Nghiªn cøu sö dông kÕt hîp enzyme trong chiÕt t¸ch vµ lµm ...
167
BỘ GIÁO DỤC VÀ ĐÀO TẠO VIỆN HÀN LÂM KHOA HỌC VÀ CÔNG NGHỆ VIỆT NAM HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ HOÀNG THỊ BÍCH Nghiªn cøu sö dông kÕt hîp enzyme trong chiÕt t¸ch vμ lμm giμu mét sè s¶n phÈm nguån gèc thiªn nhiªn LUẬN ÁN TIẾN SĨ HOÁ HỌC HÀ NỘI - 2017
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Nghiªn cøu sö dông kÕt hîp enzyme trong chiÕt t¸ch vµ lµm ...

BỘ GIÁO DỤC VÀ ĐÀO TẠO VIỆN HÀN LÂM KHOA HỌC
VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
HOÀNG THỊ BÍCH
Nghiªn cøu sö dông kÕt hîp enzyme trong chiÕt t¸ch
vµ lµm giµu mét sè s¶n phÈm nguån gèc thiªn nhiªn
LUẬN ÁN TIẾN SĨ HOÁ HỌC
HÀ NỘI - 2017

BỘ GIÁO DỤC VÀ ĐÀO TẠO VIỆN HÀN LÂM KHOA HỌC
VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
HOÀNG THỊ BÍCH
Nghiªn cøu sö dông kÕt hîp enzyme trong chiÕt t¸ch
vµ lµm giµu mét sè s¶n phÈm nguån gèc thiªn nhiªn
Chuyên ngành: Hóa học các hợp chất tự nhiên
Mã số: 62.44.01.17
LUẬN ÁN TIẾN SĨ HOÁ HỌC
Người hướng dẫn khoa học: 1. PGS.TS. Lê Mai Hương
2. PGS.TS. Nguyễn Quyết Chiến
HÀ NỘI - 2017

i
LỜI CAM ĐOAN
Tôi xin cam đoan đây là công trình nghiên cứu của bản thân, được hình thành
và phát triển từ những quan điểm của cá nhân tôi, dưới sự hướng dẫn của PGS.TS.
Lê Mai Hương và PGS.TS. Nguyễn Quyết Chiến, có tham khảo thêm các tài liệu đáng
tin cậy, có nguồn gốc rõ ràng. Các số liệu, kết quả trong luận án là hoàn toàn trung
thực và chưa được công bố trong bất kỳ công trình nào khác.
Tôi xin hoàn toàn chịu trách nhiệm về nội dung luận án này.
Tác giả luận án
Hoàng Thị Bích

ii
LỜI CẢM ƠN
Đầu tiên, tôi xin bày tỏ lòng biết ơn chân thành và sâu sắc nhất tới PGS. TS. Lê
Mai Hương và PGS. TS. Nguyễn Quyết Chiến, những người thầy bằng cả tâm huyết
của mình đã hướng dẫn tôi về khoa học, gợi mở cho tôi các ý tưởng nghiên cứu và chia
sẻ nhiều vấn đề của cuộc sống trong suốt thời gian tôi nghiên cứu luận án.
Tôi xin gửi lời cảm ơn tới GS. TS. Phạm Quốc Long - Viện trưởng Viện Hóa
học các Hợp chất thiên nhiên, cùng ban lãnh đạo Viện, bộ phận đào tạo của Viện Hóa
học các Hợp chất thiên nhiên đã tạo điều kiện rất nhiều cho tôi hoàn thành luận án
của mình
Tôi xin gửi lời cảm ơn tới các thầy, các anh chị phụ trách của Học Viện Khoa
học Công nghệ, đã tạo mọi điều kiện cho tôi hoàn thành luận án của mình.
Tôi xin gửi lời cảm ơn tới các anh chị em đồng nghiệp Trung tâm Nghiên cứu
và Phát triển các SPTN đã tạo điều kiện giúp đỡ tôi trong quá trình hoàn thành công
trình nghiên cứu này.
Tôi cảm ơn đề tài “Nghiên cứu ứng dụng công nghệ sinh học tạo chế phẩm
acid béo đa nối đôi (n3 - PUFA) từ nguyên liệu tự nhiên bổ sung vào thức ăn ương
nuôi một số đối tượng cá biển chủ lực” và “Nghiên cứu thành phần hóa học, điều tra
và đánh giá chất lượng tinh dầu trầm hiện đang được sản xuất ở Việt Nam”, mã số
VAST 04; đã tài trợ kinh phí.
Tôi xin gửi lời tri ân của mình tới gia đình, bạn bè, những người thân luôn
động viên để tôi có động lực trong công việc và hoàn thành công trình nghiên cứu
khoa học này.
Xin chân thành cảm ơn!
Tác giả
Hoàng Thị Bích

iii
MỤC LỤC
Trang
Lời cam đoan ............................................................................................................... i
Lời cảm ơn ................................................................................................................. ii
Mục lục ...................................................................................................................... iii
Danh mục các ký hiệu, các chữ viết tắt .................................................................... vii
Danh mục bảng ......................................................................................................... ix
Danh mục hình vẽ ....................................................................................................... x
MỞ ĐẦU .................................................................................................................... 1
CHƯƠNG 1. TỔNG QUAN TÀI LIỆU .................................................................. 3
1.1. Tổng quan về ứng dụng công nghệ enzyme - giải pháp “Hóa học xanh”
trong chiết xuất và làm giàu các sản phẩm có nguồn gốc thiên nhiên ... 3
1.1.1. “Hóa học xanh” trong chiết xuất và làm giàu các sản phẩm có nguồn
gốc thiên nhiên ............................................................................... 3
1.1.2. Cơ sở ứng dụng enzyme hỗ trợ chiết xuất các sản phẩm có nguồn
gốc thiên nhiên ............................................................................... 4
1.1.3. Một số yếu tố ảnh hưởng đến quá trình thủy phân bởi enzyme ..... 7
1.1.4. Lợi ích và khó khăn khi ứng dụng công nghệ EAE ....................... 9
1.2. Các enzyme sử dụng trong công nghệ enzyme hỗ trợ chưng cất (EAD) và
enzyme hỗ trợ chiết xuất (EAE) ............................................................. 10
1.2.1. Các enzyme phân giải cấu trúc thành tế bào nguyên liệu
lignocellulose ............................................................................... 10
1.2.2. Enzyme protease phân giải cấu trúc nguyên liệu giàu protein ..... 16
1.2.3. Enzyme lipase thủy phân lipid thành axit béo tự do .................... 19
1.3. Tình hình nghiên cứu ứng dụng công nghệ enzyme trong chiết xuất và
làm giàu các hợp chất thiên nhiên trong và ngoài nước......................... 22
1.3.1. Các nghiên cứu trên thế giới ........................................................ 22
1.3.2. Các nghiên cứu trong nước .......................................................... 26
1.4. Tổng quan về quế Cinnamomum cassia ................................................ 27

iv
1.4.1. Giới thiệu về Quế C.cassia ........................................................... 27
1.4.2. Những nghiên cứu về thành phần hóa học C. cassia ................... 29
1.4.3. Những nghiên cứu về hoạt tính sinh học C. cassia ...................... 31
1.4.4. Các phương pháp chiết xuất tinh dầu quế .................................... 33
1.5. Tổng quan về cá ngừ .............................................................................. 35
1.5.1. Sản lượng và giá trị dinh dưỡng của phụ phẩm cá ngừ ................ 35
1.5.2. Dầu cá và các axit béo không no đa nối đôi n-3 PUFA ............... 37
1.5.3. Các phương pháp chiết xuất lipid và làm giàu các axit béo không
no .................................................................................................. 39
CHƯƠNG 2. NGUYÊN VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU ..... 41
2.1. Nguyên liệu ............................................................................................ 42
2.1.1. Mẫu nguyên liệu ........................................................................... 42
2.1.2. Enzyme sử dụng trong nghiên cứu ............................................... 43
2.1.3. Bộ chủng vi sinh vật kiểm định .................................................... 44
2.1.4. Dòng tế bào .................................................................................. 45
2.2. Hóa chất, thiết bị .................................................................................... 45
2.2.1. Hóa chất ........................................................................................ 45
2.2.2. Thiết bị ......................................................................................... 46
2.3. Phương pháp nghiên cứu........................................................................ 46
2.3.1. Các phương pháp phân tích hóa sinh ........................................... 46
2.3.2. Các phương pháp phân tích thành phần hóa học ......................... 46
2.3.3. Các phương pháp xác định hoạt độ enzyme ................................ 47
2.3.4. Các phương pháp thủy phân ......................................................... 49
2.3.5. Các phương pháp xác định sản phẩm thủy phân .......................... 50
2.3.6. Phương pháp làm giàu các axit béo bằng kết tinh ure ................. 52
2.3.7. Các phương pháp nghiên cứu điều kiện tối ưu .................................. 52
2.3.8. Các phương pháp xác định hoạt tính sinh học ............................. 53
CHƯƠNG 3. THỰC NGHIỆM ............................................................................. 54
3.1. Nghiên cứu ứng dụng enzyme kết hợp trong chưng cất tinh dầu từ các bộ
phận khác nhau của cây quế Cinnamomum cassia ................................ 54

v
3.1.1. Nghiên cứu phân tích thành phần chất nền và tinh dầu của mẫu cành
lá và mẫu vỏ quế C. cassia ........................................................... 55
3.1.2. Nghiên cứu tác động của việc xử lý nguyên liệu với enzyme lên quá
trình chưng cất tinh dầu ................................................................ 55
3.1.3. Nghiên cứu tìm điều kiện tối ưu cho quá trình ứng dụng hệ enzyme
kết hợp Laccase-Htec2 ................................................................ 58
3.2. Nghiên cứu thăm dò ứng dụng hệ enzyme kết hợp Laccase-Htec2 trong
chưng cất tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna ...... 60
3.2.1. Nghiên cứu phân tích thành phần chất nền của bột gỗ gió bầu A.
crassna ........................................................................................... 60
3.2.2. Nghiên cứu thăm dò tác động của việc xử lý enzyme Laccase- Htec2
lên quá trình chưng cất tinh dầu từ bột gỗ gió bầu A. crassna ........ 60
3.3. Nghiên cứu ứng dụng enzyme kết hợp trong chiết xuất lipid và làm giàu
các axit béo n-3 PUFA từ đầu cá ngừ vây vàng Thunnus albarcares ... 61
3.3.1. Nghiên cứu thành phần chất nền và lipid tổng của một số loại đầu
cá ngừ của Việt Nam .................................................................... 62
3.3.2. Nghiên cứu ứng dụng enzyme protease trong chiết xuất lipid từ đầu
cá ngừ vây vàng T. albarcares ..................................................... 62
3.3.3. Nghiên cứu ứng dụng enzyme lipase kết hợp ure trong quá trình
làm giàu các axit béo n-3 PUFA của dầu đầu cá ngừ vây vàng T.
albacares ...................................................................................... 64
CHƯƠNG 4. KÊT QUẢ VÀ THẢO LUẬN ......................................................... 66
4.1. Kết quả nghiên cứu ứng dụng enzyme kết hợp trong chưng cất tinh dầu
từ các bộ phận khác nhau của cây quế Cinnamomum cassia................. 66
4.1.1. Kết quả phân tích các chất nền và tinh dầu của cành lá và vỏ quế C.
cassia ............................................................................................ 66
4.1.2. Tác động của việc xử lý nguyên liệu với enzyme lên quá trình chưng
cất tinh dầu từ cành lá quế C. cassia ............................................ 71
4.1.3. Tác động của việc xử lý nguyên liệu với enzyme lên quá trình chưng
cất tinh dầu từ vỏ quế C. cassia ................................................... 85

vi
4.1.4. Kết quả tìm điều kiện tối ưu cho quá trình ứng dụng hệ enzyme kết
hợp Laccase-Htec2 trong chưng cất tinh dầu từ cành lá và vỏ quế . 95
4.2. Kết quả thăm dò ứng dụng hệ enzyme kết hợp Laccase-Htec2 trong chưng
cất tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna ............. 106
4.2.1. Kết quả phân tích thành phần chất nền của bột gỗ gió bầu A.
crassna ........................................................................................ 106
4.2.2. Kết quả thăm dò tác động hệ enzyme Laccase - Htec2 lên hàm lượng
và thành phần tinh dầu trầm hương từ gỗ cây gió bầu A.
crassna ....................................................................................... 107
4.3. Kết quả nghiên cứu ứng dụng enzyme kết hợp trong chiết xuất lipid và làm
giàu các axit béo n-3 PUFA từ đầu cá ngừ vây vàng Thunnus
albarcares ............................................................................................. 111
4.3.1. Kết quả nghiên cứu các chất nền và thành phần axit béo của một số
loại đầu cá ngừ của Việt Nam .................................................... 111
4.3.2. Kết quả nghiên cứu ứng dụng enzyme trong chiết xuất lipid từ đầu
cá ngừ vây vàng T. albarcares ................................................... 115
4.3.3. Kết quả nghiên cứu ứng dụng enzyme lipase CRL trong quá trình
làm giàu các axit béo n-3 PUFA của dầu đầu cá ngừ vây vàng T.
albacares bằng phương pháp ure ............................................... 118
4.3.4. Đề xuất qui trình công nghệ ứng dụng enzyme kết hợp trong chiết
xuất lipid và làm giàu n-3 PUFA từ đầu cá ngừ vây vàng T.
albacares .................................................................................... 126
KẾT LUẬN ................................................................................................ 130
KIẾN NGHỊ ............................................................................................... 132
DANH MỤC CÁC CÔNG TRÌNH CÔNG BỐ CỦA TÁC GIẢ LIÊN
QUAN ĐẾN LUẬN ÁN ............................................................................ 133
DANH MỤC TÀI LIỆU THAM KHẢO ................................................. 134
PHỤ LỤC

vii
DANH MỤC CÁC KÝ HIỆU, CÁC CHỮ VIẾT TẮT
Ký hiệu Tiếng Anh Tiếng Việt
ALA : Alpha linolenic acid Axit Alpha linolenic
CMC : Carboxymethylcellulose Carboxymethylcellulose
DHA : Docosahexaenoic acid Axit Docosahexaenoic
DMSO : Dimethyl Sulphoxide Dimethyl Sulphoxide
DNS : 3,5-dinitrosalicylic acid 3,5-dinitrosalicylic acid
DPA : Docosapentaenoic acid Axit Docosapentaenoic
DPPH : Diphenylpicrylhydrazyl Diphenylpicrylhydrazyl
EAD : Enzyme assisted distillation Chưng cất có enzyme hỗ trợ
EAE : Enzyme assisted extraction Chiết xuất có enzyme hỗ trợ
EAMD : Enzyme assisted microway extraction Chiết xuất có enzyme kết hợp vi
sóng
EDTA : Ethylene Diamine Triacetic Acid Ethylene Diamine Triacetic Acid
EPA : Eicosapentaenoic acid Axit Eicosapentaenoic
FAO : Food and Agriculture Organization of
the United Nations
Tổ chức Lương nông Thế giới
FPH : Fish protein hydrolysate Protein thủy phân từ cá
GC : Gas chromatography Sắc ký khí
GC-MS : Gas chromatography mass spectrometry Sắc ký kết hợp khối phổ
HD : Hydrodistillation Chưng cất lôi cuốn hơi nước
Hep-G2 : Human hepatocellular carcinoma Dòng tế bào ung thư gan
HPLC : High Performanc/e liquid
chromatography
Sắc ký lỏng hiệu năng cao
Htec2 : Cellic HTec2 Enzyme Enzym Cellic HTec2
IC50 : Inhibitory concentration at 50% Nồng độ ức chế 50%
KPH : Không phát hiện
Laccase : Laccase enzyme Enzym laccase

viii
LD100 : Lethal Dose at 100% Liều lượng gây chết cho 100% số
động vật thử nghiệm
LU : Human lung adenocarcinoma Dòng tế bào ung thư phổi
MCF-7 : Michigan Cancer Foundation 7 Dòng tế bào ung thư vú
MIC : Minimum Inhibitory Concentration Nồng độ ức chế tối thiểu
MTT :(3-(4,5-Dimethylthiazol-2-yl)-2,5-
Diphenyltetrazolium Bromide
(3-(4,5-Dimethylthiazol-2-yl)-2,5-
Diphenyltetrazolium Bromide
MUFA : Monounsaturated fatty acid Axit béo không no 1 nối đôi
NO : Nitric oxid Nitơ mônôxit
PUFA : Polyunsaturated fatty acid Axit béo không no đa nối đôi
RD : Human rhabdomyosarcoma Dòng tế bào ung thư mô liên kết
SCFE : Supercritical fluid extraction Chiết Siêu tới hạn
SE : Solvent extraction Chiết xuất bằng dung môi
SFA : Saturated fatty acid Axit béo no
TCVN : Tiêu chuẩn Việt Nam
TLC : Thin layer chromatography Sắc ký lớp mỏng
TT : Thứ tự
TS : Transition state Trạng thái chuyển
USD : United States dollar Đô la Mỹ
USDA : United States Department of
Agriculture
Bộ Nông nghiệp Hoa kỳ
VSV : Vi sinh vật

ix
DANH MỤC BẢNG
Trang
Bảng 1.1. Thành phần lignocellulose trong một số loại sinh khối.........................11
Bảng 1.2. Tỉ lệ các thành phần của cá ngừ ...................................................36
Bảng 3.1. Một số thông số thủy phân nguyên liệu cành lá quế và vỏ quế..............56
Bảng 3.2. Một số thông số của quá trình thủy phân bởi enzyme protease..............62
Bảng 4.1. Kết quả phân tích chất nền của nguyên liệu ...........................................66
Bảng 4.2. Kết quả phân tích thành phần hóa học của tinh dầu ...............................67
Bảng 4.3. Kết quả khảo sát tác động của việc xử lý nguyên liệu với enzyme lên mức
độ thủy phân các chất nền và hiệu suất chưng cất tinh dầu từ cành lá quế..............72
Bảng 4.4. Hàm lượng tinh dầu thu nhận theo thời gian chưng cất mẫu cành lá quế có
và không có enzyme hỗ trợ.......................................................................................73
Bảng 4.5. Kết quả phân tích GC-MS các thành phần hóa học của tinh dầu thu được
từ cành lá quế có và không có xử lý với enzyme......................................................79
Bảng 4.6a. Kết quả thử hoạt tính kháng vi sinh vật kiểm định.................................81
Bảng 4.6b. Kết quả thử hoạt tính gây độc tế bào trên 03 dòng tế bào ung thư.........82
Bảng 4.6c. Kết quả thử khả năng ức chế sự sinh NO trên tế bào RAW 264.7.........83
Bảng 4.6d. Kết quả thử hoạt tính chống oxy hóa trên hệ DPPH...............................84
Bảng 4.7. Kết quả khảo sát tác động của việc xử lý nguyên liệu với enzyme lên mức
độ thủy phân thành tế bào và hiệu suất chưng cất tinh dầu từ vỏ quế......................85
Bảng 4.8. Hàm lượng tinh dầu theo thời gian chưng cất mẫu vỏ quế có và không có
enzyme hỗ trợ...........................................................................................................87
Bảng 4.9. Kết quả phân tích GC-MS các thành phần hóa học của tinh dầu thu được
từ vỏ quế có và không có xử lý với enzyme.............................................................93
Bảng 4.10a. Giá trị ở các mức của các yếu tố ảnh hưởng.......................................100
Bảng 4.10b. Ma trận kế hoạch hóa và kết quả thực nghiệm...................................101
Bảng 4.11a. Kết quả phân tích thành phần bột gỗ gió bầu A. crassna...................107
Bảng 4.11b. Kết quả phân tích thành phần hóa học của tinh dầu trầm hương thu được
từ gỗ cây gió bầu A. crassna bằng 2 phương pháp khác nhau.....................................107
Bảng 4.12. Thành phần các chất nền của một số loại đầu cá ngừ của Việt Nam...........111

x
Bảng 4.13. Thành phần axit béo trong các mẫu lipid thu được từ đầu một số loài cá
ngừ của Việt Nam..................................................................................................113
Bảng 4.14. Sản phẩm của quá trình chiết xuất lipid..............................................115
Bảng 4.15. Thành phần axit béo của các lipid chiết xuất từ đầu cá ngừ vây vàng
T. albacare bằng các phương pháp khác nhau....................................................116
Bảng 4.16a. Ảnh hưởng của pH đến quá trình thủy phân lipid thành axit béo tự do
của lipase CRL......................................................................................................119
Bảng 4.16b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng của yếu tố pH......120
Bảng 4.17a. Ảnh hưởng của nhiệt độ đến quá trình thủy phân lipid thành axit béo tự
do của lipase CRL..................................................................................................120
Bảng 4.17b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng của yếu tố nhiệt
độ...........................................................................................................................121
Bảng 4.18a. Ảnh hưởng tỷ lệ enzyme/cơ chất đến quá trình thủy phân lipid thành
axit béo tự do của lipase CRL...............................................................................121
Bảng 4.18b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng tỷ lệ enzyme/cơ chất
...............................................................................................................................122
Bảng 4.19a. Ảnh hưởng thời gian thủy phân đến quá trình thủy phân lipid thành axit
béo tự do của enzyme lipase..................................................................................122
Bảng 4.19b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng của thời gian thủy phân
...............................................................................................................................122
Bảng 4.20. Thành phần axit béo trước và sau khi làm giàu n-3 PUFA.................124
Bảng 4.21a. Các chỉ tiêu hóa lý của sản phẩm axit béo giàu n-3 PUFA................129
Bảng 4.21b. Các thành phần axit amin tự do của sản phẩm bột đạm hòa tan giàu axit
amin........................................................................................................................129

xi
DANH MỤC HÌNH VẼ
Trang
Hình 1.1. So sánh năng lượng của phản ứng có và không có xúc tác nhờ
enzyme ............................................................................................ 5
Hình 1.2. Chiết xuất các hoạt chất liên kết với mạng lưới lignocellulose 6
Hình 1.3. Công nghệ enzyme kết hợp dung môi sinh học chiết xuất các hợp
chất thiên nhiên ............................................................................... 7
Hình 1.4a. Các đơn vị cơ bản của lignin ....................................................... 11
Hình 1.4b. Cấu trúc của lignin ...................................................................... 12
Hình 1.5a. Công thức hóa học của cellulose ................................................ 13
Hình 1.5b. Cơ chế phân giải cellulose của hệ enzyme cellulase ................. 14
Hình 1.6a. Cấu trúc polymer xylan .............................................................. 15
Hình 1.6b. Enzyme xylanolytic liên quan đến quá trình phân giải xylan ..... 15
Hình 1.7. Phản ứng thủy phân protein của enzyme protease ........................ 17
Hình 1.8. Phân tử lipid và các vị trí thủy phân của enyme lipase ................. 20
Hình 1.9a. Cành lá và hoa C. cassia ............................................................. 27
Hình 1.9b. Cây C. cassia ............................................................................... 27
Hình 1.10a. Một số polyphenol trong quế Cinnamomum............................. 29
Hình 1.10b. Một số thành phần chính và phụ trong tinh dầu C. cassia ........ 30
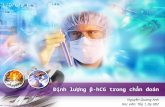
Hình 1.11. Sơ đồ chưng cất tinh dầu quế có nồi hơi riêng............................ 34
Hình 1.12. Cấu trúc của một số acid béo nhóm n-3 PUFA .......................... 38
Hình 2.1. Mẫu cành lá quế Cinnamomum cassia .......................................... 42
Hình 2.2. Mẫu vỏ quế Cinnamomum cassia ................................................. 42
Hình 2.3. Mẫu bột gỗ cây gió bầu Aquilaria crassna ................................... 42
Hình 2.4. Mẫu đầu cá ngừ vây vàng Thunnus albacares .............................. 43
Hình 2.5. Đồ thị đường chuẩn Glucose theo phương pháp DNS .................. 51
Hình 2.6. Đồ thị đường chuẩn protein .......................................................... 51
Hình 3.1. Qui trình nghiên cứu ứng dụng enzyme trong chiết xuất tinh dầu 54
Hình 3.2. Quy trình nghiên cứu ứng dụng enzyme trong chiết xuất lipid và
làm giàu n-3 PUFA từ đầu cá ngừ ................................................ 61

xii
Hình 3.3. Ba pha sản phẩm sau ly tâm .......................................................... 63
Hình 4.1a. Một số thành phần thuộc nhóm phenylpropanoid ....................... 69
Hinh 4.1b. Một số thành phần thuộc nhóm sesquiterpenoid ......................... 70
Hình 4.2. Đồ thị biểu diễn mối tương quan giữa mức độ thủy phân các chất
nền và hiệu suất chưng cất tinh dầu từ cành lá quế ...................... 72
Hình 4.3. Đồ thị biểu diễn hàm lượng tinh dầu theo thời gian chưng cất mẫu
cành lá quế có và không có enzyme hỗ trợ ................................... 74
Hình 4.4a. Sắc ký đồ của tinh dầu cành lá quế chưng cất theo cách thông
thường, nguyên liệu không qua xử lý enzyme ............................ 75
Hình 4.4b. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với enzyme
Laccase EAD .............................................................................. 76
Hình 4.4c. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với enzyme
Htec2 EAD ................................................................................. 77
Hình 4.4d. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với hệ
enzyme Laccase-Htec2 ............................................................... 78
Hình 4.5. Đồ thị biểu diễn mối tương quan giữa mức độ thủy phân thành tế
bào và tỷ lệ gia tăng tinh dầu vỏ quế ............................................ 86
Hình 4.6. Đồ thị biểu diễn hàm lượng tinh dầu theo thời gian chưng cất mẫu
vỏ quế có và không có enzyme hỗ trợ .......................................... 87
Hình 4.7a. Sắc ký đồ tinh dầu vỏ quế, nguyên liệu không xử lý enzyme ..... 89
Hình 4.7b. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với hệ
enzyme Laccase- EAD ............................................................... 90
Hình 4.7c. Sắc ký đồ tinh dầu vỏ quế, nguyên liệu qua xử lý với hệ enzyme
Htec2-EAD ................................................................................. 91
Hình 4.7d. Sắc ký đồ tinh dầu vỏ quế, nguyên liệu qua xử lý với hệ enzyme
Laccase - Htec2- EAD ................................................................ 92
Hình 4.8a. Đồ thị biểu diễn ảnh hưởng của của pH lên hàm lượng đường
khử ................................................................................................ 96
Hình 4.8b. Đồ thị biểu diễn ảnh hưởng của của nhiệt độ lên hàm lượng
đường khử ................................................................................... 97

xiii
Hình 4.8c. Đồ thị biểu diễn ảnh hưởng của tỷ lệ Laccase/cơ chất lên hàm
lượng đường khử ......................................................................... 97
Hình 4.8d. Đồ thị biểu diễn ảnh hưởng của tỷ lệ Htec2/cơ chất lên hàm
lượng đường khử ......................................................................... 98
Hình 4.8e. Bảng và đồ thị biểu diễn ảnh hưởng thời gian lên hàm lượng
đường khử ................................................................................... 99
Hình 4.9a. Mặt đáp ứng vùng tối ưu của cặp yếu tố x1x2 .......................... 104
Hình 4.9b. Mặt đáp ứng vùng tối ưu của cặp yếu tố x2x4 .......................... 104
Hình 4.9c. Mặt đáp ứng vùng tối ưu của cặp yếu tố x3x5 .......................... 104
Hình 4.10. Sơ đồ quy trình công nghệ ứng dụng hệ enzyme kết hợp Laccase-
Htec trong chưng cất tinh dầu từ cành lá quế C. cassia ............ 105
Hình 4.11. Một số thành phần đặc trưng của tinh dầu trầm hương ............ 110
Hình 4.12. Tinh thể ure kết hợp với axit béo bão hòa................................. 123
Hình 4.13. Sắc ký đồ các axit béo sau khi làm giàu ................................... 124
Hình 4.14. Sơ đồ quy trình công nghệ ứng dụng enzyme kết hợp trong chiết
xuất lipid và làm làm giàu n-3 PUFA từ đầu cá ngừ vây vàng. 126

1
MỞ ĐẦU
Ô nhiễm môi trường cũng như sức khỏe người tiêu dùng đã dẫn đến nhu cầu
cấp thiết là tìm ra các phương pháp thu nhận các sản phẩm có nguồn gốc thiên nhiên
theo hướng “Xanh” nhằm tăng năng suất, giảm thời gian, tận dụng hiệu quả nguồn
nguyên liệu và giảm ô nhiễm môi trường. Rất nhiều các nỗ lực nghiên cứu để cải
thiện năng suất cũng như chất lượng các hoạt chất, trong đó ứng dụng công nghệ
enzyme hỗ trợ quá trình chiết xuất các hợp chất thiên nhiên đang phát triển mạnh mẽ
trong vài chục năm trở lại đây và bước đầu đưa ra tính khả thi trong công nghiệp. Đây
được coi là một trong những giải pháp “Xanh” trong chiết xuất các HCTN do giảm
lượng dung môi, sử dụng các dung môi thay thế, tạo ra các sản phẩm sạch “organic”
và phế liệu của quá trình cũng ở dạng dễ phân hủy, giảm nguy cơ gây ô nhiễm môi
trường.
Nằm trong vùng khí hậu nhiệt đới gió mùa ẩm cộng với đường bờ biển trải dài
qua nhiều vĩ tuyến khác nhau nên nguồn tài nguyên thiên nhiên của Việt Nam vô
cùng phong phú với hệ thực vật đặc trưng và nguồn sinh vật biển đa dạng. Trong đó
phải kể đến cây Quế (nguồn dược liệu) và cá ngừ (nguồn lợi thủy sản) là những mặt
hàng có giá trị kinh tế, giá trị sử dụng và giá trị xuất khẩu đầy tiềm năng. Sản phẩm
dầu từ quế và cá ngừ ngày càng được chú trọng bởi chúng có giá trị cao. Tinh dầu
quế là mặt hàng xuất khẩu chiến lược, dầu cá ngừ chứa hàm lượng lớn các acid béo
không no đa nối đôi PUFAs (polyunsaturated fatty acids), đặc biệt là nhóm axit béo
không no đa nối đôi thiết yếu omega 3 như DHA và EPA có vai trò quan trọng trong
các lĩnh vực y, dược và công nghiệp thực phẩm. Tuy nhiên trong công nghiệp, quá
trình sản xuất hai loại dầu này vẫn sử dụng các phương pháp truyền thống (cất cuốn
hơi nước, ép nhiệt) chưa phát huy được hết hiệu quả chiết xuất, gây lãng phí nguồn
nguyên liệu.
Ngày nay, những tiến bộ trong công nghệ sinh học, công nghệ enzyme có khả
năng sản xuất một lượng lớn enzyme có hoạt lực cao, phổ cơ chất rộng... thì công
nghệ “XANH” ứng dụng enzyme hỗ trợ quá trình chiết xuất các hợp chất thiên nhiên
(enzyme assisted extraction- EAE) nhằm tăng hiệu suất, thay thế dung môi hữu cơ là

2
vấn đề cấp thiết [134]. Nhằm mục đích tăng hiệu suất chiết tách, phát triển hướng
nghiên cứu mới - chiết xuất và làm giàu các HCTN bằng enzyme hỗ trợ, chúng tôi
lựa chọn đề tài luận án: “Nghiên cứu sử dụng kết hợp hệ enzyme trong chiết tách
và làm giàu một số sản phẩm nguồn gốc thiên nhiên” với các đối tượng được lựa
chọn là cành lá quế, vỏ quế C. cassia, gỗ gió bầu A. crassna và phụ phẩm đầu cá ngừ.
Trong nghiên cứu này, quá trình chưng cất tinh dầu, quá trình chiết xuất lipid, làm
giàu các n-3 PUFA từ phụ phẩm đầu cá ngừ được thử nghiệm kết hợp với một số
enzyme có sẵn nhằm đánh giá hiệu quả tác động của enzyme lên chất lượng sản phẩm
tạo thành. Một số thông số tối ưu của quá trình xử lý bằng enzyme được khảo sát và
tối ưu hóa bằng qui hoạch thực nghiệm.
Mục đích nghiên cứu
Nghiên cứu ứng dụng công nghệ enzyme vào quá trình chiết xuất và làm giàu
các sản phẩm tinh dầu (từ lá và cành quế, vỏ quế Cinnamomum cassia, trầm hương
từ gỗ gió bầu Aquilaria crassna) và các axit béo n-3PUFA từ phụ phẩm đầu cá ngừ
nhằm làm tăng hiệu quả và phát triển hướng nghiên cứu “XANH” trong khai thác các
SPTN.
Nội dung chính của luận án bao gồm:
- Nghiên cứu ứng dụng enzyme kết hợp trong chưng cất tinh dầu từ các bộ
phận khác nhau của cây quế Cinnamomum cassia: tác động của việc xử lý nguyên
liệu với enzyme được đánh giá qua hiệu suất, thời gian chưng cất, chất lượng sản
phẩm tinh dầu (thành phần hóa học, hoạt tính sinh học).
- Nghiên cứu thăm dò ứng dụng hệ enzyme kết hợp Laccase-Htec2 trong
chưng cất tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna.
- Nghiên cứu ứng dụng enzyme kết hợp trong chiết xuất lipid và làm giàu các
axit béo n-3 PUFA từ đầu cá ngừ vây vàng Thunnus albarcares: tác động của enzyme
lên quá trình được đánh giá qua thành phần axit béo n-3 PUFA.

3
CHƯƠNG 1. TỔNG QUAN TÀI LIỆU
1.1. Tổng quan về ứng dụng công nghệ enzyme - giải pháp “Hóa học
xanh” trong chiết xuất và làm giàu các sản phẩm có nguồn gốc thiên nhiên
1.1.1. “Hóa học xanh” trong chiết xuất và làm giàu các sản phẩm có nguồn
gốc thiên nhiên
Mọi sinh vật, đặc biệt là thực vật trên cạn và sinh vật biển đều có chứa các
hoạt chất sinh học có khả năng ứng dụng trong thực phẩm, dược phẩm, mỹ phẩm...
Những hợp chất này được gọi là những hợp chất thiên nhiên có hoạt tính sinh học.
Phương pháp chiết xuất sử dụng dung môi hữu cơ (solvent extraction- SE) vẫn
đang được sử dụng rộng rãi trong chiết xuất các hợp chất thiên nhiên từ các nguyên
liệu khác nhau như trầm tích, đất, polymer, vi khuẩn, nấm, tảo, vi tảo và phổ biến
nhất là thực vật. Ưu điểm của phương pháp sử dụng dung môi là dễ thao tác, tuy
nhiên, các dung môi lại độc cho người và nguy hại đến môi trường. Nhận thức này
đã dẫn đến nhu cầu ngày càng tăng cho các sản phẩm được sản xuất thông qua quá
trình thân thiện với môi trường còn được gọi “Hóa học xanh” [121].
Hóa học Xanh, còn gọi là hóa học bền vững là một phần của hóa học và công
nghệ hóa học tập trung vào việc tạo ra những sản phẩm, qui trình giảm thiểu việc sử
dụng hoặc phát sinh các chất độc hại. Hóa học xanh tập trung vào phương pháp tiếp
cận công nghệ để ngăn ngừa ô nhiễm, giảm tiêu thụ nguồn tài nguyên không thể tái
tạo. Trong chiết xuất các hợp chất thiên nhiên, quá trình chiết xuất “Xanh” liên quan
đến việc sử dụng các dung môi thân thiện với môi trường sinh thái, nguyên liệu bền
vững và tái tạo, giảm chất thải gây ô nhiễm môi trường [86].
Rất nhiều phương pháp để “Xanh hóa” những qui trình công nghệ hóa học.
Những phương pháp này có thể thực hiện riêng lẻ hoặc phối hợp với các qui trình của
công nghệ hóa học, nhằm mục tiêu làm tăng hiệu suất và giảm lượng thải độc hại.
Trong báo cáo thường niên (xuất bản năm 2011) của cơ quan quản lý năng lượng và
môi trường Pháp (French Environment and Energy Management Agency) với tựa đề:
“Khó khăn khi thay thế hợp chất hữu cơ dễ bay hơi trong quá trình công nghiệp”
đã trình bày các “giải pháp khắc phục kỹ thuật, hạn chế hoặc thay thế các dung môi

4
truyền thống” [86]. Hai trong số các giải pháp đặt ra đã hướng tới ứng dụng công
nghệ enzyme với tự đề:
+ Giải pháp cơ học kết hợp tiền xử lý enzyme
+ Chiết nước kết hợp tiền xử lý enzyme
Ngày nay, những tiến bộ trong xúc tác enzyme, sự đa dạng và sẵn có của các
enzyme công nghiệp cộng với tình trạng ô nhiễm môi trường ngày càng gia tăng thì
các giải pháp công nghệ, trong đó có công nghệ chiết xuất có enzyme hỗ trợ
(enzyme assisted extraction, viết tắt là EAE) nhằm thay thế dung môi hữu cơ là vấn
đề cấp thiết [134].
1.1.2. Cơ sở ứng dụng enzyme hỗ trợ chiết xuất các sản phẩm có nguồn gốc
thiên nhiên
Enzyme là nhóm protein chuyên biệt hóa cao có vai trò và chức năng sinh học
quan trọng bậc nhất đối với tế bào và cơ thể sống. Enzyme có khả năng xúc tác với
độ đặc hiệu cơ chất lớn, xúc tác đặc hiệu các phản ứng hóa học khác khó thể thực
hiện [14].
Enzyme có tính chất ưu việt hơn hẳn các chất xúc tác hóa học, có hoạt tính xúc
tác lớn. Ở điều kiện thích hợp, hầu hết các enzyme xảy ra với tốc độ nhanh gấp 108 -
1011 lần so với phản ứng không có chất xúc tác. Bên cạnh đó, enzyme có tính đặc
hiệu cao và tác dụng trong điều kiện êm dịu, nhiệt độ thích hợp để enzyme hoạt động
30-50oC, pH trung tính và áp suất thường. Đặc biệt, hầu hết các enzyme có nguồn
gốc tự nhiên không độc. Điều này có ý nghĩa quan trọng trong công nghiệp thực phẩm
và y học [32].
Sinh vật thường được cấu tạo từ những đại phân tử tương đối trơ (polypeptid,
polysaccharid) không có khả năng hòa tan trong dung môi hữu cơ [25]. Các hợp chất
thiên nhiên có hoạt tính sinh học thường tồn tại ở dạng không tan hoặc lưu giữ trong
các bào quan dự trữ, hoặc tồn tại tại ở dạng keo kết hợp với các thành phần vách tế
bào [108]. Muốn cắt đứt các liên kết và giải phóng chúng ra khỏi tế bào cần năng
lượng hoạt hóa, tức là mức năng lượng các chất tham gia phản ứng phải đạt được để
cắt đứt các liên kết cần thiết và hình thành các liên kết mới. Về lý thuyết, tốc độ phản
ứng hóa học được xác định bởi giá trị năng lượng hoạt hóa. Năng lượng hoạt hóa
càng lớn thì tốc độ phản ứng càng nhanh và ngược lại.

5
Bất kỳ enzyme nào cũng xúc tác theo trình tự sau:
E + S ES EP E+ P
Trong đó, E là enzyme, S là cơ chất, P là sản phẩm phản ứng. ES và EP là
phức hợp enzyme với cơ chất và sản phẩm.
Sơ đồ trên Hình 1.1 cho thấy đường cong của năng lượng phản ứng có và
không có xúc tác enzyme
Hình 1.1. So sánh năng lượng của phản ứng được và không được xúc tác nhờ enzyme [14]
Ghi chú: TSc1, TSc2 và TSc3 là các trạng thái chuyển (transition state)
[ES]: tổ hợp phức hệ enzyme-cơ chất
[EP]: tổ hợp phức hệ enzyme- sản phẩm
- Con đường không có xúc tác chỉ đi qua trạng thái chuyển tiếp TS* với năng
lượng tự do Gkhông xúc tác cao, cần mức năng lượng hoạt hóa cao hơn.
- Con đường đi qua phản ứng xúc tác enzyme đi qua các trạng thái chuyển tiếp
TSc1, TSc2 và TSc3, với năng lượng tự do Gcó xúc tác thấp. Chất xúc tác làm giảm năng
lượng hoạt hóa của phản ứng hóa học, nó chỉ tham gia vào các giai đoạn trung gian
mà không tham gia trực tiếp vào phản ứng. Năng lượng liên kết do các tương tác yếu
tạo ra ở trạng thái chuyển tiếp được sử dụng để làm căng hoặc uốn gấp khúc cơ chất,
tạo điều kiện cho enzyme tiếp xúc với cơ chất dễ dàng. Chính vì vậy, enzyme tạo
được thế năng đặc biệt có lợi nhất về mặt năng lượng để thực hiện phản ứng, tạo ra
vô số sản phẩm trong điều kiện nhiệt độ thấp, áp suất thường.

6
Trái với phương pháp dung môi, tách các HCTN nhờ vào mức độ phân cực của
chúng, phương pháp ứng dụng enzyme sử dụng nước là dung môi đặc biệt để hòa tan,
phá vỡ các rào cản ngăn cản sự giải phóng hoạt chất. Lúc này, các hoạt chất đang bị
lưu giữ trong cấu trúc khép kín được giải phóng một cách tự nhiên (Hình 1.2).
Hình 1.2. Chiết xuất các hoạt chất liên kết với mạng lưới lignocellulose [91]
Hình 1.2 miêu tả quá trình chiết xuất hoạt chất liên kết với mạng lưới
lignocellulose thành tế bào thực vật (chủ yếu bao gồm cellulose, hemicellulose và
lignin). Quá trình phân giải chất nền (matrix, ma trận) của thành tế bào đã thúc đẩy
quá trình giải phóng các hoạt chất được dễ dàng và thuận lợi.
Quá trình chiết xuất lipid dưới tác dụng của enzyme protease cũng được áp
dụng nhiều trên nhiều đối tượng sinh vật biển và các hạt có dầu (cấu trúc nguyên liệu
chủ yếu là protein và lipid). Các phân tử lipid thường bị lưu giữ trong chất nền protein
của nguyên liệu. Các enzyme protease thủy phân protein màng tế bào cũng như bên
trong tế bào chất thành các peptid nhỏ hơn và các axit amin, từ đó nới lỏng sự toàn
vẹn của cấu trúc, thúc đẩy quá trình giải phóng dầu khỏi chất nền protein khi kết hợp
với các phương pháp ly tâm hoặc gia nhiệt [128]. Toàn bộ quá trình thủy phân diễn
ra trong điều kiện nhẹ nhàng ở nhiệt độ thấp (40-60oC) làm cho các chất có hoạt tính
sinh học gần như không bị thay đổi

7
Do tính chất kỵ nước của các cấu tử không hòa tan vào trong nước nên sau xử
lý enzyme, biện pháp cơ học hoặc vật lý được sử dụng để phân tách các hoạt chất ra
khỏi hỗn hợp. Ngày nay, các ứng dụng của công nghệ enzyme trong chiết xuất các
HCTN rất đa dạng, sự kết hợp enzyme với các dung môi sinh học thay thế dung môi
độc hại đã khiến cho EAE thực hiện trên cả các nguyên liệu không phải cây chứa dầu
đã mang đến sự đa dạng cho các sản phẩm được hình thành (Hình 1.3).
Hình 1.3. Công nghệ enzyme kết hợp dung môi sinh học chiết xuất các hợp chất
thiên nhiên
1.1.3. Một số yếu tố ảnh hưởng đến quá trình thủy phân bởi enzyme
Tốc độ thủy phân bằng enzyme chịu ảnh hưởng của rất nhiều yếu tố [32]:
Ảnh hưởng của nồng độ enzyme: khi nồng độ enzyme thấp, lượng cơ chất
lớn, vận tốc thủy phân phụ thuộc tuyến tính vào nồng độ enzyme. Khi nồng độ
enzyme tăng, tốc độ phản ứng thủy phân tăng đến một giá trị giới hạn V = Vmax thì
nếu nồng độ enzyme tiếp tục tăng, tốc độ phản ứng thủy phân bởi enzyme tăng không
đáng kể, thậm chí không tăng.
Ảnh hưởng của nồng độ cơ chất: Nồng độ cơ chất có ảnh hưởng lớn tới tốc
độ thủy phân, khi càng tăng nồng độ cơ chất, tốc độ phản ứng thủy phân càng tăng,
nhưng khi tốc độ phản ứng thủy phân đạt đến giới hạn V = Vmax, nếu tiếp tục tăng
nồng độ cơ chất, vận tốc phản ứng hầu như không tăng nữa.
Khi nồng độ cơ chất thấp, nhiều phân tử enzyme có trung tâm hoạt động tự do
và sự cung cấp hạn chế cơ chất sẽ xác định tốc độ phản ứng. Ngược lại nồng độ cơ

8
chất cao, hầu hết các trung tâm hoạt động bị chiếm lĩnh do đó lúc này số lượng phân
tử enzyme lại là yếu tố quyết định phản ứng.
Ảnh hưởng của các chất kìm hãm: chất kìm hãm (hay chất ức chế) là những
chất vô cơ hay hữu cơ mà khi có sự hiện diện của chúng, enzyme có thể bị giảm hoặc
mất hoạt tính. Với mỗi enzyme ta có các chất kìm hãm khác nhau, vì vậy, khi sử dụng
enzyme ta phải biết rõ các chất kìm hàm nó để điều chỉnh phản ứng.
Ảnh hưởng của các chất hoạt hóa: chất hoạt hóa là những chất khi có mặt
trong phản ứng có tác dụng làm tăng hoạt tính enzyme, các chất này có bản chất hóa
học khác nhau, có thể là ion kim loại, anion hoặc các chất hữu cơ. Tuy nhiên, các chất
hoạt hóa chỉ có tác dụng giới hạn nồng độ xác định. Khi dùng quá nồng độ cho phép,
hoạt độ enzyme này sẽ giảm.
Ảnh hưởng của nhiệt độ: bản chất của enzyme là protein nên khi tăng hay
giảm nhiệt độ thường có thể ảnh hưởng tới hoạt tính xúc tác của enzyme, enzyme thể
hiện hoạt tính cao nhất ở một giới hạn nhiệt độ thích hợp nhất định. Thông thường
đối với đa số enzyme thì nhiệt độ thích hợp nằm trong khoảng từ 40 - 50oC, ở nhiệt
độ lớn hơn 70oC đa số enzyme bị mất hoạt tính. Do vậy, nhiệt độ 70oC gọi là nhiệt
độ tới hạn của enzyme.
Tốc độ của phản ứng tăng lên cùng với sự tăng của nhiệt độ. Nhưng khi vượt
quá phạm vi nào đó, các phản ứng được enzyme xúc tác bị ảnh hưởng do sự biến tính
của phân tử protein-enzyme. Kết quả này phụ thuộc vào nhiệt độ tối thích của enzyme,
là nhiệt độ mà tại đó tốc độ phản ứng enzyme đạt cực đại. Mỗi enzyme có nhiệt độ
tối thích khác nhau. Sự khác nhau này tùy thuộc vào nguồn gốc của các enzyme, tùy
theo từng điều kiện từng sự khác nhau về tính nhạy cảm với nhiệt độ của phân tử
protein-enzyme.
Ảnh hưởng của pH: pH có ảnh hưởng mạnh mẽ đến hoạt tính của enzyme vì
pH ảnh hưởng đến mức độ ion hóa cơ chất. Đa số enzyme có khoảng pH thích hợp
trong vùng trung tính từ 5-7, một số enzyme protease, pH thích hợp nằm trong vùng
axit (pepsin,…) hoặc nằm trong vùng kiềm (trypsin, subtilin,...). Với từng enzyme,
giá trị pH thích hợp có thể thay đổi khi nhiệt độ, loại cơ chất,... thay đổi.

9
Ảnh hưởng của thời gian thủy phân: thời gian thủy phân cần thích hợp để
enzyme phân cắt các liên kết trong cơ chất, tạo thành các sản phẩm cần thiết của quá
trình thủy phân nhằm đảm bảo hiệu suất thủy phân cao, chất lượng sản phẩm tốt. Thời
gian thủy phân dài, ngắn khác nhau tùy thuộc vào từng loại enzyme, nồng độ cơ chất,
pH, nhiệt độ, sự có mặt của chất hoạt hóa, ức chế... Trong thực tế, thời gian thủy phân
phải xác định bằng thực nghiệm và kinh nghiệm thực tế cho từng quá trình thủy phân
cụ thể.
Ngoài ra, còn một số các yếu tố khác ảnh hưởng đến tốc độ phản ứng như tỷ
lệ nước/cơ chất, tốc độ khuấy,...
1.1.4. Lợi ích và khó khăn khi ứng dụng công nghệ EAE
Lợi ích của công nghệ EAE
- Công nghệ EAE ảnh hưởng đến năng suất và chất lượng sản phẩm
Trong thực tế, các phương pháp chiết xuất truyền thống thường sử dụng
phương pháp vật lý như ép, cất... chưa cho hiệu quả cao. Công nghệ EAE thường cho
kết quả tăng hiệu suất do tác động phân giải thành tế bào của enzyme đã giải phóng
dễ dàng hơn các hoạt chất vốn bị lưu giữ trong cấu trúc chất nền thành tế bào (thực
vật) hay chất nền protein tăng tính thấm của màng tế bào do đó dẫn đến năng suất cao
hơn khi chưng cất. [65], [81], [94], [161]. Mặt khác, enzyme thường hoạt động trong
điều kiện nhiệt độ thường 40-50oC, pH trung tính 5-7 nên ít ảnh hưởng đến hoạt chất
chiết xuất, cho chất lượng sản phẩm thường cao hơn so với các phương pháp truyền
thống (sử dụng hóa chất, nhiệt độ, áp suất cao...).
- Công nghệ EAE cho các sản phẩm phụ có giá trị gia tăng
Bên cạnh các sản phẩm chính của quá trình, EAE còn có các sản phẩm thủy
phân có giá trị gia tăng như các protein, polysaccharid, polyphenol... Quá trình thu
nhận các sản phẩm này không những không ảnh hưởng đến chất lượng của sản phẩm
chính mà còn có tác động có lợi giúp cho quá trình giải phóng hoạt chất thuận lợi
hơn, giảm sự biến chất của sản phẩm.
- Công nghệ EAE tác động lên thời gian chiết
Các phương pháp sử dụng dung môi hữu cơ cần thời gian ngâm chiết kéo dài
nếu thiếu sự hỗ trợ của siêu âm, vi sóng, soxhlet... Enzyme có cường lực xúc tác lớn,

10
tốc độ phản ứng nhanh, chỉ vài phút (ribonucelase), vài giờ (protease,...). Sau quá
trình thủy phân, các hoạt chất đã được giải phóng ở dạng tự do hoặc liên kết lỏng lẻo
với cơ chất, chỉ cần một tác động nhỏ như năng lượng nhiệt hay ly tâm cũng đủ để
phân tách chúng khỏi hỗn hợp. Do đó, thời gian chiết xuất nhanh hơn so với không
xử lý enzyme.
- Công nghệ EAE thân thiện với môi trường
EAE đã giảm thiểu việc sử dụng dung môi, giảm lượng phế thải, chuyển hóa
phế liệu thành dạng đơn giản, có thể tận dụng triệt để nguồn nguyên liệu, giảm các
nguy cơ gây ô nhiễm môi trường.
Khó khăn khi ứng dụng công nghệ EAE
Công nghệ EAE còn hạn chế về khả năng thương mại và công nghệ như:
- Giá các enzyme là tương đối đắt khi xử lý khối lượng lớn nguyên liệu thô;
- Chế phẩm enzyme có sẵn có thể không hoàn toàn thủy phân thành tế bào
thực vật, hạn chế hiệu suất chiết xuất các hợp chất; Vì vậy, tìm được hệ enzyme thích
hợp để phân giải cơ chất là hết sức quan trọng.
- Enzyme dễ chịu ảnh hưởng của điều kiện môi trường. Ở quy mô khác nhau,
enzyme có sự biến động về điểm hoạt động tối ưu như như tỷ lệ phần trăm oxy hòa
tan, nhiệt độ và tỷ lệ enzyme/cơ chất là khác nhau. Do đó, ở qui mô công nghiệp,
kiểm soát các yếu tố phản ứng enzyme là yếu tố then chốt quyết định hiệu quả của
quá trình xử lý bằng enzyme.
Hiệu suất chiết của các hợp chất hòa tan trong nước phụ thuộc vào tất cả công
nghệ (enzyme, cơ học,...) để làm tăng khả năng hòa tan vào nước của chúng. Do đó,
sử dụng enzyme hỗ trợ phải hiểu biết đầy đủ về thành phần thành tế bào, hoạt tính
enzyme và ảnh hưởng của điều kiện tiền xử lý và điều kiện thực nghiệm để đảm bảo
việc xử lý bằng enzyme được hiệu quả. Tuy nhiên, nếu những hạn chế trên có thể
được khắc phục, xử lý bằng enzyme dựa trên một qui trình tối ưu sẽ không chỉ làm
tăng năng suất chiết, mà còn tăng chất lượng sản phẩm khi qui trình chiết xuất được
thực hiện ở nhiệt độ thấp, áp suất thường.
1.2. Các enzyme sử dụng trong công nghệ enzyme hỗ trợ chưng cất (EAD)
và enzyme hỗ trợ chiết xuất (EAE)
1.2.1. Các enzyme phân giải cấu trúc thành tế bào nguyên liệu lignocellulose

11
Một số thành phần lignocellulose của một vài nguyên liệu thực vật được
Kumar, 2009 khảo sát [110] và chỉ ra trong Bảng 1.1:
Bảng 1.1. Thành phần lignocellulose trong một số loại sinh khối
Nguyên liệu
lignocellulose
Hàm lượng (%)
Cellulose Hemicellulose Lignin
Gỗ cứng 40-55 24-40 18-25
Gỗ mềm 45-50 25-35 25-35
Cỏ 25-40 35-50 10-30
Rơm lúa mì 30 50 15
Lá 15-20 80-85 0
Bảng 1.1 cho thấy, thành phần lignocellulose ở các cây thân gỗ gồm khoảng 40-
45% (trọng lượng khô) cellulose, 24-40% hemicellulose (ít pectin hơn vì các khoảng
trống giữa các sợi cellulose nhỏ) và lignin (chiếm tới 18-25% trọng lượng khô). Vì vậy,
cấu trúc tế bào ở gỗ cứng bền vững hơn rất nhiều so với lá không có hoặc rất ít lignin.
Do sự phức tạp của cấu trúc thực vật nên sự phân giải của lignocellulose là một quá trình
đa enzyme liên quan đến sự biến đổi thủy phân và oxy hóa.
1.2.1.1. Quá trình oxy hóa loại lignin
Lignin là một hợp chất cao phân tử đặc biệt của thực vật, thường tập trung ở
những mô hóa gỗ, là chất kết dính tế bào, làm tăng độ bền cơ học, chống thấm nước
qua vách tế bào mô xylem, ngăn cản sự xâm nhập của vi sinh vật. Lignin là sản phẩm
ngưng tụ của 3 thành phần chủ yếu gồm trans--coumaryl, trans-coniferyl, và trans-
sinapyl alcohol theo tỷ lệ khác nhau [39].
Hình 1.4a. Các đơn vị cơ bản của lignin [39]
Các polymer này có các liên kết ngang (cross-linked) với nhau tạo nên mạng

12
lưới polymer - lignin.
Hình 1.4b. Cấu trúc của lignin [39]
Lignin tạo liên kết hóa học với hemicellulose và ngay cả với cellulose (nhưng
không nhiều). Độ bền hóa học của những liên kết này phụ thuộc vào bản chất liên
kết, cấu trúc hóa học của lignin và các gốc đường tham gia liên kết. Carbon alpha
(Cα) trong cấu trúc phenyl propane là nơi có khả năng tạo liên kết cao nhất với khối
hemicellulose. Ngược lại, các đường nằm ở mạch nhánh như arabinose, galactose, và
4-O-methylglucuronic acid là các nhóm thường liên kết với lignin. Các liên kết có
thể là ether, ester (liên kết với xylan qua 4-O-methyl-D-glucuronic acid), hay
glycoside (phản ứng giữa nhóm khử của hemicellulose và nhóm OH phenolic của
lignin) [63].
Các ligninase là các enzyme phân hủy lignin. Ba enzyme quan trọng được
công nhận rộng rãi có khả năng oxy hóa lignin là LiP (lignin peroxidase), MnP
(mangan peroxidase) và Laccase (polyphenol oxidase) [55]. Nấm đảm
Basidiomycetes (nấm mục trắng -white rot fungy) và nấm mục nâu là nhóm sản sinh
hiệu quả nhất của các ligninase.

13
Phân hủy lignin được khởi động thông qua việc sản xuất oxy tự do gốc tự do
nấm mục trắng (white rot fungy) và nấm mục nâu (brown rot fungy). Các gốc oxy
được sản xuất thông qua các phản ứng Fenton và sau đó tấn công các phần lignin của
vách tế bào thực vật và tạo điều kiện cho các protein enzyme khác xâm nhập cấu trúc
lignocellulose [63].
Trong số 3 enzyme, laccase (EC 1.10.3.2, p- diphenol oxidase) là enzyme oxy
hóa khử có khả năng oxy hóa diphenol và các hợp chất có liên quan, sử dụng oxy
phân tử làm chất nhận điện tử. Không giống như lignin peroxidase và peroxidase
mangan, laccase có lợi thế hơn peroxidase trong việc sử dụng oxy như một cofactor,
có khả năng xúc tác cho quá trình oxy hóa của các hợp chất hữu cơ trong sự vắng mặt
của H2O2 hoặc Mn2+, cho phép laccase được áp dụng hiệu quả trong công nghiệp, bao
gồm tẩy trắng trong công nghiệp giấy, loại màu của thuốc nhuộm, loại bỏ các loại
thuốc diệt cỏ từ ngũ cốc cây trồng và phân hủy sinh học và bioconversion thực phẩm
và chất thải nông nghiệp [95]. Chính quá trình oxy hóa mở vòng thơm của các enzyme
phân giải lignin sẽ tạo điều kiện thuận lợi cho các enzyme cellulase và hemicellulase
tấn công, phân giải các bó sợi cellulose, hemicellulose vốn bị lignin che chắn [39].
1.2.1.2. Quá trình phân giải cellulose
Cellulose là polymer được cấu tạo từ các mắt xích β-D-Glucose liên kết với
nhau bởi liên kết β-1,4 glucoside, có công thức cấu tạo là (C6H10O5)n hay
[C6H7O2(OH)3]n trong đó n có thể nằm trong khoảng 5000-14000, là thành phần chủ
yếu cấu tạo nên vách tế bào thực vật [79].
Hình 1.5a. Công thức hóa học của cellulose [79]
Cellulase là enzyme phân hủy các phần cellulose. Cơ chế tác dụng của
cellulase không hoàn toàn rõ ràng, Zhang & cs. xem xét cơ chế hóa học được chấp

14
nhận rộng rãi nhất là sự hiệp đồng của 3 loại enzyme chính.
Hình 1.5b. Cơ chế phân giải cellulose của hệ enzyme cellulase [80]
+ Enzyme nội bào Endoglycanase hay 1,4-β-D-glucan glucanohydrolase (EC
3.2.1.4): Enzyme nội bào endoglycanase hoặc 1,4-β-D-glucan glucanohydrolase là
enzyme thủy phân nội bào liên kết 1,4-β-D-glucosidic trong phân tử cellulose bởi tác
dụng ngẫu nhiên trong chuỗi polymer hình thành các đầu chuỗi khử tự do và các
chuỗi oligosaccharide ngắn. Các endoglucanase không thể thủy phân cellulose tinh
thể hiệu quả nhưng nó sẽ phá vỡ các liên kết tại khu vực vô định hình tương đối dễ
tiếp cận.
+ Exoglucanase: exoglucanase gồm cả 1,4-beta-D-glucan glucanohydrolase
(EC 3.2.1.74), giải phóng D-glucose từ β-glucan và cellodextrin và 1,4-beta-D-glucan
cellobiohydrolase (EC 3.2.1.91) mà giải phóng D-cellobiose. Tỷ lệ thủy phân của
enzyme cellobiohydrolase ngoại bào bị hạn chế bởi sự sẵn có các đầu chuỗi cellulose.
+ β-glucosidase hay β-D-glucoside glucohydrolase (EC 3.2.1.21): giải phóng
phân tử D-glucose từ đường cellodextrin hòa tan và một loạt các glucoside khác.
1.2.1.3. Quá trình phân giải hemicellulose
Khác với cellulose, hemicellulose là một loại polymer phức tạp và phân nhánh,
độ trùng hợp khoảng 70 đến 200 đơn phân. Hemicellulose chứa cả đường 6 carbon
(gồm glucose, mannose và galactose) và đường 5 carbon (gồm xylose và arabinose)
và một vài acid [110].

15
Ở thực vật bậc cao, thành phần cơ bản của hemicellulose là xylan liên kết với
nhau bằng liên kết β-1,4-D-xylopyranose. Đa số phân tử xylan chứa nhiều nhóm ở
trục chính và chuỗi bên. Các gốc thay thế chủ yếu trên khung chính của xylan là các
gốc acetyl, arabinosyl và glucuronosyl. Các nhóm này có đặc tính liên kết tương tác
cộng hóa trị và không hóa trị với lignin, cellulose và các polymer khác.
Hình 1.6a. Cấu trúc polymer xylan [110]
Hemicellulase là enzyme phân hủy hemicellulose. Do tính chất biến động
nhiều hơn, có nhiều enzyme hoạt động trên hemicellulose. Xylanase và mannanase
là những enzyme phân giải liên kết β-1,4 trong xylan và mannan và giải phóng
oligosaccharide mà có thể thủy phân tiếp thành xylose và mannose bởi β-xylosidase
và β-mannosidase [39].
Hình 1.6b. Enzyme xylanolytic liên quan đến quá trình phân giải xylan
Ac: nhóm acetyl; α-Araf: α-arabinofuranose; α-4-O-Me-GlcA: α-4-O-
methylglucuronic acid
Enzyme acetylxylan esterase là enzyme có thể cắt đứt giữa các nhóm acetyl

16
với 2 hoặc 3 vị trí của gốc xylose và góp phần đóng vai trò thủy phân xylan trong
tự nhiên.
Vi sinh vật phân giải hemicellulose: hemicellulose khác cellulose vì chứa cả
đường pentose và hexose và cũng thường chứa axit uronic. Những vi khuẩn có khả
năng phân giải cellulose thì cũng có khả năng sử dụng hemicellulose. Tuy nhiên,
không phải tất cả các loài sử dụng được hemicellulose đều có khả năng thủy phân
cellulose. Một số loài VSV phân giải hemicellulose là Butyrivibrio fibrisovens,
Lachnospira multiparus và Bacteroides ruminicola…
Enzyme phân giải cellulose, hemicellulose là nhóm enzyme ứng dụng rất nhiều
trong công nghiệp (chỉ sau protease) với các chế phẩm enzyme dạng bột, dịch [14].
Novozyme là công ty sản xuất enzyme lớn với một số sản phẩm thủy phân cellulose và
hemicellulose như Cellic CTec2, Cellic Ctec3, Cellic Htec2 (cellulase và xylanase),
Cellulase 1.5 LFG, Novozyme 188 (β-glucosidase), Novozyme NS50013 (Cellulase),
Novozyme NS5001 (β-glucosidase), Novozyme NS50012 (hỗn hợp glucosidase,
xylanase). Cellic Htec2 là hỗn hợp đa enzyme bao gồm cellulase và xylanase đã được
ứng dụng rất nhiều trong chuyển hóa cellulose của rơm rạ, rong biển thành bioethanol
cho hiệu quả thủy phân cao, phổ cơ chất rộng [29].
1.2.2. Enzyme protease phân giải cấu trúc nguyên liệu giàu protein
Protease hay peptidase (EC.3.4.) là nhóm enzyme thủy phân có khả năng cắt
mối liên kết peptide (-CO~NH-) trong các phân tử polypeptide, protein và một số cơ
chất tương tự thành các axit amin tự do hoặc các peptide phân tử thấp.

17
H2N - CH - CO - NH - CH - CO - ... - NH - CH - COOH
H2N - CH - COOH + H2N - CH - CO.... NH - CH - COOH
Hình 1.7. Phản ứng thủy phân protein của enzyme protease
Đa số enzyme thủy phân (hydrolase) không có nhóm ngoại. Trong trung tâm
hoạt động của chúng có chứa gốc axit amin đặc hiệu. Đối với hydrolase thường chứa
hai nhóm chức. Ví dụ: + Vòng imidazol của histidin + Nhóm hydroxyl (một số axit
amin: serine, threonine) Sự tương tác giữa hai nhóm đặc hiệu (-OH, - imidazol) đã hình
thành tâm ái nhân. Xung quanh trung tâm hoạt động của hydrolase còn chứa nhiều
các axit amin, vai trò của serine có chứa nhóm - OH có tác động rất lớn làm thay đổi
trung tâm hoạt động theo hướng có lợi cho hoạt động xúc tác của enzyme. Do cấu
trúc bậc 3 của phân tử protein-enzyme mà nhóm hydroxyl của serine và vòng
imidazol của histidin gần nhau, tạo ra liên kết hydroxyl giữa gốc - OH của serine với
nitơ bậc 3 của histidin. Nhờ quá trình đó mà nhóm hydroxyl xuất hiện tính chất ái
nhân và có thể tương tác được với liên kết nhị dương của cơ chất.
Các enzyme protease sử dụng phân tách các liên kết peptid đã được ứng dụng
trong công nghiệp thực phẩm từ rất lâu đời. Chủ yếu là protein thực vật và sữa. Từ
những năm 1960, protease được ứng dụng để thủy phân các protein từ cá nhằm hướng
tới nguồn dinh dưỡng giá rẻ. Các protein thủy phân từ cá (FPH- fish protein
hydrolysate) đã được tách chiết bằng phương pháp này đã rất thành công trong sản
xuất thức ăn gia súc, sản xuất protein cô đặc bổ sung vào nước mắm ... [109]. Quá
trình thủy phân protein gắn liền sự giải phóng lipid trong dịch thủy phân nên phương
pháp ly tâm thường được sử dụng để tách lipid. Phế liệu đầu từ cá ngừ đại dương
ngoài protein có giá trị (axit amin thiết yếu 30-35%) còn chứa hàm lượng lipid cao
với các axit béo không no thiết yếu (EPA, DHA 18-22%) [12], [138].
R1 R2 Rx
R1
H2O
R2 Rx

18
Phân loại: Protease được phân làm 2 loại, endopeptidase và exopeptidase.
- Exopeptidase được phân thành 2 loại dựa vào vị trí tác động trên mạch polipeptid.
+ Aminopeptidase: xúc tác thủy phân liên kết peptide ở đầu N tự do của chuỗi
polypeptid để giải phóng ra một axit amin, một dipeptide hoặc tripeptid.
+ Carboxypeptidase: xúc tác thủy phân liên kết peptid ở đầu C của chuỗi
polypeptide và giải phóng ra một axit amin hoặc một dipeptide.
- Endopeptidase gồm serinproteinase và cystein protease
+ Serin proteinase: là những protein chứa nhóm -OH của gốc serine trong trong
tâm hoạt động và có vai trò đặc biệt quan trọng đối với hoạt động xúc tác của enzyme.
Nhóm này bao gồm hai nhóm nhỏ: chymotrypsin và subtilisin. Nhóm chymotrypsin
bao gồm các enzyme động vật như chymotrypsin, trypsin và elastase. Nhóm subtilisin
bao gồm hai loại enzyme vi khuẩn như subtilisin Carsberg, subtilisin BPN. Các serine
protease thường hoạt động mạnh ở vùng kiềm tính và thể hiện tính đặc hiệu cơ chất
tương đối rộng.
+ Cysteine protease: các protease chứa nhóm -SH trong trung tâm hoạt động.
Cystein proteinase bao gồm các protein thực vật như papain, bromelain, một vài
protein động vật và kí sinh trùng. Các cystein proteinase thường hoạt động ở vùng
pH trung tính và có tính đặc hiệu cơ chất rộng.
+ Aspatic proteinase: hầu hết các aspartic proteinase thuộc nhóm pepsin có
chứa nhóm carboxyl trong trung tâm hoạt động và thường hoạt động mạnh ở pH trung
tính bao gồm các enzyme tiêu hóa như pepsin, chymosin, cathepsin, renin.
+ Metalloproteinase: là nhóm proteinase tìm thấy ở vi khuẩn, nấm mốc cũng
như các VSV bậc cao, thường hoạt động ở vùng pH trung tính và hoạt độ giảm mạnh
dưới tác dụng của EDTA.
Có thể thu nhận protease từ rất nhiều nguồn khác nhau như động vật, thực vật
hay vi sinh vật. Tuy nhiên, enzyme thực vật và enzyme từ vi sinh vật là hai nhóm
enzyme được ứng dụng nhiều trong công nghiệp. Một số chế phẩm enzyme công
nghiệp thường được sử dụng trong chế biến các sản phẩm giàu protein [32].
- Các chế phẩm enzyme có nguồn gốc thực vật
+ Papain và chymopapain: là hai chế phẩm enzyme chứa chủ yếu là protease

19
và đều nhận được từ mủ nhựa đu đủ Carica Papaya (L) (Caricaceae). Hàm lượng
chymopapain trong nhựa đu đủ lớn hơn papain. Chế phẩm papain có hoạt tính cao
trong phân giải amit và ester, có khả năng chịu nhiệt cao hơn so với các enzyme khác,
nhiệt độ hoạt động tới hạn là 80oC. Trong khi đó, chymopapain xúc tác thủy phân các
mối liên kết peptid và ester.
+ Physin: thu được từ nhựa quả sung. Trung bình quả xanh trọng lượng từ 10-
15g có thể thu được 100-200 mg physin. Physin tương đối ổn định ở pH 6-8.
+ Bromelain: Enzyme này có nhiều trong lá và vỏ dứa, cũng thủy phân casein
tốt như papain, nhưng đối với hemoglobin vận tốc tăng lên 4 lần. Bromelain là nhóm
endoprotease có khả năng phân cắt các liên kết peptid nội phân tử protein để chuyển
phân tử protein thành các đoạn nhỏ gọi là peptid. Thành phần chủ yếu của Bromelain
có chứa nhóm sulfhydryl thủy giải protein. Trọng lượng phân tử 33.200- 33.500 Da,
bromelain trích ly từ thân dứa có điểm đẳng điện pI 9.55 trong khi trích từ quả lại là
protein acid có điểm đẳng điện pI 4.6.
- Các chế phẩm enzyme có nguồn gốc từ vi sinh vật
+ Alcalase 2.4l, Neutrase 0.5l, Esperase 7.5l, Protamex, Novozyme FM2.0L:
chúng là các chế phẩm enzyme chứa chủ yếu là endopeptidase thu nhận từ vi khuẩn.
Các chế phẩm này được sử dụng để cải thiện các tính chất công nghệ, dinh dưỡng và
hương vị của protein.
+ Flavourzym MG, Kojizyme đây là hỗn hợp của enzyme exo- và endo peptidase
thu nhận từ nấm mốc. Các chế phẩm này dùng để tăng cường thủy phân protein.
Quá trình chiết xuất lipid bằng enzyme thủy phân đạt hiệu quả liên quan đến
việc lựa chọn được loại enzyme protease phù hợp nhằm đảm bảo chất lượng của bột
đạm thủy phân và lipid
1.2.3. Enzyme lipase thủy phân lipid thành axit béo tự do
Các axit béo không no đa nối đôi chuỗi dài n-3 PUFA, đặc biệt là EPA, DHA
có vai trò rất lớn đối với đời sống sức khỏe con người và vật nuôi. Tuy nhiên, dầu cá
bị hạn chế vì chúng chứa một lượng đáng kể các axit béo no và cholesterol [92]. Có
rất nhiều phương pháp làm giàu n-3 PUFA nhưng lipase là phương án được chú ý

20
gần đây bởi chúng phản ứng ở điều kiện nhiệt độ thường, hạn chế sự biến tính các
axit béo mạch dài, đồng thời giảm thiểu lượng dung môi hóa chất sử dụng, thân thiện
với môi trường sinh thái.
Phản ứng thủy phân lipid là một trong những bước kỹ thuật quan trọng để thu
được các axit béo tự do, là yếu tố then chốt dẫn đến hiệu quả quyết định khi kết tinh
ure. Để làm giàu các axit béo không no đa nối đôi PUFA, nâng cao chất lượng sản
phẩm dầu từ đầu cá ngừ, dầu cá thường được thủy phân thành các axit tự do bằng
cách đun nóng với tác nhân axit hoặc kiềm, sau đó kết tinh với ure trong cồn. Phương
pháp này hiệu quả song tốn một lượng hóa chất, gây ô nhiễm môi trường.
Lipase hydrolase (EC 3.1.1.3) là enzyme xúc tác thủy phân triglycerid thành
di, mono glycerid hoặc glyxerol và các axit béo nhờ hoạt động trên bề mặt phân pha
dầu, nước. Lipase là enzyme linh hoạt có thể sử dụng xúc tác nhiều loại phản ứng
khác nhau, có khả năng chịu kiềm, chịu nhiệt. Chính vì thế, lipase được ứng dụng
rộng rãi trong công nghiệp như công nghiệp thực phẩm, công nghiệp hóa học, công
nghiệp mỹ phẩm, công nghiệp da, trong y dược và các ngành công nghiệp khác.
Lipase xúc tác phản ứng thủy phân cắt đứt lần lượt các liên kết α- ester chứ không cắt
cùng lúc 3 liên kết. Quá trình xúc tác thường chậm hơn so với các enzyme khác như
protease, carbohydradase.
Hình 1.8. Phân tử lipid và các vị trí thủy phân của enyme lipase
Enzyme được kích hoạt tại giao diện dầu-nước do mở nắp cấu tạo của lipase
tại giao diện của hệ thống [20, 21]. Cơ chế liên quan đến sự gắn kết của nhóm acyl
(RCOO -) trên bề mặt điện tích dương (NH3 +) và sự gắn kết của ion hydro (H+) trên
bề mặt mang điện tích âm (COO-). Nhờ vậy giải phóng các axit béo tự do. Lipase
hydrolase khác với lipase esterase ở điểm chỉ thuỷ phân cơ chất không tan trong nước

21
và hoạt lực được tăng cường khi ở bề mặt phân chia pha cơ chất - nước (interfacial
activation). Vì vậy, hoạt lực tối ưu của lipase chỉ được thể hiện trong hệ nhũ tương,
khi đó thì diện tích tiếp xúc giữa cơ chất và enzyme sẽ tăng lên rất nhiều. Ngoài ra,
không những xúc tác cho hệ nhũ tương bình thường (dầu trong nước), lipase còn hoạt
động mạnh ở những hệ nhũ tương là nước trong dầu và dầu hoà tan trong dung môi
hữu cơ.
Lipase thu từ các nguồn khác nhau và có đặc trưng khác nhau.
- Lipase thu nhận từ động vật: Lipase thu nhận từ những cơ quan, mô của một
số loài động vật có vú đã được nghiên cứu rất nhiều. Bản chất của chúng là những
glycoprotein, phân tử lượng 50 kDa, không có hoặc ít hoạt tính đối với phospholipid.
Lipase của động vật có vú bền ở pH thấp, được hoạt hóa bởi muối mật và đặc hiệu tại
vị trí sn-3 của cơ chất. Lipoprotein lipase người có chức năng thủy phân
triacylglycerol trong chylomicron. Để có hoạt tính thì enzyme này kết hợp với
apolipoprotein C-II để tạo thành dimer và được hoạt hóa bởi heparin. Tuy nhiên,
lipase trong gan thực hiện chức năng chuyển hóa lipoprotein nhưng không liên kết
với apo C-II. Lipase có trong sữa mẹ được hoạt hóa bởi muối mật để giúp trẻ sơ sinh
tiêu hóa chất béo có trong sữa.
- Lipase từ thực vật: Lipase từ thực vật không được chú ý nhiều so với những
nguồn thu nhận khác. Tuy nhiên, gần đây, loại enzyme này đã bắt đầu được quan tâm
và nghiên cứu khá phổ biến. Trong đó, lipase từ những hạt có dầu là được quan tâm
nhất. Những enzyme này nếu khác nhau từ nguồn nguyên liệu thu nhận thì đặc hiệu
về cơ chất, pH tối ưu, khả năng phản ứng với sulfuhydryl, tính kỵ nước cũng khác
nhau. Những enzyme này có quan hệ mật thiết với triacylglycerol có trong bản thân
hạt dầu đó và chỉ được tổng hợp trong quá trình nảy mầm của hạt.
- Lipase từ vi sinh vật: Đây là nguồn enzyme được quan tâm và sản xuất nhiều
nhất theo quy mô công nghiệp. Khác với thực vật và động vật, vi sinh vật được cấu
tạo từ một tế bào, chính vì vậy mà nó có những ưu điểm hơn hẳn động vật và thực
vật. Vi sinh vật có khả năng sinh tổng hợp enzyme rất lớn trong một khoảng thời gian
ngắn, hoạt tính của enzyme cao hơn hoạt tính của enzyme được tổng hợp từ động vật

22
và thực vật; và ta hoàn toàn có thể điều khiển tốc độ sinh tổng hợp enzyme trong khi
sản xuất.
Lipase được thu nhận từ vi sinh vật bao gồm vi khuẩn, nấm men và nấm mốc
là những enzyme ngoại bào có tính chất gần giống lipase tuyến tụy. Các loài nấm
mốc có khả năng sinh tổng hợp lipase như: Aspergillus spp., Mucor spp., Rhizopus
spp., Penicillium spp., Geotrichum spp. Đối với nấm men gồm những loài Torulopis
spp., Candida spp. Và vi khuẩn có khả năng sinh tổng hợp lipase bao gồm:
Pseudomonas spp., Achromobacter spp., Staphylococcus spp. Loài vi sinh vật được
sử dụng sản xuất lipase theo quy mô công nghiệp chủ yếu là nấm sợi.
Dựa vào tính đặc hiệu của enzyme lipase, mục đích của quá trình thủy phân
theo nhiều phương thức khác nhau. Lipase được sử dụng để gia tăng hàm lượng các
axit béo n-3 PUFA (EPA, DHA) trong dầu bằng việc thủy phân các thành phần axit
béo no, lipase từ Chromobacterium viscosum, Pseudomonas sp. lại giải phóng tất cả
các loại axit béo [92].
1.3. Tình hình nghiên cứu ứng dụng công nghệ enzyme trong chiết xuất
và làm giàu các hợp chất thiên nhiên trong và ngoài nước
1.3.1. Các nghiên cứu trên thế giới
Có rất nhiều các nghiên cứu đối với các loại cây có dầu [162]. Gần đây, một
số ứng dụng enzyme được sử dụng hỗ trợ tách tinh dầu và các thành phần dễ bay hơi
như:
- Freese & Binnings, (1993) đã sử dụng của enzyme trong quá trình chiết xuất
dầu và tinh dầu từ gừng, tỏi, hạt tiêu đã được báo cáo là tăng năng suất của dầu lên
30-50% so với đối chứng không xử lý enzyme [90].
- Sowbhagya et al. (2009) đã xử lý enzyme cellulase, pectinase, protease và
Viscozyme trước khi chưng cất tinh dầu từ tỏi làm tăng hàm lượng từ (0,39-0,51%)
so với đối chứng (0,28%). Tác giả cũng chứng minh rằng enzyme tạo thuận lợi cho
việc khai thác dầu tỏi, dẫn đến sự gia tăng hiệu suất và ít thay đổi hương vị hay tính
chất hóa lý của dầu [161]. Ngoài ra, Sowbhagya (2011) cũng chứng minh hiệu quả

23
của các enzyme hỗ trợ tách tinh dầu hạt cần tây (Apium graveolens L.), hạt thì là
(Cuminum cyminum L.). Enzyme cellulase, hemicellulase, pectinase, protease và hỗn
hợp cellulase + hemicellulase đều làm tăng sản lượng tinh dầu hạt cần tây (22-27%),
tinh dầu hạt thì là 18-22% so với đối chứng không sử dụng enzyme. Tác giả cũng cho
rằng, enzyme không làm thay đổi bất kỳ đặc tính cảm quan, tính chất vật lý hoặc tính
chất hóa học của tinh dầu. Tuy nhiên, có sự gia tăng đáng kể về thành phần limomene
trong tinh dầu cần tây (82.2%) khi xử lý với cellulase so với đối chứng (63.9%) [163],
[164].
- Theo Boulila A. et al. (2015) tiền xử lý lá Nguyệt Quế (Laurus nobilis L.)
bằng enzyme cellulase, hemicellulase, xylanase và hỗn hợp các enzyme đã tăng hiệu
suất thu nhận tinh dầu lần lượt là 243, 227, 240.54 và 0.48% tương ứng với các
phương án xử lý. Các kết quả phân tích GC-MS cũng cho thấy quá trình xử lý bằng
enzyme không những không làm biến đổi thành phần các hợp chất dễ bay hơi mà
còn làm giàu các thành phần monoterpenoid trong tinh dầu, từ đó làm gia tăng hoạt
tính chống oxy hóa của sản phẩm thông qua thử nghiệm với DPPH và ABTS [58].
- Sayantani Dutta and Paramita Bhattacharjee (2015) đã kết hợp α-amylase với
CO2 siêu tới hạn đã tăng hiệu suất nhựa dầu hạt tiêu lên 2.13 lần khi không xử lý
enzyme trong khi xử lý bằng enzyme (1,25 lần). Quá trình kết hợp enzyme và CO2
đã tăng hiệu suất chiết 53% cũng làm giàu hàm lượng piperin có trong hỗn hợp so
với đối chứng không sử dụng enzyme [154].
Như vậy, vai trò của enzyme ngoài hỗ trợ quá trình chiết tách, làm giảm thời
gian và năng lượng còn gia tăng hàm lượng chất chính trong sản phẩm.
Trên đối tượng sinh vật biển, quá trình chiết xuất lipid và làm giàu các axit
béo không no đa nối đôi PUFA nhờ enzyme hỗ trợ cũng được nghiên cứu.
- Năm 2002, Liaset và cộng sự đã sử dụng enzyme Protamex để thủy phân phế
phụ phẩm của cá hồi sau khi fillet, lượng lipid thu được sau quá trình ly tâm xấp xỉ
77% lượng lipid tổng số có trong nguyên liệu ban đầu và phần lipid này có chứa hàm
lượng EPA và DHA cao [117]. Tác giả cũng cho thấy phương pháp thủy phân bằng
enzyme thu được dầu cá giàu hàm lượng ω-3 với hiệu suất thu hồi cao (khoảng 77%)

24
cũng như thu được nhiều thành phần khác có giá trị như: các peptid và các axit amin
không thay thế [118].
- Linder et al., (2005) đã sử dụng các enzyme protease thương mại khác nhau
là Alcalase, Neutrase và Flavourzyme để tách chiết dầu thô từ đầu cá hồi và enzyme
lipase Aspergillus oryzae (Novozym SP 398) để thủy phân dầu thành các axit béo và
các acylglycerol. Hiệu suất thu hồi dầu cao nhất (17,4% sau 2h) đạt được khi sử dụng
Alcalase, gần với hiệu suất thu hồi dầu đạt được bằng phương pháp Blight and Dyer
(20%), quá trình thủy phân bằng enzyme lipase đạt được 45% trong 24 giờ đã làm
giảm các axit béo no SFA 27,2% xuống 20,2% và làm giàu các axit béo không no đa
nối đôi PUFA từ 41,6% lên 46,5% trong đó DHA từ 9,9% đến 11,6% và EPA từ 3,6%
đến 5,6% [120].
- Dumay và cộng sự (2006) đã nghiên cứu hoàn thiện quy trình thu hồi lipid
và phospholipid từ nội tạng cá mòi sử dụng các enzyme thương mại là Flavourzyme,
Protamex, Alcalase. Quá trình thủy phân tiến hành trong 24h kết hợp khuấy đảo liên
tục. Kết quả nghiên cứu cho thấy hàm lượng lipid tổng số trong các phần chất lỏng
sau khi ly tâm (phần dầu và phần dung dịch) tăng lên (ít nhất 85% các lipid đã được
xác định có trong nguyên liệu có mặt trong các pha này). Quá trình giúp cải thiện
sự thu hồi lipid đối với các ứng dụng mang tính thương mại. Thêm vào đó, các phần
lipid này giàu các phosphoslipid hơn lipid được chiết bằng phương pháp hóa học cổ
điển, đặc biệt là sau khi thủy phân bằng Alcalase [83].
- Laplante và cộng sự (2009) đã tiến hành nghiên cứu so sánh hiệu quả tách
chiết thu hồi dầu từ cá thu và cá trích theo 2 phương pháp tách chiết nhiệt độ thấp là
phương pháp tách chiết bằng CO2 siêu tới hạn và phương pháp thủy phân bằng
enzyme Protamex. Trong đó, quá trình thủy phân được tiến hành ở các điều kiện: pH
7, nhiệt độ 45oC, thời gian thủy phân 2,5 giờ, tỷ lệ enzyme / nguyên liệu là 0,01%
(w/w), tỷ lệ nước / nguyên liệu là 1/1 (w/w). Kết quả cho thấy hiệu suất thu hồi dầu
từ hai loại cá này đạt được khi sử dụng phương pháp thủy phân cao hơn so với phương
pháp chiết CO2 siêu tới hạn, hay nói cách khác, quá trình tách chiết bằng phương
pháp thủy phân thể hiện hiệu quả có thể so sánh hoặc tốt hơn tách chiết dầu bằng CO2

25
siêu tới hạn. Bên cạnh đó, chi phí sản xuất đối với phương pháp thủy phân lại thấp
hơn, do đó phương pháp này cũng có thể là một lựa chọn tốt hơn khi sản xuất dầu ở
quy mô công nghiệp [108].
- Mbatia (2011) đã sử dụng các loại protease (Protex 30L và Bromelain) để
giải phóng dầu từ phế liệu cá rô sông Nile (Lates niloticus) và cá hồi (Salmon salar).
Hiệu suất thu hồi dầu tương ứng đối với đầu cá rô sông Nile nước ấm và đầu cá hồi
nước lạnh lần lượt là 11,2% và 15,7% khối lượng nguyên liệu. Trong khi đó, việc
tách chiết dầu bằng dung môi hữu cơ từ hai loại nguyên liệu này tương ứng lần lượt
là 13,8% và 17,6%. Kết quả nghiên cứu cho thấy việc bổ sung nước trong quá trình
thủy phân bằng enzyme làm giảm hiệu suất thu hồi dầu và thời gian thủy phân tốt
nhất cho việc thu hồi dầu là 2 giờ [128].
Nhìn chung, các nghiên cứu thu hồi dầu cá từ các phế liệu của quá trình sản
xuất trên thế giới hiện nay khá đa dạng và ngày càng mở rộng không những về đối
tượng nguyên liệu, phương pháp tách chiết thu hồi dầu mà đi sâu vào khai thác, phát
triển các khía cạnh mới và các lĩnh vực khác nhau của công nghệ sản xuất và tinh chế
dầu cá.
Nối tiếp các công trình nghiên cứu thu hồi dầu thô từ các phế liệu của các loại
cá khác nhau sau quá trình chế biến, các quá trình làm giàu các axit béo không no đa
nối đôi PUFA bằng enzyme lipase cũng được nghiên cứu.
Gámez - Meza và cộng sự (2003) đã nghiên cứu làm tăng hàm lượng DHA và
EPA của dầu cá mòi bằng phương pháp thủy phân và tạo phức với ure. Nghiên cứu
cho thấy sự kết hợp của phương pháp thủy phân bằng enzyme hoặc hóa chất kết hợp
với quá trình tạo phức với ure là một phương pháp hứa hẹn để thu được các axit béo
chưa bão hòa omega-3 nồng độ cao từ dầu cá mòi [92].
Liu và cộng sự (2006) đã nghiên cứu quá trình dùng enzyme để tách chiết dầu
dựa trên việc thủy phân protein từ đầu cá ngừ, sau đó tiến hành bước tạo phức với
ure, thu được hỗn hợp DHA và EPA với độ tinh khiết 85,02% và hiệu suất 25,01%
[122].
Tóm lại, trên thế giới, các nghiên cứu ứng dụng công nghệ enzyme ứng dụng

26
tiền xử lý các nguyên liệu thiên nhiên như động, thực vật, tảo... để chiết xuất các chất
có hoạt tính sinh học. Các nghiên cứu cũng chỉ ra rằng sử dụng phương án enzyme
bước đầu đã cho hiệu quả cao và tiềm năng ứng dụng trong công nghiệp, trở thành
một trong những công cụ quan trọng của hóa học xanh, cho phép tạo ra các công nghệ
mới sử dụng các hóa chất thân thiện với môi trường, tiết kiệm về năng lượng và chi
phí. Ứng dụng công nghệ enzyme thường cho kết quả là giảm thời gian chiết, giảm
thiểu việc sử dụng các dung môi và tăng năng suất và chất lượng của sản phẩm.
1.3.2. Các nghiên cứu trong nước
Ở Việt Nam, các nghiên cứu ứng dụng enzyme trong chiết xuất các hợp chất
thiên nhiên cũng được quan tâm trong khoảng 20 năm trở lại đây.
Nguyễn Văn Chung (2004) đã sử dụng enzyme α-amylase trong quá trình
chưng cất tinh dầu hồ tiêu đã làm tăng hiệu suất tinh dầu 8% [4].
Lưu Thị Lệ Thủy (2008) đã nghiên cứu xây dựng quy trình công nghệ thu nhận
dầu từ hạt bí đỏ bằng phương pháp enzyme Alcalase và Viscozyme cho hiệu suất lần
lượt 75,7 và 70,3% so với chiết bằng hexan. Sản phẩm dầu tạo ra có chất lượng tương
đương dầu tinh khiết, có giá trị dinh dưỡng cao (hàm lượng axít béo omega-6 > 50%)
[34].
Nguyễn Thị Minh Nguyệt [23] sử dụng enzyme protease, hemicellulase và
carbohydradase trong chiết tách dầu béo và các thành phần của cám gạo. Hiệu suất
thu dầu cám chỉ đạt 77,5% so với phương pháp chiết bằng dung môi hexan nhưng
ngoài dầu béo, sản phẩm thu được là protein có trong cám gạo 75,4% và xơ hòa tan
10,4%. Năm 2007, nhóm nghiên cứu cũng đã tách chiết và làm giàu các axit béo
không no đa nối đôi omega-3 và omega-6 bằng enzyme để ứng dụng trong công
nghiệp thực phẩm và thực phẩm chức năng.
Ứng dụng enzyme trong chiết xuất các HCTN vẫn là hướng mới mẻ đối với các
nghiên cứu trong nước. Chủ yếu các nghiên cứu trên một số loại cây có dầu. Tuy nhiên,
hoạt chất sinh học chứa trong các nguyên liệu thực vật thân gỗ rất đa dạng và phong
phú, sử dụng enzyme để phá vỡ cấu trúc thành tế bào chứa lignocellulose thì chưa có
công trình nghiên cứu nào đề cập, đặc biệt là ứng dụng trên các loại cây chứa tinh dầu,
nguồn nguyên liệu rất giá trị ở các vùng nhiệt đới như Việt Nam.

27
Chiết xuất lipid từ đầu cá ngừ vây vàng cũng đã có một số nghiên cứu trong
nước ứng dụng, tuy nhiên vẫn còn ở dạng sơ khai, chưa đi sâu ứng dụng. Tối ưu hóa
quá trình kép sử dụng hệ enzyme protease và lipase tiền xử lý vào 2 giai đoạn chiết
xuất và làm giàu các axit béo không no đa nối đôi omega-3 từ đầu cá ngừ vây vàng
cho tiềm năng ứng dụng cao trong công nghiệp khi tạo được sản phẩm dầu cá giàu
omega3 đạt tiêu chuẩn thực phẩm. Vì vậy, việc nghiên cứu các vấn đề nêu trên là rất
cần thiết và mang ý nghĩa thực tiễn cao.
1.4. Tổng quan về quế Cinnamomum cassia
1.4.1. Giới thiệu về Quế C.cassia
Quế Cinnamomum cassia còn có tên gọi khác là Quế Đơn, Quế bì, Quế quan,
Quế thanh... Loài này được (Nees & T.Nees) J.Presl miêu tả khoa học đầu tiên năm
1825.
Đặc điểm thực vật: là loại cây lâu năm, thân gỗ, chỉ phát triển ở một số vùng
nhất định có khí hậu nhiệt đới gió mùa. Cây trưởng thành có kích thước trung bình
12 - 17 m, lá mọc so le, dài và cứng, cuống to, hoa mọc thành chùy ở kẽ những lá
phía trên, quả hình trứng, thuôn, phía dưới có đài tồn tại hoặc nguyên hoặc hơi chia
(Hình 1.9a và 1.9b).
Hình 1.9a. Cành lá và hoa C. cassia
Hình 1.9b. Cây Cinnamomum cassia
Nguồn gốc và phân bố: Loài quế Đơn (Cinnamomum cassia) là cây nguyên
sản ở Việt Nam, hiện gặp và phân bố tự nhiên tại một số khu rừng ẩm nhiệt đới ở Việt
Nam (Cúc Phương, núi Đinh...). Đây là loài Quế quen thuộc đã được gây trồng và sử

28
dụng từ lâu đời ở nước ta. Có thể gặp Quế đơn mọc dải rác hoặc gây trồng trên những
diện tích lớn tại nhiều địa phương: Yên Bái, Quảng Ninh, Hà Tây, Ninh Bình, Thanh
Hóa, Nghệ An, Quảng Trị, Quảng Nam, Quảng Ngãi, Khánh Hòa, Bà Rịa - Vũng Tàu
(núi Đinh)... (Lã Đình Mỡi, 2001) [21].
Công dụng: Từ thời thượng cổ, trước Công nguyên hàng thế kỷ, người Trung
Quốc cũng như cha ông ta đã biết dùng Quế để chữa bệnh như điều trị mãn tính viêm
phế quản, liệt dương, khó thở, thấp khớp...; Trong Tây y, Quế và tinh dầu quế được
sử dụng làm thuốc kích thích tăng khả năng tuần hoàn, tiêu hóa, hô hấp, bài tiết... Quế
cũng được coi là chất kích thích, tăng nhu động ruột và gây co bóp tử cung. Quế
cassia là một thuốc khử nấm, chống dị ứng, ung thư dạ dày, tiêu chảy, làm đổ mồ hôi,
kháng những chất gây đột biến như benzopyren và cyclphosphamid.
Bột quế, tinh dầu Quế được dùng phổ biến để làm phụ gia thực phẩm, gia vị
và hương liệu do chúng có hoạt tính chống đầy hơi, chất chống oxy hóa và chất bảo
quản [146].
Sản xuất và tiêu thụ:
Đến năm 1998, diện tích rừng quế ở nước ta đạt khoảng 61.820 ha (trong đó
có 19.743 ha có thể khai thác) với trữ lượng ước tính khoảng 29.000 - 30.000 tấn
vỏ. Hàng năm Việt Nam xuất khẩu khoảng 1.500 - 2.000 tấn vỏ và 5 - 7 tấn tinh dầu
quế. Theo thống kê của FAO (1998) tổng diện tích quế đến tuổi khai thác tại Quảng
Đông và Quảng Tây (Trung Quốc) vào khoảng 35.000 ha với sản lượng ước chừng
28.000 tấn. Trung Quốc là nước sản xuất, tiêu dùng và xuất khẩu Quế Đơn lớn nhất.
Nước ta là quê hương của loài Quế đơn (C. cassia). Tổng diện tích Quế đơn ở
nước ta vào khoảng 16.000 ha trong đó riêng huyện Văn Yên, Yên Bái, C. cassia
trồng ở 27/27 xã và thị trấn với tổng diện tích khoảng 15.000 ha, trong đó trên 50%
diện tích có thể sẵn sàng cho vỏ cây sau thu hoạch với sản lượng quế vỏ 5.160
tấn/năm, gỗ quế 25.800 m3 và các loại lá và cành (60 - 70 000 tấn/năm) [142].
Vỏ, tinh dầu vỏ, tinh dầu lá của Quế đơn đều là những sản phẩm có giá trị
thương mại cao. Sức tiêu thụ của thị trường thế giới vào khoảng 20 - 30.000 tấn
quế/năm. Các nước nhập khẩu nhiều quế là Hoa Kỳ, Pháp, Canada, Nhật Bản,
Singapor, Ấn Độ và Hồng Kông... Giá mua bán trên thị trường thay đổi trong khoảng

29
3.000 - 4.000 USD/tấn quế vỏ, 52.000-100.000 USD/tấn tinh dầu từ quế vỏ và 30-
35.000 USD/tấn tinh dầu từ lá quế [21].
1.4.2. Những nghiên cứu về thành phần hóa học C. cassia
Các hợp chất chính chủ yếu được chiết xuất và xác định trong vỏ quế gồm:
polyphenols và các phenol dễ bay hơi (2-hydroxycinnamaldehyde, dẫn xuất
cinnamaldehyde), flavan-3-ols [155].
Trong số các polyphenol của quế có chứa vanillic, caffeic, galic, protocatechuic,
p-coumaric và ferulic acid (Hình 1.10a) (Nabavi S.F, 2015) [136].
Vanilic acid
Galic acid
Protocatechuic acid
Caffeic acid
Ferulic acid
-coumaric acid
Hình 1.10a. Một số polyphenol trong quế Cinnamomum
Với các hợp chất dễ bay hơi, thành phần hóa học của tinh dầu phụ thuộc vào
bộ phận thực vật mà nó chiết xuất. Ở C. cassia, thành phần chính là trans-
Cinnamaldehyde (70-90%),phụ thuộc vào phương pháp chiết (chưng cất bằng hơi
nước có hàm lượng cao hơn chiết Soxhlet, ngoài ra còn có coumarin, cinnamyl acetat,
2-methoxycinamaldehyde, benzaldehyde, cinnamyl alcohol [21]. Một số thành phần
chính và phụ trong tinh dầu quế C. cassia (Hình 1.10b).
trans- Cinnamaldehyde
Coumarin
Cinnamyl acetat

30
Methoxycinamaldehyde
Benzaldehyde
Cinnamyl alcohol
Hình 1.10b. Một số thành phần chính và phụ trong tinh dầu C. cassia
Tinh dầu Quế thuộc tinh dầu phenylpropanoid (C6-C3) do cinnamaldehyde
(hay aldehyde cinnamic) là thành phần quan trọng nhất, quyết định chất lượng tinh
dầu Quế, chiếm từ 70-95% tinh dầu, tạo nên mùi hương đặc trưng của quế [13]. Ngoài
phenylpropanoid và terpen, trong tinh dầu chứa còn chứa các hợp chất dễ bay hơi
khác.
Tinh dầu từ vỏ có màu vàng nâu nhạt, sánh, vị cay, thơm ngọt, nóng, nặng hơn
nước. Tinh dầu từ lá thường có màu nâu đậm và giá trị tinh dầu thường thấp hơn so
với tinh dầu từ vỏ. Thành phần tinh dầu luôn có sự biến động về số lượng giữa các
loài trong cùng một chi, tập quán canh tác, vị trí địa lý, thời gian canh tác...
Tinh dầu cất từ vỏ và cành lá của quế đơn C. cassia có thành phần tương tự
nhau, chủ yếu là cinnamaldehyde (70-90%), ngoài ta còn có coumarin, cinnamyl
acetat, 2-methoxycinamaldehyde, benzaldehyde... với rất ít hoặc không có eugenol.
Trong khi đó, tinh dầu từ vỏ của C. zeylanicum chứa 60-80% cinnamaldehyde và sấp
xỉ 2% eugenol còn tinh dầu từ lá của loài này rất giầu eugenol (70-75%) [22].
Tinh dầu vỏ C. cassia ở Trung Quốc khá phức tạp, có gần 100 hợp chất và
hiện nay đã phát hiện được 93 hợp chất, trong đó nhiều nhất là E-cinnamaldehyde
(65.5%), các hợp chất khác có hàm lượng đáng kể lần lượt là coumarin (8.7%),
cinnamyl acetat (3.6%), methoxycinnamaldehyd (2.7%), benzaldehyde (0.9%)...
Tinh dầu vỏ Quế đơn (C. cassia) được sản xuất từ Australia thì thành phần hóa
học gồm khoảng 40 hợp chất, trong đó chủ yếu là cinnamaldehyde (87,0%), tiếp đến
là benzaldehyd (4,7%), 2-phenylethanol (2,5%), 3- phenylpropanal (2,0%), 1,8-
cineol (0,7%), 4-ethylguaiacol (0,5%), ethyl cinnamat (0,4%), cuminaldehyd (0,4%),
chavicol (0,3%) và coumarin (0,3%); các thành phần còn lại chỉ có hàm lượng không
đáng kể hoặc vết [140].

31
Ở nước ta, khi phân tích các mẫu tinh dầu quế đơn C. cassia khác nhau đã cho
thấy chúng dao động trong những giới hạn nhất. Hàm lượng E-cinnamaldehyde từ
80-95%, ngoài ra còn có các hợp chất khác như cinnamyl acetat, cinnamyl alcohol,
coumarin, benzyl benzoat (Lã Đình Mỡi, 2001) [21].
Tinh dầu lá quế đơn trồng tại Trung Quốc, Ping Li và cộng sự (2015) đã xác
định được 15 hợp chất, trong đó nhiều nhất là cinnamaldehyde (74,1%), 2-
methoxycinnamaldehyd (10,5%), cinnamyl acetat (6,6%), coumarin (1,2%),
benzaldehyd (1,1%), các hợp chất còn lại có hàm lượng không đáng kể [140].
Tinh dầu lá C. cassia sinh trưởng tại Australia, theo Ravindran (2004), ngoài
thành phần chính là cinnamaldehyde (77,2%) còn có tới trên 30 hợp chất khác, trong
đó đáng chú ý là coumarin (15,3%), benzaldehyd (1,2%), 4-ethylguaiacol (0,8%),
ethyl cinnamate (0,4%). Các thành phần còn lại thường không đáng [146].
Quế đơn đem từ Việt Nam qua trồng bên Trung Hoa có tên là C. cassia
macrophyllum: số lượng tinh dầu chiết xuất nói chung vượt hẳn quế bản xứ (%): 2,0
(so với 1,98) ở vỏ; 0.36 (so với 0.69) ở cành; 1,96 (so với 0.37) ở lá; cinnamaldehyde
sản xuất cũng lớn hơn (%): 61,20 (so với 52,92) ở vỏ; 77,34 (so với 64.75) ở lá. Người
Trung Hoa đánh giá quế ta tốt hơn quế họ để sản xuất thuốc chữa bệnh.
Tinh dầu lá quế đơn C. cassia trồng tại Yên Bái thu vào 2 vụ trong năm (tháng
5 và tháng 9), Lê Tùng Châu và cộng sự (1988) đã nhận thấy hàm lượng
cinnamaldehyde ở các mẫu thu ở tháng 5 thường rất cao (90%), còn các mẫu thu vào
tháng 9 thường thấp hơn (chỉ từ 50-80%) (Lã Đình Mỡi, 2001) [21].
Như vậy, hàm lượng E-cinnamaldehyde cũng như các thành phần khác trong
tinh dầu vỏ và tinh dầu lá quế đơn cũng luôn biến động dưới tác động của các yếu tố
di truyền, điều kiện môi trường và thời điểm thu hái.
1.4.3. Những nghiên cứu về hoạt tính sinh học C. cassia
Các nghiên cứu về hoạt tính sinh học của quế in vitro và in vivo cho thấy quế
có nhiều lợi ích sức khỏe và hoạt tính sinh học như kháng khuẩn, chất chống oxy hóa,
chống đái tháo đường, chống khối u, hạ huyết áp, cholesterol và bảo vệ dạ dày [137]
[76]. Theo Domadia P. et al,(2007), trans-cinnamaldehyde sử dụng trong dược phẩm

32
như thuốc chống đầy hơi, cảm lạnh, chống tập kết tiểu cầu, chống gây đột biến và
diệt khuẩn. Và những sản phẩm tự nhiên này hầu như không độc hại, không gây đột
biến gene và không có chất gây ung thư (Bickers D., 2005) [53]
Hoạt tính kháng khuẩn: Tinh dầu là nguồn kháng sinh tự nhiên tiềm năng và đã
được sử dụng rất nhiều trong y học dân tộc từ lâu đời [84], [125]. Nó ức chế hoàn toàn
sự phát triển của nhiều vi khuẩn gây bệnh như Staphylococcus, Micrococcus, Bacillus,
Enterobacter, Helicobacter pylori, Mycobacterium smegmatis [43], [130]. Trong số
các vi khuẩn đường ruột ở người, cinnamaldehyde có hoạt tính ức chế sự phát triển
của các vi khuẩn có hại như Cl. pergringens và B. fragilis hơn là các vi khuẩn có lợi
như Bifidobacteria và L. acidophilus [114]. Huang Y.F. & cs., (2014) cho rằng
thành phần của tinh dầu có thể kết hợp với bề mặt tế bào vi sinh vật và sau đó thâm
nhập vào lớp phospholipid của màng tế bào chất và enzyme liên kết màng, cho phép
chúng tích lũy trong màng tế bào, làm biến đổi cầu trúc và làm tăng tính thấm của
màng tế bào, phá hủy các thành phần nội bào, dẫn đến sự chết của tế bào [100].
Hoạt tính kháng u: Cao chiết từ vỏ quế đã cho thấy khả năng ức chế sự gia
tăng của một số dòng thế bào ung thư ở người, bao gồm ung thư vú, ung thư gan, ung
thư bạch cầu, ung thư buồng trứng và ung thư phổi trên các nghiên cứu in vitro và in
vivo [96]. Các tác giả đã cho thấy rằng, các hợp chất này có hoạt tính chống tăng sinh,
gây apoptosis thông qua sự kích hoạt p53 và ít có tác dụng gây độc tế bào trên các tế
bào bình thường bởi rất nhiều các báo khác nhau bao gồm tế bào nguyên sợi thường
(normal fibroblasts), tế bào lympho chuột chính (primary mouse lymphocytes), MCF-
10 (dòng tế bào biểu mô vú bình thường) và tế bào nguyên sợi chuột (normal rat
fibroblasts- F2408 cells) [169]. Tác giả Huang T.C. &cs., (2007) cho thấy thành phần
cinnamaldehyde trong tinh dầu lá quế Cinnamomum osmophloeum đã tác động gây
apotosis thông qua sự phá vỡ màng ty thể, kích thích sản sinh ROS, suy giảm
glutathione và kích hoạt caspase trên dòng tế bào ung thư bạch cầu người K562 [99].
Hoạt tính chống oxy hóa. Nhiều loại gia vị trong đó Quế được biết đến có hoạt
tính chống oxy hóa mạnh. Các hợp chất phenolic trong các nguyên liệu thực vật này
có mối quan hệ mật thiết với hoạt tính chống oxy hóa của chúng. Cao chiết từ vỏ C

33
zaylanicum có giá trị TEAC (107.69 mmol / 100 g) và tổng hàm lượng phenolic
(11.90 g of GAE/100g), trong đó vỏ C. cassia có giá trị TEAC (61.75 mmol /100 g)
và tổng hàm lượng phenolic (6.34galic), (Shan B., 2005) [155]. Hoạt tính chống oxy
hóa từ lá của 5 loài Cinnamomum (C. burmanni, C. cassia, C. pauciflorum, C. tamala
và C. zeylanicum) đã được nghiên cứu. Kết quả cho thấy C. zeylanicum cho hàm
lượng phenolic cao nhất trong khi C. burmanni có hàm lượng flavonoid cao nhất. Các
thử nghiệm hoạt tính chống oxy hóa của dịch chiết ethanol và chiết lỏng siêu tới hạn
sCO2 từ vỏ, chồi và lá C. cassia cho thấy cao chiết ethanol của vỏ quế có hoạt động
chống oxy hóa mạnh nhất so với các bộ phận khác (Chao L., 2008) [69].
Hoạt tính kháng viêm: Các đại thực bào có liên quan đến tình trạng viêm mãn
tính bằng cách sản xuất chất trung gian gây viêm khác nhau bao gồm các cytokine,
chemokine, interferon, các yếu tố kích thích tạo dòng (colony-stimulating factors-
CSF), lysozymes, protease, các yếu tố tăng trưởn, eicosanoids, và nitric oxide (NO).
Trong số này, Nitric oxide (NO) được tổng hợp bởi men tổng hợp NO cảm
ứng (inducible nitric oxide synthase-iNOS) tại nội mô và cơ trơn mạch máu dưới
tác dụng của các cytokine gây viêm. NO sinh ra quá mức bởi iNOS tạo ra rất nhiều
bệnh lý bao gồm hen suyễn, viêm khớp, bệnh đa xơ cứng, viêm đại tràng, bệnh vẩy
nến, rối loạn thoái hóa thần kinh, phát triển khối u và thải ghép sốc nhiễm khuẩn [66].
Tác dụng ức chế sự sản sinh NO của cao chiết vỏ quế C. cassia trên tế bào
macrophage RAW 264.7 đã được Lee &cs., 2002 khảo sát [112], các nghiên cứu
tiếp theo của nhóm nghiên cứu cho thấy một số thành phần trong tinh dầu lá quế C.
cassia như Cinnamaldehyde, Cinnamyl aclcohol có khả năng ức chế sản sinh NO
trong khi rất ít hoặc không có hiệu lực ức chế được quan sát thấy ở một số thành
phần khác như Cadinene, Cinnamic acid, α-Copaene, Eugenol, Limonene [112].
1.4.4. Các phương pháp chiết xuất tinh dầu quế
Tinh dầu quế là loại tinh dầu đắt tiền trên thị trường vì khó chiết xuất so với
đa số các loại tinh dầu khác, giá nguyên liệu cao và chất lượng tinh dầu phụ thuộc
vào từng loại nguyên liệu.
Phương pháp chưng cất hơi nước: Cũng như các loại tinh dầu khác, dầu quế
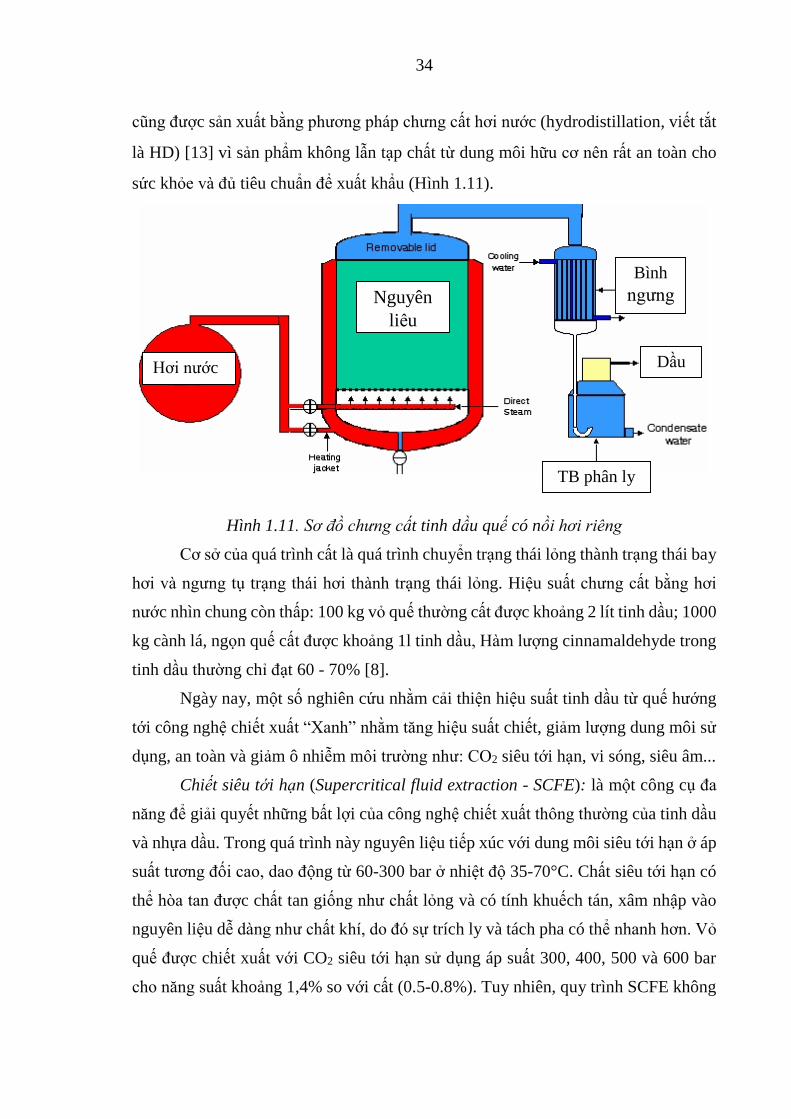
34
cũng được sản xuất bằng phương pháp chưng cất hơi nước (hydrodistillation, viết tắt
là HD) [13] vì sản phẩm không lẫn tạp chất từ dung môi hữu cơ nên rất an toàn cho
sức khỏe và đủ tiêu chuẩn để xuất khẩu (Hình 1.11).
Hình 1.11. Sơ đồ chưng cất tinh dầu quế có nồi hơi riêng
Cơ sở của quá trình cất là quá trình chuyển trạng thái lỏng thành trạng thái bay
hơi và ngưng tụ trạng thái hơi thành trạng thái lỏng. Hiệu suất chưng cất bằng hơi
nước nhìn chung còn thấp: 100 kg vỏ quế thường cất được khoảng 2 lít tinh dầu; 1000
kg cành lá, ngọn quế cất được khoảng 1l tinh dầu, Hàm lượng cinnamaldehyde trong
tinh dầu thường chỉ đạt 60 - 70% [8].
Ngày nay, một số nghiên cứu nhằm cải thiện hiệu suất tinh dầu từ quế hướng
tới công nghệ chiết xuất “Xanh” nhằm tăng hiệu suất chiết, giảm lượng dung môi sử
dụng, an toàn và giảm ô nhiễm môi trường như: CO2 siêu tới hạn, vi sóng, siêu âm...
Chiết siêu tới hạn (Supercritical fluid extraction - SCFE): là một công cụ đa
năng để giải quyết những bất lợi của công nghệ chiết xuất thông thường của tinh dầu
và nhựa dầu. Trong quá trình này nguyên liệu tiếp xúc với dung môi siêu tới hạn ở áp
suất tương đối cao, dao động từ 60-300 bar ở nhiệt độ 35-70°C. Chất siêu tới hạn có
thể hòa tan được chất tan giống như chất lỏng và có tính khuếch tán, xâm nhập vào
nguyên liệu dễ dàng như chất khí, do đó sự trích ly và tách pha có thể nhanh hơn. Vỏ
quế được chiết xuất với CO2 siêu tới hạn sử dụng áp suất 300, 400, 500 và 600 bar
cho năng suất khoảng 1,4% so với cất (0.5-0.8%). Tuy nhiên, quy trình SCFE không
Hơi nước
Nguyên
liệu
Dầu
Bình
ngưng
TB phân ly

35
được thực hiện cho sản xuất quế thương mại hoặc sản xuất nhựa oleoisin do quá trình
này là rất tốn kém và sản phẩm không khác biệt nhiều về chất lượng từ các sản phẩm
chiết xuất dung môi.
Phương pháp vi sóng hỗ trợ (Microway assisted hydrodistillation -MAHD):
Jeyaratnam N. et al. (2016) đã sử dụng vi sóng hỗ trợ quá trình chưng cất tinh dầu vỏ
quế (MAHD). Phổ GC-MS cho thấy phương pháp vi sóng tăng 9% các hợp chất chứa
oxy so với phương pháp thủy chưng cất (HD). Hơn nữa, MAHD tiết kiệm năng lượng
hơn, thân thiện với môi trường khi giảm phát thải CO2 tổng thể 59% so với HD. Ngoài
ra, các nghiên cứu hoạt tính gây độc tế bào của tinh dầu MAHD có giá trị LC50 (51,2
mg/l) thấp hơn so với HD (68,9 mg/l) [137].
Phương pháp siêu âm: Ping Li et al (2015) đã sử dụng siêu âm để hỗ trợ quá
trình chưng cất tinh dầu vỏ quế. Quá trình siêu âm cũng có tác dụng lớn vào sản lượng
dầu. Hiệu suất tinh dầu quế có thể đạt tới 14,8034%, trong đó trans- cinnamaldehyde
là 82,62% bằng phân tích HPLC [140]. Tuy nhiên phương pháp này cũng khó áp
dụng ở qui mô công nghiệp.
Phương pháp enzyme hỗ trợ (enzyme assisted distillation- EAD) hoặc enzyme
hỗ trợ kết hợp với vi sóng (enzyme assisted microwave distillation -EAMD): đến nay
chưa được ứng dụng trong trích ly tinh dầu quế hay những cây cho tinh dầu từ thân
và vỏ thân (Long não) hay những cây cho tinh dầu từ rễ, thân rễ (Hương bài, Pơ Mu,
Thiên niên kiện,...) hay tinh dầu thuộc thành phần khó bay hơi như nhựa hay sáp
(Trầm hương)...
1.5. Tổng quan về cá ngừ
1.5.1. Sản lượng và giá trị dinh dưỡng của phụ phẩm cá ngừ
Cá ngừ đại dương là loại cá lớn thuộc họ Cá bạc má (Scombridae), chủ yếu
thuộc chi (Thunnus), sinh sống ở vùng biển ấm, cách bờ độ 185 km trở ra, bao gồm
cá ngừ vây vàng Thunnus albacares, cá ngừ mắt to Thunnus obesus, cá ngừ bò
Thunnus tonggol. Một số loài nằm trong các chi khác thuộc họ Scombridae như cá
ngừ ồ Auxis rochei, cá ngừ chù Auxix thazad, cá ngừ chấm Euthynnus affinis , cá ngừ
vằn Katsuwonus pelamis... Ở nước ta, nhóm loài di cư đại dương (cá ngừ vây vàng
và cá ngừ mắt to) là những loài kích thước lớn 70-200 cm, khối lượng 1,6-64kg, còn
lại là nhóm loài có kích thước nhỏ 20-70 cm, khối lượng từ 0.5-4kg, di cư trong phạm

36
vi địa lý hẹp.
Vùng biển Việt Nam có trữ lượng cá ngừ ước tính 600 000 tấn, trong đó khả
năng cho phép khai thác cá ngừ vằn hơn 220 000 tấn/năm (theo đánh giá nguồn lợi
và trữ lượng cá ngừ ở biển Việt Nam giai đoạn 2011-2013 do Viện Nghiên cứu Hải
sản thực hiện), cá ngừ vây vàng, mắt to hơn 45 000 tấn, khả năng cho phép khai thác
từ 17000 - 22 000 tấn/năm. Từ năm 2012, nghề câu tay kết hợp với ánh sáng phát
triển, năng suất khai thác vượt trội, số lượng tăng mạnh nên năng suất đạt 16234 tấn
Năm 2011, trong khi sản lượng khai thác cá ngừ toàn cầu đạt khoảng 4,76 triệu
tấn, thì lượng sản phẩm cá ngừ đóng hộp gần 2 triệu tấn. Chất thải rắn hoặc các phụ
phẩm được thải ra từ sản xuất cá ngừ đóng hộp (bao gồm đầu, bộ xương, nội tạng,
mang, phần thịt màu sẫm, vây bụng và da) có thể chiếm khoảng 65% lượng nguyên
liệu ban đầu. Các số liệu báo cáo trong ngành sản xuất thịt cá ngừ cũng cho thấy các
phế phẩm, phụ phẩm chiếm khoảng 40% tổng nguyên liệu ban đầu.
Bảng 1.2. Tỉ lệ các thành phần của cá ngừ
Đầu (%) Xương (%) Vây, vẩy (%) Nội tạng (%) Thịt cá (%)
20 8 1 11 60
Về sản lượng khai thác cá ngừ đại dương, theo FAO tổng sản lượng khai thác
cá ngừ đại dương trên Thế Giới đã tăng lên không ngừng từ những năm 1950 và
ước đạt 393.134 tấn và năm 2010 là 4.246.558 tấn. Riêng ở Việt Nam tổng sản
lượng cá ngừ đại dương đánh bắt được đã không ngừng tăng lên từ những năm 2007
đến 2011. Theo Nguyễn Anh Tuấn và cộng sự (2011) tổng sản lượng cá trong năm
2007 đạt 19979.2 tấn và đến năm 2011 tăng lên đến 22938,1 tấn [38]. Trong khi trữ
lượng ước tính vượt trên 100.000 tấn bao gồm cả hai loài nói trên. Như vậy, theo
Bảng 1.2, mỗi năm 8000 tấn phụ phẩm cá ngừ đại dương, trong đó riêng đầu cá ngừ
đại dương 4000 tấn/năm. Đây là nguồn phụ phẩm rất có giá trị, đầu và xương cá ngừ
còn chứa rất nhiều các cơ thịt, vì vậy có thể lợi dụng quá trình thuỷ phân để tạo ra
những sản phẩm có ích như thức ăn chăn nuôi, có thể sử dụng sản phẩm thủy phân
trong việc sản xuất bột nêm gia vị, bột đạm dinh dưỡng, nước mắm… [11]. Theo
Bougatef Ali (2012), tổng axit amin thu được từ quá trình thủy phân protein đầu cá
ngừ vây xanh (Thunnus thynnus) bằng enzyme Alcalase khoảng 35%, trong đó các
axit amin thiết yếu chiếm 50,52% axit amin tổng số. Các protein thủy phân từ cá ngừ

37
có hoạt tính oxy hóa cao rất hữu ích trong thực phẩm và dinh dưỡng [57].
Đầu và bộ xương cá ngừ còn có hàm lượng canxi và photpho thích hợp để bổ
xung canxi vào thức ăn chăn nuôi hoặc bổ sung vào thực phẩm phục vụ cho con
người. Xương, bong bóng, da cá ngừ là nguyên liệu tốt dùng để sản xuất keo cá có
chất lượng cao. Gelatin trong công nghiệp thực phẩm dùng làm nguyên liệu phụ trong
sản xuất bánh điểm tâm, lạp xưởng, đồ hộp, kem cốc, chất ổn định và chất nhũ hoá
trong thực phẩm, và rất nhiều công dụng trong công nghiệp.
Ngoài ra trong phụ phẩm cá ngừ, đặc biệt là các loài cá ngừ đại dương có chứa
hàm lượng lipid cao với nhiều các axit béo không no đa nối đôi thiết yếu có giá trị [12]
[57] [138] [157]. Hàm lượng lipid trong đầu cá ngừ đại dương dao động trong khoảng
10-15% như cá ngừ vây vàng Thunnus albacares (13.5%) [138], đầu cá ngừ mắt to
Thunnus obesus (14.2%) [30], đầu cá ngừ vây xanh Thunnus thynnus (10.4%) [57]. So
với các phế liệu đầu cá ngừ kích thước nhỏ như cá ngừ sọc dưa Sarda orientalis
(8.02%), cá ngừ chấm Euthynnus affinis (5.5%) [30] và một số phế liệu đầu cá hồi
(3.57%), đầu cá basa (3,41%), đầu cá tra (3.15%) [17], cho thấy phụ phẩm đầu cá ngừ
là nguồn nguyên liệu tiềm năng sản xuất dầu cá và các sản phẩm giàu các axit béo
không đo đa nối đôi n-3 PUFA phục vụ trong công nghiệp thực phẩm và y dược.
1.5.2. Dầu cá và các axit béo không no đa nối đôi n-3 PUFA
Lipid (chất béo, dầu cá) trong cá chủ yếu là các triglycerid, do các axit béo bậc
cao kết hợp với glycerin. Ngoài ra, còn có thành phần không phải là glycein gọi là
chất không xà phòng. Ở nhiệt độ thường, phần lớn chất béo bày ở thể lỏng nên còn
gọi là dầu cá [16].
Axit béo không no đa nối đôi PUFA có vai trò quan trọng trong các lĩnh vực y,
dược và công nghiệp thực phẩm. Đây là những axit béo thiết yếu (essential fatty acid),
không tự sinh tổng hợp được trong cơ thể người mà phải bổ sung qua con đường thức ăn.
Nếu thiếu hụt các axit béo này thì sẽ gây ra mất cân bằng, làm rối loạn hệ thống miễn dịch
và chống viêm của cơ thể, dẫn đến hàng loạt bệnh tật phát sinh.
Các axit béo đa nối đôi mạch siêu dài nhóm omega-3 tiêu biểu được đưa ra
làm đại diện gồm có Alpha linoleic acid (ALA C18:3n-3 ), Eicosapentaenoic axit
(EPA C20:5 n-3), Docosahexaenoic axit (DHA C22:6 n-3). Ngoài ra, còn một thành

38
phần trung gian của hai axit béo này là Docosapentaenoic axit (DPA C22:5 n-3). Các
axit này có đặc thù riêng, sản phẩm chứa đồng thời cả ba loại axit béo này với hàm
lượng lớn thường rất hiếm gặp, chủ yếu chúng có nguồn gốc từ vi tảo hoặc một số
loài sinh vật biển đầu chuỗi thức ăn.
Alpha linolenic acid (C18:3 n-3)
Eicosapentaenoic acid (C20:5 n-3)
Docosapentaenoic acid (C22:5 n-3)
Docosahexaenoic acid (C22:6 n-3)
Hình 1.12. Cấu trúc của một số acid béo nhóm n-3 PUFA
Alpha linoleic acid (ALA) được tạo thành từ quá trình khử no của linoleic acid
dười tác dụng của enzyme 15-desaturase. Con người không tự tổng hợp được acid
này trong cơ thể do không có 15-desaturase, vì vậy chúng phải cung cấp qua chế độ
ăn. ALA ở dạng ester glycerol trong một số loài thực vật như dầu lanh, dầu tía tô, dầu
hạt cải và dầu đậu tương.
Trong cơ thể Eicosapentaenoic acid (EPA) được xem là axit béo thiết yếu sẽ
chuyển hoá thành các chất sinh học quan trọng như prostaglandin, leucotrien… Loại
prostaglandin này có tác dụng ức chế sự đông vón tiểu cầu, giảm và phòng ngừa hình
thành huyết khối, đồng thời có thể giảm bớt lượng cholesterol, giảm bớt triglyceride
trong máu làm giảm độ nhớt dính của máu, giữ cho tuần hoàn được thông thoáng.
EPA còn tác dụng làm giảm tình trạng xơ vữa động mạch. Vì vậy EPA có tác dụng
tốt đối với việc phòng ngừa và chữa trị các bệnh tim mạch do xơ vữa mạch Akoh C,
2008, [41] [Chow Ching K. 2008, [74].
Docosahexaenoic acid (DHA) là thành phần quan trọng của phospholipid
màng tế bào, đặc biệt là não bộ, võng mạc. Thành phần của não là chất béo và DHA
chiếm khoảng 1/4 lượng chất béo này. Chúng cần thiết cho sự phát triển hoàn thiện

39
chức năng nhìn của mắt và sự phát triển hoàn hảo của hệ thần kinh, não bộ ở giai
đoạn đầu đời. Với người lớn, DHA có tác dụng bảo vệ tim mạch do làm giảm
Cholesterol và Triglycerid máu. DHA liều cao còn có khả năng ức chế và tiêu diệt tế
bào ung thư trên thực nghiệm [Đặng Diễm Hồng, 2016] [9] [Akoh C.C et al., 2008]
[41], [Makrides M. et al., 2010] [127].
Docosapentaenoic acid (DPA) chứa hai chất đồng phân, cụ thể là DPAn-3 và
DPAn-6, trong đó có mặt chủ yếu trong sinh vật biển. DPAn-3 được tìm thấy chủ yếu
trong dầu cá và dầu hải cẩu, trong khi một số loại dầu vi tảo chứa đáng kể lượng
DPAn-6 [107]. DPA đồng phân là trung gian của sự trao đổi chất và sinh tổng hợp
các PUFA. Các nghiên cứu cho rằng, tỷ lệ DPA và DHA làm giảm đáng kể nguy cơ
biến tính mạch vành cấp tính, trong khi đó không có mối liên quan giữa tỷ lệ của EPA
và nguy cơ biến cố mạch vành cấp. DPA có thể hiệu quả hơn EPA hoặc DHA trong
một số ứng dụng trị liệu, chẳng hạn như giữ thành động mạch mềm và mảng bám tự
do và giúp đỡ để chữa lành các mạch máu bị hư hỏng.
Lipid (dầu) cá ngừ chứa hàm lượng cao các axit béo không no đa nối đôi n-3
PUFA (khoảng 6% EPA, 25% DHA và khoảng 3% đồng phân DPA [96], cao hơn
các dầu cá khác. Linder et al., (2005) công bố trong dầu cá ngừ chứa 5,1% EPA,
18,8% DHA và nhiều axit béo không no khác rất có lợi cho sức khỏe con người. Dầu
mắt cá ngừ chứa hàm lượng lipid tổng là 22,4%, trong đó chủ yếu là axit béo không
no (PUFA) không có khả năng sinh cholesterol.
EPA và DHA có lẽ là các axit béo thương mại thành công nhất có nguồn gốc
từ dầu cá. Mặc dù bắt đầu phát triển khá chậm từ năm 2000, thị trường omega-3 hiện
nay đã có những tăng trưởng rất đáng kể. Theo một số nghiên cứu thị trường, trong
năm 2010 nhu cầu toàn cầu cho các thành phần của omega-3 là khoảng 1,595 tỷ USD
[38].
1.5.3. Các phương pháp chiết xuất lipid và làm giàu các axit béo không no
Phương pháp chiết xuất lipid tổng
Chiết lipid tổng là một trong những bước quan trọng nhất trong toàn bộ qui
trình bởi hàm lượng lipid tổng ban đầu và thành phần các hoạt chất trong lipid tổng
sẽ ảnh hưởng trực tiếp tới hiệu suất qui trình.
Trong nghiên cứu, chiết xuất lipid từ sinh vật biển có những đặc trưng riêng

40
biệt so với các sinh vật trên cạn do trong cơ thể sinh vật biển chứa hàm lượng nước
cao (>70%) và có nhiều muối khoáng. Mặt khác, lipid từ sinh vật biển có nhiều axit
béo không no đa nối đôi nên phương pháp đồi hỏi phải xử lý mẫu một cách thận trọng
để giảm sự oxy hóa tới mức tối thiểu [a10]. Tuy không tan trong nước mà chỉ tan
trong dung môi hữu cơ nhưng lipid tổng vẫn khó chiết vì một số dung môi không hòa
tan được các lipid phân cực (chủ yếu là phospholipid). Chính vì vậy, phương pháp
chiết và tinh chế lipid phải đáp ứng các điều kiện nhanh và hiệu quả.
Rất nhiều qui trình chiết xuất lipid nhưng qui trình của Blight & Dyer (1959),
hỗn hợp CHCl3 và CH3OH được sử dụng rộng rãi như các dung môi chiết lipid và
quan sát giản đồ CHCl3-CH3OH-H2O kéo theo những giả thiết việc chiết lipid thuận
lợi nhất đem lại hiệu quả cao. Phương pháp Blight & Dyer nổi trội hơn các phương
pháp chiết khác là thực hiện trong thời gian ngắn, hiệu suất cao, có thể tái sử dụng
dung môi và tránh được các thao tác có hại.đã được áp dụng cho nghiên cứu, sàng
lọc, khảo sát lipid và axit béo trên đối tượng sinh vật biển. Tuy nhiên, nếu triển khai
ở qui mô công nghiệp, tồn dư dung môi trong sản phẩm cần quan tâm.
Chiết xuất lipid bằng phương pháp ép nhiệt có tính an toàn vì không phải sử
dụng các dung môi hữu cơ ảnh hưởng có hại cho sức khỏe con người, tuy nhiên hiệu
suất chiết xuất lipid thấp hơn phương pháp dung môi do còn dư lại một lượng lớn
lipid còn lại trong nguyên liệu. Phương pháp ly tâm được sử dụng kết hợp đã cải thiện
được hiệu quả chiết. Tuy nhiên, phương pháp này phù hợp với nguyên liệu là hạt
chứa dầu hoặc nội quan sinh vật biển có chứa hàm lượng dầu cao trong nguyên liệu,
đồng thời nhiệt độ cao làm bất hoạt các enzyme thủy phân có trong nguyên liệu, làm
giảm tác động của enzyme lên lipid chiết xuất được.
Tao và cộng sự (2010) đã nghiên cứu tách chiết dầu từ mắt cá ngừ vây vàng bằng
phương pháp tách chiết siêu tới hạn. Kết quả cho thấy dầu cá thô thu được có chỉ số
peroxyt là 2,33 mg/kg, chỉ số iot là 192,19% và chất không xà phòng hóa là 0,77%.
Ngày nay, phương pháp chiết xuất lipid dưới tác dụng của enzyme protease
đang được áp dụng nhiều trên nhiều đối tượng sinh vật biển và các hạt có dầu. Các
enzyme protease thủy phân protein màng tế bào hay màng bào quan thành các peptide
nhỏ hơn và các amino axit, thúc đẩy quá trình giải phóng dầu. Ưu điểm chính đối với
phương pháp tách chiết dầu bằng enzyme là nó có thể được tiến hành dưới các điều

41
kiện nhẹ nhàng như: nhiệt độ thấp (< 60oC) nên hạn chế đến mức tối thiểu sự oxy hóa
các axit béo không bão hòa đa nối đôi (PUFA), không sử dụng dung môi hữu cơ và
quá trình thủy phân tiến hành trong một khoảng thời gian ngắn (Mbatia, 2011) [128].
Phương pháp làm giàu axit béo không no đa nối đôi PUFA
Phương pháp hóa học là phương pháp đang được sử dụng phổ biến hiện nay,
lipid tổng được thủy phân bởi tác nhân kiềm hoặc axit tạo thành các axit béo tự do.
Sau đó, hỗn hợp aixt béo PUFA được tách khỏi hỗn hợp khi cho kết tinh với ure trong
etanol. Việc phát hiện khả năng tạo phức kết tinh với ure các hợp chất hydrocarbon
mạch thẳng trong dung môi etanol hoặc metanol được thực hiện bởi Bengen F. và các
cộng sự (1940) như một cuộc cách mạng trong công nghiệp hóa dầu. Kỹ thuật này rất
hiệu quả trong việc làm sạch các axit béo không no đa nối đôi từ lipid tổng. Tuy
nhiên, quá trình thủy phân lipid bằng kiềm hoặc axit thành các axit béo tự do ở nhiệt
độ 70-80oC trong thời gian dài đã làm đổi màu lipid tổng, đồng thời tiêu tốn lượng
lớn kiềm và nước thải ra môi trường sau thủy phân, gây ô nhiễm môi trường sinh thái.
Tách trên cột Silica gel, tráng bạc AgNO3, phù hợp trên quy mô nghiên cứu
lượng nhỏ cho phân tích thí nghiệm, tuy nhiên trong triển khai với quy mô lớn cho
ứng dụng sản xuất thì không phù hợp vì AgNO3 có giá thành rất cao.
Dùng Sắc kí lỏng cao áp pha đảo C18, tuy nhiên phương pháp này sẽ tốn lượng
lớn dung môi tinh khiết, làm cho chi phí lớn dẫn tới giá thành sản phẩm sẽ cao.
Sử dụng hệ enzyme đặc hiệu Lipase để thủy phân lipid thành các axit béo tự
do. Phương pháp này đã được Gloria &cs., 2010 sử dụng để làm tăng hàm lượng
omega 3 axit (EPA và DHA) từ dầu cá hồi bằng enzyme lipase cố định. Một số nghiên
cứu khác nhờ sử dụng hệ enzyme lipase cũng đã được trình bày bởi Fernandez-
Lafuente R &cs., (1998), Godoy CA &cs., (2010)... Ưu điểm của phương pháp sử
dụng enzyme là hạn chế được việc sử dụng dung môi, hóa chất, ít gây độc hại. Sự kết
hợp phương pháp thủy phân lipase với phương pháp kết tinh ure trong làm giàu các
axit béo không no PUFA cũng đã được đề cập bởi một số tác giả. Nghiên cứu cho
thấy sự kết hợp của phương pháp thủy phân bằng enzyme kết hợp với quá trình tạo
phức với ure là một phương pháp hứa hẹn để thu được các axit béo chưa bão hòa
omega-3 nồng độ cao.
CHƯƠNG 2. NGUYÊN VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU

42
2.1. Nguyên liệu
2.1.1. Mẫu nguyên liệu
- Cành lá quế và vỏ quế Cinnamomum cassia
Hình 2.1. Mẫu cành lá quế C. cassia Hình 2.2. Mẫu vỏ quế C. cassia
Mẫu cành lá quế (Hình 2.1), vỏ quế (Hình 2.2) thu hái ở xã Tân Đồng, huyện
Trấn Yên, tỉnh Yên Bái. Mẫu được thu ở cây có độ tuổi 12 năm trồng và được thu hái
vào 3/2015. Các mẫu nghiên cứu được PGS. TS. Trần Huy Thái, Viện Sinh thái và
Tài nguyên sinh vật định danh là loài Cinnamomum cassia (L.) J.Presl thuộc họ
Lauraceae. Nguyên liệu được hong khô trong bóng râm, xay nghiền thành bột, kích
thước 1 - 1,5 mm và đóng gói trong túi ni lông hút chân không, bảo quản ở nơi kín
gió, tránh ánh sáng trực tiếp cho đến khi sử dụng.
- Bột gỗ gió bầu Aquilaria crassna
Hình 2.3. Bột gỗ cây gió bầu Aquilaria crassna

43
Bột của gỗ cây gió bầu Aquilaria crassna trồng ở Phú Quốc. Mẫu được cung
cấp bởi kỹ sư lâm nghiệp Nguyễn Thoan, giám đốc công ty TNHH Lâm Viên, Xuân
Đỉnh, Bắc Từ liêm, Hà Nội.
- Phụ phẩm đầu cá ngừ
Hình 2.4. Mẫu đầu cá ngừ vây vàng Thunnus albacares
08 mẫu đầu cá ngừ (đã bỏ mắt và mang) của 08 loài cá ngừ có các kích cỡ
khác nhau được cung cấp bởi các công ty chế biến thủy hải sản tại thành phố Nha
Trang, tỉnh Khánh Hòa và thành phố Tuy Hòa, tỉnh Phú Yên. Đầu cá ngừ vây vàng
được cung cấp bởi công ty TNHH JK FISH Nha Trang, Khánh Hòa. Các mẫu nguyên
liệu được cấp đông ở -18oC trước khi chuyển về cơ sở nghiên cứu. Nguyên liệu được
đánh số ký hiệu, bảo quản ở -4oC tại tủ đông của Trung tâm Nghiên cứu và phát triển
các SPTN, Viện Hóa học các HCTN.
2.1.2. Enzyme sử dụng trong nghiên cứu
+ Enzyme Cellic Htec2 (Novozyme-Đan Mạch) là hỗn hợp đa enzyme bao
gồm cellulase và xylanase, phổ cơ chất rộng. Enzyme hoạt động ở nhiệt độ 40 - 600C
(tối ưu 500C), pH 4,5 - 6,0 (tối ưu 5,5). Hoạt tính xác định được: cellulase 2800 UI/g
theo phương pháp của Wood, (1987) [171], xylanase 1500 UI/g theo phương pháp
của Bailey, (1992) [48].
+ Enzyme laccase là enzyme thô thu từ canh trường nuôi cấy nấm Ganoderma
lucidum. Chủng giống Ganoderma lucidum được cung cấp bởi Viện Di truyền nông
nghiệp, cấy truyền và giữ giống ở 40C trên môi trường thạch PDA. Môi trường lên men
bao gồm 50% PDA lỏng, 10 g glucose, 2 g MgSO4·7H2O and 3 g KH2PO4. Bổ sung

44
0,05% chất cảm ứng ABTS (2,2'-di--azino 3-ethyl-benzothiazolin-sulphonate). Hoạt
tính xác định được là 178 UI/ml theo phương pháp của Ding et al. (2012) [77].
+ Nhóm enzyme protease từ vi sinh vật sử dụng hai enzyme Protamex và
Alcalase mua của hãng Novozyme Đan Mạch; Protamex được sản xuất nhờ quá trình
lên men chìm chủng Bacillus amyloliquefaciens thường được dùng để phá vỡ liên kết
peptide. Đây là một protease trung tính hoạt động tối ưu trong khoảng pH 5,5 - 7,5;
nhiệt độ tối ưu: 45 - 550C. Hoạt tính Protamex theo khuyến cáo của nhà sản xuất là
1,5 AU/g. Alcalase là enzyme protease kiềm được sản xuất từ Bacillus licheniformis.
Alcalase có tính đặc hiệu cơ chất rất rộng, hoạt động giữa pH 7,5 và 9,0, tối ưu ở 8,5;
nhiệt độ hoạt động tối ưu 45 và 650C với hoạt tính mạnh nhất vào khoảng 550C. Hoạt
tính Alcalase theo khuyến cáo của nhà sản xuất là 1.5 AU/ml.
+ Nhóm enzyme Protease từ thực vật sử dụng hai enzyme Bromelain và
Papain, là enzyme trong nước, cung cấp bởi công ty Biogreen. Hoạt tính khuyến cáo
của nhà sản xuất là 2000 UI/mg.
+ Enzyme Lipase CRL của Novozyme được sản xuất từ quá trình lên men của
nấm men Candida rugosa, là hỗn hợp đa enzyme với 5 loại isoenzyme (chủ yếu là
hydrolase và esterase), khả năng thủy phân ngẫu nhiên liên kết ester của glycerol và
axit béo và hoạt tính theo khuyến cáo của nhà sản xuất > 2000 UI/mg. Hoạt động ở
pH 6 - 8 (tối ưu pH 7,0), nhiệt độ 30 - 40°C (tối ưu 35°C).
2.1.3. Bộ chủng vi sinh vật kiểm định
Vi khuẩn Gram (-): Escherichia coli ATCC 25922, Pseudomonas aeruginosa
(ATCC 25923; Vi khuẩn Gram (+): Bacillus subtillis ATCC 11774, Staphylococcus
aureus subsp. aureus ATCC 11632; Nấm sợi: Aspergillus niger 439, Fusarium
oxysporum M42; Nấm men: Candida albicans ATCC 7754, Saccharomyces
cerevisiae SH 20.
Môi trường duy trì và bảo tồn giống nuôi cấy vi sinh vật kiểm định: Saboraud
Dextrose Broth (Sigma-Aldrich, Mỹ) cho nấm men và nấm sợi. Trypcase Soya Broth
(Sigma-Aldrich, Mỹ) cho vi khuẩn Gram (-) và Gram (+).
Môi trường thí nghiệm: Eugon Broth (Difco, Mỹ) cho vi khuẩn và Mycophil

45
(Difco, Mỹ) cho nấm.
Các chủng VSV kiểm định đang được lưu giữ tại phòng Sinh học thực nghiệm,
Viện Hóa học các Hợp chất thiên nhiên, Viện Hàn lâm Khoa học và Công nghệ Việt
Nam.
2.1.4. Dòng tế bào
- Đại thực bào macrophage RAW 264.7 do GS. J. M. Pezzuto, Trường Đại học
Hawaii và GS. Jeanette Maier, trường Đại học Milan, Italia cung cấp, đang được lưu
giữ tại phòng thử nghiệm sinh học, Viện Công nghệ Sinh học, Viện Hàn lâm Khoa
học và Công nghệ Việt Nam. Dòng tế bào RAW264.7 được nuôi cấy trong môi trường
DMEM với thành phần kèm theo gồm 2 mM L-glutamine, 10 mM HEPES, và 1,0
mM sodium pyruvate, ngoài ra bổ sung 10% fetal bovine serum - FBS (GIBCO). Tế
bào được cấy chuyển sau 3 - 5 ngày với tỉ lệ (1:3) và nuôi trong tủ ấm CO2 ở điều
kiện 37oC, 5% CO2.
- 03 dòng tế bào ung thư bao gồm dòng Hep-G2 (Human hepatocellular
carcinoma - Ung thư gan), dòng LU-1 (Human lung adenocarcinoma - Ung thư biểu
mô phổi) và dòng RD (Human rhabdomyosarcoma - Ung thư mô liên kết). Các dòng
tế bào ung thư đang được lưu giữ tại phòng Sinh học thực nghiệm, Viện Hóa học các
hợp chất thiên nhiên, Viện Hàn lâm khoa học và Công nghệ Việt Nam.
2.2. Hóa chất, thiết bị
2.2.1. Hóa chất
+ Các loại dung môi: ethanol (99,5 %), methanol (99,5 %), axeton (99%).
+ Các thuốc thử: DNS (Signa), ABTS, Congo red.
+ Các hóa chất phân tích: bản mỏng Silica gel 60 F254 của Merck, NaOH,
Na2SO4, HCl, KOH, Ure, CMC, xylan từ gỗ mềm (Sigma), xylose, glucose, Casein
sodium caseinat (Sigma), Trichloroacetic acid (TCA) (Trung Quốc).
+ Dung dịch thuốc thử định tính enzyme: Congo đỏ, Thuốc thử Lugon
+ Lipopolysaccharides (LPS) từ E. coli của Sigma Chemical Co. (St. Louis,
MO, USA). Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum
(FBS) của hãng Life Technologies, Inc., (Gaithersburg, MD, USA). Sodium nitrite,
sulfanilamide, N-1-napthylethylenediamine dihydrochloride and dimethyl sulphoxide

46
(DMSO) của Sigma Chemical Co. (St. Louis, MO, USA).
+ Các dung dịch đệm
+ Một số hóa chất khác
2.2.2. Thiết bị
Thiết bị chủ yếu sử dụng trong nghiên cứu: thiết bị chưng cất tinh dầu quy mô
100 g nguyên liệu/mẻ và 500 g nguyên liệu/mẻ, thiết bị cô quay chân không Buchi
(Thụy sỹ), thiết bị sắc ký khí GC-MS6890N Aligent Technologies (Mỹ), máy ly tâm
Universal 32, tủ sấy tự động khống chế nhiệt độ KOTTERMANN (Đức), Tủ lạnh, Tủ
lạnh đông Alaska (Việt Nam), máy xay, máy khuấy, bể ổn nhiệt (Memmert), máy so
màu UV-VIS Double Beam, Model UVD-3200 Labomed, máy khuấy, máy khuấy từ
gia nhiệt, pH meter (713- pH Meter Metrohm)....
2.3. Phương pháp nghiên cứu
2.3.1. Các phương pháp phân tích hóa sinh
2.3.1.1. Một số phương pháp phân tích trên mẫu thực vật
- Phương pháp xác định hàm ẩm được theo TCVN 1867:2001.
- Phương pháp xác định hàm lượng cellulose theo phương pháp Kurshner [5].
- Phương pháp xác định hàm lượng hemicellulose theo phương pháp Bromua-
Bromat biến tính (Tương ứng với tiêu chuẩn TAPPI T223) [5].
- Phương pháp xác định hàm lượng lignin Klason xác định theo phương pháp
biến tính Komarov [5].
- Phương pháp xác định hàm lượng chất trích ly trong cồn [5].
- Phương pháp xác định hàm lượng tinh dầu theo Dược điển Việt Nam IV [2].
2.3.1.2. Một số phương pháp phân tích trên mẫu cá ngừ
- Phương pháp xác định hàm lượng lipid tổng theo Blight & Dyer [56].
- Phương pháp xác định hàm lượng protein tổng theo Kjeldahl TCVN
4328:2001.
- Phương pháp xác định hàm lượng tro theo TCVN 5253-1990.
- Phương pháp xác định chỉ số axit theo TCVN 6127:1996.
- Phương pháp xác định chỉ số xà phòng hóa theo TCVN 6126: 2007.
- Phương pháp xác định chỉ số peroxit theo TCVN 6121 : 2010.
2.3.2. Các phương pháp phân tích thành phần hóa học

47
2.3.2.1. Phương pháp xác định thành phần tinh dầu bằng GC-MS
Thành phần hóa học của tinh dầu quế được xác định trên máy sắc ký khí Agilent
HP mode 7890A gas chromatograph trang bị cột tách mao quản (60 m x 0.25 mm x
0.25 µm) Hewlett Packard HP5-MS (5% phenyl methyl siloxane), kết nối với máy đo
phổ khối Agilent 5975C mass spectrometer. Các điều kiện phân tích sắc ký bao gồm:
nhiệt độ injector và detector là 250oC; chương trình nhiệt độ từ 60 đến 240°C (tốc độ
4°C/phút); khí mang là He (lưu lượng 1.0 ml/phút); lượng mẫu đo là 1 µl; tỉ lệ chia
dòng là 100:1. Máy đo phổ khối chạy ở chế độ EI-MS, năng lượng ion hóa là 70 eV. Các
chất được nhận dạng bằng cách so sánh phổ khối lượng và chỉ số thời gian lưu của chúng
với các dữ liệu có trong các ngân hàng dữ liệu GC-MS bao gồm NIST 08, Wiley 09 và
HPCH1607. Phần mềm xử lý số liệu phổ Mass Finder 4.0.
Thành phần hóa học của tinh dầu trầm hương cũng được xác định trên máy
sắc ký khí Agilent HP mode 7890A gas chromatograph tương tự như tinh dầu quế
nhưng có sự khác biệt là trang bị cột tách mao quản (60 m x 0.25 mm x 0.25 µm)
Hewlett Packard DB- XLB kết nối với máy đo phổ khối Agilent 5975C mass
spectrometer và chương trình chạy sắc ký nhiệt độ từ 40oC (tăng 20o/phút), giữ ở
140oC (trong 5 phút), tiếp tục tăng lên 2700C (tốc độ 4°C/phút).
2.3.2.2. Phương pháp xác định thành phần axit béo
Mẫu được đo trên hệ thống thiết bị sắc ký khí và phổ ký liên hợp GC/MS của
hãng Agilent Technologies HP 6890N trang bị cột mao quản DB‐ Wax (60 m × 0,25
mm, 0,25 µm) và DB‐ 23 (60 m × 0,25 mm, 0,25 µm). Agilent Technologies HP
6890N ghép nối với khối phổ Agilent 5973 Network Mass Selective Detector. Chạy
theo chương trình nhiệt độ: 140oC/5 phút, 165oC/10 phút, sau đó tăng nhiệt độ lên
190oC (tốc độ 5oC/phút) và giữ ở 190oC/55 phút. Tăng nhiệt độ lên 280oC, nhiệt độ
nguồn ion 210oC, khí mang He, tốc độ dòng 1,5ml/phút. Tỷ lệ chia dòng 30:1. Máy
đo phổ khối chạy ở chế độ EI-MS, năng lượng ion hóa là 70 eV.
Các axit béo được nhận dạng bằng cách so sánh phổ khối lượng và chỉ số thời
gian lưu của chúng với các dữ liệu có trong các ngân hàng dữ liệu GC-MS bao gồm
NIST 08, FAME DB23.L Wiley 09 và HPCH1607. Phần mềm xử lý số liệu phổ Mass
Finder 4.0.
2.3.3. Các phương pháp xác định hoạt độ enzyme

48
2.3.3.1. Phương pháp xác định hoạt độ enzyme cellulase
Hoạt tính cellulase trong dịch nuôi cấy được định lượng bằng phương pháp
carboxymethy cellulase (CMC) theo Wood, (1987) [171].
Cụ thể: 0,9 ml CMC 1% trong 0,2M đệm acetate (pH 5,0) để ở 50°C trong 20
phút ở bể ổn nhiệt. 0,1 ml dịch enzyme đã được hòa loãng các cấp độ khác nhau được
bổ sung vào hỗ hợp phản ứng và ủ ở 50oC trong bể ổn nhiệt trong 20 phút. Mẫu trắng
không chứa enzyme cũng được làm tương tự. Hàm lượng đường khử được sản sinh
trong quá trình phản ứng được xác định bằng thuốc thử DNS (Miller, 1959) ở bước
sóng 540 nm trong máy so màu (ELICO- SL 164). Một đơn vị hoạt tính enzyme
cellulase được xác định bằng lượng enzyme cần thiết để giải phóng 1 µmole đường
glucose trong mỗi phút
CMCase = (TN- Mkt)/20
Trong đó: + TN: Nồng độ đường glucose trong mẫu thí nghiệm (µmole)
+ Mkt: Nồng độ đường glucose trong mẫu đối chứng (µmole)
+ 20: Thời gian phản ứng (phút)
2.3.3.2. Phương pháp xác định hoạt tính xylanase (Bailey, 1992) [48]
Cơ chất: Xylan 1% trong đệm Na-citrat 0.05 M (pH 5.3). Đồng nhất 1 g xylan
trong 80 ml đệm ở 60oC trên bếp khuấy từ gia nhiệt đến khi đồng nhất. Dừng gia
nhiệt, tiếp tục khuấy trong điều kiện mát qua đêm. Bổ sung đệm định mức đến 100
ml.
Phản ứng gồm 0,9 ml cơ chất xylan 1% trong đệm Na-citrat 0,05 M để ở nhiệt
độ 50oC ở bể ổn nhiệt, thêm 100 μl dịch enzyme đã hòa loãng ở cấp độ khác nhau
được bổ sung vào hỗn hợp phản ứng và ủ ở 50oC trong 5 phút (300 s). Mẫu trắng
không chứa enzyme cũng được làm tương tự. Hàm lượng đường khử được giải phóng
trong quá trình phản ứng được xác định bằng thuốc thử DNS (Miler, 1959) ở bước
sóng 540 nm trong máy so màu (ELICO - SL 164). Một đơn vị hoạt tính enzyme
xylanase được xác định bằng lượng enzyme cần thiết để giải phóng 1 µmole đường
xylose trong mỗi phút ở điều kiện phản ứng.
Xylanase (UI/ml) = (MTN - Mkt)/20

49
Trong đó: + MTN: Nồng độ mol đường xylose trong mẫu thí nghiệm (µmole)
+ MKT: Nồng độ mol đường xylose trong mẫu đối chứng (µmole)
+ 20: Thời gian phản ứng (phút)
2.3.3.3. Xác định hoạt tính enzyme Laccase (Ding, 2012) [77]
Dung dịch phản ứng enzyme gồm: 850 µl dung dịch đệm sodium acetate 0,1
M pH 5, 100 µl ABTS 1 mM. Ủ hỗn hợp này ở 27oC trong 10 phút. Bổ sung 50 µl
enzyme. Trộn đều hỗn hợp phản ứng so sánh với mẫu kiểm chứng sau mỗi phút phản
ứng. Phản ứng kiểm chứng sử dụng nước khử ion thay cho enzyme. Một đơn vị hoạt
độ laccase là lượng enzyme cần thiết để xúc tác oxy hóa 1µmol ABTS trong 1 phút
ở 27oC trong dung dịch đệm sodium acetate pH 5.
Hoạt tính laccase được xác định dựa trên sự oxy hóa ABTS (2,2- azino-bis 3-
ethylbenzothi azoline - 6- sulfonic acid) thành hợp chất được hấp thụ ánh sáng mạnh
tại bước sóng 420 nm. Hỗn hợp phản ứng gồm 800 µl đệm acetate 0.5M, pH 5,0; 100
µl ABTS 5mM, 100 µl dịch enzyme. Phản ứng được ủ trong 10 phút ở 40oC. TCA
50% (v/v) được sử dụng làm chất ngừng phản ứng.
Laccase (U/ml) = [A*(106/e420*d)* V/v*F]/t
Trong đó: - A: sự chênh lện giá trị hấp thụ ánh sáng ở 420 nm
- 106/e420*d: sự chuyển đổi sang µmol cơ chất/ ml (εATBS, 420
nm = 3600 M-1)
- d: chiều dài đường sáng đi qua dung dịch theo mm (10mm)
- V: tổng thể tích (1000 µl)
- v: thể tích mẫu (100 µl)
- v/v: độ pha loãng của mẫu thí nghiệm (11)
- F: độ pha loãng ban đầu
- t: thời gian phản ứng (10 phút)
2.3.4. Các phương pháp thủy phân
2.3.4.1. Phương pháp thủy phân với dung môi là nước (áp dụng cho enzyme
cellulase, hemicellulase, protease...)
Nguyên liệu sau sơ chế được bổ sung nước theo tỷ lệ nhất định. Quá trình thủy
phân diễn ra trong bể ổn nhiệt có khuấy cơ học bằng khuấy từ có điều nhiệt. Khi nhiệt
độ trong bình thủy phân phù hợp với nhiệt độ từng enzyme hoạt động thì bổ sung

50
enzyme với tỷ lệ thích hợp. Thời gian thủy phân được tính tại thời điểm bổ sung
enzyme. Sau thủy phân, hỗn hợp thủy phân được gia nhiệt đến 80oC để bất hoạt
enzyme kết thúc quá trình thủy phân.
2.3.4.2. Phương pháp thủy phân trong hệ dung môi hai pha (áp dụng cho
enzyme lipase) theo Fernádez Lorente G. và cs. (2011) [88]
Phản ứng thủy phân được thực hiện trong hệ dung môi hai pha với tỷ lệ pha
nước - pha hữu cơ 1:1 (v/v), tỉ lệ dầu - dung môi 3:7 (w/v). Pha nước chuẩn bị với
đệm ở các dải pH từ 5 - 9. Phản ứng được ủ ở dải nhiệt độ 20 – 50oC và luôn được
khuấy trộn (500 vòng/phút) để đạt được sự phân tán đồng nhất của các pha. Ở các
khoảng thời gian xác định (0 h, 2 h, 4 h ...) mẫu được tách ra bằng ly tâm. Theo
Fernádez Lorente G (2010), 95% axit béo tự do có mặt trong pha dung môi. Vì vậy,
chỉ số axit được xác định sau khi loại toàn bộ dung môi [32].
- Chỉ số axit: Chỉ số axit của dầu cá trước và sau thủy phân được xác định là
số mg KOH cần dùng để trung hòa hết acid béo tự do có trong 1 g chất béo [22].
- Chỉ số xà phòng hóa là số mg KOH dùng để trung hòa hết glycerid và axit
béo tự do có trong 1 g chất béo [22].
Hiệu suất thủy phân =
Chỉ số axit (dầu thủy phân) - Chỉ số axit (dầu chưa thủy phân)
x 100% Chỉ số xà phòng hóa - Chỉ số axit (dầu chưa thủy phân)
2.3.5. Các phương pháp xác định sản phẩm thủy phân
2.3.5.1. Định lượng đường khử bằng DNS (theo Miller, 1959) [129]
Nguyên lý: Dựa trên khả năng hydro hóa cơ chất CMC của enzyme Cellulase
để biến đổi thành đường Glucose. Glucose phản ứng tạo màu với thuốc thử DNS.
Cường độ màu của hỗn hợp phản ứng tỉ lệ thuận với nồng độ đường Glucose trong
một phạm vi nhất định. Sản phẩm sau phản ứng được xác định bằng phương pháp
đo quang ở bước sóng 540 nm. Dựa theo đồ thị đường chuẩn của Glucose tinh khiết
với thuốc thử DNS ta sẽ dễ dàng tính được hàm lượng đường sinh ra trong mẫu thí
nghiệm.

51
Hình 2.5. Đồ thị đường chuẩn Glucose theo phương pháp DNS
2.3.5.2. Định lượng protein- axit amin bằng Coomasssie Brilliant Blue G-250
(theo Bradford, 2009) [59]
Phương pháp Bradford là phương pháp dựa vào sự thay đổi màu xảy ra khi
Coomasssie Brilliant Blue G-250 liên kết với protein (màu xanh) trong dung dịch
axit. G250 liên kết chặt chẽ với các protein, tương tác với các nhóm kỵ nước và các
nhóm mang điện tích dương trên phân tử protein. Các protein khi phản ứng xanh
Coomassie sẽ hình thành hợp chất màu có khả năng hấp thụ ánh sáng ở bước sóng
590 nm, G250 thường được sử dụng để nhuộm đối với những protein có trọng lượng
phân tử polypeptid thấp [22].
Hình 2.6. Đồ thị đường chuẩn protein
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 10 20 30 40 50 60 70
OD
54
0 n
m
Glucose (µg/ml)
y = 1.19x + 0.1996
R2 = 0.9826
y = 0.0149x + 0.0348
R² = 0.9805
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 10 20 30 40 50 60 70
OD
59
5n
m
Nồng độ BSA (µg/ml)

52
2.3.6. Phương pháp làm giàu các axit béo bằng kết tinh ure [11]
Hỗn hợp axit béo FFA được hòa tan dung môi theo tỷ lệ nhất định ure - cồn
3:16, FFA/ure = 1:3. Sau đó, dung dịch thu được để kết tinh ở nhiệt độ 4oC trong thời
gian là 18 - 20 giờ. Sau quá trình kết tinh, hỗn hợp pha rắn và pha lỏng được lọc qua
giấy lọc tại nhiệt độ kết tinh, rửa lại bằng dung môi có nhiệt độ kết tinh, sau đó tách
pha rắn và pha lỏng. Khối lượng hai pha thu được xác định sau khi làm bay hơi dung
môi. Thành phần axit béo của pha lỏng được phân tích trên máy GC-MS.
2.3.7. Các phương pháp nghiên cứu điều kiện tối ưu cho quá trình thủy phân
2.3.7.1. Xử lý số liệu thực nghiệm bằng phần mềm Exel
Quá trình thủy phân được khảo sát một số các yếu tố ảnh hưởng theo phương
pháp tối ưu một yếu tố, giữ nguyên các thông số, chỉ thay đổi 1 thông số. Từ đó, đánh
giá hiệu quả của quá trình thủy phân. Số liệu được xử lý bằng chương trình phần mềm
Microsoft excel.
2.3.7.2. Xử lý số liệu thực nghiệm bằng phương pháp bình phương tối thiểu [7]
Phương pháp tối ưu hóa để lựa chọn một đường khớp nhất cho một dải dữ liệu
ứng với cực trị của tổng các sai số thống kê giữa đường khớp và dữ liệu.
2.3.7.3. Tối ưu hóa bằng phương pháp qui hoạch hóa thực nghiệm [7]
Số thực nghiệm để tìm mô hình hóa thực nghiệm bậc 2 tâm trực giao Box-
Wilson được tính toán theo công thức:
N = 2n ± 2n + No
Trong đó: 2n: số thực nghiệm điểm sao
No: số thực nghiệm ở điểm tâm, thường lấy N0 = 1
Các bước tiến hành qui hoạch hóa thực nghiệm bậc 2 tâm trực giao:
- Xác định và mã hóa các điều kiện tối ưu thực nghiệm (pH, nhiệt độ, tỷ lệ
E/S, thời gian ...).
- Lập ma trận thực nghiệm mã hóa của mô hình thực nghiệm bậc 2 tâm trực giao.
- Tính hệ số hồi qui của phương trình hồi qui bậc 2 tâm trực giao.
- Đánh giá tính có ý nghĩa của hệ số hồi qui.
- Đánh giá tính phù hợp của mô hình theo phương trình hồi qui bậc 2 tâm trực
giao.

53
2.3.8. Các phương pháp xác định hoạt tính sinh học
- Phương pháp xác định hoạt tính kháng VSV kiểm định: theo phương pháp
mô tả bởi Vanden Berghe & Vlietinck (1991) [170].
- Phương pháp xác định khả năng gây độc tế bào (cytotoxicity): theo phương
pháp của Likhitayawuid và cộng sự [119] đang triển khai tại Viện nghiên cứu ung
thư Quốc gia Mỹ (NCI).
- Phương pháp xác định hoạt tính kháng viêm: thông qua khả năng ức chế sản
sinh NO của tế bào macrophage RAW 264.7 theo phương pháp của Lee et al., (2009)
[112].
- Phương pháp xác định hoạt tính chống oxi hóa: được xác định bằng thử
nghiệm trên hệ DPPH (1,1-diphenyl-2-picrylhydrazyl), dựa trên nguyên tắc DPPH có
khả năng tạo ra các gốc tự do bên trong dung dịch ethanol bão hòa theo phương pháp
Blois M S. (1958) [60].

54
CHƯƠNG 3. THỰC NGHIỆM
3.1. Nghiên cứu ứng dụng enzyme kết hợp trong chưng cất tinh dầu
từ các bộ phận khác nhau của cây quế Cinnamomum cassia
Quá trình nghiên cứu chiết xuất tinh dầu quế có enzyme hỗ trợ được thực
hiên theo sơ đồ sau:
Hình 3.1. Qui trình nghiên cứu ứng dụng enzyme trong chiết xuất tinh dầu
Laccase
Htec 2
Nguyên liệu cành lá quế, vỏ
quế
Xay, nghiền
Sơ chế
Xác định thành
phần sinh hóa
Cellulose
Pentose
Ligin
HC trích ly
Thủy phân
Tinh dầu (EAD)
Tinh dầu (HD)
Cất cuốn
hơi nước
Laccase +
Htec 2
Xác định hàm
lượng đường
Xác định hàm
lượng lignin
Hàm lượng tinh dầu
Thành phần hóa học
Hoạt tính sinh học
Đánh giá
Thời gian chưng cất
Cất cuốn
hơi nước

55
3.1.1. Nghiên cứu phân tích thành phần chất nền và tinh dầu của mẫu cành
lá và mẫu vỏ quế C. cassia
3.1.1.1. Phân tích thành phần chất nền của mẫu cành lá và mẫu vỏ quế C. cassia
Mỗi mẫu thí nghiệm lấy 5 g bột nguyên liệu đem phân tích hàm ẩm theo TCVN
1867:2001. Từ đó, xác định hàm lượng hàm lượng cellulose, pentose, hàm lượng
ligin, hàm lượng các chất trích ly theo khối lượng khô tuyệt đối.
3.1.1.2. Xác định hàm lượng và thành phần tinh dầu của mẫu cành lá và
mẫu vỏ quế C. cassia
- Định lượng tinh dầu bằng phương pháp cất cuốn hơi nước (Dược điển Việt
Nam IV [2].
Thí nghiệm 100 g bột cành lá quế, vỏ quế ngâm với nước ở nhiệt độ phòng, tỷ
lệ nguyên liệu - nước 1:5, thời gian ngâm 24 h (cành lá quế), 72 h (vỏ quế). Nguyên
liệu cho vào bình cầu dung tích 2 l, bổ sung thêm nước cất sao cho khối lượng cất
chiếm 50% thể tích bình. Tinh dầu thu được nhờ hệ thống cất tinh dầu Clevenger, sản
phẩm thu được được làm khan bằng Na2SO4.
Hàm lượng tinh dầu (%) = Khối lượng tinh dầu (g)
x 100 Khối lượng nguyên liệu (g)
- Thành phần hóa học của tinh dầu được xác định bằng GC-MS (mục 2.3.2.1).
3.1.2. Nghiên cứu tác động của việc xử lý nguyên liệu với enzyme lên quá
trình chưng cất tinh dầu
3.1.2.1. Nghiên cứu mối tương quan giữa mức độ thủy phân và hiệu suất thu
tinh dầu quế C. cassia
Bột cành lá quế, vỏ quế bổ sung thêm nước theo tỷ lệ tương ứng là 1:5 và 1:7,
ủ qua đêm ở nhiệt độ phòng. Sau 24 h ủ đối với cành lá quế và 72 h ủ đối với vỏ quế,
điều chỉnh bình thủy phân đến nhiệt độ thích hợp cho enzyme hoạt động thì bổ sung
enzyme. Quá trình thủy phân diễn ra trong bể ổn nhiệt, tốc độ khuấy đảo nguyên liệu
120 vòng/phút. Một số thông số thủy phân nguyên liệu cành lá quế và vỏ quế chỉ ra
ở Bảng 3.1.

56
Bảng 3.1. Một số thông số thủy phân nguyên liệu cành lá quế và vỏ quế
TT Thông số quá trình Phương án bổ sung enzyme
Laccase Htec2 Laccase-Htec2
1 pH 3-8 3-8 3-8
2 Nhiệt độ 30-60oC 30-60oC 30-60oC
3 Tỷ lệ E/S 0.2-1 ml/g 0.5-2%
Laccase: 0.2-1,0 ml/g,
Htec2 0.5-2%
4 Tốc độ khuấy 120 vòng/phút 120 vòng/phút 120 vòng/phút
5 Thời gian thủy phân 6 h 6 h 6 h
Các thông số sau đây được xác định trong mỗi thực nghiệm:
* Hàm lượng đường khử trong nguyên liệu sau khi xử lý enzyme (µg/ml).
Các thí nghiệm với 10 g nguyên liệu cho vào bình tam giác 250 ml, bổ sung
50 ml nước. Khử trùng ở nhiệt độ 120oC/30 phút. Quá trình thủy phân diễn ra trên
máy khuấy từ gia nhiệt (IKA -Đức) với các thông số theo các phương án xử lý (Bảng
3.1). Sau 6 h thủy phân, 10 ml dịch sau thủy phân được ly tâm ở 6000 vòng/phút
trong 15 phút. Dịch thủy phân được xác định hàm lượng đường khử bằng phương
pháp DNS trên máy so màu UV-VIS Double Beam, Model UVD-3200 Labomed đo
ở bước sóng 540 nm. Hàm lượng đường khử sinh ra trong quá trình thủy phân được
tính toán theo đường chuẩn glucose theo mục 2.3.3.1.
Hàm lượng đường khử = Hàm lượng đường sau thủy phân - Hàm lượng đường khử
trước thủy phân (μg/ml).
* Hàm lượng lignin trong nguyên liệu sau khi xử lý enzyme (% tính trên khối
lượng nguyên liệu khô).
Toàn bộ phần cặn không tan sau ly tâm được rửa nhiều lần bằng nước cất, sấy
khô đến trọng lượng không đổi ở nhiệt độ 60oC. Xác định hàm lượng lignin còn lại
trong nguyên liệu.
* Hàm lượng tinh dầu (% tinh dầu tính trên khối lượng nguyên liệu khô gió).
Mẫu thí nghiệm với 100 g nguyên liệu/mẻ trong cốc có mỏ 1000 ml, bổ sung
thêm 500 ml nước. Quá trình thủy phân diễn ra trong bể ổn nhiệt (Memmert- Đức),
khuấy đảo hỗn hợp bằng máy khuấy cơ (IKA- Đức), tốc độ 120 vòng/phút với các
thông số theo các phương án xử lý (Bảng 3.1).

57
Sau thủy phân, hỗn hợp nguyên liệu được đem đi chiết xuất tinh dầu bằng
phương pháp chưng cất lôi cuốn hơi nước trên bộ cất Clevenger [2]. Sự gia tăng hàm
lượng tinh dầu so với đối chứng không sử dụng enzyme được xác định theo công
thức:
Tỷ lệ gia tăng hàm lượng tinh dầu %= ℎ𝑒−ℎ0
ℎ0
Trong đó: he (%): Hàm lượng tinh dầu theo phương pháp enzyme hỗ trợ
h0 (%): Hàm lượng tinh dầu không sử dụng enzyme
3.1.2.2. Nghiên cứu thời gian cất tinh dầu
Hàm lượng tinh dầu của mẫu xử lý enzyme thu được sau khi chưng cất 2h, 3h,
4h... được xác định. Mẫu không enzyme cũng được tiến hành song song.
3.1.2.3. Nghiên cứu thành phần hóa học của tinh dầu
Thành phần hóa học của các mẫu tinh dầu tinh dầu khi xử lý bằng các phương
pháp khác nhau được xác định bằng phương pháp GC-MS trên máy sắc ký khí Agilent
HP mode 7890A gas chromatograph trang bị cột tách mao quản (60 m x 0.25 mm x
0.25 µm) Hewlett Packard HP5-MS (5% phenyl methyl siloxane), kết nối với máy đo
phổ khối Agilent 5973 mass spectrometer.
3.1.2.4. Nghiên cứu đánh giá hoạt tính sinh học của tinh dầu
Mẫu tinh dầu có sử dụng và không sử dụng enzyme được xác định một số hoạt
tính kháng vi sinh vật, hoạt tính kháng u, hoạt tính chống oxy hóa (tại phòng Sinh
học thực nghiệm, Viện Hóa học các HCTN), hoạt tính kháng viêm tại phòng Thử
nghiệm sinh học, Viện Công nghệ Sinh học.
* Xác định hoạt tính kháng vi sinh vật kiểm định
Trước khi tiến hành thí nghiệm, các chủng kiểm định được hoạt hóa và pha
loãng theo tiêu chuẩn McFarland 0,5 (khoảng 108 VSV/ml). Các phiến thí nghiệm
trong tủ ấm 37oC/24 giờ cho vi khuẩn và 30oC/48 giờ đối với nấm sợi và nấm men.
Sau đó đọc kết quả và tính giá trị ức chế tối thiểu MIC.
* Xác định khả năng gây độc tế bào
Sau khi tế bào được hoạt hóa phát triển đến pha log sẽ được sử dụng cho thử
test với các chất thử đã chuẩn bị sẵn ở các thang nồng độ khác nhau. Mỗi giếng thử

58
gồm tế bào, môi trường nuôi cấy và mẫu thử, được ủ trong tủ ấm CO2 ở 37oC/48
trong72h. Sau cố định tế bào và nhuộm bằng thuốc nhuộm SRB, đọc kết quả trên
máy đọc Elisa bước sóng 515 nm, từ đó xác định tỉ lệ CS (Cell survival, tế bào sống
sót). Giá trị IC50 được xác định bằng phần mềm TableCurve v4.0 (Systat Software
Inc.).
* Xác định khả năng ức chế sản sinh NO của tế bào macrophage RAW 264.7
Tế bào RAW 264.7 được ủ mẫu nghiên cứu ở các nồng độ khác nhau trong
2h trước khi được kích thích sản sinh yếu tố NO bằng LPS (1μg/mL) trong 24h.
Một số giếng không được ủ mẫu mà chỉ sử dụng dung dịch pha mẫu được coi là đối
chứng âm, chứng dương là cardamonin. Nitrite (NO2-), được xem là chỉ thị cho việc
tạo NO, sẽ được xác định nhờ bộ Griess Reagent System (Promega Cooperation,
WI, USA). Hàm lượng nitrite của từng mẫu thí nghiệm được xác định nhờ vào
đường cong hàm lượng chuẩn NaNO2 và được so sánh % với mẫu chứng âm (LPS).
* Xác định hoạt tính chống oxi hóa bằng DPPH
Hoạt tính chống oxy hóa của các dịch chiết được xác định bằng thử nghiệm
trên hệ DPPH (1,1-diphenyl-2-picrylhydrazyl), dựa trên nguyên tắc DPPH có khả
năng tạo ra các gốc tự do bền trong dung dịch ethanol bão hòa. Khả năng hấp thụ của
DPPH ở 515 nm được đo bằng máy đọc ELISA sau khi nhỏ DPPH vào dung dịch
mẫu thử trên phiến vi lượng 96 giếng. Kết quả các thử nghiệm được thể hiện là giá
trị trung bình của 3 phép thử lặp lại ± độ lệch chuẩn (p ≤ 0,05). Giá trị SC50
(Scavenging Concentration at 50% - nồng độ trung hòa được 50% gốc tự do của
DPPH) sẽ được xác định nhờ vào phần mềm máy tính TableCurve v4.0.
3.1.3. Nghiên cứu tìm điều kiện tối ưu cho quá trình ứng dụng hệ enzyme
kết hợp Laccase-Htec2 trong chưng cất tinh dầu từ cành lá và vỏ quế
3.1.3.1. Khảo sát một số yếu tố ảnh hưởng đến quá trình thủy phân
* Thí nghiệm khảo sát ảnh hưởng của pH
Thực hiện 07 thí nghiệm, mỗi mẫu 10g bột nguyên liệu cho vào bình tam giác
250 ml, bổ sung 50 ml nước cất và ngâm nước qua đêm. Giữ cố định các thông số
của quá trình như nhiệt độ 40oC, bổ sung enzyme theo Bảng 3.1, thay đổi các giá trị

59
pH từ 3,0-8,0 (bước nhảy 1) bằng axit HCl 0.01N và NaOH 0.01M. Chỉnh pH bằng
máy đo pH.
Sau 6 giờ thủy phân, tiến hành đo hàm lượng lượng đường khử bằng phương
pháp DNS. Hàm lượng đường sinh ra càng lớn, khả năng thủy phân càng hiệu quả,
lựa chọn pH 5 thích hợp cho các nghiên cứu tiếp theo.
* Thí nghiệm khảo sát ảnh hưởng của nhiệt độ
Thực hiện 07 thí nghiệm, mỗi mẫu thí nghiệm 10g bột nguyên liệu cho vào
bình tam giác 250 ml, bổ sung 50 ml nước cất và ngâm nước qua đêm. Giữ cố định
các thông số của quá trình như pH 5, bổ sung enzyme theo Bảng 3.1, thay đổi các giá
trị nhiệt độ từ từ 30- 60oC, bước nhảy 5oC.
Sau 6 giờ thủy phân, tiến hành đo hàm lượng lượng đường khử. Hàm lượng
đường sinh ra càng lớn, khả năng thủy phân càng hiệu quả, lựa chọn nhiệt độ 450C
thích hợp cho các nghiên cứu tiếp theo.
* Thí nghiệm khảo sát ảnh hưởng của tỷ lệ enzyme /cơ chất
Ảnh hưởng của tỷ lệ Laccase / cơ chất
Thực hiện 06 thí nghiệm, mỗi mẫu thí nghiệm được thực hiện với 10g bột
nguyên liệu cho vào bình tam giác 250 ml, bổ sung 50 ml nước cất và ngâm nước qua
đêm. Giữ cố định các thông số của quá trình như pH 5, nhiệt độ 45oC, bổ sung enzyme
Htec2 1%, thay đổi tỷ lệ enzyme laccase bổ sung: 0.1 - 0.7 ml/g cơ chất. Tương tự,
xác định hàm lượng đường khử sau 6 giờ thủy phân.
Ảnh hưởng của tỷ lệ Htec2 / cơ chất
Thực hiện 06 thí nghiệm, mỗi mẫu thí nghiệm được thực hiện với 10g bột
nguyên liệu cho vào bình tam giác 250 ml, bổ sung 50 ml nước cất và ngâm nước qua
đêm. Giữ cố định các thông số của quá trình như pH 5, nhiệt độ 45oC, bổ sung enzyme
Laccase phù hợp, thay đổi tỷ lệ enzyme Htec2 bổ sung: 0.5% - 1,75% (w/w), bước
nhảy 0.25%. Tương tự, xác định hàm lượng đường khử sau 6 giờ thủy phân, kết quả
lựa chọn tỷ lệ enzyme phù hợp.
* Thí nghiệm khảo sát ảnh hưởng của thời gian thủy phân

60
100 g bột nguyên liệu cho vào cốc có mỏ 1000 ml, bổ sung 500 ml nước cất
và ngâm nước qua đêm. Giữ cố định các thông số của quá trình như pH 5, nhiệt độ
45oC, bổ sung enzyme Laccase, Htec 2 phù hợp. Ở các thời điểm khác nhau của quá
trình thủy phân, hút 5 ml dịch thủy phân đem ly tâm và xác định hàm lượng đường
khử sinh ra. Lựa chọn khoảng thời gian phù hợp mà cho hàm lượng đường khử sinh
ra là lớn nhất.
3.1.3.2. Tối ưu hóa bằng qui hoạch hóa thực nghiệm
Sau khi khảo sát lựa chọn được vùng tối ưu ảnh hưởng đến mức độ thủy phân
cành lá quế bởi hệ enzyme Laccase-Htec2, để tìm điều kiện tối ưu chính xác trên cơ
sở mô hình tính toán, chúng tôi áp dụng phương pháp qui hoạch thực nghiệm. Các
bước thực hiện theo mục 2.3.7.2.
3.2. Nghiên cứu thăm dò ứng dụng hệ enzyme kết hợp Laccase-Htec2 trong
chưng cất tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna
3.2.1. Nghiên cứu phân tích thành phần chất nền của bột gỗ gió bầu A.
crassna
Thành phần chất nền của bột gỗ gió bầu A. crassna cũng được phân tích tương
tự như đối với mẫu cành lá quế và vỏ quế được trình bày ở mục 3.11.
3.2.2. Nghiên cứu thăm dò tác động của việc xử lý enzyme Laccase- Htec2
lên quá trình chưng cất tinh dầu từ bột gỗ gió bầu A. crassna
Hai thực nghiệm được tiến hành song song, trong đó bột gỗ gió bầu đều được
ngâm trong nước (tỉ lệ bột gỗ - nước là 1:7) trong 2 tuần. Tiếp theo, mẫu đối chứng
được chưng cất ngay sau khi ủ, mẫu nghiên cứu được xử lý với hệ enzyme Laccase-
Htec2 ở pH 5.2, nhiệt độ 440C, tốc độ khuấy 120 vòng/phút, tỷ lệ enzyme Laccase/S
0.7 ml/g, Htec2 / cơ chất 1.5% và thời gian thủy phân 8 giờ rồi đem cất lôi cuốn hơi
nước trong thiết bị Clevenger để thu lấy tinh dầu.
Hàm lượng tinh dầu sau chưng cất được xác định tương tự như đối với nguyên
liệu quế. Thành phần tinh dầu từ gỗ gió bầu A. crassna có và không có enzyme hỗ
trợ được xác định bằng GC-MS.
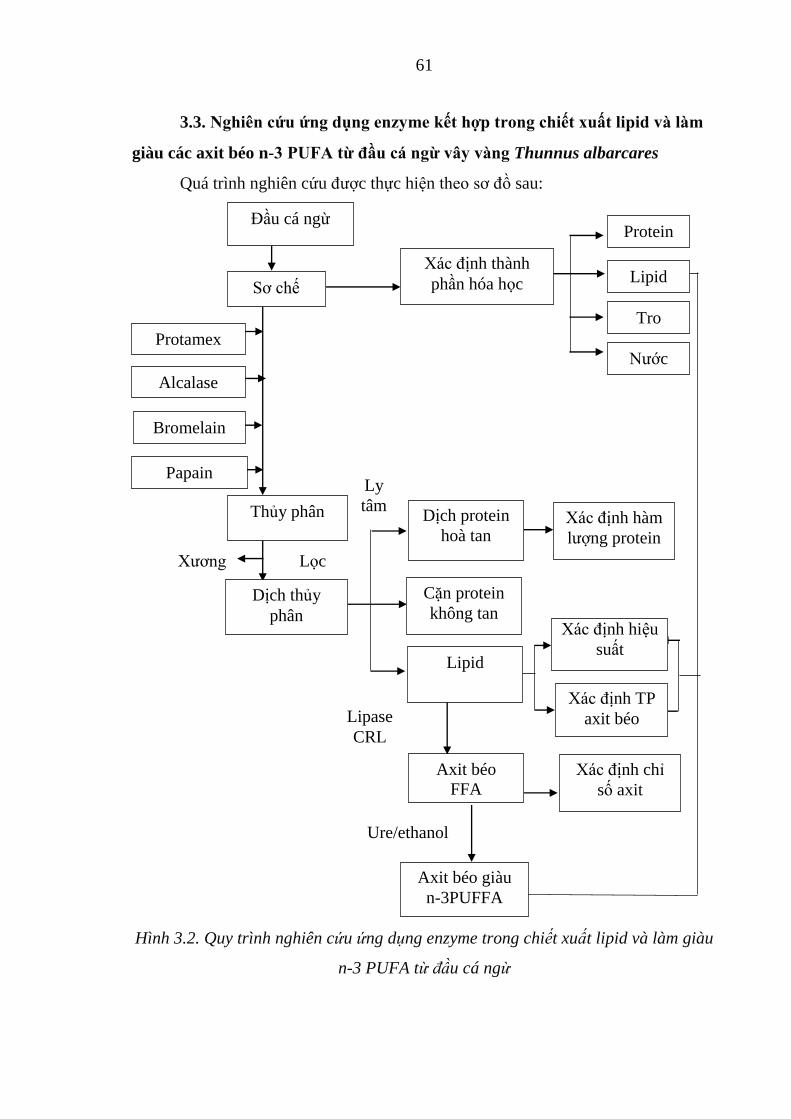
61
3.3. Nghiên cứu ứng dụng enzyme kết hợp trong chiết xuất lipid và làm
giàu các axit béo n-3 PUFA từ đầu cá ngừ vây vàng Thunnus albarcares
Quá trình nghiên cứu được thực hiện theo sơ đồ sau:
Hình 3.2. Quy trình nghiên cứu ứng dụng enzyme trong chiết xuất lipid và làm giàu
n-3 PUFA từ đầu cá ngừ
Đầu cá ngừ
Xác định thành
phần hóa học Sơ chế
Protamex
Thủy phân
Alcalase
Xác định hiệu
suất
Protein
Lipid
Tro
Bromelain
Xác định TP
axit béo
Lọc Xương
Dịch thủy
phân
Nước
Papain Ly
tâm
Lipid
Dịch protein
hoà tan
Cặn protein
không tan
Xác định hàm
lượng protein
Axit béo
FFA
Axit béo giàu
n-3PUFFA
Lipase
CRL
Xác định chỉ
số axit
Ure/ethanol

62
3.3.1. Nghiên cứu thành phần chất nền và lipid tổng của một số loại đầu cá
ngừ của Việt Nam
3.3.1.1. Nghiên cứu thành phần chất nền.
08 mẫu đầu cá ngừ thuộc 08 loài cá ngừ (đã bỏ mang, mắt) được rửa sạch, chặt
nhỏ và xay nhuyễn thành hỗn hợp đồng nhất. Sau đó phân tích hàm lượng nước, tro,
protein tổng, lipid tổng.
3.3.1.2. Nghiên cứu khảo sát thành phần axit béo
Thành phần axit béo của các mẫu lipid được xác định bằng GC-MS.
Hệ số đánh giá = % lipid tổng * % n-3 PUFA
3.3.2. Nghiên cứu ứng dụng enzyme protease trong chiết xuất lipid từ đầu
cá ngừ vây vàng T. albarcares
3.3.2.1. Mối tương quan giữa mức độ thủy phân và hiệu suất chiết xuất lipid
Mỗi mẫu thí nghiệm 100 g đầu cá ngừ đã xay nhỏ, bổ sung thêm nước, đánh
tan để đồng nhất mẫu. Quá trình thủy phân được thực hiện trong cốc có mỏ. Nâng
nhiệt độ lên 80-90oC trong 15 phút để diệt vi sinh vật. Sau đó, hạ nhiệt độ về nhiệt độ
thủy phân thì bổ sung enzyme. Các enzyme được sử dụng để đánh giá mức độ thủy
phân đầu cá ngừ bao gồm 02 enzyme có nguồn gốc từ vi sinh vật (Protamex và
Alcalase) và 02 enzyme có nguồn gốc từ thực vật (Bromelain và Papain). Một số yếu
tố ảnh hưởng đến quá trình thủy phân và chiết xuất lipid được khảo sát sơ bộ và chỉ
ra ở Bảng 3.2
Bảng 3.2. Một số thông số của quá trình thủy phân bởi enzyme protease
Thông số Protamex Alcalase Bromelain Papain
Tỷ lệ NL/nước 1:1-1:5
1:2
1:1-1:5
1:2
1:1-1:5
1:2
1:1-1:5
1:2
pH 4-7
(6.5)
5-9
(8.0)
4-7
(5.0)
4-7
(5.0)
Nhiệt độ (30-60)
45oC
(30-60)
50oC
(30-60)
50oC
(30-60)
50oC
Tốc độ khuấy 200 rpm 200 rpm 200 rpm 200 rpm
Tỷ lệ E/S 0.5-1.5%
(0.5)
0.5-1.5%
(0.5)
0.5-1.5%
(1%)
0.5-1.5%
(1%)
Thời gian thủy phân 6h 6h 6h 6h

63
Sau quá trình thủy phân, enzyme được bất hoạt bằng cách nâng nhiệt độ lên
85-90oC trong 15 phút trong bể ổn nhiệt. Sau đó, hỗn hợp được lọc qua rây để tách
riêng phần xương và dịch lọc. Phần dịch lọc được đem ly tâm lạnh 4oC ở tốc độ 10.
000 vòng/phút trong 30 phút. Sau khi ly tâm thu được 3 phần: phần dầu, phần protein
hòa tan và cặn protein (Hình 3.3).
Hình 3.3. 3 pha sản phẩm sau ly tâm
Để đánh giá hiệu quả của quá trình, 2 hàm mục tiêu được xác định là hàm
lượng dầu giải phóng (thu được ở pha dầu) và hàm lượng protein hòa tan.
- Phần dầu được rửa bằng n-hexan hoặc nước muối 0.9% ở 35-40oC làm khan
bằng Na2SO4 sau đó cô quay loại dung môi.
Hàm lượng lipid (%) = Khối lượng lipid
x 100 Khối lượng nguyên liệu
Hiệu suất chiết xuất lipid (%) = Hàm lượng lipid
x 100% Hàm lượng lipid tổng
- Phần dịch thủy phân hòa tan: lượng protein hòa tan thu hồi được tính toán
dựa trên khối lượng protein hòa tan (theo phương pháp Bradford) so với khối lượng
protein trong nguyên liệu ban đầu (theo phương pháp Kjeldahl)
% thu hồi protein hòa tan = Khối lượng protein hòa tan
x 100% Khối lượng protein trong nguyên liệu
Lipid
Protein hòa
tan
Cặn protein
không tan

64
3.3.2.2. Nghiên cứu so sánh thành phần axit béo
Các mẫu lipid chiết xuất được bằng các enzyme khác nhau được phân tích
bằng GC-MS và so sánh với kết quả phân tích mẫu thu được bằng phương pháp Blight
& Dyer. Kết quả lựa chọn enzyme thích hợp dựa trên hệ số đánh giá:
Hệ số đánh giá = % lipid x % n-3 PUFA
3.3.3. Nghiên cứu ứng dụng enzyme lipase kết hợp ure trong quá trình làm
giàu các axit béo n-3 PUFA của dầu đầu cá ngừ vây vàng T. albacares
3.3.3.1. Nghiên cứu thủy phân lipid bằng enzyme lipase CRL
- Trước quá trình thủy phân, lipid tách từ đầu cá ngừ bằng phương pháp
enzyme được xác định một số chỉ số axit, xà phòng hóa. Kết quả thu được dầu từ đầu
cá ngừ có chỉ số axit 0.43 mg KOH/g, chỉ số xà phòng hóa 198.45 (mg KOH/g).
- Phản ứng thủy phân được thực hiện trong hệ dung môi hai pha với tỷ lệ pha
nước - pha hữu cơ 1:1 (v/v), tỉ lệ dầu - dung môi 3:7 (w/v). Pha nước chuẩn bị với
đệm ở các dải pH từ 5-9. Phản ứng được ủ ở dải nhiệt độ 20-50oC và luôn được khuấy
trộn (200 vòng/phút) để đạt được sự phân tán đồng nhất của các pha. Ở các khoảng
thời gian xác định (0h, 2h, 4h...) mẫu được tách ra bằng ly tâm. Theo Fernádez
Lorente G (2010) [88], 95% axit béo tự do có mặt trong pha dung môi. Vì vậy, chỉ số
axit được xác định sau khi loại toàn bộ dung môi.
Hiệu suất thủy phân = Chỉ số axit (dầu thủy phân) - Chỉ số axit (dầu chưa thủy phân)
x 100% Chỉ số xà phòng hóa - Chỉ số axit (dầu chưa thủy phân)
- Sau quá trình thủy phân, hỗn hợp axit béo tự do (FFA) được loại dung môi
và thực hiện kết tinh bằng ure như mô tả ở mục 2.3.6.
3.3.3.2. Khảo sát một số yếu tố ảnh hưởng đến quá trình thủy phân
Quá trình thủy phân dầu đầu cá ngừ thành các axit béo tự do bởi enzyme lipase
từ C. rugosa được khảo nghiệm và các điều kiện pH, nhiệt độ, thời gian cho các thử
nghiệm xác định yếu tố cụ thể được đo trực tiếp sau khi vô hoạt enzyme với 3 ml
ethanol 99,5%.
Phản ứng thủy phân lipase là phản ứng bậc 1. Các yếu tố được khảo sát là đơn
biến, khảo sát yếu tố nào thì thay đổi yếu tố đó trong khoảng thời gian nhất định (6
giờ) và giữ nguyên các yếu tố khác. Các yếu tố pH (5,0-9,0) bước nhảy 0.5, nhiệt độ
(20- 50oC) bước nhảy 5oC, tỷ lệ enzyme/cơ chất (0.2-1,2%) bước nhảy 0.2.

65
Kết quả khảo sát được đánh giá dựa trên độ tăng chỉ số axit, hiệu suất thủy phân
dầu được tính toán. Các kết quả tính toán thực nghiệm được so sánh trên phần mềm xử
lý xây dựng phương trình hồi quy theo phương pháp bình phương tối thiểu [6].
3.3.3.3. Kết tinh FFA bằng ure/ethanol
Hỗn hợp axit béo sau thủy phân được bổ sung ure: cồn = 3:16, FFA/ure = 1:3.
Sau đó, dung dịch thu được để kết tinh ở nhiệt độ 4oC trong thời gian là 18-20 giờ.
Sau quá trình kết tinh, hỗn hợp pha rắn và pha lỏng được lọc qua giấy lọc tại nhiệt độ
kết tinh, rửa lại bằng dung môi có nhiệt độ kết tinh, sau đó tách pha rắn và pha lỏng.
Khối lượng hai pha thu được xác định sau khi làm bay hơi dung môi.
Thành phần axit béo của dầu sau kết tinh được phân tích trên máy GC-MS.

66
CHƯƠNG 4. KÊT QUẢ VÀ THẢO LUẬN
4.1. Kết quả nghiên cứu ứng dụng enzyme kết hợp trong chưng cất tinh
dầu từ các bộ phận khác nhau của cây quế Cinnamomum cassia
Trong thực tiễn, tinh dầu quế ở Việt Nam được chiết xuất chủ yếu từ cành và lá
cây quế Cinnamomum cassia bằng chưng cất lôi cuốn hơi nước. Chỉ có một lượng nhỏ
tinh dầu được chưng cất từ vỏ cây vì nó có giá trị thương mại cao hơn rất nhiều. Trong
luận án này chúng tôi nghiên cứu tác động của việc tiền xử lý bằng enzyme tới hiệu
quả chưng cất tinh dầu của cả hai loại nguyên liệu này. Các mẫu nghiên cứu là cành +
lá và vỏ quế C. cassia thu từ huyện Văn Yên, tỉnh Yên Bái, một trong những huyện có
truyền thống trồng quế và có sản lượng quế lớn nhất nước ta.
4.1.1. Kết quả phân tích các chất nền và tinh dầu của cành lá và vỏ quế C. cassia
4.1.1.1. Hàm lượng các chất nền của cành lá và vỏ quế C. cassia
Kết quả phân tích các thành phần chất nền của hai loại nguyên liệu nêu trên
được trình bày trên Bảng 4.1.
Bảng 4.1. Kết quả phân tích chất nền của nguyên liệu chưng cất tinh dầu quế
(hàm lượng chất tính theo % khối lượng khô tuyệt đối)
TT Chỉ tiêu phân tích Đơn vị Cành lá quế Vỏ thân quế
1 Cellulose % 24,74 ± 0,10 20,36 ± 0,10
2 Hemicellulose % 25,50 ± 0,10 14,02 ± 0,10
3 Lignin % 16,28 ± 0,20 25,78 ± 0,20
4 Các hợp chất trích ly (cồn) % 21,02 ± 0,10 29,55 ± 0,10
Kết quả trên Bảng 4.1 cho thấy mẫu cành, lá và mẫu vỏ quế mang đặc trưng
của nhóm cây thân gỗ với thành phần chủ yếu là lignocellulose, chiếm hơn 60% khối
lượng nguyên liệu khô. Hàm lượng các thành phần cấu trúc gồm cellulose,
hemicellulose và lignin xác định được ở mẫu cành, lá có các giá trị tương ứng là
24,74%, 25,50% và 16,28%; ở mẫu vỏ quế là 20,36%, 14,02% và 25,78%.
Tỷ lệ tương quan giữa 3 thành phần trong nguyên liệu cho thấy, cành lá quế
có tổng cellulose + hemicellulose lớn hơn vỏ quế (50,24%, so với 34,38%), còn vỏ
quế có nhiều lignin hơn cành lá quế (25,78% so với 16,28%). Kumar (2009) cho rằng
lignin tồn tại chủ yếu ở những mô hóa gỗ như vỏ cây, thân cây, rơm rạ mà tồn tại ít

67
trong lá [110]. Đặc điểm cấu trúc này cho thấy cấu trúc thành tế bào vỏ quế bền vững
hơn so với cành lá quế và cả hai loại mẫu nguyên liệu nghiên cứu đều là những nguyên
liệu khó bị phân giải bởi enzyme hơn các bộ phận khác của cây như lá, hoa, hay quả.
4.1.1.2. Hàm lượng và thành phần hóa học của tinh dầu từ cành lá và vỏ quế
C. cassia
Theo phương pháp truyền thống, các nguyên liệu được ngâm ủ tự nhiên với nước
trước khi chưng cất lấy tinh dầu. Cành lá quế được ủ trong 24 h (tỉ lệ nguyên liệu / nước
là 1:5). Vỏ quế được ủ trong 72 h (tỉ lệ nguyên liệu / nước là 1:7). Hàm lượng tinh dầu
được định lượng bằng phương pháp cất lôi cuốn hơi nước trong thiết bị Clevenger. Hàm
lượng tinh dầu cành lá quế, vỏ quế thu được bằng chưng cất hơi nước tương ứng là 0,69
± 0,02% , 2,87 ± 0,04% tinh dầu (tính theo nguyên liệu khô gió).
Tác giả Nguyễn Thị Tâm & cs. (1997) công bố hàm lượng tinh dầu của lá quế
C. cassia trồng tại Yên Bái dao động trong khoảng 0,14 - 1,04% và của vỏ quế trong
khoảng 1 - 3%. Huang & cs. (2014) công bố hàm lượng tinh dầu vỏ cành quế C.
cassia trồng tại Trung Quốc là 2,76 - 3,11% [100]. Điều đó cho thấy, mẫu nghiên cứu
đạt chất lượng để cho các thực nghiệm tiếp theo.
Thành phần hóa học của các mẫu tinh dầu được xác định bằng phương pháp
GC-MS. Các kết quả được tổng hợp ở Bảng 4.2. Các chất được nhận dạng dựa trên
sự phù hợp của phổ khối và chỉ số thời gian lưu (RI, chỉ số Kovats) của chúng với
các dữ kiện trong ngân hàng dữ liệu của máy.
Bảng 4.2. Kết quả phân tích thành phần hóa học của tinh dầu cành lá và tinh dầu
vỏ quế C. cassia (hàm lượng chất được tính theo % diện tích của mỗi peak)
TT Chất RI CTPT % trong
TD cành, lá
% trong
TD vỏ
1 Benzaldehyde 968 C7H6O 1,62 0,83
2 Salicylaldehyde 1053 C7H6O2 0,45
3 Benzenepropanal 1172 C9H12 1,59 2,39
4 Benzofuran <2-methyl> 1189 C8H6O 5,90
5 Cinnamaldehyde <Z-> 1230 C9H8O 0,75 0,64
6 Cinnamaldehyde <E-> 1289 C9H8O 67,74 53,99

68
TT Chất RI CTPT % trong
TD cành, lá
% trong
TD vỏ
7 Cinnamyl alcohol <E-> 1317 C9H10O 0,57 0,15
8 Copaene <a-> 1391 C15H24 0,68 13,86
9 Elemene <cis-b-> 1405 C15H24 0,29
10 Sativene 1409 C15H24 0,13
11 Caryophylene <b-> 1439 C15H24 0,26 0,37
12 Bergamotene <a-trans-> 1447 C15H24 0,16
13 Cinnamyl acetate 1455 C11H12O2 17,20 0,27
14 Coumarin 1457 C9H6O2 0,88 0,30
15 Humulene <a-> 1473 C15H24 0,55
16 Caryophyllene 1480 C15H24 0,13
17 Cadina-1(6),4-diene <trans-> 1489 C15H24 0,47
18 Muurolene <g-> 1491 C15H24 0,29 1,40
19 Amorphene <a-> 1495 C15H24 0,20
20 Selinene <b-> 1506 C15H24 0,18
21 Amorphene <g-> 1513 C15H24 1,17
22 Muurolene<a-> 1515 C15H24 3,20
23 Bisabolene <b> 1518 C15H24 0,27
24 Curcumene <b-> 1521 C15H22 0,18
25 Cadinene <g-> 1531 C15H24 0,17 0,26
26 Cadinene <d-> 1537 C15H24 0,56 7,69
27 Calamenene <cis-> 1540 C15H22 1,17
28 Zonarene 1542 C15H24 1,09
29 Bisabolene <E-g-> 1542 C15H24 0,19
30 Methoxycinnamaldehyde <(E)-o> 1543 C10H10O2 1,09
31 Cadina-1,4-diene <trans-> 1549 C15H24 2,29
32 Himachalene <a-dehydro-ar-> 1554 C15H24 0,4
33 Calacorene <a-> 1561 C15H20 0,54
34 Nerolidol <E-> 1569 C15H26O 0,31

69
TT Chất RI CTPT % trong
TD cành, lá
% trong
TD vỏ
35 Axenol (Gleenol) 1600 C15H26O 0,21
36 Caryophyllene oxide 1605 C15H24O 0,12
37 Tetradecanal 1614 C14H28O 0,16
38 Cubenol <1,10-di-epi-> 1646 C15H26O 0,91
39 Muurolol <epi-a-> (=T-Muurolol) 1659 C15H26O 1,33
40 Eudesmol <7-epi-a> 1662 C15H24O 0,68
41 Cadinol <a-> 1672 C15H26O 0,11
42 Bisabolol <a-> 1696 C15H26O 0,18
Tổng 99,41 99,11
Tổng các hydrocarbon 3,87 38,26
Tổng các HC chứa oxy 95,54 60,85
Nhóm Phenylpropanoid 88,73 58,83
trans- Cinnamaldehyde 67,74 53,99
Nhóm Sesquiterpenoid 2,42 40,38
Sesquiterpen hydrocarbon 1,99 35,87
Sesquiterpen chứa oxy 0,43 4,51
Hình 4.1a và 4.1b trình bày cấu trúc của một số phenylpropanoid và
sesquiterpenoid quan trọng trong các mẫu tinh dầu nêu trên Bảng 4.2.
Benzenepropanal
(C9H10O)
trans-Cinnamaldehyde
(C9H8O)
cis-Cinnamaldehyde
(C9H8O)
Cinnamyl alcohol
(C9H10O)
Coumarin
(C9H6O2)
o-Methoxy-
cinamaldehyde
Hình 4.1a. Một số thành phần thuộc nhóm phenylpropanoid

70
Copaene
(C15H24)
Muurolene
(C15H24)
Calamenene
(C15H22)
Viridiflorene
(C15H24)
Cadinene
(C15H24)
Bisabolene
( C15H24)
Nerolidol
(C15H26O)
Caryophyllene oxide
( C15H24O)
Bisabolol
(C15H26O)
Hinh 4.1b. Một số thành phần thuộc nhóm sesquiterpenoid
Kết quả GC- MS phân tích thành phần hóa học của tinh dầu cành lá quế và
tinh dầu vỏ quế C. cassia cho thấy:
- Khoảng 28 hợp chất được tìm thấy trong tinh dầu cành lá quế C. cassia, trong
đó trans-Cinnamaldehyde là hợp chất chính (69,74%), theo sau là Cinnamyl acetate
(17,2%), 2-Methyl-benzofuran (5,9%), Benzaldehyde (1,62%), Benzenepropanal
(1,59%), Ngoài ra, một số các thành phần khác cũng được xác định với hàm lượng
nhỏ hơn 1%.
- Khoảng 38 hợp chất được phát hiện trong tinh dầu vỏ quế C. cassia, trong
đó trans-Cinnamaldehyde là hợp chất chính (53,99%), theo sau là Copaene (13,86%),
Cadinene (7,69%), Benzenepropanal (2,39%). Ngoài ra, một số các thành phần khác
cũng được xác định với hàm lượng nhỏ hơn 1%.
- Nhóm phenylpropanoid (C6-C3) là nhóm chất chủ yếu trong cả tinh dầu cành
lá quế (71,53%) và vỏ quế (57,47%), trong đó trans-Cinnamaldehyde đều là thành
phần chính với hàm lượng tương ứng là 68,49% và 53,99%.

71
- Tinh dầu cành lá quế có tổng các hợp chất chứa oxy cao hơn tinh dầu vỏ quế
(95,54% so với 60,85%). Hàm lượng Cinnamyl acetate (17,2%) của nó cao hơn hẳn
tinh dầu vỏ quế (0,27%).
- Tinh dầu vỏ quế có tổng các sesquiterpenoid cao hơn hẳn tinh dầu cành lá
quế (40,38% so với 2,71%), gồm các sesquiterpen hydrocacbon là chủ yếu. Một thành
phần đáng chú ý của tinh dầu vỏ quế là o-Methoxy-cinnamaldehyde (1,09%).
Tóm lại, phenylpropanoid là nhóm hợp chất chủ yếu trong tinh dầu cành lá
quế và vỏ quế C. cassia thu từ Văn Yên, Yên Bái, trong đó trans-Cinnamaldehyde là
thành phần chính. Tổng của các hợp chất hydrocarbon, của các hợp chất chứa oxy,
của các phenylpropanoid, và của các sesquiterpenoid có sự khác nhau rõ rệt giữa
nguyên liệu cành lá quế và vỏ quế.
4.1.2. Tác động của việc xử lý nguyên liệu với enzyme lên quá trình chưng
cất tinh dầu từ cành lá quế C. cassia
Bốn dãy thí nghiệm sau đây đã được thực hiện để khảo sát tác động của việc
xử lý nguyên liệu chưng cất với enzyme lên quá trình chưng cất tinh dầu từ cành lá
quế C. cassia, đồng thời để chọn ra phương thức xử lý nguyên liệu thích hợp nhất:
- Dãy thí nghiệm 1: Nguyên liệu sau khi ngâm ủ được đem đi chưng cất lôi
cuốn hơi nước trực tiếp để chiết xuất lấy tinh dầu, không qua xử lý với enzyme.
- Dãy thí nghiệm 2: Nguyên liệu sau khi ngâm ủ được xử lý với enzyme
Laccase (là enzyme thô phân giải lignin thu được từ canh trường nuôi cấy nấm
Ganoderma lucidum) trước khi đem đi chưng cất.
- Dãy thí nghiệm 3: Nguyên liệu sau khi ngâm ủ được xử lý với enzyme Htec2
(là hỗn hợp đa enzyme bao gồm cellulase và xylanase) trước khi đem đi chưng cất.
- Dãy thí nghiệm 4: Nguyên liệu sau khi ngâm ủ được xử lý với hệ enzyme kết
hợp gồm Laccase và Htec2 trước khi đem đi chưng cất.
Các thông số sau đây được xác định trong mỗi thực nghiệm:
- Hàm lượng đường khử trong nguyên liệu sau khi xử lý enzyme (µg/ml).
- Hàm lượng lignin trong nguyên liệu sau khi xử lý enzyme (% tính trên khối
lượng nguyên liệu khô tuyệt đối).
- Hàm lượng tinh dầu (% tinh dầu tính trên khối lượng nguyên liệu khô gió).

72
- Thành phần hóa học của các tinh dầu thu được.
- Hoạt tính sinh học của các tinh dầu thu được.
Hai thông số đầu dùng để đánh giá tác động của enzyme lên mức độ thủy phân
thành tế bào của nguyên liệu. Hàm lượng tinh dầu dùng để đánh giá hiệu quả chiết
xuất. Thành phần hóa học và hoạt tính sinh học dùng để đánh giá tính chất và chất
lượng của các tinh dầu thu được.
4.1.2.1. Tác động của việc xử lý nguyên liệu với enzyme lên hiệu suất thu hồi
tinh dầu và mức độ thủy phân các chất nền của cành lá quế
Các kết quả thu được về tác động của việc xử lý nguyên liệu với enzyme lên
hiệu suất thu hồi tinh dầu và mức độ thủy phân các chất nền của nguyên liệu được
trình bày trên Bảng 4.3 và biểu diễn trên Hình 4.2.
Bảng 4.3. Kết quả khảo sát tác động của việc xử lý nguyên liệu với enzyme lên mức
độ thủy phân các chất nền và hiệu suất chưng cất tinh dầu từ cành lá quế
Enzyme Đường khử
(µg/ml)
Lignin
(%)
Tinh dầu
(%)
Gia tăng tinh
dầu (%)
Đối chứng 0 17,43 ± 0,24 0,69 ± 0,07 0
Laccase thô 3,12 ± 0,2 14,21 ± 0,21 0,74 ± 0,03 7,29
Htec2 18,13 ± 0,21 15,32 ± 0,15 0,80 ± 0,04 16,88
Laccase thô - Htec2 78,14 ± 0,13 13,01 ± 0,24 0,98 ± 0,04 41,70
Hình 4.2. Đồ thị biểu diễn mối tương quan giữa mức độ thủy phân các chất nền và
hiệu suất chưng cất tinh dầu từ cành lá quế
0
10
20
30
40
50
0
5
10
15
20
25
30
Đối chứng Laccase Htec2 Laccase + Htec2 Hàm
lượ
ng
đư
ờn
g kh
ử (
μg/
ml)
Tỷ lệ
gia
tăn
g ti
nh
dầu
,h
àm lư
ợn
g lig
nin
(%
)
Phương án xử lý enzyme
Cành lá quế Tinh dầu LigninGlucose

73
Các kết quả nêu trên cho thấy quá trình tiền xử lý cành lá quế bằng enzyme đã
làm tăng đáng kể hiệu suất thu hồi tinh dầu (tăng 31,5% so với mẫu đối chứng không
xử lý enzyme). Sự gia tăng của hiệu suất tinh dầu có tương quan tỉ lệ thuận với sự gia
tăng của hàm lượng đường khử (0 so với 58.34 μg/ml) trong dung dịch phản ứng cũng
như sự giảm thiểu của hàm lượng lignin trong nguyên liệu (17.43 so với 13.01%) là
các bằng chứng cụ thể cho tác động của hoạt động enzyme đến sự gia tăng này.
Các kết quả nêu trên cũng chỉ ra rằng, việc sử dụng kết hợp enzyme Laccase
và Htec2 cho hiệu quả cao hơn khi sử dụng riêng rẽ từng enzyme. Điều đó gợi ý cho
ta về khả năng có tác động hiệp đồng của các enzyme nghiên cứu.
4.1.2.2. Tác động của việc xử lý nguyên liệu với enzyme lên thời gian chưng
cất tinh dầu cành lá quế
Tác động của việc xử lý nguyên liệu với enzyme lên thời gian chưng cất tinh
dầu từ cành lá quế C. cassia được khảo sát bằng cách theo dõi hàm lượng tinh dầu
thu được theo thời gian chưng cất mẫu nguyên liệu được xử lý với hệ enzyme kết hợp
Laccase-Htec (Mẫu enzyme kết hợp). Để so sánh, một thí nghiệm tương tự được tiến
hành với nguyên liệu chưng cất không qua xử lý enzyme (Mẫu cất truyền thống). Kết
quả thu được được trình bày trên Bảng 4.4 và Hình 4.3.
Bảng 4.4. Hàm lượng tinh dầu thu nhận theo thời gian chưng cất mẫu cành lá quế
có và không có enzyme hỗ trợ
Thời gian (giờ)
Phương pháp
Hàm lượng tinh dầu (%)
2 3 4 5 6 7 8
Enzyme kết hợp 0,41 0,73 0,87 0,98 0,97 0,96 0,96
Cất truyền thống 0,06 0,28 0,47 0,60 0,69 0,67 0,69

74
Hình 4.3. Đồ thị biểu diễn hàm lượng tinh dầu theo thời gian chưng cất mẫu cành
lá quế có và không có enzyme hỗ trợ
Đối với mẫu cành lá quế đã xử lý enzyme, sau 2 h đầu đã thu được hơn 50%
lượng tinh dầu, sau 5 h lượng tinh dầu đạt đến cực đại. Trong khi đó, đối với phương
pháp không sử dụng enzyme sau 2 h đầu mẫu đối chứng chỉ đạt 9 - 10%, sau 8 h
lượng tinh dầu mới đạt đến cực đại. Quá trình xử lý cành lá quế bằng hỗn hợp enzyme
Laccase và Htec2 đã làm giảm đáng kể thời gian cất tinh dầu so với đối chứng không
sử dụng enzyme.
Có thể giải thích rằng, quá trình thủy phân bởi hệ enzyme Laccase-Htec2 đã
làm phá vỡ một phần cấu trúc thành tế bào cành lá quế, mối liên kết cellulose-
hemicellulose-lignin trong hệ lignocellulose được nới lỏng, tạo ra các lỗ hổng trên
thành tế bào, làm tăng tính thấm và các hợp chất dễ bay hơi như tinh dầu dễ dàng giải
thoát ra khỏi tế bào khi có tác dụng nhiệt.
4.1.2.3. Tác động của việc xử lý nguyên liệu với enzyme lên thành phần hóa
học của tinh dầu cành lá quế
Thành phần hóa học của các tinh dầu thu được từ cành lá quế C. cassia theo 4
phương án như đã nêu ở trên được xác định bằng phương pháp GC-MS. Các kết quả
thu được được tổng hợp trên Bảng 4.5. Các sắc ký đồ tương ứng được trình bày trên
các Hình 4.4a, b, c, và d.
0
0.2
0.4
0.6
0.8
1
1.2
0 1 2 3 4 5 6 7 8
Hàm
lượ
ng
tin
h d
ầu (
%)
Thời gian chưng cất (giờ)
Tinh dầu EADTinh dầu HD

75
Hình 4.4a. Sắc ký đồ của tinh dầu cành lá quế chưng cất theo cách thông thường, nguyên liệu không qua xử lý enzyme
1: Benzaldehyde (1,62%), 2: Benzenepropanal (1.59%), 3: 2-Methyl-benzofuran (5.9%), 4: trans-cinamaldehyde (69.74%), 5: Cinnamyl
acetate (17.2%).
1 2
4
3
5

76
Hình 4.4b. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với enzyme Laccase EAD
1: Benzaldehyde (1,93%), 2: Benzenepropanal (1,64%), 3: 2-Methyl-benzofuran (4,52%), 4: trans-cinamaldehyde (68,96%), 5: Cinnamyl
acetate (15,58%).
1 2
4
3
5

77
Hình 4.4c. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với enzyme Htec2 EAD
1: Benzaldehyde (1,77%), 2: Benzenepropanal (1,14%), 3: 2-Methyl-benzofuran (1,64%), 4: trans-cinamaldehyde (70,54%), 5: Cinnamyl
acetate (21,02%).
1 2
4
3
5

78
Hình 4.4d. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với hệ enzyme Laccase-Htec2
1: Benzaldehyde (0,69%), 2: Benzenepropanal (0,78%), 3: 2-Methyl-benzofuran (2,44%), 4: trans-cinamaldehyde (85,6%), 5: Cinnamyl
acetate (1,34%).
1 2
4
3 5

79
Bảng 4.5. Kết quả phân tích GC-MS các thành phần hóa học của tinh dầu thu được
từ cành lá quế có và không có xử lý với enzyme
(hàm lượng chất được tính theo % diện tích của mỗi peak)
TT Chất
Hàm lượng (%)
Không
enzyme Laccase Htec2
Laccase+
Htec2
1 Benzaldehyde 1,62 1,93 1,77 0,69
2 Salicylaldehyde 0,45 0,51 0,87 0,61
3 Unknown 0,64
4 Benzenepropanal 1,59 1,64 1,14 0,78
5 Benzofuran <2-Methyl> 5,9 4,52 1,64 2,44
6 cis-Cinnamaldehyde 0,75 0,78 0,4 1,23
7 Phenylethyl acetate 0,11
8 trans-Cinnamaldehyde 69,74 68,96 70,54 85,6
9 Unknown 0,16
10 Cinnamyl alcohol 0,57 0,58 0,17 0,29
11 α-Copaene 0,68 0,8 0,32 0,66
12 Caryophylene <b-> 0,26 0,3 0,26
13 Cinnamyl acetate 17,2 15,58 21,02 1,34
14 Coumarin 0,88 0,84 0,65 1,78
15 Caryophyllene <9-epi-(E)> 0,13 0,17 0,26
16 γ-Muurolene 0,29 0,35 0,17 0,47
17 Viridiflorene - 0,28 0,16
18 α-Muurolene - 0,18 0,2
19 β-Bisabolene 0,11
20 γ-Cadinene 0,17 0,21 0,3
21 δ-Cadinene 0,56 0,73 0,12 0,63
22 cis-Calamenene - 0,11 0,21
23 Methoxycinnamaldehyde 0,23
24 γ-Bisabolene 0,19 0,23
25 Nerolidol 0,31 0,34 0,22 0,37

80
26 Caryophyllene oxide 0,12 0,13 0,14
27 Bisabolol 0,13
Tổng 99,41 99,42 99,14 99,81
Tổng các HC hydrocarbon 3,58 5,13 1,75 4,04
Tổng các HC chứa oxy 95,54 94,17 97,39 94,85
Nhóm Phenylpropanoid 88,73 88,38 94,03 91,25
trans-Cinnamaldehyde 69,74 68,96 70,54 85,6
Nhóm Sesquiterpen 2,42 3,83 0,83 3,9
Sesquiterpen hydrocarbon 1,99 3,36 0,61 3,26
Sesquiterpen chứa oxy 0,43 0,47 0,22 0,64
So sánh kết quả phân tích GC-MS của cả 4 phương án xử lý cho thấy, quá
trình tiền xử lý cành lá quế bằng enzyme đã không làm thay đổi bản chất của tinh
dầu, các mẫu tinh dầu đều có thành phần chính là cinnamaldehyde và có cùng số
lượng các thành phần quan trọng khác.
Ít có sự biến động về tổng các hydrocarbon và tổng các hợp chất chứa oxy.
Thành phần tinh dầu cành lá quế ở cả 4 phương án xử lý đều chứa chủ yếu là các hợp
chất có oxy (94,17 - 97,39%), còn lại là các hợp chất hydrocarbon (1,75 - 5,13%).
Nhóm phenylpropanoid (C6-C3) vẫn là nhóm chất chính trong cả 4 phương án
xử lý, trong đó trans-cinnamaldehyde là thành phần chính có hàm lượng đạt từ
68,49% đến 86,83%.
Tinh dầu thu được bằng cách xử lý nguyên liệu với các enzyme Laccase và
HTec2 riêng rẽ không gây ra biến động lớn về thành phần hóa học, nhưng việc xử lý
nguyên liệu với hệ enzyme kết hợp Laccase-Htec2 đã làm thay đổi rõ rệt nhất thành
phần % của một số chất quan trọng trong tinh dầu. Cụ thể như sau:
- Làm giàu một số chất mong muốn trong tinh dầu quế C. cassia như: hàm
lượng cinnamaldehyde tăng từ 69,74% lên 85,60%, o-methoxy-cinnamaldehyde từ
0% lên 0,23%.
- Làm giảm hàm lượng một số chất khác, đặc biệt cinnamyl acetat giảm từ
17,2% xuống 1,34%, cinnamyl alcohol giảm từ 0,57% xuống 0,29%. Sự tăng và giảm
của cinnamaldehyde và cinnamyl acetat có thể có mối liên hệ mật thiết với nhau.

81
Quá trình thủy phân bằng hệ enzyme kết hợp Laccase-Htec2 cho hiệu suất và
chất lượng tinh dầu cao hơn so với phương pháp thông thường hoặc xử lý bằng
enzyme riêng rẽ. Điều này cho thấy việc sử dụng hỗn hợp các hệ enzyme có thể có
các tác động hiệp đồng giữa các enzyme riêng rẽ. Chính vì vậy, trong các nghiên cứu
tiếp theo, chúng tôi đã nghiên cứu chọn các điều kiện tối ưu cho quá trình này.
4.1.2.4. Tác động của việc xử lý nguyên liệu với enzyme lên hoạt tính sinh học
của tinh dầu cành lá quế
Để khảo sát tác động của việc xử lý nguyên liệu cành lá quế với enzyme lên
hoạt tính sinh học của sản phẩm thu được, chúng tôi đã sử dụng 2 mẫu sau:
- Mẫu 1 (ký hiệu là tinh dầu HD) là tinh dầu thu được bằng chưng cất lôi cuốn
hơi nước cành lá quế không được tiền xử lý với enzyme.
- Mẫu 2 (ký hiệu là tinh dầu EAD) là tinh dầu thu được bằng chưng cất lôi
cuốn hơi nước cành lá quế được tiền xử lý với hệ enzyme kết hợp Laccase-Htec2.
Kết quả xác định hoạt tính kháng vi sinh vật kiểm định
Hai mẫu tinh dầu cành lá quế C. cassia được thử nghiệm hoạt tính với 8 chủng
vi sinh vật kiểm định. Kết quả được trình bày tại Bảng 4.6a.
Bảng 4.6a. Kết quả thử hoạt tính kháng vi sinh vật kiểm định
Ký hiệu mẫu
Nồng độ ức chế tối thiểu (MIC, g/ml)
Vi khuẩn
Gr(-)
Vi khuẩn
Gr(+) Nấm mốc Nấm men
E.
coli
P.
aer
ug
ino
sa
B.
sub
tili
s
S. a
ure
us
A.
nig
er
F. o
xysp
oru
m
S. ce
revi
sia
e
C. a
lbic
an
s
Chứng dương 5,50 11,00 7,10 14,38 23,13 11,56 5,78 11,56
Tinh dầu HD 400 (-) 400 100 200 (-) 400 400
Tinh dầu EAD 400 (-) 400 100 200 (-) 400 400
Ghi chú: Chứng dương là Streptomycin cho vi khuẩn Gr(+), Tetracylin cho vi
khuẩn Gr(-), Nystatin cho nấm sợi và nấm men.
Kết quả cho thấy với nồng độ thử nghiệm ban đầu 400 µg/ml, hai mẫu tinh
dầu HD và EAD đều có hoạt tính kháng 6/8 chủng vi sinh vật kiểm định. Ở các nồng

82
độ pha loãng tiếp theo, hai mẫu tinh dầu đều biểu hiện hoạt tính kháng S. aureus với
MIC = 100 μg/ml, và kháng A. niger với MIC = 200 μg/ml. Huang và cs. (2014) cho
rằng mức độ kháng khuẩn của tinh dầu từ Cinnamomum cassia trồng tại Trung Quốc
có khả năng kháng mạnh các vi khuẩn Gram dương S. aureus với MIC 2.5 mg/ml, B.
subtilis 5.0 mg/ml và E. coli 10 mg/ml. Các nghiên cứu chỉ ra rằng hoạt tính kháng
vi sinh vật phổ rộng của tinh dầu quế chủ yếu là do thành phần cinamaldehyde. Tuy
nhiên, trong thử nghiệm này sự chênh lệch về hàm lượng cinamaldehyde của 2 mẫu
tinh dầu đã không thấy có sự biến động về hoạt tính sinh học.
Kết quả xác định hoạt tính gây độc tế bào
Hoạt tính gây độc tế bào của hai mẫu tinh dầu HD và EAD được thử nghiệm
trên 03 dòng tế bào ung thư người, bao gồm dòng Hep-G2 (Human hepatocellular
carcinoma - Ung thư gan), dòng LU-1 (Human lung adenocarcinoma - Ung thư biểu
mô phổi) và dòng RD (Human rhabdomyosarcoma - Ung thư mô liên kết) với chất
đối chứng dương là ellipticine pha trong DMSO. Kết quả được chỉ ra ở Bảng 4.6b.
Bảng 4.6b. Kết quả thử hoạt tính gây độc tế bào trên 03 dòng tế bào ung thư
Ký hiệu mẫu
Nồng
độ đầu
(g/ml)
Tỉ lệ CS (%)
trên các dòng tế bào IC50 (g/ml)
Hep-G2 LU-1 RD Hep-
G2 LU-1 RD
Dung môi (DMSO) 100,00 100,00 100,00 - - -
Chứng dương 5 1,340,8 3,660,9 3,871,1 0,29 0,31 0,33
Tinh dầu HD 40 3,620,7 65,601,4 14,811,5 29,25 >40 6,01
Tinh dầu EAD 40 3,780,3 32,671,7 6,10,5 12,89 25,95 2,34
Ghi chú: Chứng dương là chất chuẩn ellipticine, pha trong dung môi DMSO.
Kết quả thử nghiệm ở Bảng 4.6b cho thấy sản phẩm tinh dầu thu được bằng 2
phương pháp chiết xuất, có và không có xử lý enzyme, đều có hoạt tính gây độc tế
bào với nồng độ thử nghiệm ban đầu (40 g/ml). Ở các nồng độ thử nghiệm tiếp theo,
tinh dầu HD biểu hiện hoạt tính gây độc tế bào với 02 dòng tế bào ung thư gan và ung
thư biểu mô liên kết (Hep-G2 và RD) với giá trị IC50 lần lượt là 29,25 và 6,01 g/ml,

83
trong khi tinh dầu EAD biểu hiện hoạt tính gây độc tế bào đối với 3 dòng tế bào ung
thư (Hep-G2, LU-1 và RD) với giá trị IC50 lần lượt là 12,89, 25,95 và 2,34 g/ml.
Đặc biệt, tinh dầu EAD có hoạt tính khá mạnh trên dòng ung thư biểu mô phổi với
giá trị IC50 là 2,34 g/ml.
Nhiều các công trình công bố cao chiết từ vỏ quế đã cho thấy khả năng ức chế
sự gia tăng của một số dòng thế bào ung thư ở người [96] đồng thời ít có tác dụng
gây độc tế bào trên các tế bào bình thường [169]. Tác giả Huang T.C. &cs., (2007)
cho thấy thành phần cinnamaldehyde trong tinh dầu lá quế Cinnamomum
osmophloeum đã tác động gây độc tế bào thông qua sự phá vỡ màng ty thể, kích thích
sản sinh ROS, suy giảm glutathione và kích hoạt caspase trên dòng tế bào ung thư
bạch cầu người K562 [99]. Có thể sự gia tăng của thành phần chất chính trong tinh
dầu lá quế cinnamaldehyde từ 69,74 lên 85,60% khi sử dụng enzyme Laccase+Htec2
hỗ trợ có thể dẫn đến sự gia tăng hoạt tính gây độc tế bào của tinh dầu EAD trên 03
dòng tế bào ung thư thử nghiệm đều cao hơn so với tinh dầu HD.
Kết quả xác định hoạt tính kháng viêm
Phép thử hoạt tính kháng viêm của 2 mẫu tinh dầu HD và EAD được đánh giá
thông qua sự ức chế sinh NO bởi LPS trên đại thực bào macrophage RAW 264.7.
Bảng 4.6c. Kết quả thử khả năng ức chế sự sinh NO trên tế bào RAW 264.7
Nồng độ
(µg/ml)
% ức chế NO Nồng độ
(µg/ml)
% ức chế NO
Tinh dầu HD Tinh dầu EAD Cardamonin
100 53,2 0,5 65,8 0,4 10 102,7 0,5
50 74,8 0,5 82,0 0,3 2 84,2 0,3
25 85,6 0,6 87,4 0,4 0,4 41,4 0,4
12,5 55,0 0,8 85,6 0,5 0,08 20,2 0,3
6,25 29,7 0,7 58,6 0,8 - -
IC50 11,0 ± 0,8 - IC50 0,5 ± 0,1
Kết quả trên Bảng 4.6c chỉ ra rằng, sau 24 h cả hai mẫu tinh dầu đều thể hiện
hoạt tính ức chế đại thực bào sản sinh NO ở các khoảng nồng độ từ 6,25 đến 100

84
μg/ml (P < 0.05 với LPS đối chứng âm), tinh dầu EAD có biểu hiện hoạt tính kháng
viêm mạnh hơn tinh dầu HD. Nguyên nhân có sự chênh lệch đó có thể được giải thích
là do sự biến động của một số thành phần chính trong tinh dầu. Kết quả khảo sát một
số thành phần của lá quế lên khả năng ức chế NO của tác giả Lee H.S. (2009) cho
thấy Cinnamaldehyde, Cinnamyl aclcohol có khả năng ức chế sản sinh NO trong khi
rất ít hoặc không có hiệu lực ức chế được quan sát thấy ở một số thành phần khác như
Cadinene, Cinnamic acid, α-Copaene, Eugenol, Limonene. Đối chiếu với bảng 4.5
kết quả phân tích thành phần hóa học lá quế cho thấy trong tinh dầu EAD có sự gia
tăng của hàm lượng Cinnamaldehyde từ 69,74 lên 85,60%, nhưng Cinnamyl alcohol
giảm từ 0,57 xuống 0,29. Chính vì vậy, độ chênh lệch giữa hàm lượng
Cinnamaldehyde trong tinh dầu EAD đã làm gia tăng hoạt tính ức chế sản sinh NO
so với tinh dầu HD.
Tuy nhiên, tương quan giữa nồng độ và hoạt tính tỏ ra không tuyến tính, đặc
biệt ở vùng 25 đến 100 μg/ml, trong khi chất đối chứng dương là cardamonin hoạt
động ổn định trong thí nghiệm này. Chính vì vậy, cần có những nghiên cứu bổ sung
nhằm làm sáng tỏ kết quả nghiên cứu trên.
Kết quả xác định hoạt tính chống oxi hóa
Kết quả xác định hoạt tính chống oxi hóa trên hệ DPPH của các mẫu thử được
trình bày ở Bảng 4.6d.
Bảng 4.6d. Kết quả thử hoạt tính chống oxy hóa trên hệ DPPH
Kí hiệu mẫu Nồng độ (g/ml) Tỉ lệ trung hòa
gốc tự do (%) SC50 (µg/ml)
Chứng (+) 25 87,22 0,53 11,31
Chứng (-) - 0,0
Tinh dầu HD 100 10,34 0,7 > 100
Tinh dầu EAD 100 4,33 0,5 > 100
Ghi chú: Chứng dương là chất chuẩn ascorbic acid.
Kết quả được nêu trong Bảng 4.6d cho thấy, hai mẫu tinh dầu thử nghiệm chưa
thể hiện khả năng trung hòa gốc tự do của DPPH ở nồng độ nghiên cứu.
Nhận xét chung:
Các kết quả thử nghiệm nêu trên cho thấy, hai mẫu tinh dầu có gam hoạt tính

85
sinh học tương tự như nhau. Việc xử lý cành lá quế C. cassia bằng hệ enzyme kết
hợp Laccase-Htec2 không làm thay đổi đáng kể hoạt tính sinh học của sản phẩm tinh
dầu thu được. Sự gia tăng quan sát thấy được của hoạt tính kháng viêm và gây độc tế
bào của mẫu EAD so với mẫu Tinh dầu HD có thể liên quan đến hàm lượng
cinnamaldehyde cao hơn của nó.
Mặt khác, các kết quả thử nghiệm này một lần nữa chứng minh cho khả năng
sử dụng tinh dầu quế C. cassia nói chung như là một hoạt chất tự nhiên để tăng cường
khả năng kháng khuẩn, kháng nấm, kháng viêm và chống u.
4.1.3. Tác động của việc xử lý nguyên liệu với enzyme lên quá trình chưng
cất tinh dầu từ vỏ quế C. cassia
4.1.3.1. Tác động của việc xử lý nguyên liệu với enzyme lên hiệu suất thu hồi
tinh dầu và mức độ thủy phân các chất nền của vỏ quế
Các mẫu vỏ quế trước khi chưng cất tinh dầu được ngâm ủ với nước trong 72
h. Trong phương án 01 (đối chứng), nguyên liệu sau khi ngâm ủ được đem đi cất trực
tiếp để lấy tinh dầu. Trong phương án 02, 03, và 04, nguyên liệu sau khi ngâm ủ được
xử lý với các enzyme tương ứng là Laccase, Htec2, cũng như với hệ enzyme kết hợp
Laccase-Htec trước khi cất. Mức độ thủy phân thành tế bào được xác định cùng với
hiệu suất thu nhận tinh dầu. Kết quả chỉ ra ở Bảng 4.7 và Hình 4.5.
Bảng 4.7. Kết quả khảo sát tác động của việc xử lý nguyên liệu với enzyme lên mức
độ thủy phân thành tế bào và hiệu suất chưng cất tinh dầu từ vỏ quế
STT Enzyme Đường khử
(µg/ml)
Lignin
(%)
Tinh dầu
(%)
Gia tăng
tinh dầu
(%)
1 Đối chứng 0 25,78 ± 0,19 2,87 ± 0,05 -
2 Laccase thô 5,43 ± 0,18 22,54 ± 0,20 2,99 ± 0,04 4,35
3 Htec2 10,23 ± 0,24 24,12 ± 0,18 3,03 ± 0,04 5,55
4 Laccase+Htec2 38,34 ± 0,22 20,34 ± 0,22 3,28 ± 0,06 14,57

86
Hình 4.5. Đồ thị biểu diễn mối tương quan giữa mức độ thủy phân thành tế bào và
tỷ lệ gia tăng tinh dầu vỏ quế
Cũng tương tự như cành lá quế, kết quả trên Bảng 4.7 và Hình 4.5 cho thấy
mối tương quan rõ rệt giữa mức độ thủy phân thành tế bào và tỷ lệ gia tăng tinh dầu
vỏ quế C. cassia. Lần lượt từ phương án 01 đến 04, hàm lượng đường khử gia tăng
từ 0 lên đến 38,34 µg/ml, còn hàm lượng lignin giảm từ 25,78 xuống còn 20,34%.
Phương án 04, kết hợp 2 enzyme Laccase và Htec2 cho hiệu quả thủy phân cao nhất
với hàm lượng đường khử là 38,34 µg/ml và tỷ lệ gia tăng hiệu suất tinh dầu đạt
14,57%.
So sánh kết quả giữa hai nguyên liệu cành lá quế và vỏ quế cho thấy:
- Việc xử lý nguyên liệu chưng cất với enzyme riêng rẽ và với hệ enzyme kết
hợp Laccase-Htec2 đều làm gia tăng hiệu suất thu hồi tinh dầu so với phương án đối
chứng không sử dụng enzyme. Mức độ gia tăng hàm lượng tinh dầu tỷ lệ thuận với
mức độ thủy phân nguyên liệu bởi enzyme, trong đó hàm lượng đường khử tăng và
hàm lượng lignin trong nguyên liệu giảm.
- Các phương án xử lý cho thấy hệ enzyme kết hợp Laccase+Htec2 cho tác
động hiệp đồng trong quá trình thủy phân. Sử dụng riêng rẽ Laccase và Htec2 chưa
đem lại hiệu quả mong muốn.
- Quá trình xử lý cành lá quế bằng hệ enzyme Laccase+Htec2 cho mức độ thủy
phân cao hơn so với vỏ quế. Điều này phù hợp với các kết quả phân tích thành phần
0
10
20
30
40
50
0
5
10
15
20
25
30
ĐC Laccase thô Htec2 Laccase thô + Htec2
Hàm
lượ
ng
đư
ờn
g kh
ử (
µg/
ml)
Tỷ lệ
gia
tăn
g ti
nh
dầu
, lig
nin
(%
)Vỏ quế
Gia tăng tinh dầu Lignin Đường khử

87
chất nền của nguyên liệu (Bảng 4.1), trong đó nguyên liệu cành lá quế chứa chủ yếu
là cellulose và hemicellose, hàm lượng lignin thấp, khả năng tác dụng enzyme dễ
dàng hơn so với vỏ quế.
4.1.3.2. Tác động của việc xử lý nguyên liệu với enzyme lên thời gian chưng
cất tinh dầu vỏ quế
Tác động của việc xử lý nguyên liệu với enzyme lên thời gian chưng cất tinh
dầu từ vỏ quế C. cassia được khảo sát bằng cách theo dõi lượng tinh dầu thu được
theo thời gian chưng cất mẫu nguyên liệu được xử lý với hệ enzyme kết hợp Laccase-
Htec2 (Mẫu enzyme kết hợp). Để so sánh, một thí nghiệm tương tự được tiến hành
với nguyên liệu chưng cất không qua xử lý enzyme (Mẫu cất truyền thống). Kết quả
thu được được trình bày trên Bảng 4.8 và Hình 4.6.
Bảng 4.8. Hàm lượng tinh dầu theo thời gian chưng cất mẫu vỏ quế có và không có
enzyme hỗ trợ
Thời gian (h)
Phương pháp
Hàm lượng tinh dầu (%)
2 3 4 5 6 7 8 9
Enzyme kết hợp 0,88 1,68 2,72 3,25 3,28 3,27 3,25 3,23
Cất truyền thống 0,59 0,90 1,07 1,74 2,14 2,57 2,87 2,85
Hình 4.6. Đồ thị biểu diễn hàm lượng tinh dầu theo thời gian chưng cất mẫu vỏ quế
có và không có enzyme hỗ trợ
0
0.5
1
1.5
2
2.5
3
3.5
0 1 2 3 4 5 6 7 8 9 10
Hàm
lượ
ng
tin
h d
ầu (
%)
Thời gian chưng cất (giờ)
Vỏ quế Tinh dầu EADTinh dầu HD

88
Cũng tương tự như cành lá quế, kết quả khảo sát trên vỏ quế cho thấy hệ
enzyme Laccase + Htec2 có vai trò tích cực trong việc giảm thời gian chưng cất tinh
dầu quế. Tại các thời điểm khác nhau của quá trình chưng cất: 2h, 3h, 4h... lượng tinh
dầu thu từ mẫu vỏ quế xử lý bằng enzyme đều cao hơn so với không xử lý enzyme.
Sau 6h chưng cất, lượng tinh dầu đạt cực đại (3,28%) trong khi đó sau 8h mẫu không
xử lý enzyme mới đạt cực đại (2,87%).
4.1.3.3. Kết quả đánh giá tác động của các hệ enzyme lên thành phần hóa học
tinh dầu vỏ quế
Thành phần hóa học của các tinh dầu thu được từ vỏ quế C. cassia theo 4
phương án như đã nêu ở trên được xác định bằng phương pháp GC-MS. Các kết quả
thu được được tổng hợp trên Bảng 4.9. Các sắc ký đồ tương ứng được trình bày trên
các Hình 4.7a, b, c, và d.

89
Hình 4.7a. Sắc ký đồ tinh dầu vỏ quế, nguyên liệu không xử lý enzyme
1: Benzenpropanal (2,39%), 2: trans-Cinnamaldehyde (53,99%), 3: Copaene (13,86%), 4: Muurolene (3,2%), 5: Cadinene 7,69%, 6:
Calamenene (1,17%), 7: Cadina-1,4-diene (2,29%)
1
2
5
3
6
7
4

90
Hình 4.7b. Sắc ký đồ tinh dầu cành lá quế, nguyên liệu qua xử lý với hệ enzyme Laccase- EAD
1: Benzenpropanal (1,88%), 2: trans-Cinnamaldehyde (59,22%), 3: Copaene (5,78), 4: Muurolene (3,64%), 5: Cadinene (6,41%), 6:
Calamenene (2,82%), 7: Cadina-1,4-diene (1,23%),
1
2
4
3 5
6 7

91
Hình 4.7c. Sắc ký đồ tinh dầu vỏ quế, nguyên liệu qua xử lý với hệ enzyme Htec2-EAD
1: Benzenpropanal (2,01%), 2: trans- Cinnamaldehyde (58,36%), 3: Copaene (5,91%), 4: Muurolene (3,83%), 5: Cadinene (6,56%), 6:
Calamenene (2,6%), 7: Cadina-1,4-diene (1,23%)
1
2
3
4
5
6
7

92
Hình 4.7d. Sắc ký đồ tinh dầu vỏ quế, nguyên liệu qua xử lý với hệ enzyme Laccase - Htec2- EAD
1: Benzenpropanal (2,18%), 2: trans-Cinnamaldehyde (67,6%), 3: Copaene (11,72%), 4: Muurolene (2,14%), 5: Cadinene (4,76%), 6:
Calamenene (1,05%), 7: Cadina-1,4-diene (1,17%).
1
2
3
4
5
6
7

93
Bảng 4.9. Kết quả phân tích GC-MS các thành phần hóa học của tinh dầu thu được
từ vỏ quế có và không có xử lý với enzyme
(hàm lượng chất được tính theo % diện tích của mỗi peak)
TT Chất
Hàm lượng (%)
Không
enzyme Laccase Htec2 Lac+Htec2
1 Benzaldehyde 0,83 0,74 0,7 0,86
2 Benzenepropanal 2,39 1,88 2,01 2,18
3 Cinnamaldehyde <Z-> 0,64 0,51 0,51 0,77
4 Cinnamaldehyde <E-> 53,99 59,22 58,36 67,6
5 Cinnamyl alcohol <E-> 0,15 0,12
6 Copaene <a-> 13,86 5,78 5,91 11,72
7 Elemene <cis-b-> 0,29 0,16 0,17 0,23
8 Sativene 0,13 0,27 0,21
9 Caryophylene <E-> 0,37 0,21 0,23 0,27
10 Bergamotene <a-trans-> 0,16 0,12
11 Cinnamyl acetate <E-> 0,27 0,19 0,15 0,31
12 Coumarin 0,30 1,16 1,21
13 Humulene <a-> 0,55 0,32 0,33 0,41
14 Cadina-1(6),4-diene <trans-> 0,47 0,22 0,3 0,17
15 Muurolene <g-> 1,40 1,16 1,19 0,81
16 Amorphene <a-> 0,20 0,18 0,20
17 Selinene<b-> 0,18 0,13 0,13
18 Amorphene <g-> 1,17
19 Muurolene <a-> 3,20 3,64 3,83 2,14
20 Bisabolene <b-> 0,27 0,21 0,21 0,20
21 Curcumene <b-> 0,18 - -
22 Cadinene <g-> 0,26 0,28 0,27 0,18
23 Cadinene <d-> 7,69 6,41 6,56 4,76
24 Calamenene <cis-> 1,17 2,82 2,60 1,05

94
TT Chất
Hàm lượng (%)
Không
enzyme Laccase Htec2 Lac+Htec2
25 Zonarene 1,09
26 Methoxycinnamaldehyde <(E)-o-> 1,09 3,01 3,49
27 Cadina-1,4-diene <trans-> 2,29 1,09 1,23 1,17
28 Himachalene <a-dehydro-ar-> 0,40 0,67 1,37
29 Calacorene <a-> 0,54 0,86 0,91 0,27
30 Axenol (Gleenol) 0,21 0,51 0,46 0,19
31 Tetradecanal 0,16 0,30 0,36
32 Cubenol <1,10-di-epi-> 0,91 1,58 1,32 0,85
33 Muurolol <epi-a-> (=T-Muurolol) 1,33 2,2 1,96 0,90
34 Eudesmol <7-epi-a> 0,68
35 Cadinol <epi-a-.> 1,24 1,12 0,42
36 Cadinol <a-> 0,11 0,40 0,27
37 Cadalene 0,23 0,21
38 Bisabolol <a-> 0,18 0,41 0,44
Tổng 99,11 97,99 96,64 99,28
Tổng hydrocarbon 38,26 26,52 26,29 27,26
Tổng các HC chứa oxy 60,85 71,47 70,35 72,02
Tổng Phenylpropanoid 58,83 65,97 65,73 70,98
trans-cinnamaldehyde 53,99 59,22 58,36 67,6
Tổng sesquiterpenoid 39,29 30,98 29,85 27,44
Sesquiterpen hydrocarbon 35,87 24,64 24,28 25,08
Sesquiterpen chứa oxy 3,42 6,34 5,57 2,36
So sánh thành phần GC-MS của cả 4 phương án xử lý cho thấy quá trình tiền
xử lý vỏ quế bằng enzyme đã không làm thay đổi bản chất của tinh dầu quế, các mẫu
nghiên cứu đều có thành phần chính là cinnamaldehyde và có cùng số lượng các thành
phần quan trọng khác.

95
Ít có sự biến động về tổng các hydrocarbon và tổng các hợp chất chứa oxy,
thành phần tinh dầu vỏ quế ở cả 4 phương án xử lý đều chứa chủ yếu là các hợp chất
có oxy (60 - 72%), các hợp chất hydrocarbon (28 - 36%).
Phenylpropanoid vẫn là nhóm chất chính trong cả 4 phương án xử lý và
Cinnamaldehyde là thành phần chính với hàm lượng từ 54,63% đến 68,37%. Cũng
tương tự như tinh dầu cành lá quế, tinh dầu thu được nhờ xử lý với các enzyme
Laccase và Htec2 riêng rẽ ít có sự biến động về thành phần hóa học so với hệ enzyme
Laccase-Htec2. Việc xử lý nguyên liệu với hệ enzyme kết hợp Laccase-Htec2 làm
gia tăng hàm lượng cinnamaldehyde từ 54,63% lên 68,37%.
Tuy nhiên, so sánh sự biến động thành phần hóa học của hai tinh dầu cành lá
quế và vỏ quế cho thấy tinh dầu cành lá quế có sự biến động mạnh mẽ hơn. Có thể
mức độ thủy phân nguyên liệu của cành lá quế bởi hệ enzyme kết hợp Laccase-Htec2
cao hơn so với vỏ quế dẫn đến hiệu quả khác biệt.
4.1.4. Kết quả tìm điều kiện tối ưu cho quá trình ứng dụng hệ enzyme kết
hợp Laccase-Htec2 trong chưng cất tinh dầu từ cành lá và vỏ quế
Các kết quả nghiên cứu ở các phần trên cho thấy việc sử dụng hệ enzyme kết hợp
Laccase-Htec để xử lý cành lá và vỏ quế C. cassia trước khi chưng cất có tác động rất
tốt đến hiệu suất thu hồi, chất lượng và thời gian chưng cất tinh dầu. Vì thế, mục tiêu của
nội dung nghiên cứu này là tìm ra các điều kiện tối ưu cho quá trình này.
Các kết quả thử nghiệm trình bày ở Bảng 4.2 (trang 63) cho thấy sự gia tăng
hàm lượng tinh dầu tỷ lệ thuận với mức độ thủy phân nguyên liệu (đánh giá thông
qua sự gia tăng hàm lượng đường khử và sự sụt giảm lignin). Vì thế trong nội dung
nghiên cứu này chúng tôi tiến hành khảo sát các yếu tố ảnh hưởng để xác định điều
kiện tối ưu cho quá trình thủy phân các nguyên liệu chưng cất tinh dầu quế. Chúng
tôi lựa chọn hàm lượng đường khử sinh ra sau khi xử lý nguyên liệu với enzyme làm
chỉ tiêu đánh giá hiệu quả thủy phân vì nó được xác định bằng phương pháp so màu
của Miller với thuốc thử DNS. Phương pháp này cho kết quả nhanh, dễ thao tác và
dễ kiểm soát.

96
4.1.4.1. Kết quả nghiên cứu các yếu tố ảnh hưởng đến quá trình thủy phân các
nguyên liệu chưng cất quế với hệ enzyme kết hợp Laccase-Htec2
Kết quả khảo sát ảnh hưởng của pH
Chúng tôi thực hiện 7 thí nghiệm khảo sát trong vùng pH từ 3 - 8 sử dụng hệ
đệm thích hợp, giữ cố định các điều kiện khác như tỷ lệ nguyên liệu/nước 1:5, nhiệt
độ 45oC, thời gian thủy phân 6 h. Các kết quả thu được thể hiện trong Hình 4.8a.
Hình 4.8a. Đồ thị biểu diễn ảnh hưởng của của pH lên hàm lượng đường khử
Độ pH của môi trường phản ứng là một trong những yếu tố quan trọng ảnh
hưởng đến hoạt động của enzyme trong quá trình thủy phân, mỗi enzyme có một
khoảng hoạt động tối thích riêng. Enzyme Laccase hoạt động tốt nhất trong giải pH
3,5 - 4,5, trong khi enzyme Htec2 hoạt động tốt nhất trong khoảng pH 4,5 - 5,5. Tuy
nhiên khi phối hợp hai enzyme Laccase và Htec2, kết quả khảo sát cho thấy pH 4,5
- 5 cho khả năng thủy phân cao nhất, vì thế được lựa chọn cho các thử nghiệm tiếp
theo.
Kết quả khảo sát ảnh hưởng của nhiệt độ
Kết quả khảo sát ảnh hưởng của nhiệt độ đến quá trình thủy phân thể hiện
trong Hình 4.8b cho thấy, nhiệt độ càng tăng hiệu quả thủy phân càng cao và đạt cực
đại ở 45oC, nếu tiếp tục tăng nhiệt độ đến 50 - 55oC thì hiệu quả thủy phân có xu
hướng giảm.
0
10
20
30
40
50
60
70
3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0 7,5 8,0
Hàm
lượ
ng
đư
ờn
g kh
ử (
μg/m
l)
pH
Cành lá quế Vỏ quế

97
Hình 4.8b. Đồ thị biểu diễn ảnh hưởng của của nhiệt độ lên hàm lượng đường khử
Enzyme laccase từ G. luccidum có khoảng nhiệt độ hoạt động tối thích ở 35 -
45oC, thấp hơn so với enzyme Htec2 (ở 45 - 55oC). Trong các nghiên cứu trước đây
enzyme Htec2 thường được chúng tôi sử dụng ở nhiệt độ 50oC để thủy phân cellulose
trong rơm rạ và rong biển. Tuy nhiên, khi kết hợp Htec2 với enzyme Laccase, nhiệt độ
50oC không phát huy cao nhất khả năng hoạt động của enzyme này. Các nỗ lực thử
nghiệm riêng rẽ cho lần lượt các enzyme hoạt động ở nhiệt độ tối ưu cũng không đem
lại hiệu quả hơn phương án bổ sung enzyme đồng thời. Bên cạnh đó, nguyên liệu xử
lý là các nguyên liệu chứa tinh dầu, là những hợp chất dễ bay hơi ở nhiệt độ cao (>
50oC). Chính vì vậy, chúng tôi lựa chọn nhiệt độ 45oC để thủy phân cành lá quế, vỏ
quế vừa đảm bảo quá trình thủy phân hiệu quả, vừa hạn chế sự tác động của nhiệt độ
lên chất lượng tinh dầu sau khi chưng cất.
Kết quả khảo sát ảnh hưởng của tỷ lệ enzyme/cơ chất
Ảnh hưởng của tỷ lệ enzyme Laccase/cơ chất và Htec2/cơ chất được thực hiện
theo mô tả ở phần thực nghiệm 3.5.2. Kết quả được chỉ ra ở Hình 4.8c và 4.8d.
Hình 4.8c. Đồ thị biểu diễn ảnh hưởng của tỷ lệ Laccase/cơ chất lên hàm lượng đường khử
0
10
20
30
40
50
60
70
30,0 35,5 40,0 45,0 50,0 55,0 60,0
Hàm
lư
ợng đ
ườ
ng k
hử
(μ
g/m
l)
Nhiệt độ (oC)
Cành lá quế (LQ)
Vỏ quế (VQ)Cành lá quế Vỏ quế
0
20
40
60
80
0,2 0,3 0,4 0,5 0,6 0,7 0,8 1,0
Hàm
lượ
ng
đư
ờn
g (μ
g/m
l)
Tỷ lệ E laccase/S (ml/g)
Cành lá quế Vỏ quế
Cành lá quế Vỏ quế
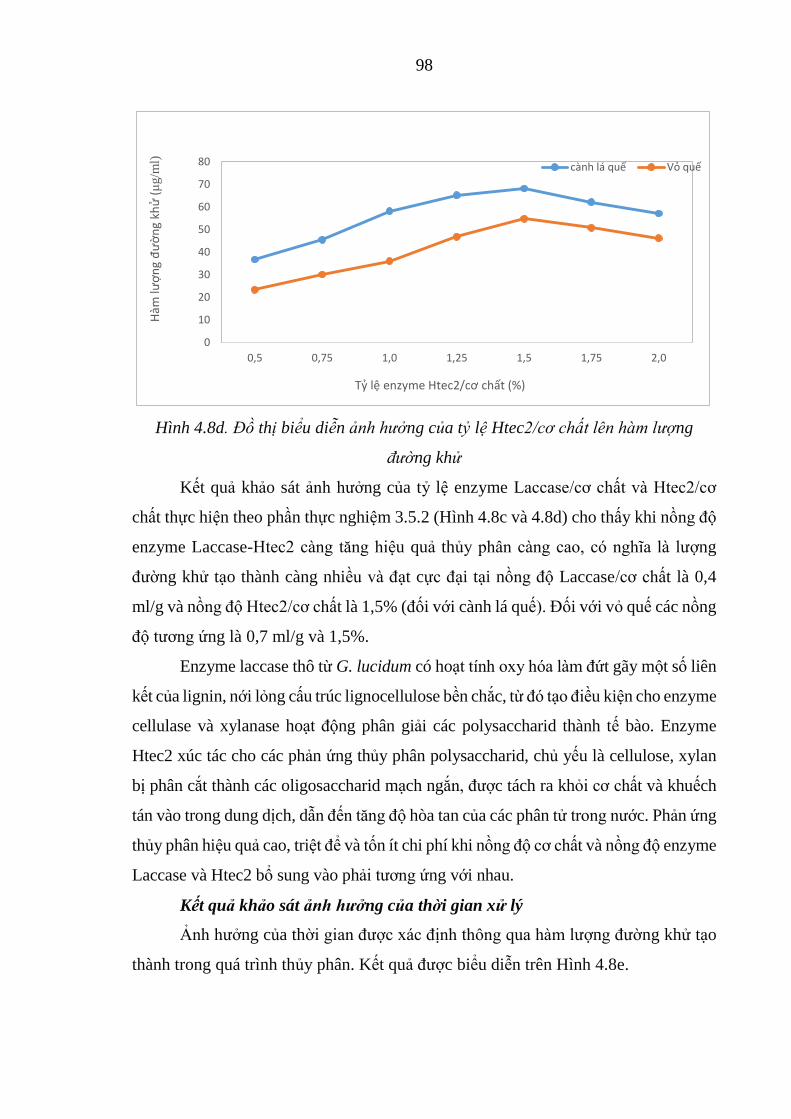
98
Hình 4.8d. Đồ thị biểu diễn ảnh hưởng của tỷ lệ Htec2/cơ chất lên hàm lượng
đường khử
Kết quả khảo sát ảnh hưởng của tỷ lệ enzyme Laccase/cơ chất và Htec2/cơ
chất thực hiện theo phần thực nghiệm 3.5.2 (Hình 4.8c và 4.8d) cho thấy khi nồng độ
enzyme Laccase-Htec2 càng tăng hiệu quả thủy phân càng cao, có nghĩa là lượng
đường khử tạo thành càng nhiều và đạt cực đại tại nồng độ Laccase/cơ chất là 0,4
ml/g và nồng độ Htec2/cơ chất là 1,5% (đối với cành lá quế). Đối với vỏ quế các nồng
độ tương ứng là 0,7 ml/g và 1,5%.
Enzyme laccase thô từ G. lucidum có hoạt tính oxy hóa làm đứt gãy một số liên
kết của lignin, nới lỏng cấu trúc lignocellulose bền chắc, từ đó tạo điều kiện cho enzyme
cellulase và xylanase hoạt động phân giải các polysaccharid thành tế bào. Enzyme
Htec2 xúc tác cho các phản ứng thủy phân polysaccharid, chủ yếu là cellulose, xylan
bị phân cắt thành các oligosaccharid mạch ngắn, được tách ra khỏi cơ chất và khuếch
tán vào trong dung dịch, dẫn đến tăng độ hòa tan của các phân tử trong nước. Phản ứng
thủy phân hiệu quả cao, triệt để và tốn ít chi phí khi nồng độ cơ chất và nồng độ enzyme
Laccase và Htec2 bổ sung vào phải tương ứng với nhau.
Kết quả khảo sát ảnh hưởng của thời gian xử lý
Ảnh hưởng của thời gian được xác định thông qua hàm lượng đường khử tạo
thành trong quá trình thủy phân. Kết quả được biểu diễn trên Hình 4.8e.
0
10
20
30
40
50
60
70
80
0,5 0,75 1,0 1,25 1,5 1,75 2,0
Hàm
lượ
ng
đư
ờn
g kh
ử (
μg/m
l)
Tỷ lệ enzyme Htec2/cơ chất (%)
cành lá quế Vỏ quế

99
Hình 4.8e. Bảng và đồ thị biểu diễn ảnh hưởng thời gian lên hàm lượng đường khử
Kết quả khảo sát ảnh hưởng của thời gian xử lý thể hiện ở Hình 4.8e cho thấy
thời gian cũng là một trong những yếu tố quyết định đến hiệu quả thủy phân. Thời
gian càng dài thì hiệu quả thủy phân càng tăng và đạt cực đại ở 8 h. Khi đó hàm lượng
đường khử ở mẫu lá quế là 78,14 μg/ml, ở mẫu vỏ quế là 61,14 μg/ml. Nếu tiếp tục
kéo dài thời gian, hiệu quả thủy phân không tăng, ngược lại hàm lượng đường còn
giảm.
Thời gian kéo dài thì enzyme có điều kiện để cắt mạch triệt để, tăng thời gian
tiếp xúc giữa enzyme và cơ chất, dẫn đến sự biến đổi sâu sắc của cơ chất. Tuy nhiên,
độ bền của enzyme ở nhiệt độ xác định cũng nằm trong khoảng giới hạn. Nếu kéo dài
thời gian thủy phân quá mức thì hoạt tính của enzyme cũng giảm đồng thời các vi
sinh vật lạ phát triển, hoạt động sinh ra nhiều sản phẩm thứ cấp. Còn nếu thời gian
thủy phân ngắn, sự phân cắt cellulose chưa triệt để, hiệu suất thủy phân kém. Các số
liệu và phân tích trên cho thấy thủy phân lignocellulose trong các mẫu cành lá quế,
vỏ quế thì thời gian 8 h là thích hợp.
Nhận xét chung:
Các kết quả nêu trên cho thấy các điều kiện sau đây là tốt nhất cho quá trình
phân giải thành tế bào cành lá và vỏ quế bằng hệ enzyme Laccase-Htec2:
- pH = 4,5 - 5,0
- Nhiệt độ thủy phân: 45oC
0
20
40
60
80
100
2h 4h 6h 8h 10h 12h
Hàm
lượ
ng
đư
ờn
g kh
ử (
μg/
ml)
Thời gian thủy phân
Cành lá quế Vỏ quế

100
- Nồng độ Laccase/cơ chất là 0,4 ml/g và nồng độ Htec2/cơ chất là 1,5% (đối
với cành lá quế). Đối với vỏ quế các nồng độ tương ứng là 0,7 ml/g và 1,5%.
- Thời gian thủy phân: 8 giờ
4.1.4.2. Tối ưu hóa quá trình thủy phân cành lá quế với hệ enzyme kết hợp
Laccase-Htec2 bằng phương pháp qui hoạch thực nghiệm
Để tìm điều kiện tối ưu chính xác của quá trình thủy phân trên cơ sở mô hình
tính toán, chúng tôi áp dụng phương pháp qui hoạch thực nghiệm [7].
Qua phân tích các yếu tố ảnh hưởng, chọn pH là (Z1) , nhiệt độ T là (Z2), thời
gian t là (Z3), tỷ lệ Laccase/cơ chất là (Z4) , và tỷ lệ Htec2/cơ chất là (Z5).
Chọn kế hoạch trực giao bậc 2 Box- Wilson để mô tả vùng gần đường và tìm
điều kiện tối ưu. Với các yếu tố k=5, số thí nghiệm tại tâm n0 =1 ta có cánh tay đòn
α = 1,546. Các giá trị của các yếu tố ảnh hưởng ở các mức tương ứng được thể hiện
trên Bảng 4.10a
Bảng 4.10a. Giá trị ở các mức của các yếu tố ảnh hưởng
Yếu tố -α -1 0 +1 + α
pH 4,23 4,5 5,0 5,5 5,77
T, 0C 37,3 40 45 50 52,7
t, h 4,91 6 8 10 10,09
Laccase E /S (ml/g) 0,245 0,3 0,4 0,5 0,555
Htec2 /S (%) (g/100g) 1,1 1,25 1,5 1,75 1,89
Trong đó các biến x1, x2, x3, x4, x5 là biến mã hóa mô tả tương ứng với các
biến thực Z1, Z2, Z3, Z4, Z5.
Tổng số thí nghiệm theo kế hoạch trực giao bậc 2 là:
N = 22-1 +2.k + n0 = 25-1 + 2.5 + 1 =27
Ma trận kế hoạch hóa và kết quả thực nghiệm ở Bảng 4.10b với 27 thí nghiệm
chính không nhân kế hoạch là 25-1 với hệ thức phát sinh là x5 = x1 *x2*x3*x4.

101
Bảng 4.10b. Ma trận kế hoạch hóa và kết quả thực nghiệm
STT x0 x1 x2 x3 x4 x5 x1x2 x1x3 x1x4 x1x5 x2x3 x2x4 x2x5 x3x4 x3x5 x4x5 x’1 x’
2 x’3 x’
4 x’5 y1 ŷ1 y2 ŷ2
1 + - - - - + + + + - + + - + - - 0,23 0,23 0,23 0,23 0,23 65,208 64,589 0,942 0,938
2 + + - - - - - - - - + + + + + + 0,23 0,23 0,23 0,23 0,23 64,19 65,542 0,953 0,944
3 + - + - - - - + + + - - - + + + 0,23 0,23 0,23 0,23 0,23 67,284 68,322 0,941 0,931
4 + + + - - + + - - + - - + + - - 0,23 0,23 0,23 0,23 0,23 69,598 69,275 0,943 0,937
5 + - - + - - + - + + - + + - - + 0,23 0,23 0,23 0,23 0,23 71,18 71,040 0,946 0,942
6 + + - + - + - + - + - + - - + - 0,23 0,23 0,23 0,23 0,23 71,494 71,993 0,945 0,948
7 + - + + - + - - + - + - + - + - 0,23 0,23 0,23 0,23 0,23 70,588 71,273 0,936 0,935
8 + + + + - - + + - - + - - - - + 0,23 0,23 0,23 0,23 0,23 72,57 72,255 0,947 0,941
9 + - - - + - + + - + + - + - + - 0,23 0,23 0,23 0,23 0,23 72,576 72,090 0,945 0,944
10 + + - - + + - - + + + - - - - + 0,23 0,23 0,23 0,23 0,23 72,89 73,042 0,947 0,950
11 + - + - + + - + - - - + + - - + 0,23 0,23 0,23 0,23 0,23 71,984 72,323 0,936 0,938
12 + + + - + - + - + - - + - - + - 0,23 0,23 0,23 0,23 0,23 73,966 73,275 0,946 0,943
13 + - - + + + + - - - - - - + + + 0,23 0,23 0,23 0,23 0,23 73,88 74,031 0,940 0,949
14 + + - + + - - + + - - - + + - - 0,23 0,23 0,23 0,23 0,23 75,844 74,984 0,951 0,954
15 + - + + + - - - - + + + - + - - 0,23 0,23 0,23 0,23 0,23 74,956 74,273 0,939 0,942
16 + + + + + + + + + + + + + + + + 0,23 0,23 0,23 0,23 0,23 75,27 75,226 0,946 0,947
17 + -1,546 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,23 1,62 -0,77 -0,77 -0,77 74,594 72,618 0,962 0,947
18 + +1,546 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,23 1,62 -0,77 -0,77 -0,77 75,714 74,091 0,966 0,959
19 + 0 -1,546 0 0 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 1,62 -0,77 -0,77 74,654 72,362 0,965 0,958
20 + 0 +1,546 0 0 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 1,62 -0,77 -0,77 76,322 74,078 0,941 0,947
21 + 0 0 -1,546 0 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 1,62 -0,77 73,27 72,169 0,939 0,948
22 + 0 0 +1,546 0 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 1,62 -0,77 77,438 77,306 0,969 0,955
23 + 0 0 0 -1,546 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 -0,77 -0,77 74,872 71,294 0,930 0,957
24 + 0 0 0 +1,546 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 -0,77 -0,77 78,872 78,055 0,976 0,967
25 + 0 0 0 0 -1,546 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 -0,77 1,62 77,136 77,749 0,962 0,971
26 + 0 0 0 0 +1,546 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 -0,77 1,62 76,482 77,749 0,965 0,971
27 + 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,23 -0,77 -0,77 -0,77 -0,77 78,118 77,749 0,976 0,971

102
Trong bảng 4.10b, để ma trận kế hoạch hóa có tính trực giao thì các cột 𝑥𝑗2
(j=1.5) được biến đổi tuyến tính thành 𝑥𝑗′ theo công thức:
𝑥𝑗′ = 𝑥𝑗
2 - 𝑥𝑗2
= 𝑥𝑗2
- ∑ 𝑥𝑗𝑖
2𝑛𝑖=1
𝑁 (1)
Cũng trong bảng này thông số tối ưu hóa thứ nhất y1 là hàm lượng đường khử
(μg/ml) và thông số tối ưu hóa thứ hai y2 là hàm lượng tinh dầu (%).
Để xác định phương sai tái sinh đã tiến hành 3 thí nghiệm lặp ở điều kiện:
pH=5,
T= 45oC, τ = 8h, Laccase/S = 0.4, Htec2/S = 1,89 đã nhận được kết quả:
- Đối với y1: 77,71 77,71 và 76.62
- Đối với y2: 0,972 0,973 và 0,968
Các công thức tình để tìm mô hình theo tài liệu []:
bj = ∑ 𝑥𝑗𝑖 𝑦𝑖
𝑁𝑖=1
∑ 𝑥𝑗𝑖2𝑁
𝑖=1
; j = 1, k̅̅̅̅̅ (2)
𝑆𝑏𝑗
2 = 𝑆𝑡𝑠
2
∑ 𝑥𝑗𝑖2𝑁
𝑖=1
(3)
ŷ = 𝑏𝑜′ + ∑ 𝑥𝑖
𝑛𝑖=1 + ∑ 𝑏𝑢𝑗𝑥𝑢𝑥𝑗
𝑘𝑢,𝑗=1 + ∑ 𝑏𝑗𝑗𝑥𝑗
′𝑘𝑗=1 (4)
Hay chuyển hàm số ŷ về dạng thông thường
ŷ = 𝑏𝑜′ + ∑ 𝑏𝑗𝑥𝑗
𝑘𝑗=1 + ∑ 𝑏𝑢𝑗𝑥𝑢𝑥𝑗
𝑘𝑢,𝑗=1 + ∑ 𝑏𝑗𝑗𝑥𝑗
2𝑘𝑗=1 (5)
thì:
bo = 𝑏𝑜′ - 𝑏11𝑥1
2̅̅ ̅ - ……. - 𝑏𝑘𝑘𝑥𝑘2̅̅ ̅ (6)
Ý nghĩa của các hệ số kiểm định theo tiêu chuẩn Student, còn tính tương hợp
của mô hình với kết quả thực nghiệm kiểm định theo tiêu chuẩn Fisher.
Chúng tôi đã sử dụng phần mềm Design Expert 7.0 như là phần mềm viết bằng
ngôn ngữ thuật toán FORTRAN để xử lý số liệu ở bảng 4.10b và nhận được kết quả
sau:
ŷ1 = 79,728 + 0,476 x1 + 0,555x2 + 1,662 x3 + 2,187 x4 + 0,439 x1x5 - 0,436 x2x3 -
0,436 x2x4 - 0,689 x3x4 - 1,979 x12 - 1,839 x2
2 - 1,825 x32 - 1,26 x4
2 -1,286 x52 (7)
Với: Sts2 = 0,2984
S𝑏𝑗= 0,09001; S𝑏𝑢𝑗
=0,09657; S𝑏𝑗𝑗= 0,16158

103
t1 = 5,288; t2 =6,161; t3 = 18,447; t4 = 24,275; t15 = 4,542; t23 = 4,519; t24 =
4,519; t34 = 7,131; t11= 12,245; t22 = 11,381; t33 = 11,728; t44 = 7,799; t55 = 7,962 đề
lớn hơn tpr(f) = t0,05(2) = 4,3
𝑠𝑑ư2 =
∑ (𝑦1𝑖−ŷ1𝑖)2𝑛𝑖=1
𝑁−𝑙 =
∑ (𝑦1𝑖−ŷ1𝑖)2𝑛𝑖=1
27−14 = 3,07501
F= 𝑠𝑑ư
2
𝑠𝑡𝑠2 =
3,07501
0,2984 = 10,304 < Fp (f1, f2) = F0,05 (13,2) = 19,4
Giá trị tính toán ŷ1theo phương tính (7) được đưa vào bảng 2.
2. Đối với hàm lượng tinh dầu y2:
�̂�2 = 0,97498 + 0,00285 x1 - 0,00347 x2 + 0,00209 x3 + 0,00328 x4 - 0,00373
x12 - 0,00833 x2
2 - 0,00791 x32 - 0,00833 x4
2 - 0,00394 x52 (8)
Với Sts2 = 7.10-6
sbj=4,3626.10-4
sbuj=4,6771.10-4
sbjj= 7,8253.10-4
t1 =6,528; t2 =7,954; t3 =4,785; t4 =7,514; t11=4,769; t22=10,647; t33=10,112;
t44= 10,647; t55=5,031
đều lớn hơn tp (f)=4,3
𝑠𝑑ư2 =0,00012
F = 𝑠𝑑ư
2
𝑠𝑡𝑠2 =
0,00012
0,7 .10−6 = 17,467 < Fp (f1, f2) = F0.05 (17,2) = 19,45
Giá trị tính toán ŷ2 theo phương tính (8) được đưa vào bảng 2.
Như vậy đã nhận hai phương tính hồi quy (7) và (8) trường hợp với thực
nghiệm. Có thể nhận thấy rằng cả 5 yếu tố đều có ảnh hưởng tới các thông số đầu ra,
tuy nhiên ở phương toán (7) có ảnh hưởng của 4 cặp tương tác. Sử dụng phần mềm
Design Expert 7.0 ta có hình 3D mô tả mặt đáp ứng theo hai yếu tố nhiệt độ và thời
gian cho ŷ1như sau:

104
Hình 4.9a. Mặt đáp ứng vùng tối ưu của cặp yếu tố x1x2
Hình 4.9b. Mặt đáp ứng vùng tối ưu của cặp yếu tố x2x4
Hình 4.9c. Mặt đáp ứng vùng tối ưu của cặp yếu tố x3x5
Tuy nhiên cần giải bài toán tối ưu nhiều mục tiêu (trong trường hợp này là 2:
hàm lượng đường khử và hiệu suất tinh dầu) với nhiều yếu tố ảnh hưởng (trong trường
hợp này là 5 yếu tố: pH, nhiệt độ T, thời gian t, tỷ lệ enzyme laccase/cơ chất, tỷ lệ
enzyme Htec2/cơ chất.
Tìm hàm lượng tinh dầu lớn nhất
ŷ2 = 0,97498 + 0,00285 x1 - 0,00347 x2 + 0,00209 x3 + 0,00328 x4 -
0,00373 𝒙𝟏𝟐 - 0,00833 𝒙𝟐
𝟐 - 0,00791 𝒙𝟑𝟐 - 0,00833 𝒙𝟒
𝟐 - 0,00394 𝒙𝟓𝟐 (9)
Với các điều kiện ràng buộc: -1.546 ≤ xj ≤ 1.546 (j = 1-5)
ŷ1 70 μg/ml
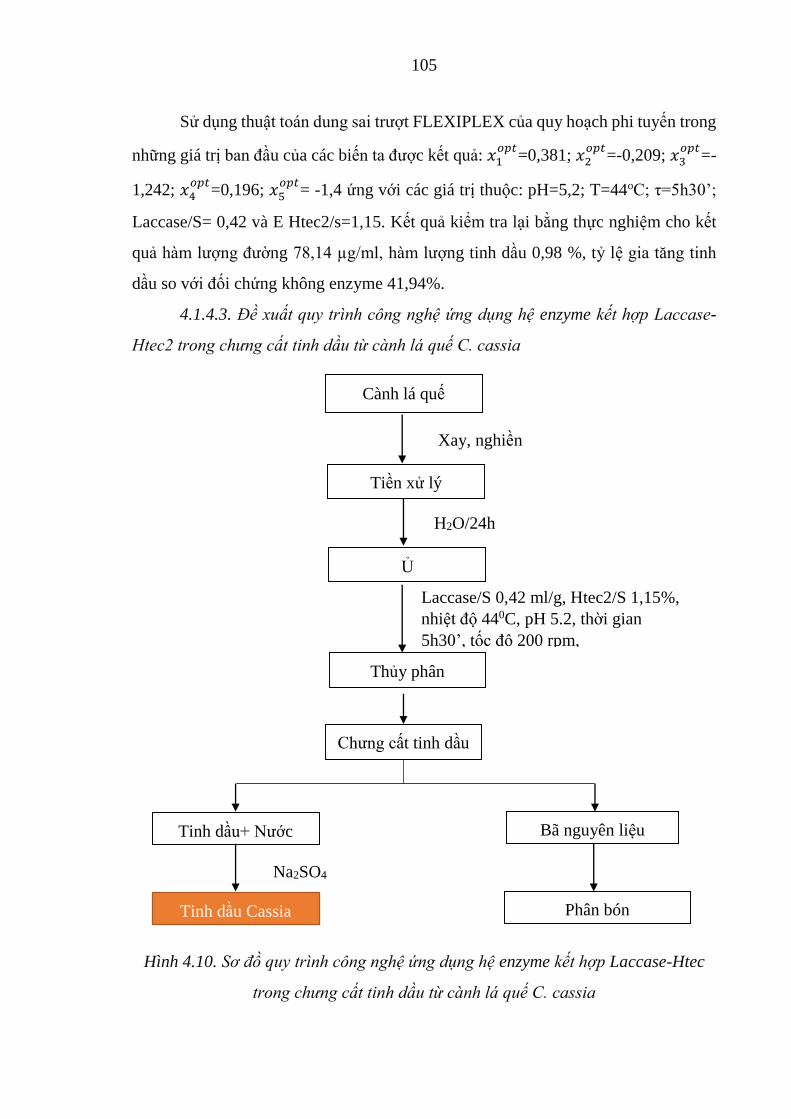
105
Sử dụng thuật toán dung sai trượt FLEXIPLEX của quy hoạch phi tuyến trong
những giá trị ban đầu của các biến ta được kết quả: 𝑥1𝑜𝑝𝑡
=0,381; 𝑥2𝑜𝑝𝑡
=-0,209; 𝑥3𝑜𝑝𝑡
=-
1,242; 𝑥4𝑜𝑝𝑡
=0,196; 𝑥5𝑜𝑝𝑡
= -1,4 ứng với các giá trị thuộc: pH=5,2; T=44oC; τ=5h30’;
Laccase/S= 0,42 và E Htec2/s=1,15. Kết quả kiểm tra lại bằng thực nghiệm cho kết
quả hàm lượng đường 78,14 µg/ml, hàm lượng tinh dầu 0,98 %, tỷ lệ gia tăng tinh
dầu so với đối chứng không enzyme 41,94%.
4.1.4.3. Đề xuất quy trình công nghệ ứng dụng hệ enzyme kết hợp Laccase-
Htec2 trong chưng cất tinh dầu từ cành lá quế C. cassia
Hình 4.10. Sơ đồ quy trình công nghệ ứng dụng hệ enzyme kết hợp Laccase-Htec
trong chưng cất tinh dầu từ cành lá quế C. cassia
Cành lá quế
Tiền xử lý
H2O/24h
Laccase/S 0,42 ml/g, Htec2/S 1,15%,
nhiệt độ 440C, pH 5.2, thời gian
5h30’, tốc độ 200 rpm,
Thủy phân
Tinh dầu+ Nước
Ủ
Xay, nghiền
Chưng cất tinh dầu
Bã nguyên liệu
Phân bón
Na2SO4
Tinh dầu Cassia

106
Trên cơ sở các kết quả nghiên cứu nêu trên, chúng tôi đề xuất một qui trình
công nghệ ứng dụng hệ enzyme kết hợp Laccase-Htec2 trong chưng cất tinh dầu từ
cành lá quế C. cassia.
Thuyết minh quy trình
1. Nguyên liệu: cành lá quế sau khi thu hái được phơi khô trong bóng râm và
đưa về bảo quản trong kho.
2. Tiền xử lý nguyên liệu: nguyên liệu cành lá quế chặt nhỏ và xay nghiền tới
kích thước lọt sàng 3 mm.
3. Ủ nguyên liệu: nguyên liệu trước khi xử lý enzyme bổ sung thêm nước theo
tỷ lệ 1:5, ngâm ủ trong 24 giờ ở nhiệt độ thường nhằm gia tăng sự hút nước của
nguyên liệu.
4. Thủy phân: Nguyên liệu cho vào nồi hai vỏ có hệ thống điều nhiệt, điều
chỉnh pH bằng axit HCl loãng về pH 5,2. Gia nhiệt đến khi hỗn hợp trong bình thủy
phân đạt 45oC thì bắt đầu bổ sung enzyme theo tỷ lệ Laccase 0,42 ml/g, Htec2 1,15%.
Trong suốt quá trình phản ứng, nhiệt độ được duy trì ở 45oC và quá trình được kiểm
soát bằng hàm lượng đường khử sinh ra. Thời gian phản ứng là 5 giờ 20 phút, được
tính từ khi bổ sung enzyme.
3. Chưng cất tinh dầu bằng cất cuốn hơi nước: Hỗn hợp sau thủy phân được
cho vào nồi có lắp hệ thống chưng cất hơi nước kiểu Clevenger. Gia nhiệt đến nhiệt
độ sôi để hơi nước cuốn theo tinh dầu, dẫn qua hệ thống làm mát thu được hỗn hợp
tinh dầu và nước. Quá trình chưng cất diễn ra khoảng 6 h để chiết triệt để tinh dầu
trong nguyên liệu. Tinh dầu quế nặng hơn nước nên sẽ chìm xuống dưới. Tách phần
tinh dầu khi hỗn hợp tinh dầu và nước phân tách hoàn toàn thành 2 pha.
4. Phần tinh dầu được thu và làm khan bằng Na2SO4 cho tinh dầu thành phẩm.
Bã nguyên liệu sau chưng cất tinh dầu có thể bổ sung vi sinh vật phân hủy để
làm phân hữu cơ.
4.2. Kết quả thăm dò ứng dụng hệ enzyme kết hợp Laccase-Htec2 trong
chưng cất tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna
4.2.1. Kết quả phân tích thành phần chất nền của bột gỗ gió bầu A. crassna
Kết quả xác định các thành phần kiến tạo nên khung cấu trúc của gỗ gió bầu
(Bảng 4.11a) cho thấy gỗ gió bầu mang các đặc trưng của nguyên liệu chứa
lignocellulose bao gồm cellulose 34,75%, hemicellulose 15,27% và lignin 29,10%. Hàm

107
lượng cellulose và lignin cao cho thấy thành tế bào của nguyên liệu rất bền chắc, gây khó
khăn cho chưng cất tinh dầu. Điều đó giải thích vì sao trong công nghệ truyền thống, bột
gỗ gió bầu thường được ủ ướt một thời gian trước khi đưa đi chưng cất.
Bảng 4.11a. Kết quả phân tích thành phần bột gỗ gió bầu A. crassna
(hàm lượng tính theo % khối lượng khô tuyệt đối)
TT Chỉ tiêu phân tích Hàm lượng (%)
1 Cellulose 34,75 ± 0,21
2 Lignin 29,10 ± 0,18
3 Pentose 15,27 ± 0,20
4 Các hợp chất trích ly (cồn) 8,32 ± 0,13
4.2.2. Kết quả thăm dò tác động hệ enzyme Laccase - Htec2 lên hàm lượng
và thành phần tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna
Dựa trên các kết quả thu được khi nghiên cứu ứng dụng enzyme trong chưng
cất tinh dầu quế, chúng tôi đã chọn hệ enzyme Laccase-Htec2 để áp dụng cho việc
chưng cất tinh dầu trầm hương.
Kết quả sau 3 ngày chưng cất, mẫu đối chứng (mẫu không có enzyme) cho
0,024% tinh dầu (tính theo khối lượng mẫu khô gió), mẫu nghiên cứu (mẫu có
enzyme) cho 0,032% tinh dầu, tỷ lệ gia tăng tinh dầu đạt 33,3%.
Thành phần hóa học của các mẫu tinh dầu (Bảng 4.11b) được xác định bằng
phương pháp GC-MS, trong đó việc nhận dạng chất dựa trên sự phù hợp của phổ khối
và chỉ số thời gian lưu (RI, chỉ số Kovats) với các dữ kiện trong ngân hàng dữ liệu
của máy.
Bảng 4.11b. Kết quả phân tích thành phần hóa học của tinh dầu trầm hương thu
được từ gỗ cây gió bầu A. crassna bằng 2 phương pháp khác nhau
STT Chất CTCT RI
Mẫu không
enzyme
(%)
Mẫu có
enzyme
(%)
1 2-Allyl-4-methylphenol C10H12O 1288 0,42
2 Copaene <a-> C15H24 1409 0,56
3 Curcumene <ar-> C15H22 1507 0,15
4 Chamigrene <b-> C15H24 1509 0,24

108
STT Chất CTCT RI
Mẫu không
enzyme
(%)
Mẫu có
enzyme
(%)
5 Muurola-4(14),5-diene <trans-> C15H24 1513 0,12
6 Ionol C15H24O 1519 0,71 2,97
7 Muurolene <a-> C15H24 1537 0,47
8 Selinene <a-> C15H24 1543 0,13
9 Dihydroagarofuran <cis-> C15H26O 1550 0,11
10 Cadinene <d-> C15H24 1552 0,52
11 Calamenene <trans-> 1566 0,41
12 n-Dodecanoic acid C12H24O2 1568 0,12
13 Nerolidol <Z-> C15H26O 1579 0,32
14 Calacorene <a-> C15H20 1590 0,28
15 Agarofuran <a-> C14H22O 1550 0,17
16 Unknown 1628 1,50
17 Salvial-4(14)-en-1-one C15H24O 1666 0,23
18 Eudesmol <10-epi-g-> C15H26O 1676 0,41 1,27
19 Eudesmol <g-> C15H26 1682 0,58 2,72
20 Agarospirol C15H26O 1685 0,23 0,72
21 Guaiol C15H26O 1688 0,50
22 Hinesol C15H26O 1690 0,16 0,56
23 Unknown 1693 1,46
24 Unknown 1699 1,88
25 Valerianol C15H26O 1704 0,89 2,59
26 Cadinol <a-> C15H26O 1710 0,36
27 Eudesmol <b-> C15H26O 1714 0,57 1,61
28 Intermedeol <neo-> C15H26O 1716 0,86 2,08
29 Unknown 1727 1,81
30 Cadalene C15H18 1733 0,82

109
STT Chất CTCT RI
Mẫu không
enzyme
(%)
Mẫu có
enzyme
(%)
31 Eudesma-4(15),7-dien-1b-ol C15H24O 1739 0,83
32 Eudesma-3,11-dien-8-one C15H22O 1746 0,63
33 Unknown 1750 2,26
34 Cadina-1(10),4-dien-8a-ol C15H24 1759 1,40 1,96
35 Unknown 1768 1,49
36 n-Tetradecanoic acid C14H28O2 1771 0,81
37 Unknown 1774 3,59
38 Calamenen-10-one <10-nor> C14H18O 1781 0,43 0,64
39 Isonaviculol 1795 1,08 1,98
40 Unknown 1800 3,07
41 Unknown 1803 1,13
42 Costol <a-> 1824 0,78 1,39
43 Neopetasone C15H22O 1829 0,11 0,76
44 Unknown 1832 2,21
45 Unknown 1842 1,31 2,54
46 Unknown 1857 1,80
47 n-Pentadecanoic acid C15H30O2 1873 2,41
48 Unknown 1891 2,11 3,33
49 Nootkatone C15H22O 1896 0,57 0,86
50 Cyclohexadecanolide C16H30O2 1957 1,97 0,72
51 n-Hexadecanoic acid C16H32O2 1979 47,54 19,45
52 Ethyl palmitate C18H36O2 2004 0,76
53 Unknown 2065 1,21
54 Unknown 2153 1,23
55 Oleic acid C18H34O2 2157 6,44 3,44
56 Unknown 2163 4,32

110
STT Chất CTCT RI
Mẫu không
enzyme
(%)
Mẫu có
enzyme
(%)
57 n-Octadecanoic acid C18H36O2 2178 1,16
58 Ethyl oleate C20H38O2 2181 0,61
59 Diisooctyl phthalate C24H38O4 2566 0,17
Tổng 81,52 80,49
Tổng các dẫn xuất của axit béo 59,85 22,89
Tổng các Terpen hydrocarbon 0 3,70
Tổng các Terpen chứa oxy 8,78 24,76
Tổng các chất khác 2,14 1,64
Tổng các chất chưa xác định được có
hàm lượng > 1%
10,75 27,50
Một số thành phần đặc trưng của tinh dầu trầm hương (Hình 4.11).
Hình 4.11. Một số thành phần đặc trưng của tinh dầu trầm hương
Các kết quả trên Bảng 4.11b cho thấy:
- Việc xử lý bột gỗ gió bầu với hệ enzyme Laccase-Htec2 để phá hủy cấu trúc
lignocellulose trước khi chưng cất làm tăng hiệu suất thu hồi tinh dầu từ 0,024% lên
0,032%.

111
- Hàm lượng của các thành phần có giá trị, mang mùi trầm hương đặc trưng
(Bảng 4.11b) cũng tăng lên rõ rệt, cụ thể cis-dihydroagarofuran (0,11%, đối chứng
0%), α-agarofuran (0,17%, đối chứng 0%), agarospirol (0,72%, đối chứng 0,23%),
hinesol (0,56%, đối chứng 0,16%), valerianol (2,59%, đối chứng 0,89%),
neopetasone (0,76%, đối chứng 0,11%), tổng các terpen hydrocarbon (3,70%, đối
chứng 0%), tổng các terpen chứa oxy (24,76%, đối chứng 8,78%).
- Các axit béo và dẫn xuất của chúng là các thành phần không mong muốn
trong tinh dầu trầm hương. Trong mẫu nghiên cứu, hàm lượng của chúng chỉ còn
22,89%, ít hơn hẳn so với đối chứng là 59,85%.
- Tổng số các chất chưa được xác định (unknown) trong kết quả phân tích tinh
dầu trầm hương là rất cao, vì thế cần có các nghiên cứu hóa học tiếp theo để làm rõ.
4.3. Kết quả nghiên cứu ứng dụng enzyme kết hợp trong chiết xuất
lipid và làm giàu các axit béo n-3 PUFA từ đầu cá ngừ vây vàng Thunnus
albarcares
4.3.1. Kết quả nghiên cứu các chất nền và thành phần axit béo của một số loại
đầu cá ngừ của Việt Nam
4.3.1.1. Kết quả nghiên cứu các chất nền
Kết quả phân tích hàm lượng protein, lipid, nước, tro của 08 mẫu đầu (đã bỏ
mang, mắt) của 08 loài cá ngừ của Việt Nam được chỉ ra ở Bảng 4.12:
Bảng 4.12. Thành phần các chất nền của một số loại đầu cá ngừ của Việt Nam
Mẫu Hàm lượng (%)
Nước Lipid Protein* Tro
Cá ngừ ồ
Auxis rochei 61,67 ± 0,23 4,50 ± 0,24 17,3 ± 0,38 9,43 ± 0,46
Cá ngừ chù
Auxis thazard 62,44 ± 0,13 5,10 ± 0,32 18,32 ± 0,26 8,99 ± 0,28
Cá ngừ chấm
Euthynnus affinis 59,39 ± 0,12 5,50 ± 0,14 18,23 ± 0,25 8,98 ± 0,48
Cá ngừ bò
Thunnus tonggol 54,72 ± 0,11 11,71 ± 0,32 17,98 ± 0,33 9,22 ± 0,21

112
Mẫu Hàm lượng (%)
Nước Lipid Protein* Tro
Cá ngừ sọc dưa
Sarda orientalis 55,82 ± 0.21 8,01 ± 0,24 18,75 ± 0,41 7,45 ± 0,32
Cá ngừ vằn
Katsuwonus
pelamis
59,50 ± 0,32 7,21 ± 0,21 17,30 ± 0,45 10,21 ± 0,35
Cá ngừ vây vàng
Thunnus albacares 56,32 ± 0,36 14,80 ± 0,28 17,67 ± 0,43 9,31 ± 0,37
Cá ngừ mắt to
Thunnus obesus 57,45 ± 0,43 14,20 ± 0,48 17,78 ± 0,37 8,13 ± 0,38
Kết quả trên Bảng 4.12 cho thấy:
- Hàm lượng protein của các mẫu dao động trong khoảng 17 - 20%, tương ứng
với 40 - 50% protein tính theo lượng chất khô. Phụ phẩm đầu cá ngừ có hàm lượng
protein cao và chứa nhiều các axit amin thiết yếu, nên đã có nhiều nghiên cứu thủy
phân nguồn protein này [12], [101], [101].
- Hàm lượng lipid của các mẫu dao động trong khoảng 4,5 - 14,8%. Kết quả
cho thấy hàm lượng lipid tăng dần theo kích thước loài, cao nhất là nhóm cá ngừ đại
dương có kích thước lớn (500 – 2.000 mm) như cá ngừ vây vàng (14,8%), cá ngừ mắt
to (14,2%) và thấp nhất ở các loài cá ngừ kích thước nhỏ (140 - 310 mm) như cá ngừ
ồ (4,5%), cá ngừ chù (5,1%) . So sánh với một số phế liệu đầu từ một số loài cá khác,
một số phế liệu đầu cá hồi (3,57%), đầu cá basa (3,41%), đầu cá tra (3,15%) (Phạm
Quốc Long, 2013) [17] cho thấy đây là nguồn nguyên liệu có thể sản xuất dầu cá phục
vụ trong công nghiệp thực phẩm và y dược.
4.3.1.2. Kết quả phân tích thành phần axit béo
Để tìm kiếm nguồn nguyên liệu giàu các axit béo không no n-3 PUFA, đặc
biệt là EPA, DHA và DPA, thành phần các axit béo trong các lipid tổng thu được
đã được phân tích bằng phương pháp GC-MS. Kết quả được trình bày trên Bảng
4.13.

113
Bảng 4.13. Thành phần axit béo trong các mẫu lipid thu được từ đầu một số loài cá
ngừ của Việt Nam (hàm lượng tính bằng % diện tích của các peak)
TT Axit béo Ngừ ồ Ngừ
chù
Ngừ
chấm
Ngừ
vằn
Ngừ
bò
Sọc
dưa
Vây
vàng
Mắt
to
1 14:00 2,82 0,83 0,74 3,50 3,14 1,73 2,70 3,33
2 15:00 0,91 0,71 0,73 1,20 1,12 0,94 0,76 1,14
3 16:1n-9 5,93 0,23 2,38 0,23 0,22
4 16:1n-7 2,05 6,34 4,76 3,08 5,34 5,19
5 16:00 22,12 24,24 23,12 18,43 19,45 20,04 13,78 22,86
6 17:1n-9 1,09 0,52 1,13 1,12 0,84 0,82 0,76
7 17:1n-7 0,92 0,64 0,82 0,83 0,64 0,82
8 17:00 0,93 1,46 1,47 1,74 1,53 1,65 0,80 1,38
9 18:4n-3 0,52 0,54 0,96
10 18:2n-6 2,13 1,33 1,43 1,09 1,19 1,12 1,43 1,34
11 18:1n-9 22,76 16,36 16,42 16,27 13,09 12,35 14,87 10,10
12 18:1n-7 3,12 3,83 3,75 3,24 2,29 3,23 2,71 3,17
13 18:00 6,11 11,46 11,86 7,72 8,74 12,61 4,33 8,39
14 19:00 0,23 0,71 0,31 0,51 0,37 0,52 0,17
15 20:5n-3
(EPA) 4,54 4,55 4,74 5,89 6,31 5,22 8,36 5,94
16 20:4n-6 1,94 4,32 4,42 3,22 2,71 4,36 3,11 2,12
17 20:4n-3 0,31 0,61 0,56 0,35 0,20
18 20:2n-6 0,30 0,31 0,59 0,44 0,36 0,84
19 20:1n-9 1,23 1,06 0,93 1,61 1,09 0,91 2,72 0,47
20 20:00 0,34 0,18 0,31 0,43 0,48 0,52 0,24 0,37
21 22:6n-3
(DHA) 20,33 21,25 18,98 20,26 23,44 22,86 27,75 27,30
22 22:5n-3 0,48 0,75 0,56 2,12 2,04 1,75 3,04 1,60
23 22:3n-6 1,18 1,10 2,53 0,82 2,26 0,73 0,20
24 22:4n-6 0,89 1,72 1,80 1,22 2,71 1,94 2,00
25 22:1n-9 0,24 1,02 0,40
26 22:00 0,50 0,23 0,35

114
TT Axit béo Ngừ ồ Ngừ
chù
Ngừ
chấm
Ngừ
vằn
Ngừ
bò
Sọc
dưa
Vây
vàng
Mắt
to
27 23:00 0,20
28 24:1n-9 0,73 0,72 0,25 0,84 1,60 0,40
29 24:00:00 0,27 0,38
30 unknow 0,22 0,25 0,40 0,50 2,70
Tổng 100 100 100 100 100 100 100 100
Tổng SFA 33,44 39,59 39,04 33,76 35,45 38,59 22,74 37,43
Tổng MUFA 35,05 24,78 25,97 29,65 24,21 22,63 28,86 19,33
Tổng PUFA 31,49 35,63 34,77 36,34 39,94 38,28 48,36 40,50
n-3 PUFA 25,35 26,86 24,28 29,4 32,89 30,18 37,82 34,84
EPA+DHA+DPA 25,35 26,55 24,28 28,27 31,79 29,83 39,15 34,84
Hệ số đánh giá* 114,08 136,99 133,54 217,30 343,98 263,12 559,74 494,73
Ghi chú: Hệ số đánh giá dựa trên hàm lượng lipid tổng * hàm lượng n-3 PUFA
Bảng 4.13 cho thấy có khoảng 30 loại axit béo từ C14 - C24 được tìm thấy trong
lipid tổng từ phụ phẩm đầu của 08 loài cá ngừ. Trong đó thành phần axit béo no SFA
chiếm 22,74 - 39,59%, axit béo không no một nối đôi MUFA chiếm 19,33 - 35,05%,
và axit béo không no đa nối đôi PUFA chiếm 31,49 - 48,36%. Đáng chú ý là phụ
phẩm đầu cá ngừ đại dương kích thước lớn (cá ngừ vây vàng và cá ngừ mắt to) có
hàm lượng các acid béo không no đa nối đôi n-3 PUFA cao, lần lượt là 48,36 và
40,5%. Đây là nhóm axit béo có vai trò quyết định chất lượng của lipid với các thành
phần chủ yếu là EPA, DHA và DPA.
So sánh thành phần n-3 PUFA của phụ phẩm đầu cá ngừ với các loài cá khác,
chỉ số này tương đương hoặc vượt trội so với một số loài cá nước mặn khác được
Phạm Quốc Long và cs. (2013) [17] công bố như cá Nục 27,75%, cá Đồng 27,88%,
cá Cơm 23,72% và một số phế liệu cá như đầu cá tra 9,76%, đầu cá basa 10,34%, đầu
cá hồi 12,14% (tác giả Linder M., 2005 [120], công bố hàm lượng n-3 PUFA của đầu
loài cá hồi họ nghiên cứu là 31,7%).
Qua hệ số đánh giá được xác định dựa trên hàm lượng lipid tổng và hàm lượng
n-3 PUFA cho thấy 03 loài cá ngừ có triển vọng trong việc sử dụng làm nguyên liệu
để sản xuất dầu cá chất lượng cao giàu n-3 PUFA là cá ngừ bò, cá ngừ vây vàng, cá

115
ngừ mắt to với hệ số đánh giá cao, lần lượt là 343,98; 559,74 và 494,73; trong đó cao
nhất là cá ngừ vây vàng Thunnus albarcares. Trong khuôn khổ luận án, chúng tôi lựa
chọn đầu cá ngừ vây vàng Thunnus albarcares có hệ số đánh giá cao nhất làm đối
tượng nghiên cứu chiết xuất lipid và làm giàu các hợp chât n3-PUFA.
4.3.2. Kết quả nghiên cứu ứng dụng enzyme trong chiết xuất lipid từ
đầu cá ngừ vây vàng T. albarcares
4.3.2.1. Tác động của việc xử lý enzyme lên hiệu suất chiết xuất lipid và mức
độ thủy phân nguyên liệu.
Enzyme protease được thử nghiệm để thủy phân đầu cá ngừ vây vàng Thunnus
albacare đều là các chế phẩm protease thương mại thuộc hai nhóm enzyme protease
từ vi sinh vật (Protamex và Alcalase) và từ thực vật (Bromelain và Papain).
Sau thủy phân, hỗn hợp phản ứng được ly tâm cho ta 2 sản phẩm mục tiêu là
lipid (dầu cá) và dịch protein hòa tan giàu axit amin. Kết quả thu nhận lipid và protein
hòa tan từ quá trình thủy phân bằng enzyme protease khác nhau được trình bày trên
Bảng 4.14, có so sánh với sản phẩm thu được bằng phương pháp ép nhiệt và phương
pháp chiết xuất với dung môi (phương pháp Blight & Dyer).
Bảng 4.14. Sản phẩm của quá trình chiết xuất lipid
Phương pháp Sản phẩm
Lipid (%) Protein hòa tan (%)
Ép nhiệt 3,22 ± 0,34 Dạng thô
Blight & Dyer 14,80 ± 0,28 Loại bỏ
Protamex 9,19 ± 0,14 3,71 ± 0,14
Alcalase 7,84 ± 0,33 3,63 ± 0,26
Bromelain 10,23 ± 0,42 4,12 ± 0,15
Papain 6,71 ± 0,43 3,14 ± 0,34
Kết quả Bảng 4.14 cho thấy:
- So với phương pháp truyền thống (ép nhiệt), phương pháp thủy phân bằng
enzyme protease đã cải thiện đáng kể quá trình tách dầu từ 3,22% lên 6,71 – 10,23%
(tăng 2 - 3 lần), đồng thời thu được sản phẩm protein hòa tan giàu axit amin tự do có
giá trị dinh dưỡng cao hơn khi sử dụng phương pháp ép nhiệt.
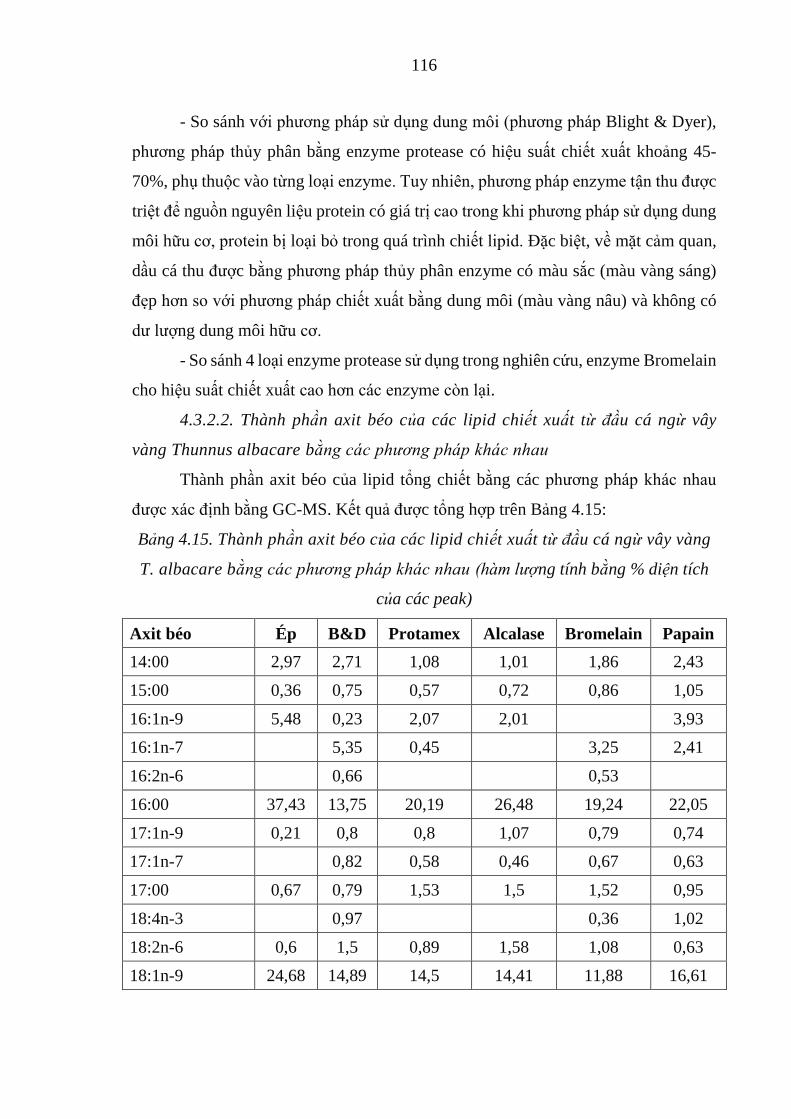
116
- So sánh với phương pháp sử dụng dung môi (phương pháp Blight & Dyer),
phương pháp thủy phân bằng enzyme protease có hiệu suất chiết xuất khoảng 45-
70%, phụ thuộc vào từng loại enzyme. Tuy nhiên, phương pháp enzyme tận thu được
triệt để nguồn nguyên liệu protein có giá trị cao trong khi phương pháp sử dụng dung
môi hữu cơ, protein bị loại bỏ trong quá trình chiết lipid. Đặc biệt, về mặt cảm quan,
dầu cá thu được bằng phương pháp thủy phân enzyme có màu sắc (màu vàng sáng)
đẹp hơn so với phương pháp chiết xuất bằng dung môi (màu vàng nâu) và không có
dư lượng dung môi hữu cơ.
- So sánh 4 loại enzyme protease sử dụng trong nghiên cứu, enzyme Bromelain
cho hiệu suất chiết xuất cao hơn các enzyme còn lại.
4.3.2.2. Thành phần axit béo của các lipid chiết xuất từ đầu cá ngừ vây
vàng Thunnus albacare bằng các phương pháp khác nhau
Thành phần axit béo của lipid tổng chiết bằng các phương pháp khác nhau
được xác định bằng GC-MS. Kết quả được tổng hợp trên Bảng 4.15:
Bảng 4.15. Thành phần axit béo của các lipid chiết xuất từ đầu cá ngừ vây vàng
T. albacare bằng các phương pháp khác nhau (hàm lượng tính bằng % diện tích
của các peak)
Axit béo Ép B&D Protamex Alcalase Bromelain Papain
14:00 2,97 2,71 1,08 1,01 1,86 2,43
15:00 0,36 0,75 0,57 0,72 0,86 1,05
16:1n-9 5,48 0,23 2,07 2,01 3,93
16:1n-7 5,35 0,45 3,25 2,41
16:2n-6 0,66 0,53
16:00 37,43 13,75 20,19 26,48 19,24 22,05
17:1n-9 0,21 0,8 0,8 1,07 0,79 0,74
17:1n-7 0,82 0,58 0,46 0,67 0,63
17:00 0,67 0,79 1,53 1,5 1,52 0,95
18:4n-3 0,97 0,36 1,02
18:2n-6 0,6 1,5 0,89 1,58 1,08 0,63
18:1n-9 24,68 14,89 14,5 14,41 11,88 16,61

117
Axit béo Ép B&D Protamex Alcalase Bromelain Papain
18:1n-7 4,78 2,73 3,34 3,75 2,97 6,93
18:00 9,85 4,28 14,33 15,77 11,56 6,69
19:00 0,19 0,59 0,46 0,43 0,16
20:4n-6 0,95 3,1 6,25 5,76 4,41 2,17
20:5n-3 2,42 8,41 4,27 3,47 5,47 9,51
20:3n-6 0,2 0,31
20:4n-3 0,79 0,86 0,62
20:2n-6 0,98 0,72 0,4 0,35
20:1n-9 0,69 2,75 2,12 0,68 1,76
20:00 0,07 0,22 0,63 0,32 0,37 0,21
22:4n-6 0,43 1,91 2,12 2,04 2,27 0,69
22:6n-3 7,34 27,81 16,78 15,29 22,9 16
22:5n-3 1,06 3,00 0,81 0,8 1,83
22:00 0,57 0,27 0,38
22:3n-6 1,22 0,9
22:1n-9 1,02
23:00 0,25 0,66 0,18
24:1n-9 0,37 2,62 0,23 1,53
24,0 0,65 0,39
unknow 0,01 0 1,03 1,33 1,25
Tổng 100 100 100 100 100 100
Tổng SFA 51,35 22,69 40,39 47,19 38,12 33,54
Tổng MUFA 35,84 28,96 26,48 21,93 21,77 33,01
Tổng PUFA 12,82 48,35 32,1 30,88 40,11 33,45
n-3 PUFA 12,82 40,98 24,86 19,56 31,42 27,15
Hệ số đánh giá 34,84 606,5 228,46 158,82 321,42 182.17
Ghi chú: B&D: chiết xuất lipid theo phương pháp Blight &Dyer
Kết quả phân tích nêu trên Bảng 4.15 cho thấy:
- Phương pháp sử dụng dung môi hữu cơ (Blight & Dyer) vẫn là phương pháp
hiệu quả nhất trong chiết xuất lipid từ sinh vật biển, cho tổng các axit béo n-3 PUFA

118
là 40,98%, trong khi chiết xuất nhờ protease, hàm lượng n-3 PUFA dao động 19,56-
31,42%, thấp nhất 19,56% khi sử dụng Alcalase, cao nhất 31,42% khi sử dụng
Bromelain. Điều đó cho thấy trong quá trình xử lý với enzyme có thể đã xảy ra hiện
tượng hydro hóa, oxy hóa, hoặc các hiện tượng khác làm biến đổi chất lượng của dầu.
- So với phương pháp chiết xuất truyền thống (ép nhiệt), phương pháp tách
dầu bằng enzyme đã làm tăng chất lượng dầu khi cho tỷ lệ các axit béo không no đa
nối đôi n-3 PUFA tăng cao hơn gấp 2 - 3 lần (từ 10,82% lên 20 - 30%). Có thể thấy
rằng, các axit béo không no đa nối đôi rất nhạy cảm với nhiệt độ. Phương pháp ép
nhiệt diễn ra ở nhiệt độ cao (trên 1000C) đã làm biến đổi chất lượng của lipid, các axit
béo no (SFA) chiếm 51.35%, trong khi phương pháp thủy phân thực hiện ở nhiệt độ
50oC nên ít ảnh hưởng đến chất lượng của dầu.
- So sánh 4 loại enzyme protease sử dụng trong nghiên cứu, các enzyme từ
thực vật (Bromelain và Papain) cho tỷ lệ các axit béo không no n-3 PUFA cao hơn
hai enzyme từ vi sinh vật (Protamex và Alcalase). Các enzyme protease từ thực vật
như papain, bromelain ít được sử dụng hơn các enzyme Protamex, Alcalase trong
công nghệ thủy phân protein để thu axit amin, nhưng lại được nghiên cứu nhiều để
xử lý nguyên liệu trong chiết xuất các loại dầu béo từ cá hay hạt chứa dầu, do các
enzyme này ngoài khả năng xúc tác còn là một dược liệu có khả năng kháng viêm và
chống oxy hóa. Mặt khác, các enzyme này có khả năng chịu nhiệt cao hơn các enzyme
từ vi sinh vật, hạn chế sự phát triển vi sinh vật trong quá trình thủy phân, đồng thời
có tác dụng như là chất chống oxy hóa tự nhiên hạn chế quá trình oxy hóa lipid trong
quá trình thủy phân.
- So với 3 enzyme còn lại, Bromelain cho hệ số đánh giá cao nhất và được lựa
chọn cho các nghiên cứu tiếp theo.
Qua quá trình khảo sát, chúng tôi lựa chọn các điều kiện tối ưu cho quá trình
thủy phân đầu cá ngừ vây vàng để thu nhận dầu béo và axit amin hòa tan như sau: Sử
dụng Bromelain làm enzyme thủy phân; Tỷ lệ nguyên liệu/nước là 1:2; Tỷ lệ
enzyme/cơ chất là 1%; Nhiệt độ thủy phân 50oC; Tốc độ khuấy là 200 vòng/phút;
Thời gian thủy phân là 6 giờ.
4.3.3. Kết quả nghiên cứu ứng dụng enzyme lipase CRL trong quá trình làm

119
giàu các axit béo n-3 PUFA của dầu đầu cá ngừ vây vàng T. albacares bằng
phương pháp ure
Lipid tổng sau khi chiết xuất bằng enzyme Bromelain được thủy phân bởi
enzyme lipase CRL từ nấm men Candida rugosa thành các axit béo tự do. Sau đó, sử
dụng phương pháp tạo phức với Ure/ethanol để làm giàu các axit béo không no đa
nối đôi n-3 PUFA.
4.3.3.1. Khảo sát điều kiện thủy phân lipid thành các axit béo tự do bằng
enzyme lipase CRL
Quá trình thủy bằng lipase CRL trong hệ dung môi hai pha n-hexan và nước
được thực hiện theo phương pháp của Fernádez và cs. (2011). Sau đây là các kết quả
khảo sát một số yếu tố ảnh hưởng đến quá trình này (pH, nhiệt độ, tỷ lệ enzyme/cơ
chất, thời gian thủy phân). Số liệu thực nghiệm được đánh giá dựa trên phương pháp
bình phương tối thiểu.
Ảnh hưởng của pH
Quá trình thủy phân có sự thay đổi giá trị pH từ 5,0 đến 9,0 cho thấy lipase
CRL hoạt động thích hợp trong môi trường kiềm pH 7,5 - 8,5,trong đó pH 8,0 cho
hiệu quả thủy phân cao nhất với sản phẩm có chỉ số axit là 98.4 mg KOH/g và hiệu
suất thủy phân đạt 49.47% (Bảng 4.16a).
Bảng 4.16a. Ảnh hưởng của pH đến quá trình thủy phân lipid thành axit béo tự do
của lipase CRL
TT pH Chỉ số axit (mg KOH/g) Mức độ thủy phân (%)
1 5,0 78,2 39,32
2 5,5 80,5 40,47
3 6,0 84,7 42,58
4 6,5 89,9 45,20
5 7,0 92,5 46,51
6 7,5 90,7 45,60
7 8,0 98,4 49,47
8 8,5 89,7 45,10
9 9,0 82,7 41,58
Phương trình hồi qui (2.1) biểu diễn ảnh hưởng của yếu tố pH có dạng parabol
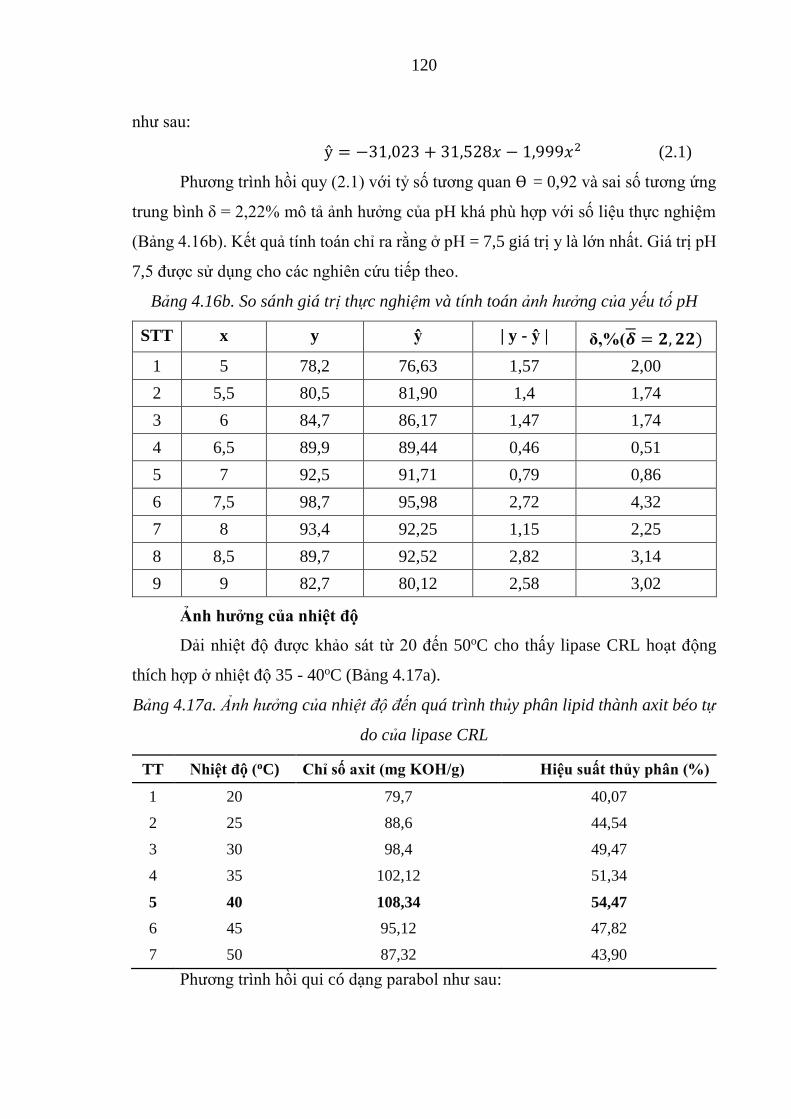
120
như sau:
ŷ = −31,023 + 31,528𝑥 − 1,999𝑥2 (2.1)
Phương trình hồi quy (2.1) với tỷ số tương quan ϴ = 0,92 và sai số tương ứng
trung bình δ = 2,22% mô tả ảnh hưởng của pH khá phù hợp với số liệu thực nghiệm
(Bảng 4.16b). Kết quả tính toán chỉ ra rằng ở pH = 7,5 giá trị y là lớn nhất. Giá trị pH
7,5 được sử dụng cho các nghiên cứu tiếp theo.
Bảng 4.16b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng của yếu tố pH
STT x y ŷ | y - ŷ | δ,%(𝜹 = 𝟐, 𝟐𝟐)
1 5 78,2 76,63 1,57 2,00
2 5,5 80,5 81,90 1,4 1,74
3 6 84,7 86,17 1,47 1,74
4 6,5 89,9 89,44 0,46 0,51
5 7 92,5 91,71 0,79 0,86
6 7,5 98,7 95,98 2,72 4,32
7 8 93,4 92,25 1,15 2,25
8 8,5 89,7 92,52 2,82 3,14
9 9 82,7 80,12 2,58 3,02
Ảnh hưởng của nhiệt độ
Dải nhiệt độ được khảo sát từ 20 đến 50oC cho thấy lipase CRL hoạt động
thích hợp ở nhiệt độ 35 - 40oC (Bảng 4.17a).
Bảng 4.17a. Ảnh hưởng của nhiệt độ đến quá trình thủy phân lipid thành axit béo tự
do của lipase CRL
TT Nhiệt độ (oC) Chỉ số axit (mg KOH/g) Hiệu suất thủy phân (%)
1 20 79,7 40,07
2 25 88,6 44,54
3 30 98,4 49,47
4 35 102,12 51,34
5 40 108,34 54,47
6 45 95,12 47,82
7 50 87,32 43,90
Phương trình hồi qui có dạng parabol như sau:

121
ŷ = 61,988 + 1,616x – 0,029x2 (2.2)
Bảng 4.17b cho thấy phương trình hồi quy (2.2) với tỷ số tương quan ϴ =
0,938 và sai số tương quant rung bình 𝛿 = 3,05% mô tả ảnh hưởng của nhiệt độ khá
phù hợp số liệu thực nghiệm. Kết quả tính cũng chỉ ra ở T = 400C giá trị y là lớn nhất.
Bảng 4.17b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng của yếu tố nhiệt độ
STT x y Ŷ |∆| Δ,%(𝜹 = 𝟑, 𝟎𝟓%)
1 20 79,7 82,77 3,07 3,68
2 25 88,6 90,83 2,23 2,58
3 30 98,4 102,58 4,18 4,71
4 35 102,12 103,06 0,94 1,14
5 40 108,34 109,75 1,41 1,79
6 45 95,12 95,87 0,75 0,99
7 50 87,32 90,56 3,24 4,8
8 55 79,7 82,71 3,01 4,51
Ảnh hưởng tỷ lệ enzyme/cơ chất
Bảng 4.18a. Ảnh hưởng tỷ lệ enzyme/cơ chất đến quá trình thủy phân lipid thành
axit béo tự do của lipase CRL
TT Tỉ lệ E/S Chỉ số axit (mg KOH/g) Hiệu suất thủy phân (%)
1 0,2 88,5 44,49
2 0,4 91,9 46,20
3 0,6 108,3 54,45
4 0,8 106,5 53,54
5 1,0 102,2 51,38
6 1,2 96,78 48,66
7 1,4 90,45 45,48
Tỉ lệ E/S được khảo sát từ 0,2 đến 1,4, kết quả cho thấy tỉ lệ E/S cho hiệu quả
thủy phân cao nhất ở trong khoảng 0,6 đến 0,8. Kết quả tính toán cho phương trình
hồi quy ở dạng parabol như sau:
ŷ = 79,346 + 49,706x - 31,038x2 (2.3)

122
Phương trình hồi quy (2.3) với ϴ = 0,94 và 𝛿=1,09% mô tả khá tốt số liệu thực
nghiệm. Kết quả chỉ ra rằng giá trị ŷ lớn nhất ở tỉ số E/S bằng 0,8 khá phù hợp với
kết quả thực nghiệm.
Bảng 4.18b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng tỷ lệ enzyme/cơ chất
STT x y ŷ |∆| δ
1 0,2 88,5 88,04 0,46 0,51
2 0,4 91,9 94,26 2,36 2,57
3 0,6 108,3 108,89 0,59 0,31
4 0,8 106,5 109,24 2,74 0,76
5 1 102,2 106,01 3,81 0,19
6 1,2 96,78 95,12 1,66 1,76
7 1,4 90,45 92,34 1,89 0,75
Ảnh hưởng thời gian thủy phân
Ảnh hưởng của thời gian tới kết quả thủy phân được biểu diễn trên Bảng 4.19a.
Bảng 4.19a. Ảnh hưởng thời gian thủy phân đến quá trình thủy phân lipid thành
axit béo tự do của enzyme lipase
TT Thời gian (giờ) Chỉ số axit (mgKOH/g) Hiệu suất thủy phân (%)
1 2 78,3 40,47
2 4 98,3 50,80
3 6 119,2 61,60
4 8 126,8 65,53
5 10 140,5 72,61
6 12 149,8 77,42
7 14 147,1 76,02
Phương trình hồi quy có dạng parabol như sau:
ŷ = 42,999 + 13,630x - 0,365x2 (2.4)
Với ϴ = 0,997 và 𝛿 = 1,50%, kết quả tính hoàn toàn phù hợp với thực nghiệm
(Bảng 4.19b). Sau quá trình thủy phân ở 40oC, pH 7,5, tỷ lệ enzyme/cơ chất là 0,8%
và thời gian thủy phân là 12 giờ, chỉ số axit đo được là 149,8 mg KOH/g, tỷ lệ axit
béo đã bị thủy phân là 77,42%.
Bảng 4.19b. So sánh giá trị thực nghiệm và tính toán ảnh hưởng của thời gian thủy phân

123
STT x y ŷ |∆| δ, 𝜹=1.5%
1 2 78,3 79,61 1,31 2,95
2 4 98,3 99,66 1,36 0,84
3 6 119,2 116,14 3,06 2,20
4 8 126,8 124,06 2,74 1,75
5 10 140,5 139,41 1,09 2,42
6 12 149,8 149,20 0,6 0,24
7 14 147,1 149,05 1,95 1,59
Theo kết quả khảo sát của chúng tôi trên cùng nguyên liệu dầu đầu cá ngừ,
cùng loại enzyme lipase CRL, và với các điều kiện phản ứng tương tự, quá trình thủy
phân bởi enzyme lipase CRL trong hệ hai pha dung môi / nước đã cho hiệu quả cao
hơn so với hệ nhũ hóa dầu / nước (số liệu không trình bày ở đây).
Kết quả chúng tôi thu được ở trên có khác biệt so với các kết quả do Bhandari
và cs. công bố năm 2013, trong đó họ đạt hiệu suất thủy phân 86.5% sau 24 giờ phản
ứng ở 35°C [3].
4.3.3.2. Kết quả làm giàu các axit béo n-3 PUFA của dầu đầu cá ngừ vây vàng
bằng phương pháp ure
Các axit béo n-3 PUFA của dầu đầu cá ngừ vây vàng được làm giàu bằng
phương pháp kết tinh với ure trong ethanol dựa trên hiện tượng urê tạo thành tinh thể
với các axit béo bão hòa và kết tủa ở nhiệt độ thấp (Hình 4.12).
Hình 4.12. Tinh thể ure kết hợp với axit béo bão hòa
Ethanol và urê được trộn thành một hỗn hợp đồng nhất với các axit béo. Axit

124
béo sẽ hình thành phức với urê và tủa xuống khi được làm lạnh. Axit béo bão hòa có
mạch thẳng hình thành phức dễ dàng hơn hẳn các axit béo có nối đôi. Số liên kết đôi
càng nhiều thì sự tạo thành phức càng khó khăn hơn. Khi tiến trình kết tinh hoàn
thành, axit béo no và một phần axit béo không no một nối đôi tạo phức, trong khi đó
axit béo đa nối đôi, có phân nhánh, hoặc có vòng sẽ ở lại trong dung dịch.
Kết quả phân tích thành phần axit béo sau khi làm giàu n-3 PUFA được chỉ ra
ở Hình 4.13 và Bảng 4.20.
Hình 4.13. Sắc ký đồ các axit béo sau khi làm giàu
Bảng 4.20. Thành phần axit béo trước và sau khi làm giàu n-3 PUFA
TT Axit béo Trước kết tinh Sau kết tinh
1 14:00 1,86 0,24
2 15:00 0,86 0,08
3 16:1n-9 1,83
4 16:1n-7 3,25
5 16:2n-6 0,53
6 16:00 19,24 4,79
7 17:1n-9 0,79 1,11
8 17:1n-7 0,67 0,47
9 17:00 1,52 0,14
10 18:4n-3 0,36
11 18:2n-6 1,08 1,22
12 18:1n-9 11,88 20,34

125
TT Axit béo Trước kết tinh Sau kết tinh
13 18:1n-7 2,97 4,11
14 18: 2n-3 0,30
15 18:00 11,56 1,26
16 19:00 0,43
17 20:4n-6 4,41 6,85
18 20:5n-3 (EPA) 5,47 8,75
19 20:4n-3 0,86 0,42
20 20:2n-6 0,4 0,29
21 20:1n-9 0,68 0,55
22 20:00 0,37
23 22:4n-6 2,27 0,96
24 22:6n-3 (DHA) 22,9 43,75
25 22:5n-3 (DPA) 1,83 2,54
26 22:0 0,38
27 23:0 0,25
28 24:1n-9 1,53
29 24:0 0,39
Tổng 100 100
Tổng SFA 36,56 5,50
Tổng MUFA 21,77 26,59
Tổng PUFA 40,11 67,48
n-3 PUFA 31,42 58,30
Nhận xét chung:
- Thay vì sử dụng kiềm (NaOH) để thủy phân lipid tổng thành các axit béo tự
do trước khi kết tinh ure, quá trình thủy phân lipid tổng bằng enzyme Lipase CRL có
lợi thế hơn phương pháp hóa học là nhiệt độ thủy phân thấp (400C), tốn ít dung môi
hóa chất và giảm thiểu sự nguy hại đối với môi trường.
- Phương pháp kết tinh bằng ure sau khi thủy phân bằng lipase CRL đã làm
giảm các axit béo no (SFA) từ 36,56% xuống 5,5% và làm giàu n-3 PUFA từ 31,13%
lên 58,3%, trong đó EPA từ 5,47 lên 8,59%, DHA từ 22,9 lên 46,70% và DPA từ 1,83
lên 2,68%.
Các nghiên cứu làm giàu n-3 PUFA của tác giả Linder và cs. (2005) [120] sử

126
dụng enzyme lipase Aspergillus oryzae (Novozym SP 398) để thủy phân dầu từ đầu
cá hồi đã làm giảm các axit béo no SFA từ 27,2% xuống 20,2% và làm giàu các axit
béo không no đa nối đôi PUFA từ 41,6% lên 46,5% trong đó DHA từ 9,9% đến 11,6%
và EPA từ 3,6% đến 5,6%). Phương pháp sử dụng enzyme lipase CRL kết hợp với
kết tinh ure của chúng tôi như nêu ở trên cho sản phẩm có mức độ làm giàu các n-3
PUFA cao hơn.
4.3.4. Đề xuất qui trình công nghệ ứng dụng enzyme kết hợp trong chiết xuất
lipid và làm giàu n-3 PUFA từ đầu cá ngừ vây vàng T. albacares
Các kết quả khảo sát nêu trên cho phép chúng tôi đề xuất một qui trình công
nghệ ứng dụng enzyme kết hợp trong chiết xuất lipid và làm làm giàu n-3 PUFA từ đầu
cá ngừ vây vàng T. albacares . Sơ đồ công nghệ được biểu diễn trên Hình 4.14.
Hình 4.14. Sơ đồ quy trình công nghệ ứng dụng enzyme kết hợp trong chiết xuất lipid
và làm làm giàu n-3 PUFA từ đầu cá ngừ vây vàng.
Thuyết minh qui trình

127
- Nguyên liệu: phụ phẩm đầu cá ngừ đảm bảo không bị ươn, thối, bốc mùi khó
chịu, được cấp đông ở -180C, bảo quản trong kho đông lạnh đến khi chế biến.
- Giai đoạn 1: Sơ chế nguyên liệu
+ Đầu cá ngừ sau khi rã đông được rửa sạch tạp chất, cát sạn.
+ Xay: Mục đích của xay là phá vỡ kết cấu tế bào, giúp cho lipid tự do nằm
trong các tế bào và mô dễ dàng thoát ra ngoài. Tế bào bị phá vỡ làm tăng diện tích tiếp
xúc của nguyên liệu với enzyme để rút ngắn thời gian thủy phân và quá trình thủy phân
được triệt để hơn. Quá trình xay nhỏ được thực hiện trên máy xay trục vít.
+ Diệt khuẩn: Nguyên liệu cho vào thiết bị thủy phân kết cấu kiểu nồi hai vỏ,
có khuấy và điều khiển nhiệt độ. Sau khi bổ sung nước (tỷ lệ nước / nguyên liệu là
2:1), hỗn hợp được gia nhiệt đến 900C trong 10 phút để diệt vi sinh vật có trong
nguyên liệu, hạn chế tối đa khả năng phát triển các vi sinh vật gây bệnh không mong
muốn trong quá trình thủy phân đồng thời chuyển hóa một phần cấu trúc của nguyên
liệu, khiến quá trình thủy phân diễn ra thuận lợi.
- Giai đoạn 2: Thủy phân hỗn hợp đầu cá ngừ
+ Bổ sung enzyme Bromelain theo tỷ lệ 1% so với trọng lượng nguyên liệu,
chỉnh pH đến 6,5; tốc độ khuấy 200 vòng/ phút ở 450C trong 6 giờ. Giữ ổn định nhiệt
độ, tốc độ khuấy trong suốt quá trình thủy phân.
+ Bất hoạt enzyme: Để kết thúc quá trình thủy phân, enzyme được bất hoạt
bằng cách nâng nhiệt hỗn hợp thủy phân lên 900C trong 15 phút.
- Giai đoạn 3: Lọc:
+ Hỗn hợp sau thủy phân đem lọc qua rây để tách xương và phần dịch lọc.
- Giai đoạn 4: Ly tâm:
+ Dịch thủy phân được ly tâm trên hệ thống ly tâm lỏng - lỏng ở áp suất 1atm,
tốc độ 600 l/giờ thu được 3 phần: phần dầu, phần protein hòa tan và cặn protein thô.
+ Phần dầu được lọc rửa bằng nước muối ấm (0,5%; 35 - 40oC) thu được
dầu thô.
+ Phần protein hòa tan được đông khô hoặc sấy phun cho sản phẩm bột đạm
hòa tan giàu axit amin.

128
+ Phần cặn protein không tan (chứa protein chưa thủy phân, peptide không
tan…) được sấy ở nhiệt độ 60 – 70oC cho sản phẩm bột cá không tan dùng trong chăn
nuôi.
- Giai đoạn 5: Thủy phân dầu bằng enzyme lipase
+ Dầu từ đầu cá ngừ cho vào thiết bị đa năng, sau đó bổ sung n-hexan theo tỷ
lệ (1:1). Lượng nước sau khi chỉnh pH được bổ sung vào hỗn hợp với tỷ lệ pha dung
môi / nước (1:7). Sau khi nhiệt độ hỗn hợp đạt 400C, bổ sung enzyme lipase CRL
theo tỷ lệ 0,8%, khuấy đều hỗn hợp. Giữ ổn định nhiệt độ 400C trong 10 giờ, tốc độ
khuấy 200 vòng/phút. Chỉ số axit được xác định trong quá trình thủy phân để kiểm
soát và đánh giá hiệu suất quá trình thủy phân.
- Giai đoạn 6: Lọc rửa các axit béo
+ Sau quá trình thủy phân, hỗn hợp các axit béo được lọc qua giấy lọc
Whatman No.2 để loại bỏ enzyme. Sau khi rửa với nước ấm, hỗn hợp axit béo được
làm khan với Na2SO4, cô quay loại dung môi.
- Giai đoạn 7: Kết tinh ure
+ Hỗn hợp axit béo được hòa tan trong dung môi theo tỷ lệ ure / cồn (3:16),
FFA / ure (1:3). Sau đó, dung dịch thu được để kết tinh ở nhiệt độ 40C trong thời gian
là 18 - 20 giờ. Sau quá trình kết tinh, hỗn hợp pha rắn và pha lỏng được lọc qua giấy
lọc tại nhiệt độ kết tinh, rửa lại bằng dung môi có nhiệt độ kết tinh, sau đó tách pha
rắn và pha lỏng.
Các sản phẩm của qui trình:
Các sản phẩm của qui trình gồm: axit béo giàu n-3 PUFA, bột đạm hòa tan
giàu axit amin và bột cá không tan dùng trong chăn nuôi. Thành phần hóa học của
sản phẩm axit béo giàu n-3 PUFA được thể hiện trên Bảng 4.20, các chỉ tiêu hóa lý
được chỉ ra trên Bảng 4.21a. Thành phần axit amin tự do của sản phẩm bột đạm hòa
tan giàu axit amin được chỉ ra ở Bảng 4.21b.

129
Bảng 4.21a. Các chỉ tiêu hóa lý của sản phẩm axit béo giàu n-3 PUFA
Tỷ trọng d20 0,85-0,87 g/ml
Điểm đông đặc 18- 20 oC
Chỉ số khúc xạ 1,458
Chỉ số axit < 0,5 mg KOH/g
Chỉ số peroxit 9,86
Chỉ số iot 145 g I/100 g
Chỉ số xà phòng hóa 160 mg KOH/g
Bảng 4.21b. Các thành phần axit amin tự do của sản phẩm bột đạm hòa tan giàu
axit amin
TT Axit amin Hàm lượng
(µg/100 g) TT Axit amin
Hàm lượng
(µg/100 g)
1 Threonin* 4524 10 Serine 4979.5
2 Valine* 29267 11 Glutamic
acid 6201.3
3 Methionin* 33615 12 Proline 7020
4 Isoleucine* 62825 13 Glycine 4439.5
5 Leucine* 9234.5 14 Alanine KPH
6 Phenylalanine* 46276 15 Tyrosine 71267
7 Tryptophan* 16 Lysine 37661
8 Histidine* 46497 17 Arginin 55952
9 Aspatic acid 4552.1
Tổng axitmin tự do 424 309.84
Axit amin thiết yếu* 232 237.4
Axit amin không thiết yếu 192 072.5
Tóm lại, quy trình chiết xuất lipid và làm giàu các n-3 PUFA ngoài sản phẩm
axit béo không no tương đương với các sản phẩm nhập ngoại (ví dụ DHA selco) còn
cho các sản phẩm có giá trị gia tăng là bột đạm hòa tan giàu axit amin thiết yếu
(232,24 mg/100 g sản phẩm) và bột cá dùng trong chăn nuôi. Qui trình công nghệ này
cho phép tận thu triệt để nguồn nguyên liệu phụ phẩm đầu cá ngừ, hạn chế ô nhiễm
môi trường. Riêng phương pháp chiết xuất dầu béo từ đầu cá ngừ vây vàng đã được
chúng tôi đăng ký giải pháp hữu ích.

130
KẾT LUẬN
Trong luận án này, một số enzyme được nghiên cứu ứng dụng nhằm hỗ trợ
cho quá trình chiết xuất tinh dầu từ cành lá, từ vỏ quế Cinnamomum cassia và từ gỗ
gió bầu Aquilaria crassna cũng như quá trình chiết xuất dầu béo và làm giàu các
axit n-3 PUFA từ đầu cá ngừ vây vàng Thunnus albacares đã thu được kết quả như
sau:
1. Luận án đã thành công khi sử dụng hệ enzyme Laccase-Htec2 để xử lý các
nguyên liệu cành lá và vỏ quế trước khi chưng cất cho hiệu suất thu hồi tinh dầu tăng
lên rõ rệt, tỷ lệ gia tăng tinh dầu cành lá quế đạt 41,7%, vỏ quế đạt 14,57%, thời gian
chưng cất tinh dầu được rút ngắn. Đặc biệt, quá trình xử lý bằng hệ enzyme Laccase-
Htec2 đã làm gia tăng hàm lượng chất chính cinnamaldehyde trong tinh dầu cành lá
quế từ 69,74% lên 85,60%, tinh dầu vỏ quế từ 54,63% lên 68,37% và không làm thay
đổi một số hoạt tính sinh học của tinh dầu cành lá quế C.cassia so với đối chứng.
2. Luận án đã sử dụng phương pháp qui hoạch thực nghiệm để tính toán điều
kiện tối ưu cho quá trình thủy phân cành lá quế trước khi chưng cất ở pH 5,2, nhiệt
độ 44oC, tỷ lệ Laccase/cơ chất 0,42 ml/g, tỷ lệ Htec2/cơ chất 1,15%, thời gian thủy
phân 5 giờ 30 phút. Trên cơ sở các kết quả thu được, một qui trình công nghệ chưng
cất tinh dầu cành lá quế có enzyme hỗ trợ đã được đề xuất.
3. Luận án đã thử nghiệm thăm dò ứng dụng của hệ enzyme Laccase-Htec2 hỗ
trợ quá trình chưng cất tinh dầu trầm hương từ gỗ cây gió bầu Aquilaria crassna cho
tỷ lệ gia tăng tinh dầu là 33,33%, các thành phần có giá trị của tinh dầu (cis-
dihydroagarofuran, α-agarofuran, agarospirol, hinesol, và neopetasone) tăng lên, các
thành phần axit béo gây cản trở giảm đi rõ rệt.
4. Luận án đã thành công khi chiết xuất lipid và làm giàu n-3 PUFA từ đầu cá
ngừ vây vàng Thunnus albacares bằng hệ enzyme kép Bromelain và Lipase CRL. Qui
trình chiết xuất lipid bằng enzyme Bromelain cho hiệu suất chiết xuất lipid 69%, hàm
lượng n-3 PUFA 31,42%. Qui trình làm giàu n-3 PUFA bằng enzyme Lipase CRL kết
hợp kết tinh ure đã tăng hàm lượng n-3 PUFA từ 31,42% lên 65,08%. Từ đó, cho phép
đề xuất một qui trình công nghệ ứng dụng kết hợp hai enzyme Bromelain và Lipase

131
CRL trong việc chiết xuất lipid và làm giàu n-3 PUFA từ đầu cá ngừ vây vàng. Ngoài
dầu cá giàu axit béo n-3 PUFA đạt chất lượng thương phẩm, qui trình này còn cho các
sản phẩm có giá trị gia tăng, tận dụng triệt để nguồn nguyên liệu đồng thời giảm thiểu
dung môi hóa chất và ô nhiễm môi trường.
5. Các kết quả nghiên cứu nêu trên là các nghiên cứu ứng dụng enzyme đầu
tiên ở Việt Nam trong việc chiết xuất tinh dầu từ cành lá, vỏ quế C. cassia và từ gỗ
gió bầu A. crassna. Qui trình ứng dụng Bromelain trong việc chiết xuất lipid từ đầu
cá ngừ vây vàng T. albarcares đã được đăng ký giải pháp hữu ích.

132
KIẾN NGHỊ
1. Quy trình chưng cất tinh dầu cành lá quế, vỏ quế nhờ enzyme hỗ trợ bước
đầu cho ưu thế hơn so với phương pháp thông thường trên phương diện gia tăng hiệu
suất, nâng cao chất lượng tinh dầu, giảm thời gian chưng cất và vẫn giữ nguyên được
hoạt tính sinh học... chỉ dừng lại ở qui mô thí nghiệm. Cần thử nghiệm ở qui mô sản
xuất tại địa phương nhằm ổn định công nghệ và chất lượng sản phẩm, đồng thời đánh
giá chi phí tiêu hao năng lượng lẫn giá thành nhằm nâng cao tính ứng dụng khi áp
dụng vào thực tiễn sản xuất.
2. Do nhu cầu cấp bách của thực tiễn sản xuất, phương pháp xử lý gỗ gió bầu
trước khi chưng cất cần được nghiên cứu gấp và sâu hơn.
3. Các kết quả nghiên cứu của luận án cho thấy việc ứng dụng enzyme hỗ trợ
các quá trình chiết xuất các hợp chất thiên nhiên mang lại hiệu quả rõ rệt. Vì thế cần
mở rộng phạm vi nghiên cứu sang các loại nguyên liệu khác như tảo biển và các lớp
hợp chất thiên nhiên có hoạt chất sinh học có ứng dụng trong thực phẩm, dược phẩm,
y học.

133
DANH MỤC CÁC CÔNG TRÌNH CÔNG BỐ CỦA TÁC GIẢ
LIÊN QUAN ĐẾN LUẬN ÁN
1. Hoang Thi Bich, Le Tat Thanh, Tran Quoc Toan, Nguyen Huy Tung, Le Xuan
Duy, Nguyen Van Tuyen Anh, Dinh Thi Thu Thuy, Tran Thi Tuyen, Do
Trung Sy, Le Mai Huong, Nguyen Quyet Chien (2016), “Investigation of
enzyme treatments to assist extraction of essential oil from the leaves and
branches of Cinnamomum cassia collected in Yenbai province”, Journal
of Science and Technology, 54 (2C), pp.479-485.
2. Le Tat Thanh, Le Xuan Duy, Tran Quoc Toan, Hoang Thi Bich, Pham Thi Bien,
Pham Thu Hue, Dinh Nguyet Thu, Pham Quoc Long (2016), “Survey on
total lipid content and composition of fatty acids from head and viscera of
tuna”, Journal of Science and Technology, 54 (2C), pp.486-492.
3. Lê Tất Thành, Hoàng Thị Bích, Trần Quốc Toàn, Nguyễn Thị Biển, Nguyễn Quang
Tùng (2016), “Ảnh hưởng của một số yếu tố tới quá trình thủy phân dầu đầu
cá ngừ bằng enzyme lipase từ Candida rugosa”, Tạp chí Khoa học và Công
nghệ-trường Đại học Công nghiệp Hà Nội, (số 36-2016), tr.67-70.
4. Hoàng Thị Bích, Nguyễn Văn Tuyến Anh, Phạm Minh Quân, Phạm Thu Huế,
Nguyễn Quang Tùng, Tô Xuân Thắng, Trần Quốc Toàn, Lê Tất Thành
(2017), “Xây dựng quy trình ứng dụng enzyme và các kĩ thuật phối hợp
để thu nhận dầu cá giàu các axit béo không no đa nối đôi EPA, DHA, DPA
từ phụ phẩm chế biến cá ngừ vây vàng Thunnus albacares”, Tạp chí Khoa
học và Công nghệ Biển, tập 17, (01), 2017, tr.95-102.
5. Hoàng Thị Bích, Nguyễn Quyết Chiến, Lê Tất Thành, Đinh Thị Thu Thủy, Đỗ Thị
Thảo, Hoàng Kim Chi, Trần Thị Như Hằng, Trần Thị Hồng Hà, Lê Mai Hương
(2017), “Khảo sát hoạt tính sinh học tinh dầu cành lá quế cinnamomum cassia
thu nhận từ phương pháp enzyme kết hợp cất cuốn hơi nước”, Tạp chí Dược
học, tháng 5/2017, (493), tr.12-15.
6. Hoang Thi Bich, Le Mai Huong, Nguyen Quyet Chien, Đinh Thi Thu Thuy, Le Tat
Thanh, Đo Trung Sy, Pham Hong Hai (2017), “Optimization of
lignocellulose hydrolysis by (Laccase and Cellic Htec2) enzyme system
assisted essential oil distillation from Cinnamomum cassia leaves and
branches”, Journal of Science and Technolog,y (Chấp nhận đăng 01/2017).

134
DANH MỤC TÀI LIỆU THAM KHẢO
Tài liệu tiếng Việt
1. Bộ Khoa học Công nghệ và Môi trường (1990), Tiêu chuẩn Việt Nam, Hà Nội.
2. Bộ Y tế (2009), Dược điển Việt Nam IV, Nxb Hà Nội.
3. Trương Thị Mỹ Chi, Nguyễn Thị Thu Hương (2010), Nghiên cứu tác dụng chống
oxi hóa in vitro và in vivo của một số loài nấm linh chi (Ganoderma
lucidum) và nấm vân chi (Trametes versicolor), Tạp chí Y học thành phố
Hồ Chí Minh, tập 14, (02), tr.135-141.
4. Nguyễn Văn Chung (2004), Nghiên cứu hoàn thiện công nghệ và thiết bị chưng
cất và trích ly nhựa dầu hồ tiêu có sử dụng enzyme, Đề tài KC 04-07.
5. Lê Quang Diễn, Doãn Thái Hòa (2013), “Ứng dụng enzyme trong sản xuất bột
giấy hiệu suất cao từ thân cây ngô - Phần I, Hiệu quả của lignin
peroxidase”, Tạp chí Khoa học và Công nghệ các trường Đại học kỹ thuật,
(92), tr.142-146.
6. Nguyễn Lân Dũng, Nguyễn Đình Quyến, Phạm Văn Ty (1998), Vi sinh vật học,
Nxb Giáo dục, Hà Nội, 537 tr.
7. Phạm Hồng Hải, Ngô Kim Chi (2007), Xử lí số liệu và quy hoạch thực nghiệm trong
nghiên cứu hóa học, Nxb Khoa học tự nhiên và Công nghệ, Hà Nội.
8. Nguyễn Ngọc Hạnh, Mai Thành Chí (2006), Nghiên cứu quy trình công nghệ chiết
xuất tinh dầu tiêu, quế và trầm bằng CO2 lỏng siêu tới hạn và sản xuất
piperine từ phế phẩm của tiêu, Tp Hồ Chí Minh.
9. Phạm Hoàng Hộ (1999), Cây cỏ Việt Nam, quyển I, Nxb Trẻ, Tp. Hồ Chí Minh.
10. Đặng Diễm Hồng (2016), Nghiên cứu qui trình tách chiết dầu sinh học giàu axit béo
omega 3 và omega 6 (EPA, DHA, DPA) từ sinh khối vi tảo biển dị dưỡng,
Chương trình công nghệ sinh học trong chế biến, Bộ Công thương.
11. Lại Mai Hương (2004), Khảo sát thành phần lipid trong một số loài cá biển Việt
Nam và nghiên cứu phương pháp làm giàu acid béo không no từ phế phụ
phẩm của ngành chế biến hải sản, Đề tài của Trường Đại học Bách Khoa
Tp Hồ Chí Minh.

135
12. Nguyễn Thị Mỹ Hương (2011), “Sử dụng sản phẩm thuỷ phân protein từ đầu cá
ngừ trong thức ăn cho tôm”, Tạp chí khoa học công nghệ thuỷ sản, Đại
học Nha Trang, (1), tr.99-108.
13. Văn Ngọc Hướng (2003), Hương liệu và ứng dụng, Nxb Khoa học và kỹ thuật,
Hà Nội.
14. Lê Gia Hy, Đặng Tuyết Phương (2010), Enzyme vi sinh vật và chuyển hóa sinh học -
Nguyên lý và ứng dụng, Nxb Khoa học tự nhiên và Công nghệ, Hà Nội.
15. Dương Minh Lam, Trương Thị Chiên (2013), “Nghiên cứu một số đặc tính sinh
học của chủng nấm đảm Trametes maxima CPB30 sinh laccase ứng dụng
trong xử lí màu nước ô nhiễm do thuốc nhuộm”, Tạp chí sinh học, 35(4),
tr.477-483.
16. Phạm Quốc Long, Châu Văn Minh (2005), Lipid và các axit béo hoạt tính sinh
học có nguồn gốc thiên nhiên, Nxb Khoa học và Kỹ thuật, 213tr.
17. Phạm Quốc Long (chủ nhiệm) (2013), Nghiên cứu quy trình công nghệ sản xuất
thuốc chữa bệnh viêm khớp dạng thấp và một số thực phẩm chức năng từ
nguyên liệu sinh vật biển, Đề tài Bộ Công thương, Hà Nội.
18. Vũ Ngọc Lộ, Đỗ Chung Võ, Nguyễn Mạnh Pha, Lê Thúy Hạnh (1996), Những
cây tinh dầu Việt Nam (Khai thác, chế biến, ứng dụng), Nxb Khoa học và
Kỹ thuật, Hà Nội, 181tr.
19. Đỗ Tất Lợi (2006), Những cây thuốc và vị thuốc Việt Nam, Nxb Y học - Nxb
Thời đại, Hà Nội.
20. Nguyễn Đức Lượng (1996), Nghiên cứu tính chất một số vi sinh vật có khả năng
tổng hợp cellulase cao và ứng dụng chúng trong công nghệ xử lý chất thải
hữu cơ, Luận án phó tiến sĩ Khoa học và kỹ thuật, Trường Đại học Bách
khoa Hà Nội.
21. Lã Đình Mỡi, Lưu Đàm Cư, Trần Minh Hợi, Nguyễn Thị Thủy, Nguyễn Thị
Phương Thảo, Trần Huy Thái, Ninh Khắc Bản (2001), Tài nguyên thực
vật có tinh dầu ở Việt Nam, tập 1, Nxb Nông nghiệp Hà Nội.
22. Nguyễn Văn Mùi (2001), Thực hành hóa sinh học, Nxb Đại học Quốc gia Hà
Nội, 173tr.

136
23. Nguyễn Thị Minh Nguyệt, Trần Nguyễn Mỹ Châu, Võ Bửu Lợi, Đặng Thị Thanh
Hương (2009), Nghiên cứu sử dụng enzyme trong chiết tách dầu béo và
các thành phần của cám gạo, Đề tài Bộ Công thương, Hà Nội.
24. Thái Lâm Phát (2008), Nghiên cứu tinh luyện dầu cá, Luận văn thạc sỹ, Trường
Đại học Bách Khoa TP. Hồ Chí Minh.
25. Nguyễn Kim Phi Phụng (2007), Phương pháp cô lập hợp chất hữu cơ, Nxb Đại
học quốc gia Hồ Chí Minh.
26. Đào Mạnh Sơn, Nguyễn Viết Nghĩa (2006), “Hiện trạng nguồn lợi và tình hình
khai thác cá ngừ đại dương ở Việt Nam”, Tạp chí Thủy sản, (7), tr.20 -
23.
27. Đỗ Trung Sỹ, Hoàng Thị Bích, Đỗ Quang Kháng, Ngô Quốc Anh (2015),
“Nghiên cứu phương pháp thủy phân phế thải rong nâu sử dụng kết hợp
acid và enzyme”, Tạp chí Hóa học, 54 (2e), tr.121-127.
28. Đỗ Trung Sỹ, Ngô Quốc Anh, Hoàng Thị Bích, Trần Quốc Toàn, Lê Tất Thành,
Nguyễn Huy Tùng, Đặng Thu Thảo, Lê Mai Hương (2014), Xác định hàm
lượng đường glucose trong dịch thủy phân cellulose có trong rong biển
bởi nấm mốc ASPERGILLUS TERRIUS AF67, Hội nghị khoa học toàn
quốc về sinh học biển và phát triển bền vững lần thứ hai (2014), tr.711-
721.
29. Đỗ Trung Sỹ, Trần Đình Toại, Lê Tất Thành, Hoàng Thị Bích, Nguyễn Văn
Tuyến Anh, Nguyễn Đình Tuyến, Phạm Quốc Long (2012), Nghiên cứu
quy trình thủy phân cá tạp và phế liệu thủy sản bằng enzyme để sản xuất
sản phẩm cao đạm giàu axit amin ứng dụng trong y, dược, Hội nghị quốc
tế Biển Đông 2012, Nha Trang, 12-14/9/2012, tr.520-528.
30. Lê Tất Thành, Hoàng Thị Bích, Trần Quốc Toàn, Nguyễn Thị Biển, Nguyễn
Quang Tùng (2016), Ảnh hưởng của một số yếu tố tới quá trình thủy phân
dầu đầu cá ngừ bằng enzyme lipase từ Candida rugosa.
31. Trần Đình Thắng (2016), Hợp chất thiên nhiên, Nxb Đại học Vinh.
32. Đặng Thị Thu, Lê Ngọc Tú, Tô Kim Anh, Phạm Thu Thủy, Nguyễn Xuân Sâm
(2004), Công nghệ enzyme, Nxb Khoa học và Kỹ thuật, Hà Nội, 304 tr.

137
33. Đặng Thị Thu, Ngô Tiến Hiền, Quyền Đình Thi, Phùng Thị Thủy, Hoàng Lan,
Đỗ Biên Cương, Thiều Linh Thùy Và Nguyễn Thị Sánh (2004), Nghiên
cứu công nghệ sản xuất một số loại dầu béo bằng lipaza, đề tài nhành của
nghiên cứu cấp Nhà nước, mã số: KC 04-07, Viện Công nghệ Thực phẩm,
Hà Nội.
34. Lưu Thị Lệ Thủy (chủ nhiệm) (2008), Nghiên cứu xây dựng qui trình công nghệ
sản xuất dầu từ hạt bí đỏ bằng phương pháp enzyme, Đề tài Bộ Công
thương, Hà Nội.
35. Trần Đình Toại, Trần Thị Hồng (2007), “Tương lai ứng dụng Enzyme trong xử
lý phế thải”, Tạp chí Khoa học Đại học Quốc gia Hà Nội, Khoa học Tự
nhiên và Công nghệ, (23), tr.75-85
36. Trần Đình Toại, Phạm Hồng Hải, Nguyễn Bá Kiên, Hoàng Thị Bích, Đỗ Trung
Sỹ (2011), “Nghiên cứu tối ưu hoá quá trình thuỷ phân cellulose tách từ
rơm rạ thành đường tan của nấm mốc Aspergillus terrius để sản xuất
ethanol- nhiên liệu sinh học”, Tạp chí Khoa học và Công nghệ, (6), tr.38-
42.
37. Trung tâm Thông tin thủy sản (2012), Báo cáo tình hình sản xuất 8 tháng đầu
năm 2012, Hà Nội.
38. Nguyễn Anh Tuấn (2011), Tận dụng nguyên liệu còn lại trong chế biến thủy sản,
Báo cáo hội thảo.
Tài liệu tiếng Anh
39. Abdul Khalil H.P.S., Siti Alwani, M., and Mohd Omar, A. K (2006), Cell walls
of tropical fibers, BioResources 1(2), pp.220-232.
40. Ademola Monsur Hammed A. M, Irwandi Jaswir, Azura Amid, Zahangir Alam,
Tawakalit Tope, Asiyanbi-H & Nazaruddin Ramli (2013), Enzymeatic
Hydrolysis of Plants and Algae for Extraction of Bioactive Compounds,
Food Reviews International, 29(4), pp.352-370.
41. Akoh C.C and Min D.B (2008), Food lipids: chemistry, nutrition, and
biotechnology, 3th ed., CRC Press, Taylor and Francis Group.

138
42. Ali Bougatef, Rafik Balti, Anissa Haddar, Kemel Jellouli, Nabil Souissi, and
Moncef Nasri (2012), Protein Hydrolysates from Bluefin Tuna (Thunnus
thynnus) Heads as Influenced by the Extent of Enzymeatic Hydrolysis,
Biotechnology and Bioprocess Engineering 17, pp.841-852.
43. Ali SM, Khan AA, Ahmed I, Musaddiq M, Ahmed KS, Polasa H, et al (2005),
Antimicrobial activities of eugenol and cinnamaldehyde against the human
gastric pathogen Helicobacter pylori, Ann Clin Microbiol Antimicrob; 4, pp.20.
44. Anastas, Paul T.; Warner, John C. (1998). Green chemistry: theory and practice.
Oxford [England]; New York: Oxford University Press.
45. Asger M., Sharif Y., and Bhatti H. N. (2010), Enhanced production of ligninolytic
enzymes by Ganoderma lucidum IBL-06 using lignocellulosic agricultural
wastes, Inter. J. Chem. Reactor Eng. 8 (1), pp.1-17.
46. Ashgari J., Touli C. K. and Mazaheritehrani M. (2010), Microwave assisted
hydrodistillation of essential oils from Echinophora platyloba DC, Journal
of Medicinal Plants Research, vol. 6 (28), pp. 4475-4480, 2010.
47. Awika, J.M. et al. (2003), Screening methods to measure antioxidant activity of
sorghum (Sorghum bicolor) and sorghum products, J. Agric. Food Chem,
(51), pp.6657-6662.
48. Bailey M.J., Biely P.and Poutanen Kaisa (1992), Interlaboratory testing of methods
for assay of xylanase activity, Journal of Biotechnology, (23), pp.257-270.
49. Bakker, M.E., Gerritsen, A.F., (1989), A suberized layer in the cell wall of
Mucilage cells of Cinnamomum, Ann. Bot. 63, 441-448.
50. Bradford, MM. A rapid and sensitive for the quantitation of microgram
quantitites of protein utilizing the principle of protein-dye binding.
Analytical Biochemistry 72: 248-254. 1976.
51. Baldrian P. (2006), Fungal laccases - occurrence and properties, FEMS
Microbiol Rev 30(2): 215-42.
52. Bhandari K., Chaurasia S.P & Dalai A.K (2013), Hydrolysis of tuna fish oil using
candida rugosa lipase for producing fatty acids containing DHA,
International Journal of Applied and Natural Sciences (IJANS), Vol. 2,
Issue 3, July 2013, pp.1-12.

139
53. Bickers D, Calow P, Greim H, Hanifin JM, Rogers AE, Saurat JH, et al (2005),
A toxologic and dermatologic assessment of cinnamyl alcohol,
cinnamaldehyde and cinnamic acid when used as fragrance ingredients.
The RIFM expert panel. Food Chem Toxicol; 43, pp.799-836.
54. Biesalski, H.K. et al. (2009), Bioactive compounds: definition and assessment of
activity. Nutrition 25, 1202-1205, Denny, A.R. and Buttriss, J.L. (2007)
Plant Foods and Health: Focus on Plant Bioactives, EuroFIR Synthesis
Report No. 4, EuroFIR Project Management Office/British Nutrition
Foundation, London.
55. Blanchette R.A. (1991), Delignification by wood-decay fungi, Annu. Rev,
Phytopathol, (29), pp.381-398.
56. Blight E. G., and Dyer W. J, (1959), A rapid method of total lipid extration and
purification, Canad. J. Biochem. Physiol, 37, pp. 911.
57. Bougatef A., Balti R., Haddar A., Jellouli K., Nabil Souissi, and Moncef Nasri
(2012), Protein Hydrolysates from Bluefin Tuna (Thunnus thynnus) Heads
as Influenced by the Extent of Enzymeatic Hydrolysis, Biotechnology and
Bioprocess Engineering 17: pp.841-852.
58. Boulila A., Hassen I., Haouari L., Mejri F., Amor I. B., Casabianca H., Hosni K.
(2015), Enzyme-assisted extraction of bioactive compounds from bay
leaves (Laurus nobilis L.), Industrial Crops and Products 74, pp.485-493.
59. Bradford M.M. (1976), Rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding, Anal. Biochem., (72), pp.248-254.
60. Blois, M.S. (1958), Antioxidant Determinations by the Use of a Stable Free
Radical. Nature, 181, 1199-1200..
61. Calinescu, I., Gavrila, A.I., Ivopol, M., Ivopol, G.C., Popescu, M., Mircioaga, N.,
(2014), Microwave assisted extraction of essential oils of enzymeatically
pretreated lavender (Lavandula angustifolia Miller), Cent. Eur. J. Chem,
(12), pp.829-836.

140
62. Camarero, S.; Ibarra, D.; Martínez, M.J.; Martinez, A.T (2005), Lignin-Derived
Compounds as Efficient Laccase Mediators for Decolorization of
Different Types of Recalcitrant Dyes, Appl. Environ. Microbiol, 71),
pp.1775-1784.
63. Camarero, S.; Martínez, M.J.; Martínez, A.T (2014), Understanding lignin
biodegradation for the improved utilization of plant biomass in modern
biorefineries, Biofuels Bioprod. Biorefin, (8), pp.615-625.
64. Cater, C.M., R.D. Hagenmaier, and K.F. Mattil (1974), Aqueous extraction- an
alternative oilseed milling process, Journal of the American Oil Chemists'
Society, 1974. 54(4): p. 137-141.
65. Cintra, O., Lopez-Munguia, A. and Vernon, J. (1986), Coconut oil extraction by
a new enzymeatic process, J. Food Sci., (51), pp.695-697.
66. Coussens, L.M.; Werb, Z (2002), Inflammation and cancer, Nature 2002, 420,
860-867.
67. Chang, K.-S., Tak, J.-H., Kim, S.-I., Lee, W.-.J., Ahn, Y.-J., (2006), Repellency
of Cinnamomum cassia bark compounds and cream containing cassia oil
to Aedes aegypti (Diptera: Culicidae) under laboratory and indoor
conditions. Pest Manag. Sci. 62, 1032-1038.
68. Chang, S.-T., Chen, P.-F., Chang, S.-C., (2001), Antibacterial activity of leaf
essential oils and their constituents from Cinnamomum osmophloeum. J.
Ethnopharmacol. 77, 123-127.
69. Chao L, Yang RF, Fu XF (2008), Ultrasonic-assisted supercritical CO2
extraction of volatile oil from compound recipe of clove and cinnamon, J
S China Univ Technol 36:67-71.
70. Chao LK, Hua K-F, Hsu H-Y et al (2005), Study on the antiinflammatory activity
of essential oil from leaves of Cinnamomum osmophloeum, J Agric Food
Chem 53:7274-7278
71. Cheng, S.-S., Chua, M.-T., Chang, E.-H., Huang, C.-G., Chen, W.-J., Chang, S.-
T., (2009), Variation in insecticidal activity and chemical compositions of
leaf essential oils from Cryptomeria japonica at different ages, Bioresour.
Technol. 100, 465-470.

141
72. Chiozza L. (1856), On the artificial production of cinnamon oil, Comptes rendus
(in French), (42), pp. 222-227.
73. Choi, J, Lee, K.-T., Ka, H., Jung, W.-T., Jung, H.-J., Park, H.-J., (2001),
Constituents of the essential oil of the Cinnamomum cassia stem bark and
the biological properties.
74. Chow Ching K. (2008), Fatty acids in foods and their health implication 3th ed.,
CRC Press, Taylor and Francis Group.
75. Chu, Q.-G., Hu, Z.-H., (1999), Comparative anatomy of oil cells and mucilage
cells in the leaves of the Lauraceae in China. Acta Phytotaxon, Sin. 37 (6),
529-540 (In Chinese with an English abstract).
76. Denys J. Charles (2013), Antioxidant Properties of Spices, Herbs and Other
Sources, Norway, IA, USA, 589 pp.
77. Ding Z., Peng L., Chen Y., Zhang L., Gu Z., Shi G. and Zhang K. (2012),
Production and characterization of thermostable laccase from the
mushroom, Ganoderma lucidum, using submerged fermentation, African
Journal of Microbiology Research 6 (6), pp. 1147-1157.
78. Dobozi, S., Halasz, A., Kovacs, K. E., & Szacks, G. (1988), Enhancement of
mustard oil yield by cellulolytic pretreatment, Applied Microbiology
Biotechnology, 29(1), pp. 39-43
79. Doi R. H. (2008), Cellulases of Mesophilic Microorganisms: Cellulosome and
Noncellulosome Producers, Ann. N. Y. Acad. Sci. 1125, pp.267-279.
80. Domadia P., Swarup S., Bhunia A., Sivaraman J., Dasgupta D. (2007), Inhibition
of bacterial cell division protein FtsZ by cinnamaldehyde, Biochemical
pharmacology 74, pp.831-840.
81. Dominquez, H., Nunez, M. J. and Loma, Y. M. (1994), Enzymeatic treatment to
enhance oil extraction from fruits and oil seeds: a review, Food Chem,
(49), pp.271-286.
82. Dumas J., Peligot E. (1834), On cinnamon oil, the hippuric acid and sebacic acid,
Organic chemistry reseach, (57), pp.305-34.

142
83. Dumay, J., Donnay-Moreno, C., Barnathan, G., Jaouen, P., Bergé, J.P., (2006),
Improvement of lipid and phospholipid recoveries from sardine (Sardina
pilchardus) viscera using industrial proteases, Process Biochemistry 41,
pp.2327-2332.
84. Eur J Gastroenterol Hepatol (1996), Jones FA. Herbs-useful plants. Their role in
history and today.;8, pp.1227-31.
85. Farid Chemat and Jochen Strube (Eds.) (2015), Green extraction of natural
products: theory and practice.
86. Farid Chemat and Maryline Abert Vian (Eds) (2016), Alternative solvents for
Natural products extraction.
87. Farnet AM, Gil G, Ferre E. (2008), Effects of pollutants on laccase activities of
Marasmius quercophilus, a white-rot fungus isolated from a
mediterranean schlerophyllous litter, Chemosphere 70(5): 895-900.
88. Fernandez G. L, Betancor L., Alfonso V., Carrascosa J., Guisan J.M (2011),
Release of Omega 3 Fatty acids by the Hydrolysis of fish oil catalyzed by
Lipase Immobilized on hydrophobic supports, J. Am Oil Chem Soc 88,
pp.173-1178.
89. Folin, O. and Ciocalteau, P. (1927), J.Biol.Chem, 73, pp.627.
90. Freese, S. and Binning, R. (1993), Enzymeatic extraction of spice aromas,
Gordian 93(11), pp.171-175.
91. G. Joana Gil-Chávez, José A. Villa, J. Fernando Ayala-Zavala, J. Basilio Heredia,
David Sepulveda, Elhadi M. Yahia,and Gustavo A. González Aguilar
(2013), Technologies for Extraction and Production of Bioactive
Compounds to be Used as Nutraceuticals and Food Ingredients:An
Overview, Food Science and Food Safety, Vol.12.
92. Gámez-Meza, N., et al., (2003), Concentration of eicosapentaenoic acid and
docosahexaenoic acid from fish oil by hydrolysis and urea complexation,
Food Research International 36, pp.721-727.
93. Gardossi, L. et al. (2009), Guidelines for reporting of biocatalytic reactions,
Trends Biotechnol. 28, pp.171-180.

143
94. Golmakani, M. T., & Rezaei, K. (2008), Comparison of microwave-assisted
hydrodistillation with the traditional hydrodistillation method in the
extraction of essential oils from Thymus vulgaris L., Food Chemistry,
(109), pp.925-930.
95. Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases
(2010), A never-ending story. Cell. Mol. Life Sci, (67), pp.369-385.
96. Hamidpour R., Hamidpour M., Hamidpour S., Shahlari M. (2015), Cinnamon
from the selection of traditional applications to its novel effects on the
inhibition of angiogenesis in cancer cells and prevention of Alzheimer's
disease, and a series of functions such as antioxidant, anticholesterol,
antidiabetes, antibacterial, antifungal, nematicidal, acaracidal, and
repellent activities, Journal of Traditional and Complementary Medicine
5, pp.66-70.
97. Hammed A.M, Jaswir I., Amid A., Alam Z., Asiyanbi T.T, ramli N. (2013),
Enzymeatic Hydrolysis of Plants and Algae for Extraction of Bioactive
Compounds, Food Reviews International, (29), pp.352-370.
98. Hongyan Mu, Jun Jin, Dan Xie, Xiaoqiang Zou, Xiaosan Wang, Xingguo Wang,
Qingzhe Jin (2016), Combined Urea Complexation and Argentated Silica
Gel Column Chromatography for Concentration and Separation of
PUFAs from Tuna Oil: Based on Improved DPA Level, J Am Oil Chem
Soc 93, pp.1157-1167.
99. Huang T.C, Fu H.Y., Ho C. T., Tan D. , Huang Y.T., Pan M.H (2007), Induction
of apoptosis by cinnamaldehyde from indigenous cinnamon Cinnamomum
osmophloeum Kaneh through reactive oxygen species production,
glutathione depletion, and caspase activation in human leukemia K562
cells, Food Chemistry 103 (2007) 434–443
100. Huang Y.F., Huang J.W., Tao L., Zhang Y. M., (2005), Chemical components
of essential oils of Cinnamomum cassia Presl, in different growth year,
Acta Sci. Nat. Univ. Sunyatseni 44 (1), pp.82-85.

144
101. Huong Thi My Nguyen, Raul Perez- Galvez, Jean Pascal Berge (2012), Effect
of diets containing tuna head hydrolysates on the surcival and growth of
shrimp Penaeus vannamei, Aquaculture, pp. 324-325.
102. Huong Thi My Nguyen (2013), Protein and lipid recovery from tuna head using
industrial protease, J. Sci. & Devel., Vol. 11, No. 8: 1150-1158.
103. Jeyaratnam, N., Nour, A.H., Akindoyo, J.O., (2016), Comparative study
between Hydrodistillation and Microwave-Assisted Hydrodistillation for
extraction of Cinnamomum Cassia oil, J. Eng. Appl. Sci, 11 (4), pp.2647-
2652.
104. Jeyaratnam, N., Nour, A.H., Akindoyo, J.O., (2016), The potential of Microwave
Assisted Hydrodistillation in extraction of essential oil from Cinnamomum
cassia (cinnamon), J. Eng. Appl. Sci. 11 (4), pp.2179-2183.
105. Jiang, Z.-H. and Argyropoulos, D.S (1999), Isolation and Characterization of
Residual Lignins in Kraft Pulps, Journal of Pulp and Papar Science, Vol.
25 (1),pp. 25-29.
106. Jiao Jiao, Yu-Jie Fu, Yuan-Gang Zu, Meng Luo, Wei Wang, Lin Zhang, Ji Li
(2012), Enzyme-assisted microwave hydro-distillation essential oil from
Fructus forsythia, chemical constituents, and its antimicrobial and
antioxidant activities, Food Chemistry, (134), pp.235-243.
107. Kaur Gunveen, DavidCameron-Smith Manohar Garg, Sinclair Andrew J
(2011), Docosapentaenoic acid (22:5n-3): a review of its biological
effects, Progr Lipid Res 50, pp.28-34.
108. Kim, Y.J. et al. (2005), Phenolic extraction from apple peel by cellulases from
Thermobifida fusca. J. Agric, Food Chem, (53), pp.9560- 9565.
109. Kristinsson H. G. and Rasco B. A. (2000), Fish Protein Hydrolysates:
Production, Biochemical, and Functional Properties, Critical Reviews in
Food Science and Nutrition, 40(1), pp.43-81.
110. Kumar, P.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. (2009), Methods for
pretreatment of lignocellulosic biomass for efficient hydrolysis and
biofuel production. Ind. Eng. Chem. Res., 48, pp.3713-3729.

145
111. Laplante, S., Souchet, N., and Bryl, P., (2009), Comparison of low-temperature
processes for oil and coenzyme Q10 extraction from mackerel and
herring, Eur. J., Lipid Sci. Technol., 111, pp.135-141.
112. Lee H.S., Kim BS, and Kim MK (2002) Suppression effect of Cinnamomum
cassia bark-derived component on nitric oxide synthase. J Agric Food
Chem 50, 7700- 7703.
113. Lee H. S (2009), Chemical Composition of Cinnamomum cassia Leaf Oils and
Suppression Effect of Cinnamyl Alcohol on Nitric Oxide Synthase, J.
Korean Soc. Appl. Biol. Chem. 52(5), pp.480-485.
114. Lee HS, Ahn YJ (1998), Growth-inhibiting effects of Cinnamomum cassia bark
derived materials on human intestinal bacteria, J Agric Food Chem;46,
pp.8-12.
115. Leskovac V. (2003), Comprehensive enzyme kinetics, Kluwer Academic
Publishers: New York, 1st edn, ISBN 978-0306467127.
116. Li, Y.-q., Kong, D.-x., Huang, R.-s., Liang, H.-l., Xu, C.-g., Wu, H., (2013),
Variations in essential oil yields and compositions of Cinnamomum cassia
leaves at different developmental stages, Ind. Crops Prod, (47), pp.92-101.
117. Liaset, B., Julshamn, K., Espe, M., (2003), Chemical composition and
theoretical nutritional evaluation of the produced fractions from enzymeic
hydrolysis of salmon frames with Protamex, Process Biochemistry 38,
pp.1747-1759.
118. Liaset, B., Nortvedt, R., Lied, E., Espe, M., (2002), Studies on the nitrogen
recovery in enzymeic hydrolysis of Atlantic salmon (Salmo salar, L.)
frames by Protamex protease, Process Biochemistry 37, pp.1263-1269.
119. Likhitayawuid K., Angerhofer C.K. (1993), “Cytotoxic and antimalarial
bisbenzylisoquinoline alkaloids from Sephania evecta”, Jounal of Natural
Products, 56 (1), pp.30-38.
120. Linder, M., Fanni, J., Parmentier, M., (2005), Proteolytic extraction of salmon
oil and PUFA concentration by lipases, Marine Biotechnology, 7(1),
pp.70-75.
121. Linthorst J. A.(2010), An overview: origins and development of green

146
chemistry, Found Chem, (12), pp.55-68.
122. Liu, S., Zhang, C., Hong, P., Ji, H., (2006), Concentration of docosahexaenoic
acid (DHA) and eicosapentaenoic acid (EPA) of tuna oil by urea
complexion: Optimization of process parameters, Journal of Food
Engineering, 73, pp.203, 209.
123. Lucchesi ME, Chemat F, Smadja J (2004), Solvent-free microwave extraction
of essential oil from aromatic herbs: comparison with conventional hydro-
distillation, J Chromatogr A 1043:323-327.
124. Madzak C, Mimmi MC, Caminade E, Brault A, Baumberger S, Briozzo P,
Mougin C, Jo- livalt C (2006), Shifting the optimal ph of activity for a
laccase from the fungus Trametes versicolor by structure-based
mutagenesis, Protein Eng Des Sel, 19(2), pp.77-84.
125. Magiatis P, Skaltsounis AL, Chinou I, Haroutounian SA (2002), Chemical
composition and in vitro antimicrobial activity of the essential oils of three
Greek Achillea species, Z Naturforsch C;57, pp.287-90.
126. Maheshwari M. L., Jain T. C., Bates RB, Battacharyya S.C., (1993), Terpenoids
XLI. Structure and absolute configuration of alpha- agrofuran, beta-
agarofuran and dihydroagarofuran, Tetrahedron, (19), pp.1079-1019.
127. Makrides M, Gibson RA, McPhee AJ, Yelland L, Quinlivan J, Ryan P. (2010),
Effect of DHA supplementation during pregnancy on maternal depression
and neurodevelopment of young children: a randomized controlled trial,
JAMA. 304 (15), pp.1675 -83.
128. Mbatia, N.B., (2011), Valorisation of fish waste biomass through recovery of
nutritional lipids and biogas, Doctoral thesis, Lund University, Sweden.
129. Miller G. L (1959), Use of dinitrosalicylic acid reagent for determination of
reducing sugar, Anal. Chem. 3, pp. 426-428.
130. Moleyar V, Narasimham P (1992), Antibacterial activity of essential oil
components, Int J Food Microbiol;16, pp. 337-42.
131. Moller JKS, Lindberg Madsen H, Aaltonen T, Skibsted LH (1999), Dittany
(Origanum dictamnus) as a source of water-extractable antioxidants,

147
Food Chem 1999, 64:215-219.
132. Mondal, K., I. Roy, and M.N. Gupta (2004), Enhancement of catalytic efficiencies
of xylanase, pectinase and cellulase by microwave pretreatment of their
substrates, Biocatalysis and Biotransformations, (22), pp. 9-16.
133. Mukhopadhayay M., (2000), Natural Extracts Using Supercritical Carbon
Dioxide. CRC Press Florida.
134. Munish Puri, Deepika Sharma and Colin J. Barrow (2012), Enzyme assisted
extraction of bioactives from plants, Trends in Biotechnology, Vol. 30,
No. 1, pp. 37-44.
135. Murugesan K, Nam I, Kim Y, Chang Y. (2007), Decolorization of reactive dyes
by a ther-mostable laccase produced by Ganoderma lucidum in solid state
culture, Enzyme Microb Technol, 40(7), pp. 1662-1672.
136. Nabavi S.F, Lorenzo A. D, Izadi M., Sánchez E.S, Daglia M. and Nabavi S.M.,
(2015), Antibacterial Effects of Cinnamon: From Farm to Food, Cosmetic
and Pharmaceutical Industries, Nutrients, 7(9), pp.7729-7748.
137. Nitthiyah Jeyaratnam, Abdurahman Hamid Nour, Ramesh Kanthasamy, Azhari
Hamid Nour, A.R. Yuvaraj, John Olabode Akindoyo (2016), Essential oil
from Cinnamomum cassia bark through hydrodistillation and advanced
microwave assisted hydrodistillation, Industrial Crops and Products, (92),
pp.57-66.
138. Nguyen H.T.M., Sylla K. S. B., Randriamahatody Z., Donnay-Moreno C.,
Moreau J., Tran L.T, Berge J. P. (2011), Enzymeatic hydrolysis of
yellowfin tuna (Thunnus albacares) by products using Protamex protease,
Food Technology and Biotechnology 49 (1), pp.48-55.
139. Pereira E. B., Castro, moraes F. F. D., Morase F. F.D and Zanin G.M (2001),
Kinetic Studies of Lipase from Candida rugosa- A Comparative Study
Between Free and Immobilized Enzyme onto Porous Chitosan Beads,
Applied Biochemistry and Biotechnology, Vol. 91-93, pp.739-752.
140. Ping Li, Lin Tian, Tao Li (2015), Study on Ultrasonic-Assisted Extraction of
Essential Oil from Cinnamon Bark and Preliminary Investigation of Its

148
Antibacterial Activity.
141. Presti, M. L., Ragusa, S., Trozzi, A., Dugo, P., Visinoni, F., Fazio, A., (2005),
A comparison between different techniques for the isolation of rosemary
essential oil, Journal of Separation Science, (28), pp.273-280.
142. Pham Thanh Loan, Ngo The Long, Cao Van, Nguyen Thi Kim Thom, Nguyen
Đac Trien (2015), Building the models of Intergrated Pest Management
(IPM) for Cinnamomum cassia in Van Yen district, Yen Bai province,
Journal of Agricultural Technology 2015 Vol. 11(8): 2469-2480.
143. Qin Chen, megan N. Marshall, Scott M. Geib (2012), Effects of laccase on
lignin depolymerization and enzymeatic hydrolysis of ensied corn stover,
Bioresourse
144. Ramadan, M.F. et al. (2009), Chromatographic analysis for fatty acids and
lipid-soluble bioactives of Derris indica crude seed oil, J. Chromatogr.
(70), pp.103-108.
145. Ranalli, A., Lucera, L., Contento, S., Simone, N. and Re, P. del. (2004),
Bioactive constituents, flavours and aroma of virgin oils obtained by
processing olives with a natural enzyme extract, European Journal of
Lipid Science and Technology. 106(3), pp.187-197.
146. Ravindran, P., Nirmal Babu, K., Shylaja, M., (2004), C Innamon and Cassia:
the Genus Cinnamomum, Medicinal and Aromatic Plants-industrial
Profiles v. 36. CRC Press, Boca Raton.
147. Rosenthal, A., D.L. Pyle, and K. Niranjan (1996), Aqueous and enzymeatic
processes for edible oil extraction, Enzyme and Microbial Technology,
(19), pp. 403-429.
148. Rui Wang, Ruijiang Wang, Bao Yang (2009), Extraction of essential oils from
five cinnamon leaves and identification of their volatile compound
compositions, Innovative Food Science and Emerging Technologies 10,
pp. 289-292.
149. Salampessy J., Phillips M, Seneweera S, Kailasapathy K (2010), Release of
antimicrobial peptides through bromelain hydrolysis of leatherjacket

149
(Meuchenia sp.) insoluble proteins, Food Chemistry 120, pp. 556-560.
150. Salgado-Roman, M., Botello-Alvarez, E., Rico-Martinez, R., Jimenez-Islas, H.,
Cardenas-Manriquez, M., & Navarrete-Bolanos, J. L. (2008), Enzymeatic
treatment to improve extraction of capsaicinoids and carotenoids from
chili (Capsicum annuum) fruits, Journal of Agriculture Food Chemistry,
56(21), pp.10012-10018.
151. Salina H. F., Shima A.R., Masniza M., Faeizah H. N (2013), Enzyme assisted
aqueous extraction and phenolic antioxidants of onion oil, International
Journal of Science, Environment and Technology, Vol. 2, No 5, pp.949 - 955.
152. Sarot Cheenpracha, Eun-Jung Park, Bahman Rostama, John M. Pezzuto and
Leng Chee Chang (2010). Inhibition of Nitric Oxide (NO) Production in
Lipopolysaccharide (LPS)-Activated Murine Macrophage RAW 264.7
Cells by the Norsesterterpene Peroxide, Epimuqubilin A, Mar. Drugs
2010, 8, pp.429-437.
153. Sathivel, S., Smiley, S., Prinyawiwatkul, W., and Bechtel, J.P., (2005),
Functional and Nutritional Properties of Red Salmon (Oncorhynchus
nerka) Enzymeatic Hydrolysates, Journal of Food Science - Vol. 70,
pp.401-406.
154. Sayantani Dutta and Paramita Bhattacharjee (2015), Enzyme-assisted
supercritical carbon dioxide extraction of black pepper oleoresin for
enhanced yield of piperine-rich extract, Journal of Bioscience and
Bioengineering Vol. 120 No. 1, pp.17-23.
155. Shan B., Yizhong Z. C., Sun M., and Corke H., (2005), Antioxidant Capacity of
26 Spice Extracts and Characterization of Their Phenolic Constituents, J.
Agric. Food Chem., 2005, 53 (20), pp. 7749-7759.
156. Shilei Geng, Zhaoxue Cui, Xinchao Huang a, Yufen Chen, Di Xu c, Ping Xiong
(2011), Variations in essential oil yield and composition during
Cinnamomum cassia bark growth, Industrial Crops and Products, (33),
pp.248-252.
157. Shimada, Y., Maruyama, K., Sugihara, A. Moriyama, S., Tominaga, Y. (1997),
Purifi cation of docosahexaenoic acid from tuna oil by a two-step

150
enzymeatic method: hydrolysis and selective esterifi cation, Journal of the
American Oil Chemists’ Society.74, pp.1441-1446.
158. Sibel .K, N. Sedef, K. Nural, S. Sahin, S. Gulum and B. Beste (2012),
Microwave-assaited hydrodistillation of essential oil from rosemary,
Journal of Food Science Technology.
159. Slizyté, R., Dauksas, E., Falch, E., Rustad, T., Storro, I., (2005), Yield and
composition of different fractions obtained after enzymeatic hydrolysis of cod
(Gardus morhua) by-products, Process Biochemistry, (40), pp.1415-1424.
160. Slizyté, R., Rustad, T., Storro, I., (2005), Enzymeatic hydrolysis of cod (Gardus
morhua) by-products - Optimization of yield and properties of lipid and
protein fraction, Process Biochemistry, (40), pp.3680-3692.
161. Sowbhagya H.B., Purnima K. T., Florence S. P., Rao A.G., Srinivas P. (2009),
Evaluation of enzyme-assisted extraction on quality of garlic volatile oil,
Food Chemistry 113, pp. 1234-1238.
162. Sowbhagya H. B. & Chitra V. N. (2010), Enzyme-Assisted Extraction of
Flavorings and Colorants from Plant Materials, Critical Reviews in Food
Science and Nutrition, 50:2, pp.146-161.
163. Sowbhagya H.B., Srinivas P., Krishnamurthy N. (2010), Effect of enzymes on
extraction of volatiles from celery seeds, Food Chemistry, (120), pp.230-234.
164. Sowbhagya H.B., Srinivas P., Purnima K.T., Krishnamurthy N. (2011),
Enzyme-assisted extraction of volatiles from cumin (Cuminum cyminum
L.) seeds, Food Chemistry, (127), pp.1856-1861.
165. Srebotnik E., Hammel K. E. (2000), Degradation of nonphenolic lignin by the
laccase/1-hydroxybenzotriazole system, Journal of Biotechnology (81),
pp.179-188.
166. Sun J. X. and Sun R. C. (2004), Isolation and characterization of cellulose from
sugarcane bagasse, Journal Polymer Degradation and Stability,Vol 84,
Issue 2, pp. 331-339.
167. Thomas M, Wood and K. Mahalingeshwara Bhat (1998), Methods in
Enzymeology, Vol. 160, 8, pp.7-112.

151
168. Tran Thanh Truc, Le Kim Thanh and Nguyen Van Muoi (2008), Effect of pH
and temperature on activity of bromelain in pineapple fruit, The
1st Conference Food Science and Technology Mekong delta, March 20-
22, pp.35-138.
169. Unlu M., Ergene E., Unlu G.V., Zeytinoglu H.S., Vural N. (2010), Composition,
antimicrobial activity and in vitro cytotoxicity of essential oil from
Cinnamomum zeylanicum Blume (Lauraceae), Food and Chemical
Toxicology 48, 3274-3280.
170. Vanden Bergher D. A., and Vlietinck (1991), “Screening methods for
Antibacterial and Ativiral Agent from Higher Plants”, Methods in Plant
biochemistry, Academic Press., USA, Vol. 6, pp.113-117.
171. Wood T. M. and Bhat K. M (1988), Methods for measuring cellulose activities,
Methods in Enzymeology 160, pp. 87-112.
172. Yoshii Eiichi, Koizumi Toru, Oribe Takiko (1987), The structure of
agarotetrol: a novel highly oxygenated chromone from agarwood (Jinko),
Tetrahedron Letters, (41), pp.3921-3924.
173. Zhongyang Ding, Lin Peng, Youzhi Chen, Liang Zhang, ZhenghuaGu, Guiyang
Shi and Kechang Zhang (2012), Production and characterization of
thermostable laccase from the mushroom, Ganoderma lucidum, using
submerged fermentation, African Journal of Microbiology Research, Vol.
6 (6), pp.1147-1157.

PHỤ LỤC