
Nuclear BAG1 expression inhibits apoptosis in colorectal adenoma-derived epithelial cells
11
Apoptosis 2005; 10: 301–311 C 2005 Springer Science + Business Media, Inc. Manufactured in The Netherlands. Nuclear BAG-1 expression inhibits apoptosis in colorectal adenoma-derived epithelial cells J. D. Barnes, N. J. Arhel, S. S. Lee, A. Sharp, M. Al-Okail, G. Packham, A. Hague, C. Paraskeva and A. C. Williams Cancer Research UK Colorectal Tumour Biology Research Group, Department of Pathology and Microbiology, School of Medical Sciences, University of Bristol, Bristol, UK(J. D. Barnes, N. J. Arhel, C. Paraskeva, A. C. Williams); Department of Oral and Dental Science, University of Bristol, Lower Maudlin Street, Bristol, UK (S. S. Lee, A. Hague); Cancer Research UK Oncology Unit, Cancer Science Division, University of Southampton School of Medicine, Southampton General Hospital, Southampton, UK (A. Sharp, M. Al-Okail, G. Packham) BAG-1 is an anti-apoptotic protein that is frequently deregulated in a variety of malignancies including col- orectal cancer. There are three isoforms: BAG-1L is lo- cated in the nucleus, BAG-1M and BAG-1S are located both in the nucleus and the cytoplasm. In colon cancer, the expression of nuclear BAG-1 is associated with poorer prognosis and is potentially a useful predictive factor for distant metastasis. However, the function of BAG-1 in colonic epithelial cells has not been studied. Having pre- viously shown a predominant nuclear localisation of BAG- 1 in adenoma-derived cell lines, 1 we wanted to determine the function of nuclear BAG-1 in these non-tumourigenic cells, to identify whether nuclear BAG-1 was implicated in tumour progression in the colon. In the current report we established that nuclear BAG-1 inhibits apoptosis in a col- orectal adenoma-derived cell line. We demonstrate that apoptosis induced by γ -radiation or the vitamin D ana- logue EB1089 in the non-tumourigenic human colorectal adenoma-derived S/RG/C2 cell line, was preceded by a decrease in nuclear and an increase in cytoplasmic BAG- 1 expression. This change in subcellular localisation of BAG-1 was due to the redistribution of the BAG-1M iso- form. In addition, we have shown that the maintenance of high nuclear BAG-1 through enforced expression of the nuclear localised BAG-1L isoform enhanced cellu- lar survival after γ -radiation or exposure to EB1089. Fur- thermore the expression of cytoplasmic BAG-1S isoform fused with a nuclear localisation signal protected against γ -radiation induced apoptosis. This demonstrates that nuclear localisation of the BAG-1 protein confers a sur- vival advantage in colorectal adenoma-derived cells and that nuclear BAG-1 could potentially be an important sur- vival factor in colorectal carcinogenesis. Keywords: apoptosis; BAG-1; chemoprevention; colon cancer. Correspondence to: A. C. Williams, Cancer Research UK Col- orectal Tumour Biology Research Group, Department of Pathol- ogy and Microbiology, University of Bristol, School of Medical Sci- ences, Bristol BS8 1TD, U.K. Tel.: +44-117-9287892; Fax: +44- 117-928-7896, e-mail: [email protected] Introduction The Bcl-2 associated athanogene (BAG-1) was discovered as a novel regulator of apoptosis through its ability to bind to Bcl-2. 2 Recent studies have shown BAG-1 to be a multifunctional protein involved in differentiation, cell cycle and apoptosis. 3 The human BAG-1 gene is lo- cated on chromosome 9, 4 and encodes three major BAG- 1 isoforms, BAG-1S (p36), BAG-1M (p46) and BAG- 1L (p50), generated via alternate translation mechanisms from the same mRNA. 5–7 The different BAG-1 isoforms have distinct subcellular localisations within the cell. The smaller of the isoforms BAG-1S, is preferentially located in the cytoplasm, the BAG-1M isoform is detected in both the nuclear and cytoplasmic compartments, and the BAG- 1L isoform is predominantly located in the nucleus. 5–9 The difference in the subcellular localisation of the BAG-1 isoforms is thought to be conferred at least in part by a nu- clear localisation signal (NLS) present in the N-terminus of the BAG-1L isoform, but absent in the BAG-1S and truncated in the BAG-1M isoform. 5–7,9 BAG-1 has previously been shown to suppress apop- tosis in various cell types although the precise mecha- nism remains to be fully established. 3 BAG-1 has been shown to interact with various cellular proteins in differ- ent subcellular compartments, for example: nuclear hor- mone receptors and Rb in the nucleus; 1,9,10 tyrosine- kinase receptors for the platelet-derived or hepatocyte growth factors on the outer cell membrane; 11 Raf-1 ki- nase in the cytosol 12,13 and the Bcl-2 protein at the inner mitochondrial membrane. 2 Among these diverse protein interactions, it is likely that several are mediated via BAG- 1‘s ability to function as a nucleotide exchange factor for the 70 kD heat-shock chaperone protein. 14 The subcel- lular localisation of BAG-1 can be altered under certain physiological conditions: the BAG-1S and BAG-1M iso- forms have been shown to shuttle to the nucleus following heat shock 15,16 or when bound to liganded glucocorticoid Apoptosis · Vol 10 · No 2 · 2005 301
Transcript of Nuclear BAG1 expression inhibits apoptosis in colorectal adenoma-derived epithelial cells

Apoptosis 2005; 10: 301–311C© 2005 Springer Science + Business Media, Inc. Manufactured in The Netherlands.
Nuclear BAG-1 expression inhibits apoptosis incolorectal adenoma-derived epithelial cells
J. D. Barnes, N. J. Arhel, S. S. Lee, A. Sharp, M. Al-Okail, G. Packham, A. Hague, C. Paraskevaand A. C. Williams
Cancer Research UK Colorectal Tumour Biology Research Group, Department of Pathology and Microbiology, Schoolof Medical Sciences, University of Bristol, Bristol, UK (J. D. Barnes, N. J. Arhel, C. Paraskeva, A. C. Williams);Department of Oral and Dental Science, University of Bristol, Lower Maudlin Street, Bristol, UK (S. S. Lee, A. Hague);Cancer Research UK Oncology Unit, Cancer Science Division, University of Southampton School of Medicine,Southampton General Hospital, Southampton, UK (A. Sharp, M. Al-Okail, G. Packham)
BAG-1 is an anti-apoptotic protein that is frequentlyderegulated in a variety of malignancies including col-orectal cancer. There are three isoforms: BAG-1L is lo-cated in the nucleus, BAG-1M and BAG-1S are locatedboth in the nucleus and the cytoplasm. In colon cancer,the expression of nuclear BAG-1 is associated with poorerprognosis and is potentially a useful predictive factorfor distant metastasis. However, the function of BAG-1 incolonic epithelial cells has not been studied. Having pre-viously shown a predominant nuclear localisation of BAG-1 in adenoma-derived cell lines,1 we wanted to determinethe function of nuclear BAG-1 in these non-tumourigeniccells, to identify whether nuclear BAG-1 was implicated intumour progression in the colon. In the current report weestablished that nuclear BAG-1 inhibits apoptosis in a col-orectal adenoma-derived cell line. We demonstrate thatapoptosis induced by γ -radiation or the vitamin D ana-logue EB1089 in the non-tumourigenic human colorectaladenoma-derived S/RG/C2 cell line, was preceded by adecrease in nuclear and an increase in cytoplasmic BAG-1 expression. This change in subcellular localisation ofBAG-1 was due to the redistribution of the BAG-1M iso-form. In addition, we have shown that the maintenanceof high nuclear BAG-1 through enforced expression ofthe nuclear localised BAG-1L isoform enhanced cellu-lar survival after γ -radiation or exposure to EB1089. Fur-thermore the expression of cytoplasmic BAG-1S isoformfused with a nuclear localisation signal protected againstγ -radiation induced apoptosis. This demonstrates thatnuclear localisation of the BAG-1 protein confers a sur-vival advantage in colorectal adenoma-derived cells andthat nuclear BAG-1 could potentially be an important sur-vival factor in colorectal carcinogenesis.
Keywords: apoptosis; BAG-1; chemoprevention; coloncancer.
Correspondence to: A. C. Williams, Cancer Research UK Col-orectal Tumour Biology Research Group, Department of Pathol-ogy and Microbiology, University of Bristol, School of Medical Sci-ences, Bristol BS8 1TD, U.K. Tel.: +44-117-9287892; Fax: +44-117-928-7896, e-mail: [email protected]
Introduction
The Bcl-2 associated athanogene (BAG-1) was discoveredas a novel regulator of apoptosis through its ability tobind to Bcl-2.2 Recent studies have shown BAG-1 tobe a multifunctional protein involved in differentiation,cell cycle and apoptosis.3 The human BAG-1 gene is lo-cated on chromosome 9,4 and encodes three major BAG-1 isoforms, BAG-1S (p36), BAG-1M (p46) and BAG-1L (p50), generated via alternate translation mechanismsfrom the same mRNA.5–7 The different BAG-1 isoformshave distinct subcellular localisations within the cell. Thesmaller of the isoforms BAG-1S, is preferentially locatedin the cytoplasm, the BAG-1M isoform is detected in boththe nuclear and cytoplasmic compartments, and the BAG-1L isoform is predominantly located in the nucleus.5–9
The difference in the subcellular localisation of the BAG-1isoforms is thought to be conferred at least in part by a nu-clear localisation signal (NLS) present in the N-terminusof the BAG-1L isoform, but absent in the BAG-1S andtruncated in the BAG-1M isoform.5–7,9
BAG-1 has previously been shown to suppress apop-tosis in various cell types although the precise mecha-nism remains to be fully established.3 BAG-1 has beenshown to interact with various cellular proteins in differ-ent subcellular compartments, for example: nuclear hor-mone receptors and Rb in the nucleus;1,9,10 tyrosine-kinase receptors for the platelet-derived or hepatocytegrowth factors on the outer cell membrane;11 Raf-1 ki-nase in the cytosol12,13 and the Bcl-2 protein at the innermitochondrial membrane.2 Among these diverse proteininteractions, it is likely that several are mediated via BAG-1‘s ability to function as a nucleotide exchange factor forthe 70 kD heat-shock chaperone protein.14 The subcel-lular localisation of BAG-1 can be altered under certainphysiological conditions: the BAG-1S and BAG-1M iso-forms have been shown to shuttle to the nucleus followingheat shock15,16 or when bound to liganded glucocorticoid
Apoptosis · Vol 10 · No 2 · 2005 301

J.D. Barnes et al.
receptor.17 BAG-1 has also been reported to move fromthe nucleus to the cytoplasm during neuronal and epi-dermal differentiation.7,18 Furthermore BAG-1 reloca-tion is seen following the overexpression of cellular pro-teins such as: the expression of the human papillomavirusE7 oncoprotein,1 the human SINA-homologous (Siah-1A) protein,19 the pro-Heparin binding EGF-like growthfactor20 and the Bcl-2 protein.2 Therefore the subcellularlocalisation of BAG-1 may represent a dynamic mech-anism for regulating the biological functions of BAG-1within the cell.
BAG-1 protein expression is frequently deregulated incancer, including carcinomas of the breast, lung, cervix,colon, oesophagus and larynx.21 In the colon, it has beenreported that there is differential BAG-1 localisation inthe normal colonic tissue, with higher levels of nuclearlocalised BAG-1 protein at the bottom of the colonic cryptwhich correlates with Bcl-2 expression, in the zone ofproliferation.7 Furthermore, BAG-1 protein expression ishigher in tumours than in normal colonic epithelium.7
Although overexpression of both the BAG-1S and BAG-1L isoforms have been shown to inhibit apoptosis inducedby a wide variety of agents in C33A cervical carcinomas22
and ZR-75-1 and MCF7 breast cancer cells,16,23 there isno data available on the effect of increased BAG-1 on cellsurvival in colonic epithelial cells. Recently it has beenreported that nuclear BAG-1 expression reflects poorerprognosis and is potentially a useful predictive factor fordistant metastasis in patients suffering from colorectalcancer.24 This suggests that nuclear BAG-1L could beimportant in the development of colorectal cancer.
The function of BAG-1 in colonic epithelial cells hasnot been previously investigated. Using the adenoma-derived S/RG/C2 cell line,1 we investigated the functionof BAG-1 in this non tumourigenic cell line, to determinewhether nuclear BAG-1 has a role in early colorectal tu-mourigenesis. Having established endogenous BAG-1 ex-pression and subcellular localisation, we analysed whetherthe expression of nuclear BAG-1L enhanced cellular resis-tance to either γ -radiation or EB1089 induced apoptosis.In addition, we studied the importance of BAG-1 localisa-tion by expressing either the nuclear localised or excludedBAG-1S isoform and assessing survival after γ -radiation.This study establishes that nuclear localised BAG-1 con-fers a survival advantage on adenoma derived cells, andsuggests that the nuclear localisation of BAG-1, whichallows cells to evade apoptosis, may be important in pro-moting tumourigenesis in the colon.
Materials and methods
Cell line and cell culture
The S/RG/C2 cell line is a clonogenic non-tumourigenicderivative of an adenoma-derived cell line.25,26 S/RG/C2
cells were grown in Dulbecco‘s Modified EaglesMedium (DMEM, Gibco), supplemented with penicillin100 units/ml, streptomycin 100 µg/ml, hydrocortisone1 µg/ml, insulin 0.2 units/ml, glutamine 2 mM and20% foetal bovine serum (FBS) (referred to as 20% DMEMmedium). NIH3T3 mouse fibroblasts and MCF7 humanbreast cancer cells were maintained in DMEM supple-mented with 10% (v/v) fetal calf serum, 2 mM glutamine,100 units/ml penicillin and 0.2 units/ml streptomycin.
Transfection of cells
The adenoma-derived cell line S/RG/C2 was stablytransfected with pcDNA3-derived expression plasmids(Invitrogen, UK) encoding human BAG-1L, or BAG-1Sfused to the SV40 large T-antigen NLS (NLS-BAG-1S).10 Empty pcDNA3 vector (Invitrogen) was usedas a negative control. Plasmid NES-BAG-1S encodeshuman BAG-1S fused at its N-terminus to a nuclearexport sequence (NES) derived from MAPKK,27 andwas generated by PCR amplification of the humanBAG-1S open reading frame using primers p36-NES5′CCCAAGCTTGAAGAGATGGTGGACCTCCAAA-AGAAGCTGGAGGAGCTGGAGCTGAATCGGAG-CCAGGAGGTG and p. 36-3′CCGCTCGAGTGCTAC-ACCTCACTCGGCCAGGGC and cloning the productin to pcDNA3 (Invitrogen) that had been linearised usingHindIII and XhoI. Transfection was carried out usinglipofectamine 2000, according to the manufacturer’sinstructions (Invitrogen, UK). Transfected cells wereselected using 400 µg/ml G418 (Sigma, UK) in 20%DMEM. Single colonies were picked and resulting linesmaintained with 200 µg/ml G418 selection medium.The overexpression of BAG-1 proteins was confirmedby western blotting analysis. NIH3T3 and MCF7 cellswere transfected using Fugene 6 (Roche) according tothe manufactures instructions.
Gamma-radiation and EB1089 treatment
Cells were exposed to 5 Gy γ -radiation from a137Cs source(Gravaton Generator) at a dose of 0.33 Gy per minute,a dose previously shown to induce significant apoptosisin the S/RG/C2 cell line.28EB1089 was kindly providedby L. Binderup (Department of Chemical Research, LeoPharmaceuticals, Denmark). Cultures were treated withEB1089 at a concentration of 1 × 10−7 M, a dose pre-viously reported to induce apoptosis in the S/RG/C2 cellline.29
Western blotting
Adherent cells were collected by trypsinization and cen-trifugation, and resuspended (1× 106 cells per sample) in
302 Apoptosis · Vol 10 · No 2 · 2005

Nuclear BAG-1 inhibits apoptosis in colonic adenoma cells
1× sample buffer (10% glycerol, 63 mM Tris HCL, pH6.8, 5% 2-mercaptoethanol, 2% SDS, 0.025% bromophe-nol blue) and boiled for 5 min. Protein samples wereresolved on 12.5% polyacrylamide gel by SDS-PAGE.BAG-1 protein was detected by using monoclonal anti-body (G3E2) which recognizes all three BAG-1 isoforms8
and a horseradish peroxidase-conjugated goat-anti-mouseantibody (1:1000 dilution). Bound immunocomplexeswere visualized using the Amersham enhanced chemi-luminescence (ECL) detection system, according to themanufacturer’s protocol.
GFP-tagged BAG-1 isoforms
BAG-1 isoforms BAG-1S, BAG-1M and BAG-1L werecloned into pEGFP-N1 and pEGFP-C1 vectors (Clon-tech, UK) using a 5′ EcoRI and a 3′ BamHI site. Forwardprimers for cloning into the pEGFP-N1 vector are 5′-CAGAATTCGGATGAAGAAGAAAACCCGGC-3′ forBAG-1M, 5′-CAGAATTCGCATGGCTCAGCGCG-GGGGG-3′ for BAG-1L; common reverse primer is 5′-CAGGATCCTCGGCCAGGGCAAAGTTTG-3′ . For-ward primers for cloning into the pEGFP-C1 vector are5′-CAGAATTCGATGAAGAAGAAAACCCGGCG-3′
for BAG-1S, 5′-CAGAATTCGATGAATCGGAGCC-AGGAGG-3′ for BAG-1M, 5′-CAGAATTCCATGGC-TCAGCGCGGGGGG-3′ for BAG-1L; common re-verse primer is 5′-CAGGATCCTCACTCGGCCAGG-GCAAAG-3′ . Primers were synthesised by MWG-Biotech AG.
The S/RG/C2 cell line was transiently transfected withexpression plasmids encoding pEGFP-tagged BAG-1L,BAG-1M and BAG-1S, using Transfast reagent (Promega,UK) according to the manufacturer’s instructions. Thesubcellular localisation of EGFP-tagged isoforms was as-sessed using confocal microscopy at 24 hr and 48 hr fol-lowing irradiation. Each image represents a stacked im-age of approximately 20 slices at 0.5 microns thicknessthrough the cell. Quantification of EGFP intensities wascarried out using the Inverted Leica TCS-NT (Leica Mi-crosystems Wetchar, Germany) confocal laser scanningmicroscope with Leica TCS-NT software for 2D and 3Dimage analysis. The total nuclear and cytoplasmic EGFPintensity from a single slice through an individual cell wascalculated, and used to establish the cytoplasmic EGFPintensity as a proportion of the total. The percentage in-crease in cytoplasmic EGFP was then calculated betweencontrol and irradiated cells. This was determined for 20individual cells from 10 separate experiments.
Confocal microscopy
Cells were grown on collagen Type IV-coated glass cover-slips in 6 well petri dishes, which were supplemented with
20% DMEM medium. Cells were fixed and permeabilisedby incubation with 4% (w/v) paraformaldehyde contain-ing 0.2% (v/v) Triton X-100. BAG-1 was visualised usingthe polyclonal anti-BAG-1 (TB3) antibody which recog-nizes all 3 BAG-1 isoforms,8 and anti-rabbit ALEXA-546(equivalent to TRITC wavelength) (Sigma, UK). Cellswere mounted with Vectashield mounting medium withDAPI (1.5 ug/ml, Vector Labs, Bulingame, CA) to allownuclei to be visualised. Cells were visualised by confo-cal microscopy (as described earlier). The localisation ofBAG-1S, BAG-1L and NES and NLS fusion proteins wasanalysed in MCF7 cells as previously described.10
Luciferase reporter assay
Cells were transiently transfected with either the vita-min D reporter plasmid DR3-TA-Luc or with the controlplasmid pTA-Luc (Clontech, Oxford, UK). All transfec-tions included the Renilla luciferase vector pRL-SV40(Promega, UK) as an internal control for transfection effi-ciency. Reporter activity was measured using the Dual Lu-ciferase reporter assay system (Promega, UK). Data werecalculated as the ratio between DR3-TA-Luc or pTA-Lucand pRL-SV40 activity and represented as a fold increasebetween the control and EB1089-treated cultures for eachof the triplicate experiments.
Assessment of apoptosis
The induction of apoptosis was measured by determiningthe proportion of total cells that had detached from the ep-ithelial monolayer and that were floating in the medium,shown to be apoptotic as previously described using acri-dine orange to detect morphological apoptosis and PARPcleavage28,29.
Statistical analysis
Statistical analysis was carried out using the SPSS soft-ware. A Dunnett‘s t-test was performed to calculatethe significance of the protection from apoptosis inboth BAG-1L and NLS/NES-BAG-1S transfection exper-iments.
Results
Relocation of BAG-1 protein after γ -radiation incolonic epithelial cells
To address the potential role of BAG-1 in control ofcell survival in colonic epithelial cells, we examined
Apoptosis · Vol 10 · No 2 · 2005 303
J.D. Barnes et al.
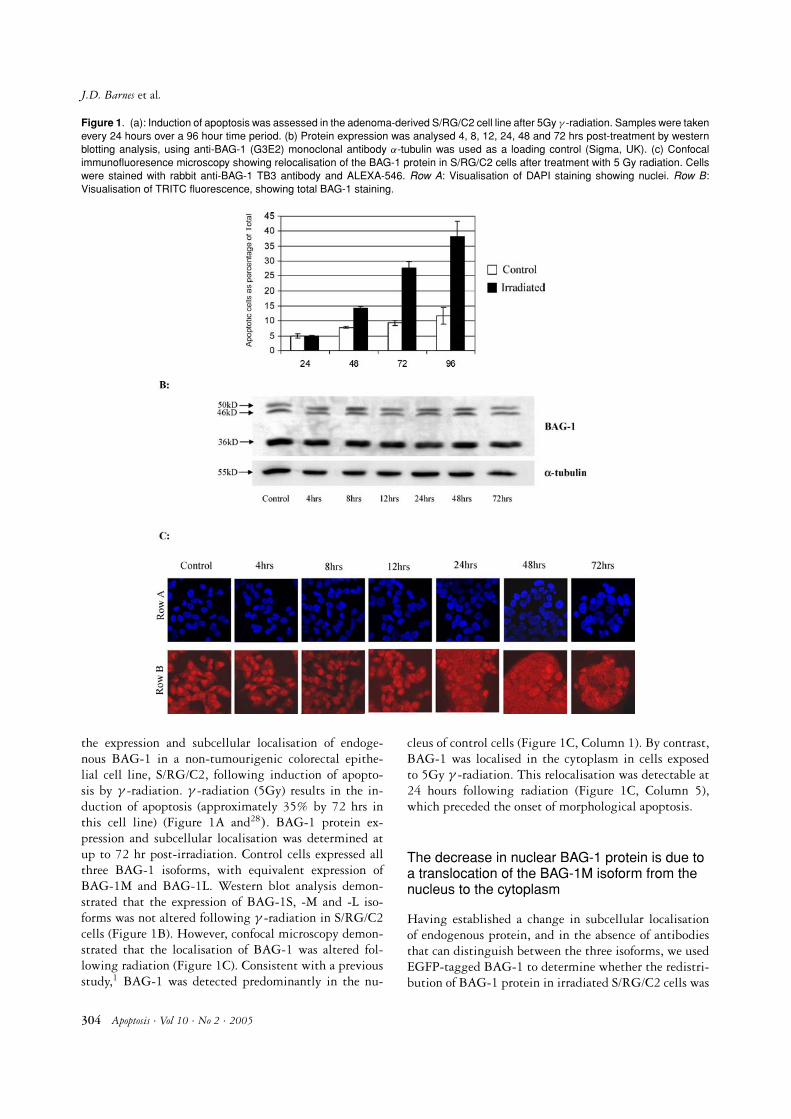
Figure 1. (a): Induction of apoptosis was assessed in the adenoma-derived S/RG/C2 cell line after 5Gy γ -radiation. Samples were takenevery 24 hours over a 96 hour time period. (b) Protein expression was analysed 4, 8, 12, 24, 48 and 72 hrs post-treatment by westernblotting analysis, using anti-BAG-1 (G3E2) monoclonal antibody α-tubulin was used as a loading control (Sigma, UK). (c) Confocalimmunofluoresence microscopy showing relocalisation of the BAG-1 protein in S/RG/C2 cells after treatment with 5 Gy radiation. Cellswere stained with rabbit anti-BAG-1 TB3 antibody and ALEXA-546. Row A: Visualisation of DAPI staining showing nuclei. Row B:Visualisation of TRITC fluorescence, showing total BAG-1 staining.
the expression and subcellular localisation of endoge-nous BAG-1 in a non-tumourigenic colorectal epithe-lial cell line, S/RG/C2, following induction of apopto-sis by γ -radiation. γ -radiation (5Gy) results in the in-duction of apoptosis (approximately 35% by 72 hrs inthis cell line) (Figure 1A and28). BAG-1 protein ex-pression and subcellular localisation was determined atup to 72 hr post-irradiation. Control cells expressed allthree BAG-1 isoforms, with equivalent expression ofBAG-1M and BAG-1L. Western blot analysis demon-strated that the expression of BAG-1S, -M and -L iso-forms was not altered following γ -radiation in S/RG/C2cells (Figure 1B). However, confocal microscopy demon-strated that the localisation of BAG-1 was altered fol-lowing radiation (Figure 1C). Consistent with a previousstudy,1 BAG-1 was detected predominantly in the nu-
cleus of control cells (Figure 1C, Column 1). By contrast,BAG-1 was localised in the cytoplasm in cells exposedto 5Gy γ -radiation. This relocalisation was detectable at24 hours following radiation (Figure 1C, Column 5),which preceded the onset of morphological apoptosis.
The decrease in nuclear BAG-1 protein is due toa translocation of the BAG-1M isoform from thenucleus to the cytoplasm
Having established a change in subcellular localisationof endogenous protein, and in the absence of antibodiesthat can distinguish between the three isoforms, we usedEGFP-tagged BAG-1 to determine whether the redistri-bution of BAG-1 protein in irradiated S/RG/C2 cells was
304 Apoptosis · Vol 10 · No 2 · 2005
Nuclear BAG-1 inhibits apoptosis in colonic adenoma cells
due to alterations in the localisation of a specific BAG-1isoform. Confocal microscopy was used to determine thelocalisation of EGFP-tagged BAG-1 isoforms in untreatedcontrol and γ -irradiated cultures. The intensity of theEGFP fluorescence in the nucleus and cytoplasm of trans-fected cells was quantified using Leica TCS-NT software.
(A)
(B)
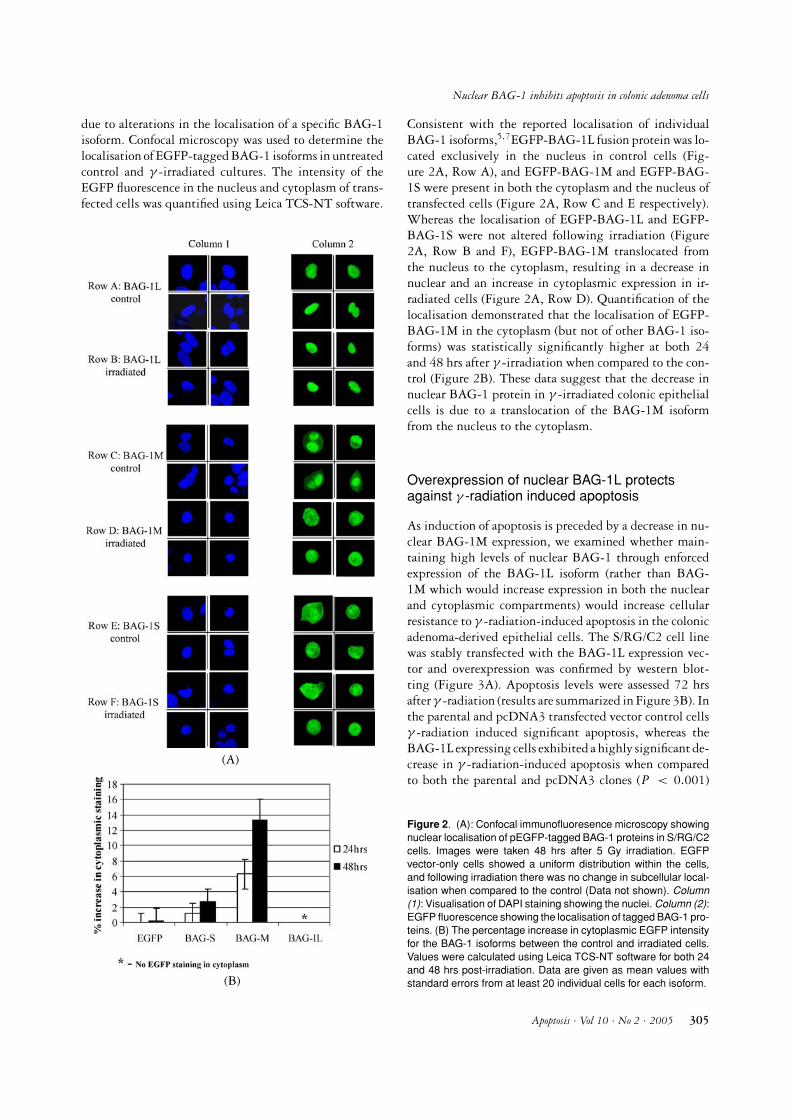
Consistent with the reported localisation of individualBAG-1 isoforms,5,7EGFP-BAG-1L fusion protein was lo-cated exclusively in the nucleus in control cells (Fig-ure 2A, Row A), and EGFP-BAG-1M and EGFP-BAG-1S were present in both the cytoplasm and the nucleus oftransfected cells (Figure 2A, Row C and E respectively).Whereas the localisation of EGFP-BAG-1L and EGFP-BAG-1S were not altered following irradiation (Figure2A, Row B and F), EGFP-BAG-1M translocated fromthe nucleus to the cytoplasm, resulting in a decrease innuclear and an increase in cytoplasmic expression in ir-radiated cells (Figure 2A, Row D). Quantification of thelocalisation demonstrated that the localisation of EGFP-BAG-1M in the cytoplasm (but not of other BAG-1 iso-forms) was statistically significantly higher at both 24and 48 hrs after γ -irradiation when compared to the con-trol (Figure 2B). These data suggest that the decrease innuclear BAG-1 protein in γ -irradiated colonic epithelialcells is due to a translocation of the BAG-1M isoformfrom the nucleus to the cytoplasm.
Overexpression of nuclear BAG-1L protectsagainst γ -radiation induced apoptosis
As induction of apoptosis is preceded by a decrease in nu-clear BAG-1M expression, we examined whether main-taining high levels of nuclear BAG-1 through enforcedexpression of the BAG-1L isoform (rather than BAG-1M which would increase expression in both the nuclearand cytoplasmic compartments) would increase cellularresistance to γ -radiation-induced apoptosis in the colonicadenoma-derived epithelial cells. The S/RG/C2 cell linewas stably transfected with the BAG-1L expression vec-tor and overexpression was confirmed by western blot-ting (Figure 3A). Apoptosis levels were assessed 72 hrsafter γ -radiation (results are summarized in Figure 3B). Inthe parental and pcDNA3 transfected vector control cellsγ -radiation induced significant apoptosis, whereas theBAG-1L expressing cells exhibited a highly significant de-crease in γ -radiation-induced apoptosis when comparedto both the parental and pcDNA3 clones (P < 0.001)
Figure 2. (A): Confocal immunofluoresence microscopy showingnuclear localisation of pEGFP-tagged BAG-1 proteins in S/RG/C2cells. Images were taken 48 hrs after 5 Gy irradiation. EGFPvector-only cells showed a uniform distribution within the cells,and following irradiation there was no change in subcellular local-isation when compared to the control (Data not shown). Column(1): Visualisation of DAPI staining showing the nuclei. Column (2):EGFP fluorescence showing the localisation of tagged BAG-1 pro-teins. (B) The percentage increase in cytoplasmic EGFP intensityfor the BAG-1 isoforms between the control and irradiated cells.Values were calculated using Leica TCS-NT software for both 24and 48 hrs post-irradiation. Data are given as mean values withstandard errors from at least 20 individual cells for each isoform.
Apoptosis · Vol 10 · No 2 · 2005 305
J.D. Barnes et al.
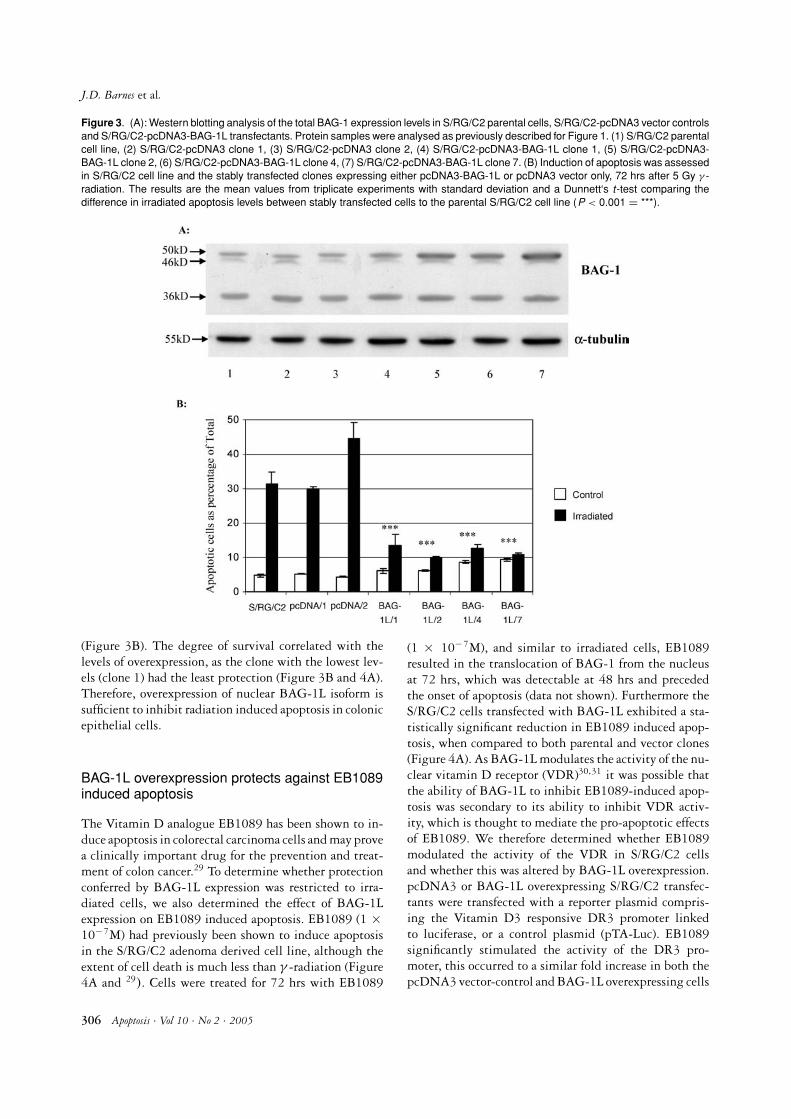
Figure 3. (A): Western blotting analysis of the total BAG-1 expression levels in S/RG/C2 parental cells, S/RG/C2-pcDNA3 vector controlsand S/RG/C2-pcDNA3-BAG-1L transfectants. Protein samples were analysed as previously described for Figure 1. (1) S/RG/C2 parentalcell line, (2) S/RG/C2-pcDNA3 clone 1, (3) S/RG/C2-pcDNA3 clone 2, (4) S/RG/C2-pcDNA3-BAG-1L clone 1, (5) S/RG/C2-pcDNA3-BAG-1L clone 2, (6) S/RG/C2-pcDNA3-BAG-1L clone 4, (7) S/RG/C2-pcDNA3-BAG-1L clone 7. (B) Induction of apoptosis was assessedin S/RG/C2 cell line and the stably transfected clones expressing either pcDNA3-BAG-1L or pcDNA3 vector only, 72 hrs after 5 Gy γ -radiation. The results are the mean values from triplicate experiments with standard deviation and a Dunnett‘s t-test comparing thedifference in irradiated apoptosis levels between stably transfected cells to the parental S/RG/C2 cell line (P < 0.001 = ***).
(Figure 3B). The degree of survival correlated with thelevels of overexpression, as the clone with the lowest lev-els (clone 1) had the least protection (Figure 3B and 4A).Therefore, overexpression of nuclear BAG-1L isoform issufficient to inhibit radiation induced apoptosis in colonicepithelial cells.
BAG-1L overexpression protects against EB1089induced apoptosis
The Vitamin D analogue EB1089 has been shown to in-duce apoptosis in colorectal carcinoma cells and may provea clinically important drug for the prevention and treat-ment of colon cancer.29 To determine whether protectionconferred by BAG-1L expression was restricted to irra-diated cells, we also determined the effect of BAG-1Lexpression on EB1089 induced apoptosis. EB1089 (1 ×10−7M) had previously been shown to induce apoptosisin the S/RG/C2 adenoma derived cell line, although theextent of cell death is much less than γ -radiation (Figure4A and 29). Cells were treated for 72 hrs with EB1089
(1 × 10−7M), and similar to irradiated cells, EB1089resulted in the translocation of BAG-1 from the nucleusat 72 hrs, which was detectable at 48 hrs and precededthe onset of apoptosis (data not shown). Furthermore theS/RG/C2 cells transfected with BAG-1L exhibited a sta-tistically significant reduction in EB1089 induced apop-tosis, when compared to both parental and vector clones(Figure 4A). As BAG-1L modulates the activity of the nu-clear vitamin D receptor (VDR)30,31 it was possible thatthe ability of BAG-1L to inhibit EB1089-induced apop-tosis was secondary to its ability to inhibit VDR activ-ity, which is thought to mediate the pro-apoptotic effectsof EB1089. We therefore determined whether EB1089modulated the activity of the VDR in S/RG/C2 cellsand whether this was altered by BAG-1L overexpression.pcDNA3 or BAG-1L overexpressing S/RG/C2 transfec-tants were transfected with a reporter plasmid compris-ing the Vitamin D3 responsive DR3 promoter linkedto luciferase, or a control plasmid (pTA-Luc). EB1089significantly stimulated the activity of the DR3 pro-moter, this occurred to a similar fold increase in both thepcDNA3 vector-control and BAG-1L overexpressing cells
306 Apoptosis · Vol 10 · No 2 · 2005

Nuclear BAG-1 inhibits apoptosis in colonic adenoma cells
Figure 4. (A): Fold increase in apoptosis between the control andEB1089 (1× 10−7 M) treated in the parental S/RG/C2 cell line andstably transfected with pcDNA3-BAG-1L or pcDNA3 after 72 hrstreatment. The results are the mean values and standard devi-ation from triplicate experiments, a Dunnett‘s t-test was used tocompare the difference in apoptosis levels between EB1089 treat-ment of stable clones to the parental cell line (P < 0.01 = ** andP < 0.001 = ***). (B) Dual luciferase assay from the S/RG/C2cells transiently transfected with either pcDNA3 or pcDNA3-BAG-1L (See materials and methods). This data represents the foldincrease in the reporter activity between pTA-Luc and DR3-TA-Luc for both the control and EB1089 treated in both cell lines, andis representative of three independent experiments.
(Figure 4B). Therefore, the survival effect of BAG-1L isnot mediated via a direct effect on the ability of EB1089to activate the VDR in these cells.
Expression of NLS-BAG-1S protects whileNES-BAG-1S fails to protect against γ -radiationinduced apoptosis
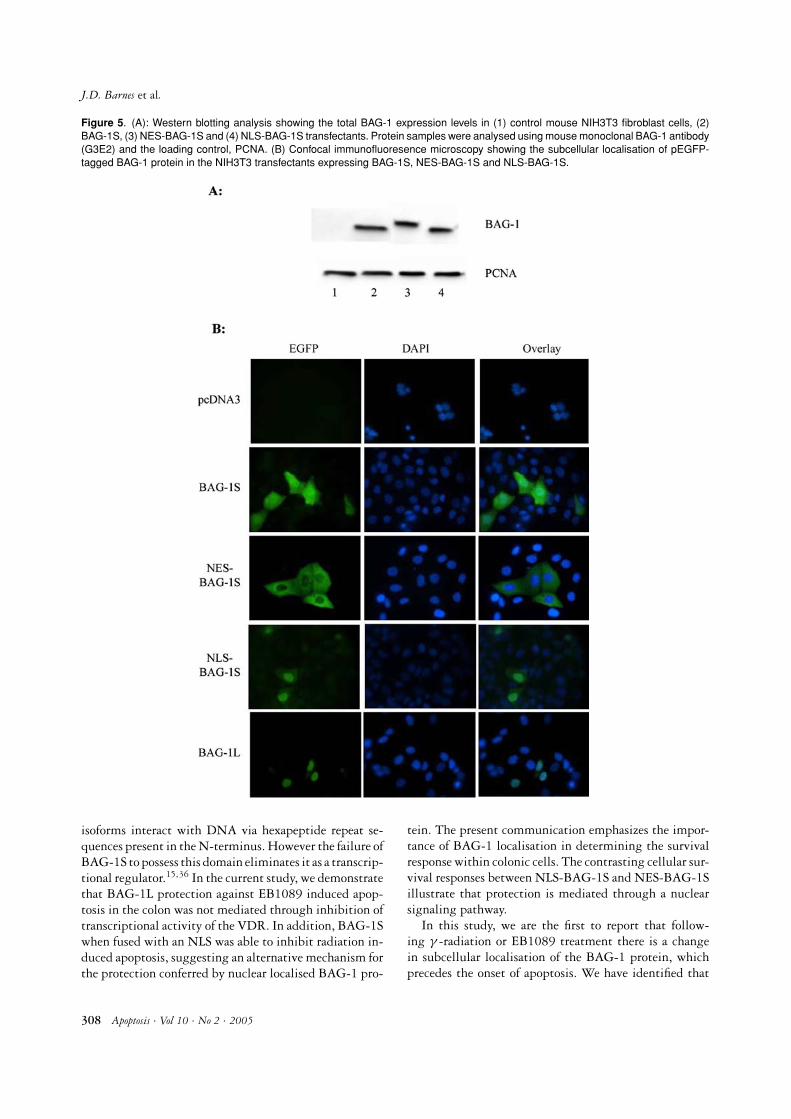
Using expression plasmids encoding BAG-1S fused atits N-terminus to a heterologous nuclear localisation se-quence (NLS)10 or a nuclear export sequence (NES), wedetermined the importance of the localisation of BAG-1 on cell survival. Transfection of mouse NIH3T3 fi-broblasts demonstrated that endogenous BAG-1 and fu-sion BAG-1S proteins were expressed at equivalent levels(Figure 5A). We used NIH3T3 cells for this analysis sincethese cells are readily transfectable and using a humanBAG-1 specific antibody, we were able to avoid detectionof mouse endogenous BAG-1, which would confound theanalysis of subcellular localisation of transfected BAG-
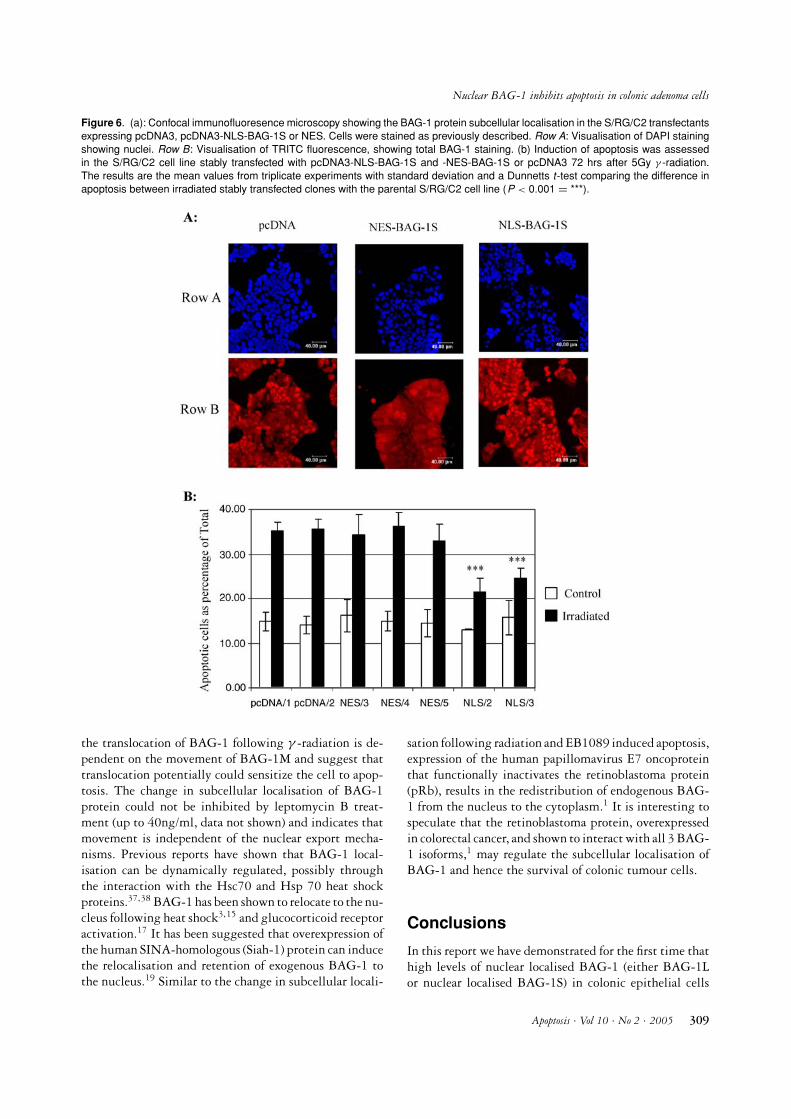
1 protein. When overexpressed NES-BAG-1S was effec-tively excluded from the nucleus, whereas NLS-BAG-1S was localised similar to BAG-1L: in the nucleus(Figure 5B). S/RG/C2 cells were stably transfected withthese expression vectors and clones were isolated (Fig-ure 6A). Cells were irradiated with 5Gy and apoptosiswas assessed after 72 hrs. The results shown in Figure6B illustrate that the NES-BAG-1S overexpressing cloneswere as sensitive to γ -radiation induced apoptosis as theparental and vector only cells. However, enforced expres-sion of NLS-BAG-1S (Clone 2 and 3) conferred a signifi-cant resistance to apoptosis when treated with γ -radiation(P < 0.001). Therefore nuclear localisation of BAG-1Sprotein is able to confer protection against γ -radiationinduced apoptosis. These results support above data, thatit is the nuclear localisation of the BAG-1 protein that isimportant for its anti-apoptotic function in these colonicepithelial cells.
Discussion
BAG-1 expression has been shown, to be altered in a vari-ety of malignancies when compared to normal epithelialcells.32 Interestingly in colon cancer, BAG-1 is overex-pressed and nuclear BAG-1 expression is associated withpoor survival.7,24 In the current study we have shown thatthe maintenance of high levels of nuclear BAG-1 expres-sion, through either BAG-1L or nuclear localised BAG-1S, caused resistance to γ -radiation induced apoptosis.This is the first study to demonstrate that nuclear BAG-1inhibits the induction of apoptosis in colorectal adenoma-derived cells, and suggests that nuclear localised BAG-1expression may confer a survival advantage on adenomacells in vivo, which could be important in the early stagesof colorectal carcinogenesis.
Although BAG-1 has been shown to modulate thefunction of cytoplasmic proteins Bcl-2 and Raf-1,2,12 themechanisms by which nuclear localised BAG-1 may pro-tect against apoptosis remains to be elucidated. A keytarget for nuclear BAG-1 is the modification of nuclearhormone receptors (NHR). BAG-1 has been implicatedin NHR regulation through the alteration of receptor ac-tivity and has subsequently been suggested to be involvedin promoting cellular survival.33 The larger BAG-1 pro-teins (BAG-1L and in some cases BAG-1M) have beenreported to bind to and/or alter the transactivation of avariety of NHRs such as vitamin D, glucocorticoid, an-drogen and oestrogen receptors.10,30,31,34,35 However, thesmaller BAG-1S isoform failed to deactivate the Vita-min D receptor30 or alter the function of either the oe-strogen or androgen receptors.9,10 A second mechanismby which nuclear BAG-1 could enhance cell survival isthrough the transcriptional regulation of genes achievedby direct binding to DNA. Both BAG-1L and BAG-1M
Apoptosis · Vol 10 · No 2 · 2005 307
J.D. Barnes et al.
Figure 5. (A): Western blotting analysis showing the total BAG-1 expression levels in (1) control mouse NIH3T3 fibroblast cells, (2)BAG-1S, (3) NES-BAG-1S and (4) NLS-BAG-1S transfectants. Protein samples were analysed using mouse monoclonal BAG-1 antibody(G3E2) and the loading control, PCNA. (B) Confocal immunofluoresence microscopy showing the subcellular localisation of pEGFP-tagged BAG-1 protein in the NIH3T3 transfectants expressing BAG-1S, NES-BAG-1S and NLS-BAG-1S.
isoforms interact with DNA via hexapeptide repeat se-quences present in the N-terminus. However the failure ofBAG-1S to possess this domain eliminates it as a transcrip-tional regulator.15,36 In the current study, we demonstratethat BAG-1L protection against EB1089 induced apop-tosis in the colon was not mediated through inhibition oftranscriptional activity of the VDR. In addition, BAG-1Swhen fused with an NLS was able to inhibit radiation in-duced apoptosis, suggesting an alternative mechanism forthe protection conferred by nuclear localised BAG-1 pro-
tein. The present communication emphasizes the impor-tance of BAG-1 localisation in determining the survivalresponse within colonic cells. The contrasting cellular sur-vival responses between NLS-BAG-1S and NES-BAG-1Sillustrate that protection is mediated through a nuclearsignaling pathway.
In this study, we are the first to report that follow-ing γ -radiation or EB1089 treatment there is a changein subcellular localisation of the BAG-1 protein, whichprecedes the onset of apoptosis. We have identified that
308 Apoptosis · Vol 10 · No 2 · 2005
Nuclear BAG-1 inhibits apoptosis in colonic adenoma cells
Figure 6. (a): Confocal immunofluoresence microscopy showing the BAG-1 protein subcellular localisation in the S/RG/C2 transfectantsexpressing pcDNA3, pcDNA3-NLS-BAG-1S or NES. Cells were stained as previously described. Row A: Visualisation of DAPI stainingshowing nuclei. Row B: Visualisation of TRITC fluorescence, showing total BAG-1 staining. (b) Induction of apoptosis was assessedin the S/RG/C2 cell line stably transfected with pcDNA3-NLS-BAG-1S and -NES-BAG-1S or pcDNA3 72 hrs after 5Gy γ -radiation.The results are the mean values from triplicate experiments with standard deviation and a Dunnetts t-test comparing the difference inapoptosis between irradiated stably transfected clones with the parental S/RG/C2 cell line (P < 0.001 = ***).
the translocation of BAG-1 following γ -radiation is de-pendent on the movement of BAG-1M and suggest thattranslocation potentially could sensitize the cell to apop-tosis. The change in subcellular localisation of BAG-1protein could not be inhibited by leptomycin B treat-ment (up to 40ng/ml, data not shown) and indicates thatmovement is independent of the nuclear export mecha-nisms. Previous reports have shown that BAG-1 local-isation can be dynamically regulated, possibly throughthe interaction with the Hsc70 and Hsp 70 heat shockproteins.37,38 BAG-1 has been shown to relocate to the nu-cleus following heat shock3,15 and glucocorticoid receptoractivation.17 It has been suggested that overexpression ofthe human SINA-homologous (Siah-1) protein can inducethe relocalisation and retention of exogenous BAG-1 tothe nucleus.19 Similar to the change in subcellular locali-
sation following radiation and EB1089 induced apoptosis,expression of the human papillomavirus E7 oncoproteinthat functionally inactivates the retinoblastoma protein(pRb), results in the redistribution of endogenous BAG-1 from the nucleus to the cytoplasm.1 It is interesting tospeculate that the retinoblastoma protein, overexpressedin colorectal cancer, and shown to interact with all 3 BAG-1 isoforms,1 may regulate the subcellular localisation ofBAG-1 and hence the survival of colonic tumour cells.
Conclusions
In this report we have demonstrated for the first time thathigh levels of nuclear localised BAG-1 (either BAG-1Lor nuclear localised BAG-1S) in colonic epithelial cells
Apoptosis · Vol 10 · No 2 · 2005 309

J.D. Barnes et al.
confers resistance to apoptotic signals. As nuclear BAG-1confers an advantage in-vitro, our hypothesis suggests thathigh levels in colorectal tumours may allow cells to evadeapoptosis. This raises a possible novel therapeutic targetin colorectal cancer, as disruption of the nuclear localisedBAG-1 in the tumour (for example as shown by1), wouldsensitize colonic epithelial cells to apoptosis and henceincrease the efficacy of current therapeutic regimes.
Acknowledgments
We would like to thank Dr Mark Jepson and Alan Leardfrom the School of Medical Sciences Cell Imaging Facilityfor their assistance.
This work was funded by Cancer Research UK, BreastCancer Campaign and Citrina Foundation. Majed Al-Okail was supported by a British Council Post-doctoralSummer Research Award. We thank the Medical ResearchCouncil for providing an Infrastructure Award to estab-lish the School of Medical Sciences Cell Imaging Facilityat the University of Bristol.
References
1. Arhel NJ, Packham G, Townsend PA, et al. The retinoblastomaprotein interacts with BAG-1 in human colonic adenoma andcarcinoma derived cell lines. Int J Cancer 2003; 106: 364–371.
2. Takayama S, Sato T, Krajewski S, et al. Cloning and functionalanalysis of BAG-1: A novel Bcl-2-binding protein with anti-cell death activity. Cell 1995; 80: 279–284.
3. Townsend PA, Cutress RI, Sharp A, Brimmell M, PackhamG. BAG-1: A multifunctional regulator of cell growth andsurvival. Biochim Biophys Acta 2003; 603(2): 83–98.
4. Takayama S, Kochel K, Irie S, et al. Cloning of cDNAs encodingthe human BAG1 protein and localisation of the human BAG1gene to chromosome 9p12. Genomics 1996; 35: 494-498.
5. Packham G, Brimmell M, Cleveland JL. Mammalian cells ex-press two differently localized BAG-1 isoforms generated byalternative translation initiation. Biochem J 1997; 328(Pt 3):807–813.
6. Yang X, Chernenko G, Hao Y, et al. Human BAG-1/RAP46protein is generated as four isoforms by alternative translationinitiation and overexpressed in cancer cells. Oncogene 1998; 17:981–989.
7. Takayama S, Krajewski S, Krajewska M, et al. Expression andlocation of Hsp70/Hsc-binding anti-apoptotic protein BAG-1and its variants in normal tissues and tumor cell lines. CancerRes 1998; 58: 3116–3131.
8. Brimmell M, Burns JS, Munson P, et al. High level expres-sion of differentially localized BAG-1 isoforms in some oestro-gen receptor-positive human breast cancers. Br J Cancer 1999;81(6): 1042–1051.
9. Knee DA, Froesch BA, Nuber U, Takayama S, Reed JC.Structure-function analysis of BAG-1 proteins. Effects on an-drogen receptor transcriptional activity. J Biol Chem 2001; 276:12718–12724.
10. Cutress RI, Townsend PA, Sharp A, et al. The nuclear BAG-1 isoform, BAG-1L, enhances oestrogen-dependent transcrip-tion. Oncogene 2003; 22: 4973–4982.
11. Bardelli A, Longati P, Albero D, et al. HGF receptor associateswith the anti-apoptotic protein BAG-1 and prevents cell death.EMBO J 1996; 15: 6205–6212.
12. Wang HG, Takayama S, Rapp UR, Reed JC. Bcl-2 interactingprotein, BAG-1, binds to and activates the kinase Raf-1. ProcNatl Acad Sci USA 1996; 93: 7063–7068.
13. Song J, Takeda M, Morimoto RI. Bag1-Hsp70 mediates a phys-iological stress signalling pathway that regulates Raf-1/ERKand cell growth. Nat Cell Biol 2001; 3: 276–282.
14. Hohfeld J, Jentsch S. GrpE-like regulation of the hsc70 chap-erone by the anti-apoptotic protein BAG-1. EMBO J 1997; 16:6209–6216.
15. Zeiner M, Niyaz Y, Gehring U. The hsp70-associating proteinHap46 binds to DNA and stimulates transcription. Proc NatlAcad Sci USA 1999; 96: 10194–10199.
16. Townsend PA, Cutress RI, Sharp A, Brimmell M, Packham G.BAG-1 prevents stress-induced long-term growth inhibition inbreast cancer cells via a chaperone-dependent pathway. CancerRes 2003; 63: 4150–4157.
17. Schneikert J, Hubner S, Martin E, Cato AC. A nuclear actionof the eukaryotic cochaperone RAP46 in downregulation ofglucocorticoid receptor activity. J Cell Biol 1999; 146: 929–940.
18. Kermer P, Krajewska M, Zapata JM, et al. Bag1 is a regulatorand marker of neuronal differentiation. Cell Death Differ 2002;4: 405–413.
19. Matsuzawa S, Takayama S, Froesch BA, Zapata JM, Reed JC.p53-inducible human homologue of Drosophila seven in absen-tia (Siah) inhibits cell growth: Suppression by BAG-1. EMBOJ 1998; 10: 2736–2747.
20. Lin J, Hutchinson L, Gaston SM, Raab G, Freeman MR. BAG-1 is a novel cytoplasmic binding partner of the membrane formof heparin-binding EGF-like growth factor: A unique role forproHB-EGF in cell survival regulation. J Biol Chem 2001; 276:30127–30132.
21. Sharp A, Crabb SJ, Cutress RI, Brimmell M, Wang XH,Packham G, Townsend PA. BAG-1 in carcinogenesis. ExpertRev Mol Med 2004; 1–15.
22. Yang X, Hao Y, Ding Z, Pater A. BAG-1 promotes apoptosisinduced by N-(4-hydroxyphenyl)retinamide in human cervicalcarcinoma cells. Exp Cell Res 2000; 256: 491–499.
23. Kudoh M, Knee DA, Takayama S, Reed JC. Bag1 proteinsregulate growth and survival of ZR-75-1 human breast cancercells. Cancer Res 2002; 62: 1904–1909.
24. Kikuchi R, Noguchi T, Takeno S, Funada Y, Moriyama H,Uchida Y. Nuclear BAG-1 expression reflects malignant po-tential in colorectal carcinomas. Br J Cancer 2002; 87: 1136–1139.
25. Paraskeva C, Finerty S, Mountford RA, Powell SC. Specific cy-togenetic abnormalities in two new human colorectal adenoma-derived epithelial cell lines. Cancer Res 1989; 49: 1282–1286.
26. Williams AC, Browne SJ, Yeudal WA, et al. Molecular eventsincluding p53 and k-ras alterations in the in vitro progressionof a human colorectal adenoma cell line to an adenocarcinoma.Oncogene 1993; 8: 3063–3072.
27. Henderson BR, Eleftheriou A. A comparison of the activity, se-quence specificity, and CRM1-dependence of different nuclearexport signals. Exp Cell Res 2000; 256: 213–224.
28. Bracey TS, Miller JC, Preece A, Paraskeva C. Gamma-radiation-induced apoptosis in human colorectal adenoma andcarcinoma cell lines can occur in the absence of wild type p53.Oncogene 1995; 12; 2391–2396.
29. Diaz GD, Paraskeva C, Thomas MG, Binderup L, Hague A.Apoptosis is induced by the active metabolite of vitamin D3
310 Apoptosis · Vol 10 · No 2 · 2005

Nuclear BAG-1 inhibits apoptosis in colonic adenoma cells
and its analogue EB1089 in colorectal adenoma and carcinomacells: possible implications for prevention and therapy. CancerRes 2000; 60: 2304–2312.
30. Guzey M, Takayama S, Reed JC. BAG1L enhances trans-activation function of the vitamin D receptor. J Biol Chem 2000;275: 40749–40756.
31. Witcher M, Yang X, Pater A, Tang SC. BAG-1 p50 isoforminteracts with the vitamin D receptor and its cellular overex-pression inhibits the vitamin D pathway. Exp Cell Res 2001;265(1): 167–173.
32. Cutress RI, Townsend PA, Brimmell M, Bateman AC, HagueA, Packham G. BAG-1 expression and function in human can-cer. Br J Cancer 2002; 87: 834–839.
33. Cato AC, Mink S. BAG-1 family of cochaperones in the mod-ulation of nuclear receptor action. J Steroid Biochem Mol Biol2001; 78, 379–388.
34. Kullmann M, Schneikert J, Moll J, et al. RAP46 is a nega-tive regulator of glucocorticoid receptor action and hormone-induced apoptosis. J Biol Chem 1998; 273: 14620–14625.
35. Froesch BA, Takayama S, Reed JC. BAG-1L protein enhancesandrogen receptor function. J Biol Chem 1998; 273: 11660–11666.
36. Niyaz Y, Zeiner M, Gehring U. Transcriptional activation bythe human Hsp70-associating protein Hap50. J Cell Sci 2001;114: 1839–1845.
37. Sondermann H, Ho AK, Listenberger LL, et al. Prediction ofnovel BAG-1 homologs based on structure/function analysisidentifies Snl1p as an Hsp70 co-chaperone in Saccharomycescerevisiae. J Biol Chem 2002; 277, 33220–33227.
38. Briknarova K, Takayama S, Brive L, et al. Structural analysis ofBAG1 cochaperone and its interactions with Hsc70 heat shockprotein. Nat Struct Biol 2001; 4: 349–352.
Apoptosis · Vol 10 · No 2 · 2005 311




















