Extinción Selectiva de Vertebrados Marinos del Neógeno en el Pacifico de Sudamérica.
46
UNIVERSIDAD CATÓLICA DEL NORTE FACULTAD DE CIENCIAS DEL MAR DEPARTAMENTO BIOLOGÍA MARINA Extinción Selectiva de Vertebrados Marinos del Neógeno en el Pacifico de Sudamérica. Jaime Villafaña Navea Profesor Guía: Marcelo Rivadeneira 2010
Transcript of Extinción Selectiva de Vertebrados Marinos del Neógeno en el Pacifico de Sudamérica.

UNIVERSIDAD CATÓLICA DEL NORTE
FACULTAD DE CIENCIAS DEL MAR
DEPARTAMENTO BIOLOGÍA MARINA
Extinción Selectiva de Vertebrados Marinos del Neógeno en el
Pacifico de Sudamérica.
Jaime Villafaña Navea
Profesor Guía: Marcelo Rivadeneira
2010

I
Extinción Selectiva de Vertebrados Marinos del Neógeno en el Pacifico de
Sudamérica.
Por: Jaime Villafaña Navea
Departamento Biología Marina
Fecha: Marzo 2011
Aprobado Comisión de Calificación
_____________________
Decano Facultad Ciencia del Mar
Ernesto Cortés Pizarro
_________________________________
Marcelo Rivadeneira
Profesor Guía
_ ______________________
Pilar Haye Molina
Profesor Corrector
_ ______________________
Carlos Gaymer Garcia
Profesor Corrector
Tesis entregada como un requisito para obtener el título de
Biólogo Marino en la Facultad de Ciencias del Mar. Sede Coquimbo.
2010

II
UNIVERSIDAD CATOLICA DEL NORTE
FACULTAD DE CIENCIAS DEL MAR
Departamento de Biología Maria
Extinción Selectiva de Vertebrados Marinos del Neógeno en el Pacifico de
Sudamérica.
Actividad de Titulación presentada
para optar al Título de Biólogo Marino
Jaime Villafaña Navea
Coquimbo, Diciembre 2010

III
Resumen
Los intensos cambios climáticos y oceanográficos globales observados durante el
Neógeno (últimos 23 millones de años) afectaron significativamente los patrones de
diversificación de biotas marinas. A pesar del rico registro fósil de vertebrados
Neógenos en la costa Pacífica de Sudamérica, a la fecha no existen estudios que den
cuenta los patrones de diversificación de los taxa. El objetivo de este estudio fue
evaluar el recambio faunístico de vertebrados marinos y los patrones de selectividad
de la extinción de diferentes géneros. Se estimó el rango estratigráfico y tamaño
corporal de 87 géneros de vertebrados marinos (peces, mamíferos y aves) presentes
durante los últimos 23 Ma en la costa Pacífica de Sudamérica. La información fue
obtenida a partir de una exhaustiva búsqueda de literatura, incluyendo más de 200
artículos publicados. Se estimaron las tasas de diversificación de géneros (diversidad,
originación y extinción) para cada taxa durante el neógeno, y se evaluó el grado de
selectividad en la extinción del Neógeno tardío respecto del rango geográfico y
tamaño corporal usando modelos lineares generalizados mixtos. Se observó un
incremento en la diversidad de géneros durante el Mioceno, la que fue seguida de una
fuerte caída hacia el Mioceno tardío-Plioceno temprano, producto de una extinción
masiva de ca. 80 % de los géneros. La extinción fue selectiva respecto del rango
geográfico, pero no del tamaño corporal; géneros con una distribución geográfica más
amplia tuvieron menos probabilidades de extinción. Las causas de esta enorme
extinción se relacionarían con los cambios oceanográficos y ecológicos inducidos por
el establecimiento del sistema de Humboldt actual. Un estudio más profundo de las
dinámicas macroevolutivas ayudaría a entender el origen de la fauna actual de
vertebrados en la región.

IV
Abstract
The intense climatic and oceanographic global changes observed during the Neogene
(last 23 Myr) deeply profoundly the diversification patterns of the marine biota.
Despite Neogene outcrops along the Pacific coast of South America harbor a rich
marine vertebrate fossil record, to date there are no studies aimed to at understanding
diversification patterns of these taxa. The goal of this study was to evaluate the faunal
turnover of marine vertebrates during the Neogene along the Pacific coast of South
America, as well as the existence of selectivity patterns for the extinction of genera.
We estimated the stratigraphic ranges, geographic ranges, and body size of 87 genera
of marine vertebrates (elasmobranchs, aves, and mammals) recorded during the last
23 Myr along the Pacific coast of South America. The information was gathered
from an exhaustive literature compilation, including more than 200 published articles.
We estimated the diversification rates (diversity, origination and extinction) for each
taxa across the Neogene, was evaluated the degree of selectivity of a late Neogene
extinction according to the geographic range and body size using generalized linear
mixed models. The diversity of genera increased during the Miocene, followed by an
abrupt drop towards the late Miocene-early Pliocene, coeval with an intense
extinction that affected ca. 80 % of genera. The extinction was selective according to
the geographic range, but not to body size; widespread genera had less probabilities
of extinction. The causes of this large extinction may be related to the oceanographic
and ecological changes induced by the onset of cold conditions of the modern
Humboldt Current. A deeper study of the macroevolutionary dynamics would help to
understand the origin of the vertebrate fauna at the region.
.

V
AGRADECIMIENTOS.
Me gustaría agradecer en primer lugar a mi profesor guía, el Dr. Marcelo
Rivadeneira, por su gran apoyo durante toda mi formación académica. No solo
contribuyo en mi desarrollo académico si no que también en el ámbito personal.
Agradezco su comprensión y sus consejos durante todo momento, además de la
confianza y tiempo brindado a mi persona. Gracias por confiar en mí y apoyarme
durante estos tres años en que los cuales trabajamos.
A mis profesores correctores, la Dra. Pilar Haye y el Dr. Carlos Gaymer por
su apoyo durante el desarrollo de esta tesis. Gracias por sus críticas constructivas y
por darse el tiempo de ayudarme en cualquier momento. Agradezco su interés por
ayudarme a mejorar y crecer en el tiempo. A todos los profesores de la carrera de
biología Marina, por los conocimientos entregados y apoyo.
Quiero agradecer a mi familia por su apoyo incondicional en todos estos cinco
cortos años de estudio. Muchas gracias por confiar en mis capacidades y apoyarme en
los momentos más difíciles.
Por ultimo quiero agradecer a todos mis amigos, los cuales apoyaron y
confiaron en cada momento de mi formación universitaria. A mis amigos del colegio,
compañeros de la carrera y amigos de la vida. Muchas gracias a todos ustedes por
siempre creer que podría lograr esto a pesar de las dificultades de la vida.
Dedico esta tesis a toda mi familia (madre y hermanos) y amigos. A mi tío
Hernán Guerrero por su confianza de años y por su ayuda en los momentos más
complicados.
Todas las personas que agradezco son parte fundamental de este logro, sin
ellos no hubiera podido llevarlo a cabo. Gracias a todos ustedes, les demostrare que
seguiré avanzando y creciendo como persona.

VI
ÍNDICE GENERAL
1. Introducción………………….………………………………………………….....1
1.1Hipótesis……………………………………………………………….…..6
1.2 Objetivo general………………………………………………………….6
1.3 Objetivo especifico…………………………………………………….…6
2. Materiales y Métodos……………….……………………………………..….…...7
2.1 Obtención de datos……………………………………………………….7
2.2 Análisis de datos………………………………………………………….8
3. Resultados………………………………………………………….……………...10
4. Discusión……………………………………………………………………….....13
5. Conclusiones.………………………………………………………………...........17
6. Referencias………….……………………………………...……………..............18
7. Tablas………………………………………………………………………..….....26
8. Figuras…………………………………………………………………………….32

VII
LISTADO DE TABLAS
Tabla 1: Principales eventos tectónicos, climáticos y oceanográficos, asociados a los
cambios en composición faunística (moluscos) observados durante los 30 millones de
años en el Pacífico Sudamérica……………………………………………………26
Tabla 2: Ocurrencias fósiles de vertebrados marinos del neógeno en el Pacífico de
Sudamérica. ……………………………………………………………………….27
Tabla 3: Coeficientes de correlación entre los pares de taxa, según los patrones de
diversificación obtenidos………………………………………………………….29
Tabla 4. Selectividad de la extinción de vertebrados marinos del Neógeno, usando
modelos lineares generalizados, con y sin incluir efectos filogenéticos…………30
Tabla 5. Porcentajes de extinción de géneros y especies durante el Neógeno en
sistemas marinos………………………………………………………………….31

VIII
LISTADO DE FIGURAS
Figura 1: Patrones de diversificación de vertebrados marinos del Neógeno en el
Pacífico de Sudamérica a nivel de género y especie...…………………………..32
Figura 2: Patrones de diversificación de vertebrados marinos del
Neógeno en el Pacífico de Sudamérica por niveles de precisión………………..33
Figura 3: Patrones de diversificación de vertebrados marinos del Neógeno en el
Pacífico de Sudamérica a nivel de taxa (aves, mamíferos y peces)………………34
Figura 4: Tasa de extinción de vertebrados marinos durante el Neógeno a nivel de
taxa (peces, mamíferos y aves), comparado con los géneros de vertebrados marinos
presentes actualmente………………………………………………………….....35
Figura 5: Rango promedio de géneros de vertebrados marinos del Neógeno en el
Pacífico de Sudamérica extintos y sobrevivientes…………………………….….36
Figura 6: Cambios paleoclimáticos y oceanográficos, asociados con la diversidad de
géneros de vertebrados marinos del Neógeno en el Pacifico de Sudamérica...…..37

1
1. INTRODUCCIÓN
La extinción es una fuerza primordial que moldea las dinámicas
macroevolutivas de las especies (Jablonski 2004). Se estima que más del 99% de
todas las especies que alguna vez existieron están actualmente extintas (Raup, 1986).
La desaparición de especies puede ocurrir en cualquier escala de tiempo, esto puede
suceder en periodos relativamente largos de estabilidad que alternan con eventos de
extinción de corta duración (Mckinney, 1997). La mayoría de los episodios de
extinción son biológicamente selectivos, el análisis posterior de las víctimas y
sobrevivientes ofrece la mayor posibilidad de dilucidar la causa de estos
acontecimientos (Raup, 1986).
La extinción selectiva de especies, o también llamada vulnerabilidad relativa
(Mckinney, 1997), ha provocado un aumento del interés en las áreas de la ecología y
la paleontología. Se han propuesto diferentes definiciones de selectividad, en el
contexto de la teoría de selección de multiniveles (Jablonski, 2008b). Todo esto con
el fin de discutir los posibles enfoques hacia una formalización mas completa de la
selectividad de especies. La selectividad se refiere a la originación o persistencia de
especies debido a la interacción con el ambiente (Pimm et al., 1988). Se han sugerido
rasgos biológicos individuales como el tamaño corporal, el tipo de dieta, la
fecundidad, tasa de reproducción, tiempo de vida y movilidad como factores que
podrían determinar extinción o sobrevivencia de determinadas especies (Mckinney,
1997). En géneros de mamíferos terrestres como el Glyptodon y Megatherium, la
mayoría de los autores concuerdan que el 80% de los géneros extintos de este periodo
poseían un tamaño corporal grande, sin embargo existe un debate que esto se podría
atribuir principalmente a tasas de reproducción bajas (Johnson, 2002). El cambio en
el tipo de dieta a finales del Pleistoceno también habría afectado la sobrevivencia de
la familia Gomphotheriidae en la región pampeana de Argentina (Sanchez, 2003). Es
así como, la gran mayoría de los estudios realizados en mamíferos e invertebrados
se han centrado desde principios y finales del Pleistoceno (2.6 – 0.01 Ma) han
tomando como indicador general de selectividad al tamaño corporal (Roy et al.,
2001, 2002). Aunque el tamaño corporal ha sido ampliamente estudiado como un

2
indicador de la extinción de especies, se han encontrado discrepancias entre los
diferentes grupos de organismos y riesgos de extinción asociados a este indicador
(Blackburn & Gaston 1994). Motivo por el cual no se puede asociar con certeza como
un patrón universal de extinción de especies, debido a la diversidad de ambientes y
taxa estudiados (Mckinney, 1997). Estudios sobre la extinción de moluscos en el
cretácico demuestra que el tamaño corporal no era un factor determinante en la
sobrevivencia de estos organismos, cobrando mayor fuerza el tamaño del rango
geográfico (Jablonsky, 1987; Jablonski & Hunt, 2006; Jablonski, 2008a; Krug et al.,
2008, 2009). En el caso de estudios en vertebrados de nuestro planeta, la extinción de
estos organismos se asocia directamente a la expansión poblacional del hombre y la
actividad de la caza (Barnosky et al., 2004; Barnosky, 2008). La gran mayoría de los
registros que se conocen sobre las causas de extinciones de vertebrados en
Sudamérica corresponden a vertebrados terrestres del Cuaternario (García, 1999;
Steadman et al., 2005; Koch & Barnosky, 2006). Además del fuerte cambio climático
provocado en este periodo (Roberts et al., 2001; Guthrie, 2006).
Gran parte de los taxa pertenecientes al Neógeno (últimos 23 Ma) se vieron
afectados significativamente por los intensos cambios climáticos y oceanográficos
de este período (Tsuchi, 2002). Se ha sugerido que la activación de la surgencia
costera y condiciones anóxicas de este periodo provocaron la pedida del 66 % de
géneros de moluscos en la costa Pacifica de Sudamérica, probando un descenso de la
diversidad hacia la actualidad (Rivadeneira & Marquet 2007). Es así como el
Neógeno se caracteriza principalmente por ser una edad que poseía un clima frío. Las
temperaturas de este periodo fueron de 5º menos en comparación con el Paleógeno,
debido a la glaciación de los casquetes polares (Zachos et al., 2001). Con respecto al
Pacífico de Sudamérica, en el Neógeno se reconocen tres episodios cálidos,
aproximadamente hace 15,5 Ma, 11,5 Ma y 5,7 Ma, además de un intenso periodo
frío entre los 15 y 23 Ma (Tsuchi, 2002). Intensos cambios tectónicos, climáticos, y
oceanográficos en la costa Pacífica de Sudamérica ocurrieron durante los últimos 30
Ma (Tabla 1), destacándose la hiperaridéz del desierto de Atacama y el cierre del
Istmo de Panamá a mediados del Plioceno (5,3 Ma). Además que en el Mioceno (23

3
Ma) la cordillera de los Andes presentaba el 50% de su altura actual (Gregory-
Wodzicki, 2000), características que habrían activado el sistema de corrientes de
Humboldt actual (Logan, 1968). Los eventos climáticos de este periodo se
caracterizan por grandes glaciaciones tanto en el Mioceno (Glaciación Antártica Este
y Oeste) como en el Plioceno (Glaciación Magallánica), además de un periodo
denominado “Optimo climático del Neógeno” (Zachos et al., 2001; Tsuchi, 2002). En
cuanto a los eventos oceanográficos, se activa la surgencia costera en Perú y Chile, el
sistema de la corriente de Humboldt a principios del Plioceno y la intensidad actual
de surgencia se presenta a finales de este periodo, 2.6 Ma aproximadamente
(Zinmeister, 1978; Ibaraki, 1997). En el Neógeno la composición faunística de Perú y
Chile comienza por una fauna subtropical, cambiando de ahí en adelante a una fauna
temperada fría (Zinmeister, 1978; McCulloch et al., 2000). Este periodo se
caracteriza por una gran diversificación de diferentes especies de mamíferos, aves y
peces, sin registrarse grandes extinciones hasta llegar al límite Plioceno-Pleistoceno
(2,6 Ma), periodo en el cual comienzan a desaparecer géneros de vertebrados con
tamaños corporales superiores a los actuales (Barnosky 2008).
A nivel sudamericano, una de las regiones con la mayor riqueza de fósiles
de restos de vertebrados marinos del Neógeno ha sido la ciudad de Pisco
(Departamento de Ica), ubicada en el centro sur de Perú (Marocco & De Muizon,
1988) y la localidad de Bahía inglesa (Chile) (Chávez, 2008). A nivel de grupos de
vertebrados, las aves son el grupo que presenta una mayor riqueza fósil,
principalmente diferentes géneros de pingüinos, destacándose la familia
Spheniscidae. Los registros de esta familia y otros grupos de aves han sido asignados
al Mioceno superior - Plioceno inferior de Chile (Walsh, 2001; Walsh & Hume, 2001;
Acosta et al., 2002; Clarke et al., 2003; Emslie & Guerra, 2003; Stucchi, 2003;
Acosta & Canto, 2005; Acosta & Stucchi, 2005; Acosta, 2006; Acosta et al., 2006;
Walsh & Suárez, 2006; Acosta et al., 2007; Chavez, 2007a, b; Gohlich, 2007; Acosta
& Tambussi, 2008; Jadwiszcak, 2009) y Plioceno inferior de Peru (Stucchi, 2002;
Clarke et al., 2003; Stucchi et al., 2003; Stucchi & Urbina, 2004; Chavez et al., 2007;
Stucchi, 2007). La mayoría de los de los registros fósiles de Chile y Perú no difieren

4
demasiado en cuanto diversidad de especies, encontrándose diferencias a nivel
especifico. Los generos Daption y Eudyptula solo se han encontrado en la localidad
de Bahía Inglesa (Chavez, 2008). En el caso de Perú, los únicos géneros presentes en
la localidad de Pisco son Morus y Ramphastosula (Stucchi, 2004).
Con respecto a los restos fósiles de peces, se pueden destacar algunos grupos
de tiburones y rayas de Perú (formación de Pisco) y Chile (formación Coquimbo y
Bahía Inglesa), pertenecientes a los géneros Carcharias, Carcharocles, Isurus,
Notorynchus y Myliobatis (Nolf, 2002; Apolín et al., 2004; Suárez et al., 2004; Staig,
2006; Cione & Barla, 2008), además de algunos estudios anteriores sobre las
características fósiles de los peces en Perú y Chile (Long, 1993).
Sobre el registro fósil de mamíferos de Sudamérica, se pueden destacar restos
de ballenas del Plioceno temprano en Perú (De Muizon, 1993; De Muizon et al.,
2004a; Bouetel & De Muizon, 2006) y manatíes del Mioceno tardío en Chile
(Bianucci et al., 2006). Además se han realizado algunas descripciones sobre los
patrones craneales del delfín Brachydelphis mazeasi (Gutstein et al., 2009). Registros
de restos fósiles de focas y lobos marinos del Neógeno han sido encontrados en Chile
y Perú (De Muizon, 1978, 1981; Walsh & Naish, 2002). Uno de los géneros mas
estudiados en Perú corresponde a los perezosos marinos, generalmente del genero
Thalasssocnus (De Muizon & Mcdonald, 1995; De Muizon et al., 2004a, b; Pujos &
Salas, 2004).
A pesar de este rico registro fósil y a la existencia de un fuerte recambio
faunístico durante el Neógeno en Chile y Perú, a la fecha no existen estudios actuales
sobre los patrones de diversificación (Originación y extinción) y selectividad de
extinción durante el Mioceno-Plioceno de géneros de vertebrados marinos. Diversos
estudios plantean la existencia de marcados cambios en la diversidad de taxa de
vertebrados e invertebrados durante el Neógeno (Rivadeneira & Marquet, 2007;
Jadwiszcak, 2009; Canto et al., 2010) y la existencia de eventos de extinción (Chávez
et al., 2007). No obstante dichas dinámicas no han sido evaluadas de manera formal.
Del mismo modo, los patrones de selectividad han sido escasamente abordados. La
mayoría de los trabajos sobre selectividad han sido evaluados en invertebrados,

5
atribuyendo la selectividad de extinción al tamaño del rango geográfico (Jablonsky,
1987). Moluscos que poseen rangos geográficos más amplios han sido menos
propensos a la extinción que aquellos con un rango más restringido (Jablonski &
Raup 1995; Payne & Finnegan 2007). En algunos estudios en vertebrados se ha
planteado que especies de pingüinos de mayor tamaño corporal habrían tenido
mayores probabilidades de extinción (Clarke, et al., 2007).
La hipótesis que se planteo en este trabajo es que la diversidad de género de
vertebrados marinos del Neógeno decae del Mioceno al Plioceno debido a una fuerte
extinción. Además que esta extinción fue selectiva de acuerdo al tamaño corporal y
rango geográfico de los grupos estudiados en este trabajo (Peces, aves y mamíferos)
Tanto el descenso de la diversidad en la transición Mioceno-Plioceno y la
selectividad ha sido estudiado en Moluscos (Rivadeneira & Marquet, 2007;
Jablonsky, 1987). De acuerdo a las hipótesis antes descritas, se plantearon como
objetivos estimar las tasas de recambio faunístico durante el Mioceno-Plioceno de los
diferentes grupos de vertebrados Marinos y determinar patrones de selectividad de
extinción. La finalidad principal de este trabajo es esclarecer las posibles causas de
extinción durante los últimos 23 Millones de años. Se tomo en cuenta características
como el tamaño corporal y el rango geográfico como indicadores de selectividad de
extinción, ambos ampliamente estudiados anteriormente. Dichos análisis podrían
contribuir a entender y predecir la desaparición estos taxa de vertebrados marinos en
la actualidad.

6
1.1 Hipótesis
Hipótesis específica 1: La diversidad de géneros de vertebrados decae del Mioceno
al Plioceno debido a una fuerte extinción durante el Mioceno-Plioceno.
Hipótesis específica 2: La extinción Mioceno-Plioceno de los diferentes taxa de
vertebrados marinos fue selectiva de acuerdo al tamaño corporal y rango geográfico.
1.2 Objetivos generales
El objetivo general de este estudio es analizar el recambio faunístico de
vertebrados marinos (aves, peces y mamíferos) del Neógeno (Mioceno-Plioceno) en
la costa Pacífica de Sudamérica.
1.3 Objetivos específicos
Objetivo específico N°1: Estimar las tasas de recambio faunístico durante el
Mioceno-Plioceno de los diferentes grupos de vertebrados marinos.
Objetivo específico N°2: Determinar los patrones de selectividad en la extinción de
diferentes grupos marinos durante el Mioceno-Plioceno en el Pacifico de Sudamérica,
en relación al tamaño corporal y rango geográfico.

7
2. MATERIALES Y MÉTODOS
2.1 Obtención de datos
Los datos del presente trabajo fueron obtenidos mediante una exhaustiva
revisión bibliográfica correspondiente al registro fósil de vertebrados marinos (peces,
aves y mamíferos) del Neógeno (Mioceno-Plioceno) en Sudamérica, principalmente
en las localidades de Chile y Perú (Tabla 2. Anexo). Los sectores correspondientes a
Chile son: Antofagasta (23º37‟26‟‟), Bahía Inglesa (27º21'28' S), Colchagua
(34º41‟00‟‟), Concón (32º57‟56‟‟S), Constitución (32º20‟21‟‟), Coquimbo
(29º58‟16‟‟), Curanilahue (37º28‟19‟‟), Isla Mocha (38º21‟32‟‟), Matanzas
(71º52‟15‟‟), Rapel (33º56‟57‟‟) y Tongoy (30º15‟08‟‟). En el caso de Perú la gran
mayoría de los restos fósiles estudiados pertenecen a la ciudad de Pisco (13º42‟55‟‟)
ubicada a 290 km al sudeste de Lima. Los demás registros fósiles corresponden a las
localidades de: Arequipa (16º09‟25‟‟), La Huaca (4º54‟40‟‟), Yauca (15º40‟18‟‟) y
Zamaca (14º39‟12‟‟). Toda la recopilación de datos se realizó dentro de estos dos
países con sus localidades descritas anteriormente.
La búsqueda de la información fue realizada por medio de lectura
bibliográfica de artículos, tesis y revistas de investigación. Mediante esta revisión se
recopilaron los rasgos estratigráficos de especies y géneros de vertebrados marinos
del neógeno en el Pacifico de Sudamérica. Además para cada género y especie se
selecciono información sobre el tamaño corporal y rango geográfico. Con respecto al
tamaño corporal, este se estimó de acuerdo a información entregada por los autores
de cada investigación, o si no con respecto a similitudes de especies o géneros
estimados por otros autores. En el caso del rango geográfico de las diferentes especies
y géneros de vertebrados, esto se llevó a cabo mediante la distribución más amplia
que presenten, es decir la localidad más al sur y más al norte en las cuales se hayan
registrado.

8
2.2 Análisis de datos
Se realizaron dos tipos de análisis, en base a información obtenida del registro fósil
de vertebrados marinos (peces, aves y mamíferos) del Pacífico de Sudamérica: a)
evaluación de patrones temporales de diversificación (originación y extinción de
géneros), y b) análisis de selectividad de la extinción
Las tasas de diversificación (originación y extinción de géneros), fueron
estimadas utilizando métodos estándar en análisis paleoecológicos (Foote & Miller,
2007), empleando las siguientes fórmulas:
100total
extExt
Mio
MioPlioMio
Para calcular la tasa extinción de géneros durante la transición Mioceno-
Plioceno ( ExtPlioMio ) se dividió la cantidad de géneros que se extinguieron en el
Mioceno y no pudieron llegar al plioceno ( extMio ) por la cantidad total de géneros
que habían antes que ocurriera la extinción ( totalMio ), posteriormente este dato se
multiplicará por 100 para así obtener el porcentaje de extinción de vertebrados
marinos. Para obtener la tasa de originación se realizará un procedimiento análogo
siguiendo la siguiente fórmula:
100. total
Orig
OrigPlio
PlioPlioMio
Los análisis fueron realizados en tres diferentes grupos, de acuerdo al grado de
precisión de las edades del registro fósil. En orden creciente: a) Nivel de precisión 1:
Todos aquellos datos que no especificaban la edad del registro fósil, solo
consideraban las edades para la formación. b) nivel de precisión 2: Datos ingresados
por Paleobiology Database, y c) Nivel de precisión 3: datos revisados en los trabajos

9
y los cuales se tenia seguridad de una edad especifica para el registro fósil. Este
análisis se realizo con el fin de comprobar si los patrones eran robustos
independientes del nivel de precisión.
Se evaluó la selectividad de rasgos ecológicos y de historia de vida de los
géneros a la extinción de fines del Neógeno (ver resultados). Se utilizaron modelos
lineares generalizados y modelos lineares generalizados mixtos (GLMM; Blackburn
& Duncan, 2001). En este análisis se considero que la extinción es una variable
binomial (1=extingue, 0=sobrevive), relacionada aditivamente con el tamaño corporal
y el rango geográfico de los diferentes géneros de vertebrados marinos. En orden a
controlar por posibles efectos filogénéticos, los análisis incluyeron la taxonomía
como efecto aleatorio, con familias anidadas dentro de clases (Blackburn & Duncan
2001; Bolker et al., 2008; Sodhi et al., 2008). El análisis se llevó a cabo utilizando la
librería lmer del software R (Team, 2010).

10
3. RESULTADOS.
Se obtuvo información para un total de 87 géneros y 124 especies de
vertebrados marinos del Neógeno de la costa pacífica de Sudamérica. Los patrones de
diversificación fueron evaluados a nivel de especie y genero. En primer lugar, la
diversidad de taxa fue aumentando gradualmente en el tiempo, hasta a llegar a un
máximo aproximadamente a finales del Mioceno-tardío (7.2-5.3). Posterior a este
periodo la diversidad disminuyo drásticamente (Fig.1.A). Con respecto a la tasa de
extinción, se detectaron siete diferentes pulsos de extinción (Fig.1.B). Los cambios
más considerables ocurrieron a los 6 Ma aprox. (final del Mioceno). El mayor pulso
de extinción ocurrió a los 4 Ma aprox. (mitad del Plioceno) con una extinción del
77% de los géneros y el ultimo a los 3 Ma aprox. (final del Plioceno). En el caso de
los porcentajes de originación, se pueden observar un total de cinco pulsos de
originación durante el Neógeno. Los dos pulsos mayores pulsos de originación
correspondieron a un 72% y 28% de originación de géneros de vertebrados marinos
del Neógeno (Fig 1c), ambas pertenecientes al Mioceno-medio (16 Ma.). Las tasas de
extinción y originación a nivel de especie resultaron similares a las de nivel de
géneros. A la vez, se calcularon los patrones de diversificación para cada grupo de
datos (Precisión 1, Precisión 2 y Precisión 3). Al analizar las curvas obtenidas en los
diferentes grupos de datos, se puede apreciar un patrón similar en cuanto a la
diversidad de géneros, ya que en los tres aumenta a partir del Mioceno-medio (16
Ma) y decae a finales del Mioceno-tardío (Fig. 2). Sin embargo, la principal
diferencia se destaca en el criterio de precisión 2, en el cual la mayor diversidad se
encuentra en un breve periodo de tiempo, entre los 5-4 Ma., correspondiente al
Plioceno-temprano (Fig. 2. B). Los pulsos de extinción también variaron entre los
diferentes criterios de precisión, siendo mayores en el criterio de precisión 2 (6 en
total), uno menos que el criterio de precisión 3 y ambos mayores en comparación con
el nivel de precisión 1 (Fig. 2. A-C). Por ultimo, en los pulsos de originación, al igual
que la tasa de extinción, el criterio de precisión 2 es el que presenta mayor número de
pulsos (durante todo el Neógeno), seguido por el criterio de precisión 3 y 1 (Fig. 3. G-

11
I). Los eventos de extinción combinados removieron ca. 88% de los géneros
existentes durante el Mioceno-tardío.
A nivel de taxa mayores (peces, mamíferos y aves), los patrones de
diversificación son similares (Fig. 3). Los peces presentan una mayor diversidad de
géneros en el periodo de 11-6 Ma aprox. (Mioceno-tardío). Los mamíferos alcanzaron
su máxima diversidad a los 5 Ma aprox. (Final Mioceno-tardío). Por último, las aves
con mayor diversidad en los 11-8 Ma aprox. En el caso de las tasas de extinción entre
taxa, los que presentan mayores porcentajes son los peces y mamíferos con un 100 %
y 89 % respectivamente en el Plioceno-temprano (5.3-3.6 Ma), periodo en el cual
también las aves presentan su mayor porcentaje de extinción de géneros. Con
respecto a la tasa de originación, las aves con un 100% son las que tienen el
porcentaje de originación de géneros más alto, seguidos por los mamíferos (70%) y
peces (50%). Todos estos cambios ocurrieron durante el Mioceno-temprano. Los
valores de correlación obtenidos demuestran relaciones positivas en las curvas de
diversidad de géneros y tasa de extinción, además de una relación negativa en la tasa
de originación. Los valores de r se pueden observar en la tabla 3. No se observan
mayores diferencias con respecto al porcentaje de extinción entre los diferentes
grupos de vertebrados marinos del Neógeno (Fig. 4), las aves presentaron el menor
porcentaje de extinción (71%) y el mayor perteneció a los peces (94%), este ultimo
valor fue similar al de los mamíferos (91%). Estos resultados fueron comparados con
la presencia de estos géneros en la actualidad, mostrando diferencias significativas en
los peces. Actualmente solo un 12% de los géneros de peces están extintos, a
diferencia del 94% obtenido por el registro fósil (Fig. 4). Las aves presentan un 35%
contrastado con el 70% obtenido por el registro fósil. No obstante, los géneros de
mamíferos analizados en este trabajo son los que están más extintos en la actualidad
(78%), cercano al 90% esperado por el registro fósil.
Finalmente, el análisis de GLMM para evaluar la selectividad de la extinción
de géneros, arrojó resultados significativos solo con respecto al rango geográfico
(p=0.04). Los géneros de vertebrados marinos que poseían rangos geográficos más

12
restringidos fueron susceptibles a la extinción que aquellos géneros sobrevivientes
con rangos más amplios (Fig. 5). Los resultados no cambiaron drásticamente al
incluir o excluir efectos filogenéticos. El otro rasgo evaluado en el análisis de
selectividad fue el tamaño corporal, el cual no presento valores significativos. Los
valores obtenidos se pueden observar en la tabla 4.

13
4. DISCUSIÓN.
El registro fósil de vertebrados marinos del Neogenoo ha sido ampliamente
estudiado en la costa Pacífica de Sudamérica (De Muizon & DeVries 1985; Chavez
2008; Canto et al., 2010). Sin embargo, a la fecha no se han sido realizado estudios
sobre los patrones de diversificación de los taxa. Este estudio es la primera
aproximación para dilucidar los patrones de diversificación de estos organismos
durante los últimos 23 Ma.
Las tasas a las que se originan o extinguen especies dependerán en gran
medida de la naturaleza de las especies, o el tipo de especiación que puedan
desarrollar (Fowler et al., 1982). Los patrones de diversificación a través del Neógeno
resultaron sumamente similares a nivel de género y especie. Esto se debe
principalmente a la baja cantidad de especies por género. Spheniscus y
Thalasssocnus fueron dos de los géneros que presentaron una mayor cantidad de
especies definidas (5 especies). Los demás géneros solo presentaban una o especies
indeterminadas, debido a las malas condiciones de algunos fósiles o transcripción
poco precisa de las edades asignadas a los organismos en estudio. Es así como se
decidió realizar los posteriores análisis solo a nivel genérico.
La diversidad de géneros fue aumentando exponencialmente en tiempo,
llegando a la ser máxima en el Mioceno- tardío. Posterior a este periodo, la diversidad
de géneros disminuyo drásticamente. Los pulsos de originación se presentaron
durante todo el Neógeno. El máximo pulso ocurrió en el Mioceno medio (74%). La
tasa de originación de géneros de vertebrados marinos obtenida en este trabajo
ocurrieron en periodos relativamente cálidos y son sincrónicos con los patrones de
especiación reportados para vertebrados e invertebrados (Jablonsky, 1987; Nieto et
al., 2003; Del Rio, 2006). Faunas de macromamiferos de la península ibérica
aumentaron su diversidad a partir del Mioceno-inferior, relacionándolo
principalmente con los cambios climáticos del periodo (Nieto et al., 2003). Moluscos
del genero Nodipecten presentaron un gran aumento en la diversidad de especies en el
Mioceno-medio, periodo asociado al “optimo climático del Neógeno” (Del rio, 2006).
El mayor pulso de extinción de géneros de vertebrados marinos ocurrió en el Plioceno

14
temprano (77%). Estos pulsos de extinción se asociarían al establecimiento
definitivo de las celdas de surgencia del sistema frío de Humboldt (Tabla 1 y Fig. 6).
Estudios recientes demuestran una extinción de similar intensidad en moluscos del
Pacifico de Sudamérica, atribuibles al menos en parte a la anoxia generada como
consecuencia del establecimiento del sistema de Humboldt (Rivadeneira & Marquet,
2007; Kiel & Nielsen, 2010). Otros trabajos han concluido que la abrupta caída o
extinción de géneros a finales del Neógeno, se debe principalmente al descenso en las
temperaturas y a los fuertes cambios paleooceanográficos del periodo (Del Rio, 2006;
Cione et al. 2007; Andrés & Daniel 2008). El efecto combinado de los pulsos de
extinción habría removido mas del 85% de los géneros de vertebrados del Neógeno
en la costa Pacifica de Sudamérica. Esta extinción constituye uno de los eventos de
mayor intensidad conocida para el Neógeno para otras localidades (Tabla 5).
No se apreciaron diferencias significativas en las tendencias de los patrones de
diversificación de vertebrados marinos durante el Neógeno a nivel de taxa (Peces,
mamíferos y aves). El análisis demostró correlaciones positivas entre los taxa (Tabla
3). Los tres taxa analizados presentaron un aumento de la diversidad de géneros desde
el Mioceno-medio hasta llegar a ser máxima a finales del Plioceno-temprano. La
única diferencia es la cantidad de géneros registrados para cada taxa, siendo las aves
el grupo que presenta la menor cantidad e géneros analizados en este trabajo.
Las máximas extinciones de géneros de vertebrados marinos del Neógeno se
presentaron a finales del Plioceno-temprano. Los mamíferos fueron los que
presentaron una mayor cantidad de pulsos de extinción, además de desarrollarse estos
pulsos en tiempos anteriores a los demás taxa. Los peces fueron los que presentaron
las extinciones mas elevadas. Esto no se repitió en los pulsos de originación, donde
los mayores pulsos correspondieron a los mamíferos. En general no se aprecian
diferencias significativas en los patrones de diversificación (originacion y extinción)
entre los diferentes taxa (peces, aves y mamíferos). Al realizar la comparación entre
la cantidad de géneros extintos (registro fósil) contra la cantidad de esos géneros que
se encuentran actualmente en Sudamérica, si se observaron diferencias entre los taxa.
Los peces fueron el grupo que presento mayores diferencias en cuanto a los

15
porcentajes de extinción. Esto se puede deber a la falta de información del registro
fósil para el Plioceno. Además se debe considerar que estos géneros que se
registraron extintos, pudieron originarse en otras localidades no estudiadas. Los
mamíferos presentaron una extinción de 90% de los géneros, la cual es similar la
condición actual de estos géneros en Sudamérica (78% de extinción), demostrando
que si se pueden obtener una estimación cercana de el porcentaje de géneros extintos
a través de una exhaustiva revisión bibliográfica del registro fósil de vertebrados
marinos.
Los patrones de diversificación de vertebrados marinos del Neógeno, son
robustos independientes del nivel de precisión analizado. El único set de datos que
presenta mayores diferencias es el nivel de precisión 2. Esto podría deberse a una
transcripción poco precisa de las edades asignadas a cada registro fósil. Este análisis
finalmente demostró que es necesario llevar a cabo una revisión exhaustiva de las
diferentes asignaciones de edades realizadas por otros autores, ya que así se evitaría
omitir información y se obtendrían datos más cercanos a la realidad.
Referente a la selectividad, algunos autores postulan que es posible explicar
la estructura y funcionamiento de los ecosistemas en base a la teoría de extinción
selectiva (Fowler et al. 1982). Se debe destacar que cualquier característica que
aumente las probabilidades de supervivencia dependerá al tipo de extinción que sea
vinculada. Estudios han demostrado que el tamaño del rango geográfico no conferiría
mucha resistencia a una gran extinción en masa, pero si a una extinción de fondo
(Jablonski 2008). Finalmente, nuestro análisis de selectividad revelo que los taxa
fueron selectivos al tamaño del rango geográfico, es decir, los géneros de vertebrados
marinos que poseían rangos geográficos más restringidos fueron más susceptibles a la
extinción que aquellos con rangos mucho más amplios. Este resultado se han repetido
par otros organismos, como moluscos y algunos géneros de vertebrados terrestres
(Jablonski, 1987; Jablonski & Raup 1995; Payne & Finnegan 2007). Al evaluar el
tamaño corporal entre los diferentes taxa, no se obtuvieron resultados significativos.
Comparaciones entre el tamaño corporal y otro rasggo biológico han demostrado
resultados similares, en los cuales el tamaño corporal no seria un indicador de

16
selectividad a la extinción. (Johnson, 2002; Sanchez, 2003).Sin embargo, se han
obtenido diferentes resultados en cuanto a las probabilidades de extinción y el tamaño
corporal (Blackburn & Gaston 1994). Estas discrepancias se deben a la variedad de
taxa, habitad y escalas espaciales diferentes en los cuales suceden las extinciones
(Mckinney 1997).

17
5. CONCLUSIÓN.
Los resultados de este trabajo indican que los intensos cambios climáticos y
oceanográficos observados durante el Neógeno a escala global y regional habrían
tenido un profundo efecto en los patrones de diversificación de los taxa, obteniéndose
una de las mayores extinciones registradas hasta el día de hoy para el Neógeno,
llegando a extinguirse mas del 80% de los géneros registrados a través del registro
fósil de vertebrados marinos en el pacifico de Sudamérica, sin embargo estos
resultados pueden estar subestimados y ese sumamente importante seguir realizando
estudios sobre este tema.
Finalmente, nuestro estudio demostró que la extinción de todos estos taxa fue
selectiva solo al tamaño del rango geográfico, e independiente al tamaño corporal,
otra de las características evaluadas en este trabajo.

18
6. REFERENCIAS.
Andrés, M. & M. Daniel (2008). "Registro del género Gazella en el Neógeno
español." Stvdia Geologica Salmanticensia 8: 17-26.
Acosta, H. C. (2006) Pingüinos fósiles (Pygoscelis calderensis sp. nov.) en la
Formación Bahía Inglesa (Mioceno Medio-Plioceno), Chile. Revista
geologica de Chile 33:327-338.
Acosta, H. C. & Canto, J. (2005) Primer registro de cráneos asignados a
Palaeospheniscus (Aves, Spheniscidae) procedentes de la Formación Bahía
Inglesa (Mioceno Medio-Tardío), Chile. Revista Chile de Historia Natural.,
78, 489-495.
Acosta, H. C. & Stucchi, M. (2005) Nuevos restos de Spheniscidae (Aves,
Sphenisciformes) procedentes de la costa del Perú. . Revista Española de
Paleontología., 20 (1), 1–5.
Acosta, H. C., Canto, J. & Tambussi, P. C. (2006) Pinguinos (Aves, Spheniscidae) en
Coquimbo (Mioceno Medio - Plioceno Tardío), Chile y su vinculacion con las
corrientes oceanicas. Revista Española de Paleontología., 21(2), 115-121
Acosta, H. C. & Tambussi, P. C. (2008) South American fossil penguins: a systematic
update. . Oryctos, 7, 109-127.
Acosta, H. C., Fritis, O., Tambussi, C.P, Quinzio, A.L. (2002) Nuevos restos de
pingüinos (Aves Spheniscidae) en la Formación Bahía Inglesa (Mioceno
superior – Plioceno inferior) de Chile. , 16.
Acosta, H. C., Tambussi, P. C., Donato, M. & Cozzuol, A. M. (2007) A new Miocene
penguin from Patagonia and its phylogenetic relationships. Acta
Palaeontologica Polonica 52(2), 299–314.
Apolín, J., González, G. & Jean-Noel., M. (2004) Seláceos del Mioceno Superior de
Quebrada Pajaritos (Piura, Perú).
Barnosky, D. A., Koch, L. P., Feranec, S. R., Wing, L. S. & Shabel, B. A. (2004)
Assessing the Causes of Late Pleistocene Extincctions on the Continents.
Science, 306.

19
Barnosky, D. A. (2008) Megafauna biomass tradeoff as a driver of Quaternary and
future extinctions., 105.
Bianucci, G., Sorbi, S., Suárez, E. M. & Landini, W. (2006) The southernmost
sirenian record in the eastern Pacific Ocean, from the Late Miocene of Chile.,
5, 945-952.
Blackburn, T. M. & K. J. Gaston (1994). "Animal body size distributions: patterns,
mechanism, and implications." Trends in Ecology & Evolution 9: 471-474.
Blackburn, Y. M. & Duncan, R. P. (2001) Determinants of establishment succes in
introduced birds. . Nature, 414, 195-197.
Bolker, B. M., M. E. Brook, et al. (2008). "Generalized linear mixed models: a
practical guide for ecology and evolution." Trends in Ecology & Evolution
24: 127-135.
Bouetel, V. & De Muizon, C. (2006) The anatomy and relationships of Piscobalaena
nana (Cetacea, Mysticeti), a Cetotheriidae s.s. from the early Pliocene of Peru.
Geodiversitas, 28(2).
Canto, J., J. Yañez & Rovira, J. (2010). "Estado actual del conocimiento de los
mamíferos fósiles de Chile." Estudios Geológicos.
Cione, L. A., Mennucci, J. & Santalucita, F. (2007). "Extinción local de tiburones del
genero Carcharias Rafinesque, 1810 (Elasmobranchii, Odontaspididae) en el
Océano Pacifico oriental." Andean Geology 34.
Cione, L. A. & Barla, J. M. (2008) Causes and contrasts in current and past
distribution of the white shark (Lammniformes: Carcharodon carcharias) off
southeastern Southe America. Revista del Museo Argentino de Ciencias
Naturales., 10(2), 175-184.
Clarke, A. J., Olivero, B. E. & Puerta, P. (2003) Description of the Earliest Fossl
Penguin from South America and First Paleogene Vertebrate Locality of
Tierra del Fuego, Argentina.
Clarke, J.D., Ksepka & Stucchi, M. (2007). "Paleogene equatorial penguins challenge
the proposed relationship between biogeography, diversity, and Cenozoic
climate change." PNAS 104 11545-11550

20
Chávez, F. M., M. Stucchi & Urbina M. (2007). "El registro de Pelagornithidae
(Aves: Pelecaniformes) y la avifauna neógena del Pacífico sudeste." Bulletin
de l‟Institut Français d‟Études Andines. 36(2): 175-197.
Chávez, F. M. (2007a) Sobre la presencia de Paraptenodytes y Palaeospheniscus
(Aves: Sphenisciformes) en la Formación Bahía Inglesa, Chile. Revista Chile
de historia Natural., 80, 255-259.
Chávez, F. M. (2007b) Fossil Birds of Chile and Antartic Peninsula., 65, 551-572.
Chávez, F. M., Stucchi, M. & Urbina, M. (2007) El registro de Pelagornithidae
(Aves: Pelecaniformes) y la avifauna neógena del Pacífco sudeste. Bulletin de
l‟Institut Français d‟Études Andines., 36(2), 175-197.
Chávez, F. M. (2008). "La Ornitofauna de la formación Bahía Inglesa, Caldera,
Chile." (Tesis de grado - Universidad Austral de Chile).
De Muizon, C. (1978) Arctocephalus (Hydractos) lomasiensis subgen. Nov. et sp.
nov, un nouvel Otariidae du Miopliocène de Sacaco (Pèrou). Bull. Int. Fr. Et. ,
7(34), 169 -188.
De Muizon, C. (1981) Les Vertebres fossiles de la Formation Pisco (Perou). Premiére
Partie: deux nouveaux Monachinae (Phocidae: Mammalia) du Pliocene
Inferior de Sud-Sacaco. Trav. Inst. Fr. Et. Andines, 22, 1-160.
De Muizon, C. and J. DeVries (1985). "Geology and paleontology of late Cenozoic
marine deposits in the Sacaco area (Peru)." Geologische Rundschau. 74: 547-
563.
De Muizon, C. (1993) Walrus-like feeding adaptation in a new cetacean from the
pliocene of Peru. Nature, 365.
De Muizon, C. & Mcdonald, G. H. (1995) An aquatic sloth frm the Pliocene of Peru.
Nature, 375.
De Muizon, C., Mcdonald, G. H., Salas, R. & Urbina, M. (2004a) The youngest
species of the aquatic sloth Thalassocnus and a reassessment of the
relationships of the Nothrothere sloths (Mammalia: Xenarthra). Journal of
Vertebrate Paleontology., 24(2), 387-397.

21
De Muizon, C., Mcdonald, G. H., Salas, R. & Urbina, M. (2004b) The evolution of
feeding adaptations of the aquatic sloth Thalasocnus. Journal of Vertebrate
Paleontology., 24(2), 398-410.
Del Rio, C. J. (2006). "El Género Nodipecten Dall (Bivalvia, Pectinidae) en el
Neógeno de la Patagonia (Argentina)." Scielo 43: 745-756.
Emslie, S. D. & Guerra, C. (2003) A new species of penguin (Spheniscidae:
Spheniscus) and other birds from the late Pliocene of Chile. . Proceedings of
the Biological Society of Washington, 116 (2), 308–316.
Foote, M. & Miller, A. I. (2007) Principles of Paleontology, 214-215.
Fowler, C. W., A. James, et al. (1982). "Selective extinction and speciation: their
influence on the strcutture and functioning of coumminities and ecosystems."
The American Naturalist 119.
García, A. (1999) La extinción de la megafauna pleistocénica en los Andes Centrales
Argentino-Chilenos. Revista Española de Antropología Americana., 29, 9-30.
Gohlich, U. B. (2007) The oldest record of the extant penguin genus Spheniscus – a
new species from the Miocene of Peru. Acta Palaeontologica Polonica, 52 (2),
285–298.
Gregory-Wodzicki, K. M. (2000) Uplift history of the central and northern andes: a
review. Geological Society of America Bulletin., 112, 1091-1105.
Guthrie, R. D. (2006) New carbon dates link climatic change wuth human colozation
and pelistoce extinctions. Nature, 441.
Gutstein, S. C., Cozzuol, A. M., Vargas, O. A., Suárez, E. M., Schultz, L. C. &
Rubilar-Rogers, D. (2009) Patterns of skull variation of Brachydelphis
(Cetace, Odontoceti) from the Neogene of the Southeastern Pacific,. Journal
of Mammalogy., 90(2), 504-519.
Ibaraki, M. (1997) Closing of the central American seaway and Neogene Coastal
upwelling along the Pacific coast of America. Tectonophysics, 281, 99-104.
Jablonski, D. (1987) Heritability at the Species Level: Analysis of Geographic
Ranges of Cretaceous Mollusks. Science: 238:360-363.

22
Jablonski, D. and D. M. Raup (1995). "Selectivity of end-Cretaceous marine bivalve
extinctions. ." Science 268: 389-391.
Jablonski, D. (2004). "Extinction: past and present." Nature 427.
Jablonski, D. & Hunt, G. (2006) Larval Ecology, Geographic Range, and Species
Survivorship in Cretaceous Mollusks: Orgnismic versus Species-Level
Explanations. The American Naturalist, 168, 556-564.
Jablonski, D. (2008a) Extinction and the spatial dymanics of biodiversity. PNAS,
105, 11528-11535.
Jablonski, D. (2008b) Species Selection: Theory and Data. Annual Review of
Ecology and Evolution, and Systematics., 39, 501-24.
Jadwiszcak, P. (2009). "Penguin past: The current state of knowledge." Polish polar
research. 30: 3-28.
Johnson, C. N. (2002). "Determinants of loss of mammal species during the Late
Quaternary „megafauna‟ extinctions: life history and ecology, but not body
size." The Royal Society 269: 2221-2227.
Kiel, S. and S. Nielsen (2010). "Quaternary origin of the inverse latitudinal diversity
gradient among southern Chilean mollusks." Geology 38: 955-958.
Koch, L. P. & Barnosky, D. A. (2006) Late Quaternary Extinctions: State of the
Debate. Annual Review of Ecology, Evolution, and Systematics., 37, 215-50.
Krug, Z. A., Jablonski, D. & Valentine, J. W. (2008) Species–genus ratios reflect a
global history of diversification and range expansion in marine bivalves. The
Royal Society.
Krug, Z. A., Jablonski, D. & Valentine, J. W. (2009) Sigbature of the End-Cretaceous
Mass Extinction in the Modern Biota. Science, 323, 767-771.
Logan, R.F (1968) Causes, climates, and distribution of deserts. En: Brown Jr. GW
(ed) Desert Biology: 21-50. Academic Press, New York.
Long, D. J. (1993) Late Miocene and Early Pliocene fish assemblages from the north
central coast of Chile. 1. Tertiary Research, 4, 117–126.
Marocco, R. & De Muizon, C. (1988). Los vertebraods del Neogeno de la costa sur
del Perú: Ambiente Sedimentario y Condiciones de Fosilizacion., 2, 105-117.

23
McCulloch, R.D., Bentley, M.J., Purves, R.S., Hulton, N.T.J., Sudgen, D.E &
Clapperton, C.M. (2000). Climatic inferences from glacial and
paleoecological evidence of the last glacial termination, southern America.
Journal of Quaternary Science, 15: 409-417.
Mckinney, L. M. (1997) Extinction Vulnerability and Selectivity: Combining
Ecological and Paleontological Views. Annual Review of Ecology, Evolution,
and Systematics., 28, 495-516.
Nieto, M., Hortal. J., Martinez. C. & Morales. J. (2003). "Variaciones en la riqueza
de macromamíferos de España durante el neógeno". Graellsia 59: 299-317.
Nolf, D. (2002) Fossil record and paleobiography of Steindachneria (Pisces,
Gadiformes). 237, 89-95.
Payne, L. J. and S. Finnegan (2007). "The effect of geographic range on extinction
risk during background and mass extinction." PNAS.
Pimm, L. S., Jones, L. H. & Diamond, J. (1988) On the risk of extinction. The
American Naturalist, 132, 757-785.
Pujos, F. & Salas, R. (2004) A systematic reassessment and Paleogeographic review
of fossil Xenarthra from Peru., 33(2), 331-377.
Raup, M. D. (1986) Biological extinction in earth history. Science, 231, 1528-1533.
Rivadeneira, M. M. and P. A. Marquet (2007). "Selective extinction of late Neogene
bivalves on the temperate Pacific coast of South America." Paleobiology 33:
455-468.
Roberts, G. R., Flannery, F. T., Ayliffe, K. L., Yoshida, H., Olley, M. J., Prideaux, J.
G., Laslett, M. G., Baynes, A., Smith, A. M., Rhys, J. & Smith, L. S. (2001)
New Ages for the Last: Australian Megafauna: Continent-Wide Extinction
About 46.000 Years Ago. Science, 292.
Roy, K., Jablonski, D. & Valentine, J. W. (2001) Climate change, species range limits
and body size in marine bivalves. Ecology letters, 4, 366-370.
Roy, K., Jablonski, D. & Valentine, J. W. (2002) Body size and invasion succes in
marine bivalves. Ecology letters, 5, 163-167.

24
Sanchez, B., Prado, J.L., Alberdi, M.T. (2003) Paleodieta, ecología y extinción de los
gonfoterios (Proboscidea).Coloquios de Paleontologia 1:617-325.
de la Región Pampeana (Argentina)
Sodhi, N. S., D. Bickford, et al. (2008). "Measuring the Meltdown: Drivers of Global
Amphibian Extinction and Decline." Plos one 3.
Staig, F. (2006) Descripción de elasmobranquios fósiles de la formación Coquimbo
(Mioceno - Plioceno), IV Región, Chile / 27-30.
Steadman, W. D., Martin, S. P., Macphee, E. D. R., Jull, T. J. A., Mcdonald, G. H.,
Woods, A. C., Iturralde-Vinent, M. & Hodgins, L. W. G. (2005)
Asynchronous extinction of late Quaternary sloths on continents and islands.
PNA, 102.
Stucchi, M. (2002) Una nueva especie de Spheniscus (Aves: Spheniscidae) de la
formación Pisco, Perú. Boletin de la Sociedad Geologica del Perú., 94, 17-24
Stucchi, M. (2003) Los piqueros (Aves: Sulidae) de la Formación Pisco, Perú. Boletin
de la Sociedad Geologica del Perú, 95, 75-91.
Stucchi, M., Urbina, M. & Giraldo, A. (2003) Una nueva especie de Spheniscidae del
Mioceno Tardio de la Formación de Pisco, Perú. Boletin del Instituto francés
de Estudios Andinos., 32, 361-375.
Stucchi, M. & Urbina, M. (2004) Ramphastosula (Aves, Sulidae): A new genus from
the early Pliocene of the Pisco formation, Peru. Journal of Vertebrate
Paleontology., 24(4), 974-978.
Stucchi, M. (2007) Los Pinguinos de la formacion Pisco (Neógeno), Perú.
Suárez, M. E., Lamilla, J. & Marquardt, C. (2004) Peces Chimaeriformes
(Chondrichthyes, Holocephali) del Neógeno de la Formación Bahía Inglesa
(Región de Atacama, Chile). Rev. geol. Chile.
Team, R. D. C. (2010) R: A language and environment for statistical computing, edn.
R foundation for Statistical Computing.
Tsuchi, R. (2002) Neogene evolution of surface marine climate in Pacific and notes
on related events,. Revista Mexicana de Ciencias Geológicas, 19, 260-270.

25
Walsh, A. S. (2001) A new Neogene marine avian assemblage from north-central
Chile. Journal of Vertebrate Paleontology., 21(3), 484–491.
Walsh, A. S. & Hume, P. J. (2001) A new Neogene marine Avian assemblage from
north-central Chile. Journal of Vertebrate Paleontology., 21(3), 484-491.
Walsh, A. S. & Naish, D. (2002) Fossil Seals from late Neogene deposits in south
America: A new Pinniped (Carnivora, Mammalia) assemblage from Chile.
Palaeontology, 45, 821-842.
Walsh, A. S. & Suárez, E. M. (2006) New Penguin remains from the Pliocene of
Northern Chile. Historical Biology, 18(2), 115-126.
Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K. (2001) Trends, Rhythms,
and Aberrations in Global Climate 65 Ma to Present., 292.
Zinmeister, W. J. (1978) Effect of formation of the west antartic ice sheet on shallow-
water marine faunas of Chile. Antartic Journal U.S.A, 13, 25-26.
Tabla 1. Principales eventos tectónicos, climáticos, oceanográficos, asociados a los
cambios en composición faunística (moluscos), observados durante los 30
millones de años en la costa pacifica de Sudamérica.

26
Fuentes. 1) Zachos et al. (2001), 2) Heiki et al. (1983), 3) Gregory-Wodzicki (2000),
4) Tsuchi (1997), 5) Hartley (2003), 6) Inferido de (7), 7) Clapperton (1994),
8) McCulluch et al. (2000), 9) Moy et al. (2002), 10) Zinmeister (1978), 11)
Herm (1969), 12) DeVries & frassinetti (2003), 13) Martinez-Pardo (1990).
Ma Época Eventos Tectónicos Eventos Climáticos Eventos Oceanográficos Comp. faunística
0 Holoceno ENSO moderno (9) ENSO moderno (9)
0,01 Final última deglaciación (8)
1 Pleistoceno Fauna moderna (16)
2 Formación fiordos(6) Extinción masiva (16)
3 Hiperaridez (5) Glaciación Magallánica (7) Fauna fría, Chile(16)
4 Plioceno Cierre Istmo de Panamá (4)
5 Glaciación Antártica Oeste (1) Humboldt Moderno (10)
6 Surgencia costera, Chile(4)
7 Hiatus (18)
8
9
10 50% altura actual Andes (5) Glaciación Antártica Este (1)
11
12
13 Mioceno
14 "Codo de Arica"(3) Surgencia costera, Perú (4) Fauna fría, Perú (17)
15 "Optimo climático " (1) "Optimo climático"(1)
16
17
18
19
20 Fauna subtropical (16)
21
22
23
24
25 Orogenesis Andes (5)
26
27 Oligoceno
28
29
30
31 Apertura Paso Drake (1)
32
Tabla 2. Ocurrencias fósiles de vertebrados marinos del Neógeno en el Pacífico de
Sudamérica. A: Aves, P: Peces y M: Mamíferos.

27
País Latitud localidad Edad Taxa Referencia
Perú 13º42'55" Pisco Mioc-Plioceno A Stucchi, 2002
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno A Acosta & Canto 2005
Perú 13º42'55" Pisco Mioceno-Plioceno A, P y M Marocco & De Muizon 1988
Perú 16º09'25'' Arequipa Mioceno-Plioceno M De Muizon, 1978
Perú 16º09'25'' Arequipa Mioceno-Plioceno A, P y M De Muizon, 1985
Perú 16º09'25'' Arequipa Mioceno-Plioceno M De Muizon, 1981
Perú 13º42'55" Pisco Mioceno A Stucchi et al 2003
Perú 13º42'55" Pisco Mioceno-Plioceno A Stucchi 2007
Perú 13º42'55" Pisco Plioceno A Stucchi & Urbina 2004
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno A Walsh & Hume 2001
Chile 32º57'67'' Concon Mioceno-Plioceno M Valenzuela 1994
Chile 27º21'28'' Bahía Inglesa Plioceno A Walsh & Suárez 2006
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno A Acosta et al 2006
Perú 16º09'25'' Arequipa Mioceno-Plioceno A,P y M De Muizon 1983
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno A Acosta et al 2002
Perú 14º39'12'' Zamaca Mioceno-Plioceno A Acosta et al 2005
Chile 35º20'21'' Constitucion Plioceno M Canto, 2010
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno A Chavez, 2008
Chile 27º21'28'' Bahía Inglesa Mioceno A Chavez, 2007
Chile 27º21'28'' Bahía Inglesa Plioceno A Emslie, 2003
Perú 13º42'55" Pisco Mioceno A Gohlich, 2007
Perú 13º42'55" Pisco Mioceno-Plioceno A Stucchi, 2003
Perú 14º13'18'' Chilcatay Mioceno A Stucchi & DeVries 2003
Perú 13º42'55" Pisco Mioceno-Plioceno A Cheneval, 1993
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno P Long, 1993
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno P Suárez, 2004
Chile 29º58'16'' Coquimbo Mioceno-Plioceno A Acosta, 2006
Perú 4º04'50'' Piura Mioceno P Apolin, 2004
Chile 71º52'15'' Matanzas Mioceno P Nolf, 2002
Chile 29º58'16'' Coquimbo Mioceno-Plioceno P Staig, 2006
Perú 16º09'25'' Arequipa Plioceno M De Muizon, 1995
Perú 16º09'25'' Arequipa Plioceno M De Muizon, 2004
Perú 13º42'55" Pisco Mioceno M Pujos & Salas 2004
(Continuación)Tabla 2. Ocurrencias fósiles de vertebrados marinos del Neógeno en
el Pacífico de Sudamérica. A: Aves, P: Peces y M: Mamíferos.

28
País Latitud localidad Edad Taxa Referencia
Chile 27º21'28'' Bahía Inglesa Mioceno M Bianucci et al 2006
Perú 13º42'55" Pisco Plioceno M Bouetel, 2006
Perú 13º42'55" Pisco Mioceno M Gutstein, 2009
Chile 27º21'28'' Bahía Inglesa Mioceno M Gutstein, 2009
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno M Walsh, 2002
Perú 13º42'55" Pisco Plioceno M De Muizon, 1993
Perú 13º42'55" Pisco Mioceno M y P Colbert, 1994
Perú 13º42'55" Pisco Mioceno-Plioceno
A, P y
M De Muizon, 1983
Perú 13º42'55" Pisco Mioceno-Plioceno
A, P y
M De Muizon, 1985
Perú 13º42'55" Pisco Mioceno-Plioceno
A, P y
M Demere & Berta, 2005
Perú 13º42'55" Pisco Mioceno-Plioceno
A, P y
M McDonald & Muizon, 2002
Perú 13º42'55" Pisco Mioceno M Dunba & Marty, 1990
Perú 13º42'55" Pisco Mioceno-Plioceno
A, P y
M Doming & Aguilera, 2008
Chile 27º21'28'' Bahía Inglesa Mioceno-Plioceno P Doming & Aguilera, 2008
Perú 13º42'55" Pisco Plioceno M Esperante & Brand, 2008
Perú 13º42'55" Pisco Mioceno-Plioceno
A, P y
M Kraus, 1998
Tabla 3. Coeficientes de correlación entre los pares de taxa, según los patrones de
diversificación obtenidos. En todos los casos los valores fueron significativos (p <
0.05)
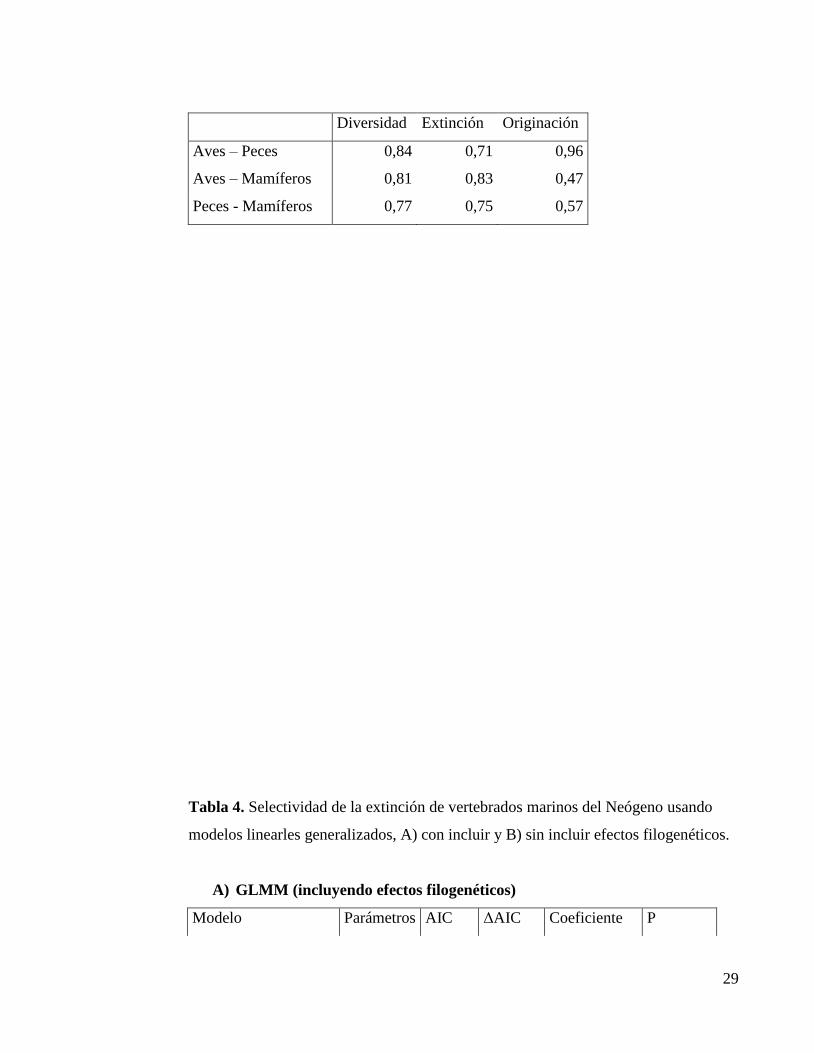
29
Diversidad Extinción Originación
Aves – Peces 0,84 0,71 0,96
Aves – Mamíferos 0,81 0,83 0,47
Peces - Mamíferos 0,77 0,75 0,57
Tabla 4. Selectividad de la extinción de vertebrados marinos del Neógeno usando
modelos linearles generalizados, A) con incluir y B) sin incluir efectos filogenéticos.
A) GLMM (incluyendo efectos filogenéticos)
Modelo Parámetros AIC ΔAIC Coeficiente P
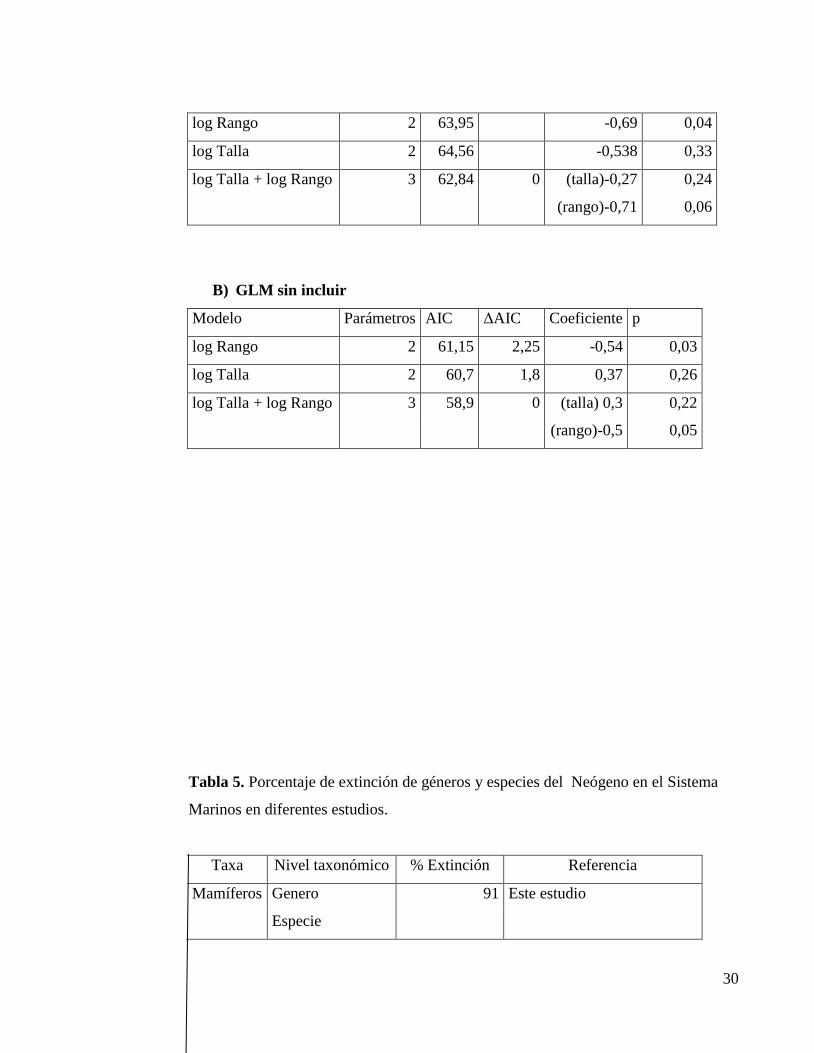
30
log Rango 2 63,95 -0,69 0,04
log Talla 2 64,56 -0,538 0,33
log Talla + log Rango 3 62,84 0 (talla)-0,27 0,24
(rango)-0,71 0,06
B) GLM sin incluir
Modelo Parámetros AIC ΔAIC Coeficiente p
log Rango 2 61,15 2,25 -0,54 0,03
log Talla 2 60,7 1,8 0,37 0,26
log Talla + log Rango 3 58,9 0 (talla) 0,3 0,22
(rango)-0,5 0,05
Tabla 5. Porcentaje de extinción de géneros y especies del Neógeno en el Sistema
Marinos en diferentes estudios.
Taxa Nivel taxonómico % Extinción Referencia
Mamíferos Genero 91 Este estudio
Especie

31
Moluscos Especie 66 Rivadeneira & Marquet 2007
Moluscos Especie 70 Allmon et al. 1993
Moluscos Especie 34 Smith & Roy 2006
Moluscos Especie 70 Stanley 1986
Moluscos Genero 64 Tod et al 2002
Moluscos Genero 85 Del Rio 2007
Aves Genero 71 Este estudio
Especie
Peces Genero 94 Este estudio
Especie
A
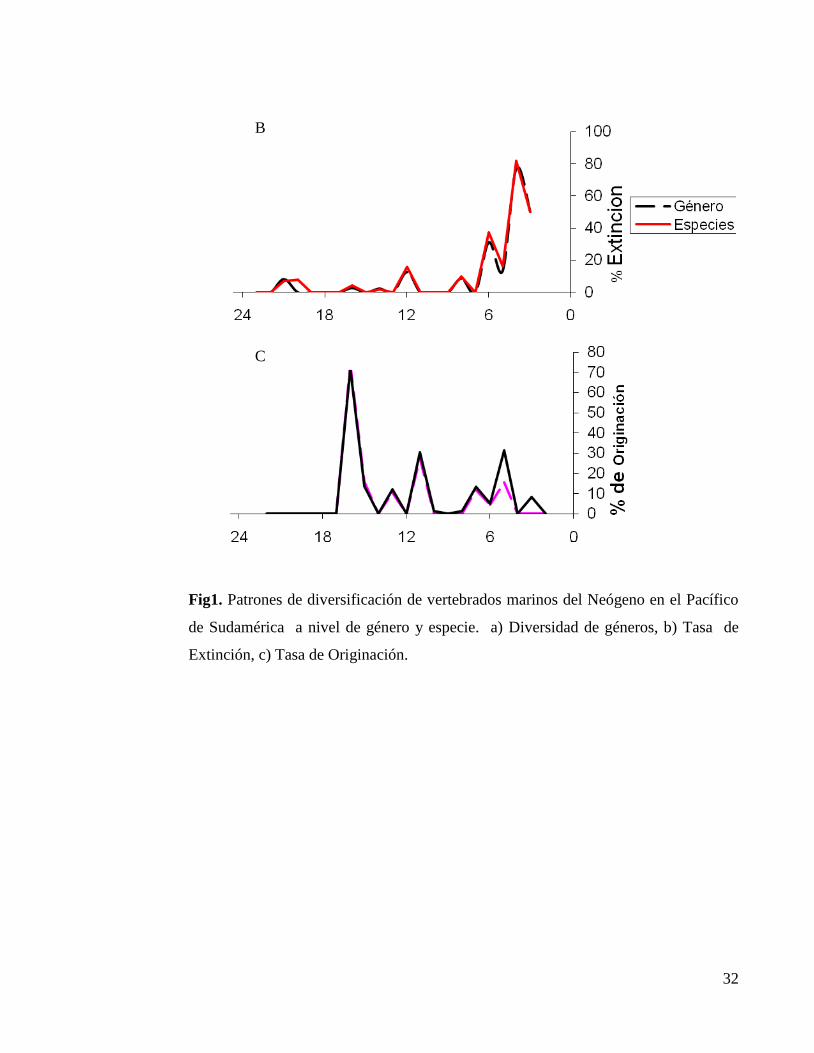
32
Fig1. Patrones de diversificación de vertebrados marinos del Neógeno en el Pacífico
de Sudamérica a nivel de género y especie. a) Diversidad de géneros, b) Tasa de
Extinción, c) Tasa de Originación.
B
C

33
Fig2. Patrones de diversificación de vertebrados marinos del Neógeno en el Pacifico
de Sudamérica por niveles de precisión: Edad formación (EF), Edad Paleobiology
Database (EPBDB) y Edad revisada (ER). Para cada criterio se estimo: Diversidad
de géneros (A, B y C), Tasa de Extinción (D, E y F) y Tasa de Originación (G, H e
I).
Millones de años

34
Fig3. Patrones de diversificación de vertebrados marinos del Neógeno en el Pacífico
de Sudamérica a nivel de taxa (Aves, mamíferos y peces): a) Diversidad de géneros,
b) Tasa de Extinción, c) Tasa de Originación.
C
A
B

35
Fig. 4. Tasa de extinción de vertebrados marinos del Neógeno en el Pacífico de
Sudamérica a nivel de taxa (peces, mamíferos y aves), comparado con los géneros
presentes actualmente.

36
Fig5. Rango promedio de géneros de vertebrados marinos del Neógeno en el Pacífico
de Sudamérica extintos y sobrevivientes.

37
Fig. 6. Cambios paleoclimáticos y oceanográficos, asociados con la diversidad de
géneros de vertebrados marinos del Neógeno en el Pacifico de Sudamérica. A)
Concentración de oxigeno, B) Nivel del Mar y C) Curva diversidad de géneros.