
First dryolestoid (Mammalia, Dryolestida) dentary from the Coniacian of Patagonia, Argentina
Upload
independentCategory
view
3download
0
Original article
New fossil records of Tapirus (Mammalia, Perissodactyla) from Brazil,with a critical analysis of intra-generic diversity assessments based
on lower molar size variability§
Nouveaux fossiles de Tapirus (Mammalia, Perissodactyla) du Brésil, avec une analysecritique des estimations de diversité intra-génériques basées sur la variabilité de taille
des molaires inférieures
Fernando A. Perini, João A. Oliveira, Leandro O. Salles *, Carlos R. Moraes Neto,Patrícia G. Guedes, Luiz Flamarion B. Oliveira, Marcelo Weksler
Museu Nacional/UFRJ, Departamento de Vertebrados, Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brazil
Received 10 September 2010; accepted 11 February 2011
Available online 12 October 2011
Abstract
A large set of South American fossils belonging to the genus Tapirus has been described on the basis of differences in size and proportions oflower molariform teeth. Nevertheless, the reliability of dental proportions for the diagnosis of fossil tapir species is controversial. In this paper, wedescribe new fossil material of Tapirus from the Quaternary of Serra da Bodoquena, Southwestern Brazil, comparing it to other fossil and extantspecimens of the genus by means of multivariate morphometric analyses of lower molariform teeth linear dimensions. The results of PrincipalComponent Analyses indicate that some of the extant and extinct material attributed to Tapirus fall within the range of variation in size andproportions of lower molariform teeth exhibited by recent species of the genus. Therefore, part of the fossil material attributed to new species or toTapirus sp. may be referable to the extant species Tapirus terrestris. We conclude that the sole use of lower molariform teeth size and proportions toerect new species of Tapirus may not be reliable, and therefore we advocate caution when describing fossil tapirs exclusively based on thesecriteria.# 2011 Published by Elsevier Masson SAS.
Keywords: South America; Quaternary; Dentition; Tapirus; Taxonomy; Multivariate analysis
Résumé
De nombreux fossiles sud-américains du genre Tapirus ont été décrits à partir de la différence des tailles et des proportions des dentsmolariformes inférieures. Néanmoins, la fiabilité des proportions dentaires pour la diagnose des espèces de tapirs fossiles est controversée. Danscet article, de nouveaux fossiles attribués au genre Tapirus sont décrits du Quaternaire de la Serra da Bodoquena, sud-ouest brésilien. L’analysemorphométrique multivariée du dentaire nous permet de constater une large part de variation individuelle non imputable à l’origine géographiquedes échantillons. En outre, une série d’analyses en composante principale indique que la taille et les proportions des dents molariformes inférieureschez quelques espèces actuelles et éteintes de Tapirus s’insèrent dans la variation observée pour l’espèce actuelle Tapirus terrestris. Enconséquence, de nombreux échantillons attribués à de nouvelles espèces, ou simplement à Tapirus sp., pourraient, en réalité, appartenir àT. terrestris. Nous invitons à plus de prudence à l’égard des descriptions de nouvelles espèces de Tapirus basées sur les seules dents jugales.# 2011 Publié par Elsevier Masson SAS.
Mots clés : Amérique du Sud ; Quaternaire ; Dentition ; Tapirus ; Taxonomie ; Analyse multivariée
Available online at
www.sciencedirect.com
Geobios 44 (2011) 609–619
§ Corresponding editor: Gilles Escarguel.* Corresponding author.
E-mail address: [email protected] (L.O. Salles).
0016-6995/$ – see front matter # 2011 Published by Elsevier Masson SAS.doi:10.1016/j.geobios.2011.02.005

F.A. Perini et al. / Geobios 44 (2011) 609–619610
1. Introduction
The genus Tapirus Brisson, 1762 is currently the onlyrepresentative of the family Tapiridae and includes four extantspecies (Grubb, 2005): T. terrestris (Linnaeus, 1758), T. indicusDesmarest, 1819, T. bairdii (Gill, 1865) and T. pinchaque(Roulin, 1829). Several additional fossil species are recognizedfor the genus, with the earlier records dating from the Mioceneof Europe and North America (Guérin and Eisenmann, 1994;Colbert and Schoch, 1998). In South America, the first recordsof tapirs appear at the onset of the Great American BioticInterchange, with remains dating from the early Pleistocene ofArgentina (Tonni, 1992; Ferrero and Noriega, 2007). Althoughcurrently represented by three extant species in this continent(T. bairdii, T. pinchaque, T. terrestris), the genus seems to haveharbored a considerably higher diversity in the South AmericanPleistocene (Holanda and Cozzuol, 2006; Ferrero and Noriega,2007). Seven extinct species have been described so far:T. tarijensis Ameghino, 1902, T. cristatellus Winge, 1906,T. dupuyi Ameghino, 1916, T. australis Rusconi, 1928,T. rioplatensis Cattoi, 1957, T. oliverasi Ubilla, 1983 andT. mesopotamicus Ferrero and Noriega, 2007. In addition,unnamed forms identified only as ‘‘Tapirus sp.’’ are frequentlymentioned in the literature (Ubilla, 1983; Sedor et al., 2004;Holanda and Cozzuol, 2006).
On the course of the exploration of a rich Holo-Pleistocenemammal fauna associated to the Bodoquena Karst, MatoGrosso do Sul State (Brazil) (Salles et al., 2006), new Tapirusremains were unveiled. As usual for the genus, most of thefossils consisted of dental and mandibular fragments, but someupper dentition and postcranial material were also recovered.During the identification process, the difficulties and incon-sistencies of the use of dental variability as diagnosticcharacters for South American tapirs became evident.
Although skull characters are clearly diagnostic for differenttapir species, with some authors even suggesting that they mightindicate generic differentiation in modern species (Hershkovitz,1954), the genus Tapirus is known to have uniform dentalmorphology (Colbert and Schoch, 1998). This problematic issuehas not prevented the description of new fossil species basedexclusively on dental attributes, mainly because detachedmandibles and lower teeth comprise the majority of the fossilremains of Tapirus (Ray, 1964; Ubilla, 1983; Agenbroad andDowns, 1984; Tonni, 1992; Tong, 2005; Holanda and Cozzuol,2006). In fact, most of the fossil forms were described solely onthe basis of mandibular fragments and lower molariform teeth: ofthe seven extinct South American species, only two were basedon material other than lower dentition and mandible fragments,namely T. cristatellus and T. mesopotamicus. In addition, mostcomparisons with fossil material are undertaken with limitedsample sizes. The shortcomings of such sample limitations areevident when the extant T. terrestris is taken as a reference forcomparisons, since this species shows a highly variable body sizeand a wide geographic distribution (Hershkovitz, 1954).Discussing the taxonomy of fossils tapirs, Simpson (1945)had already warned that most size-based dental differences mightnot be a reliable character information for diagnostic purposes.
Thereby, the present study is undertaken with two aims:
� to provide descriptions of the new fossil material of Tapirusrecovered from Southwestern Brazil;� to critically analyze the dental morphometric criteria
frequently employed to distinguish and diagnose new fossilspecies attributed to this genus in South America.
2. Material and methods
2.1. Geological setting and associated fauna
The Serra da Bodoquena (198 480–228 160 S, 568 320–578 240
W) is located at the southeastern part of the Mato Grosso do SulState, harboring the municipalities of Jardim, Bonito, BelaVista, Porto Murtinho, and Bodoquena (Fig. 1). A more detaileddescription of its geological and phytogeographic settings canbe found in Boggiani and Coimbra (1995) and Salles et al.(2006). Few studies concerning the Quaternary period havebeen conducted in the region, namely the restricted studies ofAlmeida (1965) and the chartered speleological studies ofMendes (1957), Gnaspini et al. (1994), and Ayub et al. (1996).There are hundreds of caves in the Serra da Bodoquena Karst,but no more than 50 have been formally described and mapped(Gnaspini et al., 1994; Ayub et al., 1996).
The fossil tapirs were recovered from two underwater caves,namely ‘‘Caverna do Japonês’’ (218 350 6300 S, 568 390 5900 W)and ‘‘Nascente do Formoso’’ (218 150 3500 S, 568 380 2600 W).Age of specimens presumably ranges from latest Pleistocene tomid-Holocene. Dating of the collagen extracted from a groundsloth from a dry cave of the Bodoquena Karst resulted in an ageestimate of 9960 � 60 BP (unpublished result), but the materialfrom the submerged caves likely represent a mixed assemblagefrom different horizons. Numerous fossil mammals typical ofthe South American Quaternary have been found in the clayeysedimentary layers associated to the tapir fragments heredescribed; these include xenarthrans (Dasypus, Euphractus,Propaopus, Pampatherium, Glossotherium, Mylodonopsis,Eremotherium and Glyptodon), carnivores (Smilodon,Panthera, Leopardus, Chrysocyon, Protocyon and Pteronura),mastodons (Haplomastodon), horses (Equus), deer (Mazamaand Ozotoceros), peccaries (Tayassu), llamas (Palaeolama),and members of South American endemic ungulate orders(Macrauchenia and Toxodon) (Salles et al., 2006; Perini et al.,2009). All collected material is housed at the Mammalcollection of the Museu Nacional (MN) at Rio de Janeiro,Brazil.
2.2. Taxonomic sampling and measurements
A set of 17 measurements typically employed in thecharacterization of fossil tapir species (Simpson, 1945; Cattoi,1951; Ubilla, 1983; Tonni, 1992) was taken from the lowercheek teeth of fossil and recent tapir specimens (Fig. 2). Allmeasurements were obtained with 150 mm digital calipers with0.02 mm accuracy.

Fig. 1. Map of the Serra da Bodoquena region (Mato Grosso do Sul, CentralBrazil), indicating the location of the two fossiliferous limestone caves ex-plored: Caverna do Japonês and Nascente do Formoso.
F.A. Perini et al. / Geobios 44 (2011) 609–619 611
In addition to the new fossil material reported below, westudied the lower cheek teeth of all recent species of Tapirus:T. terrestris (n = 98), T. indicus (n = 11), T. bairdii (n = 10) andT. pinchaque (n = 6). Examined specimens and respectivecollections are listed in the Appendix A. We also included in theanalyses the measurements of South American fossil tapirsprovided by the following authors: Ameghino (1902),T. tarijensis; Winge (1906), T. cristatellus and T. terrestris;Cattoi (1951), T. dupuyi; Ubilla (1983), T. oliverasi, T. australis,T. rioplatensis, Tapirus sp. 1, Tapirus sp. 2, and Tapirus sp. 4;Tonni (1992), T. terrestris; Holanda and Cozzuol (2006),Tapirus sp. 5, Tapirus sp. 6, Tapirus sp. 7, and T. terrestris.
Of these fossil species, some are known from very sparsematerial. Tapirus tarijensis was described by Ameghino (1902)
based only on a lower premolar series from the Pleistocene ofTarija, Argentina. Tapirus oliverasi is known from a lowermolar series, while T. australis and T. rioplatensis are onlyknown from a single lower molar each. The scarcity of thematerial and the absence of a comprehensive study of lowermolariform variation in extant forms enforce doubts about thesespecies validity. Tapirus dupuyi is another species known onlyfrom a single tooth, at times considered a m2, a m1 or even apm4 (Cattoi, 1951; Tonni, 1992). We follow Tonni (1992) andconsider the type material of T. dupuyi to be a pm4. Tapiruscristatellus is the only extinct species of Tapirus usuallyrecognized for the Brazilian Quaternary. It was described fromLagoa Santa karst by Winge (1906), and is known from cranialand postcranial remains. Winge (1906) listed a series ofcharacteristics, besides its larger size, to differentiate it fromT. terrestris, such as a narrower sagittal crest, less developedfrontal sinuses and differences in the structure of the humerusand tibia. Despite these differences, its validity has also beencontested (Paula Couto, 1979). Tapirus mesopotamicus,although known from a very complete skull, has, ironically,limited lower teeth material (Ferrero and Noriega, 2007) andwas not included in the present analysis. Lastly, the record ofTapirus sp. from Venezuela, reported by Rincón et al. (2009) assimilar to the North American T. webbi, was not available forthis study.
Tapirs tend to maintain the deciduous premolars until arelatively older age (Simpson, 1945); many specimens witherupted m1, m2, and in some cases m3, still keep the dp4. Thedistinction between adult and juvenile specimens on the basisof teeth eruption and wear is, therefore, a difficult task. Weclassified the premolars as deciduous or permanent on the basisof eruption sequence, wear, morphology, and the presence ofunerupted permanent teeth; in addition, a one-way analysis ofvariance (Anova) was employed to compare means of the eightpremolars measurements between samples of teeth previouslyclassified as permanent or deciduous.
Previous studies failed to detect significant sexual variationin the size of molariform teeth in T. terrestris (Simpson, 1945;Tonni, 1992), and presumably secondary sexual dimorphism isalso inexpressive in other species of Tapirus. As most of ourtapir samples lacked reference to gender, males and femaleswere pooled together in order to enhance sample sizes formorphometric analyses.
2.3. Morphometric analyses
Specimens lacking up to six measurements had missing dataestimated by the Expectation-Maximization method (Dempsteret al., 1977), a maximum-likelihood procedure. This methodinitially replaces missing values with the mean for each variableand then re-estimates means and covariances based on theavailable data as well as the missing data inputs. It then re-estimates the missing values from these parameters, andcontinues iteratively until the parameter estimates converge onfinal values (Strauss et al., 2003).
Three sampling schemes were employed in order to avoidestimating a large number of missing variables for very

Fig. 2. Occlusal schematic representation of a tapir right hemimandible (anterior to the right) showing the 17 measurements taken: Lp2: length of second premolar;Pp2: posterior width of second premolar; Lp3: length of third premolar; Ap3: anterior width of third premolar; Pp3: posterior width of third premolar; Lp4: length offourth premolar; Ap4: anterior width of fourth premolar; Pp4: posterior width of fourth premolar; Lm1: length of first molar; Am1: anterior width of first molar; Pm1:posterior width of first molar; Lm2: length of second molar; Am2: anterior width of second molar; Pm2: posterior width of second molar; Lm3: length of third molar;Am3: anterior width of third molar; Pm3: posterior width of third molar.
F.A. Perini et al. / Geobios 44 (2011) 609–619612
incomplete specimens, such as the fossil ones. Three datasetswere prepared, by selectively eliminating morphometricvariables that had a large number of missing values.
In the first sampling scheme, a Principal ComponentAnalysis (PCA; Strauss, 2010) for all measurements from allavailable Recent specimens of Tapirus terrestris was carried outto assess main trends of intraspecific quantitative variation incheek teeth within this widespread species. The resultingdistribution of scores provided an outline for limits ofintraspecific variation of molariform tooth characters againstwhich the variation represented in other extant and fossilspecies representatives could be compared. All values were log-transformed prior to the analyses. Eigenvalues and eigenvectorswere computed from variance-covariance matrices. Individualscores on principal components were portrayed as biplots andgrouped a posteriori with respect to the populations of origin.
In the second sampling scheme, we evaluated thediscrimination between extant species of Tapirus by meansof a Discriminant Analysis (Strauss, 2010) for the same set ofdental variables used in the first sampling scheme. Weperformed this analysis to evaluate the validity of dentalcharacters in the discrimination of species in the genus based onthe best available samples. This analysis is based onassumptions of multivariate normality, equal covariancematrices of groups, and independence of error in terms ofresiduals (Neff and Marcus, 1980). Because the sample size ofT. terrestris was much larger than those of the remainingspecies, we considered the larger geographic samples ofT. terrestris as distinct groups in this analysis. This criterionprovided a balanced sample design with respect to the samplesof T. pinchaque, T. indicus and T. bairdii, while allowing anobjective evaluation of the intraspecific discrimination amonggeographical samples assigned to T. terrestris.
The last sampling scheme included all extant and fossilTapirus specimens. Because premolars were missing for themajority of fossil specimens, we only considered molar
measurements for this analysis; this automatically excludedT. tarijensis and T. dupuyi, which are exclusively known frompremolars. Only specimens presenting values for at least five ofthe nine molar measurements had the missing values estimatedby the EM algorithm, and were thus considered in this analysis.PCA was chosen for a comprehensive comparison includingfossil specimens, due to the fact that this method has adistribution-free ordination and does not rely on assumptions ofmultivariate normality or equal covariance structure; suchconditions are usually not accomplished when taxonomicalentries are represented by single specimens. Given thisrestriction, the morphometric affinities of fossil specimens wereassessed by their a posteriori probabilistic allocation to the largersamples of extant species, based on the frequency of shortestMahalanobis distances estimated from 1000 bootstrap iterations.
All statistical analyses, including the EM missing dataestimation procedure, were implemented as algorithms writtenfor MatLab version 4.2 (Mathworks, 1994) by RichardE. Strauss, available at http://www.faculty.biol.ttu.edu/Strauss/Matlab/Matlab.htm as of August 2010.
3. Results
3.1. Description of quaternary tapirs
A total of 12 fragments, representing dental, cranial, andpostcranial remains were recovered and attributed to the genusTapirus. Although the second part of this study is centered onthe lower dentition variation, all recovered material is describedbelow. Measurements of the fossil material described areprovided in Suppl. Table S1.
Order PERISSODACTYLA Owen, 1848Family TAPIRIDAE Gray, 1821Genus Tapirus Brisson, 1762

Table 1F-values and associated probabilities ( p) for the Anovas for differences betweenmeans of deciduous and permanent premolar measurements.
Var F p
Lp2 14.034 < 0.001 (***)Ap2 1.9518 > 0.05 (ns)Lp3 0.6705 > 0.05 (ns)Ap3 24.6144 < 0.001 (***)Pp3 30.3526 < 0.001 (***)Lp4 4.4600 < 0.05 (*)Ap4 29.2408 < 0.001 (***)Pp4 37.2834 < 0.001 (***)
Variable abbreviations are explained in the text; probabilities were classified as:* = marginally significant; *** = highly significant; ns = not significant.
F.A. Perini et al. / Geobios 44 (2011) 609–619 613
Description:Craniodental: MN57057, a mandible (Fig. 3a). The left and
right dentaries are separated at the mandibular symphysis, andthe mandible lacks the entire anterior alveolar border. Exceptfor the second premolar, which is lacking in both dentaries, thecheek teeth series is complete. The mandible is also broken atthe vertical rami, missing the articular, coronoid, and angularprocesses. Overall, although cracked, the mandible is very wellpreserved. MN57056, a fragment of a left ramus of a mandible(Fig. 3b), broken at the distal portion, at the anterior margin ofthe fourth premolar and at the proximal portion, and missing thearticular, coronoid and angular processes. The surface of thebone shows signs of heavy wear. MN57060, a mandible of ajuvenile specimen (Fig. 3c), almost complete; the right ramus isbroken only at the tip of the coronoid process, but the left ramuslacks the articular, coronoid and angular processes. The tip ofthe mandible at the mandibular symphysis is also broken,exposing the alveoli of the canines and incisors. An uneruptedincisor is seen inside one of the alveoli. The deciduouspremolars and two unerupted first molars are also preserved.MN57059, a mandible of a juvenile specimen (Fig. 3d). Thismandible is in great part complete, but both rami are broken,lacking the articular, coronoid and articular processes. The tipof the mandible is also broken, and the entire piece shows somewear and cracks. The mandible preserves the deciduouspremolars, and the unerupted first molars are visible inside thealveoli. MN57054, a permanent right lower premolar (Fig. 3e).This isolated tooth is well preserved, but lacks the roots.MN57055, an entire left lower second molar (Fig. 3f).MN57050, a left maxilla (Fig. 3g) broken at the anteriorextremity, near the insertion of the first premolar andposteriorly at the level of the second molar, which is alsobroken. The three premolars and the first molar are preserved.The medial face of the maxilla is damaged, and the lateralsurface shows signs of wear. MN57049, a left maxilla (Fig. 3h).This fragment is heavily damaged, but the two last molars andthe alveoli of first molar and last two premolars are in goodcondition. MN57051, a fragment of left maxilla (Fig. 3i). Thisfragment is poorly preserved, lacking all the teeth andconserving only part of the alveolus.
Postcranium: MN57058, a left humerus (Fig. 3j) broken atthe anterior and proximal portions and lacking the medial andpart of the lateral tuberosity, but otherwise intact, preservingmany of the features and scars on its surface. MN57052, a righthumerus (Fig. 3k). This bone, which probably belonged to ajuvenile, is broken at extremities, lacking both articularportions. MN57053, a left fragment of a hip bone (Fig. 3l).This piece preserves the acetabulum, part of the acetabularbranch of the ischium, the shaft of the ilium, and part of thewing of the ilium.
3.2. Morphometric analyses
The descriptive statistics of molariform teeth measurements,based on available samples of recent species of Tapirus, aresummarized in Suppl. Tables S2 and S3. Because highlysignificant differences were revealed between samples of
deciduous and permanent premolars for five of eightmeasurements (Table 1), all subsequent analyses, includingthe estimation of missing values, were performed separately forsamples of deciduous and permanent teeth. The data set ofdeciduous teeth was reduced to the eight premolar variables,while the permanent dataset included 17 measurements (eightfrom premolars and nine from molars). Our results thereforesupport the contentions of Holanda and Cozzuol (2006) to treatpermanent and deciduous dentitions separated.
Because deciduous teeth were less represented in theavailable samples, usually determining a reduction of variablesto avoid singular covariance matrices, only the resultsconcerning the permanent dataset are reported below. It isalso important to note that the multivariate analyses includingfossils were based only on the nine molar tooth measurements.When analyzed separately, deciduous teeth provided similarresults to those of permanent teeth.
In the analysis of geographic variation of Tapirus terrestris,the first three principal components (PCs) accounted for 76.5%of the total variation. The plot of PC1 and PC2 centroids and95% confidence ellipses of geographic group scores reveal anotable superimposition; the sample from Colombia was theonly one showing distinct scores (Fig. 4a). PC1 loadings wereall positive, suggesting that it can be interpreted as a generalsize axis. The PC2 loadings reveal greater contrasts betweensubsets of dental characters, but still no obvious geographicstructure emerges from the score plot.
The discriminant analysis including all four extant species,T. bairdii, T. indicus, T. pinchaque, and T. terrestris (this lastrepresented by its largest geographic samples, also identified asdistinct groups) revealed that the two first discriminantfunctions account for more than 70% of the total variationamong groups. Four distinct groups of scores are revealed withrespect to the first two discriminant axes (Fig. 4b): the lowerscores in DF1 belong to samples referred to T. terrestris; anopposite group refers to larger scores of T. indicus, while a thirdgroup refers to T. pinchaque and T. bairdii, represented by theintermediate scores in DF1. Also intermediate with respect toDF1, but completely distinct in relation to DF2 is the sample ofT. terrestris from western Colombia, which constitutes thefourth group. A vector plot of character correlations with these
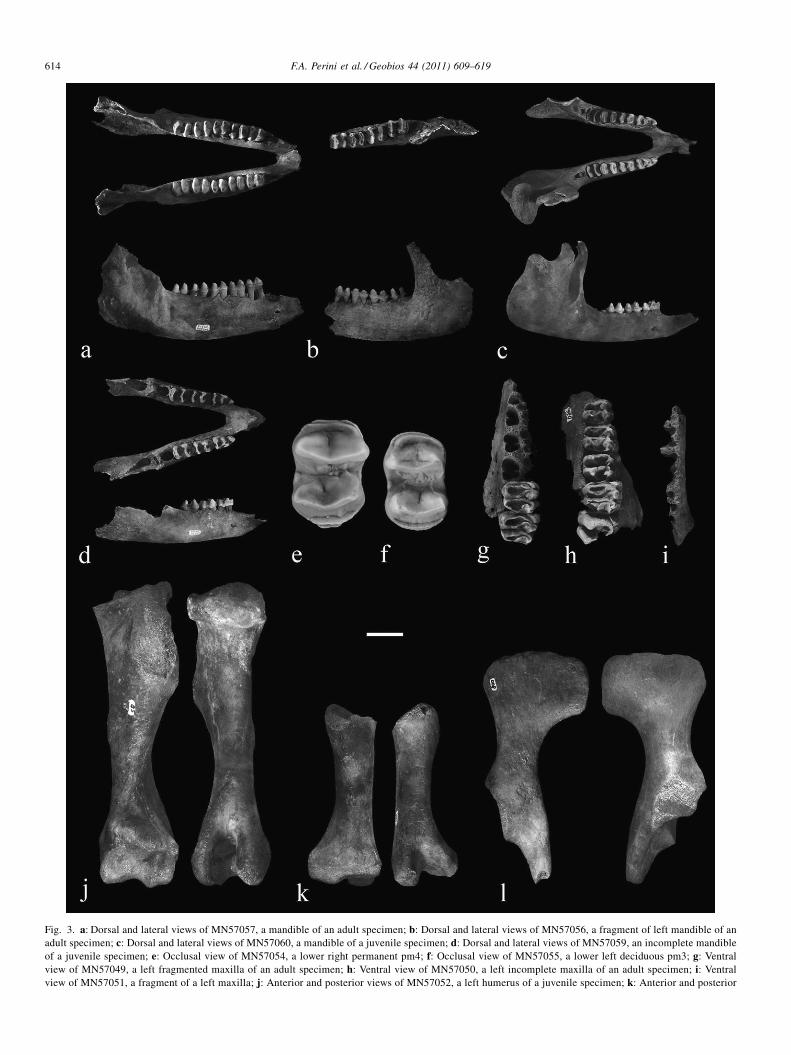
Fig. 3. a: Dorsal and lateral views of MN57057, a mandible of an adult specimen; b: Dorsal and lateral views of MN57056, a fragment of left mandible of anadult specimen; c: Dorsal and lateral views of MN57060, a mandible of a juvenile specimen; d: Dorsal and lateral views of MN57059, an incomplete mandibleof a juvenile specimen; e: Occlusal view of MN57054, a lower right permanent pm4; f: Occlusal view of MN57055, a lower left deciduous pm3; g: Ventralview of MN57049, a left fragmented maxilla of an adult specimen; h: Ventral view of MN57050, a left incomplete maxilla of an adult specimen; i: Ventralview of MN57051, a fragment of a left maxilla; j: Anterior and posterior views of MN57052, a left humerus of a juvenile specimen; k: Anterior and posterior
F.A. Perini et al. / Geobios 44 (2011) 609–619614

Table 2Frequency values for probabilistic allocations of fossil specimens to extant species based on shortest Mahalanobis distances estimated from 1000 bootstrap iterations.CO: Colombia; BR-MT: Brazil, Mato Grosso; BR-PA: Brazil, Pará; PE: Peru; BR-SP: Brazil, São Paulo.
T. terrestris T. terrestris T. terrestris T. terrestris T. terrestris T. indicus T. bairdii T. pinchaque
CO BR-MT BR-PA PE BR-SP
T. terrestris (Tonni, 1992) 0.0210 – – – 0.005 0.4845 0.2997 0.1898
T. oliverasi – – – 0.001 – 0.998 0.001 –
T. australis – – – – – 1 – –
T. rioplatensis – – – – – 1 – –
Tapirus sp. 1 (Holanda and Cozzuol, 2006) – 0.3327 – – 0.023 0.002 0.6074 0.0350
Tapirus sp. 2 (Holanda and Cozzuol, 2006) 0.7662 – – – 0.001 – 0.0060 0.2268
T. cristatelus – – – 0.2038 0.013 0.5984 0.1828 0.002
T. terrestris (Winge, 1906) – – – 0.6164 0.0909 0.2188 0.0709 0.003
Tapirus sp. MN57057 – 0.4845 – – – 0.0599 0.4505 0.005
Tapirus sp. MN57056 – – – 1 – – – –
F.A. Perini et al. / Geobios 44 (2011) 609–619 615
two first discriminant functions reveals that premolar charactersare those which most contribute to the interspecific distinction.
Unfortunately, most fossil specimens are solely representedby molar teeth, restricting comparisons including fossils tomolar teeth measurements. In the PCA of extant and fossil tapirspecimens, the first three principal components summarizedmore than 86% of the total variation. The analysis reveals alarge superimposition of scores onto the first two principalcomponents; scores of some fossil specimens, especially thoseidentified only at the genus level, are superimposed with thosedelimiting scores of extant samples of Tapirus terrestris(Fig. 4c); conversely, the scores of T. cristatellus, T. australis,T. oliverasi and T. rioplatensis fall outside the convex hulls ofextant species. The 95% confidence ellipses around extanttaxon centroids included most fossil forms, but T. oliverasi andT. rioplatensis presented very distinct scores. The firstcomponent accounted for 69.5% of the variation, with originalvariables loading positively to it, thus allowing its interpretationas a latent general size vector. As in the previous analyses,character loadings in PC1 are not of particularly highmagnitude, but samples are restricted to adult (i.e., withpermanent dentition) individuals.
In a further attempt to gauge the morphometric variationrepresented by fossil specimens in relation to the larger samplesof extant species, a classification procedure based on boot-strapped Mahalanobis distances was implemented in the samemolar tooth data subset. A similar but less well resolveddiscrimination between samples of extant taxa was obtained ina DFA considering only molar teeth (not shown). Frequencyvalues for probabilistic allocations of fossil specimens to extantspecies based on 1000 bootstrap iterations (Table 2) show thatlargest fossil forms (T. oliverasi, T. australis, T. rioplatensis) are
views of MN57058, a distal portion of a right humerus of a juvenile specimen; l:bar: a–d, 4 cm; e–f, 1 cm; g–l, 3 cm.
more similar to T. indicus, the largest extant tapir. Tapiruscristatellus and the fossil specimens referred to T. terrestris byTonni (1992) also allocate mostly to T. indicus, but lessconfidently. The form referred by Holanda and Cozzuol (2006)as Tapirus sp. 1 was classified in most bootstrap iterations asT. bairdii (60%), but was also associated to the sample ofT. terrestris from Mato Grosso (33%), while Tapirus sp. 2 of thesame authors was mostly (76%) associated to the distinctpopulation of T. terrestris from Colombia. The fossil form fromLagoa Santa identified as T. terrestris (Winge, 1906) wasmostly allocated to the extant sample of this species from Peru(61%), but was also associated to T. indicus (21%). Finally, thefossils reported in the present study showed distinct allocationsto extant forms, one of them clustering to the sample ofT. terrestris from Peru in 100% of the bootstrap iterations, andthe other presenting similar probabilities of classification eitherwithin the T. terrestris sample from Mato Grosso (48%) andwithin the sample of T. bairdii (45%). Note, however, that thesamples of T. terrestris, T. pinchaque and T. bairdii presentedsimilar scores in the first discriminant axes, implying thatallocation frequencies with respect to these species should notbe overweighted.
4. Discussion
No clear pattern of geographic variation in T. terrestrisemerged from the PCA, which rather revealed an almostcomplete overlap of confidence ellipses of scores along PC1.An incipient geographic structuring, suggested by the contrastbetween the lower scores of Colombian (including othersamples from northern South America) and the larger scoresfrom southern Brazil, is weakened by the highly variable and
Dorsal and ventral views of MN57053, a left fragment of a hip bone. Scale
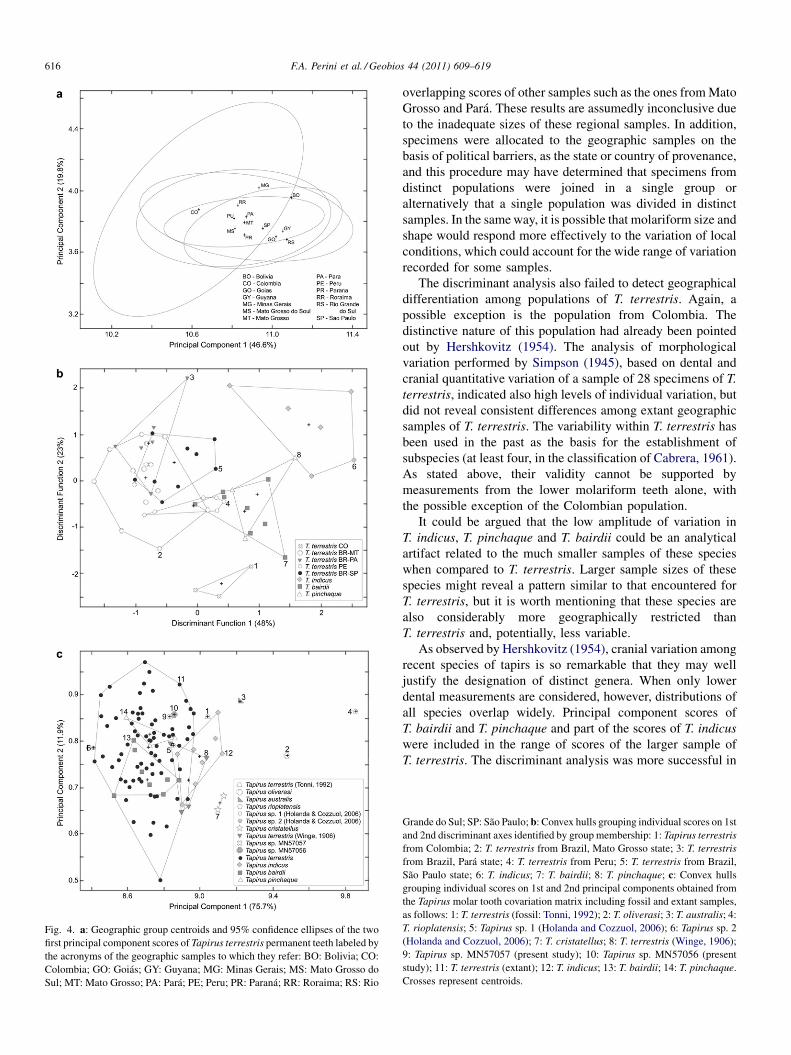
Fig. 4. a: Geographic group centroids and 95% confidence ellipses of the twofirst principal component scores of Tapirus terrestris permanent teeth labeled bythe acronyms of the geographic samples to which they refer: BO: Bolivia; CO:Colombia; GO: Goiás; GY: Guyana; MG: Minas Gerais; MS: Mato Grosso doSul; MT: Mato Grosso; PA: Pará; PE; Peru; PR: Paraná; RR: Roraima; RS: Rio
F.A. Perini et al. / Geobios 44 (2011) 609–619616
overlapping scores of other samples such as the ones from MatoGrosso and Pará. These results are assumedly inconclusive dueto the inadequate sizes of these regional samples. In addition,specimens were allocated to the geographic samples on thebasis of political barriers, as the state or country of provenance,and this procedure may have determined that specimens fromdistinct populations were joined in a single group oralternatively that a single population was divided in distinctsamples. In the same way, it is possible that molariform size andshape would respond more effectively to the variation of localconditions, which could account for the wide range of variationrecorded for some samples.
The discriminant analysis also failed to detect geographicaldifferentiation among populations of T. terrestris. Again, apossible exception is the population from Colombia. Thedistinctive nature of this population had already been pointedout by Hershkovitz (1954). The analysis of morphologicalvariation performed by Simpson (1945), based on dental andcranial quantitative variation of a sample of 28 specimens of T.terrestris, indicated also high levels of individual variation, butdid not reveal consistent differences among extant geographicsamples of T. terrestris. The variability within T. terrestris hasbeen used in the past as the basis for the establishment ofsubspecies (at least four, in the classification of Cabrera, 1961).As stated above, their validity cannot be supported bymeasurements from the lower molariform teeth alone, withthe possible exception of the Colombian population.
It could be argued that the low amplitude of variation inT. indicus, T. pinchaque and T. bairdii could be an analyticalartifact related to the much smaller samples of these specieswhen compared to T. terrestris. Larger sample sizes of thesespecies might reveal a pattern similar to that encountered forT. terrestris, but it is worth mentioning that these species arealso considerably more geographically restricted thanT. terrestris and, potentially, less variable.
As observed by Hershkovitz (1954), cranial variation amongrecent species of tapirs is so remarkable that they may welljustify the designation of distinct genera. When only lowerdental measurements are considered, however, distributions ofall species overlap widely. Principal component scores ofT. bairdii and T. pinchaque and part of the scores of T. indicuswere included in the range of scores of the larger sample ofT. terrestris. The discriminant analysis was more successful in
Grande do Sul; SP: São Paulo; b: Convex hulls grouping individual scores on 1stand 2nd discriminant axes identified by group membership: 1: Tapirus terrestrisfrom Colombia; 2: T. terrestris from Brazil, Mato Grosso state; 3: T. terrestrisfrom Brazil, Pará state; 4: T. terrestris from Peru; 5: T. terrestris from Brazil,São Paulo state; 6: T. indicus; 7: T. bairdii; 8: T. pinchaque; c: Convex hullsgrouping individual scores on 1st and 2nd principal components obtained fromthe Tapirus molar tooth covariation matrix including fossil and extant samples,as follows: 1: T. terrestris (fossil: Tonni, 1992); 2: T. oliverasi; 3: T. australis; 4:T. rioplatensis; 5: Tapirus sp. 1 (Holanda and Cozzuol, 2006); 6: Tapirus sp. 2(Holanda and Cozzuol, 2006); 7: T. cristatellus; 8: T. terrestris (Winge, 1906);9: Tapirus sp. MN57057 (present study); 10: Tapirus sp. MN57056 (presentstudy); 11: T. terrestris (extant); 12: T. indicus; 13: T. bairdii; 14: T. pinchaque.Crosses represent centroids.

F.A. Perini et al. / Geobios 44 (2011) 609–619 617
separating the modern species of Tapirus, but still revealed awide overlap, especially between T. pinchaque and T. bairdii.These results corroborate the conclusions of Ray and Sanders(1984) that the modern species of Tapirus overlap widely inmolariform measurements. The size trend documented amongSouth American species, based mainly on body length andmass, with T. pinchaque being the smallest, T. terrestrisintermediate, and T. bairdii the largest (Hershkovitz, 1954), isnot reflected in lower molariform size and proportions. Similarmolar sizes indicate, for example, that T. bairdii has small teethrelative to its size, when compared with T. terrestris.
Pleistocene tapirs are considered dentally conservative, bothin qualitative traits and morphometric terms (Ferrero andNoriega, 2007). Our results, which refer only to lowermolariform teeth, indicate that a considerable part of thevariation sampled in fossil specimens is included in the range ofvariation of extant forms. The taxonomic status of several fossilspecies described exclusively on the basis of dental material,therefore, needs to be re-evaluated. Among Argentineanspecies, Tonni (1992) already stated that several size featuressupporting the distinction of some described taxa such asT. australis were in fact highly variable, and partiallyoverlapping with the range of variation of T. terrestris. Weagree with this claim, notwithstanding our results suggest thatthe molar tooth attributed to T. australis, here considered to be am3 (following Ubilla, 1983) is in fact larger than the m3encountered in our samples of T. terrestris. Tapirus rioplatensisand T. oliverasi also seem to have considerably larger lowermolars than those of any modern species.
The only described extinct Brazilian species, T. cristatellushas been considered a junior synonym of T. terrestris by PaulaCouto (1979) and Tonni (1992), despite Winge’s (1906) listing ofa series of cranial and postcranial qualitative differences betweenT. cristatellus and T. terrestris besides size. Our study providesevidence that T. cristatellus is indeed larger than modernT. terrestris, and, coupled with the qualitative morphologicaldifferences, suggests its recognition as a valid taxon. The otherSouth American species known from cranial material,T. mesopotamicus is also easily distinguished from extantspecies by an extensive list of apomorphies (Ferrero and Noriega,2007). Although mandibular material from T. mesopotamicus isvery limited, measurements of its upper dentition revealed thatmolariform teeth were comparable in size to T. terrestris.
Of the two South American fossil species that were notincluded in the analyses, T. tarijensis is rarely present incomparisons with other fossil Tapirus, since it is known by apremolar series and, as mentioned, most other fossil species areknown only from molars. Premolar measurements provided byHolanda and Cozzuol (2006) show that the dimensions ofT. tarijensis are slightly larger than in their sample ofT. terrestris. However, the same measurements fall withinthe range of our larger sample of T. terrestris (Suppl. Table S2).With regard to T. dupuyi, the measurements provided by Cattoi(1951) indicate that it is slightly larger than the largestcorresponding teeth of our sample of T. terrestris.
As indicated above, multivariate scores of some fossilsanalyzed here are also included within the range exhibited by
extant specimens of T. terrestris. Holanda and Cozzuol (2006)reported a new fossil species of tapir from Rondonia, Brazil,based on a gracile juvenile mandible, the first record of a smallspecies of Tapirus (i.e. smaller than T. terrestris) from Brazil.One of the specimens described here (MN57060), conformswell to the mandible described by these authors, but it fitswithin the variation observed for juveniles of T. terrestris.Under those circumstances, caution is advised while recogniz-ing and describing new species exclusively based on size.Holanda and Cozzuol (2006) also concluded that the remainsfound in the Brazilian state of Acre consisted of at least two newundescribed species of Tapirus. Our results suggest that, basedon the size and proportions of lower teeth, these remainsactually fall within the variation observed for T. terrestris.Holanda and Cozzuol’s (2006) Tapirus sp. 2, notwithstandingapproaching the limits of lower molar variation of T. terrestris,is also included within its 95% confidence interval. In suchcases, the use of lower dental variation as diagnostic characterfor the recognition of new extinct tapir species does not seemjustified.
Based on the above considerations, we value therefore thatthe actual number of fossil tapir species recognized for SouthAmerica is quite likely to have been inflated by the routine ofinadequate evaluation of sizes and proportions, nearlyexclusively based on lower molars. On the other hand, thepresence of modern species in the fossil record, especiallyT. terrestris, may have been underestimated. This uncertaintyregarding the identities of species present in the SouthAmerican Quaternary, together with the high number ofunidentifiable Tapirus remains, may have led to misleadingpremises regarding the evolution of the genus in SouthAmerica. For example, de Thoisy et al. (2010) attributed thegap of almost 2 My between the earliest described fossilreferable to T. terrestris and the time of origin of the speciesestimated by molecular clock to the incompleteness of the fossilrecord or to an overestimation of its age by molecular studies.However, remains of Tapirus are known from the Ensenadan,ca. 2 My B.P. (Tonni, 1992; Holanda and Cozzuol, 2006), and,as stated above their relationships with extant species demandfurther investigation.
5. Conclusions
Dental size and proportions are frequently used as the soleindicators for identification of fossil tapir species, given thepaucity of alternative material. The use of these criteria,however, has often been questioned (Sellards, 1918; Simpson,1945; Ray, 1964; Ferrero and Noriega, 2007), and our resultscorroborate this point of view. More complete remains ofTapirus including cranial and postcranial material are fairly rarein the South American fossil record, but they seem essential toilluminate the variation observed so far in the more commonfossil remains of lower dentition.
Size and proportions of lower molars are insufficientparameters to describe and identify fossil species of Tapirus.Our evaluation is that these sources of character information donot offer grounds to build a clear perception of the number of

F.A. Perini et al. / Geobios 44 (2011) 609–619618
species of Tapirus represented in the South American fossilrecord. We then advocate here a cautious approach for theclassification of incomplete fossil material attributed to Tapirusspecimens known only from mandibular remains and lowermolariform teeth.
Some South American fossil species, as T. australis,T. rioplatensis and T. oliverasi, does seem to be significantlylarger in lower molariform dimensions than any extant speciesof the genus. However, it must be underlined that T. australisand T. rioplatensis are known from a single lower molar each,so more complete material is desirable to reinforce their statusas distinct species. In the case of the Brazilian T. cristatellus,cranial and postcranial materials are available; it is thenindicated that a taxonomical revision of this species shouldconsider these other osteological sources together with toothinformation.
At the current stage of understanding of tapir’s morphologyand evolutionary history, we are inclined to conclude thatpresumably a great portion of the Quaternary forms of SouthAmerican tapirs display molariform morphometric variationwithin the range of the currently widespread speciesT. terrestris, and may not differ significantly from it. Hence,lower molariform tooth fossil remains of South American tapirsdisplaying size and proportions included in the range ofvariation of extant forms should be referred to Tapirus sp.unless different sets of evidence point otherwise.
Acknowledgements
We would like to acknowledge all the contributions fromCECAV-IBAMA and also from the Serra da BodoquenaNational Park. Thanks to Jair da Silva for support with theillustrations and photographs. We also would like to thank thecurators and staff from the following museums that kindlyallowed us to analyze the specimens under their care: AMNH(Rob Voss, Nancy Simmons and Eileen Westwig), MZUSP(Mario de Vivo, Juliana Gualda), MNRJ (Stella Franco); and tothe reviewers of a previous version of this paper, Drs R.C.Hulbert, C. Guérin, and another anonymous reviewer forhelpful comments. Thanks to Drs H-w Tong and M.W. Colbertfor critical comments on the manuscript. Thanks also to M.Tavares (MZUSP) for the help with the French translations ofthe abstract and to D. Gettinger for revising the Englishlanguage. This work could not be developed without thefinancial support from CNPq and FAPERJ.
After this paper was accepted for publication, a new speciesof fossil Tapirus, T. rondoniensis, was described by Holandaet al., (2011) from the state of Rondonia, Brazil. The newspecies was based on a near complete skull, but there were nolower teeth available. In addition, a newly accepted paper byHolanda and Rincón, (in press) describes new Pleistocenematerial of Tapirus from Venezuela, identified as T. terrestrisand Tapirus sp., the latter indicated as potentially related toNorth American forms of Tapirus. These authors alsoperformed a PCA, comparing their material with fossil andextant species mostly considering North American species. In
what concerns the South American species, their results are,nevertheless, mostly in agreement with our own.
Appendix A
Examined specimens. Abbreviations: AMNH: AmericanMuseum of Natural History, New York, USA; MN: MuseuNacional, Rio de Janeiro, Brazil; MZUSP: Museu de Zoologiada Universidade de São Paulo, Brazil.
Tapirus terrestris. Colombia: AMNH14690 (Magdalena),AMNH23547 (Magdalena), AMNH117646 (Magdalena),AMNH142280 (Meta). Guyana: AMNH36199, AMNH140493.Brazil: Pará: MZUSP10715, MZUSP20033, AMNH95130-95133, AMNH95755, AMNH96130-96131; Roraima: MN70698-70699; Goiás: MN1700, MN4898, MZUSP6139; MatoGrosso: MN865, MN867, MN11976, MN32708, MN64153,MN64435, MN64437, MN64627, MN64652, MN64572, MN-JFV269, MZUSP7005-7007, MZUSP 3727, AMNH393-394,AMNH 36663; Mato Grosso do Sul: MZUSP3818, AMNH120996; Minas Gerais: MN868; São Paulo: MZUSP3232,MZUSP3265-3266, MZUSP3268, MZUSP3726, MZUSP3757-3758, MZUSP5701, MZUSP6575, MZUSP9712, MZUSP9714,MZUSP20034, MZUSP31983; Paraná: MZUSP3728; RioGrande do Sul: MZUSP106; MZUSP3269. Peru: AMNH71728-71731, AMNH75328, AMNH76149, AMNH76452.Bolivia: AMNH217150, AMNH 246974. Unknown locality:MN18, MN147, MN586-587, MN599, MN864, MN866,MN869, MN1601, MN1604-1605, MN1607, MN1624, MN1655, MN57061-57062, MN57064-57071, MN64215, MN71597-71599, MZUSP22421-22422, MZUSP29085, MZUSP31840.
Tapirus indicus. Burma: AMNH54960. Thailand: AMNH54657. Unknown locality: MN57063, AMNH14106, AMNH35661, AMNH77875, AMNH130108, AMNH167309,AMNH167844, AMNH201799, AMNH80077.
Tapirus bairdii. Mexico: AMNH206834, AMNH208259.Honduras: AMNH80075-80076. Nicarágua: AMNH29455,AMNH29526, AMNH29827, AMNH35000. Unknown local-ity: AMNH70396, AMNH130104.
Tapirus pinchaque. Colombia: AMNH149332. Ecuador:AMNH70521, AMNH149370, AMNH182073. Unknownlocality: AMNH149424, AMNH169931.
Appendix B. Supplementary data
Supplementary data (Tables S1–S3) associated with thisarticle can be found, in the online version, at doi:10.1016/j.geobios.2011.02.005.
References
Agenbroad, L.D., Downs, W.R., 1984. A robust tapir from Northern Arizona.Journal of the Arizona-Nevada Academy of Science 19, 91–99.
Almeida, F.F.M., 1965. Geologia da Serra da Bodoquena (Mato Grosso), Brasil.Boletim da Divisão de Geologia e Mineralogia (DNPM) 219, 1–96.
Ameghino, F., 1902. Notas sobre algunos mamiferos fosiles nuevos o pococonocidos Del Valle de Tarija. Anales Del Museo Nacional de Historianatural de Buenos Aires 3, 225–261.

F.A. Perini et al. / Geobios 44 (2011) 609–619 619
Ayub, A., Sallun Filho, W., Ferreira, N.B., Abreu, A.E.S., Teixeira, L.G.P.,Eleuterio, T.S.H., 1996. Caracterização Geo-Espeleológica Preliminar dasCavernas da Porção Central da Serra da Bodoquena, na Região de Bonito.MS Report, Instituto de Geociências, Universidade de São Paulo (unpub-lished).
Boggiani, P.C., Coimbra, A.M., 1995. Quaternary limestones of the Pantanalarea, Brazil. Anais da Academia Brasileira de Ciências 67, 343–349.
Cabrera, A., 1961. Catalogo de los mamiferos de America Del Sur. Revista delMuseo Argentino de Ciencias naturales ‘‘Bernardino Rivadavia’’ CienciasZoológicas 4, 309–732.
Cattoi, N., 1951. El ‘‘status’’ de Tapirus dupuyi (C. Amegh.). Comunicacionesdel Instituto Nacional de Investigación de las Ciencias Naturales. MuseoArgentino de Ciencias naturales ‘‘Bernardino Rivadavia’’ Ciencias Zooló-gicas 2, 103–112.
Colbert, M.W., Schoch, R.M., 1998. Tapiroidea and moropomorphs. In: Janis,C.M., Scott, K.M., Jacobs, L.L. (Eds.), Evolution of tertiary mammals ofNorth America. Terrestrial carnivores, ungulates, and ungulate-like mam-mals, Vol. 1. Cambridge University Press, Cambridge, pp. 569–582.
Dempster, A.P., Laird, N.M., Rubin, D.B., 1977. Maximum likelihood estima-tion from incomplete data via the EM algorithm. Journal of the RoyalStatistical Society 39, 1–38.
de Thoisy, B., Silva, A.G., Ruiz-García, M., Tapia, A., Ramírez, O., Arana, M.,Quse, V., Paz-y-Miño, C., Tobler, M., Pedraza, C., Lavergne, A., 2010.Population history, phylogeography, and conservation genetics of the lastNeotropical mega-herbivore, the lowland tapir (Tapirus terrestris). BMCEvolutionary Biology 10, 1–16.
Ferrero, B.S., Noriega, J.I., 2007. A new Upper Pleistocene tapir fromArgentina: remarks on the phylogenetics and diversification of NeotropicalTapiridae. Journal of Vertebrate Paleontology 27, 504–511.
Gnaspini, P., Trajano, E., Sánchez, L.E., 1994. Província Espeleológica da Serrada Bodoquena. MS: exploração, topografia e biologia, Espeleo-Tema.Revista da Sociedade Brasileira de Espeleologia 17, 19–44.
Grubb, P., 2005. Order Perissodactyla. In: Wilson, D.E., Reeder, D.M.(Eds.), Mammal species of the world – A taxonomic and geographicreference. The Johns Hopkins University Press, Baltimore, pp. 629–636.
Guérin, C., Eisenmann, V., 1994. Les tapirs (Mammalia, Perissodactyla) duMiocène supérieur d’Europe occidentale. Geobios 27, 113–127.
Hershkovitz, P., 1954. Mammals of Northern Colombia. Preliminary report no.7. Tapirs (Genus Tapirus), with a systematic review of American species.Proceedings of the United States National Museum 103, 465–496.
Holanda, E.C., Cozzuol, M.A., 2006. New records of Tapirus from the latePleistocene of Southwestern Amazonia, Brazil. Revista Brasileira dePaleontologia 9, 193–200.
Holanda, E.C., Ferigolo, J., Ribeiro, A.M., 2011. New Tapirus species (Mam-malia: Perissodactyla: Tapiridae) from the upper Pleistocene of Amazonia,Brazil. Journal of Mammalogy 92 (1), 111–120.
Holanda, E.C., Rincón, A.D., in press. Tapirs from the Pleistocene of Venezuela.Acta Palaeontologica Polonica, doi:10.4202/app.2011.0001
Mendes, J.C., 1957. Grutas Calcárias na Serra da Bodoquena, Mato Grosso.Boletim Paulista de Geografia 25, 70–77.
Neff, N.A., Marcus, L.A., 1980. Survey of multivariate methods for systematics.Neff and Marcus, New York (privately published).
Paula Couto, C., 1979. Tratado de Paleomastozoologia. Academia Brasileira deCiências, Rio de Janeiro.
Perini, F.A., Guedes, P.G., Moraes Neto, C.R., Fracasso, M.P.A., Cardoso, K.B.,Duhá, D., Salles, L.O., 2009. Carnivores (Mammalia, Carnivora) from theQuaternary of Serra da Bodoquena, Mato Grosso do Sul, Brazil. Arquivosdo Museu Nacional 67, 119–128.
Ray, C.E., 1964. Tapirus copei in the Pleistocene of Florida. Quarterly Journalof the Florida Academy of Sciences 27, 59–66.
Ray, C.E., Sanders, A.E., 1984. Pleistocene tapirs in the eastern UnitedStates. Special Publications of the Carnegie Museum of natural History8, 283–315.
Rincón, A.D., Parra, G.E., Prevosti, F.J., Alberdi, M.T., Bell, C.J., 2009. Apreliminary assessment of the mammalian fauna from the Pliocene-Pleis-tocene El Breal de Orocual locality, Monagas State, Venezuela. Museum ofNorthern Arizona Bulletin 65, 593–620.
Salles, L.O., Cartelle, C., Guedes, P.G., Boggiani, P., Janoo, A., Russo, C.A.M.,2006. First report on Quaternary Mammals from the Serra da Bodoquena,Mato Grosso do Sul, Brazil. Boletim do Museu Nacional 521, 1–12.
Sedor, F.A., Born, P.A., Santos, F.M.S., 2004. Fósseis pleistocênicos deScelidodon (Mylodontidae) e Tapirus (Tapiridae) em cavernas paranaenses(PR, sul do Brasil). Acta Biológica Paranaense 33, 121–128.
Sellards, E.H., 1918. The skull of a Pleistocene tapir including description of anew species and a note on the associated fauna and flora. Florida GeologicalSurvey Annual Report 10, 57–70.
Simpson, G.G., 1945. Notes on Pleistocene and recent tapirs. Bulletin of theAmerican Museum of Natural History 86, 37–81.
Strauss, R.E., 2010. Discriminating groups of organisms. Lecture Notes in EarthSciences 124, 73–91.
Strauss, R.E., Atanassov, M.N., Oliveira, J.A., 2003. Evaluation of the princi-pal-component and expectation-maximization methods for estimating miss-ing data in morphometric studies. Journal of Vertebrate Paleontology 23,284–296.
Tong, H., 2005. Dental characters of the Quaternary tapirs in China, theirsignificance in classification and phylogenetic assessment. Geobios 38,139–150.
Tonni, E.P., 1992. Tapirus Brisson, 1762 (Mammalia, Perissodactyla) en elLujanense (Pleistoceno Superior-Holoceno inferior) de la provincia deEntre Rios, Republica Argentina. Ameghiniana 29, 3–8.
Ubilla, M., 1983. Sobre la presencia de tapirs fosiles en el Uruguay (Mammalia,Perissodactyla, Tapiridae). Revista de la Facultad de Humanidades yCiencias. Serie Ciencias de la Tierra 1, 85–104.
Winge, H., 1906. Jordfundne og nulevende Hoydyr (Ungulata) fra Lagoa Santa,Minas Gerais, Brasilien. E Museo Lundii 3, 3–239.
Copyright © 2022 FDOKUMEN




















