCUELA DE BIOLOGÍA
Tesis de Licenciatura en Ciencias Biológicas
Nuevos Restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del
Neógeno del Sur de la provincia de Salta. Implicancias Sistemáticas.
FRANCISCO BARRIOS
Director: Dr. Jaime Powell
Co-Director: Dr. Fernando Lobo
2011

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 2
“Mi sabiduría viene de esta tierra”……J. M. Castilla.
A mi vieja
A mis hermanos/as
A mis amigos/as
A la Universidad
A mi país.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 3
ÍNDICE GENERAL
I. INTRODUCCIÓN……………………………………………………………………… 4 I.A. Reseña Histórica y Taxonómica…………………………………………………… 4 I.B. Registro Fósil de Alligatoridae en Sudamérica…………………………………… 9 I.C. Diversidad Actual de Alligatoridae en Argentina……………………………… 12 I.D. Antecedentes………………………………………………………………………... 13 I.E. Objetivos…………………………………………………………………………….. 20 II. MARCO GEOLÓGICO Y GEOGRÁFICO DEL ÁREA DE HALLAZGO……… 21 II.A. Procedencia Geográfica…………………………………………………………… 21 II.B. Procedencia Estratigráfica y Edad……………………………………………….. 21 III. MATERIALES Y MÉTODOS………………………………………………………. 23 III.A. Nomenclatura……………………………………………………………………... 23 III.B. Materiales…………………………………………………………………………. 23 III.C. Preparación y Comparación……………………………………………………... 24 III.D. Análisis Filogenético……………………………………………………………… 25 IV. RESULTADOS……………………………………………………………………….. 28 IV.A. Sistemática Paleontológica……………………………………………………….. 28 IV.B. Descripción de MAS-PALEO-RdlF 001/2011…………………………………... 29 IV.B.1. Cráneo……………………………………………………………………….... 29 IV.B.2. Cavidades del cráneo…………………………………………………………. 31 IV.B.3. Huesos craneales……………………………………………………………... 34 IV.B.3.1. Norma dorsal…………………………………………………………….. 34 IV.B.3.2. Norma lateral……………………………………………………………. 38 IV.B.3.3. Norma occipital………………………………………………………….. 40 IV.B.3.4. Norma palatal……………………………………………………………. 42 IV.B.4. Mandíbula…………………………………………………………………….. 43 IV.B.5. Dentición……………………………………………………………………… 47 IV.C. Comparaciones…………………………………………………………………… 50 IV.C.1. Comparaciones genéricas…………………………………………………….. 50 IV.C.2. Comparaciones específicas…………………………………………………… 56 IV.D. Análisis Filogenético……………………………………………………………… 61 IV.D.1. Posición filogenética del caimán de Rosario de la Frontera………………... 62 V. DISCUSIÓN…………………………………………………………………………..... 64 V.A. Consideraciones Taxonómicas……………………………………………………. 64 V.B. Aspectos Paleoclimáticos-ambientales y Paleozoogeográficos………………...... 77 VI. CONCLUSIONES……………………………………………………………………. 85 VII. AGRADECIMIENTOS……………………………………………………………... 90 VIII. BIBLIOGRAFÍA CITADA………………………………………………………... 92 APÉNDICE I……………………………………………………………………………… 105 APÉNDICE II……………………………………………………………………………... 153 APÉNDICE III…………………………………………………………………………….. 154 APÉNDICE IV…………………………………………………………………………….. 162 APÉNDICE V……………………………………………………………………………… 164 APÉNDICE VI…………………………………………………………………………….. 165

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 4
I. INTRODUCCIÓN
A fines del año 1998, investigadores encabezados por el Dr. Jaime Powell de la
Facultad de Ciencias Naturales e Instituto Miguel Lillo de la Universidad Nacional de
Tucumán, fueron alertados por el Grupo de Investigaciones del Museo Histórico
Arqueológico “Profesor Juan Carlos Rivas” de Rosario de la Frontera (provincia de
Salta), sobre el descubrimiento de unos huesos encontrados por un pescador de la zona
en una de las riberas del río Rosario de dicha localidad. Los investigadores de la citada
institución, exhumaron los restos de un cocodrilo que tendría aproximadamente cinco
millones de años (5 Ma.) de antigüedad, edad inferida a partir de su ubicación
estratigráfica.
Dicho ejemplar, motivo de este trabajo, consiste en un cráneo casi completo,
mandíbula completa y restos postcraneales (varias vértebras, piezas de las cinturas,
costillas, algunos huesos de las extremidades y osteodermos), constituyendo el fósil más
completo del grupo encontrado en la zona, y de aspecto similar a las especies que viven
en la actualidad en la parte noreste del país. El material se encuentra provisoriamente
alojado en la Colección Paleontología Vertebrados Lillo, pero será ingresado a la
Colección de Paleontología del Museo de Antropología de Salta bajo el acrónimo y
número MAS-PALEO-RdlF 001/2011.
El hallazgo de este fósil constituye un aporte significativo a la herpetofauna
cenozoica de la región, ya que plantea interesantes interrogantes sobre la sistemática y
distribución del grupo en nuestro país. Su estudio permitirá también determinar el clima
y el ambiente donde se desarrolló por comparación con los ambientes donde viven las
especies actuales.
I.A. Reseña Histórica y Taxonómica:
Los cocodrilos o crocodilianos, al igual que las aves, pertenecen al gran grupo de
los Archosauria el cual incluye también a varios taxa extintos, incluidos los famosos
pterosaurios y dinosaurios (Fig. 1) (Brochu, 2001b; Benton, 2005; Nesbitt, 2011).
Recientes análisis filogenéticos alteraron la clasificación de los crocodilianos (Brochu,
2003; Martin & Benton, 2008; Brochu, et al., 2009). Los animales previamente

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 5
llamados crocodilianos son ahora nombrados crocodilomorfos, y el término
“crocodilianos” está restringido al ancestro común de los taxa actuales y todos sus
descendientes (Molnar, 1993; Brochu, 2003; Martin & Benton, 2008).
Clado Crocodylomorpha
El registro fósil de Crocodylomorpha tiene una larga historia documentada desde
del Triásico tardío (hace más de 220 Ma), originados a partir de una rama primitiva de
arcosaurios. Su posterior desarrollo y diversificación ocurrió en el Cretácico y Terciario,
ello es evidente en estratos para ese tiempo de casi todos los continentes que brindaron
diversos y extraños grupos, especialmente distribuidos en regiones de latitud medias-
tropicales a ecuatoriales (Gasparini, 1981; Molnar, 1993; Brochu, 2003). Los cocodrilos
fósiles, especialmente aquellos con representantes actuales, son considerados buenos
indicadores de condiciones ambientales específicas (Sill, 1968; Taplin & Grigg, 1989;
Markwick, 1998).
Tradicionalmente, los cocodrilos fósiles y actuales que constituían el orden
Crocodylia (sensu Kälin, 1955), y que ahora conforman el clado Crocodylomorpha,
eran agrupados en tres subórdenes: Protosuchia, Mesosuchia y Eusuchia. Con las
filogenias recientes, estas agrupaciones pierden su status taxonómico (Brochu, 2003).
Aun así, la posición de la narina interna en el paladar y la forma del cuerpo vertebral
(Fig. 3) siguen siendo útiles herramientas para diferenciar los protosuquianos y
mesosuquianos del monofiletico y moderno Eusuchia (Molnar, 1993).
El registro fósil evidencia una gran diversidad de taxa dentro del gran clado de
los crocodilomorfos (Fig. 2). Los géneros sobrevivientes de cocodrilos sólo son una
pequeña parte de la gran diversidad pasada del grupo, la cual es revelada por varios
géneros fósiles (Brochu, 2003; Martin, 2008). De esto se desprende que las formas
actuales de cocodrilos que pertenecen al clado Eusuchia, de hábitos anfibios, son un
relicto de la gran variedad de tipos adaptativos que desarrollaron durante el Mesozoico
y Cenozoico (Kälin, 1955; Sill, 1968; Bonaparte, 1971; Buffetaut, 1979; Gasparini,
1981; Molnar, 1993 y 2010; Sereno et al., 2001; Brochu, 2001a y b; Krause et al., 2003;
Benton, 2005; Jouve et al., 2005; Pol & Gasparini, 2007; Oliveira, 2008; Sereno &
Larsson, 2009; Nesbitt, 2011); desde los netamente marinos longirostrinos

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 6
Thalattosuchia del Jurásico, a los pequeños brevirostrinos continentales Notosuchia del
Cretácico (Gasparini, 1971), y los grandes predadores terrestres “zifodontes” (de dientes
aserrados) Sebecosuchia del Cretácico y Terciario de Sudamérica (Gasparini, 1972)
(Fig. 4). El territorio argentino ha brindado la posibilidad única en América del Sur de
conocer fragmentos importantes de la historia evolutiva de los crocodilomorfos
(Gasparini, 1981).
Clado Crocodylia
La taxonomía filogenética para Crocodylia reconoce tres “stem-based groups”
indicados por los sufijos “oidea” (Fig. 2). Un stem-based group/clade es aquel definido
por todos los taxa que comparten un ancestro común más reciente con un taxon
particular que con otro; es el clado más inclusivo que contiene al ancestro común y
todos sus descendientes, originado de una rama particular (internodo) en un árbol
filogenético (de Queiroz & Gauthier, 1990). Los stem-based clade dentro de Crocodylia
son, Gavialoidea (Gavialis y todos los crocodilianos relacionados que a Alligator o
Crocodylus), Alligatoroidea (Alligator y todos los taxa más emparentados que a
Crocodylus o Gavialis), y Crocodyloidea (Crocodylus y todos los taxa relacionados que
a Alligator o Gavialis). Este sistema de nomenclatura filogenética también usa nombres
de “crown group” indicado por el sufijo “idae”, para los grupos diferenciados por el
último ancestro común de los miembros recientes (Alligatoridae, Crocodylidae, y
Gavialidae) (Brochu, 1999a y 2003; Brochu, et al., 2009). Un crown group/clade es un
clado derivado del ancestro común más reciente de los representantes vivientes; los
crown groups son subconjuntos de un node-based group. Los node-based groups/clades
son originados de un nodo particular de un árbol filogenético, y son los clados menos
inclusivo que contienen al ancestro común y todos sus descendientes (de Queiroz &
Gauthier, 1990).
En la actualidad, el clado Crocodylia (sensu Brochu 1999a, 2003) es el grupo
taxonómico más pequeño dentro de Reptilia e incluye unas 23 especies de aligátores,
caimanes, cocodrilos y gaviales; aún así son los reptiles de mayor talla. Todas estas
especies están distribuidas en los tres grandes linajes de Alligatoridae, Crocodylidae, y
Gavialidae (Fig. 5). Estos crown groups que habrían divergido al final del Mesozoico
(Brochu, 2003), se diferencian en gran medida por características craneales. Los

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 7
cocodrilos se encuentran asociados con regiones de latitudes medias-tropicales a
ecuatoriales (Brochu, 2003; Martin, 2008). Todos los integrantes del grupo son de
hábitos anfibios y comparten características morfológicas, anatómicas y fisiológicas, las
cuales los hacen más adaptados al agua que a la tierra (Grigg & Gans, 1993; Martin,
2008).
Aunque la mayoría de las poblaciones de cocodrilos sufrieron la sobrecaza y la
pérdida de hábitat durante el siglo veinte, algunas permanecen críticamente en peligro, y
otras son verdaderos logros en biología de la conservación y se convirtieron en
importantes recursos económicos (Brochu, 2003; Larriera & Imhof, 2006; Martin,
2008).
Gavialidae
El clado Gavialidae contiene sólo dos géneros y dos especies (Gavialis
gangeticus y Tomistoma schlegelii) que están restringidas al Sudeste Asiático. Se define
filogenéticamente como “el último ancestro común de G. gangeticus y T. schlegelii y
todos sus descendientes” (Brochu, 2003). Sin embargo la ubicación taxonómica del
falso gavial (T. schlegelii) es objeto de debate entre los especialistas, sobre si realmente
es un gaviálido o un crocodílido; datos morfológicos apoyan la primera hipótesis
mientras que la información molecular la segunda (Densmore & Owen, 1989; Molnar,
1993; Poe, 1996; Brochu 1997a, 1999b, 2001 y 2003; Lutzoni et al., 2000; Harshman et
al., 2003; Vélez-Juarbe et al., 2006, Martin, 2008). A pesar que hoy en día habitan la
cuenca del Ganges, Pakistán y Birmania (Fig. 6A), son conocidos en depósitos
terciarios de Asia, África, Europa, América del Norte y América del Sur (Brochu,
2003).
Crocodylidae
El grupo más diverso en especies es el clado Crocodylidae, que incluye dos
géneros y doce especies distribuidas en las regiones tropicales de América, África, Asia
y Australia (Fig. 6B) (Molnar, 1993; Vélez-Juarbe, 2006; Martin, 2008). Se define el
clado como “el último ancestro común de Crocodylus niloticus y Osteolaemus tetraspis
y todos sus descendientes” (Brochu, 2003). Diferenciado ya en el Cretácico tardío,

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 8
ocurren en todo el Hemisferio Norte durante el Terciario (Sill, 1968; Brochu, 2003;
Mead et al., 2006).
Alligatoridae
El clado Alligatoridae es el más diverso en géneros con respecto a los otros dos;
está conformado por Alligator, Caiman, Melanosuchus y Paleosuchus. Definido como
“Alligator mississippiensis y Caiman crocodilus y todos los crocodilianos más
relacionados a ellos que a Gavialis gangeticus o Crocodylus niloticus” (Brochu, 2003).
El grupo incluye al menos ocho especies vivientes, las dos especies de Alligator
disyuntas geográficamente (Alligator mississippiensis del Sudeste de América del Norte
y A. sinensis del Este de China) que conforman el clado Alligatorinae (todos los
aligatóridos más emparentados a Alligator mississippiensis que a Caiman crocodilus)
(Brochu, 2003), y a los caimanes de todo Centro y Sudamérica (Fig. 6C) que
constituyen el grupo monofilético Caimaninae (grupo de especies más relacionadas a
Caiman crocodilus que a Alligator mississippiensis) (Brochu, 2003). Las especies que
conforman el clado de los caimaninos son, el gran caimán negro de la Cuenca del Río
Amazonas (Melanosuchus niger), el yacaré ñato u overo (Caiman latirostris), el yacaré
negro (C. yacare), los caimanes enanos o de frente lisa (Paleosuchus palpebrosus y P.
trigonatus), y el complejo Caiman crocodilus (babilla o caimán de anteojos) que está
formado por varias subespecies (C. c. crocodilus, C. c. apaporiensis, C. c. fuscus y C. c.
chiapasus). Brochu (1999a) propone el nombre Jacarea al grupo que incluye al ancestro
inmediato de Caiman crocodilus, Caiman yacare, Caiman latirostris, Melanosuchus
niger, y a todos sus descendientes; este clado contiene a todos los caimanes, con la
excepción de los dos caimanes enanos Paleosuchus palpebrosus y P. trigonatus.
Los Alligatoridae sudamericanos son considerados tradicionalmente inmigrantes
de América del Norte (Aguilera et al. 2006). Los alligatorinos más tempranos son
conocidos del Paleoceno Inferior, lo que demuestra que la separación entre alligatorinos
y caimaninos ocurrió por ese tiempo, o incluso antes en el Cretácico Tardío (Brochu,
1999a y 2003; Bona, 2007). Unos pocos restos de caimanes son conocidos del
Paleoceno y Eoceno de Norteamérica (Brochu, 1999a y 2010).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 9
El registro fósil de los caimanes es el menos completo que cualquier otro linaje
de crocodiliano, y solo un puñado de caimanes son conocidos de depósitos pre-
Mioceno, pocos de los cuales fueron incluidos en análisis filogenéticos (Brochu, 2003 y
en prensa; Bona, 2007). Es evidente que considerando el registro fósil en el esquema de
relaciones de los Alligatoridae, estas son más complejas y en algunos casos poco
resueltas (Fig. 7).
I.B. Registro Fósil de Alligatoridae en Sudamérica:
Numerosos fragmentos atestiguan el predominio de los Eusuchia en el registro
fósil sudamericano del Cenozoico, pero son escasos los cráneos o fragmentos que
permitan determinaciones confiables a nivel genérico y aún específico (Gasparini,
1981). Los crocodilomorfos cenozoicos en América del Sur son Caimaninae, gaviales
Gryposuchinae y mesosuquianos Sebecidae (Gasparini, 1981; Brochu, 2003; Aguilera et
al. 2006); restos muy fragmentarios de dudosa asignación son considerados
pertenecientes a Crocodylidae (Patterson, 1936; Báez & Gasparini, 1979). De todos
estos taxa, los gryposuquinos y sebecidos están extintos y solo los aligatóridos
caimaninos están presentes en la actualidad en casi todo el continente, mientras que los
crocodílidos están restringidos al norte. Los restos de cocodrilos cenozoicos en territorio
argentino son relativamente abundantes, pero en calidad y variedad comparativamente
pobres.
En América del Sur, el primer gavial aparece en depósitos del Oligoceno Tardío-
Mioceno Temprano de la parte norte del continente, y en el Mio-Plioceno los gaviales
sudamericanos desaparecen del registro fósil (Rovereto, 1912; Langston, 1965; Sill,
1968 y 1970; Báez & Gasparini, 1979; Gasparini, 1968 y 1981; Taplin & Grigg, 1989;
Kraus, 1998; Brochu & Rincón, 2004; Vélez Juarbe et al., 2006; Salas Gismondi et al.,
2007; Riff & Aguilera, 2008).
Otros crocodilimorfos conocidos en Sudamérica durante gran parte del Terciario
son las formas pertenecientes a los Sebecidae, que se registran desde el Paleoceno hasta
el Mioceno. Fueron peculiares cocodrilos predadores de hábitos terrestres de los
bosques tropicales, y quizás estaban en la cima de la cadena trófica de esos ambientes

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 10
(Langston, 1965; Báez & Gasparini, 1979; Gasparini, 1972 y 1981; Salas Gismondi et
al., 2007; Molnar, 2010).
Los Alligatoridae están pobremente representados en el Paleógeno de
Sudamérica, y hasta el momento no han sido registrados en rocas del Cretácico Tardío
de este continente (Bona, 2007). A Caimaninae son asignados la mayoría de los restos
de Crocodylomorpha sudamericanos cenozoicos. El registro fósil de los caimanes
muestra que fue diverso tanto en especies como en adaptación ecológica, pero
relativamente pobre y fragmentario, con pocas localidades que brindaron restos de estos
crocodilianos (Fig. 8). El registro más antiguo es del Paleoceno del Estado de Rio de
Janeiro (Brasil), y consiste de fragmentos recuperados de la Cuenca de Itaboraí
(Gasparini, 1981; Aguilera et al., 2006).
Uno de los taxa formalmente descripto es el caimanino basal Necrosuchus
ionensis Simpson, 1937 del Paleoceno Temprano de la Formación Salamanca del este
de Patagonia, uno de los registros más antiguo de aligatóridos en América del Sur
(Simpson, 1937; Pascual & Odreman Rivas, 1971; Báez & Gasparini, 1979; Gasparini,
1981; Gasparini et al., 1986; Brochu 1999a y en prensa; Aguilera et al., 2006). Este y
otros registros de Argentina son demasiado fragmentarios como para proveer un mayor
conocimiento de la temprana evolución del grupo.
Eocaiman al igual que Necrosuchus, representa uno de los pocos taxones de
aligatóridos caimaninos conocidos del Paleógeno de América del Sur, y quizás es la
forma más basal del grupo. Este género se encuentra registrado en Patagonia en el
Paleoceno Inferior (Formación Salamanca) por E. palaeocenicus Bona 2007, en el
Eoceno Tardío por E. cavernensis Simpson 1933, y en el Mioceno Medio de Colombia
por Eocaiman sp. (Simpson, 1933 y 1937; Langston, 1965; Pascual & Odreman Rivas,
1971; Báez & Gasparini, 1979; Gasparini, 1981; Gasparini et al., 1986; Brochu 1999a,
2003; Aguilera et al., 2006; Bona, 2007). La presencia de estas especies da sustento a la
hipótesis de una temprana diversificación de los Caimaninae en el Cenozoico
sudamericano (Brochu, 1999a; Bona, 2007).
El origen de las especies de caimanes actuales es paleontológicamente incierto
(Brochu, 2003). El linaje Caimaninae incluye al caimán enano Paleosuchus que es
prácticamente desconocido en el registro fósil a pesar de la ocurrencia del grupo ya en

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 11
el Paleógeno; restos muy fragmentarios fueron asignados a ese género (Salas Gismondi
et al., 2007). El registro Oligoceno de los caimanes es muy fragmentario, y la posición
filogenética de estos restos es incierta (Báez & Gasparini, 1979; Chiappe, 1988;
Aguilera et al., 2006). Caiman tremembensis del Oligoceno de Brasil quizás sea la
especie más antigua del género (Chiappe, 1988).
Sin dudas, la mayor diversidad de cocodrilos en Sudamérica ocurre durante el
Mioceno-Plioceno (Rovereto, 1912; Langston, 1965; Báez & Gasparini, 1979;
Gasparini, 1981; Gasparini et al., 1986; Taplin & Grigg, 1989; Cione et al., 2000;
Sánchez Villagra & Aguilera, 2006; Salas Gismondi et al., 2007). Hay varias especies
descriptas de diferentes formaciones que compartían la mayoría de los géneros; todos
los morfotipos actuales y otros extintos coexistieron (Aguilera Villagra & Aguilera,
2006). El Nettosuchidae Mourasuchus de hocico largo, ancho, plano y con un gran
número de dientes pequeños, es uno de los morfotipos crocodilianos extintos más
extraños de todos los tiempos. Los netosúquidos eran un grupo endémico de ambientes
tropicales mésicos del continente sudamericano, desde el Mioceno Tardío al Pleistoceno
más temprano; Orthogenysuchus olseni del Eoceno temprano, aparentemente representa
un netosúquido de América del Norte (Langston, 1965 y 1966; Báez & Gasparini, 1979;
Bocquentin & Souza Filho, 1990; Brochu 1999a, 2003). Otro extraño caimán es el
gigante Purussaurus de Brasil, Colombia y Venezuela, con cráneo de hasta casi un
metro y medio de longitud (Gervais, 1876; Mook, 1921a; Langston, 1965; Aguilera et
al., 2006; Salas Gismondi et al., 2007). Aunque estrechamente relacionado a los
netosúquidos, es considerado un anti-netosúquido ya que tiene un hocico ancho, alto,
con narina externa extremadamente grande en relación al tamaño del hocico, y dientes
robustos. Algunos especímenes de Purussaurus fueron muy grandes que alcanzaron una
longitud corporal de 15 m; Mourasuchus también alcanzó grandes y similares
dimensiones (Brochu, 1999a y 2003).
En el Plioceno del Este Salta se registra un género y una especie existente
(Caiman latirostris) (Patterson, 1936); restos del Mio-Plioceno de Brasil y Venezuela
fueron asignados a esta especie (Hsiou & Fortier, 2007), demostrando que a fines del
Neógeno C. latirostris tenía una distribución geográfica mucho más extensa que en la
actualidad. El reciente Caiman yacare es conocido desde el Plioceno de Argentina
(Gasparini, 1981), y Melanosuchus (M. fisheri) del Mioceno Tardío de Venezuela

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 12
(Medina, 1976). Además, registros fragmentarios relacionados a Caiman de edad
Mioceno-Plioceno son conocidos de la provincia de Entre Ríos (Argentina) (Rovereto,
1912; Patterson, 1936; Báez & Gasparini, 1979; Gasparini, 1981; Cione et al., 2000;
Piña & Argañaraz, 2000).
Algunos restos hallados en Paraná, que no se apartan de Caiman o del género
aliado Melanosuchus, indican la presencia de formas gigantes. La variedad y el tamaño
de algunos ejemplares, sugieren que predominaban condiciones climático-ambientales
óptimas en latitudes donde hoy habitan sólo dos especies de mediano tamaño (Rovereto,
1912; Pascual & Rivas, 1971; Báez & Gasparini, 1979; Gasparini, 1981; Gasparini et al.
1986; Brochu, 1999a; Cione et al., 2000; Piñas & Argañaraz, 2000; Aguilera et al.,
2006). Cuatro especies de yacarés (los actuales Caiman yacare y C. latirostris, y los
extintos C. australis y C. lutescens), el netosúquido Mourasuchus nativus, y el gaviálido
Gryposuchus neogaeus, muestran la variedad de formas y también las condiciones para
la proliferación de esos reptiles en el “Mesopotamiense” (Mio-Plioceno) del Paraná
(Rovereto, 1912; Gasparini, 1968 y 1981; Báez & Gasparini, 1979; Pascual & Rivas,
1971; Gasparini et al., 1986; Bocquentin & Souza Filho, 1990; Cione et al., 2000; Piñas
& Argañaraz, 2000). A partir del Pleistoceno el registro fósil de Alligatoridae en
Sudamérica es escaso, y está constituido por restos muy fragmentarios (Hsiou & Fortier,
2007).
La gran diversidad de la fauna de cocodrilos del Cenozoico en Sudamérica es
notable. Las especies son referibles a cinco clados (Sebecidae, Crocodylidae,
Alligatoridae, Gavialidae, Nettosuchidae), pero en la actualidad solo dos de esos grupos
están presentes (Crocodylidae y Alligatoridae). La evidencia disponible sugiere que los
grupos más conspicuos fueron los Sebecidae y Alligatoridae, este último estuvo
representado por todo el Cenozoico y es el clado crocodiliano más importante hoy en
día (Báez & Gasparini, 1979; Gasparini, 1981; Brochu 1999a, 2003; Aguilera et al.,
2006; Bona, 2007; Hsiou & Fortier, 2007; Martin, 2008).
I.C. Diversidad Actual de Alligatoridae en Argentina:
En Argentina se citan dos especies vivientes de caimanes, Caiman yacare y C.
latirostris (Burmeister, 1880; Piña et al., 2004; Larriera & Imhof, 2006; Campos et al.,

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 13
2010; Verdade et al., 2010). Para C. latirostris algunos autores consideran dos
subespecies, C. latirostris latirostris y C. latirostris chacoensis (Cei, 1993); sin
embargo estas nominaciones están en desuso (Crea et al., 1999; Verdade & Piña, 2006 y
2007).
La presencia de un aligatórido sugiere cuerpos de agua, vegetación, y un clima
subtropical a templado cálido y húmedo (Gasparini, 1981). El yacaré overo (Caiman
latirostris) habita los humedales de Sudamérica, y su área de distribución comprende el
noreste de Argentina, el sur de Brasil, Paraguay, Uruguay, y Bolivia (Fig. 9A). El
yacaré negro (C. yacare) comparte un amplio sector de su área de distribución con la de
C. latirostris, habitando los humedales del sur y del este de Bolivia, noreste de
Argentina, sur y centro-oeste de Brasil, y Paraguay (Fig. 9B) (Schmidt, 1928; Prado,
2001; Piña et al., 2004; Larriera & Imhof, 2006; Verdade & Piña, 2006 y 2007; Campos
et al., 2010; Verdade et al., 2010).
En Salta hay poblaciones de las dos especies citadas para la Argentina, siempre
en el extremo noreste de la provincia. Poblaciones de Caiman latirostris (C. l.
chacoensis según Cei, 1993) se encuentran sobre las cuencas del río Pilcomayo, y del
rio Bermejo; mientras que poblaciones marginales de C. yacare habitan la cuenca del
río Pilcomayo, en el departamento Rivadavia (Cei, 1993; Piña et al., 2004; Larriera &
Imhof, 2006).
I. D. Antecedentes:
El primer trabajo en el que se mencionó por primera vez un caiman fósil es el de
Bravard (1858), quien nominó los fragmentos exhumados de las capas del Paraná (Entre
Ríos) erróneamente como Crocodylus australis (=Caiman australis). En 1876, Gervais
describió a Dinosuchus terror (=Purussaurus brasiliensis) a partir de una gran vértebra
hallada en el Amazonas (Brasil) (Plioceno?). Burmeister (1884) hizo una revisión del
material descripto por Bravard, pero mantuvo la asignación taxonómica propuesta por
este autor. Scalabrini en 1887, mencionó a varias especies de reptiles fósiles del Paraná
entre ellas la de Bravard, y propuso a Crocodylus paranensis y Alligator paranensis
como nuevos taxa.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 14
Ambrosetti (1887) describió nuevos restos de cocodrilos fósiles del Paraná, y
propuso a Proalligator australis como nuevo género para la especie de Bravard (1858),
considerando que se trata de un aligatórido y no un crocodílido como se pensaba.
Posteriormente, Rovereto (1912) revisó el material que describieron Bravard (1858),
Burmeister (1884), Scalabrini (1887), y Ambrosetti (1887). En su trabajo re-describió y
propuso los caimanes fósiles Alligator australis (=Caiman australis), A. lutescens (=C.
lutescens), y A.? ameghinoi; las especies Crocodylus paranensis y Alligator paranensis
de Scalabrini (1887) fueron reunidas bajo el mismo nombre (Alligator australis).
Restos aislados y transportados que fueron encontrados en el río Purus (Brasil),
permitieron a Mook (1921a) definir un nuevo género y especie de cocodrilo fósil al que
nominó Brachygnathosuchus braziliensis. Fragmentos óseos y diente de mamífero
extinto asociados determinaron una edad pliocena para el fósil.
Simpson en 1933, describió a Eocaiman cavernensis a partir de un cráneo y
mandíbulas incompletos provenientes del Eoceno Temprano (“Notopstylops Beds”) de
las barrancas del Lago Colhué Huapí de la provincia de Chubut. El registro confirma la
presencia de aligatóridos en el Paleógeno de América del Sur, siendo uno de los más
antiguos y más austral de este continente. En aspecto es considerado una forma
evolutivamente moderna teniendo en cuenta que es del Terciario Temprano. El autor no
descartaba la posibilidad de que haya sido el ancestro de Caiman y Jacare
(=Melanosuchus), y también señaló posibles relaciones entre Eocaiman y el aligatórido
basal Allognathosuchus del Eoceno de América del Norte. Sugirió que Eocaiman y
Allognathosuchus son parte de un grupo de origen geográfico y filogenético común;
evidencia para una conexión faunística y geográfica entre Norte y Sudamérica cerca del
inicio del Terciario.
Patterson (1936) dio a conocer un fragmento rostral bien preservado que refirió a
Caiman latirostris. El fósil fue hallado en Quebrada Aguas Blancas (Salta), varios
kilómetros (“250 miles west of the present range of latirostris”) fuera del rango de
distribución de la especie en la actualidad, junto con restos de mamíferos fósiles que
permitieron datar una edad pleistocena. El autor también realizó un breve análisis de los
Crocodylia fósiles de Sudamérica encontrados hasta ese entonces, y consideró entre
otras cosas que posiblemente Dinosuchus, Brachygnathosuchus y Purussaurus sean
sinónimos al igual que las especies ameghinoi y lutescens de Rovereto.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 15
Simpson (1937) describió a Necrosuchus ionensis proveniente de la Formación
Salamanca (Paleoceno Temprano) de Patagonia, y lo refirió a los Leidyosuchinae
descriptos para América del Norte. A pesar que se hallaron también restos
postcraneales, la descripción se realizó a partir de un fragmento mandibular anterior de
la rama derecha. Según Simpson, Necrosuchus ionensis está estrechamente relacionado
a Leidyosuchus del Cretácico Tardío-Paleoceno de América del Norte, y consideró que
representan un grupo de origen común en el Cretácico. A pesar que los asignó como
perteneciente a los Crocodylidae, reconoció ciertas similitudes de ambos taxa con los
Alligatoridae. El autor también hizo un resumen taxonómico de los Crocodylia
sudamericanos del Cretácico y Eoceno.
Langston (1965) proporciono nuevos taxa de Crocodylomorpha del Cenozoico
de Colombia, con descripciones detalladas y comentarios sobre sus relaciones con otros
fósiles de Sudamérica, la validez de las entidades taxonómicas hasta entonces descriptas
fue discutida. Los caimaninos que describió fueron Eocaiman sp. (el registro más
septentrional del género), Nettosuchus atopus, Balanerodus logimus, Caiman cf. C.
lutescens, y Caiman neivensis (=Purussaurus neivensis). Descartó la asignación
genérica de Alligator lutescens de Rovereto (1912) y lo asignó al género Caiman,
reconoció similitudes entre esta especie y Caiman yacare o C. crocodilus, y también
consideró que los fragmentos de C. lutescens del Paraná descripto por Rovereto no
serían del mismo animal. Destacó semejanzas entre Nettosuchus atopus, los caimanes y
Brachygnathosuchus braziliensis de Mook (1921a), considero que esta especie es
sinónima de Dinosuchus terror (Gervais, 1846). Reconoció también relaciones entre
Notocaiman stromeri y Eocaiman. En 1966, el mismo autor sinonimizó Nettosuchus y
Mourasuchus y los incluyó en una nueva familia, los Nettosuchidae.
Medina en 1976 describió a Melanosuchus fisheri, proveniente de la Formación
Urumaco (Mioceno Tardío) del norte de Venezuela. Esta especie constituye el primer y
único registro fósil de éste género de caimán.
Gasparini (1981) hizo una revisión de los Crocodylomorpha del territorio
argentino, éste análisis se completa con comentarios sobre su evolución, sus relaciones
filogenéticas, su clasificación, sus implicancias estratigráficas, y aportes al
conocimiento de diferentes ambientes del pasado sobre la base del registro de estos
reptiles. Ella descartó como válida la especie Notocaiman stromeri de la Formación Rio

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 16
Chico (Paleoceno Medio-Tardío) de Chubut, pero que no deja de ser un registro valioso
de aligatórido para ese tiempo y a altas latitudes. Citó una placa dérmica procedente de
la Formación Maíz Gordo de la Quebrada del Puesto en Mina Aguilar (Jujuy),
constituyendo el único elemento que demuestra la presencia de cocodrilos en esa área
durante el Paleoceno Tardío. Consideró a Caiman lutescens como sinónimo de C.
latirostris. Refirió restos a Caiman cf. C. yacare y a Caiman sp. provenientes del
“Mesopotamiense” (Mio-Plioceno) del Paraná.
Bocquentin Villanueva (1984) dio a conocer un nuevo Nettosuchidae y lo
nominó Mourasuchus arendsi, un peculiar cocodrilo “hocico de pato” proveniente de
sedimentitas de la Formación Urumaco del Estado de Falcón, Venezuela. La edad
inferida para esos sedimentos es Mioceno Tardío (Huayqueriense).
Chiappe en 1988 describió al pequeño caimán Caiman tremembensis, cuyos
restos fueron exhumados de sedimentos de la Formación Tremembé (Oligoceno) del
Estado de São Paulo, Brasil. La especie corresponde al registro más antiguo del género.
Según el autor, el pequeño tamaño de éste caimán sugiere que vivió en condiciones
menos favorables que las especies actuales, y que la escasez de cocodrilos fósiles en el
Oligoceno de Patagonia responden a un descenso de la temperatura desde ese periodo.
Bocquentin & Souza Filho (1990) a partir de restos más completos del Mioceno
Tardío del Estado de Acre (Brasil), consideraron a Carandaisuchus del Paraná sinónimo
de Mourasuchus del centro-norte de Sudamérica. De este modo, “Carandaisuchus”
nativus es considerado una especie de Mourasuchus siendo el registro más meridional
del género. Según los autores, la extensión del rango paleogeográfico de Mourasuchus
también confirma la existencia de conexiones faunísticas en el Mio-Plioceno entre la
cuenca superior del Río Amazonas y el Río Paraná.
Brochu (1999a) hizo una revisión de los Alligatoroidea, la taxonomía y
relaciones filogenéticas de los integrantes son reconsideradas, y su estudio se completa
con comentarios sobre paleobiogeografía del grupo. De sus resultados se destacan la
monofilia de los Alligatoridae como así también la de Alligatorinae y Caimaninae; a
pesar de tener una distribución muy disyunta, las dos especies actuales de Alligator
están estrechamente relacionadas; los caimanes Caiman latirostris, C. lutescens y
Melanosuchus forman una tricotomía no resuelta; Leidyosuchus y que su relativo

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 17
Necrosuchus ionensis son aligatoroideos basales; Eocaiman es un Caimaninae basal; y
la afinidad de los netosúquidos y de Purussaurus a los caimanes. Consideró a partir de
varias evidencias que los caimanes y aligátores divergieron en Norteamérica durante el
Cretácico Tardío, y que un simple evento de dispersión en o cerca del límite Cretácico-
Terciario es suficiente para explicar la presencia de caimanes en Sudamérica. El extenso
análisis es cita permanente en trabajos posteriores de otros autores.
Piña & Argarañaz (2001) dieron a conocer fragmentos mandibulares de Caiman
sp. para el Mioceno Superior-Plioceno de la Formación Ituzaingó (Entre Ríos). Los
fragmentos fueron comparados con los correspondientes de las especies actuales de la
región y C. lutescens de Rovereto (1912), concluyendo que esta última es una especie
válida más relacionada a C. yacare que a C. latirostris.
Starck & Anzótegui (2001) discutieron las causas que podrían haber sido
responsables del cambio climático ocurrido en el Mioceno Tardío, mediante el análisis
estratigráfico, sedimentológico y paleontológico de distintas formaciones del Cenozoico
aflorantes en el noroeste argentino, en particular en la zona del Valle Calchaquí donde
es muy evidente tal cambio. En el trabajo citaron un fragmento mandibular de Caiman
sp. que fue extraído de estratos de la Formación Palo Pintado de edad mamífero
“Huayqueriense-Montehermosense” (Mioceno Superior-Plioceno), y otros restos de
mamíferos, tortugas Chelidae, peces, moluscos, plantas, y algas provenientes del Valle
Calchaquí.
Aguilera et al. (2006) describieron y nominaron a Purussaurus mirandai de la
Formación Urumaco (Mioceno Superior) Venezuela. El análisis filogenético, usando la
matriz de datos de Brochu (1999a), soporta la asociación de las especies de Purussaurus
y confirma la relación de grupos hermanos del género con los Nettosuchidae.
Salas Gismondi et al. (2007) presentaron nuevos registros de caimaninos en la
región del Amazonas peruano para el Mioceno Medio. Los fragmentos fueron asignados
a nivel genérico, y destacaron la misma asociación con la de La Venta en Colombia, y
otras regiones tales como el Acre de Brasil, Urumaco de Venezuela, y en alguna medida
el Paraná. Estas similares asociaciones diversas y dispersas de crocodilianos también
fueron comentada por Sánchez Villagra & Aguilera (2006).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 18
Hsiou & Fortier (2007) dieron a conocer un fragmento de Caiman sp. para el
Pleistoceno (edad mamífero Lujanense) del Estado de Rio Grande do Sul, Brasil. Esta
pieza constituye el primer y único registro de caimanes para el Pleistoceno de América
del Sur. El fósil fue exhumado de depósitos lacustres de una planicie costera donde
también se hallaron restos de mamíferos, fragmentos de Testudine y del Teiidae
Tupinambis. El análisis filogenético que se realizo usando la matriz de Brochu (1999a),
dio sustento a la relación del fósil con el género Caiman.
Bona (2007) describió Eocaiman palaeocenicus, una especie de caimanino
basal, a partir de restos mandibulares provenientes de los niveles superiores de la
Formación Salamanca (Paleoceno Inferior) de la provincia del Chubut, Argentina. Para
resolver su sistemática también utilizó el análisis filogenético de los Alligatoroidea
propuesto por Brochu (1999a). La presencia de esta especie en Paleoceno Inferior de
Patagonia, da sustento a la hipótesis de una diversificación temprana de los Caimaninae
en América del Sur, y constituye uno de los pocos caimanes conocidos del Paleógeno de
este continente.
Numerosos son los autores que estudiaron y citaron caimanes fósiles
provenientes de Sudamérica y el territorio argentino en particular, hasta el momento en
Salta sólo dos fueron las localidades (Quebrada Aguas Blancas y Valle Calchaquí) que
brindaron fósiles de estos cocodrilos. Sin embargo, mucho de ese material es muy
fragmentario y escaso que requieren revisión para una adecuada determinación
taxonómica, principalmente los provenientes de los estratos del Paraná. También es de
remarcar que no se hicieron comparaciones genéricas y específicas entre caimanes
fósiles y sus relativos actuales, en algunos casos estas comparaciones sólo estuvieron
restringidas a unos pocos taxa.
En este trabajo se presenta una descripción detallada de la osteología craneal y
ubicación sistemática del fósil recuperado en Rosario de la Frontera, comparándolo con
los caimanes vivientes en Argentina (Caiman latirostris y C. yacare) y las otras formas
sudamericanas. El estudio se completa con un análisis filogenético a partir de los datos
originales de Brochu (1999a), y con comentarios sobre el paleoambiente y la
paleobiogeografía del grupo a partir de la información que brinda el fósil, teniendo en
cuenta que estos reptiles tienen requerimientos ambientales particulares, que hacia el

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 19
final del Neógeno la distribución geográfica de los crocodilianos en Sudamérica era
mucho más amplia, y que el rango actual de dispersión de las citadas especies no llega
hasta Rosario de la Frontera. En este contexto, es significativo el hallazgo ya que
representa el primer registro de un caimán del Neógeno del Sur de la provincia de Salta,
y su estudio contribuye al conocimiento de la herpetofauna pasada desde un punto de
vista paleobiogeográfico, paleoambiental y filogenético.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 20
I. E. Objetivos:
Describir la anatomía craneal del caimán (Alligatoridae, Caimaninae) fósil
de Rosario de la Frontera del sur de Salta, Argentina.
Comparar la anatomía craneal del caimán fósil con la de los caimanes
actuales y fósiles sudamericanos a partir de las descripciones tomadas de
bibliografía.
Comparar la anatomía craneal con la correspondiente de las especies Caiman
latirostris y C. yacare que habitan al noreste de Salta, Argentina.
Ubicar el material en la jerarquía taxonómica.
Determinar las relaciones filogenéticas del caimán fósil de Rosario de la
Frontera con otros caimanes fósiles y actuales de Sudamérica.
Realizar una revisión del registro fósil de los crocodilianos con énfasis en su
paleodistribución y aspectos paleoambientales.
Inferir condiciones paleoambientales y paleoclimáticas para el área del
hallazgo desde un punto de vista paleozoogeográfico.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 21
II. MARCO GEOLÓGICO Y GEOGRÁFICO DEL ÁREA DE HALLAZGO
II. A. Procedencia Geográfica:
El fósil fue exhumado de sedimentitas del margen derecho del río Rosario-
Horcones (25º 43’ S, 64º 52’ O), a unos 13 km al este de la ciudad de Rosario de la
Frontera, y cerca del paraje Horcones (departamento de Rosario de la Frontera) (Fig.
10). La localidad de Rosario de la Frontera se encuentra a 177 km de la capital de Salta,
al sur de la provincia. Las vías de acceso desde la ciudad de Salta hasta Rosario de la
Frontera son las rutas nacionales 9 y 34, caminos que están totalmente pavimentados;
desde allí se transita un camino secundario en dirección noreste hasta el lugar del
hallazgo. Esta área se encuentra emplazada en la llanura de la porción oriental de las
Sierras de Metán, meridional a las Sierras Subandinas (Sistema de Santa Bárbara)
(González Villa, 2002).
No hay registros históricos ni prehistóricos de estos reptiles en el área de Rosario
de la Frontera. El yacaré overo y negro (Caiman latirostris y C. yacare
respectivamente) están restringidos a los humedales orientales de la ecorregión del
Chaco Húmedo, de la Selva Paranaense, de los Esteros del Iberá, y del Delta e Islas del
Paraná (Burkhart et al., 1999). El rango de distribución actual de estos caimanes no
alcanza el sur de la provincia de Salta, sólo se los encuentra en los ambientes acuáticos
del noreste.
II. B. Procedencia Estratigráfica y Edad:
Grupo Orán. Subgrupo Jujuy. Formación Piquete
Las unidades terciarias aflorantes en el área del hallazgo corresponden al Grupo
Orán (Neógeno) que está conformado por los subgrupos Metán y Jujuy (Gebhard et al.,
1974; González Villa, 2002; Salfity & Monaldi, 2006). El Subgrupo Jujuy está expuesto
en todo el margen derecho del río Rosario en el sector Rosario-Horcones, y está
integrado de base a techo por las formaciones Guanaco y Piquete (Gebhard et al., 1974;
González Villa, 2002). Las sedimentitas de donde fueron exhumados los restos del

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 22
caimán fósil, corresponden a los niveles superiores de la Formación Piquete que tiene
predominio en el río Rosario (González Villa, 2002).
La Formación Piquete, al igual que la Formación Guanaco, consta de depósitos
pardos y pardos rojizos que consisten en intercalaciones de conglomerados, areniscas,
areniscas limosas, limolitas, y delgados niveles de caliza. En la Formación Piquete, la
matriz generalmente es areno-limosa de color pardo rojizo claro a rojizo, característica
que la diferencia de la unidad precedente (Gebhard et al., 1974; González Villa, 2002).
Ambiente de Sedimentación
El ambiente de depositación de la Formación Piquete es del tipo de
sedimentación fluvial, que corresponde en su mayor parte a conos aluviales proximales,
que gradan en posiciones distales a depósitos de ríos entrelazados y de llanuras de
inundación (Gebhard et al., 1974; Salfity & Monaldi, 2006). La participación de
areniscas limosas y calizas estaría relacionada con sistemas lacustres de baja energía
(González Villa, 2002).
Edad
Los datos cronológicos y paleontológicos conocidos sobre la Formación Piquete
son escasos (González Villa, 2002). Estudios magnetoestratigráficos indican una edad
pliocena para la Formación Piquete, entre 5 y 1,3 Ma aproximadamente (Gebhard et al.,
1974; Salfity & Monaldi, 2006). La Formación India Muerta de Tucumán,
correlacionable en parte con la Formación Piquete, contiene fósiles del Plioceno
(Pascual & Odreman Rivas, 1973; Powell, comunicación personal) confirmando la
antigüedad inferida para el caimán fósil de Rosario de la Frontera.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 23
III. MATERIALES Y MÉTODOS
Las abreviaturas institucionales usadas en este trabajo son las siguientes: AC-
UNSa, cátedra de Anatomía Comparada, Universidad Nacional de Salta; BDIV-UNSa,
cátedra de Biodiversidad IV (Vertebrados), Universidad Nacional de Salta; CP-UNSa,
cátedra de Paleontología, Universidad Nacional de Salta; CV-IML, cátedra de
Vertebrados, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Tucumán; EC-
IML, cátedra de Embriología y Anatomía Comparadas, Facultad de Ciencias Naturales
e Instituto Miguel Lillo, Tucumán; FML, Fundación Miguel Lillo, Tucumán; FMNH,
The Field Museum of Natural History, Chicago; MACN, Museo Argentino de Ciencias
Naturales “Bernardino Rivadavia”, Ciudad Autónoma de Buenos Aires; MAS-PALEO,
Museo de Antropología de Salta, Colección Paleontología; MLP, Museo La Plata, La
Plata; PVL, Colección Paleontología Vertebrados, Facultad de Ciencias Naturales e
Instituto Miguel Lillo, Tucumán; RdlF, Rosario de la Frontera.
III. A. Nomenclatura:
La anatomía y nomenclatura del esqueleto craneal, fueron estudiadas teniendo en
cuenta el trabajo de Iordansky (1973) sobre el cráneo de los crocodilianos; también se
consideraron las medidas e índices craneales propuestos por el mismo autor. Las
abreviaturas de los elementos craneales se encuentran en el Apéndice II, y la topología
de los mismos esta graficada en la Fig. 11. La nomenclatura de los géneros y especies,
como así también su sistemática, corresponde a la propuesta por Brochu (1999a y
2003).
III. B. Materiales:
El ejemplar estudiado (MAS-PALEO-RdlF 001/2011) consta de dos fragmentos
rostrales correspondientes a la mitad derecha e izquierda del hocico, la caja craneal, y
las dos ramas mandibulares. La mitad derecha del hocico está completamente articulada
e integra, la mitad izquierda tiene el premaxilar desarticulado y carece del
ectopterigoides. No se conservó ninguno de los nasales ni pterigoides, y los palatinos
están incompletos y aislados.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 24
La caja craneal se conservó casi integra pero separada de los fragmentos
rostrales. La rama mandibular izquierda es la más completa, y articulada en su totalidad.
La rama derecha está dividida en un fragmento anterior formado solo por el dentario
carente del esplenial, y un fragmento posterior (angular, surangular y articular)
parcialmente articulado.
III. C. Preparación y Comparación:
El material fósil fue preparado para su estudio en el Laboratorio de
Paleontología Vertebrados del Instituto Miguel Lillo (Tucumán). El fósil había sido
limpiado en 1998 luego de su extracción, pero para tener más claridad de las suturas
craneales, se removió el sedimento residual, se ensamblaron y pegaron algunas piezas.
La limpieza se realizó bajo lupa con pequeños cinceles, púas y percutor neumático, con
la supervisión del Técnico del Laboratorio, Juan Manuel Ballesteros.
Una vez preparado se le tomaron detalladas medidas con un calibre Vernier de
precisión 0,02 mm. Los caracteres de interés taxonómico fueron fotografiados con una
cámara fotográfica digital para su posterior análisis y comparación.
Para el estudio morfológico comparativo, se tuvieron en cuenta las descripciones
sobre caimanes fósiles y actuales hechas por Burmeister (1880), Rovereto (1912), Mook
(1921a, c y d, 1923), Schmidt (1928), Simpson (1933, 1937), Patterson (1936), Kälin
(1955), Medem (1955, 1958, 1960), Langston (1965, 1966), Iordansky (1973), Medina
(1976), Bocquentin Villanueva (1984), Chiappe (1988), Crea et al. (1989), Bocquentin
& Souza Filho (1990), Cei (1993), Piña & Argañaraz (2000), Aguilera et al. (2006),
Verdade & Piña (2007), Hsiou & Fortier (2007), Bona (2007), Rueda Almonacid et al.
(2007), y Brochu (1999a y en prensa).
Se revisó un cráneo del crocodílido Crocodylus niloticus, perteneciente a la
cátedra de Paleovertebrados del Instituto Miguel Lillo, como único representante del
grupo externo (Gavialis gangeticus y Crocodylus acutus). Se le tomaron también
fotografías que se usaron para la comparación con los aligatóridos.
Se revisaron también cráneos de Caiman latirostris de las cátedras de Anatomía
Comparada y Paleontología General de la Universidad Nacional de Salta, de las
cátedras de Vertebrados y Embriología y Anatomía Comparadas del Instituto Miguel

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 25
Lillo de Tucumán. Los cráneos de Caiman yacare revisados provienen de las cátedras
de Biodiversidad IV de la Universidad Nacional de Salta y de Embriología y Anatomía
Comparadas del Instituto Miguel Lillo de Tucumán.
El Dr. Diego Pol del Museo Egidio Feruglio de Trelew (Chubut), cedió
gentilmente material fotográfico de Caiman latirostris del Field Museum of Natural
History de Chicago (EE.UU.), y de Caiman yacare del Museo de La Plata. Del mismo
modo, Santiago Javier Nenda de la División Herpetológica del Museo Argentino de
Ciencias Naturales “Bernardino Rivadavia”, aportó fotografías de cráneos de las
mismas especies. En ambos casos, el material brindado sirvió para la revisión y
comparación de caracteres con el caimán fósil.
III. D. Análisis Filogenético:
La metodología adoptada para determinar las relaciones filogenéticas entre el
fósil y los demás taxa fue la Cladística, siguiendo el principio de parsimonia como
criterio de optimalidad, que selecciona la topología con el menor número de
transformaciones de carácter necesarias para explicar la variación observada (Farris,
1983).
Para resolver la sistemática de MAS-PALEO-RdlF 001/2011 y proponer una
hipótesis de relación filogenética entre el fósil y los demás grupos de cocodrilos, éste
fue incluido en un análisis filogenético utilizando datos del trabajo sobre la filogenia de
los Alligatoroidea propuesto por Brochu (1999a), incluyendo y/o eliminando taxa y/o
caracteres según trabajos posteriores presentados por el mismo autor (2000, 2004a y b,
2006, 2007, 2010, en prensa).
El análisis fue restringido a los Alligatoridea Caimaninae, y para ello se
incluyeron como grupo interno a todas las especies del clado provenientes de América
del Sur consideradas por Brochu (1999a, en prensa), más una especie del clado
Alligatorinae: Alligator mississippiensis. Los estados fueron polarizados para una mejor
comparación con un grupo externo integrado por la especie americana Crocodylus
acutus del clado Crocodylidae, y el asiático Gavialis gangeticus del clado Gavialidae.
De esta forma se tuvieron en cuenta los tres grandes crown group de Crocodylia:
Gavialidae, Crocodylidae y Alligatoridae (sensu Brochu, 2003). El grupo interno esta

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 26
formado por 15 especies de aligatóridos como terminales, incluyendo el fósil presentado
en este trabajo.
A partir de la lista original de 164 caracteres discretos propuestos por Brochu
(1999a) para 67 terminales, sólo se incluyeron aquellos caracteres relacionados con la
osteología craneal, los caracteres no informativos relacionados con tejido blando,
postcráneo y escutelación, fueron excluidos. Además se sumaron seis nuevos caracteres
(104-109) obtenidos del análisis del fósil, de material actual y de bibliografía, se incluyó
también el carácter (103) propuesto por Bona (2007). La lista completa de caracteres
utilizados en el análisis y la definición de sus estados se encuentra en el Apéndice III.
Para el análisis filogenético se construyó una matriz de 109 caracteres craneales
discretos y 17 terminales. Todos los caracteres utilizados recibieron el mismo peso, los
caracteres multiestados fueron considerados no aditivos. Para los caracteres que no se
pudieron tomar datos, debido a que no se cuenta con una muestra suficiente o no se
disponen material para su estudio, y en el caso de un fósil en el cual no se preservaron
partes relevantes o está en muy mal estado de conservación, se los ingreso en la matriz
con el símbolo (?). La matriz de datos se presenta en el Apéndice IV.
La matriz fue analizada utilizando el programa TNT (Tree analysis using New
Technology, Goloboff et al., 2003). La elección del software TNT se debió a la
velocidad que tiene el programa para realizar y comparar resultados aplicando
diferentes opciones de análisis, en poco tiempo (Goloboff et al., 2008b).
A través del comando “Tradicional Search”, se realizaron búsquedas heurísticas
con permutación de ramas TBR, “Tree Bisection Reconnection”. Para cada búsqueda se
hicieron 500 réplicas de secuencias de adición aleatoria, guardando 50 árboles por
réplica; es decir, 500 secuencias de adición con la opción de encontrar la/las topología/s
más parsimoniosa/s hasta 50 veces. De esta manera se puede encontrar con mayor
probabilidad el/los árbol/es más parsimonioso/s.
Se realizaron corridas con Pesos Iguales y Pesos Implicados. Éste último
procedimiento de pesado utiliza una función cóncava con constante K pesando en contra
las homoplasias (Goloboff, 1993). La constante K varia de cero a infinito, siendo
menores los valores que hacen a esta función más estricta. En éste trabajo se realizaron

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 27
análisis utilizando valores de 3-7 para K, los cuales representan un rango de valores de
la constante que incluye un valor débil entre los más estrictos (3) y un valor fuerte entre
los menos estrictos (7) (Goloboff, 1993; Quinteros, 2011).
A fin de obtener una medida de soporte de los resultados obtenidos, se realizaron
pruebas de Bootstrap y de Symmetric Resampling. En ambos casos se realizó una
búsqueda de árboles con 500 replicas, y una probabilidad de deleción de 0,33 en el
último caso.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 28
IV. RESULTADOS
IV. A. Sistemática Paleontológica:
CROCODYLIA Gmelin, 1789 (sensu Benton & Clark, 1988)
ALLIGATORIDAE Cuvier, 1807 (sensu Norell et al., 1994)
CAIMANINAE Norell, 1988 (sensu Brochu, 2003)
Género Caiman Spix, 1925
Caimán latirostris (Daudin, 1802)
Material. MAS-PALEO-RdlF 001/2011, fragmentos craneales y mandibulares
asociados con esqueleto postcraneal parcial.
Procedencia. Río Rosario-Horcones (25º 43´ S y 64º 52´ O), departamento de Rosario
de la Frontera, provincia de Salta, Argentina.
Ocurrencia. Sedimentitas rojizas de la Formación Piquete (Plioceno), Subgrupo Jujuy,
Grupo Orán.
Diagnosis. Hocico tan largo como ancho, cresta preorbital y rostral prominente, 18
alvéolos dentarios, 5 alvéolos premaxilares, 13 alvéolos maxilares, parietal excluido del
borde posterior de la tabla craneal, profundas fosetas en el margen lateral del paladar
para los grandes dientes mandibulares 1º y 4º pero sin llegar a perforarlas, 4º diente
maxilar caniniforme, dientes cónicos gruesos, fenestras palatales con estrecha
participación del pterigoides en su borde posterior, fenestras supratemporales reducidas,
contacto nasal-frontal, y lacrimal mayor al prefrontal, son el conjunto de características
que diagnostican a la especie (Mook, 1921c; Schmidt, 1928).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 29
IV.B. Descripción de MAS-PALEO-RdlF 001/2011:
IV.B.1. Cráneo
Forma general
El cráneo es corto y ancho, de contorno subtriangular y aplanado dorso-
ventralmente, hocico tan largo como ancho. El cráneo es acinético. Los márgenes
laterales que llevan los dientes son ondulados, tanto vertical como horizontalmente,
formando tres arcos convexos y dos arcos cóncavos intermedios, con una constricción
bien marcada a nivel de la sutura premaxilar-maxilar. El borde dorsal de la rama
mandibular también tiene este patrón particular de ondulación, los arcos convexos del
borde dorsal de la mandíbula están opuestos a los arcos cóncavos del margen lateral de
la mandíbula superior, y viceversa. El borde anterior del hocico es redondeado y un
poco abultado, el extremo anterior de la mandíbula inferior en oclusión permanece
posterior a ese borde.
La mandíbula es larga, ancha y baja, aunque más alta en su extremo posterior.
La sínfisis mandibular se extiende hasta el 5º alveolo del dentario. La mandíbula carece
de un proceso coronoides y de un prearticular, pero tiene un gran proceso retroarticular.
Los dientes son de implantación tecodonta y sólo están en el premaxilar, maxilar y
dentario. En oclusion, la hilera de dientes de la mandíbula inferior es medial a la hilera
de dientes de la mandíbula superior como en todos los aligatóridos, diferencia que lo
distinguen de los crocodílidos y gaviálidos en los cuales las hileras ocluyen linealmente.
Los cuadrados están muy inclinados por lo cual la unión mandibular esta desplazada
posteriormente.
La parte dorsal y postorbital del techo craneal es plana (tabla craneal), y muestra
salientes posterolaterales. La tabla craneal es de contorno rectangular, más ancho que
larga, mucho más ancha en el margen posterior, y cóncava en vista occipital. Las barras
postorbitales, aunque no conservadas en su totalidad, son más profundas que los huesos
adyacentes superficiales. Como en todos los arcosaurios, la región temporal es del tipo
diápsido con dos tipos de fenestras una superior y otra inferior. Las fenestras post-
temporales son reducidas, pero a diferencia de algunos arcosaurios tiene unas fenestras
mandibulares externas y otras palatales. El espacio interfenestral de las fenestras

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 30
supratemporales es más estrecho que el espacio interobital. La región órbito-temporal de
la caja craneal esta osificada por un par de lateroesfenoides que protegen el encéfalo a
nivel del foramen oval. Característica compartida con todos los crocodiliano es la
ausencia de hueso supratemporal, postfrontal, septomaxilar, y epipterigoides; también
carece de foramen pineal o parietal. La superficie occipital es vertical (tabla occipital).
Carácter exclusivo de los eusuquianos, es el desarrollo del paladar secundario
óseo formado por los procesos palatales de los huesos premaxilar, maxilar, palatinos y
pterigoides. La consecuencia del paladar secundario es la extensión posterior del pasaje
nasal y su separación de la cavidad bucal, una especialización del cráneo crocodiliano.
El pasaje nasal, que esta constituido por los huesos del paladar secundario, recorre toda
la longitud del hocico y termina sobre el margen posterior central de los flancos
pterigoides en una narina o coana interna, ventral a la base de la caja craneal; los
pterigoides y por tanto las narinas internas no se preservaron. El premaxilar es
expandido postero-dorsalmente y rodea la narina externa separándola del maxilar. La
sutura premaxilar-maxilar sobre el paladar no es transversa sino aguda anteriormente.
La sutura palatino-pterigoidea aparentemente es transversa. La lista completa de
medidas e índices craneales se encuentra en el Apéndice V.
Ornamentación
El cráneo está fuertemente ornamentado, y en menor medida las mandíbulas. La
superficie craneal está ornamentada con depresiones subcirculares profundas
(esculturación craneal), consecuencia de osificaciones dérmicas secundarias bien
desarrolladas (osteodermos). Estas osificaciones, bien marcadas en los individuos
adultos, se adhieren firmemente a la superficie de los huesos premaxilares, maxilares,
nasales, frontales, prefrontales, lacrimales, yugales, parietal, escamosales, postorbitales,
y cuadradoyugales, de igual manera a los dentarios, surangulares y angulares de las
mandíbulas.
Una considerable cresta interorbital constituye cada límite supraorbital. Este par
de crestas se extiende hacia delante desde la esquina anterior de las órbitas, sobre los
huesos prefrontales, lacrimales, y maxilares, terminando en una expansión sobre el 4º
alvéolo maxilar. Las dos crestas contrarias están conectadas a nivel de la esquina

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 31
anterior de las órbitas, por una cresta transversal anterior a la región interorbital y sobre
el extremo anterior del frontal. El borde posterior de esta cresta es una ligera elevación
de la región interorbital cóncava, mientras que el borde anterior sobresale fuertemente.
En su conjunto, estas crestas forman una U abierta bien característica que constituye la
cresta preorbital o rostral.
Una cresta poco elevada se extiende hacia delante cerca de la narina externa
sobre la sutura premaxilar-maxilar-nasal. Entre esta cresta y la mitad posterior de cada
borde lateral de la narina externa, que es elevada y rugosa, hay una leve depresión.
IV.B.2. Cavidades del cráneo
Fenestras supratemporales (fst)-Fig. 12
Las fenestras supratemporales son pequeñas, pares, de forma oval irregular y
levemente diferentes una de la otra, la fenestra supratemporal derecha es más alargada
que la izquierda. Están a nivel de los huesos circundantes en un plano horizontal sobre
el techo craneal, y separadas por un puente relativamente estrecho. Delimitadas por el
parietal, postorbital y escamosal.
Fenestras infratemporales (fit)-Fig. 12
Estas cavidades pares son pequeñas, pero más grandes que las fenestras
supratemporales, y triangulares en contorno. Está relativamente más completa la
fenestra infratemporal izquierda. Sobresalen en vista dorsal por el borde lateral de la
tabla craneal. Están limitadas por el postorbital y su rama descendente, el yugal y su
rama ascendente, y por el cuadradoyugal (la rama ascendente del cuadradoyugal no se
preservo, pero se observa su superficie de contacto sobre el cuadrado).
Órbitas (or)-Fig. 12
Las órbitas son de tamaño mediano, un poco más grande que las fenestras
infratemporales, de contorno subtriangular y desplazadas latero-dorsalmente. Tienen un
margen medial redondeado y un margen lateral un poco cóncavo, el margen posterior es
casi recto. La órbita izquierda es la más completa y se extiende aproximadamente hasta
el 13º alvéolo maxilar. Están delimitadas por el yugal, lacrimal, prefrontal, frontal,

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 32
postorbital, y la barra postorbital (constituida por la rama ascendente del yugal y la rama
descendente del postorbital). El espacio interobital es cóncavo y relativamente amplio,
comparado al espacio interfenestral. Están unidas por su esquina anterior por la cresta
transversal del relieve rostral.
Narina externa (ne)-Fig. 12
La narina externa es más ancha que larga, localizada sobre la superficie dorsal
del hocico cerca de su extremo anterior, y rodeada por el premaxilar, de esta forma el
maxilar es excluido de la narina. Los nasales no se han preservado, pero el contorno de
su sutura indica que integraban el margen posterior de esta cavidad. La porción
premaxilar del margen posterior medial se curva hacia adelante levemente, y quizás
junto con el extremo anterior de los nasales formaban una conspicua proyección hacia
delante, pero sin llegar a formar un septo internarial. Los márgenes del premaxilar
alrededor de la narina externa son engrosados, de forma tal que la abertura está por
encima del nivel de los otros huesos adyacentes del hocico.
Foramen incisivo o premaxilar (fi)-Fig. 13
El foramen incisivo se forma entre los procesos palatales del premaxilar, por
debajo de la narina externa. Los bordes laterales de esta cavidad están curvados, más
aun en su porción posterior, estos bordes se unen anteriormente en una terminación
aguda. Su borde posterior no pasa el límite del 5º alvéolo premaxilar.
Fenestras palatales o suborbitales (fpal)-Fig. 13
Estas aberturas pares son de gran tamaño, de contorno subtriangular y anchas en
su parte posterior. Sus márgenes lo constituyen los palatinos, ectopterigoides, maxilares
y pterigoides (aunque no están preservados). El margen medio formado por los
palatinos es muy curvado, el margen lateral formado por los proceso de los maxilares y
ectopterigoides es recto. La fenestra palatal derecha es la más completa, y su esquina
anterior alcanza el 7º alvéolo maxilar.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 33
Narina interna o coana (ni)-Fig. 13
Esta cavidad no se preservo, pero se puede inferir que se localizaba en una
posición posterior en el paladar (debido a la extensión del pasaje nasal por el paladar
óseo secundario), en el centro del margen posterior de los amplios flancos pterigoideos
(que tampoco se preservaron), y antero-ventral al cóndilo occipital. La ubicación de la
narina interna descripta es una característica compartida por todos los eusuchianos (Fig.
38).
Fenestras post-temporales (fpt)-Fig. 15
Estas cavidades pares reducidas y estrechas, se ubican lateralmente al
supraoccipital entre los escamosales y por encima de los exoccipitales, debajo del borde
posterior de la tabla craneal superficial. Están muy separadas una de la otra por el
supraoccipital.
Fosa del aductor mandibular (fad)-Figs. 17 y 18
Es una abertura amplia (en sentido antero-posterior) y alta de la superficie
postero-medial de la rama mandibular. La esquina anterior de esta fosa está formada por
el coronoides, el margen ventro-medial por el angular, el margen posterior por el
articular, y el margen dorso-lateral por el surangular. Constituye la amplia entrada al
canal mandibular que estrecha hacia delante. El canal mandibular (=primordial o
meckeliano) está cerrado lateralmente por el dentario y medialmente por el esplenial. La
fosa del aductor mandibular mejor preservada es la de la rama mandibular izquierda.
Foramen intermandibular caudal (fic)-Fig. 17
Es una pequeña abertura de forma oval un poco estirada, que ubica en la cara
medial o lingual de la rama mandibular por debajo de la fosa del aductor mandibular. El
borde anterior esta formado por el esplenial, y el borde posterior lo constituye dorso-
ventralmente el angular.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 34
Fenestra mandibular externa (fme)-Fig. 16
Es una característica de todos los arcosaurios, de gran tamaño y de contorno
triangular. El margen dorsal es levemente curvado, los márgenes posterior y ventral son
casi rectos que entre ambos un ángulo mayor a los 90º. Se ubica sobre la pared lateral de
la fosa del aductor mandibular comunicándola con el exterior. Esta delimitada en su
esquina anterior y gran parte de su margen dorsal por el dentario, la esquina posterior
por el surangular, y el margen ventral por el angular.
Foramen magnum (fm)-Fig. 15
Es una gran abertura ubicada en el centro de la tabla occipital por debajo del
supraoccipital, y por encima del cóndilo. Esta rodeado lateralmente por los exoccipitales
y ventralmente por el basioccipital. De contorno subromboidal, y similar en diámetro al
cóndilo occipital.
IV.B.3. Huesos craneales
En esta sección, los huesos del cráneo se describirán según la región a la que
pertenecen, como sigue: 1) dorsal, 2) lateral o mejilla, 3) occipital, y 4) basal o palatal.
IV.B.3.1 Norma dorsal:
Premaxilares (pm)-Figs. 12, 13 y 14
Estos huesos están bien desarrollados y son considerablemente más anchos que
largos. Rodean a la narina externa, y en esta región son más abultados. Cada uno tiene
un gran proceso ascendente posterior a la narina, estos procesos están acuñados entre los
nasales y maxilares, separando a este último de la narina externa. Contactan en la mayor
parte de su lado posterior con el maxilar. La sutura premaxilar-maxilar se extiende a
ambos lados sobre la superficie del hocico en sentido oblicuo, hasta el nivel del 3º
alvéolo maxilar. Presentan un proceso palatal que se unen en la línea media y
constituyen la porción más anterior del paladar secundario óseo, el foramen incisivo se
ubica en la sutura palatal entre premaxilar. Sobre el paladar óseo, la sutura premaxilar-
maxilar es transversa ahusándose hacia delante.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 35
Cada premaxilar contiene cinco alvéolos, cada uno equidistante del otro. El
premaxilar derecho conserva sólo el diente 3º y 4º, ambos funcionales y completos, el
izquierdo conserva todos los dientes, parcialmente completos y funcionales. El diente 4º
derecho esta hipertrofiado (diente caniniforme), aunque el correspondiente izquierdo no
es funcional o está poco desarrollado su alvéolo es el de mayor diámetro.
Los procesos palatales de los premaxilares llevan un par de fosetas grandes por
detrás del 1º y 2º alvéolo, que hacen de estuche para recibir al 1º gran diente del
dentario cuando las mandíbulas están cerradas, pero sin llegar a perforarlas. A pesar que
estos dientes del dentario no se preservaron, el tamaño se deduce por el diámetro de los
alvéolos. Similares fosetas están situadas en la región de la sutura premaxilar-maxilar,
que sirven para recibir al mayor 4º diente del dentario en oclusión. Otras pequeñas
fosetas para recibir a los contrapuestos mandibulares, también están presentes detrás de
los restantes alvéolos.
Nasales (n)-Fig. 12
Ningunos de estos elementos están preservados, pero en base al espacio entre los
maxilares, se puede observar que eran huesos relativamente cortos y anchos, su ancho
máximo es inmediatamente anterior a la sutura nasal-lacrimal. Los bordes laterales de
los nasales convergen anteriormente de forma gradual, desde el extremo anterior de la
sutura nasal-lacrimal al extremo posterior de la sutura premaxilar-nasal; la terminación
anterior de los bordes laterales de los nasales entre los premaxilares, converge
agudamente.
Su porción anterior llega a participar del margen posterior de la narina externa
en un proceso internarial, pero sin llegar a formar un verdadero septo. En su porción
posterior suturan con los lacrimales, prefrontales, y estrechamente con el extremo
anterior agudo del frontal. Las suturas nasal-lacrimal y nasal-prefrontal son
aproximadamente iguales en longitud.
Lacrimales (la)-Fig. 12
Solo está preservado parcialmente el lacrimal izquierdo, y un fragmento del
derecho suturando con el yugal. Son grandes, y su amplitud es mayor que su longitud.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 36
Se localizan en la esquina anterior de las órbitas, la cual constituye el límite posterior.
Están perforados en su margen postero-medial por el conducto lacrimal. Medialmente
hacen contacto con los nasales y prefrontales, lateralmente con los maxilares y yugales.
La sutura lacrimal-prefrontal se extiende a través de la cresta transversal cerca de la
esquina anterior de la órbita. La sutura con los yugales está sobre las prolongaciones
anteriores de ellos en el margen inferior de las órbitas.
Prefrontales (prf)-Fig. 12
Están preservados, tanto el izquierdo como el derecho, en su totalidad. Consisten
en placas alargadas sobre la superficie dorsal medial del hocico (próximos a las órbitas),
y procesos descendentes (pilares prefrontales) en la parte interna del paladar; solo se
aprecia la base de estos pilares a ambos lados de las crestas craneales del pasaje nasal en
la cara interna del frontal (Fig. 39C). La placa dorsal del prefrontal es puntiaguda
anteriormente, y encaja entre los lacrimales y nasales; los lacrimales impiden el
contacto maxilar-prefrontal. Los prefrontales están expandidos superficialmente el
proceso anterior del frontal, haciendo de este una terminación aguda que levemente
parece alcanzar los nasales. Forman parte el borde antero-medial de las órbitas. La
sutura con los nasales es aproximadamente igual en longitud a la sutura nasal-lacrimal.
La cresta transversa atraviesa los prefrontales, y es interrumpida medialmente en el
contacto de estos con el frontal.
Frontal (fr)-Fig. 12
Es un hueso impar y alargado, que forma el techo de la parte anterior de la caja
craneal. Su ancho máximo está en el espacio interorbital, siendo ahí sus bordes curvados
medialmente, pero es poco angosto por el desplazamiento medial de las órbitas. Los
laterales del proceso anterior permanecen estrechos a los prefrontales, pero alcanza la
parte posterior de los nasales, el proceso anterior comprende el puente entre la región
interorbital y el hocico. El extremo anterior del proceso agudo separa los extremos
posteriores de los nasales. Posteriormente, el frontal está firmemente suturado al
parietal, y en menor medida a los postorbitales, ventralmente a los lateroesfenoides. La
sutura con el parietal y los postorbitales es en forma de una amplia U, siendo casi recta
sobre el parietal. Está situado a una considerable distancia anterior a las fenestras

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 37
supratemporales. La cresta craneal frontal se extiende a los lados de la superficie ventral
del mismo, como un par de crestas descendentes que protegen el tracto olfativo de los
bulbos olfativos del encéfalo.
Parietal (p)-Fig. 12
Al igual que el frontal es un hueso impar, que comprende la mayor parte del
techo de la caja craneal. Es una amplia placa subrectangular que articula anteriormente
con el frontal (en una línea ligeramente curva), por su esquina anterior con los
postorbitales, y a los escamosales por su esquina posterior, posteriormente hace
contacto (en sutura ligeramente convexa) con el supraoccipital. El contacto con los
postorbitales y escamosales es en línea oblicua. No alcanza el borde posterior de la tabla
craneal. Forma una gran porción del margen medio y anterior de cada fenestra
supratemporal, siendo estrecho entre ellas.
Postorbitales (po)-Fig. 12
Los postorbitales constituyen las placas dorsales horizontales que forman las
esquinas antero-laterales de la tabla craneal postorbital, sus procesos descendentes se
unen con los procesos ascendentes de los yugales y forman las barras postorbitales. Los
procesos descendentes están desplazados un poco hacia adentro de la superficie craneal,
empezando desde la superficie ventral de la placa postorbital dorsal. No se descarta, a
pesar de estar un poco deteriorada esa región, el proceso descendente posterior adicional
que se extendería postero-ventralmente, desde la base del proceso anterior a lo largo del
margen posterior de la fenestra infratemporal. Articulan por su esquina antero-lateral
con el frontal, lateralmente con el parietal formando parte del borde lateral de las
fenestras supratemporales, y por su lado posterior contactan con los escamosales en
línea recta. Son relativamente pequeños y subcuadrados en contorno.
Escamosales (esc)-Figs. 12 y 15
Son huesos son grandes consistentes en una amplia placa dorsal horizontal sobre
la tabla craneal, que se extiende latero-posteriormente por encima de la región del oído
externo formando una profunda cavidad (receso ótico externo). Se expanden
posteriormente obliterando en parte las fenestras post-temporales de la tabla occipital.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 38
Contacta ampliamente la superficie anterior de los procesos paraoccipitales y la
superficie postero-dorsal del cuadrado posterior al oído medio. Suturan anteriormente
con los postorbitales, medialmente con el parietal y supraoccipital, latero-ventralmente
con el cuadrado, y postero-ventralmente con el paraoccipital; constituyen el borde
posterior de las fenestras supratemporales.
IV.B.3.2. Norma lateral:
Maxilares (m)-Figs. 12, 13 y 14
Los maxilares forman la parte principal del hocico. Sus procesos palatales, en
unión en la línea media, forman una considerable porción del paladar secundario óseo.
Tanto el derecho como el izquierdo están relativamente completos. Separados de las
órbitas por los yugales y lacrimales. Sobre la superficie dorsal tienen un par de
abultamientos por encima del alvéolo del 4º diente maxilar (diente caniniforme), que
constituye el extremo anterior de la cresta rostral lateral del relieve osteodérmico, llevan
la principal porción de la prominente cresta preorbital en forma de U. El límite posterior
de cada maxilar en la superficie dorsal, es transversal en su porción interna, la porción
externa (debajo de la órbita y en el límite dental) es oblicua y su extremo posterior se
curva hacia arriba.
Cada maxilar contiene trece alvéolos, de los cuales el 4º es el más grande. Sólo
se preservaron en el maxilar derecho los dientes 3º, 6º, 7º, 8º, 11º, y 12º (no todos
completos y funcionales), en el maxilar izquierdo se conservaron los dientes 1º, 2º, 3º,
4º, 5º, 6º, 8º, 10º, 11º y 12º, parcialmente completos y desarrollados. El espacio entre los
alvéolos es aproximadamente igual, excepto entre el 5º premaxilar-1º maxilar (el mayor
de toda la serie), y entre el 4º-7º maxilar. Los procesos palatales muestran una serie de
forámenes, relacionados con órganos sensoriales y vasos sanguíneos, dispuestos
medialmente a la hilera de alvéolos. La ondulación (festonamiento) del margen lateral,
tanto vertical como horizontal, es más profunda a nivel de la sutura premaxilar-maxilar,
y en menor medida en el espacio entre el 5º-7º diente maxilar.
Dorsalmente contactan en amplia sutura paralela al plano sagital con los nasales.
Por el borde posterior contactan con los lacrimales y yugales, anteriormente con el
premaxilar. En el lado palatal contactan con su porción posterior a los palatinos, y con

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 39
sus ramas laterales a los ectopterigoid. Forman la esquina anterior y parte del borde
lateral de las fenestras palatales.
Yugales (y)-Figs. 12 y 14
Son huesos relativamente elongados que se extienden a lo largo de los laterales
del dermatocráneo, desde la esquina anterior de las órbitas hasta el nivel de los cóndilos
mandibulares del cuadrado; son los huesos de las mejillas. A cada yugal le falta un
fragmento de su porción anterior. La parte principal de los yugales (proceso anterior y
posterior) constituye de una placa, cuya su superfície se extiende latero-dorsalmente.
Los procesos anteriores forman la pared lateral de las órbitas, mientras que los procesos
posteriores forman el arco temporal inferior, una curvatura pronunciada que constituye
el límite inferior de la fenestra infratemporal. El proceso yugal anterior cubre
ampliamente la parte posterior de los maxilares, y se extiende hacia adelante alcanzando
el 9º alvéolo maxilar; el contacto dorsal con los lacrimales es breve. Los procesos
posteriores suturan oblicuamente con los cuadradoyugales. Sólo se preservaron la base
de los procesos ascendentes dorsales que formaban la parte inferior de la barra
postorbital, aunque en la izquierda un poco más conservada está inclinada medialmente
respecto a la placa yugal.
Cuadradoyugales (cy)-Fig. 12
Los cuadradoyugales son placas pequeñas encajadas entre los yugales y los
cuadrados en la esquina postero-lateral del cráneo. Cada cuadradoyugal consiste en una
placa y un delgado proceso ascendente (incompleto en ambos lados) que se extiende a
lo largo del margen posterior de la fenestra infratemporal, quizás alcanzando el
postorbital o escamosal, esto se deduce por la superfície de contacto sobre el cuadrado.
Cuadrados (c)-Figs. 12, 13 y 15
Los cuadrados están en íntimo contacto con los huesos de la caja craneal, tienen
una superfície articular distal transversalmente ancha (cóndilos mandibulares). Los
cóndilos mandibulares están marcadamente inclinados postero-ventralmente, y
desplazados hacia atrás más allá del nivel del cóndilo occipital. La terminación interna
de los cuadrados se extiende anterior y medialmente a la fosa temporal del cráneo. La

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 40
superfície medial del cuadrado está en intimo contacto con la superfície lateral de la caja
craneal, posterior al foramen oval. Por su superfície antero-dorsal están firmemente
suturados con los escamosales, y por su lado posterior con la superfície anterior de los
procesos paraoccipitales. La extensión anterior de los cuadrados y la expansión postero-
lateral de los escamosales, ubican el oído medio dentro de una cavidad profunda, ventral
a estos últimos y anterior a los procesos paraoccipitales. Sobre la superficie antero-
ventral de los cuadrados hay un par de crestas que sirven la insercion de músculos
mandibulares.
Los cóndilos mandibulares de los cuadrados están en la esquina postero-lateral
del cráneo, y la superficie condilar se proyecta postero-ventralmente. Los dos cóndilos
del cuadrado son diferentes en tamaño, el cóndilo externo o lateral es mayor al interno o
medio. Un pequeño foramen aéreo se abre sobre la superfície postero-dorsal de los
cuadrados encima del cóndilo interno, que se conecta con el foramen aéreo del articular
de la mandíbula inferior.
IV.B.3.3. Norma occipital:
Supraoccipital (so)-Figs. 12 y 15
El supraoccipital es una placa relativamente triangular que se ubica en la parte
dorsal de la tabla occipital. Este hueso tiene una proyección dorso-medial que alcanza la
superfície dorsal de la tabla craneal y contacta por detrás al parietal formando una
pequeña placa cóncava de contorno subrectangular, de esta manera separa el parietal del
margen posterior de la tabla craneal. Articula lateralmente con los escamosales, tanto en
la tabla craneal como occipital, delimitando por debajo las fenestras post-temporales.
Ventralmente contacta con los exoccipitales, finalizando en una punta muy cerca del
borde dorsal del foramen magnum.
El límite dorsal de los laterales del supraoccipital, ventral a las rudimentarias
fenestras post-temporales, forman un par de de procesos dirigidos posteriormente
llamados procesos postoccipitales; estos procesos no estan completamente preservados
(Fig. 40). En esos procesos se insertarían los tendones del M. spinalis capitis.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 41
Exoccipitales (eo)-Fig. 15
Son huesos amplios que ocupan la mayor parte de la tabla occipital. Forman la
pared posterior de la caja craneal, confluentes con los opistóticos, y constituyen los
laterales del cóndilo occipital. Los procesos paraoccipitales, formados por la fusión de
los laterales de los exoccipitales y los paraoccipitales, están en íntimo contacto con los
cuadrados. El deprimido y amplio pasaje cranio-cuadrado, está representado por el
estrecho canal que se extiende entre los procesos paraoccipitales y los cuadrados. El
canal cranio-cuadrado se extiende desde la superfície occipital, ventral a la parte lateral
de los procesos paraoccipitales, hasta la cavidad del oído medio. Este canal provee el
pasaje para la rama principal del nervio VII, la arteria orbitotemporal, y la vena cefálica
lateral.
Los procesos paraoccipitales son verticales, sus margenes ventrales están
medialmente continuados por crestas por encima de las aberturas posteriores de los
pasajes cranio-cuadrados. Debajo de estas crestas, la superficie de los exoccipitales es
cóncavas y muestra sobre sus largos procesos descendentes (laterales al basioccipital)
unas tres aberturas, un pequeño foramen medio para el nervio craneal XII, lateralmente
un foramen vago relativamente grande, y ventralmente un foramen carótido posterior o
lateral que permite la entrada de la arteria carótida interna a la cavidad del oido medio.
Por encima del foramen magnum, los exoccipitales se unen excluyendo al
supraoccipital del límite dorsal de ésta abertura. Los extremos ventrales de los
exoccipitales (procesos descendentes) no llegan al margen ventral del basioccipital,
permanecen dorsal a éste. Contactan en la esquina latero-dorsal de la tabla occipital con
los escamosales.
Basioccipital (bo)-Fig. 15
El basioccipital, de contorno rectangular, se localiza sobre la superfície ventral
de la tabla occipital como una placa vertical (proceso basioccipital), ventral al cóndilo
occipital; este último está formado en su totalidad por el extremo dorsal del
basioccipital. Un leve engrosamiento se extiende a lo largo de los márgenes ventral y
lateral de la placa basioccipital, este engrosamiento forma la tuberosidad basioccipital.
La placa occipital lleva una cresta media, la tuberosidad y esta cresta sirven como

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 42
superfície de unión para los tendones del M. basioccipitovertebralis (=M. rectus capitis
anterior) y el M. occipitalistransversalis profundus (=M. longissimus capitis). El
basioccipital forma la parte posterior del piso de la caja craneal, lateralmente contacta
con los procesos descendentes de los exoccipitales, y su cara anterior está en íntimo
contacto con el basiesfenoides.
IV.B.3.4. Norma palatal:
Palatinos (pal)-Fig. 13
Los dos palatinos están casi completos y en contacto en la línea media.
Comprenden el piso y la pared lateral del pasaje nasal, posterior al proceso palatal de los
maxilares. Son más anchos en su porción anterior, donde articulan con los procesos
palatales posteriores de los maxilares en forma de una amplia U invertida, más estrechos
en la región que constituye el límite medial de las fenestras palatales. Su extremo
posterior se expande lateralmente y contacta con los pterigoides (que no se preservaron)
aparentemente en línea recta, su extremo anterior alcanza el nivel del 6º alvéolo maxilar.
Pterigoides (pt)-Fig. 13
No se preservó ninguno de estos elementos, no se puede saber la proporción que
ocupaban en el márgen posterior de la fenestras palatales. Habrían estado formado por
amplios y altos flancos o alas pterigoideas, conteniendo en su margen posterior a la
narina interna (condición eusuchiana). Habrían tenido contacto anterior con los
palatinos y las fenestras palatales, y lateralmente con los ectopterigoides.
Ectopterigoides o transversales (ec)-Figs. 13 y 14
Sólo se preservó casi en su totalidad el ectopterigoides derecho. Son huesos
cortos de forma irregular que llevan tres procesos. El proceso anterior, relativamente
corto y agudo, se extiende anteriormente a lo largo de los márgenes laterales de las
fenestras palatales próximos a la hilera de alvéolos maxilares, llega a forman la mitad
posterior del margen lateral de las fenestras. El proceso descendente (más largo y
agudo), de dirección postero-ventral, estaría en contacto con la esquina antero-lateral de
las alas pterigoideas. El corto proceso ascendente se extiende a la base de la barra
postorbital, contacta con el yugal y quizás con el proceso descendente de los

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 43
postorbitales. La sutura de los ectopterigoides con los maxilares alcánza el nivel del 10º
alvéolo maxilar.
Basiesfenoides (bs)-Fig. 13
Están pobremente preservados y diferenciados. Los basiesfenoides, así como el
basioccipital, se extienden ventralmente. Los cuadrados, y los pterigoides a pesar de no
estar preservados, contactarían con la superfície anterior del basiesfenoides ventral y
lateralmente; la parte principal del basiesfenoides estaría oculta por los pterigoides. En
vista ventral aparece como una lámina que forman el margen anterior del foramen de
Eustaquio, anterior a la tuberosidad del basioccipital.
Lateroesfenoides (ls)-Fig. 13
Al igual que el basiesfenoides, estos elementos están muy deteriorados. Los
lateroesfenoides forman la parte anterior de las paredes laterales de la caja craneal, por
delante del foramen oval. Postero-lateralmente contactan con los cuadrados,
dorsalmente parecen alcanzar el frontal, y quizás el parietal. Los extremos antero-
dorsales de los dos lateroesfenoides y el techado del frontal, definen el foramen
redondeado para el tracto olfativo del encéfalo.
III.B.4. Mandíbula-Figs. 16, 17 y 18
A continuacion se listan y describen los elementos que constituyen las
mandíbulas.
Dentarios (d)
Los dentarios ,preservados en su totalidad, comprenden la porción más grande y
anterior de la longitud mandibular. Son más bajos en su extremo anterior, cada uno esta
ramificado posteriormente en dos procesos (uno dorsal y otro ventral) que forman la
esquina anterior de la fenestra mandibular externa. Se articulan por su extremo anterior
formando una corta sínfisis mandibular de sección oblicua, suavemente inclinada hacia
abajo. La ancha y baja sínfisis mandibular se extiende hacia atrás hasta el nivel del 5º
alvéolo. El proceso ventral del extremo posterior contacta con el angular por debajo de

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 44
la fenestra mandibular externa, el proceso dorsal contacta con el surangular.
Lingulamente articulan con el esplenial formando la pared lateral del canal mandibular.
Cada dentario contiene dieciocho alvéolos, en la rama mandibular izquierda se
conservaron doce dientes (3º, 4º, 5º, 6º, 8º, 9º, 10º, 12º, 13º, 14º, 16º, y 18º), en la rama
derecha sólo once (5º, 6º, 7º, 9º, 11º, 12º, 14º, 15º, 16º, 17º, y 18º). El 4º alvéolo es el
más grande de toda la serie mandibular, seguido en tamaño por el 1º. El 4º diente de la
rama izquierda (caniniforme), a pesar de estar quebrado en el ápice es el de mayor
tamaño. El 4º diente encaja en una foseta formada sobre la sutura premaxilar-maxilar en
el márgen lateral del paladar. El espacio interalveolar entre el 1º y 4º alvéolo es
equidistante, entre el 4º y 9º es más estrecho y subigual, desde el 9º es mayor al de los
precedentes pero no supera al de los primeros cuatros alvéolos. Medial a la hilera de
alvéolos se observan una serie de forámenes u orificios dentales. Desde el 5º alvéolo, las
ramas mandibulares se estrechan y mantienen la mismo ancho a lo largo de la hilera de
alvéolos.
El festonamiento del borde dorsal de los dentarios presenta dos arcos cóncavos
bien marcados, uno entre el 1º y 4º alvéolo y otro entre el 4º y 12º, detrás de éste último
alvéolo hay una ligera concavidad hasta el final de la hilera. Dos arcos convexos
pronunciados también forman parte del festonamiento mandibular, uno sobre el 4º y el
otro sobre 12º alvéolo. La altura mandibular a nivel del 1º y 4º alvéolo es ligeramente
igual a la altura a nivel del 11º y 12º.
Surangulares (san)
Los surangulares son largos, aplanados e inclinados en sentido medial. La cara
interna sirve para la inserción del M. aductor mandibular externus superficialis. Forman
la esquina postero-dorsal de la fenestra mandibular externa, ventral a ésta esquina
contactan con el angular en toda su longitud posterior, la superfície postero-medial
sutura con el articular formando la pared lateral de la fosa glenoidea (=fosa articular), y
se extienden hacia atrás hasta el extremo posterior del proceso retroarticular.
Constituyen el límite latero-dorsal de la fosa del aductor mandibular. La porción antero-
medial contacta con la rama dorsal del coronoides y con el esplenial. Una pequeña
faceta o superfície de articulación sobre la sutura surangular-coronoides, sirve para la

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 45
inserción de la aponeurosis coronoides (conjunto de fibras del tendón del aductor
mandibular). El extremo anterior de los surangulares, tambien llamado proceso
surangular, está bifurcado en un extremo dorsal y otro ventral, subiguales en longitud se
apoyan sobre el dentario. El extremo dorsal del proceso surangular se extiende hacia
delante contactando con el último alvéolo de la hilera alveolar. La lámina de la sutura
surangular-articular, en la fosa del aductor, es ventral al foramen lingual de la superfície
medial del surangular (Figs. 46A y B).
Angulares (an)
Son huesos relativamentes largos, con un extremo anterior agudo ventralmente
acuñado entre el dentario y el esplenial, se extienden desde el nivel de la porción
posterior de la hilera de alvéolos hasta el extremo posterior del proceso retroarticular,
subiguales en longitud a los surangulares. La superfície dorsal del extremo anterior
agudo es cóncava, comprende el piso de la fosa del aductor mandibular y la porción
posterior del canal meckeliano. Las paredes de esa concavidad están formadas por un
par de crestas longitudinales (cresta medial y lateral). La cresta medial forma el margen
medio de la fosa del aductor, y rodea posteriomente al foramen intermandibular caudal
contactando con el esplenial y el proceso postero-ventral del coronoides. La cresta
lateral está en íntimo contacto con la cara interna del proceso ventral del extremo
posterior del dentario, y constituye el límite ventral de la fenestra mandibular externa.
Una cresta superficial se extiende en sentido antero-posterior a lo largo del borde
ventral curvandose posteriormente, ésta cresta separa la superfície lateral ornamentada
de la superfície medial lisa. La extremo posterior contacta dorsalmente con el
surangular, y dorso-medialmente con el articular.
Espleniales (esp)
Los espleniales son amplias placas delgadas y subtriangulares alargadas, que
forman una porción considerable del aspecto medial de las ramas mandibulares. Están
medialmente sobre los dentarios, y delimitan la pared medial del canal mandibular, el
cual contiene principalmente el cartílago de Meckel. La porción posterior del canal
mandibular es muy amplia, y por allí entrarían la rama mandibular del nervio craneal
V, vasos sanguíneos, y el M. intramandibularis. El extremo anterior de los espleniales

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 46
se extiende hacia la sínfisis mandibular pero sin llegar alcanzarla, terminando en una
punta dorsal al canal meckeliano o mandibular a nivel del 4º alvéolo. Se extienden
ventro-dorsalmente de forma gradual, contactando con el margen superior la hilera de
alvéolos, a partir del 14º constituye el límite medial de la porción posterior de la hilera
de alvéolos. El amplio extremo posterior contacta con el coronoides y el angular en
igual proporción, dorsalmente sutura con el surangular en menor medida.
En el extremo posterior de los espleniales, sobre el coronoides y angular
respectivamente, hay dos forámenes, el superior foramen intermandibular medio y el
inferior foramen intermandibular caudal. En cada caso, los espleniales constituyen el
límite anterior, los coronoides y angulares el límite posterior. El centro de cada esplenial
esta perforado por un pequeño foramen intermandibular oral. Por estos forámenes se
abren paso las ramificaciones de la rama mandibular del nervio V.
Coronoides (cor)
Son huesos relativamente pequeños restringuidos a la superfície medial
mandibular. No forman un proceso coronoides. Cada coronoides esta curvado hacia
atrás en un arco formando la esquina anterior de la fosa del aductor mandibular, y la
curvatura anterior esta en contacto con el esplenial. Los extremos del arco constituyen
dos procesos de diferente longitud, el proceso postero-dorsal forma la porción anterior
de la faceta en la cual se une la aponeurosis coronoides y contacta levemente con el
surangular, el proceso postero-ventral más largo forma la porción anterior del límite
medial de la fosa del aductor y sutura en toda su longitud ventral con el angular. Los
coronoides rodean posteriormente el foramen intermandibular medio, y no alcanzan a
formar parte del límite dorsal del foramen intermandibular caudal, con el cual están
separados por el angular y el esplenial.
Articulares (ar)
Son huesos relativamente anchos y robustos. Forman la fosa articular o
glenoidea de la unión mandibular, la cual se ubica antero-dorsalmente contrapuesta a los
cóndilos del cuadrado cuando las mandíbulas están cerradas. Una protuberancia media
divide la fosa en dos depresiones (una lateral y otra medial) donde articulan los cóndilos
lateral y medial del cuadrado, el borde dorsal del surangular constituye el límite lateral

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 47
de la fosa glenoidea, una cresta transversal posterior separa la fosa del proceso
retroarticular. Un pequeño foramen aereo se ubica en la superfície dorso-medial del
articular sobre la cresta transversal y posterior a la fosa glenoidea, que se conecta con el
foramen aereo del cuadrado.
El proceso retroarticular (proyección posterior del articular) muestra una fuerte
curvatura dorsal, es relativamente ancho, y el borde medial es convexo. El extremo
antero-ventral del articular forma una pequeña faceta, desde la cual se extiende
anteriormente el cartílago de Meckel a lo largo de la parte inferior de la fosa del aductor
y por el canal mandibular.
IV.B.5. Dentición
Los dientes son típicos tecodontes, profundamente implantados en los alvéolos
de las crestas alveolares. Estas crestas se extienden a lo largo de los márgenes laterales y
anterior del hocico, como asi tambíen a lo largo del borde dorsal del dentario. Como en
todos los crocodilomorfos, carece de dientes palatales. En oclusión, la hilera de alvéolos
de la mandíbula inferior permanecen medial a la hilera de la mandíbula superior.
Los dientes son pseudoheterodontes, ya que difieren en tamaño y ligeramente en
morfología. El diámetro de los alvéolos varía con la ondulación de los márgenes
mandibulares, los alvéolos más grandes están localizados en la porción central de los
arcos convexos, mientras que los más pequeños ocupan el centro de los arcos cóncavos
del festonamiento mandibular. Los alvéolos más grande de la mandíbula superior están
opuestos a los alvéolos más pequeños de la mandíbula inferior, y viceversa. Los
alvéolos más grandes de la mandíbula superior son el 4º premaxilar, el 4º > 8º > 9º
maxilar, en la mandibula inferior son el 1º, 4º, 12º > 13º > 11º, siendo el 4º el más
grande de está serie.
Aparte del festonamiento de los márgenes mandibulares, la pseudoheterodoncia
también está dada por la diferenciación en coronas cónicas y agudas de los grandes
dientes anteriores, y las bajas coronas romas de los pequeños dientes posteriores, todos
los dientes están curvados medialmente. Los dientes preservados son básicamente
cónicos, pero hay un ligera constricción entre la cilíndrica raíz de paredes delgadas, y

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 48
las puntiagudas o redondeadas coronas de paredes gruesas, es más pronunciada la
constricción en los dientes posteriores.
Los dos dientes conservados del premaxilar derecho están enteros, son
funcionales y agudos, presentan la superficie medial algo aplanada, y poseen una suave
carena antero-posterior. Los dientes del premaxilar izquierdo son iguales en morfología,
pero el 4º es un diente de reemplazo o no funcional que apenas asoma la cresta alveolar,
el 3º tiene el ápice roto. Todos los alvéolos premaxilares son de sección circular. En
orden decreciente, el diámetro de los alvéolos premaxilares es el siguiente: 4º > 3º > 5º
> 2º > 1º.
De los trece alveólos maxilares, en el maxilar derecho sólo se conservaron seis
dientes, de ellos el 3º y 8º son de reemplazo, 6º, 7º y 11º son funcionales y están
completos, el 12º esta quebrado. Todos presentan una carena antero-posterior y un
aplanamiento medial. El único de corona roma es el diente 11º, todos los demas son
conicos y agudos. Excepto los alvéolos 11º-13º que tienen contorno elíptico, muy
próximos uno del otros hasta con límites confluentes, todos los otros son circulares y
separados equidistantemente. En el maxilar izquierdo los dientes 1º, 2º, 6º y 8º están
partidos en la base de la corona, el 3º (parcialmente completo) junto son el 4º y 5º (que
están completos) son funcionales, agudos, recurvados, simétricos, y tienen una carena
antero-posterior, el dientes 10º a pesar de tener el ápice quebrado, parece ser de corona
roma junto con los dientes 11º y 12º que estan completos son funcionales, asimétricos, y
presentan una leve carena en sentido antero-posterior. También son de contorno
circular, con límites definidos a una cierta distancia uno del otro, excepto los tres
últimos alvéolos que son elípticos y casi confluentes. En los dos maxilares, el orden
decreciente de los alvéolos es el siguiente: 4º > 3º > 9º > 8º > 2º > 1º > 5º > 6º > 10º > 7º
> 11º > 12º > 13º.
En la mandíbula inferior el dentario derecho conserva once dientes, de los cuales
el 5º está partido en el ápice, el 6º, 7º, 9º, 11º, 12º y 14º están completos, son
funcionales, cónicos, agudos, recurvados, y simetricos en sentido labio-lingual, el 15º y
17º también están completos y funcionales pero son de coronas romas ligeramente
recurvados y asimétricos con la cara lingual menos amplia, el 16º y 18º son dientes de
reemplazo que poco se asoman. En el dentario izquierdo, los dientes 3º, 4º, 6º, 13º y 14º

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 49
están rotos en el ápice, los dientes 4º, 5º, 8º, 10º, y 12º son cónicos y agudos, los dientes
16º y 18º presentan corona roma, el 9º diente no es funcional. Todos los dientes
presentan la carena antero-posterior, ligeramente aplanados en la cara lingual, de
seccion circular, con la excepción de los tres que son elípticos. Los últimos tres alvéolos
son casi confluentes, los demás alvéolos son equidistantes, aunque entre el 1º y el 4º
alvéolo el espacio interalveolar es mayor. El diámetro de los alvéolos en orden
decreciente es el siguiente: 4º > 1º > 12º > 13º > 11º > 3º > 2º >5º = 15º > 14º > 10º > 6º
= 7º > 16º = 17º = 18º > 8º = 9º.
Fórmula dentaria
El conteo de dientes es unilateral, como no todos los dientes estan conservados,
se registró el número de alvéolos por hemimandíbula superior e inferior. Se contó el
número de alvéolos premaxilares (pm), maxilares (m), y dentarios (d). Para el caimán
fósil de Rosario de la Frontera, la fórmula dentaria es la siguiente:
pm (5) + m (13)
d (18)

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 50
IV.C. Comparaciones:
A diferencia de los Alligatoridae, los Gavialidae (Fig. 19) se caracterizan por un
largo y delgado hocico no festoneado con unos dientes supernumerarios (adaptación
para una dieta piscívora), presencia de una larga sínfisis esplenial y unas fenestras
supratemporales tan grandes como las órbitas, entre otras características (Iordansky,
1973; Mook, 1921c). Los Crocodylidae (Fig. 20) también poseen un hocico largo, pero
robusto, con perforación premaxilar de los dientes mandibulares, hendidura de oclusión
entre premaxilar y maxilar diferenciada desde temprana edad, un esplenial con
terminación anterior ventral al canal meckeliano (Fig. 42A), un 5º alvéolo maxilar
mayor, son algunas de las características que los distinguen de los aligatóridos (Mook,
1921c).
El material descripto fue comparado con aligatóridos caimaninos, tanto fósiles
como actuales, del Cenozoico de Sudamérica. Para ello se tuvieron en cuenta las
descripciones de bibliografía de trabajos previos, y en algunos casos se revisaron
cráneos y material fotográfico. También se comparó con Alligator, del Hemisferio
Norte, por ser el único representante actual de los Alligatorinae, y el género tipo de la
familia. A pesar de la falta de consenso en la taxonomia de los Crocodylia, y
Alligatoridae en particular, a principios del siglo pasado (Mook & Mook, 1940) de
donde provienen gran parte de los trabajos revisados, aquí se utilizara la taxonomia de
dominio actual que considera para los aligátoridos vivientes, los géneros Alligator
(mississipiensis y sinensis), Paleosuchus (trigonatus y palpebrosus), Melanosuchus
(niger), y Caiman (crocodilus, yacare y latirostris), con sus especies respectivas.
IV.C.1. Comparaciones genéricas
El género Alligator Cuvier, 1807 (A. mississippiensis en particular) (Fig. 21)
(Mook, 1921c y 1923; Kälin, 1955; Iordansky, 1973; Brochu, 1999a), de amplio hocico
de bordes laterales más o menos paralelos y ligero festonamiento, presenta grandes
diferencias con MAS-PALEO-RdlF 001/2011. Las características más notables y
exclusivas del género, es la presencia un septo internarial que divide la narina externa
(formado por una larga prolongacion anterior de los nasales y una pequeña proyección
posterior del margen anterior de ésta abertura formado por el premaxilar), y la ausencia

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 51
de un supraoccipital sobre la tabla craneal. El número de alvéolos del fósil está dentro
del rango de este género. Una amplia pero estrecha sínfisis se extiende hasta el 4º
alvéolo, narina externa con hendidura lateral, prefrontal mayor que el lacrimal alcanza
al maxilar impidiendo el contacto lacrimal-nasal, frontal con su extremo anterior
alcanza la porción posterior de los nasales en compleja sutura, ausencia de una cresta
preorbital (Alligator sinensis sólo presenta una cresta transversa interobital), órbitas
mayores a las fenestras infratemporales, fenestras supratemporales contenidas en unas
fosas, espacio interfenestral palatal mucho más amplio, palatinos con bullas palatinas
bien desarrolladalos, angular prolongado más allá del límite anterior del foramen
intermandibular caudal, foramen lingual sobre la sutura surangular-angular (Fig. 46D),
fenestra mandibular externa grande (foramen intermandibular caudal visible
lateralmente en forma completa), surangular truncado a nivel del borde dorsal de la fosa
articular (Fig. 44D), son características que diferencian Alligator del fósil. Coinciden en
tener un 4º alvéolo mandibular grande, cuyo diente encaja en una foseta sobre el piso
del hocico entre la sutura premaxilar-maxilar.
El género Necrosuchus Simpson, 1937 (Fig. 22) (Simpson, 1937; Kälin, 1955;
Brochu, 1999a y en prensa), presenta características que lo diferencian del material
descripto, una mandíbula relativamente larga, estrecha a través de la sínfisis (sin una
notable expansión a nivel del 4º alvéolo), dentario delgado ahusado anteriormente, y
una sínfisis mandibular prolongada hasta el nivel del 4º alvéolo. Ambos comparten el
mismo número de alvéolos, 18 en el dentario, pero hay pequeñas diferencias en el
diámetro de estos, los primeros dos alvéolos son grandes y subiguales al 4º, el cual es
tan grande como el 13º. Al menos los dos únicos diente preservados, 5º y 15º, llevan
numerosas carenas o crestas longitudinales que irrádian desde el ápice, son agudos y
recurvados lingualmente. Por monotipía, las características genéricas son válidas para la
diágnosis de Necrosuchus ionensis Simpson, del Paleoceno de Patagonia.
Eocaiman Simpson, 1933 (Fig. 23) (Simpson, 1933; Kälin, 1955; Langston,
1965; Brochu, 1999a; Bona, 2007), se distingue del material descripto principalmente
por un proceso anterior de los palatinos de contorno subrectangular, pterigoides
formando parte del margen posterior de las fenestras palatales en gran proporción,
cresta preorbital e interorbital leve, órbitas grandes con espacio interorbital reducido,
mandíbula inferior baja pero amplia en el sector anterior, altura mandibular a nivel del

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 52
1º y 4º alvéolo no menor a la del nivel del 12º, una amplia sínfisis de contorno
semicircular que alcanza el 5º o 6º alvéolo, y por tener mayor número de alvéolos
(aproximadamente 14 alvéolos maxilares y 19-20 alvéolos mandibulares). Posiblemente
el 4º alvéolo maxilar, 4º y 13º alvéolo dentario sean los más grandes de la serie, estos
dos últimos ocluyen dentro de fosetas en el paladar como en el caimán del río Rosario-
Horcones. El género ésta presente en el registro fósil desde el Paleoceno Inferior hasta
el Mioceno Medio, con dos especies descriptas hasta el momento, Eocamian
palaeocenicus Bona (Paleoceno Inferior) y E. cavernensis Simpson (Eoceno Tardío),
ambas de Patagonia, y fragmentos aislados (Mioceno Medio) de Colombia.
Con Paleosuchus Gray (1862) (Fig. 24) (Mook, 1921c; Schmidt, 1928; Kälin,
1955; Medem, 1958; Cei, 1993; Brochu, 1999a), las diferencias son marcadas. Este
género de caimanes de talla pequeña, se diagnostica por palpebrales (huesos de los
párpados) completamente osificados, fenestra supratemporal cerrada por expansión
medial de los huesos superficiales adyacentes (postorbital, escamosal, y parietal),
parietal alcanzando el límite posterior de la tabla craneal, supraoccipital triangular
pequeño, cráneo moderadamente alargado, hocico estrecho con un canto rostral
considerable (Fig. 52F), y foramen incisivo pequeño. Presenta 4 alvéolos en cada
premaxilar, y entre 15 a 16 alvéolos maxilares, 3º y 4º premaxilar y maxilar son los de
mayor diámetro, cada rama mandibular contiene hasta 22 alvéolos. Los dientes son
comprimidos lateralmente, en principal los posteriores. El frontal en su proyección
anterior es similar al presente en Alligator. En el género, el esplenial presenta una doble
perforación para la rama mandibular del nervio craneal V (Fig. 43C), el coronoides está
perforado y cubre el foramen intermandibular medio, el maxilar no tiene un proceso
posterior sobre el lacrimal, los palatinos desarrollan unos largos procesos laterales que
forman el margen anterior medio de la fenestra palatal, el proceso anterior de los
palatinos es de laterales paralelos con cara anterior invaginada, el festonamiento
mandibular es leve, y no hay una cresta preorbital e interorbital. Esta última
característica le dá el nombre de “caimanes de frente lisa” a las dos especies del género,
Paleosuchus trigonatus (Schneider) y P. palpebrosus (Cuvier), ambas encontradas
principalmente en las cuencas Amazónica y del Orinoco. No hay registro fósil de éste
género.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 53
Las especies del género Purussaurus Barbosa-Rodrigues, 1892 (Fig. 26) (Mook,
1921a; Kälin, 1955; Langston, 1965; Brochu, 1999a; Aguilera et al., 2006), son
caimanes gigantes diferenciados por un cráneo alargado, ancho y robusto, con una gran
apertura narial rodeada por una fosa formada por el canto rostral, nasales
extremadamente reducidos, prefrontales alargados mayores a los lacrimales, sin
contacto nasal-lacrimal, nasales y frontal separados por los prefrontales unidos
medialmente, frontal con una corta proyección anterior que no llega a pasar el límite
anterior de la órbitas que son pequeñas, tabla craneal reducida, fenestras
supratemporales contenidas en fosas, escamosales posteriormente altos y alargados, y
por un supraoccipital pequeño que no excluye el parietal del margen posterior de la
tabla craneal que es cóncavo profundo. La fórmula dentaria difiere de la del caimán
fósil, presenta mayor número de alvéolos, entre 14-15 maxilares y 21 mandibulares, los
primeros cincos alvéolos premaxilares y dentarios son los más grande de toda la serie
mandibular, y subiguales en tamaño. Otras diferencias son, foramen incisivo grande y
oval, sínfisis mandibular ancha y alta, el esplenial se extiende hasta el 8º alvéolo,
proceso retroarticular proyectado posteriormente y no postero-dorsalmente, fenestra
mandibular externa es grande con foramen intermandibular caudal visible lateralmente,
espacio interfenestral palatal amplio, sutura palatino-pterigoides ligeramente ondulada,
y suave festonamiento del hocico. Hasta el momento sólo hay tres especies descriptas,
Purussaurus brasiliensis Barbosa-Rodrigues, P. neivensis (Mook) y P. mirandai
Aguilera, Riff & Bocquentin-Villanueva, todas del Mio-Plioceno de la cuenca
Amazónica.
El caimán hocico de pato Mourasuchus Price, 1964 (Fig. 27) (Langston, 1965;
Bocquentin-Villanueva, 1984), presenta grandes diferencias con el fósil descripto, la
característica más notable es el hocico extremadamente largo, amplio, aplanado, de
laterales paralelos, sin festonamiento, y extremo anterior redondeado. Al igual que
Purussaurus, el extremo anterior del frontal es rudimentario y la tabla craneal muy
reducida. En éste género, la esculturación craneal es ligera, las fenestras
supratemporales son considerablemente reducidas, las órbitas son circulares pero de
menor tamaño con respecto a las fenestras infratemporales, la narina externa y el
foramen incisivo son grandes y subcirculares aunque más ancha la primera, el extremo
anterior de los palatinos esta muy dilatado, la sutura palato-pterigoidea es sinuosa, las

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 54
son mandíbulas extremadamente alargadas y delgadas en proporción al hocico
formando un arco mandibular amplio, la sínfisis mandibular y porción sinfisial son muy
reducidas. Relacionado con el alargamiento del hocico, los nasales son largos y
delgados, con un extremo anterior que se ahusa gradualmente hasta terminar en una
punta sin alcanzar la narina externa. Otra diferencia bien marcada es el gran número de
alvéolos, 5 premaxilares, 35-41 maxilares, y 44 mandibulares, cuyas dimensiones
disminuyen gradualmente de adelante hacia atrás. Los cinco primeros alvéolos son los
más grandes, aunque 4º o 5º son ligeramente de sección mayor. Los espacios
interalveolares son subiguales y mayores en la porción anterior, los posteriores son
reducidos. Una característica distintiva de Mourasuchus, es la presencia de dos o cuatro
perforaciones premaxilares, que reciben los dientes más grande de la mandíbula inferior
en oclusión. El esplenial se extiende anteriormente hasta el nivel del 10º alvéolo, y está
separado ampliamente de la sínfisis. Las órbitas presentan un reborde anterior
engrosado y alto, el espacio interorbital es estrecho. Los prefrontales son alargados,
mayores a los lacrimales, y unidos en la línea media separan los nasales del frontal, no
hay sutura lacrimal-nasal. El género tiene registro para el Mio-Plioceno de Brasil,
Colombia, y Argentina, con Mourasuchus amazonensis Price, M. nativus Gasparini, M.
atopus (Langston), M. arendsi Bocquentin-Villanueva.
Muy relacionado a Caiman latirostris con el cual comparte muchas
características, el género Melanosuchus Gray, 1862 (Fig. 28) (Mook, 1921c; Schmidt,
1928; Kälin, 1955; Iordansky, 1973; Medina, 1976; Cei, 1993; Brochu, 1999a) presenta
caracteres que no lo distinguen del fósil; sin embargo, hay diferencias. El caimán negro
es un taxón de mayor tamaño que se caracteriza por, cráneo alargado, hocico con
extremo anterior más puntiagudo que redondeado, sin hendidura bien marcada a nivel
de la sutura premaxilar-maxilar, cresta preorbital poco notoria que recuerda más a una V
que a una U, órbitas relativamente grandes, palpebrales parcialmente osificados, espacio
interorbital menor al espacio interfenestral superior, foramen incisivo alargado, sutura
palatino-pterigoidea cóncavo-convexa profunda, procesos posteriores de los palatinos
estrechos, y los amplios procesos anteriores de los palatinos se extienden hacia delante
alcanzando el 4º o 5º alvéolo maxilar. Sin embargo, la diferencia más notable y
autoapomorfia de éste género, es el vómer romboidal expuesto en el paladar sobre la
línea media entre la sutura premaxilar-maxilar por debajo del foramen incisivo. Con

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 55
respecto al número y diámetro de los alvéolos, no hay diferencias entre Melanosuchus y
fósil descripto. La única especie actual del género es Melanosuchus niger (Spix) que
habita la cuenca Amazónica, y no hay registro de su presencia en el pasado geológico;
el género esta presente desde el Mioceno tardío de Venezuela con M. fisheri Medina.
Con respecto a Caiman Spix, 1825 (Mook, 1921c; Schmidt, 1928; Kälin, 1955;
Iordansky, 1973; Cei, 1993; Brochu, 1999a), tanto las dimensiones como la morfología
craneal y mandibular del material descripto se relacionan con éste género. Se identifica
principalmente por la siguiente combinación de caracteres: palpebrales parcialmente
osificados, reborde anterior osificado (cresta pre- e interorbital) muy acentuado, hocico
con extremo anterior redondeado, borde lateral vertical y horizontal del hocico con
festonamiento bien marcado, constricción en el borde lateral del hocico a nivel de la
sutura premaxilar-maxilar, narina externa subcircular, lacrimal mayor al prefrontal
contactando anteriormente con los nasales, maxilar con proceso posterior en el lacrimal,
foramen incisivo grande, más largo que ancho, agudo en su parte anterior y redondeado
posteriormente, grandes fenestras palatales subtriangulares o romboidales, fenestras
supratemporales notorias pero pequeñas (éstas aberturas pueden estar obstruidas en el
adulto), separadas por un puente interfenestral considerable, el parietal no alcanza el
límite posterior de la tabla craneal separado por un supraoccipital considerable. La serie
alveolar esta formada por 5 alvéolos premaxilares, 13-14 maxilares, y entre 15 y 20
alvéolos en cada rama mandibular. Los prefrontales a veces se expanden medialmente
hasta llegar a separar el frontal de los nasales. En Caiman y Melanosuchus, el
postorbital tiene un proceso posterior descendente adicional al de la barra postorbital,
que se extiende postero-ventralmente, desde la base del proceso anterior a lo largo del
margen posterior de la fenestra infratemporal, separando el cuadrado y el escamosal de
ésta fenestra, éste proceso también está presente en Alligator pero es muy corto y
delgado. El relieve del hocico presenta el mismo conjunto de crestas observadas en el
fósil descripto, como asi también la presente en el género Melanosuchus. El aspecto
global craneal descripto en el fósil es coincidente con el observado en Caiman, por lo
tanto, el material de Rosario de la Frontera es asignado a éste género; sin embargo, se
han encontrado diferencias consideradas de nivel específico. El género tiene registro
desde el Oligoceno con Caiman tremembensis Chiappe, con su apogeo en el Mio-
Plioceno registrandose a C. niteroiensis Souza-Filho & Bocquentin-Villanueva, C.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 56
brevirostris Souza-Filho, C. lutescens (Rovereto), C. australis (Burmeister), y algunos
representates vivientes. Las especies actuales son C. latirostris (Daudin), C. yacare
(Daudin) y C. crocodilus (Linnaeus).
IV.C.2. Comparaciones específicas
Para la comparación específica se considerará las especies que habitan el
extremo sur de Sudamérica, Caiman latirostris y C. yacare, la especie tipo del complejo
C. crocodilus (C. crocodilus crocodilus) de Centro-Norte de Sudamérica, y las especies
fósiles C. tremembensis, C. australis, y C. lutescens. No se poseen datos de Caiman
brevirostris ni de C. niteroiensis del Brasil. La separación subespecífica del complejo
Caiman crocodilus no esta bien fundamentada, según Busack & Pandya (2001). No se
consideraran las subespecies Caiman latirostris latirostris y C. l. chacoensis, ya que
según Crea et al. (1989), los caracteres craneales que las separaban varian entre los
individuos, e incluso dentro de un mismo individuo.
La especie Caiman tremembensis Chiappe, 1988 (Fig. 29) (Chiappe, 1988),
representa una forma de talla pequeña (el holotipo corresponde a un fragmento del
dentario de longitud máxima de 61 mm), que tambien difiere del material descripto en
tener un hocico angosto, esto lo vincula a C. yacare, C. crocodilus y C. australis; sin
embargo, en estas tres especies la sínfisis es plana y amplia. La sínfisis mandibular en el
caimán de Tremembé es corta (se extiende hasta la mitad del 4º alvéolo), baja y
suavemente inclinada hacia abajo como la del fósil y Caiman latirostris, si bien estos
último son forma de hocico ancho, ambas comparten la sínfisis corta, baja y de sección
oblicua; de esta manera C. tremembensis se encuentra más relacionado con el yacaré
ñato. Sin embargo, la sínfisis de Caiman tremembensis es algo más baja y más corta que
la de C. latirostris. El festonamiento no es muy marcado, esto lo diferencia del fósil
descripto y las demás especies de caimanes.
Con respecto a Caiman lutescens Rovereto, 1912 (Fig. 31), conocido a partir de
fragmentos craneales, rostrales y mandibulares del Paraná, de La Venta en Colombia, y
de Urumaco en Venezuela (Rovereto, 1912; Patterson, 1936; Langston, 1965; Brochu,
1999a; Sánchez Villagra & Aguilera, 2007), hay diferencias notables. Esta especie es de
tamaño relativo mayor que cualquier otra del género, presenta un hocico angosto con la

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 57
región anterior más redondeada y abultada (que en vista lateral pareciera tener un
hocico arqueado hacia arriba), narina externa de diámetro mayor que cualquier otra
especie de Caiman, hendidura marginal pronunciada a nivel de la sutura premaxilar-
maxilar, los premaxilares separan ligeramente los nasales del borde posterior de la
narina externa, el esplenial se detiene a una distancia relativamente mayor antes de la
sínfisis, región sinfisial más angosta que la del fósil y Caiman latirostris, parecida a la
de C. crocodilus o C. yacare, las fenestras palatales son pequeñas con un puente
interfenestral amplio con respecto a otros caimanes y el fósil, lacrimal separado del
nasal por un amplio prefrontal, como en Alligator, grandes órbitas con espacio
interorbital mayor al espacio interfenestral superior, fenestras supratemporales estrechas
e irregulares en contorno, caja craneal alta, borde posterior de la tabla craneal cóncavo
profundo como en Purussaurrus y Mourasuchus. El relieve rostral es similar al
descripto para el fósil, con cresta pre- e interobital marcadas, también presente en
Caiman latirostris. En cuanto a las relaciones de ancho y longitud rostral, recuerda más
a Caiman crocodilus que a C. latirostris.
Caiman australis (Burmeister, 1884) (Fig. 30) (Rovereto, 1912; Patterson, 1936;
Chiappe, 1988), es conocido a partir de unos restos rostrales y mandibulares aislados
muy fragmentarios. La diferencia más notable es el el largo y estrecho hocico, lo cual lo
relaciona a los vivientes Caiman yacare y C. crocodilus, que al caimán de hocico ancho
C. latirostris. Otras diferencias con el fósil son, extremo anterior del hocico redondeado
con narina externa en abultamiento premaxilar (no tan marcado como en Caiman
lutescens), sínfisis mandibular plana y amplia, región sinfisial estrecha al igual que el
arco mandibular, lo cual se corresponde con un hocico angosto, y espacios
interalveolares muy estrechos. La morfología dental es igual a la de Caiman yacare.
Caiman crocodilus (Linnaeus, 1758) (Fig. 32) es muy similar en aspecto a C.
yacare, con el cual está estrechamente relacionado, incluso está última era considerada
una subespecie de aquella, o ambas bajo la misma especie Caiman sclerops (Mook,
1921c y d; Schmidt, 1928; Medem, 1955 y 1959; Iordansky, 1973; Brochu, 1999a;
Rueda-Almonacid et al., 2007). El cráneo de Caiman crocodilus, comparado con el
fósil descripto, es estrecho y alargado; en Caiman crocodilus apaporiensis el hocico es
mucho más angosto y largo (Fig. 25). Desde el nivel del 1º alvéolo maxilar al 4º el
hocico se ensancha rapidamente, posterior a esto hay una leve constricción la cual es

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 58
notable en el espacio entre el 5º-6º alvéolo, desde éste punto hacia atrás el cráneo se
ensancha regularmente, pero no tanto como en el fósil y Caiman latirostris. El
festonamiento vertical es mucho más marcado, a cada lado del hocico, una escotadura
se extiende desde 4º alvéolo premaxilar al 3º maxilar, siendo más profunda a nivel de la
sutura premaxilar-maxilar, lateral a la foseta que recibe al 4º diente mandibular, la
superfície superior del hocico es ligeramente cóncava. En su gran mayoría, el 1º diente
mandibular perfora los premaxilares en los adultos, y en algunos casos esas aberturas
son confluentes con la apertura narial. El 4º diente mandibular tambien perfora el piso
del paladar en su correspondiente foseta, en su mayoría perforandola, incluso
abriendose externamente siendo visible el diente en vista lateral como en Crocodylus
(Figs. 41D y 49E), este foramen puede estar en el premaxilar o sobre la sutura
premaxilar-maxilar. Los nasales usualmente entran en la narina externa en una corta
proyección, pero en algunos casos quedan excluidos por los premaxilares; la unión
postero-dorsal premaxilar es más notoria en los individuos adultos. El extremo posterior
de los nasales pueden llegar a tener un leve contacto con el frontal, pero el
ensanchamiento superficial de los prefrontales uniéndose medialmente, evita dicho
contacto, el extremo anterior del frontal no es tan agudo como el del fósil. Cada maxilar
presenta un gran abultamiento encima del 4º alvéolo, siendo prominente en los adultos.
La cresta interorbital o transversal es más marcada que la preorbital o rostral y forman
un leve relieve en U. Las órbitas son grandes, más en sentido longitudinal, la tabla
craneal presenta un margen posterior transversalemente recto, las fenestras
supratemporales en los adultos son pequeñas y pueden llegar a estar obliteradas por los
huesos circundantes, en Caiman crocodilus apaporiensis son grandes y contenidas en
unas fosas. Las fenestras palatales son más romboidales que triangulares, los pterigoides
participan en una pequeña proporción en el límite posterior de dichas fenestras, y el
proceso caudal de los palatinos no son tán expandidos lateralmente como en el caimán
de Rosario de la Frontera. Las mandíbulas son considerablemente festoneada,
especialmente en los individuos más viejos. En promedio, presenta un mayor número de
alvéolos, 5 alvéolos premaxilar, 14-15 maxilares, y 19-20 mandibulares, pero en
dimensiones no hay diferencias con el fósil descripto. Una característica bien notoria, es
la morfología de los dientes que son más puntiagudos, menos recurvados y de coronas
estriadas, como en Caiman yacare.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 59
Las especies vivientes Caiman latirostris y C. yacare que habitan el Sudeste de
América del Sur, y cuyo rango de distribución alcanza la parte Noreste de Salta, son
comparadas a continuación con el caimán fósil de Rosario de la Frontera. De estas dos
especies, no sólo se dispusó de información bibliográfica y gráfica, sino que también se
pudo revisar y medir material en mano.
Caiman yacare (Daudin, 1802) (Fig. 33) (Burmeister, 1880; Mook, 1921c y d;
Schmidt, 1928; Medem, 1959; Cei, 1993; Brochu, 1999a; Rueda-Almonacid et al.,
2007), al igual que el precedentemente descripto C. crocodilus y a diferencia del fósil,
tiene un cráneo de perfil triangular en vista dorsal, un hocico relativamente largo y
estrecho con su parte media un poco deprimida, extremo anterior redondeado y
ensanchado (no tanto como en el de Caiman lutescens y C. australis), una leve y
reducida cresta en U, órbitas subtriangulares y no redondeadas, el 1º y 4º diente
mandibular normalmente perforan la mandíbula superior en los adultos (Figs. 41C, 49C
y D, y 50C), los pterigoides entran en las fenestras palatales por un estrecho proceso en
zigzag (Fig. 53C), que a menudo es tan delgado que a primera vista pareciera que los
palatinos y los ectopterigoides estuvieran unidos, separando los pterigoides del borde
posterior de las fenestras, pero en algunos casos lo están. Esta especie difiere de su
emparentado Caiman crocodilus (que tiene palatinos de laterales subparalelos), en tener
procesos posteriores palatinos muy dilatados lateralmente, formando una curvatura
central en la sutura palatino-pterigoides; en el material fósil, estos procesos estan
deteriorados y no se puede saber la forma de la sutura palatino-pterigoides, a pesar de
que pareciera ser recta. El 4º diente mandibular también puede quedar expuesto
lateralmente en oclusión, produciendo una dentición pseudo-crocodilida. Poca
diferencia hay con el material descripto en la fórmula dentaria, tiene en promedio 19/18
alvéolos (5 premaxilares, 14-15 maxilares y 18-20 mandibulares), mayor cantidad que
el comparado y menor que crocodilus. La sutura premaxilar-maxilar en el paladar es
siempre aguda o curvada hacia adelante, el foramen incisivo es más pequeño que el del
fósil. Dorsalmente, los premaxilares se unen evitando la entrada de los nasales en la
narina externa. En esa especie, como en Caiman crocodilus, los prefrontales pueden
unirse medialmente evitando el contacto frontal-nasal, el festonamiento vertical y
horizontal son bien marcados, con una constricción relativamente profunda a nivel de la
sutura premaxilar-maxilar, y un abultamiento maxilar encima del 4º alvéolo. El ancho

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 60
anterior del hocico entre el 1º alvéolo maxilar, es igual o un poco más grande que el
ancho anterior de la tabla craneal, esta relación es mayor en el fósil. Las fenestras
supratemporales son moderadamente mayores a las observadas para Caiman latirostris
y el caimán de Rosario de la Frontera. Como en crocodilus, y a diferencia del fósil, los
dientes son muy agudos, de coronas altas y levemente estriadas. Hay dos fosetas
profundas mediales entre el 5º y 7º alvéolo maxilar, que reciben en oclusión a los
relativamente grandes dientes 10º y 11º mandibulares; éstas cavidades también están
presentes en crocodilus. Al igual que el hocico, el arco mandibular es estrecho.
Así como entre Caiman crocodilus y C. yacare no hay grandes diferencias (pero
difieren con el fósil), lo mismo ocurre con el material descripto y el yacaré ñato. El
yacaré ñato Caiman latirostris (Daudin, 1802) (Fig. 34), como su nombre lo índica, se
caracteriza por un hocico tan largo como ancho (Burmeister, 1880; Rovereto, 1912;
Mook, 1921c; Schmidt, 1928; Patterson, 1936; Crea et al., 1989; Cei, 1993; Brochu,
1999a; Verdade, 2000; Verdade & Piña, 2006 y 2007), característica compartida con el
fósil.
Caiman latirostris, al igual que el fósil descripto, es una forma de cráneo corto,
pesado, robusto, ancho y deprimido. Reconocible de C. yacare por la punta del hocico
gruesa no tán estrangulada, menos abultada y redondeada, y por la cresta ósea pre- e
interorbital en U muy prominente, que desde la región antero-orbital se extiende
lateralmente hacia la superfície dorsal de los maxilares, dilatandose y perdiendo sobre el
4º alvéolo. El ancho a la altura del límite orbital anterior, es igual o aún mayor que la
distancia desde dicho borde hasta la punta del hocico. Las fenestras palatales son de
contorno subtriangular o romboidal, con puente palatino interfenestral posterior
estrecho, palatinos con procesos postero-laterales amplios, pero no tan dilatados como
en yacare, la participación de los pterigoides en el borde posterior de las fenestras es
mínima. Las órbitas son subcirculares a subtriangulares, la cresta ósea transversa está
interrumpida medialmente por el contacto entre los prefrontales y el frontal, éste último
elemento alcanza con su proceso anterior agudo a los nasales. El yacare ñato es similar a
Melanosuchus niger con el cual está muy relacionado, pero difiere entre otras
características, en que no tiene un vómer expuesto en el paladar, cresta preorbital muy
marcada, órbitas más pequeñas en proporción al cráneo, y un hocico redondeado más
que agudo. La tabla craneal es relativamente grande, con fenestras supratemporales

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 61
pequeñas, circulares u ovales, pero núnca obliteradas. La narina externa es de contorno
circular, más grande que la de Caiman yacare y Melanosuchus, los nasales entran en
ella en forma de una pequeña proyección aguda, el foramen incisivo es amplio, como
así también los premaxilares, una ligera constricción marca la porción lateral de la
sutura premaxilar-maxilar. La mandíbula es corta y ancha, con región sinfisial amplia
que alcanza el 4º o 5º alvéolo. La sínfisis mandibular es baja, y de sección oblicua,
inclinada postero-ventralmente. En cuanto a la morfología y dimensiones de los dientes
y alvéolos no hay diferencias, pero la coronas de los dientes llegan a tener estrias,
carácter poco dilucidado en el fósil, quizás por una cuestión de edad. Esta especie tiene
una serie alveolar de 5 alvéolos premaxilares, 13-14 maxilares y 17-20 mandibulares, en
promedio 18/18; el fósil esta dentro de este rango. Ninguno de los dientes mandibulares
perfora la mandíbula superior (Fig. 41A). El festonamiento rostral y mandibular es bien
marcado, pero no tanto como en Caiman yacare. Como en el caimán fósil, la hilera de
orificios mediales a los alvéolos de la mandíbula superior, son más profundos que en
cualquiera de la especies tratadas anteriormente.
En base a lo descripto y comparado, se desprende que el caimán fósil encontrado
en Rosario de la Frontera pertenece a la especie Caiman latirostris (Daudin). El
conjunto de caracteristicas craneales (dientes inferiores de oclusión medial, sin
perforación premaxilar, supraoccipital grande sobre la tabla craneal, dientes cónicos
gruesos, hocico ancho con cresta rostral prominente, etc.), previamente descriptas en la
sección anterior, confirman esta asignación genérica y específica.
IV.D. Análisis Filogenético:
Como resultado del análisis de parsimonia de la matriz de 17 terminales y 109
caracteres usando el programa T.N.T., para Pesos Iguales se encontraron dos árboles
igualmente parsimoniosos de 176 pasos, con Índice de Consistencia (CI) de 0, 830 e
Índice de Retención (RI) de 0,800 (Fig. 35). Con Pesos Implicados también se
obtuvieron dos topologías igualmente óptimas (Figs. 36A y B) para valores de K que
variaron de 3 hasta 7. Los valores de ajuste (Fit) respectivos fueron los siguientes: K3=
6,85000; K5= 4,68452; K7= 3,56389. Los datos analizados bajo Pesos Implicados
presentan grupos mejor soportados (Goloboff et al., 2008a).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 62
Tanto con Pesos Iguales como con Pesos Implicados, las medidas de soporte
Boostrap y Symmetric Resampling arrojaron la misma topología de consenso, más
resuelta que la del Consenso Estricto (Fig. 36C), y congruente con resultados obtenidos
en trabajos previos de otros autores (Densmore & Owen, 1989; Poe, 1996; Brochu,
1997a y b , 1999a y b, 2001a, 2003, 2004b, 2010, en prensa; Lutzoni et al., 2000;
Harshman et al., 2003; Gatesy et al., 2003; Aguilera et al., 2006; Bona, 2007; Hsiou &
Fortier, 2007; Riff & Aguilera, 2008). En este trabajo se utilizara la topología
encontrada con las medidas de soporte para la discusión, las topologías encontradas con
Pesos Iguales, Pesos Implicados, y Consenso Estricto se obtuvieron y sólo se presentan
en el presente trabajo a título informativo. El cladograma con sus correspondientes
valores de soporte (una medida de confiabilidad de los resultados, valores mayores al
50%, mejor soporte) y respectivos nodos (terminales y puntos de ramificación) se
encuentra expresado en la (Fig. 37); los valores de soporte que se tendrán en cuenta para
la discusión en la siguiente sección del trabajo, serán los obtenidos bajo Symmetric
Resampling. Las sinapomorfias de todos los nodos y grupos se encuentran listadas en el
Apéndice VI.
IV.D.1. Posición filogenética del caimán de Rosario de la Frontera
El clado Alligatoridae es un grupo monofilético que incluye a Alligator,
Paleosuchus, Caiman y Melanosuchus, y varios otros taxa más extintos (Eocaiman,
Necrosuchus, Mourasuchus y Purussaurus), con alto valor de soporte (89%) y
soportado por 20 sinapomorfias.
Los Caimaninae están soportados por 8 sinapomorfias, los géneros Paleosuchus,
Mourasuchus, Purussaurus, Caiman y Melanosuchus constituyen este gran clado. Las
relaciones de los basales Eocaiman y Necrosuchus no son del todo claras, a pesar de las
afinidades del primero con los yacarés. En esté análisis, Alligator, Eocaiman y
Necrosuchus forman una tricotomía no resuelta.
Las especies del género Paleosuchus (P. trigonatus y P. palpebrosus)
conforman un grupo monofiletico con gran soporte (90%), soportado por 7
sinapomorfias, son basales a todos los Caimaninae. Los extraños Mourasuchus y
Purussaurus están más relacionados entre si que con cualquier otro caimán, forman un

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 63
grupo monofiletico soportado por 4 sinapomorfias. Los Nettosuchidae, representados
por el género Mourasuchus, están diagnosticados con 11 sinapomorfias.
El fósil MAS-PALEO-RdlF 001/2011 está relacionado a los Jacarea (sensu
Brochu, 1999a) en general, y a las especies del género Caiman en particular,
confirmando su asignación genérica. Más afín a los actuales Caiman latirostris y
Melanosuchus niger que a cualquier otro caimán, a pesar de la politomía que forma con
C. lutescens, C. latirostris y Melanosuchus.
Por otro lado, se confirma la parafilia de Caiman, las dos especies del género
Melanosuchus son taxa hermanos que forman parte del crown-group “Caiman” (sensu
Brochu, 1999a). Hay débil soporte entre los taxa y perdida de resolución dentro del
género Caiman, con la excepción de los clados C. crocodilus + C. yacare y
Melanosuchus niger + M. fisheri, que en todos los casos constituyen grupos
monofileticos que son consistentes en todos los análisis.
Tanto Caiman latirostris como MAS-PALEO-RdlF 001/2011, no presentan
autoapomorfias para su diagnosis. Las sinapomorfias que los caracterizan son ambiguas
o simplesimorficas, la ocurrencia en dos o más taxa de un grupo monofilético de un
carácter plesimórfico o estado de carácter (Kitching et al., 1998). Sólo el ancho del
hocico es un carácter de valor específico, el carácter cresta rostral prominente no es
exclusivo sino compartido con Caiman lutescens y Melanosuchus, y muchas otras
características son de un nivel jerárquico más inclusivo. La asignación específica es
soportada por la descripción comparativa realizada precedentemente entre las especies
del género.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 64
V. DISCUSIÓN
V.A. Consideraciones Taxonómicas:
El análisis descriptivo-comparativo y filogenético del ejemplar fósil, permiten
referir los restos como correspondiente a la actual especie de aligatórido Caiman
latirostris. La característica distintiva de la especie es el hocico ancho y amplio, con una
cresta preorbital prominente, característica considerada por varios autores (Burmeister,
1880; Mook, 1921c; Schmidt, 1928; Patterson, 1936; Cei, 1993; Verdade & Piña, 2006
y 2007). Además, la descripción de C. latirostris dada por Mook (1921c), corresponde
con la del espécimen presentado.
Necrosuchus + Eocaiman + Caimaninae
Simpson (1937) describió a Necrosuchus ionensis, del Paleoceno Temprano de
la Formación Salamanca de Patagonia (Figs. 8 y 61), como perteneciente a los
Crocodylidae; la descripción se basó en un fragmento anterior del dentario derecho. Sin
embargo, una reexaminación en trabajos posteriores (Brochu, 1997b, 2010, en prensa)
determina una estrecha relación con los Caimaninae en una posición basal, confirmada
en este trabajo.
Necrosuchus no presenta autoapomorfias que lo diagnostiquen, el carácter
alvéolos mandibulares 13º y 14º mayores posteriores al 4º, es una plesiomorfia
compartida con Eocamian y el aligatorino Alligator, el esplenial terminando antes de la
sínfisis mandibular, con una punta anterior dorsal al canal meckeliano (Carácter 6,
estado 1), es una sinapomorfia de Alligatoridae (Brochu, 1999a), considerada
erróneamente como una sínfisis esplenial por Simpson (Brochu, en prensa). Según
Brochu (1999a), una sínfisis esplenial esta presente ancestralmente en Alligatoroidea, la
exclusión del esplenial de la sínfisis mandibular con un extremo dorsal al canal de
Meckel, es característica de miembros derivados de Alligatorinae y Caimaninae.
Otro de los Caimaninae basal, junto con Necrosuchus, es Eocaiman, cuyas
relaciones tampoco son del todo claras. Originalmente fue descripto por Simpson
(1933) como Eocaiman cavernensis para el Eoceno Temprano de Patagonia. El género

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 65
tiene registro desde el Paleoceno patagónico (Gasparini, 1981; Bona, 2007), hasta el
Mioceno de Colombia (Langston, 1965) (Figs. 8 y 61).
Muchas de las sinapomorfias listadas en este análisis, y que caracterizan a
Eocaiman (cresta preorbital moderada y unida medialmente, superficie prefrontal dorsal
con excrecencia, borde maxilar amplio en la fenestra palatal), son compartidas con los
caimanes más modernos. Sin embargo, una mandíbula baja en su porción anterior por
debajo del nivel del 12º alvéolo, sínfisis extendiéndose por detrás del 6º u 8º alvéolo,
cara anterior de los palatinos de contorno rectangular, son características que permiten
diferenciar el género (Simpson, 1933; Bona, 2007).
Simpson (1933) da una diagnosis del género, que es la misma para la especie
cavernensis, algunas de la cuales ya fueron nombradas en el párrafo anterior y otras son
compartidas con el género Caiman, con el cual esta muy relacionado. Además, entre los
caracteres propuestos menciona los pterigoides formando gran parte del borde posterior
de las fenestras palatales, y las narinas internas ubicadas más cerca del borde anterior de
los pterigoides que del posterior; carácter que no fue considerado por Brochu (1999a) en
su análisis. Aparentemente, está condición diagnostica la región palatal de Eocaiman
cavernensis, y difiere de la presente en caimaninos actuales (Bona, 2007), en donde los
pterigoides participan en una pequeña proporción en la formación del borde posterior de
las fenestras palatales, y las coanas se encuentran más cerca del borde posterior de los
pterigoides que del borde anterior (Fig. 38A). Lamentablemente, el material de Rosario
de la Frontera (MAS-PALEO-RdlF 001/2011) no ha preservado los pterigoides, y por lo
tanto no se puede hacer observaciones respecto a su participación en las fenestras
palatales ni mucho menos las relaciones de las coanas dentro de ellos.
En todos los análisis realizados, siempre Eocaiman esta más emparentado a los
derivados Caimaninae actuales que Necrosuchus, esto es válido por los caracteres
derivados que comparten (cresta preorbital, borde maxilar en la fenestra palatal, etc.).
En el análisis de Brochu (2010), Necrosuchus esta más relacionado a los caimanes que
Eocaiman. El material tipo de Eocaiman y Necrosuchus es muy fragmentario, y las
partes relevantes son desconocidas. Por lo tanto, un gran porcentaje de caracteres no
fueron registrados en estos taxones, resultando en una perdida de información y
resolución sobre sus relaciones con los demás caimaninos, como también lo sugiere

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 66
Brochu (1999a). El descubrimiento de ejemplares más completos pertenecientes a estos
taxa, ayudará a resolver su posición dentro del esquema de relaciones de Alligatoroidea.
Caimaninae
No caben dudas sobre la monofilia de Paleosuchus, y las diferencias que los
separan de los demás Caimaninae. Las sinapomorfias recuperadas en este trabajo para
estos peculiares caimanes enanos de frente lisa, esplenial con doble perforación
posterior, premaxilar con 4 alvéolos en toda la ontogenia, coronoides perforado,
palpebrales completamente osificados, y proyección de los palatinos en las fenestras
palatales, también son consideradas por Brochu (1999a). Este autor también incluye una
serie de características diagnósticas para el género, lacrimal en amplio contacto con el
nasal y maxilar, sin proceso maxilar posterior, parietal no excluido del borde posterior
de la tabla craneal, las cuales no fueron recuperadas en este análisis. Los caracteres
dientes posteriores comprimidos lateralmente y contorno anterior hendido en los
palatinos, fueron recuperados como sinapomorfias del género en el presente trabajo. Sin
embargo, la morfología del extremo anterior de los palatinos (Carácter 48, estado 1) es
una característica compartida con Mourasuchus, al igual que el contacto lacrimal-nasal-
maxilar (una reversión dentro de Alligatoroidea, según Brochu, 1999a); pasa lo mismo
en Purussaurus con la participación del parietal en el borde posterior de la tabla craneal
(Carácter 87, estado 1).
A pesar que es un carácter recurrente para diferenciar Paleosuchus de los
restantes caimanes (Mook, 1921c; Schmidt, 1928; Kälin, 1955; Medem, 1958;
Iordansky, 1973; Cei, 1993), las fenestras supratemporales cerradas a la madurez
(Carácter 82, estado 2), no soporta la monofilia del género; la apertura de estas fenestras
es un carácter que varía con la edad del individuo (Mook, 1921d; Kälin, 1955;
Iordansky, 1973). Medem (1958), en su descripción de las especies del género, comenta
que en juveniles de P. palpebrosus las fenestras no son evidentes, y que en juveniles y
algunos adultos de P. trigonatus, estas cavidades son diferenciadas. Mook, en su trabajo
de 1921d, comenta que en un individuo senil de Caiman crocodilus, las fenestras
supratemporales están prácticamente cerradas.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 67
Muchos de los caracteres de Paleosuchus son modificaciones de las
sinapomorfias de los caimaninos. El coronoides no solo rodea al foramen
intermandibular medio, sino que llega a obliterarlo, localizándolo en una posición
central, sinapomorfias del género sugerida por Brochu (1999a); esta condición también
se observó en una de las ramas mandibulares de Caiman latirostris (FMNH 9713) (Fig.
42F), y en C. yacare (MACN 8265).
A pesar de estar ya diferenciado temprano en el Terciario, como lo sugieren las
filogenias, hasta el momento no hay registro fósil de las especies del género
Paleosuchus (Brochu, 2003). Un pequeño fragmento de dentario derecho, proveniente
del Mioceno Medio del Amazonas Peruano (Figs. 8 y 61), fue asignado tentativamente a
Paleosuchus por la morfología de los dientes (Salas-Gismondi et al., 2007).
Mourasuchus + Purussaurus + Jacarea
En éste análisis, los caracteres que soportan el clado Mourasuchus +
Purussaurus + Jacarea (sensu Brochu, 1999a), están relacionados con la presencia de
una cresta preorbital, y modificaciones de ellas.
Mourasuchus + Purussaurus
El clado Mourasuchus + Purussaurus, soportado por un proceso premaxilar
dorsal largo, prefrontales unidos medialmente y más largos que el lacrimal, y por una
corta punta frontal anterior, es muy consistente al igual que el clado Paleosuchus en
todos los análisis realizados. Sin embargo, no es un grupo con alto soporte. Del
conjunto de sinapomorfias listadas para este clado, solo la extensión anterior del frontal
no sobrepasando las órbitas, es exclusiva del grupo (Langston, 1966; Bocquentin-
Villanueva, 1984; Bocquentin & Souza Filho, 1990; Aguilera et al., 2006); carácter no
considerado por Brochu (1999a) en su análisis de los Alligatoroidea.
El proceso premaxilar dorsal largo es una característica compartida con los
crocodilimorfos longirostrinos (Mook, 1921c; Kälin, 1955; Gasparini, 1968; Sill, 1970;
Iordansky, 1973; Kraus, 1998; Brochu, 1997a, 2003, 2004a; Sereno et al., 2001; Riff &
Aguilera, 2008). La unión de los prefrontales, evitando el contacto nasal-frontal,
también está presente, como una autoapomorfia según este análisis, en el yacaré negro

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 68
Caiman yacare (Medem, 1955, 1960; Brochu, 1999a; Aguilera et al., 2006). El tamaño
del prefrontal mayor al lacrimal, aparece en Alligator y algunos aligatoroideos basales
(Brochu, 1999a, 2004b; Aguilera et al., 2006), como así también en Caiman lutescens
(Langston, 1965). En la mayoría de los crocodilianos, el lacrimal es mayor al prefrontal;
sin embargo, Brochu (1999a) señaló que las crías de Alligator y Caiman tienen un
prefrontal mayor, pero avanzada la ontogenia ocurre una reversión del carácter en los
caimanes, teniendo adultos con un lacrimal mayor.
De las dos sinapomorfias propuestas por Brochu (1999a) para el peculiar caimán
hocico de pato Mourasuchus, sólo el carácter nasales excluidos de la narina externa y en
contacto con los premaxilares, es una característica exclusiva del grupo (Langston,
1966; Bocquentin Villanueva, 1984), como lo demuestra este análisis. En un cráneo de
Caiman yacare se observó el mismo estado (Fig. 49C), Mook (1921c) y Medem (1955),
describen la misma característica para individuos seniles de Caiman crocodilus.
Otras sinapomorfias recuperadas en el análisis, son la presencia de unos
escamosales muy dilatados formando unos pseudo-cuernos (Bocquentin & Souza Filho,
1990; Brochu, en prensa), lo más parecido a ellos se puede observar en la Fig. 57C que
corresponde al crocodílido Crocodylus niloticus; también una sínfisis mandibular muy
estrecha es característica de Mourasuchus (Langston, 1965, 1966; Bocquentin & Souza
Filho, 1990).
Brochu (1999a) da una serie de caracteres ambiguos (homodoncia, ausencia de
festonamiento, reborde preorbital elevado) para Mourasuchus, que aquí son recuperadas
como sinapomorfias; sin embargo, son compartidas con Gavialis gangeticus de hocico
largo y delgado. En el caso de la sutura surangular-angular contactando en la esquina
posterodorsal de la fenestra mandibular externa, es una característica con Alligator
mississippiensis, y la forma anterior de los palatinos con Paleosuchus como se dijo
anteriormente. Una narina externa extremadamente ancha, es una característica
distintiva de los Nettosuchidae (Langston, 1965, 1966; Brochu, 1999a) que no fue
recuperada como sinapomorfia por el análisis filogenético. El género Mourasuchus
tiene registro para el Mio-Plioceno, principalmente en el norte de Sudamérica
(Langston, 1966); para Argentina se citó a M. nativus en el Paraná (Bocquentin &

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 69
Souza Filho, 1990). Ninguna de las especies del género fue incluida en un análisis
filogenético, Brochu (1999a) sólo considera Mourasuchus spp (Figs. 8 y 61).
Muchos de los caracteres que diferencian al gigante Purussaurus del Mio-
Plioceno sudamericano (Figs. 8 y 61) (proceso retroarticular orientado posteriormente y
no postero-dorsalmente, surangular sin extensión hasta el extremo posterior del proceso
retroarticular, fenestra mandibular externa grande, parietal alcanzado el borde posterior
de la tabla craneal, presencia de una fosa alrededor de la fenestra supratemporal), son
reversiones dentro de Crocodylia y Caimaninae en particular (Aguilera et al., 2006).
A pesar que Brochu (1999a) no presentó una lista de sinapomorfias para
Purussaurus. Aquí se presentan como características distintivas del género, una fenestra
mandibular externa grande (carácter compartido con Alligator mississippiensis), un
canto rostral formando una fosa nasal, y una tabla craneal con borde posterior
profundamente curvado, características también observadas por Aguilera et al. (2006);
sin embargo, estas dos últimas características nunca fueron incluidas en una filogenia, y
son presentadas como nuevos caracteres en el presente trabajo.
En las capas del Paraná (Argentina), hay registro de restos de caimanes de gran
talla asignados a Caiman (Rovereto, 1912; Patterson, 1936; Gasparini, 1981), que
quizás serían afines al gran Purussaurus, pero hasta el momento nunca fueron
reexaminados e incluidos en una filogenia. Esta afinidad ya fue notada por algunos
autores de principio del siglo XX como Patterson (1936), quien reconoció la posibilidad
de que Caiman paranensis (Alligator? ameghinoi de Rovereto, 1912) sea sinónimo de
Purussaurus brasiliensis, y que la diferencia de tamaño se deba a variación
ontogenética.
Jacarea
El clado Jacarea (Nodo 28) es un grupo monofilético robusto, constituido por las
especies del parafiletico género Caiman, y las de Melanosuchus. Las sinapomorfias
exclusivas que soportan el grupo son: la presencia de una lámina articular ventral al
foramen lingual dentro de la fosa del aductor, y una amplia y larga plataforma maxilar
lateral a la fenestra palatal; sinapomorfias también indicadas por Brochu (1999a). El
festonamiento marcado del hocico, según este análisis es una sinapomorfia de Jacarea,

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 70
pero es un carácter muy ambiguo dentro de Crocodylia y denota un morfotipo
generalizado (Brochu, 2001a). Una cresta ósea que une las órbitas (carácter presentado
en este trabajo) no es recuperada como apomorfia del grupo, sin embargo fue
considerada por Schmidt (1928) y Mook (1921c) (bajo el género Jacare), característica
compartida por todas las especies de Caiman, en el cual se incluía a Melanosuchus
niger (Caiman o Jacare niger respectivamente); Brochu (1999a) no considero este
carácter en su análisis.
Caiman crocodilus + C. yacare
Dentro del Nodo 29 (Caiman crocodilus + C. yacare), la única especie que no
posee autoapomorfias es C. crocodilus. Ambas especies comparten una prolongación
lingual del surangular alcanzando el último alvéolo mandibular, y dos profundas fosetas
de oclusión medial entre el 7º y 8º alvéolo maxilar (Fig. 50C), características también
comentadas por Brochu (1999a). En ninguno de los cráneos de Caiman yacare revisado
se pudo constatar la presencia de la prolongación del surangular que caracteriza al
clado.
Aunque no fue considerado en el análisis filogenético, pero si en la comparación
especifica, el extremo anterior del hocico es dilatado en los adultos de las dos especies
en relación a otros caimanes, como lo expresó Brochu (1999a); esto se pudo constatar
en uno de los cráneos de Caiman yacare (MLP s/n) (Fig. 49D). La participación o
exclusión de los nasales de la narina externa (Fig. 49C y D), no es un carácter de
variación específica sino que varía con la edad, esto fue sugerido por Mook (1921d)
para C. crocodilus, y por Rovereto (1912) para C. latirostris.
El agrupamiento de Caiman crocodilus y C. yacare, ambos de hocico angosto,
también esta evidenciado por estudios de patrones de crecimiento ontogenético y
morfométrico del hocico, que los separan de la especie hermana de hocico ancho C.
latirostris (Monteiro et al., 1997; Monteiro & Soares, 1997; Monteiro & Lessa, 2000).
Esta separación también está fundada por una mayor preferencia del yacaré ñato en una
dieta constituida por alimentos más duros (Monteiro & Lessa, 2000); de la observación
personal se pudo ver que Caiman yacare presenta dientes más delgados y agudos que C.
latirostris (Figs. 41A y C).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 71
Según Brochu (1999a), y los resultados del análisis filogenético de este trabajo,
la especie Caiman yacare se caracteriza por unos prefrontales unidos en la línea media,
separando los nasales del frontal (Carácter 60, estado 1); este estado también es
observado en el clado Mourasuchus + Purussaurus, como se dijo anteriormente. A
pesar de que es una característica diagnostica del yacaré negro (Medem, 1960; Brochu,
1999a), se observó en un cráneo de LCD = 209, 74 mm (BDIV-UNSa) un contacto
nasal-frontal (Fig. 52D), y en otro de LCD = 191,00 mm (25 EC-IML) una separación
entre esos elementos (Fig. 52C). Según Medem (1960), adultos de razas
Centroamericanas de Caiman crocodilus, raramente presentan unos prefrontales unidos
en la línea media como en C. yacare, mientras que no es lo común en poblaciones de
crocodilus del Amazonas; sin embargo constituyen poblaciones distantes
geográficamente del rango del yacaré negro. Estudios detallados de este carácter en
Caiman yacare, determinaran si se trata de un carácter ontogenético, polimórfico o de
variación geográfica.
Otros caracteres que diagnostican Caiman yacare, según el trabajo aquí
presentado, son la particular forma de la sutura palatino-pterigoides, y la reducida o nula
participación del pterigoides en el borde posterior de las fenestras palatales (Fig. 53C).
Ninguno de estos caracteres fue antes tenido en cuenta en un análisis filogenético. Sin
embargo, la exclusión de los pterigoides como característica diagnostica de Caiman
yacare fue citada por Schmidt (1928), Medem (1960) y Iordansky (1973).
La especie Caiman australis del Mio-Plioceno del Paraná, aunque es una forma
de mayor tamaño, está relacionada a C. yacare o quizás C. crocodilus como lo
sugirieron Rovereto (1912) y Gasparini (1981). En cuanto a la especie enana Caiman
tremembensis del Oligoceno de Brasil (el registro más antiguo del género), tiene
afinidades con C. yacare o C. crocodilus por el estrecho arco mandibular, pero por la
morfología de la sínfisis recuerda más a C. latirostris, según Chiappe (1988). Sin
embargo, Caiman australis y C. tremembensis nunca fueron incluidos en un análisis
filogenético, y restos más completos ayudaran a entender sus afinidades.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 72
Caiman latirostris + C. lutescens + Melanosuchus
Una prominente cresta preorbital, que extiende desde las órbitas hasta el nivel
del 4º alvéolo maxilar, es la única sinapomorfia de Caiman latirostris + C. lutescens +
Melanosuchus, según resultados obtenidos en la filogenia de Brochu (1999a) y en el
presente trabajo. Una apomorfia ambigua, y plesiomórfica (a nivel de Jacarea) que
caracteriza el clado, es la sutura frontoparietal lineal entre las fenestras supratemporales
(Brochu, 1999a), que difieren de las especies hermanas Caiman crocodilus + C. yacare
de sutura curvada; una sutura frontoparietal lineal también esta presente en
Mourasuchus y Paleosuchus. En cráneos revisados de Caiman latirostris, se pudo
observar tanto una sutura lineal como curvada, por lo tanto el carácter debería ser
reconsiderado y ver que tan variable es en las especies.
El nuevo estado (2) del carácter 37, hocico muy curvado, es la única apomorfia
exclusiva de Caiman lutescens según este análisis. Otra sinapomorfia listada para esta
especie son los prefrontales mayores a los lacrimales, que impiden el contacto nasal-
lacrimal como ya se comento previamente. A pesar de estar emparentados, las
relaciones con C. latirostris y Melanosuchus no están resueltas y no tienen soporte.
La reconstrucción de Caiman lutescens utilizada en este trabajo, es la misma que
presentó y describió Langston en 1965 como C. cf. C. lutescens. Ésta fue realizada por
la combinación de fragmentos encontrados en localidades distantes temporal y
geográficamente, por una tabla craneal (Fig. 59C) del Plioceno del Paraná descripta por
Rovereto (1912) como Alligator lutescens (el fragmento mandibular presentado aquí
también corresponde a esta especie), y un hocico parcial (Fig. 59B) del Mioceno Medio
de Colombia presentado por Langston (1965). El análisis filogenético de Brochu
(1999a) se basó en el fragmento asignado como C. cf. C. lutescens, pero que él
consideró Caiman lutescens. Patterson (1936), creyó que los fragmentos descriptos por
Rovereto, tanto los asignados a A. lutescens como a A. australis y A.? ameghinoi,
pertenecían a un juvenil de Caiman paranensis, un gigante de más de 6 m de longitud
aproximadamente; C. paranensis es considerado un nomen vanum por Langston (1965).
Langston no solo reconoció la posibilidad de que el material de La Venta
(Colombia) no sea la misma especie descripta por Rovereto (1912), sino que también

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 73
los fragmentos craneales que comprenden el material tipo de Alligator lutescens, no
estuvieran asociados. La tabla craneal esquematizada por Rovereto, con borde posterior
profundamente cóncavo, recuerda mucho a la de Purussaurus o Mourasuchus; mientras
que el hocico con cresta rostral prominente (Fig. 59A), como en Melanosuchus y
Caiman latirostris, es similar a la forma de La Venta. Sin embargo, las proporciones de
la narina externa del ejemplar de Colombia, es mayor al de la forma descripta por
Rovereto. De esta manera, y al igual como lo considero Langston (1965) y Brochu
(1999a), es posible que el fósil de La Venta y el del Paraná sean especies totalmente
diferentes que deberían ser reinterpretados.
Gasparini (1981) refirió Caiman lutescens (Alligator lutescens + C. cf. C.
lutescens) como perteneciente al actual C. latirostris. A juzgar por la tabla craneal
descripta por Rovereto de borde posterior cóncavo profundo, es muy distinta de la del
yacaré ñato en la cual es curvada pero no profunda (Figs. 56E). En cuanto a la
configuración del fragmento rostral de Alligator lutescens, éste no difiere de la de
Caiman latirostris y MAS-PALEO-RdlF 001/2011, y al igual que este último, es de
destacar el mayor tamaño que cualquiera de los cráneo revisados de la actual especie.
La diferencia más notable, es el gran tamaño de la narina externa con respecto a la
observada en Caiman latirostris. Por otro lado, el fragmento anterior de dentario
derecho descripto por Rovereto como perteneciente a Alligator lutescens, es de
dimensiones mayores que cualquier especie actual de Caiman, y de arco mandibular
angosto como C. yacare y C. crocodilus; Langston (1965) también reconoció
similitudes en el contorno del hocico entre C. cf. C. lutescens y C. yacare o C.
crocodilus. La sinonimia entre C. latirostris y C. cf. C. lutescens considerada por
Gasparini, no es aceptada por las razones ya expuestas, como también lo consideró
Brochu (1999a); pero la asignación del fragmento rostral de A. lutescens a C. latirostris
es probable, de ser así seria el registro más antiguo de la especie en el “Conglomerado
Osilífero” de la Formación Ituzaingó (Mioceno Tardío) (Figs. 8 y 61) (Cione et al.,
2000).
La marcada curvatura del hocico de Caiman lutescens (C. cf. C. lutescens),
quizás sea una consecuencia de la dilatación del extremo anterior descripta para C.
yacare o C. crocodilus, observación también considerada por Brochu (1999a). De esta
forma, a pesar de la estrecha relación con C. latirostris y Melanosuchus, C. lutescens es

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 74
un taxón formado por un mosaico de fragmentos de entidades diferentes, que debería ser
reconsiderado. Restos más completos determinaran la validez y relación de caimán
fósil.
El yacaré ñato Caiman latirostris no tiene apomorfias que lo caractericen, según
los resultados de éste análisis y el de Brochu (1999a). Morfológicamente, como se
comprobó en la comparación genérica, es muy distinto del emparentado Melanosuchus
de hocico más largo que ancho, y con exposición palatal del vómer sobre la sutura
premaxilar-maxilar (Mook, 1921c; Schmidt, 1928; Cei, 1993; Brochu, 1999a). Las dos
especies de Melanosuchus conocidas hasta el momento, M. fisheri (extinta) y M. niger
(actual), son unas de las pocas entre los crocodilianos en las cuales el vómer es visible
externamente (Iordansky, 1973; Medina, 1976; Brochu, 1999a). Este carácter (Carácter
40, estado 1) quizás sea de variación específica más que genérica como lo sugirió Mook
(1921c). Por lo expresado, Poe (1997) propuso sinonimizar Melanosuchus con Caiman.
En la región donde en Melanosuchus se exhibe el vómer, en MAS-PALEO-RdlF
001/2011 hay un espacio romboidal que probablemente represente un daño tafonómico
o postmortem. La configuración de la sutura premaxilar-maxilar del fósil de Rosario de
la Frontera es similar a la del ejemplar de Caiman latirostris descripto por Patterson
(1936), aguda anteriormente y no es recta, en donde no se observa un vómer; de esta
manera ambos fósiles estarían relacionados. Además, la forma de los procesos
posteriores de los palatinos y su contacto con los pterigoides, es muy diferente entre M.
niger y C. latirostris.
Los individuos adultos de Melanosuchus, género muy afín a Caiman latirostris,
alcanzan una Longitud Craneal Dorsal (LCD) de 50, 8 cm (Medina, 1976), con un
máximo de longitud total que excede poco los 4 m (Rovereto, 1912). Según Burmeister
(1880) y Verdade & Piña (2006 y 2007) a partir de datos históricos, el yacaré ñato es un
crocodiliano de tamaño medio que alcanzaba una longitud máxima de 3,5 m, pero en la
actualidad animales mayores a los 2 m son raros en la naturaleza. El cráneo más grande
de C. latirostris que se midió personalmente fue de LCD=28,1 cm, mientras que el fósil
del río Rosario-Horcones presenta un LCD=36,3 cm (Apéndice V). De esta manera, y a
juzgar por la talla que alcanzan los individuos de Melanosuchus, el gran tamaño del
fósil de Rosario de la Frontera estaría dentro de los límites históricos de variación

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 75
individual y etaria aceptados para Caiman latirostris, alcanzando los 3 m e incluso un
poco más. Burmeister (1880) da una edad adulta senil para individuos de 3 m o más.
Según Grigg & Gans (1993) en los cocodrilos de edad senil, el reemplazo dentario
disminuye hasta detenerse, llegando a encontrarse individuos desdentados o anodontes.
En el fósil descripto, se pudo observar dientes de reemplazos o no funcionales, que poco
asoman por los alvéolos; de este modo no sería un individuo geronte. Considerando que
el yacaré ñato presenta un dimorfismo sexual, como todos los crocodilianos, en el cual
los machos son generalmente de tamaño corporal y craneal mayor que el de las hembras
(Grigg & Gans, 1993; Verdade, 2000, 2003; Verdade & Piña, 2007), se concluye que
MAS-PALEO-RdlF 0001/2011 pertenece a un individuo adulto macho de la especie
Caiman latirostris.
La edad adulta para el fósil descripto también se confirma por la marcada
esculturación craneal, prominente cresta preorbital, órbitas relativamente pequeñas,
marcado festonamiento rostral y mandibular, hocico más largo en relación a la región
postorbital, distancia mayor entre fenestras supratemporal y menor tamaño de ella por
ensanchamiento de los elementos de la tabla craneal, agudas crestas sobre la superficie
antero-ventral del cuadrado, extremo anterior del hocico más redondeado que anguloso
(Fig. 49B), y el menor tamaño relativo de la narina externa (Mook, 1921d; Kälin, 1955;
Iordansky, 1973). Esta última característica es evidente por el ancho premaxilar anterior
a la narina (Fig. 49B), similar a lo observado en las figuras brindadas por Patterson
(1936) en su fósil de Caiman latirostris, y el fragmento rostral de Caiman lutescens
descripto por Rovereto (1912). La extensión de la sínfisis mandibular en casi todos los
cráneos de C. latirostris revisados (Longitud Craneal desde 9,6 a 28, 5 cm) no mostró
variación con la edad como lo sugirieron Mook (1921d) y Chiappe (1988), siempre se
extendía hasta el 4º o 5º alvéolo, con una excepción en un cráneo de 28,6 cm (FMNH
9713) en el cual la sínfisis alcanzaba el 6º alvéolo, en el fósil de Rosario de la Frontera
llega hasta el 5º.
A juzgar por la figura presentada por Patterson (1936) sobre el fragmento rostral
(Figs. 60A y B) perteneciente a Caiman latirostris del Pleistoceno del Este de Salta, la
forma de la narina externa es muy distinta con respecto a la de los cráneos revisados de
la misma especie. En estos cráneos, y en el fósil descripto, la narina externa es circular,
tan ancha como larga, no aplastada y más ancha como la del fósil de Patterson

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 76
recuperado de los Estratos del Chaco superior; según Pascual & Odreman Rivas (1973)
esas capas serían más antiguas y tendrían una edad pliocena. El tamaño de los alvéolos
maxilares en el fósil de Quebrada Aguas Blancas también es diferente, aparentemente el
mayor es el 3º alvéolo, mientras que en las formas actuales y MAS-PALEO-RdlF
001/2011 de la misma especie, lo es el 4º. El fósil descripto por Patterson debería ser
revisado.
En cuanto al fósil presentado por Starck & Anzótegui (2001), que corresponde a
un fragmento mandibular (Figs. C y D) proveniente de la Formación Palo Pintado
(Mioceno Tardío-Plioceno) del Valle Calchaquí, asignado provisoriamente como
Caiman sp., es de aspecto similar al observado en Caiman latirostris. De ser así,
correspondería a uno de los registros más antiguos de la especie, quizás el único, si es
que el fragmento rostral de Caiman lutescens proveniente del Paraná mantiene su status
taxonómico.
En base a lo discutido, la especie Caiman latirostris, al igual que C. yacare,
tiene registro desde el Mioceno Tardío-Plioceno, habiéndose encontrado en capas del
Paraná (Entre Ríos) (Rovereto, 1912; Patterson, 1936; Báez & Gasparini, 1979;
Gasparini, 1981; Cione, 2000), Quebrada Aguas Blancas de Salta (Patterson, 1936), y
quizás Valle Calchaquí (Starck & Anzótegui, 2001) (Figs. 8 y 61). Restos fragmentarios
aislados asignados a la misma especie, provenientes del Mio-Plioceno en Brasil, y
quizás Venezuela (Hsiou & Fortier, 2007), requieren confirmación. Sin embargo, los
restos exhumados en Rosario de la Frontera constituyen el material más completo para
C. latirostris, encontrado hasta ahora en el Cenozoico de Argentina y de Sudamérica en
general.
El registro fósil de Caimaninae del Cenozoico de Sudamérica es muy diverso,
fragmentario, escaso y disperso (Figs. 8 y 61) (Gervais, 1876; Mook, 1921a; Simpson,
1933 y 1937; Patterson, 1936; Langston, 1965 y 1966; Medina, 1976; Gasparini, 1981;
Bocquentin Villanueva, 1984; Chiappe, 1988; Bocquentin & Souza Filho, 1990;
Brochu, 1999a, 2003; Piña & Argañaraz, 2000; Verdade & Piña, 2006; Aguilera et al.,
2006; Hsiou & Fortier, 2007; Bona, 2007). Diverso en cuanto son varios los taxones
descriptos, principalmente extintos, lo que demuestra que la distribución y diversidad
actual, no es más que un relicto de la diversificación sufrida por el grupo en el Terciario,

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 77
como lo sugieren Simpson (1933 y 1937), Sill (1968), y Brochu (1999a y 2003).
Fragmentario, por que los restos recuperados son piezas craneales y mandibulares
parciales, y en contadas excepciones, casi completos. Escaso y disperso, ya que son
pocas las localidades fosilíferas que brindaron restos de caimanes fósiles, y muy
distantes geográficamente. Para muchos de estos restos, debería revisarse su posición
taxonómica.
V.B. Aspectos Paleoclimáticos-ambientales y Paleozoogeográficos:
Los cocodrilos junto con las aves son los únicos miembros sobrevivientes del
gran grupo de los arcosaurios, que fueron abundantes y diversos por todo el mundo
durante el Mesozoico. Los Eusuchia, diferenciados en el Cretácico Tardío, tuvieron su
principal momento de expansión en el Terciario; esta expansión ocurrió durante la
primera parte de este periodo cuando el clima era más cálido, reduciéndose con el
enfriamiento en la parte terminal. La actual distribución de los Crocodylia es un relicto
de temprana radiación del Terciario (Sill, 1968; Brochu, 2003). En la actualidad
comprenden 8 géneros distribuidos por latitudes bajas desde los 30º hacia el Ecuador
(Markwick, 1998).
El actual límite de distribución de los cocodrilos esta condicionado
principalmente por la temperatura y el clima (Markwick, 1998). El amplio alcance
latitudinal del grupo a principios del Terciario fue debido a un clima más cálido hacia
los polos (Sill, 1968; Taplin & Grigg, 1989). Los crocodilianos son sensibles a la
fluctuación climática y su distribución está relacionada con estos cambios que
ocurrieron durante el Terciario, cambios que también afectaron las comunidades de
mamíferos y la paleoflora (Sill, 1968; Pascual & Ortiz Jaureguizar, 1990). Por lo tanto,
los cocodrilos son excelentes indicadores de condiciones paleoclimáticas (Sill, 1968;
Gasparini, 1981; Taplin & Grigg, 1989; Markwick, 1998).
Markwick (1998) hace mención del “uniformitarismo taxonómico” a la
interpretación del paleoclima terrestre usando los datos fósiles, en analogía de la
tolerancia climática de las especies vivientes relacionadas, y corroborada por otros datos
geológicos. A partir de la paleodistribución de los fósiles climático-sensibles, es posible
inferir la distribución espacial del paleoclima (Gasparini, 1981; Gasparini et al., 1986;
Pascual, 1986; Albino, 1994; Cione et al., 1995). El uniformitarismo taxonómico es

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 78
válido para los cocodrilos, teniendo en cuenta que son de morfología relativamente
conservativa (Markwick, 1998; Gans & Grigg, 1993). Es de destacar que estos reptiles
no son “termómetros geológicos”, pero pueden aportar interesantes datos al
conocimiento paleoclimático y paleoambiental de un área (Gasparini, 1981).
Los aligatóridos, al carecer de glándula de excreción de sal, están restringidos a
los ambientes acuáticos (Taplin & Grigg, 1989; Gans & Grigg, 1993). Estar sumergidos
y contar con cuerpos de agua permanente, los protegen de las temperaturas extremas
(Gans & Grigg, 1993). Por lo tanto, la ausencia o presencia de estos cocodrilos puede
ser reflejo de las condiciones hidrológicas locales, más que de la temperatura. Según
Piña & Argañaraz (2000), como así también lo sugirió Gasparini (1981), la presencia de
un aligatórido sugiere un clima subtropical a templado cálido y húmedo, y lugares
vegetados donde el agua no escasee. La topografía también afecta la hidrología y la
temperatura, y así influye en la distribución de los crocodilianos (Markwick, 1998).
Los aligatóridos, originarios del Hemisferio Norte, alcanzaron Sudamérica
temprano en el Terciario ocupando la mayoría de los nichos (Taplin & Grigg, 1989;
Brochu, 1999a y 2003; Bona, 2007). Los aligátores y los caimanes divergieron
aparentemente en el Terciario Temprano, como lo confirman datos de secuencia de α-
globina de Alligator y Caiman (Densmore & Owen, 1989; Brochu, 1997a). Desde
entonces, los caimanes experimentaron una gran radiación adaptativa y dominaron la
fauna crocodiliana del Sudamérica (Sill, 1968; Gasparini et al., 1986; Taplin & Grigg,
1989).
El linaje de los caimanes invadió Sudamérica antes del Eoceno, cuando el
relativamente “moderno” Eocaiman y el basal Necrosuchus aparecieron en Patagonia
(Simpson, 1933 y 1937; Langston, 1965; Gasparini, 1981; Gasparini et al., 1986; Bona,
2007; Brochu, 2010, en prensa). Restos de vertebrados (peces, cocodrilos Alligatoridae,
serpientes Boidae, tortugas acuáticas Pelomedusidae) registrados en el Paleoceno
patagónico, como así también registro florístico, no solo confirman el predominio de
zonas boscosas sino más específicamente, indican al menos clima subtropical con una
exuberante vegetación y abundante agua (Pascual & Odreman Rivas, 1971; Pascual &
Ortiz Jaureguizar, 1990; Pascual, 2006). La mayoría de este material es muy
fragmentario, pero esos restos son muy interesantes desde un punto de vista

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 79
paleoclimático, pues ellos indican condiciones húmedas subtropicales, en latitudes
alrededor de los 45ºS (Báez & Gasparini, 1979; Gasparini, 1981; Gasparini et al., 1986).
Cocodrilos de edad Eocena han sido exhumados en el noroeste argentino (Salta y
Jujuy), noroeste de Mendoza y este del Chubut; en las tres áreas aparecen mesosuquios
sebécidos, sumándose en la última, los eusuquios aligatóridos (Simpson, 1933 y 1937;
Gasparini, 1981; Bona, 2007). Restos de dipnoi Lepidosirenidae presentados por
Fernández et al. (1973), de la Formación Lumbreras en la puna jujeña, indican un
ambiente tropical con marcada estacionalidad, condiciones que se extendían por el norte
argentino durante el Eoceno. El Paleoceno Tardío-Eoceno temprano del noroeste
argentino, también brindó restos de tortugas Pelomedusidae y Teiidae (Gasparini et al.,
1986), lo cual da cuenta del característico clima del Paleógeno Temprano en la porción
austral del continente, muy diferente al actual; inferencia confirmada también por
mamíferos fósiles (Pascual & Odreman Rivas, 1971; Pascual & Ortiz Jaureguizar, 1990;
Pascual, 2006).
En Patagonia, la ausencia de algunos reptiles sensibles al clima (tales como los
boideos y cocodrilos) después del Eoceno Temprano, es testimonio dramático del
cambio climático en ese tiempo (Gasparini, 1981; Gasparini et al., 1986; Pascual, 1986;
Albino, 1994). El único registro que confirma la presencia de cocodrilos en el
Oligoceno Temprano de la Argentina, son de Sebecidae de la provincia de Chubut
(Gasparini, 1981). Entre el Eoceno Medio y el Oligoceno Temprano hay un descenso
del nivel del mar y de la temperatura; el descenso total de la temperatura en ese tiempo
es el más precipitado en todo el Cenozoico, quizás relacionado a un englazamiento de la
Antártida (Pascual & Odreman Rivas, 1971 y 1973; Pascual & Ortiz Jaureguizar, 1990;
Pascual, 2006). Según Báez & Scillato Yané (1979), durante el Oligoceno Temprano la
distribución de los cocodrilos del extremo austral sudamericano sufrió una retracción,
posiblemente como consecuencia del deterioro climático existente en el Oligoceno
Tardío. Luego de este mínimo de temperatura, existió un moderado ascenso hacia fines
del Oligoceno y Mioceno Temprano, que no fue lo suficiente como para permitir el
repoblamiento de Patagonia por parte de los Crocodylia, como lo afirma Chiappe
(1988). A partir del Mioceno Medio, las condiciones climático-ambientales cambian
drásticamente en Patagonia, coincidente con la fase diastrófica Quechua responsable de
la elevación de la cordillera andinopatagónica, y el consecuente inicio de la

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 80
desertización de esa región, y de otros eventos geológicos y biogeográficos vinculados
(Báez & Scillato-Yané, 1979; De la Fuente, 1992). Todos estos cambios determinaron
la retracción hacia el norte de muchos elementos de la biota.
El registro de cocodrilos terciarios en Argentina se interrumpe hasta el Mioceno
Tardío-Plioceno (Rovereto, 1912; Patterson, 1936; Langston, 1965; Sill, 1970;
Gasparini, 1968 y 1981; Piña & Argañaraz, 2000; Cione et al., 2000). De esta
antigüedad existe en Paraná (Entre Ríos) un rico yacimiento con vertebrados, en el
mismo se ha podido identificar la presencia de Caiman australis, C. yacare, C.
latirostris, probablemente C. lutescens, Gryposuchus (Gavialidae), y Mourasuchus
(Nettosuchidae). Según Pascual & Odreman Rivas (1971), Gasparini (1981), y Cione et
al. (2000), el material de Paraná esta re-depositado, y probablemente no todas las
especies citadas hayan convivido, pero dan una idea de la composición faunística en esa
región para el Mio-Plioceno.
Ciertas evidencias demuestran que en el Mioceno Tardío-Plioceno Temprano
pudo haber comenzado el proceso de diferenciación de las actuales áreas biogeográficas
(Pascual & Odreman Rivas, 1973). Tal inferencia es confirmada por el patrón
paleobiogeográfico de Caimaninae en Sudamérica, que se repite en las tortugas
dulceacuícolas Chelidae (De la Fuente, 1992), peces cíclidos, serrálmidos y clupéidos
(Cione et al., 1995), serpientes Colubridae y Boidae, y otros herpetozoos, resultando en
la diversidad y abundancia actual (Albino & Quintana, 1992; Cei, 1993; Albino, 1994).
Uno de los ejemplos más paradigmáticos está representado por la Formación Ituzaingó
(Mioceno Tardío-Plioceno) de la Mesopotamia argentina, cuya paleofauna indica
claramente una caracterización paleoclimática y paleozoogeográfica similar a la que
presenta en la actualidad (Cione et al., 2000). El “Mar Paranaense” constituyó una
conspicua característica geográfica en tiempos del Mioceno Tardío, y quizás modelador
de los patrones biogeográficos actuales. Esta transgresión Atlántica, se extendió sobre
gran parte de la región Chaco-Pampeana y el oeste de la Mesopotamia, desde el noreste
de Patagonia al sur del Paraguay. En Patagonia, con la excepción de la región más
septentrional, el clima fue frio y semiárido a árido durante la mayor parte del Plioceno,
acentuándose en el Cuaternario (Báez & Scillato Yané, 1979).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 81
Los cambios en el Cenozoico Tardío son la culminación de procesos que
iniciaron a fines del Mioceno; es sabido que el Neógeno Tardío fue un tiempo de
diastrofismo muy activo conocido como fase Diaguita. La fase Diaguita de la orogenia
Andina produjo el levantamiento de las Sierras Pampeanas del este, haciendo sombra de
lluvia las regiones occidentales con una creciente aridización de ellas (Pascual &
Odreman Rivas, 1973; Pascual & Ortiz Jaureguizar, 1990; Pascual &; Pascual, 2006).
El análisis de Starck & Anzótegui (2001) confirma un relevante cambio
climático ocurrido aproximadamente a fines del Mioceno en el Valle Calchaquí, desde
ambientes bajo condiciones de extrema aridez con la ausencia casi total de fósiles, a
ambientes de condiciones húmedas con cuerpos de agua vegetados con una rica
asociación de fósiles (vertebrados incluyendo restos de caimán, vegetales y
palinomorfos). Los ambientes áridos fueron reemplazados en el Mioceno Tardío y
Plioceno por ambientes más húmedos en esta parte del noroeste argentino. Cerca de la
localidad de Cafayate fueron exhumados de niveles inferiores de la Formación Anta
(Mioceno Medio), restos de peces dulceacuícolas que vivieron bajo condiciones
ambientales benignas (Cione et al., 1995). Los peces de la Quebrada La Yesera indican
que los cuerpos de agua que habitaban se encontraban a baja altura sobre le nivel del
mar, muy por debajo de la altitud en que se encuentra la localidad en la actualidad (más
de 1500 msnm). Antes del ascenso de las Sierras Pampeanas y de la Cordillera, cuerpos
lénticos y lóticos de baja altura se extendían en el oeste con una biota diferente a la
actual, que probablemente desembocaban en el gran engolfamiento paranaense (Cione
et al., 1995).
Las regiones a lo largo del lado este de los Andes, por entonces más bajas y más
cálidas que en la actualidad, constituyo un importante sendero para la migración en
tiempos del Plioceno (Zurita, 2007). Hay evidencia paleontológica de un avance mucho
más hacia el oeste de faunas similares a las del Río Paraná, lo que habría coincidido con
un avance de una fauna originalmente de un ambiente más húmedo que dio origen a la
fauna herpetológica chaqueña adaptada a condiciones climáticas más rigurosas. Hay una
tendencia general de cambio climático desde el Plioceno hacia un régimen más frio y
seco (Báez & Scillato Yané, 1979). Eventos tectónicos del Plio-Pleistoceno acentuaron
la diferenciación de la región Subandina con considerables consecuencias climáticas y
fisiográficas (Zurita, 2007).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 82
Los cambios composicionales registrados en las sucesivas comunidades de
vertebrados cenozoicos, reflejan variaciones ambientales regionales y climáticas, y así
patrones de cambios climáticos (Pascual & Odreman Rivas, 1971; Pascual & Ortiz
Jaureguizar, 1990; Pascual, 2006).
El hábitat natural de los yacarés son los ríos, los pantanos y los esteros de las
selvas subtropicales, donde predomina el clima cálido y húmedo sin grandes
oscilaciones térmicas, y abundantes lluvias; también habitan las zonas de transición
entre la selva subtropical y el bosque templado, con áreas más abiertas y arboledas más
dispersas (Burkart et al., 1999; Rueda Almonacid et al., 2007).
Las dos especies actuales de caimanes (de hábitos anfibios) que habitan el país,
viven asociadas a cuerpos permanentes de agua donde pasan la mayor parte del tiempo
(Larriera & Imhof, 2006). Al contrario del yacaré negro, Caiman latirostris prefiere
ambientes con vegetación exuberante y sistemas lénticos, tales como planicies de
inundación, pantanales y manglares, y presenta una de las más amplias distribuciones en
términos latitudinales entre los crocodilianos, ocurriendo desde los 5ºS a los 34ºS,
Caiman yacare actualmente se encuentra a latitudes menores de 29ºS (Piña et al., 2004;
Verdade & Piña, 2006 y 2007). Se ha referido que la mayor incursión latitudinal por
parte de C. latirostris, en contraste a su congénere C. yacare, se debe a una mayor
tolerancia a las bajas temperaturas, como lo afirma Piña & Argañaraz (2000).
Ambas especies comparten gran parte de su distribución, pero el yacaré overo
llega más al sur que el yacaré negro, como se comentó en el párrafo anterior. El rango
de Caiman yacare se superpone al de C. latirostris en el Paraguay inferior. Cuando
estas especies están en simpatría, se reparten el ambiente de tal forma que el yacaré
negro es más visible, ya que ocupa los ambientes donde el agua corre, es menos
vegetada y accesible, mientras que C. latirostris se encuentra en ambientes de muy
difícil acceso y muy vegetados (Medem, 1960; Piña et al., 2004; Larriera & Imhof,
2006; Verdade & Piña, 2006 y 2007; Rueda Almonacid et al., 2007; Campos et al.,
2010; Verdade et al., 2010). La divergencia de hábitats de las dos especies se da en
otros aspectos ecológicos, como ser los recursos tróficos, el yacaré overo por la solidez
de sus mandíbulas hace pensar en una adaptación a la durofagia (Iordansky, 1973), con
componente dietario de crustáceos, peces, moluscos y tortugas acuáticas (Cei, 1993),

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 83
mientras que Caiman yacare se alimenta esencialmente de peces y crustáceos (Medem,
1960; Cei, 1993; Rueda Almonacid et al., 2007). Las poblaciones más abundantes de
Caiman latirostris en Argentina, se encuentran en los grandes esteros asociados a las
planicies de inundación de los ríos de llanura, como el río Paraná, el Uruguay y el
Salado (Larriera & Imhof, 2006).
Las evidencias paleofaunísticas sugieren que las condiciones paleoambientales
del noroeste de la Argentina durante el Terciario Tardío (últimos 5 Ma), fueron más
benignas que las actualmente registradas en el área considerada en este estudio. El área
del hallazgo está situada principalmente en el área del Chaco Semiárido del noroeste
argentino (Burkhart et al., 1999), fuera del rango de distribución geográfica y altitudinal
de la especie Caiman latirostris. El registro aquí presentado incrementa el rango de
distribución horizontal de la especie, hasta Rosario de la Frontera (Salta) en tiempos del
Plioceno. De este modo de infiere que el rango de dispersión geográfica de C. latirostris
a fines del Neógeno, se extendía mucho más hacia el Oeste del territorio argentino, en
contraste al patrón zoogeográfico que se observa en la actualidad.
La posición del rio Rosario-Horcones en la actualidad, a unos 700 m por encima
del nivel del mar bajo condiciones semiáridas a templadas, obviamente es el efecto del
diastrofismo cenozoico Andino. Sin embargo, la ocurrencia altitudinal más elevada
registrada por yacaré overo fue de 600 m en Brasil y 800 m en Argentina (Verdade &
Piña, 2006 y 2007).
La litología de la Formación Piquete y la información paleoclimática-ambiental
que brinda el fósil descripto (uniformitarismo taxonómico), indican que la sedimentitas
que lo contenían se depositaron en una amplia llanura de inundación (González Villa,
2002; Salfity & Monaldi, 2006), bajo condiciones ambientales similares a las
predominante hoy en el área donde Caiman latirostris aun vive, limitado a los
humedales del noreste argentino, ambientes de clima cálido y húmedo con cuerpos de
agua permanentes, asociados a flora y fauna acuática diversa; por lo menos en le sector
de Rosario de la Frontera. Lógico es inferir que tales fueron las condiciones existentes
en el área del hallazgo en tiempos del Plioceno durante la depositación de la Formación
Piquete, actualmente a una altura de 700 msnm con condiciones climático-ambientales
incompatibles con las exigencias ecológicas de los caimanes.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 84
Asi como Chiappe (1988) sugirió que el pequeño tamaño de Caiman
tremembensis (del Oligoceno de Brasil) se debió a las condiciones climáticas poco
favorables en las que vivió, y teniendo en cuenta que los cocodrilos actuales de gran
talla están relacionados a ambientes de clima cálido (Grigg & Gans, 1993) (ej.
Melanosuchus del Amazonas, hasta más de 4 m). La inferencia de un ambiente de clima
cálido para el Plioceno en el área del hallazgo (de actual clima templado), es válida por
el gran tamaño que alcanzó el fósil descripto de Caiman latirostris (más de 3, 5 m), con
respecto al tamaño que alcanzan los individuos actuales de la misma especie (hasta 2,0
m).
La actual especie sudamericana Caiman latirostris tiene registro, con seguridad,
desde el Plioceno (Estratos del Chaco Superior de Quebrada Aguas Blancas y
Formación Piquete de Rosario de la Frontera), o quizás el Mioceno Tardío si se
confirma que pertenece a esta especie el fragmento mandibular del Valle Calchaquí
presentado por Starck & Anzótegui (2001), o si pasa lo mismo con el fragmento rostral
de C. lutescens del Paraná; habitando los humedales que se extendían desde la
Mesopotamia hacia el oeste. Sin embargo, es posible afirmar que la escasez en el
registro fósil de Crocodylia por estas latitudes para fines del Neógeno, se debiera a la
falta de prospecciones. Los cambios climáticos y ambientales del Cuaternario, asociados
a eventos tectónicos, determinaron la retracción hacia el noreste de la biota relacionada
a esos ambientes acuáticos.
En este sentido, éste hallazgo provee una valiosa información sobre la evolución
de los ambientes naturales de esta parte de Argentina, en relación con los modificadores
procesos del diastrofismo cenozoico. Esta distribución geográfica concuerda con los
datos brindados por los diferentes autores citados anteriormente sobre las
deteriorizaciones climáticas ocurridas durante el Terciario, y en cierto modo, confirma
el hecho de considerar los crocodilianos como “termómetros geológicos”.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 85
VI. CONCLUSIONES
En un intento de resumir los resultados obtenidos y discutidos en el presente
trabajo, podemos concluir:
Desde una perspectiva taxonómica y morfológica
A partir del análisis descriptivo, comparativo y filogenético, se pudo determinar
que el material fósil del río Rosario-Horcones (Rosario de la Frontera),
pertenece a la actual especie de Caimaninae Caiman latirostris, constituyendo el
registro más completo y mejor preservado para el Plioceno de Salta (Argentina)
y de Sudamérica.
El material descripto pertenece a un individuo adulto de gran talla alcanzando
quizás los 4 m, tamaño similar alcanzado por individuos adultos de
Melanosuchus con el cual esta muy relacionado; individuos actuales de la misma
especie no superan los 2 m.
Los cocodrilos presentan dimorfismo sexual, los machos son de mayor tamaño
que las hembras, esto hace presumir que el ejemplar estudiado se trataba de un
individuo macho adulto.
La edad adulta del fósil también es soportada por la marcada esculturación
craneal y ensanchamiento de los huesos superficiales.
Desde una perspectiva filogenética
En este trabajo se presenta por primera vez una comparación y revisión genérica
y específica de caimanes fósiles y actuales sudamericanos.
A pesar de tener una posición basal dentro del esquema de relaciones de
Caimaninae, del conjunto de sinapomorfias, Eocaiman esta más relacionado a
los actuales caimanes derivados que Necrosuchus.
La gran participación del pterigoides en el borde posterior de la fenestra palatal
y la posición anterior de la coana, son características exclusivas de Eocaiman
que deberían ser consideradas en un análisis filogenético.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 86
Material más completo de los Caimaninae basal, Eocaiman y Necrosuchus,
ayudarán a resolver sus relaciones, y a determinar sinapomorfias para el último
caso.
Muchos de los caracteres de Paleosuchus (fenestras supratemporales cerradas,
coronoides perforados, palpebrales completamente osificados) son
modificaciones extremas de las sinapomorfias de los caimanes más derivados.
Los resultados de este análisis soportan la monofilia de Purussaurus y
confirman la posición como grupo hermano de Mourasuchus (Nettosuchidae)
previamente considerada por otros autores.
A pesar de estar estrechamente relacionados a los caimanes actuales,
Purussaurus y Mourasuchus son morfotipos de gran tamaño muy especializados
restringidos al Mio-Plioceno.
El nuevo carácter sobre la corta extensión anterior del frontal, que nunca fue
considerado en una filogenia, es una sinapomorfia exclusiva del clado
Mourasuchus + Purussaurus.
Muchas de las sinapomorfias consideradas para Mourasuchus (homodoncia,
ausencia de festonamiento, reborde orbital anterior), son compartidas con
Gavialidae.
La gran mayoría de las características propuestas para diferenciar a Purussaurus
son reversiones dentro de Crocodylia, y Caimaninae en particular. Sin embargo,
los caracteres, narina externa contenida en una fosa y borde posterior de la tabla
craneal cóncavo profundo, son apomorfias exclusivas del género según este
análisis.
El clado Jacarea (Caiman + Melanosuchus) es un grupo débilmente soportado
que requiere revisión.
Caiman crocodilus no presenta apomorfias que lo diferencien de su relativo C.
yacare; el carácter (prefrontales unidos en la línea media) usado para diferenciar
a ambas especies, es un carácter ontogenético.
De los seis caracteres nuevos propuestos en este trabajo, dos de ellos (nula
participación del pterigoides en la fenestra palatal y procesos posteriores de los
palatinos muy dilatados relacionados con una sutura palatino-pterigoides muy
ondulada) fueron recuperados como diagnósticos de Caiman yacare entre las
tres únicas sinapomorfias recuperadas.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 87
Caiman latirostris, C. lutescens y Melanosuchus están muy emparentados, pero
sus relaciones no están resueltas, y sólo están soportados por el carácter cresta
rostral prominente.
La validez de la especie Caiman lutescens es dudosa, una redescripción y
revisión ayudará determinar su validez taxonómica, y a resolver la politomía de
los caimanes de cresta rostral prominente.
El carácter vómer expuesto en el paladar de las dos especies de Melanosuchus
sería de variación específica, y ambas deberían ser consideradas pertenecientes
al género Caiman, por la topología que ocupan dentro de Jacarea.
Una revisión y descripción detallada de la morfología craneal, y estudios sobre
variación ontogenética, individual y geográfica, permitirán determinar
sinapomorfias del yacaré overo o ñato, Caiman latirostris. Lo mismo es
aplicable para las ambiguas especies C. yacare y su relativo C. crocodilus.
El ancho y robusto hocico de gruesos dientes de Caiman latirostris es una
especialización a la durofagia; el grácil hocico de dientes muy agudos de C.
yacare esta relacionado con una dieta principalmente de peces.
Caiman lutescens (Alligator lutescens + C. cf. C. lutescens) es una especie
“mosaico” formada por fragmentos de individuos diferentes, de localidades
distantes y de distinta edad.
Caiman lutescens no es sinónimo de C. latirostris por la razones ya expuestas,
pero el fragmento rostral de A. lutescens es similar al de esta especie.
Entre los caimanes de cresta rostral prominente (Caiman latirostris + C.
lutescens + Melanosuchus), C. latirostris no presenta sinapomorfias que lo
soporten, sin embargo es muy diferente de sus relativos en tener un hocico tan
ancho como largo.
El material fragmentario de Caiman australis muestra similitudes con C. yacare
o C. crocodilus, mientras que el de C. tremembensis estaría más relacionado con
C. latirostris; sin embargo, C. australis y C. tremembensis nunca fueron
incluidos en una filogenia, restos más completos ayudaran a resolver sus
relaciones.
Gran parte del material proveniente de las capas del Paraná (Mio-Plioceno),
como asi también de otras localidades de Sudamérica, debería ser revisado e
incluido en un análisis filogenético.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 88
Desde un punto de vista paleoclimático-ambiental y paleobiogeográfico
La historia paleobiogeográfica de los caimanes es similar a la de varios otros
taxones (Squamata, Pelomedusidae, etc.) con requerimientos de ambientes de
clima cálido y húmedo.
Los cocodrilos, de hábitos anfibios y de ambientes de clima cálido y húmedo,
son buenos indicadores paleoclimático-ambientales, como fuera sugerido por
varios autores.
El registro fósil muestra que la distribución geográfica de los caimanes en
Argentina a principios del Neógeno, era mucho más extensa y alcanzaba los
46ºS de latitud, donde las condiciones climáticas eran más benignas. Sufren una
retracción hacia el norte a partir del Eoceno, resultado del englazamiento de la
Antártida y la orogenia andina. A fines del Neógeno, y durante el Cuaternario,
sufren otra retracción pero hacia el noreste, también como consecuencia de la
orogenia andina y los cambios climático-ambientales asociados, quedando
restringidos a su actual distribución en los humedales del noreste, y con límite
latitudinal de 34ºS.
El fósil motivo de este trabajo fue exhumado de sedimentitas de la Formación
Piquete (Plioceno), relacionada con llanuras de inundación, que están expuestas
sobre el río Rosario-Horcones de Rosario de la Frontera.
La localidad de procedencia del material fósil está fuera del rango de
distribución geográfica de la especie Caiman latirostris, que esta restringida a
humedales bien vegetados con una rica fauna acuática asociada del noreste de la
provincia de Salta, y de Argentina.
El ambiente de depositación de la Formación Piquete sería equivalente a los
humedales del noreste argentino de clima cálido y húmedo donde habita Caiman
latirostris y su congénere C. yacare, inferencia establecida a partir de los
requerimientos ecológicos de éstas especies actuales (uniformitarismo
taxonómico), y corroborada por datos geológicos de la citada formación; por lo
menos en el sector de Rosario de la Frontera. Sin embargo, la escasez de restos
fósiles de la Formación Piquete, quizás es debida a la falta de prospección.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 89
A fines del Neógeno, el rango de distribución geográfica del yacaré ñato se
extendía hacia el sudoeste de la provincia, incursionando lo que hoy comprende
el Valle Calchaquí.
La presencia del yacaré ñato en Rosario de la Frontera en tiempos del Plioceno,
constituye el segundo registro, el más occidental de la especie, y representa el
primer Caimaninae descripto para el Neógeno Tardío del Sur de Salta
La especie Caiman latirostris tiene registro desde el Plioceno (5,3-1,8 Ma.).
Restos de esta especie se exhumaron de los Estratos del Chaco Superior de
Quebrada Aguas Blancas y de la Formación Piquete de los márgenes del río
Rosario-Horcones (Rosario de Frontera) de la provincia de Salta.
Otros restos provenientes del Mioceno Tardío de las capas del Paraná o de la
Formación Palo Pintado del Valle Calchaquí, como asi también de otras
localidades de Sudamérica, fueron asignados como C. cf. C. latirostris y
requieren confirmación.
Salta es la tercera localidad argentina, después de Paraná y Chubut, en brindar
caimanes fósiles, y hasta el momento es la única que brindo restos del actual
Caiman latirostris.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 90
VII. AGRADECIMIENTOS
Quiero dejar constancia de mi agradecimiento a las siguientes personas e instituciones que permitieron llevar a cabo este trabajo:
A Fernando Lobo, por haberme contactado y ayudado en la búsqueda de tema de tesis, y brindarme lugar de trabajo en el Laboratorio de Anatomía Comparada. Por su constante orientación y estímulo en la realización de este trabajo, y las valiosas críticas y sugerencias efectuadas a un primer manuscrito. A pesar de que no es su campo de estudio, sin él este trabajo paleontológico no se hubiera realizado. Gracias Lobo!!!
A Jaime Powell (Jimmy), por la gentileza de ofrecer el material motivo de este trabajo; el ideólogo del tema de tesis. Por su buena predisposición, a pesar de tener muchas cosas encimas, cuando estuve por Tucumán y en los constantes mails que eran nuestro medio de discusión para ir puliendo el trabajo. Por su asesoramiento brindado en los aspecto geológicos y geográficos del lugar del hallazgo. A pesar de todo, siempre sabio en sus correcciones y opiniones sobre el manuscrito. Gracias Jimmy!!!
A Federico Arias por las fructíferas, constantes y largas charlas, por su permanente apoyo y asesoramiento que brindó a mi trabajo, y por su lectura crítica a una parte del manuscrito. Aprendí mucho trabajando con vos. Gracias Fede!!!
A Sebastián Quinteros por su ayuda y sugerencias efectuadas en el análisis filogenético, parte fundamental del trabajo; y siempre dándome una mano a pesar de mis molestias. Gracias Sebas!!! Y Soledad Valdecantos por sus valiosos consejos y estar ante cualquier necesidad, al igual que la profe Virginia. Gracias Sole!!! Gracias Profe!!!
A Diego Pol, del MEF (Trelew), quien gentilmente me facilitara material fotográfico y bibliografía, imprescindibles para el desarrollo de este estudio. A Santiago Nenda, del MACN, quien también brindó material fotográfico.
A Christopher Brochu, de la University of Iowa, por su excelente predisposición, quien desinteresadamente facilito algunos trabajos de su autoría y de gran consulta.
A Sonia Kretzschmar, del Instituto de Herpetología de la Fundación Miguel Lillo, por el préstamo de material y bibliografía de dicha institución.
A la gente del Laboratorio de Paleontología de Vertebrados, Judith, Virginia, Pablo y Daniel, que siempre me recibieron hospitalariamente durante mis visitas. Al Técnico del Laboratorio, Juan Ballesteros, por asesorarme y brindarme ayuda en la preparación del fósil.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 91
A Ernestina Teisaire (cátedra de Embriología y Anatomía Comparadas), Ricardo Montero (cátedra de Vertebrados) de la Facultad de Ciencias Naturales e Instituto Miguel Lillo de Tucumán, y Juan Manuel (cátedra de Biodiversidad IV), Josefina Aris (cátedra de Paleontología General) de la Universidad Nacional de Salta, por facilitarme material de dichas cátedras, imprescindibles en este trabajo.
Al Tribunal Evaluador, Josefina Aris, Juan Manuel Díaz Gómez y Rosa Vera, por sus útiles comentarios y correcciones que mejoraron el manuscrito.
A Jorge Barrientos, quien dedico parte de su tiempo para facilitarme papers difíciles de conseguir, y que fueron de gran ayuda. Gracias Chart!!! A María López por sus charlas y aliento que me ayudaron anímicamente. Gracias Mary!!!
A mi familia, y en principal a mi madre, por su esfuerzo y contención, por bancarme en todo, sin lo cual nada de esto hubiese sido posible. Gracias Ma!!!
Y a mis amigos/as y todos aquellos que ayudaron, directa o indirectamente, a la realización de este trabajo. Gracias totales!!!

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 92
VIII. BIBLIOGRAFÍA CITADA
AGUILERA, O.A.; RIFF, O. & J. BOCQUENTIN VILLANUEVA. 2006. A new giant
Purussaurus (Crocodyliformes, Alligatoridae) from the Upper Miocene Urumaco
Formation, Venezuela. Journal of Systematic Palaeontology 4 (3): 221-232.
ALBINO, A.M. 1994. Estado actual del registro de escamados extinguidos de América
del Sur y sus implicancias paleoambientales. Cuadernos de Herpetología 8 (1): 1446-
154.
ALBINO, A.M. & C.A. QUINTANA. 1992. Los Colubridae (Reptilia: Serpentes) del
Chapadmalalense y Uquiense (Plioceno tardío-Pleistoceno temprano?) de la provincia
de Buenos Aires, Argentina. Ameghiniana 29 (2): 125-133.
AMBROSETTI, J. 1890. Observaciones sobre los reptiles fósiles oligocenos de los
terrenos terciarios antiguos del Paraná. Boletín de la Academia Nacional de Ciencias de
Córdoba 10(4): 409-426.
BÁEZ, A.M. & GASPARINI, Z.B. 1979. The South American Herpetofauna: An
evaluation of the fossil record. En: W.E. DUELLMAN (Ed.). The South American
Herpetofauna: its Origin, Evolution, and Dispersal. Museum of Natural History,
University of Kansas, Monograph 7: 29-54.
BÁEZ, A.M. & G.J. SCILLATO YANÉ. 1979. Late Cenozoic environmental changes
in temperate Argentina. En: W.E. DUELLMAN (Ed.). The South American
Herpetofauna: its Origin, Evolution, and Dispersal. Museum of Natural History,
University of Kansas, Monograph 7: 141-156.
BENTON, M.J. 2005. Vertebrate Palaeontology. Third Edition. Blackwell Publishing
455p.
BOCQUENTIN VILLANUEVA, J. 1984. Un nuevo Nettosuchidae (Crocodilia,
Eusuchia) proveniente de la Formación Urumaco (Mioceno superior), Venezuela.
Ameghiniana 21 (1): 3-8.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 93
BOCQUENTIN, J.C. & J.P. SOUZA FILHO. 1990. O crocodiliano sul-americano
Carandaisuchus como sinonímia de Mourasuchus (Nettosuchidae). Revista Brasileira
de Geociências 20 (1-4): 230-233.
BONA, P. 2007. Una nueva especie de Eocaiman Simpson (Crocodylia, Alligatoridae)
del Paleoceno inferior de Patagonia. Ameghiniana 44 (2): 435-445.
BONAPARTE, J.F. 1971. Los tetrápodos del sector superior de la Formación Los
Colorados, La Rioja, Argentina. (Triásico Superior). Opera Lilloana 22: 184p.
BRAVARD, A. 1858. Monografía de los terrenos marinos terciarios de las cercanías del
Paraná. Diario Oficial de Gobierno “El Nacional Argentino” (no visto).
BROCHU, C.A. 1997a. Morphology, fossils, divergence timing, and the phylogenetic
relationships of Gavialis. Systematic Biology 46 (3): 479-522.
BROCHU, C.A. 1997b. A review of “Leidyosuchus” (Crocodyliformes, Eusuchia) from
the Cretaceous through Eocene of North America. Journal of Vertebrate Paleontology
17 (4): 679-697.
BROCHU, C.A. 1999a. Phylogenetics, taxonomy, and historical biogeography of
Alligatoroidea. Society of Vertebrate Paleontology Memoir 6: 9-100.
BROCHU, C.A. 1999b. Taxon sampling and Reverse Successive Weighting. Systematic
Biology 48 (4): 808-813.
BROCHU, C.A. 2000. Phylogenetic relationships and divergence timing of Crocodylus
based on morphology and the fossil record. Copeia 2000 (3): 657-673.
BROCHU, C.A. 2001a. Crocodylian snouts in space and time: phylogenetic approaches
toward adaptative radiation. American Zoologist 41: 564-585.
BROCHU, C.A. 2001b. Progress and future directions in archosaur phylogenetics.
Journal of Palaeontology 75 (6): 1185-1201.
BROCHU, C.A. 2003. Phylogenetic approaches toward crocodylian history. Annual
Revue of Earth Planet Science 31: 357-397.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 94
BROCHU, C.A. 2004a. A new late Cretaceous gavialoid crocodylian from eastern
North America and the phylogenetic relationships of thoracosaurs. Journal of
Vertebrate Paleontology 24 (3): 610-633.
BROCHU, C.A. 2004b. Alligatorinae phylogeny and the status of Allognathosuchus
Mook, 1921. Journal of Vertebrate Paleontology 24 (4): 857-873.
BROCHU, C.A. 2006. A new miniature horned crocodile from the Quaternary of
Aldabra Atoll, Western Indian Ocean. Copeia 2006 (2): 149-158.
BROCHU, C.A. 2007. Morphology, relationships, and biogeographical significance of
an extinct horned crocodile (Crocodylia, Crocodylidae) from the Quaternary of
Madagascar. Zoological Journal of the Linnean Society 150: 835-863.
BROCHU, C.A. 2010. A new alligatorid from the lower Eocene Green River Formation
of Wyoming and the origin of caimans. Journal of Vertebrate Paleontology 30 (4):
1109-1126.
BROCHU, C.A. En prensa. Phylogenetic relationships of Necrosuchus ionensis
Simpson 1937 and the early history of caimanines. Zoological Journal of the Linnean
Society.
BROCHU, C.A. & A.D. RINCÓN. 2004. A gavialoid crocodylian from the Lower
Miocene of Venezuela. En: SÁNCHEZ VILLAGRA, M.R. & J.A. CLACK (Eds.).
Fossils of the Miocene Castillo Formation, Venezuela: Contributions on Neotropical
palaeontology. Special Papers in Palaeontology 71: 61-80.
BROCHU, C.A.; WAGNER, J.R.; JOUVE, S.; SUMRRAL, C.D. & L.D. DENSMORE.
2009. A correction corrected: Consensus over the meaning of Crocodylia and why its
matters. Systematic Biology 58 (5): 537-543.
BUFFETAUT, E. 1979. Evolución de los Crocodilios. Investigación y Ciencia 39: 88-
99.
BURKART, R.; BÁRBARO, N.; SÁNCHEZ, R.O. & D.A. GÓMEZ. 1999. Eco-
regiones de la Argentina. Administración de Parques Nacionales, 43p.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 95
BURMEISTER, D.G. 1880. Reseña de los crocodilinos de la República Argentina.
Anales Sociedad Científica Argentina 9: 241-251.
BURMEISTER, H. Examen crítico de los mamíferos y reptiles fósiles denominados por
don Augusto Bravard. Ibidem 3: 95-174 (no visto).
BUSACK, S.D. & S. PANDYA. 2001. Geographic variation in Caiman crocodilus and
Caiman yacare (Crocodylia: Alligatoridae): systematic and legal implications.
Herpetologica 57 (3): 294-312.
CAMPOS, Z.; LLOBET, A. PIÑA, C.I. & W.E. MAGNUSSON. 2010. Yacare Caiman
Caiman yacare. En: MANOLIS, S.C. & C. STEVENSON (Eds.). Crocodiles. Status
Survey and Conservation Action Plan. Third Edition. Crocodile Specialist group:
Darwin, 23-28.
CEI, J.M. 1993. Reptiles del noroeste, nordeste y este de la Argentina. Monografie XIV.
Museo Regionales di Historia Naturali di Torino.
CIONE, A.L.; VERGANI, G.; STARCK, D. & R. HERBST. 1995. Los peces del
Mioceno de la Quebrada de La Yesera, provincia de Salta, Argentina. Su valor como
indicadores ambientales y su antigüedad. Ameghiniana 32 82): 129-140.
CIONE, A.L.; AZPELICUETA, M.M; BOND, M.; CARLINI, A.A.; CASCIOTTA,
J.R.; COZZUOL, M.A.; DE LA FUENTE, M.; GASPARINI, Z.B.; GOIN, F.J.;
NORIEGA, J.I.; SCILLATO YANÉ, G.J.; SOIBELZON, L.; TONNI, E.P.; VERZI, D.
& M.G. VUCETICH. 2000. Miocene vertebrates from Entre Ríos province, eastern
Argentina. En: ACEÑOLAZA, F.G. & R. HERBST (Eds.). El Neógeno de Argentina.
Revista del Instituto Superior de Correlación Geológica. Serie de Correlación
Geológica 14: 191-237.
CHIAPPE, L.M. 1988. Un nuevo Caiman (Crocodylia, Alligatoridae) de la Formación
Tremembé (Oligoceno), Estado de São Paulo, Brasil, y su significado paleoclimático.
Paula-Coutiana 3: 49-66.
CREA, M.; MERLER, J. & R. QUINTANA. 1989. Contribución a la Sistemática de
Caiman latirostris (Daudin, 1802) (Crocodylia, Alligatoridae). Anales del Museo de
Historia Natural de Valparaíso 20: 75-80.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 96
DENSMORE, Ll.D. & R.D. OWEN. 1989. Molecular systematics of the Order
Crocodilia. American Zoologist 29: 831-841.
DE QUEIROZ, K. & J. GAUTHIER. 1990. Phylogeny as a central principle taxonomy:
Phylogenetic definitions of taxon names. Systematic Zoology 39 (4): 307-322.
FARRIS. J.S. 1983. The logical basis of phylogenetic analysis. Advances in Cladistics
2: 1-36.
FERNÁNDEZ, J.; BONDESIO, P. & R. PASCUAL. 1973. Restos de Lepidosiren
paradoxa (Osteichthyes, Dipnoi) de la Formación Lumbrera (Eogeno, ¿Eoceno?) de
Jujuy. Ameghiniana 10 (2): 152-172.
FUENTE, M. DE LA. 1992. Las tortugas Chelidae del Terciario superior y Cuaternario
del territorio argentino. Ameghiniana 29 (3): 211-229.
GASPARINI, Z.B. 1968. Nuevos restos de Rhamphostomopsis neogaeus (Burm.)
Rusconi 1933, (Reptilia, Crocodilia) del “Mesopotamiense” (Plioceno medio-superior)
de Argentina. Ameghiniana 5 (8): 299-311.
GASPARINI, Z.B. 1971. Los Notosuchia del Cretácico de América del Sur como nuevo
infraorden de los Mesosuchia (Crocodilia). Ameghiniana 8 (1): 83-103.
GASPARINI, Z.B. 1972. Los Sebecosuchia (Crocodilia) del territorio argentino.
Consideraciones sobre su “status” taxonómico. Ameghiniana 9 (1): 23-34.
GASPARINI, Z.B. 1981. Los Crocodilia fósiles de la Argentina. Ameghiniana 18 (3-
4): 177-205.
GASPARINI, Z.B. & E. BUFFETAUT. 1980. Dolichochampsa minima, n. g. n. sp., a
representative of a new family of eusuchian crocodiles from the late Cretaceous of
northern Argentina. Neues Jahrb. Geol. Paläontol. Mh. 5: 257-271.
GASPARINI, Z.B.; DE LA FUENTE, M. & O. DONADÍO. 1986. Los reptiles
cenozoicos de la Argentina: implicancias paleoambientales y evolución biogeográfica.
Actas IV Congreso Argentino Paleontología y Bioestratigrafía (Simposio “Evolución de
los Vertebrados Cenozoicos de América del Sur”) 2: 119-130.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 97
GATESY, J.; AMATO, G.; NORELL, M.; DESALLE, R. & C. HAYASHI. 2003.
Combined support for wholesale taxic atavism in gavialine crocodylians. Systematic
Biology 52 (3): 403-422.
GEBHARD, J.A.; GIUDICI, A.R. & J.O. GASCON. 1974. Geología de la comarca
entre el Río Juramento y Arroyo Las Tortugas, provincia de Salta y Jujuy, República
Argentina. Revista de la Asociación Geológica Argentina 24 (3): 359-375.
GERVAIS, M.P. 1876. Crocodile gigantesque fossile du Bresil. Journal de Zoologie 5:
232-236.
GOLOBOFF, P.A. 1993. Estimating character weights during tree search. Cladistics 9:
83-91.
GOLOBOFF, P.A.; FARRIS, J.S. & K.C. NIXON. 2003. T.N.T.: Tree Analysis Using
New Technology. Program and documentation, available from the authors, and at
www.zmuc.dk/public/phylogeny
GOLOBOFF, P.A.; CARPENTER, J.M.; SALVADOR ARIAS, J. & D.R. MIRANDA
ESQUIVEL. 2008a. Weighting against homoplasy improves phylogenetic analysis of
morphological data sets. Cladistics 24: 758-773.
GOLOBOFF, P.A.; FARRIS, J.S. & K.C. NIXON. 2008b. TNT, a free program for
phylogenetic analysis. Cladistics 24: 774-786.
GONZÁLEZ VILLA, R.E. 2002. El Subgrupo Jujuy (Neógeno) entre los 24º-26º LS y
64º-66º LO, tramo centro-austral de la cadena subandina argentina, provincias de Salta
y Jujuy. Facultad de Ciencias Naturales, Universidad Nacional de Salta, Tesis
Doctoral, 136p, inédita.
GRIGG, G. & C. GANS. 1993. Morphology & physiology of the Crocodylia. En:
GALSBY, CH.J.; ROSS, G.J.B. & P.L. BEESLEY (Eds.). Fauna of Australia 2A
Amphibia & Reptilia, Capítulo 40: 326-343.
HARSHMAN, J.; HUDDLESTON, C.J.; BOLLBACK, J.P.; PARSONS, T.J. & M.J.
BRAUN. 2003. True and false gharials: A nuclear gene phylogeny of Crocodylia.
Systematic Biology 52 (3): 386-402.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 98
HSIOU, A.S. & D.C. FORTIER. 2007. Primeiro registro de Caiman (Crocodylia,
Alligatoridae) para o Pleistoceno do Estado do Rio Grande do Sul, Brasil. Gaea 3 (1):
37-44.
IORDANSKY, N.N. 1973. The skull of the Crocodilia. En: GANS, C. & T. PARSONS
(Eds.). Biology of the Reptilia 4: 201-260.
JOUVE, S.; IAROCHÉNE, M.; BOUYA, B. & M. AMAGHZAZ. 2005. A new
dyrosaurid crocodyiform from the Palaeocene of Morocco and a phylogenetic analysis
of Dyrosauridae. Acta Palaeontologica Polonica 50 (3): 581-594.
KÄLIN, J.A. 1955. Crocodilia. En: PIVETEAU, J. (Ed.). Traité de Paléontologie 5:
695-784. Masson et Cie., Paris.
KITCHING, I.J.; FOREY, P.L.; HUMPHRIES, C.J. & D.M. WILLIAMS. 1998.
Cladistics. The Theory and Practice of Parsimony Analysis. Second Edition. Oxford
University Press.
KRAUS, R. 1998. The cranium of Piscogavialis jugaliperforatus n. gen., n. sp.
(Gavialidae, Crocodylia) from the Miocene of Peru. Paläontologische Zeitschrift 72 (3-
4): 389-406.
KRAUSE, D.W.; O’CONNOR, R.P.M.; ROGERS, K.C.; SAMPSON, S.D.;
BUCKLEY, G.A. & R.R. ROGERS. 2006. Late Cretaceous terrestrial vertebrates from
Madagascar: implications for Latin American biogeography. Annals of the Missouri
Botanical Garden 93: 178-208.
LANGSTON, W. 1965. Fossil crocodylians from Colombia and the Cenozoic History
of the Crocodilia in South America. University of California Publication of Geological
Sciences 52: 157p.
LANGSTON, W. 1966. Mourasuchus Price, Nettosuchus Langston, and the family
Nettosuchidae (Reptilia: Crocodilia). Copeia 1966 (4): 882-885.
LARRIERA, A. & A. IMHOF. 2006. Proyecto Yacaré. Cosecha de huevos para cría en
granjas del género Caiman en Argentina. En: BOLKOVIC, M.L. & D. RAMADORI

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 99
(Eds.). Manejo de Fauna Silvestre en la Argentina. Programa se uso sustentable.
Dirección de Fauna Silvestre, Secretaria de Ambiente y Desarrollo sustentable, 51-64.
LUTZONI, F.; WAGNER, P.; REEB, V. & S. ZOLLER. 2000. Integrating ambiguously
aligned regions of DNA sequences in phylogenetic analyses without violating positional
homology. Systematic Biology 49 (4): 628-651.
MARKWICK, P.J. 1998. Fossil crocodilians as indicators of late Cretaceous and
Cenozoic climates: implications for using palaeontological data in reconstructing
palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology 137: 205-271.
MARTIN, J.E. 2008. Un aperçu de l’évolution des Crocodiliens au Mésozoïque.
Bulletin de la Sociéte Géologique de Normandie et des Amis du Muséum du Havre 95
(5): 179-192.
MARTIN, S. 2008. Global diversity of crocodiles (Crocodilia, Reptilia) in freshwater.
Hydrobiologia 595: 587-591.
MARTIN, J.E. & M.J. BENTON. 2008. Crown clades in Vertebrate Nomenclature:
Correcting the Definition of Crocodylia. Systematic Biology 57 (1): 173-181.
MEAD, J.I.; CUBERO, R.; VALERIO ZAMORA, A.L.; SWIFT, S.L.; LAURITO, C.
& L.D. GÓMEZ. 2006. Plio-Pleistocene Crocodylus (Crocodylia) from southwestern
Costa Rica. Studies on Neotropical Fauna and Environment 41 (1): 1-7.
MEDEM, F.J. 1955. A new subspecies of Caiman sclerops from Colombia. Fieldiana:
Zoology 37: 339-344.
MEDEM, F.J. 1958. The crocodilian genus Paleosuchus. Fieldiana: Zoology 39 (21):
227-247.
MEDEM, F.J. 1960. Notes on the Paraguay caiman, Caiman yacare Daudin.
Mitteilungen aus dem Soologischen Museum in Berlin 36: 129-142.
MEDINA, C.J. 1976. Crocodilians from the late Tertiary of northwestern Venezuela:
Melanosuchus fisheri sp. nov. Breviora 438: 1-14.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 100
MOLNAR, R.E. 1993. Biogeography and phylogeny of the Crocodylia. En: GALSBY,
CH.J.; ROSS, G.J.B. & P.L. BEESLEY (Eds.). Fauna of Australia 2A. Amphibia &
Reptilia, Capítulo 42: 1-11.
MOLNAR, R.E. 2010. A new reconstruction of the skull of Sebecus icaeorhinus
(Crocodyliformes: Sebecosuchia) form the Eocene of Argentina. Brazilian
Geographical Journal: Geosciences and Humanities research médium 1 (2): 314-330.
MONTEIRO, L.R.; CAVALCANTI, M.J. & H.J.S. SOMMER. 1997a. Comparative
ontogenetic shape change in the skull of Caiman species (Crocodylia, Alligatoridae).
Journal fo Morphology 231: 53-62.
MONTEIRO, L.R. & M. SOARES. 1997. Allometric analysis of the ontogenetic
variation and evolution of the skull in Caiman Spix, 1825 (Crocodylia: Alligatoridae).
Herpetologica 53 (1): 62-69.
MONTEIRO, L.R. & L.G. LESSA. 2000. Comparative analysis of cranial suture
complexity in the genus Caiman (Crocodylia, Alligatoridae). Revista Brasileira de
Biologia 60 (4): 689-694.
MOOK, C.C. 1921a. Brachygnathosuchus braziliensis, a new fossil crocodilian from
Brazil. Bulletin of the American Museum of Natural History 44: 43-49.
MOOK, C.C. 1921b. The dermo-supraoccipital bone in the Crocodilia. Bulletin of the
American Museum of Natural History 44: 101-103.
MOOK, C.C. 1921c. Skull characters of recent Crocodilia, with notes on the affinities
of the recent genera. Bulletin of the American Museum of Natural History 44: 123-268.
MOOK, C.C. 1921d. Individual and age variations in the skull of recent Crocodilia.
Bulletin of the American Museum of Natural History 44: 51-66.
MOOK, C.C. 1923. Skull characters of Alligator sinensis Fauvel. Bulletin of the
American Museum of Natural History 48: 553-562.
MOOK, C.C. & G.E. MOOK. 1940. Some problems in crocodilian nomenclature.
American Museum Novitates 1098: 1-10.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 101
NESBITT, S.J. 2011. The early evolution of archosaurs: relationships and the origin of
major clades. Bulletin of the American Museum of Natural History 352: 292p.
OLIVEIRA, C.E.M. 2008. Associações ce ovos de crocodilomorfos do formação
Adamantina, grupo Bauru, cretáceo superior, na região de Jales-SP. Universidade
Estadual Paulista. Instituto de Geociências Exatas. Tesis Doctoral 99p.
PASCUAL, R. 1986. Evolución de los Vertebrados Cenozoicos: sumario de los
principales hitos. Actas IV Congreso Argentino Paleontología y Bioestratigrafía
(Simposio “Evolución de los Vertebrados Cenozoicos de América del Sur”) 2: 209-218.
PASCUAL, R. 2006. Evolution and geography: The biogeographic history of South
American land mammals. Annals of the Missouri Botanical Garden 93: 209-230.
PASCUAL, R. & O.E. ODREMAN RIVAS. 1971. Evolución de las comunidades de
los vertebrados del Terciario argentino. Los aspectos paleozoogeográficos y
paleoclimáticos relacionados. Ameghiniana 7 (3-4): 372-412.
PASCUAL, R. & O. ODREMAN RIVAS. 1973. Las unidades estratigráficas del
Terciario portadoras de mamíferos. Su distribución y sus relaciones con los
acontecimientos diastróficos. Actas V Congreso Geológico Argentino 3: 293-338.
PASCUAL, R. & E. ORTIZ JAUREGUIZAR. 1990. Evolving climates and mammal
fauna in Cenozoic South America. Journal of Human Evolution 19: 23-60.
PATTERSON, B. 1936. Caiman latirostris from the Pleistocene of Argentina, and a
summary of South American Cenozoic Crocodilia. Herpetologica 1 (29): 43-54.
PIÑA, C.I. & B. ARGAÑARAZ. 2000. Presencia del género Caiman (Crocodylia:
Alligatoridae) en la Formación Ituzaingó (Mioceno superior-Plioceno), Entre Ríos,
Argentina. En: ACEÑOLAZA, F.G. & R. HERBST (Eds.). El Neógeno de Argentina.
Revista del Instituto Superior de Correlación Geológica. Serie de Correlación
Geológica 14: 255-261.
PIÑA, C.I.; LARRIERA, A. & P. SIROSKI. 2004. Cocodrilos en la Región Litoral:
especies, distribución geográfica, modo de vida. En: ACEÑOLAZA, F.G.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 102
(Coordinador). Temas de la Biodiversidad del Litoral fluvial argentino. Instituto
Superior de Correlación Geológica, Miscelánea 12: 317-322.
POE, S. 1996. Data set incongruence and the phylogeny of crocodilians. Systematic
Biology 45 (4): 393-414.
POL, D. & Z.B. GASPARINI. 2007. Crocodyliformes. En: GASPARINI, Z.B.; CORIA,
R. & L. SALGADO (Eds.). Patagonian Mesozoic Reptiles. Indiana University Press.
Capítulo 5: 116-142.
QUINTEROS, A. S. 2011. Filogenia Morfológica y Molecular del grupo alticolor
(Iguania: Liolaemidae). Evolución de Caracteres del Dimorfismo Sexual. Facultad de
Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tesis
Doctoral, 193p., inédita.
RIFF, D. & O.A. AGUILERA. 2008. The world’s largest gharials Gryposuchus:
description of G. croizat n. sp. (Crocodylia, Gavialidae) from the upper Miocene
Urumaco Formation, Venezuela. Paläontologische Zeitschrift 82 (2): 178-195.
RODRÍGUEZ M., M.A. 2000. Cocodrilos (Archosauria: Crocodylia) de la región
Neotropical. Biota Colombiana 1 (2): 135-140.
ROVERETO, C. 1912. Los cocodrilos fósiles en las capas del Paraná. Anales del Mueso
Nacional de Buenos Aires 15 (22), Serie 3: 339-368.
RUEDA ALMONACID, J.V.; CARR, J.L.; MITTERMEISTER, R.A.; RODRÍGUEZ
MAHECHA, J.V.; MAST, R.B.; VOGT, R.C.; RHODIN, A.G.J.; OSSA VELÁSUQEZ,
J. DE LA; RUEDA, J.N. & C.G. MITTERMEISTER. 2007. Las tortugas y los
cocodrilianos de los países andinos del trópico. Serie de guías tropicales de campo 6.
Conservación Internacional. Editorial Panamericana, Formas e Impresos. 538p.
SALAS GISMONDI, R.; ANTONIE, P.O.; BABY, P.; BRUSSET, S.; BENAMMI, M.;
ESPURT, N.; DE FRANCESCHI, D.; PUJOS, F.; TEJADA, J. & M. URBINA. 2007.
Middle miocene crocodiles from the Fitzcarrald Arch, Amazonian Peru. En: DÍAZ
MARTÍNEZ, E. & I. RÁBANO (Eds.). 4th European Meeting on the Palaeontology
and Stratigraphy of Latin America. Cuadernos del Museo Geominero 8: 355-360.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 103
SALFITY, J.A. & C.R. MONALDI. 2006. Hoja Geológica 2566-IV: Metán (provincia
de Salta). Programa Nacional de Cartas Geológicas de la República Argentina.
Servicio Geológico Minero Argentino, Boletín 319, 74p.
SÁNCHEZ VILLAGRA, M.R. & O.A. AGUILERA. 2006. Neogene vertebrates from
Urumaco, Falcón State, Venezuela: diversity and significance. Journal of Systematic
Palaeontology 4 (3): 213-220.
SCALABRINI, P. 1887. Cartas Científicas. Museo Provincial de Entre Ríos (no visto).
SCHMIDT, K.P. 1928. Notes on South American caimans. Field Museum of Natural
History, Zoological Series 12 (17): 205-231.
SERENO, P.C.; LARSSON, H.C.E.; SIDOR, C.A. & B. GADO. 2001. The giant
crocodyliform Sarcosuchus from the Cretaceous of Africa. Science 294: 1516-1519.
SERENO, P.C. & H.C.E. LARSSON. 2009. Cretaceous crocodyliforms from the
Sahara. Zookeys 28: 1-143.
SILL, W.D. 1968. The zoogeography of the Crocodilia. Copeia 1968 (1): 76-88.
SILL, W.D. 1970. Nota preliminar sobre un nuevo gavial del Plioceno de Venezuela y
una discusión de los gaviales sudamericanos. Ameghiniana 7 (2): 151-159.
SIMPSON, G.G. 1933. A new crocodilian from the Notostylops beds of Patagonia.
American Museum Novitates 623: 1-9.
SIMPSON, G.G. 1937. An ancient eusuchian crocodile from Patagonia. American
Museum Novitates 965 (29): 1-20.
STARCK, D. & L.M. ANZÓTEGUI. 2001. The late miocene climatic change-
persistence of a climatic signal through the orogenic stratigraphic record in
northwestern Argentina. Journal of South American Earth Sciences 14: 763-774.
TAPLIN, L.E. & G.C. GRIGG. 1989. Historical zoogeography of the eusuchian
crocodilians: a physiological perspective. American Zoologist 29: 885-901.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 104
TURNER, A.H. & G.A. BUCKLEY. 2008. Mahajangasuchus insignis
(Crocodyliformes: Mesoeucrocodylia) cranial anatomy and new data on the origin of
the eusuchian-style palate. Journal of Vertebrate Paleontology 28 (2): 382-408.
VÉLEZ JUARBE, J.; BROCHU, C.A. & H. SANTOS. 2006. A gharial from the
Oligocene of Puerto Rico: transoceanic dispersal in the history of a non-marine reptile.
Proceedings of the Royal Society B 274 (1615): 1245-1254.
VERDADE, L.M. 2000. Regression equations between body and head measurements in
the broad-snouted caiman (Caiman latirostris). Revista Brasileira de Biologia 60 (3):
469-482.
VERDADE, L.M. 2003. Cranial sexual dimorphism in captive adult broad-snouted
caiman (Caiman latirostris). Amphibia-Reptilia 24 (1): 92-99.
VERDADE, L.M. & C.I. PIÑA. 2006. Caiman latirostris. En: Catalogue of American
Amphibians and Reptiles. Editado por la Society for Study of Amphibians and Reptiles
833: 1-21.
VERDADE, L.M. & C.I. PIÑA. 2007. O jacaré-de-papo-amarelo (Caiman latirostris
Daudin, 1802). Herpetología no Brasil 2: 295-307.
VERDADE, L.M.; LARRIERA, A. & C.I. PIÑA. 2010. Broad-snouted Caiman Caiman
latirostris. En: MANOLIS, S.C. & C. STEVENSON (Eds.). Crocodiles. Status Survey
and Conservation Action Plan. Third Edition. Crocodile Specialist group: Darwin, 18-
22.
ZURITA, A.E. 2007. Los Hoplophorini (Xenarthra, Glyptodontidae) del “Araucanense”
(Mioceno tardío-Plioceno) del noroeste de la Argentina: sistemática, paleobiogeografía
y paleoambiente. Ameghiniana 44 (2): 257-269.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 105
APÉNDICE I
Lista de Láminas y Figuras:
Figura 1: Relaciones de los principales clados de Archosauria. Tomada de Nesbitt, 2011.
Figura 2: Principales grupos de Crocodylomorpha y sus relaciones. Tomada de Brochu et al., 2009.
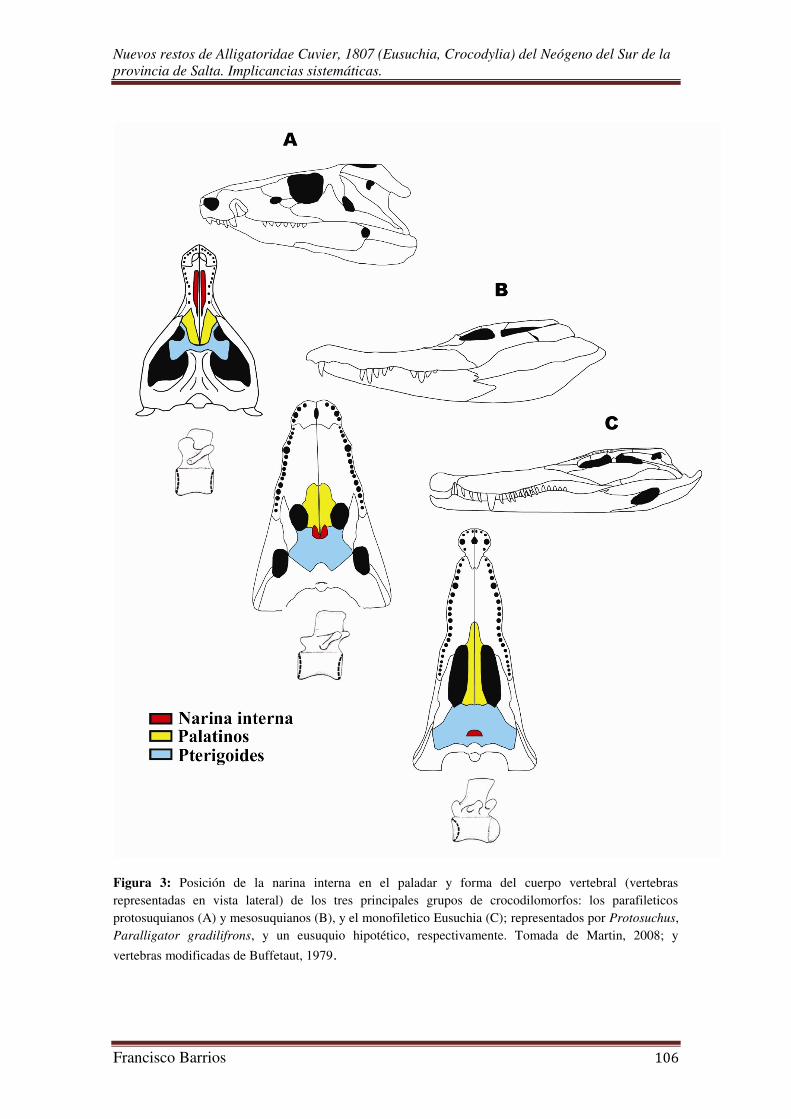
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 106
Figura 3: Posición de la narina interna en el paladar y forma del cuerpo vertebral (vertebras representadas en vista lateral) de los tres principales grupos de crocodilomorfos: los parafileticos protosuquianos (A) y mesosuquianos (B), y el monofiletico Eusuchia (C); representados por Protosuchus, Paralligator gradilifrons, y un eusuquio hipotético, respectivamente. Tomada de Martin, 2008; y
vertebras modificadas de Buffetaut, 1979.
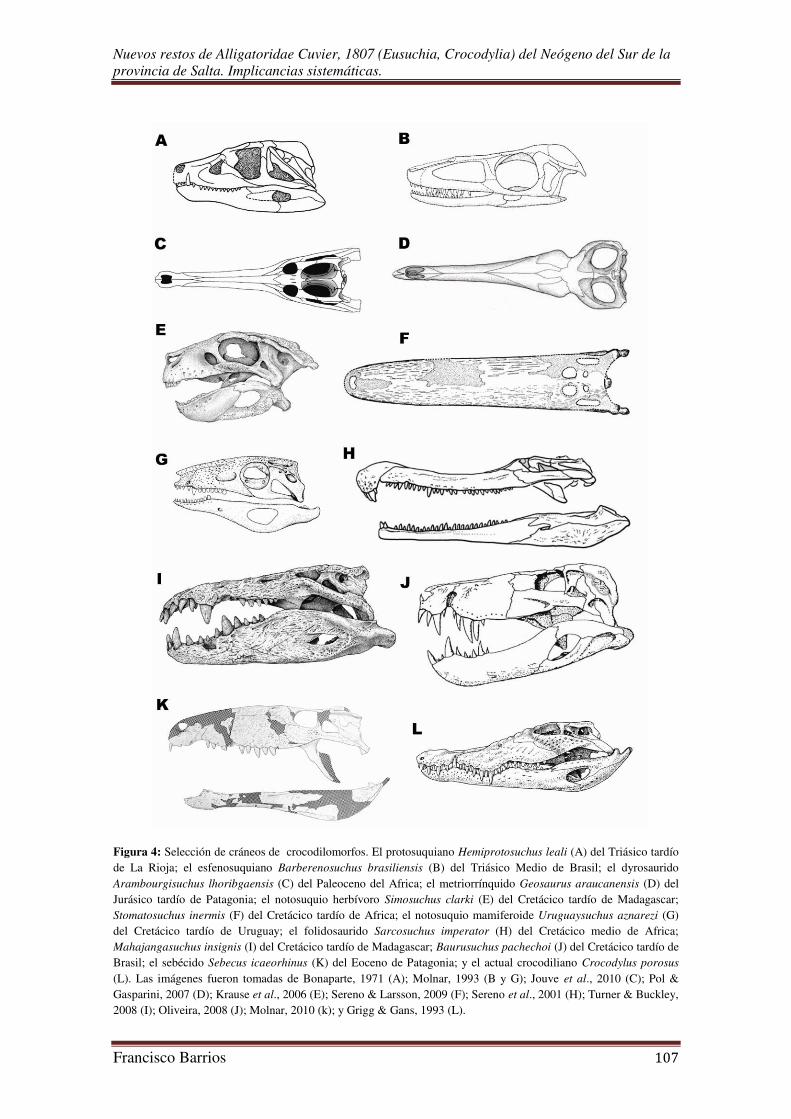
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 107
Figura 4: Selección de cráneos de crocodilomorfos. El protosuquiano Hemiprotosuchus leali (A) del Triásico tardío de La Rioja; el esfenosuquiano Barberenosuchus brasiliensis (B) del Triásico Medio de Brasil; el dyrosaurido Arambourgisuchus lhoribgaensis (C) del Paleoceno del Africa; el metriorrínquido Geosaurus araucanensis (D) del Jurásico tardío de Patagonia; el notosuquio herbívoro Simosuchus clarki (E) del Cretácico tardío de Madagascar; Stomatosuchus inermis (F) del Cretácico tardío de Africa; el notosuquio mamiferoide Uruguaysuchus aznarezi (G) del Cretácico tardío de Uruguay; el folidosaurido Sarcosuchus imperator (H) del Cretácico medio de Africa; Mahajangasuchus insignis (I) del Cretácico tardío de Madagascar; Baurusuchus pachechoi (J) del Cretácico tardío de Brasil; el sebécido Sebecus icaeorhinus (K) del Eoceno de Patagonia; y el actual crocodiliano Crocodylus porosus (L). Las imágenes fueron tomadas de Bonaparte, 1971 (A); Molnar, 1993 (B y G); Jouve et al., 2010 (C); Pol & Gasparini, 2007 (D); Krause et al., 2006 (E); Sereno & Larsson, 2009 (F); Sereno et al., 2001 (H); Turner & Buckley, 2008 (I); Oliveira, 2008 (J); Molnar, 2010 (k); y Grigg & Gans, 1993 (L).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 108
Figura 5: Esquema de relaciones de las especies vivientes y de los tres grandes grupos dentro de Crocodylia; a partir de la combinación de datos morfológicos y de secuencia. Tomada de Poe, 1996.
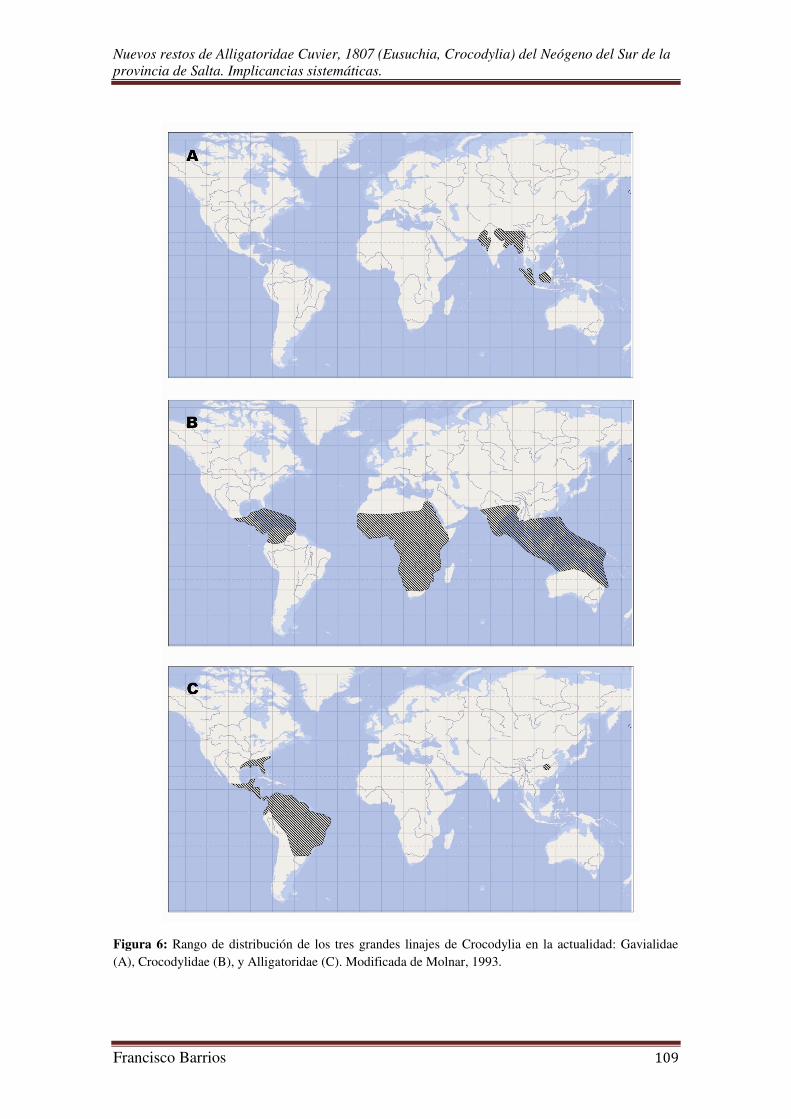
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 109
Figura 6: Rango de distribución de los tres grandes linajes de Crocodylia en la actualidad: Gavialidae (A), Crocodylidae (B), y Alligatoridae (C). Modificada de Molnar, 1993.
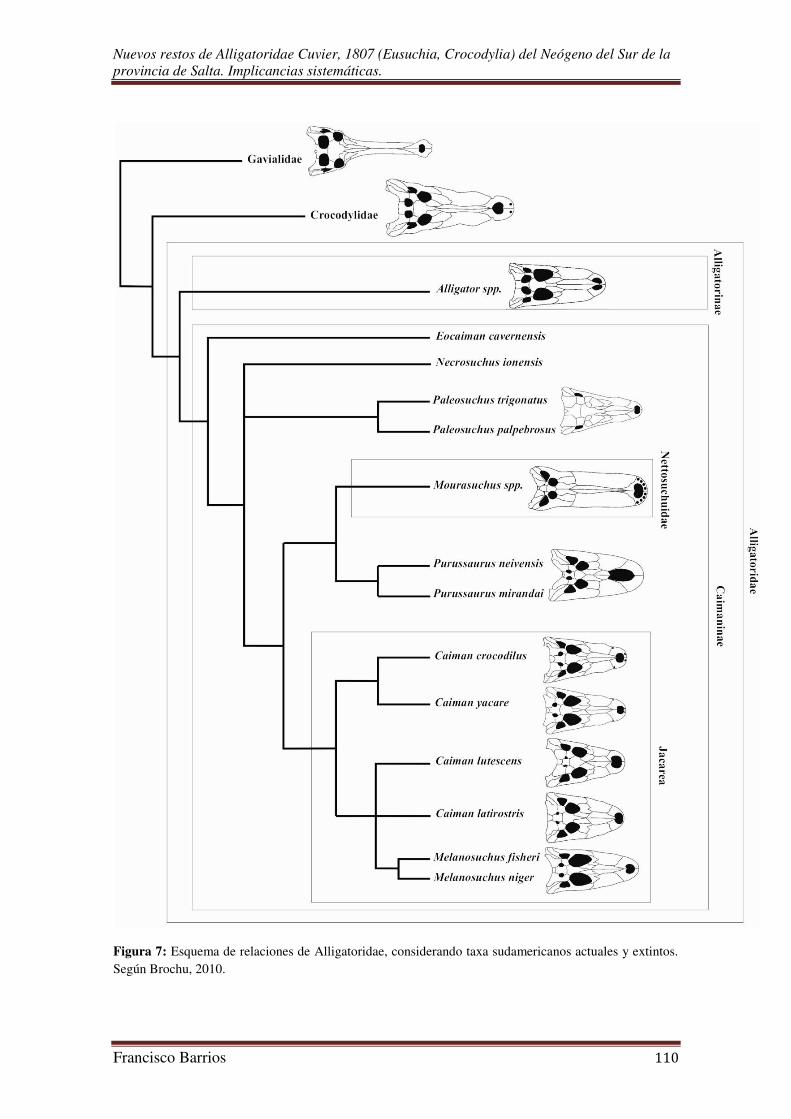
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 110
Figura 7: Esquema de relaciones de Alligatoridae, considerando taxa sudamericanos actuales y extintos. Según Brochu, 2010.
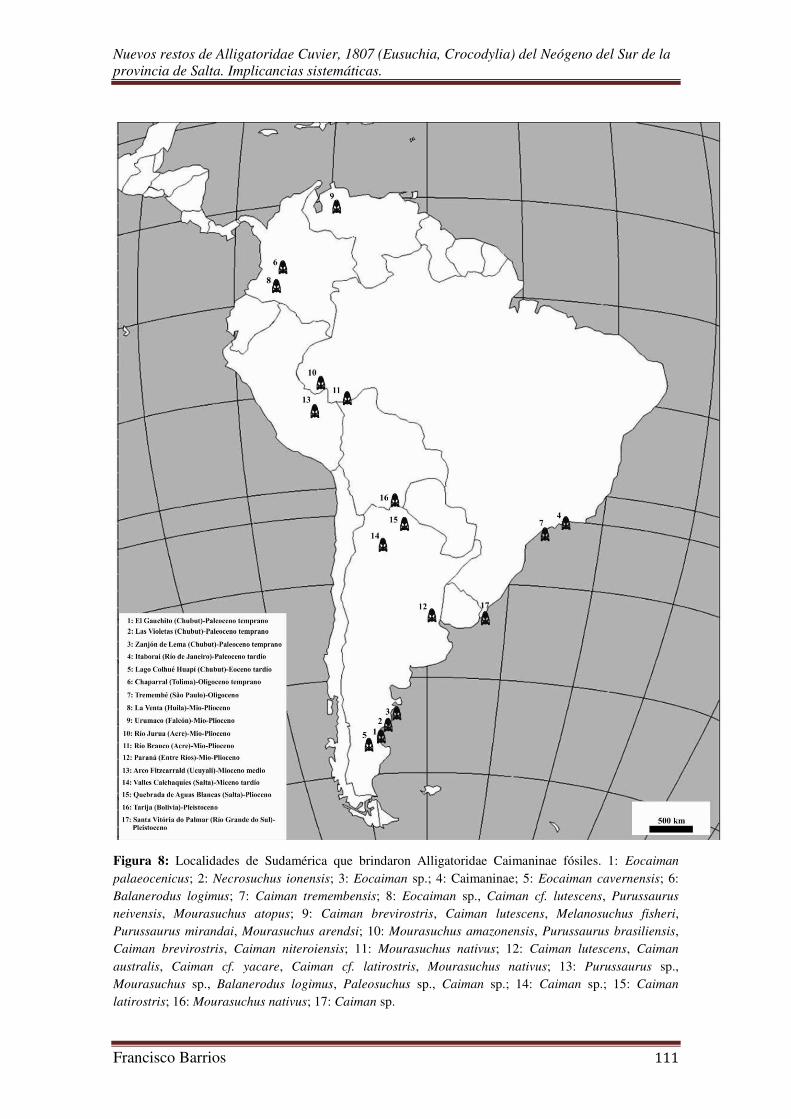
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 111
Figura 8: Localidades de Sudamérica que brindaron Alligatoridae Caimaninae fósiles. 1: Eocaiman palaeocenicus; 2: Necrosuchus ionensis; 3: Eocaiman sp.; 4: Caimaninae; 5: Eocaiman cavernensis; 6: Balanerodus logimus; 7: Caiman tremembensis; 8: Eocaiman sp., Caiman cf. lutescens, Purussaurus neivensis, Mourasuchus atopus; 9: Caiman brevirostris, Caiman lutescens, Melanosuchus fisheri, Purussaurus mirandai, Mourasuchus arendsi; 10: Mourasuchus amazonensis, Purussaurus brasiliensis, Caiman brevirostris, Caiman niteroiensis; 11: Mourasuchus nativus; 12: Caiman lutescens, Caiman australis, Caiman cf. yacare, Caiman cf. latirostris, Mourasuchus nativus; 13: Purussaurus sp., Mourasuchus sp., Balanerodus logimus, Paleosuchus sp., Caiman sp.; 14: Caiman sp.; 15: Caiman latirostris; 16: Mourasuchus nativus; 17: Caiman sp.
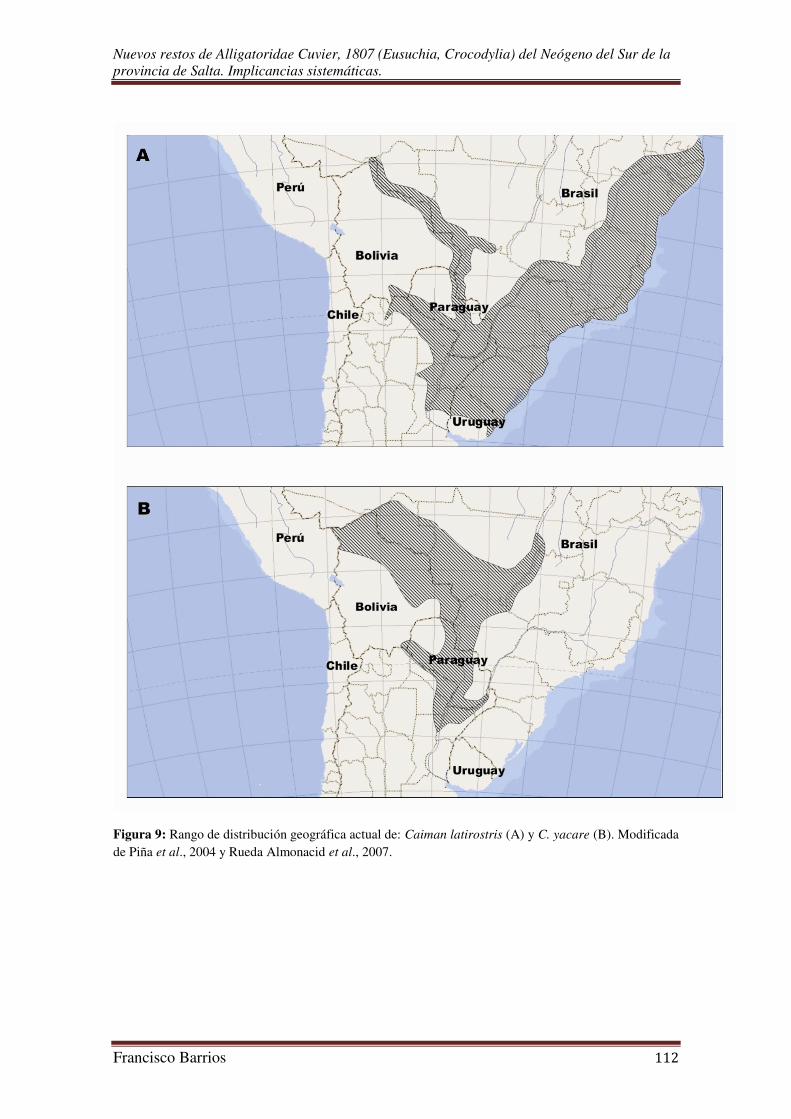
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 112
Figura 9: Rango de distribución geográfica actual de: Caiman latirostris (A) y C. yacare (B). Modificada de Piña et al., 2004 y Rueda Almonacid et al., 2007.
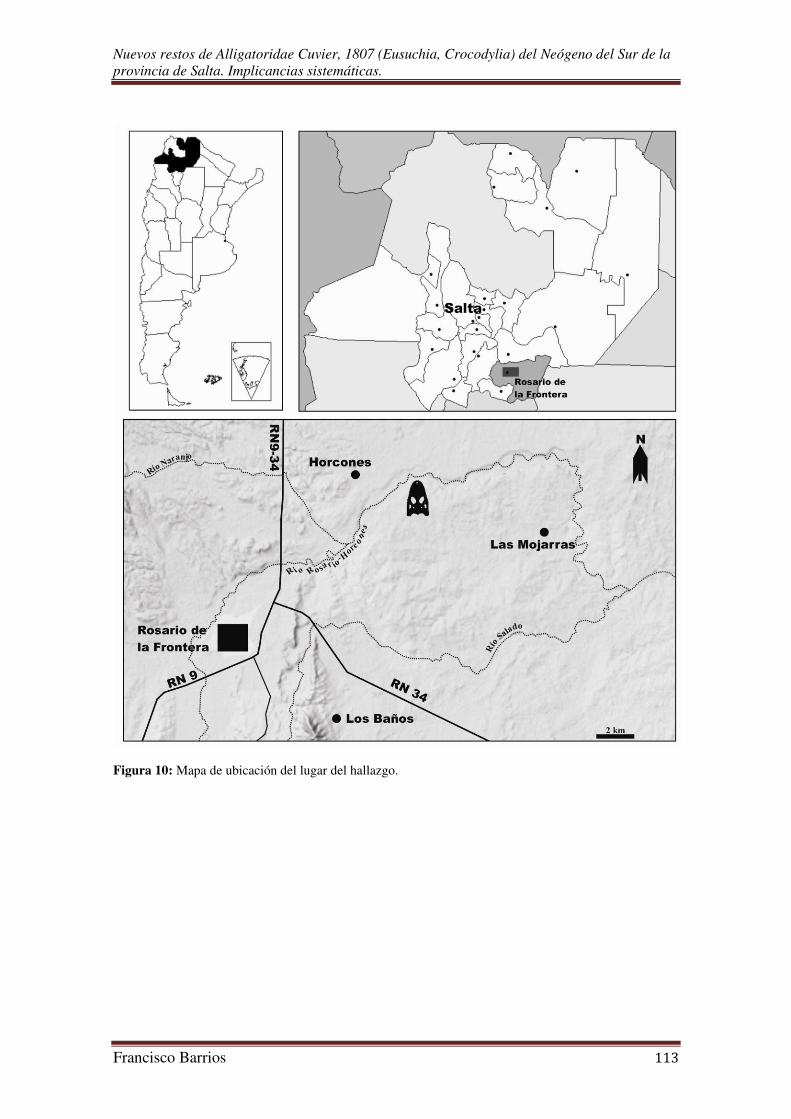
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 113
Figura 10: Mapa de ubicación del lugar del hallazgo.
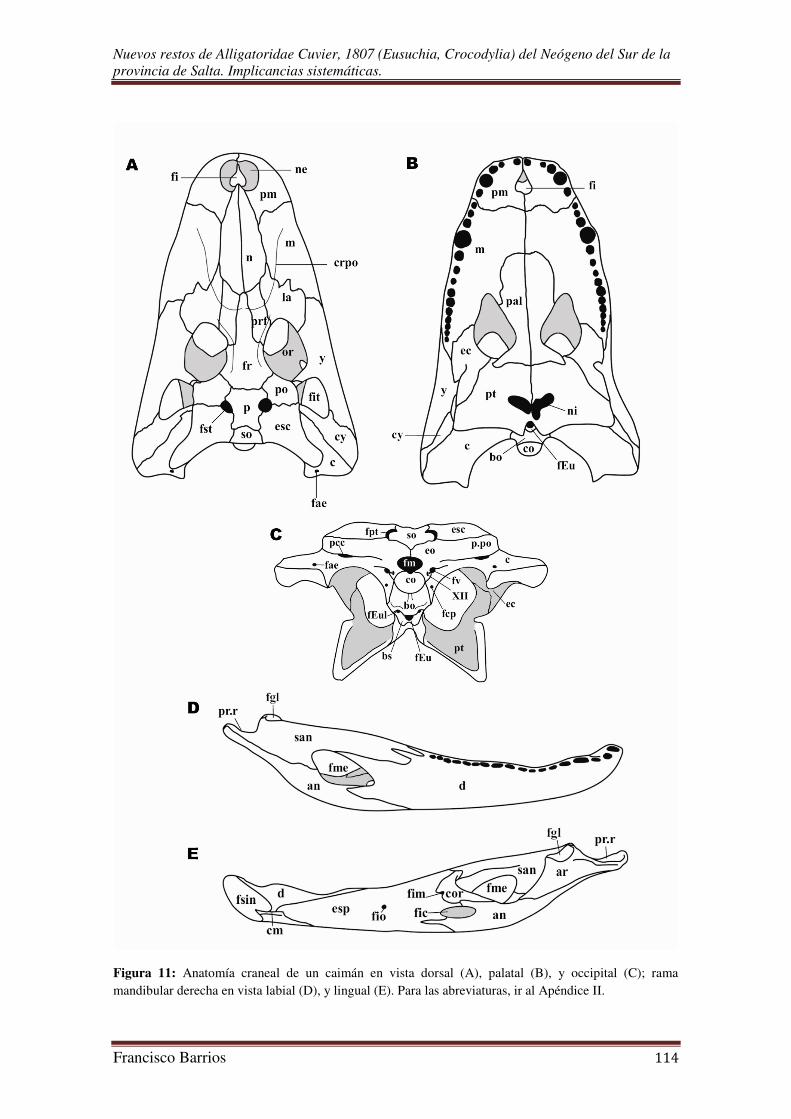
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 114
Figura 11: Anatomía craneal de un caimán en vista dorsal (A), palatal (B), y occipital (C); rama mandibular derecha en vista labial (D), y lingual (E). Para las abreviaturas, ir al Apéndice II.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 115
Figura 12: Cráneo de MAS-PALEO-RdlF 001/2011 en vista dorsal; fotografía (A) e interpretación esquemática (B). Escala de la barra=5 cm.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 116
Figura 13: Cráneo de MAS-PALEO-RdlF 001/2011 en vista ventral (palatal); fotografía (A) e interpretación esquemática (B). Escala de la barra=5 cm.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 117
Figura 14: Fragmento rostral derecho de MAS-PALEO-RdlF 001/2011 en vista lateral labial; fotografía (A) e interpretación esquemática (B). Escala de la barra=5 cm.
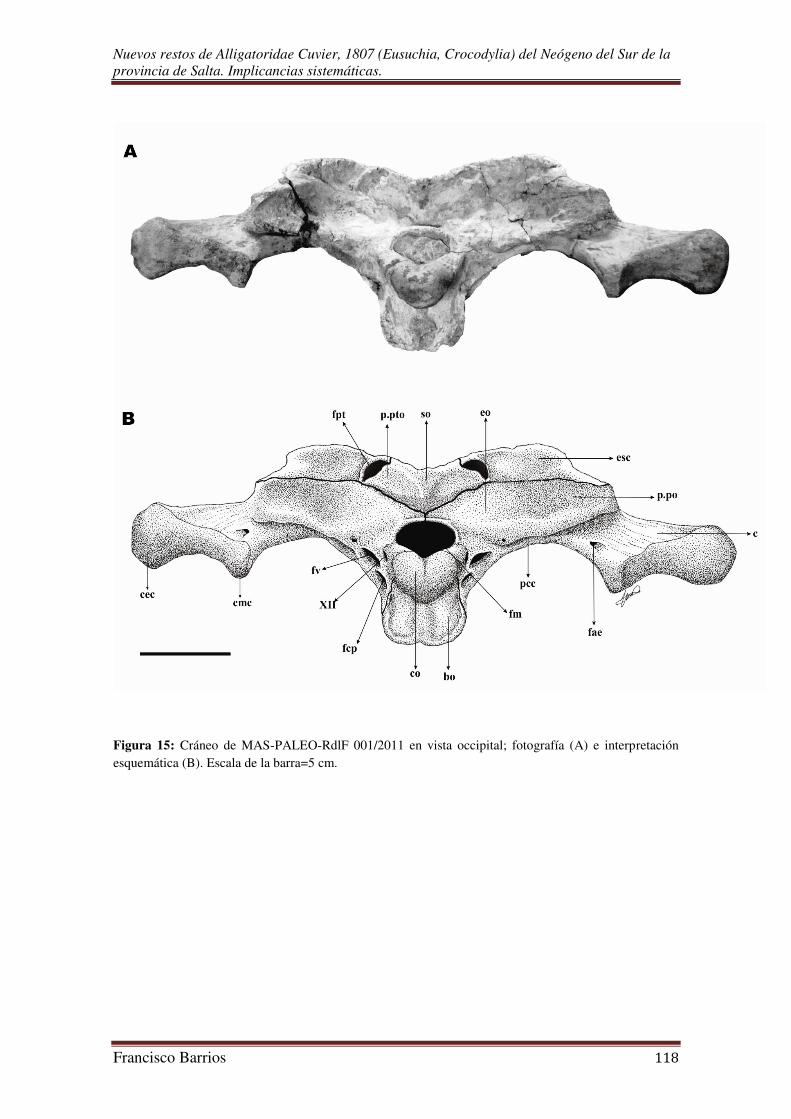
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 118
Figura 15: Cráneo de MAS-PALEO-RdlF 001/2011 en vista occipital; fotografía (A) e interpretación esquemática (B). Escala de la barra=5 cm.
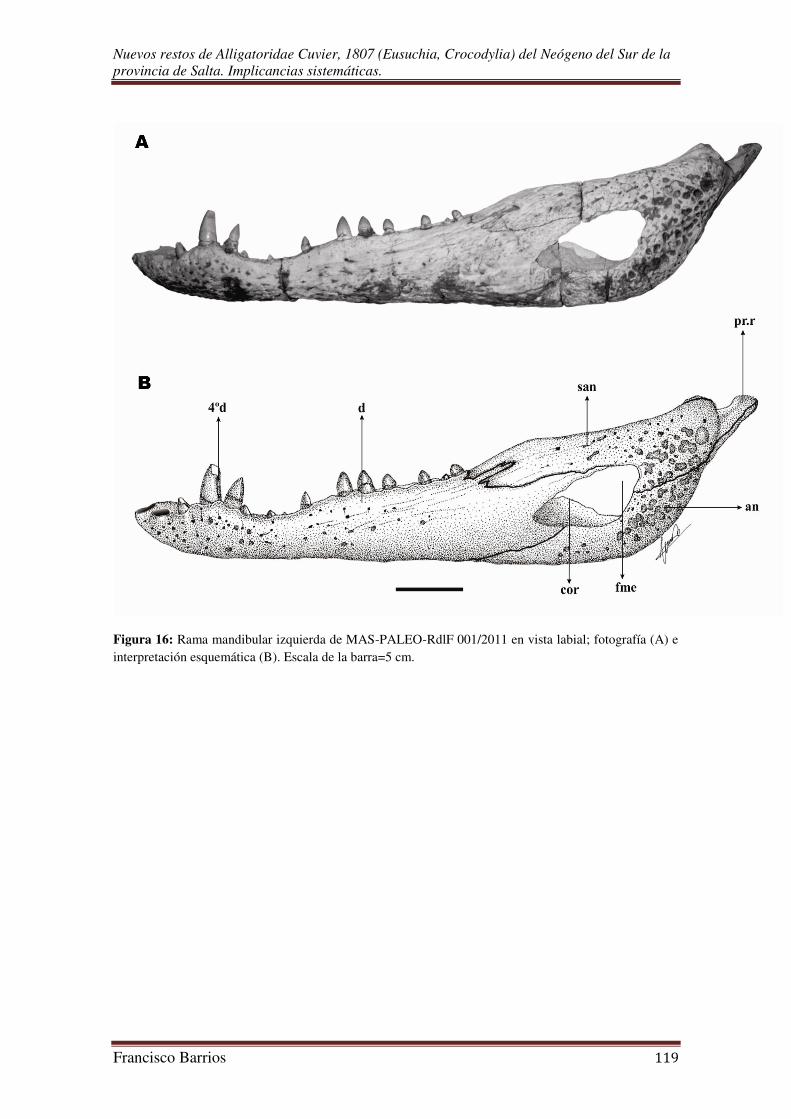
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 119
Figura 16: Rama mandibular izquierda de MAS-PALEO-RdlF 001/2011 en vista labial; fotografía (A) e interpretación esquemática (B). Escala de la barra=5 cm.
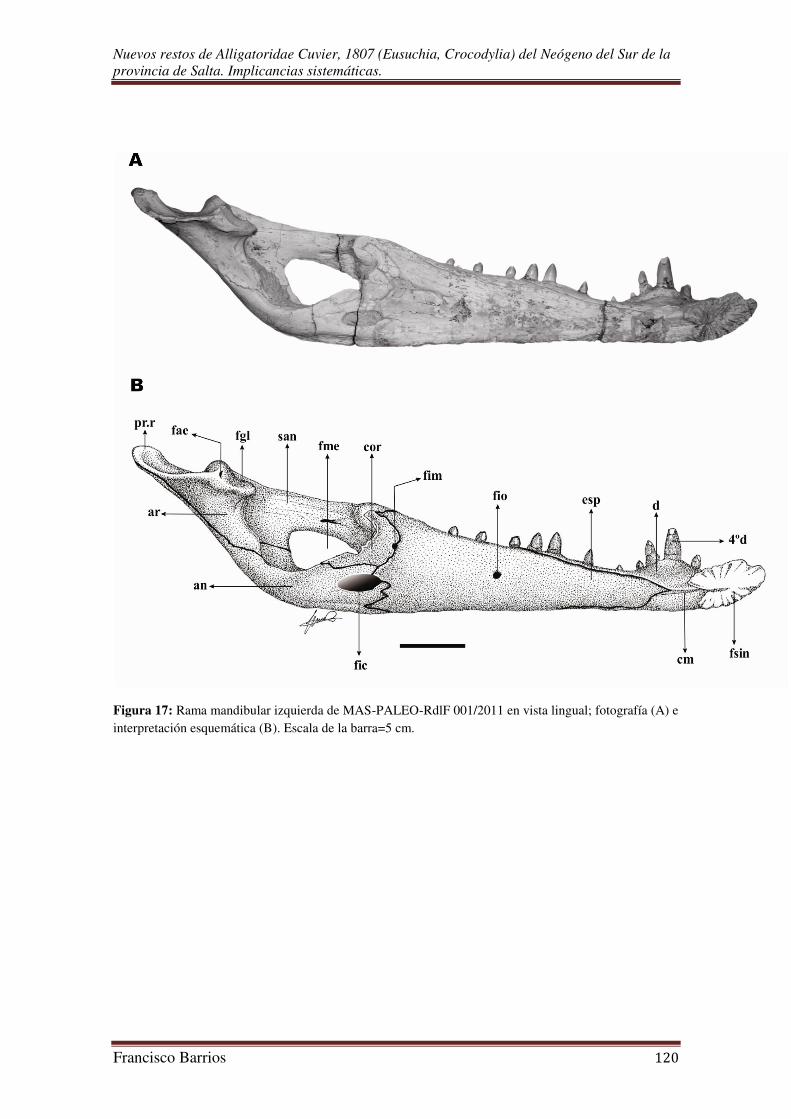
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 120
Figura 17: Rama mandibular izquierda de MAS-PALEO-RdlF 001/2011 en vista lingual; fotografía (A) e interpretación esquemática (B). Escala de la barra=5 cm.
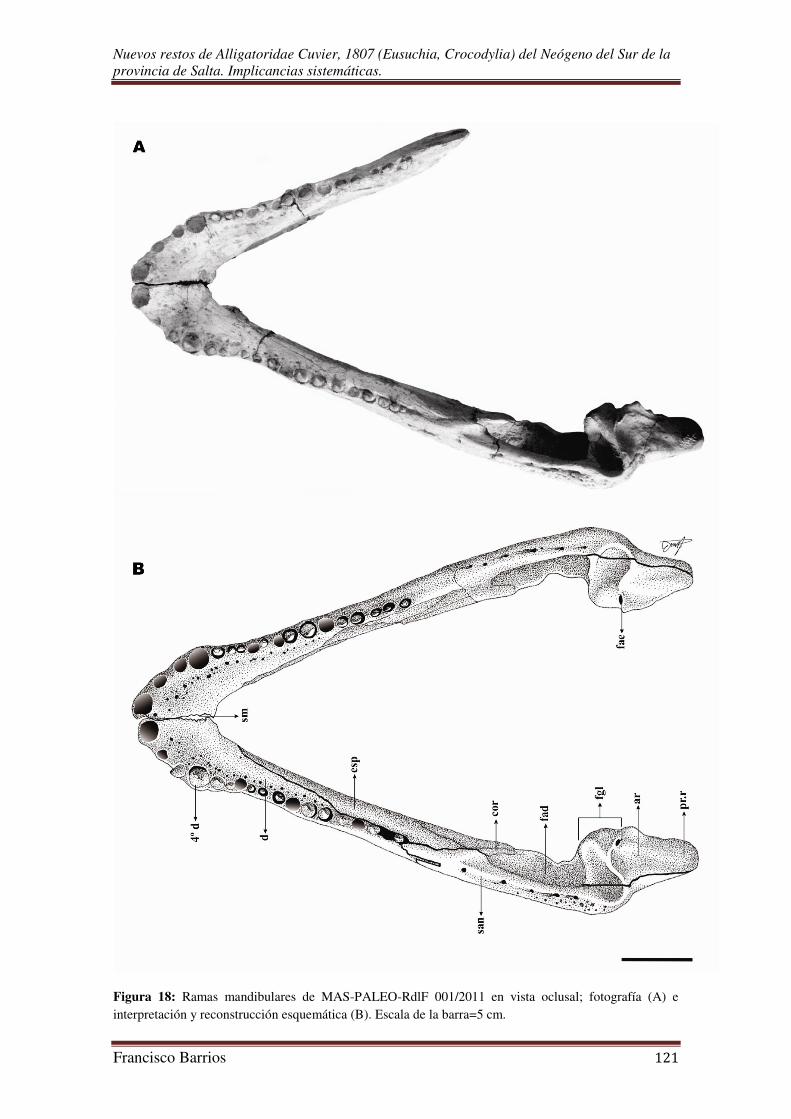
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 121
Figura 18: Ramas mandibulares de MAS-PALEO-RdlF 001/2011 en vista oclusal; fotografía (A) e interpretación y reconstrucción esquemática (B). Escala de la barra=5 cm.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 122
Figura 19: Gavialis gangeticus. Vista A dorsal, B palatal, C oclusal, y D lateral. Modificada de Mook
(1921c). Escala de la barra=5 cm.
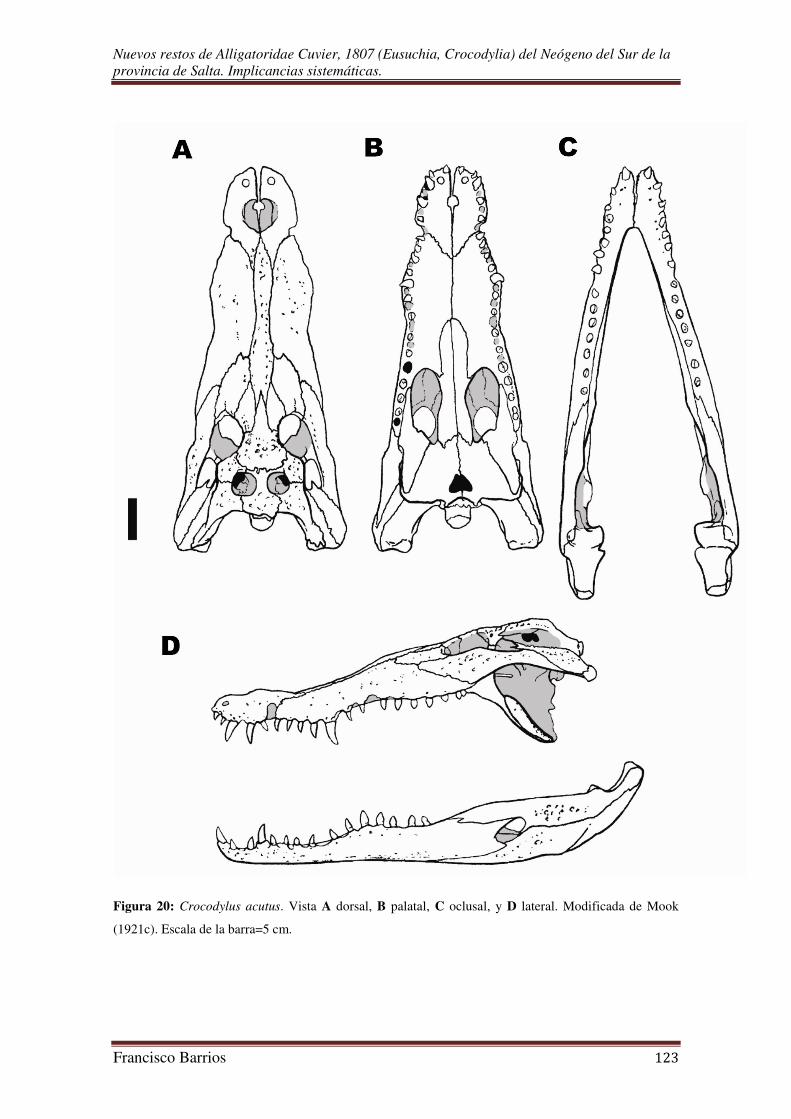
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 123
Figura 20: Crocodylus acutus. Vista A dorsal, B palatal, C oclusal, y D lateral. Modificada de Mook
(1921c). Escala de la barra=5 cm.
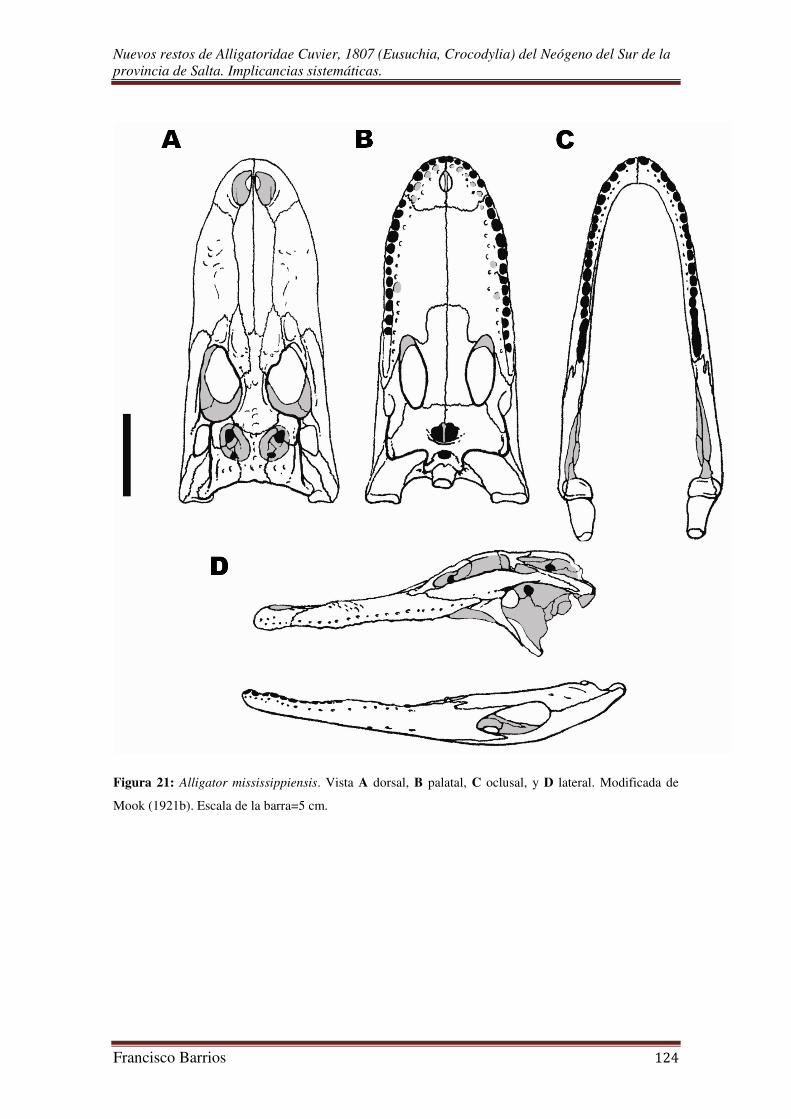
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 124
Figura 21: Alligator mississippiensis. Vista A dorsal, B palatal, C oclusal, y D lateral. Modificada de
Mook (1921b). Escala de la barra=5 cm.
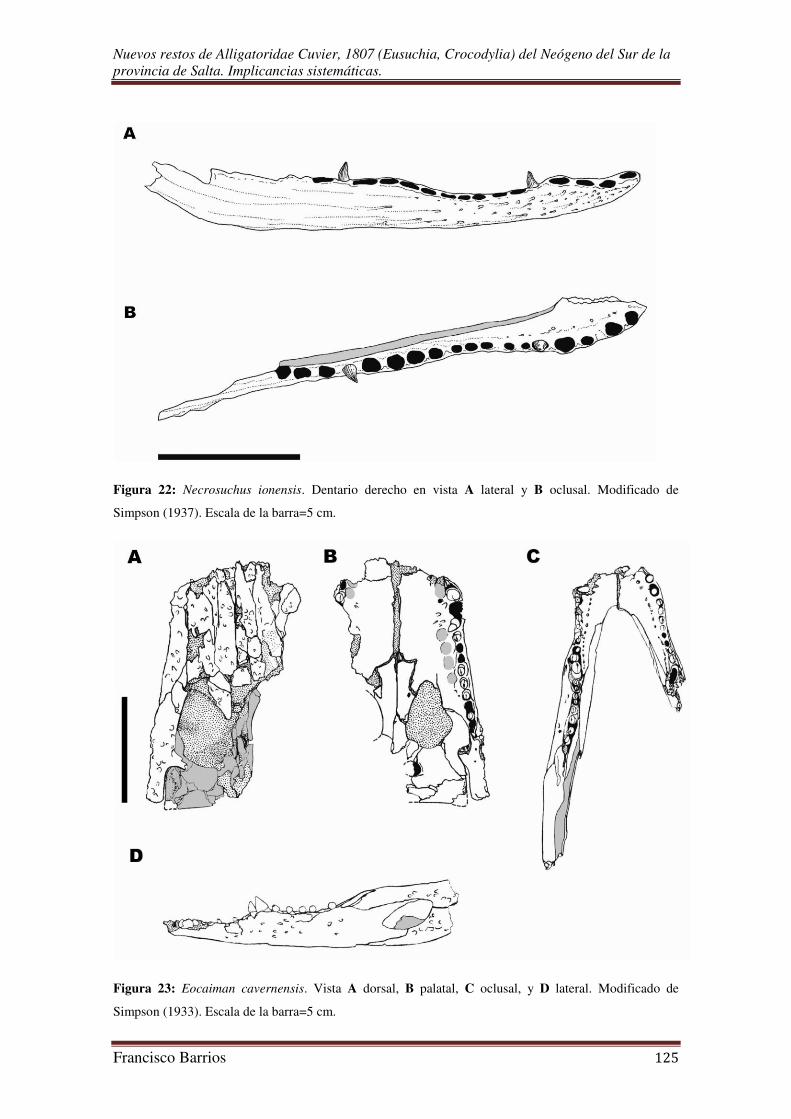
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 125
Figura 22: Necrosuchus ionensis. Dentario derecho en vista A lateral y B oclusal. Modificado de
Simpson (1937). Escala de la barra=5 cm.
Figura 23: Eocaiman cavernensis. Vista A dorsal, B palatal, C oclusal, y D lateral. Modificado de
Simpson (1933). Escala de la barra=5 cm.
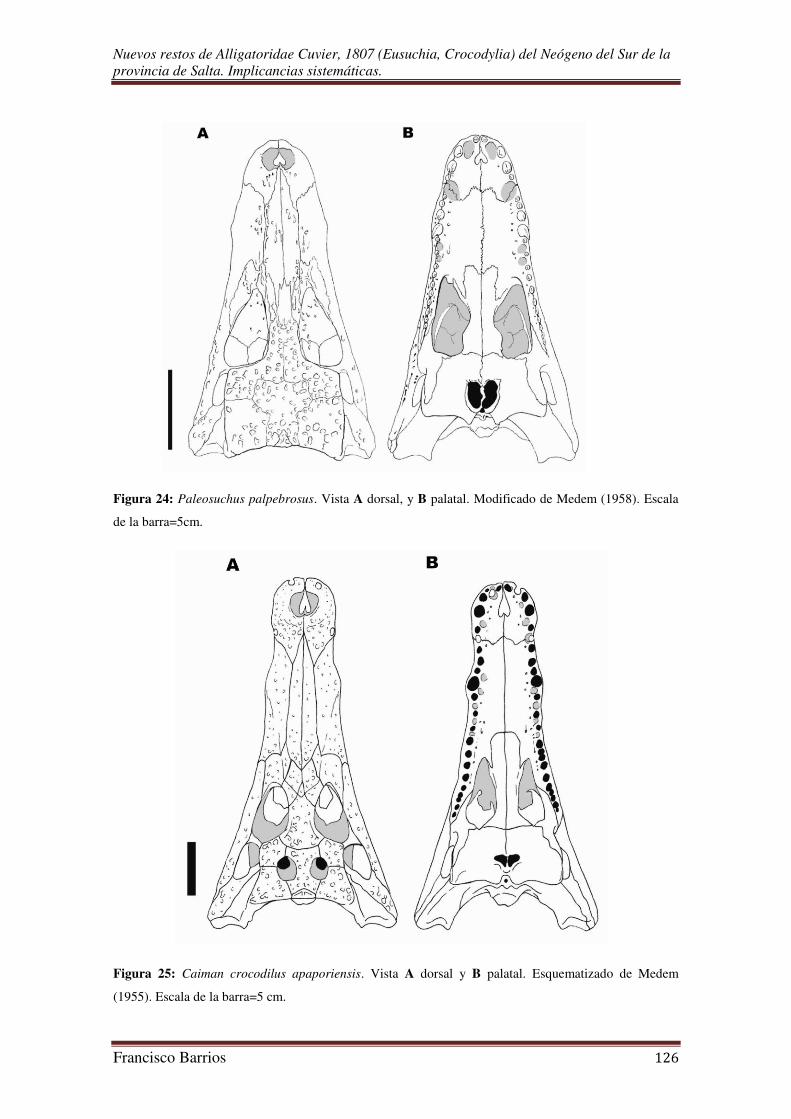
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 126
Figura 24: Paleosuchus palpebrosus. Vista A dorsal, y B palatal. Modificado de Medem (1958). Escala
de la barra=5cm.
Figura 25: Caiman crocodilus apaporiensis. Vista A dorsal y B palatal. Esquematizado de Medem
(1955). Escala de la barra=5 cm.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 127
Figura 26: Purussaurus mirandai. Vista A dorsal, B palatal, C oclusal, y D lateral. Esquematizado a
partir de Langston (1965) y Aguilera et al. (2006). Escala de la barra=50cm.
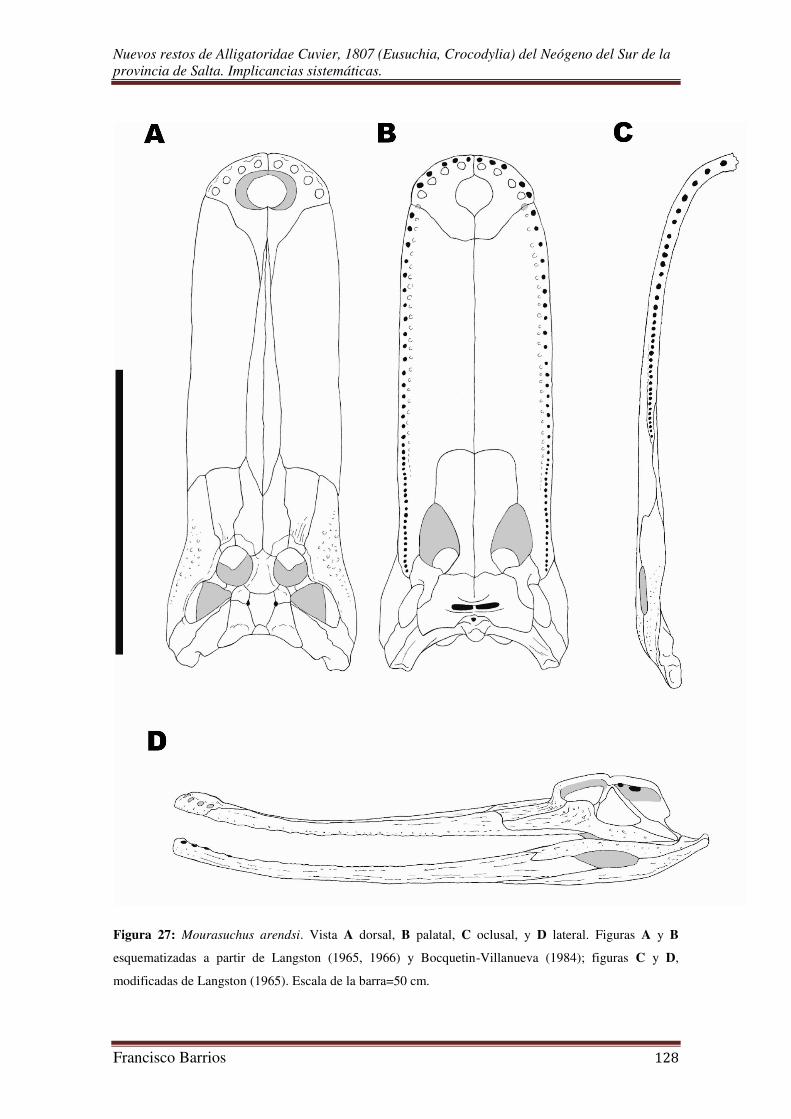
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 128
Figura 27: Mourasuchus arendsi. Vista A dorsal, B palatal, C oclusal, y D lateral. Figuras A y B
esquematizadas a partir de Langston (1965, 1966) y Bocquetin-Villanueva (1984); figuras C y D,
modificadas de Langston (1965). Escala de la barra=50 cm.
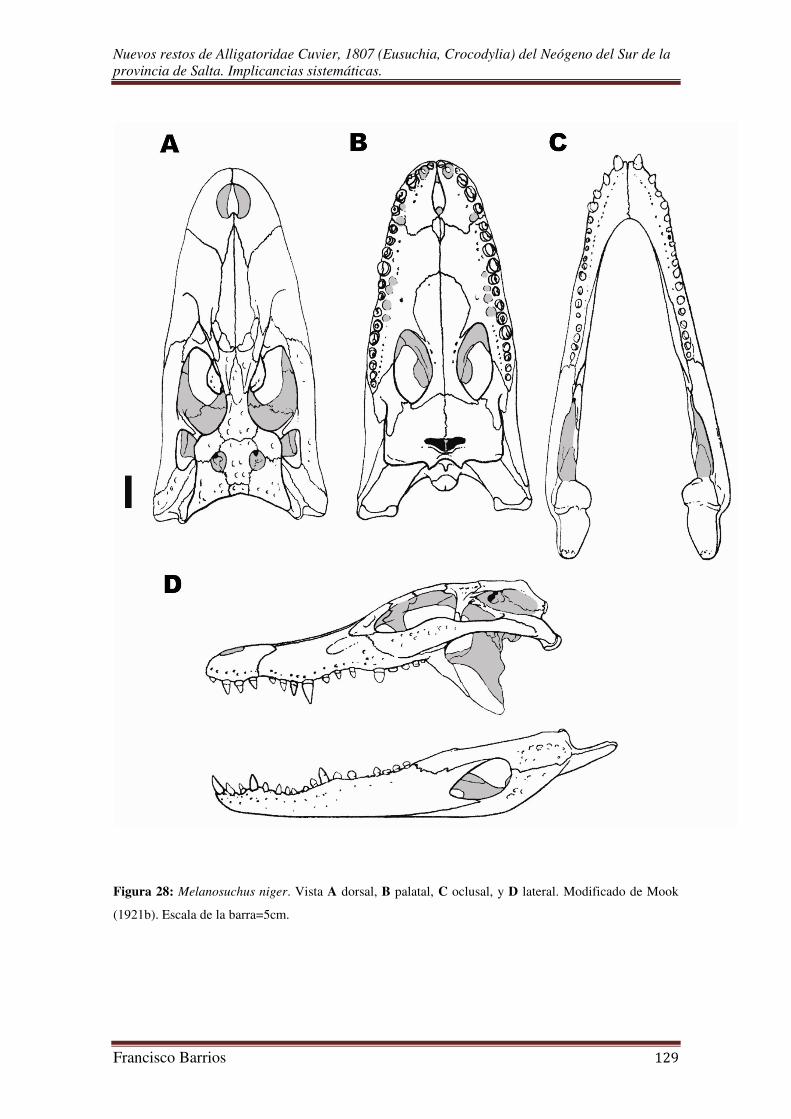
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 129
Figura 28: Melanosuchus niger. Vista A dorsal, B palatal, C oclusal, y D lateral. Modificado de Mook
(1921b). Escala de la barra=5cm.
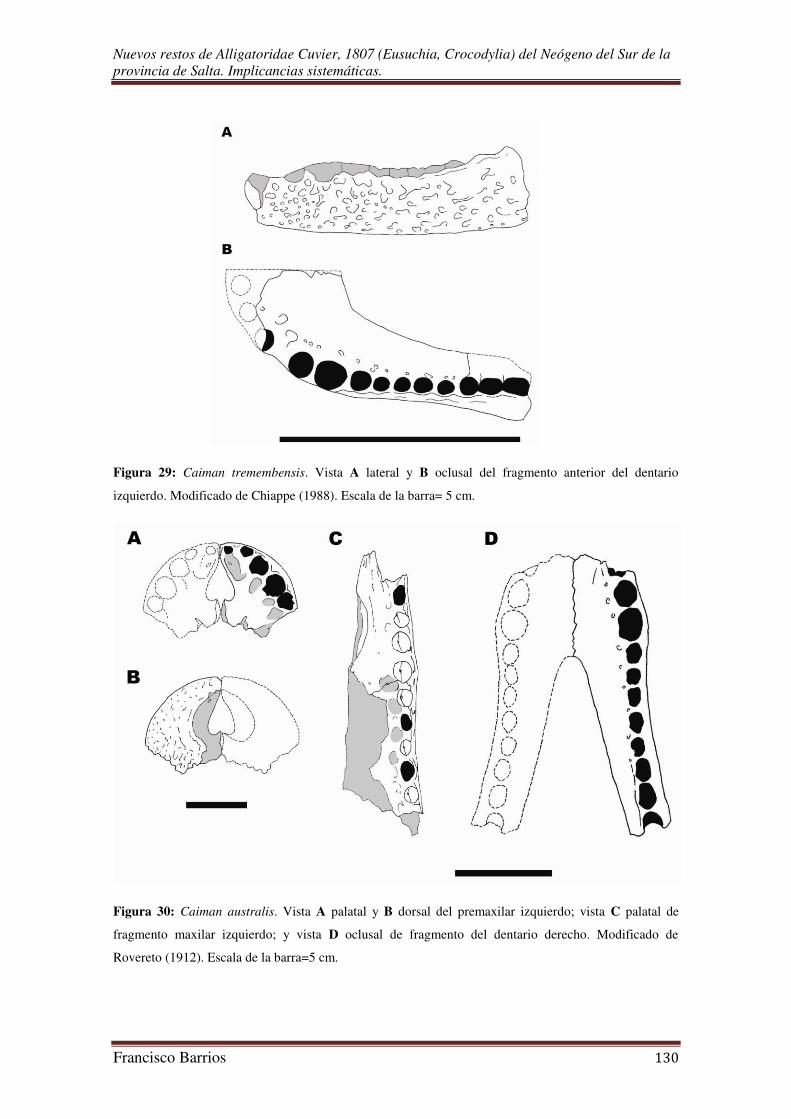
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 130
Figura 29: Caiman tremembensis. Vista A lateral y B oclusal del fragmento anterior del dentario
izquierdo. Modificado de Chiappe (1988). Escala de la barra= 5 cm.
Figura 30: Caiman australis. Vista A palatal y B dorsal del premaxilar izquierdo; vista C palatal de
fragmento maxilar izquierdo; y vista D oclusal de fragmento del dentario derecho. Modificado de
Rovereto (1912). Escala de la barra=5 cm.
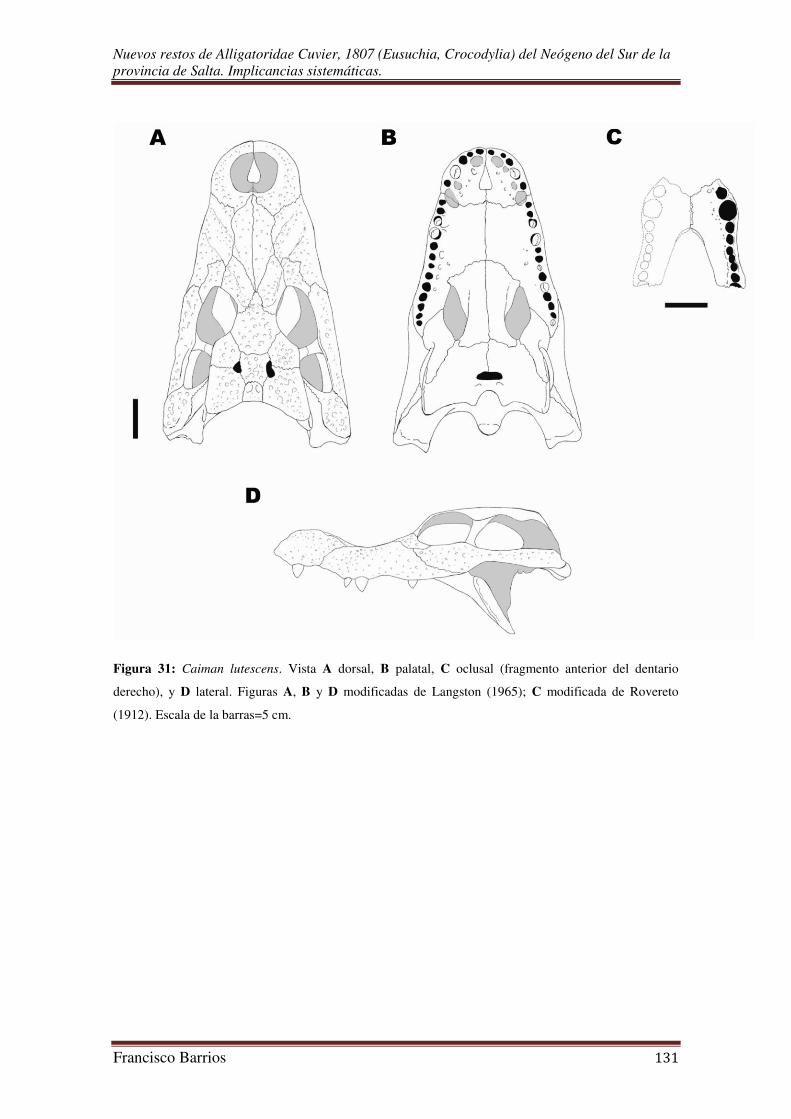
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 131
Figura 31: Caiman lutescens. Vista A dorsal, B palatal, C oclusal (fragmento anterior del dentario
derecho), y D lateral. Figuras A, B y D modificadas de Langston (1965); C modificada de Rovereto
(1912). Escala de la barras=5 cm.
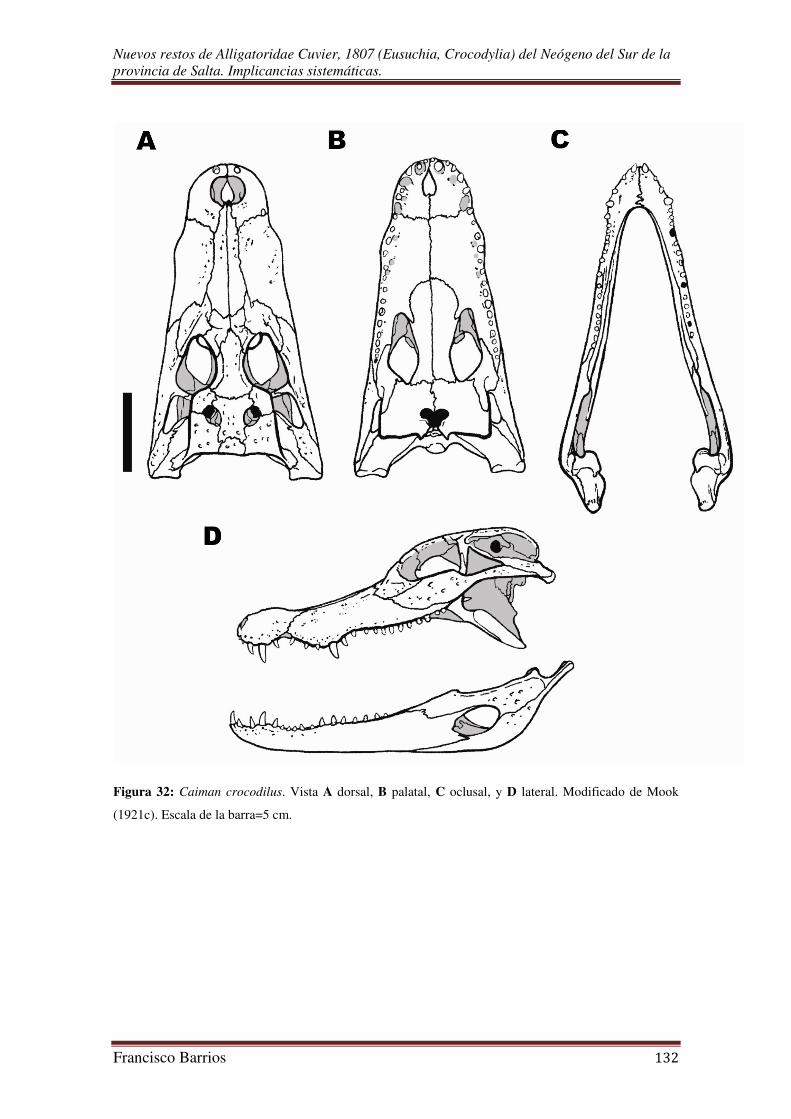
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 132
Figura 32: Caiman crocodilus. Vista A dorsal, B palatal, C oclusal, y D lateral. Modificado de Mook
(1921c). Escala de la barra=5 cm.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 133
Figura 33: Caiman yacare. Vista A dorsal, B palatal, C oclusal, y D lateral. Escala de la barra =5 cm,
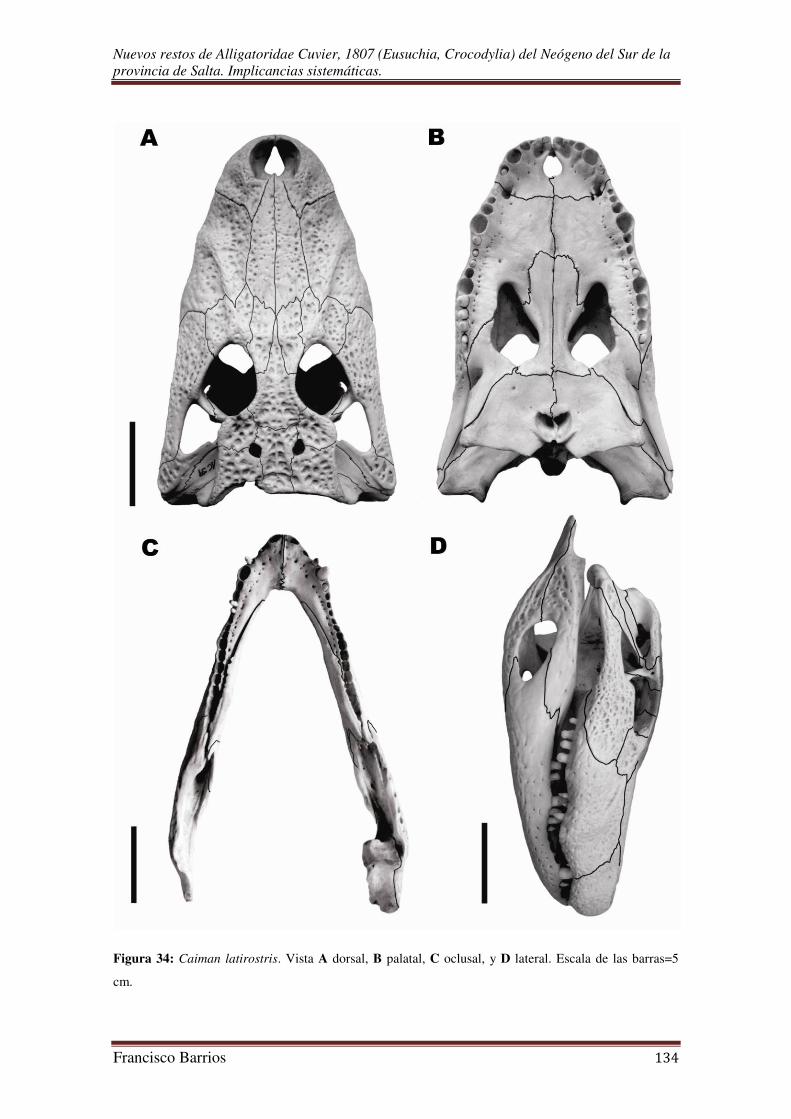
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 134
Figura 34: Caiman latirostris. Vista A dorsal, B palatal, C oclusal, y D lateral. Escala de las barras=5
cm.
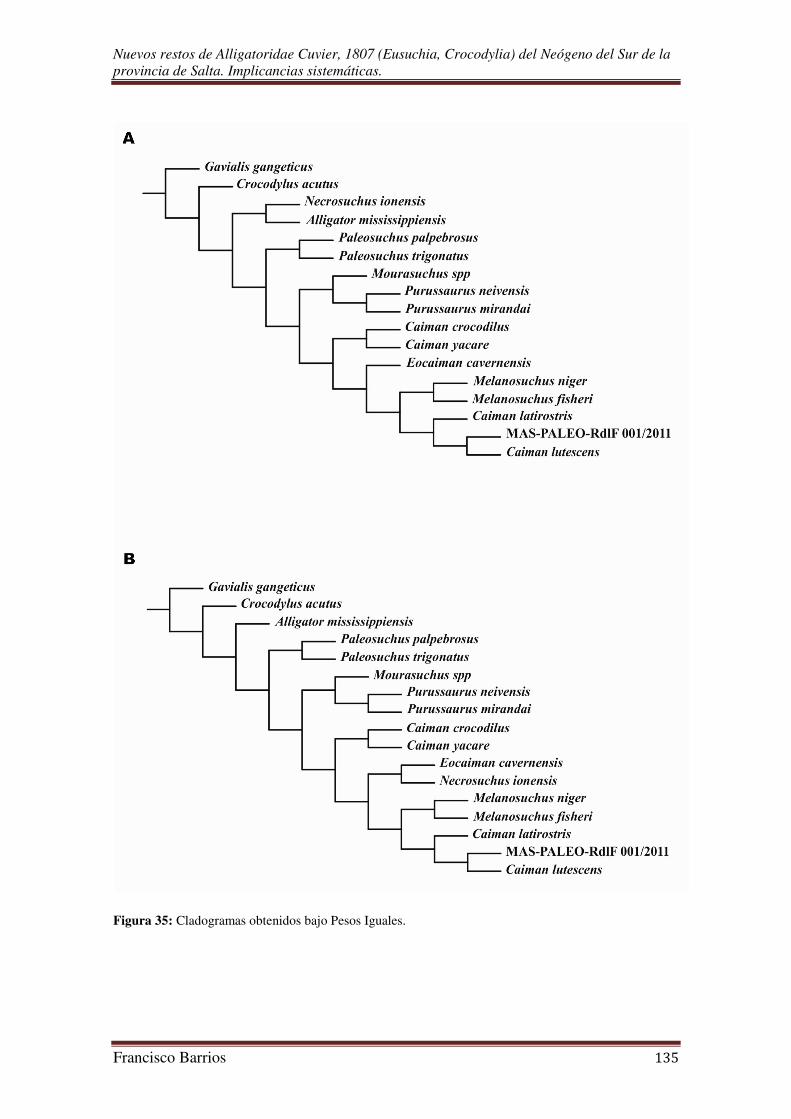
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 135
Figura 35: Cladogramas obtenidos bajo Pesos Iguales.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 136
Figura 36: Cladogramas A y B obtenidos bajo Pesos Implicados (K3-7). Consenso estricto de los dos
árboles más parsimoniosos obtenidos tanto para Pesos Iguales como para Pesos Implicados (C).
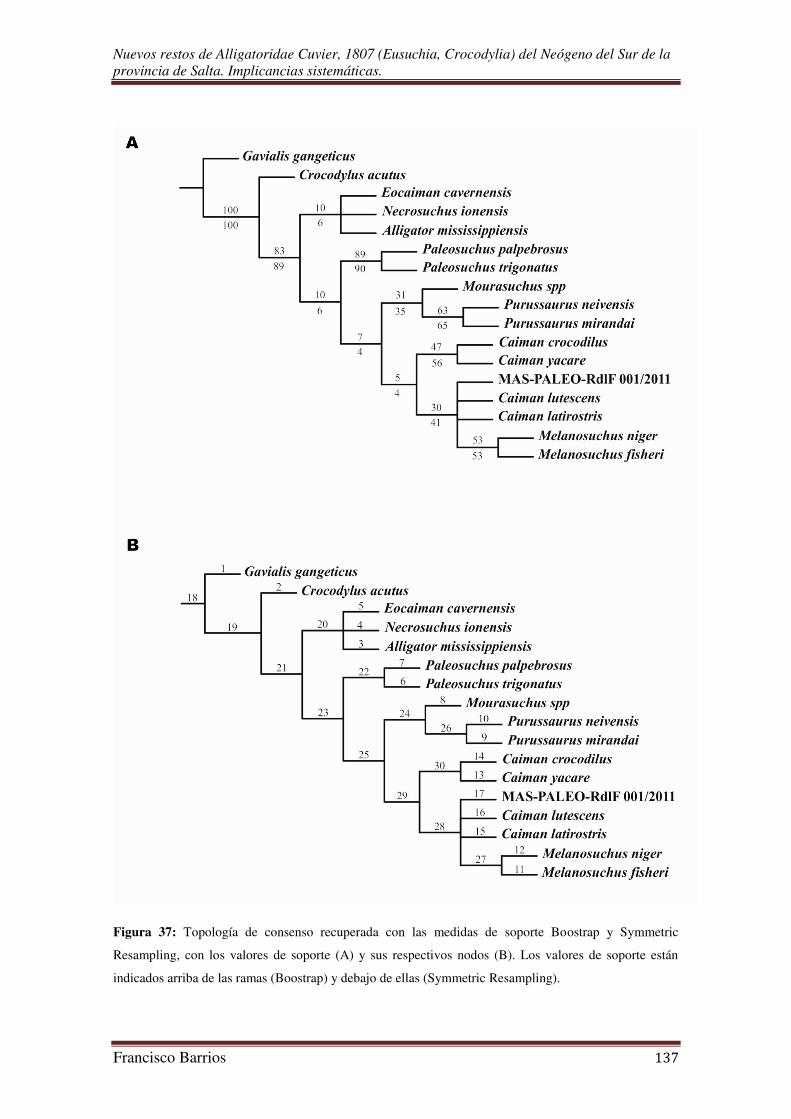
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 137
Figura 37: Topología de consenso recuperada con las medidas de soporte Boostrap y Symmetric
Resampling, con los valores de soporte (A) y sus respectivos nodos (B). Los valores de soporte están
indicados arriba de las ramas (Boostrap) y debajo de ellas (Symmetric Resampling).
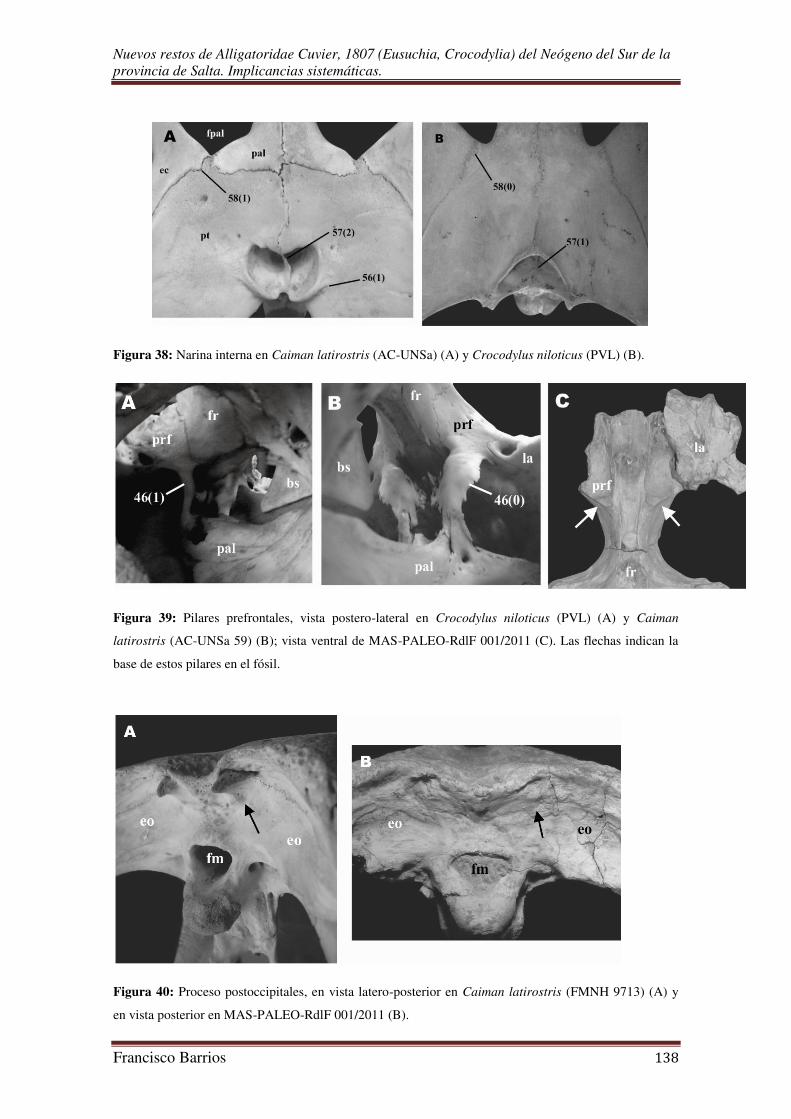
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 138
Figura 38: Narina interna en Caiman latirostris (AC-UNSa) (A) y Crocodylus niloticus (PVL) (B).
Figura 39: Pilares prefrontales, vista postero-lateral en Crocodylus niloticus (PVL) (A) y Caiman
latirostris (AC-UNSa 59) (B); vista ventral de MAS-PALEO-RdlF 001/2011 (C). Las flechas indican la
base de estos pilares en el fósil.
Figura 40: Proceso postoccipitales, en vista latero-posterior en Caiman latirostris (FMNH 9713) (A) y
en vista posterior en MAS-PALEO-RdlF 001/2011 (B).
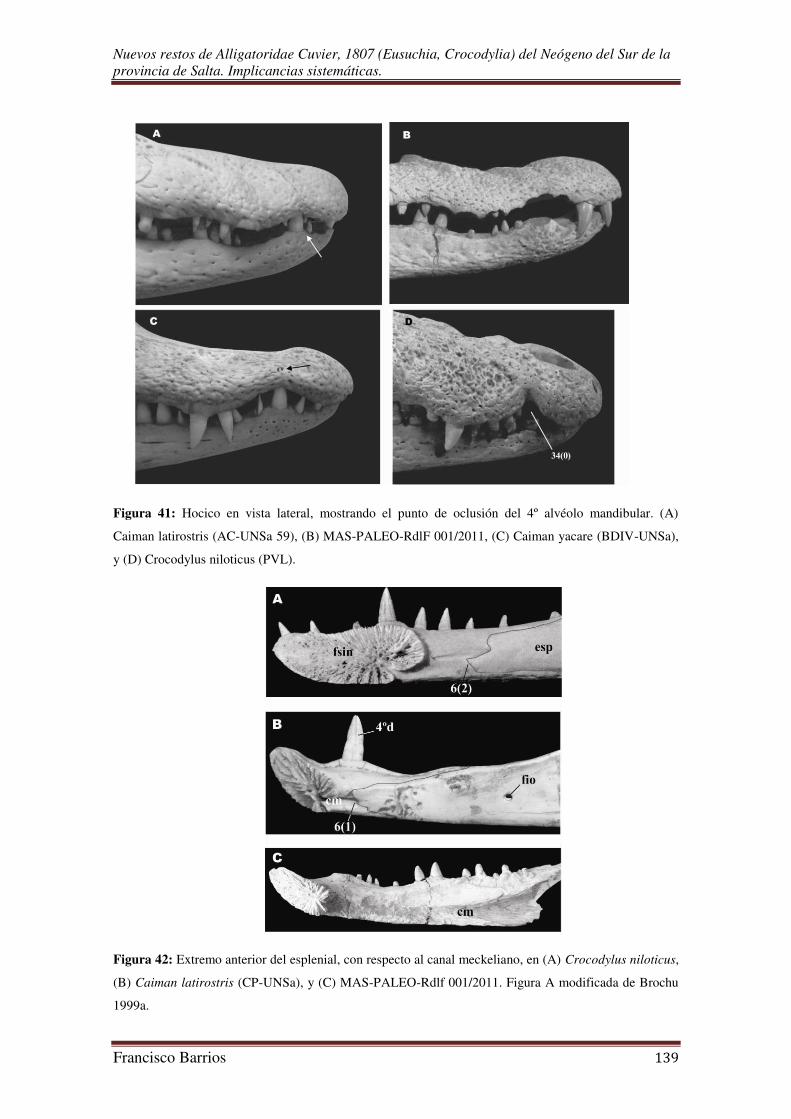
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 139
Figura 41: Hocico en vista lateral, mostrando el punto de oclusión del 4º alvéolo mandibular. (A)
Caiman latirostris (AC-UNSa 59), (B) MAS-PALEO-RdlF 001/2011, (C) Caiman yacare (BDIV-UNSa),
y (D) Crocodylus niloticus (PVL).
Figura 42: Extremo anterior del esplenial, con respecto al canal meckeliano, en (A) Crocodylus niloticus,
(B) Caiman latirostris (CP-UNSa), y (C) MAS-PALEO-Rdlf 001/2011. Figura A modificada de Brochu
1999a.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 140
Figura 43: Topología del foramen intermandibular medio y oral, y moforlogía del coronoides y de la
sutura esp-an, en vista lingual, en Caiman latirostris (AC-UNSa 59) (A) y (FMNH 9713) (F), MAS-
PALEO-RdlF 001/2011 (B), Paleosuchus trigonatus (C), Gavialis gangeticus (D) y Crocodylus niloticus
(PVL) (E). Figuras C y E modificadas de Brochu (1999a) y D de Brochu & Rincón (2004).
Figura 44: Sutura san-an y margen posterior de la fenestra mandibular externa, en vista lateral, en
Caiman latirostris (AC-UNSa 59) (A), MAS-PALEO-RdlF 001/2011 (B), Crocodylus niloticus (PVL)
(C), y Alligator mississippiensis (D). Figura D modificada de Brochu, 1999a.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 141
Figura 45: Prolongación anterior del surangular y fenestra mandibular externa, en vista lateral en Caiman
latirostris (AC-UNSa 59) (A), MAS-PALEO-RdlF 001/2011 (B), y Crocodylus acutus (C). C modificada
de Brochu, 1999a.
Figura 46: Sutura san-an en vista medial y topología del foramen lingual, en Caiman latirostris (26 EC-
IML) (A), MAS-PALEO-RdlF 001/2011 (B), Crocodylus niloticus (PVL) (C), Alligator mississippiensis
(D), y Gavialis gangeticus (E). D y E modificadas de Brochu, 1999a.
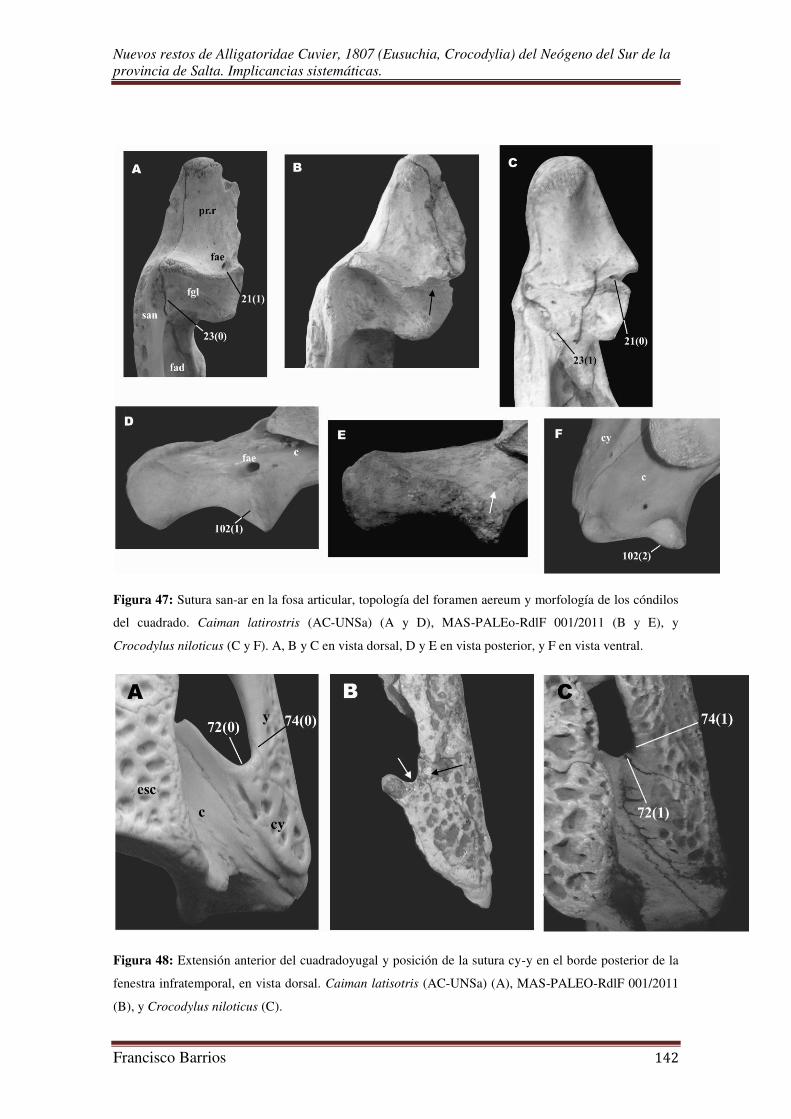
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 142
Figura 47: Sutura san-ar en la fosa articular, topología del foramen aereum y morfología de los cóndilos
del cuadrado. Caiman latirostris (AC-UNSa) (A y D), MAS-PALEo-RdlF 001/2011 (B y E), y
Crocodylus niloticus (C y F). A, B y C en vista dorsal, D y E en vista posterior, y F en vista ventral.
Figura 48: Extensión anterior del cuadradoyugal y posición de la sutura cy-y en el borde posterior de la
fenestra infratemporal, en vista dorsal. Caiman latisotris (AC-UNSa) (A), MAS-PALEO-RdlF 001/2011
(B), y Crocodylus niloticus (C).
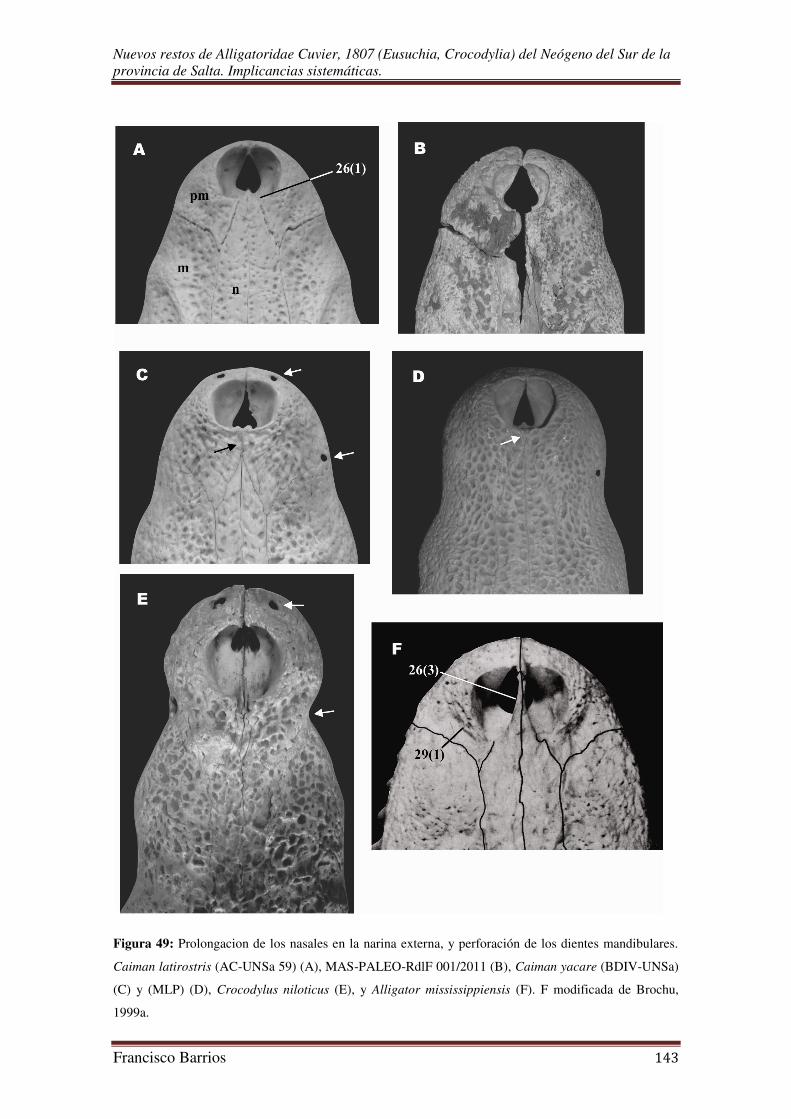
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 143
Figura 49: Prolongacion de los nasales en la narina externa, y perforación de los dientes mandibulares.
Caiman latirostris (AC-UNSa 59) (A), MAS-PALEO-RdlF 001/2011 (B), Caiman yacare (BDIV-UNSa)
(C) y (MLP) (D), Crocodylus niloticus (E), y Alligator mississippiensis (F). F modificada de Brochu,
1999a.
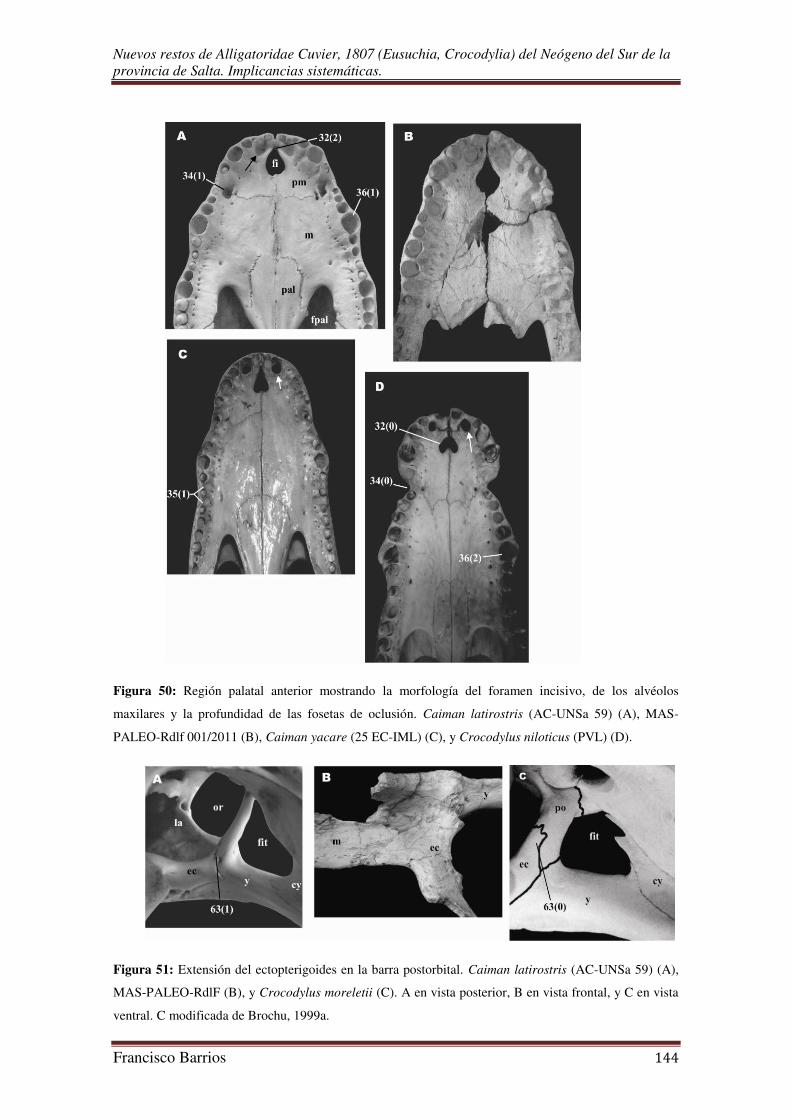
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 144
Figura 50: Región palatal anterior mostrando la morfología del foramen incisivo, de los alvéolos
maxilares y la profundidad de las fosetas de oclusión. Caiman latirostris (AC-UNSa 59) (A), MAS-
PALEO-Rdlf 001/2011 (B), Caiman yacare (25 EC-IML) (C), y Crocodylus niloticus (PVL) (D).
Figura 51: Extensión del ectopterigoides en la barra postorbital. Caiman latirostris (AC-UNSa 59) (A),
MAS-PALEO-RdlF (B), y Crocodylus moreletii (C). A en vista posterior, B en vista frontal, y C en vista
ventral. C modificada de Brochu, 1999a.
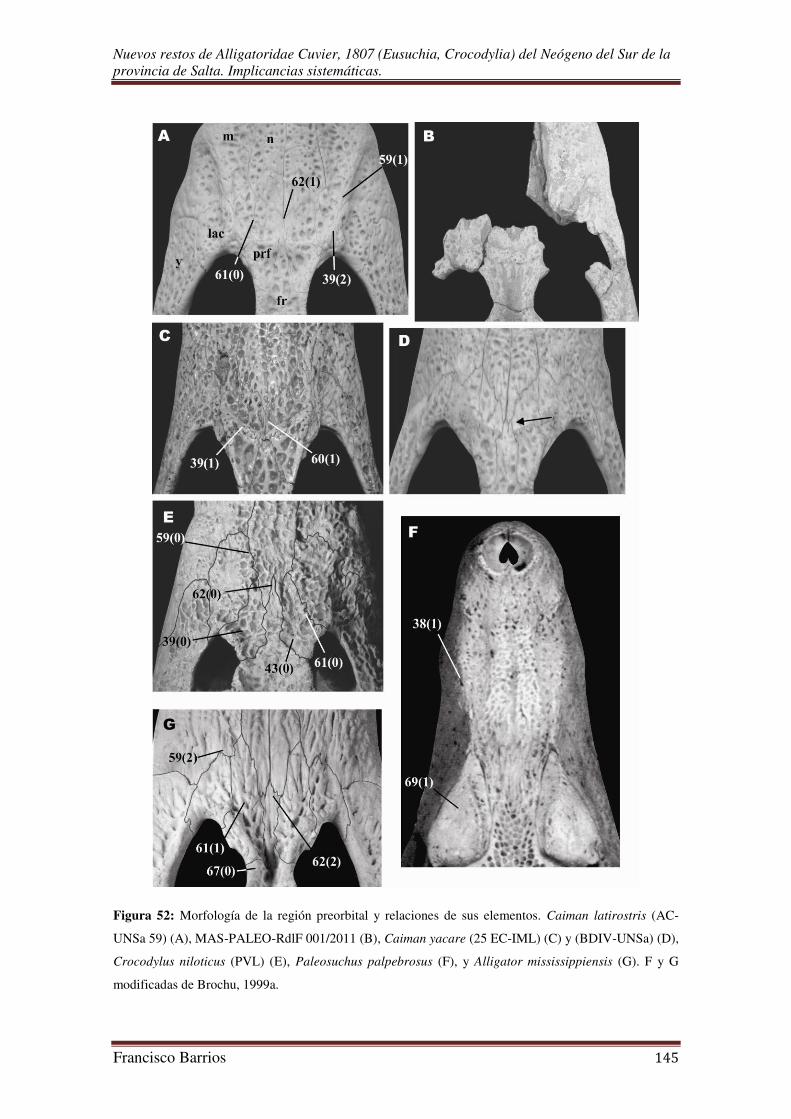
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 145
Figura 52: Morfología de la región preorbital y relaciones de sus elementos. Caiman latirostris (AC-
UNSa 59) (A), MAS-PALEO-RdlF 001/2011 (B), Caiman yacare (25 EC-IML) (C) y (BDIV-UNSa) (D),
Crocodylus niloticus (PVL) (E), Paleosuchus palpebrosus (F), y Alligator mississippiensis (G). F y G
modificadas de Brochu, 1999a.
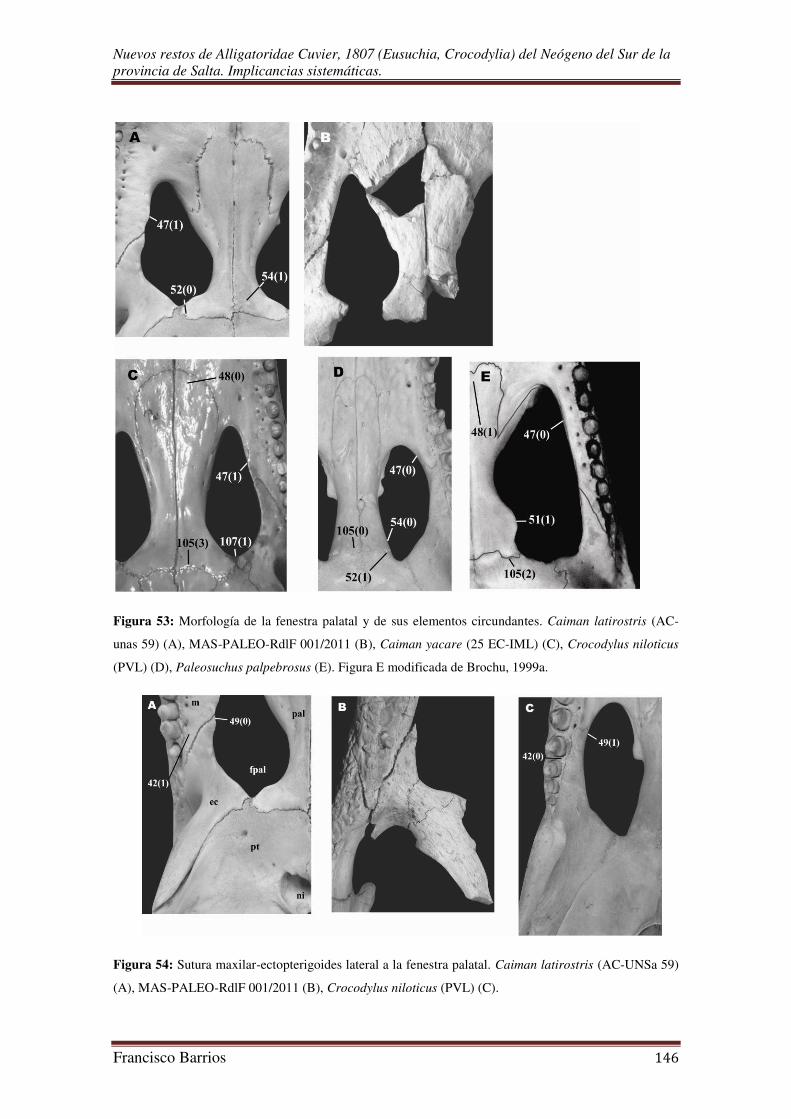
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 146
Figura 53: Morfología de la fenestra palatal y de sus elementos circundantes. Caiman latirostris (AC-
unas 59) (A), MAS-PALEO-RdlF 001/2011 (B), Caiman yacare (25 EC-IML) (C), Crocodylus niloticus
(PVL) (D), Paleosuchus palpebrosus (E). Figura E modificada de Brochu, 1999a.
Figura 54: Sutura maxilar-ectopterigoides lateral a la fenestra palatal. Caiman latirostris (AC-UNSa 59)
(A), MAS-PALEO-RdlF 001/2011 (B), Crocodylus niloticus (PVL) (C).
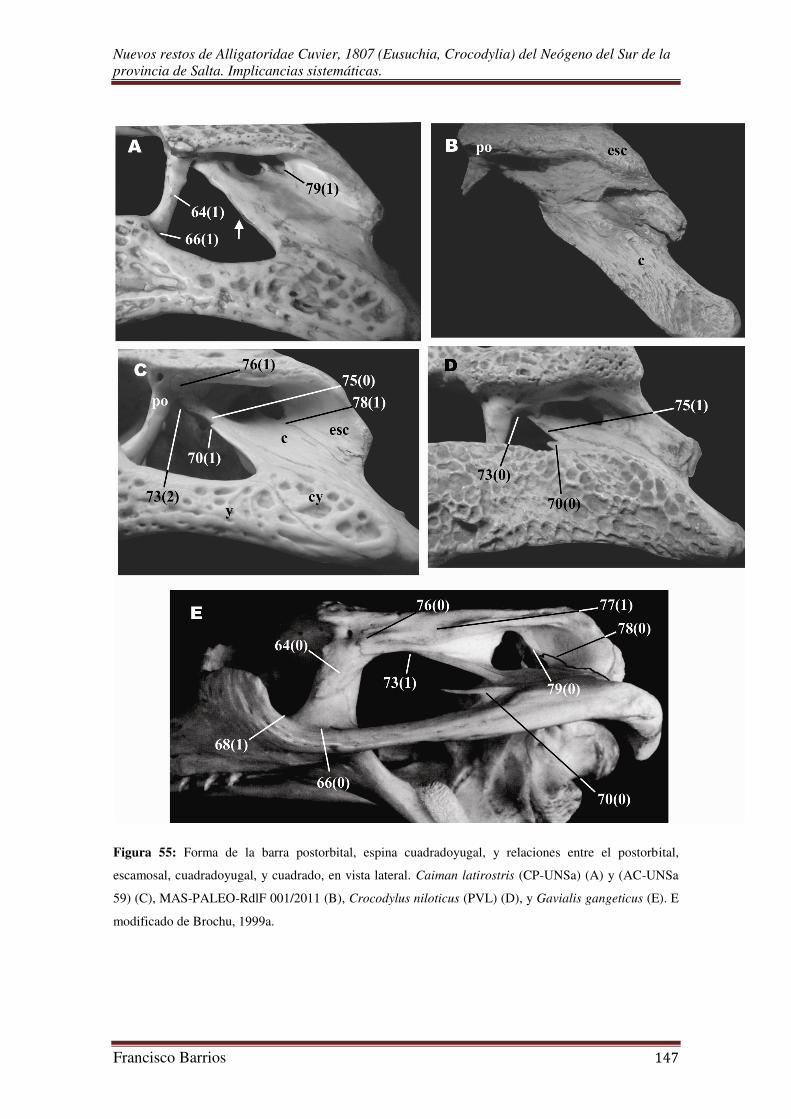
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 147
Figura 55: Forma de la barra postorbital, espina cuadradoyugal, y relaciones entre el postorbital,
escamosal, cuadradoyugal, y cuadrado, en vista lateral. Caiman latirostris (CP-UNSa) (A) y (AC-UNSa
59) (C), MAS-PALEO-RdlF 001/2011 (B), Crocodylus niloticus (PVL) (D), y Gavialis gangeticus (E). E
modificado de Brochu, 1999a.
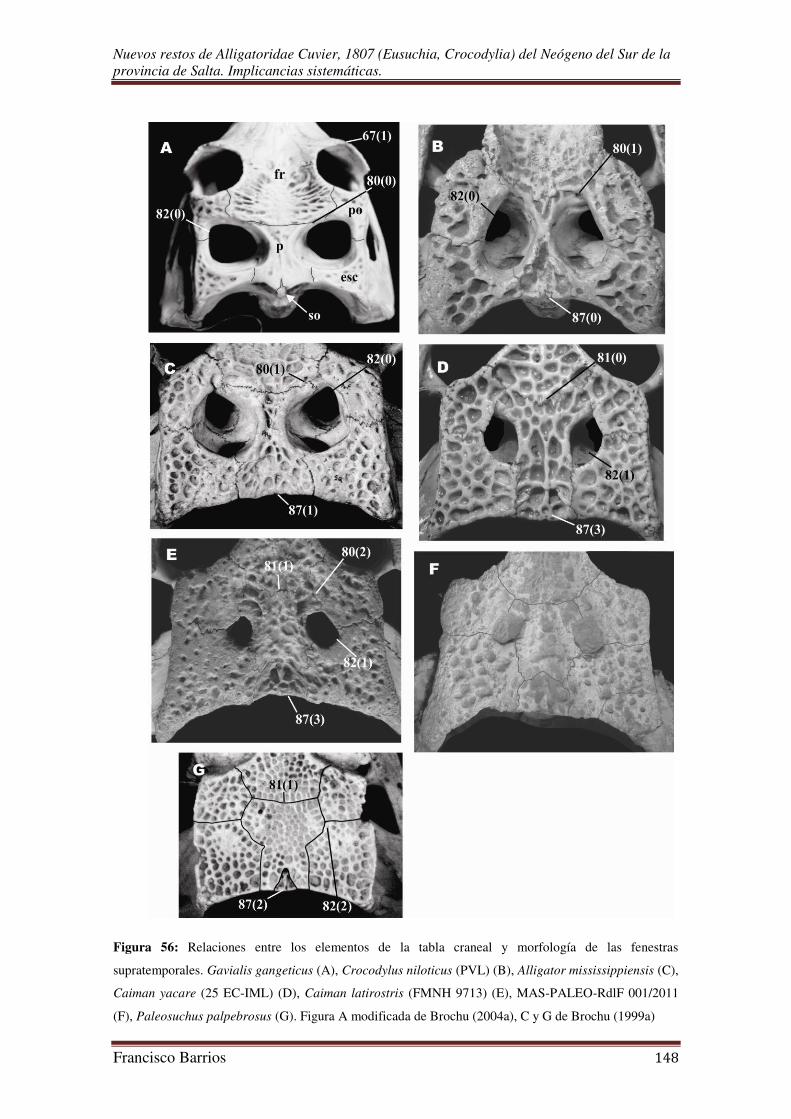
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 148
Figura 56: Relaciones entre los elementos de la tabla craneal y morfología de las fenestras
supratemporales. Gavialis gangeticus (A), Crocodylus niloticus (PVL) (B), Alligator mississippiensis (C),
Caiman yacare (25 EC-IML) (D), Caiman latirostris (FMNH 9713) (E), MAS-PALEO-RdlF 001/2011
(F), Paleosuchus palpebrosus (G). Figura A modificada de Brochu (2004a), C y G de Brochu (1999a)
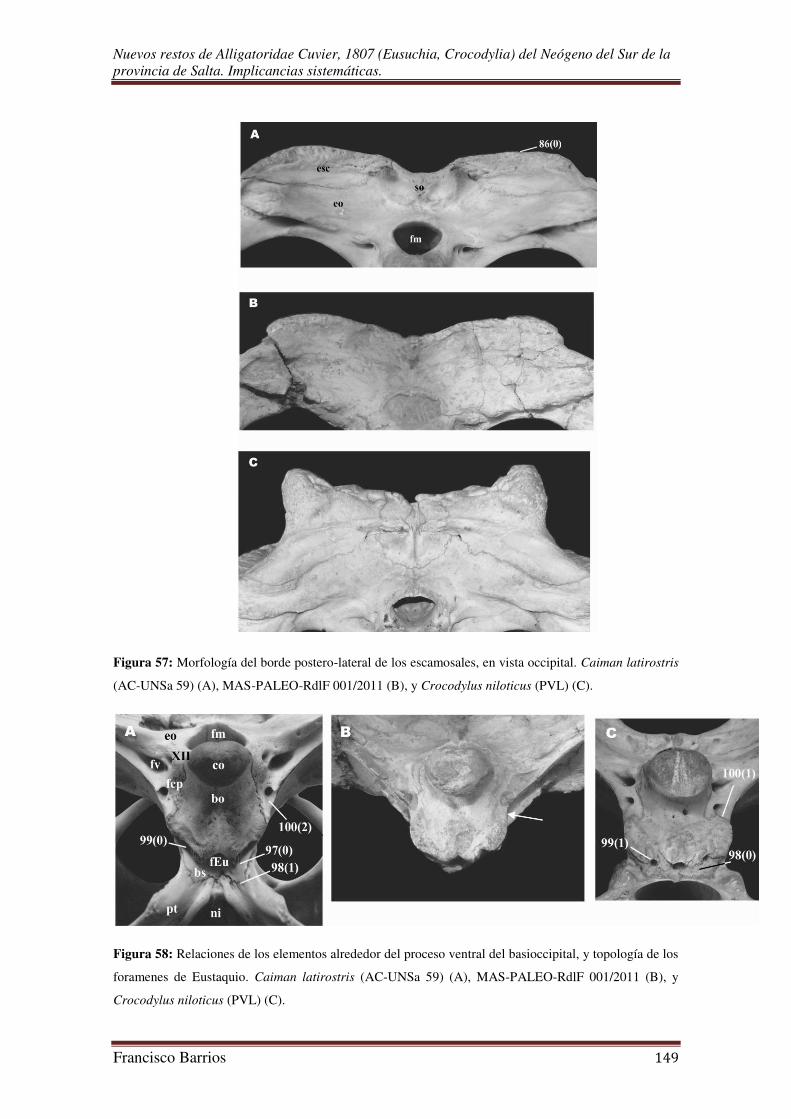
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 149
Figura 57: Morfología del borde postero-lateral de los escamosales, en vista occipital. Caiman latirostris
(AC-UNSa 59) (A), MAS-PALEO-RdlF 001/2011 (B), y Crocodylus niloticus (PVL) (C).
Figura 58: Relaciones de los elementos alrededor del proceso ventral del basioccipital, y topología de los
foramenes de Eustaquio. Caiman latirostris (AC-UNSa 59) (A), MAS-PALEO-RdlF 001/2011 (B), y
Crocodylus niloticus (PVL) (C).
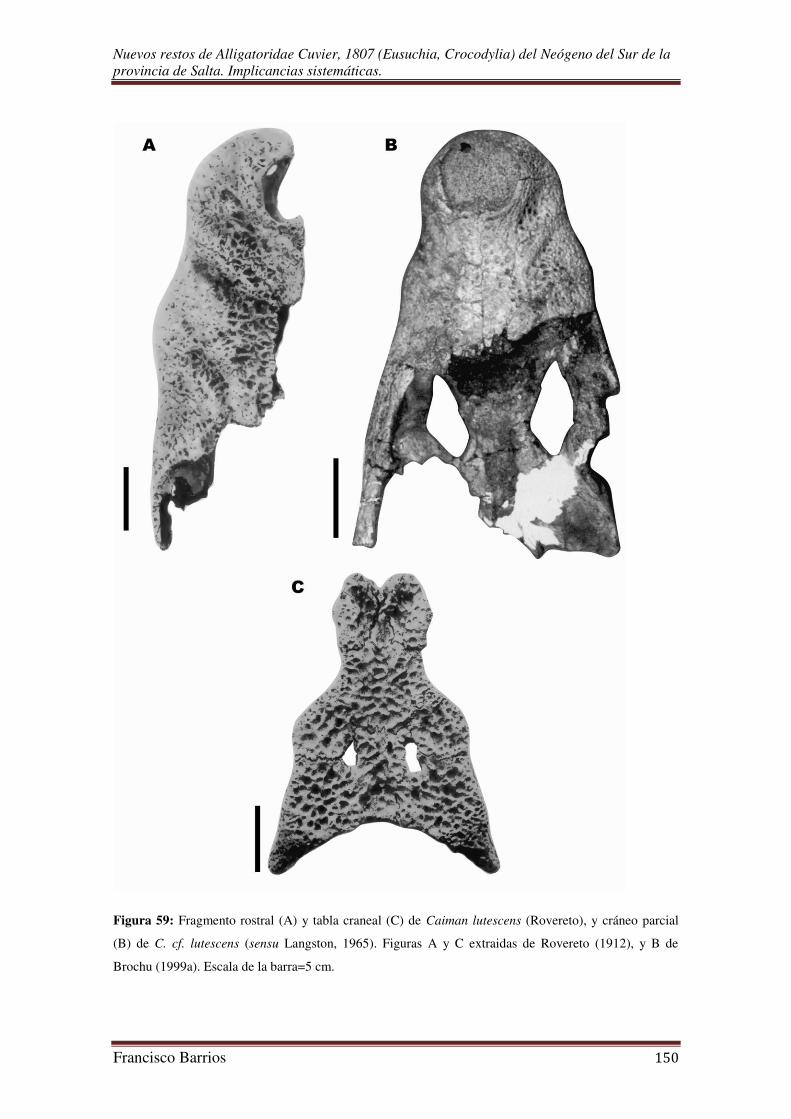
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 150
Figura 59: Fragmento rostral (A) y tabla craneal (C) de Caiman lutescens (Rovereto), y cráneo parcial
(B) de C. cf. lutescens (sensu Langston, 1965). Figuras A y C extraidas de Rovereto (1912), y B de
Brochu (1999a). Escala de la barra=5 cm.
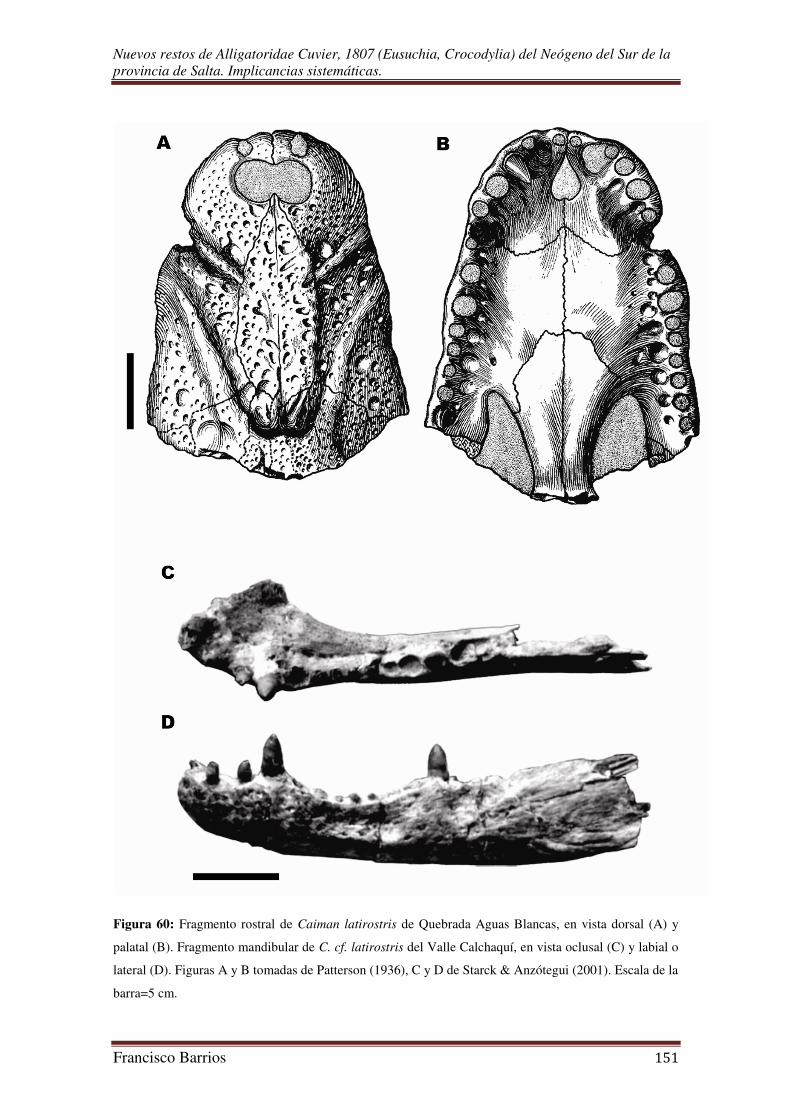
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 151
Figura 60: Fragmento rostral de Caiman latirostris de Quebrada Aguas Blancas, en vista dorsal (A) y
palatal (B). Fragmento mandibular de C. cf. latirostris del Valle Calchaquí, en vista oclusal (C) y labial o
lateral (D). Figuras A y B tomadas de Patterson (1936), C y D de Starck & Anzótegui (2001). Escala de la
barra=5 cm.
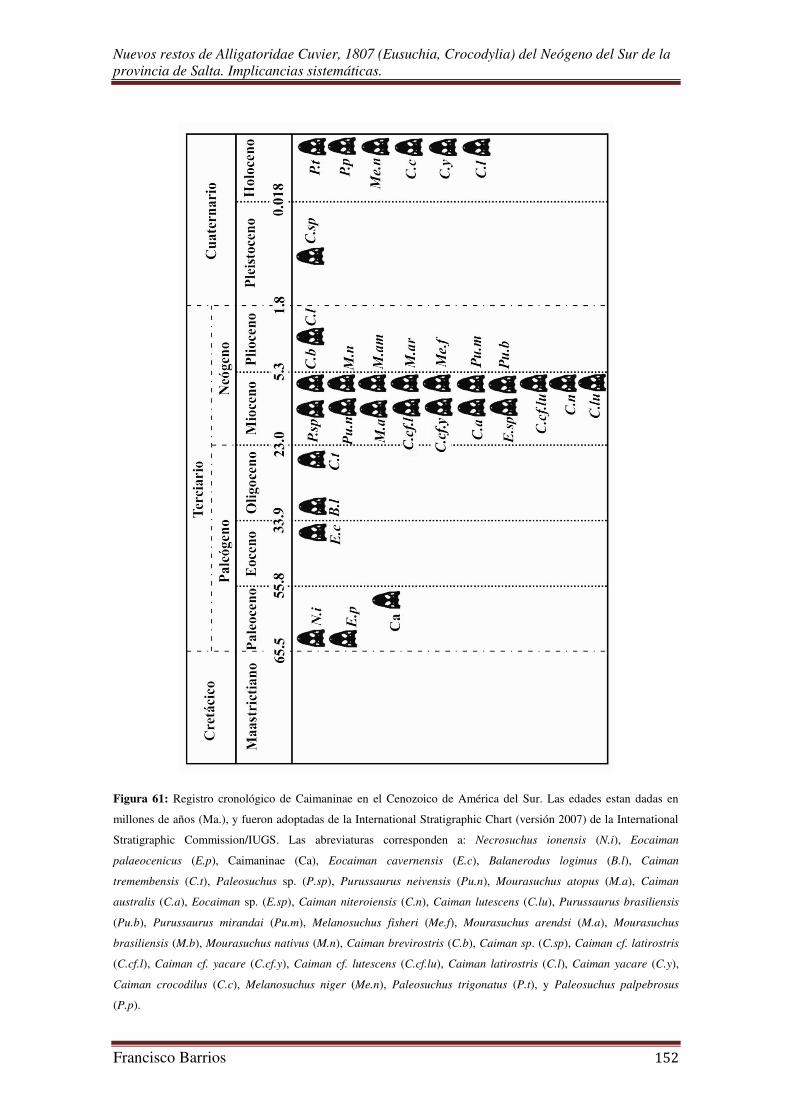
Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 152
Figura 61: Registro cronológico de Caimaninae en el Cenozoico de América del Sur. Las edades estan dadas en
millones de años (Ma.), y fueron adoptadas de la International Stratigraphic Chart (versión 2007) de la International
Stratigraphic Commission/IUGS. Las abreviaturas corresponden a: Necrosuchus ionensis (N.i), Eocaiman
palaeocenicus (E.p), Caimaninae (Ca), Eocaiman cavernensis (E.c), Balanerodus logimus (B.l), Caiman
tremembensis (C.t), Paleosuchus sp. (P.sp), Purussaurus neivensis (Pu.n), Mourasuchus atopus (M.a), Caiman
australis (C.a), Eocaiman sp. (E.sp), Caiman niteroiensis (C.n), Caiman lutescens (C.lu), Purussaurus brasiliensis
(Pu.b), Purussaurus mirandai (Pu.m), Melanosuchus fisheri (Me.f), Mourasuchus arendsi (M.a), Mourasuchus
brasiliensis (M.b), Mourasuchus nativus (M.n), Caiman brevirostris (C.b), Caiman sp. (C.sp), Caiman cf. latirostris
(C.cf.l), Caiman cf. yacare (C.cf.y), Caiman cf. lutescens (C.cf.lu), Caiman latirostris (C.l), Caiman yacare (C.y),
Caiman crocodilus (C.c), Melanosuchus niger (Me.n), Paleosuchus trigonatus (P.t), y Paleosuchus palpebrosus
(P.p).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 153
APÉNDICE II
Abreviaturas anatómicas:
an: angular ar: articular bpo: barra postorbital bo: basioccipital bs: basiesfenoides c: cuadrado cec: cóndilo externo del cuadrado cm: canal de Meckel cmc: cóndilo medio del cuadrado co: cóndilo occipital cor: coronoides crc: cresta del cuadrado crpo: cresta preorbital cy: cuadradoyugal d: dentario ec: ectopterigoides eo: exoccipital esc: escamosal esp: esplenial fad: fosa del aductor mandibular fae: foramen aéreo fcp: foramen carotideo posterior o lateral fdm 1º: foseta para el 1º diente mandibular fdm 4º: foseta para el 4º diente mandibular fEu: foramen de Eustaquio medio fEul: foramen de Eustaquio lateral fgl: fosa glenoidea fi: foramen incisivo fic: foramen intermandibular caudal fim: foramen intermandibular medio fio: foramen intermandibular oral fit: fenestra infratemporal fm: foramen magnum fme: fenestra mandibular externa fo/V: foramen oval o del nervio craneal V fpt: fenestra post-temporal fr: frontal fsin: faceta sinfiseal fst: fenestra supratemporal fv: foramen vago la: lacrimal
ls: lateroesfenoides m: maxilar n: nasal ne: narina externa ni: narina interna or: órbita p: parietal pal: palatinos pcc: pasaje cranio-cuadrado pm: premaxilar po: postorbital p.po: proceso paraoccipital p.pto: proceso postoccipital prf: prefrontal pr.r: proceso retroarticular pt: pterigoides san: surangular sm: sínfisis mandibular so: supraoccipital XII: foramen del nervio craneal XII y: yugal 4ºd: cuarto diente del dentario 4º al: cuarto alvéolo

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 154
APÉNDICE III
Lista de Caracteres:
1: Sínfisis dentaria extendiéndose por detrás del 8º alvéolo (0), entre el 6º y 8º alvéolo (1), entre el 4º y 5º alvéolo (2), o muy reducida y restringida entre los dos primeros alvéolos (3). (Modificado de Brochu, 2004a, carácter 166.)
2: Dentario entre 4º y 10º alvéolo, lineal (0) o curvado (1). (Modificado de Brochu, 1999, carácter 68.)
3: Alvéolos mayores del dentario detrás del 4º, sin diferenciación (0), 11º o 12º (1), o 13º o 14º (2). (Modificado de Brochu, 2004a, carácter 167.)
4: Esplenial con perforación anterior para la rama mandibular del nervio craneal V, presente (0) o ausente (1). (Brochu, 1999, carácter 41.)
5: Rama mandibular del nervio craneal V sale por el esplenial en una única perforación anterior (0), en una única perforación posterior (1), o por doble perforación posterior (2). (Brochu, 1999, carácter 42.). Figura 43.
6: Participación del esplenial en la sínfisis mandibular; sínfisis esplenial profunda, más larga que cinco alvéolos y formando una amplia “V” (0), esplenial excluido de la sínfisis mandibular y con su extremo anterior dorsal al canal de Meckel (1), o esplenial excluido de la sínfisis mandibular con su extremo anterior ventral al canal de Meckel (2). (Modificado de Brochu, 1999, 43.). Figura 42.
7: Coronoides en la madurez limita la mitad posterior del foramen intermandibular medio (0), lo rodea completamente (1) o lo obstruye (2). (Brochu, 1999, carácter 46.). Figura 43.
8: Borde superior del coronoides muy inclinado anteriormente (0), o casi horizontal (1). (Brochu, 1999, carácter 54.). Figura 43.
9: Proceso inferior del coronoides se dobla mayormente sobre la superficie interna de la fosa meckeliana (0), o permanece principalmente sobre la superficie media de la mandíbula (1). (Brochu, 1999, carácter 55.)
10: Coronoides sin perforación (0), o con perforación posterior para el foramen intermandibular medio (1). (Brochu, 1999, carácter 56.). Figura 43.
11: Proceso esplenial entre angular y coronoides, presente (0), o ausente (1). (Brochu, 1999, carácter 59.)
12: Sutura angular-surangular en la madurez contacta la fenestra mandibular externa en el ángulo posterior (0), o en el margen ventral (1). (Brochu, 1999, carácter 47.). Figura 44.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 155
13: Procesos anteriores del surangular desiguales (0), o subiguales a iguales (1). (Brochu, 1999, carácter 48.). Figura 45.
14: Prolongación del surangular limitando lingualmente a por lo menos uno de los alvéolos de la hilera de dientes, presente (0), o ausente (1). (Brochu, 1999, carácter 61.)
15: Fenestra mandibular externa pequeña (0), grande, menos de la mitad del foramen intermandibular caudal visible lateralmente (1), muy grande, la mayor parte del foramen intermandibular caudal visible lateralmente (2). (Modificado de Brochu, 1999, carácter 62.). Figura 44.
16: Angular se extiende dorsalmente más allá del extremo anterior del foramen intermandibular caudal, punta anterior aguda (0), o no se extiende más allá del extremo anterior del foramen intermandibular caudal, punta anterior roma (1). (Brochu, 1999, carácter 66.). Figura 43.
17: Lingualmente la sutura surangular-angular se une a la punta del articular, ventralmente (0), o dorsalmente (1). (Brochu, 1999, carácter 67.). Figura 46.
18: Surangular continúa hasta la punta dorsal de la pared lateral de la fosa glenoidea (0), o truncado y no continúa dorsalmente (1). (Brochu, 1999, carácter 106.). Figura 44.
19: Sutura surangular-articular simple (0), articular con lamina anterior dorsal al foramen lingual (1), articular con lamina anterior ventral al foramen lingual (2), o con laminas arriba y abajo del foramen (3). (Modificado de Brochu, 1999, carácter 44.). Figura 46.
20: Foramen lingual para la arteria articular y nervio alveolar perfora por completo el surangular (0), o la sutura surangular-angular (1). (Modificado de Brochu, 1999, carácter 45.). Figura 46.
21: Foramen aéreo en el extremo del margen lingual del proceso retroarticular (0), o dentro del margen (1). (Brochu, 1999, carácter 49.). Figura 47.
22: Surangular finaliza anterior (0) o se extiende hasta el extremo posterior del proceso retroarticular (1). (Modificado de Brochu, 1999, carácter 51.). Figura 44.
23: Sutura surangular-articular dentro de la fosa glenoidea, orientada anteroposteriormente (0), o muy curvada lateralmente (1). (Brochu, 1999, carácter 162.). Figura 47.
24: Surco entre articular y surangular (0), o articular nivelado contra el surangular (1). (Brochu, 1999, carácter 60.)
25: Dientes y alvéolos del maxilar y/o dentario de sección circular (0), o dientes posteriores comprimidos lateralmente (1). (Brochu, 2004a, carácter 165; estado 2 eliminado.)

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 156
26: Nasales excluidos de la narina externa, nasales y premaxilares sin contacto (0), o en contacto (1); nasales en contactos con la narina externa, pero sin dividirla (2); narina externa dividida por los nasales (3). (Modificado de Brochu, 1999, carácter 95.). Figura 46.
27: Narina externa circular (0), o más ancha que larga (1). (Modificado de Brochu, 1999, carácter 161; estado 2 eliminado.)
28: Narina externa de machos adultos reproductivos similar al de las hembras (0), o desarrollan una excresencia ósea (“ghara”) (1). (Brochu en prensa, carácter 84.)
29: Superficie premaxilar lateral a la narina externa, lisa (0), o con hendidura profunda (1). (Brochu, 1999, carácter 142.)
30: Premaxilar con cinco (0), o cuatro dientes (1). (Brochu, 1999, carácter 124.)
31: Foramen incisivo, muy pequeño y subcircular (0), pequeño y en forma de “corazón” o de “lagrima” (1), grande y oval (2), o grande y circular (3). (Modificado de Brochu, 1999, carácter 124.)
32: Foramen incisivo situado lejos de la hilera de alvéolos premaxilares, a nivel del 2º o 3º alvéolo (0), apoyado sobre la hilera (1), o proyectado entre el 1º alvéolo premaxilar (2). (Brochu, 1999, carácter 153.). Figura 50.
33: Proceso premaxilar dorsal corto, no se extiende más allá del 3º alvéolo maxilar (0), o largo, alcanzando más allá del 3º alvéolo maxilar (1). (Brochu, 1999, carácter 145.)
34: 4º diente mandibular ocluye temprano en la ontogenia entre el premaxilar y maxilar, en una hendidura (0), o en una foseta (1). (Brochu, 1999, carácter 77.). Figuras 41 y 50.
35: Dientes mandibulares ocluyen lineal a la hilera de dientes maxilares (0), con foseta de oclusión entre 5º y 7º diente maxilar y los demás lingualmente (1), o todos los dientes mandibulares ocluyen lingualmente (2). (Modificado de Brochu, 1999, carácter 78.). Figura 50.
36: Alvéolos maxilares homodontos (0), el alvéolo maxilar más grande es el 4º (1), o el 5º (2). (Modificado de Brochu, 1999, carácter 89.). Figura 50.
37: Superficie del rostro con protuberancia dorsal media (0), curvada suavemente (1), curvada profundamente (2). (Modificado de Brochu, 1999, carácter 101.)
38: Canto rostral ausente o moderado (0), prominente (1), o formando una fosa nasal (2). (Modificado de Brochu, 1999, carácter 143.). Figura 52.
39: Cresta preorbital ausente (0), moderada (1), o prominente (2) a la madurez. (Modificado de Brochu, 1999, carácter 144.). Figura 52.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 157
40: Vómer totalmente oculto por el premaxilar y maxilar (0), o expuesto en el paladar en la sutura premaxilar-maxilar (1). (Brochu, 1999, carácter 125.). Figura 27B.
41: Foramen yugal medio pequeño (0), o muy grande (1). (Brochu, 1999, carácter 120.)
42: Ectopterigoides apoyado sobre la hilera de alvéolos maxilares (0), o separado por el maxilar (1). (Brochu, 1999, carácter 91.). Figura 54.
43: Superficie dorsal del prefrontal adyacente al borde orbital; lisa (0), o con un proceso protuberante discreto (1). (Brochu en prensa, carácter 107.). Figura 52.
44: Mitad dorsal del pilar prefrontal estrecho (0), o expandido anteroposteriormente (1). (Brochu, 1999, carácter 137.)
45: Proceso medio del pilar prefrontal expandido dorsoventralmente (0), o anteroposteriormente (1). (Brochu, 1999, carácter 136.)
46: Proceso medio del pilar prefrontal, amplio (0), o constreñido (1) en la base. (Brochu, 1999, carácter 138.). Figura 38.
47: Maxilar con margen medio adyacente a la fenestra palatal, lineal (0), o lleva una amplia plataforma extendiéndose en la fenestra y haciendo el margen lateral cóncavo (1). (Modificado de Brochu, 1999, carácter 105.). Figura 53.
48: Superficie anterior del proceso palatino de contorno, redondeado o puntiagudo (0), hendido anteriormente (1), o rectangular (2). (Modificado de Brochu, 1999, carácter 108.). Figura 53.
49: Proceso ectopterigoides anterior se afina a una punta (0), o bifurcado (1). (Brochu, 1999, carácter 109.). Figura 54.
50: Proceso palatino por lo general amplio anteriormente (0), o en forma de una delgada cuña (1). (Brochu, 1999, carácter 118.)
51: Márgenes laterales de los palatinos anteriormente lisos (0), o con proyección en las fenestras palatales (1). (Brochu, 1999, carácter 94.). Figura 53.
52: Sutura palatino-pterigoides cerca (0), o alejada (1) del ángulo posterior de la fenestra palatal. (Brochu, 1999, carácter 85.). Figura 53.
53: Rama pterigoidea del ectopterigoides; recta, margen posterolateral de la fenestra palatal lineal (0), o rama curvada y margen de la fenestra cóncavo (1). (Reformulado de Brochu, 1999, carácter 88.)
54: Margen lateral de los palatinos posteriormente paralelos (0), o ensanchados formando una plataforma (1). (Brochu, 1999, carácter 90.). Figura 53.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 158
55: Narina interna en la madurez proyectada posteroventralmente (0), o anteroventralmente (1). (Brochu, 1999, carácter 72.)
56: Borde posterior de la narina interna con hendidura profunda ausente (0), o presente (1). (Brochu, 1999, carácter 107.). Figura 38.
57: Narina interna, no dividida (0), con tabique por dentro de la narina (1), o con tabique sobresaliendo de la narina (2). (Brochu, 1999, carácter 152.). Figura 38.
58: Pliegue ectopterigoides-pterigoides desaparece durante la ontogenia (0), o permanece durante toda la ontogenia (1). (Brochu, 1999, carácter 116.). Figura 38.
59: Lacrimal en contacto con el nasal, con proceso posterior del maxilar ausente (0), o presente dentro del lacrimal (1); sin contacto entre lacrimal y nasal, con proceso posterior del maxilar entre lacrimal y prefrontal (2), o con proceso dentro del lacrimal (3). (Modificado de Brochu, 1999, carácter 93.). Figura 52.
60: Prefrontales separados por el frontal y nasales (0), o unidos medialmente (1). (Brochu, 1999, carácter 100.). Figura 60.
61: Lacrimal más largo que el prefrontal (0), o prefrontal más largo que el lacrimal (1). (Brochu, 1999, carácter 117, estado 2 eliminado.). Figura 52.
62: Punta anterior del frontal, aguda y en gran contacto con los nasales (0), aguda y con contacto leve o ausente con lo nasales (1), amplia y en complejo contacto con los nasales (2). (Modificado de Brochu en prensa, carácter 131.). Figura 52.
63: Ectopterigoides se extiende a lo largo de la superficie media de la barra postorbital (0), o se detiene abruptamente ventral a la barra postorbital (1). (Brochu, 1999, carácter 133.). Figura 51.
64: Barra postorbital gruesa (0), o delgada (1). (Brochu, 1999, carácter 70.). Figura 55.
65: Barra postorbital con proceso prominente, dorsoventralmente amplio, y divisible en dos espinas (0), o con proceso corto y generalmente no prominente (1). (Brochu, 1999, carácter 134.)
66: Margen ventral de la barra postorbital a nivel (0), o por dentro de superficie lateral del yugal (1). (Brochu, 1999, carácter 146.). Figura 55.
67: Margen dorsal de las órbitas elevado (0), o todo el margen orbital sobre elevado (1). (Modificado de Brochu, 1999, carácter 103.). Figuras 52 y 56.
68: Margen ventral de la órbitas, circular (0), o con prominente hendidura (1). (Brochu, 1999, carácter 139.). Figura 55.
69: Palpebrales formados de una única osificación (0), o de múltiples osificaciones (1). (Brochu, 1999, carácter 96.)

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 159
70: Espina cuadradoyugal a la madurez; prominente (0), o muy reducida o ausente (1). (Brochu, 1999, carácter 69.). Figura 55.
71: Espina cuadradoyugal baja, cerca del ángulo posterior de la fenestra infratemporal (0), o alta, entre los ángulos posterior y superior de la fenestra (1). (Brochu, 1999, carácter 114.)
72: Cuadradoyugal forma el ángulo posterior de la fenestra infratemporal (0), o la sutura cuadradoyugal-yugal se encuentra en el ángulo posterior de la fenestra (1). (Brochu, 1999, carácter 75, estado 1 eliminado.). Figura 48.
73: Postorbital en el ángulo dorsal de la fenestra infratemporal medialmente sin contacto con el cuadrado ni el cuadradoyugal (0), en contacto sólo con el cuadradoyugal (1), o en contacto tanto con el cuadrado como con el cuadradoyugal (2). (Brochu, 1999, carácter 76, estado 3 eliminado.). Figura 55.
74: Cuadradoyugal con proceso anterior a lo largo de la barra temporal inferior, prolongado (0), o moderado a ausente (1). (Brochu, 1999, carácter 83.). Figura 48.
75: Cuadradoyugal se extiende al ángulo superior de la fenestra infratemporal, cuadrado no participa en la fenestra (0), o no se extiende al ángulo y el cuadrado participa en la fenestra (1). (Brochu, 1999, carácter 80.). Figura 55.
76: Sutura postorbital-escamosal ventral a la tabla craneal, orientada ventralmente (0), o pasa medialmente (1). (Brochu, 1999, carácter 163.). Figura 55.
77: Bordes dorsal y ventral de la ranura escamosal para la musculatura de la válvula del oído externo, paralelos (0), o ranura escamosal ensanchada anteriormente (1). (Brochu, 1999, carácter 84.9. Figura 55.
78: Sutura escamosal-cuadrado en al meato auditivo externo, se extiende dorsalmente a lo largo del margen posterior (0), o solo se extiende a la esquina posteroventral (1). (Brochu, 1999, carácter 132.). Figura 55.
79: Margen posterior de la abertura ótica, lisa (0), o curvada (1). (Brochu, 1999, carácter 102.). Figura 55.
80: Sutura frontoparietal hace una modesta entrada en la fenestra supratemporal a la madurez, sin contacto postorbital y parietal en la tabla craneal (0); sutura frontoparietal sobre la tabla craneal y sin entrar en la fenestra, con leve contacto postorbital-parietal (1), o en amplio contacto postorbital-parietal sobre la tabla craneal (2). (Modificado de Brochu, 1999, carácter 81.). Figura 56.
81: Sutura frontoparietal entre las fenestra supratemporal, cóncavoconvexa (0), o lineal (1). (Brochu, 1999, carácter 86.). Figura 56.

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 160
82: Fenestra supratemporal a la madurez, con fosa, huesos dérmicos del techo craneal no sobresalen el borde (0), sobresalen el borde de la fenestra reduciendo su luz (1), o fenestra supratemporal cerrada (2). (Brochu, 1999, carácter 87.). Figura 56.
83: Pared media parietal de la fenestra supratemporal imperforada (0), o con forámenes (1). (Brochu, 1999, carácter 104.)
84: Parietal y escamosal ampliamente separados por el cuadrado en la pared posterior de la fenestra supratemporal (0), o unidos a lo largo de la pared posteior de la fenestra (1). (Brochu, 1999, carácter 131, estado 1 eliminado.)
85: Superficie de la tabla craneal a la madurez, inclinada del eje sagital (0), o plana (1). (Brochu, 1999, carácter 123.)
86: Margen posterolateral del escamosal, horizontal (0), o elevado formando un cuerno discreto (1). (Brochu en prensa, carácter 157.). Figura 57.
87: Exposición supraoccipital sobre la tabla craneal dorsal, pequeña (0), ausente (1), grande, parietal no excluido del borde posterior de la tabla (2), o muy grande, parietal excluido del borde posterior (3). (Modificado de Brochu, 1999, carácter 82.). Figura 56.
88: Pared anterior de la caja craneal lateral al rostro del basiesfenoides, con surco (0), lisa y sin surco (1). (Brochu, 1999, carácter 122.)
89: Basiesfenoides expuesto ampliamente sobre la pared de la caja craneal anterior al foramen trigémino, ausente (0), o presente (1). (Brochu, 1999, carácter 129.)
90: Amplia exposición del proótico sobre la pared externa de la caja craneal (0), o en gran medida oculto externamente por el cuadrado y el lateroesfenoides (1). (Brochu, 1999, carácter 74.)
91: Puente lateroesfenoides compuesto en su totalidad del lateroesfenoides (0), o con proceso ascendente o palatino (1). (Brochu, 1999, carácter 115.)
92: Proceso capitado del lateroesfenoides orientado hacia la línea media, lateralmente (0), o anteroposteriormente (1). (Brochu, 1999, carácter 130.)
93: Parietal con hueco comunicando con el sistema neumático (0), o sólido, sin hueco (1). (Brochu, 1999, carácter 154.)
94: Significativo proceso cuadrado ventral sobre la pared lateral de la caja craneal (0), o sutura cuadrado-pterigoides lineal desde la exposición basiesfenoides al foramen trigémino (1). (Brochu, 1999, carácter 127.)
95: Foramen carotideo lateral o posterior se abre a la madurez, lateral (0), o dorsal (1), al basiesfenoides. (Brochu, 1999, carácter 128.)

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 161
96: Proceso pterigoides, proyectado posteriormente (0), pequeño y proyectado posteroventralmente (1), o alto y prominente (2). (Modificado de Brochu, 1999, carácter 98.)
97: Basiesfenoides ventral al basioccipital, anteroposteriormente delgado (0), o ancho (1). (Brochu, 1999, carácter 113.). Figura 58.
98: Basiesfenoides no ampliamente expuesto ventral al basioccipital a la madurez, pterigoides corto ventral al abertura de Eustaquio media (0), o basiesfenoides expuesto como una amplia lamina ventral al basioccipital a la madurez, pterigoides alto ventral a la abertura de Eustaquio media (1). (Brochu, 1999, carácter 119.). Figura 58.
99: Canales de Eustaquio laterales se abren dorsal (0), o lateral (1) al canal de Eustaquio medio. (Brochu, 1999, carácter 46.). Figura 58.
100: Exoccipitales finalizan dorsal a la tuberosidad del basioccipital (0), envía un robusto proceso ventral y participa en la tuberosidad (1), o envía un delgado proceso ventral a la tuberosidad basioccipital (2). (Brochu, 1999, carácter 151.)
101: Foramen aéreo del cuadrado en el ángulo mediodorsal (0), o superficie dorsal del cuadrado (1). (Brochu, 1999, carácter 121.)
102: Cuadrado con pequeño hemicóndilo medio reflejado ventralmente (0), con pequeño hemicóndilo medio y con ranura dorsal para el foramen aerum (1), o con hemicóndilo medio expandido (1). (Brochu, 1999, carácter 112, estado 3 eliminado.). Figura 47.
103: Dentario a nivel del 1º y 4º diente más bajo que a nivel del 11º-12º (0), dentario a nivel del 1º y 4º diente igual o más alto que a nivel del 11º-12º (1). (Bona, 2007).
104: Extremo anterior del frontal, largo y alcanzando o pasando el borde anterior de las órbitas (0), corto y no alcanza el borde anterior de las órbitas (1).
105: Sutura palatino-pterigoidea, recta (0), cóncavoconvexa (1), levemente ondulada (2), o profundamente ondulada (3). Figura 53.
106: Festonamiento lateral del hocico, ausente (0), ligero (1), marcado (2).
107: Pterigoides participa del borde posterior de la fenestra palatal (0), o excluido de la fenestra palatal (1). Figura 53.
108: Borde posterior de la tabla craneal transversalmente recto a levemente cóncavo (0), o profundamente cóncavo (1).
109: Cresta preorbital uniendo las órbitas anteriormente, ausente (0), o presente (1).

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 162
APÉNDICE IV
Matriz de Estados de Caracteres:
Gavialis gangeticus 0000000000000000000000000001000010000000000000000101000000000000001100001000100010000000000000001000001000000
Crocodylus acutus 211102010001010001110111020000000002100010011100100110001000000111000001011100110000100111010111001102?001000
Alligator mississippiensis 2121110100101120013110010300101001210000010110000000111020201211110001102001011100011010011110020101111001000
Necrosuchus ionensis 212??1???????????0??????0???????????????????????????????????????????????????????????????????????????111??????
Eocaiman cavernensis 112??1?????11?1?????????0?????????21??1??11???12?000?????0?????1?1?0???????????????????????????2??????0???0?1
Paleosuchus trigonatus 2111212111111111100011011200011001210000010110010010111121000011110011102001011212?11020010110020102111021000
Paleosuchus palpebrosus 2111212111?11?111000110?1200011001210100010110010010111121000011110011102001011212?11020010110020102111021000
Mourasuchus spp. 300111?????0111?10001100011?0031112000100111??01000011112?3111?11110?110?00????211??113????1???2010?111120000
Purussaurus mirandai 2110?1??1?11?121100?1001022?002111210210?11???0000001111?13111?11100?1102001011200??103????????201?2?11121011
Purussaurus neivensis 21???1??1?11?12110001101022?0021112102100111??0000001111213111111100?110200?011200?110200101??02??02111121011
Melanosuchus fisheri ?1?????????1?1111????1?10?0?0012?1210021?11???1?0???11??2?1001?11100????????0????1??10????????02???2?11?12001

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 163
Melanosuchus niger 2111111010111111102011010200001201210021011110100000111121100111110001102001011211111030010110020102111012001
Caiman yacare 2111111010111011102011010200001201110010011110100000111121110111110001102001011201111030010110020102111032101
Caiman crocodilus 2111111010111011102011010200001201110010011110100000111121100111110001102001011201111030010110020102111022001
Caiman latirostris 2111111010111111102011010200001201210020011110100000111121100111110001102001011211111030010110020102111022001
Caiman lutescens 2????1??????????????????020?001201212020011???1000001111212011?1?100?????0??????11??103?????2???????11??22001
MAS-PALEO-RdlF 001/2011 211111101011111110201101020?0012012100200111??10000011???1100111?100?1?0?0??01?211??103??????0??0??2111022?01

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 164
APÉNDICE V
Medidas craneales y mandibulares (en cm.) de MAS-PALEO-ROSARIO DE LA FRONTERA 001/2011:
LC: Longitud craneal (desde el extremo anterior del hocico hasta la superficie posterior del cóndilo occipital) 38,16 AC: Ancho craneal (entre cóndilos externos del cuadrado) 24,55 ABH: Ancho base del hocico (a nivel del borde orbital anterior) 24,36 LH: Longitud del hocico (desde su extremo anterior hasta el borde orbital anterior) 25,03 LCD: Longitud craneal dorsal (desde el extremo anterior del hocico hasta el margen posterior del supraoccipital) 36,35 AO: Ancho orbital máximo 6,892 LO: Longitud orbital máxima 6,148 AIO: Ancho interorbital mínimo 3,440 LTC: Longitud de la tabla craneal (desde borde posterior orbital hasta el borde posterior del supraoccipital) 8,336 ATC: Ancho de la tabla craneal (entre esquinas de los escamosales) 10,49 ANE: Ancho narina externa 4,706 LNE: Longitud narina externa 4,108 LFST: Longitud máxima de fenestra supratemporal 1,898 AFST: Ancho máximo de fenestra supratemporal 1,544 ALCO: Altura de cóndilo occipital 1,962 DTCO: Diámetro transverso del cóndilo occipital 2,894 ALFM: Altura del foramen magnum 1,822 DTFM: Diámetro transverso del foramen magnum 2,590 Altura craneal (desde la superficie dorsal del parietal hasta el borde ventral del basioccipital) 8,022 AFI: Ancho máximo del foramen incisivo 2,668 LFI: Longitud máxima del foramen incisivo 3,432 ALPN: Altura del pasaje nasal a nivel del 4º alvéolo maxilar 4,260 APNPAL: Ancho del pasaje nasal a nivel del 4º alvéolo maxilar 16,23 LFPA: Longitud máxima de la fenestra palatal (entre pterigoides y maxilar) 9,384 AFPA: Ancho máximo de fenestra palatal (entre ectopterigoides y palatinos) 4,642 APT: Ancho de los pterigoides en fenestras palatales 1,256 ALFME: Altura máxima de la fenestra mandibular externa 3,944 LFME: Longitud máxima de la fenestra mandibular externa 7,690 LM: Longitud mandibular (desde el extremo anterior del dentario hasta el extremo posterior del proceso retroarticular 46,25 LD: Longitud del dentario (desde su extremo anterior hasta el borde anterior de la fenestra mandibular externa 28,62 AAM: Ancho arco mandibular a nivel del 10º alvéolo 8,002 ALSM: Altura de la sínfisis mandibular 3,026 LSM: Longitud de la sínfisis mandibular 6,716 LFIC: Longitud foramen intermandibular caudal 3,020 AHPMM: Ancho del hocico a nivel de la sutura premaxilar-maxilar 12,62 APIFD: Ancho del placa interfenestral entre fenestras supratemporales 2,796 AP: Ancho del parietal en el borde posterior de la tabla craneal 2,234 LPA: Longitud palatal (desde el borde anterior de la fenestra palatal hasta el extremo del hocico) 18,22 LM: Longitud palatal del maxilar (a lo largo de la línea media de sus procesos palatales) 8,688 LPAL: Longitud máxima de los palatinos (a lo largo de su línea media) 12,00 AIFPAL: Ancho mínimo de la placa interfenestral palatal 3,626 LPT: Longitud de los pterigoides a lo largo de su línea media ? ANI: Ancho máximo de la narina interna ? LNI: Longitud máxima de la narina interna ? APT: Ancho de los pterigoides (entre sus esquinas posteroventrales) ? ALRMA: Altura máxima de la rama mandibular a nivel de la fenestra mandibular externa 8,668 AFGL: Ancho máximo fosa glenoidea 5,414 DAM: Diámetro anteroposterior del 4º alvéolo maxilar 1,830 DTAM: Diámetro transverso del 4º alvéolo maxilar 1,562 DAD: Diámetro anteroposterior del 4º alvéolo dentario 1,822 DTD: Diámetro transverso del 4º alvéolo dentario 1,654 ACY: Ancho craneal entre yugales a nivel de la barra postorbital 23,26
Índices craneales (en %):
ARC: Ancho relativo craneal = (AC/LC)x100 64,32 ARH: Ancho relativo del hocico = (ABH/LH)x100 97,32 LRH: Longitud relativa del hocico = (LH/LC)x100 65,59 AREIO: Ancho relativo del espacio interorbital = (AIO/LO)x100 55,95 LRTC: Longitud relativa de la tabla craneal = (LTC/ATC)x100 79,48 LOR: Longitud relativa orbital = (LO/LC)x100 16,11 ARO: Ancho relativo orbital = (AO/LO)x100 112,1 ARNE: Ancho relativo de la narina externa = (ANE/(LC-LH))x100 35,84

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 165
APÉNDICE VI
Lista de Sinapomorfias:
Gavialis gangeticus:
No autapomorphies
Crocodylus acutus:
Char. 18: 0→1
Char. 19: 0→1
Char. 23: 0→1
Char. 37: 1→0
Char. 41: 0→1
Char. 46: 0→1
Char. 49: 0→1
Char. 72: 0→1
Char. 74: 0→1
Char. 75: 0→1
Char. 88: 0→1
Char. 89: 0→1
Char. 94: 0→1
Char. 95: 0→1
Char. 99: 0→1
Alligator mississippiensis:
Char. 12: 1→0
Char. 15: 1→2
Char. 18: 0→1
Necrosuchus ionensis:
No autapomorphies
Eocaiman cavernensis:
Char. 1: 2→1
Char. 39: 0→1
Char. 43: 0→1
Char. 47: 0→1
Char. 48: 0→2
Char. 103: 1→0
Char. 109: 0→1
Paleosuchus trigonatus:
No autapomorphies
Paleosuchus palpebrosus:
Char. 38: 0→1
Mourasuchus spp:
Char. 1: 2→3
Char. 2: 1→0
Char. 3: 1→0
Char. 12: 1→0
Char. 24: 1→0
Char. 26: 2→1
Char. 36: 1→0
Char. 48: 0→1
Char. 67: 0→1
Char. 86: 0→1
Char. 106: 1→0
Purussaurus mirandai:
Char. 22: 1→0
Purussaurus neivensis:
Char. 87: 3→2
Melanosuchus fisheri:
No autapomorphies

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 166
Melanosuchus niger:
No autapomorphies
Caiman yacare:
Char. 60: 0→1
Char. 105: 2→3
Char. 107: 0→1
Caiman crocodilus:
No autapomorphies
Caiman latirostris:
No autapomorphies
Caiman lutescens:
Char. 37: 0→2
Char. 59: 1→2
Char. 61: 0→1
MAS-PALEO-RdlF 001/2011:
No autapomorphies
Node 19:
No synapomorphies
Node 20:
Char. 3: 1→2
Node 21:
Char. 5: 0→1
Char. 11: 0→1
Char. 13: 0→1
Char. 15: 0→1
Char. 21: 0→1
Char. 31: 0→1
Char. 34: 0→1
Char. 35: 0→2
Char. 42: 0→1
Char. 52: 1→0
Char. 54: 0→1
Char. 55: 0→1
Char. 63: 0→1
Char. 70: 0→1
Char. 71: 0→1
Char. 78: 0→1
Char. 84: 0→1
Char. 93: 0→1
Char. 98: 0→1
Char. 101: 0→1
Node 22:
Char. 5: 1→2
Char. 10: 0→1
Char. 25: 0→1
Char. 30: 0→1
Char. 48: 0→1
Char. 51: 0→1
Char. 69: 0→1
Node 23:
Char. 9: 0→1
Char. 16: 0→1
Char. 17: 0→1
Char. 56: 0→1
Char. 58: 0→1
Char. 80: 1→2
Char. 100: 1→2
Char. 106: 0→2

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 167
Node 24:
Char. 33: 0→1
Char. 60: 0→1
Char. 61: 0→1
Char. 104: 0→1
Node 25:
Char. 39: 0→1
Char. 43: 0→1
Char. 62: 0→1
Node 26:
Char. 15: 1→2
Char. 38: 0→2
Char. 108: 0→1
Node 27:
Char. 40: 0→1
Char. 106: 2→1
Node 28:
Char. 39: 1→2
Node 29:
Char. 19: 0→2
Char. 47: 0→1
Char. 106: 1→2
Node 30:
Char. 14: 1→0
Char. 35: 2→1

Nuevos restos de Alligatoridae Cuvier, 1807 (Eusuchia, Crocodylia) del Neógeno del Sur de la provincia de Salta. Implicancias sistemáticas.
Francisco Barrios 168
















