
Mutagenicity and DNA adduct formation of PAH, nitro-PAH, and oxy-PAH fractions of atmospheric...
9
Please cite this article in press as: G.A. Umbuzeiro, et al., Mutagenicity and DNA adduct formation of PAH, nitro-PAH, and oxy- PAH fractions of atmospheric particulate matter from S˜ ao Paulo, Brazil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008), doi:10.1016/j.mrgentox.2007.12.007 ARTICLE IN PRESS +Model MUTGEN-401418; No. of Pages 9 Available online at www.sciencedirect.com Mutation Research xxx (2008) xxx–xxx Mutagenicity and DNA adduct formation of PAH, nitro-PAH, and oxy-PAH fractions of atmospheric particulate matter from S˜ ao Paulo, Brazil Gisela A. Umbuzeiro a,∗ , Alexandre Franco b , Maria Helena Martins a , F´ abio Kummrow a,c , Lilian Carvalho b , Heinz H. Schmeiser d , Jutta Leykauf d , Marie Stiborova e , Larry D. Claxton f a CETESB, Cia de Tecnologia de Saneamento Ambiental, Av. Prof. Frederico Hermann Jr. 345, 05459-900 S˜ ao Paulo, Brazil b Instituto de Qu´ ımica da Universidade de S ˜ ao Paulo, Av. Prof. Lineu Prestes 748, 05599-970 S˜ ao Paulo, Brazil c Universidade Federal de Alfenas, R. Gabriel Monteiro da Silva 714, 37130-000 Alfenas, Brazil d Division of Molecular Toxicology, German Cancer Research Center, Im Neuenheimer Feld 280, 69120 Heidelberg, Germany e Department of Biochemistry, Faculty of Science, Charles University, Albertov 2030, 128 40 Prague 2, Czech Republic f Environmental Carcinogenesis Division, US Environmental Protection Agency, B143-06, Research Triangle Park, NC 27711, USA Received 15 August 2007; received in revised form 14 December 2007; accepted 21 December 2007 Abstract Urban particulate matter (UPM) contributes to lung cancer incidence. Here, we have studied the mutagenic activity and DNA adduct-forming ability of fractionated UPM extractable organic matter (EOM). UPM was collected with a high-volume sampler in June 2004 at two sites, one at street level adjacent to a roadway and the other inside a park within the urban area of the city of S˜ ao Paulo, Brazil. UPM was extracted using dichloromethane, and the resulting EOM was separated by HPLC to obtain PAH, nitro-PAH, and oxy-PAH fractions which were tested for mutagenicity with the Salmonella strains TA98 and YG1041 with and without S9 metabolic activation. The PAH fraction from both sites showed negligible mutagenic activity in both strains. The highest mutagenic activity was found for the nitro-PAH fraction using YG1041 without metabolic activation; however, results were comparable for both sites. The nitro-PAH and oxy-PAH fractions were incubated with calf thymus DNA under reductive conditions appropriate for the activation of nitro aromatic compounds, then DNA adduct patterns and levels were determined with thin-layer chromatography (TLC) 32 P-postlabeling method using two enrichment procedures—nuclease P1 digestion and butanol extraction. Reductively activated fractions from both sites produced diagonal radioactive zones (DRZ) of putative aromatic DNA adducts on thin layer plates with both enrichment procedures. No such DRZ were observed in control experiments using fractions from unexposed filters or from incubations without activating system. Total adduct levels produced by the nitro-PAH fractions were similar for both sites ranging from 30 to 45 adducts per 10 8 normal nucleotides. In contrast, the DNA binding of reductively activated oxy-PAH fractions was three times higher and the adduct pattern consisted of multiple discrete spots along the diagonal line on the thin layer plates. However, DNA adduct levels were not significantly different between the sampling sites. Both samples presented the same levels of mutagenic activity. The response in the Salmonella assay was typical of nitroaromatics. Although, more mutagenic activity was related to the nitro-PAH fraction in the Salmonella assay, the oxy-PAH fractions showed the highest DNA adduct levels. More studies are needed to elucidate the nature of the genotoxicants occurring in S˜ ao Paulo atmospheric samples. © 2008 Elsevier B.V. All rights reserved. Keywords: Air mutagenicity; Nitroarenes; Air pollution; DNA adducts; Nitroaromatics; Salmonella microsome assay; Ames test; YG1041; TA98 1. Introduction Ambient air genotoxicants can originate from emission of fuel combustion (motor vehicle exhaust, space heating, and power generation), waste incineration and industrial processes, ∗ Corresponding author. Tel.: +55 11 31333531; fax: +55 11 31333982. E-mail address: [email protected] (G.A. Umbuzeiro). and they are also formed by atmospheric reactions [1–3]. Thus, it is important to determine the sources of ambient genotox- icants and the levels to which populations may be exposed [4]. The Salmonella/microsome assay has been used for large, multi-site, and/or time series studies, for bioassay-directed fractionation studies, for identifying the presence of specific mutagens and classes of mutagens, and for doing site- or source-comparisons for relative levels of airborne mutagens [1]. 1383-5718/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.mrgentox.2007.12.007
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Mutagenicity and DNA adduct formation of PAH, nitro-PAH, and oxy-PAH fractions of atmospheric...

M
A
aaufsmDwRww1cbnt©
K
1
fp
1d
ARTICLE IN PRESS+ModelUTGEN-401418; No. of Pages 9
Available online at www.sciencedirect.com
Mutation Research xxx (2008) xxx–xxx
Mutagenicity and DNA adduct formation of PAH, nitro-PAH, and oxy-PAHfractions of atmospheric particulate matter from Sao Paulo, Brazil
Gisela A. Umbuzeiro a,∗, Alexandre Franco b, Maria Helena Martins a, Fabio Kummrow a,c,Lilian Carvalho b, Heinz H. Schmeiser d, Jutta Leykauf d, Marie Stiborova e, Larry D. Claxton f
a CETESB, Cia de Tecnologia de Saneamento Ambiental, Av. Prof. Frederico Hermann Jr. 345, 05459-900 Sao Paulo, Brazilb Instituto de Quımica da Universidade de Sao Paulo, Av. Prof. Lineu Prestes 748, 05599-970 Sao Paulo, Brazil
c Universidade Federal de Alfenas, R. Gabriel Monteiro da Silva 714, 37130-000 Alfenas, Brazild Division of Molecular Toxicology, German Cancer Research Center, Im Neuenheimer Feld 280, 69120 Heidelberg, Germany
e Department of Biochemistry, Faculty of Science, Charles University, Albertov 2030, 128 40 Prague 2, Czech Republicf Environmental Carcinogenesis Division, US Environmental Protection Agency, B143-06, Research Triangle Park, NC 27711, USA
Received 15 August 2007; received in revised form 14 December 2007; accepted 21 December 2007
bstract
Urban particulate matter (UPM) contributes to lung cancer incidence. Here, we have studied the mutagenic activity and DNA adduct-formingbility of fractionated UPM extractable organic matter (EOM). UPM was collected with a high-volume sampler in June 2004 at two sites, onet street level adjacent to a roadway and the other inside a park within the urban area of the city of Sao Paulo, Brazil. UPM was extractedsing dichloromethane, and the resulting EOM was separated by HPLC to obtain PAH, nitro-PAH, and oxy-PAH fractions which were testedor mutagenicity with the Salmonella strains TA98 and YG1041 with and without S9 metabolic activation. The PAH fraction from both siteshowed negligible mutagenic activity in both strains. The highest mutagenic activity was found for the nitro-PAH fraction using YG1041 withoutetabolic activation; however, results were comparable for both sites. The nitro-PAH and oxy-PAH fractions were incubated with calf thymusNA under reductive conditions appropriate for the activation of nitro aromatic compounds, then DNA adduct patterns and levels were determinedith thin-layer chromatography (TLC) 32P-postlabeling method using two enrichment procedures—nuclease P1 digestion and butanol extraction.eductively activated fractions from both sites produced diagonal radioactive zones (DRZ) of putative aromatic DNA adducts on thin layer platesith both enrichment procedures. No such DRZ were observed in control experiments using fractions from unexposed filters or from incubationsithout activating system. Total adduct levels produced by the nitro-PAH fractions were similar for both sites ranging from 30 to 45 adducts per08 normal nucleotides. In contrast, the DNA binding of reductively activated oxy-PAH fractions was three times higher and the adduct pattern
onsisted of multiple discrete spots along the diagonal line on the thin layer plates. However, DNA adduct levels were not significantly differentetween the sampling sites. Both samples presented the same levels of mutagenic activity. The response in the Salmonella assay was typical ofitroaromatics. Although, more mutagenic activity was related to the nitro-PAH fraction in the Salmonella assay, the oxy-PAH fractions showedhe highest DNA adduct levels. More studies are needed to elucidate the nature of the genotoxicants occurring in Sao Paulo atmospheric samples.2008 Elsevier B.V. All rights reserved.
omat
ai
eywords: Air mutagenicity; Nitroarenes; Air pollution; DNA adducts; Nitroar
. Introduction
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
Ambient air genotoxicants can originate from emission ofuel combustion (motor vehicle exhaust, space heating, andower generation), waste incineration and industrial processes,
∗ Corresponding author. Tel.: +55 11 31333531; fax: +55 11 31333982.E-mail address: [email protected] (G.A. Umbuzeiro).
i[mfms[
383-5718/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.mrgentox.2007.12.007
ics; Salmonella microsome assay; Ames test; YG1041; TA98
nd they are also formed by atmospheric reactions [1–3]. Thus,t is important to determine the sources of ambient genotox-cants and the levels to which populations may be exposed4]. The Salmonella/microsome assay has been used for large,ulti-site, and/or time series studies, for bioassay-directed
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
ractionation studies, for identifying the presence of specificutagens and classes of mutagens, and for doing site- or
ource-comparisons for relative levels of airborne mutagens1].

IN+ModelM
2 tion R
aipciiSoPfwa
mmihma9t1g
haamdpolaPrltha
og[N[bo
tgap2dmtnbtobpoidtd
afwmnvwt3
2
2
amCTS
2
ua
TC
S
CI
ARTICLEUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Muta
Early research recognized that although carcinogenic PAHre present in air samples they could not account for the major-ty of mutagenic activity detected. The mutagenicity of airbornearticulate is due to ∼500 identified compounds from varyinghemical classes [1]. Franco [5] showed that the mutagenic activ-ty of total extracts of urban particulate matter (UPM) collectedn two different areas of Sao Paulo state was highest in thealmonella/microsome microsuspension assay in the absencef S9 for both TA98 and YG1041. The nitro-PAH and oxy-AH fractions obtained by HPLC separation were responsibleor the majority of the mutagenicity observed with both strains;hereas, the PAH-fraction gave only low levels of mutagenic
ctivity [5].Sao Paulo metropolitan area (SPMA) is a region with about 17
illion people, 2000 major industrial sites, and more than sevenillion vehicles. The 2004 estimates for levels of air pollutants
ncluded: 1.7 million metric tons/year (t/y) CO, 404 thousand t/yydrocarbons, 371 thousand t/y NOx, 63 thousand t/y particulateatter, and 38 t/y SO2 [6]. In 2004, vehicles were responsible for
bout 98% of the total emissions of CO, 97% of hydrocarbons,6% of NOx, and 55% of SO2 [6]. About 6% of the vehicles inhe SPMA used diesel fuel, vehicles using ethanol represented4.5% of the fleet and 83% used gasohol (a mixture of 78%asoline and 22% ethanol).
A complex variety of genotoxicants in ambient air pollutionave been detected [1,7], and high exposures are associated withn increased risk of cancer [8–10]. Amongst these genotoxicantsre nitro-polycyclic aromatic hydrocarbons (nitro-PAH) whichay be present on particulate matter from direct sources, such as
iesel and gasoline exhaust [11] and may be produced from gas-hase reactions of polycyclic aromatic hydrocarbons (PAH) withxides of nitrogen, with subsequent partitioning to the particu-ate phase [12]. Both their long persistence in the environmentnd the mutagenic and carcinogenic properties of certain nitro-AH has led to considerable interest in assessing their potentialisk to humans [11,13–15]. Although, environmental levels areower than unsubstituted PAH [14], it has become clear thathis is compensated for by certain nitro-PAH exhibiting muchigher direct-acting mutagenic potency in bacterial bioassaysnd in forward mutation assays based on mammalian cells [16].
The aromatic nitro-ketone 3-nitrobenzanthrone (3-NBA) isne of the most potent mutagens and a potential human carcino-en identified in diesel exhaust and ambient air particulate matter17–19]. As a likely consequence of atmospheric washout, 3-
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
BA has also been detected in rain water and surface soil18]. The uptake of 3-NBA in humans has been demonstratedy the detection of 3-aminobenzanthrone, a major metabolitef 3-NBA, in the urine of workers occupationally exposed
cfB(a
able 1ollection sites, dates, volume, mass, extractable organic matter of the samples analy
ite Initial date/time Final date/time Volu(m3
erqueira Cesar June, 17th 2004 11:00 am June 18th 2004 11:25 am 189birapuera park June 17th 2004 12:00 am June 18th 2004 12:30 am 172
PRESSesearch xxx (2008) xxx–xxx
o diesel emissions. There is clear evidence that 3-NBA is aenotoxic mutagen that forms DNA adducts after metabolicctivation through reduction of the nitro group [20–24]. Theredominant DNA adducts detected in vivo in rodents are-(2′-deoxyguanosin-N2-yl)-3-aminobenzanthrone and N-(2′-eoxyguanosin-8-yl)-3-aminobenzanthrone [22], and they areost probably responsible for the GC to TA transversion muta-
ions induced by 3-NBA in vivo [25]. These DNA adductsot only represent premutagenic lesions in DNA but may alsoe of primary importance for tumour development in targetissue [26,27]. DNA adduct formation in rats treated eitherrally, intraperitoneally, or intratracheally with 3-NBA haseen demonstrated using thin-layer chromatography (TLC) 32P-ostlabeling. In these studies the same adduct pattern consistingf multiple 3-NBA-specific DNA adducts was detected in var-ous organs of rats. All of them were primarily located in aiagonal zone on the TLC plates at approximately 45◦ to elu-ion directions D3 and D4 [20,22,28], typical of DNA adductserived from extracts of UPM and diesel exhaust [29,30].
The objective of this work was to compare the mutagenicctivity of the PAH, nitro-PAH and oxy-PAH fractions obtainedrom two UPM samples collected in Sao Paulo city in areasith different traffic conditions using the Salmonella/microsomeicrosuspension assay. Also, DNA adduct formation of the
itro-PAH and oxy-PAH fractions after reductive activation initro was examined using the 32P-postlabeling assay. Moreover,e investigated whether DNA adducts formed by the geno-
oxic environmental pollutant 3-NBA were detectable on the2P-postlabeling maps produced by the nitro-PAH fractions.
. Material and methods
.1. Sampling sites
The samples were collected at two sites: the Cerqueira Cesar street stationnd the Ibirapuera park station (Table 1). The former site is located along aajor avenue (Av. Dr. Arnaldo at the Campus of Faculdade de Saude Publica).ar and bus traffic at this site is heavy during the morning and late afternoon.he Ibirapuera park monitoring site is located at a city park in the middle of theMPA. There was no traffic within 0.3 km.
.2. Urban particulate matter collection
The urban particulate matter (UPM) was collected with Hi-Vol samplerssing quartz filters for 24 h.The samples were collected during winter time, withverage temperature of 16 ◦C (min = 11 ◦C; max = 23 ◦C) and the meteorological
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
onditions were not favorable for the pollutants dispersion on June 17th butavorable on June 18th of 2004. For other details of the samples see Table 1.efore and after sampling, the filters were maintained at controlled temperature
from 20 to 25 ◦C) and humidity (around 40%) in order to remove moisture;nd then they were weighed after 24 h. The filters were stored for 10 days as
zed
me)
Mass(mg)
Urban particulate material(UPM) concentration (�g/m3)
Extractable organicmatter (EOM)(mg/total UPM)
1.86 295.8 156 57.70.95 227.1 132 41.7

IN+ModelM
tion R
d2
2
cSiuw(
fawoT(
atD1Pfuwt
2
f
TM
D
U
P
N
O
*
ARTICLEUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Muta
escribed above and then extracted and fractionated. The tests were performed0 days after the collection date.
.3. Organic extraction and fractionation
The organic extraction of the filters was performed with 200 ml of methylenehloride (DCM) in a Soxhlet extractor at temperatures below 40 ◦C for 20 h. Theoxhlet reflux cycle time was approximately 15 min. The volume was reduced
n a rotary evaporator at 40 ◦C to 3 ml, and this was followed by evaporationnder a gentle stream of nitrogen gas until almost dryness. The residue obtainedas weighted for the calculation of the amount of extractable organic material
EOM), resuspended in 50 �L of DCM and fractionated.Only the PAH, nitro-PAH, and oxy-PAH fractions were isolated. From each
raction obtained, 75% was sent to the German cancer research center for DNA
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
dduct analysis and 25% was analyzed at CETESB using the Salmonella assay,ith the strains TA98 and YG1041 with and without S9. The strains werebtained from Dr. Larry D. Claxton, USEPA, Research Triangle Park, NC, USA.he fractionation was performed by HPLC with a silica normal phase column
type, ‘Nucleosil 100-10’; length, 250 mm; id., 4.6 mm; provided by Macherey
rtaop
able 2utagenic activity in the Salmonella/microsome microsuspension assay for the three
oses Mean of number of revertants/plate and standard devi
PM equiv/plate (�g) TA98
−S9 +S9
Mean S.D. Mean S.D.
AH fractionNegative control 22.6 5.68 23.6 4.510.5 17.0 2.83 20.0 2.831 20.0 1.41 23.5 2.125 20.5 6.36 18.5 0.7110 18.5 6.36 17.5 0.7150 21.0 0.00 21.0 4.24100 20.0 0.00 27.0 7.07Positive control 710a 995a
Potency rev/ug UPM NDb NDb
itro-PAH fractionNegative control 20.2 0.84 20.8 3.770.5 – – 18.5 0.711 – – 25.0 0.005 – – 21.0 5.6610 29.5 2.12** 17.0 7.0750 55.5 4.95** 19.5 0.71100 66.5 4.95** 25.5 0.71Blank filter 19.0 0.58 22.0 6.51Positive control 371a 800.5a
Potency rev/�g UPM 0.72 (0.63–0.80)c NDb
xy fractionNegative control 23.0 1.00 20.8 3.770.5 25.7 0.58* 20.0 1.411 25.7 2.08 24.5 10.615 25.3 1.53 18.5 4.9510 24.3 0.58 27.0 8.4950 – – 26.5 3.54100 54.7 11.37* 24.0 2.83Blank filter 21.0 8.54 18.3 2.08Positive control 371a 800.5a
Potency rev/�g UPM 0.30 (0.25–0.35)c NDb
: Significant at 5%; **: Significant at 1%.a For details see Section 2 (materials and methods).b ND: mutagenic activity not detected at the tested doses.c In parenthesis 90% confidence limits for the slope.
PRESSesearch xxx (2008) xxx–xxx 3
nd Nagel). The initial mobile phase (100% hexane) ran for ten minutes, andhen increasing quantities of DCM were added over a 40 min period until 100%CM was obtained as described by Liberti and Cicioli [31]. The flow rate wasmL per minute. The PAH fraction was collected from 7 to 17 min; the nitro-AH fraction was collected between 17 and 32 min and the oxy-PAH fractionrom 32 min to the end of the run. A blank filter was extracted and fractionatedsing the same procedure applied for the samples. A standard solution of 3-NBAas injected in parallel and the peak of the 3NBA (29 min) was found to be in
he nitro-PAH fraction.
.4. Salmonella/microsome assay
The PAH, nitro-PAH, and oxy-PAH fractions of the EOM were assayedor mutagenicity using the Salmonella typhimurium strains TA98 (his D3052,
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
fa, �bio, �uvr B, pKM101) [32] and YG1041 (a derivative of TA98, ableo produce high levels of nitroreductase and O-acetyltransferase) [33] withnd without S9 metabolic activation, in a microsuspension assay [34]. Thebtained fractions were dried under a gentle stream of pure nitrogen and appro-riate amount of dimethylsulfoxide (DMSO) was added just before testing.
fractions analyzed from Cerqueira Cesar station, Sao Paulo city, Brazil
ation (S.D.)
YG1041
−S9 +S9
Mean S.D. Mean S.D.
139.6 20.44 102.8 7.95117.0 14.14 98.0 7.07129.0 4.24 87.5 7.78118.0 11.31 93.5 10.61155.0 1.41 105.5 2.12181.0 1.41* 125.0 16.97223.0 12.73** 150.5 17.68*625a 1375a
0.93 (0.71–1.22)c 0.54 (0.39–0.70)c
95.4 3.65 85.6 6.88118.0 4.24** 75.5 0.71122.0 7.07* 78.0 1.41237.0 14.14** 89.5 13.44339.0 14.14** 103.5 10.61667.0 19.80** 221.5 10.61**709.0 39.60** 349.0 15.56**116.3 4.16 98.3 4.73665.5a 364a
25.3 (23.8–26.9)c 2.72 (2.49–2.94)c
95.4 3.65 85.6 6.8881.0 0.00 84.5 3.5496.0 7.07 81.0 2.83
137.5 12.02* 95.0 4.24185.5 6.36** 108.5 6.36*374.0 7.07** 108.5 6.36*556.5 101.12** 158.0 12.73**108.0 15.13 98.0 5.57665.5a 364a
6.0 (5.27–6.72)c 0.5 (0.31–0.70)c

IN+ModelM
4 tion R
Bewbf0sttmuANc0iO2p
sap
2c
tPdct
TM
D
U
P
N
O
*
ARTICLEUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Muta
ecause we were not able to weigh the EOM of each fraction, doses werexpressed in equivalent amounts of UPM. The EOM for both tested filtersas around 30% (Table 1) therefore direct comparisons of the results cane made. Overnight cultures of strains (around 109 cells/mL) concentrated 5-old by centrifugation (10,000 × g at 4 ◦C for 10 min) were resuspended into.015 M sodium phosphate buffer. 50 �L of cell suspension, 50 �L of 0.015 Modium phosphate buffer or S9 mix, and 5 �L of the sample were addedo a tube and incubated at 37 ◦C for 90 min without shaking. After incuba-ion, 2 mL of molten agar was added, and the mixture was poured onto a
inimal agar plate. Colonies were counted after 66 h of incubation at 37 ◦Csing an automatic colony counter. The metabolic activation was provided byrochlor 1254 induced Sprague Dawley rat liver S9 mix (MolTox, Boone,C), which was prepared at a concentration of 4% v/v. For TA98 the positive
ontrols were 0.125 �g/plate of 4-nitroquinoline-oxide (4NQO) (Acros), and
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
.625 �g/plate of 2-aminoanthracene (2AA) (Sigma–Aldrich), both dissolvedn DMSO. For YG1041 the positive controls were 10 �g/plate of 4-nitro--phenylenediamine (4NOP) (ICN Biomedicals Inc), and 0.0312 �g/plate ofAA. The doses tested varied from 0.5 to 100 �g of UPM equivalents perlate.
caPw(
able 3utagenic activity in the Salmonella/microsome microsuspension assay for the three
oses Mean number of revertants/plate and standard deviat
PM equiv/plate (�g) TA98
−S9 +S9
Mean S.D. Mean S.D.
AH fractionNegative control 22.6 5.68 23.6 4.51
0.5 – – 19.0 5.661 20.5 0.71 19.0 0.005 20.0 4.24 27.0 2.8310 24.5 2.12 22.5 3.5450 20.5 0.71 22.0 1.41100 22.5 2.12 19.5 2.12
Positive control 710a 995a
Potency rev/(�g) UPM NDb NDb
itro-PAH fractionNegative control 20.2 0.84 26.3 3.51
0.5 20.5 2.12 24.0 3.611 23.0 2.83 24.3 2.525 – – 22.7 0.5810 25.5 2.12* 25.5 0.7150 42.5 2.12** 36.7 8.74100 59.5 2.12** 39.0 11.14
Blank filter 19.0 0.58 22.0 6.51Positive control 371.0a 800.5a
Potency rev/(�g) UPM 0.40 (0.36–0.44)c 0.17 (0.10–0.23)c
xy-PAH fractionNegative control 20.2 0.84 20.8 3.77
0.5 18.5 0.71 18.0 1.411 16.0 4.24 21.0 1.415 20.5 2.12 21.5 3.5410 18.0 0.00 22.0 4.2450 29.5 0.71** 22.0 2.83100 35.5 2.12** 21.5 0.71Blank filter 21.0 8.54 18.3 2.08
Positive control 371a 800.5a
Potency rev/(�g) UPM 0.18 (0.14–0.21)c NDb
: Significant at 5%; **: Significant at 1%.a For details see Section 2 (materials and methods).b ND: mutagenic activity not detected at the tested doses.c In parenthesis 90% confidence limits for the slope.
PRESSesearch xxx (2008) xxx–xxx
The data were analyzed with the Salanal computer program using the Bern-tein model [35]. Samples were considered positive when a significant differencemong the tested doses and the negative control (ANOVA) and a significantositive dose response were observed.
.5. In vitro DNA adduct formation by activation with rat liverytosol
Cytosol from rat liver was prepared as described previously [24] and used ashe activating system for the in vitro incubations. The dried nitro-PAH and oxy-AH fractions from the HPLC separation were dissolved in 50 �l DMSO. Theeaerated and argon-purged incubation mixtures, in a final volume of 750 �L,onsisted of 50 mM Tris-HCl buffer (pH 7.4), containing 0.2% Tween 20, cofac-ors for cytosolic enzymes (1 mM NADPH; 2 mM acetyl-CoA), rat hepatic
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
ytosolic protein (1 mg), 10 �L of nitro-PAH or oxy-PAH fraction (in DMSO),nd 0.5 mg of calf thymus DNA. The reaction was initiated by adding nitro-AH or oxy-PAH fraction and carried out at 37 ◦C for 3 h. The same incubationsere performed with 3-NBA dissolved in DMSO at different concentrations
0.75 nM, 7.5 nM, 75 nM, 750 nM).
fractions analyzed from Ibirapuera park, Sao Paulo city, Brazil
ion (S.D.)
YG1041
−S9 +S9
Mean S.D. Mean S.D.
139.6 20.44 102.8 7.95112.0 24.04 104.5 3.54125.5 9.19 97.0 11.31128.0 8.49 101.5 2.12121.5 4.95 97.5 0.71139.0 1.41 105.0 4.24142.5 3.54 106.0 1.41625a 1375a
NDb NDb
95.4 3.65 85.6 6.88100.0 14.14 80.5 2.12115.0 8.49* 92.0 1.41218.0 8.49** 96.5 6.36345.5 3.54** 138.0 2.83*724.0 7.07** 285.0 45.25**844.0 22.63** 545.5 12.02**116.0 4.16 98.0 4.73665.5a 364a
25.1 (23.4–26.8)c 4.4 (4.1–4.7)c
95.4 3.65 85.6 6.88123.0 4.24** 114.5 17.68126.5 23.33 – –170.0 2.83** 130.5 3.54*179.5 0.71** 154.0 0.00293.0 2.83** 203.5 12.02**632.0 217.79* 316.0 22.63**108.0 15.13 98.0 5.57665.5a 364a
4.7 (4.0–5.4)c 6.8 (5.29–8.3)c

IN PRESS+ModelM
tion Research xxx (2008) xxx–xxx 5
aucwS
2
Pt(wfD2plm
3
3
wsvt
3
mdaCmtftsgfitttd(taer
3
P
Fig. 1. Mutagenic potency in number of revertants per �g of urban particulateMatter of each station with TA98 and YG1041 without and with S9 (Blackfor PAH, Grey for nitro-PAH and White for oxy-PAH fractions; 1 & 2 (TA98–S9 Cerqueira Cesar and Ibirapuera respectively), 3 & 4 (TA98 + S9 CerqueiraCIr
DtcaafutN[
NmptiNtt7cttrsu
dia(
ARTICLEUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Muta
After 3 h in the dark, reactions were stopped by the addition of phenol;nd DNA was isolated and determined as described [23] and stored at −20 ◦Cntil DNA adduct analysis. Control incubations were performed either withoutytosol, without nitro-PAH- or oxy-PAH-fractions, or 3-NBA. The dose unitsere the same as already described for the Salmonella/microsome assay (seeection 2.4)
.6. DNA adduct analysis
DNA adduct analysis was by 32P-postlabeling using enrichment by nuclease1 digestion and butanol extraction as previously described [36,37]. Separa-
ion of 32P-labeled adducts was carried out on PEI-cellulose thin layer platesMacherey and Nagel, Duren, Germany). Chromatographic conditions usedere: D1, 1.0 M sodium phosphate, pH 6.5; D2 was omitted; D3, 3.5 M Li
ormate, 8.5 M urea, pH 3.50; D4, 0.8 M LiCl, 0.5 M Tris, 8.5 M urea, pH 8.0;5, 1.7 M sodium phosphate, pH 6.0. Normal nucleotides were separated in80 mM (NH4)2SO4 and 50 mM NaH2PO4, pH 6.5. Quantitative analysis waserformed using a Canberra Packard instant imager. Adduct levels were calcu-ated in units of relative adduct labelling (RAL) which is the ratio of counts per
inute (cpm) of adducted nucleotides to cpm of total nucleotides in the assay.
. Results and discussion
.1. Site characteristics
Although the Ibirapuera park station was selected because itas anticipated to be less influenced by emissions from traffic
ources, the mass collected, the total suspended particles (TSP)alues, and the extractable organic mass (EOM) were similar forhe two sites (Table 1).
.2. Salmonella/microsome assay
Because the two filters had very similar levels of particulateatter, comparisons of the results of the bioassays can be made
irectly (Tables 2 and 3). The PAH fraction showed very lowctivity (<1.0 revertants/�g UPM) for YG1041 for the Cerqueiraesar station (Table 2), and the Ibirapuera sample showed noutagenic activity associated with its PAH fraction under the
ested conditions (Table 3). TA98 revealed no mutagenic activityor the PAH fractions from either sites. The nitro-PAH frac-ions and oxy-PAH fractions of both sampling stations showedimilar mutagenic activity (Fig. 1). The decrease of the muta-enicity with S9 was evident in both samples for the nitro-PAHraction (Fig. 1). This response and the fact that the mutagenic-ty increased with YG1041 in relation to TA98 are indicativehat the compounds that are causing the mutagenicity belong tohe nitroaromatics [1,33,38,39]. In contrast, the mutagenicity ofhe Ibirapuera oxy-PAH fraction sample with YG1041 did notecrease with the addition of S9, but showed a slight increaseFig. 1), indicating the presence of compounds that need S9o be activated, such as aromatic amines or others with similarctivity. If this change in mutagenicity is due to the increased lev-ls in ozone concentrations that generate new indirect mutagensemains to be determined, but this possibility cannot be ruled out.
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
.3. DNA adduct formation
Because mutagenic activity was predominantly in the nitro-AH and oxy-PAH fractions, these fractions were used to induce
d3
Nu
esar and Ibirapuera respectively), 5 & 6 (YG1041–S9 Cerqueira Cesar andbirapuera respectively) and 7 & 8 (YG1041 + S9 Cerqueira Cesar and Ibirapueraespectively).
NA adduct formation in in vitro incubations and analyzed byhe 32P-postlabeling method. It is known that nitro aromaticompounds identified in organic extracts of both diesel exhaustnd ambient air particulate matter exert their strong mutagenicctivity in the Salmonella mutagenicity assay [1,38–40]. There-ore, both fractions were activated after redutive conditionsnder anaerobic conditions using rat liver cytosol together withhe appropriate cofactors, a system that effectively activates 3-BA to the formation of DNA adducts as shown previously
23].As an initial step in establishing the limit of detection of 3-
BA as 3-NBA specific DNA adducts by the 32P-postlabelingethod, anaerobic incubations with calf thymus DNA in the
resence of rat cytosol, NADPH, acetyl-CoA and concentra-ions ranging from 0.75 to 750 nM were performed. As shownn Fig. 2, 7.5–750 nM concentrations of 3-NBA produced 3-BA-specific DNA adducts that were clearly detectable with
he 32P-postlabeling method. DNA adduct formation was foundo be dose dependent (7.5 nM: 12.3 ± 2.1; 75 nM: 108.3 ± 14.6;50 nM: 1500 ± 125.1 per 108 normal nucleotides). A 0.75 nMoncentration (Fig. 2A) resulted in an adduct level of lesshan 1 adduct per 108 normal nucleotides in CT-DNA onhe border of the detection limit. Therefore 7.5 nM whichepresents 7.5 pmol 3-NBA in the in vitro incubations was con-idered as the detection limit for the 32P-postlabeling methodsed.
In EOM from ambient particles worldwide, 3-NBA has beenetected ranging from 1 to 40 fmol/m3 [23,41,42]. Assum-ng a moderate concentration of 5 fmol/m3 of 3-NBA in their of Sao Paulo would result in a total amount of 9 pmol1800 m3 × 5 fmol) of 3-NBA in the EOM. Therefore, given a
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
etection limit of the procedure used (incubation and subsequent2P-postlabeling method) of 7.5 pmol determined with pure 3-BA it is clear that only high air concentrations of 3-NBA arenambiguously detectable.

ARTICLE IN PRESS+ModelMUTGEN-401418; No. of Pages 9
6 G.A. Umbuzeiro et al. / Mutation Research xxx (2008) xxx–xxx
Fig. 2. Autoradiographic profiles of 3-NBA-DNA adducts obtained from anaerobic in vitro incubations of calf thymus DNA by cytosolic activation with differentdoses of 3-NBA (A) 0.75 nM; (B) 7.5 nM; (C) 75 nM and (D) 750 nM. The butanol enrichment procedure of the 32P-postlabeling method was used. Origins in theb min;C ium8
tzNb(uotp
3N3
vs
Ffoes
ottom left corner were cut off before exposure. Autoradiography was (A) 20hromatographic conditions: D1, 1 M sodium phosphate, pH 6.5; D3, 3.5 M lith.0; D5, 1.7 M sodium phosphate, pH 6.0.
As shown in Fig. 3, reductively activated nitro-PAH frac-ions from both sampling sites produced a diagonal radioactiveone on the thin layer plates with both enrichment procedures.o such diagonal radioactive zone was observed when incu-ations with the nitro-PAH fraction from an unexposed filterFig. 3E) or with both nitro-PAH fractions but without cytosol
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
sed as a negative control were analyzed. Total adduct levelsf the diagonal radioactive zones formed by the nitro-PAH frac-ions from the street and park were similar with both enrichmentrocedures (Fig. 4). Although, according to the retention time,
alwfi
ig. 3. 32P-postlabeling DNA adduct profiles of DNAs modified in vitro by rat cytorom either the Cerqueira Cesar street station A, C, F, H or Ibirapuera park station Bxy-PAH fraction of extracts from Cerqueira Cesar street station without rat liver cyxtraction (but) version of the postlabeling assay as indicated. The areas enclosed by tpots along the diagonal line (F–I) used for calculation of RALs.
(B) 10 min; and (C, D) 1 min. Arrows in A indicate faint DNA adduct spots.formate, 8.5 M urea, pH 3.50; D4, 0.8 M LiCl, 0.5 M Tris–HCl, 8.5 M urea, pH
-NBA expected to be eluted in the nitro-PAH fraction the 3-BA-specific DNA adduct pattern was not detectable on the
2P-postlabeling maps.In comparison to the nitro-PAH fractions, reductively acti-
ated oxy-PAH fractions produced multiple discrete radioactivepots along the diagonal line on the thin layer plates (Fig. 3) and
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
round three-times higher total adduct levels (Fig. 4) when ana-yzed in the same way. No adduct spots along the diagonal zoneere observed with the oxy-PAH fraction from an unexposedlter or without activating system (Fig. 3J). Similar multiple
sol mediated reduction of nitro-PAH and oxy-PAH fractions of filter extracts, D, G, I Ibirapuera park station as indicated. E: extract from a blank filter; J:
tosol DNA adducts were enriched by either the nuclease P 1 (NP 1) or butanolhe boxes represent the diagonal radioactive zone (A–D) and the discrete adduct
ARTICLE IN PRESS+ModelMUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Mutation Research xxx (2008) xxx–xxx 7
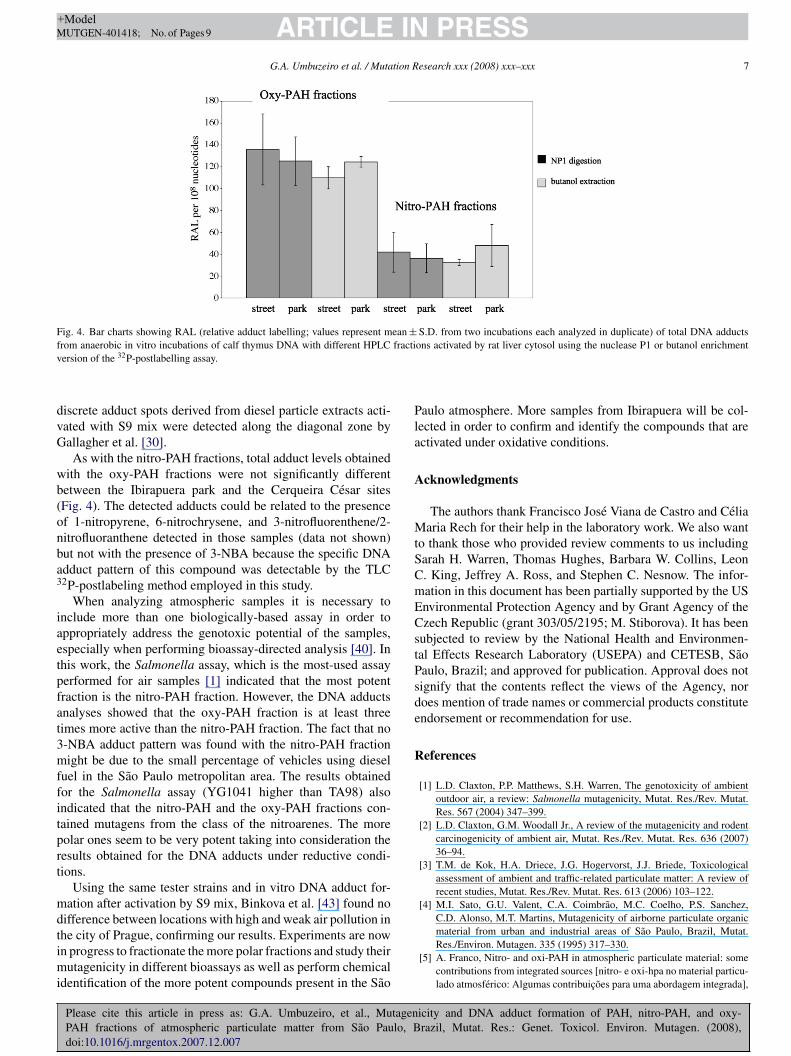
Fig. 4. Bar charts showing RAL (relative adduct labelling; values represent mean ± S.D. from two incubations each analyzed in duplicate) of total DNA adductsf fractiv
dvG
wb(onba3
iaetpfat3mffitprt
mdtimi
Pla
A
MtSCmECstPsde
R
rom anaerobic in vitro incubations of calf thymus DNA with different HPLCersion of the 32P-postlabelling assay.
iscrete adduct spots derived from diesel particle extracts acti-ated with S9 mix were detected along the diagonal zone byallagher et al. [30].As with the nitro-PAH fractions, total adduct levels obtained
ith the oxy-PAH fractions were not significantly differentetween the Ibirapuera park and the Cerqueira Cesar sitesFig. 4). The detected adducts could be related to the presencef 1-nitropyrene, 6-nitrochrysene, and 3-nitrofluorenthene/2-itrofluoranthene detected in those samples (data not shown)ut not with the presence of 3-NBA because the specific DNAdduct pattern of this compound was detectable by the TLC2P-postlabeling method employed in this study.
When analyzing atmospheric samples it is necessary tonclude more than one biologically-based assay in order toppropriately address the genotoxic potential of the samples,specially when performing bioassay-directed analysis [40]. Inhis work, the Salmonella assay, which is the most-used assayerformed for air samples [1] indicated that the most potentraction is the nitro-PAH fraction. However, the DNA adductsnalyses showed that the oxy-PAH fraction is at least threeimes more active than the nitro-PAH fraction. The fact that no-NBA adduct pattern was found with the nitro-PAH fractionight be due to the small percentage of vehicles using diesel
uel in the Sao Paulo metropolitan area. The results obtainedor the Salmonella assay (YG1041 higher than TA98) alsondicated that the nitro-PAH and the oxy-PAH fractions con-ained mutagens from the class of the nitroarenes. The moreolar ones seem to be very potent taking into consideration theesults obtained for the DNA adducts under reductive condi-ions.
Using the same tester strains and in vitro DNA adduct for-ation after activation by S9 mix, Binkova et al. [43] found no
ifference between locations with high and weak air pollution in
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
he city of Prague, confirming our results. Experiments are nown progress to fractionate the more polar fractions and study their
utagenicity in different bioassays as well as perform chemicaldentification of the more potent compounds present in the Sao
ons activated by rat liver cytosol using the nuclease P1 or butanol enrichment
aulo atmosphere. More samples from Ibirapuera will be col-ected in order to confirm and identify the compounds that arectivated under oxidative conditions.
cknowledgments
The authors thank Francisco Jose Viana de Castro and Celiaaria Rech for their help in the laboratory work. We also want
o thank those who provided review comments to us includingarah H. Warren, Thomas Hughes, Barbara W. Collins, Leon. King, Jeffrey A. Ross, and Stephen C. Nesnow. The infor-ation in this document has been partially supported by the USnvironmental Protection Agency and by Grant Agency of thezech Republic (grant 303/05/2195; M. Stiborova). It has been
ubjected to review by the National Health and Environmen-al Effects Research Laboratory (USEPA) and CETESB, Saoaulo, Brazil; and approved for publication. Approval does notignify that the contents reflect the views of the Agency, noroes mention of trade names or commercial products constitutendorsement or recommendation for use.
eferences
[1] L.D. Claxton, P.P. Matthews, S.H. Warren, The genotoxicity of ambientoutdoor air, a review: Salmonella mutagenicity, Mutat. Res./Rev. Mutat.Res. 567 (2004) 347–399.
[2] L.D. Claxton, G.M. Woodall Jr., A review of the mutagenicity and rodentcarcinogenicity of ambient air, Mutat. Res./Rev. Mutat. Res. 636 (2007)36–94.
[3] T.M. de Kok, H.A. Driece, J.G. Hogervorst, J.J. Briede, Toxicologicalassessment of ambient and traffic-related particulate matter: A review ofrecent studies, Mutat. Res./Rev. Mutat. Res. 613 (2006) 103–122.
[4] M.I. Sato, G.U. Valent, C.A. Coimbrao, M.C. Coelho, P.S. Sanchez,C.D. Alonso, M.T. Martins, Mutagenicity of airborne particulate organic
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
material from urban and industrial areas of Sao Paulo, Brazil, Mutat.Res./Environ. Mutagen. 335 (1995) 317–330.
[5] A. Franco, Nitro- and oxi-PAH in atmospheric particulate material: somecontributions from integrated sources [nitro- e oxi-hpa no material particu-lado atmosferico: Algumas contribuicoes para uma abordagem integrada],

IN+ModelM
8 tion R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
ARTICLEUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Muta
Departamento de Quımica Ambiental, Instituto de Quımica (Departmentof Ambient Chemistry, Institute of Chemistry), Universidade de Sao Paulo,Sao Paulo, Brazil, 2006, pp. 1–104.
[6] CETESB, Relatorio de qualidade do ar no estado de Sao Paulo 2004,Available from http://www.cetesb.sp.gov.br/Ar/relatorios.asp, (2005).
[7] S.A. Kyrtopoulos, P. Georgiadis, H. Autrup, N.A. Demopoulos, P. Farmer,A. Haugen, K. Katsouyanni, B. Lambert, S. Ovrebo, R. Sram, G.Stephanou, J. Topinka, Biomarkers of genotoxicity of urban air pollu-tion. Overview and descriptive data from a molecular epidemiology studyon populations exposed to moderate-to-low levels of polycyclic aromatichydrocarbons: The Aulis project, Mutat. Res./Genet. Toxicol. Environ.Mutagen. 496 (2001) 207–228.
[8] A.J. Cohen, H.R. Anderson, B. Ostro, K.D. Pandey, M. Krzyzanowski, N.Kunzli, K. Gutschmidt, A. Pope, I. Romieu, J.M. Samet, K. Smith, Theglobal burden of disease due to outdoor air pollution, J. Toxicol. Environ.Health A 68 (2005) 1301–1307.
[9] D. Krewski, R. Burnett, M. Jerrett, C.A. Pope, D. Rainham, E. Calle, G.Thurston, M. Thun, Mortality and long-term exposure to ambient air pol-lution: Ongoing analyses based on the American cancer society cohort, J.Toxicol. Environ. Health. A 68 (2005) 1093–1109.
10] C.A. Pope III, R.T. Burnett, M.J. Thun, E.E. Calle, D. Krewski, K.Ito, G.D. Thurston, Lung cancer, cardiopulmonary mortality, and long-term exposure to fine particulate air pollution, JAMA 287 (2002) 1132–1141.
11] H. Tokiwa, Y. Ohnishi, Mutagenicity and carcinogenicity of nitroarenesand their sources in the environment, Crit. Rev. Toxicol. 17 (1986) 23–60.
12] H.A. Bamford, D.Z. Bezabeh, S. Schantz, S.A. Wise, J.E. Baker, Deter-mination and comparison of nitrated-polycyclic aromatic hydrocarbonsmeasured in air and diesel particulate reference materials, Chemosphere50 (2003) 575–587.
13] IARC monographs on the evaluation of carcinogenic risks to humans.Diesel and gasoline engine exhausts and some nitroarenes. InternationalAgency for Research on Cancer, IARC Monogr. Eval. Carcinog. RisksHum. 46 (1989) 1–458.
14] IPCS, Selected nitro- and nitro–oxy-polycyclic aromatic hydrocarbons,International Programme on Chemical Safety (IPCS), Geneva, Switzerland,2003, p. 480.
15] USEPA, Health assessment document for diesel engine exhaust,EPA/600/8-90/057F, National Center for Environmental Assessment,Office of Research and Development, US. Environmental ProtectionAgency, Washington, D.C., 2002, p. 669.
16] V. Purohit, A.K. Basu, Mutagenicity of nitroaromatic compounds, Chem.Res. Toxicol. 13 (2000) 673–692.
17] E. Nagy, M. Zeisig, K. Kawamura, Y. Hisamatsu, A. Sugeta, S. Adachi,L. Moller, DNA adducts and tumour formations in rats after intratrachealadministration of the urban air pollutant 3-nitrobenzanthrone, Carcinogen-esis 26 (2005) 1821–1828.
18] V.M. Arlt, 3-Nitrobenzanthrone, a potential human cancer hazard in dieselexhaust and urban air pollution: A review of the evidence, Mutagenesis 20(2005) 399–410.
19] T. Enya, H. Suzuki, T. Watanabe, T. Hirayama, Y. Hisamatsu, 3-Nitrobenzanthrone, a powerful bacterial mutagen and suspected humancarcinogen found in diesel exhaust and airborne particulates, Environ. Sci.Technol. 31 (1997) 2772–2776.
20] V.M. Arlt, C.A. Bieler, W. Mier, M. Wiessler, H.H. Schmeiser, DNAadduct formation by the ubiquitous environmental contaminant 3-nitrobenzanthrone in rats determined by (32)p-postlabeling, Int. J. Cancer93 (2001) 450–454.
21] V.M. Arlt, M. Stiborova, A. Hewer, H.H. Schmeiser, D.H. Phillips, Humanenzymes involved in the metabolic activation of the environmental con-taminant 3-nitrobenzanthrone: evidence for reductive activation by humanNADPH: Cytochrome P450 reductase, Cancer Res. 63 (2003) 2752–2761.
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
22] V.M. Arlt, H.H. Schmeiser, M.R. Osborne, M. Kawanishi, T. Kanno, T.Yagi, D.H. Phillips, T. Takamura-Enya, Identification of three major DNAadducts formed by the carcinogenic air pollutant 3-nitrobenzanthrone inrat lung at the C8 and N2 position of guanine and at the N6 position ofadenine, Int. J. Cancer 118 (2006) 2139–2146.
[
PRESSesearch xxx (2008) xxx–xxx
23] V.M. Arlt, M. Stiborova, C.J. Henderson, M.R. Osborne, C.A. Bieler, E.Frei, V. Martinek, B. Sopko, C.R. Wolf, H.H. Schmeiser, D.H. Phillips,Environmental pollutant and potent mutagen 3-nitrobenzanthrone formsDNA adducts after reduction by NADPH quinone oxidoreductase andconjugation by acetyltransferases and sulfotransferases in human hepaticcytosols, Cancer Res. 65 (2005) 2644–2652.
24] M. Stiborova, H. Dracinska, J. Hajkova, P. Kaderabkova, E. Frei, H.H.Schmeiser, P. Soucek, D.H. Phillips, V.M. Arlt, The environmental pol-lutant and carcinogen 3-nitrobenzanthrone and its human metabolite3-aminobenzanthrone are potent inducers of rat hepatic cytochromes P4501A1 and -1A2 and NADPH Quinone oxidoreductase, Drug Metab. Dispos.34 (2006) 1398–1405.
25] V.M. Arlt, L. Zhan, H.H. Schmeiser, M. Honma, M. Hayashi, D.H. Phillips,T. Suzuki, DNA adducts and mutagenic specificity of the ubiquitous envi-ronmental pollutant 3-nitrobenzanthrone in Muta mouse, Environ. Mol.Mutagen. 43 (2004) 186–195.
26] C.A. Bieler, M.G. Cornelius, M. Stiborova, V.M. Arlt, M. Wiessler, D.H.Phillips, H.H. Schmeiser, Formation and persistence of DNA adductsformed by the carcinogenic air pollutant 3-nitrobenzanthrone in target andnon-target organs after intratracheal instillation in rats, Carcinogenesis 28(2007) 1117–1121.
27] C.A. Bieler, M.G. Cornelius, R. Klein, V.M. Arlt, M. Wiessler, D.H.Phillips, H.H. Schmeiser, DNA adduct formation by the environmentalcontaminant 3-nitrobenzanthrone after intratracheal instillation in rats, Int.J. Cancer 116 (2005) 833–838.
28] M.R. Osborne, V.M. Arlt, C. Kliem, W.E. Hull, A. Mirza, C.A. Bieler, H.H.Schmeiser, D.H. Phillips, Synthesis, characterization, and 32P-postlabelinganalysis of DNA adducts derived from the environmental contaminant 3-nitrobenzanthrone, Chem. Res. Toxicol. 18 (2005) 1056–1070.
29] R.C. Gupta, Enhanced sensitivity of 32P-postlabeling analysis of aromaticcarcinogen: DNA adducts, Cancer Res. 45 (1985) 5656–5662.
30] J.E. Gallagher, M.J. Kohan, M.H. George, J. Lewtas, Improvement in thediagnostic potential of 32P -postlabeling analysis demonstrated by the selec-tive formation and comparative analysis of nitrated-PAH-derived adductsarising from diesel particle extracts, Carcinogenesis 12 (1991) 1685–1691.
31] A. Liberti, P. Ciccioli, High resolution chromatographic techniques forthe evaluation of atmospheric pollutants HRC & CC, J. High Resolut.Chromatogr. Commun. 9 (1986) 492–501.
32] D.M. Maron, B.N. Ames, Revised methods for the Salmonella muta-genicity test, Mutat Res./Environ. Mutagen. Relat. Subj. 113 (1983) 173–215.
33] Y. Hagiwara, M. Watanabe, Y. Oda, T. Sofuni, T. Nohmi, Specificityand sensitivity of Salmonella typhimurium YG1041 and YG1042 strainspossessing elevated levels of both nitroreductase and acetyltransferaseactivity, Mutat. Res./Environ. Mutagen. Relat. Subj. 291 (1993) 171–180.
34] N.Y. Kado, D. Langley, E. Eisenstadt, A simple modification of theSalmonella liquid-incubation assay. Increased sensitivity for detectingmutagens in human urine, Mutat. Res./Environ. Mutagen. Relat. Subj. 121(1983) 25–32.
35] L. Bernstein, J. Kaldor, J. McCann, M.C. Pike, An empirical approach tothe statistical analysis of mutagenesis data from the Salmonella test, Mutat.Res./Environ. Mutagen. Relat. Subj. 97 (1982) 267–281.
36] V.M. Arlt, H. Glatt, E. Muckel, U. Pabel, B.L. Sorg, H.H. Schmeiser,D.H. Phillips, Metabolic activation of the environmental contaminant3-nitrobenzanthrone by human acetyltransferases and sulfotransferase,Carcinogenesis 23 (2002) 1937–1945.
37] V.M. Arlt, B.L. Sorg, M. Osborne, A. Hewer, A. Seidel, H.H. Schmeiser,D.H. Phillips, DNA adduct formation by the ubiquitous environmental pol-lutant 3-nitrobenzanthrone and its metabolites in rats, Biochem. Biophys.Res. Commun. 300 (2003) 107–114.
38] D.M. DeMarini, L.R. Brooks, S.H. Warren, T. Kobayashi, M.I. Gilmour,
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
P. Singh, Bioassay-directed fractionation and Salmonella mutagenicity ofautomobile and forklift diesel exhaust particles, Environ. Health Perspect.112 (2004) 814–819.
39] P. Singh, D.M. DeMarini, C.A. Dick, D.G. Tabor, J.V. Ryan, W.P. Linak, T.Kobayashi, M.I. Gilmour, Sample characterization of automobile and fork-

IN+ModelM
tion R
[
[
[
ARTICLEUTGEN-401418; No. of Pages 9
G.A. Umbuzeiro et al. / Muta
lift diesel exhaust particles and comparative pulmonary toxicity in mice,Environ. Health Perspect. 112 (2004) 820–825.
40] C.H. Marvin, L.M. Hewitt, Analytical methods in bioassay-directed investi-gations of mutagenicity of air particulate material, Mutat. Res./Rev. Mutat.
Please cite this article in press as: G.A. Umbuzeiro, et al., MutagenPAH fractions of atmospheric particulate matter from Sao Paulo, Bdoi:10.1016/j.mrgentox.2007.12.007
Res. 636 (2007) 4–35.41] P.T. Phousongphouang, A.J. Grosovsky, D.A. Eastmond, M. Covarrubias, J.
Arey, The genotoxicity of 3-nitrobenzanthrone and the nitropyrene lactonesin human lymphoblasts, Mutat. Res./Genet. Toxicol. Environ. Mutagen.472 (2000) 93–103.
[
PRESSesearch xxx (2008) xxx–xxx 9
42] N. Tang, R. Taga, T. Hattori, K. Tamura, A. Toroba, R. Kizu, K. Hayakawa,Determination of atmospheric nitrobenzanthrones by high-performanceliquid chromatography with chemiluminescence detection, Anal. Sci. 20(2004) 119–123.
icity and DNA adduct formation of PAH, nitro-PAH, and oxy-razil, Mutat. Res.: Genet. Toxicol. Environ. Mutagen. (2008),
43] B. Binkova, M. Cerna, A. Pastorkova, R. Jelinek, I. Benes, J. Novak, R.J.Sram, Biological activities of organic compounds adsorbed onto ambientair particles: Comparison between the cities of Teplice and Prague duringthe summer and winter seasons 2000–2001, Mutat. Res./Fundam. Mol.Mech. Mutagen. 525 (2003) 43–59.




















