
Arabidopsis Immunophilin-like TWD1 Functionally Interacts with Vacuolar ABC Transporters

Biochem. J. (2014) 460, 223–235 (Printed in Great Britain) doi:10.1042/BJ20140010 223
Novel role of a family of major facilitator transporters in biofilmdevelopment and virulence of Candida albicansAbdul Haseeb SHAH*, Ashutosh SINGH*, Sanjiveeni DHAMGAYE*, Neeraj CHAUHAN†, Patrick VANDEPUTTE‡1,Korivi Jyothiraj SUNEETHA§, Rupinder KAUR§, Pranab K. MUKHERJEE‖, Jyotsna CHANDRA‖, Mahmoud A. GHANNOUM‖,Dominique SANGLARD‡, Shyamal K. GOSWAMI* and Rajendra PRASAD*2
*Membrane Biology Laboratory, School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, India†Public Health Research Institute and Department of Microbiology and Molecular Genetics, New Jersey Medical School, Rutgers, The State University of New Jersey, 225 Warren Street,Newark, NJ 07103, U.S.A.‡Institut de Microbiologie, University of Lausanne and University Hospital Center, Lausanne, Switzerland§Laboratory of Fungal Pathogenesis, Centre for DNA Fingerprinting and Diagnostics, Hyderabad, Andhra Pradesh, India‖Center for Medical Mycology, Department of Dermatology, Case Western Reserve University, Cleveland, OH 44106, U.S.A.
The QDR (quinidine drug resistance) family of genes encodestransporters belonging to the MFS (major facilitator superfamily)of proteins. We show that QDR transporters, which are localized tothe plasma membrane, do not play a role in drug transport. Hence,null mutants of QDR1, QDR2 and QDR3 display no alterationsin susceptibility to azoles, polyenes, echinocandins, polyaminesor quinolines, or to cell wall inhibitors and many other stresses.However, the deletion of QDR genes, individually or collectively,led to defects in biofilm architecture and thickness. Interestingly,QDR-lacking strains also displayed attenuated virulence, butthe strongest effect was observed with qdr2Δ, qdr3Δ and inqdr1/2/3Δ strains. Notably, the attenuated virulence and biofilmdefects could be reversed upon reintegration of QDR genes.
Transcripts profiling confirmed differential expression of manybiofilm and virulence-related genes in the deletion strains ascompared with wild-type Candida albicans cells. Furthermore,lipidomic analysis of QDR-deletion mutants suggests massiveremodelling of lipids, which may affect cell signalling, leading tothe defect in biofilm development and attenuation of virulence.In summary, the results of the present study show that QDRparalogues encoding MFS antiporters do not display conservedfunctional linkage as drug transporters and perform functions thatsignificantly affect the virulence of C. albicans.
Key words: Candida albicans, lipid homoeostasis, quinidine drugresistance (QDR) transporters, virulence.
INTRODUCTION
Increased incidences of drug-resistant fungal pathogens havestimulated the identification of novel targets for combating fungalinfections in addition to the development of broad-specificityantifungal drugs. Development of antifungal therapy has been achallenge due to the limited availability of novel targets, owing tothe similarity of the metabolic pathways in pathogenic fungi andtheir higher eukaryote hosts, such as humans. Candida albicans,which represents one of the most common species of pathogenicyeasts, affects immunocompromised patients and frequentlyshows decreased susceptibility to antifungals resulting in MDR(multidrug resistance) [1,2]. Although there are continuous effortstowards development of new antifungals with novel targets, thepersistence of MDR remains a serious clinical issue. Variousmechanisms that contribute to the development of azole resistancehave been reported. These include point mutations in ERG11(ergosterol biosynthesis 11) or overexpression of its gene product,the target enzyme of azoles (i.e. lanosterol 14α-demethylase)[3,4], overexpression of the drug efflux pump-encoding genes
CDR1 (Candida drug resistance 1), CDR2 and MDR1, whichbelong to the ABC (ATP-binding cassette) and MFS (majorfacilitator superfamily) transporters [5,6]. The use of morerecent antifungals such as echinocandins has also resulted inthe emergence of resistance, although with still low incidence[7]. Fungal biofilms, which are three-dimensional complexstructures formed by sessile fungal cells in the presence of ECM(extracellular matrix), is a predominant morphological form ofCandida responsible for enhanced drug resistance [8].
Among all the different strategies adopted by C. albicansto acquire MDR, drug transporters play important roles in thedevelopment of MDR in sessile as well as in planktonic cells[8,9]. The major transporters responsible for MDR in C. albicansof the ABC proteins are Cdr1p and Cdr2p and of the MFS proteinsis Mdr1p [6,10,11]. High level of expression of any of these genesalone or in combination contributes to increased efflux of azolesand to resistance [12].
Computational analysis of the C. albicans genome revealed 95putative MFS proteins [13]. The Saccharomyces cerevisiae DHA1(drug/H+ antiporter 1) and DHA2 subfamily of MFS transporters
Abbreviations: ABC, ATP-binding cassette; BBN, barban; CDR, Candida drug resistance; CFU, colony-forming unit; CRD, congo red; CSLM,confocal scanning laser microscopy; Cy3, indocarbocyanine; Cy5, indodicarbocyanine; DHA, drug/H+ antiporter; DRM, detergent-resistant microdomain;FAR, farnesol; HBSS, Hank’s buffered salt solution; IPC, inositolphosphorylceramide; MDR, multidrug resistance; MFS, major facilitator superfamily;MG, Malachite Green; MIPC, mannosylinositolphosphorylceramide; NR, nile red; Pdr, pleiotropic drug resistance; PG, phosphatidylglycerol; PGL,phosphoglyceride; PI, phosphatidylinositol; PM, plasma membrane; Pma1, PM ATPase1; PS, phosphatidylserine; QDR, quinidine drug resistance; QDS,quinidine sulfate salt; QSM, quorum sensor molecule; R6G, rhodamine 6G; RT-PCR, reverse transcription–PCR; SE, sterol ester; SL, sphingolipid; WT,wild-type; XTT, 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide; YNB, yeast nitrogen base.
1 Present address: Groupe d’Etude des Interactions Hote-Pathogene, UPRES-EA 3142, Universite d’Angers, Angers, France.2 To whom correspondence should be addressed (email [email protected]).Complete transcriptome data for the QDR genes can be accessed from GEO dataset under accession number GSE41535.
c© The Authors Journal compilation c© 2014 Biochemical Society
Bio
chem
ical
Jo
urn
al
ww
w.b
ioch
emj.o
rg

224 A.H. Shah and others
are responsible for drug resistance and responses to chemicalstress. The S. cerevisiae DHA1 family members QDR1 (quinidineresistance 1), QDR2 and QDR3 were shown to be responsible forresistance against the drug quinidine, the herbicide BBN (barban),anti-cancerous compounds and several commonly used azoleantifungals [14–16]. These drug transporters, although related instructure and function, show specificity towards their functionalproperties [17–19]. Among Candida species, the only QDRreported for drug resistance is QDR2 of Candida glabrata, whichis implicated in resistance against imidazoles and is governed bythe transcription factor Pdr1 (pleiotropic drug resistance 1) [20].
Recently, QDR1, a putative quinidine drug resistance gene, wasidentified in one of our genome expression profiling studies as ahighly up-regulated gene in C. albicans cells treated with MG(Malachite Green) [21]. We began to explore the possible role ofthe QDR1, along with its two paralogues QDR2 and QDR3, inovercoming MG-induced stress and to address their potential rolein resistance against various other antifungal compounds or stressconditions. In the present study, we demonstrate that the presenceof QDR genes is not associated with resistance against MG orany other tested compounds including widely used antifungalagents. Genome expression, lipidomic and phenotypic profilingalong with virulence assays suggest that QDR genes are possiblefactors in C. albicans pathogenesis.
MATERIAL AND METHODS
Materials
Drugs and chemicals including MG, R6G (rhodamine 6G),NR (nile red), AMB (amphotericin B), TM (tunicamycin),NYS (nystatin), KTC (ketoconazole), VOR (voriconazole),MIC (miconazole), CTR (clotrimazole), CFW (calcoflourwhite),R123 (rhodamine 123), BBN (barban), CRD (congo red),SDS, QDS (quinidine sulfate salt), CSF (caspofungin), BER(berberine), ITC (itraconazole), NaAsO2, LiCl and CsCl, werefrom Sigma. Fluconazole was generously provided by Ranbaxy.NaCl, CuSO4·5H2O, MnCl2, CdCl2 and CoCl2·6H20 wereobtained from Qualigens. Ascorbic acid was purchased fromSRL. The oligonucleotides used in the present study, as listedin Supplementary Table S1 (http://www.biochemj.org/bj/460/bj4600223add.htm), were from Sigma Genosys. Anti-GFPmonoclonal antibody and anti-mouse secondary antibody werepurchased from Santa Cruz Biotechnology. 3H-radiolabelled FAR(farnesol) was from American Radiolabeled Chemicals. Cy3(indocarbocyanine)- and Cy5 (indodicarbocyanine)-labelled UTPwas obtained from GE Healthcare, and the SuperScript II reversetranscriptase enzyme, the oligo(dT)18 primer and the randomprimer were obtained from Invitrogen. dATP, dGTP, dTTP anddCTP were bought from Sigma.
Growth media and strains
Media chemicals were obtained either from Difco or HiMedia.Plasmids were maintained in Escherichia coli DH5α cells.E. coli cells was cultured in LB medium (Difco) to whichampicillin was added (0.1 mg/ml). WT (wild-type) and QDR-deletion mutants were cultured in YNB (yeast nitrogenbase) minimal medium broth or on YNB with 2 % agarplates. For selection of transformants in QDR gene deletions,nourseothricin (clonNAT; Werner BioAgent) was used inYPD at a concentration of 200 μg/ml. The strains used inthe present study are shown in Supplementary Table S2(http://www.biochemj.org/bj/460/bj4600223add.htm).
Drug susceptibility tests
The susceptibility of mutants to various drugs was tested bythe broth microdilution method according to CLSI (Clinicaland Laboratory Standards Institute) and serial dilution assays asdescribed previously [22,23].
Transport assays
NR accumulation by the WT and mutants was measured usinga FACSort flow cytometer (Becton Dickson ImmunocytometrySystems). Data were analysed using the CellQuest software(Becton Dickinson Immunocytometry Systems). NR was usedat a final concentration of 7 mM as described previously [24].R6G efflux was determined essentially as described previously[25]. FAR accumulation was performed using [3H]FAR(trans,trans-FAR [1-3H]; specific activity 50–60 Ci/mmol).Exponential phase cells (∼1.5 D) were washed twice with PBS.Cells were resuspended as a 5 % cell suspension in PBS andFAR was added to a final concentration of 20 nM. The cellsuspension containing FAR was incubated at 30 ◦C for 40 minwith continuous shaking. Then, 100 μl of this cell suspensionwas filtered rapidly and washed twice with ice-cold PBS on aMillipore manifold filter assembly using a 0.45-μm pore sizecellulose nitrate membrane filter. The filter discs were dried andput in cocktail ‘O’ to enhance the counts and the radioactivitywas measured in a liquid scintillation counter (Tri-Carb 2900TRLiquid Scintillation Analyzer; Packard).
QDR plasmid constructions
In order to delete QDR genes in C. albicans, the SAT1-flipper system was used. QDR1 deletion was achieved by PCRamplification of the 5′-UTR with primers QDR1-Kpn and QDR1-Xho with SC5314 genomic DNA as the template and insertionof the obtained fragment into XhoI-KpnI digested pSFS2 [26] toyield pVP62. Next, the 3′-UTR was obtained by PCR with primersQDR1-SacII and QDR1-SacI and insertion of the obtainedfragment in SacI-SacII-digested pVP62 to yield pVP63. pVP63was finally digested with KpnI-SacI to release the disruptioncassette for QDR1 inactivation. To re-introduce QDR1 in thedeletion mutant, the entire QDR1 gene was amplified with primersQDR1-Xhorev and QDR1-KpnI and introduced into pVP63digested with XhoI-KpnI, thus replacing the 5′-UTR. The plasmidpVP73 was obtained and digested with KpnI-SacI to release thereversion cassette.
For QDR2 and QDR3 deletions, 5′- and 3′-UTR regionswere cloned sequentially as in the case of QDR1 deletion. Re-introduction of QDR2 and QDR3 was also performed similarlyas done for QDR1.
Strain constructions
The C. albicans isolate SC5314 was used for the strainconstructions. Deletion of QDR1, QDR2 and QDR3 were achievedby transformation of C. albicans with KpnI-SacI linear fragmentsfrom the plasmids pVP63, pVP75 and pVP107 respectively.Transformations of C. albicans were carried out as describedin [27]. After isolation of nourseothricin-resistant isolates onYPD plates with 200 μg/ml nourseothricin, the SAT1 markerwas excised as described in [28]. Nourseothricin-susceptibleisolates were used to inactivate the second alleles of each gene.Re-introduction of intact genes in single or multiple deletion
c© The Authors Journal compilation c© 2014 Biochemical Society

Role of QDR transporters in Candida albicans 225
backgrounds were performed at respective genomic loci and wereverified by Southern blots (results not shown).
The C. albicans QDR1, QDR2 and QDR3 genes were clonedin pABC3-GFP vector (obtained from Professor Richard Cannon,University of Otago, Dunedin, New Zealand) and integrated intothe PDR5 locus of S. cerevisiae strain AD(1-8)u− in which sevenmajor transporters have been deleted [29]. The genes were clonedinto the vector utilizing its PacI, NotI or HindIII multiple cloningsites as described previously [30]. After cloning into pABC3-GFP, the cassette containing the required genes, was digested byAscI and integrated into AD(1-8)u− using the lithium acetatetransformation protocol [25] and plated on to SDu− (syntheticdefined media lacking urea) agar plates. The colonies obtainedafter incubating the plates at 30 ◦C were confirmed by Southernhybridization. The overexpressing strains were named ADQDR1-GFP, ADQDR2-GFP and ADQDR3-GFP and contained QDR1,QDR2 and QDR3 respectively.
Raft isolation and immunodetection of QDR proteins
Rafts were prepared as described previously [31]. PMs (plasmamembranes) and rafts was prepared from QDR-overexpressingstrains and immunodetected as described previously [25]. Pma1p(PM ATPase1) was used as loading control and was probed usinganti-Pma1p polyclonal antibody.
RNA isolation and cDNA synthesis
WT and QDR mutant strains were cultured in YNB dextrosemedia up to the mid-exponential phase of growth at 30 ◦C. Cellswere harvested and centrifuged at 3220 g for 5 min at 4 ◦C and theresulting cell pellet was washed three times with PBS. The finalcell pellet was used as a starting material for RNA preparationas well as for other experiments. RNA isolation was carried outby using the Qiagen RNeasy® mini kit as per the manufacturer’sspecifications, except that 300 μl of acid-washed 0.4–0.6 mmglass beads (Sigma) were used during cell lysis. RNA waseluted using DEPC milliQ water. RNA obtained was quantitatedspectrophotometrically and separated by electrophoresis in adenaturing formaldehyde agarose gel. A 10 μg aliquot of purifiedtotal RNA was used to synthesize cDNA using a protocoldescribed previously (http://www.transcriptome.ens.fr/sgdb/protocols/labelling/yeast.php). We used a direct-labelling methodto label cDNAs with 1 μM Cy3 and Cy5 dyes (AmershamPharmacia). Labelled cDNA was purified using Qiaquick PCRpurification columns with a final elution volume of 30 μl ofDNase/RNase-free water. Labelled cDNA was mixed withhybridization buffer (5 × SSC, 50% formaldehyde and 0.2%SDS pre-heated to 70 ◦C) at a 1:1 ratio and applied immediatelyon to a microarray slide. Slides were covered with 22 × 60 mmcoverslips, sealed in Corning hybridization chambers and allowedto incubate for 12 h at 42 ◦C. Slides were washed and scannedusing Scan Array Express microarray scanner from PerkinElmer.Each experimental condition was independently repeated threetimes including a dye swap (WT against qdr1Δ, WT againstqdr2Δ, WT against qdr3Δ and WT against qdr1/2/3Δ).
Scanning and data analysis
Raw data obtained from the scanner were quantified using theadaptive circle method and normalized using LOWESS (locallyweighted scatterplot smoothing). Background correction wasundertaken and the ratio between the WT and qdrΔ strainswas converted into Log2 ratios and was averaged for the three
replicates. The annotations used for the ORFs were obtained fromhttp://candida.bri.nrc.ca/L2L/. The threshold value was set to 2-fold to filter out only significantly affected genes. GO clusterswere found using the GO term finder or GO slim mapper optionat http://www.candidagenome.org [32]. The microarrays used inthe present study along with complete transcriptome data can beaccessed from GEO dataset under accession number GSE41535.
RT-PCR
RT-PCR (reverse transcription–PCR) was done using theRevertAidTM H Minus kit (MBI; Fermentas). Briefly, 1 μg ofisolated RNA was treated with DNase and thereafter primed witholigo(dT)18 and random primers for cDNA synthesis at 42 ◦Cfor 60 min. The reverse transcription reaction was terminatedby heating at 70 ◦C for 5 min. The synthesized cDNA product(1 μl) was directly used for PCR amplification (50 μl) using gene-specific forward and reverse primers (Supplementary Table S1).The amplified products were gel electrophoresed and quantitatedrelative to their expression in control strains.
Morphogenetic studies and biofilm formation
Hyphae were induced on solid and liquid media as describedpreviously [33]. Briefly, hyphae were induced in WT andmutants strains on solid spider medium and on liquid YPDbroth containing 10% FBS or on liquid N-acetyl glucosaminemedium. The exponential phase cells were starved and therequired number of cells was applied in liquid broth or solidagar plates and incubated at 37 ◦C as described previously [33].The ability of Candida isolates to form biofilms was evaluatedby metabolic activity, dry mass assay and confocal microscopy,as described previously [34]. Briefly, Candida cells were allowedto form biofilms for 48 h on silicone elastomer catheter discs.The amount of biofilm formed was assayed colorimetricallyusing the XTT [2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide; Sigma-Aldrich] metabolic activityassay in which XTT is converted by metabolically active cellsinto a red formazan product [34]. In addition, the effect of fungalsupernatants on the morphology and architecture of the formedbiofilms was evaluated using CSLM (confocal scanning lasermicroscopy) [34].
Murine model of hematogenously disseminated candidiasis
C. albicans strains SC5314, qdr1Δ, qdr2Δ, qdr3Δ andqdr1/2/3Δ, and corresponding revertant strains were grown inYPD broth at 30 ◦C to stationary phase. Cells were harvestedby centrifugation (3220 g for 5 min at 4◦C), washed twice inPBS and suspended to a density of 1 × 107 CFU (colony-formingunits)/ml before use. For animal experiments, we followedpreviously published methods [35]. Groups of ten female BALB/cmice (18–20 g each; Charles River Laboratories) were injectedintravenously via the lateral tail vein with 0.1 ml (1 × 106 CFU)of each strain. All mice were observed twice daily forsigns of morbidity and, if moribund, animals were killed by CO2
inhalation. Concomitantly, each C. albicans strain was used toinoculate five additional mice. Mice from each group were killedby CO2 inhalation at 24 and 48 h post-infection, and the kidneysfrom mice infected with each strain were removed, weighed andhomogenized in 5 ml of PBS. Homogenates were diluted in PBS,and aliquots were plated on to YPD agar supplemented with50 μg of streptomycin per ml to prevent bacterial growth. Plateswere incubated at 30 ◦C for 48 h, and the numbers of CFU/g of
c© The Authors Journal compilation c© 2014 Biochemical Society

226 A.H. Shah and others
tissue were then quantitated. The animal studies were conductedat the Public Health Research Institute, Rutgers University, NewJersey, U.S.A. All animals were housed and handled accordingto the guidelines set forth by the Institutional Animal Care andUse Committee (IACUC). Animal protocols were reviewed andapproved by the IACUC of Rutgers University, New Jersey,U.S.A. The animal experiments were conducted in facilitiesaccredited by the Association for Assessment and Accreditation ofLaboratory Animal Care (AAALAC) and were in full compliancewith the United States Animal Welfare Act (Public Law 98-198).
Adherence assay
WT and mutants Candida cells were grown overnight in YNBmedium. Cells were washed with HBSS (Hank’s buffered saltsolution) and diluted to 250 cells/ml of buffer. Of this cell dilution,1.5 ml was applied to six-well tissue culture plates and incubatedat 37 ◦C for 30 min. The medium was aspirated from wells andthe cells were then washed with HBSS. Each well was overlaidwith 3 ml of sabouraud dextrose agar. Plates were incubated at37 ◦C for 18–24 h and CFUs were counted for each strain.
Lipidomic analysis
Lipids were extracted from C. albicans cells using a slightmodification of the method of Bligh and Dyer as describedpreviously [36,37]. The methodology of MS-based lipidomicsemployed in the present study is consistent with our earlier work.PGLs (phosphoglycerides), SLs (sphingolipids) and SEs (sterolesters) were quantified exactly as described previously [36,38].
Statistical analysis
In order to determine the statistical significance of the differencesin virulence and yeast tissue counts, Student’s t tests wereperformed. P < 0.05 was considered statistically significant.
RESULTS
We demonstrated previously that WT C. albicans cells weresusceptible to a commonly used antifungal dye in the fishindustry, MG, and that its effect was independent of knownantifungal targets. Transcriptional profiling in response toMG treatment revealed that out of 204 responsive genes,QDR1 was among the most prominently induced (3.5-fold)genes [21]. According to data from the Candida GenomeDatabase (http://www.candidagenome.org), the C. albicansgenome has three QDR genes annotated as QDR1 (orf19.508),QDR2 (orf19.6992) and QDR3 (orf19.136), that show greaterthan 60% similarity to their closely related counterparttransporters from S. cerevisiae. Alignment of C. albicansQDR transporters with homologous MFS transporters fromother fungi show that QDR transporters contain the typicalantiporter motif (Figure 1A, and Supplementary Figure S1at http://www.biochemj.org/bj/460/bj4600223add.htm) and areclosely related to them based on the phylogenetic analysis(Figure 1B). The protein sequence alignment of QDR genes withclosely related MFS transporters showed few additional conservedresidues (boxed) preceding the antiporter motif whose functionalrelevance remains unexplored (Supplementary Figure S1). Thepresent study explores the role of all three members of the QDRfamily of genes in C. albicans.
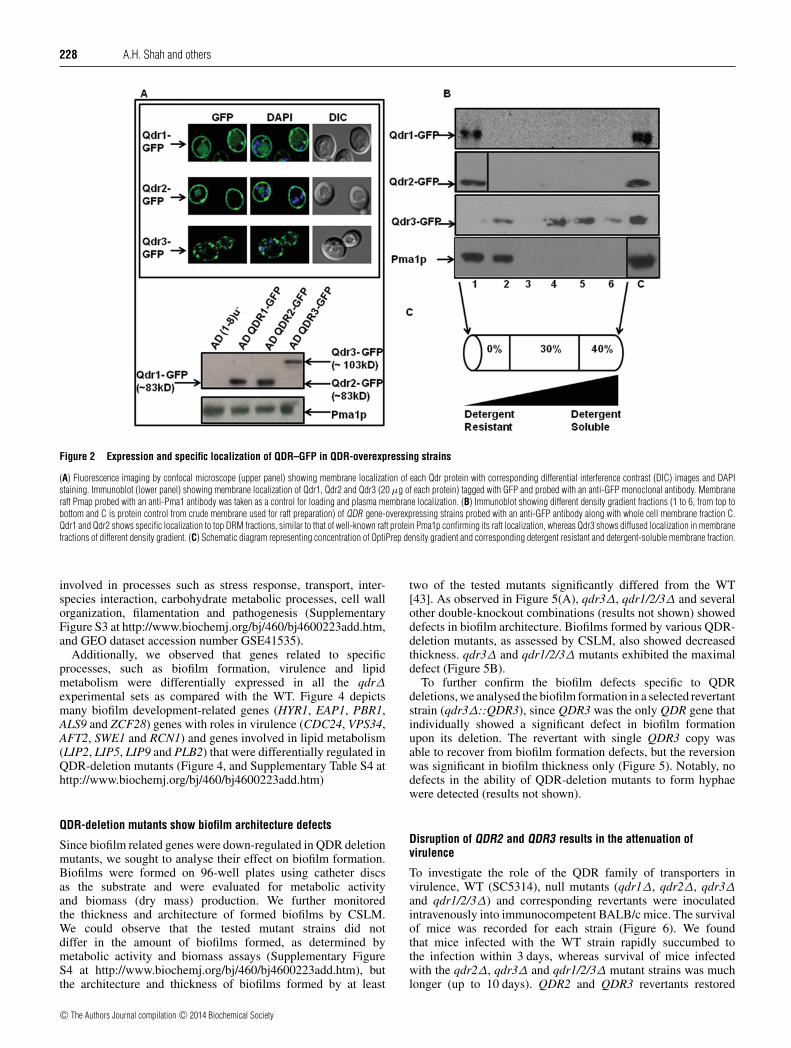
All QDR proteins, except Qdr3, are selectively localized withinmembrane rafts
We cloned QDR genes as GFP-tagged proteins stablyoverexpressed from a genomic PDR5 locus in a S. cerevisiaeAD(1-8)u− host enabling high expression levels [29]. A singlecopy integration of each transformant at the PDR5 locus wasconfirmed by Southern hybridization (results not shown). Theexpression and localization of QDR proteins was confirmedby monitoring GFP fluorescence using a confocal microscopeand was further confirmed by Western blot analysis using anti-GFP monoclonal antibodies. All of the three QDR proteinswere membrane localized as observed from localization of theirrespective GFP-tagged proteins or by the immunoblotting of thecorresponding membrane fractions using an anti-GFP monoclonalantibody (Figure 2A). Notably, the confocal images revealedsome trapped intracellular fluorescence due to an overexpressionof these proteins. The exact intracellular localization of trappedQDR proteins is not very apparent from these confocal images;however, DAPI fluorescence (blue) of cellular nuclei when mergedwith GFP fluorescence (green) ruled out any inclusion of QDRproteins within the nuclear region (Figure 2A).
Membrane rafts are DRMs (detergent-resistant microdomains)of the cell membrane that are enriched in SLs and ergosterol.These microdomains act as platforms to seed proteins that haveroles in cell signalling, virulence and protein sorting [39,40].We explored whether QDR proteins have any preference forsuch microdomains, also called ‘membrane rafts’. Our earlierstudies have shown that unlike ABC transporters of C. albicans,Mdr1, which belongs to the MFS family, is not localized withinthe microdomains of the PM [31]. Heterologously expressedGFP-tagged Qdr proteins were checked for their localizationin membrane rafts by Western blot analysis using monoclonalantibodies against GFP-tagged proteins. The results shown inFigure 2(B) show that Qdr1 and Qdr2 are preferentially localizedwithin DRMs of the PM. The distribution of Qdr1 and Qdr2 wassimilar to another well-known raft protein, Pma1p, which wasconfined to the DRM fraction. Qdr3, in contrast, appeared tobe a non-raft protein because it showed a uniform distributionthrough PM fractions (Figure 2B). Notably, in contrast withthe Qdr3 protein, the Qdr1 and Qdr2 proteins contain putativepalmitoylation sites commonly observed in raft proteins whichalso support their preferential localization to raft domains [41,42].
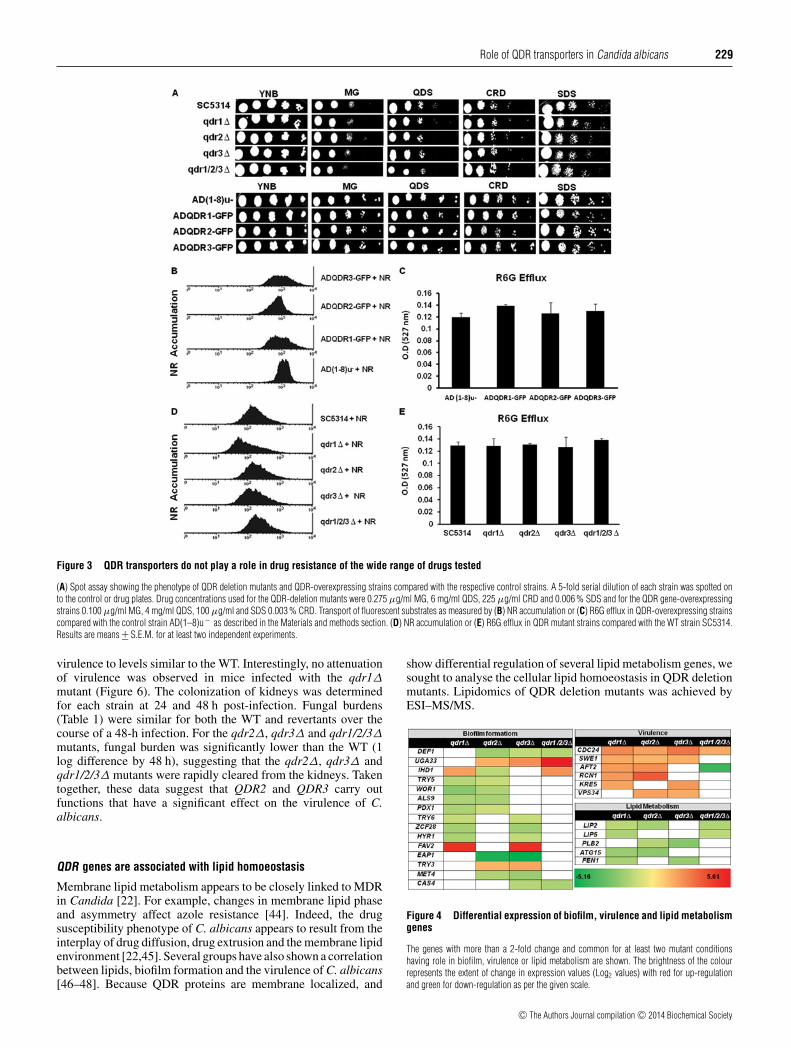
QDR transporters do not contribute to drug resistance
As noted earlier, the expression of QDR1 was highly up-regulated in the presence of MG. However, drug susceptibilityspot assays suggested that the levels of QDR1 had noeffect on the susceptibility to MG. Using the SAT1-flippercassette method homozygous nulls of all the three QDRs wereconstructed [21]. The deletion or overexpression of QDR1,QDR2 and QDR3 did not cause any change in susceptibilitiesto different drugs (Figure 3A, and Supplementary Table S3 athttp://www.biochemj.org/bj/460/bj4600223add.htm). To furtherexplore the role of these putative transporters, we performedextensive susceptibility tests with strains lacking individual QDRor their combination. From phenotypic studies, it was apparentthat qdr1�, qdr2�, qdr3� and qdr1/2/3� mutants did notexhibit altered susceptibilities to any of the tested compounds,including azoles, polyenes, echinocandins, polyamines, cellwall biosynthesis inhibitors etc. The mutants did not responddifferently than the WT to cellular stresses such as high salt,pH, oxidative, temperature and heavy metals (Figure 3A andSupplementary Table S3). Taken together, these data indicate that
c© The Authors Journal compilation c© 2014 Biochemical Society

Role of QDR transporters in Candida albicans 227
Figure 1 QDR transporters of C. albicans
(A) QDR transporters of C. albicans and other closely related transporters from S. cerevisiae showing the presence of an antiporter motif typical to DHA family of MFS transporters. (B) Phylogeneticrelationship of QDR transporters to yeast DHA1 family of MFS transporters of known function showing the C. albicans QDR transporters closest to QDR transporters from S. cerevisiae. Phylogramwas generated using Phylogeny.fr (http://www.phylogeny.fr/version2_cgi/index.cgi). Value at nodes represents bootstrap values signifying confidence levels.
QDR genes in contrast with their homologues in other yeastsappeared to have different roles in C. albicans.
QDR transporters lack transport activity to standard substrates
Several fluorescent compounds act as substrates of MFS or ABCtransporters and have been frequently used to measure transportkinetics. For example, the ABC and MFS transporters of C.albicans, Cdr1p and Mdr1p respectively, can efflux fluorescentcompounds such as R6G and NR [24,25]. We exploited thesesubstrates to explore whether QDR proteins could transport R6Gor NR. We observed that none of the overexpressing strains orQDR mutants differ in their ability to efflux NR or R6G, implyingthat these compounds are not the transport substrates of QDR(Figures 3B–3E). Accumulation of β-estradiol, which is also asubstrate of MDR transporters, was also not influenced by thepresence or absence of QDR proteins (results not shown). Thusthe lack of transport activity to these substrates corroborated withsusceptibility data. These results suggest that Candida QDR pro-teins, in contrast with their homologues in other yeast, do not ap-pear to be involved in the transport of a variety of tested substrates.
Transcripts profiling of QDR deletion mutants reveal differentialregulation of genes associated with biofilm formation, virulenceand lipids homoeostasis
To explore the possible role of QDR genes in cellular andmolecular processes, we carried out gene profiling analysis ofqdr1�, qdr2�, qdr3� and qdr1/2/3� mutants and comparedthem with their parental strain SC5314. The genes weredifferentially expressed more than 2-fold in the QDR-deletionmutants were assigned a GO-Slim term and were annotatedon the basis of their biological function [32]. For qdr1Δ, atotal of 169 genes showed significantly increased expressionas compared with the WT, whereas 231 genes were down-regulated. Similarly, for qdr2�, qdr3� and qdr1/2/3�, thenumber of genes showing significant increase in expressionwas 192, 419 and 230 respectively, whereas 366, 381 and 269genes were down-regulated compared with the WT. Few ofthe genes which were differentially regulated in two or moremutant strains were randomly selected and validated by RT-PCR(Supplementary Figure S2 at http://www.biochemj.org/bj/460/bj4600223add.htm). Some of the genes that generally showeddifferential expression among the QDR-deletion mutants were
c© The Authors Journal compilation c© 2014 Biochemical Society
228 A.H. Shah and others
Figure 2 Expression and specific localization of QDR–GFP in QDR-overexpressing strains
(A) Fluorescence imaging by confocal microscope (upper panel) showing membrane localization of each Qdr protein with corresponding differential interference contrast (DIC) images and DAPIstaining. Immunoblot (lower panel) showing membrane localization of Qdr1, Qdr2 and Qdr3 (20 μg of each protein) tagged with GFP and probed with an anti-GFP monoclonal antibody. Membraneraft Pmap probed with an anti-Pma1 antibody was taken as a control for loading and plasma membrane localization. (B) Immunoblot showing different density gradient fractions (1 to 6, from top tobottom and C is protein control from crude membrane used for raft preparation) of QDR gene-overexpressing strains probed with an anti-GFP antibody along with whole cell membrane fraction C.Qdr1 and Qdr2 shows specific localization to top DRM fractions, similar to that of well-known raft protein Pma1p confirming its raft localization, whereas Qdr3 shows diffused localization in membranefractions of different density gradient. (C) Schematic diagram representing concentration of OptiPrep density gradient and corresponding detergent resistant and detergent-soluble membrane fraction.
involved in processes such as stress response, transport, inter-species interaction, carbohydrate metabolic processes, cell wallorganization, filamentation and pathogenesis (SupplementaryFigure S3 at http://www.biochemj.org/bj/460/bj4600223add.htm,and GEO dataset accession number GSE41535).
Additionally, we observed that genes related to specificprocesses, such as biofilm formation, virulence and lipidmetabolism were differentially expressed in all the qdrΔexperimental sets as compared with the WT. Figure 4 depictsmany biofilm development-related genes (HYR1, EAP1, PBR1,ALS9 and ZCF28) genes with roles in virulence (CDC24, VPS34,AFT2, SWE1 and RCN1) and genes involved in lipid metabolism(LIP2, LIP5, LIP9 and PLB2) that were differentially regulated inQDR-deletion mutants (Figure 4, and Supplementary Table S4 athttp://www.biochemj.org/bj/460/bj4600223add.htm)
QDR-deletion mutants show biofilm architecture defects
Since biofilm related genes were down-regulated in QDR deletionmutants, we sought to analyse their effect on biofilm formation.Biofilms were formed on 96-well plates using catheter discsas the substrate and were evaluated for metabolic activityand biomass (dry mass) production. We further monitoredthe thickness and architecture of formed biofilms by CSLM.We could observe that the tested mutant strains did notdiffer in the amount of biofilms formed, as determined bymetabolic activity and biomass assays (Supplementary FigureS4 at http://www.biochemj.org/bj/460/bj4600223add.htm), butthe architecture and thickness of biofilms formed by at least
two of the tested mutants significantly differed from the WT[43]. As observed in Figure 5(A), qdr3Δ, qdr1/2/3Δ and severalother double-knockout combinations (results not shown) showeddefects in biofilm architecture. Biofilms formed by various QDR-deletion mutants, as assessed by CSLM, also showed decreasedthickness. qdr3Δ and qdr1/2/3Δ mutants exhibited the maximaldefect (Figure 5B).
To further confirm the biofilm defects specific to QDRdeletions, we analysed the biofilm formation in a selected revertantstrain (qdr3Δ::QDR3), since QDR3 was the only QDR gene thatindividually showed a significant defect in biofilm formationupon its deletion. The revertant with single QDR3 copy wasable to recover from biofilm formation defects, but the reversionwas significant in biofilm thickness only (Figure 5). Notably, nodefects in the ability of QDR-deletion mutants to form hyphaewere detected (results not shown).
Disruption of QDR2 and QDR3 results in the attenuation ofvirulence
To investigate the role of the QDR family of transporters invirulence, WT (SC5314), null mutants (qdr1Δ, qdr2Δ, qdr3Δand qdr1/2/3Δ) and corresponding revertants were inoculatedintravenously into immunocompetent BALB/c mice. The survivalof mice was recorded for each strain (Figure 6). We foundthat mice infected with the WT strain rapidly succumbed tothe infection within 3 days, whereas survival of mice infectedwith the qdr2Δ, qdr3Δ and qdr1/2/3Δ mutant strains was muchlonger (up to 10 days). QDR2 and QDR3 revertants restored
c© The Authors Journal compilation c© 2014 Biochemical Society
Role of QDR transporters in Candida albicans 229
Figure 3 QDR transporters do not play a role in drug resistance of the wide range of drugs tested
(A) Spot assay showing the phenotype of QDR deletion mutants and QDR-overexpressing strains compared with the respective control strains. A 5-fold serial dilution of each strain was spotted onto the control or drug plates. Drug concentrations used for the QDR-deletion mutants were 0.275 μg/ml MG, 6 mg/ml QDS, 225 μg/ml CRD and 0.006 % SDS and for the QDR gene-overexpressingstrains 0.100 μg/ml MG, 4 mg/ml QDS, 100 μg/ml and SDS 0.003 % CRD. Transport of fluorescent substrates as measured by (B) NR accumulation or (C) R6G efflux in QDR-overexpressing strainscompared with the control strain AD(1–8)u− as described in the Materials and methods section. (D) NR accumulation or (E) R6G efflux in QDR mutant strains compared with the WT strain SC5314.Results are means +− S.E.M. for at least two independent experiments.
virulence to levels similar to the WT. Interestingly, no attenuationof virulence was observed in mice infected with the qdr1Δmutant (Figure 6). The colonization of kidneys was determinedfor each strain at 24 and 48 h post-infection. Fungal burdens(Table 1) were similar for both the WT and revertants over thecourse of a 48-h infection. For the qdr2Δ, qdr3Δ and qdr1/2/3Δmutants, fungal burden was significantly lower than the WT (1log difference by 48 h), suggesting that the qdr2Δ, qdr3Δ andqdr1/2/3Δ mutants were rapidly cleared from the kidneys. Takentogether, these data suggest that QDR2 and QDR3 carry outfunctions that have a significant effect on the virulence of C.albicans.
QDR genes are associated with lipid homoeostasis
Membrane lipid metabolism appears to be closely linked to MDRin Candida [22]. For example, changes in membrane lipid phaseand asymmetry affect azole resistance [44]. Indeed, the drugsusceptibility phenotype of C. albicans appears to result from theinterplay of drug diffusion, drug extrusion and the membrane lipidenvironment [22,45]. Several groups have also shown a correlationbetween lipids, biofilm formation and the virulence of C. albicans[46–48]. Because QDR proteins are membrane localized, and
show differential regulation of several lipid metabolism genes, wesought to analyse the cellular lipid homoeostasis in QDR deletionmutants. Lipidomics of QDR deletion mutants was achieved byESI–MS/MS.
Figure 4 Differential expression of biofilm, virulence and lipid metabolismgenes
The genes with more than a 2-fold change and common for at least two mutant conditionshaving role in biofilm, virulence or lipid metabolism are shown. The brightness of the colourrepresents the extent of change in expression values (Log2 values) with red for up-regulationand green for down-regulation as per the given scale.
c© The Authors Journal compilation c© 2014 Biochemical Society

230 A.H. Shah and others
Figure 5 Architecture and thickness of biofilms formed by various QDR-deletion mutants
(A) Architecture of biofilms formed by each QDR-deletion mutant and revertant strain for qdr3�. qdr3Δ and qdr1/2/3Δ showed mixture of yeast and hyphal cells with less profuse matrix and thusshow architectural defects. Since only QDR3 deletion individually shows biofilm architecture defects in addition to qdr1/2/3Δ, we included only qdr3� revertant strain. (B) Biofilm thickness inthe corresponding strains also show significant decrease in qdr3� and qdr1/2/3Δ, which was reverted to some extent in the qdr3�-revertant strain. Results are means +− S.E.M. for at least twoindependent experiments.
Significant variation among several PGL classes
The lipidome analysis targeted nine major PGLs previouslyreported in C. albicans [36]. Although there was nosignificant change in overall PGL content, several PGL classes,including PI (phosphatidylinositol), PS (phosphatidylserine), PG(phosphatidylglycerol) and lysoPGLs were accumulated in QDR-deletion mutants. PI, which is an important precursor lipid forcomplex SLs, exhibited as much as 2-fold higher accumulationin QDR-deletion mutants compared with the WT, whereas the PS(an amino-PGL) level was increased only marginally by 1.2-foldin the QDR-deletion mutants. The content with regard to PG,a mitochondria-specific PGL, was 1.7-fold higher in the QDR-deletion mutants. The amount of lyso-lipids, such as lysoPC andlysoPE, were elevated in the QDR-deletion mutants by 2-
and 2.5-fold respectively (Figures 7A–7D). Surprisingly, the levelof PC and PE, two major PGLs in the PM, remained unalteredin the QDR-deletion mutants. Apparently, alteration in PGLs wasrestricted to only few select classes of lipids. Additionally, theobserved lipid changes were most prominent in the QDR2 andQDR3 mutants.
In our analyses, the nine determined PGLs could be furtherdifferentiated based on differences in their composition and thedegree of unsaturation of their fatty acids. Lipid profiling by ESI–MS/MS provided the ration of number of acyl carbons/number ofacyl carbon–carbon double bonds, which is referred to as the lipidspecies [36]. The data revealed that of the over 200 PGL speciesanalysed, almost 40 PGL species were significantly altered inat least two QDR-deletion mutants, qdr2Δ and qdr3Δ. However,
Figure 6 QDR2 and QDR3 mutants show attenuation in virulence
Survival curve of mice infected with QDR-deletion mutants show attenuation in virulence in qdr2�, qdr3� and qdr1/2/3Δ as the mice infected with mutant strains survived up to 10 dayspost-infection compared with the WT Candida strains where all the mice succumbed to infection within the first 3 days. Virulence was restored when the respective copies of three QDR genes werereverted back to mutants.
c© The Authors Journal compilation c© 2014 Biochemical Society

Role of QDR transporters in Candida albicans 231
Figure 7 Lipid composition of QDR-deletion mutants
Data are represented as nmol per mg of dry lipid mass (total normalized lipid signal), where a signal of 1 is equal to the signal obtained from 1 nmol of the corresponding internal standard. TheFigure includes only those lipid classes (inset) and species from (A–D) PLGs, (E) SLs and (F) SEs, where the difference in amounts was significant by P < 0.05 in at least two mutants, namelywhen lipid amounts in the WT were compared with the Δqdr1, Δqdr2 or �qdr3 mutant strains. The inset represents the total signal for each species which was calculated by adding the signals formolecular lipid species in that group. Error bars indicate +− S.D. (n�4 for four or more independent analyses).
only minimal changes in PGL species were observed in the qdr1Δmutant. It is evident from Figure 7 that compositional changesin lipid species of any PGL class are reflected in the overallalteration in the amounts of each. For example, high PG levelsresult from an accumulation of PG 32:1, 34:3, 34:2, 34:1, 36:3
Table 1 Kidney fungal burden of QDR-deletion mutant and revertant strains
C. albicans strains Log10 CFU/g of kidney tissue (mean +− SD) P value
24 h 48 hSC5314 6.23 +− 0.21 6.33 +− 0.17 0qdr1Δ 6.25 +− 0.37 6.4 +− 0.32 0.667qdr1Δ::QDR1 6.3 +− 0.14 6.12 +− 0.21 0.567qdr2Δ 5.3 +− 0.2 5.1 +− 0.27 0.011qdr2Δ::QDR2 6.0 +− 0.4 6.26 +− 0.47 0.394qdr3Δ 5.6 +− 0.26 5.5 +− 0.32 0.009qdr3Δ::QDR3 6.18 +− 0.31 6.0 +− 0.37 0.206qdr1/2/3Δ 5.21 +− 0.28 5.3 +− 0.19 0.004
and 36:1 species. Among other notable changes, we observedthat saturated PGL and odd chain fatty acyl-containing PGLswere accumulated in the QDR-deletion mutants (SupplementaryTable S5 at http://www.biochemj.org/bj/460/bj4600223add.htm).Overall, these data suggest that PGLs undergo major remodellingupon deletion of QDR2 and QDR3 from the membraneand the majority of these observed changes could bereversed in their revertant strains (Supplementary Table S6 athttp://www.biochemj.org/bj/460/bj4600223add.htm).
QDR affects membrane raft constituents
SLs are one of the major constituents of membrane microdomainsor ‘lipid rafts’ and are critical in cell signalling and biofilmformation and as virulence determinants in C. albicans[31,49]. In our analysis, we targeted three major complexSLs previously reported in C. albicans [36], and found thatthese SLs are accumulated (as much as 4.4-fold) in the QDR-deletion mutants (Figure 7E). This SL accumulation could be
c© The Authors Journal compilation c© 2014 Biochemical Society

232 A.H. Shah and others
attributed to the accumulation of subsequent SL intermediatemetabolites, specifically IPC (inositolphosphorylceramide) andMIPC (mannosylinositolphosphorylceramide) (Figure 7E). Ofnote, M(IP)2C (mannosyldiinositolphosphorylceramide), themost complex SL in Candida, was found in trace amounts in onlythe qdr2Δ mutants. The increased content of inositol-containingSLs also coincided with an increase in PI levels (Figure 7C).Concededly, accumulation of the SL molecular species, namelyIPC (44:0-3) and MIPC (44:0-3), was observed in the QDRdeletion mutants (Figure 7E).
Sterol, which is another major component of lipid raftintegrity, was depleted (as much as 1.7-fold) in the testedQDR-deletion mutants (Figure 7F). This depletion in sterolcontent could be attributed to decreased levels of sterol pathwayintermediates, namely lanosterol, zymosterol, episterol, fecosteroland ergosterol. Notably, the changes in these rafts associated withSLs were specific because they could be reversed in the QDR-revertant strains (Supplementary Table S6). However, reversal ofthe sterol levels was not observed in the QDR revertants.
QSM (quorum sensor molecule) levels are not affected by QDR
FAR, a QSM, is a precursor for the synthesis of sterolsin C. albicans, which blocks the morphological transitionand biofilm development in Candida [50,51]. Since QDR-deletion mutants showed changes in biosynthetic intermediatesof sterol biosynthesis, we speculated whether FAR is transportedby QDR proteins and its levels then may affect biofilmdevelopment. For this, we examined the accumulation of H3-radiolabelled FAR. As shown in Supplementary Figure S5A(at http://www.biochemj.org/bj/460/bj4600223add.htm), FARaccumulated in C. albicans. However, there was no majordifference in the accumulation of FAR in either QDR-deletionmutant or QDR-overexpressing strains (Supplementary FigureS5A and results not shown), thus suggesting that biofilm defectsin QDR mutants was not due to a role of QDRs in FAR transport.
In another set of related experiments, we addressed theexpression levels of QDR genes in response to intracellularFAR in mutants defective in FAR synthesis (dpp3Δ) [52]. Theexpression analysis revealed no significant expression differencesin any of the QDR in dpp3Δ strains, thus highlighting that the
FAR production and QDR gene expression are not associated(Supplementary Figure S5B).
DISCUSSION
Among the 95 MFS putative proteins that are present in theC. albicans genome, QDR1, QDR2 and QDR3 belong tothe DHA1 family of antiporters. QDR homologues are welldescribed in other yeast species where they are involved inresistance to quinidines, azoles, anti-cancerous compounds andherbicides [14–16]. Although CgQDR2 from C. glabrata confersPdr1-dependent resistance to quinidine and many imidazole drugs[20], the QDRs of S. cerevisiae seem to have varied functions,including maintenance of metal ion homoeostasis and responseto oxidative and polyamine stresses [17–19].
In the present study, we showed that although the threeQDR transporters (QDR1, QDR2 and QDR3) possess the typical‘antiporter motif’ (GX8GX3GPX2G2) of the MFS superfamily ofproteins, apparently they do not possess the expected transportfunction. This conclusion was drawn given that none of the testedsubstrates, including drugs, steroids, fluorescent dyes, glucoseand amino acids, were transported by any of the three QDRproteins. Furthermore, neither the null mutants for the QDRsnor heterologous host S. cerevisiae cells overexpressing theseproteins exhibited any effect on susceptibility to several drugs(including quinidine), MG, metal salts, pH or osmotic stress.Of note, although MG could induce QDR1 expression [21], itsoverexpression or deletion did not affect MG susceptibility.
Notably, however, the deletion of QDRs, either individuallyor collectively, led to defects in biofilm developmentand attenuation of virulence in a mouse infection model(Figure 8). The defects observed either in biofilm formationor in virulence could be reverted by the re-introduction ofWT genes. Additionally, adherence, which is an importantfactor contributing to virulence and colonization, remainedunaffected in QDR-deletion mutants (Supplementary TableS7A at http://www.biochemj.org/bj/460/bj4600223add.htm).Neither was the survival of different QDR-deletion mutantscompromised in the presence of mammalian THP1 macrophages(Supplementary Table S7B), thus excluding the contribution ofthese processes to the QDR-dependent attenuation of virulenceor biofilm development in Candida. To rule out the possibility
Figure 8 Illustration of the role of QDR transporters and probable path of action employed
QDR transporters localized in cell membrane may sense and/or transport extracellular signals and subsequently transfer the signals from extracellular environment to bring about the expression ofgene involved in maintaining virulence, biofilm formation, lipid homoeostasis etc. Knocking out of these genes results in changes in lipid homoeostasis and alteration in biofilm formation. Thesechanges lead to defects in formation of biofilm with decrease in virulence thus evoking combinatorial effects in lipid environment to cope with the observed changes.
c© The Authors Journal compilation c© 2014 Biochemical Society

Role of QDR transporters in Candida albicans 233
of whether polar effects of the gene deletion are contributing tophenotypes, we considered the genomic context of QDR genes.It was observed that QDR1 is immediately followed by the YDJ1gene (an uncharacterized putative co-chaperone), whereas GAP2(amino acid permease) is downstream of QDR2 and EXO84 genewhich encodes for a subunit of exocyst complex follows QDR3.However, the downstream genes are neither overlapping to QDRgene sequences nor are directly related to any of the phenotypesobserved in the QDR mutants. Moreover, our transcriptomic dataalso did not reveal any differential expression of these downstreamgenes. Thus the observed phenotypic defects in mutant strainsseem to be the direct effects of QDR gene deletion (SupplementaryFigure S6 at http://www.biochemj.org/bj/460/bj4600223add.htm)
Membrane transporters belonging to the MFS and ABCfamilies are present in large numbers in the genome and performa variety of physiological functions. The present study suggests anovel role for the QDR family of the MFS transporters in biofilmdevelopment and virulence of C. albicans. In this context, it isworth mentioning a recent study in C. albicans, which showedthat an MFS protein, GIT3 (transporter of glycerophosphodiestermetabolites), was required for full virulence in a mouse modelof disseminated candidiasis [53]. Another recent study, utilizingvirulence assays, demonstrated that the multidrug transporterabcB from Aspergillus fumigatus, belonging to the ABCsuperfamily of proteins, was a possible factor required for normalpathogenesis [54]. We have found previously that an ABC halftransporter orf19.3120 contributes to virulence given that a C.albicans lacking this transporter was attenuated in virulence ina mouse model of infection (N.K. Khandelwal and R. Prasad,unpublished work). Interestingly, orf19.3120 is similar to thehuman ABCG2 transporter, which is not only involved in MDR,but is also implicated in AML (acute myeloid leukaemia).Together, the data suggest that, in addition to their typical transportfunction, membrane transporters can also be important virulencedeterminants.
To examine the cause of the virulence attenuation andbiofilm defects, we performed transcripts profiling with differentmutant strains. Our transcriptome analysis revealed that severaldifferentially regulated genes were related to biofilm developmentand virulence (Figure 4). Our transcriptional profiling of QDR-deletion mutants also revealed differential regulation of lipidmetabolism genes, including those encoding for lipases (LIP2,LIP5 etc.), and phospholipases (PLB2 and SPO1) (Figure 4).This coincided well with the extensive lipidomic analysis of thepresent study, which highlighted a massive remodelling of lipidsin the QDR-deletion mutants with compositional alterations inacylglycerides. Lipidomic analysis has previously shown that thefluctuation in lipid composition affect biofilm and raft formationand the defects in the genes leading to the formation of lipidsaffects biofilm formation [49]. Decrease in sterol levels in biofilmsat different stages of growth has also been observed [55].Likewise, in Candida species, lipases and phospholipases areimplicated in host interaction, pathogenesis and virulence. Forinstance, lipase mutants of C. albicans and Candida parapsilosisdisplay attenuated virulence, decreased fungal burden and rapidclearance from different organs compared with the respectiveWTs [56,57]. Lipase secretion by Candida species is importantfor macrophage infection and dendritic cells also phagocytoseC. parapsilosis lipase mutants efficiently and generate potentresponses [58,59]. The phospholipases also affect virulenceby damaging the host membranes and phospholipase-encodinggenes such as PLB1 and PLB5, are implicated in virulenceand pathogenicity [60–63]. The present study found evidenceof down-regulation of phospholipase genes, accompanied withthe accumulation of lysoPGLs, which strongly suggests that
phospholipases, along with lipases, could affect QDR-dependentvirulence in C. albicans.
Of note, in several unrelated transcriptional studies, QDR1 hasbeen shown to be differentially regulated, but no specific role orfunction has been assigned to either of the QDR transportersin C. albicans. QDR1 expression was found to be higher inbiofilms compared with planktonic cells and in white cell forms ascompared with opaque cells [64,65]. The expression of QDR1 wasdown-regulated in a mutant lacking the biofilm regulator ZAP1(CSR1) [66]. Furthermore, QDR1 expression is differentiallyregulated under some stress conditions [67,68]. Collectively, thesedata thus suggest that QDRs are involved in several physiologicalresponses.
We were unable to ascertain the role of QDR proteins astransporters. We tested several compounds and metabolites thatare commonly transported by MFS proteins. However, none werefound to be substrates of the QDRs. Transport measurements mayrequire some additional conditions and testing other potentialsubstrates. Notably, changes in pH, salt and temperature stressesdid not reveal any measurable change in transport. In summary,the nature of the transported substrates/signal molecules remainselusive. However, the present study provides strong evidenceof the existence of a family of MFS transporters that play animportant role in the growth of Candida in the host.
AUTHOR CONTRIBUTION
Abdul Haseeb Shah, Ashutosh Singh, Sanjiveeni Dhamgaye, Neeraj Chauhan, PranabK. Mukherjee, Mahmoud A. Ghannoum, Dominique Sanglard and Rajendra Prasaddesigned the experiments; Abdul Haseeb Shah, Ashutosh Singh, Sanjiveeni Dhamgaye,Korivi Jyothiraj Suneetha, Patrick Vandeputte, Neeraj Chauhan, Jyotsna Chandra andPranab K. Mukherjee performed the experiments; Abdul Haseeb Shah, Ashutosh Singh,Sanjiveeni Dhamgaye, Neeraj Chauhan, Pranab K. Mukherjee, Rupinder Kaur andRajendra Prasad analysed the data with inputs from the other authors; Neeraj Chauhan,Pranab K. Mukherjee, Rupinder Kaur, Dominique Sanglard, Shyamal K. Goswami andRajendra Prasad contributed reagents/materials; and Abdul Haseeb Shah, Ashutosh Singh,Sanjiveeni Dhamgaye and Rajendra Prasad wrote the paper with input from the otherauthors.
ACKNOWLEDGEMENT
We thank Dr Ritu Pasrija for guidance with raft preparation.
FUNDING
The work was supported, in part, by the Indian Department of Biotechnology[grant numbers BT/01/CEIB/10/III/02, BT/PR 14879/BRB 10/885/2010 andBT/PR13641/Med/29/175/2010 (to R.P.)], the NIH (National Institutes of Health) [grantnumbers R21EY021303 and R21AI074077 (to P.K.M.) and RO1DE17846 (to M.A.G.)] andthe OHARA (Oral HIV AIDS Research Alliance) [grant number BRS-ACURE-S-11-000049-110229]. A.H.S. acknowledges CSIR (Council of Scientific and Industrial Research) Indiafor a senior research fellowship.
REFERENCES
1 Ostrosky-Zeichner, L., Casadevall, A., Galgiani, J. N., Odds, F. C. and Rex, J. H. (2010) Aninsight into the antifungal pipeline: selected new molecules and beyond. Nat. Rev. DrugDiscov. 9, 719–727 CrossRef PubMed
2 Odds, F. C., Brown, A. J. and Gow, N. A. (2003) Antifungal agents: mechanisms of action.Trends Microbiol. 11, 272–279 CrossRef PubMed
3 Morio, F., Loge, C., Besse, B., Hennequin, C. and Le Pape, P. (2010) Screening for aminoacid substitutions in the Candida albicans Erg11 protein of azole-susceptible andazole-resistant clinical isolates: new substitutions and a review of the literature. Diagn.Microbiol. Infect. Dis. 66, 373–384 CrossRef PubMed
c© The Authors Journal compilation c© 2014 Biochemical Society

234 A.H. Shah and others
4 Flowers, S. A., Barker, K. S., Berkow, E. L., Toner, G., Chadwick, S. G., Gygax, S. E.,Morschhauser, J. and Rogers, P. D. (2012) Gain-of-function mutations in UPC2 are afrequent cause of ERG11 upregulation in azole-resistant clinical isolates of Candidaalbicans. Eukaryot. Cell 11, 1289–1299 CrossRef PubMed
5 Cannon, R. D., Lamping, E., Holmes, A. R., Niimi, K., Baret, P. V., Keniya, M. V., Tanabe,K., Niimi, M., Goffeau, A. and Monk, B. C. (2009) Efflux-mediated antifungal drugresistance. Clin. Microbiol. Rev. 22, 291–321 CrossRef PubMed
6 Pasrija, R., Banerjee, D. and Prasad, R. (2007) Structure and function analysis ofCaMdr1p, a major facilitator superfamily antifungal efflux transporter protein of Candidaalbicans: identification of amino acid residues critical for drug/H+ transport. Eukaryot.Cell 6, 443–453 CrossRef PubMed
7 Walker, L. A., Gow, N. A. and Munro, C. A. (2010) Fungal echinocandin resistance.Fungal Genet. Biol. 47, 117–126 CrossRef PubMed
8 Mukherjee, P. K. and Chandra, J. (2004) Candida biofilm resistance. Drug Resist.Updates 7, 301–309 CrossRef PubMed
9 Ramage, G., Bachmann, S., Patterson, T. F., Wickes, B. L. and Lopez-Ribot, J. L. (2002)Investigation of multidrug efflux pumps in relation to fluconazole resistance in Candidaalbicans biofilms. J. Antimicrob. Chemother. 49, 973–980 CrossRef PubMed
10 Prasad, R., De Wergifosse, P., Goffeau, A. and Balzi, E. (1995) Molecular cloning andcharacterization of a novel gene of Candida albicans, CDR1, conferring multipleresistance to drugs and antifungals. Curr. Genet. 27, 320–329 CrossRef PubMed
11 Sanglard, D., Ischer, F., Monod, M. and Bille, J. (1997) Cloning of Candida albicansgenes conferring resistance to azole antifungal agents: characterization of CDR2, a newmultidrug ABC transporter gene. Microbiology 143, 405–416 CrossRef PubMed
12 Prasad, R. and Kapoor, K. (2005) Multidrug resistance in yeast Candida. Int. Rev. Cytol.242, 215–248 CrossRef PubMed
13 Gaur, M., Puri, N., Manoharlal, R., Rai, V., Mukhopadhayay, G., Choudhury, D. andPrasad, R. (2008) MFS transportome of the human pathogenic yeast Candida albicans.BMC Genomics 9, 579 CrossRef PubMed
14 Nunes, P. A., Tenreiro, S. and Sa-Correia, I. (2001) Resistance and adaptation to quinidinein Saccharomyces cerevisiae: role of QDR1 (YIL120w), encoding a plasma membranetransporter of the major facilitator superfamily required for multidrug resistance.Antimicrob. Agents Chemother. 45, 1528–1534 CrossRef PubMed
15 Vargas, R. C., Tenreiro, S., Teixeira, M. C., Fernandes, A. R. and Sa-Correia, I. (2004)Saccharomyces cerevisiae multidrug transporter Qdr2p (Yil121wp): localization andfunction as a quinidine resistance determinant. Antimicrob. Agents Chemother. 48,2531–2537 CrossRef PubMed
16 Tenreiro, S., Vargas, R. C., Teixeira, M. C., Magnani, C. and Sa-Correia, I. (2005) Theyeast multidrug transporter Qdr3 (Ybr043c): localization and role as a determinant ofresistance to quinidine, barban, cisplatin, and bleomycin. Biochem. Biophys. Res.Commun. 327, 952–959 CrossRef PubMed
17 Rıos, G., Cabedo, M., Rull, B., Yenush, L., Serrano, R. and Mulet, J. M. (2013) Role of theyeast multidrug transporter Qdr2 in cation homeostasis and the oxidative stress response.FEMS Yeast Res. 13, 97–106 CrossRef PubMed
18 Vargas, R. C., Garcıa-Salcedo, R., Tenreiro, S., Teixeira, M. C., Fernandes, A. R., Ramos,J. and Sa-Correia, I. (2007) Saccharomyces cerevisiae multidrug resistance transporterQdr2 is implicated in potassium uptake, providing a physiological advantage toquinidine-stressed cells. Eukaryot. Cell 6, 134–142 CrossRef PubMed
19 Teixeira, M. C., Cabrito, T. R., Hanif, Z. M., Vargas, R. C., Tenreiro, S. and Sa-Correia, I.(2011) Yeast response and tolerance to polyamine toxicity involving the drug:H+
antiporter Qdr3 and the transcription factors Yap1 and Gcn4. Microbiology 157,945–956 CrossRef PubMed
20 Costa, C., Pires, C., Cabrito, T. R., Renaudin, A., Ohno, M., Chibana, H., Sa-Correia, I.and Teixeira, M. C. (2013) Candida glabrata drug:H+ antiporter CgQdr2 confersimidazole drug resistance, being activated by transcription factor CgPdr1. Antimicrob.Agents Chemother. 57, 3159–3167 CrossRef PubMed
21 Dhamgaye, S., Devaux, F., Manoharlal, R., Vandeputte, P., Shah, A. H., Singh, A.,Blugeon, C., Sanglard, D. and Prasad, R. (2012) In vitro effect of malachite green onCandida albicans involves multiple pathways and transcriptional regulators UPC2 andSTP2. Antimicrob. Agents Chemother. 56, 495–506 CrossRef PubMed
22 Mukhopadhyay, K., Kohli, A. and Prasad, R. (2002) Drug susceptibilities of yeast cells areaffected by membrane lipid composition. Antimicrob. Agents Chemother. 46,3695–3705 CrossRef PubMed
23 Clinical and Laboratory Standards Institute (CLSI) (2008) Reference Method for BrothDilution Antifungal Susceptibility Testing of Yeasts, 3rd edition, Clinical and LaboratoryStandards Institute, Wayne
24 Ivnitski-Steele, I., Holmes, A. R., Lamping, E., Monk, B. C., Cannon, R. D. and Sklar, L. A.(2009) Identification of Nile red as a fluorescent substrate of the Candida albicansATP-binding cassette transporters Cdr1p and Cdr2p and the major facilitator superfamilytransporter Mdr1p. Anal. Biochem. 394, 87–91 CrossRef PubMed
25 Shukla, S., Saini, P., Smriti Jha, S., Ambudkar, S. V. and Prasad, R. (2003) Functionalcharacterization of Candida albicans ABC transporter Cdr1p. Eukaryot. Cell 2,1361–1375 CrossRef PubMed
26 Reuss, O., Vik, A., Kolter, R. and Morschhauser, J. (2004) The SAT1 flipper, anoptimized tool for gene disruption in Candida albicans. Gene 341,119–127 CrossRef PubMed
27 Coste, A. T., Karababa, M., Ischer, F., Bille, J. and Sanglard, D. (2004) TAC1,transcriptional activator of CDR genes, is a new transcription factor involved in theregulation of Candida albicans ABC transporters CDR1 and CDR2. Eukaryot. Cell 3,1639–1652 CrossRef PubMed
28 Eddouzi, J., Parker, J. E., Vale-Silva, L. A., Coste, A., Ischer, F., Kelly, S., Manai, M. andSanglard, D. (2013) Molecular mechanisms of drug resistance in clinical Candidaspecies isolated from Tunisian hospitals. Antimicrob. Agents Chemother. 57,3182–3193 CrossRef PubMed
29 Decottignies, A., Grant, A. M., Nichols, J. W., de Wet, H., McIntosh, D. B. and Goffeau, A.(1998) ATPase and multidrug transport activities of the overexpressed yeast ABC proteinYor1p. J. Biol. Chem. 273, 12612–12622 CrossRef PubMed
30 Lamping, E., Monk, B. C., Niimi, K., Holmes, A. R., Tsao, S., Tanabe, K., Niimi, M.,Uehara, Y. and Cannon, R. D. (2007) Characterization of three classes of membraneproteins involved in fungal azole resistance by functional hyperexpression inSaccharomyces cerevisiae. Eukaryot. Cell 6, 1150–1165CrossRef PubMed
31 Pasrija, R., Panwar, S. L. and Prasad, R. (2008) Multidrug transporters CaCdr1p andCaMdr1p of Candida albicans display different lipid specificities: both ergosterol andsphingolipids are essential for targeting of CaCdr1p to membrane rafts. Antimicrob.Agents Chemother. 52, 694–704 CrossRef PubMed
32 Boyle, E. I., Weng, S., Gollub, J., Jin, H., Botstein, D., Cherry, J. M. and Sherlock, G.(2004) GO::TermFinder-open source software for accessing Gene Ontology informationand finding significantly enriched Gene Ontology terms associated with a list of genes.Bioinformatics 20, 3710–3715 CrossRef PubMed
33 Prasad, T., Saini, P., Gaur, N. A., Vishwakarma, R. A., Khan, L. A., Haq, Q. M. and Prasad,R. (2005) Functional analysis of CaIPT1, a sphingolipid biosynthetic gene involved inmultidrug resistance and morphogenesis of Candida albicans. Antimicrob. AgentsChemother. 49, 3442–3452 CrossRef PubMed
34 Chandra, J., Mukherjee, P. K. and Ghannoum, M. A. (2008) In vitro growth and analysis ofCandida biofilms. Nat. Protoc. 3, 1909–1924 CrossRef PubMed
35 Desai, C., Mavrianos, J. and Chauhan, N. (2011) Candida albicans SRR1, a putativetwo-component response regulator gene, is required for stress adaptation,morphogenesis, and virulence. Eukaryot. Cell 10, 1370–1374CrossRef PubMed
36 Singh, A., Prasad, T., Kapoor, K., Mandal, A., Roth, M., Welti, R. and Prasad, R. (2010)Phospholipidome of Candida: each species of Candida has distinctive phospholipidmolecular species. OMICS 14, 665–677 CrossRef PubMed
37 Bligh, E. G. and Dyer, W. J. (1959) A rapid method of total lipid extraction andpurification. Can. J. Biochem. Physiol. 37, 911–917 CrossRef PubMed
38 Singh, A. and Prasad, R. (2011) Comparative lipidomics of azole sensitive and resistantclinical isolates of Candida albicans reveals unexpected diversity in molecular lipidimprints. PLoS ONE 6, e19266 CrossRef PubMed
39 Insenser, M., Nombela, C., Molero, G. and Gil, C. (2006) Proteomic analysis ofdetergent-resistant membranes from Candida albicans. Proteomics 6,S74–S81 CrossRef PubMed
40 Bagnat, M., Keranen, S., Shevchenko, A., Shevchenko, A. and Simons, K. (2000) Lipidrafts function in biosynthetic delivery of proteins to the cell surface in yeast. Proc. Natl.Acad. Sci. U.S.A. 97, 3254–3259 CrossRef PubMed
41 Levental, I., Lingwood, D., Grzybek, M., Coskun, U. and Simons, K. (2010) Palmitoylationregulates raft affinity for the majority of integral raft proteins. Proc. Natl. Acad. Sci. U.S.A.107, 22050–22054 CrossRef PubMed
42 Smotrys J, E. and Linder, M, E. (2004) Palmitoylation of intracellular signaling proteins:regulation and function. Annu. Rev. Biochem. 73, 559–587 CrossRef PubMed
43 Peyton, B. M. (1996) Effects of shear stress and substrate loading rate on Pseudomonasaeruginosa biofilm thickness and density. Water Res. 30, 29–36 CrossRef
44 Kohli, A., Smriti Mukhopadhyay, K., Rattan, A. and Prasad, R. (2002) In vitro low-levelresistance to azoles in Candida albicans is associated with changes in membrane lipidfluidity and asymmetry. Antimicrob. Agents Chemother. 46,1046–1052 CrossRef PubMed
45 Pasrija, R., Prasad, T. and Prasad, R. (2005) Membrane raft lipid constituents affect drugsusceptibilities of Candida albicans. Biochem. Soc. Trans. 33,1219–1223 CrossRef PubMed
46 Seneviratne, C. J., Jin, L. and Samaranayake, L. P. (2008) Biofilm lifestyle of Candida: amini review. Oral Dis. 14, 582–590 CrossRef PubMed
47 Chen, Y. L., Montedonico, A. E., Kauffman, S., Dunlap, J. R., Menn, F. M. and Reynolds,T. B. (2010) Phosphatidylserine synthase and phosphatidylserine decarboxylase areessential for cell wall integrity and virulence in Candida albicans. Mol. Microbiol. 75,1112–1132 CrossRef PubMed
c© The Authors Journal compilation c© 2014 Biochemical Society

Role of QDR transporters in Candida albicans 235
48 Noble, S. M., French, S., Kohn, L. A., Chen, V. and Johnson, A. D. (2010) Systematicscreens of a Candida albicans homozygous deletion library decouplemorphogenetic switching and pathogenicity. Nat. Genet. 42, 590–598CrossRef PubMed
49 Lattif, A. A., Mukherjee, P. K., Chandra, J., Roth, M. R., Welti, R., Rouabhia, M. andGhannoum, MA. (2011) Lipidomics of Candida albicans biofilms revealsphase-dependent production of phospholipid molecular classes and rolefor lipid rafts in biofilm formation. Microbiology 157, 3232–3242CrossRef PubMed
50 Nickerson, K. W., Atkin, A. L. and Hornby, J. M. (2006) Quorum sensing in dimorphicfungi: farnesol and beyond. Appl. Environ. Microbiol. 72, 3805–3813CrossRef PubMed
51 Han, T. L, Cannon, R. D. and Villas-Boas, S. G. (2011) The metabolic basis of Candidaalbicans morphogenesis and quorum sensing. Fungal Genet. Biol. 48,747–763 CrossRef PubMed
52 Navarathna, D. H., Hornby, J. M., Krishnan, N., Parkhurst, A., Duhamel, G. E. andNickerson, K. W. (2007) Effect of farnesol on a mouse model of systemic candidiasis,determined by use of a DPP3 knockout mutant of Candida albicans. Infect. Immun. 75,1609–1618 CrossRef PubMed
53 Bishop, A. C., Ganguly, S., Solis, N. V., Cooley, B. M., Jensen-Seaman, M. I., Filler, S. G.,Mitchell, A. P. and Patton-Vogt, J. (2013) Glycerophosphocholine utilization by Candidaalbicans: role of the GIT3 transporter in virulence. J Biol. Chem. 288,33939–33952 CrossRef PubMed
54 Paul, S., Diekema, D. and Moye-Rowley, W. S. (2013) Contributions of Aspergillusfumigates ATP-binding cassette transporter proteins to drug resistance and virulence.Eukaryot. Cell 12, 1619–1628 CrossRef PubMed
55 Mukherjee, P. K., Chandra, J., Kuhn, D. M. and Ghannoum, M. A. (2003) Mechanism offluconazole resistance in Candida albicans biofilms: phase-specific role of effluxpumps and membrane sterols. Infect. Immun. 71, 4333–4340CrossRef PubMed
56 Gacser, A., Stehr, F., Kroger, C., Kredics, L., Schafer, W. and Nosanchuk, J. D. (2007)Lipase 8 affects the pathogenesis of Candida albicans. Infect. Immun. 75,4710–4718 CrossRef PubMed
57 Trofa, D., Soghier, L., Long, C., Nosanchuk, J. D., Gacser, A. and Goldman, D. L. (2011) Arat model of neonatal candidiasis demonstrates the importance of lipases as virulencefactors for Candida albicans and Candida parapsilosis. Mycopathologia 172,169–178 CrossRef PubMed
58 Nagy, I., Filkor, K., Nemeth, T., Hamari, Z., Vagvolgyi, C. and Gacser, A. (2011)In vitro interactions of Candida parapsilosis wild type and lipase deficientmutants with human monocyte derived dendritic cells. BMC Microbiol. 11,122 CrossRef PubMed
59 Nemeth, T., Toth, A., Szenzenstein, J., Horvath, P., Nosanchuk, J. D., Zsuzsanna, G., Toth,R., Papp, C., Hamari, Z., Vagvolgyi, C. and Gacser, A. (2013) Characterization of virulenceproperties in the C. parapsilosis sensu lato species. PLoS ONE 8,e68704 CrossRef PubMed
60 Ghannoum, M. A. (2000) Potential role of phospholipases in virulence and fungalpathogenesis. Clin. Microbiol. Rev. 13, 122–143 CrossRef PubMed
61 Kohler, G. A., Brenot, A., Haas-Stapleton, E., Agabian, N., Deva, R. and Nigam, S. (2006)Phospholipase A2 and phospholipase B activities in fungi. Biochim. Biophys. Acta 1761,1391–1399 CrossRef PubMed
62 Leidich, S. D., Ibrahim, A. S., Fu, Y., Koul, A., Jessup, C., Vitullo, J., Fonzi, W., Mirbod, F.,Nakashima, S., Nozawa, Y. and Ghannoum, M. A. (1998) Cloning and disruption ofcaPLB1, a phospholipase B gene involved in the pathogenicity of Candida albicans. J.Biol. Chem. 273, 26078–26086 CrossRef PubMed
63 Theiss, S., Ishdorj, G., Brenot, A., Kretschmar, M., Lan, C. Y, Nichterlein, T., Hacker, J.,Nigam, S., Agabian, N. and Kohler, G. A. (2006) Inactivation of the phospholipase B genePLB5 in wild-type Candida albicans reduces cell-associated phospholipase A2 activityand attenuates virulence. Int. J. Med. Microbiol. 296, 405–420 CrossRef PubMed
64 Garcıa-Sanchez, S., Aubert, S., Iraqui, I., Janbon, G., Ghigo, J. M. and d’Enfert, C. (2004)Candida albicans biofilms: a developmental state associated with specific and stable geneexpression patterns. Eukaryot. Cell 3, 536–545 CrossRef PubMed
65 Lan, C. Y., Newport, G., Murillo, L. A., Jones, T., Scherer, S., Davis, R. W. and Agabian, N.(2002) Metabolic specialization associated with phenotypic switching in Candidaalbicans. Proc. Natl. Acad. Sci. U.S.A. 99, 14907–14912 CrossRef PubMed
66 Finkel, J. S., Xu, W., Huang, D., Hill, E. M., Desai, J. V., Woolford, C. A., Nett, J. E., Taff,H., Norice, C. T., Andes, D. R. et al. (2012) Portrait of Candida albicans adherenceregulators. PLoS Pathog. 8, e1002525 CrossRef PubMed
67 Sellam, A., Tebbji, F., Whiteway, M. and Nantel, A. (2012) A novel role for the transcriptionfactor Cwt1p as a negative regulator of nitrosative stress in Candida albicans. PLoS ONE7, e43956 CrossRef PubMed
68 Liu, T. T., Lee, R. E., Barker, K. S., Lee, R. E., Wei, L., Homayouni, R. and Rogers, P. D.(2005) Genome-wide expression profiling of the response to azole, polyene,echinocandin, and pyrimidine antifungal agents in Candida albicans. Antimicrob. AgentsChemother. 49, 2226–2236 CrossRef PubMed
Received 2 January 2014/12 March 2014; accepted 13 March 2014Published as BJ Immediate Publication 13 March 2014, doi:10.1042/BJ20140010
c© The Authors Journal compilation c© 2014 Biochemical Society
Copyright © 2022 FDOKUMEN




















