
INCONSISTENCY IN THE ANALYSIS OF MORPHOLOGICAL DEFORMITIES IN CHIRONOMIDAE (INSECTA: DIPTERA) LARVAE
Upload
khangminh22Category
view
3download
0
Departement für integrative Biologie und
Biodiversitätsforschung
Institut für Zoologie
Blütenbesuchende Nutzinsekten (Diptera:
Syrphidae, Hymenoptera: Apis mellifera, Bombus
sp.) in Brachen unterschiedlichen Alters
Masterarbeit vorgelegt zu Erlangung des akademischen Grades
Diplom- Ingenieurin
an der Universität für Bodenkultur in Wien
Betreuer: Univ. Prof. Mag. Dr. Thomas Frank
Dr. Jörg- Alfred Salamon
Eingereicht von
Karin Kastner, Bakk. techn.
Wien, im Oktober 2010

Abstract
Flower visiting insects like honeybees, bumblebees and hoverflies are playing an important
role in our ecosystems. They are efficient pollinators of the different plants as well as of
economically interesting crops.
In our highly modified landscapes these insects are treated by many different impacts and
the number of honeybees and bumblebees is steadily decreasing.
These insects need highly structured landscapes for hiding or breeding and efficient flower
coverage for nutrition during the whole season. Most of these conditions can be provided in
seminatural habitats like fallow land, flowering strips or hedges.
In these thesis nine fallows of three different ages (young: 3-4 years, middle-old: 7-9 years
and old: 13-16 years) and their benefit for pollinators have been examined. Pollinator
abundance was measured by observing five 1x1 m2 Plots in each fallow. Each pollinator
visiting blooming plants within the quadrate has been counted. The observations took place
in May, June and July 2009 in an intensively agriculturally used area in Lower Austria, the
Marchfeld. The possible impacts of age, local factors like flower coverage, plant species
richness, the colours of the florescenses or weather conditions as well as the influence of
landscape composition in an area of 500 m around the fallows on pollinators have been
investigated.
The data obtained were interpreted with 2-factorial ANOVA, canonical correspondence
analysis (CCA), single regressions and stepwise multiple regressions with backward
selection method.
In June, middle-old fallows contained significantly less honeybees, hoverflies and
bumblebees than young and old fallows. In July, the densities of honeybees and bumblebees
were significantly lower in middle-old fallows. In all three month the numbers of flowering
plants were significantly lowest in middle-old fallows and the highest numbers of blooming
plants have been in the old fallows. Parameters within the studied fallows like flower
coverage and number of plant species were most important for the abundance of hoverflies
and bumblebees. Additionally, the influence of the surrounding landscape parameters
grassland and fields were significant for the density of hoverflies. The local weather
parameters like wind or temperature were only for the distribution of bumblebees important.
Keywords: pollinators, visitation, honeybees, hoverflies, bumblebees, fallow land, age
effects, sucession.

I
Inhaltsverzeichnis
1 Einleitung 1
2 Biologie der untersuchten Nützlingsinsekten 3
2.1 Honigbienen 3
2.2 Hummeln 4
2.3 Schwebfliegen 6
3 Untersuchungsgebiet 7
4 Material und Methodik 9
4.1 Vorbereitende Arbeiten im Freiland 9
4.2 Erstellen des Aufnahmeblattes 10
4.3 Tierbeobachtung 10
4.4 Statistik 12
4.4.1 Zweifaktorielle Varianzanalyse 13
4.4.2 Ordinationsverfahren 13
4.4.3 Regressionsanalysen 14
5 Ergebnisse 14
5.1 Einfluss des Alters der Brachen auf die Blütenbesuche: Monat Mai 15
5.2 Einfluss des Alters der Brachen auf die Blütenbesuche: Monat Juni 16
5.3 Einfluss des Alters der Brachen auf die Blütenbesuche: Monat Juli 17
5.4 Aktivitätsdichte unter Einfluss von Flächenparametern 19
5.4.1 Ordination im Juni 19
5.4.2 Ordination im Juli 22
5.5 Aktivitätsdichte unter Einfluss von Flächen- und Umgebungsparametern 23
5.5.1 Honigbienen 23
5.5.2 Schwebfliegen 24
5.5.3 Hummeln 25
5.6 Blühpflanzenanzahl 26
6 Diskussion 28
6.1 Sekundärsukzession auf Brachen: Änderung der Pflanzengesellschaft 28
6.2 Der Einfluss des Alters der Brache auf die Bestäuber 30
6.2.1 Der Einfluss von Blütenfarben auf die Bestäuber 32
6.2.2 Der Einfluss von Blühpflanzenanzahl und Blütendeckung auf die Bestäuber 34
6.2.3 Der Einfluss der strukturellen Ausstattung des Lebensraumes 38
7 Zusammenfassung 40
8 Literaturangaben 42
9 Danksagung 58

II
Abbildungsverzeichnis
Abb. 1: Apis mellifera (Honigbiene) ........................................................................................4
Abb. 2: Pyrobombus lapidarius (Steinhummel).......................................................................5
Abb. 3: Episyrphus balteatus, Sphaerophoria sp., Eristalis sp. ...............................................7
Abb. 4: Vorbereitende Arbeiten auf den Brachen. ..................................................................9
Abb. 5: Die fünf Plots innerhalb einer Brache. ......................................................................10
Abb. 6: Der Einfluss des Alters im Mai. ................................................................................16
Abb. 7: Der Einfluss des Alters im Juni.................................................................................17
Abb. 8: Der Einfluss des Alters im Juli..................................................................................18
Abb. 9: CCA Grafik für Juni ..................................................................................................21
Abb. 10: CCA Grafik für Juli .................................................................................................23
Abb. 11: Anzahl von Blühpflanzen in den drei Altersklassen der Brachen in den Monaten Juni
und Juli.................................................................................................................................27

III
Tabellenverzeichnis
Tab. 1: Die neun untersuchten Brachen: ................................................................................8
Tab. 2: F- Werte einer zweifaktoriellen Varianzanalyse (ANOVA): Mai.................................15
Tab. 3: F- Werte einer zweifaktoriellen Varianzanalyse (ANOVA): Juni. ...............................16
Tab. 4: F- Werte einer zweifaktoriellen Varianzanalyse (ANOVA): Juli. ................................18
Tab. 5: Einfache und multiple Regression für Honigbienen (A. mellifera): ............................24
Tab. 6: Einfache und multiple Regression für Schwebfliegen (Hoverflies): ...........................25
Tab. 7: Einfache und multiple Regression für Hummeln (Bumblebees): ...............................26
Tab. 8: F- Werte einer zweifachen Varianzanalyse (ANOVA): Blühpflanzenanzahl. .............27

1
1 Einleitung
Insekten wie Bienen, Schwebfliegen und Hummeln üben wichtige Funktionen in der
Natur- und Kulturlandschaft aus. Bienen und Hummeln zählen zur Bestäubergemeinschaft
und sind von hoher agrarökonomischer Bedeutung (KEVAN 1999). Ebensolche Funktionen
üben adulte Schwebfliegen aus, wobei sich die Larven einiger dieser Arten von
Blattläusen ernähren, weshalb sie in der biologischen Schädlingsbekämpfung
unverzichtbar sind (KRIEG & FRANZ 1989, MEYER et al. 2009). Diese Insekten können des
Weiteren als Bioindikatoren für ein Monitoring des Zustandes einer Agrarlandschaft
herangezogen werden. Als Reaktionen auf Intensivlandwirtschaft mit Einsatz von
Insektiziden und Herbiziden sowie dem Einbringen von Krankheiten (wie die Verroa Milbe)
oder Prädatoren in ein Ökosystem, kann es zu einem Rückgang an Diversität und Anzahl
dieser Bestäuber kommen (KEVAN 1999). Flurbereinigungen führten zu nachhaltigen
Veränderungen der ursprünglichen, räumlichen Strukturen und neben dem Verlust an
Habitaten auch zu Änderungen von lokalen Wind- oder Klimabedingungen (HENDRICKX et
al. 2007).
In den 1960er und 1970er Jahren führten Maßnahmen zur Steigerung der Flächenerträge
zu einer zunehmenden Verarmung und Vereinheitlichung der Segetalflora (auch
Ackerbegleitflora oder Ackerunkrautflora) (TRAXLER et al. 2005). Dies wiederum reduzierte
neben dem Nahrungsangebot auch die Brutmöglichkeiten für Nützlinge wie Hummeln
oder Schwebfliegen (WESTRICH 1997). Viele dieser Pflanzen (z.B. Agrostemma githago,
Capsella bursa-pastoris, Echium vulgare, Leucanthemum vulgare, Senecio vulgaris,
Ranunculus avensis, siehe auch Anhang III) stellen wichtige Trachtpflanzen dar,
besonders in nahrungsarmen Vegetationszeiten (TRAXLER et al. 2005). Beim Wegfall
dieser Pflanzen sind adulte Schwebfliegen, Hummeln und Honigbienen von
Nahrungsmangel betroffen. Ein genereller Rückgang der Anzahl an Nützlingen ist die
Folge (WEISS & STETTMER 1991).
Durch die Gemeinsame Agrarpolitik (GAP) der Staaten der Europäischen Union wurden
seit 1988/89 freiwillige Flächenstilllegungen als marktpolitische Maßnahme zur Reduktion
der landwirtschaftlichen Überproduktion eingeführt. Seit 1992 sind solche Stilllegungen
aus sozioökonomischen (subventionierte Brache) oder auch ökologischen Gründen
(Ökobrache) obligatorisch, der Ertragsentgang wird finanziell über Ausgleichszahlungen
abgegolten. Wurden ab Mitte des 20. Jahrhunderts noch brachliegende Flächen für eine
Nutzung adaptiert, bedeutete die Stilllegung das genaue Gegenteil (GREILER 1994).
Intensivackerland und Grünland wurden aus der Nutzung genommen, ein Prozess der

2
beabsichtigten, kurz- bis langfristigen Verbrachung setzte ein.
Besonders aus agrarökologischer Sicht kann diese Entwicklung positiv gesehen werden
da Brachen düngemittel- und pestizidfreie Rückzugsflächen bieten und Störungen im
Sinne von landwirtschaftlichen Tätigkeiten deutlich abnehmen (HOLZNER 2001). Besonders
Nutzinsekten profitieren von dieser Entwicklung, kann doch der Nahrungsbedarf auf den
oft blütenreichen und störungsarmen Ausgleichsflächen gedeckt werden (WEISS &
STETTMER 1991).
Die im Zuge der Sekundärsukzession auf Acker- oder Grünlandbrachen entstehenden
Änderungen der Pflanzengesellschaften und die möglichen Auswirkungen auf
blütenbesuchende Nützlingsinsekten wurden in der vorliegenden Arbeit untersucht.
Dabei stand der Vergleich zwischen jungen, mittleren und alten Brachen, die teilweise
selbstbegrünt, teilweise auch eingesät sind, im Mittelpunkt. Die dort vorhandenen
Blühpflanzen und deren Nutzung als Nektar- und Pollenquelle für Honigbienen,
Schwebfliegen und Hummeln wurden untersucht. Die Fragestellungen hierzu lauten:
1. Unterscheiden sich die drei Brachenaltersklassen hinsichtlich der Abundanz der
blütenbesuchenden Insekten signifikant voneinander?
Kommt es mit zunehmendem Alter und damit verbundenen Brachesukzessions- und
Alterungsprozessen zu einer Abnahme an Blühpflanzen und somit zu einer signifikanten
Reduktion der Abundanzen der Nützlinge?
Um den Kleinstandort (microscale, 1x1 Meter) und die unmittelbaren Lebensbedingungen
auf der jeweilig untersuchten Fläche zu charakterisieren, wurden neben biotischen
Faktoren wie Blütendeckung, Blühpflanzenzahl und Blütenfarben auch abiotisch relevante
Daten aufgenommen, die das Mikroklima beschreiben (Temperatur,
Windgeschwindigkeit). Diese Faktoren können das Vorhandensein von Insekten auf den
Brachen deutlich positiv oder auch negativ beeinflussen. In der Literatur mehrfach
nachgewiesene Präferenzen der Nützlinge für einzelne Blütenfarben (Schwebfliegen
präferieren häufig die Farbe Gelb) (RÖDER 1990) oder Pflanzenarten (langrüsselige
Hummelarten bevorzugen Klee oder Disteln) (VON HAGEN 1988) sollten sich auch auf den
betrachteten Brachen wiederfinden.
2. Wie stark beeinflussen kleinstandörtliche Faktoren auf den drei
Brachenaltersklassen das Vorhandensein welcher Tiere?
Da sich die betrachteten Flächen alle in einem agrarisch sehr intensiv genutzten
Landschaftsteil östlich von Wien befinden und die betrachteten Tiere flugfähig sind,
wurden in diesem Zusammenhang nicht nur die lokalen Lebensbedingungen auf, sondern
auch das unmittelbare Umfeld um die jeweiligen Brachen in die Untersuchung mit

3
aufgenommen.
3. Beeinflusst die umgebende Landschaftsstruktur die Abundanzen der Nutzinsekten
in den drei Altersklassen der Brachen nachweislich?
Generell sind hier Unterschiede und auch eine unterschiedlich starke Einflussnahme
zwischen ackerbaulich dominierten Flächen, Siedlungsgebieten und Grünland oder
Weinkulturen auf die Anzahl der Nützlinge zu erwarten.
Um die genannten Hypothesen zu überprüfen, wurde in der vorliegenden Arbeit eine reine
Beobachtung der Blütenbesuchsfrequenz der Nützlinge durchgeführt. Auf das Fangen der
Tiere wurde bewusst verzichtet.
2 Biologie der untersuchten Nützlingsinsekten
Bestäuber wie Honigbienen, Hummeln, Solitärbienen und andere Hymenopteren sowie
blütenbesuchende Dipteren wie Schwebfliegen bestäuben rund 80% der bekannten
Pflanzen (TACK 1983). Rund 150 Pflanzenarten werden derzeit in Europa kultiviert, rund
70 Arten davon sind auf die Aktivität von Bestäubern angewiesen (TACK 1983, VON HAGEN
1988). Im Gegensatz zur Anemogamie (Windbestäubung) wird durch die Zoogamie
(Tierbestäubung) eine zielgerichtete Übertragung des Pollens auf die Narbe sichergestellt.
Eine Co- evolutionäre Entwicklung zwischen den Insekten und den zu bestäubenden
Pflanzen lässt sich anhand bestimmter blütenmorphologischer und
bestäuberanatomischer Anpassungen deutlich erkennen (FISCHER et al. 2008). Nicht jede
bestäubte Pflanzenart ist für den Menschen von ökonomischer Bedeutung. Viele werden
als Konkurrenzpflanzen in den landwirtschaftlichen Kulturen bekämpft, sind jedoch als
potentielle Nahrungsquelle für die Bestäuber von großer Wichtigkeit (WESTRICH 1997,
DECOURTYE et al. 2007).
Der Einsatz von Herbiziden, Maßnahmen zur Flurbereinigung oder die Saatgutreinigung
führten zu einem zunehmenden Verschwinden von Ackerbeikräutern oder blütenreichen
Randzonen und somit zu einer Reduktion möglicher Trachtpflanzen. Die Verfügbarkeit
von Nahrungspflanzen während der gesamten Vegetationsperiode ist in
agrarwirtschaftlich dominierten Landschaften nicht mehr optimal gewährleistet (KEVAN
1999).
2.1 Honigbienen
Apis mellifera, die Westliche Honigbiene aus der Familie der Apidae, zählt zu den
bedeutendsten und bekanntesten Bestäubern in der Kulturlandschaft (Abb. 1) (KEVAN

4
1999). Rund 80% der fremdbestäubten Kulturarten werden von Bienen (Kultur,- und
Wildformen) bestäubt (REICHHOFF- RIEHM 1984).
Die Außentemperatur für Flug- und Sammelaktivität muss mindesten 10°C betragen, die
optimale Flugtemperatur liegt zwischen 22 und 25°C (RUTTNER 2003). Seh- und
Geruchsinn sind bei Bienen gut ausgebildet. Aufgrund ihres trichromatischen
Sehvermögens nehmen sie die Farbe rot (630 – 700nm Wellenlänge) nicht wahr,
verfügen aber neben Gelb- und Blausehen über einen Sehbereich im ultravioletten
Farbspektrum (400 - 450nm Wellenlänge) (RUTTNER 2003).
Abb. 1: Apis mellifera (Honigbiene) auf einem Natternkopf (Echium vulgare). (© Kastner)
Neben bekannten Krankheiten durch Bakterien und Viren, Parasiten (Milben wie Varroa
destructor) oder Räubern (Coleoptera, Arachnida, Diptera, Hymenoptera) sind
Honigbienen vor allem durch Insektizide oder Saatgutbeizmittel (z.B. Neonicotinoide)
bedroht (KEVAN 1999, DECOURTYE et al. 2007).
Die Wirtschaftsleistung der Honigbiene wird auf rund 2,9 Milliarden Euro geschätzt. Sie
zählt neben Rindern und Schweinen zu einem der wichtigsten Nutztiere (RUTTNER 2003;
UMWELTSTIFTUNG WWF 2010)
2.2 Hummeln
Sie gehören zur Familie der echten Bienen (Apidae), Gattung Hummeln (Bombus).
Weltweit sind derzeit 250 Hummelarten bekannt, in Europa existieren rund 30 Arten (VON
HAGEN 1988). Ihr Verbreitungsgebiet sind die gemäßigten Klimazonen nördlicher Breiten,
ein gänzliches Fehlen von autochthonen Hummeln ist in Afrika und Australien zu
bemerken (VON HAGEN 1988).
Die bevorzugten Lebensräume sind halboffenes, strukturreiches Gelände, nur wenige

5
Arten siedeln in geschlossenen Waldbiotopen (RÖSELER 2001). Sie verfügen über eine
kräftige Flugmuskulatur und können auch bei niedrigen Temperaturen (ab +5°C), bei
nassem Wetter oder Wind fliegen. Ihre Sammelaktivität ist sehr effizient, sie besuchen in
der Regel im selben Zeitraum bis zu dreimal so viele Blüten wie die Honigbienen (VON
HAGEN 1988, RÖSELER 2001). Aufgrund dieser Bestäubungsleistung werden Hummelvölker
immer häufiger für Gemüsekulturen in Glashäusern (zur Bestäubung von Tomaten oder
Paprika) (KEVAN 1999) oder in Weinbaugebieten eingesetzt (RÖSELER 2001).
Durch ihre Größe, die deutliche Behaarung und die charakteristischen Färbungen an
Thorax und Abdomen sind viele Arten gut zu erkennen (Abb. 2). In dieser Arbeit wurden
Farbtafeln, Fotografien und Zeichnungen zur Bestimmung der Tiere herangezogen (VON
HAGEN 1988).
Abb. 2: Pyrobombus lapidarius (Steinhummel) auf einer Ackerkratzdistel (Cirsium arvense). (© Kastner)
Hummeln sind wichtige Bestäuber in der Kulturlandschaft, deren Bedeutung oftmals
unterschätzt oder verkannt wird (FUSSELL & CORBET 1992). Die effiziente Bestäubung von
Pflanzen mit langen Kronröhren wie Wicken, Ackerbohnen, Raps, Ölrettich,
Sonnenblumen, Buchweizen oder Kleesaaten wird vorwiegend von langrüsseligen
Hummelarten übernommen (B. hortorum, B. ruderatus, B. pomorum, B. distinguendus, B.
subterraneus, B. sylvarum, B. pascuorum, B. ruderarius, B. humilis, B. muscorum). Der
Sammelfleiß der kurzrüsseligen Arten (B. terrestris, B. lucorum, B. hypnorum, B.
pratorum, B. lapidarius, B. soroensis, B. wurfleini) ist gleich hoch (VON HAGEN 1988,
RÖSELER 2001).
Ackerbrachen bieten für Hummeln nicht nur Nahrungsangebot, sondern auch weitgehend
störungsfreie Brutmöglichkeiten. Je älter eine Brache, desto länger liegen mögliche

6
Störungen wie ein Umbruch der Erdscholle zurück. Dies erhöht das Angebot an idealen
Nistmöglichkeiten in Form von leeren Erdbauten wie zum Beispiel verlassenen
Mäusenestern (VON HAGEN 1988). Auch dichte, horstbildende Gräser wie der Glatthafer
(Arrhenatherum elatius), ein typischer Brachezeiger, der mit steigendem Alter der Flächen
zunimmt und sehr konkurrenzstark werden kann, dient den Hummeln als Versteck für die
Nester (VON HAGEN 1988).
2.3 Schwebfliegen
Mit rund 8000 bekannten Arten weltweit und knapp 800 beschriebenen Arten europaweit
sind Syrphiden eine der größten Dipterenfamilien. Die Imagines sind Blütenbesucher, die
Larven sind in unterschiedliche Ernährungstypen klassifizierbar (RÖDER 1990). Die
bedeutendste Gruppe (ca. 37% der Arten) sind die zoophag lebenden Larven. Sie
ernähren sich meist aphidivor, also von Blattläusen (RÖDER 1990).
Aufgrund einer breiten ökologischen Amplitude, ihrer großen Verbreitung, ihres
charakteristischen Aussehens und ihrer leichten Bestimmbarkeit im Feld wurden eine Art
sowie zwei Gattungen in die vorliegende Arbeit aufgenommen:
Episyrphus balteatus: Charakteristisch ist die Wespenmimikry mit typischer gelb und
schwarz gestreifter Warntracht am deutlich segmentierten Abdomen (RÖDER 1990). Die
zwei gut sichtbaren, gelben Binden am 2. und 4. Abdominalsegment machen sie für eine
Sichtbestimmung im Feld gut geeignet (Abb. 3).
Dies ist eine sehr weit verbreitete (eurytope) Art mit großer Klimatoleranz und
ausgedehntem Flugoptimum (zwischen 18°C und 27°C). Die Individuen kennzeichnen
sich durch hohe Futtertoleranz und nutzen das vorhandene Blütenspektrum gleichmäßig
(euryanth). Die Larven sind polyphag an verschiedenen Blattläusen und gelten als sehr
gefräßig und aggressiv (bei Nahrungsmangel auch als kannibalistisch beschrieben)
(RÖDER 1990). Hinsichtlich der Abundanz ist sie die am häufigsten auftretende
Schwebfliegenart in Europa (RÖDER 1990).
Gattung Sphaerophoria sp.: Auch unter Stift,- Schmalbauch-, oder
Langbauchschwebfliegen beschrieben. Aufgrund ihres Habitus sind sie von anderen
Gattungen gut zu unterscheiden. Die Fliegen kennzeichnet das sehr schlanke,
langgestreckte, gelb - schwarz gefärbte Abdomen (Abb. 3).
Es ist eine weit verbreitete, wanderfreudige und trockenliebende Gattung der offenen
Landschaften. Die Larven zeigen ein breites Beutespektrum (zoophag, aphidivor) (RÖDER
1990).

7
Gattung Eristalis: Eine artenreiche Gattung, welche im Habitus den Bienen sehr ähnlich
ist. Markant ist ihre Abdominalzeichnung, die durch keilförmige helle oder auch dunkle
Flecken auf den ersten Segmenten auffällt (Abb. 3). In der Literatur werden sie auch als
„Scheinbienen“, „Mistbienen“ oder „Schlammfliegen“ aufgeführt. Adulte Tiere sind als
eurytop und euryanth zu bezeichnen (RÖDER 1990).
Abb. 3: Episyrphus balteatus, Sphaerophoria sp., Eristalis sp. (© Kastner)
Syrphiden haben wie die Honigbienen ein trichromatisches Farbsehen, sie verfügen über
Rezeptoren für die Lichtbereiche gelb, blau und im ultravioletten Bereich. Rote
Blütenfarben werden im Allgemeinen nicht wahrgenommen, häufig besteht eine Präferenz
zu gelben oder auch weißen Blüten. Auch in der vorliegenden Arbeit wurden die
Blühpflanzen auf den Brachen in vier Farbkategorien (rot, gelb, weiß, blau) eingeteilt, um
einen möglichen Einfluss der Blütenfarben auf die Insekten zu untersuchen.
Auf eine Bestimmung bis auf Artenniveau wurde bewusst verzichtet, da der Lebensraum
Brache und die Nutzung durch blütenbesuchende Insekten im Mittelpunkt des Interesses
stand, nicht jedoch auf artspezifische Eigenheiten eingegangen werden sollte. Der
markante, gut differenzierbare Körperbau der beschriebenen Tiere machte ein relativ
einfaches und sicheres Zählen auf den Brachen möglich.
3 Untersuchungsgebiet
In dieser Arbeit wurden neun unterschiedliche Bracheflächen untersucht. Sie befinden
sich im Marchfeld, einer Region im Osten Niederösterreichs.
Geologisch gehört dieses Gebiet zum Wiener Becken, welches im Süden durch die
Donau, im Norden durch das Weinviertler Hügelland sowie dem Höhenzug Bisamberg
und im Osten willkürlich durch die Staatsgrenze bzw. durch den namensgebenden Fluss,

8
die March, begrenzt wird. (HADATSCH et al. 2000).
Das Untersuchungsgebiet ist gekennzeichnet durch intensive, landwirtschaftliche Nutzung
mit überwiegend konventioneller Bewirtschaftungsweise. Rund 81% der
Produktionsfläche entfallen auf ackerbauliche Nutzung, wobei vorwiegend Getreide,
Gemüse, Zuckerrübe, Kartoffel, Raps oder Wein produziert werden (HADATSCH et al.
2000). Das Marchfeld gilt als strukturarme, intensiv genutzte Agrarlandschaft (HADATSCH et
al. 2000).
Die Jahresdurchschnittstemperaturen der Klimanormalperiode der Jahre 1971 bis 2000
liegen bei 9,8°C (Klimadaten der Zentralanstalt für Meteorologie, 2009). Die
durchschnittlichen Tagestemperaturen in der Vegetationsperiode von April bis Oktober
schwanken zwischen 20,5°C und 20,7°C, mit einem Maximum von 24°C im Juli. Der
mittlere Jahresniederschlag ist gering und beläuft sich auf 500 bis 600 mm (HARFLINGER
1993). Besonders in den Sommermonaten Juni, Juli und August kann es zu lang
anhaltenden Trockenperioden kommen. Die Bodenwasserbilanz im Jahre 2002 zeigte ein
Defizit von 40 bis 50 mm in den Monaten April bis September auf (STENITZER & HOESCH
2002). Eine künstliche Beregnung der Feldkulturen zur Ertragssicherung ist somit oftmals
unumgänglich.
Die in der vorliegenden Arbeit untersuchten Brachen befinden sich auf neun
verschiedenen Standorten (sites) über das Marchfeld verteilt (Tab. 1, Anhang I).
Tab. 1: Die neun untersuchten Brachen: Brachen („sites“) im Marchfeld mit Lokalisation, Namensgebung und Altersklassifizierung in Jahren sowie die im weiteren Text verwendeten Abkürzungen. (y: jung, m: mittelalt, o: alt)
9 Sites Name / Fallow code age years Schönfeld Lassee SFL y 3 Gänserndorf GD y 3 Gerasdorf GED y 3 Schönfeld Lassee SFL m 7-9 Bockfließ BF m 7-9 Untersiebenbrunn USB m 7-9 Gänserndorf GD o 13-16 Groß Engersdorf GGD o 13-16 Groß Enzersdorf GZD o 13-16

9
4 Material und Methodik
4.1 Vorbereitende Arbeiten im Freiland
Die Freilandarbeiten fanden in den Monaten April, Mai, Juni und Juli der
Vegetationsperiode 2009 statt.
Eine Ausdehnung des Untersuchungszeitraumes war nicht möglich, da die Flächen
einmal pro Jahr gemäht werden. Die Mähtermine wurden von den jeweiligen Besitzern
festgelegt, der ungefähre Zeitpunkt konnte durch Telefonate erfragt werden. In zwei
Fällen konnte ein früher Mähtermin verschoben werden, um einen vollständigen
Beobachtungszyklus auch im Juli zu gewährleisten.
Bereits im April wurden die einzelnen Brachen aufgesucht und mit dem Abstecken der
Untersuchungsflächen innerhalb der jeweiligen Brache begonnen. Mit Hilfe eines
vorgefertigten Kartierungsrahmens wurden pro Brache fünf jeweils 1x1 m große
Parzellen, sogenannte Plots, mit einfachen Bambusstäben abgesteckt. Die Fläche wurde
vorher grob vermessen, um die Plots entlang einer Geraden innerhalb der Brache optimal
zu positionieren (Abb. 4). Dies war nötig, da die Randzonen der unbewirtschafteten
Flächen von den Landwirten oftmals zum Wenden ihrer Maschinen und Traktoren
verwendet wurden bzw. sonstige Störungen im Zuge von Arbeiten auf den an die Brache
angrenzenden Äckern auftreten konnten. Die Anlage der Plots innerhalb der Brache
gewährleistete somit einen ungestörten Verbleib der Stäbe während der gesamten
Beobachtungszeit und eine übersichtliche Aufnahme der gesamten Fläche (Abb. 5).
Abb. 4: Vorbereitende Arbeiten auf den Brachen. Das Abmessen der Fläche sowie das Abstecken der Beobachtungsfelder im April mit Hilfe eines vorgefertigten 1x1 m Rahmens. (© Kastner)

10
Abb. 5: Die fünf Plots innerhalb einer Brache. Fertig abgesteckte Beobachtungsplots in der jungen Brache in Gerasdorf, Kapellerfeld, Mitte April. (© Kastner)
4.2 Erstellen des Aufnahmeblattes
Als weitere, vorbereitende Tätigkeiten wurde ein Aufnahmeblatt erstellt (Anhang II).
Neben den zu beobachtenden Tieren wurden bei jedem einzelnen Durchgang folgende
Daten flächenspezifisch erhoben:
§ Anzahl und Art der jeweils blühenden und somit für die Tiere attraktiven Pflanzen
§ Blütendeckungsgrad in Prozent auf den einzelnen Plots.
§ Temperatur
§ Windgeschwindigkeit
Die Aufzeichnung von Temperatur und Windgeschwindigkeit während des
Beobachtungszeitraumes erfolgte mit Hilfe eines Anemometers (testo 410- 1 ©, 0,4 bis 20
m/s, mit Temperaturmessung).
4.3 Tierbeobachtung
Die Tierbeobachtungen auf den neun Brachen fanden in den Monaten Mai, Juni und Juli
statt.
Um möglichst umfassende Daten zu gewinnen, wurden für die eigentliche
Beobachtungstätigkeit drei Zeitfenster über den Tag verteilt ausgewählt:

11
§ Vormittag (09:00 Uhr bis 11:00 Uhr)
§ Früher Nachmittag (13:00 Uhr bis 15:00 Uhr)
§ Später Nachmittag (15:00 Uhr bis 17:00 Uhr)
Jede der Brachen wurde an allen drei Zeitfenstern und in jedem der drei Monate besucht.
Die Reihenfolge der Plots blieb stets gleich, die Abfolge der Brachen wurde nach dem
Zufallsprinzip variiert, um eine qualitative, statistische Auswertung der Daten zu erhalten.
Diese Vorgehensweise hatte folgende Gründe: Viele Pflanzen bieten Nektar und Pollen
vor allem in den Morgenstunden und vormittags an (BARTH 1991). Auch wenn die
Tagestemperatur besonders im Monat Mai um diese Zeit noch eher niedrig ist, ist diese
Tageszeit aufgrund des bestehenden Nahrungsmaximums für die Tiere von großer
Wichtigkeit (RUPPERT 1993). Um einen großen Flüssigkeits- und Energieverlust zu
vermeiden, suchen die meisten Insekten während großer Mittagshitze geschützte,
schattige Habitate in der Krautschicht auf. Aus diesem Grund erfolgten keine
Tierbeobachtungen während der Mittagsstunden. Auch bei Schlechtwetter, Regen oder
Sturm wurde auf die Beobachtung verzichtet, da keine Nahrungssuche durch die Tiere
erfolgt.
Nach vorsichtiger Annäherung an die Beobachtungsfläche und Platznehmen auf einem
Campingstuhl im Abstand von rund 1 m zum Plot, wurde eine kurze Wartezeit von fünf
Minuten eingehalten, bevor mit der eigentlichen Beobachtung begonnen wurde. Dies
erschien wichtig, da durch die Annäherung an die Plots Insekten bei der
Nahrungsaufnahme gestört oder verscheucht werden können. Durch das Setzen auf den
Klappstuhl, das Tragen von unauffälliger Kleidung und das Einhalten des Abstandes
konnten etwaige Beeinträchtigungen reduziert werden. Auch wurde darauf geachtet,
keinen Körperschatten auf den Beobachtungsplot zu werfen.
Innerhalb dieser fünf Minuten wurde die Fläche betrachtet und folgende
Flächenparameter (im folgenden Text als FP bezeichnet) festgehalten:
§ Blütendeckungsgrad (Abschätzen des Deckungsgrades in % nach der Methodik
von Braun- Blanquet)
§ Anzahl und Art der aktuell blühenden Pflanzen
§ Blütenfarben in vier Farbkategorien (rot, gelb, weiß, blau)
Des Weiteren wurden ab dem ersten Plot mit den Messungen für Temperatur und
Windgeschwindigkeit begonnen, welche über den gesamten Beobachtungszeitraum
ermittelt und als Mittelwert notiert wurden.
Die Beobachtungsdauer pro Plot wurde auf 15 Minuten festgelegt. Ähnliche

12
Untersuchungen von RUPPERT (1993) ergaben, dass dieser Zeitrahmen durchaus
repräsentative Informationen über die Besuchsfrequenz an den Blüten lieferte. Die
absolute Blütenbeobachtungszeit pro Brache und Zeitfenster lag somit bei 75 Minuten. In
dieser Zeit wurden die Blütenbesucher qualitativ und quantitativ erfasst. Der
Gesamtzeitaufwand über die drei Monate belief sich auf rund 100 Stunden reine
Betrachtungszeit.
Es ist festzuhalten, dass es mitunter zu Mehrfachsichtungen kam, da die Tiere während
des Zeitraumes stellenweise mehrfach auf der Fläche aus- und einflogen. Verblieben sie
im Plot und nahmen sie Nahrung auf, wurden sie gezählt. Weiters ist zu betonen, dass mit
dieser Methodik die Aktivitätsdichte der Nutzinsekten auf den jeweiligen Brachen erfasst
wurde, nicht jedoch die tatsächlichen Abundanzen. Um diese festzustellen, sind
großflächigere und längerfristige Beobachtungen oder Fänge durchzuführen.
Da alle drei untersuchten Tiergruppen flugfähige Tiere sind und um die Frage nach
weiteren Einflussfaktoren zu klären, wurde auch die unmittelbare Umgebung der
jeweiligen Brache auf vorhandene Strukturen untersucht. Auf Luftbildern aus der Region
Marchfeld (Anhang I) wurde ein Umkreis von 500 m um die Brache definiert und das
gesamte Gebiet abgegangen. Mit Hilfe der Luftbilder und der in natura ermittelten
Strukturen wurde der prozentuelle Anteil an sogenannten Umgebungsparametern (im
folgenden Text als UP bezeichnet) ermittelt.
Die Einteilung der Fläche erfolgte in die Landschaftstypen:
§ Felder: fields (ackerbaulich genutzt, Halm-, Hackfrüchte, Ölsaaten,
Gemüsekulturen)
§ Grünland: grassland (Wiesen, Brachen, Weingärten, Windschutzstreifen)
§ Siedlungen: houses (Gebäude, Siedlungsgebiet, Gärten, Gewässer, Sport- und
Freizeiteinrichtungen)
Jede dieser Strukturen ist in allen neun Untersuchungsgebieten präsent. Eine genauere
Differenzierung der Umgebung (etwa in Wald, lineare Strukturen, Gewässer) war nicht
sinnvoll, da sie nicht überall vorhanden waren. Die Vergleichbarkeit untereinander wäre
dadurch nicht gegeben.
4.4 Statistik
Die statistische Auswertung der Daten erfolgte mittels der Software MS Excel 2003 ©,
SPSS for Windows©, SAS© und CANOCO©.

13
Um die Vorraussetzung für parametrische Tests (Normalverteilung, Varianzhomogenität)
zu gewährleisten wurde eine logarithmische Datentransformation der Tierdaten
durchgeführt:
X`= lg(X+1)
X = Originalwert
X` = Wert nach der Transformation
Anschließend wurden sie einer Prüfung auf Normalverteilung (Kolmogorov-Smirnov Test,
Shapiro Wilk Test) und Varianzhomogenität (Levene Statistik) unterzogen.
Die so transformierten Daten zeigten keine größeren Heterogenitäten, als man aufgrund
des Zufallsprozesses einer Zählung erwarten sollte.
4.4.1 Zweifaktorielle Varianzanalyse
Die Abundanzen der untersuchten Tierguppen wurden einer zweifaktoriellen
Varianzanalyse (ANOVA) mit SPSS© sowie SAS© unterzogen.
Diese zwei Faktoren waren:
Age (Altersstufen der Brachen, aufgeteilt in y: jung (3-4 Jahre), m: mittelalt (7-9 Jahre), o:
alt (13-16 Jahre)) und Site (die neun Lokalisationen der Brachen), welcher innerhalb des
Faktors age verschachtelt (nested) ist (siehe Tab. 1).
Im Anschluss daran wurde ein post hoc Test, der Tukey Test durchgeführt. Bei diesem
Verfahren werden in multiplen Vergleichen zwischen den Mittelwerten jene Gruppen
angezeigt, die sich signifikant voneinander unterscheiden (SACHS 1997).
4.4.2 Ordinationsverfahren
Um die Verteilung der Arten entlang von Unweltvariablen analysieren zu können, wurde
ein nicht lineares Ordinationsverfahren für die Monate Juni und Juli durchgeführt. Das
Verfahren der Wahl war eine direkte Gradientenanalyse (direct gradient analysis), die
Kanonische Korrespondenzanalyse (CCA).
Neben dem Alter und den damit verbundenen Sukzessionsprozessen der Brachen
können die jeweiligen Flächenparameter nachweislich das Vorhandensein der Nützlinge
beeinflussen. Die CCA ermöglicht eine multiple Regression, in der Zusammenhänge
zwischen den Tierarten und den Umweltfaktoren berechnet und grafisch dargestellt
werden. Entscheidungskriterium für die Annahme oder die Ablehnung eines Modells ist
auch hier der p- Wert (TER BRAAK 1988).
Zur Charakterisierung der jeweiligen Brachen wurden neben den
Blütenbesuchsbeobachtungen auch die lokalen Flächenbedingungen erhoben.

14
Die in die Ordination einfließenden Parameter waren:
§ Faktor Blütenfarben (eingeteilt in die vier Farbguppen red, yellow, white, blue)
§ Faktor Blütendeckungsgrad (flow cov in %)
§ Faktor Anzahl der blühenden Pflanzenarten (species)
4.4.3 Regressionsanalysen
Sowohl mit den Flächen- als auch den Umgebungsparametern (FP, UP) wurden jeweils
einfache und multiple Regressionsanalysen durchgeführt, um den möglichen Einfluss
eines oder mehrerer Parameter auf das Vorhandensein der Tiere zu untersuchen. Eine
multiple Regression, die alle Parameter (FP und UP) enthält, wurde ebenfalls berechnet.
Bei den angewendeten multiplen Verfahren wurde die Rückwärtsmethode gewählt. Diese
stellt eine blockweise Regressionsanalyse dar, in der zu Beginn alle unabhängigen
Variablen enthalten sind. In den folgenden Rechenschritten wird jeweils jene Variable
entfernt, welche den kleinsten partiellen Korrelationskoeffizienten aufweist. Nicht
ausgeschlossen wird der Parameter jedoch, wenn er signifikant ist (Schwellenwert für das
Signifikanzniveau bei der Elimination einer Variablen, F- Wert= 0,1) (BACKHAUS 2000).
5 Ergebnisse
Die Ergebnisse der Untersuchung werden in drei Teilen behandelt.
Die erste Fragestellung behandelte das Alter der Brachen und dessen möglichen Einfluss
auf die Nutzung durch die Insektengruppen. Die 0 - Hypothese lautete: „Das Alter der
Brache hat keinen signifikanten Einfluss auf die Anzahl blütenbesuchender Insekten“. Die
Brachen jeder Altersstufe können als störungsarme, nahrungsreiche Rückzugsgebiete in
gleicher Intensität von den Bestäubergruppen genutzt werden.
In der zweiten Fragestellung wurde neben dem Alterseffekt zusätzlich auch auf
vorhandene Korrelationen zwischen Nützlingen und lokalen Flächenparametern
eingegangen.
In der dritten Fragestellung wurde der mögliche Einfluss der Flächen- und
Umgebungsparameter auf Aktivität und Abundanz der Nützlinge mittels univariater und
multipler Regression berechnet.
Die Monate Mai, Juni und Juli wurden hierbei getrennt betrachtet, da es im Laufe der
Vegetationsperiode zu einer Änderung des Angebots an blühenden Pflanzen, also der
Nahrungsquellen, in den Brachen kam. In dieser Analyse wurde die Gesamtdichte der
Nutzinsekten betrachtet, nicht die Gattungen bzw. Arten im Einzelnen. In den drei
Monaten kam es zu insgesamt 2139 Blütenbesuchen durch die Insekten, wobei 51,66%
auf Schwebfliegen, 35,20 % auf Hummeln und 13,14% auf Honigbienen entfielen.

15
Um eine bessere Darstellbarkeit zu erzielen, wurden die Daten vor der grafischen
Aufbereitung einer Rücktransformation unterzogen.
5.1 Einfluss des Alters der Brachen auf die Blütenbesuche: Monat Mai
Der erste Beobachtungszyklus wurde von 13. bis 28. Mai 2009 durchgeführt. Insgesamt
wurden im Mai 376 Blütenbesuche gezählt. Davon entfielen 20,16% auf Honigbienen,
65,94% auf Schwebfliegen und 13,90% auf Hummeln. Während sich im Mai das Alter der
Brachen signifikant auf die Abundanz der Hummeln auswirkte, gab es keinen Alterseffekt
bei Honigbienen und Schwebfliegen (Tab. 2). Die Dichte der Hummeln nahm in der Reihe
3-4 > 13-16 > 7-9 signifikant ab, wobei in den 7-9 jährigen Brachen keine Tiere
nachgewiesen wurden (Abb. 6).
Tab. 2: F- Werte einer zweifaktoriellen Varianzanalyse (ANOVA): Mai. Auswirkung der Faktoren „Alter“ (age) und „Standort“ (site) auf die Individuenzahl der Honigbienen, Schwebfliegen und Hummeln im Mai (log transformierte Daten).
F- values May honeybees hoverflies bumblebees age 1,50 0,26 16,56*** site 4,44*** 6,36*** 12,15*** modell 3,71** 4,84*** 13,25***
*** p< 0,001, ** p<0,01, * p≤ 0,05 Die Individuendichte der Bienen war zwar sehr gering, es wurden aber in allen drei
Alterstypen der Brachen Honigbienen beobachtet. Syrphiden waren im Mai die
individuenstärkste Gruppe und konnten in allen drei Alterskategorien in annähernd
gleicher Dichte nachgewiesen werden (Abb. 6).

16
May 2009
0
1
2
3
4
honeybees hover flies bumblebees
Age / years
Ind
ivid
ua
ls /
fallo
w
3-4
7-9
13-16a
a
aa
a
ab
a
b
Abb. 6: Der Einfluss des Alters im Mai. Individuenzahlen (Mittelwerte mit Standardfehler) der Honigbienen (honeybees), Schwebfliegen (hoverflies) und Hummeln (bumblebees) in den 3 Altersklassen der untersuchten Brachen. Balken mit unterschiedlichen Buchstaben innerhalb jeder Tiergruppe unterscheiden sich signifikant voneinander. Die Daten wurden nach log Transformation rücktransformiert.
Die Dichten der Honigbienen und Schwebfliegen unterschieden sich nicht signifikant
zwischen den Altersklassen der Brachen.
5.2 Einfluss des Alters der Brachen auf die Blütenbesuche: Monat Juni
Der zweite Beobachtungszyklus wurde von 5. bis 27. Juni 2009 durchgeführt. Insgesamt
wurden im Juni 627 Blütenbesuche gezählt. Davon entfielen 13,40% auf Honigbienen,
60% auf Schwebfliegen und 34,60% auf Hummeln.
Die Dichten von Honigbienen, Syrphiden und Hummeln waren signifikant hinsichtlich des
Einflusses des Alters der Brache auf die Nutzung durch die Insekten (Tab. 3).
Tab. 3: F- Werte einer zweifaktoriellen Varianzanalyse (ANOVA): Juni. Auswirkung der Faktoren „Alter“ (age) und „Standort“ (site) auf die Individuenzahl der Honigbienen, Schwebfliegen und Hummeln im Juni (log transformierte Daten).
F- values June
honeybees hoverflies bumblebees age 10,26*** 4,63** 6,65*** site 1,63 6,07*** 4,39* modell 3,79* 5,71*** 4,96***
*** p< 0,001, ** p<0,01, * p≤ 0,05

17
Die Dichte der Honigbienen (honeybees) und Hummeln (bumblebees) war in den
mittelalten Brachen (7-9 Jahre) signifikant niedriger als in den jungen (3-4 Jahre) oder
alten Brachen (13-16 Jahre). Die Individuenzahlen der Schwebfliegen war in den alten
Brachen signifikant höher als in den jungen und mittelalten Brachen (Abb. 7).
June 2009
0
1
2
3
4
5
6
honeybees hover flies bumblebees
Age / years
Ind
ivid
ual
s /
fall
ow
3-47-913-16
a a
aa
a
b
b
b
b
Abb. 7: Der Einfluss des Alters im Juni Individuenzahlen (Mittelwerte mit Standardfehler) der Honigbienen (honeybees), Schwebfliegen (hoverflies) und Hummeln (bumblebees) in den 3 Altersklassen der untersuchten Brachen. Balken mit unterschiedlichen Buchstaben innerhalb jeder Tiergruppe unterscheiden sich signifikant voneinander. Die Daten wurden nach log Transformation rücktransformiert.
In den mittelalten Brachen kam es im Vergleich zum Mai zu einem deutlichen Rückgang
an Blütenbesuchen, vor allem bei Honigbienen und bei den Schwebfliegen. Die Hummeln
schienen erstmals in allen drei Altersklassen der Brachen auf, jedoch auch hier mit der
niedrigsten Dichte in mittelalten Brachen (Abb. 7).
5.3 Einfluss des Alters der Brachen auf die Blütenbesuche: Monat Juli
Der dritte Beobachtungszyklus wurde von 2. bis 17. Juli durchgeführt. Insgesamt wurden
im Juli 1145 Blütenbesuche gezählt. Davon entfielen 10,74% auf Honigbienen, 46,90%
auf Schwebfliegen und 42,36% auf Hummeln.
Die Abundanzen von Honigbienen und Hummeln unterschieden sich signifikant zwischen
den Altersklassen der Brachen (Tab. 4).

18
Tab. 4: F- Werte einer zweifaktoriellen Varianzanalyse (ANOVA): Juli. Auswirkung der Faktoren „Alter“ (age) und „Standort“ (site) auf die Individuenzahl der Honigbienen, Schwebfliegen und Hummeln im Juli (log transformierte Daten).
F- values July
honeybees hoverflies bumblebees age 3,32* 3,17 5,35** site 8,38*** 2,09 4,68** modell 7,11*** 2,36* 4,84***
*** p< 0,001, ** p<0,01, * p≤ 0,05
Der untersuchte Einfluss des Alters der Brachen war bei Honigbienen mit einem p- Wert
von 0,047 statistisch signifikant, jedoch wurde dies im konservativen Tukey Test nicht
angezeigt (Abb. 8). Die Individuenzahl der Bienen war in den mittelalten Brachen deutlich
niedriger als in den alten und jungen Brachen Die Aktivitätsdichte von Hummeln war in
den mittelalten Brachen (7-9 Jahre) signifikant niedriger als in jungen (3-4 Jahre) und
alten (13-19 Jahre) Brachen. Bei den Schwebfliegen zeigte sich im Juli keine Signifikanz
(Tab. 4), jedoch lag aufgrund des p-Wertes (0,054) ein Trend hinsichtlich eines Einflusses
des Alters der Brachen vor. Ähnlich wie bei Honigbienen und Hummeln waren die Dichten
in den mittelalten Brachen deutlich niedriger als in den jungen und alten Brachen (Abb. 8).
July 2009
0
1
2
3
4
5
6
7
8
9
10
11
12
honeybees hover flies bumblebees
Age / years
Ind
ivid
ual
s / f
allo
w
3-4
7-9
13-16
a
a
a
a
a
a
a a
b
Abb. 8: Der Einfluss des Alters im Juli Individuenzahlen (Mittelwerte mit Standardfehler) der Honigbienen (honeybees), Schwebfliegen (hoverflies) und Hummeln (bumblebees) in den 3 Altersklassen der untersuchten Brachen. Balken mit unterschiedlichen Buchstaben innerhalb jeder Tiergruppe unterscheiden sich signifikant voneinander. Die Daten wurden nach log Transformation rücktransformiert.

19
Zusammenfassend war auch im Juli die Anzahl an Blütenbesuchen aller drei
Insektengruppen in den mittelalten Brachen am geringsten.
Die Analyse der Daten der drei Monate ergab, dass das Alter der Brachen häufig einen
signifikanten Einfluss auf die Aktivitätsdichte von blütenbesuchenden Insekten hatte.
5.4 Aktivitätsdichte unter Einfluss von Flächenparametern
Neben dem Alter und den damit verbundenen Sukzessionsprozessen der
Vegetationsdecke wurden auf den Brachen vorhandene Bedingungen und
Habitatstrukturen hinsichtlich eines fördernden Einflusses auf das Vorhandensein von
Nützlingen untersucht. In die Ordination flossen die erhobenen Flächenparameter FP
(siehe Kapitel 4.4.2) ein. Durch das Ordinationsverfahren werden die aufgenommenen
Tierarten entlang von Achsen angeordnet. Somit finden sich Nützlinge mit ähnlichen
Ansprüchen an ihre Umwelt in nahen Positionen zueinander, sie gruppieren sich auch zu
den jeweiligen Umweltfaktoren.
Die sehr geringen Insektendichten im Mai machten eine Ordination nicht sinnvoll. Erst mit
Anstieg der Insektendichten in den Monaten Juni und Juli konnte eine interpretierbare
Ordination durchgeführt werden.
In den folgenden CCAs der Monate Juni und Juli kann bei Schwebfliegen und bei
Hummeln näher differenziert und auf das Artenniveau eingegangen werden. Auf den
Flächen beobachtete und in konstanten Abundanzen vorkommende Hummeln waren
Dunkle Erdhummel (B. terrestris), Ackerhummel (Pyrobombus lapidarius) und
Gartenhummel (B. hortorum). Diese ubiquitär vorkommenden Hummeln wurden auf allen
neun Brachen und in allen drei Beobachtungseinheiten festgestellt.
Wiesenhummel (Pyrobumbus pratorum), Waldhummel (Megabombus sylvarum),
Ackerhummel (Megabombus pascuorum floralis), Veränderliche Hummel (Megabombus
humilis) und Kuckuckshummeln wie die Keusche Schmarotzerhummel (Psithyrus vestalis)
wurden nur vereinzelt beobachtet und nicht in die Ordination aufgenommen.
5.4.1 Ordination im Juni
Die CCA erklärte 28,4% der Gesamtvarianz der Bestäubergemeinschaft.
Die Blütenfarbe red (7,4%) und der Faktor species (13,35%) wirkten sich signifikant auf
die Tiergruppen aus und erklärten 20,75% der Gesamtvarianz.
Wie im Ordinationsdiagramm (Abb. 9) ersichtlich, wurden die drei Altersklassen der
Brachen sehr gut durch Achse 2 voneinander getrennt. So finden sich junge Brachen (y)

20
sehr zentral im Koordinationsursprung, alte Brachen (o) positionieren sich im linken
Quadranten. An Achse 1, weit rechts außen liegend, gruppieren sich mittelalte Brachen
(m) (Abb. 9).
Bei der Positionierung der FP ist die Blühfarbe red (dazu zählen unter anderem Carduus
sp, Cirsium arvense, Trifolium pratense, Papaver rhoeas, Onobrychis viciaefolia) aufgrund
der Länge des berechneten Vektors ein sehr einflussreicher Faktor, jedoch besteht zu
keiner der beobachteten Arten eine enge Korrelation. Am ehesten ist noch eine positive
Korrelation mit B. hortorum und B. terrestris gegeben. Die Blühfarben yellow (dazu
zählen unter anderem Lotus corniculatus, Reseda lutea, Tragopogon pratensis, Senecio
vulgaris, Sysymbrium loeselii, Sonchus oleraceus, Solidago gigantea) und white (dazu
zählen unter anderem Achillea millefolium, Anthemis austriaca, Capsella bursa-pastoris,
Convolvulus arvensis, Erigeron annuus, Stellaria media, Tripleurospermum perforatum)
sowie der Faktor species korrelierten stark mit alten Brachen, während die weiteren FP
nicht deutlich mit einzelnen Alterklassen korrelierten (Abb. 9). Bei den Insektentaxa ist es
die Honigbiene (A. meli) die am häufigsten in alten Brachen vorkam und positiv mit den
Flächenparametern species, white und yellow korrelierte. Die Schwebfliegen der
Gattung Sphaerophoria waren positiv mit der Blühfarbe blue (dazu zählen unter anderem
Echium vulgare, Medicago sativa, Vicia villosa) assoziiert. Die zentrumsnahe
angeordneten Insekten wie B. lapidarius, Schwebfliegen der Gattung Eristalis und E.
balteatus verhielten sich indifferent und erlaubten hier keine Aussage über Präferenzen
auf den Brachen (Abb. 9). Sowohl bei der Ordination im Juni als auch im Juli sind die
Blühfarben red und blue einander gegenüber angeordnet (Abb. 9, Abb. 10).

21
A.meli.
E. balt.
Erist.
Sphaer.
B.lapi.
B.terr.
B.hort.
flow.cov.
species white
yellow
red
blue
y
m o
Axis 2
Axis 1
Abb. 9: CCA Grafik für Juni Ordinationsdiagramm einer kanonischen Korrespondenzanalyse mit Abundanzdaten von Bestäubertaxa und unterschiedlichen Umweltfaktoren in den drei Altersklassen der untersuchten Brachen (y = 3-4, m = 7-9, o = 13-16 Jahre). Abkürzungen Bestäubertaxa: B. hort. = Bombus hortorum, B. terr = Bombus terrestris, B. lapi. = Pyrobombus lapidarius, Erist. = Eristalis sp., E. balt. = Episyrphus balteatus, Sphaer. = Sphaerophoria sp., A. meli. = Apis mellifera. Blütenfarben: red = rot, yellow = gelb, white = weiß, blue = blau. FP: flow cov = Blütendeckung, species = Anzahl der Blühpflanzen.
Die CCA im Juni bestätigte deutlich den Trend der geringeren Individuendichten in den
mittelalten Brachen. Keine der Tierarten oder der FP korrelierten positiv mit mittelalten
Brachen. Die drei Vektoren für species und Blühfarben yellow und white sind eindeutig
gegenläufig positioniert, was einer negativen Korrelation entspricht.

22
5.4.2 Ordination im Juli
Die CCA erklärte 24,04 % der Gesamtvarianz der Bestäubergemeinschaft.
Der Faktor species wirkte sich signifikant aus und erklärte 8,1% der Gesamtvarianz. Der
Einfluss aller weiteren, einzelnen Parameter auf das Modell insgesamt war annähernd
gleich hoch, jedoch nicht signifikant (Varianzerklärung durch red = 8,1%, blue = 6,1%,
white, yellow und flow cov = 4%).
Wie im Ordinationsdiagramm (Abb. 10) ersichtlich, wurden die drei Altersstufen der
Brachen deutlich durch Achse 1 voneinander getrennt. Junge (y) und alte (o) Brachen
positionierten sich wiederum deutlich gegenläufig zu den mittelalten Brachen (m) (Abb.
10). Die Faktoren red, flow cov und species korrelierten stark mit den alten Brachen, die
Blühfarbe blue deutlich mit den jungen Brachen. Die Blühfarben white und yellow
korrelierten nicht mit einer der Altersklassen. Die Hummelart B. hortorum kam am
häufigsten in den alten Brachen vor und korrelierte somit mit den Faktoren red, flow cov
und species. Weitere Korrelationen ergaben sich zwischen der Hummelart B. terrestris
und der Blühfarbe white und den Schwebfliegen der Gattung Eristalis sp. mit der
Blühfarbe blue in jungen Brachen. Die Bedeutung von jungen Brachen für die
Insektenpopulationen war im Vergleich zum Vormonat höher, diese Brachen
positionierten sich nicht mehr zentrumsnahe, sondern im linken unteren Quadranten der
Ordination. Wie auch im Ordinationsdiagramm Juni positionierten sich E. balteatus und B.
lapidarius in der Nähe des Zentrums, zudem auch Schwebfliegen der Gattung
Sphaerophoria und A. mellifera.
Die mittelalten Brachen betreffend ergab sich auch im Juli ein, dem Vormonat ähnliches,
Bild. Mittelalte Brachen korrelierten mit insgesamt fünf der sechs Umweltparameter (red,
flow cov, species, yellow und white) deutlich negativ. Auch bei den Insekten gab es
keinerlei positive Korrelation mit Brachen dieser Altersklasse (Abb. 10).

23
A.meli.
E.balt.
Erist.
Sphaer.
B.lap.
B.terr.
B.hort.
flow.cov.
species
white
yellow
red
blue
y
m
o
Axis 2
Axis 1
Abb. 10: CCA Grafik für Juli Ordinationsdiagramm einer kanonischen Korrespondenzanalyse mit Abundanzdaten von Bestäubertaxa und unterschiedlichen Umweltfaktoren in den drei Altersklassen der untersuchten Brachen (y = 3-4, m = 7-9, o = 13-16 Jahre). Abkürzungen Bestäubertaxa: B. hort. = Bombus hortorum, B. terr = Bombus terrestris, B. lapi. = Pyrobombus lapidarius, Erist. = Eristalis sp., E. balt. = Episyrphus balteatus, Sphaer. = Sphaerophoria sp., A. meli. = Apis mellifera. Blütenfarben: red = rot, yellow = gelb, white = weiß, blue = blau. FP: flow cov = Blütendeckung, species = Anzahl der Blühpflanzen.
5.5 Aktivitätsdichte unter Einfluss von Flächen- und
Umgebungsparametern
Die Aktivitätsdichte der Bestäuber in Brachen unterschiedlichen Alters und unter dem
Einfluss von Flächen- und Umgebungsparametern wurde mit Hilfe einfacher und multipler
Regression untersucht. Neben Flächenparametern wie flow cov und species flossen die
zusätzlichen Parameter Windgeschwindigkeit (wind) und Temperatur (temp), sowie die
umgebenden Landschaftstypen in Form der Umgebungsparameter (UP) in die
Regressionsmodelle ein (siehe Kapitel 4.3).
Die Datengrundlage der Blütenbesuchsbeobachtungen im Mai war aufgrund der geringen
Individuenanzahl vor allem bei Honigbienen und Hummeln wenig aussagekräftig.
5.5.1 Honigbienen
Die Honigbienen korrelierten in der einfachen Regression im Mai nicht signifikant mit
einem der Einzelparameter (FP oder UP). In der multiplen Regression mit FP korrelierte
die Anzahl von A. mellifera tendenziell positiv mit species, wobei jedoch alle anderen FP
eliminiert wurden. Bei den UP ergab das Modell, welches fields und houses beinhaltet,

24
eine signifikante Korrelation mit hohem Erklärungswert (r2 = 0,79) (Tab. 5). Beide
Parameter sind negativ korreliert was bedeutet, dass sich Honigbienen im Mai nicht in
jenen Brachen fanden, in deren Umkreis die Strukturen Felder oder Siedlungen
überwiegen. Im Gesamtmodell, in welchem alle Parameter einflossen (AP), haben
wiederum die drei Parameter species, fields und houses einen sehr hohen
Erklärungswert (r2 = 0,93) (Tab. 5). Im Mai fanden sich viele Honigbienen in Brachen, die
eine hohe Anzahl an Blühpflanzen aufwiesen, jedoch siedlungsfern waren und einen
niedrigen Anteil an Feldern hatten. Weder im Juni noch im Juli konnten weitere,
signifikante Korrelationen zwischen Flächen- oder Umgebungsparametern als Einfluss auf
die Dichte der Honigbienen festgestellt werden.
Tab. 5: Einfache und multiple Regression für Honigbienen (A. mellifera): Der Einfluss von FP und UP bei einfacher und multipler Regression auf Honigbienen (A. mellifera). m R = multiple Regression, FP = Flächenparameter, UP = Umgebungsparameter, AP = alle Parameter.
Honeybees Model Parameter p-Value R2 Beta
m R, FP species 0,06 0,50 0,74 m R, UP fields 0,005 -2,00 MAY houses 0,003 0,79 -2,22 m R, AP species 0,02 0,39 fields 0,002 0,93 -1,83 houses 0,001 -2,18
5.5.2 Schwebfliegen
Schwebfliegen, die in allen drei Monaten jeweils die individuenstärkste Gruppe waren,
zeigten bei der einfachen Regression mit UP und bei den multiplen Regressionen mit FP,
UP und AP Signifikanzen.
Die multiple Regression mit den FP flow cov und species war in allen drei Monaten
signifikant. In der einfachen Regression war der FP species aufgrund der p- Werte nicht
signifikant, im multiplen Modell jedoch verblieben nach Elimination aller anderen
Einflussparameter diese beiden Parameter. Die Anzahl der Schwebfliegen korrelierte im
Mai positiv mit beiden FP, eine dichte Blütendeckung und eine hohe Anzahl an
Blühpflanzen waren für die Anzahl der Schwebfliegen bedeutend. In den Monaten Juni
und Juli korrelierten die Schwebfliegen jedoch negativ mit species und positiv mit flow
cov (Tab. 6). Eine prozentuell hohe Blütendeckung bei gleichzeitig geringer Anzahl
verschiedener Blühpflanzen auf den Brachen, war für die Dichte der Schwebfliegen in den
Monaten Juni und Juli wichtig.
25
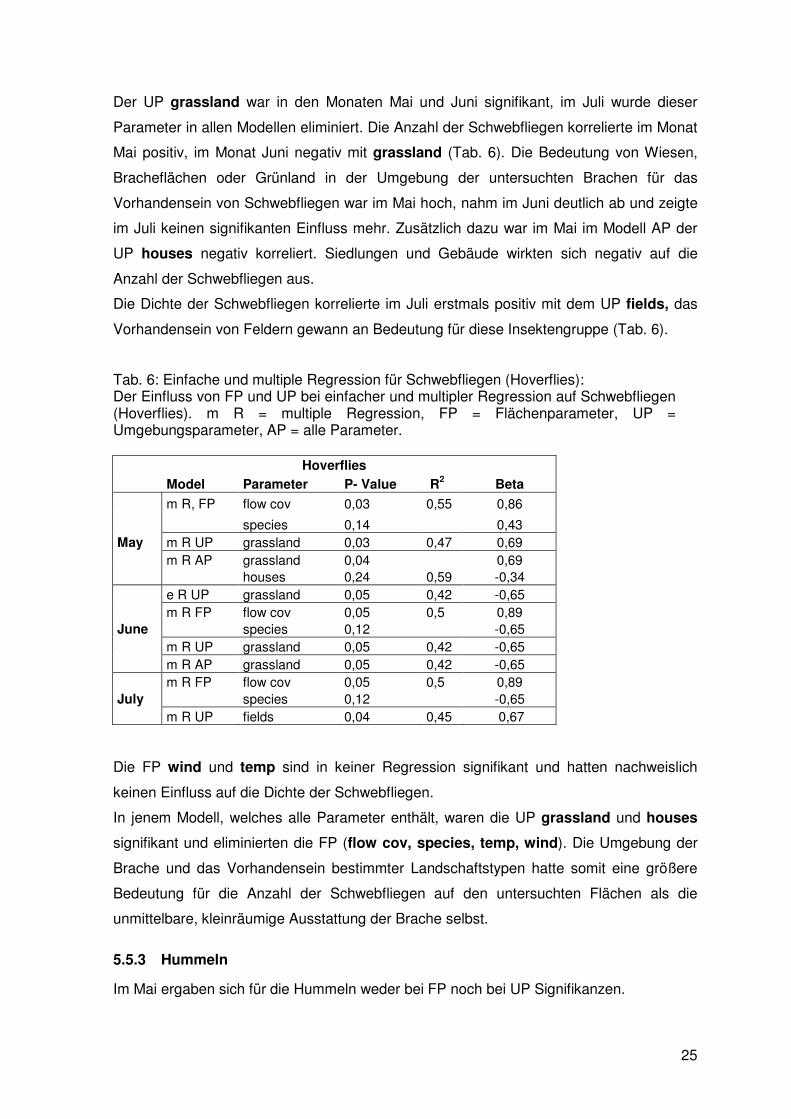
Der UP grassland war in den Monaten Mai und Juni signifikant, im Juli wurde dieser
Parameter in allen Modellen eliminiert. Die Anzahl der Schwebfliegen korrelierte im Monat
Mai positiv, im Monat Juni negativ mit grassland (Tab. 6). Die Bedeutung von Wiesen,
Bracheflächen oder Grünland in der Umgebung der untersuchten Brachen für das
Vorhandensein von Schwebfliegen war im Mai hoch, nahm im Juni deutlich ab und zeigte
im Juli keinen signifikanten Einfluss mehr. Zusätzlich dazu war im Mai im Modell AP der
UP houses negativ korreliert. Siedlungen und Gebäude wirkten sich negativ auf die
Anzahl der Schwebfliegen aus.
Die Dichte der Schwebfliegen korrelierte im Juli erstmals positiv mit dem UP fields, das
Vorhandensein von Feldern gewann an Bedeutung für diese Insektengruppe (Tab. 6).
Tab. 6: Einfache und multiple Regression für Schwebfliegen (Hoverflies): Der Einfluss von FP und UP bei einfacher und multipler Regression auf Schwebfliegen (Hoverflies). m R = multiple Regression, FP = Flächenparameter, UP = Umgebungsparameter, AP = alle Parameter.
Hoverflies
Model Parameter P- Value R2 Beta
m R, FP flow cov 0,03 0,55 0,86
species 0,14 0,43 May m R UP grassland 0,03 0,47 0,69 m R AP grassland 0,04 0,69 houses 0,24 0,59 -0,34 e R UP grassland 0,05 0,42 -0,65 m R FP flow cov 0,05 0,5 0,89 June species 0,12 -0,65 m R UP grassland 0,05 0,42 -0,65 m R AP grassland 0,05 0,42 -0,65 m R FP flow cov 0,05 0,5 0,89 July species 0,12 -0,65 m R UP fields 0,04 0,45 0,67
Die FP wind und temp sind in keiner Regression signifikant und hatten nachweislich
keinen Einfluss auf die Dichte der Schwebfliegen.
In jenem Modell, welches alle Parameter enthält, waren die UP grassland und houses
signifikant und eliminierten die FP (flow cov, species, temp, wind). Die Umgebung der
Brache und das Vorhandensein bestimmter Landschaftstypen hatte somit eine größere
Bedeutung für die Anzahl der Schwebfliegen auf den untersuchten Flächen als die
unmittelbare, kleinräumige Ausstattung der Brache selbst.
5.5.3 Hummeln
Im Mai ergaben sich für die Hummeln weder bei FP noch bei UP Signifikanzen.

26
Im Juni korrelierte die Anzahl der Hummeln mit den FP flow cov, temp und species im
multiplen Modell. Wie auch bei den Schwebfliegen waren die Parameter flow cov und
species signifikant. Für die Anzahl der Hummeln auf den Brachen waren sowohl eine
prozentuell hohe Blütendeckung als auch eine hohe Anzahl an Blühpflanzen
entscheidend. Im Modell hinzukommend war die negative Korrelation mit dem FP temp.
Hohe Temperaturen wirkten sich negativ auf die Anzahl der Blütenbesuche durch
Hummeln aus. War der FP temp bei keinem der anderen beobachteten Blütenbesucher
signifikant, ist er bei den Hummeln erstmals von Bedeutung (Tab. 7). In der multiplen
Regression AP wurde einzig der UP grassland eliminiert, die weiteren Faktoren blieben
aufgrund ihrer Signifikanz im Modell und erklärten zusammen 99% der Varianz (Tab. 7).
Auch hier war der Faktor temp negativ korreliert, ebenso korrelierte die Anzahl der
Hummeln negativ mit Gebäuden und Siedlungen (houses). Der FP wind schien erstmals
bei den Hummeln auf und ist positiv korreliert.
Tab. 7: Einfache und multiple Regression für Hummeln (Bumblebees): Der Einfluss von FP und UP bei einfacher und multipler Regression auf Hummeln (Bumblebees). m R = multiple Regression, FP = Flächenparameter, UP = Umgebungsparameter, AP = alle Parameter.
Bumblebees
Model Parameter P- Value R2 Beta flow cov 0,063 0,43 m R FP temp 0,01 0,87 -0,64 species 0,09 0,37 flow cov 0,004 1,32 June temp 0,004 -0,52 m R AP wind 0,04 0,32 species 0,01 0,99 0,4 fields 0,02 0,48 houses 0,03 -0,44
Im Juli konnten keine Korrelationen zwischen der Dichte der Hummeln und den Faktoren
UP, FP und AP festgestellt werden.
5.6 Blühpflanzenanzahl
In den vorangehenden Analysen zeigte sich, dass vor allem in den Monaten Juni und Juli
die mittelalten Brachen von weniger Bestäubern genutzt wurden als junge oder alte
Brachen.
Neben den Tierbeobachtungen wurde zum jeweiligen Aufnahmezeitpunkt auch die Anzahl
der blühenden Trachtpflanzen (species) aufgezeichnet. Da der Parameter species
sowohl in der Kanonischen Korrespondenzanalyse als auch bei den Regressionsmodellen

27
stets einflussreich war, wurde die Anzahl der blühenden Pflanzen auf den Brachen in den
Monaten Juni und Juli einer genaueren Betrachtung unterzogen. Der Vergleich der Anzahl
der Blühpflanzen mittels zweifaktorieller ANOVA zwischen den drei Alterklassen der
Brachen ist in Tab. 8 ersichtlich.
Tab. 8: F- Werte einer zweifachen Varianzanalyse (ANOVA): Blühpflanzenanzahl. Auswirkungen der Faktoren „Alter“ (age) und „Standort“ (site) auf die Anzahl an Blühpflanzen in den Brachen der Monate Juni und Juli.
F- values species
June July age 47,78*** 37,32*** site 7,91*** 8,21*** modell 17,78*** 15,49***
*** p< 0,001, ** p<0,01, * p≤ 0,05
Number of plant species
0
2
4
6
8
10
12
14
16
18
3-4 7-9 13-16 3-4 7-9 13-16 Age (years)
Nu
mb
er /
fal
low
fallow young
fallow middle
fallow old
JulyJune
a
a
a
a
b b
Abb. 11: Anzahl von Blühpflanzen in den drei Altersklassen der Brachen in den Monaten Juni und Juli. Balken mit unterschiedlicher Farbe kennzeichnen die drei verschiedenen Alterstypen der Brachen. Aufgetragen sind Mittelwert und Standardabweichung. Balken mit unterschiedlichen Buchstaben innerhalb der Monate unterscheiden sich signifikant voneinander.
Sowohl im Juni als auch im Juli war die Anzahl blühender und somit nutzbarer Pflanzen in
den mittelalten Brachen (Mittelwerte zwischen 2,93 im Juni und 2,87 im Juli) am
niedrigsten. Ein gutes Angebot an Trachtpflanzen fand sich in den jungen (zwischen 8,07
im Juni und 7,47 im Juli) und in den alten Brachen (Mittelwerte im Juni 11,67 und 10,27 im
Juli) (Abb. 11). Alte Brachen hatten in beiden Monaten die höchste Anzahl an
Blühpflanzen.

28
6 Diskussion
Im Folgenden soll der Einfluss des Alters der Brachen auf die Aktivitätsdichte von
Honigbienen, Schwebfliegen und Hummeln diskutiert werden.
In diesem Zusammenhang wird auch der Frage nachgegangen, inwieweit
flächenspezifische Parameter wie Blütendeckung, Blühpflanzenanzahl oder Blühfarben,
sowie die umgebenden landschaftlichen Strukturen die Anzahl der Insekten beeinflussen.
6.1 Sekundärsukzession auf Brachen: Änderung der Pflanzengesellschaft
Die Intensivierung der landwirtschaftlichen Produktion in den letzten Jahrzehnten führte
zu Verlusten bei Strukturheterogenität und Habitatdiversität und zu einem deutlichen und
mehrfach dokumentierten Rückgang der Biodiversität in landwirtschaftlichen Gebieten
(KOHLER et al. 2007, HAENKE et al. 2009, POTTS et al. 2009). Intensiv genutztes Ackerland ist
sehr homogen, strukturarm und stark an die anthropogenen Bewirtschaftungsziele
angepasst. Um diesen Verlusten vorzubeugen wurden unterschiedliche
Managementkonzepte zur Verbesserung der ökologischen Situation in agrarischen
Gebieten entwickelt (z.B. das österreichische Programm für umweltgerechte
Landwirtschaft ÖPUL oder Programme der gemeinsamen, europäischen Agrarpolitik
CAP) (DECOURTYE et al. 2007). Derartige Maßnahmen sind die Stilllegungen von Äckern
oder Grünland, Anlage von Brachen, Neuanlage von Hecken, Schaffung von
Feuchtbiotopen oder sonstiger Strukturen oder die Einsaat von sogenannten Nützlings-
oder Blühstreifen in Äckern (ÖPUL 2007). Durch das Anlegen von Brachen wurden intensiv
genutzte Flächen aus der Nutzung genommen und einer Eigendynamik überlassen.
Die neun betrachteten Ackerbrachen im Marchfeld bestätigen im Wesentlichen den Ablauf
einer Sekundärsukzession nach Flächenstilllegung, sie blieben aber nicht gänzlich sich
selbst überlassen. Der pflanzliche Aufwuchs wird einmal pro Jahr von den Besitzern der
Fläche teils gehächselt und gemulcht, teils auch abtransportiert (Kapitel 4.1). Im Zuge der
stattfindenden Sekundärsukzession setzten unterschiedliche, standortsabhängige
Entwicklungen ein und die Flächen entwickelten sich gemäß ihrem Potential und den
vorherrschenden biotischen und abiotischen Bedingungen (HOLZNER 2001).
In den ersten Jahren, der sogenannten ersten Besiedelungswelle, dominieren auf frisch
brach gefallenen Flächen jene einjährigen Blühpflanzen, deren Diasporen in den Böden
vorhanden sind oder über Verbreitungsmechanismen (Windverbreitung oder Verbreitung
durch Tiere) auf die Fläche gelangen (HOLZNER 2001). Im Gegensatz zu Grünlandbrachen
ist eine Ackerbrache durch einen sehr raschen Lebensformwechsel vor allem in den
ersten Vegetationsperioden gekennzeichnet. Einjährige Getreideunkräuter (Secalinetalia)

29
oder Hackfruchtunkräuter (Chenopodietalia), welche ansonst durch Herbizidspritzungen
unterdrückt werden, erscheinen zuerst (SCHMIDT 1981). Der Anteil an offenem
Mineralboden ist vor allem bei Ackerbrachen hoch, die verdämmend wirkende Schicht
alter, abgestorbener Biomasse ist noch gering (SCHMIDT 1981, HOLZNER 2001).
Untersuchungen zur Sekundärsukzession von Pflanzengesellschaften auf
selbstbegrünten Ackerbrachen zeigen vor allem in den ersten drei Jahren eine
kontinuierliche Zunahme der Pflanzenartenzahlen, jedoch einen markanten Rückgang
dieser ab dem vierten Jahr bzw. der zweiten Besiedelungswelle (ODUM 1969, GREILER
1994, HOLZNER 2001). Langzeituntersuchungen über den Sukzessionsprozess und die
Kolonisationsfähigkeit von Pflanzen durch SANDOR et al. (2009) konnten einen signifikanten
Abfall an Pflanzenarten ab der fünften Vegetationsperiode nachweisen. Die Ergebnisse
der vorliegenden Untersuchung der neuen Brachen im Marchfeld bestätigten im
Wesentlichen einen derartigen Sukzessionsverlauf. Die drei jungen Brachen (3-4 Jahre)
hatten in den Monaten Juni und Juli einen signifikant höheren Anteil an Blühpflanzen als
mittelalte Brachen (7-9 Jahre). In unmittelbarem Zusammenhang mit dem Rückgang an
Blühpflanzen ist der Rückgang an Blütenbesuchern in mittelalten Brachen zu sehen
(HEGLAND & BOEKE 2006), denn diesen Insekten fehlte es dort an Nahrung. Im Juni konnte
ein Einfluss des Alters auf die Anzahl der Blütenbesuche bei allen drei Bestäubergruppen,
also Honigbienen, Schwebfliegen und Hummeln, deutlich nachgewiesen werden. Im Juli
waren signifikant weniger Honigbienen und Hummeln in mittelalten Brachen zu finden. Bei
den Schwebfliegen konnte dieser Alterseffekt der mittelalten Brachen im Juli zwar
statistisch nicht signifikant festgestellt werden, jedoch ließ sich (mit einem p- Wert von
0,054) dieser Trend ebenfalls erkennen.
Die deutliche Abnahme an für die Bestäuber nutzbaren Blühpflanzen in mittelalten
Brachen kann mit einer Änderung der Pflanzenartengesellschaft im Zuge der zweiten
Besiedelungswelle erklärt werden. Durch das Anwachsen der Streuschicht, die Zunahme
von Zwei- und Mehrjährigen (z.B. Carduus acanthoides, Cirsium arvense, Senecio
vernalis, Solidago canadensis), also dominanten und konkurrenzstarken Pflanzen, werden
die vergleichsweise konkurrenzschwachen Einjährigen (z.B. Anthemis austriaca, Capsella
bursa- pastoris, Erigeron annuus, Galium aparine) weitgehend verdrängt. Auch
horstbildende Gräser (z.B. Arrhenatherum elatius, Apera spica venti, Calamagrostis
epigejos, Dactylis glomerata, Poa pratensis) kommen verstärkt auf und wirken aufgrund
starker Nährstoff- und Lichtkonkurrenz bestandsbildend (HOLZNER 2001). In
Untersuchungen sechsjähriger Brachen zeigte sich eine tiefgreifende Änderung der
Pflanzengesellschaft mit einem nur mehr 1%igen Deckungsgrad bei einjährigen
Blühpflanzen (SCHMIDT 1981, GREILER 1994). Untersuchungen brachgefallener Äcker durch

30
TRAMER (1975), SCHMIDT (1976), BROWN & SOUTHWOOD (1987) und KLOTZ &
SCHMIEDEKNECHT (1992) zeigen mehrfach einen derartigen Rückgang der
Pflanzenartenzahl ab dem dritten bis vierten Jahr (GREILER 1994). Eine steigende Anzahl
unterschiedlicher Pflanzenarten, ähnlich hoch jener in jungen Brachen, konnte erst wieder
ab dem zehnten Jahr der Sukzession nachgewiesen werden (SANDOR et al. 2009). Mit
fortschreitender Sukzession und im Rahmen der dritten Besiedelungswelle werden auch
relativ stabil erscheinende pflanzensoziologische Strukturen durch natürliche Ereignisse
verändert oder durch einwandernde Gehölze oder Sträucher verdrängt, wenn nicht
Maßnahmen gegen Verbuschung oder Verwaldung getroffen werden (HOLZNER 2001).
Im Rahmen von Flächenmanagementmaßnahmen (z.B. einmalige Mahd je
Vegetationsperiode, Beweidung) können Nutzbarkeit und Qualität der Brachen für die
Bestäuber erhalten werden. In diesem Fall ist es vor allem die dritte Besiedelungswelle,
welche verzögert wird. Gehölze und Stauden können trotz ihrer Konkurrenzkraft eine
Fläche nicht besiedeln, da sie nicht erstarken können (HOLZNER 2001). Von der Mahd
profitieren in diesem Fall vor allem lichtliebende Hemikryptophyten, da die Zunahme der
verdämmend wirkenden Streuschicht verlangsamt wird (SCHMIDT 1981). Auch von GREILER
(1994) konnte ein positiver Effekt von Mahd auf die Artenvielfalt der Pflanzen in
Ackerbrachen nachgewiesen werden. In dieser Untersuchung fanden sich in einmal
jährlich gemähten Brachen im Mittel 40% mehr Pflanzenarten als auf ungemähten
Brachen. Auch horstbildende Gräser wurden länger unterdrückt und die Anzahl an
Blühpflanzen sank nicht so rasch ab. Des Weiteren sind vergraste, in der Sukzession weit
fortgeschrittene Flächen nur bedingt für blütenbesuchende Insekten attraktiv.
In der vorliegenden Untersuchung konnten ähnlich gelagerte Ergebnisse gewonnen
werden. Sowohl im Juni als auch im Juli war die höchste Anzahl an Blühpflanzen jeweils
in alten Brachen (13-16 Jahre) zu finden. Durch diese Sukzessionsprozesse der
Pflanzengesellschaft kommt es aufgrund der engen Wechselwirkungen mit ihrem
Lebensraum zu Änderungen der Artenzusammensetzung bzw. der Abundanzen bei den
Insekten (GREILER 1994, HEGLAND & BOEKE 2006). Aufgrund des höheren Angebotes an
Blühpflanzen, somit auch an Nektar und Pollen, wurden vorwiegend alte und junge
Brachen durch die Bestäuber genutzt.
6.2 Der Einfluss des Alters der Brache auf die Bestäuber
Im Rahmen der zweifaktoriellen ANOVA konnte der Einfluss des Alters der Brachen auf
die drei Bestäubergruppen nachgewiesen werden. Signifikant wurde dieser Effekt im Mai
bei Hummeln, im Juni bei Honigbienen, Schwebfliegen und Hummeln und im Juli bei
Honigbienen und Hummeln. In mittelalten Brachen war die Anzahl der Blütenbesuche

31
signifikant niedriger als in jungen oder alten Brachen. Der für die Hummeln im Mai
nachgewiesen Alterseffekt sollte jedoch mit Vorsicht betrachtet werden, da die Anzahl der
Hummeln im Mai noch sehr gering war. Es kam vorwiegend zu Sichtungen von
Hummelköniginnen. Erste Hummelarbeiterinnen, deren Entwicklung zwischen 19 und 22
Tagen dauert, finden sich unter klimatisch sehr guten Bedingungen frühestens Ende Mai,
in der Regel aber erst Anfang bis Mitte Juni (VON HAGEN 1988).
Im Vergleich zum Mai war der Einfluss des Alters der Brachen im Juni deutlicher. Durch
das Auftreten der ersten Generationen bei den Schwebfliegen und das Anwachsen der
Bienen- und Hummelvölker kam es zu einer Steigerung der Tiersichtungen insgesamt. In
der Varianzanalyse wurde deutlich, dass es bei gleichzeitigem Anstieg der Abundanzen
zu einem signifikanten Rückgang der Blütenbesuche aller drei Insektengruppen in den
mittelalten Brachen kam. Diese waren für die Bestäuber weniger attraktiv, was wiederum
mit der geringeren Anzahl an Blühpflanzen, also der Ressourcenausstattung dieser
Flächen, im Zusammenhang stehen kann (WESTRICH 1997, KLEJN & VAN LANGEVELDE 2006,
ALBRECHT et al. 2007, MEYER et al. 2009). Die für Bestäuber wichtigsten Ressourcen sind
das qualitativ und quantitativ ausreichende Vorhandensein von Nahrung und Habitat,
besonders in landwirtschaftlich intensiv genutzten Gebieten (DECOURTYE et al. 2007,
EBELING et al. 2008). Durch die Bruttätigkeit bei Schwebfliegen sowie dem Anwachsen der
Völker bei Honigbienen und Hummeln steigt im Laufe der Vegetationsperiode der Bedarf
an Nektar (VON HAGEN 1988, RÖDER 1990, BARTH 1991). Diesem höheren Nahrungsbedarf
der Insekten stand eine Abnahme potentieller Nahrungspflanzen in mittelalten Brachen
gegenüber. Die meisten Frühlingsblüher (z.B. Leontodon hispidus, Capsella bursa-
pastoris…) waren im Juni bereits verblüht. In den Kulturen der Ackerflächen war die Blüte
möglicher Nahrungspflanzen (z.B. Wein oder Raps) so gut wie abgeschlossen, ebenso
die Blüte von Bäumen in den Windschutzstreifen, die sich im Marchfeld traditionell
zwischen den Kulturflächen befinden (HADATSCH et al. 2000). Mit den ersten
Herbizidspritzungen wird das Auflaufen von Ackerbeikräutern innerhalb der Kulturen
verhindert. Umso bedeutender wurde im Verlauf der Vegetationsperiode der ungestörte
Aufwuchs auf den Brachen hinsichtlich der Bereitstellung eines ganzsaisonalen
Nahrungsangebotes (WESTRICH 1997).
Mit Hilfe der Kanonischen Korrespondenzanalyse erfolgte eine weitere, detaillierte
Charakterisierung der drei untersuchten Altersklassen der Brachen (Kap. 5.4). Neben den
drei Altersklassen flossen dabei die erhobenen Flächenparameter Anzahl der
Blühpflanzen (species), Blütendeckung (flow cov) und die Blühfarben rot, blau, gelb und
weiß in die Ordination ein. Da es im Mai nur zu wenigen Tiersichtungen kam, konnte

32
keine aussagekräftige Ordination durchgeführt werden. Die CCAs von Juni und Juli
ergaben jeweils ein sehr deutliches Bild bezüglich der Ressourcenausstattung der jungen
und alten Brachen und der Nutzung durch Insekten. Sie untermauerten die Ergebnisse
der ANOVA.
Ein wichtiges Ergebnis war, dass keiner der erhobenen Flächenparameter positiv mit
mittelalten Brachen korreliert war. Sowohl im Juni als auch im Juli positionierten sich die
einflussreichen Flächenparameter (flow cov, species) deutlich entgegengesetzt zu den
mittelalten Brachen. Markant waren die positiven Korrelationen zwischen Honigbienen,
species (Blühpflanzenanzahl) und den Blühfarben weiß und gelb in alten Brachen im Juni.
Hier waren es insgesamt drei Flächenparameter, die einen Lebensraum innerhalb alter
Brachen charakterisierten, welcher besonders von Honigbienen gut genutzt werden
konnte. Im Juli boten alte Brachen eine gute Ressourcenausstattung in Form eines hohen
Blütendeckungsgrades, einer hohen Anzahl an Blühpflanzen und vielen roten Blüten. Sie
wurden von der Hummelart B. hortorum genutzt. Waren die jungen Brachen im Juni noch
zentrumsnahe angeordnet, erfolgte im Juli eine deutlichere Positionierung. In diesen
jungen Brachen fanden sich vorwiegend blaue Blüten und damit positiv korreliert die
Schwebfliegengattung Eristalis.
Auf die Ergebnisse der CCAs und die Bedeutung der einzelnen Flächenparameter für die
Bestäuber soll in den folgenden Kapiteln näher eingegangen werden.
6.2.1 Der Einfluss von Blütenfarben auf die Bestäuber
Im Juni war die Blühfarbe rot signifikant und hatte den größten Einfluss auf die
Hummelarten B. hortorum und B. terrestris. Noch deutlicher war die Korrelation zwischen
der langrüsseligen Hummelart B. hortorum und roten Blüten in alten Brachen im Juli. Zu
dieser Zeit blühten auf den Brachen unter anderem Wiesenklee (Trifolium pratense L.)
oder verschiedene Distelarten (Cirsium arvense, C. vulgare, Carduus acanthoides). Bei
diesen Arten handelt es sich um Blühpflanzen mit langen Kronröhren. Diese können
aufgrund der Anatomie ihrer Mundwerkzeuge fast ausschließlich von Hummeln genutzt
werden (VON HAGEN 1988). Auf die besondere Bedeutung von Hummeln zur Bestäubung
von Klee bzw. von Fabaceae wurde bereits 1885 von Charles Darwin hingewiesen, der
zur Steigerung der Flächenerträge bei Klee und Luzerne zu einem Import von
Hummelvölkern nach Neuseeland und Australien riet (VON HAGEN 1988). Ein deutlicher
Zusammenhang zwischen dem Vorhandensein von Blüten mit langen Kronröhren und der
Anzahl und Aktivität von Hummelarten konnte auch bei HEGLAND & BOEKE (2006)
nachgewiesen werden.
Ein interessanter, vegetationsökologischer Aspekt der CCAs beider Monate ist, dass sich

33
die Blühfarben rot und blau stets gegenüber positionieren. Einfach ausgedrückt bedeutet
dies, wo viele rote Blüten waren, fanden sich nur wenige blaue Blüten. Möglicherweise
liegt es daran, dass sich unter den neun betrachteten Flächen auch eingesäte Brachen
befanden. So wurde z.B. auf einigen, vorwiegend jungen Brachen die Saatluzerne
(Medicago sativa) gefunden, welche der Blühfarbe blau zugeordnet wurde. Rote
Blühpflanzen wiederum fanden sich als eingesäter Klee, als Futteresparsette (Onobrychis
viciifolia) oder auch als die ausdauernden und konkurrenzstarken Disteln. Letztere vor
allem im Juli in alten Brachen, wo sie sich im Rahmen der zweiten Besiedlungswelle
(Kapitel 6.1) bereits flächendeckend etablieren konnten. Dazu passend positionierte sich
in der Juli - CCA die langrüsselige Hummelart B. hortorum zu roten Blütenfarben in alten
Brachen. Von allen untersuchten Bestäubergruppen konnte diese Hummelart die Pflanzen
am besten nutzen. Auch in der Juni - CCA positioniert sich die B. hortorum in der Nähe
der Blühfarbe rot, wenn gleich nicht in dieser Deutlichkeit.
Wurden im Juni eher weiße und gelbe Blühpflanzen in alten Brachen nachgewiesen, gab
es im Juli einen Wechsel der Farben dieser Brache zu rot. Auch hier ist das Verblühen
von Frühlingsblühern und die Dominanz der Disteln ein möglicher Erklärungsgrund, ihre
Blüte begann auf den untersuchten Brachen erst im Juli (eigene Beobachtung). Dazu
konträr fanden sich blaue Blüten vorwiegend in jungen Brachen und waren für
Schwebfliegen der Gattung Eristalis attraktiv. Blaue Blüten waren auch im Juni für
Schwebfliegen nutzbar, hier war es jedoch die Gattung Sphaerophoria. Die bereits in der
Literatur als futtertolerant beschriebene Art Epishyrpus balteatus (RÖDER 1990)
positionierte sich dazu passend in beiden Monaten zentrumsnahe. Sie schien das
angebotene Blütenspektrum gleichmäßig auszunutzen und zeigte keine Farbpräferenzen.
Hinsichtlich der Farbpräferenzen bei Schwebfliegen gibt es kaum Publikationen, am
häufigsten wird eine Affinität zu gelben oder weißen Blüten beschrieben (SSYSMANK 1997).
Wie Honigbienen verfügen Schwebfliegen über ein trichromatisches Sehvermögen, was
bedeutet, dass Farben im roten Farbspektrum (630 - 700 nm) nicht wahrgenommen
werden. Bei den Eristalinen kommt es wie bei den Bienen aufgrund ihrer Größe oftmals
zu einer Präferenz für große, radiäre Blütenstände, da sie sich besser als „Landeplätze“
für die Insekten eignen (RÖDER 1990).
Im Juni und im Juli konnten keine klaren Korrelationen zwischen einem der Bestäuber und
gelben Blüten nachgewiesen werden. Im Juni waren die Honigbienen verstärkt auf weißen
Blüten zu finden, im Juli die Hummelart B. terrestris. Zur allgemeinen Bedeutung der
Blühfarben für die Bestäuber gilt es zu sagen, dass sich Honigbienen und adulte
Schwebfliegen auf der Suche nach Nahrungspflanzen, neben der Anzahl an verfügbaren
Blühpflanzen oder der Blütendeckung, auch an den Blühfarben orientieren (RUPPERT

34
1993). Eine hohe Bedeutung haben Farben für die Fernorientierung und das generelle
Auffinden potentieller Nahrungspflanzen (RUPPERT 1993). Durch die Blühfarben wird eine
Reizwirkung auf die Insekten erzeugt und eine Nahrungsquelle kann somit auch auf große
Distanz hin ausfindig gemacht werden (RÖDER 1990, RUPPERT 1993). Nimmt das
Nahrungsangebot lokal, also im unmittelbaren Aktionsradius der Bestäuber, ab, werden
mit Hilfe der Farben auch weiter entfernte Bestände lokalisiert und angeflogen (WEISS &
STETTMER 1991).
6.2.2 Der Einfluss von Blühpflanzenanzahl und Blütendeckung auf die Bestäuber
Die Nutzbarkeit der Brachen durch die Bestäuber hängt wesentlich von der Verfügbarkeit
und Menge an Nahrung ab. Diese qualitative und quantitative Ressourcenausstattung der
Brachen ließ sich am deutlichsten anhand der Flächenfaktoren Anzahl der Blühpflanzen
(species) und Blütendeckung (flow cov) darstellen. Hummeln haben einen hohen Bedarf
an Nektar und Pollen (VON HAGEN 1988), gleichzeitig ist für sie aber auch das
Vorhandensein von ungestörten Brutmöglichkeiten wichtig (HEGLAND & BOEKE 2006,
ALBRECHT et al. 2007). Auch für Honigbienen ist aufgrund der Bruttätigkeit der Bedarf an
Nektar groß, Schwebfliegen benötigen zudem eiweißreichen Pollen für die
Gonadenreifung (WEISS & STETTMER 1991).
Wie bereits in Kapitel 6.2 erwähnt, waren in der Juni - CCA die Honigbienen deutlich
positiv mit dem Flächenparameter species in alten Brachen korreliert. Im Juli konnte die
Hummelart B. hortorum dem Faktor species zugeordnet werden, es waren auch hier
wieder die alten Brachen, die eine hohe Anzahl an unterschiedlichen Blühpflanzen boten.
Zusätzlich dazu war die prozentuale Blütendeckung (flow cov) hoch. Bei der Analyse der
Anzahl der Blühpflanzen auf den Brachen konnte ebenfalls bestätigt werden, dass es die
alten Brachen waren, die in beiden Monaten die höchste Anzahl an Blühpflanzen hatten
(Kapitel 5.6, Abb. 11). Dies wirkte sich positiv auf die Anzahl der blütenbesuchenden
Insekten aus. Die Wichtigkeit eines guten Angebotes an Blühpflanzen für Bestäuber
wurde mehrfach anhand von Untersuchungen zur Bedeutung von angesäten Blühstreifen
(HAENKE et al. 2009, POTTS et al. 2009), Brachen (DECOURTYE et al. 2007) oder sogenannten
ökologischen Ausgleichsflächen (ALBRECHT et al. 2007) nachgewiesen.
Ebenso deutlich zeigten die CCAs von Juni und Juli sowie der Vergleich der Anzahl an
Blühpflanzen, dass die mittelalten Brachen über eine schlechte Ressourcenausstattung
verfügten und nicht von den Bestäubern genutzt werden konnten. Der Faktor species
korrelierte deutlich negativ mit mittelalten Brachen, im Juli waren es zudem der Faktor
flow cov sowie die Blühfarben rot, gelb und weiß.

35
Durch Regressionsmodelle zwischen den Bestäubern und Flächenfaktoren konnte der
hohe Einfluss der Parameter species und flow cov nochmals verdeutlicht werden.
Honigbienen und Schwebfliegen korrelierten im Mai positiv mit dem Faktor species,
Hummeln im Juni. Bei Schwebfliegen und Hummeln war außerdem der Faktor flow cov
auf den untersuchten Flächen einflussreich. Ein hoher Deckungsgrad an nutzbaren
Pflanzen lockte einerseits die Insekten an, andererseits verblieben sie dadurch länger auf
dieser Fläche (DECOURTYE et al. 2007) (vgl. auch Reizwirkung von Blühfarben, Kap. 6.2.1).
Der Nahrungsbedarf konnte sehr ökonomisch innerhalb kurzer Zeit, sowie auf
vergleichsweise kleiner Fläche gedeckt werden (WEISS & STETTMER 1991, HEGLAND &
BOEKE 2006). Bei einer geringen Dichte an Trachtpflanzen oder einer Verknappung des
Nahrungsangebotes müssen die Tiere größere Distanzen zurücklegen oder Suchflüge
durchführen (JAUKER et al. 2009). Honigbienen und Hummeln, also soziale Staatenbildner,
suchen in erster Linie in der unmittelbaren Umgebung ihres Baus nach Nahrung.
Diesbezüglich konnten Untersuchungen zu Flugweiten und Verteilung von Honigbienen
und Hummeln in Abhängigkeit von der Entfernung zu ihrem Habitat, eine deutliche
Abnahme der Sichtungen mit zunehmender Entfernung zum Bau nachweisen (JAUKER et
al. 2009). Finden diese Insekten jedoch in ihrem ummittelbaren Aktionsradius von rund
500 m um ihr Nest ausreichend Trachtpflanzen vor, verbleiben sie. Bei lokaler
Verknappung des Nahrungsangebotes sind bei Hummeln Flugweiten bis zu 2 km
beschrieben (RÖSELER 2002). Somit ist nicht nur die lokale Ressourcenverfügbarkeit der
Brache, sondern auch die der Umgebung für die Abundanzen der Insekten bedeutend.
Weiters ist die Verknüpfung und die räumliche Erreichbarkeit von potentiell
ressourcenreichen Habitaten für die Bestäuber wichtig (HEGLAND & BOEKE 2006, EBELING
et al. 2008, HAENKE et al. 2009, POTTS et al. 2009).
Die durchgeführten Regressionsanalysen zwischen den Bestäubern und den
Flächenparametern konnten den hohen Einfluss der Faktoren Blütendeckung und
Pflanzenarten auf die Abundanzen ebenfalls darstellen.
Honigbienen
Die Honigbienen zeigten nur im Mai Signifikanzen (Kap. 5.5.1, Tab. 5). Neben dem positiv
korrelierten Faktor species waren vor allem Umgebungsparameter (fields, houses)
signifikant und korrelierten deutlich negativ. Die Lokalisation ergiebiger Trachtquellen wird
innerhalb der Arbeiterinnen mit Hilfe des Schwänzeltanzes umgehend kommuniziert
(AKKERMANN 2004). Blütenreiche Vegetation kann somit effizient vom gesamten
Bienenvolk genutzt werden. Auch entspricht es gängiger Imkerpraxis, die Bienenstöcke zu
ergiebigen Blühpflanzenbeständen zu transportieren („Wandern“), um die Bestäubung

36
z.B. in Weingärten oder Obstkulturen sicher zu stellen (NÖ BIENENZUCHTGESETZ §6).
Dementsprechend werden diese Bienenstöcke eher in siedlungsfernen, blütenreichen
Gebieten aufgestellt. Somit kann das saisonale Angebot an Nektar und Pollen genutzt
werden und das Ergebnis sind die, je nach Blütenart und Jahreszeit unterschiedlichen
Honigsorten.
Die positive Korrelation zwischen Honigbienen und species aus dem Ergebnis der CCA
Juni konnte in der Regressionsanalyse nicht dargestellt werden.
Schwebfliegen
Bei den Schwebfliegen konnten in allen drei Monaten Ergebnisse gewonnen werden. Im
Untersuchungszeitraum waren sie die individuenreichste Gruppe, was auch damit
zusammenhing, dass hier eine Art (Episyrphus balteatus) und zwei Gattungen
(Sphaerophoria, Eristalis) zusammengefasst wurden. Im Gegensatz zur CCA wurde bei
der Regressionsanalyse kein Augenmerk auf den Einfluss der Flächen- oder
Umgebungsparameter auf die einzelnen Taxa gelegt. Diesbezüglich gibt es jedoch
Untersuchungen, die einen positiven Einfluss von Pflanzenartenzahlen, sowohl auf die
Artenvielfalt als auch auf die Abundanzen von Syrphiden nachwiesen (HEGLAND & BOEKE
2006, EBELING et al. 2008, MEYER et al. 2009, POTTS et al. 2009). In der vorliegenden
Untersuchung wurde bei der schrittweisen Regression deutlich, dass der Faktor flow cov
in allen drei Monaten eine höhere Erklärungskraft für die Abundanzen von Schwebfliegen
hatte als der Faktor species. Bei den Hummeln war es ebenfalls der Flächenparameter
flow cov, der im Juni als wichtigster Einflussfaktor für die Dichte auftrat. Ein hoher
Deckungsgrad an nutzbaren Pflanzen war für diese Insektengruppen wichtiger als eine
hohe Anzahl unterschiedlicher Pflanzenarten, die möglicherweise aufgrund
bestäuberanatomischer oder blütenmorphologischer Besonderheiten nicht optimal genutzt
werden konnten. Wie bereits im Kapitel Blütenfarben (Kap. 6.2.1) erwähnt, können nicht
alle Pflanzenarten gleichermaßen von den Bestäubern genutzt werden (VON HAGEN 1988).
War jedoch eine hohe Dichte einer nutzbaren Pflanze im Plot vorhanden, erklärte diese
aus oben bereits angeführten Gründen einen hohen Prozentsatz der Aktivität. Ähnliche
Ergebnisse konnten auch bei HEGLAND & BOEKE (2006) festgestellt werden. In dieser
Studie war die Frequenz der Blütenbesuche durch Insekten abhängig von der Dichte an
nutzbaren Blüten (flow cov) und der Faktor Pflanzenartenzahl (species) zeigte eine
negative Korrelation (HEGLAND & BOEKE 2006).
Interessant wiederum war, dass die Schwebfliegen nur im Mai positiv, in den Monaten
Juni und Juli jedoch negativ mit dem Faktor species korrelierten. Als mögliche Ursachen
sind hier wieder das Alter und der damit verbundene Sukzessionsprozess der Brachen zu

37
nennen (Kapitel 6.1). Nach dem Verblühen der Frühlingsblüher kam es im Juni und Juli in
den mittelalten Brachen zu einer zunehmenden Vergrasung und einer Abnahme an
Blühpflanzen (Kap. 5.6). Auch der Anstieg der Temperaturen, sowie eine Verschiebung
der Lokalität des Nahrungsangebotes können die Schwebfliegen in alternative Habitate,
wie Siedlungen oder Hausgärten drängen. Des Weiteren ist auch hier wieder die
Dominanz einzelner Blühpflanzen auf den Brachen zu nennen, da dadurch eine hohe
Blütendeckung bei niedriger Pflanzenartenzahl vorhanden war. Ein weiterer bedeutender
Einflussfaktor für das Vorhandensein von Schwebfliegen in einem Habitat betrifft die Brut.
Im Gegensatz zu Honigbienen und Hummeln sind Schwebfliegen bei der Reproduktion
wenig ortsgebunden (JAUKER et al. 2009). Die Oviposition erfolgt in für die Larvenstadien
passenden Mikrohabitaten, es bedarf keiner weiteren Brutpflege wie bei Honigbienen oder
Hummeln (JAUKER et al. 2009). Hierzu ist das Auftreten der Getreideblattläuse zu nennen,
da die Blattlauskolonien von Schwebfliegen mit aphidophagen Larven (Epishyrphus
balteatus, Sphaerophoria) als Eiablageplätze genutzt werden. Mehrfach wurden die
umfangreichen Wanderbewegungen von Schwebfliegen im Zusammenhang mit Eiablage
oder Nahrungssuche beschrieben (RUPPERT 1993, FRANK 1999, JAUKER et al. 2009). Dazu
passend erschien der Umgebungsfaktor fields erstmals im Juli von Bedeutung für die
Schwebfliegen (Kap. 6.2.3.).
Hummeln
Die Einflussfaktoren für die Abundanzen der Hummeln im Juli sind schwer zu
interpretieren. Es ergaben sich keine Erklärungen mit einem der einzelnen Parameter, in
der multiplen Regression hingegen wurde nur der Umgebungsparameter grassland
eliminiert. Neben den Faktoren flow cov und species waren auch temp, wind, fields und
houses signifikant (Kap. 5.5.3, Tab. 7). Hummeln wählen auf der Suche nach Nahrung
neben der Nähe zum Nest auch häufig eine Fläche aufgrund optischer Faktoren aus (VON
HAGEN 1988, HEGLAND & BOEKE 2006). Ein hoher Deckungsgrad und die damit verbundene
Signalwirkung der Farben der Blühpflanzen können Hummeln verstärkt anlocken
(HEGLAND & BOEKE 2006). In den CCAs von Juni und Juli konnte dies für die Blühfarben rot
und die Hummelarten B. terrestris und B. hortorum nachgewiesen werden.
Ebenfalls auffallend war, dass die ermittelten abiotischen Parameter wind und temp
erstmals einen Einfluss auf das Vorhandensein der Insekten zeigten. Die Anzahl der
Hummeln auf den Brachen verringerte sich mit steigenden Temperaturen. Hummeln sind
aufgrund der Physiologie ihrer Flugmuskulatur in der Lage, aktiv Wärme durch
Muskelzittern zu erzeugen und so auch bei niedrigen Tagestemperaturen auf
Nahrungssuche zu gehen (VON HAGEN 1988). Im Gegenzug dazu können sie

38
überschüssige Wärme über Flügeln oder Abdomen ableiten (VON HAGEN 1988). Durch
diese aktive Thermoregulation sind Hummeln relativ unabhängig, was die
Temperaturverhältnisse angeht. An sehr heißen Tagen jedoch besteht
Überhitzungsgefahr, da sie die durch Flugaktivität entstehende Wärme nicht ausreichend
an die Umgebung abgeben können (VON HAGEN 1988). Die für viele Hummeln kritische
Temperaturschwelle liegt bei ca. 30°C, wo hingegen trockenheitsliebende
Schwebfliegengattungen wie Sphaerophoria bei diesen Temperaturen keine Probleme
haben (VON HAGEN 1988, RÖDER 1990). Des Weiteren bleiben die Ergebnisse der
Regression bei den Hummeln schwer zu interpretieren, sind im Modell doch sechs von
sieben Einflussfaktoren signifikant.
6.2.3 Der Einfluss der strukturellen Ausstattung des Lebensraumes
In der vorliegenden Untersuchung der neun Brachen wurden neben den lokalen
Flächenparametern (flow cov, species, wind, temp) auch die umgebenden Strukturen in
Form der Umgebungsparameter (grassland, fields, houses) erhoben.
Neben der bereits diskutierten lokalen Ausstattung der Flächen ist auch eine vielfältige,
strukturreiche Landschaft ein weiterer wesentlicher Faktor für das Vorhandensein von
Syrphiden und Hummeln (SSYMANK 1997, KEVAN 1999). In der durchgeführten
Regressionsanalyse zwischen den Bestäubern und Umgebungsparametern konnten
diesbezüglich vor allem bei Schwebfliegen Ergebnisse gewonnen werden (Kap. 5.5.2,
Tab. 6). In allen drei Monaten waren es die Umgebungsparameter (grassland, fields),
welche sich auf die Dichte der Insekten auswirkten. Untersuchungen zu
Wanderbewegungen von Schwebfliegen ergaben, dass es vor allem zum Zeitpunkt der
Milchreife von Getreide (Ende Juni bis Mitte Juli) und dem zeitgleichen Auftreten großer
Blattlauskolonien zu einem starken Anstieg der Abundanzen direkt in den Feldern kam
(HAENKE et al. 2009). Im Mai profitierten die Tiere vermutlich noch von den blütenreichen
Brachen, aber zur Zeit der Eiablage wurden jene Flächen aufgesucht, die den Larven als
Habitate dienen konnten. Dazu passend verlor der Faktor grassland zwischen Mai und
Juni an Bedeutung für die Schwebfliegen und war im Juli nicht mehr signifikant. Somit war
eine wichtige Vorraussetzung für das Vorhandensein von Schwebfliegen nicht nur ein
ausreichendes Nahrungsangebot auf den Brachen selber, auch das Vorhandensein von
geeigneten Larvalhabitaten in der unmittelbaren Umgebung war entscheidend. Aufgrund
dieser vielfältigen Bedürfnisse sind Schwebfliegen sehr mobil und verlassen
ressourcenarme Flächen rascher als Honigbienen oder Hummeln, die mehr an ihr
unmittelbares Habitat gebunden sind (JAUKER et al. 2009). Zeigen Untersuchungen zur
Dispersion von Schwebfliegen innerhalb agrarisch dominierter Landschaften noch eine

39
gewisse Mobilität dieser Bestäubergruppe an, so sind die Flugweiten bei Staaten
bildenden Hummeln und Honigbienen deutlich geringer und nehmen mit steigender
Distanz zum Nest signifikant ab (JAUKER et al. 2009). Besonders innerhalb intensiv
landwirtschaftlich genutzter Gebiete sind somit ressourcenreiche Strukturen von großer
Wichtigkeit für Honigbienen und Hummeln (HAENKE et al. 2009). Dies unterstreicht die
Bedeutung von störungsarmen und ressourcenreichen Rückzugsgebieten wie
Bracheflächen, Saumstrukturen oder Windschutzstreifen, und deren Vernetzung
untereinander für die Bestäuber. Denn nur so ist die Erreichbarkeit von Nahrungspflanzen
vor allem für eher ortstreue Insekten wie Honigbienen und Hummeln gewährleistet
(KEVAN 1999). Dazu passend konnte KOHLER et al. (2007) beobachten, dass in
landwirtschaftlich intensiv genutzten Gebieten die Anzahl der Honigbienen bereits ab
einer Entfernung von 25m zu Bracheflächen deutlich abnahm, die Anzahl der
Schwebfliegen nahm ab 125m Entfernung signifikant ab.
In der vorliegenden Untersuchung deutete neben den Schwebfliegen auch die Analyse
der Dichte der Hummeln auf ähnlich gelagerte Ergebnisse hin. Im Juni waren sechs von
insgesamt sieben Einflussfaktoren signifikant. Die Ansprüche der Hummeln waren nicht
nur flächenspezifisch (hohe Blütendeckung und viele Blühpflanzen), sondern auch
umgebungsspezifisch (hoher Anteil fields, geringer Anteil houses).
Kontrastreiche und mit vielfältigen Mikro- und Makrohabitaten ausgestattete Areale zeigen
nachweislich einen positiven Effekt auf die Abundanzen unterschiedlicher Bestäuber
(SJODIN et al. 2008, MEYER et al. 2009,). Hierzu sind nicht nur Brachen (angesät oder
selbstbegrünt) zu zählen. Auch andere vernetzende Landschaftselemente wie
Windschutzstreifen, Hecken oder die besonders in den letzen Jahren geförderten
Blühstreifen neben Äckern beeinflussen die Aktivität und das Vorhandensein von
Nützlingen (JAUKER et al. 2009). Dabei sind aber auch die unterschiedlichen Ansprüche der
Insekten wie eine ausreichende Anzahl an Trachtpflanzen, Freiheit von mechanischen
oder sonstigen Störungen, Angebot an Nestmöglichkeiten oder Larvalhabitaten und die
landschaftliche Verknüpfung derartiger Areale zu beachten (HAENKE et al. 2009, JAUKER et
al. 2009, POTTS et al. 2009).

40
7 Zusammenfassung
Blütenbesuchende Insekten wie Honigbienen, Hummeln oder Schwebfliegen spielen eine
wichtige Rolle in unseren Agrarökosystem da sie effiziente Bestäuber unterschiedlichster
Pflanzen sind. Neben Wildpflanzen oder Bäumen werden durch sie auch viele
landwirtschaftlich bedeutende Kulturpflanzen bestäubt. Aphidophage Schwebfliegenlarven
sind in der biologischen Schädlingskontrolle unverzichtbar. Durch die moderne, nach
ökonomischen Zielen ausgerichtete Landwirtschaft werden diese Bestäuber von
unterschiedlichsten Gefahren (Insektizide, Flurbereinigung, Prädatoren, eingeschleppte
Krankheiten) dezimiert, die Anzahl und Vielfalt der Bestäuber ist besonders in den letzten
Jahren deutlich gesunken. Die Bedürfnisse dieser Insekten sind vielfältig. Sie brauchen
strukturreiche Landschaften für Verstecke und Brut sowie eine ausreichende Versorgung
mit Trachtpflanzen, welche über die gesamte Vegetationsperiode hinweg Nektar und
Pollen bieten.
In den Monaten Mai, Juni und Juli der Vegetationsperiode 2009 wurde auf neun Brachen
im Marchfeld, Niederösterreich, eine Zählung dieser drei Bestäubergruppen durchgeführt.
Dazu wurden innerhalb jeder Brachen fünf 1x1m2 große Plots abgesteckt und darin die
Aktivitätsdichte der Insekten mittels Blütenbesuchsbeobachtung erhoben.
Diese Brachen konnten in drei Altersklassen (jung: 3-4 Jahre, mittelalt: 7-9 Jahre und alt:
13-16 Jahre) eingeteilt werden. Es wurde der mögliche Einfluss des Alters der Brache auf
die Aktivitätsdichte von Honigbienen, Schwebfliegen und Hummeln untersucht. Neben
dem Alter der Brachen wurden auch die Einflüsse von kleinstandörtlichen Faktoren und
die des lokalen Umfeldes im Umkreis von jeweils 500 Metern um die Brache untersucht.
Zu diesem Zweck wurden einerseits sog. Flächenparameter (flower coverage, number of
plant species, temperature, wind sowie die Blühfarben rot, gelb, weiß, blau) und
andererseits sog. Umgebungsparameter (grassland, fields, houses) erhoben. Die
gewonnenen Daten wurden (nach log- Transformation) mit einer zweifaktoriellen
Varianzanalyse (ANOVA), einer Kanonischen Korrespondenzanalyse (CCA) sowie
einfachen und multiplen Regressionen (mit Rückwertsmethode) analysiert.
Im Vergleich der Altersstufen zeigte sich, dass vorwiegend junge und alte Brachen von
den Insekten genutzt wurden. Im Juni konnten in mittelalten Brachen signifikant weniger
Honigbienen, Schwebfliegen und Hummeln gezählt werden als in jungen oder alten
Brachen. Im Juli waren Honigbienen und Hummeln in signifikant niedrigeren Dichten in
Brachen mittleren Alters zu finden. Auch die Anzahl an Blühpflanzen war in diesen beiden

41
Monaten in mittelalten Brachen signifikant niedriger und führte vermutlich zu den
geringeren Dichten an Bestäubern. Die höchste Anzahl an Blühpflanzen wurde jeweils in
alten Brachen nachgewiesen. Die Kanonische Korrespondenzanalyse der Monate Juni
und Juli unterstrich die Ergebnisse der ANOVA und zeigte zudem eine negative
Korrelation der mittelalten Brachen mit den Flächenparametern flower coverage und plant
species sowie auch den Tierguppen an. Für die Abundanzen von Honigbienen und
Hummeln waren in erster Linie lokale Flächenparameter entscheidend. So waren vor
allem die Faktoren Blütendeckung (flower coverage) und Blühpflanzenanzahl (plant
species) signifikant, bei Schwebfliegen waren es neben diesen beiden
Flächenparametern auch die Umgebungsparameter grassland und fields, die einen
signifikanten Einfluss auf die Aktivitätsdichte ausübten. Die lokalen Wetterbedingungen
(wind und temperature) waren einzig bei der Gruppe der Hummeln signifikant.

42
8 Literaturangaben
AKKERMANN, R. (2004): Honigbienen. Apis mellifera L. Naturschutzbiologie,
Bienenbotanik, Bienenweide. Runge, Cloppenburg.
ALBRECHT, M., DUELLI, P., MULLER, C., KLEIJN, D. & SCHMID, B. (2007): The Swiss agri-
environment scheme enhances pollinator diversity and plant reproductive success
in nearby intensively managed farmland. Journal of Applied Ecology 44: 813- 822.
BACKHAUS, K. (2000): Multivariate Analysemethoden: eine anwendungsorientierte
Einführung. Springer, Berlin.
BARTH, F. G. (1991): Insects and Flowers, The Biology of a Partnership. Princeton
University Press, Princeton.
BERNER, U. (1979): Die Bienenweide. 3. Auflage, Ulmer, Stuttgart.
BRAUN- BLANQUET, J. (1964): Pflanzensoziologie. 3. Auflage, Springer, Wien.
EBELING, A., KLEIN A.M., SCHUMACHER, J., WEISSER, W.W. & TSCHARNTKE, T. (2008): How
does plant richness affect pollinator richness and temporal stability of flower visits?
Oikos 117: 1808- 1815.
DECOURTYE, A., LECOMPTE, P., PIERRE, J., CHAUZAT, MP. & THIEBEAU, P. (2007): Advantages
for farmers and beekeepers of introducing flowering fallows in field crop zones.
Cahiers Agricultures 16: 213- 218.
EXLER, D. (2005): Acker- Brachen im Dienste des Natur- und Grundwasserschutzes.
Diplomarbeit Universität für Bodenkultur, Wien.
FISCHER, M., OSWALD, K. & ADLER W. (2008): Exkursionsflora für Österreich, Liechtenstein,
Südtirol. Land Oberösterreich, Linz.
FRANK, T. (1999): Density of adult hoverflies (Dipt., Syrphidae) in sown weed strips and
adjacent fields. Journal of Applied Entomology 123: 351- 355.

43
FUSSELL, M. & CORBET, A. S. (1992): Flower usage by bumble- bees: a basis for forage
plant management. Journal of Applied Ecology 29: 451- 465.
GREILER, H.-J. (1993): Insektengesellschaften auf selbstbegrünten und eingesäten
Ackerbrachen. Haupt, Bern.
HEGLAND, S.G. & BOEKE, L. (2006): Relationships between the density and diversity of floral
resources and flower visitor activity in a temperate grassland community.
Ecological Entomology 31: 532- 538.
HADATSCH, S., KRATOCHVIL, R., VABITSCH, A., FREYER B. & GÖTZ, B. (2000): Biologische
Landwirtschaft im Marchfeld, Potentiale zur Entlastung des Natur- und
Landschaftshaushaltes. Umweltbundesamt, Wien.
HAENKE, S., SCHEID, B., SCHAEFER, M., TSCHARNTKE, T. & THIES, C. (2009): Increasing
syrphid fly diversity and density in sown flower strips within simple vs. complex
landscapes. Journal of Applied Ecology 46: 1106- 1114.
HARFLINGER, O. (1993): Das Klima des Marchfeldes. Universitätsverlag Wagner,
Innsbruck.
HENDRICKX, F., MAELFAIT, J.P., VAN WINGERDEN, W. et al. (2007): How landscape structure,
land- use intensity and habitat diversity affect components of total arthropod
diversity in agricultural landscapes. Journal of Applied Ecology 44: 340- 351.
HOLZNER, W. (2001): Ackerbrachen- Natur oder Unkraut- Infektionsherd? Dokumentation
der Fachtagung Ackerbrachen: Ackerbrachen- Flächenutzung mit Zukunft?
Universität für Bodenkultur, Facultas, Wien.
JAUKER, F., DIEKOETTER, T., SCHWARZBACH, F. & WOLTERS, V. (2009): Pollinators dispersal in
an agricultural matrix: opposing responses of wild bees and hoverflies to
landscape structure and distance form main habitat. Landscape Ecology 24: 547-
555.
KEVAN, P. G. (1999): Pollinators as bioindicators of the state of the environment: species,
activity and diversity. Acriculture Ecosystem & Environment 74: 373- 393.

44
KLEIJN, D. & VAN LANGEVELDE, F. (2006): Interacting effects of landscape context and
habitat quality on flower visiting insects in agricultural landscapes. Basic and
Applied Ecology 7: 201- 214.
KOHLER, F., VAN KLING, R., NOORDIJK, J. & KLEIJN, D. (2007): The influence of nature
reserves on hoverflies and bees in agricultural areas (Diptera: Syrphidae:
Hymenoptera: Aphidae s.l.). Entomologische Berichte 67: 187- 192.
KRIEG, A. & FRANZ, J.M. (1989): Lehrbuch der Biologischen Schädlingsbekämpfung. Paul
Parey Verlag, Berlin und Hamburg.
MEYER, B., JAUKER, F. & STEFFAN- DEWENTER, I. (2009): Contrasting resource- dependent
responses of hoverfly richness and density to landscape structure. Basic and
Applied Ecology 10: 178- 186.
POTTS, S.G., WOODCOCK, B.A., ROBERTS, S.P.M., TSCHEULIN, T., PILGIRM, E.S., BROWN, V.K. &
TALLOWIN J.R. (2009): Enhancing pollinator biodiversity in intensive grasslands.
Journal of Applied Ecology 46: 369- 379.
REICHHOFF- RIEHM, H. (1984): Insekten. Mosaik Verlag, München.
RÖDER, G. (1990): Biologie der Schwebfliegen Deutschlands, Diptera: Syrphidae. Erna
Bauer Verlag, Keltern- Weiler.
RÖSELER, P.-F. (2001): Der Hummelgarten, Lebensraum und Biologie der Hummeln. Triga
Verlag, Gelnhausen.
RUPPERT, V. (1993): Einfluss blütenreicher Feldrandstrukturen auf die Dichte
blütenbesuchender Nutzinsekten insbesondere der Syrphidae (Diptera:
Syrphidae). Haupt, Bern.
RUTTNER, F. (2003): Naturgeschichte der Honigbienen, Biologie, Sozialleben, Arten und
Verbreitung. Kosmos, Stuttgart.
SACHS, L. (1997): Angewandte Statistik, Anwendung statistischer Methoden. Springer,
Berlin.

45
SANDOR, B., MEINERS, S. & CADENASSO, M. (2009): Plant colonization windows in a mesic
old field succession. Applied Vegetation Sience 2: 205-212.
SCHMIDT, W. (1981): Natural and experimental succession on fallow land. Goltze,
Göttingen.
SJODIN, N.E., BENGTSONN. J. & EKBOM, B. (2008): The influence of grazing intensity and
landscape composition on the divesity and abundance of flower- visiting insects.
Journal of Applied Ecology 45: 763- 772.
SSYMANK, A. (1997): Blütenökologische Aspekte zur Ackerwildkrautflora, aufgezeigt am
Beispiel der Schwebfliegen (Diptera, Syrphidae). Bundesamt für Naturschutz,
Bonn.
STENITZER, E. & HOESCH J. (2005): Grundwasserneubildung im Marchfeld,
Lysimeteruntersuchung und Modellrechnung. 11. Gumpensteiner
Lysimetertagung, Raumberg Gumpenstein.
TACK, K.- J. (1983): Merkblatt der Arbeitsgruppe Bienenforschung an der FU Berlin:
Hummeln- Blütenbesuchende Insekten. Informationen 4, Berlin.
TER BRAAK, C. F. J. (1988): Canoco – a Fortran program for canonical community
ordination by partial detrendet canonical correspondence analysis, principle
component analysis and redundancy analysis. Technical report, TNO Inst.,
Wageningen, Netherlands.
TRAXLER, A., MINARZ, E., HÖTTINGER, H., PENNERSTOFER, J., SCHMATZBERGER, A., BANKO, G.,
PLACER, K., HADROBOLEC, M. & GAUGITSCH, H. (2005): Biodiversitäts- Hotspots der
Agrarlandschaft als Eckpfeiler für Risikoabschätzung und Monitoring von GVO.
Eigenpublikation, Umweltbundesamt Wien.
VON HAGEN, E. (1988): Hummeln bestimmen, ansiedeln, vermehren, schützen. Neumann-
Neudam, Melsungen.
WEISS, E. & STETTMER, C. (1991): Unkräuter in der Agrarlandschaft locken
blütenbesuchende Nutzinsekten an. Haupt, Bern.

46
WESTRICH, P. (1997): Zur Bedeutung des Ackersenf (Sinapsis arvensis L.) als
Nahrungsquelle von Wildbienen (Hymenoptera, Apidae). Bayrisches Landesamt für
Umwelt 142, s.l.
Internetquellen
GOOGLE MAPS ÖSTERREICH BETA: http://maps.google.at/
KARTE UND ROUTE: http://www.herold.at/routenplaner/
KLIMADATEN VON ÖSTERREICH: http://www.zamg.ac.at/fix/klima/oe71-
00/klima2000/klimadaten_oesterreich_1971_frame1.htm (03.08.2010)
LUFTBILDER: KARTEN UND GEOINFORMATION NIEDERÖSTERREICH:
http://www.intermap1.noel.gv.at/webgisatlas/(S(1ryfbo45gnariy45ftl5enfx))/init.aspx?karte
=atlas_bh
MASKE, P. & HEDERER, M. (2008): Forschung über Bienensterben.
http://www.cbgnetwork.org/2757.html, (16.05.2010).
NIEDERÖSTERREICHISCHES BIENENZUCHTGESETZ: http://www.imker-
klosterneuburg.at/content/view/31/41/ (01.08.2010).
ÖPUL 2007: Sonderrichtlinie des Bundesministers für Land- und Forstwirtschaft, Umwelt
und Wasserwirtschaft (BMLFUW) für das Österreichische Programm zur Förderung einer
umweltgerechten, extensiven und den natürlichen Lebensraum schützenden
Landwirtschaft.
http://www.ama.at/Portal.Node/ama/public?gentics.rm=PCP&gentics.pm=gti_full&p.conte
ntid=10008.47092&SRL_O4_20100527.pdf (15.09.2010).
ROUTENPLANER UND KARTEN: http://www.viamichelin.at/
WIRTSCHAFTSLEISTUNG DER HONIGBIENE: N.N.: http://www.proplanta.de/Agrar-
Nachrichten/agrar_news_themen.php?SITEID=1140008702&Fu1 (23.05.2009).
47
Anhang I
Namen und Lokalisation der neun Brachen im Marchfeld.
Rot: junge Brachen, Grün: mittelalte Brachen, Blau: alte Brachen
(Quelle: www.noe.gv.at, Modifiziert, Janet Wissuwa)

48
Brache Schönfeld Lassee (SFL j, m)
Junge (rosa) und mittelalte (rot) Brache.
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: www.noe.gv.at, Karten und Geoinformation in Niederösterreich, modifiziert)
49
Brache Gänserndorf (GD j)
Junge Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: googlemaps: http://maps.google.at/, modifiziert)

50
Brache Gerasdorf (GED)
Junge Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: Herold, Karte und Route: http://www.herold.at/routenplaner/, modifiziert)
51
Brache Bockfließ (BF)
Mittelalte Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: google maps: http://maps.google.at/, modifiziert)

52
Brache Untersiebenbrunn (USB)
Mittelalte Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle:google maps: http://maps.google.at/, modifiziert)

53
Brache Gänserndorf (GD)
Alte Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: Herold Karte und Route: http://www.herold.at/routenplaner/, modifiziert)

54
Brache Groß Engersdorf (GGD)
Alte Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: Karten und Routenplaner: http://viamichelin.at , modifiziert)
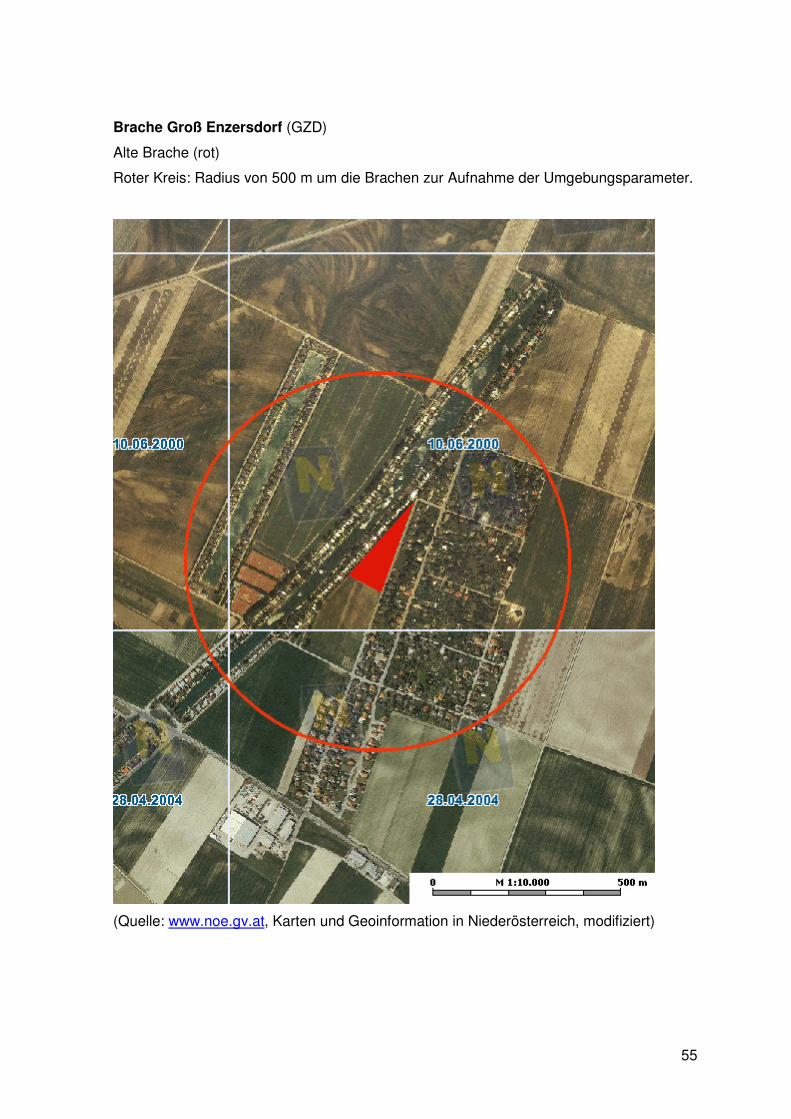
55
Brache Groß Enzersdorf (GZD)
Alte Brache (rot)
Roter Kreis: Radius von 500 m um die Brachen zur Aufnahme der Umgebungsparameter.
(Quelle: www.noe.gv.at, Karten und Geoinformation in Niederösterreich, modifiziert)
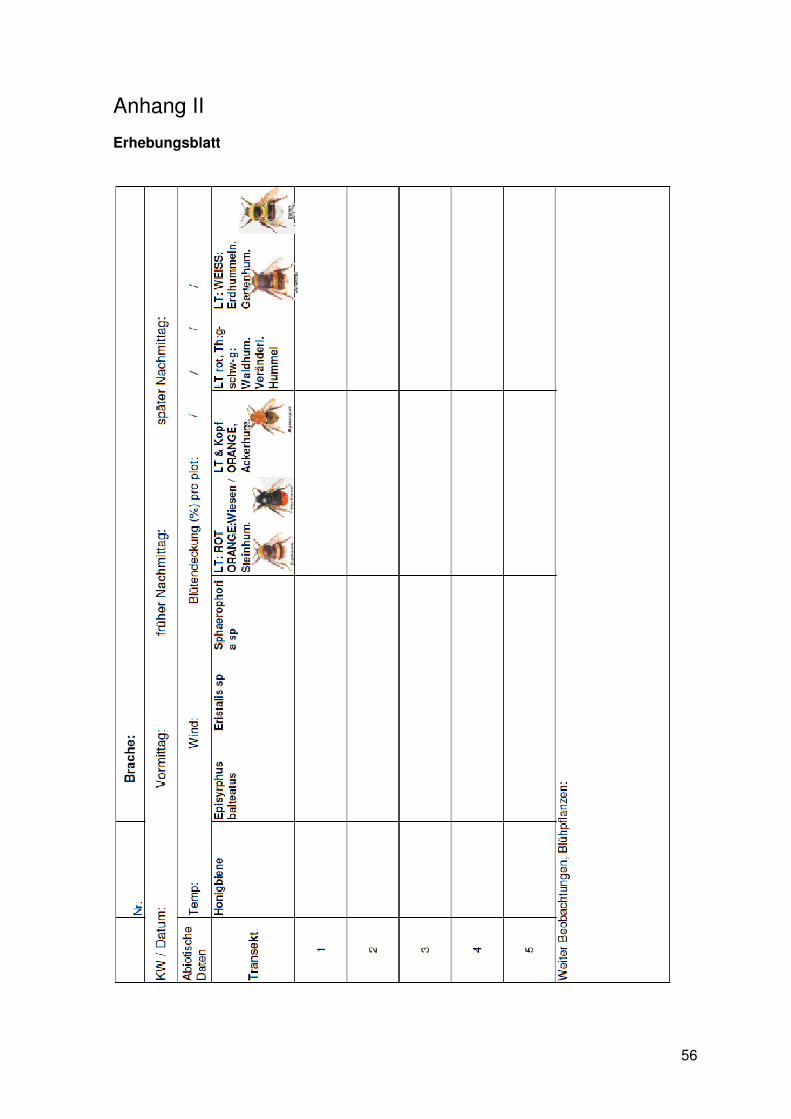
56
Anhang II
Erhebungsblatt

57
Anhang III
Liste der vorgefundenen Blühpflanzen in den neuen Brachen (ohne Gräser, Moose,
Stauden)
Achillea millefolium Medicago sativa, M. lupulina
Anthemis austriaca Melilotus officinalis
Anthriscus caucalis Onobrychis viciifolia
Arenaria serpyllifolia Papaver rhoeas
Ballota nigra Reseda lutea
Capsella bursa- pastoris Senecio vernalis
Carduus acanthoides,C. crispus Silene latifolia
Cirsium arvense, C. vulgare Solidago gigantea
Consolida regalis Sonchus oleraceus
Convolvulus arvensis Stellaria media
Crepis setosa , C. biennis Sysymbrium altissimum, S. loeselii
Daucus carota Sysymbrium orientale
Echium vulgare Taraxacum officinale
Erigeron annuus Tragopogon pratensis
Falcaria vulgaris Trifolium campestre, T. pratense, T. repens,
Fallopia vulgaris Tripleurospermum perforatum
Galium aparine, G. mollugo, G. spurium Veronica arvensis, V. persica
Geum urbanum Vicia arvensis, V. angustifolia, V. pannonica,
Lamium aplexicaule Vicia villosa
Leontodon hispidus Viola arvensis
Lotus corniculatus
(Vegetationsaufnahmen 2009: durchgeführt und mit freundlicher Genehmigung von Janet
Wissuwa)

58
9 Danksagung
An dieser Stelle möchte ich mich bei meinem Betreuer Prof. Dr. Thomas Frank sehr herzlich
für die Vergabe des Themas sowie für die stets hilfreiche Betreuung und Unterstützung
bedanken.
Bei Dr. Jörg Salamon möchte ich mich für seine Geduld sowie für die Hilfe bei den
Auswertungen bedanken.
Mein Dank richtet sich an Janet Wissuwa, mit der ich viele Stunden auf den Brachen
verbringen durfte. Für die netten Konversationen ebenso wie für die große Hilfe in
botanischen Belangen.
Bei meinem Mann, Dipl. Ing. Helmut Mayrhofer, möchte ich mich für die unglaubliche Stütze
und Hilfe, sowie den unbezahlbaren „technical support“ während der gesamten Studienzeit
danken. Auch für dich waren es oft schwierige Zeiten, in denen du mir aber stets mit
Vertrauen in meinem Weg begegnet bist. Vielen Dank!

59
Lebenslauf
Persönliche Daten Karin Maria Kastner, Bakk. Techn.
Neustiftgasse 86/1/10
1070 Wien
+43/650/4050798
geboren am 07.01.1976 in Haslach, OÖ.
Röm./kath.
Familienstand Lebensgemeinschaft
Schulbildung 1982-1986: Volksschule in St. Stefan
1986-1990: Hauptschule in Helfenberg
1990-1992: Landwirtschaftliche Berufs- und
Fachschule
1992-1993: Fachschule für Sozialberufe in Linz
1993-1996: Krankenpflegeschule am
Allgemeinen Öffentlichen Landeskrankenhaus
Rohrbach
2001: Berufsreifeprüfung am BFI Wien
2002: Sonderausbildung für Anästhesie- und
Intensivpflege

60
2004 – 2008: Bachelorstudium „Umwelt- und
Bioressourcenmanagement“ an der Universität
für Bodenkultur in Wien
Seit Februar 2008: Masterstudium
Agrarwissenschaften: Agrarbiologie
In Ausbildung: Abfallbeauftragte gemäß § 11
AWG 2002
Fremdsprachen Englisch in Wort und Schrift,
Italienisch (Grundkenntnisse)
Fähigkeiten sehr gute PC Kenntnisse (MS Office, GIS
Grundkenntnisse)
Führerschein A, B, F
Bisherige Tätigkeit 1996 bis 2006: Betriebsführung
landwirtschaftlicher Betrieb
Seit Oktober 1996: beschäftigt als DGKS am
Krankenhaus der Barmherzigen Brüder in Wien,
Intensivstation, Teilzeitbeschäftigung während
des Studiums.
Copyright © 2022 FDOKUMEN




















