
Novel MRI and fluorescent probes responsive to the Factor XIII transglutaminase activity
10
Received: 8 June 2009, Revised: 13 April 2010, Accepted: 15 April 2010, Published online in Wiley InterScience: 2010 Novel MRI and fluorescent probes responsive to the Factor XIII transglutaminase activity Lorenzo Tei a , Galit Mazooz b , Yael Shellef b , Reut Avni b , Katrien Vandoorne b , Alessandro Barge c , Vyacheslav Kalchenko d , Mark W. Dewhirst e , Linda Chaabane f , Luigi Miragoli f , Dario Longo g , Michal Neeman b * and Silvio Aime g Transglutaminases, including factor XIII and tissue transglutaminase, participate in multiple extracellular processes associated with remodeling of the extracellular matrix during wound repair, blood clotting, tumor progression and fibrosis of ischemic injuries. The aim of this work was to evaluate a novel substrate analog for transglutaminase optimized by molecular modeling calculations (DCCP16), which can serve for molecular imaging of transglutaminase activity by magnetic resonance imaging and by near-infrared imaging. Experimental data showed covalent binding of Gd–DCCP16 and DCCP16-IRIS Blue to human clots, to basement membrane components and to casein in purified systems as well as in three-dimensional multicellular spheroids. In vivo, DCCP16 showed enhancement with a prolonged retention in clots and tumors, demonstrating the ability to detect both factor XIII and tissue transglu- taminase mediated covalent binding of the contrast material. Copyright # 2010 John Wiley & Sons, Ltd. Keywords: molecular imaging; magnetic resonance imaging; near infrared imaging; transglutaminase activity; responsive contract agents; blood clotting; tumor imaging 1. INTRODUCTION Transglutaminases are a family of enzymes that catalyze extracellular covalent cross-linking of proteins, the most prominent of which are factor XIII (also known as plasma transglutaminase), which cross-links fibrin during blood clotting, and tissue transglutaminase (TG2), which catalyzes covalent cross-linking of the extracellular matrix. These enzymes partici- pate in multiple pathological processes associated with degen- erative and proliferative tissue remodeling, including wound healing, cancer, myocardial infarctions and atherothrombosis. Clotting of fibrin is a central process in all these pathologies, and thus it has become fundamental to develop imaging methods to detect endogenous transglutaminase activity associated with clotting of fibrin. Moreover, recent studies suggested the use of transglutaminase therapy to reduce the risk of rupture in myocardial infarction (1). Such therapeutic manipulation of transglutaminase activity could benefit from molecular imaging guidance. Fibrin stabilizing Factor XIII (FXIII) is an enzyme of the blood coagulation system that cross-links fibrin and is involved in many pathologies, such as thrombotic disorders, coronary artery disease, myocardial infarction and cerebrovascular diseases (2). FXIII is also involved in proangiogenetic processes (3), in some tumor development (4), in fibrin stabilization, wound healing and platelet activation (5–7). FXIII plays a key role in the final stages of blood coagulation, producing a mechanically stronger clot with high fibrinolytic resistance due to the FXIII-mediated linking of the plasmin inhibitor (a 2 -antiplasmin) to fibrin (5). Unlike other participants of the multi-enzyme coagulation cascade FXIIIa does not contribute to the process of clotting of blood, but instead stabilizes the clot by cross-linking proteins in the fibrin thrombus by forming an isopeptide bond between the g -carbonyl group of a glutamine (Gln) in one protein and the e-amino group of a lysine (Lys) residue in a nearby protein (5). The effects of TG2 in cancer are complex and not yet fully resolved. On one hand, multiple studies implicated the enhanced (www.interscience.wiley.com) DOI:10.1002/cmmi.392 Full Paper * Correspondence to: M. Neeman, Department of Biological Regulation, Weiz- mann Institute, Rehovot 76100, Israel. E-mail: [email protected] a L. Tei Dipartimento di Scienze dell’Ambiente e della Vita, Universita ` del Piemonte Orientale, Viale T. Michel 11, I-15121, Alessandria, Italy b G. Mazooz, Y. Shellef, R. Avni, K. Vandoorne, M. Neeman Department of Biological Regulation, Weizmann Institute, Rehovot 76100 Israel c A. Barge Dipartimento di Scienza e Tecnologia del Farmaco, Universita ` di Torino, Via P. Giuria 9, I-10125, Torino, Italy d V. Kalchenko Department of Veterinary Resources, Weizmann Institute, Rehovot 76100 Israel e M. W. Dewhirst Department of Radiation Oncology, Box 3455, Duke University Medical Center, Durham, NC 27710 USA f L. Chaabane, L. Miragoli CRB Bracco Imaging S.p.A., Bioindustry Park Canavese, Via Ribes 5, Colleretto Giacosa (TO), 10010, Italy g D. Longo, S. Aime Dipartimento di Chimica IFM, and Molecular Imaging Center, Universita ` di Torino, Via Nizza 52, I-10125, Torino, Italy Contrast Media Mol. Imaging (2010) Copyright # 2010 John Wiley & Sons, Ltd.
Transcript of Novel MRI and fluorescent probes responsive to the Factor XIII transglutaminase activity

Received: 8 June 2009, Revised: 13 April 2010, Accepted: 15 April 2010, Published online in Wiley InterScience: 2010
Novel MRI and fluorescent probes responsiveto the Factor XIII transglutaminase activityLorenzo Teia, Galit Mazoozb, Yael Shellefb, Reut Avnib, Katrien Vandoorneb,Alessandro Bargec, Vyacheslav Kalchenkod, Mark W. Dewhirste,Linda Chaabanef, Luigi Miragolif, Dario Longog, Michal Neemanb*and Silvio Aimeg
Transglutaminases, including factor XIII and tissue transglutaminase, participate in multiple extracellular processesassociated with remodeling of the extracellular matrix during wound repair, blood clotting, tumor progression andfibrosis of ischemic injuries. The aim of this work was to evaluate a novel substrate analog for transglutaminaseoptimized by molecular modeling calculations (DCCP16), which can serve for molecular imaging of transglutaminaseactivity bymagnetic resonance imaging and by near-infrared imaging. Experimental data showed covalent binding ofGd–DCCP16 and DCCP16-IRIS Blue to human clots, to basement membrane components and to casein in purifiedsystems as well as in three-dimensional multicellular spheroids. In vivo, DCCP16 showed enhancement with aprolonged retention in clots and tumors, demonstrating the ability to detect both factor XIII and tissue transglu-taminase mediated covalent binding of the contrast material. Copyright # 2010 John Wiley & Sons, Ltd.
Keywords: molecular imaging; magnetic resonance imaging; near infrared imaging; transglutaminase activity; responsivecontract agents; blood clotting; tumor imaging
1. INTRODUCTION
Transglutaminases are a family of enzymes that catalyzeextracellular covalent cross-linking of proteins, the mostprominent of which are factor XIII (also known as plasmatransglutaminase), which cross-links fibrin during blood clotting,and tissue transglutaminase (TG2), which catalyzes covalentcross-linking of the extracellular matrix. These enzymes partici-pate in multiple pathological processes associated with degen-erative and proliferative tissue remodeling, including woundhealing, cancer, myocardial infarctions and atherothrombosis.Clotting of fibrin is a central process in all these pathologies, andthus it has become fundamental to develop imaging methods todetect endogenous transglutaminase activity associated withclotting of fibrin. Moreover, recent studies suggested the use oftransglutaminase therapy to reduce the risk of rupture inmyocardial infarction (1). Such therapeutic manipulation oftransglutaminase activity could benefit from molecular imagingguidance.Fibrin stabilizing Factor XIII (FXIII) is an enzyme of the blood
coagulation system that cross-links fibrin and is involved in manypathologies, such as thrombotic disorders, coronary arterydisease, myocardial infarction and cerebrovascular diseases (2).FXIII is also involved in proangiogenetic processes (3), in sometumor development (4), in fibrin stabilization, wound healing andplatelet activation (5–7). FXIII plays a key role in the final stages ofblood coagulation, producing a mechanically stronger clot withhigh fibrinolytic resistance due to the FXIII-mediated linking ofthe plasmin inhibitor (a2-antiplasmin) to fibrin (5). Unlike otherparticipants of the multi-enzyme coagulation cascade FXIIIa doesnot contribute to the process of clotting of blood, but insteadstabilizes the clot by cross-linking proteins in the fibrin thrombus
by forming an isopeptide bond between the g-carbonyl group ofa glutamine (Gln) in one protein and the e-amino group of a lysine(Lys) residue in a nearby protein (5).The effects of TG2 in cancer are complex and not yet fully
resolved. On one hand, multiple studies implicated the enhanced
(www.interscience.wiley.com) DOI:10.1002/cmmi.392
Full Paper
* Correspondence to: M. Neeman, Department of Biological Regulation, Weiz-mann Institute, Rehovot 76100, Israel.E-mail: [email protected]
a L. Tei
Dipartimento di Scienze dell’Ambiente e della Vita, Universita del Piemonte
Orientale, Viale T. Michel 11, I-15121, Alessandria, Italy
b G. Mazooz, Y. Shellef, R. Avni, K. Vandoorne, M. Neeman
Department of Biological Regulation, Weizmann Institute, Rehovot 76100
Israel
c A. Barge
Dipartimento di Scienza e Tecnologia del Farmaco, Universita di Torino, Via P.
Giuria 9, I-10125, Torino, Italy
d V. Kalchenko
Department of Veterinary Resources, Weizmann Institute, Rehovot 76100
Israel
e M. W. Dewhirst
Department of Radiation Oncology, Box 3455, Duke University Medical
Center, Durham, NC 27710 USA
f L. Chaabane, L. Miragoli
CRB Bracco Imaging S.p.A., Bioindustry Park Canavese, Via Ribes 5, Colleretto
Giacosa (TO), 10010, Italy
g D. Longo, S. Aime
Dipartimento di Chimica IFM, and Molecular Imaging Center, Universita di
Torino, Via Nizza 52, I-10125, Torino, Italy
Contrast Media Mol. Imaging (2010) Copyright # 2010 John Wiley & Sons, Ltd.

expression of TG2 by tumor cells in metastasis and invasion, inresistance to chemotherapy and inhibition of autophagy, placingTG2 suppression as a candidate approach for therapy in multipletypes of cancer including ovarian carcinoma (8), pancreaticcancer (9,10), malignant melanoma (11) and breast cancer (12).On the other hand, high activity of TG2 in the tumor stromarenders it resistant to proteolytic degradation, and can impedetumor cell invasion, suggesting that TG2 over-expression couldsuppress tumor progression (13). Tissue transglutaminase canhelp in stabilization of the provisional fibrin matrix which isdeposited at the early stages of tumor angiogenesis. Elevatedactivity of transglutaminases was shown at the boundaries ofinvading tumors and several biological events such as cellularproliferation, differentiation and apoptosis are partially regulatedby these enzymes (14).Imaging probes composed of peptides extracted from
a2-antiplasmin, the primary inhibitor of plasmin-mediatedfibrinolysis, were reported for mapping the activity of FXIIIaand of other transglutaminases (15–20). These peptides wereconjugated to a Gd(III)-chelating unit to obtain magneticresonance imaging probes, to a near infrared fluorescent probe(AlexaFlor 680C2) for optical imaging purposes and to 111In-DOTAfor generating Single Photon Emission Computed Tomography(SPECT) probes for nuclear imaging (16–19). Although bothnear-infrared (NIR) imaging and SPECT offer better sensitivity,they lack soft tissue contrast and thus anatomy must be inferredfrom independent measurement using other modalities. More-over, the spatial resolution of both methods is significantly lowerthan that offered by MRI. Thus, MRI appears to provideadvantages for in vivo detection of FXIII activity for diagnosisand monitoring therapies.To widen the armory of imaging tools for this application we
combined molecular mechanics and literature data to design anewmodel peptide conjugated to imaging reporters representedby a Gd–DOTA monoamide (DOTAMA) moiety for MRI visualiza-tion or a fluorescent dye for optical detection. The position of theimaging reporter in the chain was designed to preventany steric hindrance at the enzyme active site. According tothe predictive calculations, the peptide DCCP16 was designedand in this paper we report its ability to enter the complexnetwork of a clot formation, being an efficient substrate for thecatalytic center of FXIII. Furthermore, in vivo validationwas gainedby NIR imaging and MRI mapping of transglutaminase inducedprobe retention in mouse models of tumor xenografts and bloodclotting.
2. RESULTS
2.1. Design and modeling of the imaging probe
Transglutaminases catalyze an acyl transfer reaction forming ane(g-glutamyl)lysyl (isopeptide) bond between the g-carbonylgroup of a peptide-bound Gln and the e-amino group of a Lysresidue in a nearby protein. This transglutaminase reaction causesa fibrin-to-fibrin cross-link and the assembly of the polymeric fibrinnetwork present in the clot (Fig. 1a). The cross-link can be blockedby covalent incorporation of an acyl-acceptor amine substrate(either peptidic or not) into the fibrin units (Fig. 1b) or anacyl-donor Gln-containing peptide complementary to FXIIIa-reactive Lys donor cross-linking sites of the protein (Fig. 1c).The interacting peptidic chains from two fibrin molecules reach
the catalytic center by two opposite channels (Fig. 2): one isspecific for the Gln and the latter for the amine moiety.Since no X-ray crystal structures of substrates or inhibitors
bound to FXIII were reported so far, the new model peptide wasdesigned by combining literature data and molecular modelingprocedures. It consists of two portions: one containing the Glnmoiety and one in which a Lys residue is connected to a spacerwhich protrudes the imaging reporting unit away from theprotein surface.In recent years several peptides based either on b-casein and
a2-antiplasmin for the Gln-donor substrate requirements (21,22)or on the bovine aA-crystallin for the amine-donor substraterequirements have been investigated as model systems for thetransglutaminase activity (23).It is also well established from previous studies that FXIII
and TGs are much more sensitive towards the Gln-bearingsubstrate than to amine donor Lys residues (5). Furthermore,it has been proposed that Gln substrates are able to participatein hydrophobic interactions within the enzyme cavity (24).For these reasons the first moiety consists of the previouslyidentified hexapeptide TVQQEL (25) bearing two Gln moietiesand a Val residue. The Lys substrate binds only after the Glnsubstrate reaction is complete (23,26). For this second substrate,a pentapeptide ALVKG was selected, not only because it isknown to be a good substrate for transglutaminases (27), butalso in order to exploit additional hydrophobic interactions to theFXIII surface.At this stage, molecular mechanics calculations were employed
to evaluate how to connect the two peptidic moieties allowingthe aminoacids to enter the enzyme active site and how toaccommodate the imaging reporter avoiding steric hindranceand therefore enzyme inhibition. Initial positions and confor-mations of the Gln moiety were manually set near the catalyticCys314 and stable complex structures were obtained by energyminimization with the enzyme model atoms being free toaccommodate the Gln substrate. Yee et al. (28) postulated thatCys314 can be approached by the Gln substrate macromoleculefrom the direction of the two b-barrel-domains, most likelydisplacing Tyr560 by movement of barrel 1, and that the Lyssubstrate can access the active-site cavity from the oppositedirection. To identify the residues involved in rotation of barrel 1to make the catalytic site accessible to ligands, we used a hingeprediction server (29). Results from hinge prediction showed, aspossible moving residues, Pro505, Leu506, Asn507, which belongto the loop connecting the core domain and the barrel 1domain, and residues Glu631 and Ile632 forming loop betweenbarrel 1 and barrel 2. This movement induces the displacementof barrel 1 as a result of the activation after binding of the aminesubstrate.Finally, on the basis of the remaining accessible space inside
the FXIII binding cavity, a four Gly spacer and a Lys to allow theconjugation of the DOTAMA unit were inserted in between theLys and the Gln moieties. This spacer was chosen in order to keepthe Gd-complex far enough from interfering with the activesite. The MRI probe was therefore set as GKVLAK–(DOTAMA)–GGGGTVQQEL (DCCP16, Fig. 1f). An analogous analysiswas followed when the conjugation involved the fluorescentdye. As negative control peptide, the amino acids responsiblefor the transglutaminase activity of DCCP16, i.e. Gln3, Gln4 andLys15, were replaced by alanines, thus the probe GAVLAK–(DOTAMA)–GGGGTVAAEL (DCCP16 control) was considered for invivo MRI studies.
www.interscience.wiley.com/journal/cmmi Copyright # 2010 John Wiley & Sons, Ltd. Contrast Media Mol. Imaging (2010)
L. TEI ET AL.

2.2. Synthesis and Characterization of MRI and FluorescentProbes
The solid-phase peptide synthesis of the 16-terms peptidesGKVLAKGGGGTVQQEL and GAVLAKGGGGTVAAEL was followedby the deprotection of IvDde group on the Lys11 and by theattachment of the DOTAMA chelating unit and IRIS Bluefluorescent moiety. The DOTAMA chelating moiety was selectedfor its particularly high thermodynamic and kinetic stability alsoin vivo (30). IRIS Blue is a cyanine dye that shows an intenseabsorption (extinction coefficient: e¼ 127000 M
�1 cm�1) andemission in the visible spectrum (absorption maxi-mum¼ 660 nm, emission maximum¼ 680 nm in phosphatebuffer at pH 7.4) (31). The probes (Fig. 1f and 1g) were thenpurified by HPLC-MS and characterized by ESI-MS. In the case ofthe MRI probes, the Gd(III) complexes were synthesized in waterand controlled by ESI-MS and 1H NMR relaxometry. The NMRD
profile of Gd–DCCP16 in buffer solution (PBS, pH 6.8) is reportedin Fig. 3. A relaxivity of 8.1mM
�1s�1 was measured at 0.5 T and298 K. The analysis of the NMRD profile indicated that theobserved relaxivity is essentially determined by the elongation ofthe molecular reorientational time (tR) relative to parentGd–DOTAMA systems.
2.3. In vitro analysis of factor XIII catalyzed cross-linking offibrin clots.
In order to show that Gd–DCCP16 enter in the clot fibrin network,two types of in vitro MR experimental workup were carried out.The first experiment was designed in order to allow theGd-containing peptide to enter the clot formation, whereasthe second experiment dealt with the evaluation of thenoncovalent interaction between the Gd-containing probe andthe preformed clot.
Figure 1. Different possibility of transglutaminase mediated acyl transfer reaction. Reaction between (a) fibrin units, (b) fibrin and an amino
group-containing substrate, (c) fibrin and a Gln-containing peptide, (d) the two ends of Gd–DCCP16 peptide units (Lys15/Gln3�4) forming oligomers;
(e) incorporation of the Gd–DCCP16 into the fibrin-to-fibrin cross-link. (f ) Chemical structures of Gd–DCCP16 and (g) of DCCP16-IRIS Blue.
Contrast Media Mol. Imaging (2010) Copyright # 2010 John Wiley & Sons, Ltd. www.interscience.wiley.com/journal/cmmi
IMAGING PROBES RESPONSIVE TO TRANSGLUTAMINASES

From in vitro MR experiments, a marked signal enhancementwas observed on clot preparation in which Gd–DCCP16 is part ofthe clot structure, whereas only a small enhancement wasdetectable on clots analogously prepared in the presence ofGd–HP–DO3A (ProHance, Bracco Imaging), a commerciallyavailable, nonspecific MRI contrast agent widely used in clinicalpractice. Moreover, no significant enhancement was observedwhen Gd-complex solutions (Gd–DCCP16 and ProHance) wereincubated with preformed clots (Fig. 3A, B). These data suggestthe absence of noncovalent interaction between DCCP16 and thepreformed clot, whereas DCCP16 is prompt to react with fibrinonly during the process of clot formation. Furthermore, the signalenhancement in MR images reflects the formation of covalentbonds between peptide and fibrin in which Gd–DCCP16 enters inthe clot fibrin network as fibrinogen mimetic.There are many possible linkages between the Gd–DCCP16
g-carbonyl and the e-amino groups of Gln and Lys, respectively,and the proteins in the fibrin thrombus (Fig. 1a–e). In addition tothe simple covalent bond between the peptide Gln (or Lys) andthe protein, in principle, Gd–DCCP16 may form oligomericstructures by FXIII-mediated binding of the Lys residue to the Glnresidue of a nearby peptide, as shown in Fig. 1d. These oligomericstructures may further react with fibrin and may be incorporatedin the clot. The peptide could also be inserted in between a fibrincross-linked system in two steps: first, the reaction of a Lys (or Gln)of DCCP16 with the correspondent fibrin amino acid residue and,second, the reaction of the remaining free Gln (or Lys) of thepeptide with the protein (Fig. 1e).From a 1H NMR relaxometric study, the NMRD profile of the clot
formed in the presence of Gd–DCCP16 does not show anyrelaxivity peak around 35–40MHz, as would have been expectedfor the formation of a rigid and high molecular weightprotein–Gd complex conjugate as a consequence of a markedincrease of reorientational correlation time, tR (Fig. 3C).Interestingly, the high r1p (the proton relaxation enhancementof water protons in the presence of the paramagnetic complex at1mM concentration) value (52.0mM�1 s�1 at 20MHz and 298 K)can be accounted for in terms of a lower molar fraction of waterpresent inside the clot and also by a high content of secondsphere water molecules rigidly assembled in a network of
hydrogen bonds with the protein content of the clots. The fittingprocedure suggests a ca 70% reduction of water fraction insidethe clot structure. Fitting parameters are reported in Table 1S(Supporting Information).These experiments demonstrated the potential use of DCCP16
for molecular imaging of transglutaminase activity. Thus, NIR andMRI probes were generated and applied in follow-up exper-iments for in vitro and in vivo verification of utility in models ofclotting and cancer.
2.4. In vitro NIR imaging demonstrates TG2 activity incovalent cross-linking of DCCP16-IRIS Blue
In order to verify that indeed DCCP16 is covalently linked bytransglutaminase activity, we resorted to fluorescence imaging.Covalent binding of DCCP16-IRIS Blue mediated by TG2 wasdemonstrated by incubation with N0,N0-dimethylcasein (Fig. 4).The reaction included casein 1.25mgml�1, CaCl2, Tris 0.1 M, DTT,TG2 0.05 mg ml�1 and DCCP16-IRIS Blue 0.19mM. The product wasanalyzed using a denaturing SDS gel electrophoresis, monitoringthe Iris Blue fluorescence on an IVIS100 system (Xenogen). Underthese conditions any noncovalent association of the contrastmedia with casein would dissociate. A strong fluorescent bandwas observed corresponding to molecular weight between 25and 50 kDa. Such product can only be explained by covalentbinding of more than onemoiety of DCCP16-IRIS Blue (�2 kDa) toN0,N0-dimethylcasein (180 aa; �25–30 kDa). Hence, DCCP16-IRISBlue can serve as a substrate allowing for covalent cross-linking todimethylcasein, catalyzed by TG2.
2.5. In vitro spheroid labeling: fluorescence and MRimaging
Large MCF7 spheroids, 1mm in diameter, were incubated in RPMIculture medium supplemented with DCCP16-IRIS Blue (0.012mM)at 37 8C for 48 h. The spheroids were then extensively washed inPBS and fluorescence imaging was analyzed by two photonmicroscopy (Fig. 5A; DCCP16 fluorescence in red) and using theIVIS100 system (Xenogen; Fig. 5B, fluorescence in color overlaidon a grayscale photograph). Control spheroids were incubated inRPMI for 48 h in the absence of DCCP16. Strong fluorescence wasobserved only from DCCP16 treated spheroids (Fig. 5A, B rightpanels and Movie 1 in Supporting Information), while controlspheroids showed no fluorescence (Fig. 5 A, B left panels andMovie 2 in Supporting Information). MRI measurements weredone on suspended spheroids incubated for 48 h withGd–DCCP16 (Fig. 5C, D). Contrast enhancement was observedin T1 weighted images of spheroids incubated with Gd–DCCP16(Fig. 5C right panel), relative to control spheroids (Fig. 5C, leftpanel). Considerably high R1 relaxation rate was measured in thepresence of spheroids incubated with Gd–DCCP16 relative tocontrol spheroids (Fig. 5D right and left panels respectively).
2.6. In vivo NIR imaging of factor XIII activity in woundhealing
Thrombi were induced on the right ears of CD1 nude female mice,6 weeks of age, by laser and by thermal injury (Fig. 6). DCCP16-IRISBlue (200 mM, 200 ml) was subsequently administered intrave-nously. Fluorescence was followed using a fluorescence micro-scope (Zeiss) and an IVIS100 system (Xenogen). During the first20min after administration the material was distributed evenlythroughout the entire tissue surrounding the wounds. After 24h,
Figure 2. X-ray crystal structure of Factor XIIIa (PDB code 1F13)
showing the channels through which fibrin(ogen) and DCCP16
peptidic chains access to the enzyme active site. The two DCCP16peptides are represented as ball and stick, green and cyan colored,
respectively.
www.interscience.wiley.com/journal/cmmi Copyright # 2010 John Wiley & Sons, Ltd. Contrast Media Mol. Imaging (2010)
L. TEI ET AL.
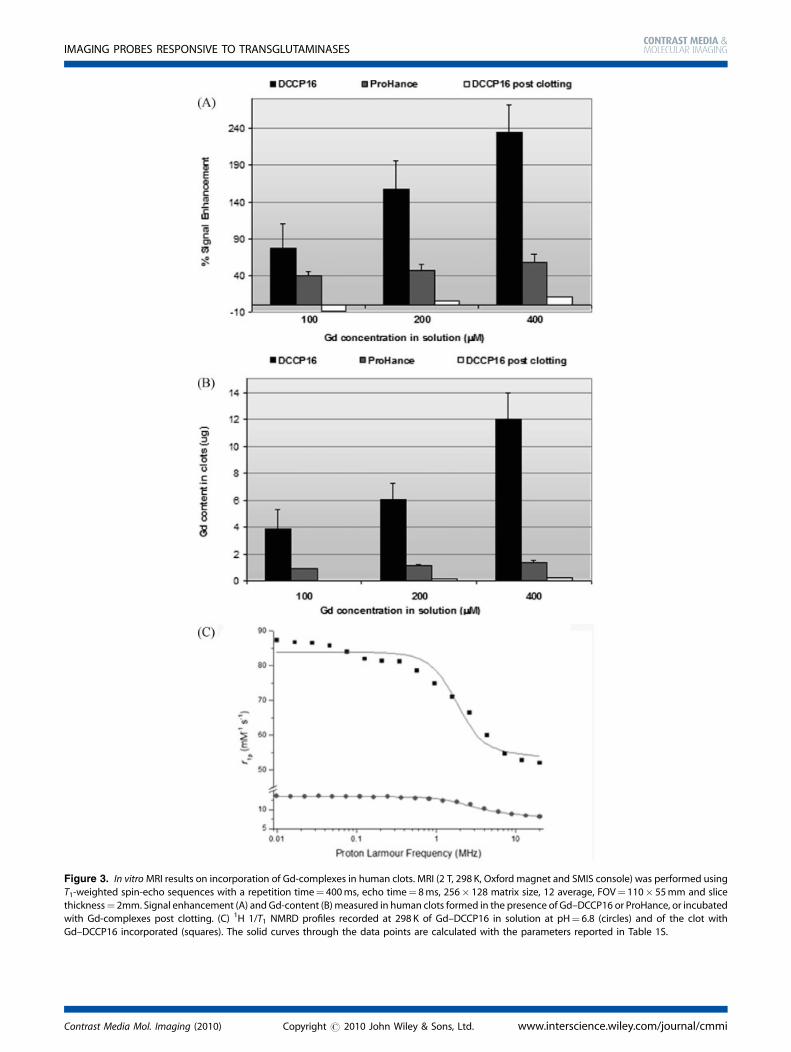
Figure 3. In vitroMRI results on incorporation of Gd-complexes in human clots. MRI (2 T, 298 K, Oxford magnet and SMIS console) was performed using
T1-weighted spin-echo sequences with a repetition time¼ 400ms, echo time¼ 8ms, 256� 128 matrix size, 12 average, FOV¼ 110� 55mm and slice
thickness¼ 2mm. Signal enhancement (A) and Gd-content (B)measured in human clots formed in the presence of Gd–DCCP16 or ProHance, or incubatedwith Gd-complexes post clotting. (C) 1H 1/T1 NMRD profiles recorded at 298 K of Gd–DCCP16 in solution at pH¼ 6.8 (circles) and of the clot with
Gd–DCCP16 incorporated (squares). The solid curves through the data points are calculated with the parameters reported in Table 1S.
Contrast Media Mol. Imaging (2010) Copyright # 2010 John Wiley & Sons, Ltd. www.interscience.wiley.com/journal/cmmi
IMAGING PROBES RESPONSIVE TO TRANSGLUTAMINASES
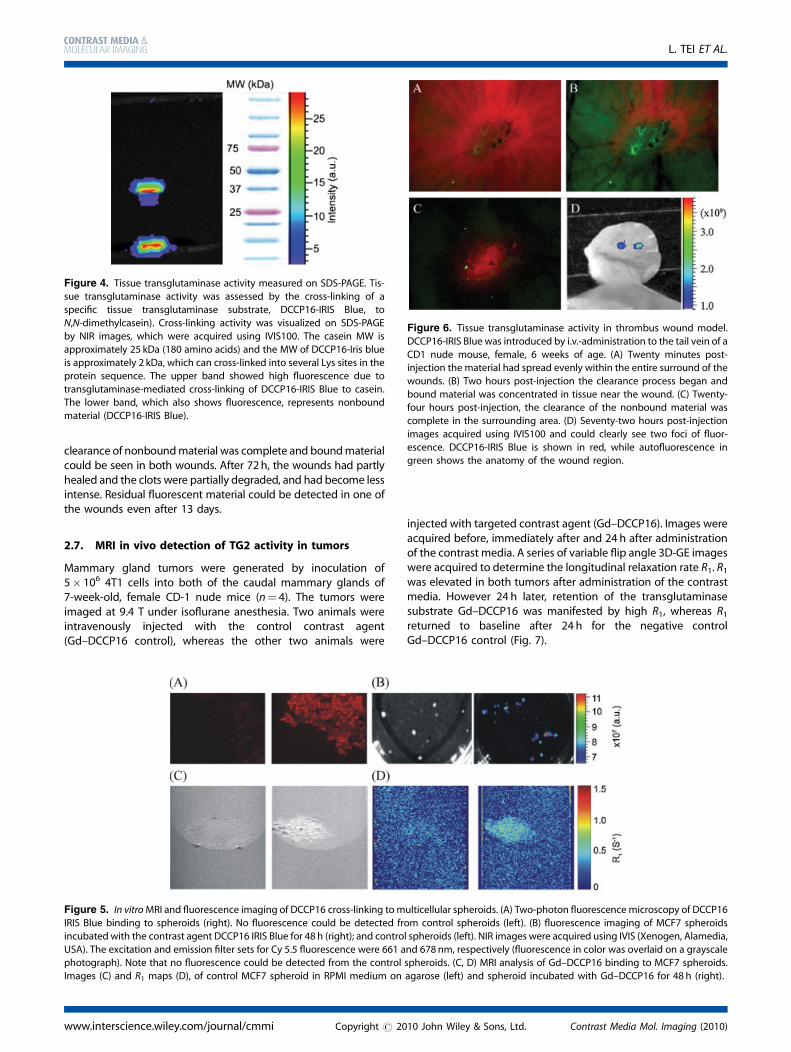
clearance of nonboundmaterial was complete and boundmaterialcould be seen in both wounds. After 72h, the wounds had partlyhealed and the clots were partially degraded, and had become lessintense. Residual fluorescent material could be detected in one ofthe wounds even after 13 days.
2.7. MRI in vivo detection of TG2 activity in tumors
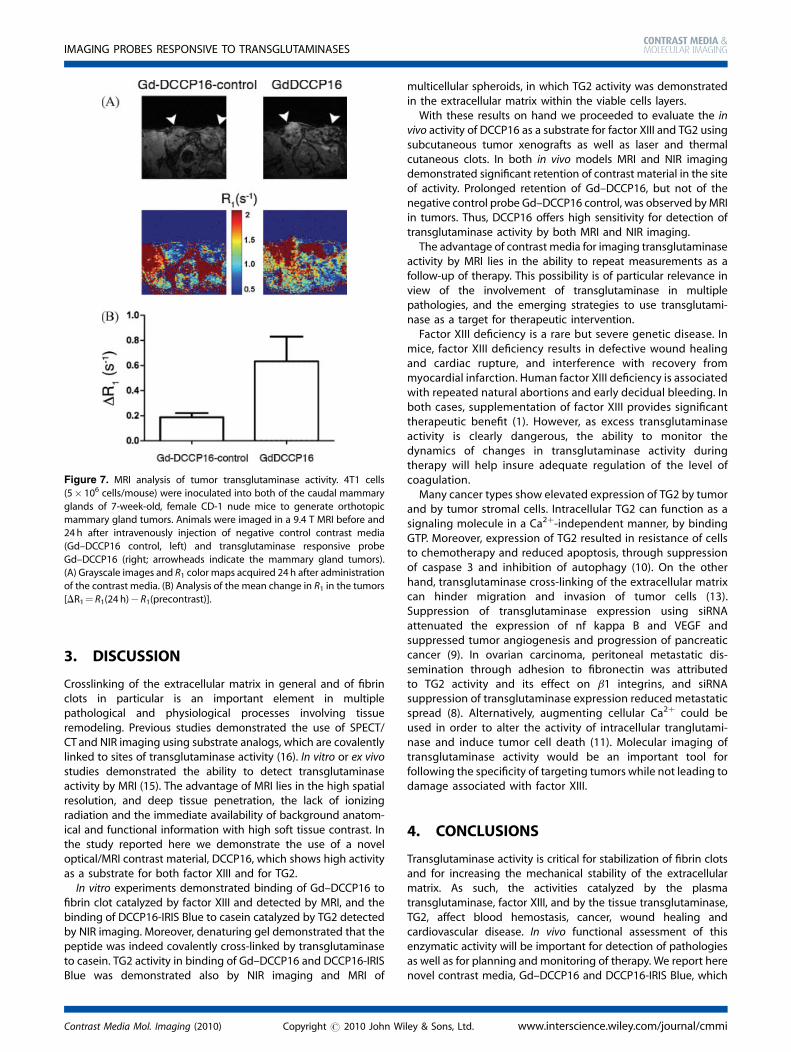
Mammary gland tumors were generated by inoculation of5� 106 4T1 cells into both of the caudal mammary glands of7-week-old, female CD-1 nude mice (n¼ 4). The tumors wereimaged at 9.4 T under isoflurane anesthesia. Two animals wereintravenously injected with the control contrast agent(Gd–DCCP16 control), whereas the other two animals were
injected with targeted contrast agent (Gd–DCCP16). Images wereacquired before, immediately after and 24 h after administrationof the contrast media. A series of variable flip angle 3D-GE imageswere acquired to determine the longitudinal relaxation rate R1. R1was elevated in both tumors after administration of the contrastmedia. However 24 h later, retention of the transglutaminasesubstrate Gd–DCCP16 was manifested by high R1, whereas R1returned to baseline after 24 h for the negative controlGd–DCCP16 control (Fig. 7).
Figure 4. Tissue transglutaminase activity measured on SDS-PAGE. Tis-sue transglutaminase activity was assessed by the cross-linking of a
specific tissue transglutaminase substrate, DCCP16-IRIS Blue, to
N,N-dimethylcasein). Cross-linking activity was visualized on SDS-PAGE
by NIR images, which were acquired using IVIS100. The casein MW isapproximately 25 kDa (180 amino acids) and the MW of DCCP16-Iris blue
is approximately 2 kDa, which can cross-linked into several Lys sites in the
protein sequence. The upper band showed high fluorescence due to
transglutaminase-mediated cross-linking of DCCP16-IRIS Blue to casein.The lower band, which also shows fluorescence, represents nonbound
material (DCCP16-IRIS Blue).
Figure 5. In vitroMRI and fluorescence imaging of DCCP16 cross-linking to multicellular spheroids. (A) Two-photon fluorescence microscopy of DCCP16
IRIS Blue binding to spheroids (right). No fluorescence could be detected from control spheroids (left). (B) fluorescence imaging of MCF7 spheroids
incubated with the contrast agent DCCP16 IRIS Blue for 48 h (right); and control spheroids (left). NIR images were acquired using IVIS (Xenogen, Alamedia,
USA). The excitation and emission filter sets for Cy 5.5 fluorescence were 661 and 678 nm, respectively (fluorescence in color was overlaid on a grayscalephotograph). Note that no fluorescence could be detected from the control spheroids. (C, D) MRI analysis of Gd–DCCP16 binding to MCF7 spheroids.
Images (C) and R1 maps (D), of control MCF7 spheroid in RPMI medium on agarose (left) and spheroid incubated with Gd–DCCP16 for 48 h (right).
Figure 6. Tissue transglutaminase activity in thrombus wound model.
DCCP16-IRIS Blue was introduced by i.v.-administration to the tail vein of a
CD1 nude mouse, female, 6 weeks of age. (A) Twenty minutes post-injection the material had spread evenly within the entire surround of the
wounds. (B) Two hours post-injection the clearance process began and
bound material was concentrated in tissue near the wound. (C) Twenty-four hours post-injection, the clearance of the nonbound material was
complete in the surrounding area. (D) Seventy-two hours post-injection
images acquired using IVIS100 and could clearly see two foci of fluor-
escence. DCCP16-IRIS Blue is shown in red, while autofluorescence ingreen shows the anatomy of the wound region.
www.interscience.wiley.com/journal/cmmi Copyright # 2010 John Wiley & Sons, Ltd. Contrast Media Mol. Imaging (2010)
L. TEI ET AL.
3. DISCUSSION
Crosslinking of the extracellular matrix in general and of fibrinclots in particular is an important element in multiplepathological and physiological processes involving tissueremodeling. Previous studies demonstrated the use of SPECT/CT and NIR imaging using substrate analogs, which are covalentlylinked to sites of transglutaminase activity (16). In vitro or ex vivostudies demonstrated the ability to detect transglutaminaseactivity by MRI (15). The advantage of MRI lies in the high spatialresolution, and deep tissue penetration, the lack of ionizingradiation and the immediate availability of background anatom-ical and functional information with high soft tissue contrast. Inthe study reported here we demonstrate the use of a noveloptical/MRI contrast material, DCCP16, which shows high activityas a substrate for both factor XIII and for TG2.In vitro experiments demonstrated binding of Gd–DCCP16 to
fibrin clot catalyzed by factor XIII and detected by MRI, and thebinding of DCCP16-IRIS Blue to casein catalyzed by TG2 detectedby NIR imaging. Moreover, denaturing gel demonstrated that thepeptide was indeed covalently cross-linked by transglutaminaseto casein. TG2 activity in binding of Gd–DCCP16 and DCCP16-IRISBlue was demonstrated also by NIR imaging and MRI of
multicellular spheroids, in which TG2 activity was demonstratedin the extracellular matrix within the viable cells layers.With these results on hand we proceeded to evaluate the in
vivo activity of DCCP16 as a substrate for factor XIII and TG2 usingsubcutaneous tumor xenografts as well as laser and thermalcutaneous clots. In both in vivo models MRI and NIR imagingdemonstrated significant retention of contrast material in the siteof activity. Prolonged retention of Gd–DCCP16, but not of thenegative control probe Gd–DCCP16 control, was observed by MRIin tumors. Thus, DCCP16 offers high sensitivity for detection oftransglutaminase activity by both MRI and NIR imaging.The advantage of contrast media for imaging transglutaminase
activity by MRI lies in the ability to repeat measurements as afollow-up of therapy. This possibility is of particular relevance inview of the involvement of transglutaminase in multiplepathologies, and the emerging strategies to use transglutami-nase as a target for therapeutic intervention.Factor XIII deficiency is a rare but severe genetic disease. In
mice, factor XIII deficiency results in defective wound healingand cardiac rupture, and interference with recovery frommyocardial infarction. Human factor XIII deficiency is associatedwith repeated natural abortions and early decidual bleeding. Inboth cases, supplementation of factor XIII provides significanttherapeutic benefit (1). However, as excess transglutaminaseactivity is clearly dangerous, the ability to monitor thedynamics of changes in transglutaminase activity duringtherapy will help insure adequate regulation of the level ofcoagulation.Many cancer types show elevated expression of TG2 by tumor
and by tumor stromal cells. Intracellular TG2 can function as asignaling molecule in a Ca2þ-independent manner, by bindingGTP. Moreover, expression of TG2 resulted in resistance of cellsto chemotherapy and reduced apoptosis, through suppressionof caspase 3 and inhibition of autophagy (10). On the otherhand, transglutaminase cross-linking of the extracellular matrixcan hinder migration and invasion of tumor cells (13).Suppression of transglutaminase expression using siRNAattenuated the expression of nf kappa B and VEGF andsuppressed tumor angiogenesis and progression of pancreaticcancer (9). In ovarian carcinoma, peritoneal metastatic dis-semination through adhesion to fibronectin was attributedto TG2 activity and its effect on b1 integrins, and siRNAsuppression of transglutaminase expression reduced metastaticspread (8). Alternatively, augmenting cellular Ca2þ could beused in order to alter the activity of intracellular tranglutami-nase and induce tumor cell death (11). Molecular imaging oftransglutaminase activity would be an important tool forfollowing the specificity of targeting tumors while not leading todamage associated with factor XIII.
4. CONCLUSIONS
Transglutaminase activity is critical for stabilization of fibrin clotsand for increasing the mechanical stability of the extracellularmatrix. As such, the activities catalyzed by the plasmatransglutaminase, factor XIII, and by the tissue transglutaminase,TG2, affect blood hemostasis, cancer, wound healing andcardiovascular disease. In vivo functional assessment of thisenzymatic activity will be important for detection of pathologiesas well as for planning and monitoring of therapy. We report herenovel contrast media, Gd–DCCP16 and DCCP16-IRIS Blue, which
Figure 7. MRI analysis of tumor transglutaminase activity. 4T1 cells
(5� 106 cells/mouse) were inoculated into both of the caudal mammary
glands of 7-week-old, female CD-1 nude mice to generate orthotopicmammary gland tumors. Animals were imaged in a 9.4 T MRI before and
24 h after intravenously injection of negative control contrast media
(Gd–DCCP16 control, left) and transglutaminase responsive probe
Gd–DCCP16 (right; arrowheads indicate the mammary gland tumors).(A) Grayscale images and R1 color maps acquired 24 h after administration
of the contrast media. (B) Analysis of the mean change in R1 in the tumors
[DR1¼ R1(24 h)� R1(precontrast)].
Contrast Media Mol. Imaging (2010) Copyright # 2010 John Wiley & Sons, Ltd. www.interscience.wiley.com/journal/cmmi
IMAGING PROBES RESPONSIVE TO TRANSGLUTAMINASES

allow detection of transglutaminase activities of factor XIII andTG2. These contrast media include a peptidic moiety optimized toserve as a transglutaminase substrate analog, and thus covalentlylinked by transglutaminase in sites of activity, allowing itsdetection by MRI or NIR imaging depending on the imagingprobe attached to the peptide. Noninvasive imaging oftransglutaminase activity by MRI, as reported here, provides animportant tool for monitoring transglutaminase targeted therapyin diverse pathologies including cancer, wound healing,myocardiac infarction and pregnancy failures associated withcongenital factor XIII deficiency.
5. EXPERIMENTAL
5.1. Molecular Calculations
Molecular mechanics calculations with an all-atom force-fieldwere performed using MOE program with an Amber99 forcefield parameter set (MOE, Molecular Operating Environment;www.chemcomp.com). A distance-dependent dielectric constantwas used. Energy minimization was carried out with an energygradient tolerance of 0.01 kcalmol�1 A�1. A nonbonded cut-off of10–12 A was used to model the protein structure.For modeling the binding, the crystal structure of FXIII (PDB
code 1F13) with the highest resolution (2.1 A) available in theProtein Data Bank (32) was used. Hydrogen atoms were added tothe model and the geometry was optimized with a multi-stepprocedure to provide the final binding model of FXIIIa. TheN-terminal activation peptide was removed to make the activesite more accessible. In order to allow the DCCP16 peptide toaccess the active-site cavity, the resulting structure wasenergy-minimized by maintaining fixed the backbone atomsof the enzyme and by allowing the side chains free to move inorder to maximize the binding interaction of the peptide. Then,400 ps of unconstrained molecular dynamics (MD) simulationswere performed in the absence of explicit water molecules butwith a distance-dependent dielectric constant. The canonicalensemble (NVT) was used with a target temperature of 300 K. Anintegration time step of 1 fs was used.
5.2. Reagents
Fmoc-protected amino acids, activators and Rink Amide resin(Advanced Biotech, Italy) were used without purification. IRIS Bluefluorophore and ProHance1 (Gd–HPDO3A) were kindly providedby Cyanine Technologies (Torino, Italy) and Bracco Imaging(Milan, Italy), respectively. Other chemicals were purchased fromSigma-Aldrich Co. DOTA(OtBu)3 was synthesized using a literatureprocedure (33).
5.3. Synthesis of Gd–DCCP16 and Gd–DCCP16 controlpeptide
The 16-terms peptides GKVLAKGGGGTVQQEL (DCCP16) andGAVLAKGGGGTVAAEL (DCCP16 control) were synthesized bysolid-phase peptide synthesis using a Rink Amide resin as solidsupport (0.2 g with loading 0.5mmol g�1) and the standardFmoc strategy. The Lys11 residue, orthogonally protected by a4,4-dimethyl-2,6-dioxocyclohex-1-ylidene (ivDde) group, wasunprotected with 2% hydrazine DMF solution. A protectedDOTA analog (1,4,7,10-tetraazacyclododecane-1,4,7-tris(aceticacid t-butyl ester)-10-acetic acid [DOTA(OtBu)3)] was coupled
to the e-NH2 group of the Lys11 using O-(7-azabenzotriazol-1-yl)-N,N,N’,N’-tetramethyluronium hexafluorophosphate (HATU)as activator and N,N- diisopropylethyl amine (DIPEA) as base. Thepeptides were then removed from the resin, purified by HPLC-MS(Waters FractionLynx using a Waters XTerra RPC18 19/50 column,HPLC-MS method 1 [see Supporting Information]) to a finalpurity> 95%. DCCP16: retention time 4.5min; obtained 48mg,25% yield; ESI-MS (Waters Micromass ZQ): calcd, 1927.7; found:1928.2 [MþHþ]. DCPP16 control: retention time 4.9min,obtained 35mg, 20% yield; ESI-MS: calcd, 1755.4; found:1756.2 [MþHþ].An equimolar amount of GdCl3 was slowly added to a 40mM
water solution of DCCP16 and DCCP16 control peptides,maintaining the pH value at 6.5 with NaOH. The mixture wasallowed to stir overnight at RT; then the pH was raised to 8.5 andthe mixture was stirred for 2 h. Centrifugation at 8000 rpm for5min at 10 8C allowed the separation of Gd(OH)3 from thesolution. The Orange Xylenol UV method (34) was used toquantify the residual free Gd3þ ion present in Gd–DCCP16 andGd–DCCP16 control, which was found to be under 0.3%(molmol�1). The final products were subjected to inductivelycoupled plasma spectrometry (ICP-MS) for the exact determi-nation of the Gd content, then lyophilized.ESI-MS: Gd–DCCP16 – calcd, 2081.4; found, 1041.2, 1041.7,
1042.2 [(Mþ 2Hþ), z¼ 2]; Gd–DCCP16 control – calcd, 1909.6;found: 956.0, 956.5 [(Mþ 2Hþ), z¼ 2]; isotopic distributionconsistent with Gd complexes.Gd–DCCP16 was characterized by acquiring its NMRD profile
on the Field Cycling Relaxometer (Stelar, Italy) operatingover an extended frequency range (0.01–20MHz). The obtained1/T1 values of proton water molecules of 1mM solution ofthe Gd-containing probe were analyzed on the basis of theestablished Solomon–Bloenbergen–Morgan equations for theinner-sphere contribution and Freed’s equations for the outersphere term (35).
5.4. Synthesis of DCCP16-IRIS Blue
A solution of dicyclohexylcarbodiimide (2.2mg,10.5mmol) inDMF (0.5ml) was added to a solution of IRIS Blue (5.0mg, 7mmol)and N-hydroxysuccinimide (1.2mg, 10.5mmol) dissolved in DMF(0.5ml). The reaction was heated to 508C and left in the darkunder stirring for 2 h. The solution obtained was used in theconjugation step without further purification. Thus, the resi-n-anchored DCCP16 peptide, after deprotection of the IvDdegroup, was reacted with the DMF solution of the activated IRISBlue fluorophore for 2 h in the dark and at room temperature.Cleavage from the resin and HPLC purification [Amersham AKTAPurifier 10/100 using a Waters XTerra RPC18 19/50 column,HPLC-MS method 2 (see Supporting Information), retention time5.37min] gave the DCCP16-IRIS Blue compound in 16% yield(2.5mg, 1.12mmol). The product was then characterized byESI-MS (DCCP16-IRIS Blue: calcd, 2230.6; found, 1116.2 [(Mþ 2Hþ),z¼ 2]).
5.5. In vitro MR imaging tests in human plasma clots
Clots were formed in a 2ml vial with 300 ml of reconstitutedHuman plasma and 300 ml of Pathromtin SL (silicon dioxideparticles, vegetable phospholipids, NaCl 2.4 g l�1, Hepes 14.3 gl�1, pH 7.6). After incubation for two minutes at 378C, with orwithout Gd–DCCP16, 300ml of CaCl2 solution was added to form
www.interscience.wiley.com/journal/cmmi Copyright # 2010 John Wiley & Sons, Ltd. Contrast Media Mol. Imaging (2010)
L. TEI ET AL.

the clot. The formed clots were washed four times with 3ml ofTBS to remove unbound Gd-complex and placed in 1.5ml vialsfor MRI measurements. Experiments were performed withGd–DCCP16 and ProHance (control) at different concentrationsof gadolinium (100–400mM).MRI was performed at 2 T (Oxford magnet and SMIS console)
using T1-weighted spin-echo sequences with a repetitiontime¼ 400ms, echo time¼ 8ms, 256� 128 matrix size, 12average, FOV¼ 110� 55mm and slice thickness¼ 2mm (258C).Maximum signal enhancement was calculated by the formula:
SI ¼ S Ið ÞGd�S Ið ÞcS Ið Þc
where S(I)Gd represents the signal intensity of treated clots andS(I)c the signal from untreated clots, respectively. The clotstreated and untreated with Gd–DCCP16 were also analyzed byfield cycling relaxometry. The obtained NMRD profiles werecompared with that obtained for an aqueous solution ofGd–DCCP16 in order to extract information on the relativemobility of the incorporated probe and on the relative amount ofwater in the clot interacting with the paramagnetic centers. Afterthe MR imaging experiments, the clots underwent ICP-MSanalysis to determine the amount of Gd(III). The latter analyticalmethodology implied a complete mineralization (concentrationHNO3, 90 8C, overnight) of clot specimens.
5.6. In vitro fluorescence imaging of covalent cross-linkingby TG2
Casein (0.04ml; 1.25mgml�1) was added to TG2 (0.002ml; 2 mgml�1) and DCCP16-IRIS Blue (0.002ml; 12mM). After incubation(45min, 37 8C) the sample was boiled and loaded on SDS-PAGEgel (12.5% along with markers). Casein cross-linked withDCCP16-IRIS Blue is expected at MW higher than 30 kDa.
5.7. In vitro analysis of TG2 activity in 3D multicellulartumor spheroids
MCF7 breast carcinoma spheroids were cultured in RPMIsupplemented with 10% FCS (Biological Industries, Beit Haemek,Israel) and antibiotics (50 units ml�1 penicillin and 50 units ml�1
streptomycin) as reported previously (15). Briefly, aggregation ofcells into spheroids was initiated by plating cells on agar-coatedplates. After 48 h the spheroids were transferred to a 500mlspinner flask (Belco, Vineland, NJ, USA); themediumwas changedevery day. Spheroids were incubated with DCCP16-IRIS Blue orGd–DCCP16. NIR images of the spheroids were acquired at lowresolution (IVIS100, Xenogen) and high resolution (2P micro-scopy; Zeiss LSM 510 META NLO, Germany; equipped with aBroad Band Mai Tai–HP–femtosecond single box tunableTi-Sapphire oscillator, with automated broad band wavelengthtuning 700–1020 nm from Spectraphysics, USA, for two photonexcitation). MRI measurements were done on a vertical bore 9.4 Tspectrometer (Bruker, Karsruhe). R1 maps were derived by singleexponential fit of spin echo images acquired at different TRvalues.
5.8. In vivo imaging of thermal clot in CD-1 mice
All animal experiments were approved by the Institutional AnimalCare and Use Committee of the Weizmann Institute. Clots wereinduced in the ears of CD-1, 6-week-old female nude mice, underanesthesia (75mg kg�1 ketamine i.p., Fort Dodge Animal Health;
3mg kg�1 xylazine i.p, XYZ-M2 V.M.D). A total of six mice werestudied, using either laser (n¼ 2) or heated needle (n¼ 4) toinduce two or three small clots localized under a microscope to asingle vessel in a single ear. The contrast agent DCCP16-IRIS Bluewas injected through the tail vain immediately after the injury(150ml, 1.8mM). Serial images were acquired using the IVIS100system with in a period of time between 24 h and 13 days afteradministration of the contrast material.In vivo NIR images were acquired using IVIS 100 (Xenogen). The
excitation and emission filter sets for 678 and 661 nmrespectively. Mice were visualized 20min to 13 days afteradministration of the contrast material.
5.9. Tumor xenografts
4T1 breast carcinoma cells were cultured in DMEM, supple-mented with 10% FCS (Biological Industries, Beit Haemek, Israel),2mM L-Gln and antibiotics (50 units ml�1 penicillin and 50 unitsml�1 streptomycin).Orthotopic mammary gland tumors were generated by
inoculation of 5� 106 4T1 cells into both of the caudal mammaryglands of 7-week-old, female CD-1 nude mice (n¼ 4). The tumorswere imaged on a 9.4 T Bruker Biospec (Bruker, Karlsruhe,Germany) using a 20mm surface coil for receiving and a linearcoil for transmitting. Two animals were intravenously injectedwith control contrast agent (Gd–DCCP16 control), whereas theother two animals were injected with the transglutaminaseresponsive probe (Gd–DCCP16). The animals were imaged beforecontrast and 24 h after contrast injection. The 3D field of view(FOV) of the image covered the lower abdomen. A series ofvariable flip angle precontrast T1-weighted 3D-GE images wereacquired to determine the endogenous precontrast R1. Imagingparameters: precontrast flip angles were 5, 15, 30, 50 and 708; TRwas 10ms; TE was 4ms; two averages; spectral width 50 kHz;matrix 256� 128� 64; zero-filled to 256� 256� 128; FOV35� 35� 17.5mm; resolution 137� 273� 137mm; acquisitiontime 163 s.
6. SUPPORTING INFORMATION
Supporting information can be found in the online version of thisarticle.
Acknowledgements
This work was supported in part by EC-FP6-projects (MEDITRANS:NMP4-CT-2006-026668; DiMI: LSHB-CT-2005-512146), and ENCI-TE-project (FP7-HEALTH-2007-A), in part by the Israel ScienceFoundation to M.N., and in part by Italian Government (FIRBRBNE03PX83_006, FIRB RBIP06293N) to S.A. EC-COST D38 Actionis acknowledged. Cyanine Technology is gratefully acknowl-edged for providing a small amount of Iris Blue dye.
REFERENCES1. Nahrendorf M, Aikawa E, Figueiredo JL, Stangenberg L, van den
Borne SW, Blankesteijn WM, Sosnovik DE, Jaffer FA, Tung CH,Weissleder R. Transglutaminase activity in acute infarcts predictshealing outcome and left ventricular remodelling: implicationsfor FXIII therapy and antithrombin use in myocardial infarction.Eur Heart J 2008; 29: 445–454.
Contrast Media Mol. Imaging (2010) Copyright # 2010 John Wiley & Sons, Ltd. www.interscience.wiley.com/journal/cmmi
IMAGING PROBES RESPONSIVE TO TRANSGLUTAMINASES

2. Ariens RA, Lai TS, Weisel JW, Greenberg CS, Grant PJ. Role of factorXIII in fibrin clot formation and effects of genetic polymorphisms.Blood 2002; 100: 743–754.
3. Dardik R, Loscalzo J, Eskaraev R, Inbal A. Molecular mechanismsunderlying the proangiogenic effect of factor XIII. Arterioscler.Thromb Vasc Biol 2005; 25: 526–532.
4. Wojtukiewicz MZ, Rucinska M, Zacharski LR. Localization of bloodcoagulation factors in situ in pancreatic carcinoma. ThrombHaemost. 2001; 86: 1416–1420.
5. Muszbek L, Yee VC, Hevessy Z. Blood coagulation factor XIII:structure and function. Thromb Res 1999; 94: 271–305.
6. Standeven KF, Ariens RA, Grant PJ. The molecular physiology andpathology of fibrin structure/function. Blood Rev 2005; 19:275–288.
7. Serrano K, Devine DV. Intracellular factor XIII cross-links plateletcytoskeletal elements upon platelet activation. Thromb Haemost.2002; 88: 315–320.
8. SatpathyM, Cao L, Pincheira RE, Emerson R, Bigsby R, Nakshatri H,Matei D. Enhanced peritoneal ovarian tumor dissemination bytissue transglutaminase. Cancer Res 2007; 67: 7194–7202.
9. Verma A, Guha S, Diagaradjane P, Kunnumakkara AB, SanguinoAM, Lopez-Berestein G, Sood AK, Aggarwal BB, Krishnan S,Gelovani JG, Mehta K. Therapeutic significance of elevated tissuetransglutaminase expression in pancreatic cancer. Clin CancerRes 2008; 14: 2476–2483.
10. Akar U, Ozpolat B, Mehta K, Fok J, Kondo Y, Lopez-Berestein G.Tissue transglutaminase inhibits autophagy in pancreatic cancercells. Mol. Cancer Res 2007; 5: 241–249.
11. Fok J, Ekmekcioglu S, Mehta K. Implications of tissue transglu-taminase expression in malignant melanoma. Mol Cancer Ther2006; 5: 1493–1503.
12. Kim DS, Park SS, Nam BH, Kim IH, Kim SY. Reversal of drugresistance in breast cancer cells by transglutaminase 2 inhibitionand nuclear factor-kappaB inactivation. Cancer Res 2006; 66:10936–10943.
13. Mangala LS, Arun B, Sahin AA, Mehta K. Tissue transglutamina-se-induced alterations in extracellular matrix inhibit tumor inva-sion. Mol Cancer 2005; 9: 4–33.
14. Griffin M, Casadio R, Bergamini CM. Transglutaminases: nature’sbiological glues. Biochem J 2002; 368: 377–396.
15. Mazooz G, Mehlman T, Lai TS, Greenberg CS, Dewhirst MW,Neeman M. Development of magnetic resonance imaging con-trast material for in vivo mapping of tissue transglutaminaseactivity. Cancer Res 2005; 65: 1369–1375.
16. Tung CH, Ho NH, Zeng Q, Tang Y, Jaffer FA, Reed GL, Weissleder R.Novel factor XIII probes for blood coagulation imaging. Chem-BioChem 2003; 4: 897–899.
17. Nahrendorf M, Hu K, Frantz S. Factor XIII deficiency causes cardiacrupture, impairs wound healing, and aggravates cardiac remo-deling in mice with myocardial infarction. Circulation 2006; 113:1196–1202.
18. Kim DE, Jaffer FA, Weissleder R, Tung CH, Schellingerhout D.Near-Infrared fluorescent imaging of cerebral thrombi andblood–brain barrier disruption in a mouse model of cerebralvenous sinus thrombosis. J Cerebr Blood Flow Metab 2005; 25:226–233.
19. Jaffer FA, Tung CH, Wykrzykowska JJ, Ho NH, Houng AK, Reed GL,Weissleder R. Molecular imaging of factor XIIIa activity in
thrombosis using a novel, near-infrared fluorescent contrastagent that covalently links to thrombi. Circulation 2004; 110:170–176.
20. Miserus RJ, Herıas MV, Prinzen L, Lobbes MB, Van Suylen RJ,Dirksen A, Hackeng TM, Heemskerk JW, van Engelshoven JM,Daemen MJ, van Zandvoort MA, Heeneman S, Kooi ME. JACCCardiovasc Imag. 2009; 2: 987–96.
21. Marinescu A, Cleary DB, Tara R, Littlefield TR, Maurer MC. Struc-tural features associated with the binding of glutamine-containing peptides to Factor XIII. Arch Biochem Biophys 2002;406: 9–20.
22. Gorman JJ, Folk JE. Structural features of glutamine substrates fortransglutaminase. Specificities of human plasma factor XIIIa andthe guinea pig liver enzyme toward synthetic peptides. J BiolChem 1981; 256: 2712–2715.
23. Grootjans JJ, Groenen PJ, de Jong WW. Substrate requirementsfor transglutaminases. Influence of the amino acid residue pre-ceding the amine donor lysine in a native protein. J Biol Chem1995; 270: 22855–22858.
24. McDonagh J, Fukue H. Determinants of substrate specificity forfactor XIII. Semin Thromb Hemost 1996; 22: 369–376.
25. Groenen PJ, Bloemendal H, de Jong WW. The carboxy-terminallysine of alpha B-crystallin is an amine-donor substrate for tissuetransglutaminase. Eur J Biochem 1992; 205: 671–674.
26. Sobel JH, Gawinowicz MA. Identification of the alpha chain lysinedonor sites involved in factor XIIIa fibrin cross-linking. J Biol Chem1996; 271: 19288–19297.
27. Groenen PJ, Smulders RH, Peters RF, Grootjans JJ, van den IjsselPR, Bloemendal H, de Jong WW. The amine-donor substratespecificity of tissue-type transglutaminase. Influence of aminoacid residues flanking the amine-donor lysine residue. Eur JBiochem 1994; 220: 795–799.
28. Yee VC, Pedersen LC, Le Trong I, Bishop PD, Stenkamp RE, TellerDC. Three dimensional structure of a transglutaminase: Humanblood coagulation factor XIII. Proc Natl Acad Sci USA 1994; 91:7296–7300.
29. Krebs WG, Gerstein M. The morph server: a standardized systemfor analyzing and visualizing macromolecular motions in a data-base framework. Nucleic Acids Res 2000; 28: 1665–1675.
30. Geninatti Crich S, Cabella C, Barge A, Belfiore S, Ghirelli C,Lattuada L, Lanzardo S, Mortillaro A, Tei L, Visigalli M, Forni G,Aime S. In vitro and in vivo magnetic resonance detection oftumor cells by targeting glutamine transporters with Gd-basedprobes. J Med Chem 2006; 49: 4926–4936.
31. Bertolino CA, Caputo G, Barolo C, Viscardi G, Coluccia S. Novelheptamethine cyanine dyes with large Stokes’ shift for biologicalapplications in the near infrared. J Fluoresc 2006; 16: 221–225.
32. Weiss MS, Metzner HJ, Hilgenfeld R. Two nonproline cis peptidebonds may be important for factor XIII function. FEBS Lett 1998;423: 291–296.
33. Eisenwiener KP, Powell P, Maecke HR. A convenient synthesis ofnovel bifunctional prochelators for coupling to bioactive pep-tides for radiometal labelling. Bioorg Med Chem Lett 2000; 10:2133–2135.
34. Barge A, Cravotto G, Gianolio E, Fedeli F. Contrast Media MolImag. 2006; 1: 184–188.
35. Aime S, Botta M, Terreno E. Gd(III)-based contrast agents for MRI.Adv Inorg Chem 2005; 57: 173–237.
www.interscience.wiley.com/journal/cmmi Copyright # 2010 John Wiley & Sons, Ltd. Contrast Media Mol. Imaging (2010)
L. TEI ET AL.




















