
New dinosaur tracks from the Dolomia Principale (Upper Triassic) of the Carnic Prealps...
18
Bollettino della Società Paleontologica Italiana, 2014, i-xviii. Modena ISSN 0375-7633 doi:10.4435/BSPI.2014.01 New dinosaur tracks from the Dolomia Principale (Upper Triassic) of the Carnic Prealps (Friuli-Venezia Giulia, NE Italy) Marco MARZOLA & Fabio Marco DALLA VECCHIA M. Marzola, GeoBioTec, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, P-2829-516 Caparica, Portugal; Museu da Lourinhã, Rua João Luís de Moura 95, 2530-158 Lourinhã, Portugal; [email protected] F.M. Dalla Vecchia, Institut Català de Paleontologia “M. Crusafont” (ICP), Grup de Recerca del Mesozoic, C/Escola Industrial 23, E-08201 Sabadell, Spain; [email protected] KEY WORDS - Dinosaur tracks, tridactyl footprints, Anchisauripus, Dolomia Principale, Parco Dolomiti Friulane, Upper Triassic, Italy. ABSTRACT - Ten new track-bearing boulders discovered in the Parco Naturale delle Dolomiti Friulane area (Carnic Prealps, NE Italy) during the last 15 years demonstrate that bipedal dinosaurs with a functionally tridactyl, digitigrade pes, within the size-class of the ichnogenus Anchisauripus Lull, 1904, were the most common trackmakers in the carbonate tidal flats of the Dolomia Principale during the latest Triassic. If the footprint morphology actually reflects the functional tridactyl condition of the trackmakers’ feet, theropods are the most suitable trackmakers. The ichnoassociation of the Parco delle Dolomiti Friulane area differs from coeval ichnoassociations in the dominance of mid-sized tridactyl footprints and the rarity of small tridactyl (Grallator Hitchcock, 1858; length <15 cm) and chirotheriid tracks. Tracks potentially produced by basal sauropodomorphs, which were the most common European dinosaurs in the late Norian-Rhaetian interval, are underrepresented. RIASSUNTO - [Nuove orme di dinosauro nella Dolomia Principale (Triassico Superiore) delle Prealpi Carniche (Friuli-Venezia Giulia, Italia nord-orientale)] - In questo lavoro vengono descritti dieci nuovi massi con orme fossili rinvenuti negli ultimi 15 anni nel Parco Naturale delle Dolomiti Friulane (Prealpi Carniche, Italia nord-orientale). Questi nuovi reperti, inquadrati nel contesto icnologico del Triassico Superiore del Parco delle Dolomiti Friulane, confermano che i dinosauri bipedi con un pes funzionalmente tridattile e digitigrado erano i più comuni autori di orme sulle piane tidali carbonatiche della Dolomia Principale durante il Triassico Superiore. Questi dinosauri producevano orme lunghe tra i 15 e i 25 centimetri, tipiche della classe dimensionale dell’icnogenere Anchisauripus Lull, 1904. Se la morfologia delle orme riflette realmente la tridattilia funzionale del pes del trackmaker, i dinosauri teropodi sono i più probabili candidati. I potenziali autori alternativi bipedi o facoltativamente tali (arcosauri pseudosuchi, dinosauromorfi non dinosauri, ornitischi basali, saurischi basali, sauropodomorfi basali) erano troppo piccoli rispetto alle orme in esame. Inoltre, quando presenti nel record fossile attuale, le loro zampe posteriori presentano un dito I sensibilmente lungo, che toccava il suolo e poteva produrre orme tetradattili. L’icnoassociazione del Parco delle Dolomiti Friulane si distingue dalle altre icnoassociazioni coeve per l’abbondanza di orme tridattili di medie dimensioni (lunghe 15-25 cm) e la rarità di quelle più piccole e delle orme chirotheriidi. Anche le impronte riferibili ai sauropodomorfi basali, i dinosauri più comuni in Europa durante il Norico superiore-Retico, sono poco rappresentate. INTRODUCTION The Parco Naturale delle Dolomiti Friulane (Natural Park of the Friulian Dolomites; PNDF) extends in the western part of the Carnic Prealps (Pordenone Province, Friuli-Venezia Giulia Autonomous Region, northeastern Italy). The Late Triassic Dolomia Principale is a formation that crops out extensively in the Park area and has yielded a relatively abundant record of tetrapod footprints (Dalla Vecchia & Mietto, 1998; Dalla Vecchia, 2002, 2006). These are usually found on the surface of large boulders fallen down from steep slopes and cliffs. Most of the footprints were initially referred to theropod dinosaurs, a few have been attributed to “prosauropods” (= basal Sauropodomorpha of recent phylogenetic analyses; e.g., Upchurch et al., 2007; Ezcurra, 2010) and to crurotarsal archosaurs (Dalla Vecchia & Mietto, 1998). Later, Dalla Vecchia (2002, 2006, 2008) emphasized that the tridactyl footprints previously referred to theropod dinosaurs could have been produced by herrerasaurids, that according to some phylogenetic hypotheses (e.g., Langer, 2004), are basal, non-theropod saurischians with a functionally tridactyl pes. The Dolomia Principale (Hauptdolomit of German- speaking authors) has yielded a relatively rich tetrapod footprint record also in the nearby Dolomites of the Veneto Region and Bolzano/Bozen Autonomous Province (Mietto, 1988; Leonardi, 2000; Belvedere et al., 2008). This paper deals with the tracks discovered in the PNDF during the last 15 years, which have not yet been reported in the literature. The aim of this study is: 1) the description of unpublished new material and comparison with similar ichnotaxa, 2) the identification of possible trackmakers and 3) the understanding of the overall ichnoassemblage and its contextualization within the latest Triassic tetrapod tracks record. PROVENANCE AND GEOLOGICAL REMARKS The new track-bearing boulders are located in the Settimana Creek Valley and in the southern slope of Mt Caserine Basse north of the Grave di Gere locality, both of Claut municipality, and in the Pezzeda Creek Valley of Cimolais municipality (Fig. 1). Geographic coordinates are reported for each boulder. Most of the lithostratigraphic units cropping out in the PNDF originated from carbonate sediments deposited published online
Transcript of New dinosaur tracks from the Dolomia Principale (Upper Triassic) of the Carnic Prealps...

Bollettino della Società Paleontologica Italiana, 2014, i-xviii. Modena
ISSN 0375-7633 doi:10.4435/BSPI.2014.01
New dinosaur tracks from the Dolomia Principale (Upper Triassic) of the Carnic Prealps (Friuli-Venezia Giulia, NE Italy)
Marco Marzola & Fabio Marco Dalla Vecchia
M. Marzola, GeoBioTec, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, P-2829-516 Caparica, Portugal; Museu da Lourinhã, Rua João Luís de Moura 95, 2530-158 Lourinhã, Portugal; [email protected]
F.M. Dalla Vecchia, Institut Català de Paleontologia “M. Crusafont” (ICP), Grup de Recerca del Mesozoic, C/Escola Industrial 23, E-08201 Sabadell, Spain; [email protected]
KEY WORDS - Dinosaur tracks, tridactyl footprints, Anchisauripus, Dolomia Principale, Parco Dolomiti Friulane, Upper Triassic, Italy.
ABSTRACT - Ten new track-bearing boulders discovered in the Parco Naturale delle Dolomiti Friulane area (Carnic Prealps, NE Italy) during the last 15 years demonstrate that bipedal dinosaurs with a functionally tridactyl, digitigrade pes, within the size-class of the ichnogenus Anchisauripus Lull, 1904, were the most common trackmakers in the carbonate tidal flats of the Dolomia Principale during the latest Triassic. If the footprint morphology actually reflects the functional tridactyl condition of the trackmakers’ feet, theropods are the most suitable trackmakers. The ichnoassociation of the Parco delle Dolomiti Friulane area differs from coeval ichnoassociations in the dominance of mid-sized tridactyl footprints and the rarity of small tridactyl (Grallator Hitchcock, 1858; length <15 cm) and chirotheriid tracks. Tracks potentially produced by basal sauropodomorphs, which were the most common European dinosaurs in the late Norian-Rhaetian interval, are underrepresented.
RIASSUNTO - [Nuove orme di dinosauro nella Dolomia Principale (Triassico Superiore) delle Prealpi Carniche (Friuli-Venezia Giulia, Italia nord-orientale)] - In questo lavoro vengono descritti dieci nuovi massi con orme fossili rinvenuti negli ultimi 15 anni nel Parco Naturale delle Dolomiti Friulane (Prealpi Carniche, Italia nord-orientale). Questi nuovi reperti, inquadrati nel contesto icnologico del Triassico Superiore del Parco delle Dolomiti Friulane, confermano che i dinosauri bipedi con un pes funzionalmente tridattile e digitigrado erano i più comuni autori di orme sulle piane tidali carbonatiche della Dolomia Principale durante il Triassico Superiore. Questi dinosauri producevano orme lunghe tra i 15 e i 25 centimetri, tipiche della classe dimensionale dell’icnogenere Anchisauripus Lull, 1904. Se la morfologia delle orme riflette realmente la tridattilia funzionale del pes del trackmaker, i dinosauri teropodi sono i più probabili candidati. I potenziali autori alternativi bipedi o facoltativamente tali (arcosauri pseudosuchi, dinosauromorfi non dinosauri, ornitischi basali, saurischi basali, sauropodomorfi basali) erano troppo piccoli rispetto alle orme in esame. Inoltre, quando presenti nel record fossile attuale, le loro zampe posteriori presentano un dito I sensibilmente lungo, che toccava il suolo e poteva produrre orme tetradattili. L’icnoassociazione del Parco delle Dolomiti Friulane si distingue dalle altre icnoassociazioni coeve per l’abbondanza di orme tridattili di medie dimensioni (lunghe 15-25 cm) e la rarità di quelle più piccole e delle orme chirotheriidi. Anche le impronte riferibili ai sauropodomorfi basali, i dinosauri più comuni in Europa durante il Norico superiore-Retico, sono poco rappresentate.
INTRODUCTION
The Parco Naturale delle Dolomiti Friulane (Natural Park of the Friulian Dolomites; PNDF) extends in the western part of the Carnic Prealps (Pordenone Province, Friuli-Venezia Giulia Autonomous Region, northeastern Italy). The Late Triassic Dolomia Principale is a formation that crops out extensively in the Park area and has yielded a relatively abundant record of tetrapod footprints (Dalla Vecchia & Mietto, 1998; Dalla Vecchia, 2002, 2006). These are usually found on the surface of large boulders fallen down from steep slopes and cliffs. Most of the footprints were initially referred to theropod dinosaurs, a few have been attributed to “prosauropods” (= basal Sauropodomorpha of recent phylogenetic analyses; e.g., Upchurch et al., 2007; Ezcurra, 2010) and to crurotarsal archosaurs (Dalla Vecchia & Mietto, 1998). Later, Dalla Vecchia (2002, 2006, 2008) emphasized that the tridactyl footprints previously referred to theropod dinosaurs could have been produced by herrerasaurids, that according to some phylogenetic hypotheses (e.g., Langer, 2004), are basal, non-theropod saurischians with a functionally tridactyl pes.
The Dolomia Principale (Hauptdolomit of German-speaking authors) has yielded a relatively rich tetrapod
footprint record also in the nearby Dolomites of the Veneto Region and Bolzano/Bozen Autonomous Province (Mietto, 1988; Leonardi, 2000; Belvedere et al., 2008).
This paper deals with the tracks discovered in the PNDF during the last 15 years, which have not yet been reported in the literature.
The aim of this study is: 1) the description of unpublished new material and comparison with similar ichnotaxa, 2) the identification of possible trackmakers and 3) the understanding of the overall ichnoassemblage and its contextualization within the latest Triassic tetrapod tracks record.
PROVENANCE AND GEOLOGICAL REMARKS
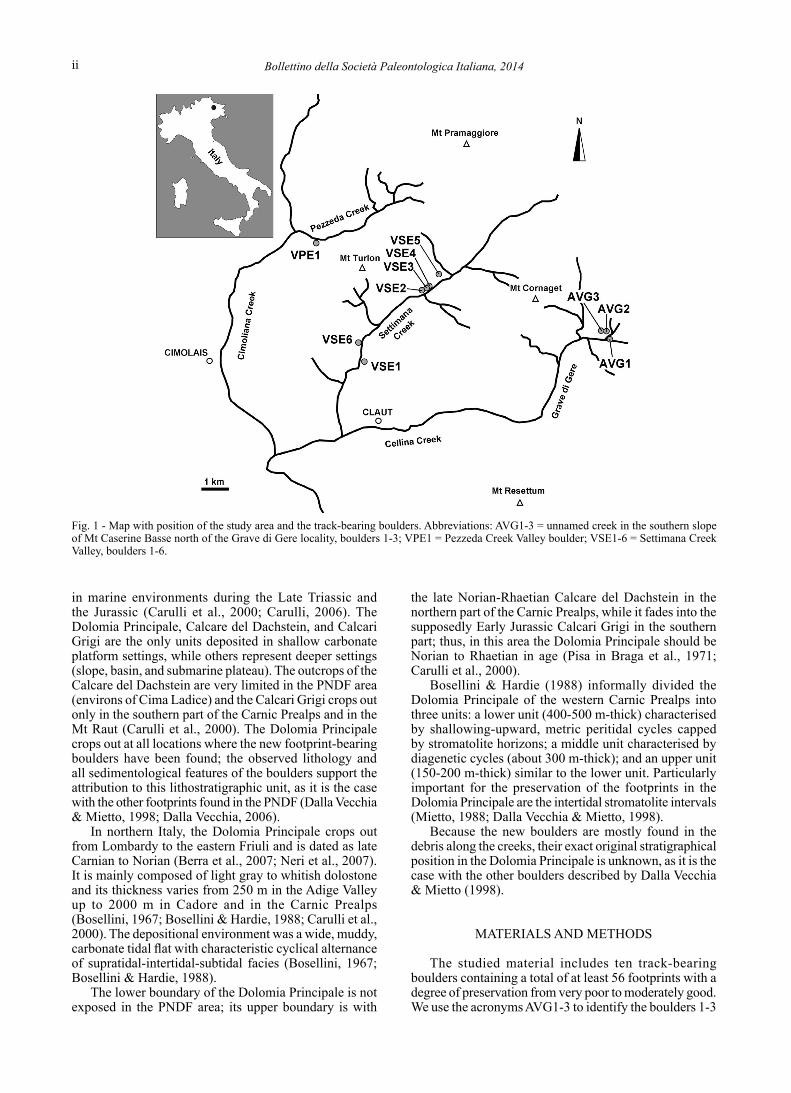
The new track-bearing boulders are located in the Settimana Creek Valley and in the southern slope of Mt Caserine Basse north of the Grave di Gere locality, both of Claut municipality, and in the Pezzeda Creek Valley of Cimolais municipality (Fig. 1). Geographic coordinates are reported for each boulder.
Most of the lithostratigraphic units cropping out in the PNDF originated from carbonate sediments deposited
published online
Bollettino della Società Paleontologica Italiana, 2014ii
in marine environments during the Late Triassic and the Jurassic (Carulli et al., 2000; Carulli, 2006). The Dolomia Principale, Calcare del Dachstein, and Calcari Grigi are the only units deposited in shallow carbonate platform settings, while others represent deeper settings (slope, basin, and submarine plateau). The outcrops of the Calcare del Dachstein are very limited in the PNDF area (environs of Cima Ladice) and the Calcari Grigi crops out only in the southern part of the Carnic Prealps and in the Mt Raut (Carulli et al., 2000). The Dolomia Principale crops out at all locations where the new footprint-bearing boulders have been found; the observed lithology and all sedimentological features of the boulders support the attribution to this lithostratigraphic unit, as it is the case with the other footprints found in the PNDF (Dalla Vecchia & Mietto, 1998; Dalla Vecchia, 2006).
In northern Italy, the Dolomia Principale crops out from Lombardy to the eastern Friuli and is dated as late Carnian to Norian (Berra et al., 2007; Neri et al., 2007). It is mainly composed of light gray to whitish dolostone and its thickness varies from 250 m in the Adige Valley up to 2000 m in Cadore and in the Carnic Prealps (Bosellini, 1967; Bosellini & Hardie, 1988; Carulli et al., 2000). The depositional environment was a wide, muddy, carbonate tidal flat with characteristic cyclical alternance of supratidal-intertidal-subtidal facies (Bosellini, 1967; Bosellini & Hardie, 1988).
The lower boundary of the Dolomia Principale is not exposed in the PNDF area; its upper boundary is with
the late Norian-Rhaetian Calcare del Dachstein in the northern part of the Carnic Prealps, while it fades into the supposedly Early Jurassic Calcari Grigi in the southern part; thus, in this area the Dolomia Principale should be Norian to Rhaetian in age (Pisa in Braga et al., 1971; Carulli et al., 2000).
Bosellini & Hardie (1988) informally divided the Dolomia Principale of the western Carnic Prealps into three units: a lower unit (400-500 m-thick) characterised by shallowing-upward, metric peritidal cycles capped by stromatolite horizons; a middle unit characterised by diagenetic cycles (about 300 m-thick); and an upper unit (150-200 m-thick) similar to the lower unit. Particularly important for the preservation of the footprints in the Dolomia Principale are the intertidal stromatolite intervals (Mietto, 1988; Dalla Vecchia & Mietto, 1998).
Because the new boulders are mostly found in the debris along the creeks, their exact original stratigraphical position in the Dolomia Principale is unknown, as it is the case with the other boulders described by Dalla Vecchia & Mietto (1998).
MATERIALS AND METHODS
The studied material includes ten track-bearing boulders containing a total of at least 56 footprints with a degree of preservation from very poor to moderately good. We use the acronyms AVG1-3 to identify the boulders 1-3
Fig. 1 - Map with position of the study area and the track-bearing boulders. Abbreviations: AVG1-3 = unnamed creek in the southern slope of Mt Caserine Basse north of the Grave di Gere locality, boulders 1-3; VPE1 = Pezzeda Creek Valley boulder; VSE1-6 = Settimana Creek Valley, boulders 1-6.

iiiM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
from an unnamed creek along the southern slope of Mt Caserine Basse just north of the Grave di Gere locality, VPE1 for the boulder from Pezzeda Creek Valley, and VSE1-6 for the boulders 1-6 from the Settimana Creek Valley (Fig. 1).
All footprints identified on those boulders appear to be tridactyl and mesaxonic; in no case the trace of another foot is associated with the tridactyl footprint to form a manus-pes couple.
Measurement of interdigital angles was carried out following Thulborn (1990, fig. 4.5). Pace length was measured as far as possible between the proximal ends of digit III traces. The height at the hip was calculated using the morphometric ratios method by Thulborn (1990, p. 251) and absolute speed using the equation of Alexander (1976). Tridactyl footprints are identified as right or left pedes according to characteristic features of digit IV. In fact, the trace of digit IV of the pes in dinosaurian tridactyl footprints often includes the imprint of a robust metatarsal pad, thus projecting more posteriorly than the trace of digit II, causing a typical posteromedial indentation (sensu Pittman, 1989). All measurements are reported in Tab. 1. Some footprints have been photographed during night with artificial illumination to increase their visibility. Track outlines were drawn onto transparent acetate film from the original material and drawings of the single footprints were traced on photographs of the cast with direct control of the morphology on the cast changing the lighting to emphasize particulars and then redrawn on transparent film. Casts were taken from eight specimens; those of VSE1-1, VSE2-1A and VSE2-1B, VSE3 and VSE5 are stored at the Museo Friulano di Storia Naturale (Udine, Italy) under repository numbers MFSN cal-073 to 077, respectively, while those of VSE2-1A, VPE1-1A, AVG1-2B and AVG1-1 are deposited at the Laboratorio “L’Occhione” of the Associazione Naturalistica Cordenonese (Cordenons, Italy) under repository numbers LOC-01 to 04. In order to illustrate the footprint morphology, photographs of the casts were taken under different illumination. The photograph of the footprint VSE5 has been elaborated to 3D by the software Agisoft PhotoScan.
Ichnological abbreviations: AII-III and AIII-IV = interdigital divarication II-III and III-IV; AII-IV = total digital divarication; ANG = pace angulation; FL = footprint length; FW = footprint width; h = height at the hip; LII-IV = length of digits II-IV prints; PL = pace length; SL = stride length; Te = toe extrusion (sensu Weems, 1992); V = absolute speed.
ICHNOTAXONOMIC REMARKS
Klein & Haubold (2007), Klein & Lucas (2010), and Klein et al. (2011) referred tridactyl mesaxonic footprints from the Upper Triassic produced by bipedal trackmakers to the ichnogenera Grallator Hitchcock, 1858 and Eubrontes Hitchcock, 1845. These ichnogenera and Anchisauripus Lull, 1904 have been diagnosed by Olsen et al. (1998) based on specimens from the Lower Jurassic of northeastern USA. Eubrontes (Olsen et al., 1998, p. 590) is characterised by a large size (>25 cm long), a “broad pes” and a total digital divarication averaging 25°-40°. Anchisauripus (Olsen et al., 1998, p. 592) is characterised
by a medium size (15-25 cm long), the trace of digit III that “projects relatively further anteriorly than in Eubrontes but not as far as in Grallator (digit III projection ratio >1.3 and <1.8)”, the footprint length/width ratio near 2, and the total digital divarication in the range 20°-35°. Grallator (Olsen et al., 1998, p. 595) is characterised by a small size (<15 cm long), the trace of digit III that “projects relatively further anteriorly ... than in Eubrontes and Anchisauripus”, the footprint length/width ratio near or greater than 2, and the total digital divarication in the range 10°-30°.
The total digital divarication depends greatly on the conditions of the substrate (Gatesy et al., 1999; Manning, 2004; Diàz-Martínez et al., 2009); this must be considered in the comparison between the type material of Grallator, Anchisauripus, and Eubrontes from the Lower Jurassic continental sandstones of North America and the footprints from PNDF. The Triassic ichnogenus Coelurosaurichnus von Huene, 1941 is considered a junior synonym of Grallator (see Leonardi & Lockley, 1995; Klein & Lucas, 2010). The Triassic ichnogenus Atreipus Olsen & Baird, 1986 is based on tridactyl mesaxonic pes imprints similar in size (pes print ranging 9-14 cm according to those authors) and morphology to Grallator that are associated with a small, tri- to tetradactyl manus imprint (see also Haubold & Klein, 2002; Safran & Rainforth, 2004; D’Orazi Porchetti et al., 2008). Haubold & Klein (2000) erected the Grallator-Atreipus plexus for grallatorid footprints of trackways produced by facultative bipedal trackmakers. The tridactyl footprints from PNDF are compared to those ichnotaxa.
Also the ichnogenus Anomoepus Hitchcock, 1848 has a tridactyl pes imprint associated with a manus imprint, but the latter is pentadactyl, the pes has a higher total digital divarication (~70°), and the trace of digit III projects only slightly beyond those of digits II and IV. Although Anomoepus is a typical Early Jurassic ichnogenus (Olsen & Rainforth, 2003; Lockley & Gierliński, 2006), it is also used for comparison and discussion here.
DESCRIPTION AND COMPARISON OF THE MATERIAL
Settimana Creek ValleyBoulDer VSE1 - (620 m a.s.l. at the confluence of
the Culisei Creek; Gauss Coordinates, obtained by GPS: 46.29°N, 12.50°E) (Fig. 2a).
This boulder contains two poorly preserved tracks that are shallow negative epireliefs surrounded by broad expulsion rims. VSE1-1 is the better preserved of the two and tridactyl (Fig. 2b-c). VSE1-2 is just a depression with a triangular outline without clear digit traces which actual morphology is concealed by the infilling by the overlaying bed; however, its posterior part has the same outline as VSE1-1, so it is probably another tridactyl footprint. VSE1-1 is about 22 cm long and moderately elongated (FL/FW = 1.57), AII-IV is moderately high (44°) and Te is about 8.5 cm. The distal portion of the impression of digit III is slightly curved and its end seems to be pointed. The traces of the outer digits do not have clearly defined margins; the left one is shorter than the right one but more posteriorly elongated. Therefore, the former probably represents the digit IV and the footprint can be identified as a left one.
Bollettino della Società Paleontologica Italiana, 2014iv
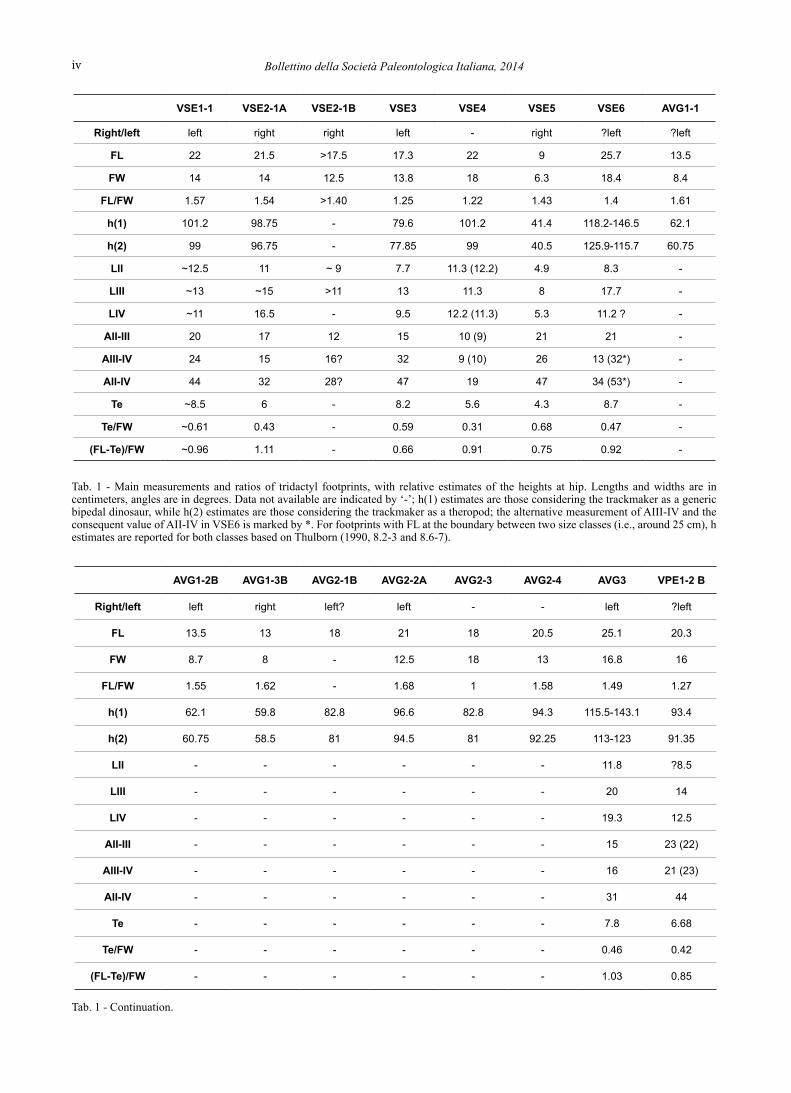
VSE1-1 VSE2-1A VSE2-1B VSE3 VSE4 VSE5 VSE6 AVG1-1
Right/left left right right left - right ?left ?left
FL 22 21.5 >17.5 17.3 22 9 25.7 13.5
FW 14 14 12.5 13.8 18 6.3 18.4 8.4
FL/FW 1.57 1.54 >1.40 1.25 1.22 1.43 1.4 1.61
h(1) 101.2 98.75 - 79.6 101.2 41.4 118.2-146.5 62.1
h(2) 99 96.75 - 77.85 99 40.5 125.9-115.7 60.75
LII ~12.5 11 ~ 9 7.7 11.3 (12.2) 4.9 8.3 -
LIII ~13 ~15 >11 13 11.3 8 17.7 -
LIV ~11 16.5 - 9.5 12.2 (11.3) 5.3 11.2 ? -
AII-III 20 17 12 15 10 (9) 21 21 -
AIII-IV 24 15 16? 32 9 (10) 26 13 (32*) -
AII-IV 44 32 28? 47 19 47 34 (53*) -
Te ~8.5 6 - 8.2 5.6 4.3 8.7 -
Te/FW ~0.61 0.43 - 0.59 0.31 0.68 0.47 -
(FL-Te)/FW ~0.96 1.11 - 0.66 0.91 0.75 0.92 -
AVG1-2B AVG1-3B AVG2-1B AVG2-2A AVG2-3 AVG2-4 AVG3 VPE1-2 B
Right/left left right left? left - - left ?left
FL 13.5 13 18 21 18 20.5 25.1 20.3
FW 8.7 8 - 12.5 18 13 16.8 16
FL/FW 1.55 1.62 - 1.68 1 1.58 1.49 1.27
h(1) 62.1 59.8 82.8 96.6 82.8 94.3 115.5-143.1 93.4
h(2) 60.75 58.5 81 94.5 81 92.25 113-123 91.35
LII - - - - - - 11.8 ?8.5
LIII - - - - - - 20 14
LIV - - - - - - 19.3 12.5
AII-III - - - - - - 15 23 (22)
AIII-IV - - - - - - 16 21 (23)
AII-IV - - - - - - 31 44
Te - - - - - - 7.8 6.68
Te/FW - - - - - - 0.46 0.42
(FL-Te)/FW - - - - - - 1.03 0.85
Tab. 1 - Main measurements and ratios of tridactyl footprints, with relative estimates of the heights at hip. Lengths and widths are in centimeters, angles are in degrees. Data not available are indicated by ‘-’; h(1) estimates are those considering the trackmaker as a generic bipedal dinosaur, while h(2) estimates are those considering the trackmaker as a theropod; the alternative measurement of AIII-IV and the consequent value of AII-IV in VSE6 is marked by *. For footprints with FL at the boundary between two size classes (i.e., around 25 cm), h estimates are reported for both classes based on Thulborn (1990, 8.2-3 and 8.6-7).
Tab. 1 - Continuation.
vM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
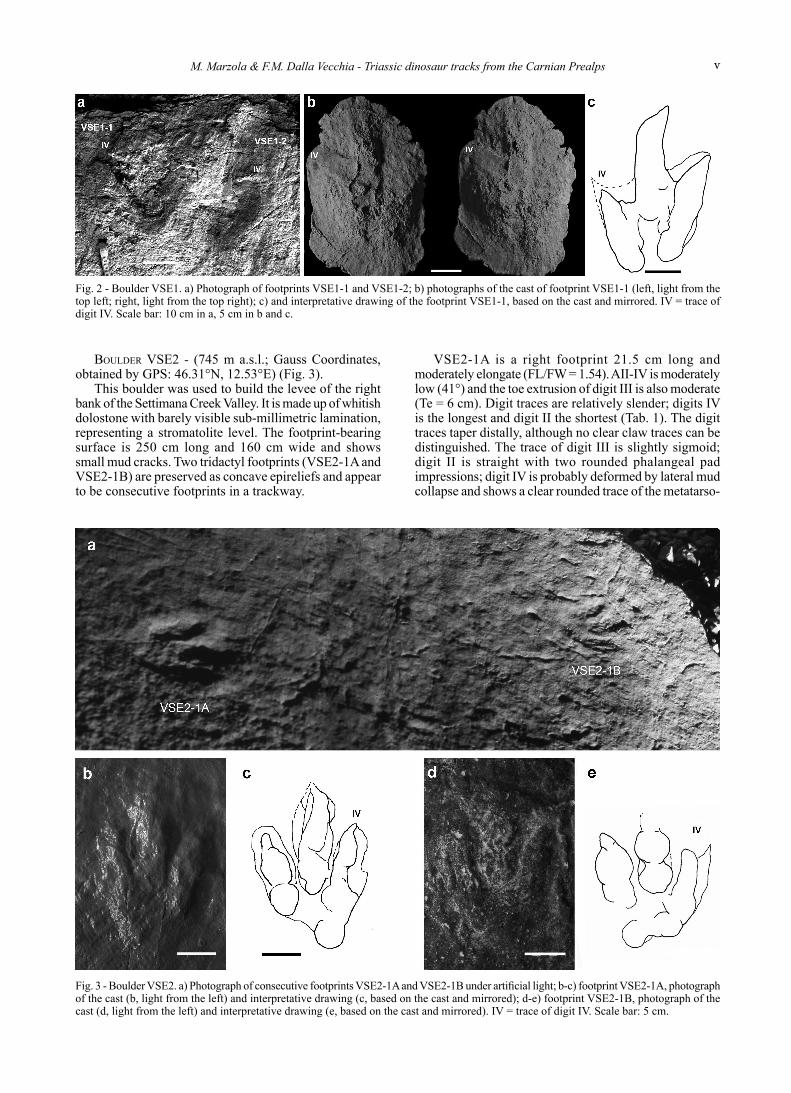
BoulDer VSe2 - (745 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.31°N, 12.53°E) (Fig. 3).
This boulder was used to build the levee of the right bank of the Settimana Creek Valley. It is made up of whitish dolostone with barely visible sub-millimetric lamination, representing a stromatolite level. The footprint-bearing surface is 250 cm long and 160 cm wide and shows small mud cracks. Two tridactyl footprints (VSE2-1A and VSE2-1B) are preserved as concave epireliefs and appear to be consecutive footprints in a trackway.
VSE2-1A is a right footprint 21.5 cm long and moderately elongate (FL/FW = 1.54). AII-IV is moderately low (41°) and the toe extrusion of digit III is also moderate (Te = 6 cm). Digit traces are relatively slender; digits IV is the longest and digit II the shortest (Tab. 1). The digit traces taper distally, although no clear claw traces can be distinguished. The trace of digit III is slightly sigmoid; digit II is straight with two rounded phalangeal pad impressions; digit IV is probably deformed by lateral mud collapse and shows a clear rounded trace of the metatarso-
Fig. 2 - Boulder VSE1. a) Photograph of footprints VSE1-1 and VSE1-2; b) photographs of the cast of footprint VSE1-1 (left, light from the top left; right, light from the top right); c) and interpretative drawing of the footprint VSE1-1, based on the cast and mirrored. IV = trace of digit IV. Scale bar: 10 cm in a, 5 cm in b and c.
Fig. 3 - Boulder VSE2. a) Photograph of consecutive footprints VSE2-1A and VSE2-1B under artificial light; b-c) footprint VSE2-1A, photograph of the cast (b, light from the left) and interpretative drawing (c, based on the cast and mirrored); d-e) footprint VSE2-1B, photograph of the cast (d, light from the left) and interpretative drawing (e, based on the cast and mirrored). IV = trace of digit IV. Scale bar: 5 cm.

Bollettino della Società Paleontologica Italiana, 2014vi
phalangeal pad. The posteromedial indentation (sensu Pittman, 1989) is evident (Fig. 3b-c).
The state of preservation of VSE2-1B is worse than VSE2-1A. Although it is apparently the subsequent trace of the latter, its congruent morphology suggests another right footprint. If this is the case, the left footprint is missing. Only the two proximal digital pad traces of digit III are evident, while the distal part of the digit is only partly preserved. This makes the footprint rather short (17.5 cm in length). The impression of digit IV is poorly defined but seems to end proximally with a rounded metatarso-phalangeal pad that is connected to another rounded pad placed in a posteromedial position. The trace of digit II is shorter and broader than that of digit IV, with two pad prints, the distal one being larger than the proximal one.
BoulDer VSe3 - (745 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.31°N, 12.53°E) (Fig. 4).
This boulder was also used to build the levee of the right bank of the Settimana Creek Valley; it is located about 40 meters upstream from VSE2. It is composed of whitish dolostone with barely visible submillimetric lamination like the boulder VSE2, but without mud cracks. The footprint-bearing surface is 250 cm long and 250 cm wide. A single tridactyl footprint of the left pes, 17.3 cm long, is preserved as a concave epirelief (Fig. 4a-b). The presence of an associated manus print can be excluded with confidence because the state of preservation is relatively good (Fig. 4a) and the surface is smooth. Digit III is straight and 130 mm long, with the impressions of three phalangeal pads, a smaller, possible phalangeal-metatarsal pad and a small, rounded, claw trace; the distal portion partly preserves the infilling by the overlying bed. Digit II is short (77 mm), with (possibly two) pad prints and a long claw trace directed anteromedially. The trace of digit IV is shallower and less defined than the other two, which are 8 mm deep at maximum; it is 95 mm long, with a broad and rounded phalangeal-metatarsal pad print and a small rounded claw trace which is directed anterolaterally.
The trace of digit II projects less posteriorly than that of digit IV, causing a posteromedial indentation (sensu Pittman, 1989) of the footprint. The footprint is scarcely elongated (FL/FW about 1.25), but digit III is much longer than the other two and largely projecting beyond their distal ends (Te is 82 mm). AII-IV is moderately high (47°).
BoulDer VSe4 - (745 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.31°N, 12.53°E) (Fig. 5).
This boulder was also used to build the levee of the right bank of the Settimana Creek Valley; it is located a few meters upstream from VSE3. It is composed of pale gray dolostone with barely visible submillimetric lamination. The footprint-bearing surface is 250 cm long and 150 cm wide. A single, poorly preserved, tridactyl and mesaxonic footprint 22 cm long is preserved as a concave epirelief (Fig. 5a). It is scarcely elongated (FL/FW is 1.22) and AII-IV is rather low (19°). The trace of digit III is straight, relatively short and distally rounded. The left outer digit trace is curved outward (Fig. 5b).
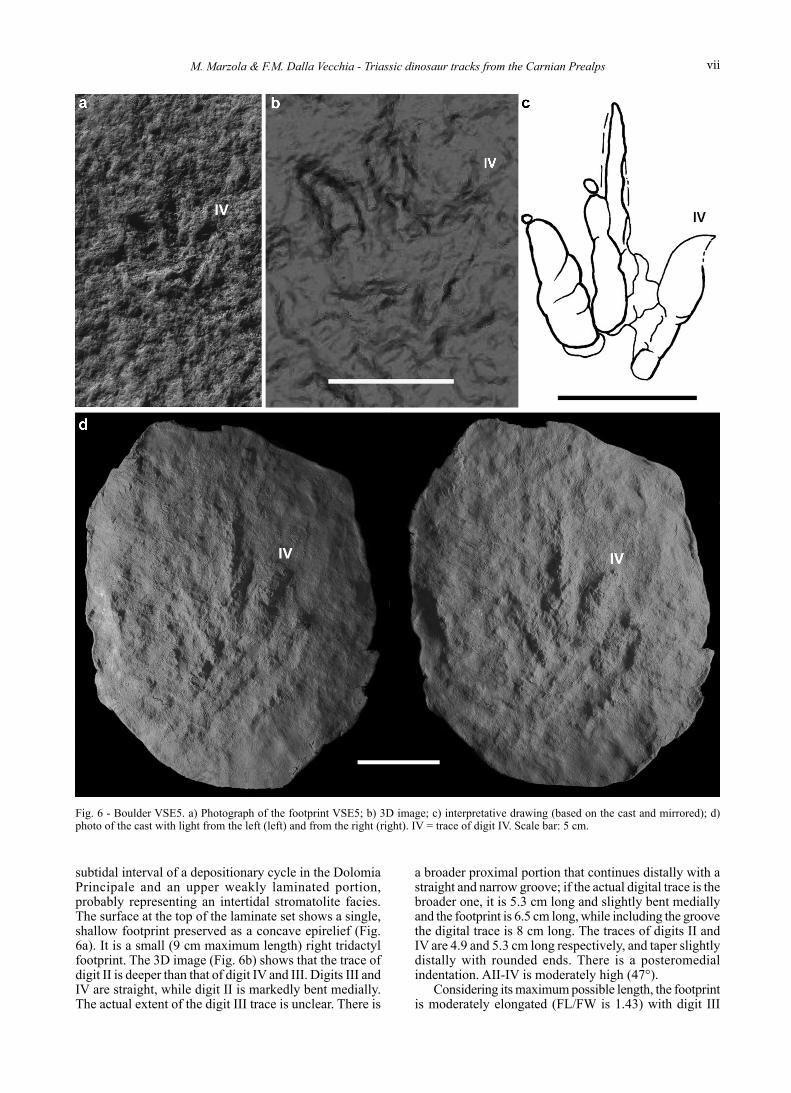
BoulDer VSe5 - (750 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.31°N, 12.53°E) (Fig. 6).
This boulder measures 70 x 60 x 60 cm and is made of a massive basal portion, probably corresponding to the
Fig. 4 - Boulder VSE3. a) Photograph of the footprint VSE3; b) photograph of the cast of the footprint (light from the left); c) interpretative drawing (based on the cast and mirrored). IV = trace of digit IV. Scale bar: 5 cm.
Fig. 5 - Boulder VSE4. a) Photograph of the footprint VSE4; b) interpretative drawing. Scale bar: 5 cm.
viiM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
subtidal interval of a depositionary cycle in the Dolomia Principale and an upper weakly laminated portion, probably representing an intertidal stromatolite facies. The surface at the top of the laminate set shows a single, shallow footprint preserved as a concave epirelief (Fig. 6a). It is a small (9 cm maximum length) right tridactyl footprint. The 3D image (Fig. 6b) shows that the trace of digit II is deeper than that of digit IV and III. Digits III and IV are straight, while digit II is markedly bent medially. The actual extent of the digit III trace is unclear. There is
a broader proximal portion that continues distally with a straight and narrow groove; if the actual digital trace is the broader one, it is 5.3 cm long and slightly bent medially and the footprint is 6.5 cm long, while including the groove the digital trace is 8 cm long. The traces of digits II and IV are 4.9 and 5.3 cm long respectively, and taper slightly distally with rounded ends. There is a posteromedial indentation. AII-IV is moderately high (47°).
Considering its maximum possible length, the footprint is moderately elongated (FL/FW is 1.43) with digit III
Fig. 6 - Boulder VSE5. a) Photograph of the footprint VSE5; b) 3D image; c) interpretative drawing (based on the cast and mirrored); d) photo of the cast with light from the left (left) and from the right (right). IV = trace of digit IV. Scale bar: 5 cm.

Bollettino della Società Paleontologica Italiana, 2014viii
trace being much longer than the other two and projecting far beyond their distal ends (Te = 4.3 cm). Considering the shorter option, the footprint would be practically as wide as long (FL/FW would be 1.03) and digit III trace would be very scarcely projecting far beyond the distal ends of the other two digits (Te = 1.5 cm).
BoulDer VSe6 - (650 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.29°N, 12.50°E) (Fig. 7).
This boulder measures 50 x 40 x 15 cm and is composed of light gray, massive dolostone. Its lower surface shows a very poorly preserved, relatively large (FL is 25.7 cm) tridactyl footprint as a convex hyporelief (Fig. 7a). It is probably a left footprint, because the trace of the left digit has a posterior phalangeal-metatarsal pad, indicating digit IV. This is one of the largest tracks found so far in the PNDF, the largest being that from the Casera Cjasavent locality (35 cm long; Dalla Vecchia & Mietto, 1998). It is scarcely elongated (FL/FW about 1.40) and the total digital divarication is 39° (if AIII-IV is measured based on the distal part of digit IV) or 53° (if AIII-IV is measured based on the chord of the whole, curved trace of digit IV). Digit III is slightly sigmoid in shape and the supposed digit IV is curved medially (Fig. 7b).
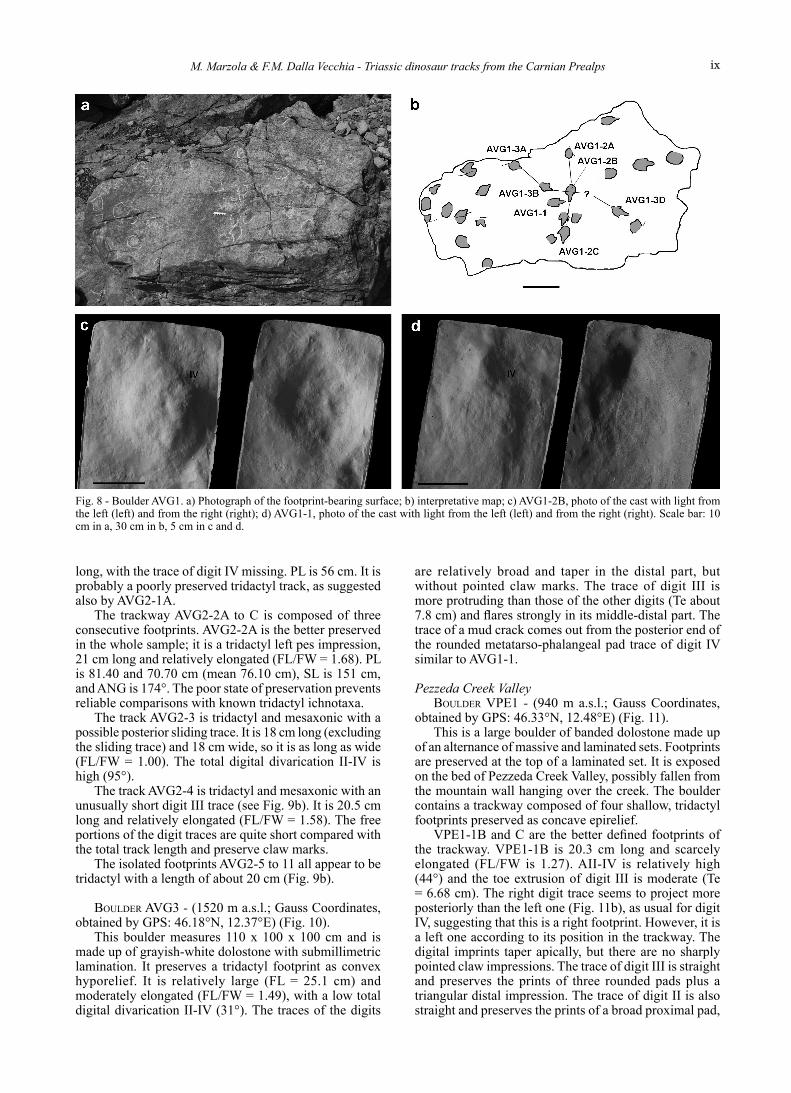
Unnamed creek, southern slope of Mt Caserine Basse BoulDer aVG1 - (1390 m a.s.l.; Gauss Coordinates,
obtained by GPS: 46.16°N, 12.35°E) (Fig. 8). This boulder is located in the “Ciolòn” locality. It
is made up of whitish dolostone with barely visible submillimetric lamination. The exposed top surface of the boulder measures 240 x 140 cm and presents several concave epireliefs among which at least a dozen can be identified as tridactyl footprints. Some of them are organized into two short trackway segments. All the footprints are poorly-preserved, and thus measurements are only indicative.
AVG1-2A to C is a trackway segment composed by three consecutive footprints. PL is 35 cm, SL is 69 cm, and ANG is 170°. AVG1-2B is the best preserved footprint; it is 13.5 cm long and moderately elongated (FL/FW = 1.55). Its outline is poorly defined, but the narrow and straight trace of digit III projecting forward is evident. The straight trace of an outer digit, which is elongated backward (Fig. 8c), can be identified as digit IV, in agreement with the position of the footprint in the trackway.
AVG1-3A to D is probably a trackway segment because AVG1-3A, AVG1-3B, and AVG1-3D are arranged in a row. However, there is no evidence of a preserved footprint between AVG1-3B and AVG1-3D that cannot be consecutive footprints because the gap between them is too large; PL between AVG1-3A and AVG1-3B is 37 cm, while the distance between AVG1-3B and AVG1-3D is 65 cm. As in AVG1-2, footprints are tridactyl and mesaxonic, but the poor state of preservation prevents further morphological description. AVG1-3B is 13 cm long and moderately elongated (FL/FW = 1.62).
AVG1-1 has the same size (FL= 13.5 cm) and the same general morphology as AVG1-2B (Fig. 8d). It is also a left footprint and moderately elongated (FL/FW = 1.61). The probable trace of a mud crack comes out from the proximal termination of the impression of digit IV.
BoulDer aVG2 - (1500 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.18°N, 12.37°E) (Fig. 9).
This boulder measures 250 x 130 x100 cm and is made up of whitish dolostone with submillimetric lamination. The lower surface of the boulder shows 18 tracks preserved as convex hyporeliefs (casts); two consecutive footprints (AVG2-1A to B) and a trackway (AVG2-2A to C) can be distinguished in the sample (Fig. 9b).
AVG2-1A preserves only its distal part, the long digit III print having a remarkably protruding relief. AVG2-1B is apparently didactyl and probably a left footprint, 18 cm
Fig. 7 - Boulder VSE6. a) Photograph of the boulder with footprint under artificial light from the left (the footprint is enlightened with a broken ellipsoid line and all the three digits are marked). b) Interpretative drawing. II = trace of digit II; III = trace of digit III; IV = trace of digit IV. Scale bar: 5 cm.
ixM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
long, with the trace of digit IV missing. PL is 56 cm. It is probably a poorly preserved tridactyl track, as suggested also by AVG2-1A.
The trackway AVG2-2A to C is composed of three consecutive footprints. AVG2-2A is the better preserved in the whole sample; it is a tridactyl left pes impression, 21 cm long and relatively elongated (FL/FW = 1.68). PL is 81.40 and 70.70 cm (mean 76.10 cm), SL is 151 cm, and ANG is 174°. The poor state of preservation prevents reliable comparisons with known tridactyl ichnotaxa.
The track AVG2-3 is tridactyl and mesaxonic with a possible posterior sliding trace. It is 18 cm long (excluding the sliding trace) and 18 cm wide, so it is as long as wide (FL/FW = 1.00). The total digital divarication II-IV is high (95°).
The track AVG2-4 is tridactyl and mesaxonic with an unusually short digit III trace (see Fig. 9b). It is 20.5 cm long and relatively elongated (FL/FW = 1.58). The free portions of the digit traces are quite short compared with the total track length and preserve claw marks.
The isolated footprints AVG2-5 to 11 all appear to be tridactyl with a length of about 20 cm (Fig. 9b).
BoulDer aVG3 - (1520 m a.s.l.; Gauss Coordinates, obtained by GPS: 46.18°N, 12.37°E) (Fig. 10).
This boulder measures 110 x 100 x 100 cm and is made up of grayish-white dolostone with submillimetric lamination. It preserves a tridactyl footprint as convex hyporelief. It is relatively large (FL = 25.1 cm) and moderately elongated (FL/FW = 1.49), with a low total digital divarication II-IV (31°). The traces of the digits
are relatively broad and taper in the distal part, but without pointed claw marks. The trace of digit III is more protruding than those of the other digits (Te about 7.8 cm) and flares strongly in its middle-distal part. The trace of a mud crack comes out from the posterior end of the rounded metatarso-phalangeal pad trace of digit IV similar to AVG1-1.
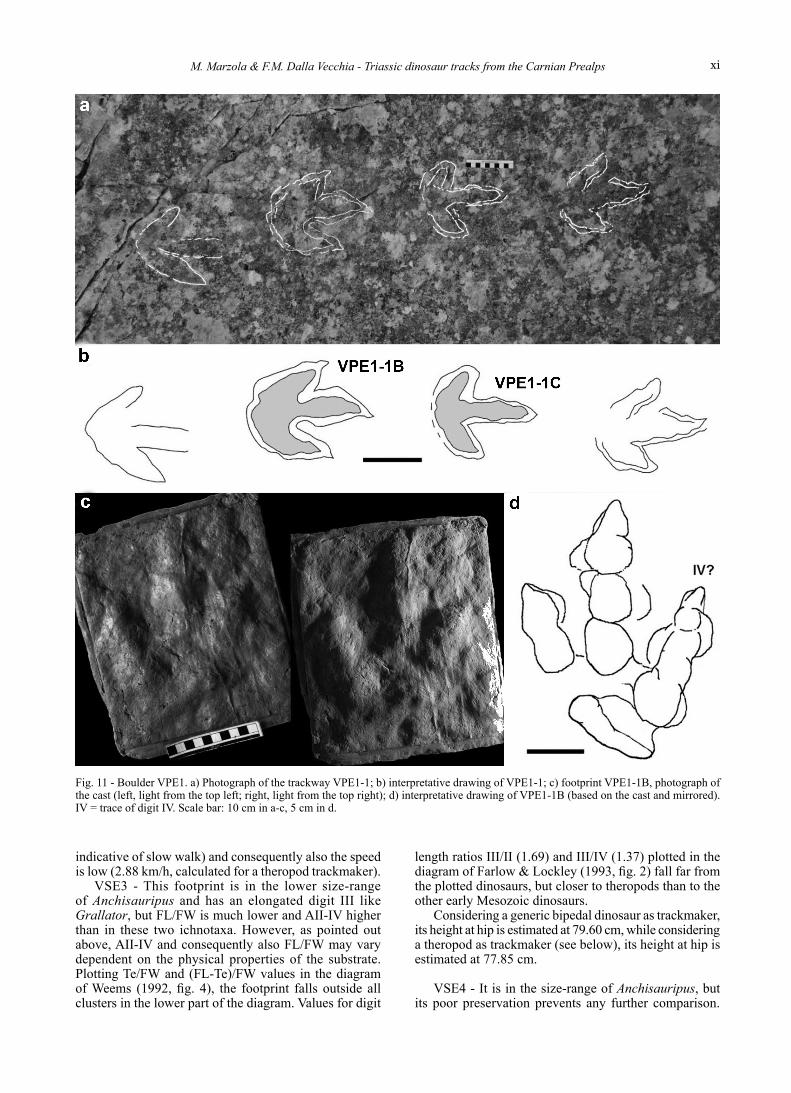
Pezzeda Creek ValleyBoulDer VPe1 - (940 m a.s.l.; Gauss Coordinates,
obtained by GPS: 46.33°N, 12.48°E) (Fig. 11). This is a large boulder of banded dolostone made up
of an alternance of massive and laminated sets. Footprints are preserved at the top of a laminated set. It is exposed on the bed of Pezzeda Creek Valley, possibly fallen from the mountain wall hanging over the creek. The boulder contains a trackway composed of four shallow, tridactyl footprints preserved as concave epirelief.
VPE1-1B and C are the better defined footprints of the trackway. VPE1-1B is 20.3 cm long and scarcely elongated (FL/FW is 1.27). AII-IV is relatively high (44°) and the toe extrusion of digit III is moderate (Te = 6.68 cm). The right digit trace seems to project more posteriorly than the left one (Fig. 11b), as usual for digit IV, suggesting that this is a right footprint. However, it is a left one according to its position in the trackway. The digital imprints taper apically, but there are no sharply pointed claw impressions. The trace of digit III is straight and preserves the prints of three rounded pads plus a triangular distal impression. The trace of digit II is also straight and preserves the prints of a broad proximal pad,
Fig. 8 - Boulder AVG1. a) Photograph of the footprint-bearing surface; b) interpretative map; c) AVG1-2B, photo of the cast with light from the left (left) and from the right (right); d) AVG1-1, photo of the cast with light from the left (left) and from the right (right). Scale bar: 10 cm in a, 30 cm in b, 5 cm in c and d.

Bollettino della Società Paleontologica Italiana, 2014x
two or three traces of smaller successive pads and a faint triangular distal trace; the proximal pad is contiguous with an elongated “heel” print. The trace of digit IV is short and broad with two pad prints and a faint, short ungual impression. The footprints seem to be lined up: the mean ANG is about 175°. Instead, mean PL and SL are quite short: 28.3 and 57 cm, respectively.
DISCUSSION
The ichnotaxonomic assignment and identification of the trackmakers of these very poorly to moderately well preserved footprints are tentative because their final morphology may be biased by the following preservation factors: 1) the manus imprints are possibly not preserved because they were more faintly impressed than the pes imprints and may have been wiped out more easily; 2) the medial and lateral digit traces (digits I and V) may be not preserved because originally they were less deeply impressed than digits II to IV (see King & Benton, 1996; Sarjeant, 1996). Extramorphological (substrate-related) influences on the track shape are extremely important but often have not been considered adequately in the studies, resulting in misidentifications as well as ichnotaxonomic oversplitting. The determination of
tetrapod ichnites should be strictly based on anatomical features that, especially in poorly preserved tracks, are hard to recognize.
Remarks on the new track-bearing bouldersVSe1 - The arrangement of the two tracks may be
explained as a partial trackway of a bipedal stationary trackmaker or could just be the result of accidental preservation of footprints left by different individuals. The preserved portion of the original surface is too small to ascertain it. No manus trace is observed on the boulder, but this could be due to the limited available surface and the poor conditions of preservation. By its size, VSE1-1 can be compared to the ichnogenus Anchisauripus, but it is less elongated and has a higher total divarication. However, as pointed out above, AII-IV as well as FL/FW may vary dependent on the conditions of the substrate. Plotting Te/FW and (FL-Te)/FW values of VSE1-1 in the diagram of Weems (1992, fig. 4), the footprint falls outside the clusters identified in that diagram.
Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 101.2 cm, while considering a theropod as trackmaker (see below), its height at hip is estimated at 99 cm.
VSe2 - An assignment of those footprints to Anchisauripus is in agreement with their size and total digit divarication, but the relative slenderness (FL/FW) and the projection of digit III are similar to those of Eubrontes (Olsen et al., 1998). Plotting Te/FW and (FL-Te)/FW values of VSE2-1A in the diagram of Weems (1992, fig. 4), the footprint falls within the margin of a cluster including Anchisauripus minusculus Hitchcock, 1858, “Eubrontes giganteus” Hitchcock, 1836, E. tuberatus (Hitchcock, 1858), and Gigandipus caudatus Hitchcock, 1856.
Values for digit length ratios III/II (1.36) and III/IV (0.91) in VSE2-1A plotted in the diagram of Farlow & Lockley (1993, fig. 2) fall between the plots of theropods (Coelophysis, Syntarsus, and Procompsognathus) and those of other early Mesozoic dinosaurs (Heterodontosaurus, Lufengosaurus, Plateosaurus, and Massospondylus).
Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 98.90 cm, while considering a theropod as trackmaker (see below), its height at hip is estimated at 96.75 cm. If the tracks were produced by the same foot, SL is 99 cm, SL/h is rather low (1-1.02,
Fig. 10 - Boulder AVG3. a) Photograph of footprint AVG3; b) interpretative drawing. IV = trace of digit IV. Scale bar: 10 cm.
Fig. 9 - Boulder AVG2. a) Photograph of the footprint-bearing surface under artificial light (from the right) in perspective view; b) interpretative map. Scale bar: 20 cm. Notice relatively short digit III and elongated heel in AVG2-4, possibly indicating incomplete chirotheriid or other tetradactyl to pentadactyl forms.
xiM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
indicative of slow walk) and consequently also the speed is low (2.88 km/h, calculated for a theropod trackmaker).
VSe3 - This footprint is in the lower size-range of Anchisauripus and has an elongated digit III like Grallator, but FL/FW is much lower and AII-IV higher than in these two ichnotaxa. However, as pointed out above, AII-IV and consequently also FL/FW may vary dependent on the physical properties of the substrate. Plotting Te/FW and (FL-Te)/FW values in the diagram of Weems (1992, fig. 4), the footprint falls outside all clusters in the lower part of the diagram. Values for digit
length ratios III/II (1.69) and III/IV (1.37) plotted in the diagram of Farlow & Lockley (1993, fig. 2) fall far from the plotted dinosaurs, but closer to theropods than to the other early Mesozoic dinosaurs.
Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 79.60 cm, while considering a theropod as trackmaker (see below), its height at hip is estimated at 77.85 cm.
VSe4 - It is in the size-range of Anchisauripus, but its poor preservation prevents any further comparison.
Fig. 11 - Boulder VPE1. a) Photograph of the trackway VPE1-1; b) interpretative drawing of VPE1-1; c) footprint VPE1-1B, photograph of the cast (left, light from the top left; right, light from the top right); d) interpretative drawing of VPE1-1B (based on the cast and mirrored). IV = trace of digit IV. Scale bar: 10 cm in a-c, 5 cm in d.

Bollettino della Società Paleontologica Italiana, 2014xii
Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 101.2 cm, while considering a theropod as trackmaker (see below), its height at hip is estimated at 99 cm.
VSE5 - If the actual morphology is that of the short digit III option, this footprint is different from any other Triassic tridactyl footprints. In the case of the groove representing the actual digit III trace, size matches Grallator, but AII-IV is too high for this ichnogenus as well as for Anchisauripus. However, AII-IV may vary dependent on the conditions of the substrate. Plotting Te/FW and (FL-Te)/FW values in the diagram of Weems (1992, fig. 4), the footprint falls outside all clusters in the lower part of the diagram. Values for digit length ratios III/II (1.63) and III/IV (1.51) plotted in the diagram of Farlow & Lockley (1993, fig. 2) fall far from the plotted dinosaurs, but closer to theropods than to the other early Mesozoic dinosaurs, like VSE3, probably because the footprint does not reflect the actual proportions of the digits. Considering the maximum footprint length and a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 41.40 cm, while considering a theropod (see below), its height at hip is estimated at 40.5 cm.
VSE6 - Size, broadness, total digital divarication and relative shortness of digit III are within the limits of the ichnogenus Eubrontes (Olsen et al., 1998). Plotting Te/FW and (FL-Te)/FW values in the diagram of Weems (1992, fig. 4), the footprint falls within the cluster of “Eubrontes giganteus”, E. divaricatus Hitchcock, 1865, E. approximatus Hitchcock, 1865, and E. platypus Hitchcock, 1858.
Values for digit length ratios III/II (2.13) and III/IV (1.58) plotted in the diagram of Farlow & Lockley (1993, fig. 2) fall far from the plotted dinosaurs, probably because the footprint does not reflect the actual proportions of the digits.
Considering a generic bipedal dinosaur as trackmaker, the height at hip is estimated at 118.20-146.50 cm (using the morphometric ratios method of Thulborn, 1990 for both small and large bipedal dinosaurs because the size boundary between the two groups is at FL = 25 cm). Considering a theropod as trackmaker (see below), its height at hip is estimated at 125.9-115.7 cm.
AVG1 - All footprints are in the size range of Grallator. Considering a generic bipedal dinosaur as trackmaker, the height at hip is estimated at 62.10, 59.80, and 62.10 cm for AVG1-2, AVG1-3 and AVG1-1, respectively; considering a theropod as trackmaker (see below), the height at hip is estimated at 60.75, 58.50 and 60.75 cm.
The gait was a slow walk according to SL/h (1.11/1.14 and 1.09/1.11 for AVG1-2 and AVG1-3, respectively) and V was 2.65/2.71 km/h and 2.50/2.57 km/h for AVG1-2 and AVG1-3, respectively.
AVG2 - The poor state of preservation of AVG2-1 prevents any comparisons with known tridactyl ichnotaxa; however, AVG2-1B is in the size-range of Anchisauripus. Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 82.80 cm, while considering a theropod as trackmaker (see below), its height at hip is estimated at 81 cm.
AVG2-2A is in the size-range of Anchisauripus, but it is slightly less elongated than footprints referred to that ichnogenus. Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 96.60 cm, while considering a theropod as trackmaker (see below), its height at hip is estimated at 94.50 cm. SL/h is 1.56/1.60 and V is 5.84/5.99 km/h.
The high total divarication and low FL/FW of AVG2-3 are unlike those of the other tridactyl footprints in the sample as well as the tridactyl ichnotaxa Grallator, Anchisauripus and Eubrontes. They are more similar to those of Anomoepus, but there is no trace of the manus associated and the state of preservation does not allow to observe the diagnostic features of this Jurassic ichnogenus (Olsen & Rainforth, 2003). Considering the maximum footprint length and a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 82.80 cm; considering a theropod as the trackmaker (see below), its height at hip is estimated at 81 cm.
If the length of the digit III trace of AVG2-4 reflects the anatomical length of the trackmaker’s digit, AVG2-4 might be an incomplete chirotheriid track or an incomplete track of another pentadactyl or tetradactyl ichnogenus (but see Klein et al., 2006; Klein & Haubold, 2007).
The isolated footprints AVG2-5 to 11 are in the size-range of Anchisauripus. The height at hip of the trackmakers is estimated at 92 cm if they were generic bipedal dinosaurs, at 90 cm if theropods.
AVG3 - This footprint is positioned at the boundary between the size-range of Eubrontes and Anchisauripus. Its total digital divarication II-IV is in the range of both ichnotaxa, but its width is larger than that of Anchisauripus.
Plotting Te/FW and (FL-Te)/FW values in the diagram of Weems (1992, fig. 4), the footprint falls within the cluster of Anchisauripus minusculus, “Eubrontes giganteus”, E. tuberatus and Gigandipus caudatus. Values for digit length ratios III/II (1.69) and III/IV (1.04) plotted in the diagram of Farlow & Lockley (1993, fig. 2) fall close to the plotted values of the theropods.
Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 115.5-143.1 cm (using the morphometric ratios method of Thulborn, 1990 for both small and large bipedal dinosaurs because the size boundary between the two groups is at FL = 25 cm); considering a theropod as the trackmaker (see below), its height at hip is estimated at 113-123 cm.
VPE1 - The tracks are in the size-range of Anchisauripus, but the relative broadness (FL/FW) of VPE1-1B, its total divarication and the moderate projection of digit III are more similar to those of Eubrontes.
Plotting Te/FW and (FL-Te)/FW values in the diagram of Weems (1992, fig. 4), the footprint falls in the lower part of the cluster of “Eubrontes giganteus”, E. divaricatus, E. approximatus and E. platypus. Values for digit length ratios III/II (1.12) and III/IV (1.65) plotted in the diagram of Farlow & Lockley (1993, fig. 2) fall outside the diagram space; if the specimen is a right footprint (with III/II = 1.65 and III/IV = 1.12) it would be close to the plotted values of the theropods.
Considering a generic bipedal dinosaur as trackmaker, its height at hip is estimated at 93.40 cm; considering a

xiiiM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
theropod as the trackmaker (see below), its height at hip is estimated at 91.35 cm. It was moving very slowly (SL/h is 0.61/0.62) and its speed was very low (V = 1.19/1.22 km/h); this contrasts with the high pace angulation (see Alexander, 1976; Demathieu, 1984).
THE IDENTIFICATION OF POSSIBLE TRACKMAKERS
Most of the new tracks from the PNDF appear to have been left by functionally tridactyl, digitigrade and bipedal trackmakers. Small to medium-sized tridactyl footprints with slender, pointed digit traces and low total digital divarication, along trackways produced by bipeds were traditionally referred to theropod dinosaurs (e.g., Haubold, 1971; Thulborn, 1990; Lockley, 1991). In the post-Triassic record, the only alternative trackmakers for those footprints are bipedal ornithischians (Thulborn, 1990; Lockley, 1991).
However, the situation is much more complicated when the Upper Triassic record is considered. Here we have to deal with a high diversity of potential bipedal trackmakers: these include dinosaurs such as basal saurischians, basal theropods, basal ornithischians and possibly basal sauropodomorphs (see Carrano, 2000; Langer, 2003; Butler et al., 2007; Langer et al., 2010; Martinez et al., 2011), non-dinosaurian dinosauromorphs (Padian, 1997; Langer et al., 2010; Piechowski & Dzik, 2010; Klein et al., 2011), and even pseudosuchian archosaurs (Nesbitt & Norell, 2006; Nesbitt, 2007; Gauthier et al., 2011). Furthermore, the systematic position of many Triassic dinosaurs is debatable: for example, Herrerasaurus ischigualastensis Reig, 1963 was considered a basal theropod by Sereno et al. (1993), Sereno & Novas (1994), Sereno (2007), Martinez et al. (2011) and Sues et al. (2011), but a basal saurischian by Langer (2004), Langer & Benton (2006), and Novas et al. (2011).
Another difficulty is the incomplete record of skeletons of potential bipedal trackmakers: usually, the autopodia of Late Triassic archosaurs are only fragmentarily preserved or completely missing, therefore, our knowledge about their anatomy and digit proportions is restricted (e.g., Walker, 1964; Casamiquela, 1967; Romer, 1972a; Krebs, 1976; Chatterjee, 1984, 1987; Long & Murry, 1995; Hunt et al., 1998; Langer et al., 1999; Sullivan & Lucas, 1999; Langer, 2004; Butler et al., 2007; Ferigolo & Langer, 2007; Galton, 2007; Irmis et al., 2007a, b; Nesbitt, 2007, 2011; Nesbitt et al., 2009, 2010; Martínez & Alcober, 2009; Ezcurra, 2010; Kammerer et al., 2011; Novas et al., 2011; Sues et al., 2011).
Tracks of the Grallator-Eubrontes type are usually assigned to theropod trackmakers (Olsen et al., 1998; Klein & Lucas, 2010). Haubold & Klein (2000, 2002), Klein & Haubold (2007), and Klein et al. (2011) refer the footprints of the Grallator-Atreipus plexus to dinosauromorphs. The Dinosauromorpha are composed of the Lagerpetidae and the Dinosauriformes, the latter including Lewisuchus admixtus Romer, 1972a, Marasuchus lilloensis (Romer, 1972b), Saltopus elginensis von Huene, 1910, the Silesauridae, and the Dinosauria (Langer et al., 2013). A dinosauromorph affinity of some Middle Triassic footprints referred to “Anchisauripus” and “Coelurosaurichnus” is
also considered by some authors (e.g., Gand & Demathieu, 2005). Atreipus from the Upper Triassic of Europe and North America has been attributed to an ornithischian dinosaur or a basal dinosauriform by Olsen & Baird (1986), to the Theropoda by Thulborn (1993), to the Ornithischia by Haubold (1986) and Weishampel (2006), to basal Dinosauriformes by Haubold & Klein (2000), and to the Dinosauriformes in general by Irmis et al. (2007a) and Nesbitt et al. (2007). Atreipus is also interpreted as a probable dinosauriform ichnite by D’Orazi Porchetti et al. (2008). As seen above, most of the tridactyl footprints preserved on the boulders from the PNDF and described herein are in the size range of Anchisauripus (Tab. 1) and do not show an associated manus imprint.
Taking the skeletal reconstruction of Herrerasaurus ischigualastensis (see Paul, 2010, p. 69) as a model for a generalized dinosauromorph bipedal trackmaker and using the hip height calculation model of Thulborn (1990, fig. 8.4) referred to generic bipedal dinosaurs (Thulborn, 1990, p. 251), the total body length of the producer of the largest footprint in our sample (VSE6, 25.7 cm long) would be in the range 4.42-5.48 m, while that of the smaller one (VSE5, 9 cm long) would be 1.55 m. Trackmakers of the most abundant footprints in our sample (the Anchisauripus-size class) would have a total body length ranging between 3.10 and 3.70 m (mean 3.53 m). Many Triassic bipedal dinosaurs (e.g., Pisanosaurus mertii Casamiquela, 1967, Eocursor parvus Butler, Smith & Norman, 2007, Eoraptor lunensis Sereno, Forster, Rogers & Monetta, 1993, Alwalkeria maleriensis [Chatterjee, 1987], Guaibasaurus canderlariensis Bonaparte, Ferigolo & Ribeiro, 1999, Staurikosaurus pricei Colbert, 1970, Eodromaeus murphi Martínez, Sereno, Alcober, Colombi, Renne, Montañez & Currie, 2011, Saturnalia tupiniquim Langer, Abdala, Richter & Benton, 1999, Panphagia protos Martínez & Alcober, 2009, and Asylosaurus yalensis Galton, 2007) have estimated body lengths ≤ 2 m (Paul, 2010; Martínez et al., 2011). Also, silesaurids (Silesaurus opolensis Dzik, 2003, Eucoelophysis baldwini Sullivan & Lucas, 1999, Technosaurus smalli Chatterjee, 1984, Sacisaurus agudoensis Ferigolo & Langer, 2007 and Diodorus scytobrachion Kammerer, Nesbitt & Shubin, 2011) (Dzik, 2003; Langer et al., 2010) and the lagerpetonids Dromomeron gregorii Irmis, Nesbitt, Padian, Smith, Turner, Woody & Downs, 2007b and D. romeri Nesbitt, Irmis, Parker, Smith, Turner & Rowe, 2009, which were probably facultative bipeds, were ≤ 2 m in length. They seem to be too small for most of the PNDF footprints.
The largest tridactyl footprints of the PNDF are in the range of Herrerasaurus ischigualastensis (total body length 4.5 m; Paul, 2010), the bipedal pseudosuchian archosaur Poposaurus gracilis Mehl, 1915 (total body length about 5 m; see Schachner et al., 2011), and the theropod Liliensternus liliensterni von Huene, 1934 (total body length 5.2 m; Paul, 2010). However, Herrerasaurus ischigualastensis has a scarcely projecting digit III and a comparatively long digit I, whose impression should be visible, at least in well-preserved footprints. Poposaurus gracilis probably was also tetradactyl (see Schachner et al., 2011). Liliensternus liliensterni has a scarcely projecting digit III, but apparently had a reduced digit I (Rowe & Gauthier, 1990). Most of the tridactyl footprints in the sample are in the size range of the herrerasaurid

Bollettino della Società Paleontologica Italiana, 2014xiv
Chindesaurus bryansmalli Long & Murry, 1995 (total body length 2.4 m; Paul, 2010) and the basal theropod Coelophysis bauri (Cope, 1887) (total body length up to 3 m; Paul, 2010) or close to it. Unfortunately, the pes of Chindesaurus bryansmalli is unknown (Long & Murry, 1995). Coelophysis bauri has a digit III projecting far beyond digits II and IV and a short digit I that barely touched the ground (Colbert, 1989). Thus, at the present state of knowledge, theropod dinosaurs are still the best trackmaker candidates for tridactyl footprints described here.
THE PNDF FOOTPRINTS WITHIN THE LATE TRIASSIC ICHNOASSOCIATIONS
Previously reported footprints from the PNDF included tridactyl and mesaxonic footprints within the Anchisauripus size class (sensu Olsen et al., 1998) (a total of three short trackways and at least 10 isolated tracks from Ciol della Fratta A, Ciol del Tramontin and Upper Susaibes Creek localities; Dalla Vecchia & Mietto, 1998, figs 5-6 and 9-11) and two consecutive tridactyl footprints within the Eubrontes size class (Dalla Vecchia & Mietto, 1998, fig. 2). Dalla Vecchia & Mietto (1998) noted the similarity of a 18 cm long, single tetradactyl footprint (Dalla Vecchia & Mietto, 1998, fig. 6) from Ciol della Fratta. A locality with the ichnospecies Sphingopus ferox Demathieu, 1966, which, however, is typical of the Middle Triassic according to Klein & Haubold (2007). Alternatively, this specimen could be a tridactyl footprint like the others present in the same surface, which is casually associated with a close but distinct depression, or an incomplete chirotheriid pes imprint.
A trackway with footprints made apparently by a bipedal trackmaker was reported by Dalla Vecchia & Mietto (1998) from Scandoler Valley (Dalla Vecchia & Mietto, 1998, figs 3-4); footprints are 18 cm long, apparently not tridactyl, with outer rotation (8°-28°) and relatively low pace angulation (140°-160°). However, because of logistic problems (the footprint-bearing surface was vertical), those authors could not observe the trackway directly and they described it from photographs.
A single 22-23 cm long track from the Upper Susaibes Creek locality is apparently pentadactyl (Dalla Vecchia & Mietto, 1998, fig. 10); Dalla Vecchia & Mietto (1998) and Dalla Vecchia (2006) noticed the resemblance with the ichnogenera Pseudotetrasauropus Ellenberger, 1972, Otozoum Hitchcock, 1847, and Evazoum Nicosia & Loi, 2003. Those ichnotaxa are, however, tetradactyl and considered extramorphological variants of Brachychirotherium Beurlen, 1950 by some authors (e.g., Klein et al., 2006; Klein & Haubold, 2007). The track from the Upper Susaibes Creek locality, if actually pentadactyl, is not a chirotheriid footprint because of the position and morphology of the digit V trace.
A relatively wide quadrupedal trackway from Ciol della Fratta B locality with pes tracks 27-28 cm long (Dalla Vecchia & Mietto, 1998, fig. 7) could be a chirotheriid trackway, but the poor preservation of the tracks does not allow a positive referral based on the identification of the diagnostic morphological features of chirotheriid footprints.
A short, quadrupedal trackway segment from the Upper Susaibes Creek locality (Dalla Vecchia & Mietto, 1998, fig. 11) might also be assigned to chirotheriids; unfortunately, only the small (13 cm long) tetradactyl trace of the manus is well preserved, while the pes prints are misshapen.
Finally, an apparently quadrupedal trackway with large tridactyl pes prints 32-40 cm long and much smaller manus traces (Dalla Vecchia & Mietto, 1998, fig. 8) was reported from Forcella delle Pregoiane (Claut) by Dalla Vecchia & Mietto (1998). However, those authors doubt about the real identity of these structures as footprints because they could not observe them directly.
The Norian-Rhaetian ichno-associations worldwide are dominated by tridactyl tracks of the Grallator-Eubrontes type (Grallator including Anchisauripus) and chirotheriid tracks referred to Brachychirotherium (sensu stricto) (Gatesy et al., 1999; Klein & Haubold, 2007; Klein & Lucas, 2010; Niedzwiedzki, 2011). The PNDF assemblage is distinguished by the rarity or possible absence of chirotheriid footprints and by the dominance of middle-sized (Anchisauripus size-class) tridactyl footprints. With one possible exception, tetradactyl footprints like Tetrasauropus Ellenberger, 1970, Pseudotetrasauropus, Otozoum, and Evazoum that might be referred to basal sauropodomorphs, are absent. On the contrary, basal sauropodomorphs are the most common dinosaurs in the Norian-Rhaetian terrestrial tetrapod associations of Europe based on skeletal remains (Weishampel et al., 2004).
The other footprint-bearing localities of the Dolomia Principale in NE Italy have yielded small to large-sized tridactyl footprints mostly referred to theropods, trackways and some tetradactyl footprints attributed to basal sauropodomorphs, a trackway referred to a basal ornithischian, and a single chirotheriid footprint (Mietto, 1988, 1991; Dalla Vecchia & Mietto, 2000; Leonardi, 2000; Belvedere et al., 2008).
A block at Mt Pelmetto locality (Belluno Dolomites, Belluno Province) preserves four trackways and some isolated tridactyl footprints belonging to the Grallator size-class, which were referred to small theropods (Mietto, 1988). The same block preserves also two trackways assigned to a small basal sauropodomorph and to a small basal ornithischian, respectively (Mietto, 1988; Leonardi, 2000). Several small to large-sized tridactyl footprints and the trackway of a quadrupedal trackmaker composed of subelliptical footprints were recorded in other blocks (Dalla Vecchia & Mietto, 2000). A “chirotheroid” footprint referred to Brachychirotherium sp. was found on the opposite side of the main block (Dalla Vecchia & Mietto, 2000, p. 330). Two large tridactyl footprints referred to Eubrontes are reported from the Tre Cime di Lavaredo (Belluno Dolomites, Belluno Province; Mietto, 1991; Leonardi, 2000) and a medium-sized tridactyl footprint is reported from Puez (western Dolomites, Bolzano/Bozen Province; Leonardi, 2000). Finally, the “Strada delle Gallerie” site (Pasubio Massif, Vicenza Province) yielded large (Eubrontes) and small (cf. Grallator) tridactyl footprints referred to theropods and tetradactyl footprints similar to Pseudotetrasauropus and Evazoum assigned to basal sauropodomorphs (Belvedere et al., 2008).

xvM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
As in the PNDF ichnoassociation, chirotheriid footprints and small (< 15 cm) tridactyl footprints seem to be uncommon, being present essentially in the Mt Pelmetto site.
The sample of the PNDF is the largest described sample of the Dolomia Principale ichnofauna and probably the most representative of its paleoichnological diversity. However, it must be kept on mind that the Dolomia Principale spans the uppermost Carnian to the Rhaetian, an interval of approximately 27 million years. Only the site of Pasubio Massif has been dated on biostratigraphic bases (conodonts) to the latest middle Norian (Belvedere et al., 2008), while the stratigraphic position of the ichnofossils from Mt Pelmetto at the base of the formation would suggest a latest Carnian age for them (Mietto, 1988). Thus, it is possible - although not demonstrated - that the vertebrate paleoichnological record of the Dolomia Principale averages different tetrapod faunas living in the area during different intervals of a wide time span, as suggested also by Belvedere et al. (2008).
CONCLUSIONS
The most common trackmakers roaming on the Late Triassic tidal flats of the Dolomia Principale in the PNDF were bipeds with a functionally tridactyl pes and a total body length around 3.5 m, occasionally around 5 m. According to the limited fossil record of skeletal autopodia of bipedal or facultative bipedal Triassic archosaurs, the best candidate trackmakers for those footprints are basal theropods. Norian-Rhaetian ichnoassociations worldwide are characterised by the presence of tridactyl mesaxonic tracks (Grallator-Eubrontes type), but also by chirotheriid tracks, which appear to be infrequent in the whole PNDF sample.
The vast majority of footprints preserved on the ten new footprint-bearing boulders described in this paper are tridactyl mesaxonic and apparently impressed by digitigrade and bipedal trackmakers. Most of these footprints range 15-25 cm in length as the ichnogenus Anchisauripus according to Olsen et al. (1998), smaller footprints are more scarce and only two specimens are slightly longer than 25 cm. Only a single track might be a chirotheriid one.
Furthermore, tracks potentially produced by basal sauropodomorphs, which were the most common dinosaurs in the Norian-Rhaetian interval, are also rare, as are tridactyl and tetradactyl footprints of small-sized dinosauromorphs.
ACKNOWLEDGEMENTS
This work is mostly the result of the Master thesis work of the corresponding author (MM), undertaken at the Università degli Studi di Ferrara during the years 2010-2012. Tutor was Benedetto Sala, co-tutors Giuseppe Muscio and the other author of this paper (FMDV). We thank Mauro Caldana, collaborator of the PNDF and President of the Associazione Naturalistica Cordenonese, for the discovery of the boulders, sharing all the information in his possess, the casting of some footprints, and guiding us to the fossil-bearing localities. We are grateful to Giuseppe Muscio, Museo Friulano di Storia Naturale of Udine, for the collaboration. Thanks to Octávio
Mateus (Universidade Nova de Lisboa - GEAL Museu da Lourinhã, Portugal) for a preliminary critical review of the manuscript and to the reviewers Massimo Bernardi (MUSE Museo delle Scienze di Trento, Italy) and Hendrik Klein (Saurierwelt Paläontologisches Museum Neumarkt, Germany) for their useful comments that improved the original manuscript.
REFERENCES
Alexander R.McN. (1976). Estimates of speeds of dinosaurs. Nature, 261: 129-130.
Belvedere M., Avanzini M., Mietto P. & Rigo M. (2008). Norian dinosaur footprints from the “Strada delle Gallerie” (Monte Pasubio, NE Italy). In Avanzini M. & Petti F. M. (eds), Italian Ichnology, Proceedings of the Ichnology session of Geoitalia 2007. Studi Trentini di Scienze Naturali, Acta Geologica, 83: 267-275.
Berra F., Delfrati L. & Ponton M. (2007). Dolomia Prinicipale. In Cita M.B., Abbate E., Aldighieri B., Balini M., Conti M.A., Falorni P., Germani D., Groppelli G., Manetti P. & Petti F.M. - Carta Geologica d’Italia 1:50.000. Catalogo delle formazioni - Unità tradizionali (1). Quaderni del Servizio Geologico d’Italia, serie III, 7: 63-72.
Beurlen K. (1950). Neue Fährtenfunde aus der fränkischen Trias. Neues Jahrbuch für Geologie und Paläontologie - Monatshefte, 1950: 308-320.
Bonaparte J.F., Ferigolo J. & Ribeiro A.M. (1999). A new early Late Triassic saurischian dinosaur from Rio Grande do Sul state, Brazil. In Tomida Y., Rich T.H. & Vickers-Rich P. (eds), Proceedings of the Second Gondwanan Dinosaur Symposium, National Science Museum Monographs,15: 89-109.
Bosellini A. (1967). La tematica deposizionale della Dolomia principale (Dolomiti e Prealpi venete). Bollettino della Società Geologica Italiana, 86: 133-169.
Bosellini A. & Hardie L.A. (1988). Facies e cicli dolomitici della Dolomia Principale delle Alpi Venete. Memorie della Società Geologica Italiana, 30: 245-266.
Braga Gp., Carloni G.C., Colantoni P., Corsi M., Cremonini G., Frascari F., Locatelli D., Monesi A., Pisa G., Sassi F.P., Selli R., Vai G.B., Zirpoli G. (1971). Note illustrative della carta geologica d’Italia alla scala 1:100.000. Fogli 4c-13. Monte Cavallino-Ampezzo. 108 pp. Nuova Tecnica Grafica, Roma.
Butler R.J., Smith R.M.H. & Norman D.B. (2007). A primitive ornithischian dinosaur from the Late Triassic of South Africa, and the early evolution and diversification of Ornithischia. Proceedings of the Royal Society B, 274: 2041-2046.
Carrano M.T. (2000). Homoplasy and the evolution of dinosaur locomotion. Paleobiology, 26: 489-512.
Carulli G.B. (ed.) (2006). Carta geologica del Friuli Venezia Giulia, scala 1:150.000, Regione Autonoma Friuli Venezia Giulia, Direzione Regionale Ambiente e Lavori Pubblici, Servizio Geologico Regionale.
Carulli G.B., Cozzi A., Longo Salvador G., Pernancic E., Podda F. & Ponton M. (2000). Geologia delle Prealpi Carniche. Pubblicazioni del Museo Friulano di Storia Naturale, 44: 1-47 + geological map at scale 1:50,000, Udine.
Casamiquela R.M. (1967). Un nuevo dinosaurio ornitisquio triásico (Pisanosaurus mertii; Ornithopoda) de la Formación Ischigualasto. Ameghiniana, 4: 47-64.
Chatterjee S. (1984). A new ornithischian dinosaur from the Triassic of North America. Naturwissenschaften, 71: 630-631.
Chatterjee S. (1987). A new theropod dinosaur from India with remarks on the Gondwana-Laurasia connection in the Late Triassic. In McKenzie G.D. (ed.), Gondwana 6: Stratigraphy, Sedimentology and Paleontology. Geophysical Monographs, 41: 183-189.
Colbert E.H. (1970). A saurischian dinosaur from the Triassic of Brazil. American Museum Novitates, 2405: 1-39.
Colbert E.H. (1989). The Triassic dinosaur Coelophysis. Bulletin of the Museum of North Arizona, 57: 1-160.

Bollettino della Società Paleontologica Italiana, 2014xvi
Cope E.D. (1887). The dinosaurian genus Coelurus. American Naturalist, 21: 367-369.
Dalla Vecchia F.M. (2002). Terrestrial reptiles in the Norian of the Carnian Pre-Alps (Friuli, NE Italy) - Paleoenvironmental implications. Memorie della Società Geologica Italiana, 57: 101-106.
Dalla Vecchia F.M. (2006). The tetrapod fossil record from the Norian-Rhaetian of Friuli (northeastern Italy). In Harris J.D., Lucas S.G., Spielmann J.A., Lockley M.G., Milner A.R.C. & Kirkland J.I. (eds), The Triassic-Jurassic terrestrial transition. New Mexico Museum of Natural History & Science Bulletin, 37: 432-444.
Dalla Vecchia F.M. (2008). The impact of dinosaur palaeoichnology in palaeoenvironmental and palaeogeographic reconstructions - the case of the Periadriatic carbonate platforms. Oryctos, 8: 89-106.
Dalla Vecchia F.M. & Mietto P. (1998). Impronte di rettili terrestri nella Dolomia Principale (Triassico Superiore) delle Prealpi Carniche (Pordenone, Friuli). Atti Ticinesi di Scienze della Terra, 7: 87-107.
Dalla Vecchia F.M. & Mietto P. (2000). L’icnosito del Monte Pelmetto (Triassico superiore, Cadore, Italia). Riassunti delle comunicazioni orali e dei posters - 80° Riunione Estiva S.G.I., Trieste 6-8 settembre 2000: 329-330.
Demathieu G. (1966). Rhynchosauroides petri et Sphingopus ferox, nouvelles empreintes de Reptiles de grès triasiques de la bordure Nord-Est du Massif Central. Comptes Rendus de l’Académie des Sciences, Série D, 263: 483-486.
Demathieu G. (1984). Une ichnofaune du Trias moyen du Bassin de Lodéve (Hérault, France). Annales de Paléontologie, 70: 247-273.
Diàz-Martínez I., Pérez-Lorente F., Canudo J.I. & Pereda-Suberbiola X. (2009). Causas de la variabilidad en icnitas de dinosaurios y su aplicación en icnotaxonomía, Actas de las IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes, Burgos, Spain. Colectivo Arqueológico-Paleontológico Salense: 207-220.
D’Orazi Porchetti S., Nicosia U., Mietto P., Petti F.M. & Avanzini M. (2008). Atreipus-like footprints and their co-occurrence with Evazoum from the upper Carnian (Tuvalian) of Trentino-Alto Adige. In Avanzini M. & Petti F. M. (eds), Italian Ichnology, Proceedings of the Ichnology session of Geoitalia 2007. Studi Trentini di Scienze Naturali, Acta Geologica, 83: 277-287.
Dzik J. (2003). A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland. Journal of Vertebrate Paleontology, 23: 556-574.
Ellenberger P. (1970). Les niveaux paleontologiques de premiere apparition des Mammiferes Primordiaux en Afrique du Sud et leur Ichnologie: Establissement de zones stratigraphiques detaillees dans le Stormberg du Lesotho, (Afrique du Sud) (Trias Superieur a Jurassique). In Haughton S.H. (ed.), Z.U.G.S., 2nd
Symposium on Gondwana Stratigraphy and Palaeontology, Pretoria, Council for Scientific and Industrial Research: 343-370.
Ellenberger P. (1972). Contribution à la classification des Pistes de Vertébrés du Trias: Les types du Stormberg d’Afrique du Sud (I). Palaeovertebrata. 104 pp. Memoire Extraordinaire.
Ezcurra M.D. (2010). A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny. Journal of Systematic Palaeontology, 8: 371-425.
Farlow J.O. & Lockley M.G. (1993). An osteometric approach to the identification of the makers of early Mesozoic tridactyl dinosaur footprints. In Lucas S.G. & Morales M. (eds), The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin, 3: 123-131.
Ferigolo J. & Langer M.C. (2007). A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Historical Biology, 19: 23-33.
Galton P.M. (2007). Notes on the remains of archosaurian reptiles, mostly basal sauropodomorph dinosaurs, from the 1834 fissure
fill (Rhaetian, Upper Triassic) at Clifton in Bristol, southwest England. Revue de Paléobiologie, 26: 505-591.
Gand G. & Demathieu G. (2005). Les pistes dinosauroides du Trias moyen francais: interprétation et réévaluation de la nomenclature. Geobios, 38: 725-749.
Gatesy S.M., Middleton K.M., Jenkins F.A. & Shubin N.H. (1999). Three-dimensional preservation of foot movements in Triassic theropod dinosaurs. Nature, 399: 141-143.
Gauthier J.A., Nesbitt S.J., Schachner E.R., Bever G.S. & Joyce W.G. (2011). The bipedal stem crocodilian Poposaurus gracilis: inferring function in fossils and innovation in Archosaur locomotion. Bulletin of the Peabody Museum of Natural History, 52: 107-126.
Haubold H. (1971). Handbuch der Paläoherpetologie. T.18. Ichnia Amphibiorum et Reptiliorum fossilium. 124 pp. G. Fischer Verlag, Stuttgart.
Haubold H. (1986). Archosaur footprints at the terrestrial Triassic-Jurassic transition. In Padian K. (ed.), The beginning of the age of dinosaurs, Cambridge University Press: 189-201.
Haubold H. & Klein H. (2000). Die dinosauroiden Fährten Parachirotherium-Atreipus-Grallator aus dem unteren Mittelkeuper (Obere Trias: Ladin, Karn, ?Nor) in Franken. Hallesches Jahrbuch für Geowissenschaften B, 22: 59-85.
Haubold H. & Klein H. (2002). Chirotherien und Grallatoriden aus der Unteren bis Oberen Trias Mitteleuropas und die Entstehung der Dinosauria. Hallesches Jahrbuch für Geowissenschaften B, 24: 1-22.
Hitchcock, E. (1836). Ornithichnology - description of the foot marks of birds, (Ornithichnites) on new Red Sandstone in Massachusetts. The American Journal of Science and Arts, 29: 307-340.
Hitchcock E. (1845). An attempt to name, classify, and describe the animals that made the fossil footmarks of New England. Proceedings of the 6th Annual Meeting of the Association of American Geologists and Naturalists, 6: 23-25.
Hitchcock E. (1847). Description of two new species of fossil footmarks found in Massachusetts and Connecticut, or, of the animals that made them. The American Journal of Science and Arts, series 2, 4: 46-57.
Hitchcock E. (1848). An attempt to discriminate and describe the animals that made the fossil footmarks of the United States, and especially of New England. Memoirs of the American Academy of Arts and Sciences, 3: 129-256.
Hitchcock E. (1856). Description of a new and remarkable species of fossil footmark, from the sandstone of Turner’s falls, in the Connecticut Valley. American Journal of Science, ser. 2, 21: 97-100.
Hitchcock E. (1858). Ichnology of New England. A report on the sandstone of the Connecticut Valley, especially its fossil footmarks. 214 pp. William White, Boston.
Hitchcock E. (1865). Supplement to the Ichnology of New England. 96 pp. Wright and Potter, Boston.
Huene F. von (1910). Ein primitiver Dinosaurier aus der mittleren Trias von Elgin. Geologie und Paläontologie, Abhandlungen, 8: 317-322.
Huene F. von (1934). Ein neuer Coelurosaurier in der thüringischen Trias. Paläontologische Zeitschrift, 16: 145-170.
Huene F. von (1941). Die Tetrapoden-Fährten im toskanischen Verrucano und ihre Bedeutung. Neues Jarhrbuch für Mineralogie, Geologie und Paläontologie Abteilung, 86: 1-34.
Hunt A.P., Lucas S.G., Heckert A.B., Sullivan R.M. & Lockley M.G. (1998). Late Triassic dinosaurs from the western United States. Geobios, 31: 511-531.
Irmis R.B., Nesbitt S.J., Padian K., Smith N.D., Turner A.H., Woody D. & Downs A. (2007b). A Late Triassic Dinosauromorph Assemblage from New Mexico and the Rise of Dinosaurs. Science, 317: 358-361.
Irmis R.B., Parker W.G., Nesbitt S.J. & Liu J. (2007a). Early ornithischian dinosaurs: the Triassic record. Historical Biology, 19: 3-22.

xviiM. Marzola & F.M. Dalla Vecchia - Triassic dinosaur tracks from the Carnian Prealps
Kammerer C.F., Nesbitt S.J. & Shubin N.H. (2011). The first silesaurid dinosauriform from the Late Triassic of Morocco. Acta Palaeontologica Polonica, 57: 277-284.
King M.J. & Benton M.J. (1996). Dinosaurs in the Early and Mid-Triassic? - The footprints evidence from Britain. Palaeogeography, Palaeoclimatology, Palaeoecology, 122: 213-225.
Klein H. & Haubold H. (2007). Archosaur footprints - potential for biochronology of Triassic continental sequences. In Lucas S.G. & Spielmann J.A. (eds), The Global Triassic. New Mexico Museum of Natural History & Science Bulletin, 41: 120-130.
Klein H. & Lucas S.G. (2010). Tetrapod footprints - their use in biostratigraphy and biochronology of the Triassic. Geological Society, London, Special Publications, 334: 419-446.
Klein H., Lucas S.G. & Haubold H. (2006). Tetrapod track assemblage of the Redonda Formation (Upper Triassic, Chinle Group) in east-central New Mexico - re-evaluation of ichnofaunal diversity from studies of new material. In Harris J.D., Lucas S.G., Spielmann J.A., Lockley M.G., Milner A.R.C. & Kirkland J.I. (eds), The Triassic-Jurassic terrestrial transition. New Mexico Museum of Natural History and Science Bulletin, 37: 241-250.
Klein H., Voigt S., Saber H., Schneider J.W., Hminna A., Fischer J., Lagnaoui A. & Brosig A. (2011). First occurrence of a Middle Triassic tetrapod ichnofauna from the Argana Basin (Western High Atlas, Morocco). Palaeogeography, Palaeoclimatology, Palaeoecology, 307: 218-231.
Krebs B. (1976). Pseudosuchia. In Charig J., Krebs B., Sues H.-D. & Westphal F. (eds), Thecodontia. Handbuch der Paläoherpetologie, 13: 40-98.
Langer M.C. (2003). The sacral and pelvic anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil). Paleobios, 23: 1-40.
Langer M.C. (2004). Basal Saurischia. In Weishampel D.B., Dodson P. & Osmólska H. (eds), The Dinosauria (2nd ed.). University of California Press: 25-46.
Langer M.C., Abdala F., Richter M. & Benton M.J. (1999). A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil. Comptes Rendus de l’Academie des Sciences, Paris: Sciences de la Terre et des planètes, 329: 511-517.
Langer M.C. & Benton M.J. (2006). Early dinosaurs: a phylogenetic study. Journal of Systematic Palaeontology, 4: 309-358.
Langer M.C., Ezcurra M.D., Bittencourt J.S. & Nova F.E. (2010). The origin and early evolution of dinosaurs. Biological Reviews, 85: 55-110.
Langer M.C., Nesbitt S.J., Bittencourt J.S. & Irmis R.B. (2013). Non-dinosaurian Dinosauromorpha. Geological Society, London, Special Publications, 379: 157-186.
Leonardi G. (2000). I dinosauri d’Italia e delle aree adiacenti. In Leonardi G. & Mietto P. (eds), Dinosauri in Italia - Le orme giurassiche dei Lavini di Marco (Trentino) e gli altri resti fossili italiani. Accademia Editoriale, Pisa-Roma: 275-295.
Leonardi G. & Lockley M.G. (1995). A proposal to abandon the ichnogenus Coelurosaurichnus von Huene, 1941, a junior synonym of Grallator E. Hitchcock, 1858. Journal of Vertebrate Paleontology, 15: 40 (A).
Lockley M.G. (1991). Tracking dinosaurs: A new look at an ancient world. 238 pp. Cambridge University Press, Cambridge.
Lockley M.G. & Gierliński G.D. (2006). Diverse vertebrate ichnofaunas containing Anomoepus and other unusual trace fossils from the Lower Jurassic of the western United States: implications for paleoecology palichnostratigraphy. In Harris J.D., Lucas S.G., Spielmann J.A., Lockley M.G., Milner A.R.C. & Kirkland J.I. (eds), The Triassic-Jurassic terrestrial transition. New Mexico Museum of Natural History and Science Bulletin, 37: 176-191.
Long R.A. & Murry P.A. (1995). Late Triassic (Carnian and Norian) Tetrapods from the Southwestern United States. New Mexico Museum of Natural History and Science Bulletin, 4: 1-254.
Lull R.S. (1904). Fossil footprints of the Jura-Trias of North America. Memoirs of the Boston Society of Natural History, 5: 461-557.
Manning P.L. (2004). A new approach to the anlysis and interpretation of tracks: examples from the dinosauria. In McIlroy D. (ed.), The Application of Ichnology to the Palaeoenvironmental and Stratigraphic Analysis. Geological Society, London, Special Publications, 228: 93-123.
Martínez R.N. & Alcober O.A. (2009). A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE, 4: 1-12, e4397.
Martínez R.N., Sereno P.C., Alcober O.A., Colombi C.E., Renne P.R., Montañez I.P. & Currie B.S. (2011). A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea. Science, 331: 206-210.
Mehl M.G. (1915). Poposaurus gracilis, a new reptile from the Triassic of Wyoming. Journal of Geology, 23: 516-522.
Mietto P. (1988). Piste di dinosauri nella Dolomia Principale (Triassico superiore) del M. Pelmetto (Cadore). Memorie della Società Geologica Italiana, 30: 307-310.
Mietto P. (1991). Impronte di dinosauri nel Triassico superiore delle Dolomiti. In Muscio G. (ed.), Dinosaurs. Il mondo dei dinosauri, Kaleidos: 83-87.
Neri C., Gianolla P., Furlanis S., Caputo R. & Bosellini A. (2007). Note Illustrative della Carta Geologica d’Italia alla scala 1:50.000, Foglio 029 Cortina d’Ampezzo. APAT - Dipartimento Difesa del Suolo-Servizio Geologico d’Italia, Roma.
Nesbitt S.J. (2007). The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History, 302: 1-84.
Nesbitt S.J. (2011). The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History, 352: 1-292.
Nesbitt S.J., Irmis R.B. & Parker W.G. (2007). A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology, 5: 209-243.
Nesbitt S.J., Irmis R.B., Parker W.G., Smith N.D., Turner A.H. & Rowe T. (2009). Hind limb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. Journal of Vertebrate Paleontology, 29: 498-516.
Nesbitt S.J. & Norell M.A. (2006). Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda). Proceedings of the Royal Society of London, Series B, Biological Sciences, 273: 1045-1048.
Nesbitt S.J., Sidor C.A., Irmis R.B., Angielczyk K.D., Smith R.M.H. & Tsuji L.A. (2010). Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira. Nature, 464: 95-98.
Nicosia U. & Loi M. (2003). Triassic footprints from Lerici (La Spezia, northern Italy). Ichnos, 10: 127-140.
Niedzwiedzki G. (2011). A Late Triassic dinosaur-dominated ichnofauna from the Tomanová Formation of the Tatra Mountains, Central Europe. Acta Palaeontologica Polonica, 56: 291-300.
Novas F.E., Ezcurra M.D., Chatterjee S. & Kutty T.S. (2011). New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 101: 333-349.
Olsen P.E. & Baird D. (1986). The ichnogenus Atreipus and its significance for Triassic biostratigraphy. In Padian K. (ed.), The Beginning of the Age of Dinosaurs, Cambridge University Press: 61-87.
Olsen P.E. & Rainforth E.C. (2003). The Early Jurassic ornithischian dinosaurian ichnogenus Anomoepus. In Le Tourneau P.M. & Olsen P.E. (eds), The Great Rift Valleys of Pangea in Eastern North America. Sedimentology, Stratigraphy, and Paleontology, 2, Columbia University Press: 314-367.
Olsen P.E., Smith J.B. & McDonald N.G. (1998). Type material of the type species of the classic theropod footprint genera Eubrontes, Anchisauripus, and Grallator (Early Jurassic,

Bollettino della Società Paleontologica Italiana, 2014xviii
Hartford and Deerfield Basins, Connecticut and Massachusetts, USA). Journal of Vertebrate Paleontology, 18: 586-601.
Padian K. (1997). Bipedality. In Currie P.J. & Padian K. (eds), Encyclopedia of Dinosaurs. Academic Press: 175-179.
Paul G. (2010). The Princeton Guide to Dinosaurs. 320 pp. Princeton University Press, Princeton & Oxford.
Piechowski R. & Dzik J. (2010). The axial skeleton of Silesaurus opolensis. Journal of Vertebrate Paleontology, 30: 1127-1141.
Pittman J.G. (1989). Stratigraphy, lithology, depositional environment, and track type of dinosaur track-bearing beds of the Gulf Coastal Plain. In Gillette D.D. & Lockley M.G. (eds), Dinosaur tracks and traces. Cambridge University Press: 135-153.
Reig O.A. (1963). La presencia de dinosaurios saurisquios en los” Estratos de Ischigualasto”(Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina). Ameghiniana, 3: 3-20.
Romer A.S. (1972a). The Chañares (Argentina) Triassic reptile fauna. XIV. Lewisuchus admixtus, gen. et sp. nov., a further thecodont from the Chañares beds. Breviora, 390: 1-13.
Romer A.S. (1972b). The Chañares (Argentina) Triassic reptile fauna. XV. Further remains of the thecodonts Lagerpeton and Lagosuchus. Breviora, 394: 1-7.
Rowe T. & Gauthier J. (1990). Ceratosauria. In Weishampel D.B., Dodson P. & Osmólska H. (eds), The Dinosauria, University of California Press: 151-168.
Safran J. & Rainforth E.C. (2004). Distinguishing the tridactyl dinosaurian ichnogenera Atreipus and Grallator: where are the latest Triassic ornithischia in the Newark Supergroup? Geological Society of America. Abstracts with Programs, 36: 96.
Sarjeant W.A.S. (1996). A re-appraisal of some supposed dinosaur footprints from the Triassic of the English Midlands. Mercian Geologist, 14: 22-30.
Schachner E.R., Manning P.L. & Dodson P. (2011). Pelvic and hind limb myology of the basal archosaur Poposaurus gracilis (Archosauria: Poposauroidea). Journal of Morphology, 272: 1464-1491.
Sereno P.C. (2007). The phylogenetic relationships of early dinosaurs: a comparative report. Historical Biology, 19: 145-155.
Sereno P.C., Forster C.A., Rogers R.R. & Monetta A.M. (1993). Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. Nature, 361: 64-66.
Sereno P.C. & Novas F. E. (1994). The skull and neck of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology, 13: 451-476.
Sues H-D., Nesbitt S.J., Berman D.S. & Henrici A.C. (2011). A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proceedings of the Royal Society B, 278: 3459-3464.
Sullivan R.M & Lucas S.G. (1999). Eucoelophysis baldwini, a new theropod dinosaur from the Upper Triassic of New Mexico, and the status of the original types of Coelophysis. Journal of Vertebrate Paleontology, 19: 81-90.
Thulborn R.A. (1993). A tale of three fingers: ichnological evidence revealing the homologies of manual digits in theropod dinosaurs. In Lucas S.G. & Morales M. (eds), The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin, 3: 461-463.
Thulborn T. (1990). Dinosaur tracks. 410 pp. Chapman & Hall, London, New York, Melbourne, Tokyo, Madras.
Upchurch P., Barrett P.M. & Galton P.M. (2007). A phylogenetic analysis of basal sauropodomorph relationships: Implications for the origin of sauropod dinosaurs, Special Papers in Palaeontology, 77: 57-90.
Walker A.D. (1964). Triassic reptiles from the Elgin area Ornithosuchus and the origin of Carnosaurs. Philosophical Transactions of the Royal Society of London, 744: 53-134.
Weems R.E. (1992). A re-evaluation of the taxonomy of Newark Supergroup saurischian dinosaur tracks, using extensive statistical data from a recently exposed tracksite near Culpeper, Virginia. In Proceedings of the 26th Forum on the Geology of Industrial Minerals - 14-18 May 1990, Virginia Division of Mineral Resources Publication, 119: 113-127.
Weishampel D.B. (2006). Another look at the dinosaurs of the East Coast of North America. Actas III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Salas de los Infantes, Burgos, Spain. Colectivo Arqueológico-Paleontológico Salense: 129-168.
Weishampel D.B., Barrett P.M., Coria R.A., Le Loeuff J., Xing X., Xijin Z., Sahni A., Gomani E.M.P. & Noto C.R. (2004). Dinosaur distribution. In Weishampel D.B., Dodson P. & Osmólska H. (eds), The Dinosauria (2nd ed.). University of California Press: 517-606.
Manuscript received 12 March 2013Revised manuscript accepted 10 December 2013Published online 19 February 2014Editor Fabio Massimo Petti




















