
Myocyte differentiation and body wall muscle regeneration in the planarian Girardia tigrina
11
&p.1:Abstract Freshwater planarians (Platyhelminthes, Tur- bellaria) show a great degree of morphological plasticity, making them a useful model for studying cell differentia- tion and pattern restoration processes during regenera- tion. Using confocal microscopy and a monoclonal anti- body specific for muscle cells (TMUS-13), we have monitored the restoration of the body wall musculature during head regeneration in whole-mount organisms. Our results show that until the 4th day of regeneration the blastema is occupied by very disorganized muscle fi- bers, that from this moment become progressively orga- nized restoring the original muscle pattern. In addition to recognizing mature muscle cells, TMUS-13 also recog- nizes differentiating myocytes, allowing us to trace the origin of newly formed muscle cells. We report that myocytes are detected in the postblastema region as ear- ly as day 1 of regeneration. This is the first demonstra- tion that, in addition to serving as a proliferative zone as previously described, overt differentiation begins in the postblastema, at least for muscle cells. We also show that the TMUS13 antigen is the myosin heavy-chain gene from planarians. &kwd:Key words Planarian · Muscle system · Myocyte differentiation · Regeneration · Myosin heavy chain &bdy: Introduction Cell differentiation and pattern formation mechanisms are among the most challenging issues in developmental biology. Many studies about cell proliferation and differ- entiation, morphogenesis and pattern formation have been done to understand embryonic development in many organisms. However, less is known about how all these processes are carried out during regeneration. Freshwater planarians (Platyhelminthes, Turbellaria, Tri- cladida) are useful organisms for studying the cellular and molecular mechanisms leading to pattern restoration because of their great regenerative power (for general re- views, see Baguñà et al. 1990, 1994). These organisms are acoelomate, triploblastic and bilaterally symmetrical, with pronounced antero-posterior and dorso-ventral po- larities. In addition to their regenerative capacities, fresh- water planarians are able to grow and degrow (literally, shrinkage of the animal due to changes in the balance be- tween cell proliferation and cell death) in volume and length in a continuous manner depending on food avail- ability and temperature (Romero 1987; Romero and Baguñà 1988; Romero and Baguñà 1991). All these pro- cesses are based upon the proliferation and differentia- tion of undifferentiated, totipotent stem-cells called neo- blasts (Brøndsted 1969; Baguñà 1973). During regenera- tion, neoblasts (20–30% of total cell number; Hay and Coward 1975; Baguñà 1976; Romero 1987) located be- low the wound epithelium proliferate to form a blastema and then differentiate into all of the cell types necessary to replace the missing structures (Baguñà and Romero 1981; Saló and Baguñà 1984). The morphological plas- ticity shown by these organisms means that the main body axes must be continuously re-specified in the adult. All these features make planarians a useful model for studying developmental processes. It has been suggested (Saló and Baguñà 1989) that pattern formation in pla- narians follows a morphallactic-epimorphic model, with an early morphallactic phase (pattern restoration in the absence of mitosis) in which pattern is set in a disto- proximal sequence within a narrow strip of tissue below the wound and a later phase of amplification by epimor- phosis (based upon cell proliferation). To examine how the original pattern is restored during regeneration we are studying restoration of the planarian muscle. Functionally, these cells are important because Edited by D.A. Weisblat F. Cebrià ( ✉ ) · M. Vispo · P. Newmark 1 · D. Bueno · R. Romero Departament de Genètica, Facultat de Biologia, Universitat de Barcelona, Av. Diagonal 645, E-08071 Barcelona, Catalonia, Spain Present address: 1 Department of Embryology, Carnegie Institution of Washington, 115 West University Parkway, Baltimore, MD 21210, USA&/fn-block: Dev Genes Evol (1997) 207:306–316 © Springer-Verlag 1997 ORIGINAL ARTICLE &roles:Francesc Cebrià · Marcelo Vispo Phillip Newmark · David Bueno · Rafael Romero Myocyte differentiation and body wall muscle regeneration in the planarian Girardia tigrina &misc:Received: 15 April 1997 / Accepted: 7 July 1997
Transcript of Myocyte differentiation and body wall muscle regeneration in the planarian Girardia tigrina

&p.1:Abstract Freshwater planarians (Platyhelminthes, Tur-bellaria) show a great degree of morphological plasticity,making them a useful model for studying cell differentia-tion and pattern restoration processes during regenera-tion. Using confocal microscopy and a monoclonal anti-body specific for muscle cells (TMUS-13), we havemonitored the restoration of the body wall musculatureduring head regeneration in whole-mount organisms.Our results show that until the 4th day of regenerationthe blastema is occupied by very disorganized muscle fi-bers, that from this moment become progressively orga-nized restoring the original muscle pattern. In addition torecognizing mature muscle cells, TMUS-13 also recog-nizes differentiating myocytes, allowing us to trace theorigin of newly formed muscle cells. We report thatmyocytes are detected in the postblastema region as ear-ly as day 1 of regeneration. This is the first demonstra-tion that, in addition to serving as a proliferative zone aspreviously described, overt differentiation begins in thepostblastema, at least for muscle cells. We also show thatthe TMUS13 antigen is the myosin heavy-chain genefrom planarians.
&kwd:Key words Planarian · Muscle system · Myocytedifferentiation · Regeneration · Myosin heavy chain&bdy:
Introduction
Cell differentiation and pattern formation mechanismsare among the most challenging issues in developmentalbiology. Many studies about cell proliferation and differ-
entiation, morphogenesis and pattern formation havebeen done to understand embryonic development inmany organisms. However, less is known about how allthese processes are carried out during regeneration.Freshwater planarians (Platyhelminthes, Turbellaria, Tri-cladida) are useful organisms for studying the cellularand molecular mechanisms leading to pattern restorationbecause of their great regenerative power (for general re-views, see Baguñà et al. 1990, 1994). These organismsare acoelomate, triploblastic and bilaterally symmetrical,with pronounced antero-posterior and dorso-ventral po-larities. In addition to their regenerative capacities, fresh-water planarians are able to grow and degrow (literally,shrinkage of the animal due to changes in the balance be-tween cell proliferation and cell death) in volume andlength in a continuous manner depending on food avail-ability and temperature (Romero 1987; Romero andBaguñà 1988; Romero and Baguñà 1991). All these pro-cesses are based upon the proliferation and differentia-tion of undifferentiated, totipotent stem-cells called neo-blasts (Brøndsted 1969; Baguñà 1973). During regenera-tion, neoblasts (20–30% of total cell number; Hay andCoward 1975; Baguñà 1976; Romero 1987) located be-low the wound epithelium proliferate to form a blastemaand then differentiate into all of the cell types necessaryto replace the missing structures (Baguñà and Romero1981; Saló and Baguñà 1984). The morphological plas-ticity shown by these organisms means that the mainbody axes must be continuously re-specified in the adult.All these features make planarians a useful model forstudying developmental processes. It has been suggested(Saló and Baguñà 1989) that pattern formation in pla-narians follows a morphallactic-epimorphic model, withan early morphallactic phase (pattern restoration in theabsence of mitosis) in which pattern is set in a disto-proximal sequence within a narrow strip of tissue belowthe wound and a later phase of amplification by epimor-phosis (based upon cell proliferation).
To examine how the original pattern is restored duringregeneration we are studying restoration of the planarianmuscle. Functionally, these cells are important because
Edited by D.A. Weisblat
F. Cebrià (✉) · M. Vispo · P. Newmark1 · D. Bueno · R. RomeroDepartament de Genètica, Facultat de Biologia,Universitat de Barcelona, Av. Diagonal 645,E-08071 Barcelona, Catalonia, Spain
Present address:1Department of Embryology, Carnegie Institution of Washington,115 West University Parkway, Baltimore, MD 21210, USA&/fn-block:
Dev Genes Evol (1997) 207:306–316 © Springer-Verlag 1997
O R I G I N A L A RT I C L E
&roles:Francesc Cebrià · Marcelo VispoPhillip Newmark · David Bueno · Rafael Romero
Myocyte differentiation and body wall muscle regenerationin the planarian Girardia tigrina
&misc:Received: 15 April 1997 / Accepted: 7 July 1997

307
they act early in regeneration, closing the wound by con-traction of the body wall in the area where the regenera-tion blastema will form (Chandebois 1980). Experimen-tally, we have obtained muscle-specific monoclonal anti-bodies (mAbs; Romero et al. 1991; Bueno 1997a) thatcan be used as markers to monitor the restoration of themuscle pattern within the blastema during regeneration,as well as for tracing the differentiation of neoblasts firstinto myocytes and then into mature muscle fibers. Phylo-genetically, Platyhelminthes are placed at the base oftriploblastic organisms (Ax 1987; Brusca and Brusca1990); thus, studying muscle cell differentiation andmuscle fiber assembly in these organisms may providesome clues about the origin of the three-dimensionallycomplex body wall musculature. Moreover, in recentyears myogenesis has become a good model for studyinghow undifferentiated cells are committed to their finalfate through cell proliferation and differentiation. Thus,studying myogenesis in planarians may be useful fortracing the evolutionary origin of the mechanisms con-trolling this process.
Previous work on muscle differentiation during regen-eration in other planarian species has utilized electronmicroscopy (Pedersen 1972; Hori 1983; Morita and Best1984). However, to get a more comprehensive view ofthe process it is necessary to analyze whole-mount prep-arations. In this report, we analyze muscle system orga-nization in intact organisms, as well as its restorationduring regeneration, using two different markers: phal-loidin (Wulf et al. 1979) and the mAb TMUS-13 (Buenoet al. 1997a). This mAb binds specifically to all musclecells of the organism and previous work suggested thatTMUS-13 also recognizes myocytes (Espinosa 1993;Bueno et al. 1997b). This capability of TMUS-13 forrecognizing differentiating myocytes makes it a goodmarker for studying muscle cell differentiation duringthe processes of cell renewal and regeneration. Finally,we also present data showing that the antigen recognizedby TMUS-13 is the myosin heavy chain.
Materials and methods
Species and culture conditions
The planarians used belong to an asexual race (class A; Ribas etal. 1989) of the species Girardia tigrina (Girard 1850; Ball 1974;De Vries and Sluys 1991) collected near Barcelona. They weremaintained in spring water in the dark at 4–6°C and fed once amonth with beef liver. Specimens chosen for the different experi-ments were starved for 15 days before use.
Regenerating organisms and terminology
For regeneration studies, organisms 9–10 mm in length were cutpostcephalically (behind the auricles; Saló and Baguñà 1984) andkept at a temperature of 17 ± 1°C. In regenerating organisms, anteri-or blastema refers to the small unpigmented mound of tissue madeof small undifferentiated cells (neoblasts) that forms and growsabove the wound in anterior (head) regenerates. Postblastema refersto the pigmented old region directly behind the blastema, and iswhere most mitotic activity occurs during regeneration (Fig. 1).
Whole-mount staining with FITC-phalloidin
Intact organisms were fixed for 15 min at room temperature (RT)in 3.7% formaldehyde (Merck). After washing for 10 min in phos-phate-buffered saline (PBS; 8 g/l NaCl, 0.2 g/l KCl, 0.2 g/lKH2PO4, 1.4 g/l Na2HPO4 2H2O pH 7.4), they were permeabilizedin 0.3% Triton X-100 (Merck) in PBS for 10 min at RT. Theywere then washed for 15 min in PBS and incubated with fluores-cein isothiocyanate(FITC)-phalloidin (Molecular Probes) diluted1:100 in PBS overnight at 4°C. After washing in PBS three timesfor 5 min, fluorescence was detected with confocal microscopy.
Whole-mount immunostaining with TMUS-13
Intact and regenerating organisms were treated with macerationsolution (Espinosa 1993) – glacial acetic acid: glycerol: PBS(1:1:13) – for 15 min at room temperature and then fixed in 4%paraformaldehyde (Merck) in PBS for 3 h at 4°C. After fixationanimals were bleached in 10% H2O2 in MeOH overnight at 4°Cand then washed twice for 5 min in MeOH, 5 min in PBST (0.3%Triton-X100 in PBS): MeOH (1:1) and 5 min in PBST. The ani-mals were blocked for 1 h at room temperature in 0.25% bovineserum albumin (BSA; Sigma) in PBST and then incubated inTMUS-13 diluted 1:10 in 0.25% BSA in PBST for 24 h at roomtemperature (shaking). The samples were then washed in 0.25%BSA in PBST for 24 h at room temperature (several changes ofthe medium) and TMUS13 was detected with secondary goat-anti-mouse conjugated to fluorescein (GAM-FITC; Sigma) diluted1:75 or Texas red (GAM-TR; Ammersham) diluted 1:100 in0.25% BSA in PBST for 24 h at room temperature (shaking). Af-ter washing in PBST for a few hours, specimens were mounted in50% glycerol in PBS; 2.3% n-propyl gallate and the fluorescencewas detected with confocal microscopy.
Confocal microscopy
Confocal laser scanning microscopy (CLSM) was performed witha Leica TCS 4D (Leica Lasertechnik, Heidelberg) adapted to aninverted microscope (Leitz DMIRB). Images were taken usingX40 (NA 1.0) or X100 (NA 1.3) Leitz Plan Fluotar objectives.FITC-phalloidin, FITC-goat anti-mouse or TR-goat anti-mousewere excited at 488 nm using a krypton-argon laser. Each imagecollected was the average of eight line scans at the standard scanrate. Images of 512 × 512 pixels were processed and printed in aShinko CHC-S445 full color printer. All the images are in thesame pseudo color (glow look-up table) for either fluorochromeused (FITC or Texas Red). In the figures where it has been neces-
Fig. 1 Dorsal view of intact and head-regenerating Girardia tigr-ina (ph pharynx)&/fig.c:

sary to distinguish the myocytes from the mature muscle fibers(both immunostained with TMUS-13), two different pseudo colors(green and red) have been assigned. In these cases, two indepen-dent 3D projections were performed from different confocal mi-croscopic sections and then superimposed.
cDNA library screening
Monoclonal antibody TMUS-13 was used to screen a G. tigrinacDNA library constructed in LAMBDA ZAP (Stratagene). Afterinduction in 10 mM Isopropyl β-D-Thiogalactopyranoside (IPTG;Sigma) IPTG for 4 h at 37°C, nitrocellulose filters (Schleicher andSchuell) were blocked in 5% non-fat dry milk in PBS overnight at4°C (shaking). After washing twice for 5 min in PBS at room tem-perature, the filters were incubated with TMUS-13 diluted 1:100in MTP (0.5% non-fat dry milk, 0.05% Tween-20 in PBS) for 24 hat 4°C (shaking). After washing three times for 10 min in MTP atroom temperature, the filters were incubated with the secondaryantibody (horseradish peroxidase-conjugated goat anti-mouse se-rum; Sigma) diluted 1:3000 in MTP at room temperature for 2 h.After washing twice for 5 min in MTP the immunolabels were de-tected using diaminobenzidine tetrahydrochloride (DAB; Sigma)0.5 mg/ml, CoCl2 0.05%, H2O2 (1:104) in PBS. The reaction wasstopped with distilled water and the filters were washed severaltimes in PBS. Eight positive clones from a screening of 5 × 105
phages were isolated and analyzed by PCR and restriction enzymedigestion; all contained cDNA inserts of equal lengths.
Sequencing of TMUS13 cDNA
In vivo excision of pBluescript from positive clones was per-formed according to manufacturer’s instructions. After transfor-mation, the TMUS13 cDNA was sequenced using M13 and M13reverse universal primers. Cycle sequencing using dye-labeled ter-minators (PrismTM Ready Reaction DyeDeoxyTM TerminatorCycle Sequencing Kit) was performed in a Perkin-Elmer 480 ther-mal cycler. Analysis of the cDNA sequence was carried outthrough the BLAST WWW Server (Altschul et al. 1990).
Southern blot analysis
Ten µg genomic DNA of G. tigrina (Garcia-Fernàndez et al. 1993)were digested with excess EcoRI or HindIII (Promega). After sep-aration on 0.8% agarose gel, the DNA was transferred to a Hy-bond-N+ membrane (Amersham) by capillary blotting, and hy-bridized with TMUS13 cDNA labeled with [32-P]dCTP by randompriming. The hybridization was carried out at 42°C under highstringency conditions: 50% formamide, 0.1% sodium dodecyl sul-fate (SDS), 5 × Denhardt’s solution (1 × Denhardt’s: 0.2 mg/mleach ficoll, bovine serum albumin and polyvinylpyrrolidone),4 × SSPE (3.6 M NaCl, 0.2 M NaH2PO4, 0.02 M ethylenediaminetetraacetic acid) and 100µg/ml denatured herring sperm DNA. Af-ter hybridization filters were washed in 0.5 × sodium sodium ci-trate (SSC), 0.1% SDS for 20 min at 65°C and exposed to X-rayfilm at –70°C using two intensifying screens.
Fusion peptide construction and western blot analysis
TMUS13 cDNA was digested with BamHI and XhoI (Promega)and cloned into pGEX-4T-1 (Pharmacia) using the DNA ligationsystem (Amersham). After transformation according to Hanahan(1983), fusion peptide expression was induced in 0.1 mM IPTG.Bacterial proteins were obtained through sonication (BransonSonifier 250), and batch purification of fusion peptide using bulkglutathione sepharose 4B was carried out according to manufac-turer’s instructions. Purified GST-TMUS13 fusion peptide was runinto a 10% SDS-PAGE (Mini Protean II; Bio-Rad) according to
Laemmli (1970). Electrophoresed proteins were transferred to ni-trocellulose membrane using a semi-dry transfer system from Bio-metra (Fast-Blot B33). Membranes were incubated with TMUS-13diluted 1:100 in PBS for 4 h at room temperature, washed in MTPfor 30 min and incubated with peroxidase-conjugated anti-mouseIgG (Sigma) for 2 h at room temperature. After washing withMTP for 1 h at RT, the membranes were visualized with DAB(Sigma).
Results
The body wall musculature in intact flatwormsas revealed by FITC-phalloidin staining
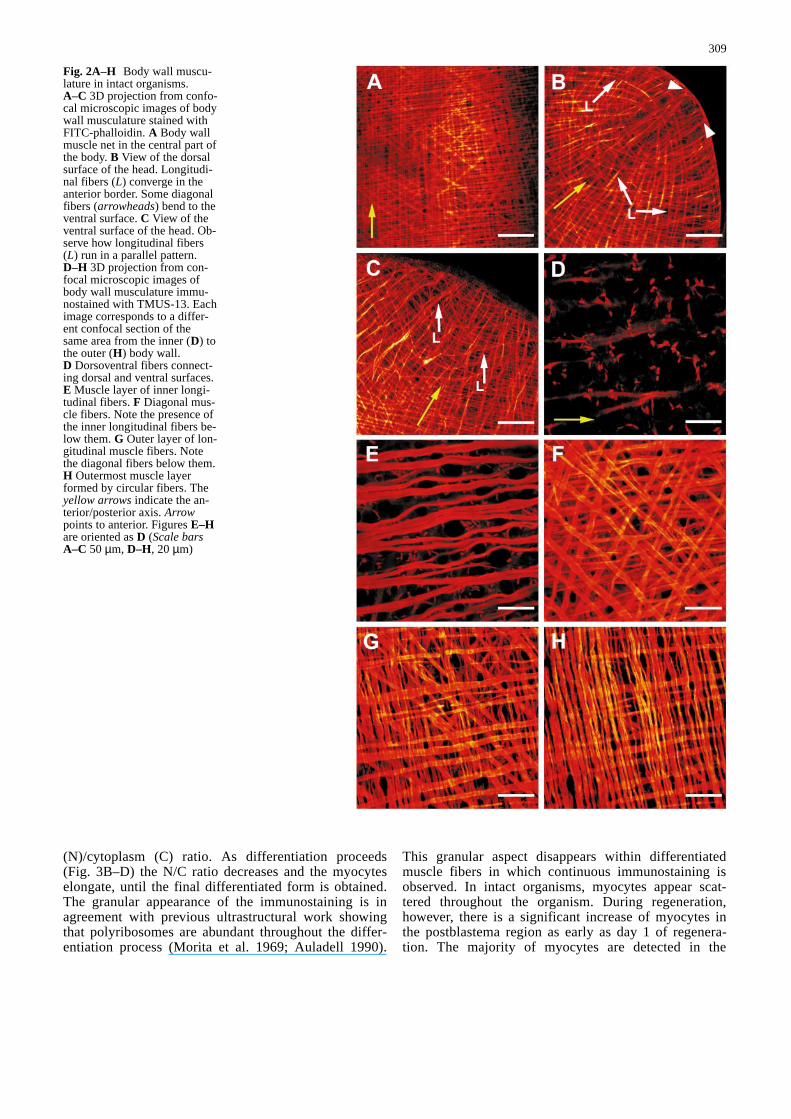
Confocal microscopic analysis of FITC-phalloidin-stained whole-mount preparations shows that the bodywall musculature of G. tigrina is formed by four subepi-dermal layers (Fig. 2A) arranged as follows (from theoutside to the inside of the body wall): an outer layer ofcircular fibers, a thin layer of longitudinal fibers, a layerof diagonal fibers and finally, an inner layer of longitudi-nal fibers. These muscle layers are compressed within athickness of 7–12µm below the basal lamina and thesame pattern (outer circular, thin longitudinal, diagonal,inner longitudinal) is observed along both the dorsal andventral surfaces. Dorsoventral muscle fibers connectthese two surfaces and are more abundant in the head,tail and lateral regions than in the central region of thebody. All of these muscle fibers are arranged in a denseand compact muscle net. Careful examination of the in-nermost longitudinal fibers in the anterior tip of the headshows that the dorsal and ventral patterns are quite dif-ferent: dorsal longitudinal fibers converge upon a centralzone near the anterior border (Fig. 2B) whereas ventralfibers run in parallel or even diverge in a fan-shaped pat-tern as they approach the anterior border (Fig. 2C). Fi-nally, Fig. 2B shows some diagonal dorsal fibers bendingto the ventral surface suggesting a continuity of these fi-bers in both surfaces.
TMUS-13 immunostaining identifies planarian myocytes
In intact organisms the body muscle pattern revealed byTMUS-13 immunostaining is identical to that describedby phalloidin staining. Figure 2D–H shows the differentlayers of muscle fibers that form the body wall muscula-ture of this species. Inner longitudinal fibers (Fig. 2E)are slightly thicker and usually appear anastomosed, incontrast with the outer layer of longitudinal fibers(Fig. 2G). Outermost circular fibers are the thinnest andmost numerous fibers (Fig. 2H). In contrast to phalloi-din staining (phalloidin binds to F-actin of mature mus-cle cells), however, TMUS-13 detects myocytes at dif-ferent stages of differentiation. These myocytes arefound in both intact and regenerating organisms. Figure3A corresponds to a myocyte in an early stage of dif-ferentiation still showing some features of neoblastmorphology such as a small size and a large nucleus
308
309
Fig. 2A–H Body wall muscu-lature in intact organisms.A–C 3D projection from confo-cal microscopic images of bodywall musculature stained withFITC-phalloidin. A Body wallmuscle net in the central part ofthe body. B View of the dorsalsurface of the head. Longitudi-nal fibers (L) converge in theanterior border. Some diagonalfibers (arrowheads) bend to theventral surface. C View of theventral surface of the head. Ob-serve how longitudinal fibers(L) run in a parallel pattern.D–H 3D projection from con-focal microscopic images ofbody wall musculature immu-nostained with TMUS-13. Eachimage corresponds to a differ-ent confocal section of thesame area from the inner (D) tothe outer (H) body wall.D Dorsoventral fibers connect-ing dorsal and ventral surfaces.E Muscle layer of inner longi-tudinal fibers. F Diagonal mus-cle fibers. Note the presence ofthe inner longitudinal fibers be-low them. G Outer layer of lon-gitudinal muscle fibers. Notethe diagonal fibers below them.H Outermost muscle layerformed by circular fibers. Theyellow arrowsindicate the an-terior/posterior axis. Arrowpoints to anterior. Figures E–Hare oriented as D (Scale barsA–C 50 µm, D–H, 20µm)&/fig.c:
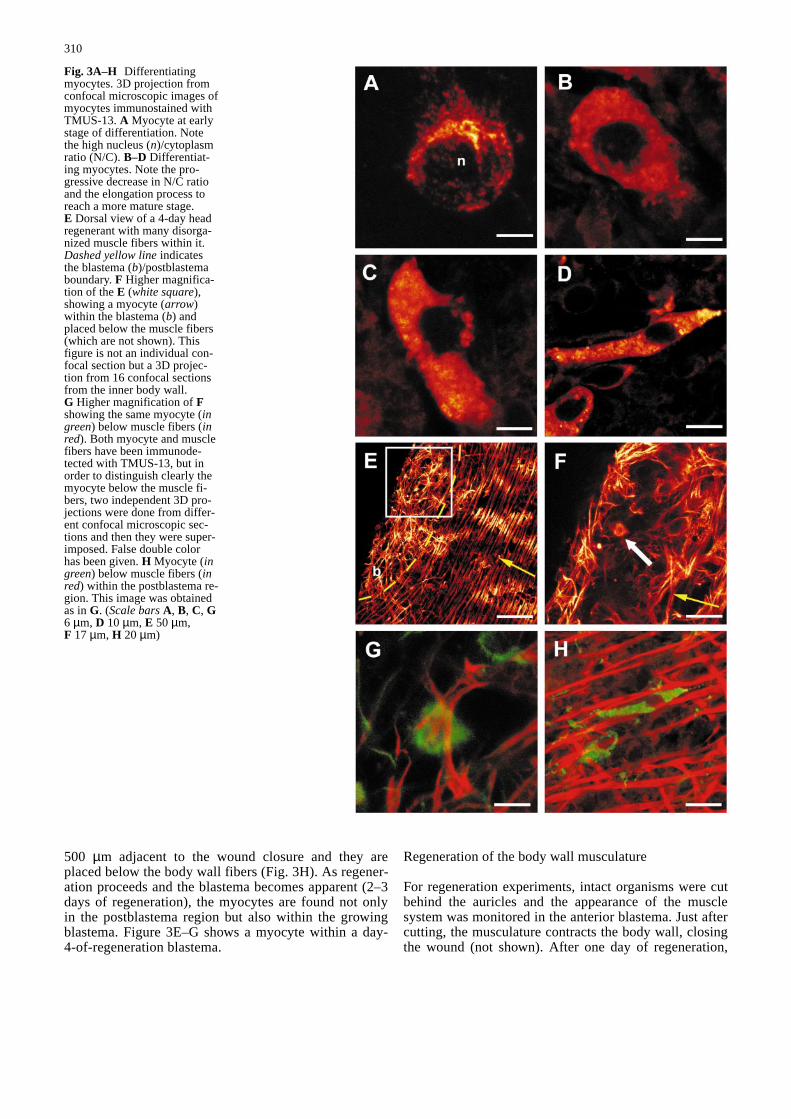
(N)/cytoplasm (C) ratio. As differentiation proceeds(Fig. 3B–D) the N/C ratio decreases and the myocyteselongate, until the final differentiated form is obtained.The granular appearance of the immunostaining is inagreement with previous ultrastructural work showingthat polyribosomes are abundant throughout the differ-entiation process (Morita et al. 1969; Auladell 1990).
This granular aspect disappears within differentiatedmuscle fibers in which continuous immunostaining isobserved. In intact organisms, myocytes appear scat-tered throughout the organism. During regeneration,however, there is a significant increase of myocytes inthe postblastema region as early as day 1 of regenera-tion. The majority of myocytes are detected in the
310
Fig. 3A–H Differentiatingmyocytes. 3D projection fromconfocal microscopic images ofmyocytes immunostained withTMUS-13. A Myocyte at earlystage of differentiation. Notethe high nucleus (n)/cytoplasmratio (N/C). B–D Differentiat-ing myocytes. Note the pro-gressive decrease in N/C ratioand the elongation process toreach a more mature stage.E Dorsal view of a 4-day headregenerant with many disorga-nized muscle fibers within it.Dashed yellow lineindicatesthe blastema (b)/postblastemaboundary. F Higher magnifica-tion of the E (white square),showing a myocyte (arrow)within the blastema (b) andplaced below the muscle fibers(which are not shown). Thisfigure is not an individual con-focal section but a 3D projec-tion from 16 confocal sectionsfrom the inner body wall.G Higher magnification of Fshowing the same myocyte (ingreen) below muscle fibers (inred). Both myocyte and musclefibers have been immunode-tected with TMUS-13, but inorder to distinguish clearly themyocyte below the muscle fi-bers, two independent 3D pro-jections were done from differ-ent confocal microscopic sec-tions and then they were super-imposed. False double colorhas been given. H Myocyte (ingreen) below muscle fibers (inred) within the postblastema re-gion. This image was obtainedas in G. (Scale barsA, B, C, G6 µm, D 10 µm, E 50 µm,F 17 µm, H 20 µm)&/fig.c:
500 µm adjacent to the wound closure and they areplaced below the body wall fibers (Fig. 3H). As regener-ation proceeds and the blastema becomes apparent (2–3days of regeneration), the myocytes are found not onlyin the postblastema region but also within the growingblastema. Figure 3E–G shows a myocyte within a day-4-of-regeneration blastema.
Regeneration of the body wall musculature
For regeneration experiments, intact organisms were cutbehind the auricles and the appearance of the musclesystem was monitored in the anterior blastema. Just aftercutting, the musculature contracts the body wall, closingthe wound (not shown). After one day of regeneration,
311
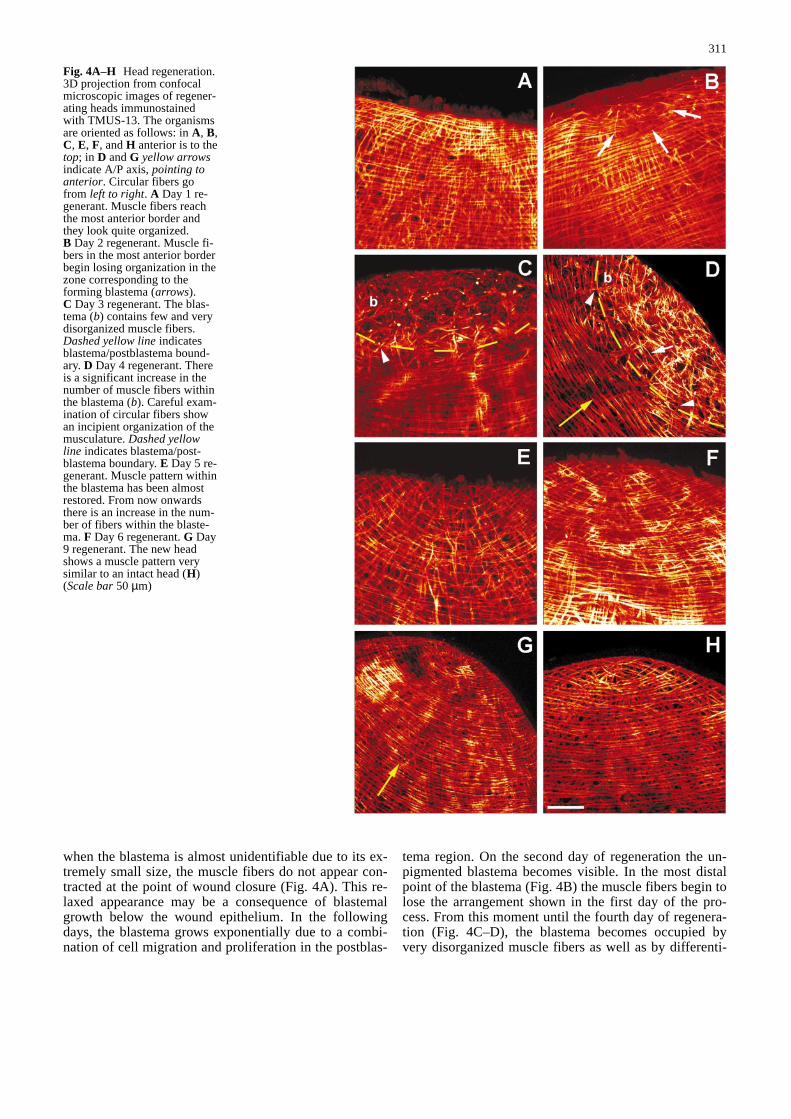
Fig. 4A–H Head regeneration.3D projection from confocalmicroscopic images of regener-ating heads immunostainedwith TMUS-13. The organismsare oriented as follows: in A, B,C, E, F, and H anterior is to thetop; in D and G yellow arrowsindicate A/P axis, pointing toanterior. Circular fibers gofrom left to right. A Day 1 re-generant. Muscle fibers reachthe most anterior border andthey look quite organized.B Day 2 regenerant. Muscle fi-bers in the most anterior borderbegin losing organization in thezone corresponding to theforming blastema (arrows).C Day 3 regenerant. The blas-tema (b) contains few and verydisorganized muscle fibers.Dashed yellow lineindicatesblastema/postblastema bound-ary. D Day 4 regenerant. Thereis a significant increase in thenumber of muscle fibers withinthe blastema (b). Careful exam-ination of circular fibers showan incipient organization of themusculature. Dashed yellowline indicates blastema/post-blastema boundary. E Day 5 re-generant. Muscle pattern withinthe blastema has been almostrestored. From now onwardsthere is an increase in the num-ber of fibers within the blaste-ma. F Day 6 regenerant. G Day9 regenerant. The new headshows a muscle pattern verysimilar to an intact head (H)(Scale bar50 µm)&/fig.c:
when the blastema is almost unidentifiable due to its ex-tremely small size, the muscle fibers do not appear con-tracted at the point of wound closure (Fig. 4A). This re-laxed appearance may be a consequence of blastemalgrowth below the wound epithelium. In the followingdays, the blastema grows exponentially due to a combi-nation of cell migration and proliferation in the postblas-
tema region. On the second day of regeneration the un-pigmented blastema becomes visible. In the most distalpoint of the blastema (Fig. 4B) the muscle fibers begin tolose the arrangement shown in the first day of the pro-cess. From this moment until the fourth day of regenera-tion (Fig. 4C–D), the blastema becomes occupied byvery disorganized muscle fibers as well as by differenti-

ating myocytes, which are also observed in the postblas-tema. On the fourth day of regeneration (Fig. 4D), themuscle net within the blastema becomes denser andmore compact than in previous days. This patterninglikely arises from the myocytes observed in earlier stagescompleting their differentiation towards mature musclecells and forming new fibers. At this stage, within theblastema there is an incipient arrangement of the fibers,which is most evident for the forming circular fibers(Fig. 4D, arrow). At the fifth day of regeneration(Fig. 4E), the muscle fibers within the blastema becomemore organized and the pattern may be considered re-stored. Once the pattern is restored the number of fibersin the blastema increases progressively in each of the fol-lowing days (Fig. 4F–G) until days 10–12, when the den-sity of muscle fibers is rather similar to that found in in-tact organisms (Fig. 4H).
Molecular characterization of TMUS-13 antigen
With the aim of characterizing the antigen recognized byTMUS-13 at the molecular level, a cDNA expression li-brary of G. tigrina was screened with TMUS-13. Theisolated partial cDNA was sequenced and found to con-tain an open reading frame of 1293 nucleotides. BLASTanalysis demonstrated that the TMUS-13 antigen is theproduct of a myosin heavy-chain (mhc) gene, the firstidentified from planarians. Using this cDNA fragment asa probe, southern blot analysis on genomic DNA from G.tigrina was performed. We observe two bands on DNAdigested with EcoRI that are due to a restriction site forthis enzyme within the partial cDNA sequence of themhcgene at bases 590–595. Two bands are also observedon DNA digested with HindIII that could be due to a re-striction site in the genomic DNA to which our probe hy-bridizes (data not shown). To show that the epitope rec-ognized by the mAb TMUS-13 is located in this clonedfragment of 431 amino acids, we have made a glutathi-one-S-transferase (GST)-MHC fusion protein. Westernblot analysis of the GST-MHC fusion protein immunode-tected with mAb TMUS-13 shows that TMUS-13 recog-nizes the 75-kDa fusion protein, as well as some truncat-ed/proteolytic fragments of the fusion protein, which arenot observed in cells expressing GST alone (data notshown). From both experiments we conclude that theepitope recognized by TMUS-13 is located in the myosinheavy-chain protein from G. tigrina.
Discussion
Body wall muscle organization
Although the body wall musculature of turbellariansconsists, in general, of an outer layer of circular fibers,an inner layer of longitudinal fibers and an intermediatelayer of diagonal fibers between them (Rieger et al.1991a), some diversity in the three-dimensional structure
of the musculature appears in different groups. Smallturbellarians like catenulids and some acoels have a thin-ner musculature formed by circular and longitudinalmuscle fibers although no real muscle net is formed(Crezée 1975; Moraczewski 1981). Larger turbellarianshave a thicker musculature usually with a diagonal layerof fibers between circular and longitudinal ones (Riegeret al. 1994; for phylogenetic origin discussion, see Rieg-er et al. 1991b). Other groups, like most polyclads havefive or six muscle layers (Prudhoe 1985). Pedersen(1972) described three muscle layers in the body wall ofG. tigrina. Whole-mount staining and confocal micro-scopic analysis have allowed us to define, for the firsttime, a fourth muscle layer in this species. This fourthlayer of thin longitudinal fibers has been considered acharacter that evolved specifically (synapomorphy) with-in freshwater planarians and it has been used as a mor-phological character to infer the phylogenetic relation-ships within the suborder Tricladida (Platyhelminthes;Sluys 1989). Our results on G. tigrina add informationabout muscle organization in the Platyhelminthes, an es-sential point if we consider that Turbellarian vermiformorganization is supposed to play a key role in ancestralBilateria (Clark 1964; Ax 1995).
Myocytes in intact organisms
As pointed out in the introduction, planarians show agreat morphological plasticity enabling them to grow anddegrow continuously depending on external conditions.
According to Romero (1987) and Romero and Baguñà(1988), these processes depend on the daily balance be-tween cell proliferation and cell death. Myocytes found inintact organisms would be necessary for maintaining sucha balance as well as for cell renewal. We can not discard,however, the idea that not all these myocytes will be usedin cell renewal. After any injury suffered by the organ-isms that would trigger the mechanisms implicated in re-generation, some of these myocytes (the ones placedcloser to the wound) could be quickly mobilized and usedin the regenerative process. In the body wall musculatureof intact organisms, some thin fibers are usually founddispersed among the more numerous and thicker fiberscharacteristic of each muscle layer (Fig. 2G–H). It istempting to speculate that these thin fibers are nascent fi-bers that with time will replace the old ones.
Myocyte differentiation and blastema formation
Two opposing theories have been used to explain the ori-gin of blastema cells in planarians: the neoblast (Wolffand Dubois 1948) and dedifferentiation theories(Flickinger 1964; Hay 1966; Chandebois 1976; for astatement of the controversy, see Slack 1980; Baguñà1981; for discussion, see Gremigni 1988; Baguñà et al.1989, 1990). A present model (based upon Saló andBaguñà 1984 and Baguñà et al. 1989) assumes that, after
312

313
Fig. 5 Amino acid similarity of Girardia tigrina myosin heavychain (MHC) with MHC proteins from other species: Platyhelmin-thes (Schistosoma mansoni, Weston et al. 1993); mollusc (Aequi-pecten irradians, Nyitray et al. 1990); insect (Drosophila melano-gaster, George et al. 1989); nematode [Caenorhabditis elegans
(unc-54), Karn et al. 1983]; chordate [Homo sapiens(embryonic),Bober et al. 1990]. Identical amino acids are indicated by horizontallines. Conserved substitutions are indicated in bold letter. (The se-quence of Gtmhcwill appear in the EMBL, DDBJ and GenBankNucleotide Sequence Databases with the accession number U91585)

314
cutting, there is an increased mitotic activity in the post-blastema region, which is considered a proliferative re-gion of neoblasts. Neoblasts would enter into the blaste-ma as undifferentiated cells and the number of cells pro-duced by mitosis within the postblastema is sufficient toexplain the growth of the blastema without requiring oth-er mechanisms, such as dedifferentiation or long-rangecell migration (Saló and Baguñà 1989). Moreover, Pe-dersen (1972) and Morita and Best (1984) did not findany evidence of de- and redifferentiation of pre-existingmuscle cells during regeneration.
Muscle cell differentiation during regeneration hasbeen studied mainly using morphological characters.Sauzin (1967), Pedersen (1972) and Morita and Best(1984) concluded that newly differentiating muscle cellswere first discernible within the blastema on the thirdday. Hori (1983) showed that, at the wound center, undif-ferentiated cells were rapidly replaced by regenerativecells including myocytes during the first 2 days after am-putation. Our results with the monoclonal antibodyTMUS-13 show that, at least for muscle cells, the post-blastema region is also a differentiation area as we havefound myocytes at different stages of differentiation atday 1 of regeneration before the blastema is visible. Theorigin of these myocytes is likely correlated with the firstmitotic peak detected within the postblastema region at4–12 h after cutting, and derived from a population ofG2 neoblasts that exists in intact organisms (Saló andBaguñà 1984). As regeneration proceeds these postblas-tema myocytes can follow two main pathways: (1) enter-ing into the blastema to form the new muscle fibers, or(2) finishing differentiation in the postblastema regionand intercalating into old fibers. Both mechanisms couldbe acting simultaneously. If myocytes enter the blastema,it would mean that the blastema grows not only by an ac-cumulation of undifferentiated cells (neoblasts) but alsoa mixture of neoblasts at different stages of differentia-tion towards different cell types necessary to restore themissing pattern. If some myocytes intercalate into old fi-bers within the postblastema the pattern of these old fi-bers would become reorganized (morphallaxis), althoughthe myocytes would come from proliferating neoblasts inaccordance with an epimorphic model. This dual mor-phallactic-epimorphic model has also been describedduring pharynx regeneration (Bueno et al. 1997b).
Another interesting point from our results concerns therole of the old fibers that close the wound. As we havedescribed, initial blastemal growth could be responsiblefor the relaxation and disorganization of the muscle fibersthat contract the body wall, observed in the first days ofregeneration. Pedersen (1972) pointed out this loosenessin structural organization, a feature that in his opinion isprobably advantageous during a period where importantcell migrations are taking place. Although from our re-sults we cannot distinguish between old and new musclefibers within the blastema and even if many of the old fi-bers are phagocytized (Morita and Best 1984), our resultsshow some muscle fibers coming from the postblastemaand entering into the blastema (Fig. 4C, D, arrowheads).
These muscle fibers might be interpreted as old fibers re-maining within the blastema. It is possible that the oldmuscle fibers (placed outside or within the blastema) playa role as a guidance or substrate for the assembly of thenewly differentiated muscle cells. Although few data con-cerning muscle patterning during regeneration are avail-able, this possible role for the old fibers in the patterningof new muscle fibers would be consistent with the embry-ological data from other organisms. For example, in Ho-ploplana inquilina(Platyhelminthes) Reiter et al. (1996)suggest that the first longitudinal and circular muscle fi-bers that appear may guide the orientation of further de-veloping fibers. In grasshopers (Ho et al. 1983), however,muscle patterning during embryogenesis requires the ini-tial presence of muscle pioneer cells with which othercells fuse to form the muscles. More data are needed tocorrelate the muscle patterning within the blastema of G.tigrina with other factors such as epithelium-mesen-chyme interactions and/or the influence of the nervoussystem. For example, in Macrostomum hystricinum mari-num (Platyhelminthes; Reiter et al 1996) migration anddifferentiation of myocytes of circular fibers appears tobe spatially restricted to the nerve cords; in Drosophilathe muscle precursors appear exclusively among cells indirect contact with either the epidermis or the central ner-vous system (Bate 1990).
Myosin heavy-chain gene
The myosin heavy-chain from G. tigrina shows a highdegree of sequence similarity with the mhc genes fromSchistosoma mansoni, a parasitic Platyhelminthes, andother species from Caenorhabditis elegansto humans.Figure 5 shows the comparison between G. tigrina MHCand other MHC proteins from different species. The par-tial MHC we have cloned corresponds to the α-helicalcoiled region (Warrick and Spudich 1987; Kiehart 1990,for a general review on myosins). The future analysis ofthe whole amino acid sequence of the G.tigrina MHCwill show at the molecular level if the ambiguous charac-teristics of planarian muscle, between smooth (by physio-logical studies; Sarnat 1984), and striated muscle (by ul-trastructural studies; MacRae 1963, 1965; Moraczewskiand Czubaj 1974; Morita 1965; Pedersen 1964), persist.
&p.2:Acknowledgements We would like to give special thanks to Sal-vador Carranza for his technical advice and Susanna Castel fromServeis Científico-Tècnics (Univ. of Barcelona) for her expert as-sistance with the confocal microscopy. We would also like tothank Estela Castillo and Emili Saló for supplying cDNA and ge-nomic libraries, respectively, and José Ramón Bayascas and NeusCols for their advice and suggestions. In addition, we acknowl-edge the two anonymous referees, whose constructive commentsand suggestions greatly improved this manuscript. F. C. has a pre-doctoral fellowship (FPI) from Ministerio de Educación y Cienciaand M. V. has a predoctoral fellowship (AECI-MUTIS) from Mi-nisterio de Asuntos Exteriores, Spain. P. N. was supported by theCancer Research Fund of the Damon Runyon-Walter WinchellFoundation, DRG-1322. This work was supported by a grantDGICYT nº PB-93-0074 of the Ministerio de Educación y Ciencia(Spain) to R.R..

315
References
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Ba-sic local alignment search tool. J Mol Biol 215: 403–410
Auladell C (1990) Anàlisi de l’heterogeneïtat estructural i funcio-nal de les cèllules indiferenciades (neoblasts) de la planàriaDugesia (G) tigrina. PhD Thesis, Universitat de Barcelona
Ax P (1987) The phylogenetic system. John Wiley & Sons, ChichesterAx P (1995) In Das System der metazoa I. Ein Lehrbuch der
Phylogenetischen Systematik. Fischer, StuttgartBaguñà J (1973) Estudios citotaxonómicos, ecológicos e hist-
ofisiologìa de la regulación morfogenética durante el crecimi-ento y la regeneración en la raza asexuada de la planaria Dug-esia mediterranea. PhD Thesis, Universitat de Barcelona
Baguñà J (1976) Mitosis in the intact and regenerating planarianDugesia mediterranea n. sp. II. Mitotic studies during regener-ation and a possible mechanism of blastema formation. J ExpZool 195: 65–80
Baguñà J (1981) Planarian neoblasts. Nature 290: 14–15Baguñà J, Romero R (1981) Quantitative analysis of cell types dur-
ing growth, degrowth and regeneration in the planarians Dugesiamediterraneaand Dugesia tigrina. Hydrobiologia 84: 181–194
Baguñà J, Saló E, Auladell C (1989) Regeneration and pattern forma-tion in planarians. III. Evidence that neoblasts are totipotent stemcells and the source of blastema cells. Development 107: 77–86
Baguñà J, Romero R, Saló E, Collet J, Auladell C, Ribas M, Riu-tort M, Garcia- Fernàndez J, Burgaya F, Bueno D (1990)Growth, degrowth and regeneration as developmental phenom-ena in adult freshwater planarians. In: Marthy, HJ (ed) Experi-mental embryology of aquatic plants and animals. (NATO-ASIseries) Plenum Press, New York, pp 129–162
Baguñà J, Saló E, Romero R, Garcia-Fernàndez J, Bueno D,Muñoz-Marmol AM, Bayascas-Ramìrez JR, Casali A (1994)Regeneration and pattern formation in planarians: cells, mole-cules and genes. Zool Sci 11: 781–795
Ball IR (1974) A contribution to the phylogeny and biogeographyof the freshwater triclads (Platyhelminthes, Turbellaria). In:Riser NW, Morse HP (ed) Biology of the Turbellarians. Mc-Graw-Hill, New York, pp 339–401
Bate M (1990) The embryonic development of larval muscles inDrosophila. Development 110: 791–804
Bober E, Buchberger-Seidl A, Braun T, Singh S, Goedde HW, Ar-nold HH (1990) Identification of three developmentally con-trolled isoforms of human myosin heavy chains. Eur J Bio-chem 189: 55–65
Brøndsted HV (1969) Planarian regeneration. Pergamon Press,Oxford London
Brusca RC, Brusca GJ (1990) Invertebrates. Sinauer, SunderlandMassachusetts
Bueno D, Baguñà J, Romero R (1997a) Cell-, tissue-, and posi-tion-specific monoclonal antibodies against the planarian Dug-esia(Girardia) tigrina. Histochem Cell Biol 107: 139–149
Bueno D, Espinosa L, Baguñà J, Romero R (1997b) Planarianpharynx regeneration in regenerating tail fragments monitoredwith cell-specific monoclonal antibodies. Dev Genes Evol206: 425–434
Chandebois R (1976) Histogenesis and morphogenesis in planari-an regeneration. Karger, Basel
Chandebois R (1980) The dynamics of wound closure and its rolein the programming of planarian regeneration. II. Distalizat-ion. Dev Growth Differ 22: 693–704
Clark RB (1964) Dynamics in metazoan evolution. The origin ofthe coelom and segments. Clarendon Press, Oxford
Crezée M (1975) Monograph of the Solenofilomorphidae (Turbel-laria: Acoela). Int Rev Ges Hydrobiol 60: 769–845
De Vries ED, Sluys R (1991) Phylogenetic relationships of the ge-nus Dugesia(Platyhelminthes, Tricladida, Paludicola). J ZoolLondon 223: 103–116
Espinosa Ll (1993) Anàlisi espaial i temporal de la regeneració dela faringe i cavitat faríngia de la planària Dugesia(G)tigrinamitjançant anticossos monoclonals. Graduate Research Pro-ject, Universitat de Barcelona
Flickinger RA (1964) Isotopic evidence for a local origin of blas-tema cells in regenerating planaria. Exp Cell Res 34: 403–406
Garcia-Fernàndez J, Baguñà J, Saló E (1993) Genomic organiza-tion and expression of the planarian homeobox genes Dth-1and Dth-2. Development 118: 241–253
George EL, Ober MB, Emerson CP Jr (1989) Functional domainsof the Drosophila melanogastermuscle myosin heavy-chaingene are encoded by alternatively spiced exons. Mol Cell Biol9: 2957–2974
Girard C (1850) A brief account of the freshwater planarians ofthe United State. Proc Boston Soc Nat Hist 3:264–265
Gremigni V (1988) Planarian regeneration: An overview of somecellular mechanisms. Zool Sci 5: 1153–1163
Hanahan D (1983) Studies on transformation of Eschirichia coliwith plasmids. J Mol Biol 166: 557–580
Hay ED (1966) Regeneration. Holt, Rinehart and Winston, NewYork
Hay ED, Coward SJ (1975) Fine structure studies on the planarianDugesia. I. Nature of the “neoblast” and other cell types in non-injured worms. J Ultrastruct Res 50: 1–21
Ho RK, Ball EE, Goodman CS (1983) Muscle pioneers: large me-sodermal cells that erect a scaffold for developing muscles andmotoneurons in grasshopper embryos. Nature 301: 66–69
Hori I (1983) Differentiation of myocytes in the regenerating pla-narian Dugesia japonica. Cell Differ 12: 155–163
Karn J, Brenner S, Barnett LD (1983) Protein structural domainsin the Caenorhabditis elegansunc-54 myosin heavy chaingene are not separated by introns. Proc Natl Acad Sci USA 80:4253–4257
Kiehart DP (1990) Molecular genetic dissection of myosin heavychain function. Cell 60: 347–350
Laemmli UK (1970) Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227: 680–685
MacRae EK (1963) Observations on the fine structure of pharyn-geal muscle in the planarian Dugesia tigrina. J Cell Biol 18:651–662
MacRae EK (1965) The fine structure of muscle in a marine tur-bellarian. Z Zellforsch Mikrosk Anat 68: 348–362
Moraczewski J (1981) The fine structure of some catenulida (Tur-bellaria, Archoophora). Zool Pol 28: 367–415
Moraczewski J, Czubaj A (1974). Muscle ultrastructure in Cat-enula lemnae(Dug) (Turbellaria, Acheaeophora). J Submi-crosc Cytol 6: 29–38
Morita M (1965) Electron microscopic studies on planarian. I.Fine structures of muscle fiber in the head of the planarianDugesia dorotocephala. J Ultrastruct Res 13: 383–395
Morita M, Best JB (1984) Electron microscopic studies of planari-an regeneration. III. Degeneration and differentiation of mus-cles. J Exp Zool 229: 413–424
Morita M, Best JB, Noel J (1969) Electron microscopy studies ofplanarian regeneration. I. Fine structure of neoblasts in Dug-esia dorotocephala. J Ultrastruct Res 27: 7–23
Nyitray L, Goodwin EB, Szent-Gyorgyi AG (1990) Nucleotide se-quence of full length cDNA for a scallop striated muscle myo-sin heavy chain. Nucleic Acids Res 18: 7158
Pedersen KJ (1964) The cellular organization of Convoluta con-voluta, an acoel turbellarian: a cytological, histochemical, andfine structural study. Z Zellforsch Mikrosk Anat 64: 655–687
Pedersen KJ (1972) Studies on regeneration blastemas of the pla-narian Dugesia tigrinawith special reference to differentiationof the muscle-connective tissue filament system. WilhelmRoux’ Arch Entwicklungsmech Org 169:134–169
Prudhoe S (1985) A monograph on polyclad Turbellaria. OxfordUniversity Press, Oxford New York
Reiter D, Boyer B, Ladurner P, Mair G, Salvenmoser W, Rieger R(1996) Differentiation of the body wall musculature in Mac-rostomum hystricinum marinumand Hoploplana inquilina(Plathelminthes), as models for muscle development in lowerSpiralia. Roux’s Arch Dev Biol 205: 410–423
Ribas M, Riutort M, Baguñà J (1989) Morphological and bio-chemical variation in populations of Dugesia (G) tigrina(Tur-bellaria, Tricladida, Paludicola) from the western mediterra-

Saló E, Baguñà J (1984) Regeneration and pattern formation inplanarians I. The pattern of mitosis in anterior and posteriorregeneration in Dugesia (G) tigrina, and a new proposal forblastema formation. J Embryol Exp Morphol 83: 63–80
Saló E, Baguñà J (1989) Regeneration and pattern formation inplanarians. II. Local origin and role of cell movements in blas-tema formation. Development 107: 69–76
Sarnat HB (1984) Muscle histochemistry of the planarian Dugesiatigrina (Turbellaria: Tricladida): Implications in the evolutionof muscle. Trans Am Microsc Soc 103: 284–294
Sauzin MJ (1967) Ëtude ultrastructurale de la différentiation aucours de la régénération de la planaire Dugesia gonocephala. II.Différenciation musculaire. Bull Soc Zool Fr 92: 613–619
Slack JMW (1980) A serial threshold theory of regeneration. JTheor Biol 82: 105–140
Sluys R (1989) Phylogenetic relationships of the triclads (Platy-helminthes, Seriata, Tricladida). Bijdr Dierk 59: 3–25
Warrick HM, Spudich JA (1987) Myosin structure and function incell motility. Annu Rev Biochem 56: 695–726
Weston D, Schmitz J, Kemp WM, Kunz W (1993) Cloning and se-quencing of a complete myosin heavy chain cDNA fromSchistosoma mansoni. Mol Biochem Parasitol 58: 161–164
Wolff E, Dubois F (1948) Sur la migration des cellules deregénération chez les planaires. Rev Suisse Zool 55: 218–227
Wulf E, Deboben A, Bautz FA, Faulstich H, Wieland T (1979)Fluorescent phallotoxin, a tool for the visualization of cellularactin. Proc Natl Acad Sci USA 9: 4498–4502
316
nean: Biogeographical and taxonomical implications. J ZoolLondon 218: 609–626
Rieger RM, Tyler S, Smith JP III, Rieger G (1991a) Turbellaria.In: Harrison FW, Bogitsh BJ (ed) Microscopic anatomy of in-vertebrates, vol 3. Wiley Liss, New York, pp 7–140
Rieger R, Salvenmoser W, Legniti A, Reindl S, Adam H, Simons-berger P, Tyler S (1991b) Organization and differentiation ofthe body-wall musculature in Macrostomum (Turbellaria,Macrostomidae). Hydrobiologia 227: 119–129
Rieger RM, Salvenmoser W, Legniti A, Tyler S (1994) Phalloidin-rhodamine preparations of Macrostomum hystricinum mari-num (Plathelminthes): morphology and postembryonic devel-opment of the musculature. Zoomorphology 114: 133–147
Romero R (1987) Anàlisi cellular quantitativa del creixement i dela reproducció a diferents espècies de planàries. PhD Thesis,Universitat de Barcelona
Romero R, Baguñà J (1988) Quantitative cellular analysis of life-cycle strategies of iteroparous and semelparous triclads. Forts-chr Zool 36: 283–289
Romero R, Baguñà J (1991) Quantitative cellular analysis ofgrowth and reproduction in freshwater planarians (Turbellaria;Tricladida). I. A cellular description of the intact organism. In-vert Reprod Dev 19: 157–165
Romero R, Fibla J, Bueno D, Sumoy L, Soriano MA, Baguñà J(1991) Monoclonal antibodies as markers of specific cell typesand regional antigens in the freshwater planarian Dug-esia(G)tigrina. Hydrobiologia 227: 73–79




















