
Revision of the Neotropical genus Cheiromyia Dyte (Diptera: Dolichopodidae
Upload
independentCategory
view
2download
0
ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2014 Magnolia Press
Zootaxa 3764 (2): 152–168
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3764.2.3
http://zoobank.org/urn:lsid:zoobank.org:pub:82E13B7A-C007-4345-B4CF-D4921194DBA2
Notes on the first instar larvae of Ctenophora and Nephrotoma
(Diptera, Tipulidae)
VIRGINIJA PODENIENE1, NIJOLE NASEVICIENE2 & SIGITAS PODENAS1, 2
1Department of Zoology, Vilnius University, Ciurlionio 21/27, 03101 Vilnius, Lithuania. E-mail [email protected] of Ecology, Nature Research Centre, Akademijos 2, 08412 Vilnius, Lithuania.
E-mail [email protected]. [email protected]
Abstract
1830 egg-larvae of 7 species belonging to long palped crane flies (Tipulidae): Ctenophora guttata Meigen, Nephrotoma
pratensis Linnaeus, N. dorsalis Fabricius, N. scurra Meigen, N. flavescens Linnaeus, N. submaculosa Edwards and N. cro-
cata Linnaeus were obtained from 22 females captured in Lithuania in 2011–2012. It took from five days to more than
three weeks for eggs to hatch. Crane flies have four instars of larvae. Second, third and the last instar larvae are very sim-
ilar, when the first instar or egg-larvae differs radically. Descriptions and illustrations of external morphology, chaetotaxy
of abdominal segments, characters of head capsules and last abdominal segments are given for the previously unknown
first instar larvae of Ct. guttata, N. crocata, N. dorsalis, N. flavescens, N. pratensis, N. scurra and poorly known N. sub-
maculosa. It was found out that difference of head capsule and last abdominal segment among the first instar larvae of
above mentioned species of genus Nephrotoma are more obvious than in last instar. During this study it was found, that
such characters as shape of apical teeth of mandible, shape of basal segment of antenna and number of sensillae, shape of
hypostomium and arrangement of sensory structures on labrum, differ among egg-larvae of Nephrotoma. It was found,
that pads on frontal part of prothorax and shape of lateral plates of egg-larvae labrum of Nephrotoma differ significantly
from that of Ctenophora and could be used as genus separating characters.
Key words: Crane fly, head capsule, egg-larva, last abdominal segment
Introduction
Long palped crane flies (Tipulidae) is one of the largest Diptera families, which counts 4276 recognized species
(Oosterbroek 2013) and what makes up almost 30 percent of the total number of all crane flies (Tipuloidea). All
crane flies have four larval instars, pupae and adult (Henning 1950) in their development. Adults of crane flies are
one of the most commonly encountered flies easily recognized because of typical general appearance and long legs.
They are short living insects with almost aphagous adult stage. The greatest part of their life crane flies spent as
larvae. Preimaginal stages of Tipulidae are not as well studied as adults. The best studied are the last instar larvae,
but even then, last instar larvae of less than 0.5 percent of all Tipulidae species are known and described at the
moment (Chiswell 1956, Gelhaus 1986, Theowald 1967, Savchenko 1983, Podeniene 2003). Knowledge of larval
ecology is mostly based on European and North American species and still is poorly understood. It is known, that
larvae of Tipulidae can develop in different aquatic and terrestrial habitats, most of them consume decaying plant
material (Pritchard 1983). Larvae of several species are serious pests.
The first instar larvae are very poorly known. The first descriptions of egg-larvae were made in the early
twentieth century. After that, the first instar larvae of a little more than 20 species were described (Gerbig 1913,
Hemminsen 1965, Savchenko 1983, Kim & Lee 2004, Lantsov 1982, 1984, 2009). It is known, that features of
head capsules, general morphology and chaetotaxy of the second and third instar larvae are very similar to the last
instar, when the first instar or egg-larvae differs drastically (Gerbig 1913, Hemmingsen 1965, Savchenko 1983).
Spiracular field of egg-larva has just lateral and ventral spiracular lobes, when in other instars, dorsal lobes are
present too. Egg-larvae have few pairs of tiny sclerites with very long bristles on dorsal part of spiracular field and
152 Accepted by V. Blagoderov: 13 Dec. 2013; published: 10 Feb. 2014

very long apical bristle on lateral lobe (Hemmingsen 1965). Differences in head capsules of last instar larvae and
egg-larvae could be found in structure of mandible, hypoharynx and hypostomium (Savchenko 1983).
Higher-level and interfamily classification of crane flies is still unresolved (Petersen et al., 2010). Although
Tipulidae in most analyses show monophyly, position of this family within Tipuloidea (Tipulomorpha) is less
certain. Phylogenetic analysis, based on larval head characters, showed close relationship between Tipulidae and
Cylindrotomidae, but limited data did not clarify this problem (Neugart et al., 2009). Relationships between genera
and subgenera of Tipulidae are also are unresolved. Morphological data from all free-living stages combined with
molecular data can provide the most consistent and supported results (Petersen et al., 2010). Morphological data
from egg-larvae are of great importance to these issues, but they were not used in any phylogenetic analyses.
Genus Nephrotoma Meigen, 1803 is the second largest genus of family Tipulidae. 424 species are distributed
in all zoogeographical regions and almost half of them can be found in Palaearctic (Oosterbroek 2013).
Nephrotoma shows great uniformity in general appearance of adults and run close to those of Tipula. Last instar
larvae of these genera are similar too. Larvae of Nephrotoma can be distinguished from Tipula just by presence of a
pair of welts on the dorsal side of the thorax. Larvae of Nephrotoma are terrestrial and can be found developing in
uppermost layer of soil, leaf litter, dung and even in rotten wood (Chiswell 1956, Podeniene 2003). Egg-larvae of
almost 20 species of genus Tipula are described or illustrated (de Jong 1925, White 1951, Hennig 1950, Foote
1963, Theowald 1967, Pritchard & Hall 1971, Savchenko 1983, Lantsov 1982, Hartman & Hynes 1982, Podeniene
2001, Kim & Lee 2004, Lantsov 2009). Just one illustrated larva of Nephrotoma sp. and short descriptions of
spiracular fields of Nephrotoma quadrifaria, N. submaculosa and N. analis are known from literature
(Hemmingsen 1965, Savchenko 1983).
Genus Ctenophora Meigen, 1803 is a small genus, which counts 31 species and 28 of them are described from
Palaearctic. Adults are large, brightly colored and usually ichneumon or wasp-like. So far it is not clear, what
features could be used for discrimination of the last instar larvae of genus Ctenophora from larvae of Tipula,
Dictenidia and Phoroctenia. Larvae of Ctenophora develop in rotten wood. Last instar larvae of only three
European species of Ctenophora are described (Chiswell 1956, Sorensen 2002) and just one egg-larva of
Ctenophora sp. is illustrated so far (Gerbig 1913, Savchenko 1983).
Material and methods
Material has been collected by N. Naseviciene in Lithuania, Vilnius and Lazdijai districts in 2011–2012. Cranefly
adults were collected with entomological net. 22 caught females laid the eggs. Those were: Ctenophora guttata (1
female), Nephrotoma pratensis (6 females), N. dorsalis (3 females), N. scurra (4 females), N. flavescens (1
female), N. submaculosa (1 female), N. crocata (6 females). Adult females have been determined by S. Podenas.
Eggs for rearing were obtained from decapitated fertilized females and larvae were subsequently reared in
laboratory in Petri dishes and incubated on damp filter paper. The filter paper was kept constantly moist but not
wet, at temperature between 18°C and 25°C. It took from five days to more than three weeks for eggs to hatch:
Ctenophora guttata, 6 days; Nephrotoma crocata, from 6 to 8 days; N. scurra, from 6 to 16 days N. submaculosa,
about 7 days; N. dorsalis, from 14 to 23 days; N. flavescens, 23 days; N. pratensis, 7 to 21 days. Totally 1830 egg-
larvae of above mentioned specimens were studied: Ctenophora guttata (93 specimens), Nephrotoma pratensis
(915 specimens), N. dorsalis (176 specimens), N. scurra (394 specimens), N. flavescens (26 specimens), N.
submaculosa (15 specimens) and N. crocata (212 specimens).
The first instar larvae were killed with boiling water and stored in 70 percent ethanol with some glycerin. The
head capsules were removed from the rest of the body and macerated in room temperature KOH overnight, then
temporary slides of whole head capsule and some parts (hypostomium, hypopharynx, mandibles, maxillae) in
concentrated glycerin were made. Last abdominal segment after measurements was cut from the rest of the body
and placed in glycerine on slide with spiracular field on top. Arrangement of setae on sixth abdominal segment
have been illustrated and described. Pictures were taken with Infinity camera, and Infinity Analyse software v 6.1.
SEM images were taken with The Quanta 250 microscope at the Laboratory of Bedrock Geology at Nature
Research Centre, Lithuania. Later, all pictures were traced and redrawn by hand.
Specimens were studied with Nikon E200 microscope. Figures of spiracular discs were prepared by Inga
Marciukaityte; the others by Nijole Naseviciene.
Material is deposited at Zoological Museum of Vilnius University.
Zootaxa 3764 (2) © 2014 Magnolia Press · 153TIPULIDAE EGG-LARVAE

Systematics
Order Diptera
Family Tipulidae
Subfamily Ctenophorinae
Ctenophora (Ctenophora) guttata Meigen, 1818
(Figs. 1–17)
First instar larvae. Length 2.5–2.6 mm; width–0.44–0.45 mm. Body covered with pale pubescence. Cuticle is
transparent.
Head capsule. Length 0.41–0.43 mm; width 0.24–0.25 mm. Head capsule is prognatous and hemicephalic,
oval in shape, slightly depressed dorsoventrally and differently from the head capsule of the last instar larvae, just
slightly sclerotised (Figs. 1, 2). Internolateralia and externolateralia are separated by incisions, which reaches
almost half-length of head capsule. Each incision is continuous with deep groove (the most sclerotised part of head
capsule) that extends almost to the base of antenna. Externolateralia are widely separated on the ventral side of
capsule (Fig. 2). Premaxillary suture separates side plates from the rest of head capsule. Side plate is wedge shaped
with anteromedially situated long seta. Hypostomium bears six large sharp teeth (Fig. 3). Basally it is fused with
ventral margins of genae and with side plates. Prementum is visible from below. It has three large sharp teeth on
anterior margin. A pair of labial palps is present in the middle part of prementum, each consisting of five peg-like
sensillae. Labial palps are separated by cushion of short bristles (Fig. 4). Frontoclypeus is fused with
internolateralia. Clypeal part of frontoclypeus is membranous and separated from labrum by deep fold. One long
seta and three sensory pits present on the anterior part of clypeus, one long and one short seta present near the base
of antenna. Three very short bristles are located near the grove on both sides of clypeus (Fig. 1). Ecdysial and
coronal sutures are not visible. Labrum is trapezoidal in shape and dorsally is composed of two wedge-shaped
sclerotised plates separated by membranous area (Fig. 5). Apical part of labrum and ephipharynx are covered with
numerous short hairs. One long seta and one short sensory peg present on membranous part on both sides of
labrum. One long, one of medium length, two short setae and three sensory pits present on anteriolateral part of
labral lobe. One long and one short seta are located on posterior part of labral lobe. Antenna is short, slightly
tapering apically. It has just one cylindrical segment, which length is almost two-thirds the width (Fig. 6). Apically
it has one large cone-shaped and six small peg-like sensillae, dorsally it has a sensory pit near the base. Mandible is
one segmented and more sclerotised than the rest of head capsule. Five sharp teeth are present on each mandible
(Fig. 7). Second dorsal and second ventral teeth are much smaller than apical, first ventral and dorsal teeth.
Prostheca or lacinia mobilis present on the dorsal side of mesal mandibular base (Fig. 7). Lacinia mobilis is arch-
shaped, with 11–12 sharp small teeth on apical part and with cushion of short hairs in the middle. Lateral margin of
mandible has two long setae near the base. A sensory pit is present at the base of dorsal side of mandible.
Mandibles operate in horizontal plane. Inconspicuous larval eye spots present below the base of mandible. Maxilla
has wedge-shaped cardo, bearing two almost equal setae near the distal end (Fig. 8). There are two sclerites on
stipes. The inner sclerite extends around inner margin of maxilla onto its dorsal surface. The outer sclerite extends
around outer margin of maxilla to its dorsal surface. There is a diamond-shaped sclerite near the base of galea (Fig.
8). Galea and lacinia are fused. Galea is covered with numerous short hairs. There are three long sensory structures
on the ventral side of galea. Lacinia bears five sharp pointed outgrooves on the outer margin.
Thorax. All thoracic segments are short and wide, covered with microscopic hairs. The first thoracical segment
has no welts (Figs. 9, 10).
Abdomen. Abdominal segments are longer than wide. They are covered with two types of hairs. Short hairs
cover lateral parts of segments and large partions of tergal and sternal parts (Figs. 11, 12, 13). Long hairs form
transversal lines on tergal and sternal parts (Figs. 11, 12). Setae are light brown. Setae D1–D4 and D6 are long,
more than five times as long as D5. Setae D2, D3 and D5, D6 are very close to each other (Fig. 14). Setae V1 and
V3 are very long. They are twice as long as V4 and almost five times as long as V5. Setae V1, V2 and V4, V5 are
very close to each other (Fig.15). Seta L3 is very long. It is slightly longer than L2, twice as long as L1 and more
than six times as long as L4. Setae L1, L2 and L3 are very close to each other (Fig. 16).
PODENIENE ET AL.154 · Zootaxa 3764 (2) © 2014 Magnolia Press

Hairs on sixth and seventh abdominal segments are longer than those on other segments. Third thoracic and
first abdominal segments are wider than longer. First, second thoracic and second–eighth abdominal segments are
longer than wide. Last or anal abdominal segment is constricted.
FIGURES 1–10. Ctenophora (Ctenophora) guttata, first instar larva: 1. General view head capsule, dorsal aspect. 2. General
view head capsule, ventral aspect. 3. Hypostomium. 4. Prementum. 5. General view of labrum. 6. Right antenna. 7. Mandible.
8. Right maxilla, ventral view. 9. The first thoracical segment, dorsal view. 10. The first thoracical segment, lateral view.
Spiracular disc. Spiracular field is surrounded by four flat and elongate round-tipped lobes (Fig. 17), which are
almost completely covered with large, wedge-shaped sclerites. Lateral pair is more prominent. Lateral lobe is
almost as long as its width at the base. It has four long stout apical bristles. Each bristle is almost five times as long
as the lobe. Length of ventral lobe is almost 1.5x its width at the base. The lobe has one long and one medium-
length apical bristles and single long bristle on inner and on outer margins. They are as long as the lobe. It also has
Zootaxa 3764 (2) © 2014 Magnolia Press · 155TIPULIDAE EGG-LARVAE

one of medium-long setae and a short bristle on outer margin. There are four tufts of longer and shorter bristles
above the each spiracle. The innermost tuft consists of three short bristles. Laterally from it, is tuft of four very long
bristles (they are almost three times as long as bristles of the innermost tuft). Further laterally, is tuft of three long
bristles and outermost is one long bristle. Spiracles are large, circular and close to each other. Distance between
them is smaller than diameter of a spiracle.
Anal field. Anus is surrounded by four short, white and fleshy anal papillae. A few long setae present on anal
segment.
Specimens examined. 93 egg-larvae from 1 female, which was captured in Vilnius, N54.68194, E25.27087, 12
VI 2011, eggs were laid on 13 VI 2011, egg-larvae hatched on 19 VI 2011.
FIGURES 11–17. Ctenophora (Ctenophora) guttata, first instar larva: 11. Abdominal segment, lateral view. 12. Abdominal
segment, dorsal view. 13. Abdominal segment, ventral view. 14. Dorsal abdominal setae (segments II–VII). 15. Ventral
abdominal setae (segments II–VII). 16. Lateral abdominal setae (segments II–VII). 17. Spiracular field.
PODENIENE ET AL.156 · Zootaxa 3764 (2) © 2014 Magnolia Press

Subfamily Tipulinae
Nephrotoma flavescens Linnaeus, 1758
(Figs. 18–32)
First instar larvae. Length 1.75–1.78 mm; width 0.29–0.30 mm. Body covered with pale pubescence. Cuticle is
transparent.
Head capsule. Length 0.36–0.38 mm; width 0.16–0.18 mm. Head capsule is prognatous and hemicephalic. It is
elongate-oval in shape, slightly depressed dorsoventrally and, differently from the head capsule of the last instar
larvae, just slightly sclerotised (Fig. 18). Internolateralia and externolateralia are separated by incisions, which
reach almost half-length of head capsule; each incision is continuous with deep groove (the most sclerotised part of
head capsule) that extends almost to the base of antenna. Externolateralia are widely separated on the ventral side
of capsule (Fig. 19). Premaxillary suture separates side plates from the rest of head capsule. Side plate is wedge-
shaped with a long anteromedial seta. Hypostomium is strongly sclerotised. It bears five sharp teeth. The
outhermost teeth (both on dorsal and ventral part) are much smaller than the other. Small incision is on posterior
part of hypostomium (Fig. 20). Basally the hypostomium is fused with ventral margins of genae and with side
plates. Prementum is visible from below. It is very similar to that of Ct. guttata. Frontoclypeus is short (4/5 times as
long as internolateralia) and fused with internolateralia. Internolateralia are separated by fold on posterior part
(Figs. 18, 19). Internolateralia are much longer than externolateralia. Clypeal part of frontoclypeus is membranous
and separated from labrum by deep fold. There are four short setae on the anterior part of clypeus. A short seta is
situated below the anterior part of clypeus towards the base of antenna. There are four short setae below the base of
antenna (Fig. 18). Ecdysial and coronal sutures are not visible. Labrum is rectangular in shape and composed of
two wedge-shaped sclerotised plates separated by membranous areas. Each labral lobe has distinct triangularly
shaped outgrowth directed outward (Fig. 21). Apical part of labrum and ephipharynx are covered with numerous
hairs. One long and one short seta present on membranous part on each side of labrum (Fig. 22). One long, one
medium-length, two short setae and one sensory pit present on anteriolateral part, two sensory pits present on
medial part, one short and one very short setae are located on posterior part of labral lobe. Antenna is short and
composed of one cone-shaped segment, almost as long as wide at base (Fig. 23). It has a large cone-shaped and a
few small peg-like apical sensillae and subapical posterodorsal sensory pit. Mandible is one-segmented and much
more sclerotised than the rest of head capsule. Five sharp teeth are present on both mandibles (Fig. 24). Apical, first
ventral and dorsal teeth are the most prominent. Second dorsal and second ventral teeth are much smaller.
Prostheca or lacinia mobilis present on the dorsal side of inner surface of mandible (Fig. 24). Lacinia mobilis is
arch-shaped, with 9–10 sharp small teeth on apical part and with cushion of long hairs in the middle. Lateral
surface of mandible has two very short setae at the base. A sensory pit is present at the base of dorsal side.
Mandibles operate in horizontal plane. Inconspicuous larval eye spot is present below the base of mandible.
Maxilla has triangularly shaped cardo, which bears two almost equal setae near the distal end (Fig. 25). There are
two sclerites on stipes. The inner sclerite extends around inner margin of maxilla onto its dorsal surface and the
outer sclerite extends around outer margin of maxilla to its dorsal surface (Fig. 25). Galea and lacinia are fused.
Galea is covered with numerous short hairs. There are three long sensory structures on the ventral side of galea.
Lacinia has three sharp pointed outgrooves on the outer margin.
Thorax. All thoracic segments are short and wide, covered with microscopic hairs. Prothorax has dorsal and
ventral pads (Figs. 26, 27).
Abdomen. Abdominal segments are longer than wide. They are covered with two types of hairs. Short hairs
cover lateral parts of segment and big portion of tergal and sternal parts. Tips of these hairs are three-branched
(Figs. 28). Long hairs form lines on tergal and sternal parts of segments. Setae are light brown. Setae D1 and D6
are very short; D4 and D5 are twice as long as D1; D2 is three times as long as D1; D3 is more than five times as
long as D1. Setae D2, D3 and D4, D5 make pairs and are very close to each other (Fig. 29). Setae V1 and V2 are
equal in length and twice as long as V3 and V4. Setae V1, V2 and V3, V4 make pairs and are close to each other
(Fig. 30). Setae L1, L2 and L3 are close to each other (Fig. 31). L2 is short; L1 is three times as long as L2 and
twice as long as L3; L4 is rather far away from other setae and its length is similar to that of L3.
Hairs on sixth and seventh abdominal segments are longer than those on other segments. Third thoracic and
first abdominal segments are wider than long. First and second thoracic and second to eighth abdominal segments
are longer than wide. Last or anal abdominal segment is constricted.
Zootaxa 3764 (2) © 2014 Magnolia Press · 157TIPULIDAE EGG-LARVAE
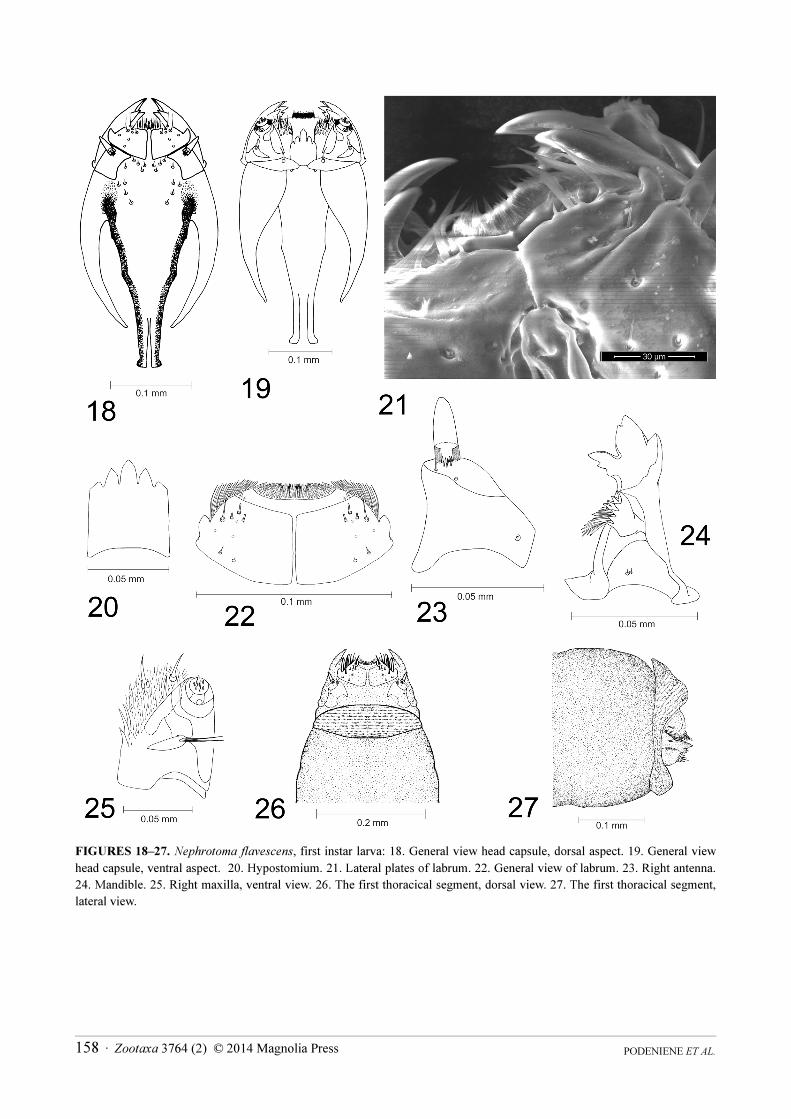
FIGURES 18–27. Nephrotoma flavescens, first instar larva: 18. General view head capsule, dorsal aspect. 19. General view
head capsule, ventral aspect. 20. Hypostomium. 21. Lateral plates of labrum. 22. General view of labrum. 23. Right antenna.
24. Mandible. 25. Right maxilla, ventral view. 26. The first thoracical segment, dorsal view. 27. The first thoracical segment,
lateral view.
PODENIENE ET AL.158 · Zootaxa 3764 (2) © 2014 Magnolia Press

FIGURES 28–32. Nephrotoma flavescens, first instar larva: 28. Abdominal segment, dorsal view. 29. Dorsal abdominal setae
(segments II–VII). 30. Ventral abdominal setae (segments II–VII). 31. Lateral abdominal setae (segments II–VII). 32.
Spiracular field.
Spiracular disc. Spiracular field is surrounded by four flat and elongate round-tipped lobes (Fig. 32). Lateral
lobe is prominent, its length almost 1.5x the width at the base. It has two long stout apical bristles. Each bristle is
almost six times as long as the lobe itself and about as long as one-third of the whole body. Three tufts of two short
bristles each present on dorsal part. There are three tufts of long bristles above the spiracles. The innermost and
outermost tufts consist of two bristles, the middle tuft consists of three bristles. Ventral lobes are almost as long as
wide at the base. A tuft of four long bristles is on the apical part of lobe and a tuft of three slightly shorter bristles is
on inner part; two tufts of two short bristles each present on the outer margin; a tuft of four bristles different in
length and a single bristle are at the base of lobe. Both pairs of lobes are almost completely covered with large,
wedge-shaped sclerites. Spiracles are large, oval and close to each other. Distance between them is almost the same
as diameter of a spiracle itself.
Anal field. Anus is surrounded by four short, white and fleshy anal papillae. There are few long setae on anal
segment (here and in other Nephrotoma species below it was difficult to establish number and arrangement of setae
due to preservation).
Specimens examined. 26 egg-larvae from 1 female, female was captured in Lazdijai district, N54.24080,
E23.51548, 23 VI 2011, eggs were laid on 24 VI 2011, egg-larvae hatched on 17 VII 2011.
Nephrotoma crocata Linnaeus, 1758
(Figs. 33–40)
Zootaxa 3764 (2) © 2014 Magnolia Press · 159TIPULIDAE EGG-LARVAE

First instar larvae. Length 1.83–1.85 mm; width 0.32–0.33 mm. Body covered with pale pubescence. Cuticle is
transparent.
Head capsule. Length 0.40–0.42 mm; width 0.20–0.22 mm. In general, is very similar to that of N. flavescens.
Differences were noticed in shape of hypostomium, mandible, antenna and arrangement of sensory structures on
labrum. Hypostomium bears five blunt teeth (Fig. 33). Mandible generally is very similar to that of N. flavescens,
but shapes of teeth are slightly different (Fig. 34). Segment of antenna is cone-shaped, but membranous apical part
is much smaller than in N. flavescens (Fig. 35). There are three small peg-like sensillae on the apex of basal
segment. Labrum in general is very similar to that of N. flavescens, but there are two long setae on membranous
part of it (Fig. 36).
Thorax. Is very similar to that of N. flavescens.
Abdomen. In general is very similar to that of N. flavescens. Setae D1, D4, D5 and D6 are very short (Fig. 37);
D3 is almost twice as long as D1; D2 is very long almost six times as long as D1. Setae D2, D3 and D4, D5 make
pairs and are very close to each other. Seta V1 is short; V4 is twice as long as V4 (Fig. 38); V3 is slightly longer
than V4; V2 is almost eight times as long as V1. Setae L1, L2 and L3 are close to each other (Fig. 39). Seta L2 is
short; L1 is three times as long as L2 and just slightly longer than L3; L4 is situated further from other setae and is
approximately as long as L3.
Spiracular disc. Spiracular field is surrounded by four flat and elongate round-tipped lobes (Fig. 40). Lateral
pair is more prominent. Lateral lobe is almost as long as wide at the base. It bears four long stout apical bristles.
Tuft of two short bristles is located on dorsal part of it and next by it is a single very short bristle. Lobe bears very
pale wedge-shape sclerite. Ventral lobe length is nearly half its width at the base. It has a tuft of four bristles at the
base. These bristles are different in length. Tuft of two short bristles is on the apical part of lobe and a tuft of three
different in length bristles on inner part of lobe. Two short bristles are on the outer margin of lobe. Lobe is almost
completely covered with large and very pale sclerites. There are three tufts of long bristles above the spiracles. The
outermost tuft consists of two bristles, the others consist of three bristles. Spiracles are large, circular and close to
each other. Distance between the spiracles is smaller than diameter of a spiracle itself.
Anal field. Anus is surrounded by four long, cone-shaped, white and fleshy anal papillae. There are a few long
setae on anal segment.
Specimens examined. 10 egg-larvae from female captured in Vilnius, N54.68194, E25.27087, 11 VI 2011, eggs
were laid on 24 VI 2011, egg-larvae hatched on 22 VI 2011; 16 egg-larvae from female captured near Lazdijai,
N54.24096, E23.51549, 23 VI 2011, eggs were laid on 23 VI 2011, egg-larvae hatched on 01 VII 2011; 39 egg-
larvae from female captured near Lazdijai, N54.24060, E23.51560, 23 VI 2011, eggs were laid on 24 VI 2011, egg-
larvae hatched on 30 VI 2011; 3 egg-larvae from female captured near Lazdijai, N54.24080, E23.51548, 23 VI
2011, eggs were laid on 28 VI 2011, egg-larvae hatched on 04 VII 2011; 76 egg-larvae from female captured near
Lazdijai, N54.24080, E23.51548, 23 VI 2011, eggs were laid on 23 VI 2011, egg-larvae hatched on 01 VII 2011; 67
egg-larvae from female captured near Lazdijai, N54.24095, E23.51552, 23 VI 2011, eggs were laid in 25 VI 2011,
egg-larvae hatched on 03 VII 2011.
Nephrotoma pratensis Linnaeus, 1758
(Figs. 41–48)
First instar larvae. Length 1.75–1.85 mm; width 0.32–0.33 mm. Body covered with pale pubescence. Cuticle is
transparent.
Head capsule. Length 0.38–0.39 mm; width 0.22–0.23 mm. In general is very similar to that of N. flavescens.
Differences were noticed in shape of hypostomium, mandible, antenna and arrangement of sensory structures on
labrum. Hypostomium bears five blunt teeth. There is a prominent incision on posterior part of hypostomium (Fig.
41). Mandible in general is very similar to that of N. flavescens, but shape of teeth is slightly different (Fig. 42).
Basal segment of antenna is cone-shaped, but membranous apical part is much smaller than in N. flavescens (Fig.
43). There are several small peg-like sensillae on the apex of basal segment. Labrum in general is very similar to
that of N. crocata (Fig. 44).
PODENIENE ET AL.160 · Zootaxa 3764 (2) © 2014 Magnolia Press
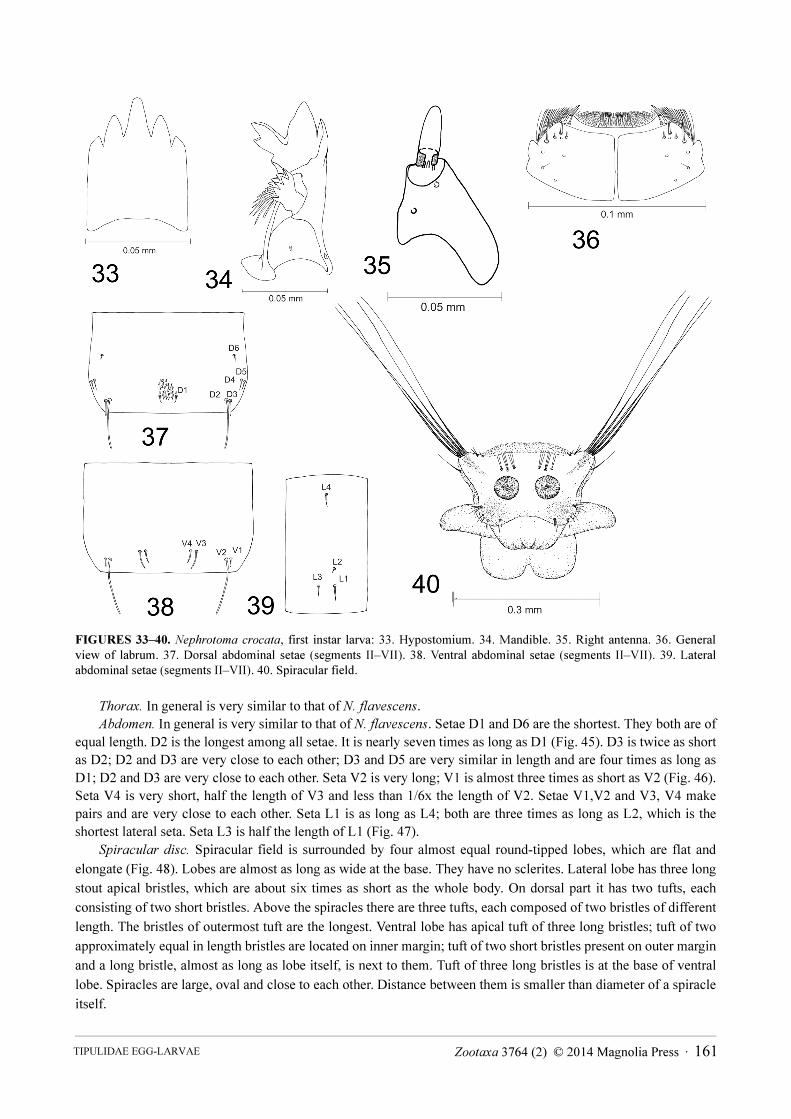
FIGURES 33–40. Nephrotoma crocata, first instar larva: 33. Hypostomium. 34. Mandible. 35. Right antenna. 36. General
view of labrum. 37. Dorsal abdominal setae (segments II–VII). 38. Ventral abdominal setae (segments II–VII). 39. Lateral
abdominal setae (segments II–VII). 40. Spiracular field.
Thorax. In general is very similar to that of N. flavescens.
Abdomen. In general is very similar to that of N. flavescens. Setae D1 and D6 are the shortest. They both are of
equal length. D2 is the longest among all setae. It is nearly seven times as long as D1 (Fig. 45). D3 is twice as short
as D2; D2 and D3 are very close to each other; D3 and D5 are very similar in length and are four times as long as
D1; D2 and D3 are very close to each other. Seta V2 is very long; V1 is almost three times as short as V2 (Fig. 46).
Seta V4 is very short, half the length of V3 and less than 1/6x the length of V2. Setae V1,V2 and V3, V4 make
pairs and are very close to each other. Seta L1 is as long as L4; both are three times as long as L2, which is the
shortest lateral seta. Seta L3 is half the length of L1 (Fig. 47).
Spiracular disc. Spiracular field is surrounded by four almost equal round-tipped lobes, which are flat and
elongate (Fig. 48). Lobes are almost as long as wide at the base. They have no sclerites. Lateral lobe has three long
stout apical bristles, which are about six times as short as the whole body. On dorsal part it has two tufts, each
consisting of two short bristles. Above the spiracles there are three tufts, each composed of two bristles of different
length. The bristles of outermost tuft are the longest. Ventral lobe has apical tuft of three long bristles; tuft of two
approximately equal in length bristles are located on inner margin; tuft of two short bristles present on outer margin
and a long bristle, almost as long as lobe itself, is next to them. Tuft of three long bristles is at the base of ventral
lobe. Spiracles are large, oval and close to each other. Distance between them is smaller than diameter of a spiracle
itself.
Zootaxa 3764 (2) © 2014 Magnolia Press · 161TIPULIDAE EGG-LARVAE
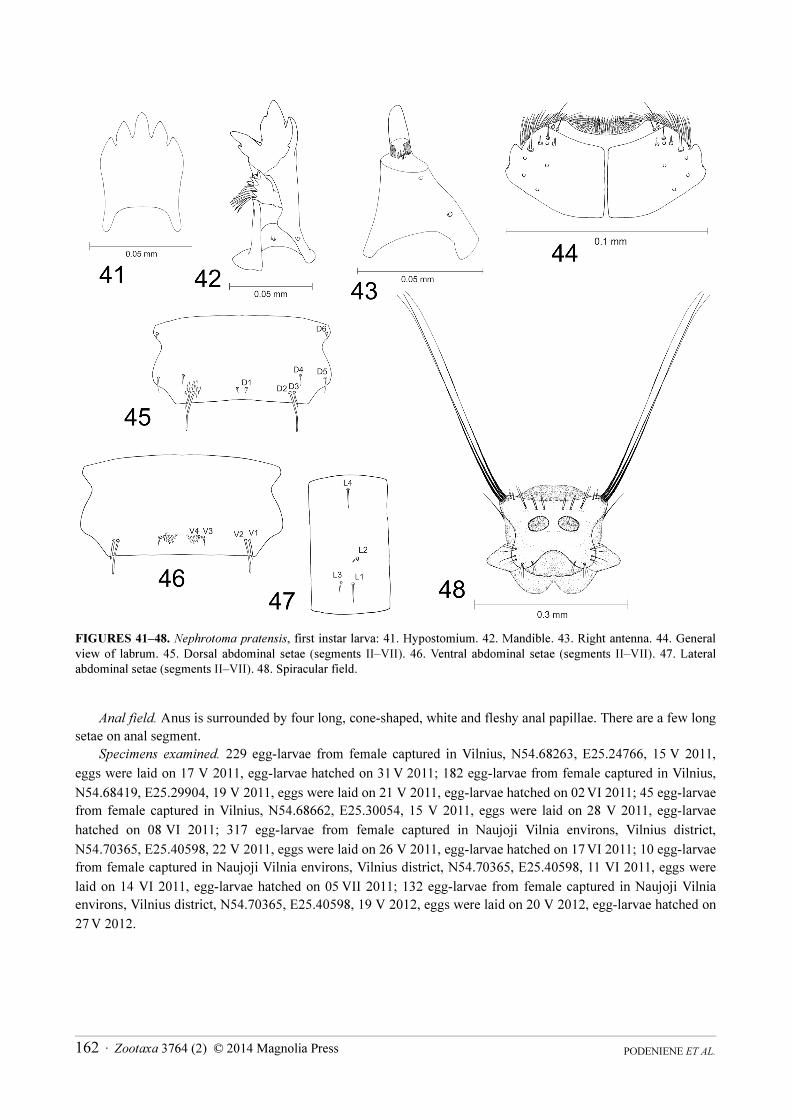
FIGURES 41–48. Nephrotoma pratensis, first instar larva: 41. Hypostomium. 42. Mandible. 43. Right antenna. 44. General
view of labrum. 45. Dorsal abdominal setae (segments II–VII). 46. Ventral abdominal setae (segments II–VII). 47. Lateral
abdominal setae (segments II–VII). 48. Spiracular field.
Anal field. Anus is surrounded by four long, cone-shaped, white and fleshy anal papillae. There are a few long
setae on anal segment.
Specimens examined. 229 egg-larvae from female captured in Vilnius, N54.68263, E25.24766, 15 V 2011,
eggs were laid on 17 V 2011, egg-larvae hatched on 31 V 2011; 182 egg-larvae from female captured in Vilnius,
N54.68419, E25.29904, 19 V 2011, eggs were laid on 21 V 2011, egg-larvae hatched on 02 VI 2011; 45 egg-larvae
from female captured in Vilnius, N54.68662, E25.30054, 15 V 2011, eggs were laid on 28 V 2011, egg-larvae
hatched on 08 VI 2011; 317 egg-larvae from female captured in Naujoji Vilnia environs, Vilnius district,
N54.70365, E25.40598, 22 V 2011, eggs were laid on 26 V 2011, egg-larvae hatched on 17 VI 2011; 10 egg-larvae
from female captured in Naujoji Vilnia environs, Vilnius district, N54.70365, E25.40598, 11 VI 2011, eggs were
laid on 14 VI 2011, egg-larvae hatched on 05 VII 2011; 132 egg-larvae from female captured in Naujoji Vilnia
environs, Vilnius district, N54.70365, E25.40598, 19 V 2012, eggs were laid on 20 V 2012, egg-larvae hatched on
27 V 2012.
PODENIENE ET AL.162 · Zootaxa 3764 (2) © 2014 Magnolia Press
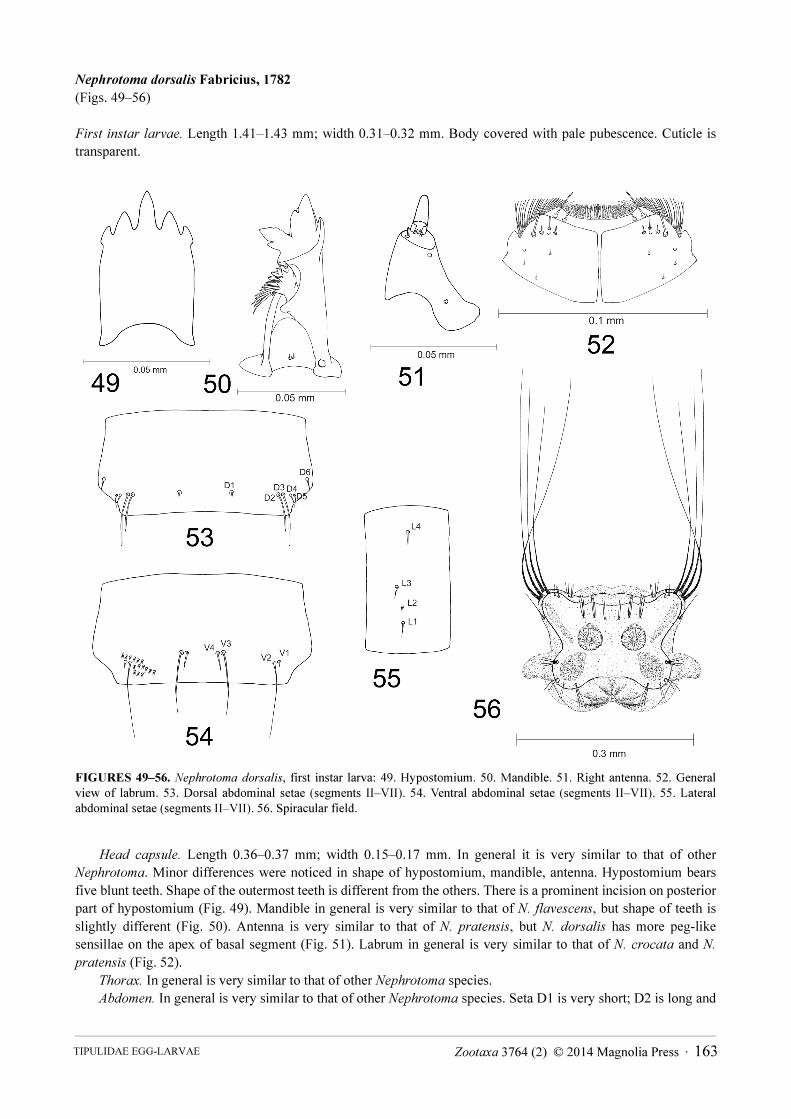
Nephrotoma dorsalis Fabricius, 1782
(Figs. 49–56)
First instar larvae. Length 1.41–1.43 mm; width 0.31–0.32 mm. Body covered with pale pubescence. Cuticle is
transparent.
FIGURES 49–56. Nephrotoma dorsalis, first instar larva: 49. Hypostomium. 50. Mandible. 51. Right antenna. 52. General
view of labrum. 53. Dorsal abdominal setae (segments II–VII). 54. Ventral abdominal setae (segments II–VII). 55. Lateral
abdominal setae (segments II–VII). 56. Spiracular field.
Head capsule. Length 0.36–0.37 mm; width 0.15–0.17 mm. In general it is very similar to that of other
Nephrotoma. Minor differences were noticed in shape of hypostomium, mandible, antenna. Hypostomium bears
five blunt teeth. Shape of the outermost teeth is different from the others. There is a prominent incision on posterior
part of hypostomium (Fig. 49). Mandible in general is very similar to that of N. flavescens, but shape of teeth is
slightly different (Fig. 50). Antenna is very similar to that of N. pratensis, but N. dorsalis has more peg-like
sensillae on the apex of basal segment (Fig. 51). Labrum in general is very similar to that of N. crocata and N.
pratensis (Fig. 52).
Thorax. In general is very similar to that of other Nephrotoma species.
Abdomen. In general is very similar to that of other Nephrotoma species. Seta D1 is very short; D2 is long and
Zootaxa 3764 (2) © 2014 Magnolia Press · 163TIPULIDAE EGG-LARVAE

six times as long as D1; D3 is the longest and eight times as long as D1 (Fig. 53); D4, D5, D6 are very similar in
length, three times as long as D1. Setae D2, D3 and D4, D5 make pairs and are very close to each other. Seta V1 is
very short; V2 is the longest ventral seta and is ten times as long as V1 (Fig. 54); V3 is eight time as long as V1; V4
is four times as long as V1. Setae V1, V2 and V3, V4 make pairs and are very close to each other. Lateral setae are
arranged in a line. Setae L1, L3 and L4 are very similar in length and are three times as long as L2 (Fig. 55). Seta
L4 is most distant from the other setae.
Spiracular disc. Spiracular field is surrounded by four flat and elongate round-tipped lobes (Fig. 56), which are
covered by sclerites. Lateral lobe is prominent, its length almost 1.5x the width at the base. It has four long stout
apical bristles. The length of each bristle is almost as long as one-fourth of the whole body. Two tufts, each
consisting of two bristles, are located on dorsal margin. Lobe bears pale, stripe-shaped sclerite. Ventral lobe is
almost as long as its width at the base. It has apical tuft of two bristles. Tuft of three long bristles is on inner margin;
tufts of two short and two long bristles are on outer margin; a tuft of four long bristles is at the base. Lobe bears
pale, wide leaf-shaped sclerite. Three tufts of long bristles are above the spiracles. The innermost tuft consists of
two bristles; others consist of three bristles. Spiracles are large, circular and close to each other. Distance between
them is smaller than diameter of a spiracle itself.
Anal field. Anus is surrounded by four long, cone-shaped, white and fleshy anal papillae. There are few long
setae on anal segment.
Specimens examined. 23 egg-larvae from female captured near Lazdijai, N54.24060, E23.51560, 23 VI 2011,
eggs were laid on 25 VI 2011, egg-larvae hatched on 14 VII 2011; 43 egg-larvae from female captured near
Lazdijai, N54.24067, E23.51526, 23 VI 2011, eggs were laid on 24 VI 2011, egg-larvae hatched on 09 VII 2011;
110 egg-larvae from female captured near Lazdijai, N54.24072, E23.51560, 23 VI 2011, eggs were laid on 25 VI
2011, egg-larvae hatched on 18 VII 2011.
Nephrotoma submaculosa Edwards, 1928
(Figs. 57–64)
First instar larvae. Length 1.59–1.60 mm; width 0.31–0.32 mm. Body is covered by pale pubescence. Cuticle is
transparent.
Head capsule. Length 0.37–0.38 mm, width 0.18–0.20 mm. In general it is very similar to that of other
Nephrotoma species. Minor differences were noticed in shape of hypostomium, mandible and antenna.
Hypostomium bears five sharp teeth. Three middle teeth are very similar is size and shape; the outermost teeth are
much smaller. There is a prominent incision on posterior part of hypostomium (Fig. 57). Mandible in general is
very similar to that of N. flavescens, but shapes of teeth are slightly different (Fig. 58). Antenna is very similar to
that of N. dorsalis, but shape of basal segment is different (Fig. 59). Labrum in general is very similar to that of N.
dorsalis, but arrangement of sensory structures is different (Fig. 60).
Thorax. In general is very similar to that of other Nephrotoma species.
Abdomen. In general is very similar to that of other Nephrotoma species. Seta D1 is very short; D2 is long and
four times as long as D1 (Fig. 61); D3 is slightly shorter than D2; D4 and D5 are very similar in length and slightly
longer than D1; D6 is the shortest seta and twice as short as D1. Setae D2, D3 and D4, D5 make pairs and are very
close to each other. Seta V1 is long; V2 is slightly longer than V1 and is the longest ventral seta (Fig. 62). Setae V3
and V4 are very similar in length and half the length of V1; V1, V2 and V3, V4 make pairs and are very close to
each other. Lateral setae are arranged in a line. Setae L1, L4 are very similar in length and three times as long as L2
(Fig. 63). Seta L3 is twice as long as L2; L4 is situated away from the other setae.
Spiracular disc. Spiracular field is surrounded by four almost equal, flat, elongate and round-tipped lobes (Fig.
64). Lobes are almost as long as their width at the base. Lateral lobe has two long stout apical bristles. Each bristle
is almost as long as one-third of the whole body. Base of the inner bristle extends into an irregular stripe-shaped
sclerite, which ends above the spiracle. A tuft of two long bristles and a tuft of three long bristles are situated above
that sclerite on dorsal part of the lobe. Above the spiracles there are three tufts, each consisting of three bristles of
different length. Ventral lobe has an apical tuft of four long bristles; three bristles, similar in length, are situated on
inner margin; a tuft of three short bristles is on outer margin; a tuft of two short bristles close to it; a tuft of four
long bristles is at the base. The lobe bears very pale oval-shaped sclerite. Spiracles are large, oval and close to each
other. Distance between them is the same as diameter of a spiracle itself.
PODENIENE ET AL.164 · Zootaxa 3764 (2) © 2014 Magnolia Press
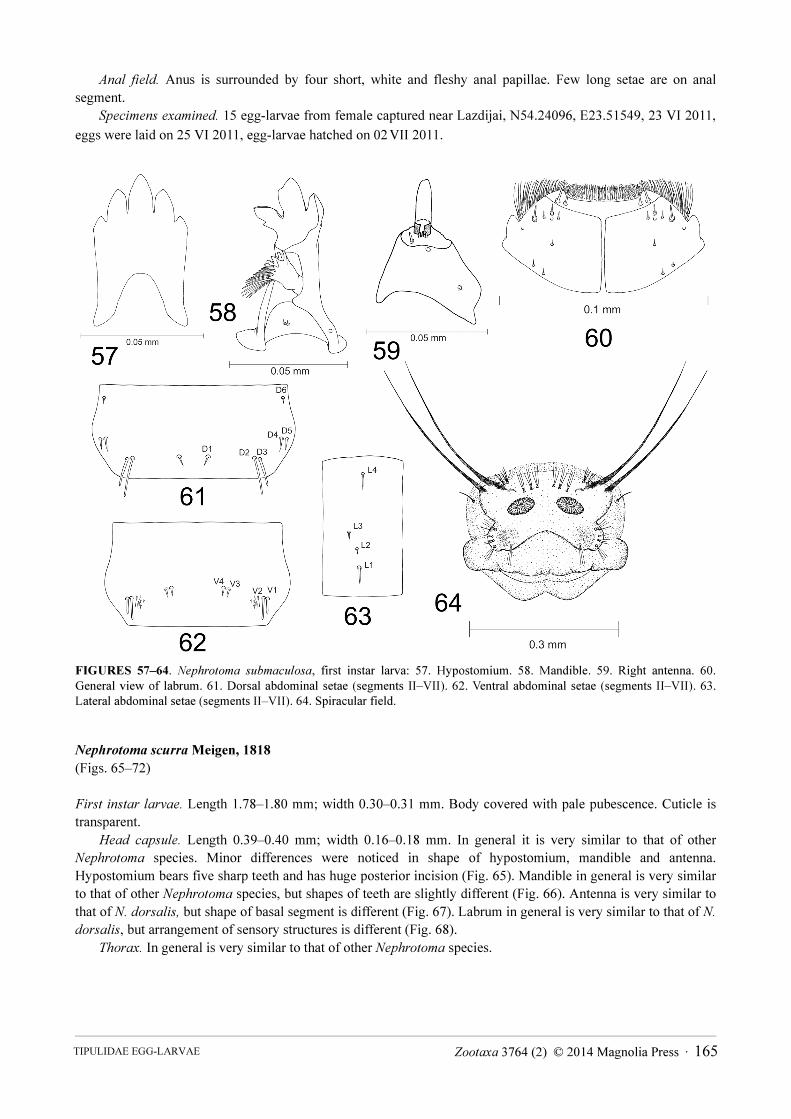
Anal field. Anus is surrounded by four short, white and fleshy anal papillae. Few long setae are on anal
segment.
Specimens examined. 15 egg-larvae from female captured near Lazdijai, N54.24096, E23.51549, 23 VI 2011,
eggs were laid on 25 VI 2011, egg-larvae hatched on 02 VII 2011.
FIGURES 57–64. Nephrotoma submaculosa, first instar larva: 57. Hypostomium. 58. Mandible. 59. Right antenna. 60.
General view of labrum. 61. Dorsal abdominal setae (segments II–VII). 62. Ventral abdominal setae (segments II–VII). 63.
Lateral abdominal setae (segments II–VII). 64. Spiracular field.
Nephrotoma scurra Meigen, 1818
(Figs. 65–72)
First instar larvae. Length 1.78–1.80 mm; width 0.30–0.31 mm. Body covered with pale pubescence. Cuticle is
transparent.
Head capsule. Length 0.39–0.40 mm; width 0.16–0.18 mm. In general it is very similar to that of other
Nephrotoma species. Minor differences were noticed in shape of hypostomium, mandible and antenna.
Hypostomium bears five sharp teeth and has huge posterior incision (Fig. 65). Mandible in general is very similar
to that of other Nephrotoma species, but shapes of teeth are slightly different (Fig. 66). Antenna is very similar to
that of N. dorsalis, but shape of basal segment is different (Fig. 67). Labrum in general is very similar to that of N.
dorsalis, but arrangement of sensory structures is different (Fig. 68).
Thorax. In general is very similar to that of other Nephrotoma species.
Zootaxa 3764 (2) © 2014 Magnolia Press · 165TIPULIDAE EGG-LARVAE
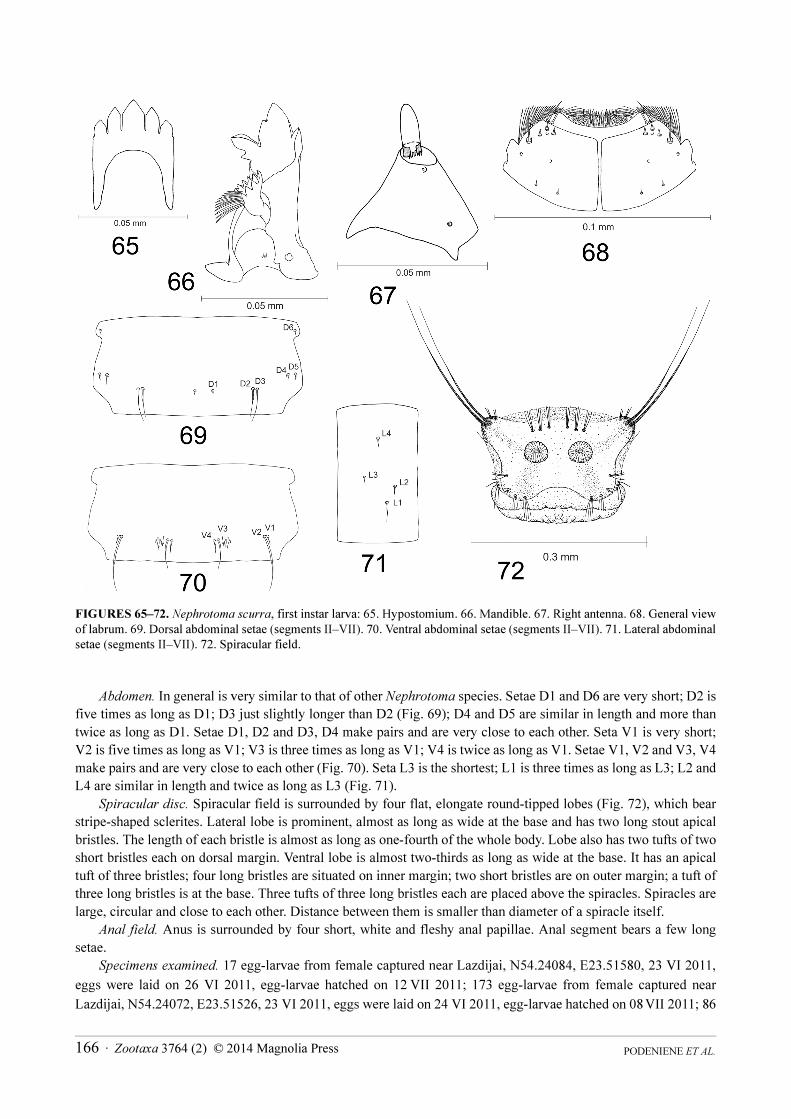
FIGURES 65–72. Nephrotoma scurra, first instar larva: 65. Hypostomium. 66. Mandible. 67. Right antenna. 68. General view
of labrum. 69. Dorsal abdominal setae (segments II–VII). 70. Ventral abdominal setae (segments II–VII). 71. Lateral abdominal
setae (segments II–VII). 72. Spiracular field.
Abdomen. In general is very similar to that of other Nephrotoma species. Setae D1 and D6 are very short; D2 is
five times as long as D1; D3 just slightly longer than D2 (Fig. 69); D4 and D5 are similar in length and more than
twice as long as D1. Setae D1, D2 and D3, D4 make pairs and are very close to each other. Seta V1 is very short;
V2 is five times as long as V1; V3 is three times as long as V1; V4 is twice as long as V1. Setae V1, V2 and V3, V4
make pairs and are very close to each other (Fig. 70). Seta L3 is the shortest; L1 is three times as long as L3; L2 and
L4 are similar in length and twice as long as L3 (Fig. 71).
Spiracular disc. Spiracular field is surrounded by four flat, elongate round-tipped lobes (Fig. 72), which bear
stripe-shaped sclerites. Lateral lobe is prominent, almost as long as wide at the base and has two long stout apical
bristles. The length of each bristle is almost as long as one-fourth of the whole body. Lobe also has two tufts of two
short bristles each on dorsal margin. Ventral lobe is almost two-thirds as long as wide at the base. It has an apical
tuft of three bristles; four long bristles are situated on inner margin; two short bristles are on outer margin; a tuft of
three long bristles is at the base. Three tufts of three long bristles each are placed above the spiracles. Spiracles are
large, circular and close to each other. Distance between them is smaller than diameter of a spiracle itself.
Anal field. Anus is surrounded by four short, white and fleshy anal papillae. Anal segment bears a few long
setae.
Specimens examined. 17 egg-larvae from female captured near Lazdijai, N54.24084, E23.51580, 23 VI 2011,
eggs were laid on 26 VI 2011, egg-larvae hatched on 12 VII 2011; 173 egg-larvae from female captured near
Lazdijai, N54.24072, E23.51526, 23 VI 2011, eggs were laid on 24 VI 2011, egg-larvae hatched on 08 VII 2011; 86
PODENIENE ET AL.166 · Zootaxa 3764 (2) © 2014 Magnolia Press

egg-larvae from female captured in Naujoji Vilnia environs, Vilnius district, N54.70365, E25.405980,
09 VIII 2011, eggs were laid on 13 VIII 2011, egg-larvae hatched on 21 VIII 2011; 118 egg-larvae from female
captured in Vilnius, N54.68194, E25.27087, 31 VII 2012, eggs were laid on 01 VIII 2012, egg-larvae hatched on
06 VIII 2012.
Discussion
Differences between the last instar larvae of genus Nephrotoma were noticed just in sclerotisation of spiracular
field and shape of anal papillae. Characters of head capsule of the last instar larvae have never been discussed or
used in any key. Arrangement and length of dorsal, ventral and lateral setae have been discussed by Chiswell
(1954), but have never been used in any key. During this study it was found that such characters as shape of apical
teeth of mandible, shape of basal segment of antenna, number of sensillae, shape of hypostomium and arrangement
of sensory structures on labrum differ among egg-larvae of genus Nephrotoma. The length of abdominal setae
differs significantly among egg-larvae of different species. The longest ventral and dorsal setae could be half the
length of the whole segment in N. crocata, N. dorsalis and N. scurra, or just as long as one-third of it in N.
flavescens, N. pratensis and N. submaculosa. Arrangement of dorsal setae is very similar in all Nephrotoma
species. In some species seta D2 is the longest dorsal seta (N. crocata, N. pratensis, N. submaculosa), when in other
species, the longest is D3 (N. dorsalis, N. flavescens, N. scurra). Ventral setae are arranged in same way in all
Nephrotoma species, but length of them is different in all species. Lateral setae could be arranged in two ways. In
some species, they all are forming a line, in others, setae L1–L3 are far apart from each other. The most significant
differences were observed in spiracular field. Number and length of lateral bristles, number of tufts, number of
bristles in tufts, number and length of apical bristles on ventral lobes and sclerotisation of spiracular field makes
spiracular field of each Nephrotoma species unique.
Egg-larvae, as well as other instar larvae of Nephrotoma, have pads on frontal part of prothorax. This character
was known as the only character separating larvae of that genus from the other genera. In egg-larvae pad is solid
and better expressed on dorsal and ventral parts of prothorax. In the last instar larvae there is a pair of pads on
dorsal side of prothorax. We found, that lateral plates of labrum of genus Nephrotoma differ significantly from that
of genus Ctenophora. This character could be used as genera separating feature as well.
During this study, we never noticed any protruding sclerotization on the dorsal side of head capsule, mentioned
by Hemmingsen (1965), which can be used as “hatching spine”. It was noticed, that egg-larvae eat their egg skins
and is quite possible, that they are using their huge mandible with sharp apical teeth for hatching.
Acknowledgements
We thank Inga Marciukaityte (Vilnius, Lithuania) for the excellent drawings. We are grateful to dr. Pjotr
Oosterbroek (Amsterdam, The Netherlands) and dr. Herman de Jong (Museum of Natural History, Leiden, The
Netherlands) for valuable information and advices. Our special thanks to dr. Gaile Zaludiene, Laboratory of
Bedrock Geology at Nature Research Centre, Lithuania, for the help when taking SEM pictures.
Literature cited
Chiswell, J.R. (1956) A taxonomic account of the last instar larvae of some British Tipulinae (Diptera: Tipulidae). Transactions
of the Royal Entomological Society of London, 108, 409–484.
http://dx.doi.org/10.1111/j.1365-2311.1956.tb01273.x
Edwards, F.W. (1928) The nematocerous Diptera of Corsica. Encyclopedie Entomologique, (B II), Diptera, 4,157–189.
Foote, B.A. (1963) Observations on the biology of Tipula footeana Alexander (Diptera: Tipulidae). Bulletin of the Brooklyn
Entomological Society, 58, 145–150.
Gelhaus, J.K. (1986) Larvae of the Crane fly genus Tipula in North America (Diptera: Tipulidae). University of Kansas Science
Bulletin, 53 (3), 121–182.
Gerbig, F. (1913) Uber Tipulidenlarven mit besonderer Berucksichtigung der Respirationsorgane. Zoologische Jahrbucher,
Abteilung fur Systematik, Geographie und Biologie der Tiere, 35, 127–181.
Zootaxa 3764 (2) © 2014 Magnolia Press · 167TIPULIDAE EGG-LARVAE

Hartman, M. & Hynes, C.D. (1982) The immature stages of Tipula simplex Doane and Tipula acuta Doane (Diptera:
Tipulidae). Pan-Pacific Entomologist, 58, 153–158.
Hemmingsen, A.M. (1965) The lotic cranefly, Tipula saginata Bergroth, and the adaptive radiation of the Tipulinae, with a test
of Dyars law. Videnskabelige Meddelelser fra Dansk Naturhistorisk Forening, 128, 93–150.
Hennig, W. (1950) Die Larvenformen der Dipteren. 2 Teil. Akademie–Verlag, Berlin, 458 pp.
Jong, W.H. de (1925) Een studie over emelten en hare bestrijding. Verslagen en Mededelingen van de Plantenziektekundige
Dienst. Wageningen, 42, 1–108.
Kim, D.S. & Lee, J.E. (2004) Immature stages of Tipula patagiata (Diptera, Tipulidae) from Korea. Korean Journal of Applied
Entomology, 43, 263–266.
Lantsov, V.I. (1982) Ecology and preimaginal developmental stages of the arctic crane-fly Tipula glaucocinerea (Diptera,
Tipulidae). Zoologicheskiy Zhurnal, 61, 1913–1916.
Lantsov, V.I. (1984) Ecology, morphology and taxonomy of arctic crane-flies of the genus Prionocera (Diptera, Tipulidae).
Zoologicheskiy Zhurnal, 63, 1196–1204.
Lantsov, V.I. (2009) The ecology, biology and larval instars of the North Caucasian population (Lake Maliy Tambukan) of
Tipula subcunctans Alexander, 1921 (Diptera: Tipulidae). In: Lantsov, V. (Ed.), Crane flies. History, taxonomy and
ecology (Diptera: Tipulidae, Limoniidae, Pediciidae, Trichoceridae, Ptychopteridae, Tanyderidae). Memorial volume
dedicated to Dr. Charles Paul Alexander (1889-1981), Dr. Bernhard Mannheims (1909-1971) and Dr. Evgeniy Nikolaevich
Savchenko (1909-1994). Zoosymposia, 3, 115–129.
Linnaeus, C. (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum caracteribus,
differentiis, synonymis, locis. Ed. 10. Salvii, Holmiae [= Stockholm], 1: i–iv, 1–824.
http://dx.doi.org/10.5962/bhl.title.542
Meigen, J.W. (1803) Versuch einer neuen Gattungs Eintheilung der europaischen zweiflugeligen Insecten. Magazin fur
Insektenkunde (Illiger), 2, 259–281.
Meigen, J.W. (1818) Systematische Beschreibung der bekannten europaischen zweiflugeligen Insekten. Aachen, 1, i–xxxvi, 1–
333.
http://dx.doi.org/10.5962/bhl.title.13731
Neugart, C., Schneeberg, K. & Beutel, R.G. (2009) The morphology of the larval head of Tipulidae (Diptera, Insecta), the
dipteran groundplan and evolutionary trends. Zoologischer Anzeiger, 248, 213–235.
http://dx.doi.org/10.1016/j.jcz.2009.10.001
Oosterbroek, P. (2013) Catalogue of the Craneflies of the World (Insecta, Diptera, Nematocera, Tipuloidea). Available from:
http://ip30.eti.uva.nl/ccw/ (accessed on 8 June 2013)
Petersen, M.J., Bertone, M.A., Wiegmann, B.M. & Courtney, G.W. (2010) Phylogenetic synthesis of morphological and
molecular data reveals new insights into the higher-level classification of Tipuloidea (Diptera). Systematic Entomology,
35, 526–545.
http://dx.doi.org/10.1111/j.1365-3113.2010.00524.x
Podeniene, V. (2001) Notes on the larvae of Lithuanian Tipulidae from the subgenera Tipula (Acutipula), T. (Beringotipula) and
T. (Platytipula). Ekologija, 2, 81–89.
Podeniene, V. (2003) Morphology and ecology of the last instar larvae of the crane flies (Diptera, Tipulomorpha) of Lithuania.
Doctoral dissertation, Vilnius University, Biomedical sciences, Zoology, 295 pp.
Pritchard, G. (1983) Biology of Tipulidae. Annual Review of Entomology, 28, 1–22.
Pritchard, G. & Hall, H.A. (1971) An introduction to the biology of craneflies in a series of abandoned beaver ponds, with an
account of the life cycle of Tipula sacra Alexander (Diptera; Tipulidae). Canadian Journal of Zoology, 49, 467–482.
http://dx.doi.org/10.1139/z71-072
Theowald, B. (1967) Familie Tipulidae, larven und Puppen. Bestimmungsbücher zur Bodenfauna Europas, 7, 1–100.
Savchenko, E.N. (1983) Crane-flies (Fam. Tipulidae), Introduction and the beginning of systematics part. Subfam.
Dolichopezinae, subfam. Tipulinae (start). Fauna USSR. Diptera, Leningrad “Nauka”, 2(1–2), 1–585.
Sorensen, L. (2002) Status of wood-living ctenophorine craneflies (Diptera: Tipulidae) in Denmarks old forests. Entomologiske
Meddelelser, 70, 129–142.
White, J.H. (1951) Observations on the life history and biology of Tipula lateralis Meig. Annals of Applied Biology, 38, 847–
858.
http://dx.doi.org/10.1111/j.1744-7348.1951.tb07855.x
PODENIENE ET AL.168 · Zootaxa 3764 (2) © 2014 Magnolia Press
Copyright © 2022 FDOKUMEN




















