
Morphological and electrophysiological characterization of abnormal cell types in pediatric cortical...
15
Morphological and Electrophysiological Characterization of Abnormal Cell Types in Pediatric Cortical Dysplasia Carlos Cepeda, 1 Raymond S. Hurst, 5 Jorge Flores-Herna ´ndez, 1 Elizabeth Herna ´ndez-Echeagaray, 1 Gloria J. Klapstein, 1 Marea K. Boylan, 1 Christopher R. Calvert, 1 Emily L. Jocoy, 1 Oanh K. Nguyen, 1 Ve ´ronique M. Andre ´, 1,2 Harry V. Vinters, 1,3 Marjorie A. Ariano, 4 Michael S. Levine, 1 * and Gary W. Mathern 1,2 1 Mental Retardation Research Center, University of California, Los Angeles, California 2 Department of Neurosurgery, University of California, Los Angeles, California 3 Department of Pathology and Laboratory Medicine, University of California, Los Angeles, California 4 Department of Neuroscience, Chicago Medical School, North Chicago, Illinois 5 Department of Pharmacology, Pharmacia Corp., Kalamazoo, Michigan The mechanisms responsible for seizure generation in cortical dysplasia (CD) are unknown, but morphologically abnormal cells could contribute. We examined the pas- sive and active membrane properties of cells from pedi- atric CD in vitro. Normal- and abnormal-appearing cells were identified morphologically by using infrared video- microscopy and biocytin in slices from children with mild to severe CD. Electrophysiological properties were as- sessed with patch clamp recordings. Four groups of abnormal-appearing cells were observed. The first con- sisted of large, pyramidal cells probably corresponding to cytomegalic neurons. Under conditions that reduced the contribution of K conductances, these cells gener- ated large Ca 2 currents and influx when depolarized. When these cells were acutely dissociated, peak Ca 2 currents and densities were greater in cytomegalic compared with normal-appearing pyramidal neurons. The second group included large, nonpyramidal cells with atypical somatodendritic morphology that could correspond to “balloon” cells. These cells did not dis- play active voltage- or ligand-gated currents and did not appear to receive synaptic inputs. The third group included misoriented and dysmorphic pyramidal neu- rons, and the fourth group consisted of immature- looking pyramidal neurons. Electrophysiologically, neurons in these latter two groups did not display significant abnormalities when compared with normal- appearing pyramidal neurons. We conclude that there are cells with abnormal intrinsic membrane properties in pediatric CD. Among the four groups of cells, the most abnormal electrophysiological properties were displayed by cytomegalic neurons and large cells with atypical morphology. Cytomegalic neurons could play an important role in the generation of epileptic activity. © 2003 Wiley-Liss, Inc. Key words: cortical dysplasia; epilepsy; cytomegalic neurons; electrophysiology; calcium imaging Focal cortical dysplasia (CD; Taylor et al., 1971) is a malformation that constitutes one of the leading causes of pediatric epilepsy (Mischel et al., 1995; Mathern et al., 1999; Schwartzkroin and Walsh, 2000; Crino et al., 2002). CDs are disorders of neuronal migration and/or cell dif- ferentiation characterized by cortical dyslamination; ec- topic neurons within white matter; and, in severe cases, the presence of cytomegalic neurons and “balloon” cells, without clear neuronal or glial differentiation, in the af- fected areas (Farrell et al., 1992; Vinters et al., 1992). It has been hypothesized that these abnormal cells could partic- ipate in the generation of epileptic activity (Spreafico et al., 1998; Kerfoot et al., 1999). Their potential as an epileptogenic substrate has been supported by the demon- stration that cytomegalic neurons express abundant gluta- mate receptor subunits (Kerfoot et al., 1999) and that these subunits are differentially coassembled (Ying et al., 1998). In addition, focal CD tissue maintained in vitro generates ictal-like activity when exposed to 4-aminopyridine, a K channel blocker (Mattia et al., 1995; Avoli et al., 1999, 2002). Previous attempts to identify cellular abnormalities in pediatric cortical tissue samples in vitro have been gener- ally unsuccessful (Dudek et al., 1995; Tasker et al., 1996). In our own studies with Lucifer yellow, an intracellular marker, we found that about 20% of labeled cells appeared abnormal (Cepeda et al., 1993). However, a correlation between abnormal morphology and cellular electrophysi- Contract grant sponsor: USPHS; Contract grant number: NS38992. *Correspondence to: Michael S. Levine, PhD, Mental Retardation Re- search Center, 760 Westwood Plaza NPI 58-258, University of California at Los Angeles, Los Angeles, CA 90095. E-mail: [email protected] Received 26 January 2003; Revised 13 February 2003; Accepted 17 Feb- ruary 2003 Journal of Neuroscience Research 72:472– 486 (2003) © 2003 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Morphological and electrophysiological characterization of abnormal cell types in pediatric cortical...

Morphological and ElectrophysiologicalCharacterization of Abnormal Cell Types inPediatric Cortical Dysplasia
Carlos Cepeda,1 Raymond S. Hurst,5 Jorge Flores-Hernandez,1
Elizabeth Hernandez-Echeagaray,1 Gloria J. Klapstein,1 Marea K. Boylan,1
Christopher R. Calvert,1 Emily L. Jocoy,1 Oanh K. Nguyen,1 Veronique M. Andre,1,2
Harry V. Vinters,1,3 Marjorie A. Ariano,4 Michael S. Levine,1* and Gary W. Mathern1,2
1Mental Retardation Research Center, University of California, Los Angeles, California2Department of Neurosurgery, University of California, Los Angeles, California3Department of Pathology and Laboratory Medicine, University of California, Los Angeles, California4Department of Neuroscience, Chicago Medical School, North Chicago, Illinois5Department of Pharmacology, Pharmacia Corp., Kalamazoo, Michigan
The mechanisms responsible for seizure generation incortical dysplasia (CD) are unknown, but morphologicallyabnormal cells could contribute. We examined the pas-sive and active membrane properties of cells from pedi-atric CD in vitro. Normal- and abnormal-appearing cellswere identified morphologically by using infrared video-microscopy and biocytin in slices from children with mildto severe CD. Electrophysiological properties were as-sessed with patch clamp recordings. Four groups ofabnormal-appearing cells were observed. The first con-sisted of large, pyramidal cells probably correspondingto cytomegalic neurons. Under conditions that reducedthe contribution of K� conductances, these cells gener-ated large Ca2� currents and influx when depolarized.When these cells were acutely dissociated, peak Ca2�
currents and densities were greater in cytomegaliccompared with normal-appearing pyramidal neurons.The second group included large, nonpyramidal cellswith atypical somatodendritic morphology that couldcorrespond to “balloon” cells. These cells did not dis-play active voltage- or ligand-gated currents and didnot appear to receive synaptic inputs. The third groupincluded misoriented and dysmorphic pyramidal neu-rons, and the fourth group consisted of immature-looking pyramidal neurons. Electrophysiologically,neurons in these latter two groups did not displaysignificant abnormalities when compared with normal-appearing pyramidal neurons. We conclude that thereare cells with abnormal intrinsic membrane propertiesin pediatric CD. Among the four groups of cells, themost abnormal electrophysiological properties weredisplayed by cytomegalic neurons and large cells withatypical morphology. Cytomegalic neurons could playan important role in the generation of epileptic activity.© 2003 Wiley-Liss, Inc.
Key words: cortical dysplasia; epilepsy; cytomegalicneurons; electrophysiology; calcium imaging
Focal cortical dysplasia (CD; Taylor et al., 1971) is amalformation that constitutes one of the leading causes ofpediatric epilepsy (Mischel et al., 1995; Mathern et al.,1999; Schwartzkroin and Walsh, 2000; Crino et al., 2002).CDs are disorders of neuronal migration and/or cell dif-ferentiation characterized by cortical dyslamination; ec-topic neurons within white matter; and, in severe cases,the presence of cytomegalic neurons and “balloon” cells,without clear neuronal or glial differentiation, in the af-fected areas (Farrell et al., 1992; Vinters et al., 1992). It hasbeen hypothesized that these abnormal cells could partic-ipate in the generation of epileptic activity (Spreafico etal., 1998; Kerfoot et al., 1999). Their potential as anepileptogenic substrate has been supported by the demon-stration that cytomegalic neurons express abundant gluta-mate receptor subunits (Kerfoot et al., 1999) and that thesesubunits are differentially coassembled (Ying et al., 1998).In addition, focal CD tissue maintained in vitro generatesictal-like activity when exposed to 4-aminopyridine, a K�
channel blocker (Mattia et al., 1995; Avoli et al., 1999,2002).
Previous attempts to identify cellular abnormalities inpediatric cortical tissue samples in vitro have been gener-ally unsuccessful (Dudek et al., 1995; Tasker et al., 1996).In our own studies with Lucifer yellow, an intracellularmarker, we found that about 20% of labeled cells appearedabnormal (Cepeda et al., 1993). However, a correlationbetween abnormal morphology and cellular electrophysi-
Contract grant sponsor: USPHS; Contract grant number: NS38992.
*Correspondence to: Michael S. Levine, PhD, Mental Retardation Re-search Center, 760 Westwood Plaza NPI 58-258, University of Californiaat Los Angeles, Los Angeles, CA 90095.E-mail: [email protected]
Received 26 January 2003; Revised 13 February 2003; Accepted 17 Feb-ruary 2003
Journal of Neuroscience Research 72:472–486 (2003)
© 2003 Wiley-Liss, Inc.

ological properties could not be established, in part be-cause of the heterogeneity of the sample population thatcontained CD and non-CD etiologies. Although insightshave been obtained from animal models of CD (Roper etal., 1995; Jacobs et al., 1996; Lee et al., 1997; Sancini et al.,1998), these models have failed to reproduce the cytome-galic neurons and “balloon” cells associated with the hu-man disease (Schwartzkroin and Walsh, 2000).
Consequently, there is almost no electrophysiologi-cal information about the passive and active membraneproperties of abnormal cells from human CD. Failure toidentify abnormal cells in vitro could be partially explainedby the fact that, until recently, visualization of recordedcells in slices was not possible. Infrared videomicroscopyand differential interference contrast (IR-DIC) optics al-low cell visualization in slices, particularly in young tissuesamples (Cepeda et al., 1999; Mathern et al., 2000). Usingthis technique, in combination with whole-cell patchclamp recordings and biocytin labeling, we provide acorrelative characterization of normal- and abnormal-appearing cells in pediatric CD tissue, based on a numberof morphological and electrophysiological properties. Weexamined, in particular, isolated Ca2� conductances, be-cause they have been shown to have a role in the gener-ation of epileptic activity (Speckmann et al., 1993). Thepresent study, which spans 5 years of data collection,characterizes abnormal cells from a relatively homoge-neous pathological population of young CD patients.
MATERIALS AND METHODS
Clinical Population and Presurgical Evaluation
The cohort consisted of 37 patients with CD and intrac-table seizures operated on at the University of California at LosAngeles (UCLA) from 1997 to 2002 (Mathern et al., 1999).Informed consent for tissue sample use for research was obtainedfrom the parents. The protocol was approved by the reviewboard of the Human Protection Research Committee at UCLA.Presurgical evaluation included clinical history, neurological ex-aminations, interictal and ictal scalp video/electroencephalography(EEG) telemetry, and neurodevelopment assessments. Neuro-imaging studies included high-resolution magnetic resonanceimaging (MRI) and 18fluoro-2-deoxyglucose positron emissiontomography (PET). The epileptogenic region for surgical resec-tion was anatomically defined based on convergent information.At surgery, electrocorticography (ECoG) further defined thebrain regions to be removed based on background slowing,location of interictal spikes and polyspikes, and ictal discharges.
Selection of Neocortical Samples for Electrophysiology
When possible, two neocortical sites per patient wereremoved for in vitro electrophysiology, representing the mostabnormal (MA) and least abnormal (LA) regions based on neu-roimaging and ECoG studies. MA samples were regions show-ing thickened cortical gyri on MRI, the greatest hypometabo-lism, and the most abnormal ECoG. LA samples were fromregions with the fewest neuroimaging and ECoG abnormalitiesbut still part of the planned surgical resection. Sample sites wereremoved microsurgically (�1.5–2.0 cm round and deep) and
directly placed in artificial cerebrospinal fluid (ACSF). CD at thetwo sample sites was confirmed by routine histopathology fromadjacent regions examined by the neuropathologist and thedegree of CD (severe or mild) classified as previously published(Mischel et al., 1995).
Electrophysiological Recordings
Immediately after resection, brain samples were placed incold oxygenated ACSF containing (in mM) NaCl 130,NaHCO3 26, KCl 3, MgCl2 5, NaH2PO4 1.25, CaCl2 1.0,glucose 10 (pH 7.2–7.4). Within 5–10 min of surgical removal,slices (350 �m) were cut (Microslicer; DSK model 1500E) andplaced in ACSF for at least 1 hr (in this solution CaCl2 wasincreased to 2 mM, and MgCl2 was decreased to 2 mM). Sliceswere constantly oxygenated with 95% O2-5% CO2 (pH 7.2–7.4, osmolality 290–300 mOsm, 32°C � 2°C). After incuba-tion, tissue sections were transferred to a custom-designedchamber attached to the fixed stage of an upright microscope(Zeiss Axioskop or Olympus model BX50WI). The slice washeld down with thin nylon threads glued to a platinum wire andsubmerged in continuously flowing oxygenated ACSF (25°C) at4 ml/min. Slices were illuminated with near-IR light (790 nm;Ealing Optics, Hollston, MA). Individual cells were visualizedwith a 40� water-immersion objective by using DIC optics.The image was detected with an IR-sensitive CCD camera(C2400; Hamamatsu, Tokyo, Japan).
Patch electrodes (3–6 M�) were filled with the followinginternal solution (in mM): Cs-methanesulfonate 125, NaCl 4,KCl 3, MgCl2 1, MgATP 5, ethylene glycol-bis(�-aminoethylether)-N,N,N�,N�-tetraacetic acid (EGTA) 9, HEPES 8, GTP1, phosphocreatine 10, and leupeptin 0.1 (pH 7.25–7.3, osmo-lality 280–290 mOsm). The rationale for selecting a Cs�-basedinternal solution included an improved voltage clamp, the in-creased flexibility to change the holding membrane potential,and the need to block K� channels to disclose Ca2� conduc-tances. In experiments in which Ca2� influx was estimated,EGTA was eliminated from the patch solution, and tetraethyl-ammonium (TEA; 20 mM) and tetrodotoxin (TTX; 1 �M)were added to the bath. The access resistances ranged from 8 to25 M�. Cells were initially held at –70 mV. Axopatch 200B or1D amplifiers were used for voltage clamp recordings. Accessresistance was compensated 60–85%. A 3 M KCl Agar bridgewas inserted between the extracellular solution and the Ag–AgCl-indifferent electrode. Liquid junction potentials (rangingfrom 3 to 5 mV) were not corrected. Basic membrane propertieswere measured by applying a 10 mV hyperpolarizing or depo-larizing voltage step and using the membrane test functionintegrated in the pClamp8 software (Axon Instruments, FosterCity, CA).
Drugs were applied in the bath or iontophoreticallythrough a five-barreled pipette (tip diameter 6–8 �m) placed15–50 �m from the recorded cell. The pipette containedN-methyl-D-aspartate (NMDA; 0.1 M, pH 8), kainate (0.1 M,pH 8) or -amino-3-hydroxy-5-methyl-4-isoxazolepropionicacid (AMPA; 0.1 M, pH 8.5) and saline for current balancingand control. Holding currents of appropriate polarity rangedbetween 15 and 20 nA.
Abnormal Cells in Human Cortical Dysplasia 473

Cell Visualization and Classification Criteria
Cells were visualized in gray matter, roughly in layersII–V. Cells with visible nuclei lying on the surface of the slicewere usually dead. Cells that yielded successful tight seals hadsmooth, regular membrane contours and a pale appearance. Incontrast, cells with rugged surfaces and dark pigmentation couldnot be patched. In addition, upon approximation of the patchpipette, the positive pressure produced a small dimple on themembrane of viable cells, indicating a soft texture. Nonviablecells were pushed away by the patch pipette, indicating a hard-ened texture. Only cells in which gigaohm seals could beobtained were examined. Pyramidal somata were easy to differ-entiate because the apical dendrite could be traced. Other typesof neurons were harder to differentiate, and a definite identityfor the cell had to await biocytin processing. The imagingprogram (Optimas; BioScan, Edmonds, WA) allowed somaticarea measurements online, so that abnormally large somata couldbe defined. Cells with somatic areas greater than 300 �m2 wereconsidered potentially abnormal. In most experiments, elec-trodes were filled with 0.2% biocytin (Sigma, St. Louis, MO) inthe internal solution to label visualized cells. After the experi-ment, the slice was fixed in 4% paraformaldehyde overnight,then processed according to published protocols (Horikawa andArmstrong, 1988).
Ca2� Imaging
In some experiments, Fura-2 pentapotassium salt (200 �M;Molecular Probes, Eugene, OR) was included in the internalsolution of the pipette. Once patched and filled with dye,neurons were imaged using a cooled CCD camera (AstroCamTE3/A/S; Perkin Elmer, Cambridge, United Kingdom) mountedon the microscope. A monochromator (SpectraMASTER; Per-kin Elmer) excited the sample at alternating wavelengths of 340and 380 nm with a 200–400-msec exposure time, at an imagerecording frequency of 0.8 Hz. Regions of interest (ROIs)approximately the size of the cell soma were designated withineach patched neuron. Average emitted light intensity of thisROI was measured for each excitation wavelength in eachframe. Software (Merlin; Perkin Elmer) calculated the ratio oflight emitted at 340 nm/380 nm. Ca2� concentrations arepresented in nanomolar units with the Grynkiewicz transforma-tion equation (Grynkiewicz et al., 1985).
Acute Neuron Dissociation
To obtain a better voltage clamp and remove influencesfrom distal dendritic processes, acute dissociation of neurons wasperformed in slices from 15 CD cases. After 1 hr of incubationin the ACSF solution, a slice was placed in a low-Ca2�–isethionate solution (in mM): 140 Na isethionate, 2 KCl,4 MgCl2, 0.1 CaCl2, 23 glucose, 15 HEPES (pH 7.4, 300 –305 mOsm). Small cortical pieces were dissected and placed inan oxygenated cell-stir chamber (Wheaton, Inc., Millville, NJ)containing papain (1–2 mg/ml; Calbiochem, La Jolla, CA) inHEPES-buffered Hank’s balanced salt solution (HBSS; Sigma) at35°C. After 20–40 min of enzyme digestion, tissue was rinsedthree times with the low-Ca2�–isethionate solution and me-chanically dissociated with a graded series of fire-polished Pas-teur pipettes. The cell suspension was plated into a Petri dishcontaining HEPES-buffered HBSS saline on the microscope
stage. Whole-cell recordings used standard electrophysiologicaltechniques. The internal solution consisted of (in mM): 180N-methyl-d-glucamine (NMDG), 40 HEPES, 2 MgCl2, 10EGTA, 12 phosphocreatine, 2 Na2ATP, 0.2 Na3GTP, 0.1 leu-peptin (pH 7.2–7.3 with H2SO4, 265–270 mOsm). The exter-nal solution consisted of (in mM): 135 NaCl, 20 CsCl, 5 BaCl2,10 glucose, 10 HEPES, 0.001 TTX (pH 7.3, 300–305 mOsm).Recordings were obtained with an Axopatch 200A amplifier.Electrode resistance was typically 2–4 M� in the bath. After sealrupture, series resistance (4–10 M�) was compensated (70–90%) and periodically monitored.
Statistical Analyses
In the text and tables, values are presented as means �standard errors (SE). Differences between means were assessedwith ANOVAs when multiple measures or groups were com-pared, followed by appropriate t-tests or t-tests alone when onlytwo groups were compared. Differences between means wereconsidered statistically significant at P 0.05.
RESULTSThe tissue originated from 37 children (20 males and
17 females) with intractable epilepsy (ages 0.19–14 years,average 3.05 years). Two patients had surgery twice, and,because �1 year had elapsed between surgeries, they wereconsidered separate cases. About half of the patients (N �19) had experienced infantile spasms. The average age ofseizure onset was 0.75 years. Pathologic findings in allcases revealed CD, from mild (N � 12) to severe (N � 27;Fig. 1). Cortical dyslamination and ectopic neurons in thewhite matter occurred in most cases. Hemimegalen-cephaly (N � 5), polymicrogyria (N � 4), and heterotopia(N � 6) also occurred. In three cases, CD was associatedwith tuberous sclerosis complex (TSC). Samples from LAand MA regions were compared, based on ECoG, PET,and MRI, in 18 cases. In 21 cases, only one cortical regionwas sampled (15 MA and 6 LA).
Most patients were under 5 years of age (N � 31),allowing optimal cell visualization with IR-DIC optics.Viable cells were encountered �30–100 �m from thesurface. In CD cases, electrophysiological data were ob-tained from 298 cells. Cells in frontal regions were sam-pled most often (N � 147), followed by temporal (N �74) and parietal (N � 69) regions. Only eight cells weresampled in the occipital region. Two hundred eleven cellswere from severe CD, and 87 were from mild CD cases,based on the pathologic findings. Two hundred cells werefrom the MA and 98 from the LA regions. More than halfof the recorded cells were morphologically examined afterbiocytin injection (N � 167). Interneurons (N � 24) andglial cells (N � 5) were occasionally sampled and could bedistinguished both morphologically and electrophysiologi-cally. Cells were recorded in presumed layers II–V, al-though it was difficult to determine their precise localiza-tion because of the severe cortical dyslamination, adefining feature of CD.
Normal-appearing pyramidal neurons (N � 183 ofthe total 298 cells) were sampled to establish a set ofnormative parameters for comparison. These neurons
474 Cepeda et al.

were recorded from mild (N � 72) or severe (N � 111)CD cases in LA (N � 69) and MA (N � 114) regions. Thedata were pooled into a single group (see Table II), be-cause there were no statistically significant differences inthe basic membrane properties of normal-appearing pyra-midal neurons, regardless of severity (mild vs. severe) anddegree of abnormality (LA vs. MA). In addition, normal-appearing pyramidal neurons (N � 23) obtained from theLA regions of non-CD cases (N � 5, Rasmussen’s en-cephalitis 2, tumor 1, stroke 1, and hippocampal sclerosis1), were also used for comparison. Membrane propertiesof normal-appearing pyramidal neurons from non-CDand CD cases were similar (see Table II).
Biocytin labeling revealed that most cells from CDcases displayed some degree of morphologic abnormalityon close examination. Thus, the term normal-appearing iscomparative and relative. Nonetheless, several cell typesdeviated significantly from the normative group, based onthe present data set. In total, 75 abnormal-appearing cells
(25% of the population) were identified and classified intofour groups (Fig. 2, Table I): cytomegalic pyramidal neu-rons, atypical nonpyramidal cells, misoriented and dys-morphic pyramidal neurons, and pyramidal neurons withimmature characteristics. Eleven cells could not be classi-fied and will not be discussed. Cytomegalic neurons andatypical cells were never observed in non-CD cases.
Cytomegalic Pyramidal NeuronsThese were pyramidal cells with very large somata
(30–40 �m diameter) found in nine severe and one mildCD cases (ages 0.2–7.85 years). They were classified ascytomegalic based on enlarged somata and often abnor-mally thick initial segments of dendrites and/or axons(Figs. 2, 3A3,A4,B2). Many cytomegalic neurons had nu-merous dendrites emanating from their cell bodies. Thesewere varicose and tortuous, with some distal tips recurvingtoward the somata. Dendritic fields were slightly shorterthan those of normal-appearing pyramidal neurons, with
Fig. 1. Immunohistochemistry of NeuN, a specific neuronal marker, innon-CD (left panels) and CD cases (right panels). A1: Low-magnification view of normal cortical cytoarchitecture (0.1 years, fron-tal). A2: Low-magnification view of a severe case of cortical dyslami-nation (CD severe, 0.68 years MA, temporal). B1: High-magnification
view of the normal cortex (layer V) shown in A1. B2: High-magnification view of severely dysplastic cortex (11.7 years MA, fron-tal) in which numerous cytomegalic neurons can be observed (arrows).[Color figure can be viewed in the online issue, which is available atwww.interscience.wiley.com.]
Abnormal Cells in Human Cortical Dysplasia 475

decreased branching and apparent reductions in spine den-sity. The somatic cross-sectional areas of cytomegalic neu-rons were significantly larger than those of normal-appearing pyramidal neurons (507 � 28 vs. 250 � 17�m2, P 0.001; measured from biocytin-filled neurons,10 cytomegalic and 10 normal-appearing from the samefive CD cases).
Twenty-two cytomegalic neurons were character-ized electrophysiologically. All but one were obtainedfrom severe CD cases. The vast majority (N � 18) were in
the MA sample. The four cells from the LA sample werefrom one severe CD case. Passive membrane propertieswere abnormal compared with those of normal-appearingcells (Table II). Cell capacitance was higher (P 0.001),reflecting the enlarged membrane area. Input resistancewas lower (P 0.002), and the time constant was longer(P 0.001).
In normal-appearing pyramidal neurons, in standardACSF and with Cs�-methanesulfonate in the patch pi-pette to block K� channels partially, a series of depolar-izing voltage commands primarily evoked inward currents(Fig. 3C1). The first large peak corresponded to the fastNa� current and was blocked by TTX (1 �M; see alsoFig. 8C1,2). The initial peak was followed by a Ca2�
current (Fig. 3C1,2), which was sensitive to cadmium(Cd2�; 100 �M, N � 35). For cytomegalic neurons in theslices, it was difficult to obtain a reasonable space clamp. Instandard ACSF, when the membrane was depolarized, bysteps, by slow ramps, or by holding at depolarized poten-tials, most of these neurons displayed repetitive, un-clamped Ca2� spikes with very slow inactivation (Fig.3D1,2,E). The Ca2� spikes were completely blocked byCd2� (N � 3; Fig. 3E). TTX and TEA (20 mM) reducedthe tendency to generate repetitive Ca2� spikes after sim-ilar voltage protocols, leaving instead very large Ca2�
currents (Fig. 4A1–3). The peak Ca2� current was signif-icantly higher in cytomegalic compared with normal-appearing neurons (P 0.001; Table II).
Intracellular Ca2� concentrations from normal-appearing and cytomegalic neurons were estimated inseven mild and three severe CD cases, as well as fivenon-CD cases, by using Fura-2 (200 �M) in the patchpipette and TTX and TEA in the bath. Baseline Ca2�
levels were slightly decreased in cytomegalic comparedwith normal-appearing neurons (69 � 12 and 82 �12 nM, respectively), but the difference was not statisti-cally significant. In contrast, depolarizing ramps, voltagesteps, and holding the membrane at depolarized potentialsfor long periods all demonstrated significantly increasedCa2� influx in the cytomegalic neurons (N � 7 cells)compared with normal-appearing neurons (N � 37 cells,20 from CD and 17 from non-CD cases; Fig. 4B1–3).Peak Ca2� currents were also significantly larger in thissubset of cytomegalic neurons when using the ramp (Fig.4A1, inset; P 0.01) and step protocols (from –40 to0 mV; P 0.001 to P 0.01; Fig. 4A2).
Although voltage clamp recordings in slices sug-gested alterations in Ca2� conductances in cytomegalicneurons, accurate measurements of Ca2� currents couldnot be obtained in slices because of space clamp limitations(Armstrong and Gilly, 1992). These limitations were ex-acerbated in cytomegalic neurons because of their largemembrane capacitance, their low input resistance, and thelarge amplitude of the macroscopic Ca2� currents. Toobtain a better space clamp and more precise measure-ments of the amplitude of Ca2� currents and currentdensities, slices from mild (N � 6) or severe CD (N � 9)cases were treated enzymatically to dissociate cortical neu-
Fig. 2. Camera lucida drawings of biocytin-filled cells illustrating themajor cell types in CD. Indicates the axon. Top left is a normal-appearing pyramidal neuron (CD severe, 0.21 years LA, frontal). Topright is a cytomegalic neuron (CD severe, 0.21 years MA, temporal).Note the thickness of the axon. Bottom left shows an atypical cell (CDsevere/TSC, 3.2 years MA, frontal). Bottom right shows a dysmorphicneuron with tortuous processes (CD severe, 1.4 years MA, frontal) anda group of immature pyramidal neurons (CD severe, 0.92 years. LA,frontal). Scale bar refers to all neurons.
TABLE I. Abnormal Cell Groups*
Cell type
CD mild CD severe
LA MA LA MA
Cytomegalic (N � 22) 0 1 4 17Atypical (N � 24) 0 0 0 24Misoriented (N � 6) 1 2 2 1Dysmorphic (N � 12) 1 1 3 7Immature (N � 11) 1 2 6 2Totals 3 6 15 51
*LA, least abnormal regions; MA, most abnormal regions.
476 Cepeda et al.

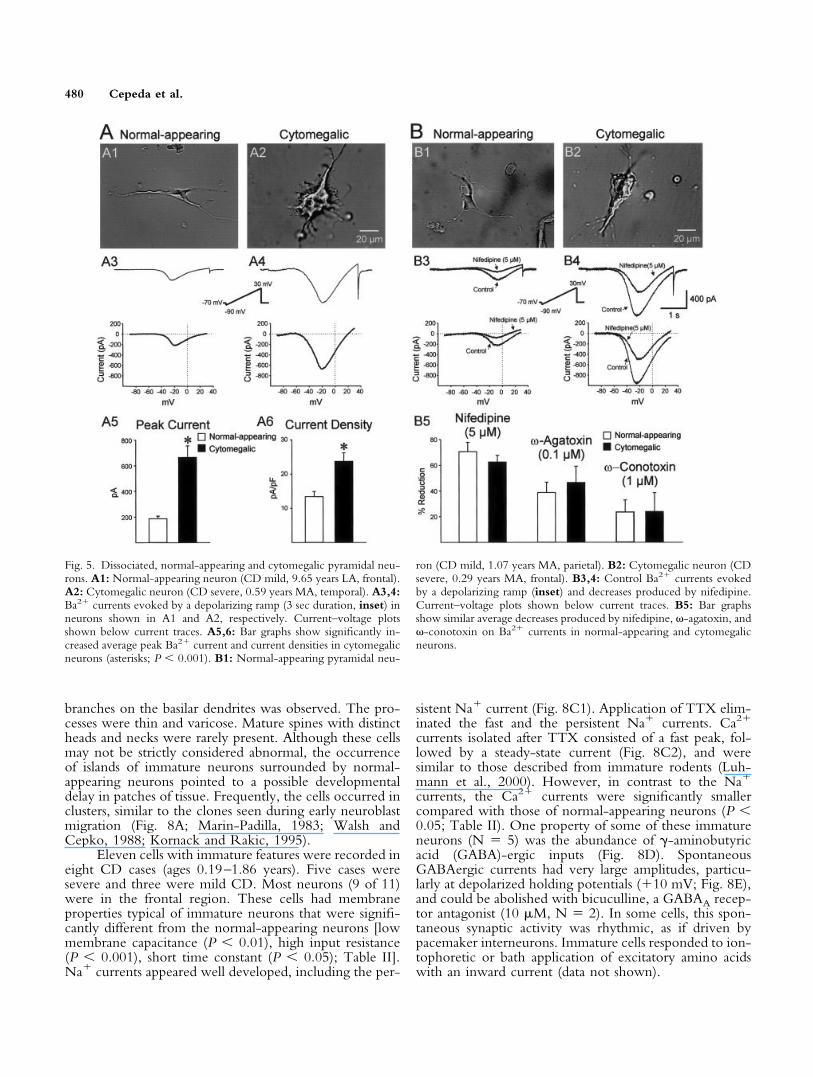
rons (Sayer et al., 1993). Cytomegalic cells (N � 13; Fig.5A2,B2) displayed higher capacitance (P 0.001), lowerinput resistance (P 0.05), and longer time constants(difference not statistically significant; Table II) thannormal-appearing neurons (N � 38; Fig. 5A1,B1). Inthese experiments, Ca2� was replaced by Ba2�. High-voltage Ba2� currents were significantly larger in cytome-galic compared with normal-appearing pyramidal neurons(P 0.001; Fig. 5A3–5). Current density was almosttwice that of normal-appearing neurons (P 0.001; Fig.5A6). To determine the type of Ca2� current, selectivechannel blockers (nifedipine, “L” type; -agatoxin,“P/Q” types; and -conotoxin, “N” type) were appliedto a subset of the dissociated cells (N � 16; 12 normal-
appearing and 4 cytomegalic). Most of the Ba2� current(�60%) was from influx through L-type channels, with asmaller contribution by P/Q (�25%)- and N (�40%)-type channels (Fig. 5B3–5). There were no significantdifferences in the percentage reduction of Ba2� current byselective toxins between cytomegalic and normal-appearing neurons (Fig. 5B5).
Atypical, Nonpyramidal CellsThese cells (N � 24) were found only in the MA
tissue samples from severe CD cases (N � 5, ages 0.59 –14 years) and in all cases associated with TSC (N � 3, ages2.97–5.46 years). The CD/TSC cases accounted for halfof the cells in this group (N � 12); the other half came
Fig. 3. Morphology and electrophysiology of normal-appearing andcytomegalic neurons. A: Normal-appearing (A1,2) and cytomegalic(A3,4) neurons obtained from the LA (frontal) and the MA (temporal)tissue samples, respectively, from a 0.21-year-old patient with severeCD (IR-DIC images). Arrows point to somata. B: Normal-appearing(B1) and cytomegalic (B2) cells after biocytin processing. B1 corre-sponds to the IR-DIC image shown in A2. B2 corresponds to the cellshown in A4. C1: Currents evoked by a step voltage protocol (from–80 to �10 mV in 10 mV increments, 400 msec duration, inset) in thenormal-appearing neuron (A2). C2: Current-voltage plot from C1taken at 300 msec. D1: Depolarizing step voltage commands (inset)evoked repetitive, unclamped Ca2� spikes in the cytomegalic neuron(A4 image). These current oscillations were not seen in normal-
appearing neurons and likely were due to space clamp limitationsproduced by increased somatic area and large current amplitude. D2:Ramp voltage command (8 sec duration, inset) also evoked repetitiveCa2� spikes in the same neuron. E: Holding the voltage at –30 mV(inset) induced repetitive, slowly inactivating spikes (top trace) thatcould be blocked by Cd2� (bottom trace). Dotted lines show expandedtime sweep of the repetitive spikes. F: Spontaneous postsynaptic cur-rents (top trace) recorded in a cytomegalic neuron from a severe CDcase (0.29 years MA, frontal). Bottom trace shows spontaneous currentsin a normal-appearing pyramidal neuron (CD severe, 0.92 years MA,parietal). [Color figure can be viewed in the online issue, which isavailable at www.interscience.wiley.com.]
Abnormal Cells in Human Cortical Dysplasia 477

from CD cases alone. Cellular morphology was very dif-ferent from that of the cytomegalic pyramidal neurons.Cell bodies were generally round, with a diameter of20–30 �m (Fig. 6A). Axons could not be distinguished inany cell. Spines were occasionally present, and two cellshad no visible spines. The processes had a wavy appear-ance, undulating widely, with sharp angles and saw-toothed patterns (Fig. 6B). Some cells had processes witha straighter course, although they were still quite distorted.With the cell body at the center, the domain of the cellwas usually spherical and concentrated within a 300–400 �m diameter. However, some processes continued forup to 700 �m from the cell body. These cells had mor-phological traits of both neurons and glia. Their neuron-like features included large soma, extensive dendritic do-main, and occasional spines. Their glial features consistedof no discernible axon, and their processes branched onlyonce or twice. Their overall morphology resembled thatof fibrillary astrocytes. Cells with similar morphology havebeen described with the use of Golgi impregnation in acase of Bourneville’s disease or TSC (Machado-Salas,1984).
Electrophysiologically, these cells were characterizedby the absence of voltage-gated Na� and Ca2� currents(Fig. 6C). In some cells (N � 3), there was a very small,fast inward current, followed by an incipient outwardcurrent. The current–voltage relationship was linearbetween – 80 and �10 mV (Fig. 6C1,2). These cellshad significantly higher input resistances than normal-appearing pyramidal cells (P 0.001; Table II) and didnot respond to application of excitatory amino acids (Fig.6D). In addition, they may not receive major synapticinputs, insofar as none of the cells examined (N � 5)displayed spontaneous synaptic currents compared withnormal-appearing cells (Fig. 6E). Atypical cells had a highinput resistance and lacked a complement of K� currents.Because Cs� was present in the patch pipette, it was notpossible to determine whether some of these cells could
have generated inwardly rectifying currents. However,given that the current–voltage relationship was linear evenimmediately after achieving the whole-cell configuration,prior to cell dialysis with Cs�, this seems unlikely, sug-gesting that these cells were electrically more inert. Theseproperties separate atypical cells from commonly foundastrocytes and other glial cells (Tse et al., 1992; Ketten-mann, 1999; Zhou and Kimelberg, 2000).
Misoriented and Dysmorphic Pyramidal NeuronsThese neurons were observed frequently in the ab-
normal tissue samples. Misorientation was common, al-though not exclusive, in tissue from young cases. Abnor-mal polarity ranged from slight rotation (30–45°) tocomplete inversion of the soma (180°) in relation to thepial surface (Fig. 7A1,3). Pyramidal neurons with dysmor-phic processes included extremely tortuous dendrites (Fig.7A4,B1,2) and axons, bifurcated or double “apical” den-drites, and other dysmorphisms. Sometimes, misorienta-tion and tortuousness occurred in conjunction.
Misoriented pyramidal neurons were found in severe(N � 3) or mild (N � 2) CD cases. Six neurons wererecorded electrophysiologically. Two had �45° somaticmisorientation, and four had complete (180°) inversion ofthe soma. These neurons were electrophysiologically sim-ilar to normal-appearing, properly oriented pyramidalcells. Basic membrane properties (Table II) and voltage-and ligand-gated currents were not significantly different(Fig. 7C1). The neurons displayed inward currents andresponded to iontophoretic or bath application of excita-tory amino acids (Fig. 7D1).
Pyramidal neurons with dysmorphic processes oc-curred both in severe (N � 5) and in mild (N � 2) CDcases, and no preferential age was observed. Overall, thesecells resembled normal-appearing pyramidal neurons, withsomatic diameters of 12–30 �m (Fig. 7A2–4) and similarsomatodendritic architecture and branching orders. How-ever, they exhibited tortuous and varicose dendrites, re-
TABLE II. Passive and Active Membrane Properties
Cell type
Cells in slices
Cap (pF) Ri (M�) Tau (msec) ICa2� (pA)
Normal, non-CD (N � 23) 183 � 16 126 � 14 2.6 � 0.2 1,340 � 370Normal, CD (N � 183) 166 � 14 172 � 13 2.9 � 0.3 1,666 � 135Cytomegalic (N � 22) 397 � 32* 71 � 7* 5.0 � 0.3* 4,008 � 604*Atypical (N � 24) 217 � 33 553 � 96* 3.6 � 1.0 0*Misoriented (N � 6) 134 � 28 142 � 35 2.3 � 0.3 1,548 � 400Dysmorphic (N � 12) 168 � 46 191 � 32 4.5 � 1.1 1,470 � 281Immature (N � 11) 64 � 7* 607 � 150* 0.9 � 0.2* 439 � 157*
Dissociated cells
Cap (pF) Ri (G�) Tau (�sec)
Normal, CD (N � 38) 15 � 1.1 1.9 � 0.2 292 � 34Cytomegalic (N � 13) 29 � 2.6** 1.2 � 0.3** 483 � 163
*Significantly different from normal-appearing cells.**Significantly different from normal-appearing dissociated cells.
478 Cepeda et al.

curving branches, and decreased spine density. The prox-imal portion of the apical dendrite had a saw-toothedpattern that sometimes extended throughout the length ofthe apical shaft (Fig. 7B). Axons were also overly tortuous.Three cells had dendrites that were significantly thickerthan normal.
Twelve cells were recorded electrophysiologically.Despite morphological abnormalities, their electrophysio-logical characteristics were similar to those of normal-appearing neurons (Table II). Only two cells with abnor-mally thick, tortuous dendrites displayed repetitive Ca2�
spikes when depolarized. Furthermore, all neurons re-sponded to iontophoretic or bath application of excitatoryamino acids (Fig. 7C2,D2).
Cells With Immature CharacteristicsThese cells had smaller somatic diameters (10–12 �m)
than other cells of similar ages and had round or pyramidalsomata. Dendrites were not elaborate, with an apical pro-cess and rudimentary basilar dendrites (Fig. 8B). Neitheroblique branches on the apical dendrite nor secondary
Fig. 4. Ca2� currents and influx in normal-appearing and cytomegalicneurons. A: In TTX and TEA, whole-cell Ca2� currents were evokedby a slow depolarizing ramp (A1), by a series of step voltage commands(–90 to �40 mV in 10 mV increments, 500 msec duration, 20 secbetween each step; A2), and by holding the membrane at –30 mV for30 sec (A3). Traces in A1–3 are from the same normal-appearing (LA,frontal area) and cytomegalic neurons (MA, parietal area). Both cellsfrom a patient with severe CD (0.92 years). Large-amplitude Ca2�
currents were evoked in the cytomegalic neuron compared with thenormal-appearing cell. Inset in A1 shows that the average peak currentwas significantly greater in cytomegalic neurons (*P 0.01). Inset inA2 shows that average currents were significantly greater at step volt-ages of –40 to 0 in cytomegalic neurons (P 0.001 to P 0.01;asterisks). B: Intracellular Ca2� measured with Fura-2 was significantly
increased in the cytomegalic neuron when the membrane was depo-larized by the ramp (B1), step voltage commands (B2), or long-duration depolarizing voltage step (B3) shown in A. Inset in B1 showsthat the average percent increase in Ca2� evoked by ramps was signif-icantly greater in cytomegalic neurons (P 0.05; asterisk). B2: Intra-cellular Ca2� increases during each voltage step of the protocol shownin A2 for the normal-appearing and cytomegalic neuron, showing amarked increase in Ca2� influx in the cytomegalic cell. Numbers abovepeaks are step voltage commands. Bottom graph in B2 shows thataverage percentage change in Ca2� influx for all cytomegalic neuronswas significantly greater than that for normal-appearing neurons atvoltage steps of –30 to �10 mV (asterisks). B3: Intracellular Ca2�
increases also were greater in cytomegalic neurons during a long-duration voltage step from –70 to –30 mV.
Abnormal Cells in Human Cortical Dysplasia 479
branches on the basilar dendrites was observed. The pro-cesses were thin and varicose. Mature spines with distinctheads and necks were rarely present. Although these cellsmay not be strictly considered abnormal, the occurrenceof islands of immature neurons surrounded by normal-appearing neurons pointed to a possible developmentaldelay in patches of tissue. Frequently, the cells occurred inclusters, similar to the clones seen during early neuroblastmigration (Fig. 8A; Marin-Padilla, 1983; Walsh andCepko, 1988; Kornack and Rakic, 1995).
Eleven cells with immature features were recorded ineight CD cases (ages 0.19–1.86 years). Five cases weresevere and three were mild CD. Most neurons (9 of 11)were in the frontal region. These cells had membraneproperties typical of immature neurons that were signifi-cantly different from the normal-appearing neurons [lowmembrane capacitance (P 0.01), high input resistance(P 0.001), short time constant (P 0.05); Table II].Na� currents appeared well developed, including the per-
sistent Na� current (Fig. 8C1). Application of TTX elim-inated the fast and the persistent Na� currents. Ca2�
currents isolated after TTX consisted of a fast peak, fol-lowed by a steady-state current (Fig. 8C2), and weresimilar to those described from immature rodents (Luh-mann et al., 2000). However, in contrast to the Na�
currents, the Ca2� currents were significantly smallercompared with those of normal-appearing neurons (P 0.05; Table II). One property of some of these immatureneurons (N � 5) was the abundance of �-aminobutyricacid (GABA)-ergic inputs (Fig. 8D). SpontaneousGABAergic currents had very large amplitudes, particu-larly at depolarized holding potentials (�10 mV; Fig. 8E),and could be abolished with bicuculline, a GABAA recep-tor antagonist (10 �M, N � 2). In some cells, this spon-taneous synaptic activity was rhythmic, as if driven bypacemaker interneurons. Immature cells responded to ion-tophoretic or bath application of excitatory amino acidswith an inward current (data not shown).
Fig. 5. Dissociated, normal-appearing and cytomegalic pyramidal neu-rons. A1: Normal-appearing neuron (CD mild, 9.65 years LA, frontal).A2: Cytomegalic neuron (CD severe, 0.59 years MA, temporal). A3,4:Ba2� currents evoked by a depolarizing ramp (3 sec duration, inset) inneurons shown in A1 and A2, respectively. Current–voltage plotsshown below current traces. A5,6: Bar graphs show significantly in-creased average peak Ba2� current and current densities in cytomegalicneurons (asterisks; P 0.001). B1: Normal-appearing pyramidal neu-
ron (CD mild, 1.07 years MA, parietal). B2: Cytomegalic neuron (CDsevere, 0.29 years MA, frontal). B3,4: Control Ba2� currents evokedby a depolarizing ramp (inset) and decreases produced by nifedipine.Current–voltage plots shown below current traces. B5: Bar graphsshow similar average decreases produced by nifedipine, -agatoxin, and -conotoxin on Ba2� currents in normal-appearing and cytomegalicneurons.
480 Cepeda et al.

DISCUSSIONWe have categorized abnormal-appearing cells in
CD and TSC cases into four major groups based on a setof common morphological and electrophysiological prop-erties: cytomegalic neurons, atypical cells, misoriented anddysmorphic neurons, and pyramidal neurons with imma-ture features. Cytomegalic neurons were found almostexclusively in severe CD cases. Atypical cells were ob-served only in severe CD and in all TSC cases. The othertwo groups occurred in both mild and severe CD. Becauseseizures occurred in all CD and non-CD cases, the mech-anisms leading to epileptogenic activity can be attributednot solely to one class of cell but to a complex interplaybetween normal- and abnormal-appearing cells as well astheir network interactions.
In immature neurons, the capacity to generate par-oxysmal activity seemed small. Unless immature cells havean increased propensity to generate synchronized activity,it would be difficult for these collections of cells to behighly epileptogenic. Notwithstanding the fact that theyreceive prominent, sometimes rhythmic GABAergic inputand that electrotonic coupling is increased in very youngneurons (Cepeda et al., 1993), their active membraneproperties per se do not show signs of abnormality, e.g.,large inward currents. However, their presence in dysplas-tic brains may reflect regional abnormalities in develop-ment and cell differentiation. This anisochrony couldcause perturbations in the integrative capabilities of re-stricted brain regions, leading to areas of increased excit-ability surrounding these cells.
Fig. 6. Morphology and electrophysiology of atypical cells A: IR-DICimages of atypical cells obtained from two cases of severe CD, one ofthem (A1) associated with TSC (3.2 years MA, frontal) and the otherwithout TSC (A2; 2.8 years MA, temporal). Arrows point to roundsomata. B: Atypical biocytin-filled cells. B1 is from the cell in A1.B2–4 are from severe CD. B2 and B4 are from the same case (1.41years MA, parietal). B3 is from a 14-year-old MA, parietal area. C1:Cell shown in A1 and B1 displayed no active currents when depolar-izing step voltage commands were applied (inset). The capacitiveartifact could not be fully compensated because of the large size of thecell. The current–voltage relationship consisted of a straight line. C2:
Cell shown in B2 also displayed no active currents to depolarizingvoltage commands (inset). Again, the current–voltage relationshipconsisted of a straight line. D: The cell shown in A1 and B1 did notrespond to iontophoretic application of excitatory amino acids. Appli-cation of NMDA or kainate (KA) at increasing iontophoretic ejectioncurrents produced no responses. E: Cell shown in B2 displayed nospontaneous postsynaptic currents (top trace). Bottom trace showsspontaneous currents typically displayed by a normal-appearing pyra-midal neuron. [Color figure can be viewed in the online issue, whichis available at www.interscience.wiley.com.]
Abnormal Cells in Human Cortical Dysplasia 481

Similarly, misoriented and dysmorphic pyramidalneurons displayed low to mild epileptogenic potential.Although misorientation and tortuousness were usuallyexpressed in different neurons, these abnormalities mightshare similar developmental origins, e.g., lack of adequateradial glia guidance or absence of appropriate chemicaltargets. The evidence indicates that misorientation per semay contribute to but is probably not a causal factorunderlying epileptic activity. For example, a mouse modelof cortical migration disorder, the Reeler mouse, withcomplete inversion of neuronal polarity, does not displayspontaneous seizures (Caviness, 1976; Schwartzkroin andWalsh, 2000). In fact, relatively small but not negligiblenumbers of improperly oriented pyramidal neurons occurin normal brains (Van der Loos, 1965). The degree ofneuronal disarray in some of our cases was very remark-able. Abnormal polarity and the existence of cells withsinuous dendrites could predispose toward aberrant wiringof the cerebral cortex, and this abnormal connectivity may
shift the balance of excitation–inhibition in cortical net-works.
Cells with atypical morphological features werecharacterized by the absence or marked decreases of activevoltage- and ligand-gated currents as well as by the appar-ent lack of spontaneous synaptic input. The electrophys-iological properties of these cells were similar, but notidentical, to those of astrocytes (Kettenmann, 1999; Zhouand Kimelberg, 2000). Determination of cellular lineagewas difficult in most cases, because the cells shared mor-phological properties of both neurons and glia (Vinters etal., 1992; Cotter et al., 1999). We previously demon-strated, using immunohistological methods in one CD/TSC case, that most of these cells were labeled by neuronal(neuro N) but not glial (glial fibrillary acidic protein)markers. Only very small numbers of cells were simulta-neously labeled by both markers (Mathern et al., 2000).Therefore, we concluded that these atypical cells weresimilar to the “balloon” or giant cells reported previously
Fig. 7. Morphology and electrophysiology of misoriented and dysmor-phic neurons. A: IR-DIC images of misoriented (A1, CD severe,0.19 years LA, frontal; A3, CD severe, 0.69 years MA, parietal) andtortuous dysmorphic (A2,4, both from CD severe, 0.19 years MA,parietal) pyramidal neurons. Thick arrows show direction of pia. Thinarrows show direction of apical dendrites. Arrow in A4 shows tortuousapical dendrite. B: Neurons with extremely tortuous processes afterbiocytin processing (B1, CD severe, 1.4 years. MA, frontal; B2, CDsevere, 0.19 years. MA, parietal). IR-DIC image of neuron in B2shown in A2. C1,2: Electrophysiology of the misoriented neuronshown in A1 and the tortuous neuron shown in A2 and B2. The tracesand graphs below show voltage-gated currents were similar to those
evoked by step voltage commands (inset) in normal-appearing pyra-midal neurons (Fig. 2C1,2). Current–voltage plots taken at 300 msec.D1,2: Misoriented and tortuous pyramidal neurons shown in A1 andB2, respectively, responded to bath application of NMDA. The mem-brane was held at –70 mV and slowly depolarized to 30 mV to blockmost voltage-gated Ca2� conductances. The faster downward rampchanged the membrane from �30 mV to –90 mV (inset). The current-voltage relationship was obtained by digitally subtracting the currentevoked by the ramp before and after bath application of NMDA (onlythe downward ramp was used). [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
482 Cepeda et al.

for TSC (Crino et al., 1996; Mizuguchi and Takashima,2001).
The fact that atypical cells were found in severe CDand in all CD/TSC cases suggests a morphological link. Ithas been proposed that CD may represent an atypical formof TSC (Vinters et al., 1992, 1999). Immunohistochemicalstudies have demonstrated that globular and atypical cellsin CD are similar to those observed in cortical tubers andshow both neuronal and astrocytic epitopes, indicating thelocal proliferation of multipotential neuroectodermal cells(Vinters et al., 1992). In contrast, single-cell analysis intubers suggests that giant cells are of neuronal lineagedespite the persistence of embryonic markers, such as
nestin (Crino et al., 1996). The recent demonstration thatradial glia are capable of generating astrocytes as well asneurons during cortical development (Malatesta et al.,2000; Alvarez-Buylla et al., 2001; Noctor et al., 2001) mayprovide important clues about the origin of these bizarrecells. It is possible to speculate that atypical cells wereoriginally radial glia that, under unspecified circumstances,had an arrest in development, preventing them fromreaching their final morphological and functional fate.
The present evidence does not support an obviousrole for atypical cells in generating paroxysmal activity, inthat they appear to lack a number of voltage-gated currentsand are insensitive to application of excitatory amino acids.
Fig. 8. Morphology and electrophysiology of immature pyramidalneurons. A: IR-DIC image shows a cluster of immature cells (whitearrows). The black arrow points to the neuron recorded and filled withbiocytin and shown as the darker neuron after biocytin processing in B(CD severe, 0.92 years LA, frontal). The second pyramidal cell in B,which is more lightly filled, was also patched but for a shorter period oftime. C: Traces correspond to inward currents evoked by a series ofdepolarizing step voltage commands (inset) in standard ACSF and afteraddition of TTX. The current–voltage relationships (taken at about70 msec) show the persistent Na� and the steady-state inward Ca2�
currents before and after addition of TTX. D: These currents also occur
during a depolarizing ramp voltage command (4 sec duration; inset).The large-amplitude, rhythmic synaptic outward currents were prob-ably mediated by activation of GABAA receptors. In this cell, thereversal potential for these currents occurred between –60 and –55 mV.After TTX, the persistent Na� current and the spontaneous synapticactivity were abolished, leaving only the Ca2� current and someminiature synaptic currents. E: Spontaneous rhythmic GABAergicactivity while the membrane was held at �10 mV before TTX (CDsevere, 0.38 years LA, frontal). [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
Abnormal Cells in Human Cortical Dysplasia 483

Although recently it has been reported that putative “bal-loon” cells could be the main culprits with regard toepileptogenicity in CD (Spreafico et al., 1998), the presentdata are not supportive of this suggestion. Furthermore,atypical cells were the only cells that did not have anyappreciable synaptic input. This fact may correlate withthe low synaptophysin immunoreactivity described fortubers and giant cells from TSC (Lippa et al., 1993; Hiroseet al., 1995) and makes it even less likely that these cellscontribute significantly or directly to seizure generation.However, the presence of “silent” cells could inducesynaptic reorganization that might lead to aberrant inner-vation of the more normal neurons. This could potentiallycontribute to hyperexcitability and seizures.
In contrast, cytomegalic neurons have propertiessuggesting that they could contribute to abnormal dis-charges in severe CD. Their large somatic size and axo-dendritic hypertrophy probably reflect a capacity for denseinnervation. In standard ACSF, they produced repetitiveCa2� spikes, without much inactivation, and TTX/TEAreduced this tendency, suggesting participation of Ca2�-activated K� conductances in the production of regener-ative spikes. Finally, dissociated cytomegalic neurons dis-played increased Ca2� current densities, suggesting up-regulation of Ca2� channel density or alterations in thekinetic properties. These results are supported by evidencefrom studies indicating that Ca2� channels may be criti-cally involved in the generation of epileptic discharges(Speckmann et al., 1993). Accordingly, paroxysmal depo-larization shifts at the single-cell level are initiated by theinflux of Ca2� ions, and L-type channel antagonists canreduce epileptiform discharges in human neocortical tissue(Straub et al., 2000). In addition, cytomegalic neuronsexpress glutamate receptor subunits abundantly (Kerfoot etal., 1999), and we have preliminary evidence indicatingthey may also have reduced Mg2� sensitivity (Flores-Hernandez et al., unpublished observations).
Cytomegalic neurons show mature features consist-ing of elaborate dendritic and axonal processes. Theirmorphological characteristics have a number of similaritiesto characteristics found in previous Golgi studies of humansubplate cells and the meganeurons of acquired CD (Ko-stovic and Rakic, 1990; Marin-Padilla, 1999). The sub-plate in higher primates and humans is the first set of cellsto migrate from the germinal matrix; these cells generatecomplex dendrites and axons and then die in the last 4–6 weeks of gestation. The loss of subplate cells correlateswith the formation of secondary cortical gyri on the sur-face of the infant brain. The subplate is four to five timesthe thickness of the cortical mantle at about 24 weeks andis composed of at least five cell types: normally shapedpyramidal neurons, inverted pyramidal cells, bipolar cells,cells with “spider”-like processes of normal size, and cy-tomegalic neurons (Mrzljak et al., 1988; Kostovic andRakic, 1990). We have observed all these cell types in oursevere CD cases. Clinically, we noticed that these cells areassociated with regions of the brain where the gyrus waslarger than expected, and they never involved primary
gyri, only secondary ones. Therefore, our findings areconsistent with the hypothesis that severe CD in humansmay consist of abnormal migration of cells in combinationwith arrest of cortical formation, i.e., preservation of sub-plate cells, and failure to reach complete differentiation.
One limitation of the present study is the use of Cs�
as the main carrier in the internal solution, which pre-cludes any solid assumptions on cell firing patterns orexcitability. However, previous studies using currentclamp and K�-based solutions were unsuccessful in dem-onstrating alterations in the firing patterns and membraneproperties of cells from pediatric CD tissue, but primarilynormal-appearing cells were examined (Wuarin et al.,1990; Cepeda et al., 1991, 1992, 1999; Dudek et al., 1995;Tasker et al., 1996). Another limitation is inherent in theslice preparation. It is often very difficult to obtain rea-sonable space clamp control in cells with extended pro-cesses (Armstrong and Gilly, 1992). This was particularlytrue for cytomegalic neurons, because of their large mem-brane area, their low input resistance, and the magnitudeof the Ca2� currents. Because of these limitations, weperformed additional experiments in dissociated neurons.These experiments confirmed the altered Ca2� conduc-tances observed in slices.
The oscillatory behavior displayed by cytomegalicneurons, as well as the impossibility of obtaining goodclamp control, could be attributed to unclamped Ca2�
spikes generated in distal dendrites. There has been a seriesof electrophysiological studies examining the crucial roleof dendrites in the production of nonlinear regenerativeresponses, including plateau potentials and bistable repet-itive firing modes, especially in motor neurons. PersistentNa� and Ca2� currents are thought to be mediated bymembrane channels located primarily on the dendrites(Carlin et al., 2000). Their dendritic location was sug-gested by the presence of clockwise hysteresis, a phenom-enon arising from the fact that the voltage-dependentchannels carrying the inward current are electrically distantfrom the soma and not under voltage clamp control (Pow-ers and Binder, 2003). Other indications of the contribu-tion of distal channels are a delayed current onset inresponse to a depolarizing voltage clamp step and a pro-longed decay following the step offset (Carlin et al., 2000).For cytomegalic neurons, we observed very long tail cur-rents following voltage clamp steps, consistent with sus-tained depolarizations in regions of the dendrites beyondvoltage clamp control. These persistent Ca2� currentscould be relevant for the generation of epileptic dis-charges.
The present electrophysiological and morphologicalstudies of CD tissue demonstrate that individual abnormal-appearing neurons have abnormal intrinsic membraneproperties that could contribute to epilepsy. By furthercharacterizing these electrophysiological features, particu-larly in cytomegalic neurons, we may be able to under-stand better a component of the basic mechanism ofepileptogenesis in children with CD.
484 Cepeda et al.

ACKNOWLEDGMENTSWe thank the young patients and their parents for
allowing use of part of the resected tissue for the presentstudy. We also thank Susan Koh, Joyce Wu, the UCLAHospital Pediatric Neurology staff, Carol Gray, andDonna Crandall for their assistance. C.C. dedicates thispaper to Dr. Robert Naquet for being a constant source ofinspiration and for his valuable criticisms on a previousversion of the manuscript.
REFERENCESAlvarez-Buylla A, Garcia-Verdugo JM, Tramontin AD. 2001. A unified
hypothesis on the lineage of neural stem cells. Nat Rev Neurosci 2:287–293.
Armstrong CM, Gilly WF. 1992. Access resistance and space clamp prob-lems associated with whole-cell patch clamping. Methods Enzymol 207:100–122.
Avoli M, Bernasconi A, Mattia D, Olivier A, Hwa GG. 1999. Epileptiformdischarges in the human dysplastic neocortex: in vitro physiology andpharmacology. Ann Neurol 46:816–826.
Avoli M, Mattia D, Olivier A. 2002. A window on the physiopathogenesisof seizures in patients with cortical dysplasia. Adv Exp Med Biol 497:123–132.
Carlin KP, Jones KE, Jiang Z, Jordan LM, Brownstone RM. 2000. Den-dritic L-type calcium currents in mouse spinal motoneurons: implicationsfor bistability. Eur J Neurosci 12:1635–1646.
Caviness VS Jr. 1976. Patterns of cell and fiber distribution in the neocortexof the reeler mutant mouse. J Comp Neurol 170:435–447.
Cepeda C, Peacock W, Levine MS, Buchwald NA. 1991. Iontophoreticapplication of NMDA produces different types of excitatory responses indeveloping human cortical and caudate neurons. Neurosci Lett 126:167–171.
Cepeda C, Radisavljevic Z, Peacock W, Levine MS, Buchwald NA. 1992.Differential modulation by dopamine of responses evoked by excitatoryamino acids in human cortex. Synapse 11:330–341.
Cepeda C, Walsh JP, Peacock W, Buchwald NA, Levine MS. 1993.Dye-coupling in human neocortical tissue resected from children withintractable epilepsy. Cereb Cortex 3:95–107.
Cepeda C, Li Z, Cromwell HC, Altemus KL, Crawford CA, Nansen EA,Ariano MA, Sibley DR, Peacock WJ, Mathern GW, Levine MS. 1999.Electrophysiological and morphological analyses of cortical neurons ob-tained from children with catastrophic epilepsy: dopamine receptor mod-ulation of glutamatergic responses. Dev Neurosci 21:223–235.
Cotter D, Honavar M, Lovestone S, Raymond L, Kerwin R, Anderton B,Everall I. 1999. Disturbance of Notch-1 and Wnt signalling proteins inneuroglial balloon cells and abnormal large neurons in focal corticaldysplasia in human cortex. Acta Neuropathol 98:465–472.
Crino PB, Trojanowski JQ, Dichter MA, Eberwine J. 1996. Embryonicneuronal markers in tuberous sclerosis: single-cell molecular pathology.Proc Natl Acad Sci USA 93:14152–14157.
Crino PB, Miyata H, Vinters HV. 2002. Neurodevelopmental disorders asa cause of seizures: neuropathologic, genetic, and mechanistic consider-ations. Brain Pathol 12:212–233.
Dudek FE, Wuarin JP, Tasker JG, Kim YI, Peacock WJ. 1995. Neuro-physiology of neocortical slices resected from children undergoing surgi-cal treatment for epilepsy. J Neurosci Methods 59:49–58.
Farrell MA, DeRosa MJ, Curran JG, Secor DL, Cornford ME, Comair YG,Peacock WJ, Shields WD, Vinters HV. 1992. Neuropathologic findingsin cortical resections (including hemispherectomies) performed for thetreatment of intractable childhood epilepsy. Acta Neuropathol 83:246–259.
Grynkiewicz G, Poenie M, Tsien RY. 1985. A new generation of Ca2�
indicators with greatly improved fluorescence properties. J Biol Chem260:3440–3450.
Hirose T, Scheithauer BW, Lopes MB, Gerber HA, Altermatt HJ, HukeeMJ, VandenBerg SR, Charlesworth JC. 1995. Tuber and subependymalgiant cell astrocytoma associated with tuberous sclerosis: an immunohis-tochemical, ultrastructural, and immunoelectron and microscopic study.Acta Neuropathol 90:387–399.
Horikawa K, Armstrong WE. 1988. A versatile means of intracellularlabeling: injection of biocytin and its detection with avidin conjugates.J Neurosci Methods 25:1–11.
Jacobs KM, Gutnick MJ, Prince DA. 1996. Hyperexcitability in a model ofcortical maldevelopment. Cereb Cortex 6:514–523.
Kerfoot C, Vinters HV, Mathern GW. 1999. Cerebral cortical dysplasia:giant neurons show potential for increased excitation and axonal plasticity.Dev Neurosci 21:260–270.
Kettenmann H. 1999. Physiology of glial cells. Adv Neurol 79:565–571.Kornack DR, Rakic P. 1995. Radial and horizontal deployment of clonally
related cells in the primate neocortex: relationship to distinct mitoticlineages. Neuron 15:311–321.
Kostovic I, Rakic P. 1990. Developmental history of the transient subplatezone in the visual and somatosensory cortex of the macaque monkey andhuman brain. J Comp Neurol 297:441–470.
Lee KS, Schottler F, Collins JL, Lanzino G, Couture D, Rao A, HiramatsuK, Goto Y, Hong SC, Caner H, Yamamoto H, Chen ZF, Bertram E,Berr S, Omary R, Scrable H, Jackson T, Goble J, Eisenman L. 1997. Agenetic animal model of human neocortical heterotopia associated withseizures. J Neurosci 17:6236–6242.
Lippa CF, Pearson D, Smith TW. 1993. Cortical tubers demonstratereduced immunoreactivity for synapsin I. Acta Neuropathol 85:449–451.
Luhmann HJ, Reiprich RA, Hanganu I, Kilb W. 2000. Cellular physiologyof the neonatal rat cerebral cortex: intrinsic membrane properties, sodiumand calcium currents. J Neurosci Res 62:574–584.
Machado-Salas JP. 1984. Abnormal dendritic patterns and aberrant spinedevelopment in Bourneville’s disease—a Golgi survey. Clin Neuropathol3:52–58.
Malatesta P, Hartfuss E, Gotz M. 2000. Isolation of radial glial cells byfluorescent-activated cell sorting reveals a neuronal lineage. Development127:5253–5263.
Marin-Padilla M. 1983. Structural organization of the human cerebralcortex prior to the appearance of the cortical plate. Anat Embryol 168:21–40.
Marin-Padilla M. 1999. Developmental neuropathology and impact ofperinatal brain damage. III: gray matter lesions of the neocortex. J Neu-ropathol Exp Neurol 58:407–429.
Mathern GW, Giza CC, Yudovin S, Vinters HV, Peacock WJ, ShewmonDA, Shields WD. 1999. Postoperative seizure control and antiepilepticdrug use in pediatric epilepsy surgery patients: the UCLA experience,1986–1997. Epilepsia 40:1740–1749.
Mathern GW, Cepeda C, Hurst RS, Flores-Hernandez J, Mendoza D,Levine MS. 2000. Neurons recorded from pediatric epilepsy surgerypatients with cortical dysplasia. Epilepsia 41:S162–S167.
Mattia D, Olivier A, Avoli M. 1995. Seizure-like discharges recorded inhuman dysplastic neocortex maintained in vitro. Neurology 45:1391–1395.
Mischel PS, Nguyen LP, Vinters HV. 1995. Cerebral cortical dysplasiaassociated with pediatric epilepsy. Review of neuropathologic featuresand proposal for a grading system. J Neuropathol Exp Neurol 54:137–153.
Mizuguchi M, Takashima S. 2001. Neuropathology of tuberous sclerosis.Brain Dev 23:508–515.
Mrzljak L, Uylings HB, Kostovic I, Van Eden CG. 1988. Prenatal devel-opment of neurons in the human prefrontal cortex: I. A qualitative Golgistudy. J Comp Neurol 271:355–386.
Abnormal Cells in Human Cortical Dysplasia 485

Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR.2001. Neurons derived from radial glial cells establish radial units inneocortex. Nature 409:714–720.
Powers RK, Binder MD. 2003. Persistent Na� and Ca2� currents in rathypoglossal motoneurons. J Neurophysiol 89:615–624.
Roper SN, Gilmore RL, Houser CR. 1995. Experimentally induceddisorders of neuronal migration produce an increased propensity forelectrographic seizures in rats. Epilepsy Res 21:205–219.
Sancini G, Franceschetti S, Battaglia G, Colacitti C, Di Luca M, SpreaficoR, Avanzini G. 1998. Dysplastic neocortex and subcortical heterotopias inmethylazoxymethanol-treated rats: an intracellular study of identifiedpyramidal neurones. Neurosci Lett 246:181–185.
Sayer RJ, Brown AM, Schwindt PC, Crill WE. 1993. Calcium currents inacutely isolated human neocortical neurons. J Neurophysiol 69:1596–1606.
Schwartzkroin PA, Walsh CA. 2000. Cortical malformations and epilepsy.Ment Retard Dev Disabil Res Rev 6:268–280.
Speckmann EJ, Straub H, Kohling R. 1993. Contribution of calcium ionsto the generation of epileptic activity and antiepileptic calcium antago-nism. Neuropsychobiology 27:122–126.
Spreafico R, Battaglia G, Arcelli P, Andermann F, Dubeau F, Palmini A,Olivier A, Villemure JG, Tampieri D, Avanzini G, Avoli M. 1998.Cortical dysplasia: an immunocytochemical study of three patients. Neu-rology 50:27–36.
Straub H, Hohling JM, Kohling R, Lucke A, Tuxhorn I, Ebner A, Wolf P,Pannek H, Oppel F, Speckmann EJ. 2000. Effects of nifedipine onrhythmic synchronous activity of human neocortical slices. Neuroscience100:445–452.
Tasker JG, Hoffman NW, Kim YI, Fisher RS, Peacock WJ, Dudek FE.
1996. Electrical properties of neocortical neurons in slices from childrenwith intractable epilepsy. J Neurophysiol 75:931–939.
Taylor DC, Falconer MA, Bruton CJ, Corsellis JA. 1971. Focal dysplasia ofthe cerebral cortex in epilepsy. J Neurol Neurosurg Psychiatry 34:369–387.
Tse FW, Fraser DD, Duffy S, MacVicar BA. 1992. Voltage-activated K�
currents in acutely isolated hippocampal astrocytes. J Neurosci 12:1781–1788.Van der Loos H. 1965. The “improperly” oriented pyramidal cell in the
cerebral cortex and its possible bearing on problems of neuronal growthand cell orientation. Bull John Hopkins Hosp 117:228–250.
Vinters HV, Fisher RS, Cornford ME, Mah V, Secor DL, De Rosa MJ,Comair YG, Peacock WJ, Shields WD. 1992. Morphological substrates ofinfantile spasms: studies based on surgically resected cerebral tissue. ChildNerv Syst 8:8–17.
Vinters HV, Park SH, Johnson MW, Mischel PS, Catania M, Kerfoot C.1999. Cortical dysplasia, genetic abnormalities and neurocutaneous syn-dromes. Dev Neurosci 21:248–259.
Walsh C, Cepko CL. 1988. Clonally related cortical cells show severalmigration patterns. Science 241:1342–1345.
Wuarin JP, Kim YI, Cepeda C, Tasker JG, Walsh JP, Peacock WJ,Buchwald NA, Dudek FE. 1990. Synaptic transmission in human neo-cortex removed for treatment of intractable epilepsy in children. AnnNeurol 28:503–511.
Ying Z, Babb TL, Comair YG, Bingaman W, Bushey M, Touhalisky K.1998. Induced expression of NMDAR2 proteins and differential expres-sion of NMDAR1 splice variants in dysplastic neurons of human epilepticneocortex. J Neuropathol Exp Neurol 57:47–62.
Zhou M, Kimelberg HK. 2000. Freshly isolated astrocytes from rat hip-pocampus show two distinct current patterns and different [K�]o uptakecapabilities. J Neurophysiol 84:2746–2757.
486 Cepeda et al.




















