
Prospects for aminoacyl-tRNA synthetase inhibitors as new antimicrobial agents
Upload
independentCategory
view
0download
0
The Plant Cell, Vol. 2, 925-939, September 1990 O 1990 American Society of Plant Physiologists
Nuclear Factors lnteract with Conserved A/T-Rich Elements Upstream of a Nodule-Enhanced Glutamine Synthetase Gene from French Bean
Brian G. forde,‘ Jacqueline Freeman, Jane E. Oliver,’ and Manuel Pineda3
Biochemistry and Physiology Department, AFRC lnstitute of Arable Crops Research, Rothamsted Experimental Station, Harpenden AL5 2JQ, United Kingdom
The gln-y gene, encoding the y subunit of glutamine synthetase in French bean (Phaseolus vugaris), is strongly induced during nodule development. We have determined the nucleotide sequence of a 1.3-kilobase region at its 5‘ end and have identified severa1 sequences common to the promoter regions of late nodulin genes from other legume species. The fi’-flanking region was analyzed for sequence-specific interactions with nuclear factors from French bean. A factor from nodules (PNF-1) was identified that binds to multiple sites between -860 and -154, and a related but distinct factor (PRF-1) was detected in extracts from uninfected roots. PNF-1 and PRF-1 bound strongly to a synthetic oligonucleotide containing the sequence of an A/T-rich 21-base pair imperfect repeat found at positions -516 and -466. The same factors also had a high affinity for a protein binding site from a soybean leghemoglobin gene and appeared to be closely related to the soybean nodule factor NAT2, which binds to A/T- rich sequences in the lbcs and nodulin 23 genes [Jacobsen et al. (1990). Plant Cell2, 85-94]. Comparison of NAT2/ PNF-1 binding sites from a variety of nodulin genes revealed the conservation of the short consensus core motif TATTTWAT, and evidence was obtained that this sequence is important for protein recognition. Cross-recognition by PNF-1 of a protein binding site in a soybean seed protein gene points to the existence of a ubiquitous family of factors with related binding affinities. Our data suggest that PNF-1 and PRF-1 belong to an evolutionarily conserved group of nuclear factors that interact with specific A/T-rich sequences in a diverse set of plant genes. We consider the possible role of these factors in coregulating the expression of gh- r and other late nodulin genes.
INTRODUCTION
The development of an effective, nitrogen-fixing symbiosis in root nodules of the Leguminosae depends on a complex series of interactions between the host plant and the invading rhizobia. The plant and the Rhizobium each re- spond by synthesizing a group of proteins (termed nodu- lins) that are characteristic of the symbiotic state (Morrison et al., 1988). There are at least 20 to 30 plant-specified nodulins, of which the best characterized and most abun- dant are the leghemoglobins, whose function is to facilitate O2 diffusion to the bacteroids. A second major adaptation is the accumulation in nodules of high levels of glutamine synthetase (GS; EC 6.3.1.2), the plant enzyme catalyzing the first step in the assimilation of the ammonia that is generated by the nitrogen-fixing bacteroids (Atkins, 1987).
In developing root nodules of French bean, the dramatic
’ To whom correspondence should be addressed. * Current address: Clinical Research Centre, Harrow, Middlesex, United Kingdom. Current address: Departamento de Bioguimica, Biologia Molec-
ular y Fisiologia, Facultad de Ciencias, 14071 -Córdoba, Spain.
increase in nodule GS activity that begins 1 O days to 12 days after infection (Lara et al., 1983) is mainly the result of the induction of a single GS gene, gln-y, one of five homologous GS genes in the French bean genome (see Forde and Cullimore, 1989). Work with chimeric gene fusions has shown that a 2-kb fragment from the 5’- flanking region of gln-y is able to direct the appropriate nodule-enhanced pattern of expression in transgenic plants of the forage legume Lotus corniculatus (Forde et al., 1989).
We are interested in the regulatory mechanisms by which gln-y and other nodulin genes are able to respond to the symbiotic interaction with Rhizobium. Functional analysis of the upstream region of the soybean leghemo- globin c3 (/bc3) gene has revealed both positive and nega- tive regulatory elements and a promoter-proximal se- quence required for nodule-specific expression (Stougaard et al., 1987, 1990). A nodule-specific nuclear factor from soybean was found to recognize short A/T-rich sequences in the /bcs promoter (Jensen et al., 1988), and related factors were identified in nodules and other tissues of

926 The Plant Cell
Sesbania roslrata and alfalfa (Metz et al., 1988). The same soybean factor also has binding sites in a second late nodulin gene, N23 (Jacobsen et al., 1990), but its function as a frans-acting regulatory factor has still not been established.
Although activation of the gln-y and leghemoglobin genes occurs at about the same stage oF nodule devel- opment (Lara et al., 1983; Bennett et al., 1989), there is evidence that the regulatory mechanisms may not be identical. In soybean, it has been found that when nodule development is disrupted by growing plants in argon: oxygen mixtures (Hirel et al., 1987), or by use of a nifA mutant of Rhizobium (Studer et al., 1987), there are differ- ential effects on the expression of leghemoglobin and GS genes; in French bean, it has been demonstrated that gln-y expression is not confined to the root nodule (Bennett et al., 1989).
In the present paper, we report the nucleotide sequence of the 5’-flanking region of the gln-y gene and the results of an analysis of sequence-specific interactions between this region and nuclear factors from French bean roots and nodules. These studies have provided evidence for the presence of a nuclear factor in nodules and a related factor in roots that bind in a sequence-specific manner to multiple A/T-rich sites in the upstream region of gln-y. The relationship between these factors and previously reported factors that bind t o A/T-rich sequences in nodulin (and non-nodulin) genes is examined and their possible function in regulating gln-y expression is discussed.
RESULTS
Nucleotide Sequence Analysis of the gh-r Gene
Figure 1 shows the nucleotide sequence of the 5‘-flanking region of gln-7 and the first 400 bp of the transcribed por- tion of the gene. Over the region of overlap with pcGS-71, a full-length cDNA clone for the y subunit of GS in French bean (Bennett et al., 1989), the sequence of exons 1 and 2 of the gene is identical with the cDNA (starting at position -68 in Figure 1). Previous studies have indicated that there is only a single 7-type GS gene in French bean (Cullimore et al., 1984; Gebhardt et al., 1986), so it is likely that the cDNA and genomic clones correspond to the same gene. Primer extension analysis was used to locate the 5’ end of the gln-y mRNA in nodules. Figure 2 shows that the major transcriptional start sites in the gln-y gene are located between positions -73 and -68, about 30 bp downstream of a canonical TATA box sequence. It has not been established whether different transcription start sites are used when the gene is expressed in other tissues of the plant.
From a comparison of the 5’-flanking regions of the
soybean N24 gene and the leghemoglobin and N23 gene families, Sandal et al. (1987) identified two short conserved sequences, AAAGAT and CTCTT, each of which is found in one or more copies within 200 bp of the start of transcription in all the genes examined, and they desig- nated these as putative nodulin consensus motifs. The same sequences were subsequently found in two leg- hemoglobin genes of the stem-nodulating legume Ses- bania (Metz et al., 1988). One copy of each motif occurs within the organ-specific element (OSE) of the soybean lbc, gene, a 37-bp sequence that is involved in directing nodule-specific expression (Stougaard et al., 1987, 1990). When the gln-y sequence was searched for homologies to the Ibc, promoter, a 15-bp sequence that is related to part of the OSE was identified about 140 bp upstream from the TATA box. Table 1 shows that similar 15-bp sequences are also found at analogous locations in the promoter regions of the soybean N23 and Sesbania glbs genes. In each case, the OSE-related segment contains a sequence that is identical (Ibc,, glbs, and N23), or almost identical (gln-y), to the AAAGAT motif.
No perfect matches to either of the nodulin consensus motifs are found within the promoter-proximal region of the gln-y gene, but one copy of each sequence does occur further upstream, and copies of both sequences (mostly in the inverted orientation) are also found downstream of the cap site (Figure 1). However, the two sequences also occur at a similar frequency in the 5’-flanking region of gln-p (Turton et al., 1988), a related GS gene that does not show the same pattern of induction during nodulation (Gebhardt et al., 1986; Bennett et al., 1989; Forde et al., 1989).
The 5’-flanking region of gln-y contains a number of repeated sequences and sequences with unusual struc- tures (Figure 1). Among these (at -516 and -466) are two imperfect copies of an extremely A/T-rich 21 -bp sequence that, as discussed below, is closely related to one of two protein binding sites identified in the soybean /bc3 pro- moter (Jensen et al., 1988).
Analysis of DNA-Protein lnteractions in the 5‘-Flanking Region of gln-r
We have carried out an in vitro study of interactions between nuclear factors from French bean and sequences in the upstream region of gln-y using the technique of gel retardation analysis. Figure 3A illustrates in diagrammatic form the two overlapping sets of restriction fragments, covering the region from -971 to -21, that were used in these experiments. Each fragment was end labeled with 32P and incubated with crude nuclear extracts from French bean roots and nodules. For each extract and for each fragment, the concentration of herring sperm DNA needed to eliminate nonspecific binding was first established (gen-

Conserved A/T-Rich Binding Sites 927
HindIII . AAGCTTGGATCATATGTAGATACATGGT~TGTGTTGTAATGAACTTACCCTAATCTAT~~AGGT~CACTAAGTCTTATATATTATT -889
GGTTTAAATTAAAATTTAGTGTAAAAGTACATAT~TCATATATGCCAAATTTTT~TTATTGTGTGAGGTTGTTTGTGTTTATGA~~ - 799
b DraI . RsaI MaeIII
-978
-888
- 798
- 708
-618
- 528
-438
-348
-258
ATTTAATTGTCATAATGCATCAATAAAAAAAAATCATTATTATAATAT~ACTTAACGAAGAAATAAAAATGAATGATAAGTAAAGTTTCG -709
TAAACATT~CTTTCTTTCAAAAAACTAAAGTGTCTGAAATTCAACCTT~CACTTGGAAATGAAATTGA~AATTCTCCAAAATATAATT~G -619
C
. SpeI AAGTTTATGCATATTTATGAACTAGTGTAATTAGAGAAGTATTCAAAG~CAAATAG~ATATTTTTC~TAATACATTAAACACTTTA~A - 529
TATAAACACGGTATACTAATTATTATATTTAATTATTGATTAGG~GAAA~AGTCTAATATTAATTAGAATATTTAATGGTGGGA -439
GAAAAATAAAAATATTTGG~AATTACCAAAGATGAAAAGGCAAATTTCAATAACCATTTATTTTATAAAAAATCTATGAAATGGCCTTTG -349
G G G T T T A A A T A A C A T ~ T T A A G C ~ A G C T T Â ~ T A A C A ~ C A A T A A A T G A A T A ~ C A T T G A A T T A T A ~ T T A T A T T T A ~ G T A T A A A A A C T - 259
---L----L-- RsaI RsaI . A s 1 - 2 - - - -A- -,
C C
a C DraI. I . MaeIII .
ATGTAGGAAAAGG~TTTTAAGATGTTT~AAATATAACAGTATTATTC~TACCAGTGAAAATACATGTGCGTATTCTGATACAGTTAAAA - i 69
DraI . - i 68 T A C C A A T T ~ T C T T T T A A A A G T T A T C T T G C A G T A T T T G T ~ T T G G A A A C T G A G A G A T A G C A T A A A C A C ~ ~ ~ C C A C ~ G C A A C A A C C ~ T - 79
o... - 78 GTATCAACGCATTGAAAGGAAGAGAAAGAGAAAATTTC~CTGTG~TCTCTGCTGAAAGTTTTGG~TTCTTGAAGATGTCATCAA~C i i
b b b M S S I
12 TCCGATCTiGTTAACCTTAACCTCTCTGACTCCACCGAGAGGGTGATCGCCGAGTATAiATGgt tag tgt t tgt tcccttcgt t t t t tcc 1 O1
102 tacactcaagtcctctgtttcccctgcacgtcaaaaccaccattgta~tgcagcatg~tacattcactcaccattgcttcttgcctaatt 191
S D L V N L N L S D S T E R V I A E Y I W
192 c a a c t a c c a t t c t c a c t t c t a g c c t t t t t c a t t t c t t t t a c c t g c g c t t a a t c a a c g c t t c a a ~ g t a t t t t c t c t a t c t t t a g G G T 281
282 TGGTGGATCTGGCATGGACATGAGGAGCAAAGCAAGAqtaatttacttci 331
Figure 1. 5’-Flanking and N-Terminal Sequences of the qln-r Gene.
The N-terminal sequence of the 7 polypeptide is written below the nucleotide sequence using the single-letter code, and intron sequences are shown in lowercase type. The locations of the intronlexon boundaries were determined by comparison with the sequence of pcGS-yl , a gln-r cDNA clone (Bennett et al., 1989). The major transcriptional start sites, as determined by primer extension, are indicated by closed circles. Numbering is from the first base of the ATG translational start codon (designated zero). A putative TATA box sequence and a sequence with homology to part of the OSE of the soybean Ibc, gene (Stougaard et al., 1987) are boxed. The two 21-bp A/T-rich repeats (see text) are indicated by dashed lines, and other inverted or direct repeats by arrows. Additional sequences referred to in the text are underlined and identified by letters: those labeled a and b match the nodulin consensus motifs identified by Sandal et al. (1987) and those labeled c conform to the TATTTWAT consensus motif (see Figure 6).
b a V b .
G G S G M D M R S K A R
erally 0.5 pg to 5 pg per assay). The binding reactions were electrophoresed on acrylamide gels to resolve the DNA-protein complexes from unbound DNA.
Figure 36 presents the results obtained when one set
of five fragments was tested with nodule extracts in the gel retardation assay. Three of the fragments (RS2, RS3, and MR3) formed complexes that migrated more slowly than free DNA (lanes d, f, and h) and that were not seen

928 The Plant Cell
A C G T
a b c d efFigure 2. Mapping of the 5' End of the gln-y mRNA by PrimerExtension.
The primer extension reactions were carried out as described inMethods using an oligonucleotide specific for the 5'-untranslatedregion of the gln-y mRNA and poly(A)+ RNA from nodules astemplate (lane e). As a control, an identical reaction was carriedout in parallel on poly(A)* RNA from roots (lane f), which is highlydepleted in gln-y mRNA (Gebhardt et al., 1986; Bennett et al.,1989). Size markers were obtained by carrying out a set of fourdideoxy sequencing reactions (lanes a to d) using the same primerand a single-stranded DNA template consisting of the sensestrand of the 5' end of the gln-y gene (see Methods). The majortranscriptional start sites, as inferred from the sizes of the majorprimer extension products, are indicated with arrows, and thelocation of the putative TATA box sequence is also shown.
in the absence of nuclear protein (lanes c, e, and g). Figure3C shows the results obtained when the binding reactionswere carried out on the second set of restriction fragments.The three fragments (AD1, AD2, and ADS) that span thesame region of the gene as RS2, RS3, and MRS also
formed complexes with factors from nuclear extracts (Fig-ure 3C, lanes a, d, and g), but the fourth fragment, AD4,did not (not shown). A distinctive feature associated withADS was the formation of a second, less abundant com-plex of lower mobility (see lane g), which was found toincrease in relative abundance at high concentrations ofnuclear protein. Both complexes appear to represent se-quence-specific interactions because they were each sen-sitive to competition by a 50-fold molar excess of theunlabeled fragment. Both complexes were also very muchmore susceptible to competition with poly(dA-dT) thanwith poly(dG • dC) (data not presented).
These observations suggested that a nodule factor wasinteracting in a sequence-specific manner with A/T-richsequences in ADS and that the formation of additionalcomplexes was due to multiple interactions between thisfactor and the DNA fragment. To investigate the possibilitythat the 21-bp A/T-rich repeats (both copies of which arepresent on ADS, and one each on RS3 and MRS) may bethe target sites for this nodule factor, we synthesized adouble-stranded oligomer (7-AT2) containing the sequenceof the second of the two repeats (see Methods) and testedits ability to compete for binding in the gel retardationassay. These experiments showed that both of the com-plexes formed with ADS could be effectively titrated out inthe presence of an excess of the 7-AT2 oligomer (Figure3C, lanes h and i). In separate competition experiments(not shown), the concentration of 7-AT2 required to obtain50% inhibition of binding to ADS was less than one-hundredth the concentration of poly(dA-dT) required toobtain the same effect. The 7-AT2 oligomer also competedeffectively for binding to AD1 (lanes b and c), to AD2 (lanese and f), and to RS2, RS3, and MRS (not shown). There-fore, it appears that a single type of binding activity wasresponsible for all of the interactions with the gln-y genethat were detected in nodule extracts in our experiments.A minimum of four binding sites for this factor would beneeded to account for the results obtained in Figure 3.Two of these sites probably correspond to the two copiesof the 21-bp A/T-rich repeat, but the exact number andposition of the other binding sites have not been deter-mined. The location of the 21-bp A/T-rich repeats closeto the ends of RS3 and MRS may help to explain whybinding to these fragments was much weaker than to ADS(Figure SB).
Under the conditions used in the present study, nobinding of nodule extracts to NR4, MR2, or AD4 wasobserved. However, Figure 3D shows that NR4 did forma complex with factors from root extracts and that thiscomplex could be competed out by an excess of theunlabeled fragment. Unlike the nodule binding activity, thisroot factor was not sensitive to the presence of largeamounts (up to 1 HQ) of poly(dA-dT) nor to competitionby a 100-fold molar excess of the 7-AT2 oligomer (notshown).

Conserved A/T-Rich Binding Sites 929
Table I. Conserved, OSE-Related Sequences in the Promoter Regions of Four Nodulin Genes
Gene Positiona Conserved Sequence
Soybean /bcs -1 35 1 1 6 A A A A 6 A 1 6 A 1 1 6 c 90 b p T A T A Sesbania glb, -196 I T A A A A A G A T T A T T G + l O0 bp--*TATA Soybean N23 -1 77 T T A T A A A 6 A T G A A A G c 53 b p T A T A French bean g/n-7 -245 11 I1 I A A 6 A 1 61 11 6-1 28 bp-+TATA Consensus T T N W A A A G A T G A T T G
a Relative to the translation initiation codon.
Related but Distinct Binding Factors Are Present in Nodules and Roots
To investigate whether the nuclear factors that recognize the A/T-rich binding sites in the gln-y gene are specific to nodules, we carried out a gel retardation experiment with the 32P-labeled y-AT2 oligomer and a crude nuclear extract from uninoculated roots. Figure 4A shows that two com- plexes were formed when the y-AT2 oligomer was incu- bated with the root extract (lane f), both of which had higher mobility than the single complex formed with the nodule extract (PNF-1, lane a). However, only one of the root complexes was effectively competed out by the pres- ente of excess unlabeled 7-AT2 oligomer (PRF-1, lanes f to j). The faster moving root complex, which was not seen in binding experiments with the AD3 fragment (see Figure 4B), was much less susceptible to competition by y-AT2 and is thought to be due to nonspecific DNA binding activity, although this has not been investigated further. Preliminary experiments with extracts from other tissues indicate that factors related to PNF-1 (Phaseolus nodule factor-1) and PRF-1 (Phaseolus root factor-1) are also present in leaves and in white cotyledons of germinating seeds.
Figure 4B shows the results obtained when we investi- gated the effects of RNase and proteinase K treatments on the root and nodule factors that bind to AD3. lnitial attempts to perform the enzyme treatments at 37OC were unsuccessful because of the temperature sensitivity of both binding factors, particularly PRF-1. When the nuclear extracts were incubated for 10 min at 25°C immediately before the binding assay, the nodule binding activity was retained (lane c), although much of the root binding activity was lost (lane 9). Both factors were resistant to a 1 O-min pretreatment with RNase A (lanes d and h) but both were sensitive to proteinase K (lanes e and i). The proteinase K digestion led to a marked increase in the mobility of the major root and nodule complexes without having any significant quantitative effect on the binding activities when compared with the preincubated controls. A slightly differ- ent result was obtained when a range of concentrations of trypsin (up to 30 ng/assay) was substituted for protein-
ase K. As the concentration of trypsin was increased above a threshold of 2.5 ng/assay, there was a progressive increase in mobility of the PNF-1 and PRF-1 complexes that, in this case, was accompanied by a progressive loss in binding activity (data not shown). At the highest trypsin concentration, the residual binding activity was about 30% of the control.
From these experiments, it is clear that PNF-1 and PRF-1 are at least partially, and probably wholly, proteinaceous. The increase in the electrophoretic mobility of the PNF-1 and PRF-1 complexes after proteinase K digestion may indicate that one of severa1 protein species involved in formation of these complexes is more susceptible than the others to proteolysis but is not required for binding. Alter- natively, partia1 proteolysis of a single DNA-binding protein could have removed an N- or C-terminal segment that is not required for binding. A number of prokaryotic and eukaryotic DNA-binding proteins have been shown to con- tain a separate DNA binding domain that is linked to the rest of the protein by a protease-sensitive peptide (Geisler and Weber, 1977; Sauer et al., 1979; Supakar et al., 1989).
PNF-1 and PRF-1 also Recognize an A/T-Rich Sequence in a Soybean Leghemoglobin Gene
The similarities already noted between the 21 -bp A/T-rich repeats and an A/T-rich site in the soybean lbc, gene led us to investigate whether there was cross-recognition between PNF-1 and the Ibc, binding sites. The target DNA used in these experiments was a double-stranded oligomer (Ib-AT1) corresponding to binding site 1 in the Ibc, gene (Jensen et al., 1988). Of the two A/T-rich binding sites in Ibc,, this sequence has the higher affinity for the binding protein from soybean nodules (Jensen et al., 1988) but unlike binding site 2 is only distantly related to the gln-y repeats. The 32P-labeled yAT2 oligomer was incubated with extracts of French bean roots and nodules in the presence of a range of concentrations of the unlabeled Ib-AT1 and yAT2 oligomers. After electrophoresis and autoradiography, the bands containing the DNA-protein complexes were excised from the gel and the amounts of labeled DNA bound were estimated by scintillation count-

930 The Plant Cell
H DRM
(-978)
S A.RR. . D MI_________IT I I T______T I____1
100 bpAD1 AD3 AD2 AD4
I/////////////////////771 \_
NR4 RS2 RS3 MRS MR2
B NR4RS2RS3 MRS MR2 C AD1 AD2 AD3 D NR4
t-
a b c d e h i j a b c d e f g h i a b c d e fFigure 3. Interaction of Nuclear Factors with the 5'-Flanking Region of gln-y.
(A) Map of the upstream region of gln-y showing the restriction fragments used as probes to study DNA-protein interactions. Thelocations of four TATTTWAT motifs are indicated by closed circles, and the two 21-bp A/T-rich repeats are indicated by arrows. Anasterisk marks the segment with homology to the /oc3 OSE (Table 1), and a right-angled arrow shows the position of the majortranscriptional start sites. A, D, H, M, R, and S refer to Accl, Oral, Hindlll, Maelll, Rsal, and Spel restriction sites, respectively. Therestriction fragments giving the strongest binding to nodule factors are indicated by the black rectangles, those binding weakly by theshaded rectangles, and the fragment that bound to a factor present in roots but not nodules by the rectangle with vertical bars.(B) Gel retardation assays performed with nodule nuclear extracts and one set of five restriction fragments spanning a 950-bp regionupstream of the gln-y gene. Binding reactions included crude nuclear extract from French bean nodules (lanes b, d, f, h, and j) or noextract (lanes a, c, e, g, and i) and 1 ng to 3 ng of the appropriate 32P-labeled DNA fragment. After electrophoresis, the gel was fixed,dried, and autoradiographed overnight (except for lanes e and f, which were exposed for 3 days to visualize the weak binding to RS3).Retarded bands resulting from binding to nodule factors are indicated by arrows. U, unbound DNA fragments.(C) Gel retardation assays performed with nodule nuclear extracts and a second set of restriction fragments from gln-y. 32P-labeled AD1,AD2, or ADS were incubated in the presence of nodule extract and a 200-fold (lanes b, e, and h) or a 500-fold (lanes c, f, and i) molarexcess of a synthetic double-stranded oligomer (7-AT2) or in the absence of the oligomer (lanes a, d, and g). The 7-AT2 oligomer containsthe sequence of the second of the two 21-bp A/T-rich repeats in the gln-y promoter (see Figure 1 and Methods).(D) Gel retardation assays performed with root extracts and the gln-y fragment NR4. Binding assays included 0.5 ng of 32P-labeled NR4,root nuclear extract (1.4 //g of protein), and a range of concentrations of unlabeled NR4: 0 ng (lane a), 2.5 ng (lane b), 5 ng (lane c), 10 ng(lane d), 20 ng (lane e), or 50 ng (lane f).

Conserved A/T-Rich Binding Sites 931
A NODULE ROOT B NODULE ROOT
*<PNF-1- ft
•M +—PRF-1 PNF-1
• «•••• •ns
u
h i j
•PRF-1
a b c
Figure 4. Analysis of Root and Nodule Factors That Bind to A/T-Rich Target Sites in gln-y.(A) Binding of root and nodule factors to the 7-AT2 oligomer. Gel retardation assays were carried out on the 32P-labeled 7-AT2 oligomerusing nuclear extracts from nodules (lanes a to e) or roots (lanes f to j) and a range of concentrations of unlabeled 7-AT2 oligomer: 0 ng(lanes a and f), 0.6 ng (lanes b and g), 1.5 ng (lanes c and h), 3 ng (lanes d and i), or 6 ng (lanes e and j). PNF-1 is the major retardedcomplex obtained with nodule extracts; PRF-1 is the major complex obtained with root extracts; ns, nonspecific binding activity.(B) Sensitivity of PNF-1 and PRF-1 to temperature, ribonuclease, and proteolysis. Binding assays contained 32P-labeled AD3 togetherwith nodule extract (6.6 ̂ g of protein, lanes b to e), or root extract (3.5 ng of protein, lanes f to i), or no protein (lane a). Before additionof the labeled DNA, the binding assays were preincubated for 10 min at 25°C with 3 ng of bovine pancreatic RNase A (lanes d and h), or3 ^g (3 to 6 x 10~5 units) of proteinase K (lanes e and i), or no enzyme (lanes c and g). Lanes b and f: controls without preincubation.Note that the mobility difference between PNF-1 and PRF-1 is less apparent with ADS than when the smaller 7-AT2 oligomer is the targetDNA.
ing. Figure 5A shows that the lb-AT1 oligomer was able tocompete for binding to PNF-1 even more efficiently thanthe 7-AT2 oligomer itself. Thus, the same nuclear factorthat interacts with the A/T-rich repeats in the gln-y genehas a high affinity for the Ibc3 binding site. Figure 5Bshows that similar results were obtained when the com-petition experiment was performed with a root extract,although in this case the binding factor (PRF-1) showed asimilar affinity for both oligomers (Figure 5B).
Comparison of A/T-Rich Binding Sites in NodulinGenes
In an attempt to identify common sequence motifs in thevarious A/T-rich binding sites that have now been identifiedin nodulin genes from several legume species, the se-quences of the gln-y A/T-rich repeats have been aligned
in Figure 6 with the sequences of A/T-rich binding sitesfrom the soybean Ibc3 and N23 genes and the Sesbaniaglb3 gene. The two gln-y sequences, which are identicalat 18 out of 21 positions, differ at only six and fivepositions, respectively, from the sequence at binding site2 of the Ibc3 gene (Figure 6). The sequences of the fourother binding sites, on the other hand, show very littlesequence similarity to the gln-y repeats and are them-selves quite diverse in structure. It is possible, neverthe-less, to identify a shorter core motif (boxed in Figure 6)with the consensus sequence TATTTWAT (where W = Aor T), which is common to all of the binding sites and formspart of an imperfect palindromic sequence of variablelength.
When the gln-y upstream region was scanned in bothorientations for sequences conforming to this consensus,we located just two additional matching sequences (at-799 and -380; Figure 1). The former sequence occurs

932 The Plant Cell
A 1 O O k 1
NODULE
6 1
I I I 1 1 I
o 1 2 3 4 5 6 ng competitor DNA
O 0 L 1
ROOT
O ' I I I I I 1
o 1 2 3 4 5 6 ng competitor DNA
Figure 5. Comparison of the Relative Affinities of PNF-1 and PRF-1 for the 7-AT2 and Ib-ATl Oligomers.
(A) Competition experiment with nodule extract. Binding reactions contained 32P-labeled y-AT2, nodule extract, and different amounts of unlabeled y-AT2 (O) or Ib-AT1 (a) oligomers. The Ib- AT1 oligomer contains the 24-bp sequence of binding site 1 in the soybean /bcs gene (see Figure 6 and Methods). After electropho- resis, the gels were fixed, dried, and autoradiographed overnight. Gel slices containing the retarded DNA-protein complexes were counted in a scintillation counter and the results (which are the average of duplicate experiments) were expressed as a percent- age of the counts obtained in the absence of competitor. (B) Competition experiment with root extract. The experiment was carried out as for (A) except that the nuclear extract was from roots.
in AD1 and RS2 (see Figure 3A) and so might account for their ability to bind to PNF-1. However, detailed mapping studies will be necessary to determine the actual PNF-1 binding site in AD1 and RS2.
PNF-1 Binds with Low Affinity to a Protein Binding Site in a Soybean Seed Protein Gene
Protein binding studies with a soybean lectin gene (lec) led to the identification of two A/T-rich sequences that interact with a factor from immature embryos (Jofuku et al., 1987). Both of these binding sites contain the sequence motif AATTTAAT (in opposite orientations), which differs at only a single position from the nodulin octanucleotide core motif (Figure 6). Jofuku and coworkers reported that the soybean embryo factor did not bind to a 1600-bp fragment from the upstream region of the Ibc, gene, but it is likely that the sensitivity of the gel retardation assay with a fragment of this size would be very low. To determine whether PNF-1 is able to recognize the lec binding sites, we synthesized a double-stranded oligomer (lec-AT2) cor- responding to the region from -153 to -132 of the lec gene, the second of the two binding sites (see Jofuku et al., 1987, and Methods). A reciproca1 competition experi- ment was then carried out in which the y-AT2 and lec-AT2 oligomers were end labeled and mixed with a nodule nuclear extract in the presence of a range of concentra-
PV gln - - f (1)
Pv g7n-7 ( 2 )
Sr g763
Gm N23 (1)
Consensus core m o t i f : rl ATTTWA
Figure 6. Alignment of A/T-Rich Binding Sites in Nodulin Genes from Three Legume Species.
The two 21-bp A/T-rich repeats from gln-y have been aligned with five A/T-rich sequences that bind to related nuclear factors from other legume species. Where binding to a particular oligo- nucleotide sequence has been demonstrated, the relevant se- quence is underlined; where the extent of the binding site was delimited by Exolll digestion, the minimal binding site is marked by an open bar; where the binding site has been mapped by footprinting, the protected region is marked by a solid bar. A conserved octanucleotide motif is boxed, and a region of imperfect dyad symmetry within each sequence is indicated by asterisks. The Ibc, and N23 binding sites were mapped by Jensen et al. (1988) and Jacobsen et al. (1990), respectively, and the g/bs site was reported by Metz et al. (1988). Pv, French bean; Gm, soy- bean; Sr, Sesbania.
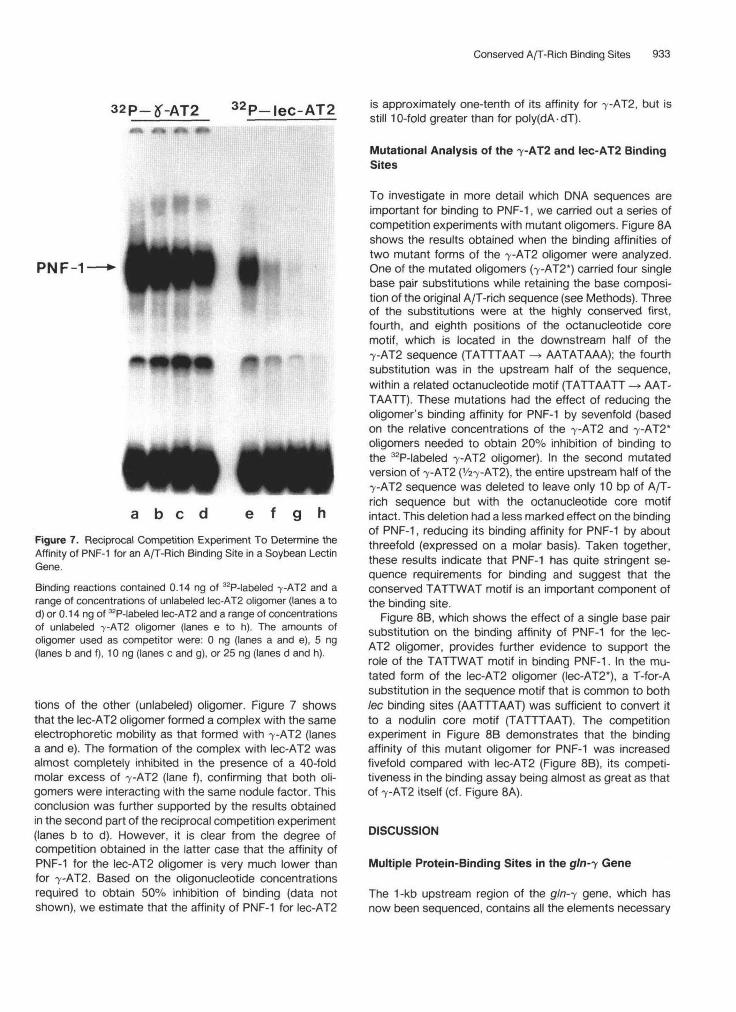
Conserved A/T-Rich Binding Sites 933
32P-lec-AT2 is approximately one-tenth of its affinity for 7-AT2, but isstill 10-fold greater than for poly(dA • dT).
Mutational Analysis of the 7-AT2 and lec-AT2 BindingSites
PNF-1
w^Pf
a b e d f g hFigure 7. Reciprocal Competition Experiment To Determine theAffinity of PNF-1 for an A/T-Rich Binding Site in a Soybean LectinGene.Binding reactions contained 0.14 ng of 32P-labeled 7-AT2 and arange of concentrations of unlabeled lec-AT2 oligomer (lanes a tod) or 0.14 ng of 32P-labeled lec-AT2 and a range of concentrationsof unlabeled 7-AT2 oligomer (lanes e to h). The amounts ofoligomer used as competitor were: 0 ng (lanes a and e), 5 ng(lanes b and f), 10 ng (lanes c and g), or 25 ng (lanes d and h).
tions of the other (unlabeled) oligomer. Figure 7 showsthat the lec-AT2 oligomer formed a complex with the sameelectrophoretic mobility as that formed with 7-AT2 (lanesa and e). The formation of the complex with lec-AT2 wasalmost completely inhibited in the presence of a 40-foldmolar excess of -y-AT2 (lane f), confirming that both oli-gomers were interacting with the same nodule factor. Thisconclusion was further supported by the results obtainedin the second part of the reciprocal competition experiment(lanes b to d). However, it is clear from the degree ofcompetition obtained in the latter case that the affinity ofPNF-1 for the lec-AT2 oligomer is very much lower thanfor 7-AT2. Based on the oligonucleotide concentrationsrequired to obtain 50% inhibition of binding (data notshown), we estimate that the affinity of PNF-1 for lec-AT2
To investigate in more detail which DMA sequences areimportant for binding to PNF-1, we carried out a series ofcompetition experiments with mutant oligomers. Figure 8Ashows the results obtained when the binding affinities oftwo mutant forms of the 7-AT2 oligomer were analyzed.One of the mutated oligomers (7-AT2*) carried four singlebase pair substitutions while retaining the base composi-tion of the original A/T-rich sequence (see Methods). Threeof the substitutions were at the highly conserved first,fourth, and eighth positions of the octanucleotide coremotif, which is located in the downstream half of the7-AT2 sequence (TATTTAAT -> AATATAAA); the fourthsubstitution was in the upstream half of the sequence,within a related octanucleotide motif (TATTAATT —»AAT-TAATT). These mutations had the effect of reducing theoligomer's binding affinity for PNF-1 by sevenfold (basedon the relative concentrations of the 7-AT2 and 7-AT2*oligomers needed to obtain 20% inhibition of binding tothe 32P-labeled 7-AT2 oligomer). In the second mutatedversion of 7-AT2 (1/?7-AT2), the entire upstream half of the7-AT2 sequence was deleted to leave only 10 bp of A/T-rich sequence but with the octanucleotide core motifintact. This deletion had a less marked effect on the bindingof PNF-1, reducing its binding affinity for PNF-1 by aboutthreefold (expressed on a molar basis). Taken together,these results indicate that PNF-1 has quite stringent se-quence requirements for binding and suggest that theconserved TATTWAT motif is an important component ofthe binding site.
Figure 8B, which shows the effect of a single base pairsubstitution on the binding affinity of PNF-1 for the lec-AT2 oligomer, provides further evidence to support therole of the TATTWAT motif in binding PNF-1. In the mu-tated form of the lec-AT2 oligomer (lec-AT2*), a T-for-Asubstitution in the sequence motif that is common to bothlee binding sites (AATTTAAT) was sufficient to convert itto a nodulin core motif (TATTTAAT). The competitionexperiment in Figure 8B demonstrates that the bindingaffinity of this mutant oligomer for PNF-1 was increasedfivefold compared with lec-AT2 (Figure 8B), its competi-tiveness in the binding assay being almost as great as thatof 7-AT2 itself (cf. Figure 8A).
DISCUSSION
Multiple Protein-Binding Sites in the gln-y Gene
The 1 -kb upstream region of the gln-y gene, which hasnow been sequenced, contains all the elements necessary

934 The Plant Cell
O O
Õ 40- * I 8 -AT2 I
2o t O1 I I I I I
O 2 4 6 8 10 ng competitor DNA
B 1
- O
C O O
L c
O0 1
6o t
the presence in this region of two types of binding site for nuclear proteins: (1) a sequence between -978 and -861 that binds a factor present in roots but not nodules, and (2) multiple binding sites between -860 and -154 that interact with related but distinct factors from roots and nodules (designated PRF-1 and PNF-1, respectively). PRF-1 and PNF-1 could be distinguished both by the electropho- retic mobility of the complexes that they formed in the gel retardation assay (Figure 4) and by their relative affinities for different A/T-rich sequences (Figure 5), but whether they represent different gene products or post-translation- ally modified forms of the same protein(s) is unknown.
No DNA-protein interactions were detected within the region of the gln-7 promoter that contains a sequence related to part of the OSE of the soybean Ibc, gene (Table 1). Jensen et al. (1988) were similarly unable to detect any nuclear factor binding to the OSE sequences in the Ibc3 gene, even though its role in conferring nodule-specific expression (Stougaard et al., 1987, 1990) make it a likely target site for transcriptional factors. The conservation of similar OSE-related sequences in the promoter region of several different types of late nodulin gene from diverse legume species (Table 1) is consistent with a possible role in the regulation of transcription, but detailed mutational analysis will be required to establish whether the OSE- related segment of the gh-y promoter is involved in con- trolling the nodule-enhanced pattern of expression of this gene.
PNF-1 1s Closely Related to the Soybean Nodule Factor NAT2
O 2 4 6 a 10 ng competitor DNA
Figure 8. Competition Experiments To Determine the Affinity of PNF-1 for Mutant Oligomers.
(A) Mutational analysis of y-AT2. The ability of the mutant oligo- mers to compete for binding of PNF-1 to the 3zP-labeled 7-AT2 oligomer was assessed by the procedure described in Figure 5. Binding reactions contained 32P-labeled 7-AT2, nodule extract, and different amounts of unlabeled y-AT2 (O), %y-AT2 (a), and
( E ) Mutational analysis of lec-AT2. As for (A), except that the unlabeled competitor oligomers were lec-AT2 (O) and lec- AT2' (O). The sequences of the double-stranded oligonucleotides used in these experiments are given in Methods.
7-AT2' (O).
for induction of gene expression in nodules of transgenic L. corniculatus plants (Forde et al., 1989; W.-J. Shen, M.S. Williamson, and B.G. Forde, unpublished results). Gel re- tardation analysis, using crude nuclear extracts from French bean roots and nodules, has now demonstrated
Two distinct nodule factors (NAT1 and NAT2) and one leaf factor (LAT1) have recently been shown to bind to several A/T-rich sites in the promoter region of the soybean N23 gene (Jacobsen et al., 1990). NATl and LAT1 were found to be related to the high-mobility group I (HMG I) protein of eukaryotic chromatin, but NAT2 differed in having more stringent sequence requirements for binding and in gen- erating a DNA-protein complex with a much lower electro- phoretic mobility. Under the standard conditions used in our binding assays, we detected only a single type of binding activity in nodule extracts. However, when herring sperm DNA was used as nonspecific competitor instead of a synthetic polynucleotide mix in binding assays with the y-AT2 oligomer, we were able to detect a second complex with a very much higher electrophoretic mobility than the PNF-1 complex (data not presented). This second binding factor in French bean nodules appears to corre- spond to NATl because it was soluble in 2% TCA and extremely sensitive to the presence of poly(dA. dT) in the binding assay, properties that are characteristic of the soybean HMG I-like proteins. PNF-1, on the other hand, resembles NAT2 and differs from the HMG I-like factors in that its binding activity is sensitive to heat and to TCA

Conserved A/T-Rich Binding Sites 935
treatment (Jacobsen et al., 1990). Taking these observa- tions together with the finding that PNF-1 has a high affinity for an NAT2 binding site from the soybean /bc3 gene, we conclude that PNF-1 is probably the homolog in French bean of NAT2.
A/T-Rich Binding Sites in Nodulin Genes Share a Common Sequence Motif
Binding sites for NAT2 and PNF-1 and for related factors in Sesbania and alfalfa (Metz et al., 1988) have now been identified in leghemoglobin genes of both soybean (Jensen et al., 1988) and Sesbania (Metz et al., 1988), in the soy- bean N23 gene (Jacobsen et al., 1990), and in the g/n-y gene of French bean (this paper). As Figure 6 shows, apart from their high A/T content and partia1 dyad sym- metry, these binding sites show remarkable sequence diversity, indicating a high degree of degeneracy in the sequence requirements for recognition. Nevertheless, a significant degree of sequence specificity is indicated by the finding that PNF-1 has very differing affinities for dif- ferent A/T-rich binding sites (Figure 7) and by the demon- stration that certain single base pair substitutions, even while maintaining the base composition of the target se- quence, can have a dramatic effect on the binding of NAT2 (Jacobsen et al., 1990) and PNF-1 (Figure 8). A 16-bp motif, WTWAATWWTTTATTTW, is shared by the soybean /bc3 and Sesbania g/b3 binding sites (Metz et al., 1988), but the binding sites subsequently identified in g/n-7 (this paper) and in the N23 gene (Jacobsen et al., 1990) conform only poorly to this consensus. However, by aligning the A/T-rich sequences as shown in Figure 6, it was possible to recognize a shorter motif with the consensus sequence TATTTWAT, which is common to all the NAT2/PNF-1 target sites so far reported (Figure 6). Experimental evi- dente that this motif is important for protein recognition was obtained by analysis of the binding affinities of mu- tated forms of the yAT2 oligomer (Figure 8A) and by the demonstration that a single base pair substitution in the AATTTAAT motif of the lec-AT2 oligomer, which generated a nodulin consensus motif TATTTAAT, greatly increased its affinity for PNF-1 (Figure 86).
Do A/T-Rich Binding Sites Have a Role in Regulating Nodulin Gene Expression?
The conservation of A/T-rich binding sites in a diverse set of late nodulin genes suggests that they may have an important part to play in regulating nodulin gene expres- sion, but the nature of this role is still uncertain. Studies carried out on a series of 5' deletion mutants of the gln-7 gene found that removal of the region between -597 and -354, which encompasses at least two PNF-1 binding sites, resulted in the loss of detectable gene expression in
nodules of transgenic Lotus plants (W.-J. Shen, M.S. Wil- liamson, and B.G. Forde, unpublished results). Similar evidence for an association between the NAT2 binding sites and a weak positive regulatory element in the /bc3 gene has been reported (Jensen et al., 1988). Although recent work has indicated that deletion of two NAT2 binding sites has no dramatic effect on the function of the /bc3 promoter in transgenic plants (Stougaard et al., 1990), the presence of additional binding sites further upstream (Jacobsen et al., 1990) leaves open the possibility that NAT2 is involved in the regulation of this gene. In the soybean N23 gene, a positive regulatory element located between -344 and -293 (Jorgensen et al., 1988) also contains an A/T-rich binding site (Stougaard et al., 1990), and a similar correlation has been noted in the Sesbania g/b3 gene (de Bruijn et al., 1989). Thus, there is accumu- lating, but still circumstantial, evidence to suggest that the A/T-rich binding sites have a role in enhancing gene expression in nodules.
Similarities between NAT2 binding sites and sequence motifs found in the vicinity of scaffold-attached regions of severa1 Drosophi/a genes (Gasser and Laemmli, 1986) led to the suggestion that NAT2 might be involved in formation of the nuclear scaffold structure (Jacobsen et al., 1990). An alternative possibility is that NAT2/PNF-1 may function by interfering with the binding of the HMG I-like proteins to A/T-rich sequences. Because the binding site require- ments for NAT2/PNF-1 are much more stringent than for the HMG I-like factors, this will only affect a certain subset of the HMG I binding sites. The effect that such competition for binding will have will clearly depend on the function of the HMG I-like proteins in plants. In mammals, HMG I may have a role in nucleosome positioning (Strauss and Var- shavsky, 1984) or as a component of the nuclear scaffold (Solomon et al., 1986). Thus, the binding of NAT2 or PNF-1 could have an indirect effect on chromatin struc- ture in the vicinity of their target promoters by excluding HMG I-like proteins. A third possibility is that NAT2 and PNF-1 are conventional trans-acting factors, exerting their influence through protein-protein interactions with the tran- scriptional complex (Mitchell and Tjian, 1989).
PNF-1 and NAT2 May Be Members of a Larger Family of Nuclear Factors
Datta and Cashmore (1989) identified a nuclear protein from pea seedlings (AT-1) that binds to A/T-rich sequences sharing the motif AATATTTTATT in a group of photoreg- ulated genes from pea, tomato, and tobacco. Similar binding sites have been mapped in a variety of seed protein genes that are expressed in developing embryos of soy- bean (Jofuku et al., 1987; Allen et al., 1989), French bean (Bustos et al., 1989; Riggs et al., 1989), and sunflower (Jordano et al., 1989). In common with the nodulin genes, these A/T-rich binding sites are often present in multiple

936 The Plant Cell
copies and are usually located between 500 bp and 1 O00 bp upstream from the start of transcription. The sequence similarities between these A/T-rich binding sites (which generally include motifs conforming to the consensus WATS-IA,-PT) and those in the nodulin genes (which con- form to the same general consensus) suggest that many of the plant factors that recognize A/T-rich sequences and occur in diverse plant tissues could have related binding requirements. In support of this hypothesis, we have dem- onstrated that the French bean nodule factor PNF-1 was able to bind to a synthetic target site for a factor from immature soybean embryos (Figure 7).
In a further parallel with the nodulin genes, the A/T-rich binding sites in other plant genes are frequently associated with positive regulatory elements (Bustos et al., 1989; Datta and Cashmore, 1989; Riggs et al., 1989). Most notably, a 55-bp A/T-rich sequence from the p-phaseolin gene, which binds to nuclear proteins from immature French bean cotyledons, was shown to enhance expres- sion from a minimal 35s promoter in transgenic tobacco, not only in seeds but also in other parts of the plant (Bustos et al., 1989). On the other hand, an AT-1 binding site in the tobacco cab-E gene is reported to reside within a negative regulatory element (Datta and Cashmore, 1989).
Thus, a picture is emerging of an evolutionarily con- served and ubiquitous group of nuclear factors having an affinity for certain A/T-rich sequences found in a wide variety of plant genes. Extrapolating from the example of PNF-1 and PRF-1, it is likely that related factors present in different tissues (or even within the same tissue) will have different sequence preferences and will, therefore, be capable of binding to a different (but possibly overlap- ping) spectrum of A/T-rich sequences. The existence of families of transcription factors with related DNA binding specificities is now well established in metazoans and in yeast (see Mitchell and Tjian, 1989). Further work will be needed to establish whether the group of plant factors that are related by their affinity for similar A/T-rich sequences are also related structurally and functionally, and to deter- mine their role, if any, in the transcriptional control of gene expression in higher plants.
METHODS
Plant Growth
Phaseolus vulgaris (French bean) var Tendergreen was grown from surface-sterilized seed under 16-hr light (20°C)/8-hr dark (16OC) cycles in a sand and grave1 mixture (1:2) as described (Cullimore et al., 1983). Seeds were inoculated with Rhizobium bguminosarum biovar phaseoh strain R3622 at the time of sowing and again 2 days later. Nodules were harvested into liquid N2 3 to 4 weeks after sowing and stored in liquid NP or at -70°C.
Roots were obtained from uninoculated plants grown for 2 weeks using nutrient solution supplemented with nitrate.
Genomic Cloning and Nucleotide Sequencing
Unless otherwise stated, standard recombinant DNA techniques were used (Maniatis et al., 1982). The isolation and restriction mapping of the French bean genomic clones XGSN-56 and XGSN- 57 and their respective subclones pN/HB68 and pN/HB57d, which contain the 5’-flanking region of gln-7, has been reported previ- ously (Forde et al., 1989). Partia1 sequence analysis of the 2.1-kb insert in pN/HB57d was carried out on both strands by the dideoxy method (Sanger et al., 1977) using T4 DNA polymerase-generated deletions in M13mp18 and M13mp19 (Dale et ai., 1985). Se- quence data were assembled on a VAX 11/750 computer using DBUTIL (Staden, 1982) and sequence analysis and comparisons were performed on a VAX 1 1 /785 computer using the Wisconsin sequence software package (Devereux et al., 1984).
Primer Extension
The extraction of poly(A)’ RNA from roots and nodules and the primer extension reactions were carried out essentially as de- scribed previously (Freeman et al., 1990). The primer consisted of a 21 mer oligonucleotide (5’-CAGCAGAGACTCTTCACAGAG- 3’), which is complementary to a gene-specific region in the 5’- untranslated region of the gln-y mRNA. Accurate size markers were generated by performing sequencing reactions with the same primer on an M13 subclone of pN/HB57d carrying the nontranscribed strand of the gh-y gene. Primer extension reac- tions were electrophoresed on 6% polyacrylamide sequencing gels in parallel with lhe size markers and the gels were fixed, dried, and autoradiographed.
DNA Probes
DNA fragments used for gel retardation analysis contained se- quences from the 5’-flanking region of gln-7 and were obtained by cleavage of pN/HB57d with combinations of restriction en- zymes (see Figure 3A). The fragments were electrophoresed in 4% acrylamide gels, excised, and electroeluted. The restriction fragments (with the exception of AD4) were then filled in using the Klenow fragment of DNA polymerase I and subcloned in the Smal site of pUC8 or pUC9. After purification of the plasmid DNAs, the inserts were excised by digestion with BamHl and EcoRl and subsequently electroeluted from 1.2% agarose gels. The AD4 fragment was obtained from pN/BA68, a derivative of pN/HB57d (Forde et al., 1989), and labeled directly without sub- cloning. To avoid the denaturation of the DNA that can occur on drying after ethanol precipitation, leading to artifacts in the gel retardation assay (Svaren et al., 1987), pellets obtained after electroelutions were washed with 75% ethanol and drained thor- oughly before redissolving directly in 10 mM Tris-HCI. pH 8. 0.1 mM EDTA.
Oligonucleotides were synthesized by phosphoramidite chem- istry using a New Brunswick Biosearch 3810 DNA synthesizer. After deprotection, the oligonucleotides were purified by electro- phoresis in a 16% acrylamide sequencing gel and recovered from

Conserved A/T-Rich Binding Sites 937
gel slices by diffusion into 0.5 M NH40Ac. The oligonucleotides were precipitated in 80% ethanol, washed, dried, and taken up in sterile distilled water. Table 2 gives the sequences of the oligo- nucleotides that were used for binding studies.
Restriction fragments and double-stranded oligonucleotides were labeled by filling in with CI-~~P-~ATP (3000 Ci/mmol; Amer- sham International), dCTP, dGTP, and TTP using Klenow, and unincorporated nucleotides were removed by chromatography in Sephadex G-50.
Nuclear Extracts
A crude preparation of root or nodule nuclei was obtained using a procedure modified from Willmitzer and Wagner (1981). Plant material (10 g to 40 g, fresh weight) was ground to a coarse powder in liquid NP using a pestle and mortar. All subsequent steps were carried out on ice and centrifugations were at 4°C. The powder was transferred to a second mortar containing ice- cold homogenization buffer (7.5 mL/g of tissue) and ground for 3 minto 4 min. Homogenization buffer consisted of 0.25 M sucrose, 10 mM NaCI, 10 mM Mes. pH 6.0, 10 mM 1,lO-phenanthroline, 0.1 5 mM spermine, 0.5 mM spermidine, 20 mM P-mercaptoetha- nol, 0.2 mM phenylmethylsulfonyl fluoride, 0.01 O/O Triton X-1 00, the pH being adjusted to 5.3 with HCI. After filtering through four layers of muslin and then through two layers of Miracloth, the homogenate was centrifuged for 15 min at 5009. The pellet was washed three times in wash buffer (homogenization buffer minus Triton X-100, 2.5 mL/g of starting material), centrifuging for 7 min at 5009 each time.
The procedure for extraction of nuclear proteins was modified from Siebenlist et al. (1984). The crude nuclear pellet was resus- pended in 1 mL to 2 mL of NPEB (1 O mM Hepes, pH 8,300 mM NaCI, 25% (w/v) glycerol, 0.1 mM EDTA, 0.1 mM EGTA, 0.5 mM spermidine, 0.1 5 mM spermine, 1 mM phenylmethylsulfonyl fluo- ride, 7 mM P-mercaptoethanol) and mixed for 2 hr at 4OC by rotation on a Multimixer. Debris was removed by centrifugation at 12,0009 for 10 min and by recentrifuging the supernatant for a further 5 min. The crude nuclear extracts were frozen in liquid N, in aliquots of 50 pL or 100 WL and stored at -7OOC. Protein
Table 2. Oligonucleotides Used in the Gel Retardation Experiments”
Ib-ATl 5 ’ - T C G A G A T A T A T T A A T A T T T T A T T T T A T A A 6 . 3 ’ 3 ’ - C T A T A T A A T T A T A A A A T A A A A T A T T C 6 A . 5 ’
7-AT2 5’ - C T A 6 A T A T T A A T T A 6 A A T A T T T A A T 6 6 - 3 ’ 3 ’ - T A T A A T T A A T C T T A T A A A T T A C C C T A 6 . 5 ’
y-AT2‘ 5 ’ . C T A 6 A A A T T A A T T A 6 A A A A T ~ T A A ~ 6 6 ~ 3 ’
’/z 7-AT2 5’ - c T E 6 A A T A T T T A A T 6 6 . 3 ’ 3’. T ~ T A A T T A A T C T T ~ T A l A T T ~ C C C T A 6 ~ 5’
3 ’ - E C T T A T A A A T T A C C C T A 6 - 5’ lec-AT2 5’ . G A T C C C 6 T A A A T T T A A A R T T l A A T 6 T T . 3’
3 ’ - 6 6 C A T T T A A A T T T T A A A T T A C A A 6 A T C - 5 ’ lec-AT2’ 5 ’ . 6 A T C C C 6 T A A A 1 T T A A l A T T T A A T 6 T 1.3 ’
3’. 6 6 C A T T T A A A T T A T A A A T T A C A A 6 A T C - 5 ’
a Underlined nucleotides indicate positions at which mutant oligo- mers differ from the parenta1 type. The oligomer containing the sequence of binding site 1 of the
soybean /bc, gene (Jensen et al., 1987) was kindly provided by Dr. Erik Jensen, University of Aarhus.
concentrations in the extracts were estimated by the method of Bradford (1976) using a kit from Bio-Rad Laboratories.
Gel Retardation Analysis
Unless’othewise stated, binding reactions consisted of 1 O pL of crude nuclear extract (about 3 pg of protein), 1 pL of 20 X mix (300 mM Hepes, pH 8, 20 mM EDTA, 100 mM dithiothreitol), 0.5 pg each of poly(dA.dT) and poly(dG.dC), 2 pg of herring sperm DNA, and 0.2 ng to 0.5 ng (2 fmol to 7 fmol) of 32P-labeled restriction fragment, in a final volume of 20 pL. When 3ZP-labeled double-stranded oligonucleotides (0.1 ng, 5 fmol) were used as probes, the herring sperm DNA was omitted. Reactions were incubated for 5 min at 24OC before addition of the labeled DNA, and the incubation was continued for 20 min. In competition experiments, the unlabeled competitor DNA was added after the 5-min preincubation and a further 5-min incubation was carried out before addition of the labeled DNA. At the end of the incubation period, 4 pL of 0.1% bromphenol blue was added and one-third of each reaction was loaded on a 5% polyacrylamide gel (8O:l acry1amide:bisacrylamide) that had been pre-electrophoresed at 175 V for 20 min. The electrophoresis buffer was 0.25 x TBE (1 X TBE is 89 mM Tris-borate, pH 8.3, 2 mM EDTA). Samples were electrophoresed at 200 V for 2 hr, the gel temperature being maintained at 13OC to 16OC by water cooling. The gel was fixed in 40% methanol, 10% acetic acid for 45 min, dried, and exposed to x-ray film with two intensifying screens for 16 hr to 72 hr at -7OOC.
ACKNOWLEDGMENTS
We are grateful to our colleagues in the Biochemistry and Physi- ology Department for their practical help and advice, particularly Jo Clark and Michele Bianchi, and to Professor Kjeld Marcker and his colleagues at the University of Aarhus for helpful discussions. We thank Julian Franklin for providing glasshouse facilities, Martin Cornelius for synthesizing oligonucleotides, and Alison Hopley for carrying out the primer extension reactions. M.P. held a Senior Training Fellowship from the European Economic Community (EEC), and this work was also supported partly by Contract No. BAP/0099/UK of the Biotechnology Action Programme of the EEC.
Received May 7, 1990; revised July 19, 1990.
REFERENCES
Allen, R.D., Bernier, F., Lessard, P.A., and Beachy, R.N. (1989). Nuclear factors interact with a soybean @-conglycinin enhancer. Plant Cell 1, 623-631.
Atkins, C.A. (1 987). Metabolism and translocation of fixed nitro- gen in the nodulated legume. Plant Soil 100, 157-1 69.

938 The Plant Cell
Bennett, M.J., Lightfoot, D.A., and Cullimore, J.V. (1989). cDNA sequence and differential expression of the gene encoding the glutamine synthetase polypeptide of Phaseolus vulgaris L. Plant MOI. Biol. 12, 553-565.
Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye-binding. Anal. Biochem. 72, 248-254.
Bustos, M.M., Guiltinan, M.J., Jordano, J., Begum, D., Kalkan, FA.; and Hall, T.C. (1989). Regulation .of. P-glucuronidase expression in transgenic tobacco plants by an A/T-rich, cis- acting sequence found upstream of a French bean P-phaseolin gene. Plant Cell 1, 839-853.
Cullimore, J.V., Lara, M., Lea, P.J., and Miflin, B.J. (1983). Purification and properties of two forms of glutamine synthetase from the plant fraction of Phaseolus root nodules. Planta 157,
Cullimore, J.V., Gebhardt, C., Saarelainen, R., Miflin, B.J., Idler, K.B., and Barker, R.F. (1984). Glutamine synthetase of fhas- eolus vulgaris L: Organ-specific expression of a multigene fam- ily. J. MOI. Appl. Genet. 2, 589-599.
Dale, R.M.K., McClure, B.A., and Houchins, J.P. (1985). A rapid single-stranded cloning strategy for producing a sequential series of overlapping clones for use in DNA sequencing: Appli- cation to sequencing the corn mitochondrial 18s rRNA. Plasmid
Datta, N., and Cashmore, A.R. (1989). Binding of a pea nuclear protein to promoters of certain photoregulated genes is modu- lated by phosphorylation. Plant Cell 1, 1069-1 077.
de Bruijn, F.J., Felix, G., Grunenberg, B., Hoffmann, H.J., Metz, B., Ratet, P., Simons-Schreier, A., Srabados, L., Welters, P., and Schell, J. (1 989). Regulation of plant genes specifically induced in nitrogen-fixing nodules: Role of cis-acting elements and trans-acting factors in leghemoglobin gene expression. Plant MOI. Biol. 13, 319-325.
Devereux, J., Haeberli, P., and Smithies, O. (1984). A compre- hensive set of sequence analysis programs for the VAX. Nucl. Acids Res. 12, 387-395.
Forde, B.G., and Cullimore, J.V. (1 989). The molecular biology of glutamine synthetase in higher plants. Oxf. Surv. Plant MOI. Biol. 6, 247-296.
Forde, B.G., Day, H.M., Turton, J.F., Shen, W.-J., Cullimore, J.V., and Oliver, J.E. (1 989). Two glutamine synthetase genes from Phaseolus vulgaris L. display contrasting developmental and spatial patterns of expression in transgenic Lotus cornicu- latus plants. Plant Cell 1, 391 -401.
Freeman, J., Marquer, A.J., Wallsgrove, R.M., Saarelainen, R., and Forde, B.G. (1 990). Molecular analysis of barley mutants deficient in chloroplast glutamine synthetase. Plant MOI. Biol.
Gasser, S.M., and Laemmli, U.K. (1986). Cohabitation of scaffold binding regions with upstream/enhancer elements of three de- velopmentally regulated genes of D. melanogaster. Cell 46,
Gebhardt, C., Oliver, J.E., Forde, B.G., Saarelainen, R., and Miflin, B.J. (1 986). Primary structure and differential expression of glutamine synthetase genes in nodules, roots and leaves of Phaseolus vulgaris. EMBO J. 5, 1429-1 435.
Geisler, N., and Weber, K. (1977). lsolation of the amino-terminal fragment of lactose repressor necessary for DNA-binding. Bio-
245-253.
13,31-40.
14,297-31 1.
521-530.
chemistry 16, 938-943. Hirel, B., Bouet, C., King, B., Layzell, D., Jacobs, F., and Verma,
D.P.S. (1 987). Glutamine synthetase genes are regulated by ammonia provided externally or by symbiotic nitrogen fixation.
Jacobsen, K., Laursen, N.B., Jensen, E.@., Marcker, A., Poul- sen, C., and Marcker, K.A. (1990). HMG I-like proteins from leaf and nodule nuclei interact with different AT motifs in soy- bean nodulin promoters. Plant Cell 2, 85-94.
Jensen, E.@., Marcker, K.A., Schell, J., and de Bruijn, F.J. (1 988). lnteraction of a nodule specific, trans-acting factor with distinct DNA elements in the soybean leghaemoglobin Ibc, 5’ upstream region. EMBO J. 7, 1265-1271.
Jofuku, K.D., Okamuro, J.K., and Goldberg, R.B. (1987). Inter- action of an embryo DNA binding protein with a soybean lectin gene upstream region. Nature 328, 734-737.
Jordano, J., Almoguera, C., and Thomas, T.L. (1989). A sun- flower helianthinin gene upstream sequence ensemble contains an enhancer and sites of nuclear protein interaction. Plant Cell
Jergensen, J.-E., Stougaard, J., Marcker, A., and Marcker, K.A. (1988). Root nodule specific gene regulation: Analysis of the soybean nodulin N23 gene promoter in heterologous symbiotic systems. Nucl. Acids Res. 16, 39-50.
Lara, M,, Cullimore, J.V., Lea, P.J., Miflin, B.J., Johnston, A.W.B., and Lamb, J.W. (1983). Appearance of a nove1 form of plant glutamine synthetase during nodule development in Phaseolus vulgaris L. Planta 157, 254-258.
Maniatis, T., Fritsch, E.F., and Sambrook, J. (1 982). Molecular Cloning: A Laboratory Manual (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory).
Metr, B.A., Welters, P., Hoffmann, H.J., Jensen, E.@., Schell, J., and de Bruijn, F.J. (1988). Primary structure and promoter analysis of leghemoglobin genes of the stem-nodulated tropical legume Sesbania rostrata: Conserved coding sequences, cis- elements and trans-acting factors. MOI. Gen. Genet. 214, 181-191.
Mitchell, P.J., and Tjian, R. (1 989). Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins. Science 245,371-378.
Morrison, N.A., Bisseling, T., and Verma, D.P.S. (1988). Devel- opment and differentiation of the root nodule: lnvolvement of plant and bacterial genes. In Developmental Biology, Vol. 5, W. Browder, ed (New York: Plenum Press), pp. 405-425.
Riggs, C.D., Voelker, T.A., and Chrispeels, M.J. (1989). Coty- ledon nuclear proteins bind to DNA fragments harboring regu- latory elements of phytohemagglutinin genes. Plant Cell 1,
Sandal, N.N., Bojsen, K., and Marcker, K.A. (1987). A family of nodule specific genes from soybean. Nucl. Acids Res. 15,
Sanger, F., Nicklen, S., and Coulson, A.R. (1 977). DNA sequenc- ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA
Sauer, R.T., Pabo, C.O., Meyer, B.J., Ptashne, M., and Black- man, K.C. (1 979). Regulatory functions of the X repressor reside in the amino-terminal domain. Nature 279, 396-400.
Siebenlist, U., Hennighausen, L., Battey, J., and Leder, P.
EM60 J. 6, 1 167-1 171.
1, 855-866.
609-621.
1507-1 51 9.
74,5463-5467.

Conserved A/T-Rich Binding Sites 939
(1 984). Chromatin structure and protein binding in the putative regulatory region of the c-myc gene in Burkitt lymphoma. Cell
Solomon, M.J., Strauss, F., and Varshavsky, A. (1986). A mam- malian high mobility group protein recognizes any stretch of six A-T base pairs in duplex DNA. Proc. Natl. Acad. Sci. USA 83,
Staden, R. (1982). Automation of the computer handling of gel reading data produced by the shotgun method of DNA sequenc- ing. Nucl. Acids Res. 10, 4731-4751,
Stougaard, J., Sandal, N.N., Gron, A., Kühle, A., and Marcker, K.A. (1987). 5’ Analysis of the soybean leghaemoglobin Ibc, gene: Regulatory elements required for promoter activity and organ specificity. EMBO J. 6, 3565-3569.
Stougaard, J., Jorgensen, J.-L., Christensen, T., Kühle, A., and Marcker, K.A. (1 990). lnterdependence and nodule specificity of cis-acting regulatory elements in the soybean leghaemoglo- bin lbc, and N23 gene promoters. MOI. Gen. Genet. 220,
Strauss, F., and Varshavsky, A. (1984). A protein binds to a satellite DNA repeat at three specific sites that would be brought
37,381 -391.
1276-1 280.
353-360.
into mutual proximity by DNA folding in the nucleosome. Cell
Studer, D., Gloudemans, T., Franssen, H.J., Fischer, H.-M., Bisseling, T., and Hennecke, H. (1987). lnvolvement of the bacterial nitrogen fixation regulatory gene (nifA) in control of nodule-specific host-plant gene expression. Eur. J. Cell Biol.
Supakar, P.C., Zhang, X.-Y., Githens, S., Khan, R., Ehrlich, K.C., and Ehrlich, M. (1989). How different DNA sequences are recognized by a DNA-binding protein: Effects of partia1 proteolysis. Nud. Acids Res. 17, 861 1-8629.
Svaren, J., Inagami, S., Lovegren, E., and Chalkley, R. (1987). DNA denatures upon drying after ethanol precipitation. Nucl. Acids Res. 15,8739-8754.
Turton, J.F., Hopley, A.P., and Forde, B.G. (1988). 5’-Flanking sequence of a glutamine synthetase gene specifying the p subunit of the cytosolic enzyme from fhaseolus vulgaris L. Nucl. Acids Res. 16, 11 367.
Willmitzer, L., and Wagner, K.G. (1981). The isolation of nuclei from tissue cultured cells. Exp. Cell Res. 135, 69-77.
37,889-901.
45,177-184.

DOI 10.1105/tpc.2.9.925 1990;2;925-939Plant Cell
B G Forde, J Freeman, J E Oliver and M Pinedaglutamine synthetase gene from French bean.
Nuclear factors interact with conserved A/T-rich elements upstream of a nodule-enhanced
This information is current as of April 8, 2013
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists
Copyright © 2022 FDOKUMEN




















