
Gleditisioxylon riojana nov. sp. en el Neógeno de Argentina: sus relaciones paleoambientales y...
12
Original article Gleditisioxylon riojana nov. sp. en el Neógeno de Argentina: sus relaciones paleoambientales y biogeográficas § Gleditisioxylon riojana nov. sp. in Neogene of Argentina: Palaeoenvironment and biogeography relationships Gleditisioxylon riojana nov. sp. dans le Néogène d’Argentine : relations paléoenvironnementales et paléogéographiques Leandro C.A. Martínez a,*,b , Rafael Rodríguez-Brizuela c a División Paleobotánica, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentina b División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque S/N, (1900) La Plata, Argentina c Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentina Recibido el 20 de agosto de 2010; Aceptado el 2 de diciembre de 2010 On-line 7 de julio de 2011 Resumen Se describe una nueva especie de leguminosa fósil del Neógeno en el noroeste de Argentina. La buena preservación de los tejidos y los caracteres diagnósticos presentes en el leño permitieron su clasificación a nivel específico. Gleditisioxylon riojana nov. sp. (Caesalpinoideae) presenta los siguientes caracteres diagnósticos: anillos de crecimiento marcados, porosidad semicircular, punteaduras ornamentadas, engrosa- mientos helicoidales en la pared de los vasos, placas de perforación simples, parénquima paratraqueal y radios 1-6 seriados. En base a los caracteres xilológicos presentes en el fósil estudiado se infieren las posibles condiciones climáticas de la Formación Toro Negro en donde se habría desarrollado dicho ejemplar. La presencia de este género, sumado a otras evidencias, sugeriría una nueva ruta para explicar la disyunción del actual del género Gleditsia L. con una única especie en el trópico y subtrópico de América del Sur. # 2011 Elsevier Masson SAS. All rights reserved. Palabras clave : Leguminosae; Caesalpinoideae; Madera fósil; Formación Toro Negro; Neógeno; La Rioja; Argentina Abstract A new fossil species of Leguminosae is described from Neogene of northwest of Argentina. The good preservation of tissues and the diagnostic characters present in this fossil wood let assigning it to a new species of Gleditisioxylon Müller-Stoll and Madel. Gleditisioxylon riojana nov. sp. (Caesalpinoideae) has diagnostic features such as: growth rings distinct, semi-annular porosity, vestured pits, helical thickenings, simple plates, paratracheal parenchyma and rays 1-6 seriate. The possible climatic conditions of Toro Negro Formation were inferred by the use of xylological characters presents in this fossil wood. The presence of Gleditsioxylon added to other data, suggest a new hypothesis to explain the disjunction of Gleditsia L. genus and the occurrence of a single extant species in tropical and subtropical South America. # 2011 Elsevier Masson SAS. All rights reserved. Keywords: Leguminosae; Caesalpinoideae; Fossil wood; Toro Negro Formation; Neogene; La Rioja; Argentina Résumé Une nouvelle espèce de Légumineuse fossile est décrite pour le Néogène du nord-est de l’Argentine. La bonne préservation des tissus et les caractères présents dans le bois permettent son attribution au genre Gleditisioxylon Müller-Stoll et Madel. Gleditisioxylon riojana nov. sp. (Caesalpinoideae) présente les caractères diagnostiques suivants : limites de cernes distinctes, semi-porosité, ponctuations intervasculaires ornées, Geobios 44 (2011) 461–472 § Editor correspondiente: Marc Philippe. * Autor correspondiente. Dirección correo electrónico: [email protected] (L.C.A. Martínez). 0016-6995/$ – see front matter # 2011 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.geobios.2010.12.002
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Gleditisioxylon riojana nov. sp. en el Neógeno de Argentina: sus relaciones paleoambientales y...

Original article
Gleditisioxylon riojana nov. sp. en el Neógeno de Argentina:sus relaciones paleoambientales y biogeográficas§
Gleditisioxylon riojana nov. sp. in Neogene of Argentina:Palaeoenvironment and biogeography relationships
Gleditisioxylon riojana nov. sp. dans le Néogène d’Argentine :relations paléoenvironnementales et paléogéographiques
Leandro C.A. Martínez a,*,b, Rafael Rodríguez-Brizuela c
a División Paleobotánica, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentinab División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque S/N, (1900) La Plata, Argentina
c Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentina
Recibido el 20 de agosto de 2010; Aceptado el 2 de diciembre de 2010
On-line 7 de julio de 2011
Resumen
Se describe una nueva especie de leguminosa fósil del Neógeno en el noroeste de Argentina. La buena preservación de los tejidos y loscaracteres diagnósticos presentes en el leño permitieron su clasificación a nivel específico. Gleditisioxylon riojana nov. sp. (Caesalpinoideae)presenta los siguientes caracteres diagnósticos: anillos de crecimiento marcados, porosidad semicircular, punteaduras ornamentadas, engrosa-mientos helicoidales en la pared de los vasos, placas de perforación simples, parénquima paratraqueal y radios 1-6 seriados. En base a los caracteresxilológicos presentes en el fósil estudiado se infieren las posibles condiciones climáticas de la Formación Toro Negro en donde se habríadesarrollado dicho ejemplar. La presencia de este género, sumado a otras evidencias, sugeriría una nueva ruta para explicar la disyunción del actualdel género Gleditsia L. con una única especie en el trópico y subtrópico de América del Sur.# 2011 Elsevier Masson SAS. All rights reserved.
Palabras clave : Leguminosae; Caesalpinoideae; Madera fósil; Formación Toro Negro; Neógeno; La Rioja; Argentina
Abstract
A new fossil species of Leguminosae is described from Neogene of northwest of Argentina. The good preservation of tissues and the diagnosticcharacters present in this fossil wood let assigning it to a new species of Gleditisioxylon Müller-Stoll and Madel. Gleditisioxylon riojana nov. sp.(Caesalpinoideae) has diagnostic features such as: growth rings distinct, semi-annular porosity, vestured pits, helical thickenings, simple plates,paratracheal parenchyma and rays 1-6 seriate. The possible climatic conditions of Toro Negro Formation were inferred by the use of xylologicalcharacters presents in this fossil wood. The presence of Gleditsioxylon added to other data, suggest a new hypothesis to explain the disjunction ofGleditsia L. genus and the occurrence of a single extant species in tropical and subtropical South America.# 2011 Elsevier Masson SAS. All rights reserved.
Keywords: Leguminosae; Caesalpinoideae; Fossil wood; Toro Negro Formation; Neogene; La Rioja; Argentina
Résumé
Une nouvelle espèce de Légumineuse fossile est décrite pour le Néogène du nord-est de l’Argentine. La bonne préservation des tissus et lescaractères présents dans le bois permettent son attribution au genre Gleditisioxylon Müller-Stoll et Madel. Gleditisioxylon riojana nov. sp.(Caesalpinoideae) présente les caractères diagnostiques suivants : limites de cernes distinctes, semi-porosité, ponctuations intervasculaires ornées,
Geobios 44 (2011) 461–472
§ Editor correspondiente: Marc Philippe.* Autor correspondiente.
Dirección correo electrónico: [email protected] (L.C.A. Martínez).
0016-6995/$ – see front matter # 2011 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.geobios.2010.12.002

épaississements spiralés, perforations terminales simples, parenchyme paratrachéal et rayons 1-6 sériés. D’après les caractères du xylème présentsdans le fossile étudié, on peut déduire les possibles conditions climatiques de la Formation Toro Negro lorsque cet exemplaire s’est développé. Laprésence de ce genre, plus d’autres évidences, suggèrent une nouvelle hypothèse pour expliquer la disjonction du genre Gleditsia L. et l’existenced’une unique espèce actuelle tropicale à subtropicale en Amérique du Sud.# 2011 Elsevier Masson SAS. Tous droits réservés.
Mots clés : Leguminosae ; Caesalpinoideae ; Bois fossile ; Formation Toro Negro ; Néogène ; La Rioja ; Argentina
L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472462
1. Introducción
La familia Leguminosae es de distribución mundial ycomprende alrededor de 720 géneros y 18,000 especies, siendola tercera familia de angiospermas en cuanto a su número derepresentantes (Lewis et al., 2005). Su registro fósil es amplio ydiverso, con hallazgos de polen y maderas probablementeafines a leguminosas a partir del Cretácico de Canadá, Siberia yColombia (Raven y Polhill, 1981). En el Cenozoico se confirmacon certeza la presencia de la familia, con abundantes restosasignados a las subfamilias Mimosoideae, Caesalpinoideae yPapillionoideae (Herendeen et al., 1992).
Se tienen registros de leños de leguminosas en Argentina apartir de los primeros estudios de maderas fósiles realizados enel Noroeste del país (O’Donell, 1938), en el Grupo Santa María(Neógeno) (Bossi y Palma, 1982). En Argentina los registros demaderas cenozoicas pertenecientes a leguminosas (Mimosoi-deae y Papillionoideae) son abundantes, en donde se handescripto 9 géneros (Herbst et al., 2007; Brea et al., 2008;Franco y Brea, 2008, 2010; Pujana, 2009; Martínez, 2010).
Fig. 1. Mapa y perfil estratigráfico de la localidad fosilífera. Quebrada de Troya,Location map and stratigraphic section of the fossiliferous locality. Quebrada de Tr
En este trabajo se describe una madera afín a la subfamiliaCaesalpinoideae procedente de la Formación Toro Negro,provincia de La Rioja, Argentina. La presencia de maderasfósiles en esta Formación fue previamente citada por RodríguezBrizuela y Tauber (2006) y Martínez y Rodríguez Brizuela(2007), no obstante hasta esta fecha, ningún estudio detalladoha sido dedicado a dicha temática.
2. Marco geológico
El material estudiado proviene de la Formación Toro Negro(Turner, 1964; Ramos, 1970), aflorante en el departamentoVinchina, al noroeste de la provincia de La Rioja, Argentina(Fig. 1). La Formación Toro Negro se encuentra representada porextensos afloramientos que junto con la infrayacente FormaciónVinchina (Turner, 1964; Ramos, 1970) conforman la denomi-nada Sierra de Los Colorados. La sección tipo de la unidad seencuentra en la Quebrada de la Troya, hacia el sector central de lasierra de Los Colorados, donde alcanza aproximadamente2000 metros de espesor aflorante. La estructura es homoclinal, de
La Rioja, Argentina (modificado de Rodríguez Brizuela y Tauber, 2006).oya, La Rioja, Argentina (modified from Rodríguez Brizuela and Tauber, 2006).

L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472 463
rumbo meridional y con inclinaciones promedio de 358 alnoroeste.
Litológicamente se compone de sedimentos continentalesaluviales (Rodríguez Brizuela, 2005) que son producto de lasedimentación sinorogénica vinculada al avance del frentetectónico andino (Ramos, 1999). Varios niveles piroclásticos seintercalan hacia el tope de la columna estratigráfica. Estosniveles fueron objeto de estudios destinados a ajustar la edadradimétrica de la unidad. Por un lado los estudios basados entrazas de fisión de Tabutt et al. (1989) arrojaron edadesabsolutas de 4,3 Ma, mientras que las edades K/Ar publicadaspor Ciccioli et al. (2005) arrojaron edades de 8,6 y 6,8 Ma. Lasmencionadas edades, conjuntamente con el análisis de laasociación faunística (Rodríguez Brizuela y Tauber, 2006),permiten asignarle un rango temporal Mioceno Superior-Plioceno Inferior.
Desde el punto de vista paleontológico, estos sedimentos ylos fósiles contenidos revelaron la existencia de un ricoecosistema y una intensa actividad biológica en las planiciesfluviales de la región. Una serie de estudios enfocados en lafauna fósil de la unidad así lo demostraron (Arcucci et al., 1999;De Iullis et al., 2004; Rodríguez Brizuela, 2004, Rodríguez yTauber, 2006).
3. Materiales y métodos
El material estudiado procede de la localidad Quebrada deTroya y se corresponde a una sección de leño silicificadohallado en el nivel fosilífero no 13 (Fig. 1). El fósil posee unamuy buena preservación de los tejidos. Su estudio se realizómediante cortes petrográficos y se confeccionaron láminaspetrográficas en los 3 cortes convencionales para el estudio delas maderas: corte transversal (CT), corte longitudinal radial(CLR) y corte longitudinal tangencial (CLT). Se utilizómicroscopio óptico (MO) Diaplan Leica para las observa-ciones y fotografías. El material fue también estudiadocon microscopio electrónico de barrido (MEB) PhilipsXL30 TMP, procediéndose a la fractura de pequeñosfragmentos en los 3 sentidos de corte anteriormentedescriptos, montaje en porta-especímenes y posterior meta-lizado con Au-Pt.
La terminología utilizada en la descripción anatómica es lapropuesta por el Glosario de términos de la AsociaciónInternacional de Anatomistas de Maderas (IAWA, 1989). En laspunteaduras ornamentadas se utilizó el criterio y clasificaciónpropuesto por Othani y Ishida (1976). Las medidas descritasson el promedio de 25 mediciones y entre paréntesis figuran susvalores extremos (mínimo y máximo). Las comparaciones seefectuaron con descripciones y claves de identificación demaderas de Metcalfe y Chalk (1950), Tortorelli (2009), Cozzo(1950, 1951). Además, se utilizaron las descripcionesanatómicas presentes en la base de datos de maderas Insidewood (InsideWood, 2004).
Los materiales se encuentran depositados en la ColecciónPaleontológica de la Facultad de Ciencias Exactas y Naturales yAgrimensura, Corrientes (UNNE) bajo los acrónimos CTES PBy CTES PMP.
4. Paleontología sistemática
MAGNOLIOPSIDA Cronquist, Takhtajan y Zimmermann,1966ROSIDAE Takhtajan, 1966FABALES Bromhead, 1838LEGUMINOSAE Jussieu, 1789 nom. cons. y nom. alt.CAESALPINOIDEAE DC., 1825Gleditsioxylon Müller-Stoll y Mädel, 1967Especie tipo: Gleditsioxylon columbiana Müller-Stoll yMädel, 1967
Gleditsioxylon riojana Martínez nov. spFigs. 2, 3 y 4Derivatio nominis: Por provenir de la provincia de La Rioja.Holotipo: CTES PB-12371, PMP-3007, PMP-3008 y PMP-
3009.Localidad tipo y horizonte: Quebrada de la Troya.
Formación Toro Negro. Mioceno Superior-Plioceno Inferior.Grupo Vinchina. La Rioja, Argentina.
Diagnosis: Leño secundario. Anillos de crecimientodistinguibles. Porosidad semicircular. Vasos dispuestos irregu-larmente o en cadenas oblicuas/radiales, de contornoscirculares a subcirculares. Vasos solitarios, en grupos, enseries de radiales cortos y radiales largos. Elementos de vasocortos, con punteaduras intervasculares alternas, pequeñas amedianas, con cavidades subcirculares, circulares u ovales, deabertura interna subcircular a oval y con ornamentos.Engrosamientos helicoidales en la pared interna de los vasos.Placas de perforación simples. Punteaduras radiovascularessimilares a las intervasculares, tílides. Radios homogéneos, decélulas procumbentes, multiseriados (2-6 seriados) y altos.Parénquima paratraqueal vasicéntrico confluente, muy abun-dante, dispuesto irregularmente, longitudinalmente en series.Parénquima terminal cristalífero. Fibras liberiformes enpaquetes distribuidos irregularmente, de pared delgada agruesa, con punteaduras simples.
Diagnosis. Secondary wood. Growth rings boundariesdistinct. Semi-annular porosity. Vessels in diagonal or randompattern. Vessels circular to subcircular in outline, solitary, inclusters or radials multiples. Vessel elements short. Intervesselpits small to medium, oval to circular, alternate and vestured.Helical thickenings in vessel elements. Tyloses. Perforationplates simple. Vessels ray-pits similar to intervessel pits. Rayshomocellular, procumbent cells, 2-6 seriate (Kribs Homo-geneous type II). Abundant axial parenchyma paratrachealvasicentric, conuent to banded, in longitudinal section instrands. Marginal parenchyma with prismatic crystals. Fibreswith simple pits from thin to thick-walled.
Descripción: Se conoce un fragmento de madera fósilsilicificada y picnoxílica, de 23 cm de largo y 5 cm de diámetro,con porosidad semicircular y anillos de crecimiento micro-scópicamente distinguibles por la presencia de parénquimaterminal y la reducción del tamaño de los vasos (Fig. 2(1, 2)).
Vasos: en el CT los vasos se disponen irregularmente oen cadenas oblicuas/radiales, se hallan solitarios (46 %), engrupos (32 %), en series de radiales cortos (18 %) y radiales

Fig. 2. Gleditsioxylon riojana nov. sp., CTES PB-12371. 1. CT. Vista general del leño. Porosidad semi-circular. Anillos de crecimiento y parénquima terminal(flechas). CTES PMP-3007. 2. CT. Vista general del leño. Disposición de los paquetes de fibras (flechas). CTES PMP-3007. 3. CT. Detalle de vasos y parénquimaparatraqueal. CTES PMP-3007. 4. CLT. Vista general de los vasos. CTES PMP-3009. 5. CLT. Vasos con placas de perforación simples (flechas). CTES PMP-3009. 6.CLT. Vaso con placa de perforación simple y engrosamientos helicoidales. CTES PMP-3009. 7. CLR. Punteaduras radiovasculares. CTES PMP-3008. 8. CLT. Tílides(flechas). CTES PMP-3009. 9. CLT. Tílide (flecha). CTES PMP-3009. Escalas = 200 mm (1–3), 100 mm (4–9).Gleditsioxylon riojana nov. sp. CTES PB-12371. 1. TS. General view. Wood semi ring-porous. Growth rings and marginal parenchyma (arrows). 2. TS. General view.Fibers (arrows). 3. TS. Details of vessels and paratracheal parenchyma. 4. TLS. General view of vessels. 5. TLS. Vessels with simple perforation plates (arrows) andalternate vestured pits. 6. TLS. Vessel with simple perforation plate and helical thickenings. 7. RLS. Vessel-ray pits. 8. TLS. Tylosis (arrows). 9. TLS. Tylosis (arrow).Scale bars = 200 mm (1–3), 100 mm (4–9).
L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472464

Fig. 3. Gleditsioxylon riojana nov. sp. CTES PB-12371. 1. CT. Hifas y conidios dentro del lumen de un vaso (flechas). CTES PMP-3007. 2. CLT. Hifas dentro dellumen de los vasos (flechas). CTES PMP-3009. 3. CLT. Radios. Aspecto general. CTES PMP-3009. 4. CLT. Detalle de los radios. CTES PMP-3009. 5. CLR. Radios.Aspecto general. Parénquima terminal (flechas). CTES PMP-3007. 6. CLR. Radios. Detalle de las células radiales. Punteaduras (flechas blancas) y gomas (flechasnegras). CTES PMP-3007. 7. CLT. Parénquima paratraqueal. CTES PMP-3009. 8. CLT. Parénquima seriado (flechas). CTES PMP-3009. 9. CLR. Parénquimaterminal (flechas). CTES PMP-3008. 10. CLT. Cristales. Escalas = 100 mm (1–5), 50 mm (7–9), 25 mm (6, 10).Gleditsioxylon riojana nov. sp. CTES PB-12371. 1. TS. Hyphae and conidia inside of vessel lumina (arrows). 2. RLS. Hyphae inside of vessels lumina (arrows). 3. TLS.Rays. General view. 4. TLS. Detail of the rays. 5. RLS. Rays. General view. Marginal parenchyma (arrows). 6. RLS. Rays. Detail of radial cells. Pits (white arrows) andgums (black arrows). 7. TLS. Paratracheal parenchyma. 8. TLS. Strand parenchyma (arrows). 9. RLS. Marginal parenchyma (arrows). CTES PMP-3009. 10. TLS.Crystals. Scale bars = 100 mm (1–5), 50 mm (7–9), 25 mm (6, 10).
L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472 465

Fig. 4. Gleditsioxylon riojana nov. sp. CTES PB-12371. 1. CLT. Elementos de vaso. Escala = 100 mm. 2. CLT. Punteaduras ornadas. Escala = 10 mm. 3. CLT.Punteaduras ornadas. Escala = 5 mm. 4. CLT. Verrugas en la pared interna de los vasos. Escala = 5 mm. 5. CLT. Fibras con punteaduras simples (flechas).Escala = 20 mm. 6. CLT. Parénquima paratraqueal disyunto (flechas). Escala = 50 mm. 7. CLT. Elemento de vaso con punteaduras ornadas, apéndice y placa deperforación simple. Escalas = 10 mm.Gleditsioxylon riojana nov. sp. CTES PB-12371. 1. TLS. Vessel element. Scale bar = 100 mm. 2. TLS. Vestured pits. Scale bar = 10 mm. 3. TLS. Vestured pits. Scalebar = 5 mm. 4. TLS. Warts on inner wall of vessels. Scale bar = 5 mm. 5. TLS. Fibers with simple pits. Scale bar = 20 mm. 6. TLS. Disjunt paratracheal parenchyma(arrows). Scale bar = 50 mm. 7. TLS. Vessel element with vestured pits, tail and simple perforation plate. Scale bar = 10 mm.
L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472466
largos (4 %). Su densidad es baja, con 20 vasos por mm2 (11-30). En el leño temprano están solitarios, agrupados o en seriesde múltiples cortos, mientras que en el leño tardío se observanen mayor proporción los solitarios, también hallándose, seriesde radiales largos, cortos y agrupados (Fig. 2(1–3)). El
contorno de los vasos es circular a subcircular, son elementosgrandes con un diámetro medio radial de 222.11 mm (35.2 mm-330 mm) y tangencial de 184.8 mm (52.8 mm-308 mm). Lasparedes de los vasos tienen un espesor de 11.55 mm (5.16 mm-18.92 mm; Fig. 2(1–3)). En el corte longitudinal tangencial los

L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472 467
vasos poseen un recorrido desde recto hasta sinuosoaxialmente, éstos son elementos bajos con una longitud mediade 170.89 mm (105.6 mm-228.8 mm; Fig. 2(4, 5) y 4(1)). Lasuperficie externa de la pared de los vasos posee punteadurasintervasculares alternas y ornamentadas, pequeñas a medianasy con un diámetro vertical de 4.95 mm (2.1 mm-7.7 mm), concavidades subcirculares, circulares u ovales (Fig. 4(2, 3, 6)),sus aberturas internas son subcirculares a ovales. Losornamentos que se ubican alrededor del orificio interno dela cavidad de la punteadura son del Tipo 1 (Fig. 4(3)), mientrasque los que se hallan cubriendo parcialmente la cavidadpertenecen a los Tipos 6 y 14 (Fig. 4(3)), los que cubrentotalmente la cavidad de la punteadura son del Tipo 12 (sensuOthani y Ishida, 1976; Fig. 4(2)). En superficie interna de losvasos se observan verrugas pequeñas y las punteaduraspresentan bordes lisos y sin ornamentos (Fig. 4(4)). También,algunos vasos tienen engrosamientos helicoidales delgados,los cuales se disponen a lo largo de todo el elemento de vaso,éstos son más visibles y se hallan en mayor frecuencia en losvasos estrechos (Fig. 2(6)). Los elementos de vaso tienenplacas de perforación simples (Fig. 2(5, 6) y 4(7)), horizontalesu oblicuas con apéndices vasculares cortos (Fig. 4(7)). En cortelongitudinal radial se observan punteaduras radiovascularesornadas y similares a las intervasculares, su forma esgeneralmente uniforme, disponiéndose en 2 ó 3 filas depunteaduras por radio (Fig. 2(7)). En la luz de algunos vasos seobservan tílides de paredes gruesas (Fig. 2(8, 9)), como asítambién hifas y conidios (Fig. 3(1, 2)).
Radios: en CT tienen recorrido recto en el sentido radial(Fig. 2(1, 2)). En el CLT su densidad es de 4 (2-6) radios pormm, son en su mayor parte 3-seriados (32 %), le siguen 4-seriados (28 %), 5 y 6-seriados (24 %) y 2 seriados (16 %). SonHomogéneos del Tipo II (sensu Kribs, 1935; Fig. 3(3, 4)), dedisposición al azar y sin estratificación (Fig. 3(3)). Los radiostienen una altura media de 675.82 mm (25 mm-1362.9 mm) y unancho medio de 50 mm (12.4 mm-96.1 mm). Su altura ennúmero de células es de 33 (4-65). Las células que los formanpresentan una altura media de 13.28 mm (4 mm 20 mm) y unespesor medio de 11.7 mm (4 mm-20 mm; Fig. 3(4)).
En CLR se evidencia que los radios son homocelularescompuestos exclusivamente por células procumbentes. En suinterior se observa la presencia de contenidos obscuros denaturaleza incierta, atribuidos a depósitos de gomas o resinas(Fig. 2(7) y 3(5, 6)). En las paredes horizontales y verticales delas células radiales se observa la presencia de punteadurasdiminutas (Fig. 3(6)).
Parénquima: en CT se observa el gran desarrollo delparatraqueal vasicéntrico confluente, muy abundante y dis-puesto irregularmente (Fig. 2(1, 2)). Axialmente, el parénquimase organiza en series de 4 (2-6) células, con una longitud mediade 50.77 mm (34.4 mm-99.76 mm; Fig. 3(8)). Se observa lapresencia de células parenquimáticas paratraqueales disyuntas(Fig. 4(6)) con punteaduras ornamentadas, similares a laspunteaduras intervasculares (Fig. 3(7) y 4(6)). Los anillos decrecimiento se distinguen por la presencia de parénquimaterminal, de 1 ó 2 células de ancho (Fig. 2(1–3)), con cristalesprismáticos en su interior (Fig. 3(9)).
Fibras: en el CT las fibras se hallan irregularmentedispuestas en paquetes y son libriformes (Fig. 2(2)), con paredsecundaria delgada a gruesa, con un diámetro medio radial de12.72 mm (3.44 mm-22.36 mm) y tangencial de 11.07 mm(3.44 mm-24.08 mm) (Fig. 2(1–3)); longitudinalmente las fibrasposeen extremos aguzados y en las caras radiales portanpunteaduras simples y minúsculas (Fig. 4(5)).
Comparaciones: El material estudiado es asignado a lafamilia Leguminosae por presentar caracteres tales comoporosidad semicircular, vasos solitarios, en series de radialescortos, largos y agrupados, de tamaño grande a mediano en CT,elementos de vaso cortos, placas de perforación simples,punteaduras intervasculares alternas y ornadas, radios multi-seriados, homocelulares, de células procumbentes y parén-quima paratraqueal vasicéntrico (Cozzo, 1950, 1951; Baretta-Kuipers, 1981). El leño de esta familia se caracteriza pormostrar cierto gran grado de variación, pudiéndose distinguirsus 3 subfamilias por medio de sus caracteres xilológicos(Mimosoideae, Caesalpinoideae y Papillionoideae; Cozzo,1951).
El material se diferencia de la subfamilia Papillionoideaepor carecer de elementos estratificados y poseer apéndicescortos; pero para poder distinguirlo de ciertos géneros deMimosoideae sólo puede hacerse en base a la presenciaconjunta de engrosamientos helicoidales y de radios mayores a500 mm (Cozzo, 1951).
Tomando en conjunto los caracteres presentes en el leñoestudiado (engrosamientos helicoidales en los elementos devaso, radios multiseriados altos mayores de 500 mm yparénquima exclusivamente seriado) la muestra tiene mayorafinidad con las Caesalpinoideae (Cozzo, 1950, 1951; Baretta-Kuipers, 1981). Las Caesalpinoideae en el presente constan conmás de 2250 especies, comprendidas en unos 171 géneros(Bruneau et al., 2008). En Argentina existen 16 génerosvivientes de Caesalpinoideae con aproximadamente unas80 especies, distribuidas principalmente en regiones de tipotemplado-cálido (Zuloaga y Morrone, 1999).
Para la identificación del taxón, se seleccionaron aquellosgéneros modernos que presenten la combinación total o parcialde caracteres diagnósticos tales como: engrosamientos en lapared de los vasos, tilosis, radios homogéneos desde 2- a 6-seriados, parénquima paratraqueal abundante y parénquimaterminal. La presencia de engrosamientos helicoidales en loselementos de vasos es una condición comparable con losgéneros Cercis L., Gleditsia L., Gymnocladus Lam. yZuccagnia Cav., nom. cons., y semejante a las estriacionesen la pared de los vasos de los géneros Parkinsonia L.,Zuccagnia y Senna birostris (Domb. ex Vog.) H.S. Irwin yBarneby (Metcalfe y Chalk, 1950; Cozzo, 1951; Othani yIshida, 1976). Además, tilosis como en la madera fósil sereporta para: Bauhinia L., Hymeanea L., Caesalpiniapulcherrima (L.) Sw., Gleditsia formosana Hay., y Melanox-ylon brauna Schott (Metcalfe y Chalk, 1950; Cozzo, 1951).
Entre los géneros que poseen radios homogéneos 2-seriados amultiseriados, al igual que en la nueva madera, se incluyen a:Acrocarpus R. Wight ex Arnott Ness., Amphimas Pierre exHarms in Engler et Prantl, Bauhinia, Burkea Hook., Caesalpinia,

L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472468
Ceratonia L., Cercidium Zul., Cercis, Copaifera L., CynometraL., Daniellia Benn., Detarium Juss., Dicorynia Benth.,Distemonanthus Benth., Eperua Aubl., Gleditsia, Gossweiler-odendron Harms., Gymnocladus, Hardwickia Roxb., Hymeanea,Melanoxylon Schott, Parkinsonia, Peltogyne Vogel., Schizolo-bium Vog., Schotia Jacq., Senna Mill., Trachylobium Hayne yZuccagnia (Cozzo, 1951; Metcalfe y Chalk, 1950). Abundanteparénquima paratraqueal en forma de bandas irregulares,comparables con el nuevo registro, en Afzelia quanzensis Welw.,Aldina Endl., Amphimas Pierre ex Dalla Torre y Harms.,Baikiaea Benth., Bauhinia, Campsiandra Benth., Cassia L.,Chidlowia Hoyle, Cynometra, Dicorynia, DistemonanthusBenth., Gleditsia, Haematoxylum L., Holocalyx Micheli,Koompassia Maingay ex Bentham, Peltogyne Vogel y Petlo-phorum (Vog.) Benth, mientras que la presencia de parénquimaterminal es una característica común en casi todos los géneros(Metcalfe y Chalk, 1950).
La estructura del leño de las Caesalpinoideae siguetendencias en 2 direcciones diferentes, una hacia la especia-lización de los radios (como sucede en las Mimosoideae) y otrahacia la estratificación de los elementos (al igual que en lasPapillionoideae) (Baretta-Kuipers, 1981). El ejemplar estu-diado tiene caracteres que lo alían más afín con lasmimosoideas, por su tendencia hacia la especialización delos radios (presencia de grandes radios homocelulares) ydebido a la ausencia de estructura estratificada, tal comosucede en los géneros de la tribu Caesalpinieae (Baretta-Kuipers, 1981). En dicha tribu se halla el género Gleditsia,quien comparte la mayor cantidad de caracteres con el materialen estudio.
Las descripciones hechas por Cozzo (1951) y Tortorelli(2009) sobre el género Gleditsia coinciden con los caractereshallados en el fósil en estudio tales como: porosidadsemicircular, vasos solitarios, agrupados, en series de múltiplescortos y largos, elementos de vaso de recorrido sinuoso yaxialmente cortos, con tabiques oblicuos, apéndices yengrosamientos helicoidales (en G. triacanthos L.); los radiosse presentan desde homogéneos a levemente heterogéneos,generalmente pluriseriados, raramente uniseriados (depen-diendo de las especies); el parénquima axial es abundante yvasicéntrico, desde bandeado corto hasta continuo, tambiénposeen parénquima terminal, cristales y células radiales consustancias amorfas.
El fósil estudiado presenta una fuerte correspondencia conGleditsia amorphoides (Griseb.) Taub., sin embargo esta últimase distingue de nuestro ejemplar por carecer de engrosamientoshelicoidales y tílides (Cozzo, 1951). No obstante, losengrosamientos helicoidales se han descripto para otrasespecies tales como: G. triacanthos, G. aquatica Marshall,G. japonica Lodd. Cat. ex W.H. Baxter, G. sinensis Lam.(Othani et al., 1983; InsideWood, 2004); y el desarrollo detilosis fue mencionado en G. formosana (Metcalfe y Chalk,1950).
El género Gleditsia a pesar de sus pocas especies presentauna amplia distribución, hallándose en el este de Asia(8 especies), este de América del Norte (2 especies), centroeste de América del Sur (1 especie), costa sur del Mar de
Caspio (1 especie) y en el noreste de la India (1 especie)(Schnabel et al., 2003). Gleditsia amorphoides es la únicaespecie autóctona de América del Sur, siendo un árbolmesoxerofítico que alcanza los 10 m de altura formando partedel estrato arbóreo bajo en diferentes comunidades de losbosques higrófilos o de la Selvas en las regiones del ParqueChaqueño, Mesopotamia y Yungas de Argentina, tambiénhabita en Bolivia, Paraguay, sur de Brasil y Uruguay (Burkart,1952; Carnevali, 1994; Zuloaga y Morrone, 1999; Tortorelli,2009).
Se consultaron las diagnosis de los géneros de maderasfósiles afines a la subfamilia Caesalpinoideae con el objetivo dehallar la filiación del fósil en cuestión. El resumen de lascomparaciones se halla expuesto y resumido en el Cuadro 1.Los géneros Pahudioxylon Chowdhury, Ghosh y Kazmi (1960),Brachystegioxylon Lakhanpal y Prakash (Lemoigne, 1978) yCrudioxylon Pons (1980) se diferencian del fósil aquí estudiadopor presentar parénquima paratraqueal aliforme o aliforme-confluente y radios 1 a 3 seriados. Afzelioxylon Louvet (1965),Berlinioxylon Müller-Stoll y Mädel (1967), CaesalpinioxylonSchenk (1890), Cassinium Prakash (1975), TetrapleuroxylonMüller-Stoll y Mädel (1967) y Peltophoroxylon Ramanujam(1960) Müller-Stoll y Mädel (1967) se distinguen de nuestroejemplar por presentar parénquima aliforme. Mientras queCynometroxylon (Chowdhury y Ghosh, 1946) y DialiumoxylonLakhanpal y Prakash (Lemoigne, 1978) distan del leño aquídescripto por presentar un abundante parénquima en bandastangenciales. Erithrophloeoxylon Müller-Stoll y Mädel (1967)difiere del fósil argentino por poseer estratificación como asítambién parénquima vasicéntrico aliforme, confluente ybandeado. Aulacoxylon (Combes) De Franceschi y De Ploëg(2003) se distingue por tener parénquima paratraquealvasicéntrico escaso y radios heterogéneos 1 a 3 seriados.Finalmente, Doroteoxylon Nishida, Nishida y Ohsawa (1989)difiere fundamentalmente de nuestro ejemplar por presentarradios heterogéneos, fibras septadas y estratificadas (Pujana,2009).
El género Gleditsioxylon Müller-Stoll y Mädel (1967) es elmás afín al leño en estudio por presentar: porosidad anular,vasos solitarios y en series de radiales cortos o en nidos conplacas de perforación simples; punteaduras intervascularesalternas de tamaño mediano. En los vasos pequeños presentaengrosamientos espiralados. El parénquima es paratraquealvasicéntrico y en el leño tardío es confluente en forma bandastangenciales que rodean a los vasos. Los radios sonhomogéneos y generalmente desde anchos a muy anchos(hasta 12 células de espesor), además en algunas especies seobserva la presencia de células cristalíferas. En la literaturase mencionan 3 especies de Gleditsioxylon. La especieG. columbianum Prakash y Barghoorn (1961) difiere denuestro material por presentar radios bajos, porosidad anular ycarecer de parénquima cristalífero; mientras que las especiesG. montanense Prakash, Barghoorn y Scott (1962) yG. paleojaponicum Müller-Stoll y Mädel (1967) se distinguenprincipalmente por sus anchos radios multiseriados de hasta10 ó 12 células de espesor. Las comparaciones se hallanresumidas en el Cuadro 2.
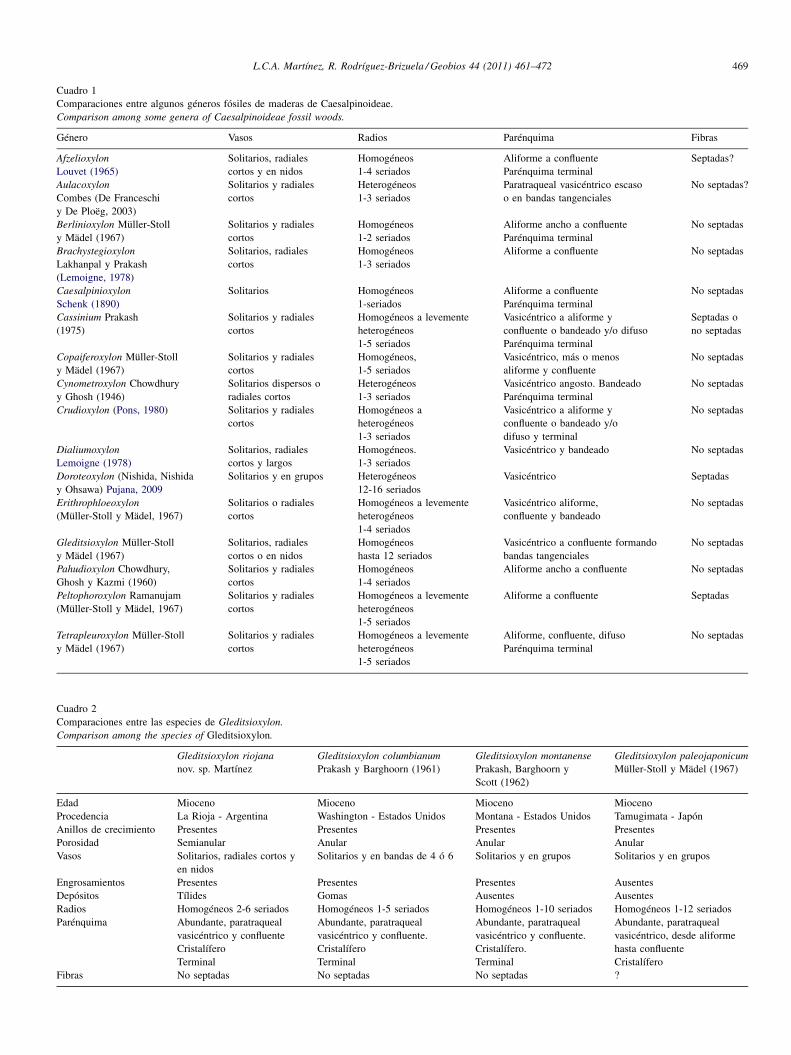
Cuadro 2Comparaciones entre las especies de Gleditsioxylon.
Comparison among the species of Gleditsioxylon.
Gleditsioxylon riojana Gleditsioxylon columbianum Gleditsioxylon montanense Gleditsioxylon paleojaponicum
nov. sp. Martínez Prakash y Barghoorn (1961) Prakash, Barghoorn yScott (1962)
Müller-Stoll y Mädel (1967)
Edad Mioceno Mioceno Mioceno MiocenoProcedencia La Rioja - Argentina Washington - Estados Unidos Montana - Estados Unidos Tamugimata - JapónAnillos de crecimiento Presentes Presentes Presentes PresentesPorosidad Semianular Anular Anular AnularVasos Solitarios, radiales cortos y
en nidosSolitarios y en bandas de 4 ó 6 Solitarios y en grupos Solitarios y en grupos
Engrosamientos Presentes Presentes Presentes AusentesDepósitos Tílides Gomas Ausentes AusentesRadios Homogéneos 2-6 seriados Homogéneos 1-5 seriados Homogéneos 1-10 seriados Homogéneos 1-12 seriadosParénquima Abundante, paratraqueal Abundante, paratraqueal Abundante, paratraqueal Abundante, paratraqueal
vasicéntrico y confluente vasicéntrico y confluente. vasicéntrico y confluente. vasicéntrico, desde aliformeCristalífero Cristalífero Cristalífero. hasta confluenteTerminal Terminal Terminal Cristalífero
Fibras No septadas No septadas No septadas ?
Cuadro 1Comparaciones entre algunos géneros fósiles de maderas de Caesalpinoideae.Comparison among some genera of Caesalpinoideae fossil woods.
Género Vasos Radios Parénquima Fibras
Afzelioxylon Solitarios, radiales Homogéneos Aliforme a confluente Septadas?Louvet (1965) cortos y en nidos 1-4 seriados Parénquima terminalAulacoxylon Solitarios y radiales Heterogéneos Paratraqueal vasicéntrico escaso No septadas?Combes (De Franceschi cortos 1-3 seriados o en bandas tangencialesy De Ploëg, 2003)Berlinioxylon Müller-Stoll Solitarios y radiales Homogéneos Aliforme ancho a confluente No septadasy Mädel (1967) cortos 1-2 seriados Parénquima terminalBrachystegioxylon Solitarios, radiales Homogéneos Aliforme a confluente No septadasLakhanpal y Prakash cortos 1-3 seriados(Lemoigne, 1978)Caesalpinioxylon Solitarios Homogéneos Aliforme a confluente No septadasSchenk (1890) 1-seriados Parénquima terminalCassinium Prakash Solitarios y radiales Homogéneos a levemente Vasicéntrico a aliforme y Septadas o(1975) cortos heterogéneos confluente o bandeado y/o difuso no septadas
1-5 seriados Parénquima terminalCopaiferoxylon Müller-Stoll Solitarios y radiales Homogéneos, Vasicéntrico, más o menos No septadasy Mädel (1967) cortos 1-5 seriados aliforme y confluenteCynometroxylon Chowdhury Solitarios dispersos o Heterogéneos Vasicéntrico angosto. Bandeado No septadasy Ghosh (1946) radiales cortos 1-3 seriados Parénquima terminalCrudioxylon (Pons, 1980) Solitarios y radiales Homogéneos a Vasicéntrico a aliforme y No septadas
cortos heterogéneos confluente o bandeado y/o1-3 seriados difuso y terminal
Dialiumoxylon Solitarios, radiales Homogéneos. Vasicéntrico y bandeado No septadasLemoigne (1978) cortos y largos 1-3 seriadosDoroteoxylon (Nishida, Nishida Solitarios y en grupos Heterogéneos Vasicéntrico Septadasy Ohsawa) Pujana, 2009 12-16 seriadosErithrophloeoxylon Solitarios o radiales Homogéneos a levemente Vasicéntrico aliforme, No septadas(Müller-Stoll y Mädel, 1967) cortos heterogéneos confluente y bandeado
1-4 seriadosGleditsioxylon Müller-Stoll Solitarios, radiales Homogéneos Vasicéntrico a confluente formando No septadasy Mädel (1967) cortos o en nidos hasta 12 seriados bandas tangencialesPahudioxylon Chowdhury, Solitarios y radiales Homogéneos Aliforme ancho a confluente No septadasGhosh y Kazmi (1960) cortos 1-4 seriadosPeltophoroxylon Ramanujam Solitarios y radiales Homogéneos a levemente Aliforme a confluente Septadas(Müller-Stoll y Mädel, 1967) cortos heterogéneos
1-5 seriadosTetrapleuroxylon Müller-Stoll Solitarios y radiales Homogéneos a levemente Aliforme, confluente, difuso No septadasy Mädel (1967) cortos heterogéneos Parénquima terminal
1-5 seriados
L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472 469

L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472470
5. Discusión
5.1. Inferencias ambientales
En base a las características anatómicas y dendrológicas seobserva que Gledisioxylon riojana nov. sp. posiblementehabría crecido en condiciones subtropicales, bajo un régimende estacionalidad marcada y abundante disponibilidad de agua.La estacionalidad estaría evidenciada por la presencia deanillos de crecimiento distinguibles, los cuales se formanante variaciones ambientales (temperatura, luz y agua;Creeber, 1977).
Además se calcularon los índices de vulnerabilidad ymesomorfía propuestos por Carlquist (1977). El Índice deVulnerabilidad (IV) es usado para indicar el grado dexeromorfismo o mesomorfía que presentaría una planta. Unavulnerabilidad baja de 1.0 (quizás 0.1-0.5) podría indicar unalto grado de xeromorfismo y el límite inferior de la verdaderamesomorfía podría ser cercano a 3.0 (Carlquist, 1977). El IVcalculado de Gledisioxylon riojana nov. sp. es de 9.24. ElÍndice de Mesomorfía (IM) es otra medida usada, donde losvalores de mesomorfía por debajo de 30.0 indicarían xer-omorfía (Carlquist, 1977) y por arriba de 1000 se observa enespecies mesofíticas que crecen en el sotobosque de las selvassubecuatoriales (Carlquist, 1982). El IM calculado enGledisioxylon riojana nov. sp. es de 1579.
El resultado de estos índices indicarían que Gledisioxylonriojana nov. sp. posiblemente fue una planta netamentemesofítica que se habría desarrollado bajo un buen régimenhídrico en condiciones cálidas. Estos resultados coinciden conlas características ecológicas en las que crece en la especieactual Gleditsia amorphoides y con las condiciones paleoe-cológicas inferidas a partir de los datos aportados por losestudios sedimentológicos en la Formación Toro Negro(Rodríguez Brizuela y Tauber, 2006).
5.2. Sobre el género Gleditsia L. y su distribución
En África es donde actualmente se encuentran mejorrepresentadas las Leguminosae, por ello se presume que podríatratarse del área dónde se sucedió la primera radiación ydiversificación de esta familia. Las relaciones entre los gruposde leguminosas indican que la familia evolucionó durante elCenozoico en África, Sudamérica y Madagascar; dispersándosea partir de estas áreas (Raven y Polhill, 1981). En estas mismasregiones geográficas es donde principalmente las caesalpinói-deas son abundantes (América del Sur, África tropical y sudestede Asia), también con un origen presumible en estas áreastropicales o subtropicales en el Cretácico Superior (Maas-trichtiano; Cowan, 1981; Raven y Polhill, 1981).
Se considera que uno de los géneros de Caesalpinoideae másantiguos es Gleditsia, esto es debido a su simple y atípicaorganización floral (Dickison, 1981; Polhill y Vidal, 1981). Losestudios filogenéticos ubican al género Gleditsia dentro delClado Umtiza siendo éste uno de los más antiguos dentro de lascaesalpinóideas, con un ‘‘crown node’’ en el Paleoceno tardío(56.5 Ma.) (Herendeen et al., 2003; Bruneau et al., 2008).
Siendo así, la antigüedad del clado Umtiza podría explicar ladisyunción continental que se observa a nivel genérico, comoasí también la vicarianza que presenta en la actualidad el géneroGleditsia (Bruneau et al., 2008).
En cuanto al registro fósil, existe evidencia material de estelinaje posterior al Eoceno, con fósiles semejantes a Gleditsia apartir del Oligoceno de América del norte (Herendeen et al.,1992; Schnabel et al., 2003). Fósiles asignables fidedignamentetanto a Gleditsia como a Gymnocladus son los hallados entre elMioceno y Plioceno de América del Norte (Prakash yBarghoorn, 1961; Axelrod, 1992, Wheeler y Baas, 1992) yAsia (Hsü, 1983; Guo y Zhou, 1992; Tao, 1992).
Basados en la evidencia material y en datos moleculares,Schnabel y Wendel (1998) y Schnabel et al. (2003) proponen unorigen Asiático para Gleditsia. Además, dichos estudiosmoleculares también determinan que la especie sudamericanaGleditsia amorphoides es la más basal dentro del clado,perteneciendo a un linaje hermano del género, que habríadivergido rápidamente a partir de su centro de origen. Gleditsiaamorphoides, posee además una distribución disyunta enrelación a las demás especies, y sus caracteres morfológicostambién la ubican como la más basal de las especies vivientes.Haciendo uso de estos datos, estos autores plantean queGleditsia amorphoides divergió a partir de un ancestro asiáticodispersándose, ya sea a través del océano Pacífico, o por lamigración del taxón cruzando América del Norte parafinalmente alcanzar América del Sur. En la última hipótesissugieren que las especies ancestrales de América del Norte seextinguieron y los representantes actuales que hoy viven allíprovendrían de Asia, a partir de una segunda radiación delgénero (Schanabel y Wendel, 1998; Schnabel et al., 2003).
Sin embargo una hipótesis no contemplada por estos autores,podría ser una situación contraria, con un probable origenSudamericano del género, ya que es allí donde habita la especiemás basal: Gleditsia amorphoides. La contraparte de estapropuesta es que se cuenta con poca evidencia de fósiles deGleditsia en América del Sur, pero esto podría deberse a la faltade datos, ya que las floras fósiles Cenozoicas en el neotrópicode Sudamérica han sido poco estudiadas. Como evidencia afavor se cuenta con maderas del Oligoceno de Patagonia,pertenecientes a Doroteoxylon (Nishida, Nishida y Ohsawa,1989) las cuales presentan una fuerte correspondencia conGleditsia y Robinia L. (Pujana, 2009).
Otro dato a destacar es la distribución actual de los génerosde Caesalpinoideae pertenecientes al Clado Umtiza, dónde apesar de su viacariancia, Tetrapterocarpon Humbert, UmtizaT.R. Sim, Arcoa Urban y Gleditsia son tropicales osubtropicales de distribución Gondwanica. A su vez el géneroUmtiza está presente sólo en África y es el más próximofilogenéticamente a los géneros Gleditsia y Gymnocladus(Herendeen et al., 2003; Bruneau et al., 2008).
Así, teniendo en cuenta estos datos podría entoncespostularse una nueva hipótesis sobre un posible origenSudamericano del género Gleditsia, quizás en el Paleógeno apartir de ancestros filiados con la especie actual Gleditsiaamorphoides. Estos antecesores habrían partido desde Américadel Sur, migrado por América del Norte hasta alcanzar Asia.

L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472 471
Extinguiéndose luego las especies norteamericanas, y a partirde una posterior diversificación de las especies asiáticas(probablemente ocurrida durante el Neógeno), éstas habríanvuelto a radiar y colonizar América del Norte. Sin embargo,para que esta hipótesis fuere justificada se necesitarán aún másestudios detallados sobre la biogeografía, el registro y origen delas Leguminosae, como así también un mayor conocimientoacerca de las floras fósiles neotropicales de Sudamérica yÁfrica.
Agradecimientos
Los autores agradecen al Dr. Rafael Herbst habernossugerido el estudio del material, como así también a las Dras.Analía Artabe y Georgina del Fueyo por sus sugerencias ycomentarios al manuscrito. A Nicole Tavil por la ayuda con elfrancés, como así también a Dr. Sergio Cevallos-Ferriz y unrevisor anónimo que mejoraron considerablemente nuestrotrabajo. Este estudio fue desarrollado bajo el marco de losproyectos ‘‘Floras e Invertebrados del Neógeno del Noroeste deArgentina: Taxonomía, Paleoasociaciones y Bioestratigrafía’’(PICTO 2007-124) y ‘‘Paleofloras, palinoestratigrafía ypalinofacies de formaciones Neógenas de los valles Calcha-quíes’’ (PI 17/F161. Res. 774/07), ambos otorgados a la Dra.Luisa. M. Anzótegui.
Referencias
Arcucci, A.B., Tambussi, C.P., Pardiñas, U.F., Petrulevicius, J.F., 1999. Nuevosrestos de vertebrados en el terciario superior del noroeste de La Rioja,Argentina. 14 Jornadas Argentinas de Paleontología de Vertebrados. Ame-ghiniana 36, 95–96.
Axelrod, D.I., 1992. Climatic pulses, a major factor in legume evolution. In:Herendeen, P.S., Dilcher, D.L. (Eds.), Advances in Legume Systematics,Part 4, The fossil record. Royal Botanic Gardens, Kew, pp. 259–279.
Baretta-Kuipers, T., 1981. Wood anatomy of Leguminosae: its relevance toTaxonomy. In: Polhill, R.M., Raven, P.H. (Eds.), Advances in LegumeSystematics. Part 2. Royal Botanical Garden, Kew, pp. 677–705.
Bossi, G.E., Palma, R.M., 1982. Reconsideración de la estratigrafía del valle deSanta María, provincia de Catamarca, Argentina. Actas del V CongresoLatinoamericano de Geología 1, 155–172.
Brea, M., Zamuner, A.B., Matheos, S.D., Iglesias, A., Zucol, A., 2008. Fossilwood of the Mimosoideae from the early Paleocene of Patagonia,Argentina. Alcheringa 32, 427–441.
Bruneau, A., Mercure, M., Lewis, G.P., Herendeen, P., 2008. Phylogeneticpatterns and diversification in the caesalpinioid legumes. Botany 86, 697–
718.Burkart, A., 1952. Las Leguminosas argentinas. Silvestres y cultivadas. Acme
Agency, Buenos Aires.Carlquist, S., 1977. Wood Anatomy of Onagraceae: additional species and
concepts. Annals of the Missouri Botanical Garden 64, 627–637.Carlquist, S., 1982. Wood Anatomy of Illicium (Illiciaceae): phylogenetic,
ecological, and functional interpretations. American Journal of Botany 69,1587–1598.
Carnevali, R., 1994. Fitogeografía de la Provincia de Corrientes. Gobierno de laProvincia de Corrientes - INTA. Edición del autor, Corrientes.
Ciccioli, P.L., Limarino, C.O., Marenssi, S.A., 2005. Nuevas edades redimé-tricas para la Formación Toro Negro en la Sierra de Los Colorados, SierrasPampeanas Noroccidentales, provincia de La Rioja. Revista de la Asocia-ción Geológica Argentina 60, 251–254.
Chowdhury, F.N.I., Ghosh, S.S., 1946. On the anatomy of Cynometroxylonindicum gen. and sp. nov. a fossil dicotyledoneous wood from Nailalung,Assam. Proc. Natl. Inst. Sci. India 12, 435–447.
Chowdhury, F.N.I., Ghosh, S.S., Kazmi, M.H., 1960. Pahudioxylon bankurensisgen. and sp. nov. a fossil wood from the Miocene bed of Bankura District,West Bengal, India. Proceedings of the Natural Institute for Sciences ofIndia 26, 22–28.
Cowan, R.S., 1981. Caesalpinoideae. In: Polhill, R.M., Raven, P.H. (Eds.),Advances in Legume Systematics. Part 1. Royal Botanic Gardens, Kew, pp.57–64.
Cozzo, D., 1950. Anatomía del leño secundario del las Leguminosas Papilio-noideas Argentinas silvestres y cultivadas. Instituto Nacional de Investiga-ción de las Ciencias Naturales y Museo Argentino de Ciencias NaturalesBernardino Rivadavia. Ciencias Botánicas 1, 222–361.
Cozzo, D., 1951. Anatomía del leño secundario del las Leguminosas Mimo-soideas y Cesalpinoideas Argentinas silvestres y cultivadas. InstitutoNacional de Investigación de las Ciencias Naturales y Museo Argentinode Ciencias Naturales Bernardino Rivadavia. Ciencias Botánicas 2, 63–146.
Creeber, G.T., 1977. Tree rings: a natural data-storage system. BiologicalReviews 52, 349–383.
De Franceschi, D., De Ploëg, G., 2003. Origine de l’ambre des faciès sparna-ciens (Éocène inférieur) du Bassin de Paris : le bois de l’arbre producteur.Geodiversitas 25, 633–647.
De Iullis, G., Ré, G.H., Vizcaino, S.F., 2004. The Toro Negro Megatheriinae(Mammalia, Xenarthra): a new species of Pyramiodontherium and a reviewof Plesiomegatherium. Journal of Vertebrate Paleontology 24, 214–227.
Dickison, W.C., 1981. The evolutionary relationships of the Leguminosae. In:Polhill, R.M., Raven, P.H. (Eds.), Advances in Legume Systematics. Part 1.Royal Botanic Gardens, Kew, pp. 35–54.
Franco, M.J., Brea, M., 2008. Leños fósiles de la Formación Paraná (MiocenoMedio), Toma Vieja, Paraná, Entre Ríos, Argentina: registro de bosquesestacionales mixtos. Ameghiniana 45, 699–717.
Franco, M.J., Brea, M., 2010. Microlobiusxylon paranaensis gen. and sp. nov.(Fabaceae, Mimosoideae) from the Pliocene-Pleistocene of Ituzaingó For-mation, Paraná Basin, Argentina. Revista Brasileira de Paleontologia 13,103–114.
Herbst, R., Brea, M., Crisafulli, A., Gnaedinger, S., Lutz, A.I., Martínez,L.C.A., 2007. La Paleoxilología en la Argentina. Historia y desarrollo,Asociación Paleontológica Argentina. Publicación Especial 11. Ameghini-ana 508 aniversario 57–71.
Guo, S.X., Zhou, Z.K., 1992. The megafossil legumes from China. In: Here-ndeen, P.S., Dilcher, D.L. (Eds.), Advances in legume systematics, Part 4,the fossil record. Royal Botanic Gardens, Kew, pp. 207–223.
Committee, I.A.W.A., 1989. IAWA list of microscopic features for hardwoodidentification. International Association of Wood Anatomists. Bulletin n. s.10, 219–332.
Herendeen, P.S., Crepet, W.L., Dilcher, D.L., 1992. The fossil history of theLeguminosae: phylogenetic and biogeographic implications. In: Herendeen,P.S., Dilcher, D.L. (Eds.), Advances in Legume Systematics, part 4. Thefossil record. Royal Botanic Gardens, Kew, pp. 303–316.
Herendeen, P.S., Lewis, G.P., Bruneau, A., 2003. Floral Morphology inCaesalpinioid Legumes: Testing the Monophyly of the ‘‘Umtiza Clade’’.International Journal of Plant Sciences. 164 (5 Supplement: Flowers:Diversity, Development, and Evolution) S393–S407.
Hsü, J., 1983. Late Cretaceous and Cenozoic vegetation in China, emphasizingtheir connections with North America. Annals of the Missouri BotanicalGarden 70, 490–508.
InsideWood, 2004-onwards. Publicado en Internet. http//insidewood/lib.nc-su.edu/
Kribs, D.A., 1935. Salient Lines of Structural Specialization in the Wood Raysof Dicotyledons. Botanical Gazette 96, 547–557.
Lemoigne, Y., 1978. Flores Tertiaires de la Haute Vallée de l’Omo (Ethiopie).Palaeontographica B 165, 80–157.
Lewis, G., Schrire, B., Mackinder, B., Lock, M. (Eds.), 2005. Legumes of theWorld. Royal Botanic Garden, Kew.
Louvet, P., 1965. Sur une nouvelle légumineuse fossile du Tinrhert, Afzelioxylonkiliani n. gen. n. sp. Comptes rendus du 90ème Congrès national des Sociétéssavantes, Section Sciences Paris 2, 317–332.

L.C.A. Martínez, R. Rodríguez-Brizuela / Geobios 44 (2011) 461–472472
Metcalfe, C.R., Chalk, L., 1950. Anatomy of the Dicotyledons. ClarendonPress, Oxford.
Martínez, L.C.A., 2010. Prosopisinoxylon anciborae nov. gen. and sp. (Legu-minosae, Mimosoideae) from the Late Miocene, Chiquimil Formation(Santa María Group) Catamarca, Argentina. Review of Palaeobotany andPalynology 158, 262–271.
Martínez, L.C.A., Rodríguez Brizuela, R., 2007. Madera de Fabaceae de laFormación Toro Negro (
?
Mioceno Superior?.) de La Rioja, Argentina.Reunión Anual de Comunicaciones de la Asociación PaleontológicaArgentina, Corrientes. Ameghiniana 44, 81–82.
Müller-Stoll, W.R., Mädel, E., 1967. Die Fossilen Leguminosen-Hölzer. EineRevision der mit Leguminosen verglichenen fossilen Hölzer und Beschrei-bungen älterer und neuer Arten. Palaeontographica B 119, 95–174.
Nishida, M., Nishida, H., Ohsawa, T., 1989. Comparison of petrified woodsfrom the Cretaceous and Tertiary of Antarctica and Patagonia. Proceedingsof the National Institute of Polar Research Symposium on Polar Biology 2,198–212.
O’Donell, C., 1938. Troncos y ramas fósiles de dicotiledóneas en el Araucar-iano de Tiopunco (Departamento de Tafí, provincia de Tucumán).Universidad Nacional de Tucumán, Cuadernos de Minería y Geología1, 26–29.
Othani, J., Ishida, S., 1976. Study on pits of wood cells using scanning electronmicroscopy. Research Bulletins of the College Experiment of Forests 33,407–435.
Othani, J., Meylan, B.A., Butterfield, B.G., 1983. Occurrence of warts in thevessel elements and fibres of New Zealand woods. New Zealand Journal ofBotany 21, 359–372.
Polhill, R.M., Vidal, J.E., 1981. Caesalpinieae. In: Polhill, R.M., Raven, P.H.(Eds.), Advances in Legume Systematics, Part 1. Royal Botanic Gardens,Kew, pp. 81–96.
Pons, D., 1980. Les bois fossiles du Tertiaire supérieur de la région deToluviejo-Corozal (Département de Sucre, Colombie). Comptes Rendusdu 105ème Congrès national des Sociétés savantes, Science I 163–182.
Prakash, U., 1975. Fossil woods from the Siwalik beds of Himachal Pradesh,India. Palaeobotanist 22, 192–210.
Prakash, U., Barghoorn, E.S., 1961. Miocene fossil woods from the Columbiabasalts of Central Washington. Journal of the Arnold Arboretum 42,165–203.
Prakash, U., Barghoorn, E.S., Scott, A.C., 1962. Fossil Wood of Robinia andGleditsia from the Tertiary of Montana. American Journal of Botany 49,692–696.
Pujana, R.R., 2009. Fossil woods from the Oligocene of southwestern Patagonia(Río Leona Formation). Atherospermataceae, Myrtaceae, Leguminosae andAnacardiaceae. Ameghiniana 46, 523–535.
Ramos, V., 1970. Estratigrafía y estructura del terciario en la sierra de LosColorados (provincia de La Rioja) República Argentina. Revista de laAsociación Geológica Argentina 25, 359–382.
Ramos, V., 1999. Los depósitos sinorogénicos terciarios de la región andina. In:Caminos, R. (Ed.), Geología Argentina, 29. Anales del Instituto deGeología y Recursos Minerales de Buenos Aires, pp. 651–682.
Raven, P.H., Polhill, R.M., 1981. Biogeography of Leguminosae. In: Polhill,R.M., Raven, P.H. (Eds.), Advances in Legume Systematics, Part 1. RoyalBotanic Gardens, Kew, pp. 27–34.
Rodríguez Brizuela, R., 2004. Registro de Vulturidae (Aves, Ciconiiformes) enel Neógeno de la provincia de La Rioja, Argentina. Revista del MuseoArgentino de Ciencias Naturales. Nueva Serie 6, 307–311.
Rodríguez Brizuela, R., 2005. Secuencias aluviales dominadas por variacioneshidrológicas: un particular caso en el Neógeno de la provincia de La Rioja,Argentina. XVI Congreso Geológico Argentino. Actas CD-ROM, artículo153.
Rodríguez Brizuela, R., Tauber, A., 2006. Estratigrafía y mamíferos fósiles de laFormación Toro Negro (Neógeno), departamento Vinchina, noroeste de laprovincia de La Rioja, Argentina. Ameghiniana 43, 257–272.
Schenk, A., 1890. Abteilung: Paläophytologie II. In: von Zittel, K.A.,Schimper,W.P., Schenk, W. (Eds.), Handbuch der Paläontologie. von R. Oldenbourg,München, Leipzig.
Schnabel, A., Wendel, J.F., 1998. Cladistic biogeography of Gleditsia (Legu-minosae) based on ndhF and rpl16 chloroplast gene sequences. AmericanJournal of Botany 85, 1753–1765.
Schnabel, A., McDonel, P.E., Wendel, J.F., 2003. Phylogenetic relationships inGleditsia (Leguminosae) based on ITS sequences. American Journal ofBotany 90, 310–320.
Tabutt, K.D., Naeser, C.W., Jordan, T.E., Cerveny, P.F., 1989. New fission trackages of Mio-Pliocene tuffs in the Sierras Pampeanas and Precordillera ofArgentina. Revista de la Asociación Geológica Argentina 44, 408–419.
Tao, J.R., 1992. The Tertiary vegetation and ora and oristic regions in China.Acta Phytotaxonomica Sinica 30, 25–43.
Tortorelli, L., 2009. Maderas y Bosques Argentinos, 2 da. Edición. OrientaciónGráfica Editora, Buenos Aires.
Turner, J.C., 1964. Descripción Geológica de la Hoja 15 C, Vinchina. Provincia deLa Rioja. Boletín de la Dirección Nacional de Geología y Minería 100, 1–81.
Wheeler, E., Baas, P., 1992. Fossil wood of the Leguminosae: a case study inxylem evolution and ecological anatomy. In: Herendeen, P.S., Dilcher, D.L.(Eds.), Advances in legume systematics, part 4, the fossil record. RoyalBotanic Gardens, Kew, pp. 281–301.
Zuloaga, F.O., Morrone, O., 1999. Catálogo de las Plantas Vasculares de laArgentina. Dicotyledoneae 2. Monographs in Systematic Botany from theMissouri Botanical Garden.




















