
Murine but Not Human Mesenchymal Stem Cells Generate Osteosarcoma-Like Lesions in the Lung
9
Murine but Not Human Mesenchymal Stem Cells Generate Osteosarcoma-Like Lesions in the Lung SUSANA AGUILAR, a,b EMMA NYE, c,d JERRY CHAN, e MICHAEL LOEBINGER, a BRADLEY SPENCER-DENE, c,d NICK FISK, e GORDON STAMP, c,d DOMINIQUE BONNET, b SAM M. JANES a a Centre for Respiratory Research, Rayne Institute, University College London, London, United Kingdom; b Hematopoietic Stem Cell Laboratory and c Experimental Pathology Laboratory, London Research Institute, Cancer Research UK, London, United Kingdom; d Department of Histopathology and e Institute of Reproductive and Developmental Biology, Department of Paediatrics, Imperial College London, Hammersmith Campus, London, United Kingdom Key Words. Tumor • Mesenchymal stem cell • Lung • Cell therapy • Osteosarcoma ABSTRACT Murine mesenchymal stem cells are capable of differentiation into multiple cell types both in vitro and in vivo and may be good candidates to use as cell therapy for diseased or damaged organs. We have previously reported a method of enriching a population of murine MSCs that demonstrated a diverse dif- ferentiation potential both in vitro and in vivo. In this study, we show that this enriched population of murine mesenchymal stem cells embolize within lung capillaries following systemic injection and then rapidly expand within, and invade into, the lung parenchyma, forming tumor nodules. These lesions rarely contain cells bearing the immunohistochemical characteristics of lung epithelium, but they do show the characteristics of immature bone and cartilage that resembles exuberant frac- ture callus or well-differentiated osteosarcoma. Our findings indicate that murine mesenchymal stem cells can behave in a manner similar to tumor cells, with dysregulated growth and aberrant differentiation within the alveolar microenvironment after four passages. We demonstrate that unlike human MSCs, MSCs from different mouse strains can acquire chromosomal abnormalities after only a few in vitro passages. Moreover, other parameters, such as mouse strain used, might also play a role in the induction of these tumors. These findings might be clinically relevant for future stem cell therapy studies. STEM CELLS 2007;25:1586 –1594 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION MSCs (also known as marrow stromal cells) are bone marrow- derived stem cells with the potential to differentiate into bone, cartilage, and fat [1, 2]. Recent studies suggest that they may have further potential, developing into muscle cells [3], neuro- nal progenitors [4], and lung and kidney epithelium [5, 6]. This raises the exciting possibility that MSCs may have therapeutic potential. MSCs can be quickly expanded in vitro and could be used to engraft into diseased tissues expressing missing or damaged genes, as has been reported for the correction of CFTR gene in cystic fibrosis in humans [6]. In vivo experiments have confirmed expectations of tradi- tional MSC differentiation by showing repair of bone and car- tilage after local injection using injury models [5, 7–9]. Intra- peritoneal injection of human MSCs in sheep strongly suggested that MSCs have the capacity to differentiate into multiple cell types [10], whereas systemic infusion of these cells leads to an even wider distribution into multiple organs, with evidence of differentiation into epithelial cells of local tissues [5, 9, 11]. After venous injection, bone marrow-derived cells (BMDC) must pass through the lungs. Crucial to the regeneration of lung epithelium or replacement of genes in genetic disorders, BMDC must engraft and differentiate into an epithelial phenotype. Previous studies suggest higher engraftment of BMDC into the lungs compared with other organs [9, 12], and evidence exists that tissue damage may be important for this engraftment after systemic delivery of both hematopoietic stem cells (HSCs) and MSCs [5, 12, 13]. For example, the engraftment of enhanced green fluorescent protein (eGFP)-labeled BMDC 1 week after endotoxin inhalation [14] or 3 weeks after inhalation of elastase [15] was seen only after a pulmonary insult but not in undam- aged controls. Murine MSC transplantation has led to both lung engraft- ment as type 2 pneumocytes and attenuation of lung damage [16]. However, others have documented that 80% of collagen 1-producing fibroblasts found at sites of bleomycin-induced fibrosis were of donor origin [17]. One reason for this may be the preselection of cells before transplant. We previously de- scribed a culture system for enrichment of murine MSCs that removes contaminating hematopoietic cells and leaves cells capable of in vitro differentiation into bone, cartilage, fat, mus- cle, and neuronal cells [5]. These cells were transduced with a lentiviral construct carrying eGFP, allowing the tracking of engrafted MSC progeny. Although we saw some evidence for differentiation of these cells into multiple cell types, including airway epithelial cells, the mice became short of breath and had to be sacrificed after 28 days. This study examines in detail the Correspondence: Dominique Bonnet, Ph.D., Hematopoietic Stem Cell Laboratory, London Research Institute, Cancer Research UK, 44 Lincoln’s Inn Fields, London WC2A 3PX, U.K. Telephone: 44-207-269-3282; Fax: 44-207-269-3581; e-mail: Dominique.bonnet@ cancer.org.uk Received November 30, 2006; accepted for publication March 7, 2007; first published online in STEM CELLS EXPRESS March 15, 2007. ©AlphaMed Press 1066-5099/2007/$30.00/0 doi: 10.1634/stemcells.2006-0762 TRANSLATIONAL AND CLINICAL RESEARCH S TEM CELLS 2007;25:1586 –1594 www.StemCells.com
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Murine but Not Human Mesenchymal Stem Cells Generate Osteosarcoma-Like Lesions in the Lung

Murine but Not Human Mesenchymal Stem Cells GenerateOsteosarcoma-Like Lesions in the Lung
SUSANA AGUILAR,a,b EMMA NYE,c,d JERRY CHAN,e MICHAEL LOEBINGER,a BRADLEY SPENCER-DENE,c,d
NICK FISK,e GORDON STAMP,c,d DOMINIQUE BONNET,b SAM M. JANESa
aCentre for Respiratory Research, Rayne Institute, University College London, London, United Kingdom;bHematopoietic Stem Cell Laboratory and cExperimental Pathology Laboratory, London Research Institute, CancerResearch UK, London, United Kingdom; dDepartment of Histopathology and eInstitute of Reproductive andDevelopmental Biology, Department of Paediatrics, Imperial College London, Hammersmith Campus, London,United Kingdom
Key Words. Tumor • Mesenchymal stem cell • Lung • Cell therapy • Osteosarcoma
ABSTRACT
Murine mesenchymal stem cells are capable of differentiationinto multiple cell types both in vitro and in vivo and may begood candidates to use as cell therapy for diseased or damagedorgans. We have previously reported a method of enriching apopulation of murine MSCs that demonstrated a diverse dif-ferentiation potential both in vitro and in vivo. In this study, weshow that this enriched population of murine mesenchymalstem cells embolize within lung capillaries following systemicinjection and then rapidly expand within, and invade into, thelung parenchyma, forming tumor nodules. These lesions rarelycontain cells bearing the immunohistochemical characteristicsof lung epithelium, but they do show the characteristics of
immature bone and cartilage that resembles exuberant frac-ture callus or well-differentiated osteosarcoma. Our findingsindicate that murine mesenchymal stem cells can behave in amanner similar to tumor cells, with dysregulated growth andaberrant differentiation within the alveolar microenvironmentafter four passages. We demonstrate that unlike human MSCs,MSCs from different mouse strains can acquire chromosomalabnormalities after only a few in vitro passages. Moreover,other parameters, such as mouse strain used, might also play arole in the induction of these tumors. These findings might beclinically relevant for future stem cell therapy studies. STEMCELLS 2007;25:1586–1594
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
MSCs (also known as marrow stromal cells) are bone marrow-derived stem cells with the potential to differentiate into bone,cartilage, and fat [1, 2]. Recent studies suggest that they mayhave further potential, developing into muscle cells [3], neuro-nal progenitors [4], and lung and kidney epithelium [5, 6]. Thisraises the exciting possibility that MSCs may have therapeuticpotential. MSCs can be quickly expanded in vitro and could beused to engraft into diseased tissues expressing missing ordamaged genes, as has been reported for the correction of CFTRgene in cystic fibrosis in humans [6].
In vivo experiments have confirmed expectations of tradi-tional MSC differentiation by showing repair of bone and car-tilage after local injection using injury models [5, 7–9]. Intra-peritoneal injection of human MSCs in sheep stronglysuggested that MSCs have the capacity to differentiate intomultiple cell types [10], whereas systemic infusion of thesecells leads to an even wider distribution into multiple organs,with evidence of differentiation into epithelial cells of localtissues [5, 9, 11].
After venous injection, bone marrow-derived cells (BMDC)must pass through the lungs. Crucial to the regeneration of lungepithelium or replacement of genes in genetic disorders, BMDC
must engraft and differentiate into an epithelial phenotype.Previous studies suggest higher engraftment of BMDC into thelungs compared with other organs [9, 12], and evidence existsthat tissue damage may be important for this engraftment aftersystemic delivery of both hematopoietic stem cells (HSCs) andMSCs [5, 12, 13]. For example, the engraftment of enhancedgreen fluorescent protein (eGFP)-labeled BMDC 1 week afterendotoxin inhalation [14] or 3 weeks after inhalation of elastase[15] was seen only after a pulmonary insult but not in undam-aged controls.
Murine MSC transplantation has led to both lung engraft-ment as type 2 pneumocytes and attenuation of lung damage[16]. However, others have documented that 80% of collagen1-producing fibroblasts found at sites of bleomycin-inducedfibrosis were of donor origin [17]. One reason for this may bethe preselection of cells before transplant. We previously de-scribed a culture system for enrichment of murine MSCs thatremoves contaminating hematopoietic cells and leaves cellscapable of in vitro differentiation into bone, cartilage, fat, mus-cle, and neuronal cells [5]. These cells were transduced with alentiviral construct carrying eGFP, allowing the tracking ofengrafted MSC progeny. Although we saw some evidence fordifferentiation of these cells into multiple cell types, includingairway epithelial cells, the mice became short of breath and hadto be sacrificed after 28 days. This study examines in detail the
Correspondence: Dominique Bonnet, Ph.D., Hematopoietic Stem Cell Laboratory, London Research Institute, Cancer Research UK, 44Lincoln’s Inn Fields, London WC2A 3PX, U.K. Telephone: 44-207-269-3282; Fax: 44-207-269-3581; e-mail: [email protected] Received November 30, 2006; accepted for publication March 7, 2007; first published online in STEM CELLS EXPRESS March15, 2007. ©AlphaMed Press 1066-5099/2007/$30.00/0 doi: 10.1634/stemcells.2006-0762
TRANSLATIONAL AND CLINICAL RESEARCH
STEM CELLS 2007;25:1586–1594 www.StemCells.com

kinetics of MSC engraftment in the lung parenchyma and theirsubsequent differentiation potential.
We found that after systemic delivery of MSCs, cells em-bolized in lung parenchyma capillaries. These cells both trans-migrated and divided over the next 24 hours. However, far fromshowing the expected local differentiation pattern, the majorityof cells expanded to form tumors of immature disorganizedbone resembling well-differentiated osteosarcoma or exagger-ated callus. However, there were no definitive features of neo-plastic transformation, such as atypical nuclei, tumor necrosis,or areas of undifferentiated sarcoma. The lesions expandedrapidly, destroying the lung parenchyma, and in several casesled to recipient death by 28 days. Importantly, we found that theformation of these tumors appears specific to murine MSCs. Thesame type of experiments using MSCs derived from human fetalblood underwent similar initial engraftment but subsequentclearance from the lungs. The precise mechanism of the tumordevelopment is not clear; however, we demonstrated that murineMSCs acquired significant chromosomal abnormalities afterfour in vitro passages, whereas we did not detect any abnormal-ities in human MSCs after six or more passages in vitro. Ourresults show that an enriched population of murine MSCs can beexpanded in vitro for cell therapy studies, but the cells arechromosomally unstable, and this may contribute, in part, totumor formation after systemic injection.
MATERIALS AND METHODS
Murine MSC Isolation, Purification, and ExpansionMSCs from NOD/SCID bone marrow were used for initial exper-iments because of the ease of culturing MSCs without the presenceof contaminating hematopoietic cells in this mouse strain [5]. Laterexperiments used MSCs from Rosa26-LacZ mice (backgroundC57Bl/6J � 129S2). These cells were transplanted into both NOD/SCID mice and syngenic B6/129S2F1 ([C57Bl/6J � 129S2] F1)mice. Bone marrow cells were collected by flushing the femurs,tibias, and iliac crests from 8–12-week-old mice with phosphate-buffered saline (PBS) supplemented with 2% fetal bovine serum(FBS) (Gibco, Paisley, U.K., http://www.invitrogen.com). Redblood cell-depleted bone marrow mononuclear cells were plated ata density of 106 cells per cm2 in murine mesenchymal medium withmurine mesenchymal supplements (Stem Cell Technologies, Van-couver, BC, Canada, http://www.stemcell.com), supplemented with100 IU/ml penicillin and 100 �g/ml streptomycin (Gibco). Half theculture medium was changed at day 3 to remove some nonadherentcells. Whole medium was subsequently replaced weekly. The cellswere grown for 2–3 weeks until almost confluent. Adherent cellswere then detached by 0.25% trypsin-EDTA and replated using a1:3 dilution factor. Subsequent passaging and seeding of the cellswas performed at a density of 5,000 cells per cm2. MSCs wereenriched at passages 2 and 3 by elimination of cells stained with ratanti-mouse CD45-CyChrome and CD11b-PE (BD Biosciences, Ox-ford, U.K., http://www.bdbiosciences.com). The negative fraction(35%–40% of adherent cells) was sorted using the FACSVantage(Becton, Dickinson and Company, Oxford, U.K., http://www.bd.com) and expanded before transduction and injection as describedbelow. At injection, MSCs were resuspended in PBS, 2% fetal calfserum with 1 mM EDTA and filtered to avoid cell aggregates.
Human Adult and Fetal MSC Sample CollectionHuman adult bone marrow cells were purchased from Stem CellTechnologies. Fetal blood collection was approved by the ResearchEthics Committee (Hammersmith and Queen Charlotte’s Hospitals)in compliance with national guidelines regarding the use of fetaltissue for research purposes (Polkinghorne Commission Recom-mendations). All women gave written informed consent for collec-tion and use of human tissues. Murine experiments were carried out
under full ethical approval from the host institutions and the appro-priate Home Office license.
Fetal blood (100 �l) was obtained by ultrasound-guided cardiacaspiration (10 weeks of gestation) before a clinically indicatedtermination of pregnancy. Fetal gestational age was determined bycrown-rump length measurement on ultrasound.
Culture of Adult and Fetal Blood MSCsFetal blood was plated in 100-mm dishes at 105 nucleated cells permilliliter, and bone marrow mononuclear cells were plated at adensity of 106 cells per cm2. Both were cultured in MSC growthmedium consisting of 10% FBS (Stem Cell Technologies) in Dul-becco’s modified Eagle’s medium (Sigma-Aldrich, Poole, Dorset,U.K., http://www.sigmaaldrich.com) supplemented with 2 mM L-glutamine, 50 IU/ml penicillin, 50 mg/ml streptomycin (Gibco-BRL, Gaithersburg, MD, http://www.gibcobrl.com) at 37°C in 5%CO2. After 3 days, nonadherent cells were removed, and the me-dium was replaced. After 10 days, adherent cells were then detachedby 0.25% trypsin-EDTA (Stem Cell Technologies) and replated ata density of 5,000 cells per cm2, expanded, and cultured to conflu-ence in 75-cm2 flasks. Adult BM-derived MSCs and fetal blood-derived MSCs underwent 4 and 5 passages, respectively, beforebeing transduced.
In Vitro Lentivirus-Mediated Gene (eGFP) Transferinto Murine and Human MSCsThe HIV-1-based self-inactivating lentiviral vector (pHRSINcPPT-SEW), carrying the eGFP reporter gene under the control of thespleen focus-forming virus long terminal repeat, was used to trans-duce murine and human adult MSCs. For transduction, 1 � 104
purified MSCs from passage 4 were seeded into individual wells ofa 12-well plate. The following day, virus particles were added at amultiplicity of infection (MOI) of 50 to murine MSCs and at anMOI of 10 to human adult MSCs, and transductions were performedfor 20 hours.
Fetal MSCs were transduced with a lentivirus encoding theeGFP reporter gene driven by hPGK at an MOI of 12.5 as previ-ously described [18].
Transduced MSCs were washed several times after virus re-moval with culture medium to avoid viral contamination beforeinfusion into mice. Flow cytometry assessment of the transducedcells at day 5 showed expression of eGFP expression �95% formurine MSCs [5], 93% for human adult MSCs, and 98% for fetalMSCs [18].
Adoptive TransfersA total of 39 NOD/SCID mice ages 8–12 weeks old received donorNOD/SCID MSCs in four independent in vivo studies. eGFP-MSCs(2 � 106), obtained 4 days post-virus removal to minimize furtherexpansion, were delivered intravenously by tail vein injection intoeach sublethally irradiated mouse (375 cGy using a 137Cs source).Pairs of mice were then sacrificed on days 1, 2, 7, 14, and 28postinfusion, and lungs were collected in three experiments. In thefourth experiment, nine NOD/SCID mice were used; five of themwere injected with eGFP-MSCs (2 � 106), and four were injectedwith nontransduced MSCs (2 � 106). The fourth experiment alsoincluded nine NOD/SCID mice that were injected with MSCs fromRosa26-LacZ mice (0.35 � 106 and 2 � 106 cells) after sublethalirradiation (375 cGy). MSCs from Rosa26-LacZ mice were alsoinjected (0.5 � 106 and 2 � 106 cells) into syngenic mice aftersublethal irradiation of 500 cGy or no irradiation. For the humanfetal MSCs, a total of 16 NOD/SCID mice were injected with 2 �106 cells after sublethal irradiation (375 cGy). Human adult MSCswere injected (1 � 106 and 2 � 106 cells) into 12 NOD/SCID miceafter sublethal irradiation of 375 cGy and into 5 NOD/SCID micewith no irradiation.
Tissue Processing and ImmunohistochemistryTissues were fixed in 10% neutral buffered formalin (NBF) andembedded in paraffin. Sections (4 �m thick) were stained with H&E
1587Aguilar, Nye, Chan et al.
www.StemCells.com
or Alizarin Red or immunostained using the avidin-biotin-peroxi-dase or alkaline phosphatase technique.
For all the mice used in the fourth experiment, lungs wereinflated with 1% paraformaldehyde, fixed in NBF, and embedded inparaffin. For human cells and the MSCs from Rosa-LacZ mice, theleft main bronchus was ligated after lung inflation, and the left lungwas excised and embedded in optimal tissue compound for frozensectioning.
The sections were immunostained with anti-eGFP antibody(rabbit polyclonal; 1:500; Molecular Probes Inc., Leiden, The Neth-erlands, http://probes.invitrogen.com), anti-Ki67 (rat monoclonal;1:25; DAKO, Cambridgeshire, U.K., http://www.dako.com), anti-AE1/AE3 (mouse monoclonal; 1:100; DAKO), anti-endomucin(V.7C7 rat IgG2a polyclonal; 1:500; generously provided by Dr.Dieter Vestweber, Institute of Cell Biology, University of Munster,Munster, Germany), anti-TTF-1 (mouse monoclonal; 1:50; Novo-castra Ltd., Newcastle upon Tyne, U.K., http://www.novocastra.co.uk), anti-osteocalcin (goat polyclonal; 1:100; Santa Cruz Biotech-nology Inc., Heidelberg, Germany, http://www.scbt.com), and anti-collagen type II (mouse monoclonal; 1:25; Biocarta, San Diego,http://www.biocarta.com).
The endomucin antibody required no antigen retrieval. ForKi67, eGFP, TTF-1, and osteocalcin, sections required microwaveantigen retrieval (sodium citrate, pH 6, for 10 minutes), and sectionswere quenched for endogenous peroxidase (with 1.6% H2O2) andwhen necessary for endogenous alkaline phosphatase (2 mM le-vamisole). For AE1/AE3, sections were treated with protease(Streptomyces griseus) antigen retrieval (0.04% in PBS at 37°C for10 minutes; Sigma-Aldrich), and for osteocalcin, sections weretreated with pepsin antigen retrieval (1 mg/ml in 50 mM Tris-HCl,pH 2, for 15 minutes). Biotinylated secondary antibodies (1:250)were used, and immunoreactivity was detected using the ABCperoxidase-based system in combination with 3,3�-diaminobenzi-dine, the Vector Blue alkaline phosphatase-based system, or acombination of both (Vector Laboratories, Burlingame, CA, http://www.vectorlabs.com), following the manufacturer’s protocol. Ab-sent and/or nonspecific primary negative controls were included.For secondary antibodies conjugated with fluorochromes, the sec-tions were incubated with Sudan Black (0.1% in 70% ethanol for 30minutes) to lower autofluorescence.
For the Alizarin Red S stain for calcium, sections were dewaxedand taken to 95% alcohol. The sections were then air-dried beforebeing placed in Alizarin solution for 1–5 minutes (1% aqueous redS [Sigma-Aldrich], pH 6.3) until the desired intensity was seen, andthey were then counterstained with methyl green (Vector Labora-tories) for 2 minutes, rinsed in water, blotted, and then rinsed inacetone for 30 seconds.
All microscopy was performed on a Nikon Eclipse E1000microscope, and all images were captured using a NikonDXM1200F digital camera (Tokyo, http://www.nikon.com).
Karyotype AnalysisMetaphase analysis was performed by using 80% confluent flasksof MSCs that were subsequently incubated with 0.1 �g/ml colcemid(Roche Diagnostics, Basel, Switzerland, http://www.roche-applied-science.com) overnight for passage 1 and for 5 hours for laterpassages and human fetal MSCs. Mitotic cells were washed in PBSand trypsinized, and the cells were incubated in hypotonic solution(75 mM KCl) for 15 minutes at 37°C and fixed in methanol/aceticacid (3:1). Fixed chromosomes spreads were stained with 200 ng/ml4,6-diamidino-2-phenylindole in 2� standard saline citrate andexamined by immunofluorescence microscope. Forty metaphases ofeach passage and cell type were counted.
RESULTS
Isolation and Characterization of Murine MSCsThe murine MSCs used in these experiments have been exten-sively characterized, as previously described [5]. Previous stud-ies have found the isolation and expansion of these cells to bestrain-dependent [19, 20], and we previously found that MSCs
from the NOD/SCID mouse strain exhibited higher cell expan-sion and survival potential after removal of the hematopoieticcells in early passages [5]. The initial experiments were con-ducted using NOD/SCID bone marrow because of this and theavailability of this strain in the laboratory. As previously re-ported [19], we observed that MSC cultures were contaminatedwith hematopoietic cells co-expressing CD45 and CD11b. Con-taminating cells consisted of 0.5%–2% B cells (CD19�), T cells(CD3�), NK cells (NK1.1�), and granulocytes (Ly-6Gr�) [5].After purification, the MSCs were transduced with eGFP-lenti-virus vector without losing their in vitro differentiation capaci-ties into adipocytes, chondrocytes, and osteoblasts (as describedpreviously [5]).
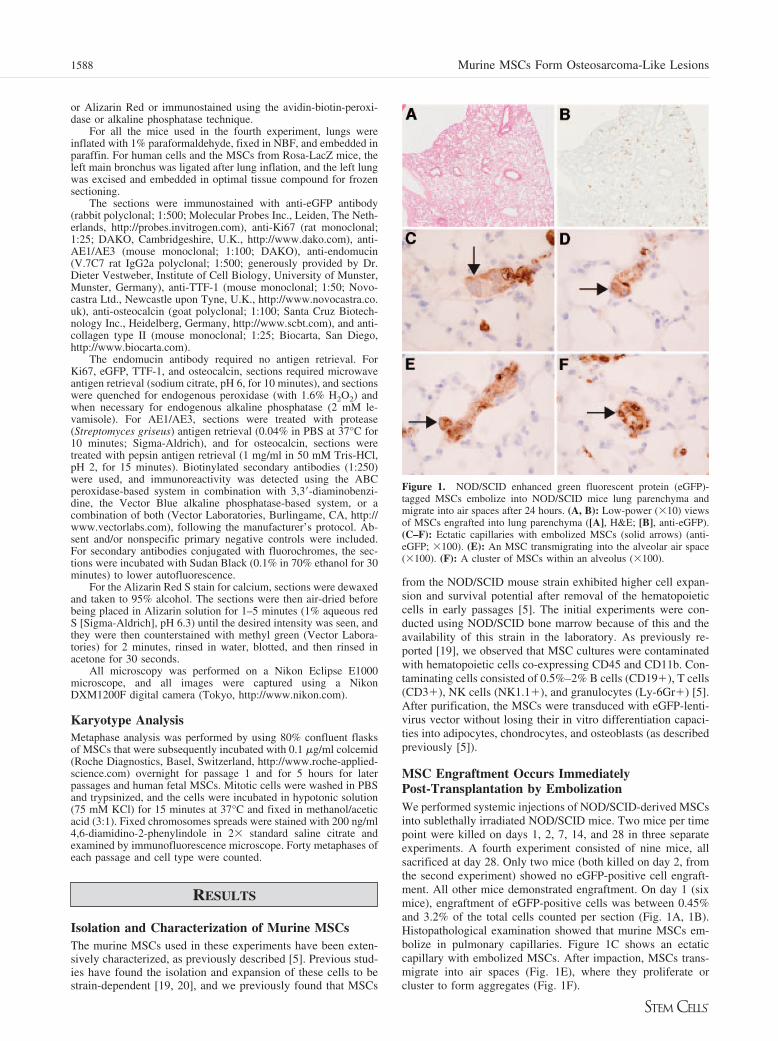
MSC Engraftment Occurs ImmediatelyPost-Transplantation by EmbolizationWe performed systemic injections of NOD/SCID-derived MSCsinto sublethally irradiated NOD/SCID mice. Two mice per timepoint were killed on days 1, 2, 7, 14, and 28 in three separateexperiments. A fourth experiment consisted of nine mice, allsacrificed at day 28. Only two mice (both killed on day 2, fromthe second experiment) showed no eGFP-positive cell engraft-ment. All other mice demonstrated engraftment. On day 1 (sixmice), engraftment of eGFP-positive cells was between 0.45%and 3.2% of the total cells counted per section (Fig. 1A, 1B).Histopathological examination showed that murine MSCs em-bolize in pulmonary capillaries. Figure 1C shows an ectaticcapillary with embolized MSCs. After impaction, MSCs trans-migrate into air spaces (Fig. 1E), where they proliferate orcluster to form aggregates (Fig. 1F).
Figure 1. NOD/SCID enhanced green fluorescent protein (eGFP)-tagged MSCs embolize into NOD/SCID mice lung parenchyma andmigrate into air spaces after 24 hours. (A, B): Low-power (�10) viewsof MSCs engrafted into lung parenchyma ([A], H&E; [B], anti-eGFP).(C–F): Ectatic capillaries with embolized MSCs (solid arrows) (anti-eGFP; �100). (E): An MSC transmigrating into the alveolar air space(�100). (F): A cluster of MSCs within an alveolus (�100).
1588 Murine MSCs Form Osteosarcoma-Like Lesions

MSC Plasticity Is Rare in the Lung DespiteExpansion of Engrafted Donor-Derived CellsWe and others have previously reported that bone marrow-derived cells have the ability to differentiate into lung epithelialcells [5, 13, 16, 17]. Examination of donor murine MSC-derivedtissue (eGFP-positive) revealed rare cells (�1 in 1,000) thatboth were eGFP-positive and expressed markers specific to lungepithelium, such as TTF-1 or AE1/AE3 (Fig. 2A–2D). Althoughwe used Sudan Black to reduce autofluorescence, we cannot ruleout the possibility that these double-stained cells are the result ofcell overlay or apoptosis, as has been described recently [21,22]. However, the majority of eGFP-positive tissue derivedfrom donor MSCs did not costain with either a lung epithelialcell nuclear marker (TTF-1) or an epithelial cell membranemarker (AE1/AE3) (Fig. 2E, 2F). Indeed, the lung epithelialarchitecture within the eGFP-positive tissue was grossly dis-torted by proliferating eGFP-positive cells.
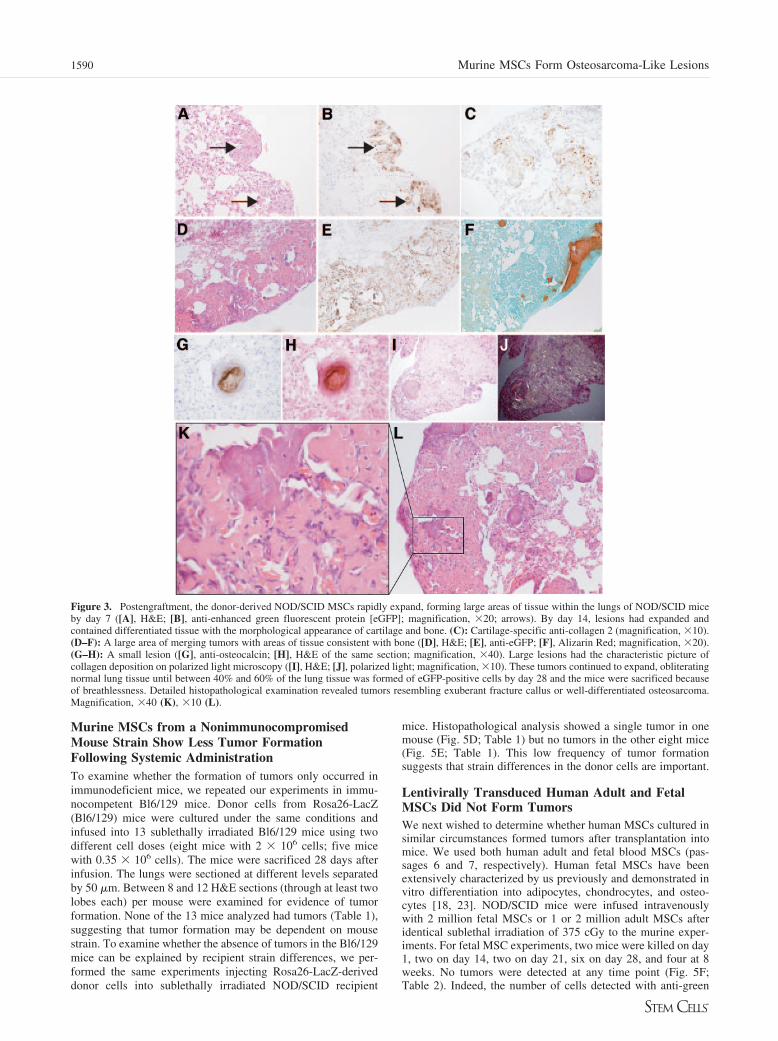
Donor-Derived Murine MSCs Form Tumors Withinthe Lung Consisting of Bone and CartilagePostengraftment, donor-derived NOD/SCID MSCs rapidly ex-panded, forming large areas of tissue within the lungs of NOD/SCID mice by day 7 (Fig. 3A, 3B). The majority of this tissuehad the morphological appearance of cartilage or bone, and byday 14, areas of lung stained clearly for cartilage-specificmarker collagen 2 (Fig. 3C) and bone markers Alizarin Red(Fig. 3F) and osteocalcin (Fig. 3G, 3H). Large areas had thecharacteristic collagen deposition of bone on polarized lightmicroscopy (Fig. 3I, 3J). The tumors continued to expand oblit-erating normal lung tissue until between 40% and 60% of thelung parenchyma contained eGFP-positive cells in 6 of 12 micesacrificed at 28 days (six of six mice from the first threeexperiments). The other nine mice (five transduced with lenti-virus-eGFP and four untransduced) had multiple nodules, butthey were of smaller size, occupying �5% of the lung paren-chymal area on tissue sections (all injected in the fourth exper-iment). These mice were injected over 2–3 minutes, in contrastto the bolus injections given previously to minimize the embo-lization of the cells within the capillaries. Detailed histopatho-logical examination revealed osteochondroblastic proliferationthat resembled the exaggerated healing response of fracturecallus or well-differentiated osteosarcoma. (Fig. 3K, 3L).
Although other organs were analyzed in the first threeexperiments, no tumor formation was detected in kidney, liver,or heart (data not shown).
Murine MSCs Grew Within Vessel Lumen,Infiltrating and Tracking Along Vessel Walls, butDid Not Costain with Endothelial MarkersReports of MSC differentiation into endothelial cells led us tosearch for donor-derived cells that costained with endothelialmarkers. MSCs were frequently seen to expand within vesselsdifferentiating into bone or cartilage (Fig. 4A). eGFP-positivecells also invaded the wall of the vessel in an apparently orga-nized manner (Fig. 4B, 4C) but did not costain for the endothe-lial markers CD31 or endomucin. Within the vessel wall, cellspositive for endothelial markers were not donor-derived (Fig.4C). eGFP-positive cells “pagetoid spread” along the vascularbasement membrane, undermining and displacing the endothe-lium (Fig. 4E, 4F), but again, these donor-derived cells did notstain for endothelial markers.
Murine MSCs Rapidly Proliferated FollowingSystemic InjectionLung epithelium divides irregularly in the steady state, with rarecells staining positively for proliferative markers. Using immu-nohistochemistry for Ki67, a marker of cellular proliferation, wedetermined that injected MSCs were proliferating followingembolization within lung parenchymal capillaries within 24hours (Fig. 4G) and continued to proliferate within the lung,explaining the rapid growth of the lesions formed (Fig. 4H).
Lentiviral Transduction of Donor Cells Does NotCause the Tumor FormationThe cells injected in the first three experiments had been trans-duced with a lentivirus expressing eGFP, allowing easy identi-fication of donor cells. To examine whether the lentiviral trans-duction led to the tumor formation, we performed a fourthexperiment, in which we injected four NOD/SCID mice withnontransduced NOD/SCID cells and five NOD/SCID mice withcells transduced with the lentivirus. All nine mice developedequivalent tumors (Fig. 5A–5C). As noted above, the tumorswere smaller than those seen in the first three experiments buthad the same morphological and differentiation characteristics,indicating that the formation of tumor was not related to virusintegration.
Figure 2. NOD/SCID-derived MSCs at 14days post-transplantation showing rare en-hanced green fluorescent protein (eGFP)-pos-itive cells costaining with epithelial markersin the lung. (A–D): Costaining of anti-eGFP(A), anti-TTF-1 (B), and 4,6-diamidino-2-phenylindole (DAPI) (C); (D): Merged imagewith double-stained cell marked with a whitearrowhead. (E, F): Shown is a large lesioncontaining eGFP-positive tissue but nocostaining with epithelial markers. (E):Green, anti-TTF-1; red, anti-eGFP; blue,DAPI. (F): Green, anti-AE1/AE3; red, anti-eGFP; blue, DAPI. Magnification, �100 (A–D), �20 (E), �40 (F).
1589Aguilar, Nye, Chan et al.
www.StemCells.com
Murine MSCs from a NonimmunocompromisedMouse Strain Show Less Tumor FormationFollowing Systemic AdministrationTo examine whether the formation of tumors only occurred inimmunodeficient mice, we repeated our experiments in immu-nocompetent Bl6/129 mice. Donor cells from Rosa26-LacZ(Bl6/129) mice were cultured under the same conditions andinfused into 13 sublethally irradiated Bl6/129 mice using twodifferent cell doses (eight mice with 2 � 106 cells; five micewith 0.35 � 106 cells). The mice were sacrificed 28 days afterinfusion. The lungs were sectioned at different levels separatedby 50 �m. Between 8 and 12 H&E sections (through at least twolobes each) per mouse were examined for evidence of tumorformation. None of the 13 mice analyzed had tumors (Table 1),suggesting that tumor formation may be dependent on mousestrain. To examine whether the absence of tumors in the Bl6/129mice can be explained by recipient strain differences, we per-formed the same experiments injecting Rosa26-LacZ-deriveddonor cells into sublethally irradiated NOD/SCID recipient
mice. Histopathological analysis showed a single tumor in onemouse (Fig. 5D; Table 1) but no tumors in the other eight mice(Fig. 5E; Table 1). This low frequency of tumor formationsuggests that strain differences in the donor cells are important.
Lentivirally Transduced Human Adult and FetalMSCs Did Not Form TumorsWe next wished to determine whether human MSCs cultured insimilar circumstances formed tumors after transplantation intomice. We used both human adult and fetal blood MSCs (pas-sages 6 and 7, respectively). Human fetal MSCs have beenextensively characterized by us previously and demonstrated invitro differentiation into adipocytes, chondrocytes, and osteo-cytes [18, 23]. NOD/SCID mice were infused intravenouslywith 2 million fetal MSCs or 1 or 2 million adult MSCs afteridentical sublethal irradiation of 375 cGy to the murine exper-iments. For fetal MSC experiments, two mice were killed on day1, two on day 14, two on day 21, six on day 28, and four at 8weeks. No tumors were detected at any time point (Fig. 5F;Table 2). Indeed, the number of cells detected with anti-green
Figure 3. Postengraftment, the donor-derived NOD/SCID MSCs rapidly expand, forming large areas of tissue within the lungs of NOD/SCID miceby day 7 ([A], H&E; [B], anti-enhanced green fluorescent protein [eGFP]; magnification, �20; arrows). By day 14, lesions had expanded andcontained differentiated tissue with the morphological appearance of cartilage and bone. (C): Cartilage-specific anti-collagen 2 (magnification, �10).(D–F): A large area of merging tumors with areas of tissue consistent with bone ([D], H&E; [E], anti-eGFP; [F], Alizarin Red; magnification, �20).(G–H): A small lesion ([G], anti-osteocalcin; [H], H&E of the same section; magnification, �40). Large lesions had the characteristic picture ofcollagen deposition on polarized light microscopy ([I], H&E; [J], polarized light; magnification, �10). These tumors continued to expand, obliteratingnormal lung tissue until between 40% and 60% of the lung tissue was formed of eGFP-positive cells by day 28 and the mice were sacrificed becauseof breathlessness. Detailed histopathological examination revealed tumors resembling exuberant fracture callus or well-differentiated osteosarcoma.Magnification, �40 (K), �10 (L).
1590 Murine MSCs Form Osteosarcoma-Like Lesions

fluorescent protein (GFP) staining reduced from approximately1 in 350 at day 1 to �1 in 5,000 at day 28 (Table 2), suggestingclearance of these cells. In the case of human adult MSCs, weobserved identical results with no engrafted cells and no tumors
detected at 4 and 8 weeks postinjection of eGFP-transducedMSCs. Similarly, no tumors were found in mice injected withnontransduced or nonirradiated adult MSCs (Table 3).
Murine MSCs Have an Abnormal Karyotype, butHuman MSCs Do NotChromosomal abnormalities are thought to be critical in tumorformation, and recent studies have identified abnormalities inmurine MSCs after high passage numbers [24–26]. To checkwhether the donor MSCs used in our study had normal chro-mosomes, we performed a karyotype analysis on all the celltypes injected, counting at least 40 metaphases per cell type andpassage. We found that in both NOD/SCID- and Rosa26-LacZ(Bl6/129)-derived MSCs, the majority of metaphases had theexpected number of chromosomes (n � 40) at passage 1 (datanot shown). Unexpectedly, at the time of injection (passage 4 or5), both strains of MSCs showed numerical chromosomal ab-normalities, with a modal chromosome number of 69 for NOD/SCID-derived MSCs and 76 for LacZ-derived cells (Fig. 6A,6B). These changes in cell karyotype may be responsible, inpart, for the tumor formation observed. However, almost all thehuman fetal MSCs (35 of 40 cells) showed the normal numberof chromosomes (n � 46) at passage 7 (Fig. 6A, 6B).
DISCUSSION
We systemically injected an enriched population of murineMSCs into mice following low-dose irradiation. The aim of our
Figure 4. NOD/SCID MSCs formed tumorswithin vessels that infiltrated vessel walls.(A): Histopathological analysis shows carti-lage tissue growing within a vessel at 14 days(magnification, �10). (B, C): Shown are en-hanced green fluorescent protein (eGFP)-pos-itive cells infiltrating the vessel wall but notexpressing the endothelial marker endomucin([B], anti-eGFP [brown]; [C], the same sec-tion, anti-endomucin [purple]; magnification,�40) (arrowheads point to two endomucin-positive cells (purple in [C]) that do notcostain with eGFP, indicated in [B], enlargedin insets). [D–F]: Shown are eGFP-positivecells extended along the vessel wall under-neath the endothelium (arrowheads) but,again, not costaining ([D, E], anti-eGFP[brown]; [F], the same section as [E], anti-endomucin [purple; shown as a thin dark-purple line on the luminal side of the endo-thelial cells in [F]). Magnification, �40 (D),�100 (E, F). Engrafted MSCs were highlyproliferative from day 1 postdelivery. (G):MSCs on day 1 postinjection. (H): a day 14lesion. Both slides (G, H) costained with anti-eGFP (blue) and anti-Ki67 (brown). Magnifi-cation, �100 (G), �20 (H).
Figure 5. Tumor formation analysis of mouse and human MSCs.MSCs from NOD/SCID mice form tumors with or without lentiviraltransduction. Magnification, �20 (A–C). (A): Enhanced green fluores-cent protein (eGFP)-positive donor cells in the tumors formed afterinjection of NOD/SCID MSCs that were transduced with lentivirus-eGFP; (B): Serial section stained with an isotype control. MSCs fromNOD/SCID mice without lentiviral transduction were also capable offorming tumors at the same level and, as expected, were negative foreGFP (C). (D): Shown is tumor formed after LacZ MSCs were injectedinto NOD/SCID mice (arrow; magnification, �40; H&E), whereas notumors were found in the other eight mice analyzed ([E]; magnification,�10; H&E). NOD/SCID mice injected with human fetal MSCs dem-onstrated no tumor formation ([F]; magnification, �10: H&E).
1591Aguilar, Nye, Chan et al.
www.StemCells.com
study was to evaluate the extent of lung parenchyma engraft-ment as epithelial cells under these conditions. Although we sawrare evidence of differentiation of MSCs into cells with stainingcharacteristics of lung epithelium, we saw widespread engraft-ment and expansion of MSCs into large tumors, showing dif-ferentiation into bone and cartilage cells with destruction ofnormal lung architecture.
We have demonstrated that not only do murine MSCs cul-tured under our conditions have high in vitro capacity forosteogenic and chondrogenic differentiation [5] but this poten-tial is retained in vivo and occurs outside of a normal biologicalmicroenvironment. On engraftment into the lungs, most likely atthe first pass following venous injection, MSCs begin prolifer-ating immediately, as indicated by immunopositivity with anti-Ki67 antibody, and then continue to proliferate and differentiateinto bone and cartilage lineages. The lesions are multiple,throughout the lungs, and expand rapidly, involving between40% and 60% of cross-sectional area of lung parenchyma at 28days, leading to recipient sacrifice due to breathlessness. Thelesions grew destructively both within alveolar spaces and ves-sels. The majority of lesions have osteoblastic predominance,but occasional tumors consist predominantly of chondrocytesand chondroblasts. Morphologically, these tumors are typical offracture callus exuberance, resembling the well-differentiatedosteosarcoma previously recognized in osteogenesis imperfecta[27].
Additional experiments using a different mouse strain, Bl6/129, revealed no tumor formation (0 of 13 mice analyzed).Hence, there does appear to be some strain dependence. Toestablish whether this was due to the genetic background of therecipient, we administered Rosa26-LacZ (Bl6/129)-derivedMSCs into NOD/SCID mice and found that only one of ninemice displayed a small tumor. These results suggest that tumorformation is dependent on both recipient strain and the strainfrom which donor cells are derived. Although both donor mu-rine strains had numerical abnormalities in karyotype, it is
possible that differences in the cytogenetic events occurringbetween the two strains could explain the differences in thetumorigenesis capacity. This hypothesis is supported by a reportthat shows a variety of cytogenetic abnormalities in MSCsderived from C57/Bl6 mice, some capable of forming sarcomasin C57/Bl6 mice but others not [28]. Moreover, another possibleexplanation could be differences in the expression of cell adhe-sion molecules that may favor the higher engraftment in the lungparenchyma of NOD/SCID-derived MSCs. Of note, all fourseparate cultures of NOD/SCID MSCs produced lung tumorsupon injection in our experiments.
Murine MSCs, although less well studied than humanMSCs, have similar in vitro differentiation potential but differ ina number of other attributes, including cell surface moleculeexpression [5, 29]. This may alter cell homing and engraftmentafter systemic injection. To assess whether human MSCs wouldalso form tumors in our model, we injected both human fetal andadult MSCs carrying a lentivirally expressed eGFP marker.Human cells engrafted into the lung parenchyma, but at a lowerlevel. Importantly, we found no evidence of tumor formationafter 2 months, and the levels of GFP-positive cells decreasedover time, with no evidence of proliferation.
Few studies have examined the effects of systemic deliveryof MSCs. Ortiz et al. reported an attenuation of bleomycin-induced inflammation and fibrosis after transplanting murineMSCs, resulting in engraftment of donor-derived type 2 pneu-mocytes, although at lower levels than those of HSC transplants[16]. A separate group reported engraftment of a plastic-adher-ent murine bone marrow population of cells as type 1 pneumo-cytes [13]. Two recent studies, however, demonstrated thatlong-term cultures of MSCs (4–5 months) contain a populationof transformed cells capable of tumor formation after systemicinjection [24, 30]. In one study, there was no apparent differ-entiation of human MSC-derived tumors within the lung paren-chyma [30], whereas in the other, murine MSC-derived tumorsappeared to undergo fibroblastic differentiation [24]. In vitro,both these cells had an abnormal karyotype and high telomeraseactivity. Similarly, in long-term culture, telomerase transducedhuman MSCs evolved spontaneous genetic changes leading totumorigenicity [31]. More recently still, a study demonstratedsarcoma-like tumors developing after injection of MSCs fromC57/Bl6 mice. These cells showed cytogenetic abnormalitiesafter nine in vitro passages that were capable of tumorigenesis.Nevertheless, the authors were unable to reproduce the resultswith subsequently derived MSCs despite demonstrating cytoge-netic abnormalities in all their MSC cultures from two mousestrains [28], indicating that the nature of the chromosomalaberrations and/or other factors might influence the capacity ofMSCs to form tumors. Importantly, the culture systems in thesedifferent studies and our own used standard culture conditionsand thus are unlikely to be responsible for the tumor formation.
Table 1. Summary of tumor nodule formation
Number of transplanted mice Donor strain Recipient strain Number of injected cells Tumoral nodules detected
35 NOD/SCID (lentivirus-eGFP) NOD/SCIDa 2 Million 35/354 NOD/SCID (nontransduced) NOD/SCIDa 2 Million 4/49 R26-LacZ NOD/SCIDa 2 Million 1/6
0.35 Million 0/313 R26-Lac Z Bl6/129b 2 Million 0/8
0.5 Million 0/30.5 Millionc 0/2
aNOD/SCID mice received a sublethal dose of irradiation, 375 cGy.bBl6/129 mice received a sublethal dose of irradiation, 500 cGy.cNonirradiated mice.
Table 2. Summary of lung engraftment and tumor formation inbone marrow transplantation of human fetal enhanced greenfluorescent protein-transduced MSCs by histology
Number oftransplanted mice
Timepostinjection
Tumor nodulesdetected
Engrafted cells(% of total cells)
2 1 Day ND 17 per slide2 7 Days ND 3–5 per slide2 21 Days ND 0–1 per slide6 4 Weeks ND �0.014 8 Weeks ND �0.01
All the mice were sublethally irradiated with 375 cGy, and 2million cells were injected.Abbreviation: ND, not detected.
1592 Murine MSCs Form Osteosarcoma-Like Lesions

Why these enriched MSCs form tumors is intriguing. Wehave found that our highly selected murine MSCs rapidly accrue
genetic changes, as demonstrated by their abnormal karyotype.It is likely that this has led to abnormal expression of genes orgenetic/epigenetic changes responsible for tumor formation. Itwould be interesting to examine this short-term-cultured MSCpopulation for the genetic abnormalities frequently found inosteosarcoma tumors and cell lines. The irradiation dose weadministrated in our experiments was not likely critical for thetumor formation, as previous reports showed that this sublethaldose is not harmful to the lung [32, 33]. Supporting these data,the transplanted human MSCs were also given a sublethal doseof irradiation and no tumors were seen.
Our data provide further evidence for the rapid acquisitionof chromosomal abnormalities and potential adverse effects ofsystemic injection of murine MSCs after a minimal in vitroculture necessary to obtain a quantity of cells required for celltherapy studies. These results stress the importance of carefulanalysis of cells used for cell therapy both before and aftertransplantation.
ACKNOWLEDGMENTS
We thank Fernando Anjos-Afonso for help with these experiments,George Elias and his team in the Cancer Research UK histopathol-ogy laboratories, Karen Groot for critical reading of the manuscript,and Denise Sheer for help with karyotyping analysis. This workwas supported by Cancer Research UK. S.M.J. is a Medical Re-search Council Clinician Scientist. D.B. and S.M.J. are joint lastauthors.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Pittenger MF, Mackay AM, Beck SC et al. Multilineage potential ofadult human mesenchymal stem cells. Science 1999;284:143–147.
2 Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietictissues. Science 1997;276:71–74.
3 Woodbury D, Schwarz EJ, Prockop DJ et al. Adult rat and human bonemarrow stromal cells differentiate into neurons. J Neurosci Res 2000;61:364–370.
4 Wakitani S, Saito T, Caplan AI. Myogenic cells derived from rat bonemarrow mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve1995;18:1417–1426.
5 Anjos-Afonso F, Siapati EK, Bonnet D. In vivo contribution of murinemesenchymal stem cells into multiple cell-types under minimal damageconditions. J Cell Sci 2004;117:5655–5664.
6 Wang G, Bunnell BA, Painter RG et al. Adult stem cells from bonemarrow stroma differentiate into airway epithelial cells: Potentialtherapy for cystic fibrosis. Proc Natl Acad Sci U S A 2005;102:186 –191.
7 Hofstetter CP, Schwarz EJ, Hess D et al. Marrow stromal cells formguiding strands in the injured spinal cord and promote recovery. ProcNatl Acad Sci U S A 2002;99:2199–2204.
8 Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells migratethroughout forebrain and cerebellum, and they differentiate into astro-cytes after injection into neonatal mouse brains. Proc Natl Acad SciU S A 1999;96:10711–10716.
9 Pereira RF, O’Hara MD, Laptev AV et al. Marrow stromal cells as asource of progenitor cells for nonhematopoietic tissues in transgenicmice with a phenotype of osteogenesis imperfecta. Proc Natl Acad SciU S A 1998;95:1142–1147.
10 Liechty KW, MacKenzie TC, Shaaban AF et al. Human mesenchymal
Table 3. Summary of lung engraftment and tumor formation in bone marrow transplantation of human adult enhanced green fluorescentprotein-transduced MSCs by histology
Number of transplanted mice Time postinjection Number of injected cells Tumor nodules detected Engrafted cells (% of total cells)
5 4 Weeksa 1 Million ND �0.013 4 Weeksa 2 Million ND �0.013 4 Weeksb 1 Million ND �0.012 4 Weeksb 2 Million ND �0.014 8 Weeksa 1 Million (nontransduced) ND �0.01
aNOD/SCID mice received a sublethal dose of irradiation, 375 cGy.bNOD/SCID receiving no irradiation.Abbreviation: ND, not detected.
Figure 6. Murine MSCs but not human MSCs show abnormal karyo-type. The analysis of 40 metaphases of murine and human MSCs at thetime of injection (P4, P5, or P7) showed numerical chromosomal ab-normalities in both NOD/SCID and LacZ MSCs but not in the humanMSCs ([A]; magnification, �100; 4,6-diamidino-2-phenylindole stain-ing). Although the majority of the human cells had an expected numberof chromosomes (n � 46), both strains of murine MSCs showedchanges in the number of chromosomes having the majority of met-aphases within the chromosome range 41–79 (B). Abbreviation: P,passage.
1593Aguilar, Nye, Chan et al.
www.StemCells.com

stem cells engraft and demonstrate site-specific differentiation after inutero transplantation in sheep. Nat Med 2000;6:1282–1286.
11 Devine SM, Cobbs C, Jennings M et al. Mesenchymal stem cells dis-tribute to a wide range of tissues following systemic infusion intononhuman primates. Blood 2003;101:2999–3001.
12 Krause DS, Theise ND, Collector MI et al. Multi-organ, multi-lineageengraftment by a single bone marrow-derived stem cell. Cell 2001;105:369–377.
13 Kotton DN, Ma BY, Cardoso WV et al. Bone marrow-derived cells asprogenitors of lung alveolar epithelium. Development 2001;128:5181–5188.
14 Yamada M, Kubo H, Kobayashi S et al. Bone marrow-derived progenitorcells are important for lung repair after lipopolysaccharide-induced lunginjury. J Immunol 2004;172:1266–1272.
15 Ishizawa K, Kubo H, Yamada M et al. Bone marrow-derived cellscontribute to lung regeneration after elastase-induced pulmonary emphy-sema. FEBS Lett 2004;556:249–252.
16 Ortiz LA, Gambelli F, McBride C et al. Mesenchymal stem cell engraft-ment in lung is enhanced in response to bleomycin exposure and ame-liorates its fibrotic effects. Proc Natl Acad Sci U S A 2003;100:8407–8411.
17 Hashimoto N, Jin H, Liu T et al.. Bone marrow-derived progenitor cellsin pulmonary fibrosis. J Clin Invest 2004;113:243–252.
18 Chan J, O’Donoghue K, de la Fuente J et al. Human fetal mesenchymalstem cells as vehicles for gene delivery. STEM CELLS 2005;23:93–102.
19 Phinney DG, Kopen G, Isaacson RL et al. Plastic adherent stromal cellsfrom the bone marrow of commonly used strains of inbred mice: Vari-ations in yield, growth, and differentiation. J Cell Biochem 1999;72:570–585.
20 Peister A, Mellad JA, Larson BL et al. Adult stem cells from bonemarrow (MSCs) isolated from different strains of inbred mice vary insurface epitopes, rates of proliferation, and differentiation potential.Blood 2004;103:1662–1668.
21 Kotton DN, Fabian AJ, Mulligan RC. Failure of bone marrow to recon-stitute lung epithelium. Am J Respir Cell Mol Biol 2005;33:328–334.
22 Chang JC, Summer R, Sun X et al. Evidence that bone marrow cells donot contribute to the alveolar epithelium. Am J Respir Cell Mol Biol2005;33:335–342.
23 Chan J, O’Donoghue K, Gavina M et al. Galectin-1 induces skeletalmuscle differentiation in human fetal mesenchymal stem cells and in-creases muscle regeneration. STEM CELLS 2006;24:1879–1891.
24 Miura M, Miura Y, Padilla-Nash HM et al. Accumulated chromosomalinstability in murine bone marrow mesenchymal stem cells leads tomalignant transformation. STEM CELLS 2006;24:1095–1103.
25 Hackett JA, Feldser DM, Greider CW. Telomere dysfunction increasesmutation rate and genomic instability. Cell 2001;106:275–286.
26 Feldser DM, Hackett JA, Greider CW. Telomere dysfunction and theinitiation of genome instability. Nat Rev Cancer 2003;3:623–627.
27 Ramı́rez N, Vilella FE, Colon M et al. Osteogenesis imperfecta andhyperplastic callus formation in a family: A report of three cases and areview of the literature. J Pediatr Orthop B 2003;12:88–96.
28 Tolar J, Nauta AJ, Osborn MJ et al. Sarcoma derived from culturedmesenchymal stem cells. STEM CELLS 2007;25:371–379.
29 Deans RJ, Moseley AB. Mesenchymal stem cells: Biology and potentialclinical uses. Exp Hematol 2000;28:875–884.
30 Rubio D, Garcia-Castro J, Martin MC et al. Spontaneous human adultstem cell transformation. Cancer Res 2005;65:3035–3039.
31 Burns JS, Abdallah BM, Guldberg P et al. Tumorigenic heterogeneity incancer stem cells evolved from long-term cultures of telomerase-immor-talized human mesenchymal stem cells. Cancer Res 2005;65:3126–3135.
32 Theise ND, Henegariu O, Grove J et al. Radiation pneumonitis in mice:A severe injury model for pneumocyte engraftment from bone marrow.Exp Hematol 2002;30:1333–1338.
33 Herzog EL, Van Arnam J, Hu B et al. Threshold of lung injury requiredfor the appearance of marrow-derived lung epithelia. STEM CELLS2006;24:1986–1992.
1594 Murine MSCs Form Osteosarcoma-Like Lesions




















