
Myofilament response to Ca2+ and Na+/H+ exchanger activity in sex hormone-related protection of...
30
1 Myofilament Response to Ca 2+ and Na + /H + Exchanger Activity in Sex Hormone Related Protection of Cardiac Myocytes from Deactivation in Hypercapnic Acidosis Tepmanas Bupha-Intr*, Jonggonnee Wattanapermpool*, James R. Peña † , Beata M Wolska † , and R John Solaro Dept. of Physiology & Biophysics, Medicine † , and Center for Cardiovascular Research, College of Medicine, University of Illinois at Chicago, IL, USA and *Department of Physiology, Faculty of Science, Mahidol University, Thailand Running title: Sex hormones, acidosis, and cardiac myocyte function Address for correspondence: R. John Solaro, PhD Department of Physiology and Biophysics College of Medicine University of Illinois at Chicago 835 S. Wolcott Ave. Chicago, IL 60612-7342 Tel: 312-996-7620 Fax: 312-996-1414 email: [email protected] Page 1 of 30 Articles in PresS. Am J Physiol Regul Integr Comp Physiol (October 12, 2006). doi:10.1152/ajpregu.00376.2006 Copyright © 2006 by the American Physiological Society.
Transcript of Myofilament response to Ca2+ and Na+/H+ exchanger activity in sex hormone-related protection of...

1
Myofilament Response to Ca2+ and Na+/H+ Exchanger Activity in Sex Hormone Related
Protection of Cardiac Myocytes from Deactivation in Hypercapnic Acidosis
Tepmanas Bupha-Intr*, Jonggonnee Wattanapermpool*, James R. Peña†, Beata M Wolska†,
and R John Solaro
Dept. of Physiology & Biophysics, Medicine†, and Center for Cardiovascular Research,
College of Medicine, University of Illinois at Chicago, IL, USA
and
*Department of Physiology, Faculty of Science, Mahidol University, Thailand
Running title: Sex hormones, acidosis, and cardiac myocyte function
Address for correspondence: R. John Solaro, PhD Department of Physiology and Biophysics College of Medicine University of Illinois at Chicago 835 S. Wolcott Ave. Chicago, IL 60612-7342 Tel: 312-996-7620 Fax: 312-996-1414 email: [email protected]
Page 1 of 30Articles in PresS. Am J Physiol Regul Integr Comp Physiol (October 12, 2006). doi:10.1152/ajpregu.00376.2006
Copyright © 2006 by the American Physiological Society.

2
Abstract Compared to sham operated controls, myofilaments from hearts of ovariectomized ( OVX) rats
demonstrate an increase in Ca2+ sensitivity with no change in maximum tension (Wattanapermpool and
Reiser, Am J Physiol 1999;277:H467-73). To test the significance of this modification in intact cells, we
compared intracellular Ca2+ transients and shortening of ventricular myocytes isolated from sham and 10
week OVX rats. There was a decrease in the peak Ca2+ transient with prolonged 50% decay time in
OVX cardiac myocytes without changes in the resting intra-cellular Ca2+ concentration. Per cent cell
shortening was also depressed and relaxation was prolonged in cardiac myocytes from OVX rats
compared to shams. Ovariectomy induced a sensitization of the myofilaments to Ca2+. Hypercapnic
acidosis suppressed the shortening of OVX myocytes to a lesser extent than that detected in shams.
Moreover, a larger compensatory increase in % cell shortening was obtained in OVX myocytes during
prolonged acidosis. The elevated compensation in cell shortening was related to a higher amount of
increase in the amplitude of the Ca2+ transient in OVX myocytes. However, these differences in Ca2+
transients and % cell shortening were no longer evident in the presence of 1µM cariporide, a specific
inhibitor of Na+/H+ exchanger type 1 (NHE1). Our results indicate that deprivation of female sex
hormones modulates the intracellular Ca2+ concentration in cardiac myocytes possibly via an increased
NHE1 activity, which may act in concert with Ca2+ hypersensitivity of myofilament activation as a
determinant of sex differences in cardiac function.
Keywords: heart, estrogen, acidosis, sarcomeric proteins, sodium, proton exchanger
Page 2 of 30

3
Introduction
Sex steroid hormones exert significant control over cardiac physiology, but the molecular and
cellular mechanisms for these effects remain unclear. Understanding these mechanisms is crucial for
translation to hormone therapy (30). Scheuer and co-workers (24,25) first demonstrated a suppression
of function in isolated heart preparation in both gonadectomized male and female rats. Moreover,
these investigators reported that the decreases in ejection fraction, peak systolic pressure, and cardiac
output in the hearts from gonadectomized rats could be normalized with hormone supplementation.
These results, which pointed to an important role of both male and female steroid hormones in
regulating cardiac function, fit with the hypothesis of a cardioprotective role of female sex hormones.
Epidemiological studies also supported this hypothesis by demonstrating that men have a higher
incidence of heart disease than pre-menopausal women (14). Echo-cardiographic assessment of
cardiac function in pre-menopausal women also demonstrated enhanced contractile function compared
to post-menopausal women (20). Evidence of improved prognosis in female heart failure patients
taking estrogen compared to those off estrogen also supports the hypothesis of the importance of sex
hormones in heart diseases (16,21).
Although, anti-apoptotic effects of estrogen have been reported (19), depression in Ca2+ fluxes
to and from the myofilaments appears to account in part for the depression of contractility in hearts of
ovariectomized (OVX) rats. Ren et al. (22) reported that myocytes from 6-week OVX rat hearts
demonstrated slowing of relaxation and contraction kinetics associated with a slower decay and
depressed peak amplitude of Ca2+ transients. Bupha-Intr and Wattanapermpool (5) reported a
depression in Ca2+ uptake rate and ATPase activity by vesicles of the cardiac sarcoplasmic reticulum
(SR) derived from hearts of rats 10 weeks following ovariectomy, compared to sham controls.
Depression in expression of SERCA at the protein and mRNA levels appeared to account for this
depression in myocyte function and Ca2+ fluxes (5). There is also evidence of sex related differences in
fluxes through the Na+,/Ca2+ exchanger (NCX). Sugishita et al. (29) reported that metabolic inhibition
induced a relatively large increase in intracellular Ca2+ in myocytes of transgenic male versus female
Page 3 of 30

4
mouse hearts over expressing (NCX). Treatment of the males with 17β-estradiol (E2) significantly
attenuated this difference. Whether these effects on intracellular Ca2+ are due directly to NCE activity
or indirectly through modification of the activity of the Na+,H+ exchanger (NHE1) remains unclear.
There is, however, evidence of acute effects of E2 on regulation of NHE1 activity. In demonstrating
that acute treatment with E2 improved recovery of hearts stressed by normoxic acidification, Anderson
et al. (2) proposed a mechanism involving in part promotion of NO synthesis that increased NHE1
activity.
Alterations in myofilament response to Ca2+ also appear to play an important role in effects of
estrogen on cardiac function. There is a reduction in ATPase rate in myofilaments isolated from hearts
of OVX rats, which is attributable to a shift in myosin isoform population from one predominant in the
alpha-myosin heavy chain to one containing significant amounts of beta-myosin heavy chain (33).
Moreover, compared to preparations from shams, ATPase activity (33) and tension (34) of
myofilaments from OVX rat hearts were more sensitive to Ca2+. These changes in myofilament
response to Ca2+ were reversed in preparations from OVX rats supplemented with E2 (36) or in animals
subjected to exercise training (4).
In experiments reported here, we have extended our studies of the relation between sex
hormones and myofilament response to Ca2+ with the objective of understanding the relevance of this
effect in isolated cardiac myocytes. We think this understanding is important in view of evidence that
an increase in myofilament response to Ca2+ is a common functional consequence of mutations in
sarcomeric proteins that are linked to familial hypertrophic myopathies (10). Moreover, the role of
myofilament response to Ca2+ has not been generally considered in the cardio-protective effects of E2
in acidosis (2) and metabolic inhibition (29) even though there is evidence that altered myofilament
response to Ca2+ is a dominant mechanism responsible for the fall in tension associated with acidosis
(27). We therefore compared the effects of hypercapnic acidosis on shortening and intra-cellular Ca2+
of myocytes isolated from sham, OVX, and OVX + E2 rat hearts. We also compared tension-pCa
relations in detergent extracted fiber bundles from these hearts. To probe for changes in myofilament
Page 4 of 30

5
response to Ca2+, we compared responses at physiological pH to responses at acidic pH Our results
confirm that cardiac myofilaments functioning in intact cardiac myocytes from OVX rat hearts are more
sensitive to Ca2+ than in control myocytes. Our data also indicate that chronic deprivation of female
sex hormones modulates the intracellular Ca2+ concentration in cardiac myocytes via an increase in
NHE1 activity, which may act in concert with the hypersensitivity of myofilament activation as a
significant determinant of sex differences in cardiac function.
Materials and Methods
Animals
Our studies employed female Sprague-Dawley rats that were OVX or sham operated at 8
weeks of age by the supplier (Harlan, Indianapolis, IN) and killed at 18 weeks of age. We randomly
divided the OVX rats into two groups, and injected them subcutaneously with 0.1 ml of corn oil with or
without 5 µg of 17β-estradiol. We did these injections three times per week for 10 weeks, as previously
described (5). We verified deficiency of ovarian sex hormones by the absence of ovarian tissues and
by the shrinkage of uterine mass (6).
Cardiac Myocyte Isolation.
At the 10 week time point, we heparinized (1,000 unit/kg body wt) the rats; and, after 30 min
anesthetized them with an intra-peritoneal injection of pentobarbital sodium (25 mg/kg body weight).
Left ventricular myocytes were isolated as previously described (31, 39, 40). The heart was rapidly
removed, the aorta was cannulated and perfused with oxygenated nominally Ca2+-free Tyrode solution
until complete cessation of beating. The perfusion solution was then switched to 25 µM CaCl2 Tyrode
solution containing 1 mg/ml BSA, 160 unit/ml collagenase type II (Boehringer Mannheim), and 0.1
mg/ml protease type XIV (Sigma, St. Louis, MO). The hearts were then perfused for 16.5 min/g wet wt
with the above solution. At the end of the perfusion period, the hearts were removed from cannula and
placed into a Petri dish containing 50 µM CaCl2 Tyrode-BSA buffer. Ventricles were cut into small
pieces and gently triturated with a pipette and filtered through a cell collector. The cell suspension was
Page 5 of 30

6
placed into centrifuge tubes and gradually increased the extra-cellular CaCl2 concentration to a final
level of 0.5 mM CaCl2. Myocytes were studied within six hours after isolation.
Intracellular Ca2+ transients, pH, and Cell Shortening.
Myocytes were loaded with 5 µM Fura 2-AM for 15 min in 0.5 mM CaCl2 Tyrode buffer with 1
mg/ml BSA and 5% fetal bovine serum for 15 minutes. After washing off the excess Fura 2-AM, the
myocytes were electrically stimulated at a frequency of 0.5 Hz for 10 to 15 min or until the shortening
was stable with perfusion of 2 mM CaCl2 Tyrode buffer at room temperature. Fura 2 fluorescence
(340:380 ratio) and cell shortening were monitored simultaneously as described in detail by Wolska et
al. (38).
To test the effect of acidic pH on intracellular Ca2+ transients and cell shortening, we perfused
the myocytes with control Krebs-Henseleit buffer (KH) saturated with 95% O2 and 5% CO2 for 15 min,
with KH saturated with 85% O2 and 15% CO2 for 20 min, and then with control KH. In these
experiments, we also determined intracellular pH with the aid of BCECF as previously described (42),
and made the measurements in the presence and absence of 1.0 µM cariporide, an inhibitor of the
NHE1 (12).
Force measurements on skinned fiber bundles
We determined the relationship between pCa ( log of the molar free Ca2+) and tension
(force/cross-sectional area) in detergent (Triton X-100) extracted fiber bundles prepared from left
ventricular papillary muscles as previously described (39,41). The fiber bundles (~150-200 µm in
diameter and 4-5 mm long) were cut in ice cold high relaxing (HR) buffer: 10 mM EGTA, 2 mM free
Mg2+, 5 mM MgATP2-, 79.2 mM KCl, 12 mM creatine phosphate, 20 mM MOPS, pH 7.0, ionic strength
0.15 M, 2.5 µg/ml pepstatin A, 1 µg/ml leupeptin and 50 µM PMSF. After mounting the fiber bundles in
an apparatus for determination of force, sarcomere length was set at 2.2 µm as determined from a laser
diffraction pattern in HR solution and cross-sectional area determined. Isometric force was recorded on
a chart recorder at different pCa at room temperature. CaCl2 concentrations in HR were varied to
achieve a range of Ca2+ concentrations as computed using a computer program and binding constants
Page 6 of 30

7
summarized by Godt and Lindley (9). Only fiber bundles that developed 85-90% of their initial maximum
tension at the end of experiment were retained for analysis. We plotted isometric tension as a
function of pCa, fitted the curves to the A. V. Hill equation as previously described (41), and expressed
the relations as a per cent of maximum tension. pCa50 (half-maximally activating pCa values) was
computed from individual Hill fits of each pCa-tension relation and then averaged.
Expression of NHE1
To determine the level of expression of the NHE-1 gene, real-time one-step quantitative RT-
PCR with SYBR Green detection (Qiagen,Valencia, CA) was performed using a LightCycler
thermocycler (Roche Diagnostics) as previously described (1). Briefly, total RNA was isolated from
ventricular tissue with the RNeasy Fibrous Tissue Mini Kit (Qiagen, Valencia, CA). The RNA was
reverse transcribed and the cDNA used to derive a PCR product for NHE-1 and ribosomal protein L7.
The PCR product was cloned into the vector, pBlue-Topo (Invitrogen, Carlsbad, CA). The plasmid
underwent amplification, DNA purification, and in vitro transcription. The resulting RNA was recovered,
quantified and used as standards in the RT-PCR reaction. Gene expression was normalized to
ribosomal protein L7. The following primers were used: NHE-1(forward) 5’-
TTTCCCCgATTTCCTTCTCT-3’; NHE-1(reverse) 5’-gCgTgTAAgACCTgggACAT-3’; Ribosomal protein
L7 (forward) 5’-gAAgCTCATCTATgAgAAggC-3’; Ribosomal protein L7 (reverse) 5’-
CAgACggAgCAgCTgCAgCAC-3’. We also determined the levels of NHE1 by Western blot analysis of
left ventricular proteins separated by 8% SDS-PAGE with anti-NHE1 antibody (Chemicon, 1:1000) using
an HRP labeled secondary antibody and visualization using ECL (Amersham Pharmacia).
Statistical analysis.
Data are presented as means ± SE. All curve fittings were achieved using GraphPad Prism.
One-way ANOVA was used for statistically analysis and Student-Newman-Kuels test for multiple
comparisons . P < 0.05 was considered the significant difference among groups.
Page 7 of 30

8
Results
Data in Table 1 summarize heart, body, and uterine weights of sham, OVX, and OVX +E2
animals. Compared to the shams, OVX rats demonstrated a significant increase in both heart and
body weight, but no change in heart weight/ body weight ratio. As expected uterine weight was
significantly lower in the OVX rats compared to shams, and increased significantly upon E2
supplementation. However, uterine weight did return fully to control levels with E2 supplementation.
We think this is due likely to a requirement for other hormones in the re-growth of the uterus.
Cell Shortening and Intracellular Calcium Transients in myocytes.
Table 2 summarizes data comparing mechanics of ventricular myocytes from sham, OVX, and
OVX + E2 rat hearts. OVX induced a change in all parameters. The per cent cell shortening was
significantly depressed by ~22%, as was +dL/dt, the maximum velocity of shortening by ~30% and –
dL/dt, the maximum velocity of re-lengthening by ~50%. The prolongation of the time to peak
amplitude of shortening and the time to 80% full relaxation (T80) also reflected a significant alteration in
myocytes shortening and re-lengthening kinetics. These changes in mechanical properties were only
partially restored by E2 supplementation. Per cent cell shortening and T80 were both significantly
different from OVX myocytes, but other parameters were not. As indicated by the data in Table 3,
which summarizes our measurements of intracellular Ca2+ transients, the depression in per cent cell
shortening induced by OVX was associated with a significant fall in the peak amplitude of the Ca2+
transients with no change in diastolic levels of Ca2+ or the time to peak amplitude. The time to 50%
decay of the Ca2+ transient (T50) was significantly prolonged by OVX and restored to control levels by
E2 supplementation. Moreover, in contrast to the findings from measurements of per cent cell
shortening, the reduction in amplitude of Ca2+ transients and prolonged time of Ca2+ decay in OVX
myocytes could be completely restored with E2 supplementation.
Previous studies demonstrated that compared to controls, ATPase rate (33) and force (34) of
myofilaments isolated from hearts of OVX rats are more sensitive to Ca2+. To determine whether the
increase in myofilament sensitivity occurred in the myocytes from OVX hearts, we performed a set of
Page 8 of 30

9
experiments in which we varied extra-cellular Ca2+ to generate a range of increasing levels of the Ca2+
transient. We determined the relation between the peak amplitude of these Ca2+ transients (expressed
as 340:380 ratio), and the extent of cell shortening. Figure 1 summarizes the data for the level of Ca2+
inducing half-maximal activation of shortening in myocytes from shams, OVX and OVX + E2 hearts.
The value of the peak amplitude of Ca2+ transient that gave rise to 50% of the maximum cell shortening
was significantly diminished in OVX myocytes (Fura-2 ratio = 2.52 ± 0.05 in sham controls, and Fura-2
ratio = 2.11 ± 0.04 in OVX myocytes), and was partially restored in the myocytes from OVX + E2 (Fura-
2 ratio = 2.30 ± 0.04). These results indicated that the previous in vitro data indicating an enhanced
myofilament response to Ca2+ associated with OVX also occurs in the intact cardiac myocytes.
Acidosis, Myofilament Tension, and Myocyte Function.
In view of the cardio-protective effects of E2 in acidosis (2) and metabolic inhibition (29); and,
inasmuch as altered myofilament response to Ca2+ is a dominant mechanism responsible for the fall in
tension associated with acidosis (27), we compared the effects of pH on pCa-tension relations of
skinned fiber bundles isolated from sham, OVX, and OVX + E2 rat hearts. We also determined the
relative effects of hypercapnic acidosis on shortening and intra-cellular Ca2+ of myocytes isolated from
sham, OVX, and OVX + E2 rat hearts.
The upper panel of Figure 2 shows relations between pCa and relative tension at pH 7.0, pH
6.8, and pH 6.5 for skinned fiber bundles from the three groups of rat hearts. These results
demonstrate the well-known effect of acidic pH to desensitize cardiac myofilaments to Ca2+. Data in the
lower panel of Figure 5 summarize the half-maximal activating pCa values (pCa50). In agreement with
previous results (34), the skinned fibers from OVX rat hearts demonstrated a significant increase in
sensitivity to Ca2+ at pH 7.0 (pCa50 = 5.84 ± 0.02) compared to fibers from shams (pCa50 = 5.77 ± 0.02).
Skinned fibers from hearts of OVX rats supplemented with E2 had a sensitivity to Ca2+ essentially the
same as controls (pCa50 = 5.75 ± 0.03). However, at pH 6.8 and 6.5 there were no significant
Page 9 of 30

10
differences in the pCa50 among the fiber bundles from the three groups. In all cases there were no
significant differences in the Hill coefficient among the fiber groups.
We also determined the relative effects of hypercapnic acidosis on shortening and intra-cellular
Ca2+ of myocytes isolated from sham, OVX, and OVX + E2 rat hearts. Intra-cellular pH fell from a value
of pH 7.25 ± 0.08 in control conditions to pH 6.65 ± 0.07 during perfusion with KH buffer saturated with
15% CO2, 85% O2. The top 3 records in Figure 3A show representative traces of myocytes shortening
before, during, and after these changes in pH. In myocytes from sham rat hearts, about 3 min after
introduction of the hypercapnic KH buffer, the extent of cell shortening fell significantly before rising to a
steady-state value after about 5 min. This steady-state level of extent of shortening remained
significantly lower (56.4 ± 3.9 % reduction) than the value before acidosis and abruptly returned to this
control level with an over-shoot after re-introduction of the control KH buffer. However, although
myocytes from OVX hearts demonstrated the initial fall in extent of shortening similar to that of the
sham control myocytes, extent of shortening rose to a steady-state level 99.8 ± 5.4 % of the value in
the control KH buffer. This resistance to hypercapnic acidosis was partially reversed (72.0 ± 2.8 %
reduction in cell shortening), in myocytes from hearts of the OVX + E2 rats but remained significantly
higher (P < 0.01) than during the control period. The lower 3 traces in Figure 3A show typical records
of myocyte shortening obtained in the presence of cariporide, an inhibitor the NHE1 exchanger.
Regardless of the source, the myocytes demonstrated a lack of biphasic response and essentially the
same extent of severe reduction in steady-state shortening with hypercapnic acidosis. Figure 3B
summarizes the change in extent of shortening at 5 min, 10 min, and with recovery from acidosis.
These data show that the significant protection of OVX myocytes from acidosis was not present in the
myocytes treated with cariporide. To determine whether cariporide might affect myofilament response
to Ca2+, we compared pCa-tension relations in the presence and absence of 1 µM cariporide. These
experiments (data not shown) demonstrated that cariporide does not exert any direct effects on
myofilament response to Ca2+.
Page 10 of 30

11
Data in Figure 4 summarize the per cent change in peak amplitude of intra-cellular Ca2+ transients in
the cardiac myocytes from the three groups in the absence and presence of cariporide before, at 5 min
and at 10 min of acidosis, and during recovery after hypercapnic acidosis. In the absence of
cariporide, the Ca2+ transient increased significantly at 10 min in myocytes from the OVX and OVX +E2
rat hearts. However, there were no changes in the peak amplitudes of the Ca2+ transients at any time
in the presence of cariporide. The data in Figures 3 and 4 indicate that the contribution of the activity
of the NHE1 and NCX in regulation of intra-cellular Ca2+ was relatively high in myocytes from OVX rat
hearts compared to shams. We therefore determined the level of expression of NHE1 message and
protein in the hearts from the three groups of rats. Figure 5A shows results of experiments, in which we
quantified the level of expression of the NHE-1 gene employing real-time quantitative RT-PCR with
SYBR Green detection. The data showed no significant differences. This was also true at the protein
level as illustrated in Figure 5B, which shows a representative Western blot indicating that there was no
significant difference in expression of NHE1 among the sham, OVX, and OVX + E2 hearts. This
determination was made three times with no difference in protein level detected.
Discussion
Data reported here are the first to report an effect of ovariectomy to enhance Ca2+-sensitivity of
myofilaments functioning in the intact cardiac myocyte. An important new finding with regard to this
altered response to Ca2+ is that although it appears to occur at pH 7.0, the sensitization did not occur at
pH 6.8 or pH 6.5. This result adds new insights with regard to the interpretation of the significance of
the altered myofilament response to Ca2+ in the physiology and pathology of the myocardium. Our data
are also the first to demonstrate a significant role of enhanced NHE1 activity in altering the response of
heart cells from OVX rats to the stress of acidosis.
There is now general agreement that altered myofilament response to Ca2+ is a significant
aspect of the integrated biology of cardiac function (28). A mechanism involving sarcomere length
dependent alterations in myofilament response to Ca2+ remains the dominant theory for the cellular
Page 11 of 30

12
basis of Starling’s Law of the heart (23). Moreover, there is substantial evidence for the functional
significance of myofilament protein phosphorylation as a significant factor controlling cardiac dynamics
(29). As illustrated by the data presented here, depressed cardiac function in acidosis also reflects an
influence of intra-cellular environment to depress myofilament response to Ca2+, a mechanism which is
dominant in determining cellular function. For example, Figures 4 and 5 summarized data showing a
severe depression in cell shortening with acidosis in the presence or absence of cariporide with little or
no change in the peak amplitude of the Ca2+ transient. Chronic increases in cardiac myofilament
response to Ca2+, which our previous (33,34) and present data indicate occur with ovariectomy, are
now also recognized to be of major significance as a cause of hypertrophy associated with mutations in
sarcomeric proteins (1012). The increased Ca2+ sensitivity of OVX cardiac myofilaments is also similar
to that reported in cardio-myopathic and heart failure models (11,37). The elevated myofilament
response to Ca2+ may induce diastolic defects leading to cell stretch and elaboration of signals
promoting hypertrophic growth (8).
The effects of chronic increases in myofilament response to Ca2+ are, however, complex
inasmuch as there is evidence of the beneficial effects of therapeutic agents, which induce myofilament
Ca2+ -sensitization (13). Moreover, hearts of transgenic mouse models with constitutive myofilament
Ca2+-sensitization associated with modification of troponin I demonstrate remarkable resistance to
acidosis (32,35), sepsis (15), and ischemia/reperfusion (3,7). An important distinction is that compared
to controls the myofilaments containing modified troponin I or treated with Ca2+-sensitizing agents are
not only more sensitive to Ca2+ they are resistant to deactivation by acidic pH (30,45), whereas the
myofilaments from OVX hearts are not.
To more clearly understand whether the increase in myofilament response to Ca2+ associated
with ovariectomy is an adaptive or a maladaptive response, we stressed myocytes with acidosis. We
chose this approach because of the dominance of depressed myofilament Ca2+ responsiveness in
acidosis, and because of its relevance to ischemia and ischemia/reperfusion injury (3). We found that
the acidosis induced depression of shortening of myocytes isolated from hearts of OVX rats was
Page 12 of 30

13
blunted compared to the controls. Although this blunting may have been due in part to the enhanced
sensitivity of the myofilaments to Ca2+, data shown in Figure 2 indicate that the sensitization is apparent
only at pH 7.0.
It is likely that this increase in Ca2+ during acidosis in myocytes from OVX hearts and not in
controls occurred as a result of increased Na exchange associated with stimulation of the NHE1 and a
resultant decrease in the efflux of Ca2+ through NCX. This could be due either to a reduced Ca2+ -efflux
or increased influx via reverse mode activity of the NCX. It is also possible that the elevation of
intracellular Ca2+ is due in part to a concerted depression in Ca2+ uptake by the SR, which was
demonstrated in earlier studies to occur in OVX rat hearts (5). Yet, inhibition of the differential effects of
acidosis on myocytes from OVX and sham hearts by cariporide demonstrated the critical role of NHE1
in this response. We did not find a difference in expression of NHE1 among the three groups studied.
We therefore conclude that compared to controls, the loss of ovarian sex hormones may induce a
sensitization of NHE1 activity to pH. This conclusion fits with studies investigating acute effects of E2,
which reported administration of E2 to hearts during ischemia and reperfusion limited the rise in Ca2+
occurring with ischemia and reperfusion (2). By use of inhibitors, these studies provided evidence that
acute administration of E2 increased NOS activity and NO release, and thereby depressed the activity
of the NHE1 resulting in a decreased intra-cellular Na and a limited rise in Ca2+ during
ischemia/reperfusion. A role for NOS has been documented as a basis for the difference in control of
NCX activity between hearts of male and female rats (6,18). Our data provide evidence that the
protective effect of E2 in ischemia and reperfusion occurs during chronic depressions in levels of
ovarian sex hormones. In this case the withdrawal of E2 would release NHE1 from inhibition by NO
resulting in an enhanced activity, which in concert with altered NCX activity led to elevated intracellular
Ca2+ Morimoto et al. (17) reported an increase in mesenteric eNOS expression associated with E2
treatment of OVX rats. An effect of chronic deprivation of ovarian sex hormones on NOS activity is an
attractive mechanism, but whether this occurs with ovariectomy awaits further investigation.
Page 13 of 30

14
Grant Support
This work was supported by NIH Grants RO1 HL 64035 (RJS), PO1 HL 62426 (RJS), and
RO1 HL 64209 and 79032 (BMW). Dr. Bupha-Intr also received support as a Golden Jubilee pre-
doctoral fellow of Mahidol University, Bangkok, Thailand.
Page 14 of 30

15
References
1. Alden KJ, Goldspink PH, Ruch SW, Buttrick PM, Garcia J. Enhancement of L-type Ca2+ current
from neonatal mouse ventricular myocytes by constitutively active PKC-BII. Am J Physiol Cell
Physiol 2002;282:C768-C774.
2. Arteaga GM, Warren CM, Milutinovic S, Martin AF, and Solaro RJ. Specific Enhancement of
Sarcomeric Response to Ca2+ Protects Murine Myocardium Against Ischemia/Reperfusion
Dysfunction. Am J Physiol Heart Circ Physiol 289:H2183-92, 2005.
3. Bupha-Intr T, Wattanapermpool J. Cardioprotective effects of exercise training on myofilament
calcium activation in ovariectomized rats. J Appl Physiol 96:1755-1760, 2004.
4. Bupha-Intr T, Wattanapermpool J. Regulatory role of ovarioan sex hormones in the calcium uptake
activity of cardiac sarcoplasmic reticulum. Am J Physiol Heart Circ Physiol Mar 3; [Epub ahead of
print] ,2006.
5. Cross HR, Murphy E, and Steenbergen C. Ca(2+) loading and adrenergic stimulation reveal
male/female differences in susceptibility to ischemia-reperfusion injury. Am J Physiol Heart Circ
Physiol 283:H481-H489, 2002.
6. Day SM, Westfall MV, Fomicheva EV, Hoyer K, Yasuda S, La Cross NC, D'Alecy LG, Ingwall
JS, and Metzger JM. Histidine button engineered into cardiac troponin I protects the ischemic and
failing heart. Nat Med 12:181-189, 2006.
7. Frey N and Olson EN. Cardiac hypertrophy: the good, the bad, and the ugly. Annu Rev Physiol
2003;65:45-79, 2003.
8. Godt RE, Lindley B. Influence of temperature upon contractile activation and isometric force
production in mechanically skinned muscle fibers of the frog. J Gen Physiol 80:279-97, 1982.
9. Gomes AV, Potter JD. Molecular and cellular aspects of troponin cardiomyopathies. Ann N Y Acad
Sci 1015:214-24, 2004.
10. Heyder S, Malhotra A, and Ruegg JC. Myofibrillar Ca2+ sensitivity of cardiomyopathic hamster
Page 15 of 30

16
hearts. Pflugers Arch 429:539-545, 1995.
11. Karmazyn M. The role of the myocardial sodium-hydrogen exchanger in mediating ischemic and
reperfusion injury. From amiloride to cariporide. Ann N Y Acad Sci. 874:326-34, 1999.
12. Kass D and Solaro RJ. Mechanisms and Use of Calcium Sensitizing Agents. Circulation 113:305-
315, 2006.
13. Kuhn FE, Rackley CE. Coronary artery disease in women. Risk factors, evaluation, treatment, and
prevention. Arch Intern Med 13;153:2626-36, 1993.
14. Layland J, Cave AC, Grieve DJ, Sparks E, Kentish JC, Warren C, Solaro RJ, and Shah AM.
Protection against endotoxaemia induced contractile dysfunction in mice with cardiac-specific
expression of slow skeletal troponin I. FASEB J 19:1137-1139, 2005.
15. Lindenfeld J, Ghali JK, Krause-Steinrauf HJ, Khan S, Adams K, Goldman S, Peberdy MA,
Yancy C, Thaneemit-Chen S, Larsen RL, Young J, Lowes B, and Rosenberg YD; BEST
Investigators. Hormone replacement therapy is associated with improved survival in women with
advanced heart failure. J Am Coll Cardiol 42(7):1238-45, 2003.
16. Morimoto K, Kurahashi Y, Shintani-Ishida K, Kawamura N, Miyashita M, Uji M. Tan N. and
Yoshida K. Estrogen replacement suppresses stress-induced cardiovascular responses in
ovariectomized rats. Am J Physiol Heart Circ Physiol 287:H1950-1956, 2004.
17. Murphy E, Cross HR, and Steenbergen C. Is Na/Ca2+ exchange during ischemia and reperfusion
beneficial or detrimental? Ann Ny Acad Sci 976:421-430, 2002.
18. Patten RD, Pourati I, Aronovitz MJ, Baur J, Celestin F, Chen X, Michael A, Haq S, Nuedling S,
Grohe C, Force T, Mendelsohn ME, and Karas RH. 17beta-estradiol reduces cardiomyocyte
apoptosis in vivo and in vitro via activation of phospho-inositide-3 kinase/Akt signaling. Circ Res
95:692-629, 2004.
19. Pines A, Fisman EZ, Shemesh J, Levo Y, Ayalon D, Kellermann JJ, Motro M, and Drory Y.
Menopause-related changes in left ventricular function in healthy women. Cardiology 80:413-
416,1992.
Page 16 of 30

17
20. Reis SE, Holubkov R, young JB, White BG, Cohn JN, and Fledman AM. Estrogen is associated
with improved survival in aging women with congestive heart failure: analysis of the vesnarinone
studies. J Am Coll Cardiol 36:529-533,2000.
21. Ren J, Hintz KK, Roughead ZK, Duan J, Colligan PB, Ren BH, Lee KJ, and Zeng H. Impact of
estrogen replacement on ventricular myocyte contractile function and protein kinase B/Akt
activation. Am J Physiol Heart Circ Physiol 284:H1800-7, 2003.
22. Rice JJ and de Tombe PP. Approaches to modeling crossbridges and calcium-dependent
activation in cardiac muscle. Prog Biophys Mol Biol 85:179-95, 2004.
23. Schaible TF, Scheuer J. Comparison of heart function in male and female rats. Basic Res Cardiol
79:402-412, 1984.
24. Schaible TF, Malhotra A, Ciambrone G, and Scheuer J. The effects of gonadectomy on left
ventricular function and cardiac contractile proteins in male and female rats.Circ Res 54:38-49,
1984.
25. Solaro R J. (2001) Modulation of cardiac myofilament activity by protein phosphorylation.
Handbook of Physiology: Section 2: The Cardiovascular System. Volume 1 The Heart (E. Page, H.
Fozzard, R. J. Solaro, Eds.) Oxford University Press, New York, pp 264-300.
26. Solaro RJ, Lee JA, Kentish JC, and Allen DG. Effects of acidosis on ventricular muscle from
adult and neonatal rats. Circ Res 63:779-87, 1988.
27. Solaro RJ and Hasenfuss G. Control of cardiac function in health and disease by mechanisms at
the level of the sarcomere. In: Molecular Mechanisms of Cardiac Hypertrophy and Failure (Walsh
RA, ed) Taylor and Francis Group (London) pp 265-279, 2005.
28. Sugishita K, Su Z, Li F, Philipson KD, and Barry WH. Gender influence [Ca2+ ]I during
metabolic inhibition in myocytes over expressing the Na+-Ca2+ exchanger. Circulation 104: 2102-
2106, 2001.
29. Turgeon JL, McDonnell DP, Martin KA, and Wise PM. Hormone therapy: physiological
complexity belies therapeutic simplicity. Science 304:1269-1273, 2004.
Page 17 of 30

18
30. Tytgat J. How to isolate cardiac myocytes. Cardiovasc Res 28:280-283, 1994.
31. Urboniene D, Dias F, Peña JR, Walker LA, Solaro RJ, and Wolska BM. Expression of Slow
Skeletal Troponin I in Adult Mouse Heart Helps to Maintain the Left Ventricular Systolic Function
During Respiratory Hypercapnia. Circ Res 97:70-77, 2005.
32. Wattanapermpool J. Increase in calcium responsiveness of cardiac myofilament activation in
ovariectomized rats. Life Sci 63:955-964, 1998.
33. Wattanapermpool J and Reiser PJ. Differential effects of ovariectomy on calcium activation of
cardiac and soleus myofilaments. Am J Physiol 277:H467-H473, 1999.
34. Wattanapermpool J, Reiser PJ, and Solaro RJ. Troponin I isoforms and differential effects of
acidic pH on soleus and cardiac myofilaments. Am J Physiol 268:C323-30, 1995.
35. Wattanapermpool J, Riabroy T, and Preawnim S. Estrogen suppement prevents the calcium
hypersensitivity of cardiac myofilaments in ovariectomized rats. Life Sci 66:533-543, 2000.
36. Wolff MR, Buck SH, Stoker SW, Greaser ML, and Mantzer RM. Myofibrillar calcium sensitivity of
isometric tension is increased in human dilated cardiomyopathies: role of altered beta-
adrenergically mediated protein phosphorylation. J Clin Invest 98:167-176, 1996.
37. Wolska BM, Averyhart-Fullard V, Omachi A, Stojanovic MO, Kallen RG, and Solaro RJ.
Changes in thyroid state affect pHi and Nai+ homeostasis in rat ventricular myocytes. J Mol Cell
Cardiol 29:2653-2663, 1997.
38. Wolska BM, Kitada Y., Palmiter KA., Westfall M.A., Johnson MD. and Solaro R J. CGP48506,
a novel benzodiazocine derivative, increases contractility of ventricular myocytes and myofilaments
by direct effects on the actin-myosin reaction. Am J Physiol (Heart) 270:H24-H32, 1996
39. Wolska BM, Vijayan K, Arteaga GM, Konhilas JP, Phillips RM, Kim R, Naya T, Leiden JM,
Martin AF, de Tombe PP, and Solaro RJ. Expression of slow skeletal troponin I in adult
transgenic mouse heart muscle reduces the force decline observed during acidic conditions. J
Physiol 536:863-70, 2001.
40. Wolska BM, Stojanovic MO, Luo W, Kranias EG. and Solaro RJ. Effect of ablation of
Page 18 of 30

19
phospholamban on dynamics of cardiac myocyte contraction and intracellular Ca2+. Am. J.
Physiol:Cell 271:C391-C397, 1996.
Page 19 of 30

20
Figure Legends
Figure 1. Effects of ovariectomy on relationship between the per cent maximum shortening
and amplitude of calcium transient (reported as the Fura-2 ratio) of cardiac myocytes isolated
from hearts of shams, OVX rats, and OVX rats supplemented with E2. Data are means ± SEM
from 10 myocytes. * Significantly different from shams, P < 0.05; # significantly different from
OVX, P < 0.05.
Figure 2. A. pCa-tension relations of skinned fiber bundles from sham, OVX and OVX + E2
rats at three different pH values. B. Comparison of pC50 values. Data are means ± SEM
from 10 preparations. * Significantly different from shams, P < 0.05; # significantly different
from OVX, P < 0.05.
Figure 3. A. Representative tracings showing effects of acidosis on shortening during twitch
contractions with and without the NHE1 inhibitor cariporide of myocytes isolated from hearts of
shams, OVX rats, and OVX rats supplemented with E2. B. Data summarizing extent of
shortening in the absence (top panel) and presence (bottom panel) of cariporide at 5 and 10
min of acidosis and following recovery in myocytes from hearts of shams, OVX rats, and OVX
rats supplemented with E2. Percent shortening under control conditions was taken as 100%.
Data are means ± SEM from 10 myocytes. * P < 0.05 = significantly different from sham.
Figure 4. Data summarizing of Ca2+ transient amplitude in the absence (top panel) and
presence (bottom panel) of cariporide at 5 and 10 min of acidosis and following recovery in
Page 20 of 30

21
myocytes from hearts of shams, OVX rats, and OVX rats supplemented with E2. Percent
shortening under control conditions was taken as 100%. Data are means ± SEM from 10
myocytes. * P < 0.05 = significantly different from sham.
Figure 5. Data demonstrating no change in expression of Na+-H+ exchanger 1 (NHE1). A.
NHE1 expression of studied in 7 separate hearts from sham-operated (SHAM), 7 from
ovariectomized (OVX) and 9 from OVX with estrogen supplementation (OVX+E2). Expression
was normalized to ribosomal protein L7 and the data are means ± SEM. B. Representative
immunoblot image of proteins and calsequestrin (CQ) of SHAM, OVX, and OVX+E2.
Page 21 of 30

22
TABLE 1. General characteristics of sham controls, ovariectomized, and ovarectomized With
estrogen supplementation rats
Parameters SHAM OVX OVX+E2
n 18 19 19
Body Weight, g 267 ± 3.3 323 ± 3.4* 251 ± 3.2
Heart Weight, g 1.11 ± 0.01 1.26 ± 0.02* 1.10 ± 0.02
HW/BW Ratio 0.41 ± 0.01 0.39 ± 0.01 0.438 ± 0.01
Uterine Weight, g 0.56 ± 0.02 0.12 ± 0.01* 0.34 ± 0.02 *#
Results are means ± SEM from each group. SHAM represents sham-operated control;
OVX, ovariectomized rats; OVX+E2, ovariectomized plus estrogen supplementation. *P< 0.05
vs SHAM, #P<0.05 vs OVX.
Page 22 of 30

23
TABLE 2. Contraction/relaxation kinetics of myocytes isolated from hearts of sham controls,
ovariectomized, and ovariectomized with estrogen supplementation rats.
Parameters SHAM OVX OVX+E2
Hearts 4 5 5
Cells, n 14 19 18
% Shortening 8.35 ± 0.39 6.50 ± 0.27* 8.23 ± 0.37#
Time to Peak Ampitude of
shortening, msec
173 ± 11 228 ± 13* 233 ± 8.9*
T80, msec 170 ± 13 334 ± 22* 236 ± 18* #
+dL/dt, µm/sec 104.3 ± 6.8 71.0 ± 4.9* 74.2 ± 5.5*
-dL/dt, µm/sec 92.0 ± 7.9 48.6 ± 4.7* 57.6 ± 5.2*
Results are mean ± SE; n = number of cells; T80, time required for cell to 80% of full
relaxation from the peak of shortening; +dL/dt, rate of cell shortening; -dL/dt, rate of cell re-
lengthening. * P< 0.05 vs SHAM; #P < 0.05 vs OVX.
Page 23 of 30

24
TABLE 3. Ca2+ transient kinetics of myocytes isolated from hearts of sham controls,
ovariectomized, and ovariectomized with estrogen supplementation rats.
Parameters SHAM OVX OVX+E2
Hearts 7 6 6
Cells, n 20 20 20
Baseline, ratio unit 1.36 ± 0.04 1.33 ± 0.04 1.37 ± 0.03
Amplitude, ratio unit 2.12 ± 0.13 1.74 ± 0.10* 2.26 ± .14#
Time to Peak, msec 95.3 ± 4.3 95.5 ± 1.9 97.7 ± 2.2
T50, msec 180 ± 10.7 232 ± 9.3* 196 ± 9.3
Results are means ± SEM; n = number of cells; baseline, 340/380 ratio at rest;
amplitude, peak of 340/380 ratio minus baseline ratio; T50, times required for cell to achieve
50% of full relaxation of Ca2+ signal to baseline ratio from peak ratio. * P< 0.05 vs SHAM.
Page 24 of 30

Figure 1. Effects of ovariectomy on relationship between the per cent maximum shortening and amplitude of calcium transient (reported as the Fura-2 ratio) of cardiac myocytes isolated from hearts of shams, OVX rats, and OVX rats supplemented with E2. Data are means ± SEM from 10 myocytes. * Significantly different from shams, P < 0.05;
# significantly different from OVX, P < 0.05.
Page 25 of 30
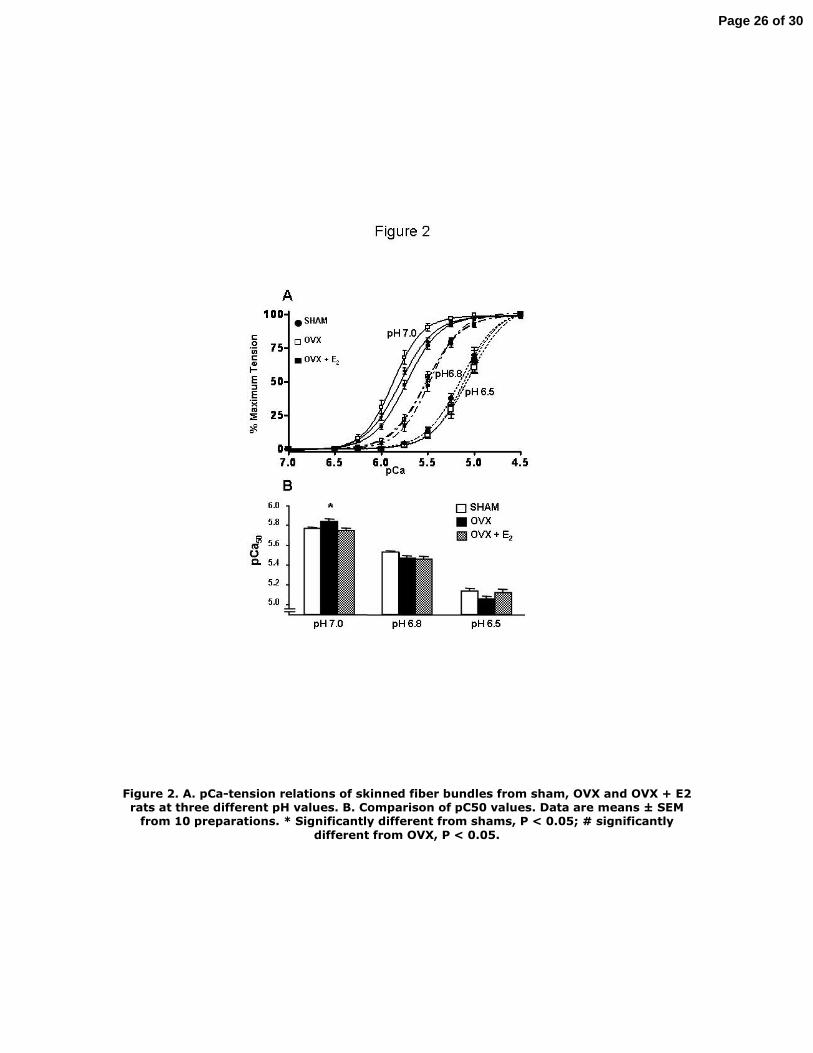
Figure 2. A. pCa-tension relations of skinned fiber bundles from sham, OVX and OVX + E2 rats at three different pH values. B. Comparison of pC50 values. Data are means ± SEM
from 10 preparations. * Significantly different from shams, P < 0.05; # significantly different from OVX, P < 0.05.
Page 26 of 30

Figure 3. A. Representative tracings showing effects of acidosis on shortening during twitch contractions with and without the NHE1 inhibitor cariporide of myocytes isolated
from hearts of shams, OVX rats, and OVX rats supplemented with E2.
Page 27 of 30

B. Data summarizing extent of shortening in the absence (top panel) and presence (bottom panel) of cariporide at 5 and 10 min of acidosis and following recovery in
myocytes from hearts of shams, OVX rats, and OVX rats supplemented with E2. Percent shortening under control conditions was taken as 100%. Data are means ± SEM from 10
myocytes. * P < 0.05 = significantly different from sham.
Page 28 of 30

Figure 4. Data summarizing of Ca2+ transient amplitude in the absence (top panel) and presence (bottom panel) of cariporide at 5 and 10 min of acidosis and following recovery in myocytes from hearts of shams, OVX rats, and OVX rats supplemented with E2. Percent shortening under control conditions was taken as 100%. Data are means ± SEM from 10
myocytes. * P < 0.05 = significantly different from sham.
Page 29 of 30
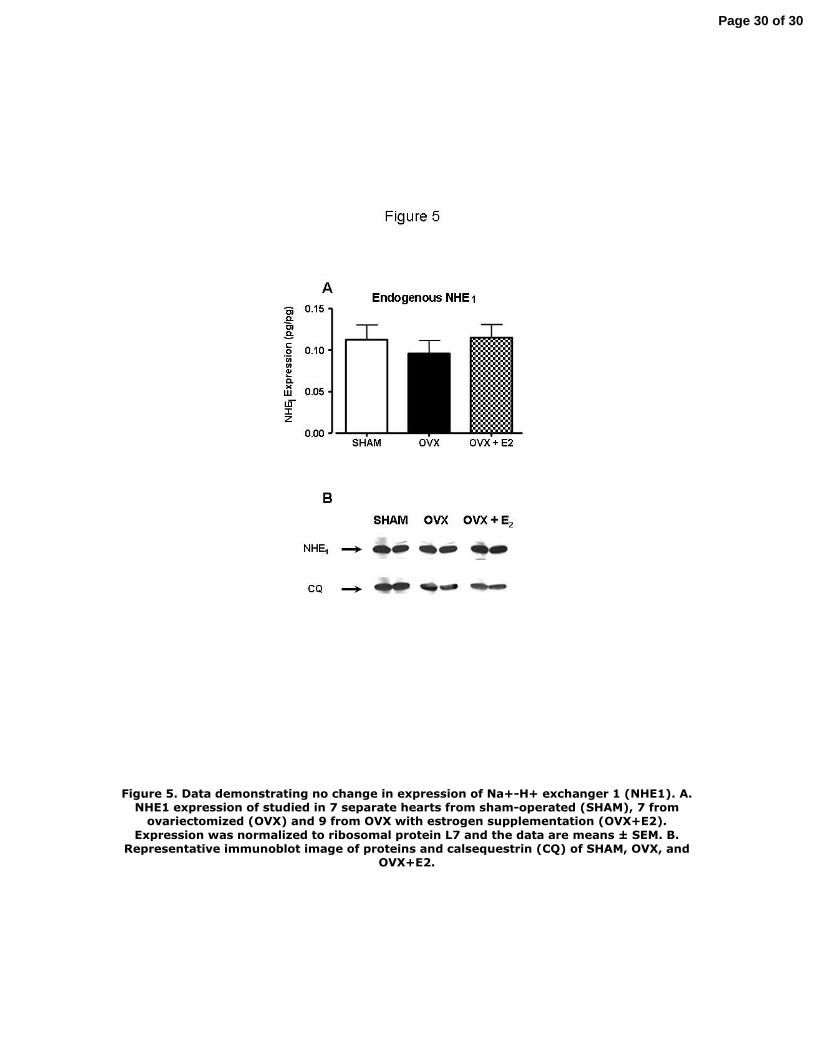
Figure 5. Data demonstrating no change in expression of Na+-H+ exchanger 1 (NHE1). A. NHE1 expression of studied in 7 separate hearts from sham-operated (SHAM), 7 from
ovariectomized (OVX) and 9 from OVX with estrogen supplementation (OVX+E2). Expression was normalized to ribosomal protein L7 and the data are means ± SEM. B.
Representative immunoblot image of proteins and calsequestrin (CQ) of SHAM, OVX, and OVX+E2.
Page 30 of 30




















