
PLANT AND MICROBIAL CONTROLS ON NITROGEN RETENTION AND LOSS IN A HUMID TROPICAL FOREST
Upload
independentCategory
view
0download
0
Plant and Soil (2005) 268: 61–74 © Springer 2005DOI 10.1007/s11104-004-0227-2
Nitrogen use efficiency of monoculture and hedgerow intercropping in thehumid tropics
Edwin C. Rowe1,2,5, Meine Van Noordwijk3, Didik Suprayogo4 & Georg Cadisch1
1Department of Agricultural Sciences, Imperial College at Wye, University of London, Wye, Ashford, TN25 5AH,UK. 2Current address: Plant Production Systems, Wageningen University, P.O. Box 430, 6700 AK Wageningen,The Netherlands. 3ICRAF – SE ASIA, P.O. Box 161, Bogor 16001, Indonesia. 4Fakultas Pertanian, Jurusan Tanah,Universitas Brawijaya, Jalan Veteran, Malang, Indonesia. 5Corresponding author∗
Received 5 January 2004. Accepted in revised form 26 April 2004
Key words: agroforestry, competition, complementarity, isotope, subsoil, uptake
Abstract
The design of productive and efficient intercropping systems depends on achieving complementarity between com-ponent species’ resource capture niches. Spatiotemporal patterns of capture and use of pruning and urea nitrogen(N) by trees and intercrops were elucidated by isotopic tracing, and consequences for nitrogen use efficiency wereexamined. During the first cropping season after applying urea-15N, maize accounted for most of the plant 15N re-covery in Peltophorum dasyrrachis (33.5%) and Gliricidia sepium (22.3%) hedgerow intercropping systems. Maizeyield was greatest in monoculture, and maize in monoculture also recovered a greater proportion of urea 15N (42%)than intercropped maize. Nitrogen recovery during active crop growth will not be increased by hedgerow inter-cropping if hedgerows adversely affect crop growth through competition for other resources. However, hedgerowsrecovered substantial amounts of 15N during both cropping cycles (e.g. a total of 13–22%), showing evidence ofspatio-temporal complementarity with crops in the spatial distribution of roots and the temporal distribution ofN uptake. The degree of complementarity was species-specific, showing the importance of selecting appropriatetrees for simultaneous agroforestry. After the first cropping season 17–34% of 15N applied was unaccounted forin the plant-soil system. Urea and prunings N were recovered by hedgerows in similar amounts. By the end ofthe second (groundnut) cropping cycle, total plant 15N recovery was similar in all cropping systems. Less N wastaken up by the maize crop from applications of labelled prunings (5–7%) than from labelled urea (22–34%), butthe second crop recovered similar amounts from these two sources, implying that prunings N is more persistentthan urea N. More 15N was recovered by the downslope hedgerow than the upslope hedgerow, demonstrating theinterception of laterally flowing N by hedgerows.
Introduction
Nitrogen management is particularly difficult on soilswhere plant rooting depth is constrained by aluminiumtoxicity, a common problem in the Ultisols whichcover 11% of the tropics (Sanchez, 1976). Hedgerowintercropping has been recommended for improvingagronomic nitrogen use efficiency (NUE) on stronglyleached soils. The labour requirements for frequentpruning to manage competition between trees and cropplants are often a constraint to farm-level applica-
∗FAX No: +31-317-484-892. E-mail: [email protected]
tion, but hedgerow intercropping can still be used asa model system for testing our understanding of theprocesses governing efficiency in any perennial inter-cropping system. The agronomic efficiency of use of amaterial may be simply defined as the ratio of outputsor yields to inputs (Huxley, 1999). The definition ofNUE that is used here is the mass of N within the cropplant per mass of N applied. Nitrogen taken up by treesin agroforestry systems may be returned to the soil andused by subsequent crops, or may be considered partof the output if these trees have direct value as timber,fodder or fruit trees.

62
The use efficiency of N in an alley cropping agro-forestry system, relative to that in a crop monoculture,depends on a variety of interactions. Trees affectN extraction rates both through their own uptake andthrough influencing crop growth and N uptake. Treesmay also affect rates of N loss, for example by increas-ing transpiration and thus reducing water throughput,or by increasing infiltration rates or the proportion ofpreferential flow (Suprayogo et al., 2002). The qualityof tree residues affects N supply dynamics to the soil(Handayanto et al., 1994). In systems where trees areprimarily included for their soil fertility benefit, bene-fits will occur only when the trees acquire resourcesthat the crop would not otherwise acquire (Cannellet al., 1996). Thus the success of the system dependson limiting competition for resources and increasingthe temporal and spatial complementarity of resourcecapture. This may be achieved in tree-crop associa-tions if tree roots are active deeper in the soil thancrop roots (Young, 1986). Deep nutrient uptake bytrees is important where reserves of nutrients occur atdepth because of weathering or prior leaching, or iftrees intercept N that would otherwise be leached (the‘safety-net’ hypothesis; van Noordwijk et al., 1996).
Isotopic labeling of materials allows the fate ofnutrients applied to agroforestry systems to be mon-itored. Recent studies have demonstrated differencesin vertical distribution of root activity between trees(Lehmann et al., 2001), and between trees and associ-ated crops (Rowe et al., 2001). Surface applications of15N have also been used to demonstrate strong compe-tition for N by Erythrina indica trees used as supportfor Piper nigrum vines (Wahid et al., 2004). Recover-ies of 15N from surface applications made across thewidth of the alley in hedgerow intercropping systemsreflect overall NUE (Rao and Shinde, 1990). Van-Lauwe et al. (1998) reported 15N budgets followingapplication of high quality Leucaena and low quality(i.e. with higher C:N ratio and greater lignin content)Dactyladenia prunings to alley cropping systems. Lit-tle of the applied N (< 10%) was taken up by themaize crop in either case. Recovery by the hedgerowswas greater (35% of that applied, over 858 days) forthe higher quality Leucaena prunings than for the lowquality Dactyladenia (7.1%). Xu et al. (1993) alsofound that the amount of 15N recovered by maize afterapplication of Leucaena prunings was small (5% after52 days) in comparison with recovery of 15N labelledammonium fertilizer (50%). Giller and Cadisch (1995)found similar low residue recoveries and concludedthat this was in many cases due to inefficient utiliza-
tion in space and time of the N released from plantresidues.
In the current study, 15N labelled materials wereapplied to a monocrop system, and to intercroppingsystems with hedgerows of Peltophorum dasyrrhachis(Miq.) Kurz and/or Gliricidia sepium L.. The locationof 15N derived from these materials was monitoredby following the size of plant and soil 15N poolsover two cropping seasons. The objectives were toexplore the dynamics of N availability, leaching andplant uptake within the systems, and to assess overallNUE. We hypothesised (H1) that hedgerow intercrop-ping systems use N more efficiently than monocropsystems, and (H2) that P. dasyrrachis hedgerow in-tercropping systems are more efficient in their use ofN than G. sepium hedgerow intercropping systems,since P. dasyrrachis root systems are spatially dis-tinct from crop root systems (Rowe et al., 2001) andthus have more opportunity to intercept N before it isleached. The rapid mineralisation of high quality (lowC:N and low polyphenol content) G. sepium pruningscompared with low quality (high C:N and polyphenol)P. dasyrrachis prunings (Handayanto et al., 1994) washypothesised (H3) to lead to greater losses of pruningsN from G. sepium systems, and mixing P. dasyrrachisprunings with G. sepium prunings was expected toslow the release of N from the G. sepium prunings andincrease overall NUE (H4).
Materials and methods
Experimental site
The experiment was carried out at the Biological Man-agement of Soil Fertility (BMSF) project site of Uni-versitas Brawijaya/ICRAF/Wye College/PT BungaMayang, near Karta, North Lampung, Sumatera, In-donesia (4◦31′ S, 104◦55′ E), on a field with a gentleslope (4%). The soil was a Plinthitic Kandiudult with asandy clay topsoil and clay accumulation at depth, par-ticularly below around 1 m (Suprayogo et al., 2002).Aluminium saturation in the subsoil approached 60%(van der Heide et al., 1992). Six cropping systemswere established in 1985–1986 to explore the effectsof hedgerows on the growth of food crops, of whichfour were included in the current study: one withno hedgerows (‘monocrop’), plus hedgerow intercrop-ping systems using Peltophorum dasyrrhachis (‘P-P’),G. sepium (‘G-G’), and a system in which hedgerowsof the latter two species alternated (‘G-P’) (van Noord-wijk et al., 1995). Hedgerows were 4 m apart and trees

63
were 50 cm apart within the row. Plastic sheets wereinstalled in the soil to 50 cm depth around monocropplots at setup, to prevent the incursion of tree roots.Two crops were generally grown per year; in the cur-rent experiment these crops were maize followed bygroundnut. Mean maize yields (t ha−1) over four crop-ping seasons in 1991–1992 were 2.30 (monocrop),2.33 (G-G), 3.32 (P-P) and 2.55 (G-P) (van Noordwijket al., 1995).
Experimental design of 15N study
During the wet season of 1997–8 the fate of fertilizerversus organic N sources was compared in the fourcropping systems studied, using 15N urea and 15N la-belled prunings. There were four replicated urea plotsper treatment but (because of limited supplies of 15Nlabelled prunings) only two labelled prunings plots perhedgerow intercropping system treatment (G-G, P-P,G-P). No labelled prunings were applied to monocropplots. Because of the danger of 15N contamination,plots were positioned at least 4 m apart, as suggestedfor these species by Rowe and Cadisch (2002).
Eight months before the start of the current exper-iment, Mucuna pruriens was sown on all treatmentplots as a green manure, but the above-ground bio-mass was removed before planting. Glyphosate herbi-cide (Roundup, Monsanto Co., St Louis, MO, USA)was applied, and thereafter the field was kept clearof weeds by regular hand weeding. On 5th Decem-ber 1997 all hedgerows were pruned. Prunings wereapplied evenly over the hedgerow and alley. Urea(30 kg N ha−1), KCl (60 kg K ha−1) and triplesuper-phosphate (60 kg P ha−1) were applied. On11th December 1997 a crop of maize was sown at aspacing of 65 × 50 cm, in 5 rows along each alley. On10th January 1998 an additional topdressing of urea(60 kg N ha−1) was applied. This crop was harvestedon 11th March 1998 and hedgerows were pruned on22nd March 1998. A crop of groundnut was sown on27th March 1998 and harvested on 1st July 1998. Anadditional application of 60 kg K and 60 kg P ha−1was made on 28th March 1998, but no N was applied.
On 10th January 1998, 15N labelled materials wereapplied to 0.5 m wide strips (‘microplots’) runningacross the 4 m wide crop alley and centred on a treeon either side (Figure 1). Urea (5.523 atom% 15N)was applied to the 15N microplots at a rate of 60 kgN ha−1. An amount of water equivalent to 10 mm ofrainfall was then added to the plot, to wash in the 15Nand reduce potential runoff losses. Unlabelled urea
(60 kg N ha−1) was applied at this time to the restof the field, including the border 0.5 m × 4 m strips oneither side of the 15N application strip. Labelled treematerial was prepared the previous year, by pruningtwo isolated trees of each species and applying 15Nenriched (10 atom% 15N) ammonium sulphate weeklyduring the regrowth period. After 77 days the labelledregrowth material was pruned, divided into leavesand stems, dried and subsamples finely ground beforeanalysis. Mean N concentrations for the two trees were3.37±0.03% and 0.95±0.03 % for G. sepium leaf andstem respectively, and 2.02±0.02% and 0.57±0.02%for P. dasyrrachis leaf and stem. Mean 15N concentra-tions were 1.52±0.25 atom% and 1.28±0.21 atom%for G. sepium leaf and stem respectively, and 2.53 ±0.10 atom% and 2.25±0.06 atom% for P. dasyrrachisleaf and stem. Material from each tree was more ho-mogeneous (e.g. leaf material from the first G. sepiumtree was 1.96 ± 0.03 atom% 15N). Labelled tree prun-ings were applied at rates of 5.9 (G. sepium) plus 4.1(P. dasyrrachis), 5.9 and 4.1 t ha−1 to G-P, G-G andP-P systems respectively, equivalent to 125, 66 and201 kg N ha−1. These rates were 25–34% of the dryweights applied following pruning on 5th December.To test whether P. dasyrrachis prunings influencedthe mineralisation of G. sepium prunings, approxi-mately the same amount of G. sepium prunings wasapplied to the G-P system as in the G-G system, alongwith P. dasyrrachis prunings in the same proportionas above; in this system only the G. sepium pruningsapplied were 15N labelled. Equal dry weights of unla-belled prunings from trees growing outside the fieldwere applied on either side of the application strip.Urea was not applied to 15N prunings application plotsor border strips.
Plant and soil sampling
Soil was sampled on two occasions for analysis ofmineral N, total N and 15N contents. Two replicatesamples were taken using a 6 cm diameter auger fromsix layers by depth (0–5, 5–20, 20–40, 40–60, 60–80and 80–100 cm), for each cropping system. Within thehedgerow intercropping systems, two replicate sam-ples were taken for each depth at different positions,0.7 m from the line of the hedgerow, and in the centre,2 m from hedgerows. Subsamples of 5 g fresh weightwere shaken for 2 h in 20 mL 2 M KCl for mineralN analysis, and stored when necessary at −18 ◦C. Am-monium and nitrate concentrations in the extracts weredetermined colorimetrically by flow injection analy-

64
Figure 1. Layout of 15N application plots. 15N was applied in a strip across the crop alley denoted by the solid line. Recovery was calculatedfor crop plants and trees in centre and border zones, denoted by dashed lines.
ses, using the methods of Alves et al. (1993) and Gineet al. (1980) respectively. An additional subsamplewas air-dried for total N and 15N analysis. Soil bulkdensity was measured for samples obtained by drivinga 10 cm diameter ×5 cm PVC tube into the side of apit at the central depth for the layer, following dryingat 105 ◦C for 2 days.
The uptake of 15N by crop plants within the 15Napplication strip (‘Centre’ plants: 2 m2) was moni-tored, as was uptake by ‘Border’ crop plants growingin two strips of 0.5 m × 4 m either side of the applica-tion strip (4 m2) (Figure 1). Because of the presumedwider spread of tree root systems, the three trees clos-est to the application strip were considered ‘Centre’trees, (corresponding to an area of 1.5 m × 4 m = 6 m2
per hedgerow). The three trees either side of thesewere sampled as ‘Border’ trees (12 m2 per hedgerow).Reported 15N recovery is the sum of recoveries calcu-lated separately for ‘Centre’ and ‘Border’ areas.
At crop harvests, crop plants were uprooted andsubdivided into suitable components. The fresh weightof each component was determined, and subsamples(50–100 g) were weighed, dried and re-weighed to de-termine dry matter content. Leaves, or pieces of othercomponents, were taken from all plants in the zone
sampled. Root samples were obtained from uprootedplants, to avoid contamination by roots from plantsfrom outside the plot. Maize plants were divided atharvest into leaf plus husk, stem, grain, cob cores, androot. Groundnut plants were divided at harvest intograin, husk, stover (leaf plus stem), and roots.
At each pruning, all regrowth was removed fromtrees and divided into leaf and stem, leaving only the75–90 cm height main trunk(s). Leaf and stem freshweight were recorded, and subsamples were dried todetermine dry weight ratios. Leaves and stems werecarefully subsampled for 15N analysis. The biomassof trunks (the portion of the stem not pruned) wasestimated from volume and density measurements.Samples of trunks for 15N analysis were obtained bydrilling half way through main stems and collectingthe resulting sawdust. The biomass of roots was esti-mated from augered samples (see below). Dry weightsof roots from all depths were summed to give totalroot weights for each species in the top 100 cm of soil.Samples of tree roots for 15N analysis were obtainedfrom the centre tree in each zone by excavating andcutting a section of root. Litterfall was collected in0.5 × 0.5 m traps situated beneath the central treeson each plot. Litter was collected and dried every

65
2 weeks until tree pruning, when all litter collectedfrom each trap was pooled and weighed. Followingsub-sampling, the remainder of the plant material wasreturned to the plot, with the exception of grain, maizecores and groundnut husks which were removed.
Crop yield (of grain or of biomass) from cen-tre and border areas was summed, and converted toyield per unit area using the total plot harvest area of1.5 m × 4 m = 6 m2. Tree biomass and N yields wereconverted to yield per unit field area on the basis ofthe area (0.5 m × 4 m = 2 m2) available to each of thetrees included in the yield measurement. Total N yieldper plot or subplot was calculated as the product of drymatter yield and proportional N content.
Root sampling took place over a three week pe-riod, 8–11 weeks after sowing maize. Samples wereobtained using an 8 cm diameter coring auger (Ander-son and Ingram, 1993). Samples were taken at threedistances from the line of the hedgerow (25, 85 and184 cm) and therefore there were three positions for P-P and for G-G, five positions for G-P, and one positionfor Monocrop. Sampling positions were halfway alonga line from expected maximum Lrv (beneath the plant)and expected minimum Lrv (furthest from plant), rel-ative to trees for the zone nearest (0–50 cm from)hedgerows, and relative to maize plants for the othersamples. Samples were taken from six depth ranges(0–5, 5–20, 20–40, 40–60, 60–80, 80–100 cm depth).From each position and depth 12 replicate sampleswere taken from different field locations. The col-lected soil samples were soaked overnight in water andwashed over sieves of 2 mm and 0.25 mm mesh size,ensuring that all washing water passed through bothsieves. Live roots of different species were separatedaccording to visual characteristics (Rowe et al., 2001)and sorted by diameter class (>2 mm and <2 mm).Root length was measured using the grid intersectmethod (Tennant, 1975). Presented Lrv values do notinclude roots greater than 2 mm in diameter.
15N recovery calculations
Plant samples were air-dried for 2–3 days in openpaper bags, then oven dried at 60 ◦C for 48 h. Soiland plant samples were ground finely and subsampleswere analysed for % N and atom % 15N content us-ing a Europa 20/20 mass spectrometer (PDZ, formerlyEuropa Scientific, Crewe, UK) coupled to a Roboprepautomated C/N analyzer.
After application, the 15N in plant and soil compo-nents (i.e., amount (µg) of 15N excess) was expressed
as percentage recovery, i.e., the percentage of theamount of 15N applied which was found in the com-ponent. The amount of 15N applied was calculatedas excess over the 15N natural abundance of totalsoil N. Natural abundance was found to differ be-tween hedgerow treatments and soil depths, and foreach treatment the mean natural abundance at 0–5, 5–20, 20–40, 40–60, 60–80 and 80–100 cm soil depth(which varied from 0.0023 to 0.0040 atom% 15N),weighted by % N content at each depth (data notshown), was used.
Subsamples of all plant components were analysedfor 15N content in the centre zone of each plot. For plotborders, the total amount of 15N was estimated fromthe amount of 15N in components containing most ofthe N (Rowe and Cadisch, 2002). Average cumulativedry weight of litter fall per 0.5 × 0.5 m littertrap was4.6 g (G. sepium) or 4.9 g (P. dasyrrachis). Cumulativelitterfall across the whole alley was estimated to con-tain only 2–5 % of the N content of trees at pruning,and was therefore disregarded in calculations of 15Nrecovery.
The amount of 15N excess in each soil volume wascalculated as the product of volume, bulk density, to-tal N proportion and 15N excess proportion, correctedfor background 15N abundance using the mean naturalabundance value for the appropriate depth/hedgerowcombination. Bulk density was 1.36, 1.39, 1.52, 1.52,1.55 and 1.50 for soil layers 0–5, 5–10, 20–40, 40–60, 60–80 and 80–100 cm, respectively. Soil volumeswere calculated as the product of layer thickness andthe application area (Centre zone). Border soil 15Nwas not measured.
To estimate the amount of 15N which remained inlitter at the time of first crop harvest, a further exper-iment was set up to measure 15N remaining in litterfollowing an equivalent application of 15N labelledprunings to the soil surface. Prunings of G. sepiumand P. dasyrrachis were taken in December 1998 fromtrees which had been labelled with 15N in the main ex-periment. Material was taken from two labelled treesper species, and also from unlabelled P. dasyrrachistrees. Prunings were separated into leaf and stem, anddried in the sun. Subsamples were taken for determi-nation of N and 15N content. Plastic pipes of 15 cmdiameter were cut into 10 cm lengths and then set5 cm deep into the soil. Prunings were applied onthe same day of the year (10th January 1999), and atthe same rates (dry leaf t ha−1 and dry stem t ha−1),as in the main experiment. The same treatments wereused (labelled G. sepium, labelled P. dasyrrachis, and

66
labelled G. sepium plus unlabelled P. dasyrrachis),with 5 replicate pipes per treatment. On March 19th
1999 (after an interval equivalent to that before sam-pling for the first 15N balance in the main experiment),surface litter remaining was carefully brushed up.15N recovery was calculated as a percentage of 15Napplied.
Statistics
Ranges are indicated in the text and figures as ± onestandard error of the mean. Yield and 15N recoverydata were analysed using linear regression, with pair-wise comparisons made using the RPAIR procedure ofGenstat (Payne et al., 1987). Root length density datawere log transformed (Log10 (Lrv cm cm−3 + 10−4))before analysis by ANOVA, to reduce heteroscedastic-ity of variance.
Results
Following 11–12 years of inputs of tree prunings, totaltopsoil N was greater in G-G and P-P systems thanafter continuous monocrop (Table 1). However, thiseffect was not evident below 5 cm depth, and wasnot found in the mixed G-P system. Large quantitiesof mineral N (310–449 kg N ha−1 in the top metre)were available for plant uptake in all systems at thebeginning of the experiment (Table 2) but less than25% was in the more mobile form of nitrate (data notshown). Analysis at the end of the field season showedan overall decline in total mineral N content in the topmetre of soil during the first cropping season. Min-eral N contents at 60–100 cm depth, however, weremaintained or increased under monocrop and in sys-tems with G. sepium, indicating a greater amount ofleaching in these than in the P-P system. The amountof mineral N present in the top metre of soil at thebeginning of the second crop was greatest in the G-Gsystem (239 kg N ha−1) and least in the systems withP. dasyrrachis (139–160 kg N ha−1).
Maize grain yield on 15N application microplotswas significantly reduced in the G-G intercroppingsystem compared to monocrop (Figure 2). At the sec-ond harvest, microplot groundnut grain and total Nyields were significantly greater in monocrop than inhedgerow intercropping systems. Grain and total Nyields were not affected by 15N application method.
Tree N yields calculated for the pruning on22nd March, after 76 and 108 days regrowth for
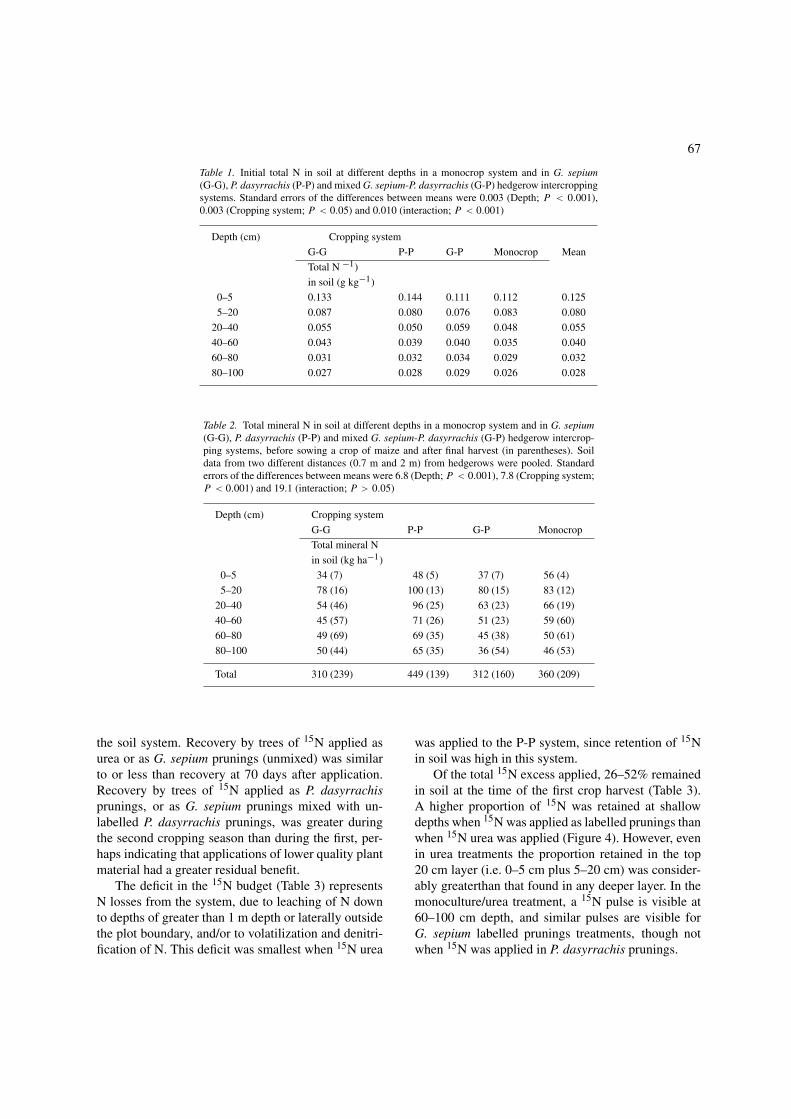
G. sepium and P. dasyrrachis respectively, showed thatthe method used for 15N application had no significanteffect on N content of regrowth and so data from thesetreatments were pooled. N yields from the G. sepiumpruning on 8th January were estimated from main plotdry matter yields multiplied by N content of prun-ings measured on 22nd March, and included in totalN yield. Input of N from prunings during and after thegrowth of the first crop was greater in the G-G systemthan in the P-P system (Figure 3). Prunings N inputin the G-P system was intermediate. Total organic Ninputs (prunings plus maize stover) during this 108 dayperiod were (G-G) 172 kg N ha−1; (P-P) 91 kg N ha−1;(G-P) 138 kg N ha−1; (Monocrop) 51 kg N ha−1.
The proportion of 15N applied as urea which wasrecovered by maize in the different cropping sys-tem treatments corresponded approximately to maizeN yield from these treatments (Table 3; Figure 2); lessurea 15N was recovered by maize in the G-G systemthan in monocrop. Total plant (maize + tree) recoveryof urea 15N was similar in these four systems. Re-covery by G. sepium hedgerows of 15N from labelledG. sepium prunings was similar to that from 15N urea,but maize recovered a smaller proportion of the 15Nfrom labelled prunings of either species than of urea15N. Total plant recovery from labelled tree pruningsapparently increased in the order P-P → G-P → G-G,corresponding to increasing mean quality of the litterapplied, but this trend was not significant.
Recovery by trees and maize of 15N from labelledG. sepium prunings was not significantly reduced bythe addition of unlabelled P. dasyrrachis prunings.Following labelled prunings applications, amounts of15N remaining in soil plus litter were similar for allhedgerow intercropping systems (Table 3). However,the proportion of this pool which remained in surfacelitter was much greater (53%) following P. dasyrrachisprunings applications than following G. sepium prun-ings applications (10%). More 15N from labelledG. sepium prunings remained in litter when thesewere applied with unlabelled P. dasyrrachis prun-ings. Amounts of 15N remaining in soil (plus litter)were smaller following application as 15N urea thanfollowing application as 15N labelled prunings. Thesmallest deficit in the 15N budget was found followingapplication of 15N urea to the P-P system.
Crop recovery of 15N at the second crop harvestwas less than that at first harvest (Table 3). The declinein crop recovery between the two dates was steeperfor urea than for labelled prunings applications, sug-gesting that labelled prunings N is more persistent in
67
Table 1. Initial total N in soil at different depths in a monocrop system and in G. sepium(G-G), P. dasyrrachis (P-P) and mixed G. sepium-P. dasyrrachis (G-P) hedgerow intercroppingsystems. Standard errors of the differences between means were 0.003 (Depth; P < 0.001),0.003 (Cropping system; P < 0.05) and 0.010 (interaction; P < 0.001)
Depth (cm) Cropping system
G-G P-P G-P Monocrop Mean
Total N −1)
in soil (g kg−1)
0–5 0.133 0.144 0.111 0.112 0.125
5–20 0.087 0.080 0.076 0.083 0.080
20–40 0.055 0.050 0.059 0.048 0.055
40–60 0.043 0.039 0.040 0.035 0.040
60–80 0.031 0.032 0.034 0.029 0.032
80–100 0.027 0.028 0.029 0.026 0.028
Table 2. Total mineral N in soil at different depths in a monocrop system and in G. sepium(G-G), P. dasyrrachis (P-P) and mixed G. sepium-P. dasyrrachis (G-P) hedgerow intercrop-ping systems, before sowing a crop of maize and after final harvest (in parentheses). Soildata from two different distances (0.7 m and 2 m) from hedgerows were pooled. Standarderrors of the differences between means were 6.8 (Depth; P < 0.001), 7.8 (Cropping system;P < 0.001) and 19.1 (interaction; P > 0.05)
Depth (cm) Cropping system
G-G P-P G-P Monocrop
Total mineral N
in soil (kg ha−1)
0–5 34 (7) 48 (5) 37 (7) 56 (4)
5–20 78 (16) 100 (13) 80 (15) 83 (12)
20–40 54 (46) 96 (25) 63 (23) 66 (19)
40–60 45 (57) 71 (26) 51 (23) 59 (60)
60–80 49 (69) 69 (35) 45 (38) 50 (61)
80–100 50 (44) 65 (35) 36 (54) 46 (53)
Total 310 (239) 449 (139) 312 (160) 360 (209)
the soil system. Recovery by trees of 15N applied asurea or as G. sepium prunings (unmixed) was similarto or less than recovery at 70 days after application.Recovery by trees of 15N applied as P. dasyrrachisprunings, or as G. sepium prunings mixed with un-labelled P. dasyrrachis prunings, was greater duringthe second cropping season than during the first, per-haps indicating that applications of lower quality plantmaterial had a greater residual benefit.
The deficit in the 15N budget (Table 3) representsN losses from the system, due to leaching of N downto depths of greater than 1 m depth or laterally outsidethe plot boundary, and/or to volatilization and denitri-fication of N. This deficit was smallest when 15N urea
was applied to the P-P system, since retention of 15Nin soil was high in this system.
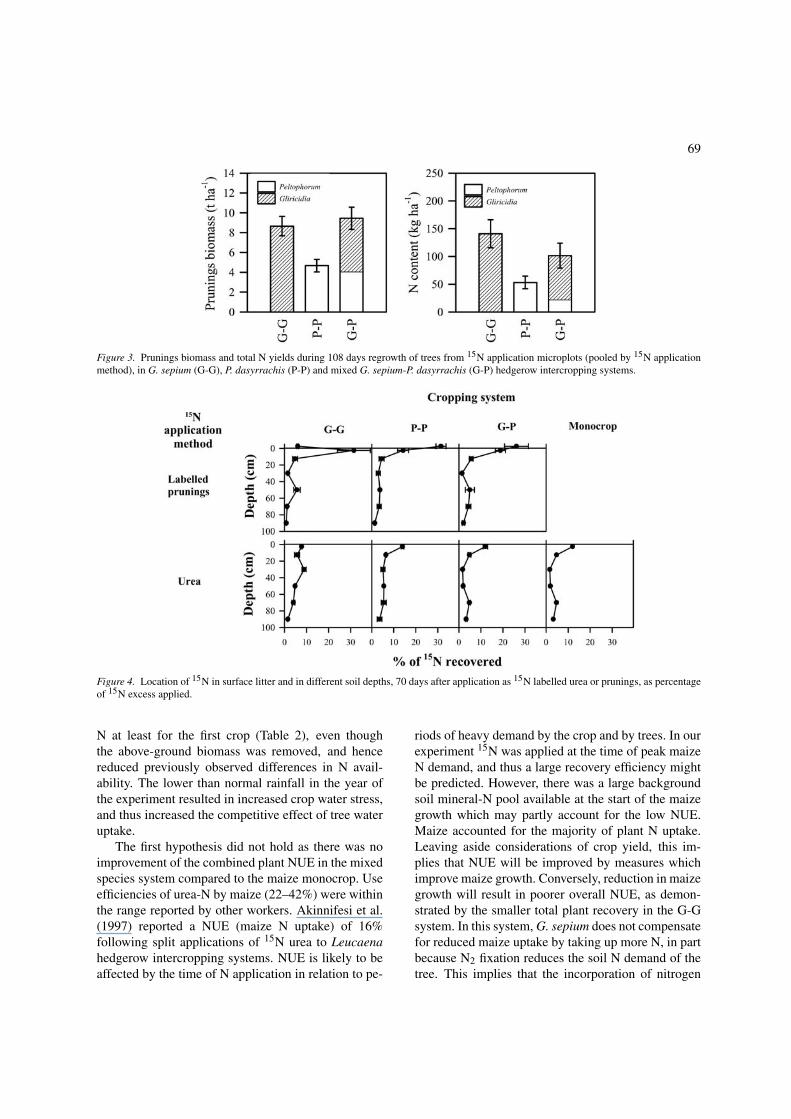
Of the total 15N excess applied, 26–52% remainedin soil at the time of the first crop harvest (Table 3).A higher proportion of 15N was retained at shallowdepths when 15N was applied as labelled prunings thanwhen 15N urea was applied (Figure 4). However, evenin urea treatments the proportion retained in the top20 cm layer (i.e. 0–5 cm plus 5–20 cm) was consider-ably greaterthan that found in any deeper layer. In themonoculture/urea treatment, a 15N pulse is visible at60–100 cm depth, and similar pulses are visible forG. sepium labelled prunings treatments, though notwhen 15N was applied in P. dasyrrachis prunings.

68
Figure 2. Grain and total crop N yield from 15N application microplots (pooled by 15N application method), in a monocrop system andin G. sepium (G-G), P. dasyrrachis (P-P) and mixed G.sepium-P. dasyrrachis (G-P) hedgerow intercropping systems. Tree prunings yieldscorrespond to 108 days regrowth. Treatments which differed significantly (P < 0.05) are indicated with different letters.
In the single tree species hedgerow treatments, re-covery of 15N was measured separately for trees eitherside of the application plot. Recovery of 15N (mean ofG. sepium and P. dasyrrachis systems) by downslopehedgerows was 4.6% and 1.2% from urea and pruningsrespectively; recovery by upslope hedgerows was less(P < 0.05), at 2.1% and 1.4%, respectively (s.e.d.between position means = 0.7), even though the slopeon this field was only around 4%. This effect was ap-parently more pronounced for 15N applied as urea thanas labelled prunings.
All species showed a rapid decline in root lengthdensity with depth (Figure 5). Species differed how-ever in their root distribution with depth (P < 0.001).The decline in log Lrv with soil depth was steeper formaize than for the trees and steeper for G. sepium thanfor P. dasyrrachis. In the top layers of soil, G. sepiumLrv was greater than P. dasyrrachis Lrv, but the twospecies had similar Lrv beneath around 60 cm soildepth. Thus P. dasyrrachis had a greater proportion
of its root system in the ‘safety-net’ layer, beneath themain crop rooting layer. The presence of G. sepiumroots apparently resulted in shallower maize root dis-tributions, with 90% and 87% of maize roots foundin the top 5 cm of soil in G–G and G–P systemscompared to 76% in P–P and 72% in monoculture(Figure 5).
Discussion
Maize yields from the P-P system were similar tothose from monoculture, but those in the G-G sys-tem were significantly reduced, presumably becauseof competition from the tree for light, water and/orother nutrients. It is not clear why the beneficial ef-fect of P. dasyrrachis hedgerows reported previouslyfrom this site by van Noordwijk et al. (1995) was notseen in this season. The residual effect of the previousMucuna cover crop appears to have provided sufficient
69
Figure 3. Prunings biomass and total N yields during 108 days regrowth of trees from 15N application microplots (pooled by 15N applicationmethod), in G. sepium (G-G), P. dasyrrachis (P-P) and mixed G. sepium-P. dasyrrachis (G-P) hedgerow intercropping systems.
Figure 4. Location of 15N in surface litter and in different soil depths, 70 days after application as 15N labelled urea or prunings, as percentageof 15N excess applied.
N at least for the first crop (Table 2), even thoughthe above-ground biomass was removed, and hencereduced previously observed differences in N avail-ability. The lower than normal rainfall in the year ofthe experiment resulted in increased crop water stress,and thus increased the competitive effect of tree wateruptake.
The first hypothesis did not hold as there was noimprovement of the combined plant NUE in the mixedspecies system compared to the maize monocrop. Useefficiencies of urea-N by maize (22–42%) were withinthe range reported by other workers. Akinnifesi et al.(1997) reported a NUE (maize N uptake) of 16%following split applications of 15N urea to Leucaenahedgerow intercropping systems. NUE is likely to beaffected by the time of N application in relation to pe-
riods of heavy demand by the crop and by trees. In ourexperiment 15N was applied at the time of peak maizeN demand, and thus a large recovery efficiency mightbe predicted. However, there was a large backgroundsoil mineral-N pool available at the start of the maizegrowth which may partly account for the low NUE.Maize accounted for the majority of plant N uptake.Leaving aside considerations of crop yield, this im-plies that NUE will be improved by measures whichimprove maize growth. Conversely, reduction in maizegrowth will result in poorer overall NUE, as demon-strated by the smaller total plant recovery in the G-Gsystem. In this system, G. sepium does not compensatefor reduced maize uptake by taking up more N, in partbecause N2 fixation reduces the soil N demand of thetree. This implies that the incorporation of nitrogen

70
Table 3. Recovery of 15N applied either as urea or labelled prunings to a monocrop system and to G. sepium (G-G),P. dasyrrachis (P-P) and mixed G. sepium-P. dasyrrachis (G-P) hedgerow intercropping systems. The location of 15N isshown in plants and in the top 1 m of soil 70 days after 15N application, and in plants 172 days (groundnut) or 180 days(G. sepium and P. dasyrrachis) after application. Values within the same row annotated with the same letter were notsignificantly different (P < 0.05)
Plant or soil pool 15N application method
Urea Labelled prunings
Cropping system
G-G P-P G-P Mon. G-G P-P G-P
a) After 70 days % of applied 15N recovered
G. sepium 11.3b 4.5a 9.3ab 3.7a
P. dasyrrachis 9.2b 3.5a 3.1a 1.6a
Maize 22.3b 33.5bc 26.5bc 42.0c 5.8a 5.1a 7.4ab
Litter 6.0a 31.4b 26.1b
Soil (0–1 m) 32.6a 40.0a 28.5a 26.0a 52.1a 27.7a 33.1a
Total soil and plant 66.2 82.7 63.0 68.0 73.2 67.3 71.9
Deficit 33.8 17.3 37.0 32.0 26.8 32.7 28.1
b) After 172 or 180 days
G. sepium 11.5a 6.0a 8.2a 12.5a
P. dasyrrachis 4.0a 3.0a 10.0a 4.0a
Groundnut 5.4a 2.5a 3.7a 5.2a 5.5a 1.8a 2.3a
Total plant 16.9c 6.5ab 12.7bc 5.2a 13.7bc 11.8abc 18.8c
fixing trees into cropping systems may lead to ineffi-ciency in N cycling. This was reflected in a decreasein simulated interception efficiency with increasing N2fixation rate (Cadisch et al., 2004). Van Noordwijk andCadisch (2002) used a series of simulations using theWaNuLCAS agroforesty model to demonstrate that ef-fective N filtering by roots depends on unfulfilled plantN demand, and is thus incompatible with maximisingthe yield of all components. In a study of N fertiliseruse in Sorghum bicolor hedgerow intercropping sys-tems, Lehmann et al. (2002) found that N leaching wasreduced under hedgerows compared to crop alleys.However the hedgerows recovered six times as muchN as the crop, indicating that there was competition,and the crop took up less N than in monoculture.
Total plant N production was considerably greaterin the P-P system than the monocrop system, due tothe additional N in P. dasyrrachis prunings. GreaterN inputs in the P-P system than under continuousmonocropping resulted, over 11–12 years, in total top-soil N levels around 50% greater than under the lattersystem. The lack of such an effect in the G-P systemmay be because the mixture of prunings from the twospecies was mineralised faster that either alone (Gart-ner and Cardon, 2004), but this is contradicted by theimproved retention of G. sepium litter N when mixed
with P. dasyrrachis litter (Table 3); it is difficult toexplain what interactions lead to this result. However,since the systems have an identical history of N fer-tilizer use and P. dasyrrachis is non-fixing, it can besaid that P. dasyrrachis trees in the P-P system mustbe meeting their N requirements from complemen-tary sources, spatially or temporally separated frommaize uptake. Some of the improved N retention maycome from delayed mineralisation of the recalcitrantP. dasyrrachis prunings (Handayanto et al., 1994).P. dasyrrachis prunings are high in polyphenols whichhave a strong protein binding capacity and led to im-mobilization of fertilizer N (Figure 4). Tree safety-netuptake, i.e. N uptake by trees from the soil layer be-neath the main crop rooting zone as a proportion ofuptake plus N leached out of this layer (Cadisch et al.,2004), must however contribute substantially to themaintenance of relatively high total soil N in the P-P system. This is facilitated by its root system with ahigh proportion of roots in the subsoil (Figure 5; Roweet al., 2001) creating an important N sink below themain crop rooting zone.
P. dasyrrachis has a higher proportion of its rootsin the subsoil than G. sepium (Figure 5; Rowe et al.,2001). This suggests that a greater proportion of itsN uptake is derived from subsoil, as concluded by

71
Figure 5. Root length density of maize (�), G. sepium (�) and P. dasyrrachis (�) at different soil depths in monocrop and 3 hedgerowintercropping systems; G. sepium (G-G), P. dasyrrachis (P-P) or alternating G. sepium and P. dasyrrachis hedgerows (G-P). Pooled means forall horizontal positions.
Rowe et al. (1999) following injections of 15N atdifferent soil depths. In the current study there wassubstantial downward movement of applied 15N; 2–10% of the 15N applied was found between 60 cm and100 cm depth after 70 days (Figure 3). The verticalseparation between P. dasyrrachis and maize N uptakezones, coupled with the rapid leaching of N, seemslikely to account for at least some of the demonstratedN uptake complementarity between the P. dasyrrachisand the crop. Mineral N contents in deeper soil in-creased during the cropping season in monocultureand in systems with G. sepium (Table 2), implying thatthese systems were more prone to leaching losses thanthe P-P system. Thus the results are consistent withthe hypothesis (H2) that P. dasyrrachis hedgerow in-tercropping systems are more efficient in their use of Nthan G. sepium hedgerow intercropping systems, sinceP. dasyrrachis root systems are spatially distinct fromcrop root systems and thus have more opportunity tointercept N before it is leached. Nutrient resources in
subsoil are accessible to deep-rooting trees, and thelarge volume of the subsoil means that these nutrientscan represent a substantial resource (Lehmann et al.,2001). However, systems need careful design if thiseffect is to be usefully exploited, since the depth oftree root activity is dependent on complex interac-tions between soil type, tree species and management(Lehmann, 2003).
Amounts of N recycled by trees were substan-tial. Above-ground inputs of organic residue N tosoil were 15 to 20 times greater in hedgerow in-tercropping systems than in monocrop. Amounts ofN remaining in soil (plus litter) after 70 days weresubstantially greater after applying prunings than af-ter applying urea (Table 3). However, inputs of N inpruning materials were substantially less available tothe next crop than N applied as urea; crop plants tookup proportionally much less N from prunings thanfrom urea applications. Use efficiency of G. sepiumprunings N over the two seasons (10.9 or 9.4% re-

72
spectively for G-G and G-P treatments) was less thanthe use efficiency range (12–29%) reported by Gillerand Cadisch (1995). Use efficiency of P. dasyrrachisprunings N (7.0%) was even lower; the large amountof P. dasyrrachis residue which remained at the sur-face (Table 3; Figure 3) suggests that this was due tothe slow mineralisation of P. dasyrrachis leaves rel-ative to G. sepium leaves. Thus greater losses of Nfrom G. sepium than from P. dasyrrachis were notobserved, and H3 was not supported. The increasedretention in litter of N from G. sepium prunings whenthese were applied with unlabelled P. dasyrrachisprunings suggests that the lower quality prunings ei-ther moderated the release of N from G. sepiumleaves, or re-immobilised the released N. This pro-vides some support for H4, though the effect did notresult in greater subsequent recovery by groundnut ofG. sepium N in the G-P system. Trees accounted for alarger proportion of total plant recovery when 15N wasapplied as prunings than after 15N urea application.This may have been because prunings 15N becameavailable later than urea 15N, at a time when maizeN uptake was declining. Overall deficits were simi-lar, suggesting that organic N is also susceptible toleaching and/or gaseous loss.
These efficiencies may somewhat underestimatethe actual amount of N taken up by plants as a resultof prunings applications, because 15N displaces andmakes available N from other soil pools (pool substi-tution; Jenkinson et al., 1985). The cumulative effectsof N release from successive applications of pruningsmay be greater than is suggested by the release, in sub-sequent cropping seasons, from a single application.However, the large amount of N remaining in litter inthe P. dasyrrachis system may be very recalcitrant. Ina pot experiment carried out by Cadisch et al. (1998),maize plants recovered only 20% of 15N applied aslabelled P. dasyrrachis prunings (finely ground andmixed with soil) over three cropping seasons, and thegreat majority of this was recovered in the first sea-son. The eventual fate of the recalcitrant N fraction isunclear. Oorts et al. (2002) observed that low qualityresidues showed most significant differences in finesilt fractions resulting in increased CEC values and thelargest carbon contents, the latter coinciding with ourobservations. It may contribute to long term soil fer-tility, whether through later mineralisation or throughfavourable effects of the organic matter on soil struc-ture (Hamblin, 1985), but it appears not to becomeavailable in substantial quantities to plants within atimespan relevant to farmers’decision-making.
Total plant 15N uptake was greater from G. sepiumprunings (low C:N and low polyphenol content) thanfrom the lower quality P. dasyrrachis prunings (highC:N and polyphenol). This is in accordance with otherobservations (Cadisch et al., 1998) as it has been wellestablished that the lignin+polyphenol:N ratio deter-mines N release from organic residues (Handayantoet al., 1994; Constantinides and Fownes, 1994). Largedifferences in losses of labelled pruning N betweenthe systems were not observed, presumably becauseonly small amounts of N were captured and recycledby trees. This is in contrast to results from a less Nrich environment where trees recycled up to 35% ofhigh quality pruning N (VanLauwe et al., 1998).
The vertical distribution of 15N in soil 70 days afterapplication (Figure 4) shows that significant amountswere retained at or near the surface, suggesting thatsome of the applied 15N was protected from leach-ing and other losses by incorporation into soil organicmatter, particularly when applied as prunings. Someof the applied 15N was converted to or remained insoluble form and was leached down the profile. Themovement of mineral N down the profile explains theconsiderably greater proportion of plant recovery ac-counted for by tree uptake during the second croppingseason – the pulse of 15N visible in the monocultureand G. sepium labelled prunings treatments is likelyto have passed beyond the range of groundnut rootswhile the crop was establishing. The total proportionof 15N retained in soil 70 days after application didnot differ between application methods or hedgerowtreatments (Table 3). However, if the N deeper in theprofile at this time is indeed more subject to leach-ing than topsoil N, as expected from the low organicN content of subsoil, greater losses might be expectedfrom urea and labelled G. sepium prunings treatmentsduring subsequent seasons.
In pot experiments with ground plant material,Handayanto et al. (1997) observed a strong nega-tive priming effect of P. dasyrrachis on the releaseof G. sepium N. In the current study, (unlabelled)P. dasyrrachis prunings appeared to moderate the re-lease of 15N from (labelled) G. sepium prunings inthe G-P system, since amounts of 15N recovered inplants and in soil were similar to those found withoutaddition of P. dasyrrachis, but significantly more 15Nremained in litter. Thus P. dasyrrachis leaves appearto have moderated rather than prevented the release ofG. sepium N, since uptake of G. sepium N by cropplants was not affected.

73
Priming effects may also occur in single speciesstands as continuous additions of prunings over timeinteract with partially decomposed litter from pre-vious applications. Cadisch et al. (1998) observedthat continuous application of high quality G. sepiumprunings had a positive priming effect on the N re-lease from previously applied 15N labelled G. sepiumprunings. The opposite occurred with polyphenol richP. dasyrrachis prunings, where a negative priming ef-fect was observed. In both occasions the priming effectwas less than 2% of the applied N. While such interac-tions can easily be observed in controlled pot studiesit is much more difficult to observed them in spatiallyvariable field experiments.
Considerable lateral leaching of applied N was in-dicated by the increased 15N uptake by downslopehedgerows relative to upslope hedgerows even on thisgentle slope (4%). This also indicates that this lateralflow can be intercepted by trees or crop plants else-where in the field, if it remains at soil depths where itis still accessible. Thus even trees planted at relativelywide spacings could have a significant filter function(or lateral ‘safety-net’). This is particularly true forsoils such as Ultisols where hydraulic conductivity de-creases with depth and so a large proportion of thewater falling in an intense rainstorm may flow laterallythrough the top layers of soil. Such a filter will operatemost effectively if it is perpendicular to the flow of N,i.e. with hedgerows along contours.
Conclusions
G. sepium hedgerows reduced crop yield, and nitrogenuse efficiency (crop N uptake during the first season)was less in the G. sepium intercropping system thanin monoculture. Thus agroforestry does not alwaysimprove resource use efficiency. The hedgerow inter-cropping systems studied do appear to recycle more Nthan in monoculture, partly because of separation inspace and time of tree root N uptake from that by cropplants. However, this advantage must be set againstpotential reductions in crop yield due to competitionfor nitrogen or other resources. Over the years of theprevious study the P. dasyrrachis system in particu-lar gave improved crop yields over monoculture, butin this experiment crop yields were greatest in mono-culture. Competition, and the labour input requred tocontrol it, is a major constraint to adoption of alleycropping systems.
Acknowledgements
This publication is an output from research projectsfunded by the EU and the Department for Interna-tional Development of the United Kingdom (R6523,Forestry Research Programme). However, the Depart-ment for International Development can accept noresponsibility for any information provided or viewsexpressed.
References
Akinnifesi F K, Kang B T, Sanginga N and Tijani-Eniola H 1997Nitrogen use efficiency and N competition between Leucaenahedgerows and maize in an alley cropping system. Nutr. Cycl.Agroecosys. 47, 71–80.
Alves J R, Boddey R M and Urquiaga S S 1993 A rapid and sensitiveflow injection technique for the analysis of ammonium in soilextracts. Commun. Soil Sci. Plant Anal. 24, 277–284.
Anderson J M and Ingram J S I 1993 Tropical Soil Biology and Fer-tility: A Handbook of Methods. CAB International, Wallingford,UK. 221 pp.
Cadisch G, Handayanto E, Malama C, Seyni F and Giller K E1998 N recovery from legume prunings and priming effects aregoverned by the residue quality. Plant Soil 205, 125–134.
Cadisch G, de Willigen P, Suprayogo D, van Noordwijk M andRowe E C 2004 Catching and competing for mobile nutrientsin soils. In Belowground Interactions in Tropical Agroecosys-tems with Multiple Plant Components. Eds. M van Noordwijk, GCadisch and C Ong. pp. 171–191. CAB International, Walling-ford, UK.
Cannell M G R, van Noordwijk M and Ong C K 1996 The centralagroforestry hypothesis: The trees must acquire resources thatthe crop would not otherwise acquire. Agrofor. Syst. 33, 1–5.
Constantinides M and Fownes J H 1994 Nitrogen mineralizationfrom leaves and litter of tropical plants: Relationship to nitro-gen, lignin and soluble polyphenol concentrations. Soil Biol.Biochem. 26, 49–55.
Gartner T and Cardon Z 2004 Decomposition dynamics in mixed-species leaf litter. Oikos 104, 230–246.
Giller K E and Cadisch G 1995 Future benefits from biological ni-trogen fixation: An ecological approach to agriculture. Plant Soil174, 255–277.
Gine M F, Bergamin-Filho H, Zagatto E A G and Reis B F 1980 Si-multaneous determination of nitrate and nitrite by flow injectionanalysis. Anal. Chim. Acta, 114, 191–197.
Hamblin A P 1985 The influence of soil structure on water move-ment, crop root growth, and water uptake. Adv. Agron. 38,95–155.
Handayanto E, Cadisch G and Giller K E 1994 Nitrogen releasefrom prunings of legume hedgerow trees in relation to quality ofthe prunings and incubation method. Plant Soil 160, 237–248.
Handayanto E, Giller K E Cadisch G 1997 Regulating N re-lease from legume tree prunings by mixing residues of differentquality. Soil Biol. Biochem. 29, 1417–1426.
Huxley P A 1999 Tropical Agroforestry. Blackwell Science, Ox-ford, UK. 371 pp.
Jenkinson D S, Fox R H and Rayner J H 1985 Interactions betweenfertilizer nitrogen and soil nitrogen – The so-called ‘priming’effect. J. Soil Sci. 36, 425–444.

74
Lehmann J 2003 Subsoil root activity in tree-based cropping sys-tems. Plant Soil 255, 319–331.
Lehmann J, Muraoka T and Zech W 2001 Root activity patterns inan Amazonian agroforest with fruit trees determined by 32P, 33Pand 15N applications. Agrofor. Syst. 52,185–197.
Lehmann J, Gebauer G and Zech W 2002 Nitrogen cycling assess-ment in a hedgerow intercropping system using 15N enrichment.Nutrient Cycl. Agroecos. 62, 1–9.
Oorts K, Merckx R, VanLauwe B, Sanginga N and Diels J 2002.Dynamics of Charge Bearing Soil Organic Matter Fractions inHighly Weathered Soils. Proceedings of the 17th World Congressof Soil Science, CD-ROM. International Soil Science Society,Bangkok, Thailand.
Payne R W, Lane P W, Ainsley A E, Bicknell K E, Digby P G N,Harding S A, Leech P K, Simpson H R, Todd A D, Verrier P J andWhite R P 1987 Genstat 5 Reference Manual. Oxford, ClarendonPress. 749 pp.
Rao K V and Shinde J E 1990 Uptake and balance of 15N labelledgreen manures and urea in tropical wetland rice culture. In Sta-ble Isotopes in Plant Nutrition, Soil Fertility and EnvironmentalStudies. Ed. International Atomic Energy Agency. pp. 317–362.IAEA, Vienna, Austria.
Rowe E C and Cadisch G 2002 Implications of heterogeneityon procedures for estimating plant 15N recovery in hedgerowintercrop systems. Agrofor. Syst. 54, 61–70.
Rowe E C, Hairiah K, Giller K E, van Noordwijk M and Cadisch G1999 Testing the safety-net role of hedgerow tree roots by 15Nplacement at different soil depths. Agrofor. Syst. 43, 81–93.
Rowe E C, van Noordwijk M, Suprayogo D, Hairiah K, GillerK E and Cadisch G 2001 Root distributions partially explain15N uptake patterns in Gliricidia and Peltophorum hedgerowintercropping systems. Plant Soil 235, 167–179.
Sanchez P A 1976 Properties and Management of Soils in theTropics. Wiley, New York. pp. 618.
Suprayogo D, van Noordwijk M, Hairiah K and Cadisch G 2002The inherent ‘safety-net’ of Ultisols: measuring and modellingretarded leaching of mineral nitrogen. Eur. J. Soil Sci. 53, 185–194.
Tennant D 1975 A test of modified line intersect method fordetermining root length. J. Ecol. 63, 995–1001.
van der Heide J, Setijono S, Syekhfani M S, Flach E N, Hairiah K,Ismunandar S, Sitompul S M and van Noordwijk M 1992 Canlow external input cropping systems on acid upland soils in thehumid tropics be sustainable? Backgrounds of the UniBraw/IBNitrogen management project in Bunga Mayang (Sunkai Selatan,Kotabumi, S. Sumatera, Indonesia). Agrivita 15, 1–10.
van Noordwijk M and Cadisch G 2002 Access and excess problemsin plant nutrition. Plant Soil 247, 25–39.
van Noordwijk M, Lawson G, Soumare A, Groot J J R and HairiahK 1996 Root distribution of trees and crops: Competition and/orcomplementarity. In Tree-crop Interactions – A PhysiologicalApproach. Eds. C K Ong and P A Huxley. pp. 319–364. CABI,Wallingford, Oxon., UK.
van Noordwijk M, Sitompul S M, Hairiah K, Listyarini E andSyekhfani M S 1995 Nitrogen supply from rotationally or spa-tially zoned inclusion of Leguminosae for sustainable maizeproduction on an acid soil in Indonesia. In Plant-soil Interac-tions at Low pH. Ed. R A Date. pp. 779–784. Kluwer AcademicPublishers.
VanLauwe B, Sanginga N and Merckx R 1998 Recovery of Leu-caena and Dactyladenia residue nitrogen-15 in alley croppingsystems. Soil Sci. Soc. Am. J. 62, 454–460.
Wahid P A, Suresh P R and George S S 2004 Absorption and par-titioning of applied 15N in a black pepper + erythrina system inKerala, India. Agrofor. Syst. 60, 143–147.
Young A 1986 The effects of trees on soils. In Amelioration ofSoil by Trees. Eds. R T Prinsley and M J Swift. pp. 10–19.Commonwealth Science Council, London.
Xu Z H, Myers R J K, Saffigna P G and Chapman A L 1993 Nitro-gen fertilizer in Leucaena alley cropping: II. Residual value ofnitrogen fertilizer and Leucaena residues. Fert. Res. 34, 1–8.
Section editor: J.K. Vessey
Copyright © 2022 FDOKUMEN




















