
Mortality patterns and skeletal physical condition of the carnivorans from the Miocene assemblage of...
16
Mortality patterns and skeletal physical condition of the carnivorans from the Miocene assemblage of Batallones-1 (Madrid Basin, Spain) M. Soledad Domingo, M. Teresa Alberdi, Beatriz Azanza and Jorge Morales With 5 figures and 4 tables Domingo, m.S., AlberDi, m.T., AzAnzA, b. & morAleS, J. (2012): Mortality patterns and skeletal physical condition of the carnivorans from the Miocene assemblage of Batallones-1 (Madrid Basin, Spain). – N. Jb. Geol. Paläont. Abh., 265: 131-145; Stuttgart. Abstract: Batallones-1 lower level assemblage contains numerous and diverse Miocene vertebrates that were deposited over time at the bottom part of a cavity. In this study, the age structure and skeletal physical condition of the four more abundant taxa, all of them carnivorans, are analysed to better delineate the taphonomic history undergone by the remains and the concentration mode of the assemblage. The prime-adult dominated profile of the two sabertoothed cats and the hyaenid can be related to a proneness of these individuals to search for resources in dangerous locations during the stressful period of dispersal. In turn, the amphicyonid profile, where juveniles are abundant, can be the result of the entrance in the cave of mothers with cubs. The demography of the analysed taxa has allowed to discard in Batallones-1 LLA mass, attritional or accidental death modes. Rather, the entrance of animals in the cavity was intentional and resulted in the entrapment of the individuals. Besides, the animals represented in Batallones-1 LLA do not show signals of poor physical condition as revealed by the comparison of the frequency of osteopathologies and broken canines in life of the extinct taxa with modern healthy counterparts. Consequently, incidence of injuries in the skeleton of these individuals can be discarded as a major cause of entrapment. Key words: age, mortality, injury, carnivores, Batallones-1, Miocene. 1. Introduction Age-frequency distributions of vertebrates from both palaeontological and archaeological sites have long been recognized as a powerful tool that provides valu- able information about the individuals’ pre- or post- depositional history (KurTén 1953, 1958; V oorhieS 1969; STiner 1990; l ymAn 1994; STeele & WeAVer 2002; STeele 2003; KAhlKe & gAuDzinSKi 2005; Ar- genTi & mAzzA 2006; lAuTerS et al. 2008). These analyses are carried out through the comparison of mortality patterns in the fossil record to analogous data coming from demographic studies on modern mammalian communities. These patterns are com- monly represented as histograms where the x axis rep- resents age classes from younger to older and the y axis represents the frequency or Minimum Number of Individuals counted per age class. Two basic types of mortality profiles have been recognized: (1) attritional profile and (2) catastrophic profile. In the attritional pattern (Fig. 1A), very young and, to a lesser extent, very old individuals are overrepresented relative to their live abundances (l ymAn 1994). This mortality type results in a bimodal frequency distribution and is also known as a ‘U-shaped’ profile (l ymAn 1994). In the catastrophic profile (Fig. 1B), also known as the ©2012 E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, Germany www.schweizerbart.de DOI: 10.1127/0077-7749/2012/0251 0077-7749/2012/0251 $ 3.75 N. Jb. Geol. Paläont. Abh. 265/2, 131–145 Article Stuttgart, August 2012
Transcript of Mortality patterns and skeletal physical condition of the carnivorans from the Miocene assemblage of...

Mortality patterns and skeletal physical condition of the carnivorans from the Miocene assemblage of Batallones-1 (Madrid Basin, Spain)
M. Soledad Domingo, M. Teresa Alberdi, Beatriz Azanza and Jorge Morales
With 5 figures and 4 tables
Domingo, m.S., AlberDi, m.T., AzAnzA, b. & morAleS, J. (2012): Mortality patterns and skeletal physical condition of the carnivorans from the Miocene assemblage of Batallones-1 (Madrid Basin, Spain). – N. Jb. Geol. Paläont. Abh., 265: 131-145; Stuttgart.
Abstract: Batallones-1 lower level assemblage contains numerous and diverse Miocene vertebrates that were deposited over time at the bottom part of a cavity. In this study, the age structure and skeletal physical condition of the four more abundant taxa, all of them carnivorans, are analysed to better delineate the taphonomic history undergone by the remains and the concentration mode of the assemblage. The prime-adult dominated profile of the two sabertoothed cats and the hyaenid can be related to a proneness of these individuals to search for resources in dangerous locations during the stressful period of dispersal. In turn, the amphicyonid profile, where juveniles are abundant, can be the result of the entrance in the cave of mothers with cubs. The demography of the analysed taxa has allowed to discard in Batallones-1 LLA mass, attritional or accidental death modes. Rather, the entrance of animals in the cavity was intentional and resulted in the entrapment of the individuals. Besides, the animals represented in Batallones-1 LLA do not show signals of poor physical condition as revealed by the comparison of the frequency of osteopathologies and broken canines in life of the extinct taxa with modern healthy counterparts. Consequently, incidence of injuries in the skeleton of these individuals can be discarded as a major cause of entrapment.
Key words: age, mortality, injury, carnivores, Batallones-1, Miocene.
1. Introduction
Age-frequency distributions of vertebrates from both palaeontological and archaeological sites have long been recognized as a powerful tool that provides valu-able information about the individuals’ pre- or post-depositional history (KurTén 1953, 1958; VoorhieS 1969; STiner 1990; lymAn 1994; STeele & WeAVer 2002; STeele 2003; KAhlKe & gAuDzinSKi 2005; Ar-genTi & mAzzA 2006; lAuTerS et al. 2008). These analyses are carried out through the comparison of mortality patterns in the fossil record to analogous data coming from demographic studies on modern
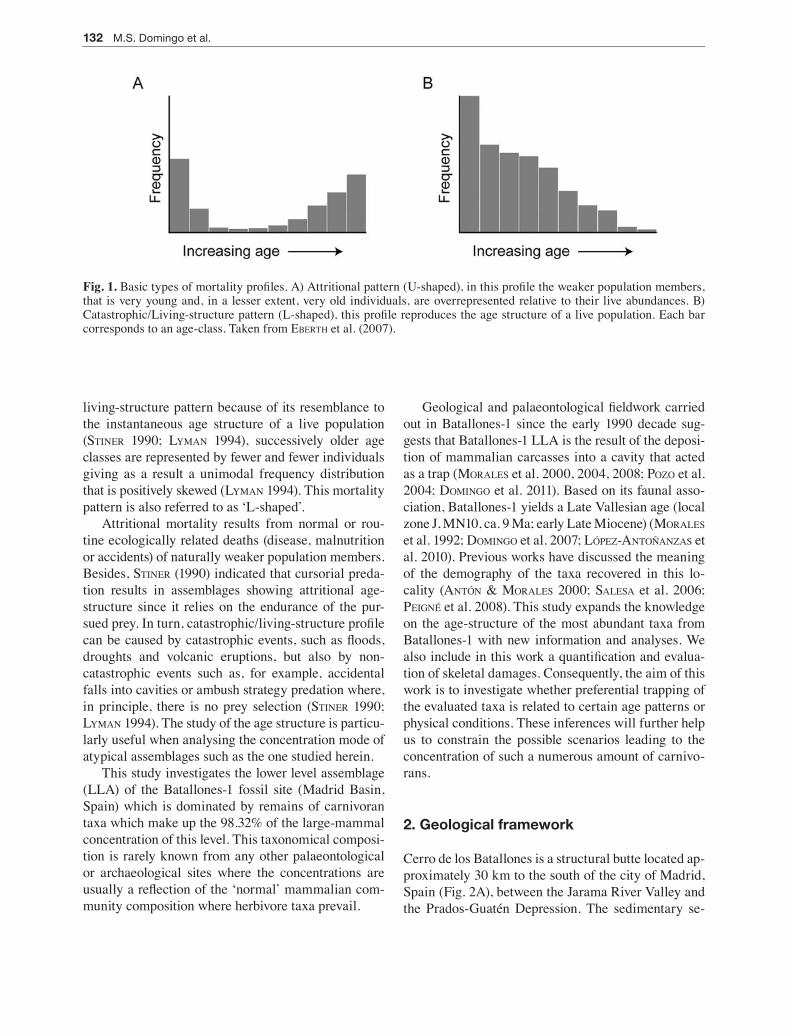
mammalian communities. These patterns are com-monly represented as histograms where the x axis rep-resents age classes from younger to older and the y axis represents the frequency or Minimum Number of Individuals counted per age class. Two basic types of mortality profiles have been recognized: (1) attritional profile and (2) catastrophic profile. In the attritional pattern (Fig. 1A), very young and, to a lesser extent, very old individuals are overrepresented relative to their live abundances (lymAn 1994). This mortality type results in a bimodal frequency distribution and is also known as a ‘U-shaped’ profile (lymAn 1994). In the catastrophic profile (Fig. 1B), also known as the
©2012 E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart, Germany www.schweizerbart.de
DOI: 10.1127/0077-7749/2012/0251 0077-7749/2012/0251 $ 3.75
N. Jb. Geol. Paläont. Abh. 265/2, 131–145 ArticleStuttgart, August 2012
132 M.S. Domingo et al.
living-structure pattern because of its resemblance to the instantaneous age structure of a live population (STiner 1990; lymAn 1994), successively older age classes are represented by fewer and fewer individuals giving as a result a unimodal frequency distribution that is positively skewed (lymAn 1994). This mortality pattern is also referred to as ‘L-shaped’.
Attritional mortality results from normal or rou-tine ecologically related deaths (disease, malnutrition or accidents) of naturally weaker population members. Besides, STiner (1990) indicated that cursorial preda-tion results in assemblages showing attritional age-structure since it relies on the endurance of the pur-sued prey. In turn, catastrophic/living-structure profile can be caused by catastrophic events, such as floods, droughts and volcanic eruptions, but also by non-catastrophic events such as, for example, accidental falls into cavities or ambush strategy predation where, in principle, there is no prey selection (STiner 1990; lymAn 1994). The study of the age structure is particu-larly useful when analysing the concentration mode of atypical assemblages such as the one studied herein.
This study investigates the lower level assemblage (LLA) of the Batallones-1 fossil site (Madrid Basin, Spain) which is dominated by remains of carnivoran taxa which make up the 98.32% of the large-mammal concentration of this level. This taxonomical composi-tion is rarely known from any other palaeontological or archaeological sites where the concentrations are usually a reflection of the ‘normal’ mammalian com-munity composition where herbivore taxa prevail.
Geological and palaeontological fieldwork carried out in Batallones-1 since the early 1990 decade sug-gests that Batallones-1 LLA is the result of the deposi-tion of mammalian carcasses into a cavity that acted as a trap (morAleS et al. 2000, 2004, 2008; Pozo et al. 2004; Domingo et al. 2011). Based on its faunal asso-ciation, Batallones-1 yields a Late Vallesian age (local zone J, MN10, ca. 9 Ma; early Late Miocene) (morAleS et al. 1992; Domingo et al. 2007; lóPez-AnToñAnzAS et al. 2010). Previous works have discussed the meaning of the demography of the taxa recovered in this lo-cality (AnTón & morAleS 2000; SAleSA et al. 2006; Peigné et al. 2008). This study expands the knowledge on the age-structure of the most abundant taxa from Batallones-1 with new information and analyses. We also include in this work a quantification and evalua-tion of skeletal damages. Consequently, the aim of this work is to investigate whether preferential trapping of the evaluated taxa is related to certain age patterns or physical conditions. These inferences will further help us to constrain the possible scenarios leading to the concentration of such a numerous amount of carnivo-rans.
2. Geological framework
Cerro de los Batallones is a structural butte located ap-proximately 30 km to the south of the city of Madrid, Spain (Fig. 2A), between the Jarama River Valley and the Prados-Guatén Depression. The sedimentary se-
Fig. 1. Basic types of mortality profiles. A) Attritional pattern (U-shaped), in this profile the weaker population members, that is very young and, in a lesser extent, very old individuals, are overrepresented relative to their live abundances. B) Catastrophic/Living-structure pattern (L-shaped), this profile reproduces the age structure of a live population. Each bar corresponds to an age-class. Taken from eberTh et al. (2007).

Mortality patterns of carnivorans from Batallones-1 133
quence of the butte consists of three units deposited in terrestrial environments during the Vallesian (early late Miocene). The two lower units are mainly formed by more than 14 m of clays of different composition while the top unit is composed of 5 m of alternations of siliciclastics and marly and carbonate beds (see fur-ther descriptions in morAleS et al. 2000, 2004, 2008; Pozo et al. 2004; Domingo et al. 2011).
A main feature of the fossil assemblages from Cerro de los Batallones is that they are included in a sedimentary matrix that discordantly cuts the three units of the butte indicating that fossil remains were deposited in cavities (Fig. 2B). In this area, clays are dominant so a pseudo-karstic process known as piping (subsurficial hydraulic erosion of fundamentally clay-ey materials) has been invoked as being responsible for the formation of the cavities (Pozo et al. 2004). The sediments that fill the cavities and where the fossils are embedded, consist of olive marls and fine to coarse siliciclastics. Pozo et al. (2004) suggested that the cavi-ties were filled by the entrance of successive clastic
material contributions probably due to episodic floods.Two types of fossiliferous assemblages (embedded
in a sedimentary matrix of similar lithology) can be distinguished in Cerro de los Batallones (Fig. 2B): 1) lower level assemblages (LLAs) characterized by be-ing embedded in the lower-middle part of the strati-graphic sequence of the butte and having a fossiliferous content dominated by carnivorans remains (%NISP - Number of Specimens Identified per taxon - carnivo-rans in Batallones-1 LLA = 98.32%) and 2) upper level assemblages (ULAs) characterized for being located towards the uppermost part of the butte sequence and having a fossiliferous content that mainly consists of mammalian herbivores remains (%NISP herbivores in Batallones-1 ULA = 80.55%) (morAleS et al. 2008; Domingo et al. 2011). Geophysical surveys carried out in the butte indicate that potentially all the fossil sites are constituted by these two types of assemblages and that they are connected by a pipe-like conduit which results in an hourglass shape of the localities in verti-cal section (Fig. 2B).
Fig. 2. A) Location of Cerro de los Batallones within the Madrid Basin. B) Geological sketch (vertical section) of Batal-lones-1 showing the upper level assemblage (ULA) and lower level assemblage (LLA) (Modified from Pozo et al. 2004 and morAleS et al. 2008). Note that fossils are embedded in sediments that cut the three units defined at the butte. Mining works eliminated most of the upper part of Batallones-1 sequence so we provide here an interpretive reconstruction (dashed lines). The drawing is not to scale.

134 M.S. Domingo et al.
The cavities reached decametric diameters and had a pseudo-circular shape in plan view. The entrance to the cavities would be located in the upper part since geophysical investigation has not detected lateral ex-tension of the sedimentary matrix that fills the cavi-ties (morAleS et al. 2008). Overall dimensions of the cavities are still not known, one of the reasons being that excavation of different localities is still under way. Batallones-1 is an almost exhausted fossil site, howev-er, the destruction of most of its ULA during the min-ing of the sepiolite layer (in Unit II of Fig. 2B) hinders our knowledge of the total dimensions of the locali-ties and of the total depth of the cavities. Still, it can be noted that the total area excavated in Batallones-1 LLA was of around 85 m2 and that in some areas a depth of more than 350 cm was excavated.
3. Material and Methods
This study is based on 6597 remains corresponding to the complete assemblage found in the first exca-vation period (1991-1993) of Batallones-1 LLA (this is ca. 35% of the total large mammal remains found throughout all the field seasons carried out in this lo-cality). All the fossils here studied are housed in the collections of the Museo Nacional de Ciencias Natu-rales-CSIC (Madrid, Spain).
Age-structure and skeletal physical condition were analysed on the four most abundant taxa which are the two sabertoothed cats Promegantereon ogygia and Machairodus aphanistus, the hyaenid Protictitherium crassum and the amphicyonid Magericyon anceps. Two common indexes were used for the quantifica-tion of remain frequencies and individuals: Number of Specimens Identified per taxon, NISP, and Minimum Number of Individuals, MNI (lymAn 1994).
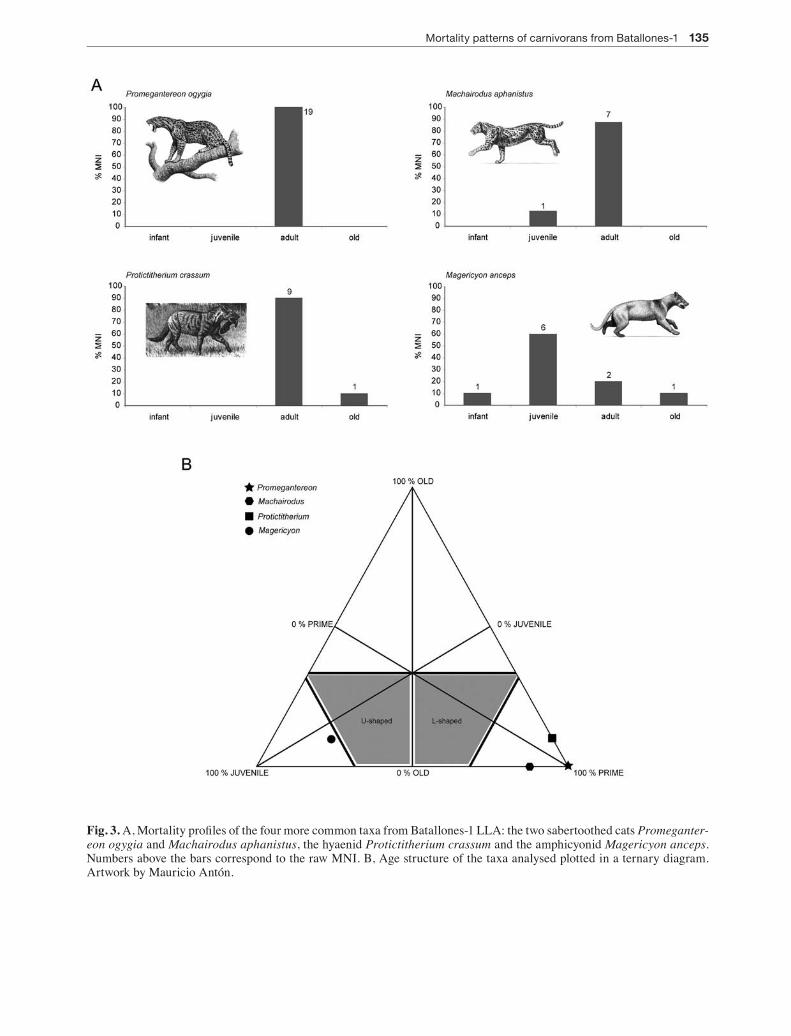
The mortality profiles are estimated based on the eruption and wear stage of in situ mandibular and maxillar teeth. Four qualitative age classes were de-fined: (1) infants, these are individuals with erupting or recently erupted deciduous dentition; (2) juveniles, these are individuals with the deciduous dentition to-tally erupted and sometimes worn and individuals with erupting permanent dentition; (3) adults, these are in-dividuals with complete permanent dentition; dentition shows no wearing or light to moderate wearing; and (4) old, these are individuals with the dentition heavily worn. The age-structure data are represented in his-tograms (Fig. 3A) and in a ternary diagram (Fig. 3B) where the corners represent 100% juveniles (infants and juveniles are here lumped together), 100% prime adult individuals and 100% old individuals (STiner
1990). Statistical analyses consisted in the construc-tion of contingency tables that were analysed using the Fisher’s exact test (significance level: α = 0.01).
4. Ecological features of the analysed carnivorans
Promegantereon ogygia was a puma-sized saber-toothed cat (SAleSA et al. 2006). Its remains represent almost 34% of the large-mammal assemblage from Batallones-1 LLA (NISP = 2231) (Domingo 2009). Another sabertoothed cat, Machairodus aphanistus, is the second more frequent taxa in Batallones-1 LLA making up 24.25% of the sample (NISP = 1600). This sabertoothed cat reached the size of a modern tiger (AnTón et al. 2004; SAleSA et al. 2006). Based on the fact that these two taxa coexisted in the mammalian community that surrounded the cavity, SAleSA et al. (2006) inferred the presence of a wooded environment. In this manner, the sympatry of the two sabertoothed cats would be facilitated in the same fashion modern felids of comparable sizes (e.g., lions and leopards) do nowadays. The most probable prey of Promegantereon ogygia would be juveniles of the equid Hipparion sp. and the suid Microstonyx sp. and adult individuals of cervids and bovids (SAleSA et al. 2006). As for Mach-airodus aphanistus, AnTón et al. (2004) stated that it is possible that horses or antelopes were the largest prey this carnivore could efficiently kill.
Protictitherium crassum was a primitive hyaenid that exhibited a size comparable to the extant African civet, Civettictis civetta (AnTón & morAleS 2000). In Batallones-1 LLA, it is represented by a NISP of 635, making up 9.63% of the large-mammal fossiliferous concentration (Domingo 2009). This hyaenid does not sport a dentition adapted to bone cracking, therefore, its diet and behaviour seem to be more similar to those of an opportunistic carnivoran such as a jackal than to a modern hyaena (AnTón & morAleS 2000; SAleSA et al. 2006).
Magericyon anceps belongs to the extinct family Amphicyionidae and represents ca. 6% of the large-mammal assemblage (NISP = 395). A size compara-ble to a tiger has been inferred for this species. Am-phicyonids’ dentitions were adapted to a carnivorous diet although their strong post-carnassial teeth point to bone-crushing capabilities as well. In the case of Magericyon anceps, Peigné et al. (2008) confirmed that the crushing function was less important in this species than in other amphicyonids, i.e., Magericyon anceps shows hypercarnivorous traits in its dentition that imply a greater consumption of meat.
Mortality patterns of carnivorans from Batallones-1 135
Fig. 3. A, Mortality profiles of the four more common taxa from Batallones-1 LLA: the two sabertoothed cats Promeganter-eon ogygia and Machairodus aphanistus, the hyaenid Protictitherium crassum and the amphicyonid Magericyon anceps. Numbers above the bars correspond to the raw MNI. B, Age structure of the taxa analysed plotted in a ternary diagram. Artwork by Mauricio Antón.
136 M.S. Domingo et al.
5. Results
5.1. Age profiles
The four carnivorans analysed here exhibit different mortality profiles among them that, furthermore, do not match any of the two classic age distribution mod-els. The two sabertoothed cats and the hyaenid exhibit an age structure dominated by the adults (Fig. 3A-B). Most of the analysed dental elements exhibit no or light wear so the age structure of these three carnivoran is mainly made up of young or prime adults. In contrast, the amphicyonid shows a mortality profile dominated by the juveniles (Fig. 3A-B).
5.2. Occurrence of bone pathologies and injuries
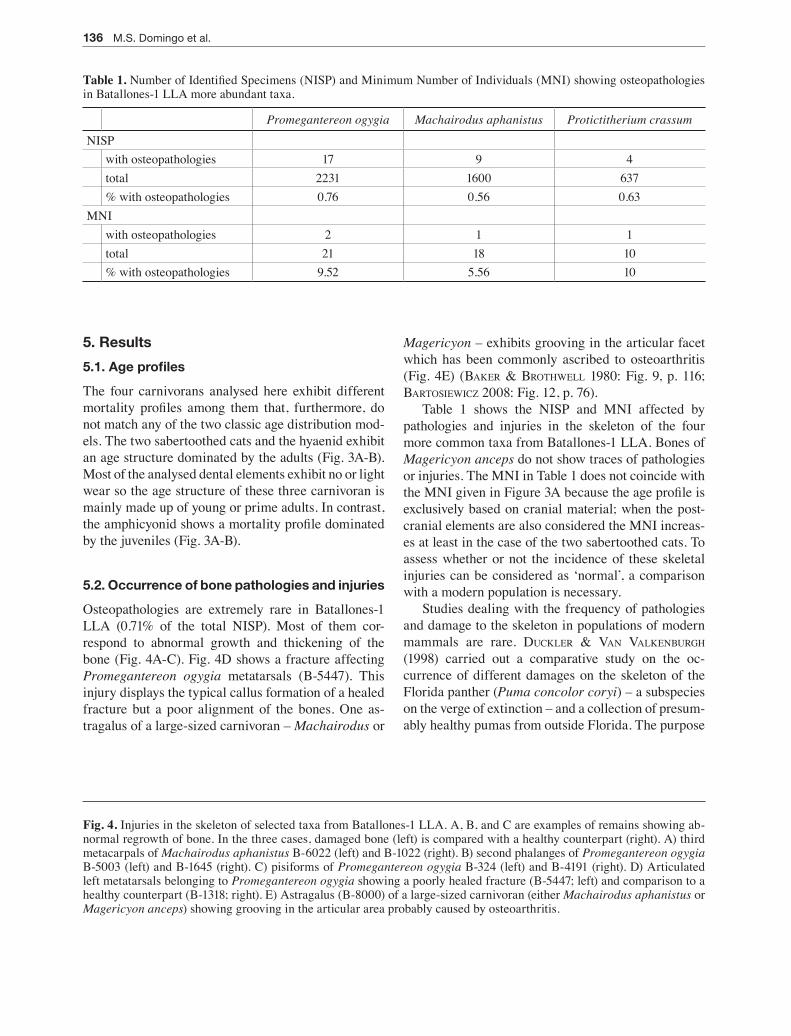
Osteopathologies are extremely rare in Batallones-1 LLA (0.71% of the total NISP). Most of them cor-respond to abnormal growth and thickening of the bone (Fig. 4A-C). Fig. 4D shows a fracture affecting Promegantereon ogygia metatarsals (B-5447). This injury displays the typical callus formation of a healed fracture but a poor alignment of the bones. One as-tragalus of a large-sized carnivoran – Machairodus or
Magericyon – exhibits grooving in the articular facet which has been commonly ascribed to osteoarthritis (Fig. 4E) (bAKer & broThWell 1980: Fig. 9, p. 116; bArToSieWicz 2008: Fig. 12, p. 76).
Table 1 shows the NISP and MNI affected by pathologies and injuries in the skeleton of the four more common taxa from Batallones-1 LLA. Bones of Magericyon anceps do not show traces of pathologies or injuries. The MNI in Table 1 does not coincide with the MNI given in Figure 3A because the age profile is exclusively based on cranial material; when the post-cranial elements are also considered the MNI increas-es at least in the case of the two sabertoothed cats. To assess whether or not the incidence of these skeletal injuries can be considered as ‘normal’, a comparison with a modern population is necessary.
Studies dealing with the frequency of pathologies and damage to the skeleton in populations of modern mammals are rare. DucKler & VAn VAlKenburgh (1998) carried out a comparative study on the oc-currence of different damages on the skeleton of the Florida panther (Puma concolor coryi) – a subspecies on the verge of extinction – and a collection of presum-ably healthy pumas from outside Florida. The purpose
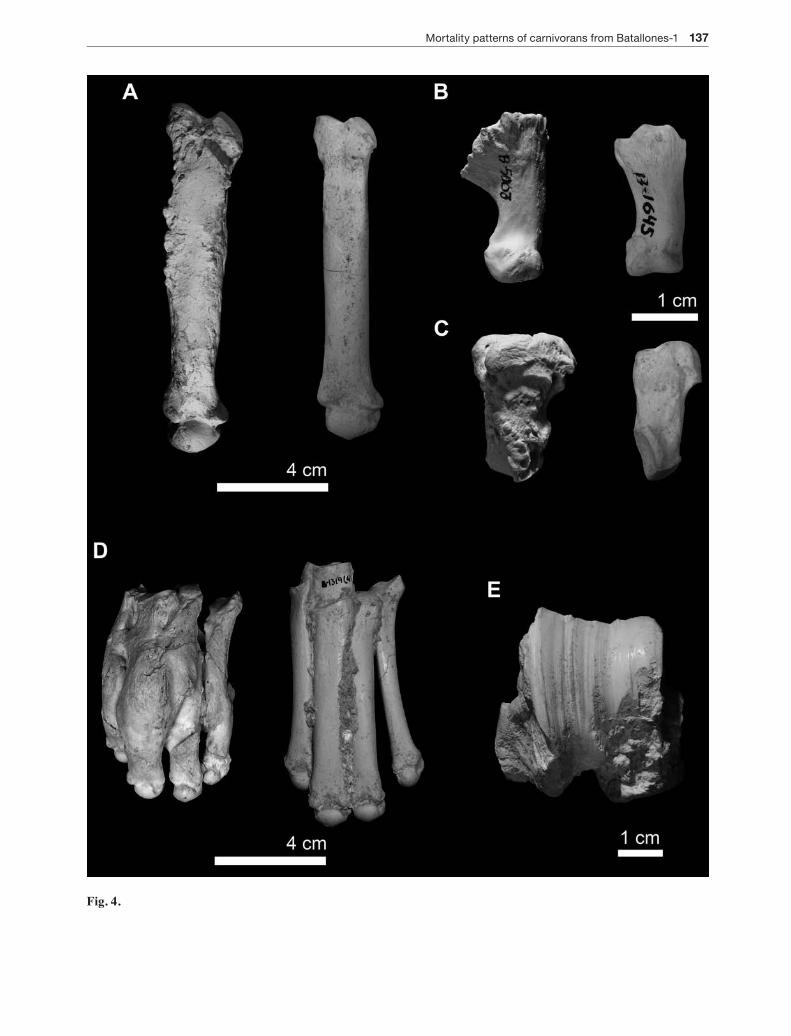
Fig. 4. Injuries in the skeleton of selected taxa from Batallones-1 LLA. A, B, and C are examples of remains showing ab-normal regrowth of bone. In the three cases, damaged bone (left) is compared with a healthy counterpart (right). A) third metacarpals of Machairodus aphanistus B-6022 (left) and B-1022 (right). B) second phalanges of Promegantereon ogygia B-5003 (left) and B-1645 (right). C) pisiforms of Promegantereon ogygia B-324 (left) and B-4191 (right). D) Articulated left metatarsals belonging to Promegantereon ogygia showing a poorly healed fracture (B-5447; left) and comparison to a healthy counterpart (B-1318; right). E) Astragalus (B-8000) of a large-sized carnivoran (either Machairodus aphanistus or Magericyon anceps) showing grooving in the articular area probably caused by osteoarthritis.
Table 1. Number of Identified Specimens (NISP) and Minimum Number of Individuals (MNI) showing osteopathologies in Batallones-1 LLA more abundant taxa.
Promegantereon ogygia Machairodus aphanistus Protictitherium crassum
NISP with osteopathologies 17 9 4 total 2231 1600 637 % with osteopathologies 0.76 0.56 0.63MNI with osteopathologies 2 1 1 total 21 18 10 % with osteopathologies 9.52 5.56 10
Mortality patterns of carnivorans from Batallones-1 137
Fig. 4.
138 M.S. Domingo et al.
of the study was to test the potential incidence of stress on the skeleton of the dwindling population. To our knowledge, this is the only available study that pro-vides data comparable to our data. Therefore, we use the values obtained by DucKler & VAn VAlKenburgh (1998) for the healthy population to check the ‘normal-ity’ of the occurrence of skeletal damage in the car-nivorans from Batallones-1 LLA. Most of the individ-uals from the healthy cougar population exhibit light or moderate dental wear so their age structure can be considered similar to the age structure of the carnivo-rans analysed in Batallones-1 LLA. In our statistical comparison, Harris lines and unknown abnormalities reported by DucKler & VAn VAlKenburgh (1998) are not considered. Rather, our comparison relies on the number of individuals affected by arthritis and frac-tures. The Fisher’s exact test reveals that there are no statistical differences in the occurrence of skeletal in-juries between the three Batallones-1 taxa analysed – Promegantereon ogygia, Machairodus aphanistus and Protictitherium crassum – and the healthy puma population (p = 1, p = 0.63 and p = 1 respectively). Therefore, the number of pathologic and injured indi-viduals from Batallones-1 LLA can be considered as ‘normal’ and inside the limits seen in modern healthy populations.
5.3. Incidence of broken canines in life
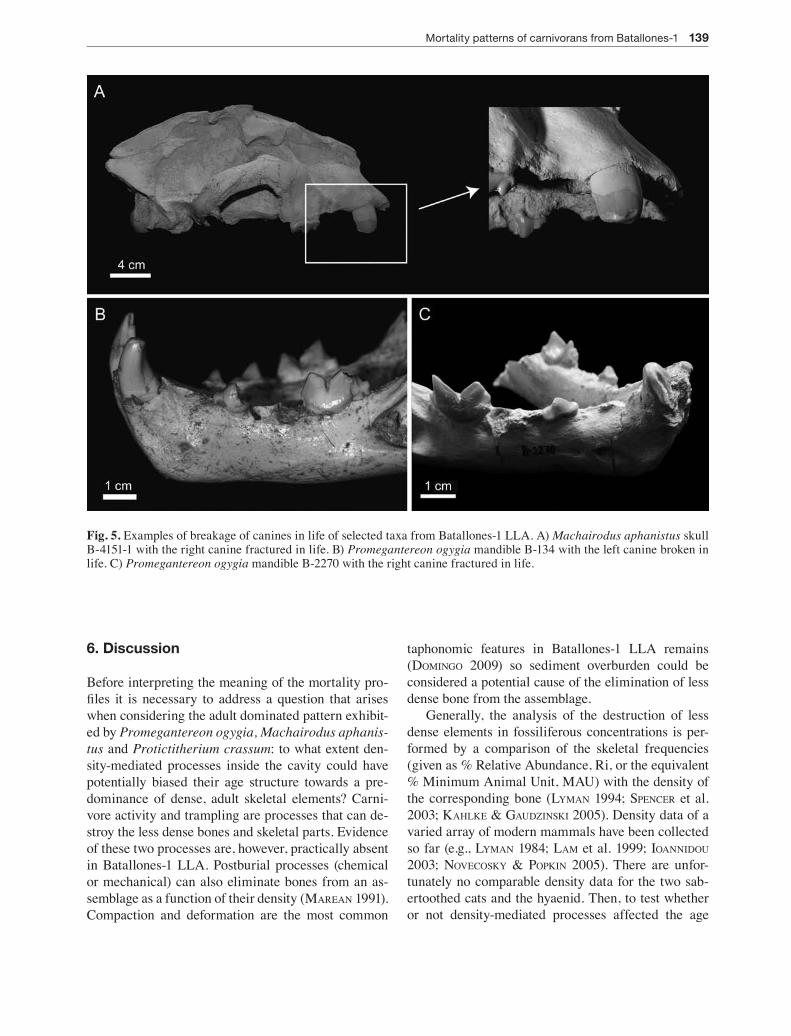
Canine teeth are the dental elements that more fre-quently break in the life of a carnivoran (VAn VAlKen-burgh 1988; VAn VAlKenburgh & herTel 1993). This is likely the reflection of their anterior position in the jaw and their elongate shape which makes them prone to higher bending stresses (VAn VAlKenburgh 1988). Based on this premise – their higher level of breakage –, we consider canines representative of dental frac-ture and, therefore, focus our analysis on this dental element. Teeth broken in life are distinguished by the worn and rounded aspect they show as a consequence of subsequent tooth-tooth or tooth-food contact in life (Fig. 5).
Table 2 quantifies the number of broken canines in life for the more abundant taxa in Batallones-1 LLA. Again, Magericyon anceps did not yield fractured ca-nines. These data were compared with available values in the literature coming from modern counterparts. A mod-ern analogue to Protictitherium crassum would be an opportunistic carnivoran such as the jackal; however, we have not found in the literature suitable data on the
breakage of the canines of jackals to establish a com-parison with this hyaenid. Therefore, our analysis con-centrates on the comparison of the two sabertoothed cats data and data for modern felids. The modern sample was taken from VAn VAlKenburgh & herTel (1993) and DucKler & VAn VAlKenburgh (1998) and includes data on breakage of canines from the lion, Panthera leo, and the cougar, Puma concolor. Again, this comparison is feasible since the age structure of the analysed taxa can be considered as similar.
Table 3 shows the Fisher’s exact test results for the compared sample of felids. There are no significant differences in the frequency of breakage of canines of the extinct vs. the extant taxa. Therefore, in principle, this result indicates that there is not a departure from ‘normality’ (i.e., there is not a higher incidence of bro-ken canines in life) in the sabertoothed cat population from Batallones-1 when compared to modern popula-tions.
Table 3. Fisher’s exact test results on the comparison of the incidence of breakage of canines in life of the two saber-toothed cats from Batallones-1 LLA and of modern felids. Puma concolor data are referred to the healthy non-endan-gered population of DucKler & VAn VAlKenburgh (1998).
Promegantereon ogygia
Machairodus aphanistus Panthera leo Puma
concolorPromegantereon ogygia p = 0.09 p = 1 p = 0.35
Machairodus aphanistus p = 0.06 p = 0.26
Table 2. Frequency of broken canines in life from Batal-lones-1 LLA. Both upper and lower canines are considered in this quantification.
Canines
broken in life
Total canines
% Canines broken in
lifePromegantereonogygia 3 69 4.35
Machairodusaphanistus 4 26 15.38
Protictitheriumcrassum 1 20 5
Mortality patterns of carnivorans from Batallones-1 139
6. Discussion
Before interpreting the meaning of the mortality pro-files it is necessary to address a question that arises when considering the adult dominated pattern exhibit-ed by Promegantereon ogygia, Machairodus aphanis-tus and Protictitherium crassum: to what extent den-sity-mediated processes inside the cavity could have potentially biased their age structure towards a pre-dominance of dense, adult skeletal elements? Carni-vore activity and trampling are processes that can de-stroy the less dense bones and skeletal parts. Evidence of these two processes are, however, practically absent in Batallones-1 LLA. Postburial processes (chemical or mechanical) can also eliminate bones from an as-semblage as a function of their density (mAreAn 1991). Compaction and deformation are the most common
taphonomic features in Batallones-1 LLA remains (Domingo 2009) so sediment overburden could be considered a potential cause of the elimination of less dense bone from the assemblage.
Generally, the analysis of the destruction of less dense elements in fossiliferous concentrations is per-formed by a comparison of the skeletal frequencies (given as % Relative Abundance, Ri, or the equivalent % Minimum Animal Unit, MAU) with the density of the corresponding bone (lymAn 1994; SPencer et al. 2003; KAhlKe & gAuDzinSKi 2005). Density data of a varied array of modern mammals have been collected so far (e.g., lymAn 1984; lAm et al. 1999; ioAnniDou 2003; noVecoSKy & PoPKin 2005). There are unfor-tunately no comparable density data for the two sab-ertoothed cats and the hyaenid. Then, to test whether or not density-mediated processes affected the age
Fig. 5. Examples of breakage of canines in life of selected taxa from Batallones-1 LLA. A) Machairodus aphanistus skull B-4151-1 with the right canine fractured in life. B) Promegantereon ogygia mandible B-134 with the left canine broken in life. C) Promegantereon ogygia mandible B-2270 with the right canine fractured in life.
140 M.S. Domingo et al.
composition of these taxa, we carried out a qualitative analysis based on the comparison of the abundances of high- and low-density long-bone ends. Specifical-ly, humeri and tibiae were surveyed since it has been demonstrated that their distal ends are denser than the proximal ends (mAreAn & ehrhArDT 1995; WolVer-Ton 2006). The MNEs and the % MNEs of the humeri and tibiae normalized to the most abundant portion are shown in Table 4. Complete or almost complete bones (only lacking some bone chips) predominate and only in two cases the denser distal part is preserved more abundantly than the proximal end (humeri of Prome-gantereon ogygia and tibiae of Protictitherium cras-sum). In the rest of the cases, either the proximal por-tion is more abundant than the distal portion or they are evenly preserved (Table 4). Therefore, the lack of juveniles individuals of the two sabertoothed cats and the hyaenid is not a taphonomic artifact. Instead, juve-niles of these taxa were not deposited in the assembla-ge. Besides, the fact that Magericyon anceps is mainly represented by remains belonging to juveniles individ-uals is another evidence to rule out the intervention of density-mediated process in the demographical com-position of Batallones-1 LLA.
In view of the age profiles of the four more com-mon taxa recovered in Batallones-1 LLA, some mor-tality causes can be ruled out and the assemblage for-mation scenario can be better constrained. Since the mortality distribution does not reflect an attritional pattern, causes leading to a preferential death of the
weakest population members (very young and old in-dividuals) must be discarded. Besides, none of the age structures adapt to the L-shaped pattern, therefore, any kind of mass event must be ruled out as the death cause for the individuals represented in the LLA of Batal-lones-1. In any case, the cavity/trap configuration of the fossil site already made it improbable that such a high and diverse amount of carnivorans died suddenly and ended up at the bottom of the cavity. Moreover, the negative Ce anomaly detected in the Rare Earth Ele-ments analysis of Batallones-1 LLA fossils suggested that the existing ponds were oxic and that, therefore, a high number of decomposing carcasses did not oc-cur at the same time ruling out mass mortality events (Domingo et al. 2011).
As previously mentioned, the L-shaped age struc-ture is not only caused by catastrophic mortality. Oth-er non-selective processes such as accidental falls in vertical cavities can produce this kind of profile since, in principle, all the animals from a live population can accidentally fall regardless of their age (KoS 2003). The mortality profiles reported in this study dismiss also the possibility that the animals fell accidentally. Besides, accidental falls would not give as a result a selection towards certain taxonomical groups as is the case in Batallones-1 where carnivorans far outnumber herbivores. From this, it can be inferred that the cavity was fairly visible so accidents were avoided. The pre-dominance of carnivorans would be due to a common attractor for this group of taxa, namely, the presence of
Table 4. Minimum Number of Elements (MNE) and %MNE for complete, proximal, distal and middle shaft sections of long bones (humerus and tibia) that usually display differential preservation.
Promegantereon ogygia Machairodus aphanistus Protictitherium crassum MNE %MNE MNE %MNE MNE %MNE Humerus Complete/almost complete 21 100 9 100 10 100 Proximal 1 4.76 9 100 1 10 Distal 2 9.52 3 33.33 1 10 Only diaphysis 0 0 0 0 0 0 Tibia Complete/almost complete 25 100 8 100 9 100 Proximal 2 8 1 12.50 1 11.11 Distal 2 8 1 12.50 2 22.22 Only diaphysis 0 0 0 0 0 0

Mortality patterns of carnivorans from Batallones-1 141
food (prey or carrion) but also water (probably during drought periods) difficultly accessible for other less agile animals.
An important feature of the fossil site is precisely the contrasting age patterns exhibited by the analysed taxa, with the two sabertoothed cats and the hyaenid showing a profile dominated by adults and the amphi-cyonid showing a profile where infants and juveniles predominate. KAhlKe & gAuzinSKi (2005) found also differing age profiles in their study of the Lower Pleis-tocene fossil assemblage of Untermassfeld (Germany). These authors suggested that the detected differences in mortality were better explained by the ecology and ethology of the taxa rather than by their death mode and this seems to be also the case in Batallones-1 LLA.
As STiner (1990) indicated, prime-dominated pat-tern is a kind of mortality that is not described in the general literature on population biology, however, it has been documented in some fossil assemblages. In her study on mortality patterns in relation to predation in archaeological contexts, STiner (1990) indicates that human hunting would be the only process capable of giving as a result such an age structure. The age of Cerro de los Batallones fossil sites prevents any kind of human action. KAhlKe & gAuzinSKi (2005) docu-mented the case where the remains from one of the cervids from the flood-derived assemblage of Unter-massfeld (Germany) exhibit a mortality profile domi-nated by the prime-adult individuals. The explanation the authors give to such a profile have an ecological/ethological component but also a geological/tapho-nomic component. Although, in general, cervids show a considerable capacity to swim, the lesser physiologi-cal capability of Cervus s.l. nestii vallonnetensis made that all age classes, and not only the weakest indi-viduals, drowned. After this, the heavy flood selected against the lighter and smaller carcasses giving as a result a thanatocoenosis dominated by the prime-adult individuals. In the case of Batallones-1, its trap con-figuration with no inferred lateral entrances makes it improbable that there was a significant loss of infant and juvenile carcasses due to water transport. Besides, the preservation of juvenile remains of Magericyon anceps and of small-sized taxa such as mustelids or lagomorphs evidence that winnowing of those smaller carcasses was improbable. Furthermore, other density-mediated biases on Batallones-1 LLA were previously discarded.
The work of WolVerTon (2006) on the age profiles of black bears in natural traps provide a better analogy to explain the prime-dominated profiles in Batallon-
es-1. In fact, SAleSA et al. (2006) already proposed the same explanation (given below), to explicate the mor-tality pattern of Promegantereon ogygia. A survey of the literature on the demography of modern carnivo-rans such as leopards, cheetahs, bobcats, foxes, wolves or bears reveals that the mortality of young adult in-dividuals is not rare (STorm 1965; bAiley 1974, 1993; PeTerSon et al. 1984; rogerS 1987; STAnDer 1997; Ke-lly et al. 1998; mArKer et al. 2003; obbArD & hoWe 2008). High mortality rates among these individuals are related to their poorer hunting and competitive abilities.
Young males of different types of carnivorans dis-perse in order to establish new territories and this dis-persal is considered to be a stressful period in their lives which can enhance mortality chances. In this context, young adults compete with resident and more experienced adults. Besides, in his study on the Af-rican leopard, bAiley (1993) pointed out that seden-tarism can also cause a preferential mortality of in-experienced females in moments of food shortage in their smaller home ranges. As a consequence, young-adult carnivoran individuals, more desperate in the search for resources, are more likely to enter into un-known habitats or dangerous locations. In this manner, the mortality patterns of the two sabertoothed cats and probably also the hyaena, as shown in the black bear mortality study in natural traps by WolVerTon (2006), could be due to an intentional entry of these inexperi-enced animals, that were in a stressful period of their lives, into a cavity where there would be availability of food or water. Furthermore, based on the absence of juveniles, AnTón & morAleS (2000) and SAleSA et al. (2006) indicate that Promegantereon ogygia (and this is potentially extendable to Machairodus aphanistus) would have a solitary lifestyle in which the cubs are hidden while the female is hunting.
The mortality profile of Magericyon anceps bor-ders on the U-shaped pattern although the very high proportion of infants/juveniles and the very low amount of old individuals makes it improbable that the mortality had an attritional origin. Again the explana-tion for such a mortality pattern can be extracted from the inferred behaviour of this extinct taxon. The high abundance of juveniles and the cavity character of the fossil site could lead to suggest that this taxon used the cave as a den. There are, however, reasons to reject this hypothesis. First, had the cave been easily accessi-ble both to entrance and egress, other carnivores could theoretically have used the cave over time for the same purpose which is not the case in view of the absence

142 M.S. Domingo et al.
of juveniles of other taxa. Besides, dens are charac-terized by containing remains of prey, many of them modified by the activity of the carnivoran (e.g., mAr-eAn & ehrhArDT 1995; DieDrich & zAK 2006; PoKineS & KerbiS PeTerhAnS 2007). These features are not present in Batallones-1 LLA, therefore, the possibility must be ruled out that Batallones-1 LLA could have worked as a den. Rather, we follow interpretations by AnTón & morAleS (2000) who stated that the high percentage of Magericyon anceps juvenile individu-als can be explained if they followed the adults during foraging activities and became trapped in the cavity at the same time. This close bond between cubs and adults, presumably females, may be compared to that observed in extant ursids.
The weakness or strength of the individuals recov-ered in fossil sites does not only depend on their age, but also on their physical condition. This is the reason why an evaluation of the skeletal traumas has been in-cluded in this investigation. The comparison of Batal-lones-1 LLA analysed taxa with modern counterparts suggests that there was not a preferential entrance into the trap of individuals weakened by injuries in their skeleton. It cannot be rejected, however, that those in-dividuals suffering from osteopathologies or broken canines in life got trapped when searching for ‘easy’ food or water. Besides, although not reflected in the fossils, it is highly possible that the trapped prime adults were suffering from bad health as is the case in many young adult carnivorans. In the analysis of the sabertoothed cats’ broken canines, the higher in-cidence of breakage in Machairodus aphanistus com-pared that of Promegantereon ogygia (Table 2) might be related to the fact that two Machairodus aphanistus individuals exhibit a moderately worn adult dentition while none of the Promegantereon individuals display this more-advanced wear stage (i.e., all the Promega-tereon individuals show non or light wear). SAleSA et al. (2006) suggested that, although Promegantereon ogygia was a solitary cat, it developed some degree of tolerance between adults as jaguars do nowadays. Then, alternatively, the lower degree of canine break-age in Promegantereon ogygia compared to Machai-rodus aphanistus can be related to the fact that indi-viduals with broken canines of the former species fed more easily in the territory and on the remains left by other individuals. Despite the absence of further de-tailed information on the social behaviour of Machai-rodus aphanistus, the higher amount of Machairodus aphanistus’ individuals with broken canines in life could be pointing towards a lower degree of tolerance
in this larger-sized sabertoothed cat and, thus, a higher probability of looking for food in the cavity.
The absence of skeletal damage in Magericyon anceps skeleton can be explained by the fact that this taxon is mainly represented by juveniles which are less prone to have suffered injuries in their shorter lifetime. This study demonstrates the importance of the analysis of the demography of the taxa recovered in a fossil site in order to constrain the reasons leading to the composition of an assemblage. In this case, the behaviour of the taxa, and not a particular death mode of a population (e.g. catastrophe, predation, diseases of the weakest age classes, accidents etc.), is a major factor in determining the age structures recorded in Batallones-1 LLA. These mortality patterns, used in combination with other taphonomic and geologic lines of evidence, will help to construct a definitive forma-tion scenario for this fossiliferous locality.
7. Conclusions
The construction of mortality profiles from the taxa represented in a fossil site is shown to be a useful tool in understanding the taphonomic histories and re-stricting the formation scenario of a locality. In this particular case, we have analysed the age structure of the four more common taxa from the carnivore-dom-inated Miocene site of Batallones-1 LLA. An impor-tant feature is that the studied taxa exhibit contrasting age patterns. On the one hand, the two sabertoothed cats and the hyaenid show mortality profiles domi-nated by prime adults. On the other hand, juveniles prevail in the amphicyonid age profile. None of these mortality patterns adapt to any of the two classically proposed models (attritional and catastrophic). Ac-cordingly, events leading to the death of the, in prin-ciple, weaker age classes and also mass deaths have been discarded. Besides, L-shaped profile can be the result of other non-selective processes such as acciden-tal falls into a cavity. The absence of an L-shaped pat-tern together with the low presence of herbivore taxa suggest that animals were not dying in the cavity as a result of accidental falls; rather the entrance would be well visible. In Batallones-1 LLA, the causes behind the mortality profiles seems to be more related to the ecology and ethology of the represented taxa than to the death mode.
The prime-adult dominated mortality pattern shown by Promegantereon ogygia, Machairodus aph-anistus and Protictitherium crassum would respond to a deliberate entrance into the cavity of individuals that

Mortality patterns of carnivorans from Batallones-1 143
are struggling due to their inexperience in obtaining resources. Stress produced during dispersal, but also sedentarism in the case of females, can take carni-vores to search for resources in dangerous places. The Magericyon anceps mortality profile can be the result of the incorporation into the cavity of females with their cubs. Once the carnivorans were at the bottom of the cave, they would have been unable to escape.
In this study, we also have investigated the skeletal physical condition of the taxa by quantifying the inci-dence of injuries in the bones and the amount of broken canines in life. The comparison of these variables with modern and healthy populations implies that Batal-lones-1 LLA taxa do not show a significantly elevated occurrence of damage in bones and teeth. Therefore, it can be concluded that there was not a preferential entrapment of individuals having poor skeletal health.
Acknowledgements
We thank mAuricio AnTón for letting us use his illustrations of Cerro de los Batallones carnivorans. Comments provided by lAurA Domingo, JAmeS nebelSicK and one anonymous reviewer improved the manuscript. JAmeS nebelSicK is also acknowledged for his editorial work. This study was funded by the Spanish Government Ministry of Science and In-novation research projects CGL2008-05813-C02-01/BTE, CGL2007-60790/BTE, and CGL2010-19116/BOS and is included in the CAM-UCM 910607 Research Group. MSD was supported by the Fundación Española para la Ciencia y la Tecnología (FECYT) and the Spanish Ministry of Educa-tion Postdoctoral Fellowship Programme.
References
AnTón, M. & morAleS, J. (2000): Inferencias paleoecoló-gicas de la asociación de carnívoros del yacimiento de Cerro Batallones. – In: morAleS, J. (Ed.): Patrimonio Paleontológico de la Comunidad de Madrid. Arqueo-logía, Paleontología and Etnografía, monográfico, 6: 190-201; Madrid (Serie de la Consejería de Educación Comunidad de Madrid).
AnTón, M., SAleSA, M.J., morAleS, J. & Turner, A. (2004): First known complete skulls of the scimitar-toothed cat Machairodus aphanistus (Felidae, Carnivora) from the Spanish Late Miocene site of Batallones-1. – Journal of Vertebrate Paleontology, 24: 957-969.
ArgenTi, P. & mAzzA, P.P.A. (2006): Mortality analysis of the Late Pleistocene bears from Grotta Lattaia, central Italy. – Journal of Archaeological Science, 33: 1552-1558.
bAiley, T.N. (1974): Social organization in a bobcat popu-lation. – Journal of Wildlife Management, 38: 435-446.
bAiley, T.N. (1993): The African leopard. Ecology and be-haviour of a solitary felid. – 429 pp; New York (Colum-bia University Press).
bAKer, J. & broThWell, D. (1980): Animal diseases in Ar-chaeology. – 235 pp; London (Academic Press).
bArToSieWicz, L. (2008): Taphonomy and palaeopathology in archaeozoology. – Geobios, 41: 69-77.
DieDrich, C.G. & záK, K. (2006): Prey deposits and den sites of the Upper Pleistocene hyena Crocuta crocuta spelaea (Goldfuss, 1823) in horizontal and vertical ca-ves of the Bohemian Karst (Czech Republic). – Bulletin of Geosciences, 81: 237-276.
Domingo, M.S. (2009): Estudio tafonómico del yacimien-to de vertebrados vallesienses (Mioceno Superior) de Batallones 1, Torrejón de Velasco, Madrid. – 507 pp; Unpublished Ph.D. dissertation, Madrid (Universidad Complutense de Madrid).
Domingo, M.S., AlberDi, M.T. & AzAnzA, B. (2007): A new quantitative biochronological ordination for the Upper Neogene mammalian localities of Spain. – Palaeogeog-raphy, Palaeoclimatology, Palaeoecology, 255: 361–376.
Domingo, M.S., Domingo, L., Sánchez, I.M., AlberDi, M.T., AzAnzA, B. & morAleS, J. (2011): New insights on the taphonomy of the exceptional mammalian fossil sites of Cerro de los Batallones (Late Miocene, Spain) based on Rare Earth Element geochemistry. – Palaios, 26: 55-65.
DucKler, G.L. & VAn VAlKenburgh, B. (1998): Osteologi-cal corroboration of pathological stress in a population of endangered Florida pumas (Puma concolor coryi). – Animal Conservation, 1: 39-46.
eberTh, D.A., rogerS, R.R. & Fiorillo, A.R. (2007): A practical approach to the study of bonebeds. – In: ro-gerS, R.R., eberTh, D.A. & Fiorillo, A.R. (Eds.): Bone-beds. Genesis, Analysis, and Paleobiological Significan-ce. 265-331; Chicago (The University of Chicago Press).
ioAnniDou, E. (2003): Taphonomy of animal bones: species, sex, age and breed variability of sheep, cattle and pig bone density. – Journal of Archaeological Science, 30: 355-365.
KAhlKe, R.-D. & gAuDzinSKi, S. (2005): The blessing of a great flood: differentiation of mortality patterns in the large mammal record of the Lower Pleistocene fluvial site of Untermassfeld (Germany) and its relevance for the interpretation of faunal assemblages from archaeo-logical sites. – Journal of Archaeological Science, 32: 1202-1222.
Kelly, M.J., lAurenSon, M.K., FiTzgibbon, C.D., collinS, D.A., DurAnT, S.M., FrAme, G.W., berTrAm, B.C.R. & cAro, T.M. (1998): Demography of the Serengeti chee-tah (Acinonyx jubatus) population: the first 25 years. – Journal of Zoology, 244: 473-488.
KoS, A.M. (2003): Pre-burial taphonomic characterisation of a vertebrate assemblage from a pitfall cave fossil de-posit in southeastern Australia. – Journal of Archaeolo-gical Science, 30: 769-779.
KurTén, B. (1953): On the variation and population dyna-mics of fossil and recent mammal populations. – Acta Zoologica Fennica, 76: 1-122.
KurTén, B. (1958): Life and death of the Pleistocene cave bear. – Acta Zoologica Fennica, 95: 1-59.
lAm, Y.M., chen, X. & PeArSon, O.M. (1999): Intertaxo-nomic variability in patterns of bone density and the differential representation of bovid, cervid, and equid elements in the archaeological record. – American An-

144 M.S. Domingo et al.
tiquity, 64: 343-362.lAuTerS, P., boloTSKy, Y.L., VAn iTTerbeecK, J. & goDe-
FroiT, P. (2008): Taphonomy and age profile of a latest Cretaceous dinosaur bone bed in far Eastern Russia. – Palaios, 23: 153-162.
lóPez-AnToñAnzAS, R., Peláez-cAmPomAneS, P., álVArez-SierrA, M.A. & gArcíA PAreDeS, I. (2010): New species of Hispanomys (Rodentia, Cricetodontinae) from the Upper Miocene of Batallones (Madrid, Spain). – Zoo-logical Journal of the Linnean Society, 160: 725-747.
lymAn, R.L. (1984): Bone density and differential survi-vorship of fossil classes. – Journal of Anthropological Archaeology, 3: 259-299.
lymAn, R.L. (1994): Vertebrate Taphonomy. – 524 pp; Cam-bridge (Cambridge University Press).
mAreAn, C.W. (1991): Measuring the post-depositional des-truction of bone in archaeological assemblages. – Jour-nal of Archaeological Science, 18: 677-694.
mAreAn, C.W. & ehrhArDT, C.L. (1995): Paleoanthropo-logical and paleoecological implications of the tapho-nomy of a sabertooth’s den. – Journal of Human Evolu-tion, 29: 515-547.
mArKer, L.L., DicKmAn, A.J., Jeo, R.M., millS, M.G.L. & mAcDonAlD, D.W. (2003): Demography of the Namibian cheetah, Acinonyx jubatus jubatus. – Biological Con-servation, 114: 413-425.
morAleS, J., cAPiTán, J., cAlVo, J.P. & SeSé, c. (1992): Nue-vo yacimiento de vertebrados del Mioceno Superior al sur de Madrid (Cerro Batallones, Torrejón de Velasco). – Geogaceta, 12, 77-80.
morAleS, J., AlcAlá, L., AmezuA, L., AnTón, M., FrAile, S., gómez, E., monToyA, P., nieTo, M., Pérez, B., SA-leSA, M.J. & Sánchez, I.M. (2000): El yacimiento del Cerro de los Batallones. – In: morAleS, J. (Ed.): Patri-monio Paleontológico de la Comunidad de Madrid. Ar-queología, Paleontología and Etnografía, monográfico 6. 179-190; Madrid (Serie de la Consejería de Educación Comunidad de Madrid).
morAleS, J., AlcAlá, L., álVArez SierrA, M.A., AnTón, M., AzAnzA, B., cAlVo, J.P., cArrASco, P., FrAile, S., gAr-cíA PAreDeS, I., gómez, E., hernánDez FernánDez, M., merino, L., meulen, A. VAn Der, mArTín eScorzA, C., monToyA, P., nieTo, M., Peigné, S., Pérez, B., Peláez-cAmPomAneS, P., Pozo, M., QuirAlTe, V., SAleSA, M.J., Sánchez, I.M., Sánchez mArco, A., SilVA, P.G., SoriA, M.D. & Turner, A. (2004): Paleontología del sistema de yacimientos de mamíferos miocenos del Cerro de los Batallones, Cuenca de Madrid. – Geogaceta, 35: 139–142.
morAleS, J., Pozo, M., SilVA, P.G., Domingo, M.S., lóPez-AnToñAnzAS, R., álVArez SierrA, M.A., AnTón, M., mArTín eScorzA, C., QuirAlTe, V., SAleSA, M.J., Sán-chez, I.M., AzAnzA, B., cAlVo, J.P., cArrASco, P., gAr-cíA-PAreDeS, I., Knoll, F., hernánDez FernánDez, M., hoeK oSTenDe, L. VAn Der, merino, L., meulen, A.J. VAn Der, monToyA, P., Peigné, S., Peláez-cAmPomAneS, P., Sánchez-mArco, A., Turner, A., AbellA, J., AlcAl-De, G.M., AnDréS, M., Demiguel, D., cAnTAlAPieDrA, J.L., FrAile, S., gArcíA yelo, B.A., gómez cAno, A.R., lóPez guerrero, P., oliVer Pérez, A. & Siliceo, G. (2008): El sistema de yacimientos de mamíferos mio-
cenos del Cerro de los Batallones, Cuenca de Madrid: estado actual y perspectivas. – Paleontologica Nova SE-PAZ, 8: 41–117.
noVecoSKy, B.J. & PoPKin, P.R.W. (2005): Canidae volume bone mineral density values: an application to sites in western Canada. – Journal of Archaeological Science, 32: 1677-1690.
obbArD, M.E. & hoWe, E.J. (2008): Demography of the black bears in hunted and unhunted areas of the boreal forest of Ontario. – Journal of Wildlife Management, 72: 869-880.
Peigné, S., SAleSA, M.J., AnTón, M. & morAleS, J. (2008): A new Amphicyonine (Carnivora: Amphicyonidae) from the Upper Miocene of Batallones-1, Madrid, Spain. – Palaeontology, 51: 943-965.
PeTerSon, R.O., WoolingTon, J.D. & bAiley, T.N. (1984): Wolves of the Kenai Peninsula, Alaska. – Wildlife Mo-nographs, 88: 3-52.
PoKineS, J.T. & KerbiS PeTerhAnS, J.C. (2007): Spotted hye-na (Crocuta crocuta) den use and taphonomy in the Ma-sai Mara National Reserve. – Journal of Archaeological Science, 34: 1914-1931.
Pozo, M., cAlVo, J.P., SilVA, P.G., morAleS, J., Peláez-cAm-PomAneS, P. & nieTo, M. (2004): Geología del sistema de yacimientos de mamíferos miocenos del Cerro de los Batallones, Cuenca de Madrid. – Geogaceta, 35: 143-146.
rogerS, L.L. (1987): Effects of food supply and kinship on social behavior, movements, and population dynamics of black bears in northeastern Minnesota. – Wildlife Monographs, 97: 1-72.
SAleSA, M.J., AnTón, M., Turner, A. & morAleS, J. (2006): Inferred behaviour and ecology of the primitive sabre-toothed cat Paramachairodus ogygia (Felidae, Machai-rodontinae) from the late Miocene from Spain. – Jour-nal of Zoology, 268: 243-254.
SPencer, L.M., VAn VAlKenburgh, B. & hArriS, J.M. (2003): Taphonomic analysis of large mammals reco-vered from the Pleistocene Rancho La Brea tar seeps. – Paleobiology, 29: 561-575.
STAnDer, P.E. (1997): Field age determination of leopards by tooth wear. – African Journal of Ecology, 35: 156-161.
STeele, T.E. (2003): Using mortality profiles to infer beha-vior in the fossil record. – Journal of Mammalogy, 84: 418-430.
STeele, T.E. & WeAVer, T.D. (2002): The modified trian-gular graph: a refined method for comparing mortali-ty profiles in archaeological samples. – Journal of Ar-chaeological Science, 29: 317-322.
STiner, M.C. (1990): The use of mortality patterns in ar-chaeological studies of hominid predatory adaptations. – Journal of Anthropological Archaeology, 9: 305-351.
STorm, G.L. (1965): Movements and activities of foxes as determined by radio-tracking. – The Journal of Wildlife Management, 29: 1-13.
VAn VAlKenburgh, B. (1988): Incidence of tooth breakage among large, predatory mammals. – The American Na-turalist, 131: 291-302.
VAn VAlKenburgh, B. & herTel, F. (1993): Tough times at La Brea: tooth breakage in large carnivores of the Late

Mortality patterns of carnivorans from Batallones-1 145
Pleistocene. – Science, 261: 456-459.VoorhieS, M.R. (1969): Taphonomy and population dyna-
mics of an early Pliocene vertebrate fauna, Knox Coun-ty, Nebraska. – Contributions to Geology, University of Wyoming, Special Paper, 1: 1-69.
WolVerTon, S. (2006): Natural-Trap ursid mortality and the Kurtén Response. – Journal of Human Evolution, 50: 540-551.
Manuscript received: October 13th, 2011. Revised version accepted by the Tübingen editor: April 4th, 2012.
Addresses of the authors:
m. SoleDAD Domingo, Museum of Paleontology, Univer-sity of Michigan, Ann Arbor, MI 48109, USA; e-mail: [email protected];m. TereSA AlberDi & Jorge morAleS, Departamento de Paleobiología, Museo Nacional de Ciencias Naturales-CSIC, 28006, Madrid, Spain;beATriz AzAnzA, Departamento de Ciencias de la Tierra, Facultad Ciencias, Universidad de Zaragoza, 50009, Zara-goza, Spain.





















