
More than meat: diversity in caprine harvesting strategies and the emergence of complex production...
26
More than meat: diversity in caprine harvesting strategies and the emergence of complex production systems during the Late Pre-Pottery Neolithic B Cheryl A. Makarewicz This paper examines the diversity of goat and sheep husbandry practices in southern Jordan during the Late Pre-Pottery Neolithic B (Late PPNB), immediately following the domestication of these animals. Although the predominant view is one of sheep and goat husbandry as a relatively simple affair focused on the production of meat through the slaughter of young adult animals, results presented here reveal a multi-faceted system that included use of specialized harvesting strategies, direct manipulation of the caprine diet through provisioning, and the disarticulation of herds. The management strategies used by herders differed between Late PPNB settlements and were variously tuned for the production of dairy, meat, ‘tender meat’, and possibly surplus animals but overall adhered to strategies designed for risk reduction. Such complexity in pastoralist behaviour, and particularly the possibility of surplus production, suggests that a re-evaluation of the role of caprine husbandry in Late PPNB economic and social structures is necessary. Keywords: animal husbandry practices, sheep and goat management systems, harvesting, zooarchaeology, Neolithic Introduction Domestic plant and animal technologies were primary features of early Neolithic subsistence systems, provid- ing households and communities with a ready and steady source of food products throughout the year. In the southern Levant, the exploitation of domesticates eclipsed the use of wild plant and animal resources during the Late PPNB (c. 7250–6700 cal BC), when domestic animal herding first coalesced with barley and wheat agriculture to create what later became the foundational blocks of agro-pastoralist subsistence economies (Horwitz et al. 1999; Kuijt and Goring- Morris 2002). For the first time in the region, sheep and goat herds became a central and ubiquitous component of Late PPNB subsistence systems, and these flocks provisioned people with a regular source of meat (Becker 2004; Horwitz et al. 1999; Makarewicz 2009). Yet, despite the newly salient role of combined sheep and goat herding in PPNB subsistence systems, surprisingly little is known about the character and range of animal husbandry practices used to manage caprine herds during the Late PPNB. For the most part, archaeologists have been content to characterize Late PPNB caprine husbandry as a relatively unsophisti- cated endeavour that consisted primarily of a simple strategy narrowly focused on the slaughter of juvenile animals in order to produce a renewable source of meat (Bar-Yosef 2000; von den Driesch and Wodtke 1997). Instead, it is likely that novel and complex food production strategies — including those related to livestock herding — were developed during the Late PPNB in order to mitigate the effects of new resource pressure on wild and domestic foodstuffs, as well as emergent social stresses, associated with the prolif- eration of large settlements within a relatively small region, hypothesized increases in population (Kuijt 2008), increased territorialization and possibly the advent of land tenure (Goring-Morris and Belfer- Cohen 2008). Rather than repetitively following a concretized configuration of static subsistence strate- gies, which would have required deep negotiation of newly pressured and contested spaces and resources, Cheryl A. Makarewicz, Institute for Prehistoric and Protohistoric Archaeology, Christian Albrechts University, Johanna-Mestorf Strasse 2– 6, Kiel 24118, Germany; email: [email protected] ßCouncil for British Research in the Levant 2013 Published by Maney 236 Levant 2013 VOL 45 NO 2 DOI 10.1179/0075891413Z.00000000027
Transcript of More than meat: diversity in caprine harvesting strategies and the emergence of complex production...

More than meat: diversity in caprineharvesting strategies and the emergence ofcomplex production systems during the LatePre-Pottery Neolithic B
Cheryl A. Makarewicz
This paper examines the diversity of goat and sheep husbandry practices in southern Jordan
during the Late Pre-Pottery Neolithic B (Late PPNB), immediately following the domestication of
these animals. Although the predominant view is one of sheep and goat husbandry as a relatively
simple affair focused on the production of meat through the slaughter of young adult animals,
results presented here reveal a multi-faceted system that included use of specialized harvesting
strategies, direct manipulation of the caprine diet through provisioning, and the disarticulation of
herds. The management strategies used by herders differed between Late PPNB settlements and
were variously tuned for the production of dairy, meat, ‘tender meat’, and possibly surplus animals
but overall adhered to strategies designed for risk reduction. Such complexity in pastoralist
behaviour, and particularly the possibility of surplus production, suggests that a re-evaluation of
the role of caprine husbandry in Late PPNB economic and social structures is necessary.
Keywords: animal husbandry practices, sheep and goat management systems, harvesting, zooarchaeology, Neolithic
Introduction
Domestic plant and animal technologies were primary
features of early Neolithic subsistence systems, provid-
ing households and communities with a ready and
steady source of food products throughout the year. In
the southern Levant, the exploitation of domesticates
eclipsed the use of wild plant and animal resources
during the Late PPNB (c. 7250–6700 cal BC), when
domestic animal herding first coalesced with barley and
wheat agriculture to create what later became the
foundational blocks of agro-pastoralist subsistence
economies (Horwitz et al. 1999; Kuijt and Goring-
Morris 2002). For the first time in the region, sheep and
goat herds became a central and ubiquitous component
of Late PPNB subsistence systems, and these flocks
provisioned people with a regular source of meat
(Becker 2004; Horwitz et al. 1999; Makarewicz 2009).
Yet, despite the newly salient role of combined sheep
and goat herding in PPNB subsistence systems,
surprisingly little is known about the character and
range of animal husbandry practices used to manage
caprine herds during the Late PPNB. For the most part,
archaeologists have been content to characterize Late
PPNB caprine husbandry as a relatively unsophisti-
cated endeavour that consisted primarily of a simple
strategy narrowly focused on the slaughter of juvenile
animals in order to produce a renewable source of meat
(Bar-Yosef 2000; von den Driesch and Wodtke 1997).
Instead, it is likely that novel and complex food
production strategies — including those related to
livestock herding — were developed during the Late
PPNB in order to mitigate the effects of new resource
pressure on wild and domestic foodstuffs, as well as
emergent social stresses, associated with the prolif-
eration of large settlements within a relatively small
region, hypothesized increases in population (Kuijt
2008), increased territorialization and possibly the
advent of land tenure (Goring-Morris and Belfer-
Cohen 2008). Rather than repetitively following a
concretized configuration of static subsistence strate-
gies, which would have required deep negotiation of
newly pressured and contested spaces and resources,
Cheryl A. Makarewicz, Institute for Prehistoric and ProtohistoricArchaeology, Christian Albrechts University, Johanna-Mestorf Strasse 2–6, Kiel 24118, Germany; email: [email protected]
�Council for British Research in the Levant 2013Published by Maney
236 Levant 2013 VOL 45 NO 2 DOI 10.1179/0075891413Z.00000000027

Late PPNB herders looked for new ways to squeeze
more out of existing resources and exploit new
resource patches and environments. In addition to
developing new animal husbandry practices that
widened herd access to graze, herders could diversify
and enhance food production, beyond the subsistence
base, by developing management strategies that
focused on accessing different types and qualities of
ante-mortem products from animals. By developing
management practices that focused on these issues,
herders could diversify and enhance food production
beyond the subsistence base.
Here, the range and diversity in sheep and goat
management strategies used by herders in an increas-
ingly crowded and built Late PPNB environment are
examined through an evaluation of taxonomic,
ageing, and biometrical data obtained from faunal
assemblages recovered from several major settlements
located in central and southern Jordan. These
zooarchaeological data provide important insights
into herd demographic composition and timing of
animal slaughter, both critical facets of animal
management systems that can be readily manipulated
by herders to achieve certain production goals, and to
tune herds to specific environmental and graze
conditions. In addition, light stable isotopic data,
which provides information on animal diet, are
examined in order to better understand the role, if
any, of fodder provisioning in Late PPNB herding
systems. Husbandry practices, such as foddering,
expand the accessibility of quality graze to animals
outside of periods of natural availability, and through
foddering, herders can maintain and increase animal
health and production.
Through detailed documentation of caprine herd-
ing strategies in use during the Late PPNB, it is
possible to build a finely textured understanding of
how households and communities manipulated and
extracted resources from their animal subsistence
base and, in turn, investigate how variation in caprine
management practices may have contributed to
broader Pre-Pottery Neolithic social and economic
transformations.
The Late PPNB
At approximately 7200 cal BC, a constellation of
exceptionally large and architecturally dense settle-
ments appeared throughout southern Jordan, herald-
ing the emergence of new social and economic
systems, subsistence technologies, and landscape
organization in the region (Fig. 1). Small settlements
containing circular architecture, typical for Middle
PPNB sites located within the Irano-Turanian zone
of southern Jordan, gave way to large (.10 ha),
agglutinative settlements characterized by two-
storey, neatly coursed, dressed stone architecture.
In the Mediterranean zone of central Jordan, where
the Middle PPNB was expressed quite differently
compared to the south, largely single-storey, isolated
rectilinear structures gave way to a denser arrange-
Figure 1 Map of Jordan indicating the location of Late PPNB sites mentioned in the text
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 237

ment of two-storey architecture during the Late
PPNB, as seen at ‘Ain Ghazal and possibly Wadi
Shu’eib. These large settlements may have served as
central places through which people, ideas, technol-
ogy, and material goods flowed, although it remains
to be established whether these settlements were
organized hierarchically as suggested by Gebel
(2004) and Bienert (2004).
The reasons for the apparently rapid coalescence
and proliferation of large Late PPNB settlements to
the east of the Jordan Valley are poorly understood.
Based on burial densities and estimated site areas, it is
likely that Late PPNB population levels were higher
than Middle PPNB ones (Bocquet-Appel 2002; Eshed
et al. 2004; Kuijt 2008). However, the reasons why
populations increased remains under debate. Some
resource stress models suggest that the Mediterranean
zone west of the Jordan Valley had become depleted of
plant and animal resources by the end of the Middle
PPNB, causing entire communities to uproot and
migrate to central and southern Jordan (Gebel 2004;
Rollefson 2010). However, these models fail to consider
the possibility that a new system of landscape organi-
zation favouring local population aggregation in south-
ern Jordan emerged during the Late PPNB. New
excavations in the Galilee, coupled with new radio-
carbon determinations and lithic techno-typological
analyses focused on establishing the cultural-chronology
of sites in these regions, strongly suggest that the
Mediterranean region was not abandoned as previously
thought. Settlements dating to the Late PPNB, both
newly established communities and ones exhibiting
occupational continuity from the Middle PPNB, are
distributed across the entire region (Barzilai and Getzov
2011; Bocquentin et al. 2011; Khalaily et al. 2008;
Milevski et al. 2008). The Mediterranean zone clearly
remained an area of human occupation during the
entirety of the Late PPNB, and the higher carrying
capacity and regenerative capabilities of Mediterranean
ecosystems make it unlikely that the area suffered a
major decline in population levels.
In addition to the proliferation of Late PPNB
settlements in central and southern Jordan, the first
domestic livestock may have appeared in the eastern
steppes (or deserts) at this time, as there is robust
evidence for intensive caprine herding in the region
during the PPNC (Early Late Neolithic in the steppic
chronological sequence) (Fujii 2009; Garrard et al.
1994; Horwitz et al. 1999). Although inadequate
chronological resolution prevents us from determining
exactly how and if these two separate events were
related (e.g. did nascent herding activities in the Azraq
Basin coincide with, or follow, the emergence of large
Late PPNB settlements?), and the underlying factors
driving this settlement process remained debated, it is
evident that people depending on domestic animal
exploitation first expanded into desert areas during the
Late PPNB (Fujii 2009). What remains unknown,
however, is the degree to which these activities were
linked to large agglutinative settlements and the socio-
economic relationship that may have existed between
the two. What does seem likely, however, is that the
more extensive use of desert areas too marginal for
intensive agricultural activity and the explosion of
large-scale settlements east of the Jordan Valley are
indicative of new pressures placed on local plant and
animal resources.
The cultural changes associated with the Late
PPNB extended beyond shifts in settlement patterns
and architectural organization to include other
developments as well. The scale of storage increased
dramatically during the Late PPNB, and extensive
storage facilities were a ubiquitous feature of Late
PPNB settlements. Consisting of interior ground
floor rooms c. 1.5 sq m in size, these storage
installations were substantial, pre-planned invest-
ments that were incorporated into the very fabric of
domestic structures. Although it is unclear exactly
what was stored within these small spaces, e.g. food,
fuel, or other goods, the proliferation of such large
storage spaces suggests an improved ability to
produce food due to new technological developments
and/or an intentional intensification in plant cultiva-
tion and animal exploitation activities. Mortuary
practices also changed markedly, so that for the first
time, some personal adornments such as bracelets
and shell and stone beads were placed with the dead,
while skull removal and skull plastering, a common
practice during the Middle PPNB that promoted
social levelling, ceased (Kuijt and Goring-Morris
2002). These new mortuary practices may indicate a
transformation in social structure to emphasize
individual household action over community social
cohesion.
Late PPNB animal exploitation strategies
Underlying these pronounced shifts in Late PPNB
settlement patterns and social organization, and
perhaps even precipitating these transformations,
are significant changes in the plant and animal
exploitation strategies used by people inhabiting
these large communities in southern Jordan. There
is a major, perhaps even threshold-crossing, shift in
subsistence approaches from strategies defined by
light management of plants and animals and some
exploitation of wild resources, to strategies heavily
Makarewicz More than meat
238 Levant 2013 VOL 45 NO 2

rooted in use-domesticated plants and animals (Kuijt
and Goring-Morris 2002). Cereal and legume agri-
culture, which included tending of domesticated
barley (Hordeum spontaneum), emmer wheat
(Triticum dicoccum), lentils (Lens culinaris), and pea
(Pisum sativum) emerged as the dominant mode of
plant exploitation, although gathering of wild cereals,
legumes, fruits, and nuts remained important plant
exploitation strategies (Makarewicz et al. 2006; Neef
2004; White and Wolff 2012).
Ungulate hunting and experimentation with goat
management, pursued during the Middle PPNB, gave
way to strategies focused on the use of domesticated
sheep and goats during the Late PPNB (Becker 2004;
Horwitz et al. 1999; Makarewicz 2009). The impor-
tance of caprine exploitation rose sharply during the
Late PPNB, so that for the first time intensive goat
and sheep herding became the dominant mode of
animal exploitation in the southern Levant (Horwitz
et al. 1999). On average, caprines comprise approxi-
mately 85% of animal remains recovered from Late
PPNB sites, compared to 45% during the preceding
Middle PPNB. Also for the first time during the Late
PPNB, morphologically domestic goats exhibiting
small body size and twisted horn-cores were herded,
as were a totally new animal technology, domestic
sheep, which were either intentionally imported or
rapidly diffused into the southern Levant from the
northern Levant (Horwitz and Ducos 1998). Nascent
management of cattle east of the Jordan Valley may
have also begun during the Late PPNB, although it
remains to be seen whether husbanded cattle derived
from local wild aurochs stocks or were imported from
the north (Becker 1991; Horwitz and Ducos 2005).
Hunting of wild game remained an important facet of
animal exploitation systems, and wild goats and
gazelle may have provided a fall-back food resource
that could be accessed by herders as they experi-
mented with developing new caprine herding strate-
gies (Makarewicz 2009). Pigs do not generally feature
in Late PPNB animal exploitation systems in south-
ern Jordan due to the absence of the well-watered
environments that they require.
The new reliance on goats and sheep during the
Late PPNB may have sparked new developments in
animal management systems that sought to enhance
the productive and reproductive potential of live-
stock. By diversifying animal husbandry practices,
herders could reduce their subsistence risk and even
increase the productive output of their animal herds
in an increasingly crowded and built environment.
Herders are involved in complex management deci-
sions that — if economically optimized — aim to
promote the long-term security of their animal herds,
while maintaining sufficient off-take of a variety of
ante- and postmortem animal products for their own
needs. Of course, environmental conditions or social
obligations may result in husbandry decisions that
are at odds with optimal herd management. In either
case, however, herders attempt to control herd
production and reproduction by applying and adjust-
ing the type, timing, and intensity of a series of
interlinked husbandry practices such as harvesting,
weaning, fodder provisioning, winter pasturage, and
mobility — all of which are detectable in the faunal
record through application of zooarchaeological and
isotopic analyses (Arbuckle et al. 2009; Balasse and
Tresset 2002; Makarewicz in press; 2009; Makarewicz
and Tuross 2006; 2009).
Zooarchaeological methodology
In order to establish the sheep and goat management
strategies in use at Late PPNB settlements east of the
Jordan Valley, sheep to goat ratios, animal age at
time of slaughter, and herd sex composition were
examined. These variables are a measure of herder
decisions regarding animal product production, herd
maintenance, and risk reduction since each taxon
exhibits different physiological, reproductive, and
productive qualities (Dahl and Hjort 1976; Redding
1981). Goats, for example, have a higher reproductive
rate, produce more milk, and adapt more easily to
marginal environmental conditions compared to
sheep, which produce fewer offspring and lower meat
and milk yields, but generate higher quality meat and
milk (Redding 1981).
The biological disparities between sheep and goats
means that strategies for herd security and produc-
tion will be different for each taxon. Consequently,
herders adjust the ratio of sheep to goat in their
flocks, and the timing of animal kill-off according to
local environmental conditions and their subsistence
needs. Models that take into consideration the
productive potential of caprines predict that survivor-
ship curves for goats will exhibit a younger kill-off
compared to curves calculated for sheep because of
the higher reproductive rate of goats (Redding 1981).
Along these lines, survivorship for a combined sheep/
goat category will indicate progressively younger kill-
off as the frequency of goat increases relative to
sheep.
In addition to baseline reproductive capacity, the
shape of survivorship curves is determined by the
production goals of herders, who accordingly slaugh-
ter sheep and goats through application of distinct
harvesting strategies, tuned for the production of
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 239

distinct ante- and post-mortem products. Payne’s
(1973) classic model derived from modern caprine
flocks optimized for the production of meat, milk and
wool predicts the intensive slaughter of juvenile males
if meat production is desired, kill-off of very young
male kids and lambs when maximizing dairy produc-
tion is the goal, and the general retention of both male
and female animals into later adulthood when fibre
production is the aim. However, herders frequently
may pursue several of these production goals simulta-
neously, or seek other production goals that do not
feature in Payne’s model, or consider other variables,
i.e. availability of fodder, pasture grounds, and labour,
that influence harvesting decisions. In addition,
herders may still achieve considerable production of
milk, meat, and/or wool even when not adhering to
optimizing harvesting strategies as described in
Payne’s model (Arbuckle et al. 2009; Makarewicz
2011; Vigne and Helmer 2007).
Additional models have been developed in an
attempt to isolate explicitly additional harvesting
strategies that herders may use for production goals
other than intensive milk, meat, and wool produc-
tion, including the ‘tender meat’ and ‘type B’ milk
models developed by Vigne and Helmer (2007). The
‘tender meat’ model indicates kill-off of young males
between the ages of 6–12 months. In contrast to the
classic ‘type A milk’ model as outlined by Payne, the
‘type B milk’ model is characterized by the delayed
kill-off of young animals throughout their first year.
This model predicts intense kill-off of young lambs
and kids between 6–12 months, and then again for
adult females between 2–4 years. Milk production
is probably one of the most challenging animal
exploitation strategies to identify in the zooarchaeo-
logical record, especially when herders are balanc-
ing multiple production goals. In addition to
taphonomic biases that may selectively destroy the
bones of young animals most likely to be culled as
part of a milk production strategy, there are a variety
of ways in which herders achieve milk production,
not all of which involve the slaughter of young
animals.
It is important to keep in mind that multiple
strategies with different production goals can pro-
duce similar mortality profiles, especially tender
meat, ‘type B milk’, and mixed meat and milk, as
herders balance between their subsistence goals,
social contexts, and environmental conditions
(Arbuckle et al. 2009). We should also remember
that the models described above provide a frame of
reference for interpreting production and manage-
ment goals from demographic data and should be
viewed as heuristic devices for understanding herder
decision-making in prehistory.
In this paper, new and previously published
taxonomic, age and biometric data sets for both
sheep and goats were (re)analysed in order to define
the range of variation in Late PPNB harvesting
practices, and to evaluate the role of animal age and
sex in shaping caprine kill-off strategies. Sheep and
goat bone and tooth specimens from each site were
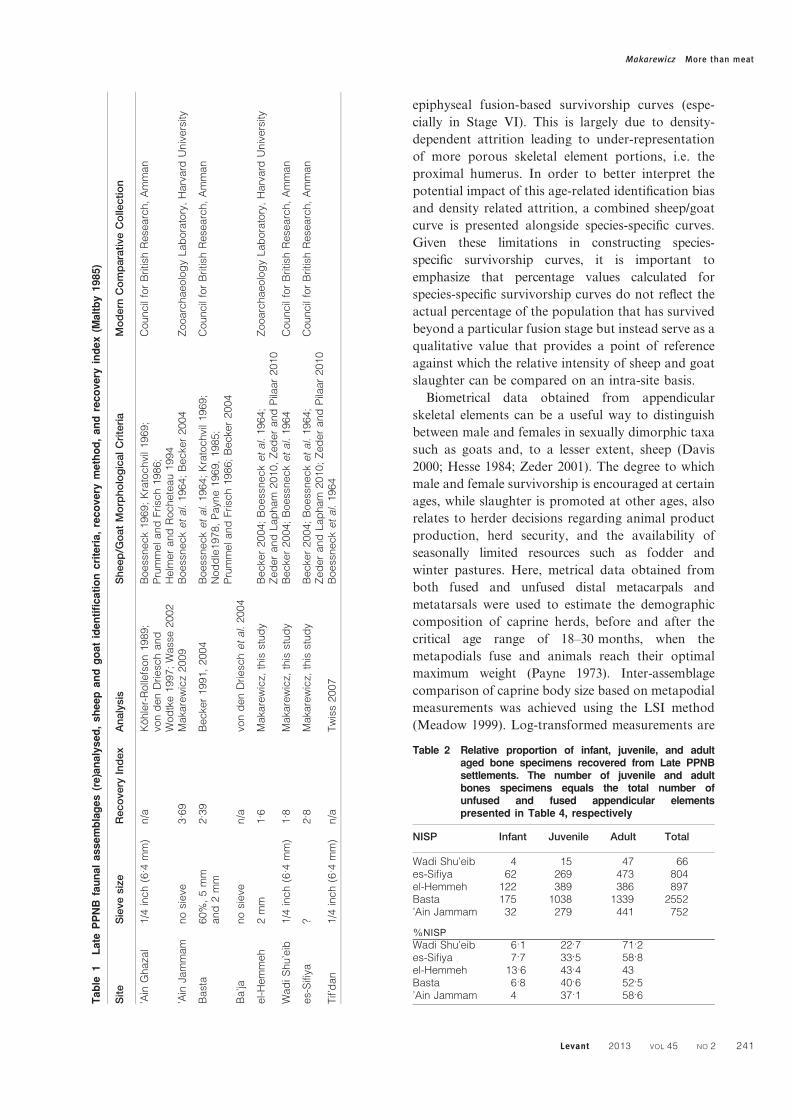
separated according to criteria described in Table 1.
The relative frequency of taxa identified for each site
was calculated based on NISP (number of individual
specimens) (Table 2). Ageing data derived from the
state of epiphyseal fusion and tooth wear and data
are routinely used to construct survivorship curves,
which establish the intensity of animal slaughter at
particular ages and the production goals of herders
(Hesse 1984; Payne 1973). Here, survivorship curves
for goats, sheep, and a combined sheep/goat category
were calculated according to the state of long bone
epiphyseal fusion according to Zeder (2006) and
mandibular tooth eruption wear according to Payne
(1973) (Tables 3 and 4). Survivorship curves based on
eruption and tooth wear data and calculated for a
combined sheep/goat category were derived from
single teeth, partial and complete mandibular rows in
the case of ‘Ain Jammam, Basta, Ba’ja and es-Sifiya.
The published zooarchaeological literature for ‘Ain
Ghazal and Ba’ja does not indicate how mandibular
ageing data were collected (von den Driesch et al.
2004; Wasse 2002).
For survivorship curves calculated according to
state of epiphyseal fusion, in order to ensure that the
youngest infant animals were not allocated to older
fusion stages, all appendicular skeletal bone speci-
mens (except the radius) displaying unfused epiphyses
and belonging to infant-aged and neonatal animals
were excluded from demographic analyses. In order
to broadly establish the harvesting decisions of
herders regarding the very youngest animals, the
relative proportions of infant-aged bone specimens to
juvenile (unfused) and adult (fused) specimens were
instead calculated (Table 5; Fig. 2).
Species-specific survivorship curves calculated for
sheep and goat are likely systematically biased
towards older kill-off due to under-representation
of unfused skeletal parts (Munson 2000). This bias is
caused by a combination of density-dependent
destruction of more fragile immature bones and
difficulties in distinguishing between the unfused
bones of sheep and goats, especially in the distal
humerus, proximal humerus and distal femur. In
some cases, there is a ‘resurrection effect’ visible in
Makarewicz More than meat
240 Levant 2013 VOL 45 NO 2
epiphyseal fusion-based survivorship curves (espe-
cially in Stage VI). This is largely due to density-
dependent attrition leading to under-representation
of more porous skeletal element portions, i.e. the
proximal humerus. In order to better interpret the
potential impact of this age-related identification bias
and density related attrition, a combined sheep/goat
curve is presented alongside species-specific curves.
Given these limitations in constructing species-
specific survivorship curves, it is important to
emphasize that percentage values calculated for
species-specific survivorship curves do not reflect the
actual percentage of the population that has survived
beyond a particular fusion stage but instead serve as a
qualitative value that provides a point of reference
against which the relative intensity of sheep and goat
slaughter can be compared on an intra-site basis.
Biometrical data obtained from appendicular
skeletal elements can be a useful way to distinguish
between male and females in sexually dimorphic taxa
such as goats and, to a lesser extent, sheep (Davis
2000; Hesse 1984; Zeder 2001). The degree to which
male and female survivorship is encouraged at certain
ages, while slaughter is promoted at other ages, also
relates to herder decisions regarding animal product
production, herd security, and the availability of
seasonally limited resources such as fodder and
winter pastures. Here, metrical data obtained from
both fused and unfused distal metacarpals and
metatarsals were used to estimate the demographic
composition of caprine herds, before and after the
critical age range of 18–30 months, when the
metapodials fuse and animals reach their optimal
maximum weight (Payne 1973). Inter-assemblage
comparison of caprine body size based on metapodial
measurements was achieved using the LSI method
(Meadow 1999). Log-transformed measurements are
Ta
ble
1L
ate
PP
NB
fau
na
la
ss
em
bla
ge
s(r
e)a
na
lys
ed
,s
he
ep
an
dg
oa
tid
en
tifi
ca
tio
nc
rite
ria
,re
co
ve
rym
eth
od
,a
nd
rec
ov
ery
ind
ex
(Ma
ltb
y1
98
5)
Sit
eS
ieve
siz
eR
eco
very
Ind
ex
An
aly
sis
Sh
eep
/Go
at
Mo
rph
olo
gic
al
Cri
teri
aM
od
ern
Co
mp
ara
tive
Co
llecti
on
’Ain
Ghazal
1/4
inch
(6. 4
mm
)n/a
Kohle
r-R
olle
fson
1989;
von
den
Driesch
and
Wod
tke
1997;
Wasse
2002
Boessneck
1969;
Kra
tochvil
1969;
Pru
mm
eland
Frisch
1986;
Helm
er
and
Rochete
au
1994
Council
for
British
Researc
h,
Am
man
’Ain
Jam
mam
no
sie
ve
3. 6
9M
akare
wic
z2009
Boessneck
et
al.
1964;
Becker
2004
Zooarc
haeolo
gy
Lab
ora
tory
,H
arv
ard
Univ
ers
ity
Basta
60%
,5
mm
and
2m
m2. 3
9B
ecker
1991,
2004
Boessneck
et
al.
1964;
Kra
tochvil
1969;
Nod
dle
1978,
Payne
1969,
1985;
Pru
mm
eland
Frisch
1986;
Becker
2004
Council
for
British
Researc
h,
Am
man
Ba’ja
no
sie
ve
n/a
von
den
Driesch
et
al.
2004
el-H
em
meh
2m
m1. 6
Makare
wic
z,
this
stu
dy
Becker
2004;
Boessneck
et
al.
1964;
Zed
er
and
Lap
ham
2010,
Zed
er
and
Pila
ar
2010
Zooarc
haeolo
gy
Lab
ora
tory
,H
arv
ard
Univ
ers
ity
Wad
iS
hu’e
ib1/4
inch
(6. 4
mm
)1. 8
Makare
wic
z,
this
stu
dy
Becker
2004;
Boessneck
et
al.
1964
Council
for
British
Researc
h,
Am
man
es-S
ifiy
a?
2. 8
Makare
wic
z,
this
stu
dy
Becker
2004;
Boessneck
et
al.
1964;
Zed
er
and
Lap
ham
2010;
Zed
er
and
Pila
ar
2010
Council
for
British
Researc
h,
Am
man
Tif’d
an
1/4
inch
(6. 4
mm
)n/a
Tw
iss
2007
Boessneck
et
al.
1964
Table 2 Relative proportion of infant, juvenile, and adultaged bone specimens recovered from Late PPNBsettlements. The number of juvenile and adultbones specimens equals the total number ofunfused and fused appendicular elementspresented in Table 4, respectively
NISP Infant Juvenile Adult Total
Wadi Shu’eib 4 15 47 66es-Sifiya 62 269 473 804el-Hemmeh 122 389 386 897Basta 175 1038 1339 2552’Ain Jammam 32 279 441 752
%NISP
Wadi Shu’eib 6.1 22.7 71.2es-Sifiya 7.7 33.5 58.8el-Hemmeh 13.6 43.4 43Basta 6.8 40.6 52.5’Ain Jammam 4 37.1 58.6
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 241
compared with those from a standard animal, a
female Ovis orientalis and the averaged measurements
of male and female Capra aegagrus, presented by
Uerpmann and Uerpmann (1994).
Previously published metacarpal and metatarsal
proximal and distal breadth data from Basta, Ba’ja,
and Tif’dan were used to document the size, age and
sex of animals harvested. Since specimens from Basta
and Ba’ja were measured before Zeder’s (2001) work
indicating that the unfused bones of juvenile males
are larger than the fused bones of females in all but
the youngest animals, only data from fused distal
metapodials were available for analysis. Despite the
absence of this valuable data set, it is still possible to
isolate distinct kill-off patterns for male and female
animals, i.e. the fused bones of small females will
dominate the metrical distribution when the harvesting
strategy focuses on the removal of young males
(Arbuckle, this volume).
Due to the high fragmentation and overall smaller
size of the faunal assemblages from el-Hemmeh, ‘Ain
Jammam, and es-Sifiya, the number of fused and
unfused metapodials producing distal breadth mea-
surements was generally low and certainly too few to
generate meaningful metrical distributions. In order
to amplify metapodial sample size, metrical data
from the metacarpus and metatarsus DVM (antero-
posterior diameter of the verticilus of the medial
condyle) and DVL (antero-posterior diameter of the
verticilus of the lateral condyle) were taken (Davis
2000). In modern wild sheep and goats from the Near
East and in PPNB caprines from southern Jordan,
there is small but consistent metrical offset of 0.2 to
0.4 mm between the DVM and DVL, with the lateral
condyle generally exhibiting a larger overall size
(Makarewicz and Arbuckle, unpublished data). Since
measurements from individual bone specimens are
grouped in intervals of 0?5 mm in order to generate
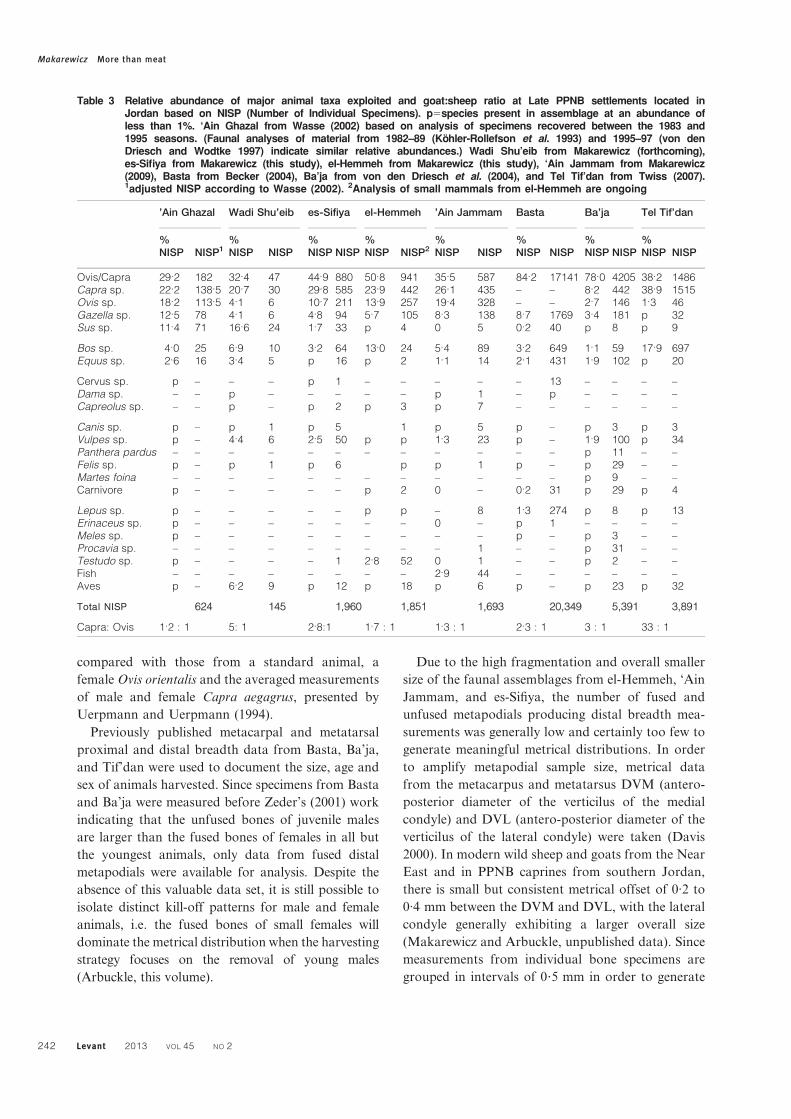
Table 3 Relative abundance of major animal taxa exploited and goat:sheep ratio at Late PPNB settlements located inJordan based on NISP (Number of Individual Specimens). p5species present in assemblage at an abundance ofless than 1%. ‘Ain Ghazal from Wasse (2002) based on analysis of specimens recovered between the 1983 and1995 seasons. (Faunal analyses of material from 1982–89 (Kohler-Rollefson et al. 1993) and 1995–97 (von denDriesch and Wodtke 1997) indicate similar relative abundances.) Wadi Shu’eib from Makarewicz (forthcoming),es-Sifiya from Makarewicz (this study), el-Hemmeh from Makarewicz (this study), ‘Ain Jammam from Makarewicz(2009), Basta from Becker (2004), Ba’ja from von den Driesch et al. (2004), and Tel Tif’dan from Twiss (2007).1adjusted NISP according to Wasse (2002). 2Analysis of small mammals from el-Hemmeh are ongoing
’Ain Ghazal Wadi Shu’eib es-Sifiya el-Hemmeh ’Ain Jammam Basta Ba’ja Tel Tif’dan
%NISP NISP1
%NISP NISP
%NISP NISP
%NISP NISP2
%NISP NISP
%NISP NISP
%NISP NISP
%NISP NISP
Ovis/Capra 29.2 182 32.4 47 44.9 880 50.8 941 35.5 587 84.2 17141 78.0 4205 38.2 1486Capra sp. 22.2 138.5 20.7 30 29.8 585 23.9 442 26.1 435 – – 8.2 442 38.9 1515Ovis sp. 18.2 113.5 4.1 6 10.7 211 13.9 257 19.4 328 – – 2.7 146 1.3 46Gazella sp. 12.5 78 4.1 6 4.8 94 5.7 105 8.3 138 8.7 1769 3.4 181 p 32Sus sp. 11.4 71 16.6 24 1.7 33 p 4 0 5 0.2 40 p 8 p 9
Bos sp. 4.0 25 6.9 10 3.2 64 13.0 24 5.4 89 3.2 649 1.1 59 17.9 697Equus sp. 2.6 16 3.4 5 p 16 p 2 1.1 14 2.1 431 1.9 102 p 20
Cervus sp. p – – – p 1 – – – – – 13 – – – –Dama sp. – – p – – – – – p 1 – p – – – –Capreolus sp. – – p – p 2 p 3 p 7 – – – – – –
Canis sp. p – p 1 p 5 1 p 5 p – p 3 p 3Vulpes sp. p – 4.4 6 2.5 50 p p 1.3 23 p – 1.9 100 p 34Panthera pardus – – – – – – – – – – – – p 11 – –Felis sp. p – p 1 p 6 p p 1 p – p 29 – –Martes foina – – – – – – – – – – – – p 9 – –Carnivore p – – – – – p 2 0 – 0.2 31 p 29 p 4
Lepus sp. p – – – – – p p – 8 1.3 274 p 8 p 13Erinaceus sp. p – – – – – – – 0 – p 1 – – – –Meles sp. p – – – – – – – – – p – p 3 – –Procavia sp. – – – – – – – – – 1 – – p 31 – –Testudo sp. p – – – – 1 2.8 52 0 1 – – p 2 – –Fish – – – – – – – – 2.9 44 – – – – – –Aves p – 6.2 9 p 12 p 18 p 6 p – p 23 p 32
Total NISP 624 145 1,960 1,851 1,693 20,349 5,391 3,891
Capra: Ovis 1.2 : 1 5: 1 2.8:1 1.7 : 1 1.3 : 1 2.3 : 1 3 : 1 33 : 1
Makarewicz More than meat
242 Levant 2013 VOL 45 NO 2
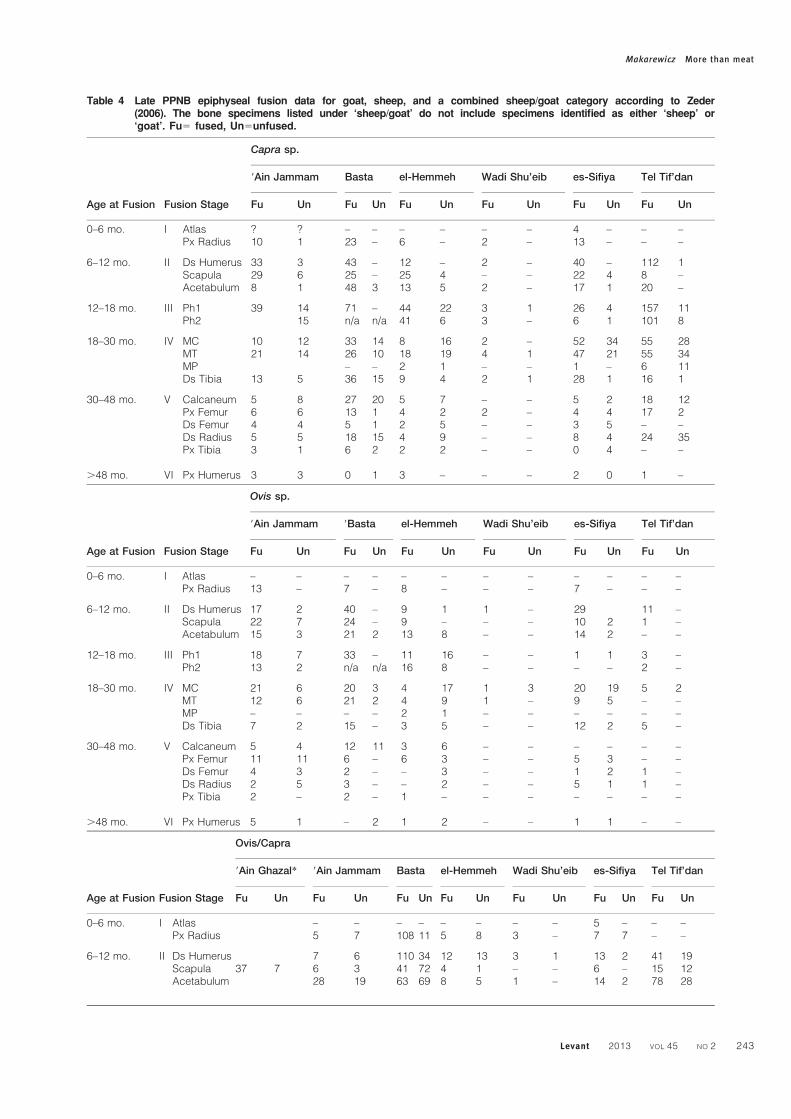
Table 4 Late PPNB epiphyseal fusion data for goat, sheep, and a combined sheep/goat category according to Zeder(2006). The bone specimens listed under ‘sheep/goat’ do not include specimens identified as either ‘sheep’ or‘goat’. Fu5 fused, Un5unfused.
Age at Fusion Fusion Stage
Capra sp.
9Ain Jammam Basta el-Hemmeh Wadi Shu’eib es-Sifiya Tel Tif’dan
Fu Un Fu Un Fu Un Fu Un Fu Un Fu Un
0–6 mo. I Atlas ? ? – – – – – – 4 – – –Px Radius 10 1 23 – 6 – 2 – 13 – – –
6–12 mo. II Ds Humerus 33 3 43 – 12 – 2 – 40 – 112 1Scapula 29 6 25 – 25 4 – – 22 4 8 –Acetabulum 8 1 48 3 13 5 2 – 17 1 20 –
12–18 mo. III Ph1 39 14 71 – 44 22 3 1 26 4 157 11Ph2 15 n/a n/a 41 6 3 – 6 1 101 8
18–30 mo. IV MC 10 12 33 14 8 16 2 – 52 34 55 28MT 21 14 26 10 18 19 4 1 47 21 55 34MP – – 2 1 – – 1 – 6 11Ds Tibia 13 5 36 15 9 4 2 1 28 1 16 1
30–48 mo. V Calcaneum 5 8 27 20 5 7 – – 5 2 18 12Px Femur 6 6 13 1 4 2 2 – 4 4 17 2Ds Femur 4 4 5 1 2 5 – – 3 5 – –Ds Radius 5 5 18 15 4 9 – – 8 4 24 35Px Tibia 3 1 6 2 2 2 – – 0 4 – –
.48 mo. VI Px Humerus 3 3 0 1 3 – – – 2 0 1 –
Age at Fusion Fusion Stage
Ovis sp.
9Ain Jammam 9Basta el-Hemmeh Wadi Shu’eib es-Sifiya Tel Tif’dan
Fu Un Fu Un Fu Un Fu Un Fu Un Fu Un
0–6 mo. I Atlas – – – – – – – – – – – –Px Radius 13 – 7 – 8 – – – 7 – – –
6–12 mo. II Ds Humerus 17 2 40 – 9 1 1 – 29 11 –Scapula 22 7 24 – 9 – – – 10 2 1 –Acetabulum 15 3 21 2 13 8 – – 14 2 – –
12–18 mo. III Ph1 18 7 33 – 11 16 – – 1 1 3 –Ph2 13 2 n/a n/a 16 8 – – – – 2 –
18–30 mo. IV MC 21 6 20 3 4 17 1 3 20 19 5 2MT 12 6 21 2 4 9 1 – 9 5 – –MP – – – – 2 1 – – – – – –Ds Tibia 7 2 15 – 3 5 – – 12 2 5 –
30–48 mo. V Calcaneum 5 4 12 11 3 6 – – – – – –Px Femur 11 11 6 – 6 3 – – 5 3 – –Ds Femur 4 3 2 – – 3 – – 1 2 1 –Ds Radius 2 5 3 – – 2 – – 5 1 1 –Px Tibia 2 – 2 – 1 – – – – – – –
.48 mo. VI Px Humerus 5 1 – 2 1 2 – – 1 1 – –
Age at Fusion Fusion Stage
Ovis/Capra
9Ain Ghazal* 9Ain Jammam Basta el-Hemmeh Wadi Shu’eib es-Sifiya Tel Tif’dan
Fu Un Fu Un Fu Un Fu Un Fu Un Fu Un Fu Un
0–6 mo. I Atlas – – – – – – – – 5 – – –Px Radius 5 7 108 11 5 8 3 – 7 7 – –
6–12 mo. II Ds Humerus 7 6 110 34 12 13 3 1 13 2 41 19Scapula 37 7 6 3 41 72 4 1 – – 6 – 15 12Acetabulum 28 19 63 69 8 5 1 – 14 2 78 28
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 243
histograms, this slight metrical difference has a
negligible impact on metrical distribution shape.
Analytical caveats
Demographic and biometric analyses are powerful
ways to uncover prehistoric animal management
strategies related to harvesting and animal product
production. It is, however, important to bear in mind
that central and southern Jordanian Late PPNB
faunal assemblages were formed in part through
hunting and herding activities, based on the exploita-
tion of three separate goat species: the domestic goat
(Capra hircus), the bezoar (C. aegagrus), and the ibex
(C. ibex). Unfortunately, these species are difficult, if
not nearly impossible, to distinguish from each other
morphologically in the appendicular skeleton. While
metrical determinations can help distinguish between
wild and domesticated animals to a certain extent,
there is marked variation in body size of early
Holocene southern Levantine wild goats (both bezoar
and ibex) that makes it difficult to definitively
separate domesticated goats from wild ones on
metrical grounds. For example, relatively small wild
animals, similar in size to Early Bronze Age
domesticates, are present at Wadi Faynan 16
(PPNA) and Wadi Mataha (Natufian), while very
large animals with body sizes exceeding that of wild
bezoar standards (see Uerpmann and Uerpmann
1994) are present at Basta and Beidha (PPNB)
(Baadsgaard 2000; Becker 2004; Carruthers 2002;
Makarewicz and Arbuckle, unpublished data).
Until the range of metrical variation in wild goat
species across different southern Levantine biomes
can be better established, goat survivorship curves
calculated for Late PPNB assemblages in this region
represent a mixture of specimens from domestic and
some wild goats and, consequently, reflect a conflation
of different exploitation strategies. It remains to be seen
exactly how wild goats were exploited during the Late
PPNB. These animals may have been hunted year-
round or on a seasonal basis, hunted through use of an
optimal acquisition strategy that focused on prime-
aged adults or via a less-restricted hunting strategy that
targeted both juvenile and adult animals. Alternatively,
morphologically wild animals may have been managed
in some way, as part of local domestication trajectories
that began in the southern Levant during the Middle
PPNB (Hecker 1982; Horwitz 2003; Makarewicz and
Tuross 2012). However, isolating how wild goats were
exploited during the Late PPNB requires a deeper
understanding of goat exploitation strategies for the
earlier PPN, particularly in southern Jordan, than is
currently available.
In addition to the analytical challenges that arise
when faced with the presence of three distinct, but
difficult to distinguish, goat species in the region,
there is a more intractable problem associated with
recovery bias when dealing with many Late PPNB
assemblages. Several faunal assemblages recovered
from Late PPNB sites suffer from significant recovery
bias due to a lack of systematic sieving of cultural
deposits removed during excavation or sieving using
screens with large mesh spacing (Table 1). It is well
known that faunal assemblages recovered without use
of a systematic sieving protocol, or with large meshed
sieves, are negatively impacted, and skeletal parts
belonging to the smaller, unfused bones of infant and
juvenile aged animals will be severely under-represented
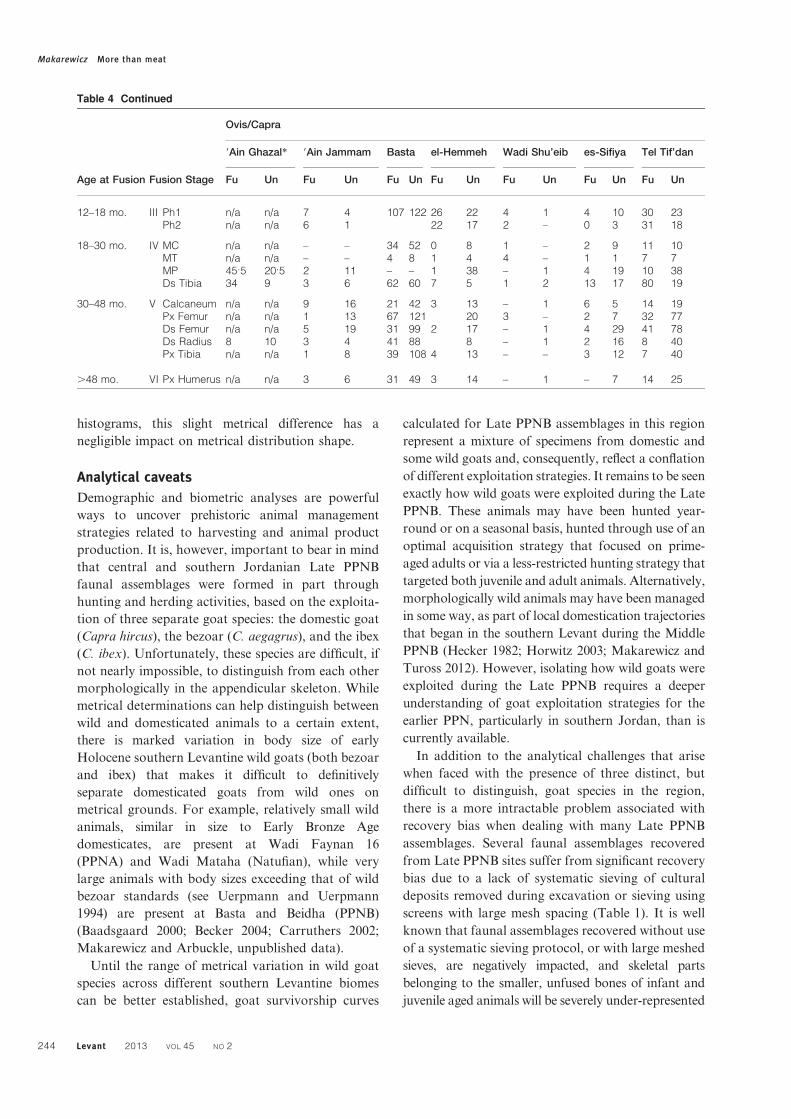
Age at Fusion Fusion Stage
Ovis/Capra
9Ain Ghazal* 9Ain Jammam Basta el-Hemmeh Wadi Shu’eib es-Sifiya Tel Tif’dan
Fu Un Fu Un Fu Un Fu Un Fu Un Fu Un Fu Un
12–18 mo. III Ph1 n/a n/a 7 4 107 122 26 22 4 1 4 10 30 23Ph2 n/a n/a 6 1 22 17 2 – 0 3 31 18
18–30 mo. IV MC n/a n/a – – 34 52 0 8 1 – 2 9 11 10MT n/a n/a – – 4 8 1 4 4 – 1 1 7 7MP 45.5 20.5 2 11 – – 1 38 – 1 4 19 10 38Ds Tibia 34 9 3 6 62 60 7 5 1 2 13 17 80 19
30–48 mo. V Calcaneum n/a n/a 9 16 21 42 3 13 – 1 6 5 14 19Px Femur n/a n/a 1 13 67 121 20 3 – 2 7 32 77Ds Femur n/a n/a 5 19 31 99 2 17 – 1 4 29 41 78Ds Radius 8 10 3 4 41 88 8 – 1 2 16 8 40Px Tibia n/a n/a 1 8 39 108 4 13 – – 3 12 7 40
.48 mo. VI Px Humerus n/a n/a 3 6 31 49 3 14 – 1 – 7 14 25
Table 4 Continued
Makarewicz More than meat
244 Levant 2013 VOL 45 NO 2

Ta
ble
5M
ort
ality
an
ds
urv
ivo
rsh
ipd
ata
for
sh
ee
p/g
oa
tb
as
ed
on
too
thw
ea
ra
cc
ord
ing
toP
ay
ne
(19
73
)
’Ain
Gh
aza
les-S
ifiy
ael-
Hem
meh
Basta
Ba’ja
’Ain
Jam
mam
MW
SA
ge
% Surv
ivo
r-sh
ip
% mo
rta-
lity
Sp
ecim
en
co
un
t
% Surv
ivo
r-sh
ip
% mo
rta-
lity
Sp
ecim
en
co
un
t
% Surv
ivo
r-sh
ip
% mort
a-
lity
Sp
ecim
en
co
un
t
% Surv
ivo
r-sh
ip
% mo
rta-
lity
Sp
ecim
en
co
un
t
% Surv
ivo
r-sh
ip
% mo
rta-
lity
Sp
ecim
en
co
un
t
% Surv
ivo
r-sh
ip
% mo
rta-
lity
Sp
ecim
en
co
un
t
A0–2
mo.
98. 5
1. 5
195. 6
4. 4
598. 4
1. 6
196. 9
3. 1
4100
00
96. 3
3. 7
2B
2–6
mo.
98. 5
00
94. 7
0. 9
195. 3
3. 1
290. 1
6. 9
998. 7
1. 3
194. 4
1. 9
1C
6–12
mo.
78. 5
20. 0
13
80. 5
14. 2
16
75
20. 3
13
73. 9
16. 2
21
88. 8
10. 0
879. 6
14. 8
8D
1–2
yrs
.52. 3
26. 2
17
67. 2
13. 3
15
54. 7
20. 3
13
58. 5
15. 4
20
63. 8
25. 0
20
48. 1
31. 5
17
E2–3
yrs
.38. 5
13. 8
932. 7
34. 5
39
34. 4
20. 3
13
46. 2
12. 3
16
––
na
40. 7
7. 4
4F
3–4
yrs
.4. 7
33. 8
22
23. 0
9. 7
11
23. 5
10. 9
738. 5
7. 7
10
5. 0
58. 8
47*
20. 3
20. 4
11
G4–6
yrs
.3. 1
1. 5
115. 9
7. 1
812. 6
10. 9
710
28. 5
37
05
47. 3
13. 0
7H
6–8
yrs
.1. 6
1. 5
10. 9
15. 0
17
6. 3
6. 3
42. 3
7. 7
10
00
7. 3
00
I8–12
yrs
.0
1. 5
10
0. 9
10
6. 3
40
2. 3
30
00
7. 4
4n
65
113
64
130
80
54
’Ain
Gh
aza
les-S
ifiy
ael-
Hem
meh
MW
SA
ge
%S
urv
ivo
rsh
ip%
mo
rtalit
yS
pecim
en
co
un
t%
Su
rviv
ors
hip
%m
ort
alit
yS
pecim
en
co
un
t%
Su
rviv
ors
hip
%m
ort
alit
yS
pecim
en
co
un
t
A0–2
mo.
98. 5
1. 5
195. 6
4. 4
598. 4
1. 6
1B
2–6
mo.
98. 5
00
94. 7
0. 9
195. 3
3. 1
2C
6–12
mo.
78. 5
20. 0
13
80. 5
14. 2
16
75
20. 3
13
D1–2
yrs
.52. 3
26. 2
17
67. 2
13. 3
15
54. 7
20. 3
13
E2–3
yrs
.38. 5
13. 8
932. 7
34. 5
39
34. 4
20. 3
13
F3–4
yrs
.4. 7
33. 8
22
23. 0
9. 7
11
23. 5
10. 9
7G
4–6
yrs
.3. 1
1. 5
115. 9
7. 1
812. 6
10. 9
7H
6–8
yrs
.1. 6
1. 5
10. 9
15. 0
17
6. 3
6. 3
4I
8–12
yrs
.0
1. 5
10
0. 9
10
6. 3
4n
65
113
64
Basta
Ba’ja
’Ain
Jam
mam
MW
SA
ge
%S
urv
ivo
rsh
ip%
mo
rtalit
yS
pecim
en
co
un
t%
Su
rviv
ors
hip
%m
ort
alit
yS
pecim
en
co
un
t%
Su
rviv
ors
hip
%m
ort
alit
yS
pecim
en
co
un
t
A0–2
mo.
96. 9
3. 1
4100
00
96. 3
3. 7
2B
2–6
mo.
90. 1
6. 9
998. 7
1. 3
194. 4
1. 9
1C
6–12
mo.
73. 9
16. 2
21
88. 8
10. 0
879. 6
14. 8
8D
1–2
yrs
.58. 5
15. 4
20
63. 8
25. 0
20
48. 1
31. 5
17
E2–3
yrs
.46. 2
12. 3
16
––
na
40. 7
7. 4
4F
3–4
yrs
.38. 5
7. 7
10
5. 0
58. 8
47*
20. 3
20. 4
11
G4–6
yrs
.10
28. 5
37
05
47. 3
13. 0
7H
6–8
yrs
.2. 3
7. 7
10
00
7. 3
00
I8–12
yrs
.0
2. 3
30
00
7. 4
4n
130
80
54
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 245

(Payne 1975). Consequently, young animals are
probably under-represented in several of the Late
PPNB faunal assemblages analysed here, and some
survivorship curves generated from epiphyseal fusion
and tooth wear and eruption data are artificially
skewed towards older kill-off. Along these lines,
previous work claiming that the predominance of
young adult caprine kill-off at Late PPNB settlement
reflects strategies for meat production may well have
drawn upon evidence that, while thought to reflect
herder harvesting choices, may be at least partially an
artefact of excavation methodology.
Sites and data
The sites investigated in this paper include Late
PPNB settlements located in central and southern
Jordan (Fig. 1). In order to examine hypothesized
variation in caprine management systems within this
region over a relatively constrained cultural-chron-
ological period, ageing and measurement data from
published and new data sets are analysed.
‘Ain Ghazal
‘Ain Ghazal is a large, multi-period Pre-Pottery
Neolithic site which reached its greatest expanse
during the Late PPNB when it extended to 14 ha.
Located on the edge of a Mediterranean ecotone,
‘Ain Ghazal is nestled within the broad slopes of the
Wadi Zarqa. The architectural layout and style of
‘Ain Ghazal share some similarities with the more
southern Late PPNB settlements. However there are
important differences, in particular the presence of
apsidal buildings and circular ‘temples’, suggesting
that ‘Ain Ghazal may have existed slightly outside the
Late PPNB interaction sphere as manifested in
southern Jordan.
Animal exploitation at ‘Ain Ghazal focused pre-
dominantly on caprine herding, with goats only slightly
outnumbering sheep (1.2:1) (Wasse 2002). Morpho-
logical analyses of goat horn cores indicates that wild
bezoar were very occasionally exploited during the Late
PPNB (Wasse 2002). In contrast to most other
contemporary settlements in the region, suid (pig)
exploitation and the hunting of wild ungulates, such
as gazelle and equids, made a significant contribution to
the overall animal-based portion of the subsistence
economy at ‘Ain Ghazal (Table 2).
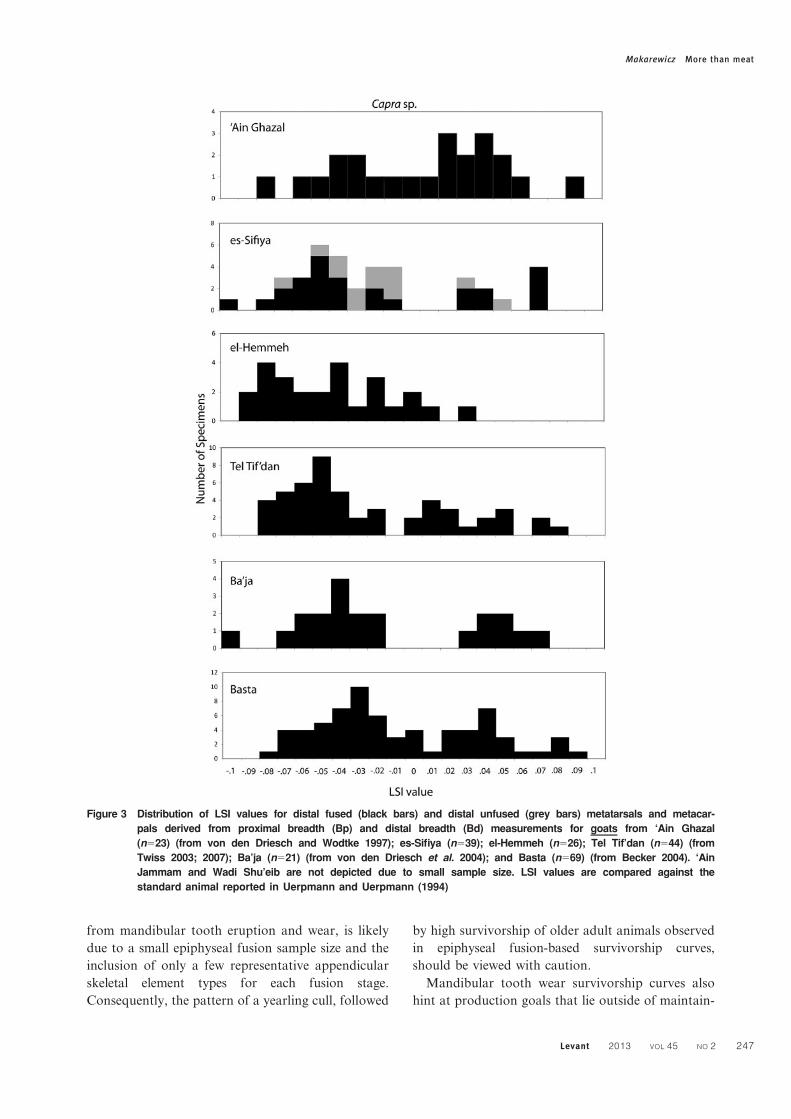
A histogram of LSI values calculated from
metapodials provides additional, if somewhat ambig-
uous information, on goat exploitation at ‘Ain
Ghazal (Fig. 3). A clear biomodal distribution is
visible, with the peak on the left representing small-
bodied domestic females. The peak on the right likely
represents larger-bodied adult males which dominate
the assemblage, although it is possible that wild
female goats are also represented here. The single
outlier value is likely from a large male wild goat. For
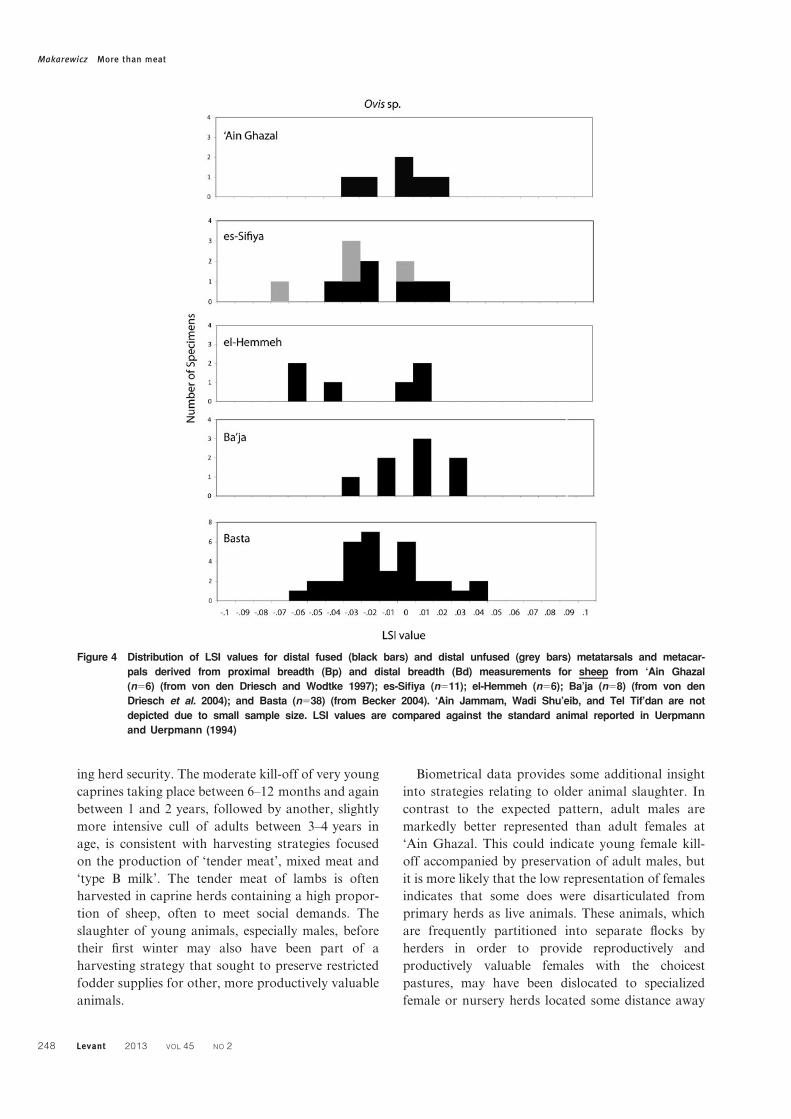
sheep, a biomodal distribution is visible, suggesting
the presence of male and females, but low sample
sizes caution against over-interpretation (Fig. 4).
Survivorship curves based on mandibular tooth
wear for a combined caprine category indicate that
animals were slaughtered primarily as juveniles and
young adults, with kill-off taking place between 6–
12 months (with only 52% of animals reaching two
years in age) and 3–4 years, with very few animals
surviving beyond this age (Fig. 5). Epiphyseal fusion-
based survivorship curves, constructed from a small
published epiphyseal fusion data set based on a few
skeletal element categories (i.e. scapula, distal tibia,
distal metapodials, distal radius) from ‘Ain Ghazal,
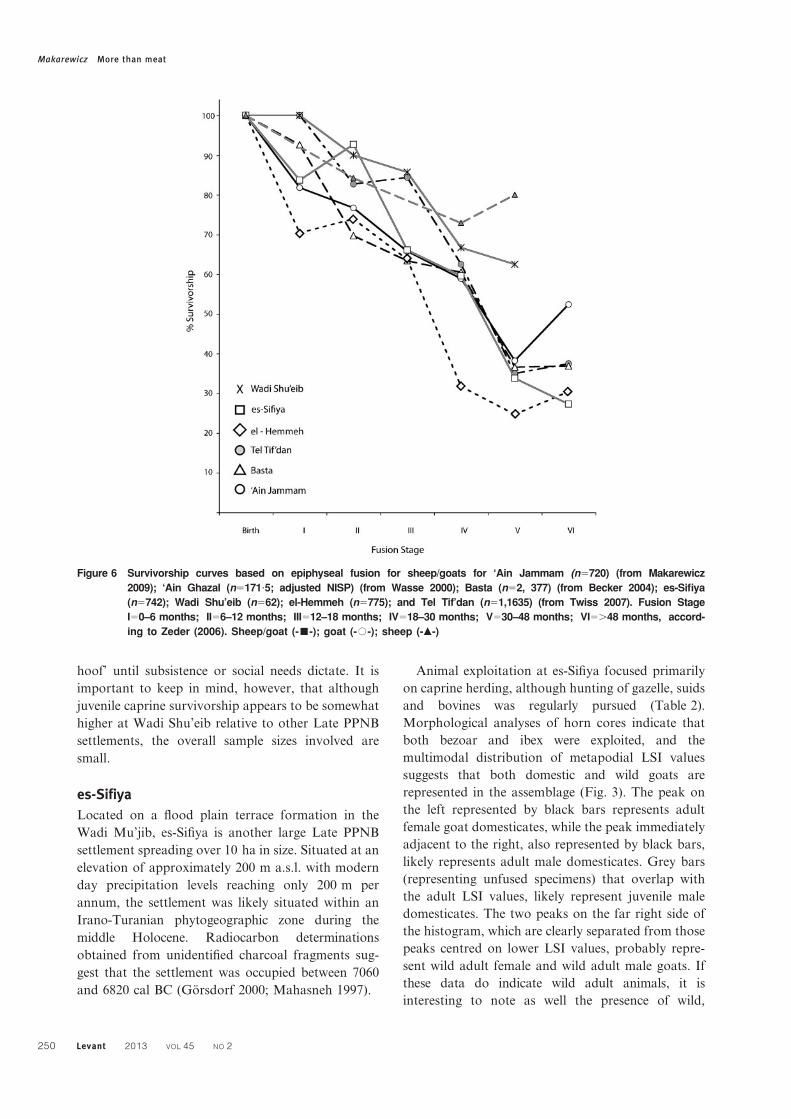
indicate a focus on yearling kill-off with 84% of
caprines surviving past 6–12 months (Fig. 6). There is
a very moderate increase in the intensity of slaughter
of slightly older animals, with c. 73% of animals
having survived Stage IV (18–30 months). Species-
specific epiphyseal fusion data was not available for
‘Ain Ghazal.
The balanced ratio of sheep to goats at ‘Ain
Ghazal is likely due in part to the location of the
settlement in a productive environment more hospi-
table to supporting sheep herds but may also indicate
that higher potential productivity, rather than risk
reduction, was pursued by herders there. The
disparity between survivorship curves calculated
from epiphyseal fusion data and those generated
Figure 2 Relative proportion of infant-, juvenile- and
adult- aged bone specimens in tripolar format.
Wadi Shu’eib (-6-); es-Sifiya (- %-); el-Hemmeh
(-e-); Basta (-n-); ‘Ain Jammam (-#-)
Makarewicz More than meat
246 Levant 2013 VOL 45 NO 2
from mandibular tooth eruption and wear, is likely
due to a small epiphyseal fusion sample size and the
inclusion of only a few representative appendicular
skeletal element types for each fusion stage.
Consequently, the pattern of a yearling cull, followed
by high survivorship of older adult animals observed
in epiphyseal fusion-based survivorship curves,
should be viewed with caution.
Mandibular tooth wear survivorship curves also
hint at production goals that lie outside of maintain-
Figure 3 Distribution of LSI values for distal fused (black bars) and distal unfused (grey bars) metatarsals and metacar-
pals derived from proximal breadth (Bp) and distal breadth (Bd) measurements for goats from ‘Ain Ghazal
(n523) (from von den Driesch and Wodtke 1997); es-Sifiya (n539); el-Hemmeh (n526); Tel Tif’dan (n544) (from
Twiss 2003; 2007); Ba’ja (n521) (from von den Driesch et al. 2004); and Basta (n569) (from Becker 2004). ‘Ain
Jammam and Wadi Shu’eib are not depicted due to small sample size. LSI values are compared against the
standard animal reported in Uerpmann and Uerpmann (1994)
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 247
ing herd security. The moderate kill-off of very young
caprines taking place between 6–12 months and again
between 1 and 2 years, followed by another, slightly
more intensive cull of adults between 3–4 years in
age, is consistent with harvesting strategies focused
on the production of ‘tender meat’, mixed meat and
‘type B milk’. The tender meat of lambs is often
harvested in caprine herds containing a high propor-
tion of sheep, often to meet social demands. The
slaughter of young animals, especially males, before
their first winter may also have been part of a
harvesting strategy that sought to preserve restricted
fodder supplies for other, more productively valuable
animals.
Biometrical data provides some additional insight
into strategies relating to older animal slaughter. In
contrast to the expected pattern, adult males are
markedly better represented than adult females at
‘Ain Ghazal. This could indicate young female kill-
off accompanied by preservation of adult males, but
it is more likely that the low representation of females
indicates that some does were disarticulated from
primary herds as live animals. These animals, which
are frequently partitioned into separate flocks by
herders in order to provide reproductively and
productively valuable females with the choicest
pastures, may have been dislocated to specialized
female or nursery herds located some distance away
Figure 4 Distribution of LSI values for distal fused (black bars) and distal unfused (grey bars) metatarsals and metacar-
pals derived from proximal breadth (Bp) and distal breadth (Bd) measurements for sheep from ‘Ain Ghazal
(n56) (from von den Driesch and Wodtke 1997); es-Sifiya (n511); el-Hemmeh (n56); Ba’ja (n58) (from von den
Driesch et al. 2004); and Basta (n538) (from Becker 2004). ‘Ain Jammam, Wadi Shu’eib, and Tel Tif’dan are not
depicted due to small sample size. LSI values are compared against the standard animal reported in Uerpmann
and Uerpmann (1994)
Makarewicz More than meat
248 Levant 2013 VOL 45 NO 2
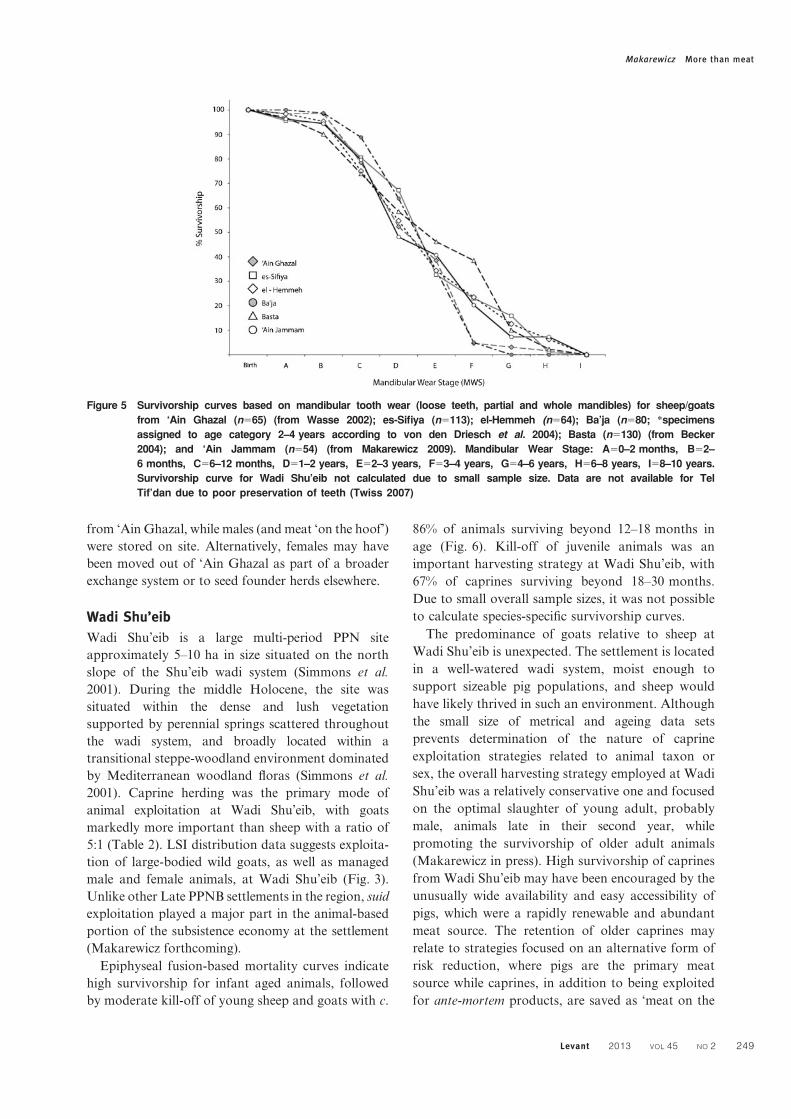
from ‘Ain Ghazal, while males (and meat ‘on the hoof’)
were stored on site. Alternatively, females may have
been moved out of ‘Ain Ghazal as part of a broader
exchange system or to seed founder herds elsewhere.
Wadi Shu’eib
Wadi Shu’eib is a large multi-period PPN site
approximately 5–10 ha in size situated on the north
slope of the Shu’eib wadi system (Simmons et al.
2001). During the middle Holocene, the site was
situated within the dense and lush vegetation
supported by perennial springs scattered throughout
the wadi system, and broadly located within a
transitional steppe-woodland environment dominated
by Mediterranean woodland floras (Simmons et al.
2001). Caprine herding was the primary mode of
animal exploitation at Wadi Shu’eib, with goats
markedly more important than sheep with a ratio of
5:1 (Table 2). LSI distribution data suggests exploita-
tion of large-bodied wild goats, as well as managed
male and female animals, at Wadi Shu’eib (Fig. 3).
Unlike other Late PPNB settlements in the region, suid
exploitation played a major part in the animal-based
portion of the subsistence economy at the settlement
(Makarewicz forthcoming).
Epiphyseal fusion-based mortality curves indicate
high survivorship for infant aged animals, followed
by moderate kill-off of young sheep and goats with c.
86% of animals surviving beyond 12–18 months in
age (Fig. 6). Kill-off of juvenile animals was an
important harvesting strategy at Wadi Shu’eib, with
67% of caprines surviving beyond 18–30 months.
Due to small overall sample sizes, it was not possible
to calculate species-specific survivorship curves.
The predominance of goats relative to sheep at
Wadi Shu’eib is unexpected. The settlement is located
in a well-watered wadi system, moist enough to
support sizeable pig populations, and sheep would
have likely thrived in such an environment. Although
the small size of metrical and ageing data sets
prevents determination of the nature of caprine
exploitation strategies related to animal taxon or
sex, the overall harvesting strategy employed at Wadi
Shu’eib was a relatively conservative one and focused
on the optimal slaughter of young adult, probably
male, animals late in their second year, while
promoting the survivorship of older adult animals
(Makarewicz in press). High survivorship of caprines
from Wadi Shu’eib may have been encouraged by the
unusually wide availability and easy accessibility of
pigs, which were a rapidly renewable and abundant
meat source. The retention of older caprines may
relate to strategies focused on an alternative form of
risk reduction, where pigs are the primary meat
source while caprines, in addition to being exploited
for ante-mortem products, are saved as ‘meat on the
Figure 5 Survivorship curves based on mandibular tooth wear (loose teeth, partial and whole mandibles) for sheep/goats
from ‘Ain Ghazal (n565) (from Wasse 2002); es-Sifiya (n5113); el-Hemmeh (n564); Ba’ja (n580; *specimens
assigned to age category 2–4 years according to von den Driesch et al. 2004); Basta (n5130) (from Becker
2004); and ‘Ain Jammam (n554) (from Makarewicz 2009). Mandibular Wear Stage: A50–2 months, B52–
6 months, C56–12 months, D51–2 years, E52–3 years, F53–4 years, G54–6 years, H56–8 years, I58–10 years.
Survivorship curve for Wadi Shu’eib not calculated due to small sample size. Data are not available for Tel
Tif’dan due to poor preservation of teeth (Twiss 2007)
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 249
hoof’ until subsistence or social needs dictate. It is
important to keep in mind, however, that although
juvenile caprine survivorship appears to be somewhat
higher at Wadi Shu’eib relative to other Late PPNB
settlements, the overall sample sizes involved are
small.
es-Sifiya
Located on a flood plain terrace formation in the
Wadi Mu’jib, es-Sifiya is another large Late PPNB
settlement spreading over 10 ha in size. Situated at an
elevation of approximately 200 m a.s.l. with modern
day precipitation levels reaching only 200 m per
annum, the settlement was likely situated within an
Irano-Turanian phytogeographic zone during the
middle Holocene. Radiocarbon determinations
obtained from unidentified charcoal fragments sug-
gest that the settlement was occupied between 7060
and 6820 cal BC (Gorsdorf 2000; Mahasneh 1997).
Animal exploitation at es-Sifiya focused primarily
on caprine herding, although hunting of gazelle, suids
and bovines was regularly pursued (Table 2).
Morphological analyses of horn cores indicate that
both bezoar and ibex were exploited, and the
multimodal distribution of metapodial LSI values
suggests that both domestic and wild goats are
represented in the assemblage (Fig. 3). The peak on
the left represented by black bars represents adult
female goat domesticates, while the peak immediately
adjacent to the right, also represented by black bars,
likely represents adult male domesticates. Grey bars
(representing unfused specimens) that overlap with
the adult LSI values, likely represent juvenile male
domesticates. The two peaks on the far right side of
the histogram, which are clearly separated from those
peaks centred on lower LSI values, probably repre-
sent wild adult female and wild adult male goats. If
these data do indicate wild adult animals, it is
interesting to note as well the presence of wild,
Figure 6 Survivorship curves based on epiphyseal fusion for sheep/goats for ‘Ain Jammam (n5720) (from Makarewicz
2009); ‘Ain Ghazal (n5171?5; adjusted NISP) (from Wasse 2000); Basta (n52, 377) (from Becker 2004); es-Sifiya
(n5742); Wadi Shu’eib (n562); el-Hemmeh (n5775); and Tel Tif’dan (n51,1635) (from Twiss 2007). Fusion Stage
I50–6 months; II56–12 months; III512–18 months; IV518–30 months; V530–48 months; VI5.48 months, accord-
ing to Zeder (2006). Sheep/goat (-&-); goat (-#-); sheep (-m-)
Makarewicz More than meat
250 Levant 2013 VOL 45 NO 2
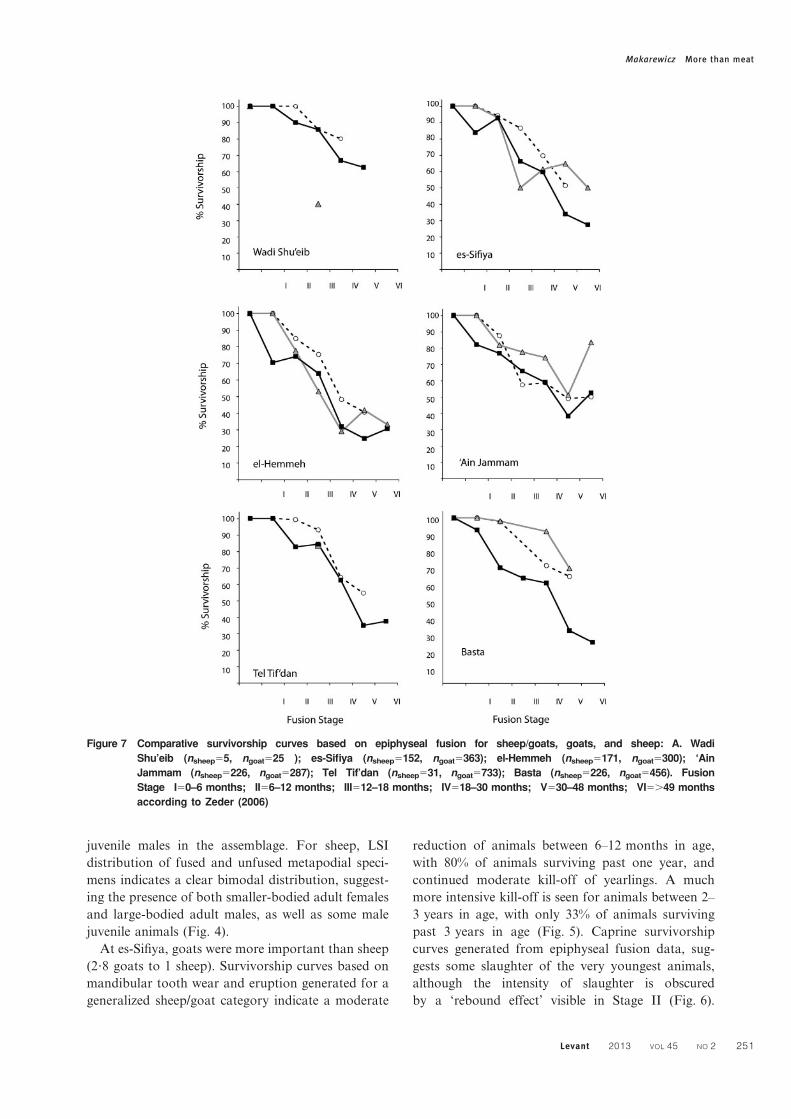
juvenile males in the assemblage. For sheep, LSI
distribution of fused and unfused metapodial speci-
mens indicates a clear bimodal distribution, suggest-
ing the presence of both smaller-bodied adult females
and large-bodied adult males, as well as some male
juvenile animals (Fig. 4).
At es-Sifiya, goats were more important than sheep
(2?8 goats to 1 sheep). Survivorship curves based on
mandibular tooth wear and eruption generated for a
generalized sheep/goat category indicate a moderate
reduction of animals between 6–12 months in age,
with 80% of animals surviving past one year, and
continued moderate kill-off of yearlings. A much
more intensive kill-off is seen for animals between 2–
3 years in age, with only 33% of animals surviving
past 3 years in age (Fig. 5). Caprine survivorship
curves generated from epiphyseal fusion data, sug-
gests some slaughter of the very youngest animals,
although the intensity of slaughter is obscured
by a ‘rebound effect’ visible in Stage II (Fig. 6).
Figure 7 Comparative survivorship curves based on epiphyseal fusion for sheep/goats, goats, and sheep: A. Wadi
Shu’eib (nsheep55, ngoat525 ); es-Sifiya (nsheep5152, ngoat5363); el-Hemmeh (nsheep5171, ngoat5300); ‘Ain
Jammam (nsheep5226, ngoat5287); Tel Tif’dan (nsheep531, ngoat5733); Basta (nsheep5226, ngoat5456). Fusion
Stage I50–6 months; II56–12 months; III512–18 months; IV518–30 months; V530–48 months; VI5.49 months
according to Zeder (2006)
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 251

Survivorship declines dramatically in fusion stage III,
with 66% of caprines surviving beyond 12–18 months
in age, although a ‘rebound effect’ obscures the pattern
somewhat. Survivorship drops markedly in older
animals, with only c. 34% of animals surviving past
30–48 months. Species-specific demographic profiles
indicate generally higher survivorship of goats over
sheep (Fig. 7). Young goats are largely retained with
87% surviving beyond 12–18 months, in contrast to
sheep, which exhibit markedly lower survivorship
beyond this age. Goats are steadily harvested as
juveniles and adult animals.
The higher abundance of goats relative to sheep
at es-Sifiya likely reflects, in part, an environmen-
tally based herding decision to maintain a higher
proportion of physiologically more robust goats in
flocks herded in a drier Irano-Turanian phytogeo-
graphic zone. The general focus on the moderate
slaughter of lambs, kids, and yearlings, and the
emphasis on adult animals aged 2–3 years, fits with
the expectations associated with tender meat and
‘type B milk’ production. There are clear differences
in the harvesting strategies applied to sheep and
goats at es-Sifiya. The higher survivorship of goats
over sheep, especially for young juvenile animals
(Fig. 7), suggests that yearling sheep were slaugh-
tered for their tender meat, while goats were
harvested for their meat as juveniles and young
adults, when it is was optimal to do so. Sex-specific
harvesting strategies enacted on goats are also
consistent with models for herd production and
herd security. Metrical data indicating low abun-
dance of large fused specimens (representing domes-
tic adult males), accompanied by large unfused
specimens (representing domestic juvenile males)
and abundant representation of small fused speci-
mens (adult females), provide additional, clear
evidence for the focused slaughter of goats during
their juvenile years while promoting adult female
survivorship (Fig. 3). The sheep metrical data,
hampered by small sample size, are more difficult
to interpret (Fig. 4). The presence of both female
and male adults, as well as young male sheep, hints
at some targeting harvesting of juvenile sheep as
strongly observed in goats.
el-Hemmeh
El-Hemmeh is a small (c. 2 ha) Late PPNB settlement
located on a dissected alluvial fan overlooking the
floodplain of the Wadi Hasa. The site is one of
several Late PPNB sites situated within the Wadi
Hasa, including Khirbet Hamman and Hamarash
located c. 10 km and 20 km downstream, respectively
(Petersen 2004). El-Hemmeh offers an opportunity to
examine caprine management strategies in use at a
smaller settlement that sits within the same broader
catchment area as larger sites. A single radiocarbon
determination from unidentified wood charcoal dates
occupation at el-Hemmeh to c. 7600 cal BC
(Makarewicz et al. 2006; Makarewicz and Austin
2006).
Animal exploitation at el-Hemmeh focused on
sheep and goat herding, with some contribution from
taurines and hunted gazelle to the overall subsistence
economy (Table 2; Makarewicz et al. 2006). Sheep
are somewhat less abundant than goats with a ratio
of 1:1?7. Log-transformed metrical data for metapo-
dials indicate a roughly bimodal distribution of values,
with the left peak clearly representing small-bodied
adult females, while the right peak, although spread
out, likely representing larger-bodied adult males
(Fig. 3). It is notable that, when compared to similar
metrical data from other Late PPNB settlements in the
region, the very largest-bodied goats are generally
missing from el-Hemmeh, suggesting that wild goat
bone specimens are minimally represented, if at all, in
the overall assemblage.
Survivorship curves constructed from mandibular
tooth wear data indicate marked kill-off of caprines
aged 6–12 months, with 75% surviving beyond one
year in age. This is followed by moderate but steady
slaughter of young animals aged 12–36 months, with
only 34% of animals surviving 3 years in age (Fig. 5).
Survivorship curves generated from epiphyseal fusion
data broadly agree with tooth wear-based curves.
Demographic profiles generated from fusion data
also indicate that a marked kill-off of infant-aged
animals was relatively high, with only c. 70% of
caprines surviving beyond 0–6 months in age (Fig. 6).
Young juveniles were slaughtered at relatively low
intensities with survivorship of c. 64% beyond 12–
18 months. The next episode of intensive slaughter
was concentrated on adult animals in their second
year, with only 32% of caprines surviving beyond 18–
30 months in age.
Species-specific survivorship curves indicate that
sheep from el-Hemmeh were slaughtered at higher
intensities relative to goats (Fig. 7). There is a very
slight difference in mortality between the youngest
kids and lambs but a more pronounced divergence in
kill-off of yearlings, with c. 75% goats surviving
beyond 12–18 months compared to 53% of sheep.
Survivorship of juvenile and young adult sheep
remains depressed, with 48% of goats surviving
beyond 18–30 months compared to 29% for sheep,
surviving beyond 18–30 months compared to 48% for
Makarewicz More than meat
252 Levant 2013 VOL 45 NO 2
goats, although a later ‘rebound’ in older sheep may
indicate a less intense kill-off of this age cohort.
The slightly higher abundance of goats relative to
sheep at el-Hemmeh likely reflects herder decisions
related to production rather than environmental
constraints, as the wetland conditions at the bottom
of the Wadi Hasa would have supported sheep
(Contreras, personal communication). However, if
flocks were herded along a transhumance mobility
pattern out of the wadi bottom to the top of the
Kerak plateau, it may have been more beneficial to
populate herds with a slightly higher proportion of
more hardy goats rather than to focus heavily on
more productive sheep. Interestingly, the youngest
kill-off of caprines is visible at el-Hemmeh compared
to other Late PPNB assemblages examined here. The
strongly defined focus on the kill-off of lambs and
kids, especially in epiphyseal fusion-based survivor-
ship curves, suggests non-intensive milk production
was regularly pursued at el-Hemmeh. The moderate
kill of young yearlings is consistent with models for
mixed meat and milk production, while the more
intensive harvesting of young adult two year olds is
consistent with optimal meat production and herd
security.
The elevated survivorship of goats over sheep as
yearlings, juveniles, and young adults especially
suggests that goats served a more important role in
maintaining everyday subsistence, while sheep,
slaughtered using non-optimizing harvesting strate-
gies for meat production, may have been killed to
meet social demands. LSI values generated for
metapodials provide some additional information
on how goats were managed at el-Hemmeh (Fig. 3).
There appears to be a preference for young male kill-
off while adult female survivorship was promoted,
although the pattern is not strongly defined and the
absence of measurable, unfused metapodials in the
distal breadth dimension poses interpretative diffi-
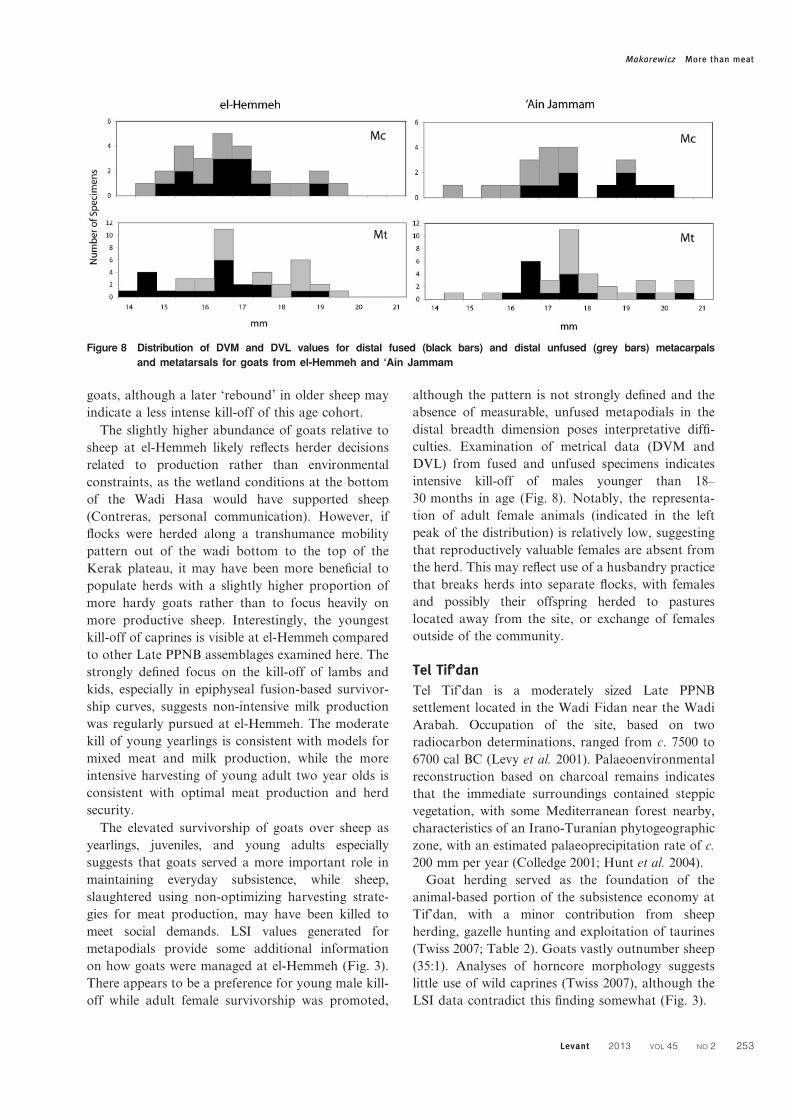
culties. Examination of metrical data (DVM and
DVL) from fused and unfused specimens indicates
intensive kill-off of males younger than 18–
30 months in age (Fig. 8). Notably, the representa-
tion of adult female animals (indicated in the left
peak of the distribution) is relatively low, suggesting
that reproductively valuable females are absent from
the herd. This may reflect use of a husbandry practice
that breaks herds into separate flocks, with females
and possibly their offspring herded to pastures
located away from the site, or exchange of females
outside of the community.
Tel Tif’dan
Tel Tif’dan is a moderately sized Late PPNB
settlement located in the Wadi Fidan near the Wadi
Arabah. Occupation of the site, based on two
radiocarbon determinations, ranged from c. 7500 to
6700 cal BC (Levy et al. 2001). Palaeoenvironmental
reconstruction based on charcoal remains indicates
that the immediate surroundings contained steppic
vegetation, with some Mediterranean forest nearby,
characteristics of an Irano-Turanian phytogeographic
zone, with an estimated palaeoprecipitation rate of c.
200 mm per year (Colledge 2001; Hunt et al. 2004).
Goat herding served as the foundation of the
animal-based portion of the subsistence economy at
Tif’dan, with a minor contribution from sheep
herding, gazelle hunting and exploitation of taurines
(Twiss 2007; Table 2). Goats vastly outnumber sheep
(35:1). Analyses of horncore morphology suggests
little use of wild caprines (Twiss 2007), although the
LSI data contradict this finding somewhat (Fig. 3).
Figure 8 Distribution of DVM and DVL values for distal fused (black bars) and distal unfused (grey bars) metacarpals
and metatarsals for goats from el-Hemmeh and ‘Ain Jammam
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 253

LSI data for Tel Tif’dan goat metapodials indicates
a multi-modal distribution, suggesting that both
domesticate and wild animals were exploited
(Fig. 3). The peak on the left represents small-bodied,
adult female domesticates which dominate the
assemblage. It is likely that the next peak, represented
by LSI values ranging from 20?02 to z0?03, largely
represents larger-bodied male domesticates, although
it is possible that wild female goats are also
represented. The two peaks to the far right likely
represent wild female and male goats.
Tooth wear data were not available due to
extremely poor preservation of teeth (Twiss 2007).
Survivorship curves generated according to epiphy-
seal fusion data indicate use of a conservative
harvesting strategy at Tel Tif’dan, compared to other
Late PPNB sites in the region, with a moderate
retention of lambs and kids (Fig. 6). Kill-off was
concentrated on young animals, with c. 83% of
animals surviving beyond 6–12 months. Slightly
older animals were retained, with no decrease in
survivorship beyond 12–18 months. Kill-off of juve-
niles and younger adults was relatively intense, with
only 63% survivorship of caprines beyond 18–
30 months and then 35% surviving past four years.
Due to the relatively low relative abundance of sheep
present at Tel Tif’dan, it is difficult to elucidate
species-specific harvesting strategies in use at the site.
Goat survivorship for animals between 12–18 months
appears to have been somewhat higher than sheep of
the same age (Fig. 7).
The almost exclusive focus on goats over sheep at
Tel Tif’dan was probably heavily influenced by local
environmental conditions. Although the settlement
was situated within an Irano-Turanian zone during
the Late PPNB, palaeoenvironmental reconstructions
suggest that local precipitation levels were quite low
and not favourable to the keeping of large sheep
flocks. Previous analyses have interpreted caprine
exploitation at Tel Tif’dan as heavily focused on herd
security and the production of meat, while recogniz-
ing that other ante-mortem products, such as milk
and hair, may have been exploited (Twiss 2007).
While the intensive slaughter of adult caprines is
almost certainly indicative of a harvesting strategy
designed for meat production (although perhaps not
optimally so, perhaps indicating a more conservative
form of harvesting that placed greater emphasis on
risk reduction), the moderate slaughter of young
juveniles (mostly goats aged 6–12 months) combined
with the intensive harvesting of adults, but retention
of females well into adulthood (as indicated in body
size distribution data), is consistent with expectations
of models for meat and some milk exploitation. In
addition, by eliminating younger animals, herders at
Tif’dan could concentrate limited winter graze and
fodder supplies on core herd animals and, conse-
quently, increase overall herd stability through
maintenance of animal body condition. In addition,
the absence of a harvest of animals in fusion stage III
(12–18 months) at Tel Tif’dan is unusual and may
indicate spring-summer movement of this age cohort
away from the settlement to summer pasture
grounds. The lower survivorship of sheep of the
same age may indicate that the meat from yearling
sheep compensated for the absence of that obtained
from goats, although additional lines of evidence,
such as skeletal element distribution data, would be
needed to further investigate this possibility.
Ba’ja
The site of Ba’ja is located on an intramontane
terrace deep within the sandstone formation of the
low mountains surrounding the Petra region. The site
is uniquely placed, hidden on the landscape and
reachable only through a narrow siq, which prevents
easy access to the settlement (Gebel and Beinert
1997). Restricted in size by its topographical
surround, Ba’ja is quite small (c. 1?5 ha) relative to
other Late PPNB settlements in the region but may
have served as a centre for specialized production and
distribution of sandstone rings (Gebel 2010; Gebel
and Beinert 1997). Based on determinations derived
from three juniper charcoal fragments, the occupa-
tional sequence at Ba’ja dates from 7500 cal BC to
6900 cal BC (Gebel 2013).
Previous work on the faunal remains from Ba’ja is
limited to the remains recovered from the 1997 season
(von den Driesch et al. 2004). Sheep and goats are the
most abundant taxa represented at Ba’ja, with goats
outnumbering sheep at a ratio of 3:1 (Table 2).
LSI transformed metapodial measurement data for
goats from Ba’ja are somewhat ambiguous (Fig. 3).
A histogram generated from LSI data indicates a
bimodal distribution, with the peak on the left likely
to represent small-bodied female domesticates, which
dominate the assemblage, while the peak on the right
represents either larger-bodied male domesticates or
wild female animals. No pattern is discernible in the
sheep LSI data (Fig. 4).
Mortality curves based on tooth wear for a
combined caprine category indicate a moderate kill-
off of animals aged 1 to 2 years, followed by a much
more intensive cull of animals slaughtered between
the ages of 3 and 4 years, with few animals surviving
Makarewicz More than meat
254 Levant 2013 VOL 45 NO 2

beyond this age (Fig. 5). Caprine epiphyseal fusion
data is not available for Ba’ja.
In their analysis, von den Driesch et al. (2004)
argued that the combination of high adult kill-off and
high proportion of male pelvic remains indicated that
sheep and goats were managed primarily for their
meat, although they do not rule out the possibility
that caprines were also being exploited for ante-
mortem products. However, a reconsideration of the
survivorship data reveals a more complex picture.
The slaughter of young caprines aged 1–2 years at
Ba’ja is consistent with a harvesting strategy focused
on the slaughter of animals for their meat. Size
distribution data indicating a slight skew towards
small adult female goats, a pattern consistent with the
moderate focus on the slaughter of juvenile male
goats, also suggests meat production and herd
security. For sheep, the absence of any discernable
pattern for female- or male-based kill-off may either
reflect lower expression of sexual dimorphism in
sheep, a harvesting strategy focused on the exclusive
kill-off of either female or male animals, or a strategy
that evenly targets both males and females (Fig. 7).
The harvesting strategy enacted on older animals
at B’aja is more difficult to interpret, in part because
tooth wear data for some older animals was conflated
into a broader age category of 2–4 years, rather than
2–3 years and 3–4 years as broken down in Payne’s
(1973) schema. However, it is clear that harvesting of
adults took place at markedly higher intensities
relative to juveniles. This unusual pattern of high
adult kill-off may indicate that Ba’ja was being
externally provisioned, either with joints of meat or
live animals.
Although the kill-off pattern seen at Ba’ja does not
fit exactly with Stein’s (1987) model representing
‘producer’ and ‘consumer’ sites, which holds that
assemblages formed by consumers should contain
young, predominantly male animals aged between 1–
3 years while animals older than 3 years are absent
altogether, Stein’s model was developed to under-
stand how herds may have been managed to
provision villages in ancient market economies.
During the Neolithic, the criteria for entering animals
into exchange may have been different. In keeping
with an overall strategy of risk reduction, the
expectation is that herders from producing sites
would off-load their older, surplus animals only
when it became clear that these animals were no
longer needed for subsistence or herd regeneration. It
may be that the predominance of older adults at Ba’ja
reflects the use of animals acquired through exchange
from external sources, perhaps necessitated by the
location of Ba’ja in a locale unsuitable for the keeping
of large caprine herds. This may also explain the
absence of foddered animals at Ba’ja — animals no
longer needed for herd maintenance are often denied
fodder in order to concentrate this limited resource
into more productive ones.
The absence of foddering in Ba’ja goats is
suggested by the restricted range and low variability
in collagen carbon isotope values (d13Cmean5218?6¡
0?4ø; d13Crange5219?2ø to 217?9ø) which are
similar to that observed for co-occurring wild, territor-
Figure 9 Carbon (d13C) and nitrogen (d15N) isotopic values for goat collagens from Ba’ja (–&–) and Basta (– q –), and
gazelle collagens from Basta (– –)
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 255

ial gazelle from nearby Basta (d13Cmean5216?6¡
1?0ø; d13Crange5215?0ø to 218?3ø. Collagen was
not well preserved in Ba’ja gazelles). This, combined
with the relatively low carbon values exhibited by Ba’ja
goats, indicating a minimal input of C4 graze in the diet
in contrast to gazelle (which preferentially consume
graze) and some goats from Basta, which are mixed
feeders seasonally provisioned with C4 annuals or
water-stressed C3 graze or browse collected during the
dry season, strongly suggests that Ba’ja goat diet
reflects primarily natural inputs (Fig. 9; Makarewicz in
press; Makarewicz and Tuross 2012). Differences
between goats and gazelle nitrogen isotope values likely
reflect isotopic variation in the underlying floral biome
and physiological differences between the two taxa.
Goat herds likely freely grazed and browsed the
mountainous landscapes surrounding Ba’ja. Instead
of employing intensive fodder provisioning strategies
similar to those enacted seasonally on some goats at
Basta, it is likely herders at Ba’ja relied solely on
open-pasture resources for their animals year-round
and encouraged goat herds to freely graze and browse
in the mountainous pasture in the vicinity of the site.
Given the logistical difficulties of moving herds and
fodder into and out of the settlement, and the general
lack of space for keeping large numbers of animals
penned on-site, it is not entirely surprising that goats
at Ba’ja were not foddered. Moreover, these isotopic
data also suggest that herders did not maintain off-
site satellite herding camps stocked with fodder and
instead employed an extensive pasturage system year-
round. The absence of foddering seen in goats
exploited at Ba’ja (regardless if these animals were
herded by the inhabitants of Ba’ja or imported from
nearby communities) suggests that these animals were
at risk on a seasonal basis, and these animals did not
feature in broader management strategies concerned
with enhancing animal body condition.
Basta
The site of Basta is a large settlement located in the
rolling steppic hills that join the Petra mountains with
the deserts to the east. Extending over 10 ha, like
other Late PPNB sites, Basta is known for its densely
packed architecture and thickly plastered walls and
floors. Based on radiocarbon determinations
obtained from unidentified charcoal fragments, the
occupational sequence at Basta spans from 7550 cal
BC to 7050 cal BC (Gebel 2013; Nissen et al. 1987).
While animal exploitation at Basta focused on
caprine herding, gazelle and equid hunting together
made a significant contribution to the overall
subsistence economy (Becker 1991; Table 2). Goats
are more abundantly represented than sheep at a
ratio of 2?3:1 (Table 2). Previous analyses suggest
that domesticated sheep and goats at Basta were
exploited primarily for their meat and possibly for
some dairy (Becker 2004). Wild goats bezoar (C.
aegagrus) were also hunted (Becker 2004).
Comparison of LSI values for fused metacarpals
and metatarsals, which fuse between 18 and
30 months, provide insight into herd demography
after the age at which animals have reached their
optimal body weight while balancing for energetic
intake. The trimodal distribution observed in mea-
surement data for goats suggests a diverse array of
exploitation strategies (Fig. 3). The low peak on the
right likely represents hunted, large-bodied wild
animals, while the other two peaks probably repre-
sent smaller-bodied male and female domesticates.
The middle peak, which represents domesticated
males allowed to survive beyond the age of metapo-
dial fusion, clearly indicates strong young male kill-
off while a high proportion of females were preserved
well into adulthood. For sheep, a bimodal distribu-
tion is visible, suggesting that both female and male
animals survived well into adulthood (Fig. 4).
Survivorship curves generated from mandibular
tooth wear for a combined sheep/goat category
indicate a relatively steady reduction of juvenile and
adult animals with concentrated kill-off of animals
between 6 and 12 months, followed by a steady
reduction of animals, and followed again by a more
intense kill-off of older animals between 4 to 6 years
in age (Fig. 5). Survivorship curves calculated from
epiphyseal fusion data suggest the moderate-low
intensity slaughter of young animals up to 6 months
in age, the focused kill-off of yearlings with only 70%
of animals surviving 6–12 months in age, accompa-
nied by a pronounced harvest of adult caprines aged
30–48 months with only 37% surviving to this age
(Fig. 6).
Bearing in mind that sheep bone specimens are less
well represented than those belonging to goats,
species-specific survivorship curves demonstrate
higher survivorship of sheep over goats, indicating
that each species was managed using different
harvesting strategies at Basta (Fig. 7). In particular,
juvenile goats were slaughtered at higher intensities
relative to sheep, with 71% of goats surviving beyond
18–30 months compared to 92% for sheep. Intensive
harvesting of sheep was delayed until animals reached
adulthood, with 70% of animals surviving beyond
30–48 months.
Animal husbandry strategies employed at Basta
extended to include manipulation of the animal diet.
Makarewicz More than meat
256 Levant 2013 VOL 45 NO 2

Wide carbon and nitrogen isotopic variation and high
d13C values visible in goats from Basta relative to co-
occurring gazelle suggests some goats, but not
necessarily all, were supplied with seasonally avail-
able C4 fodder or water-stressed C3 forage collected
during the summer months (Fig. 9; Makarewicz and
Tuross 2009; 2012). Through foddering, herders at
Basta could overcome periods of seasonally related
shortages in quality graze that would otherwise
negatively impact animal body condition, shorten
lactation duration, and increase mortality (Makarewicz
and Tuross 2006; 2009; 2012). Additional isotopic
analyses also indicate that the goats from Basta were
regularly moved to different pastures some distance, to
an area characterized by a wetter environment
(Makarewicz and Tuross 2012). The pattern of low
d18O and d15N values relative to gazelle, which
presumably were hunted within the vicinity of the
settlement, indicates that goats were ingesting forage
from outside the immediately available graze surround-
ing Basta (Makarewicz and Tuross 2012, Fig. 5).
The survivorship and body size distribution data
suggest that a complex harvesting system was in place
at Basta, one that took into account graze availability
and factors relating to risk reduction and perhaps
wealth accumulation. The intensive kill-off of year-
lings suggests that young caprines were slaughtered
before the winter months in order to conserve limited
fodder resources. The stable isotopic data, which
indicates that fodder was not necessarily applied to
all animals, further support this interpretation. The
intensive slaughter of older adult caprines may reflect
the slaughter of female animals whose progeny
production has dropped off.
The sharp divergence in harvesting strategies
applied to sheep and goats seen at Basta suggests
herders defined different subsistence and production
goals for each animal. For goats, the pronounced
harvest of juvenile and young adult goats fits well
with models for optimal meat off-take. The low
proportion of fused metapodial specimens represent-
ing male domesticates visible in LSI distribution
supports this finding and suggests that male goats
were slaughtered primarily during their first and
second years. However, for sheep, the retention of
juvenile and young adult animals, followed by an
intensive harvesting of adults, is more difficult to
explain. Although the ‘milk B’ model calls for the
slaughter of adult females when milk production
drops off, the low kill-off of infant-aged caprines
suggests that milk production was not heavily
pursued at Basta, or at least not at levels readily
detectable in survivorship curves. In addition, biome-
trical data indicate that both male and female animals
were surviving into adulthood relatively equally. As
an alternative explanation, sheep may have served as
a secondary meat source, one that was accessed only
if the meat supplied by goats was inadequate, and
served to reduce subsistence risks. In addition, the
longevity of sheep may indicate these animals
functioned as a form of storable live animal wealth
and were slaughtered only when social or political
needs dictated.
‘Ain Jammam
The settlement of ‘Ain Jamman is located near Ras
en-Naqb on a escarpment (c. 1600 m a.s.l.) over-
looking the low-lying (c. 800 m a.s.l.) Wadi Hisma
(Fino 2004). The chronological placement of ‘Ain
Jamman within the Late PPNB is difficult due to the
extremely large standard deviations seen in the two
radiocarbon dates obtained from the site (Rollefson
2005). Animal use at ‘Ain Jammam focused primarily
on sheep and goat herding, with goats slightly more
important than sheep (1?3 goats:1 sheep), but gazelle
hunting was also an important feature of the overall
animal exploitation system and provided herders with
a stable fall-back resource as they experimented with
new forms of caprine husbandry (Table 2;
Makarewicz 2009). Hunting of small game was also
pursued at ‘Ain Jammam, and aurochs and possibly
managed cattle were also exploited. Parrot fish from
the Red Sea were consumed at ‘Ain Jammam,
suggesting that smoked or salted fish were procured
via exchange or direct collection (Makarewicz 2009).
A single horn core attributed to C. ibex suggests at
least occasional exploitation of ibex at ‘Ain Jammam.
A trimodal distribution of LSI values generated from
the appendicular skeleton suggests wild goats com-
prise a very small proportion of the overall assem-
blage (Fig. 3; Makarewicz 2009). Metrical data (DVL
and DVM measurements) from metacarpals (less so
in the metatarsals) indicate a higher proportion of
smaller-bodied adult females (.24 months) relative
to larger-bodied adult males and a high proportion of
young (,24 months) males in the assemblage
(Fig. 8).
Survivorship curves calculated from tooth wear
data indicate intensive kill-off of caprines aged 6–
12 months, with only 48% of animals reaching 2
years in age (Fig. 5). Animal slaughter intensifies
again for animals 3–4 years in age, with only 20% of
herd animals reaching 4 years in age. Survivorship
curves generated from epiphyseal fusion data indicate
moderate kill-off of the youngest caprines, with 82%
surviving beyond 6 months (Fig. 6). Slaughter
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 257

intensity was relatively low for yearlings and juvenile
animals at ‘Ain Jammam. Kill-off focused on fully
grown adult animals, with only 38% of caprines
surviving beyond 30–48 months in age.
Differences in survivorship calculated for sheep
and goat indicate that different harvesting strategies
were in use for each taxon, with consistently higher
survivorship of juvenile and adult sheep relative to
goats (Fig. 7, Makarewicz 2009). Juvenile goats were
intensively harvested with only c. 57% of animals
surviving beyond 12–18 months compared to c. 78%
for sheep (Fig. 8). Sheep survivorship was promoted
well into young adulthood, followed by a pronounced
kill-off of mature adults.
There are some minor disparities between the
mandibular tooth wear and epiphyseal fusion survivor-
ship curves calculated for ‘Ain Jammam, particularly
in the youngest age categories due to small sample sizes
in the fusion stage I group, but the broad picture is the
same. The moderate kill-off of older kids/lambs aged
6–12 months, followed by intense culling of yearlings
and pronounced slaughter of adult animals (3–4 year
olds) suggests use of ‘type B milk’ and mixed meat and
milk harvesting strategies. Comparison of the DVL
and DVM measurements of fused and unfused
metacarpals and metatarsals suggests that young male
goats were a focus of slaughter. This, along with sex-
specific survivorship curves generated for both sheep
and goat indicating moderate kill-off of male kids/
lambs, may also indicate that tender meat may have
also been a production goal (Makarewicz 2009).
The same metrical data also suggests that, on
occasion, young female kids were slaughtered. Sex-
specific curves also hint at the kill-off of female
yearling goats, while female sheep were preserved as
juveniles and throughout adulthood (Makarewicz
2009). Since female goats generally do not begin to
produce offspring until their second year, it is
unusual that female goats would be slaughtered at
such a young age. This harvesting strategy may reflect
herder decisions to reduce surplus animals and
concentrate fodder stocks on older, reproductively
viable females.
Discussion
Goats were a central aspect of Late PPNB subsistence
and were exploited through application of a wide
variety of harvesting and dietary manipulation
strategies. The use of sex-based slaughtering strate-
gies at many sites reflects husbandry decisions that
were focused on the optimized production of meat
from older juvenile and adult animals, as well as the
extraction of tender meat from yearlings. Tender
meat is often required when social demands dictate
(Arbuckle et al. 2009; Russell 2012), and the frequent
use of this production strategy at many Late PPNB
settlements may indicate that it was socially advanta-
geous to do so, and served to increase social
negotiation between households during a period of
amplifying social differences.
The relatively high kill-off of lambs and kids
observed at several sites suggests milk production was
also part of Late PPNB sheep and goat production
systems. While recognizing that milk off-take may
have occurred on an irregular basis, previous research
has largely discounted the practice of dairying as a
consistent and widely practiced caprine exploitation
strategy (Becker 2004). Although additional biome-
trical analyses are needed in order to further support
their findings, Vigne and Helmer (2007) have argued
for milk exploitation at Middle PPNB Halula, citing
the high kill-off of very young lambs, and ‘type B
milk’ exploitation at Middle PPNB Aswad (i.e.
slaughter of a few lambs 2 to 6 months, followed by
intensive kill-off of animals 6–12 months in age,
followed by the moderate slaughter kill-off of older
females when their milk production decreases).
The absence of female adult goats in their
reproductive prime at some sites suggests the surplus
generation of female animals at producer sites and
may also indicate the emergence of nascent animal
exchange systems during the Late PPNB. Whether
based in subsistence concerns or socio-political
motivations, the exchange of live animals has
significant impacts on the economic and social
landscapes of both producers and consumers. The
high reproductive and productive potential of female
animals confers high value on these animals, offering
individuals who own an excess of female caprines the
opportunity to adjust their economic and social
positioning. Exchange of live animals may have been
an avenue through which widening social difference
manifested during the Late PPNB.
Sheep were preserved well into adulthood at some
settlements, a strategy that may reflect an alternative
form of subsistence risk reduction that is not
inconsistent with wealth accumulation. Domestic
animals are frequently imbued with different types
and gradations of value by the people who exploit
them and are a medium that can be transformed into
wealth and prestige (Russell 2012). Sheep, as a novel
technology that was rapidly introduced to the south-
ern Levant during the Late PPNB (Horwitz et al.
1999), were not as readily accessible as goats at many
settlements. The charismatic novelty and rarity of the
animal, combined with its enhanced productive
Makarewicz More than meat
258 Levant 2013 VOL 45 NO 2

qualities of richer milk and more tender, flavourful
meat, may have encouraged herders to retain their
sheep as long as possible. As a reservoir of wealth,
sheep may have provided a mechanism through
which herders could negotiate social and political
landscapes.
Conclusion
Sheep and goat husbandry played a pivotal but
poorly understood role in Late PPNB communities
east of the Jordan Valley. Although previous research
has solidly documented the ubiquitous importance of
caprine herding at these settlements and has provided
some insights into the broad animal production goals
of Late PPNB herders, the species-specific analyses
presented here demonstrate a much more complex
and dynamic picture of pastoralist activity during the
Late PPNB. Rather than a one-dimensional sub-
sistence activity focused solely on meat production
through use of a single harvesting strategy focused on
the kill-off of adult animals, as has been previously
suggested (Becker 2004; Twiss 2007; von den Driesch
et al. 2004; von den Driesch and Wodtke 1997; Wasse
2002), sheep and goat husbandry practices appear to
have consisted of a diverse series of interlinked
strategies that permitted herders to extract a wider
variety and quantity of products, including milk,
from their animals. By using an array of strategies
applied at different intensities and timings to various
animal categories, herders could also glean more
from limited resources such as land, water, and graze
that were likely increasingly pressured and less
productive, due to the rise in domesticate animals
use and human population packing of the landscape
hypothesized for the Late PPNB.
No less significantly, zooarchaeological analyses
such as the ones presented here provide the detail
needed to isolate potential links between subsistence
and ‘subsistence-plus’ economic activity and Late
PPNB household and community social organiza-
tion. Current models conceptualize Late PPNB
society as one where households had begun to acquire
materials beyond what was required for subsistence
and emerge as significant controllers of wealth (Byrd
1994). Although these models imply that agricultural
products were the medium through which wealth was
accumulated, pointing to the large internal storage
features in which grain could be concealed in a
manner likely to discourage sharing, the shift in social
organization during the Late PPNB also coincides
with the first time that domestic animals dominate the
subsistence spectrum, suggesting that these processes
are linked. Livestock herds represent an element of
increased control over a resource and, in the case of
the Late PPNB, possibly ownership. Moreover, in
many pastoralist and agro-pastoralist societies,
domestic animals are a store of wealth, which can
be accrued in live animals or generated through the
raw and transformed forms of meat, milk, and fibre
they produce (Lemonnier 1993; Russell 2012), and
provide a point of entry into multiple economic,
social, and political systems widely accessible to
individual households. Through refined application
of various husbandry practices to their flocks, herders
can accumulate and unload wealth relatively rapidly.
Given that herders are not uniformly skilled at
maintaining and enhancing the productive and
reproductive capabilities of their herds, the potential
for wealth differential between herding households is
high. Although the relationship between wealth
accumulation and animal-based production during
the Late PPNB certainly needs further exploration,
the emergence of new and diverse domestic livestock
herding strategies hints at the intentional creation of
new production pathways that expanded the human
subsistence and wealth possibilities.
Acknowledgements
Support for analysis of zooarchaeological materials
from el-Hemmeh, ‘Ain Jammam, es-Sifiya, and Wadi
Shu’eib was provided by the American School
for Prehistoric Research, the Department of An-
thropology at Harvard University, the Archaeology
Center at Stanford University, and the Christian-
Albrechts University, Kiel. I would like to especially
thank Ben Arbuckle for his comments clarifying the
interpretation of survivorship curves. I would like to
thank Ines Reese for her assistance with the figures,
and Graham Philip for his patience. I would also
like to thank Bill Finlayson and three anonymous
reviewers, whose comments greatly improved the
quality of this paper.
BibliographyArbuckle, B. S., Otzan, A. and Gulcur, S. (2009) The evolution of sheep
and goat husbandry in central Anatolia. Anthropozoologica 44,
129–57.
Baadsgaard, A. (2000) Subsistence change during the Epipaleolithic of
southern Jordan: a view from Wadi Mataha. Unpublished PhD
thesis, Brigham Young University.
Balasse, M. and Tresset, A. (2002) Early weaning of Neolithic domestic
cattle (Bercy, France) revealed by intra-tooth variation in nitrogen
isotope ratios. Journal of Archaeological Science 29, 853–59.
Bar-Yosef, O. (2000) The context of animal domestication in south-
western Asia. Pp. 184–94 in H. Buitenhuis, M. Mashkour,
M. Choyke and F. Poplin (eds), Archaeozoology of the Near
East IV. Groningen: ARC-Publication.
Barzilai, O. and Getzov, N. (2011) The 2010 excavation season at
Mishmar Ha’emeq in the Jezreel Valley. Neolithics 1/11, 19–22.
Becker, C. (1991) The analysis of mammalian bones from Basta, a Pre-
Pottery Neolithic site in Jordan: problems and potential.
Paleorient 17, 59–75.
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 259

— (2004) On the identification of sheep and goats: the evidence from
Basta. Pp. 219–310 in H. J. Nissen, M. Muhesien and H. G. K.
Gebel (eds), Basta I: The Human Ecology. Berlin: ex oriente.
Bienert, H. D. (2004) The Pre-Pottery B (PPNB) in Jordan: first steps
towards proto-urban societies? Pp. 21–40 in H. D. Bienert, H. G.
K. Gebel and R. Neef (eds), Central Settlements in Neolithic
Jordan. Studies in Early Near Eastern Production, Subsistence,
and Environment 5. Berlin: ex oriente.
Bocquentin, F., Barzilai, O. Khalaily, H. and Horwitz, L. (2011) The
PPNB site of Beisamoun (Hula Basin): present and past research.
Pp 197–212 in E. Healyer, S. Campbell and O. Maeda (eds), The
State of the Stone Terminologies, Continuities and Contexts in Near
Eastern Lithics: Proceedings of the Sixth PPN Conference on
Chipped and Ground Stone Artefacts in the Near East, Manchester,
3rd–5th March 2008. SENEPSE 13. Berlin: ex oriente.
Bocquet-Appel, J. P. (2002) Paleoanthropological traces of a Neolithic
demographic transition. Current Anthropology 43, 637–50.
Boessneck, J. (1969) Osteological Differences between Sheep (Ovis aries
Linne) and Goat (Capra hircus Linne). Pp. 331–58 in D. Brothwell
and E. Higgs (eds), Science and Archaeology. Praeger: New York.
Boessneck, J., Muller, H-H. and Teichert, M. (1964) Osteologische
Unterscheidungsmerkmale zwischen Schaf (Ovis aries Linne) und
Ziege (Capra hircus Linne) Kuhn Archiv 78, 1–129.
—, Mu ller, H.-H and Teichert, M. (1964) Osteologische
Unterscheidungsmerkmale zwischen Schaf (Ovis aries Linne)
und Ziege (Capra hircus Linne) Kuhn Archiv 78, 1–129.
Byrd, B. F. (1994) Public and private, domestic and corporate: the
emergence of the southwest Asian Village. American Antiquity 59,
639–66.
Carruthers, D. B. (2002) The Dana-Faynan-Ghuywayr early prehis-
toric project preliminary animal bone report on mammals from
Wadi Faynan 16. Pp. 93–97 in H. Buitenhuis, A. M. Choyke,
M. Mashkour and A. H. al-Shiyab (eds), Archaeozoology of the
Near East V. Groningen: ARC-Publication 62.
Colledge, S. (2001) Plant Exploitation on Epipaleolithic and Early
Neolithic Sites in the Levant. Oxford: BAR International Series
986.
Dahl, G. and Hjort, A. (1976) Having Herds: Pastoral Growth and
Household Economy. Stockholm: University of Stockholm.
Davis, S. J. M. (2000) The effect of castration and age on the
development of the Shetland sheep skeleton and a metric
comparison between bone of males, females, and castrates.
Journal of Archaeological Science 27, 373–90.
Eshed, V., Gopher, A., Gage, T. and Hershkovitz, I. (2004) Has the
transition to agriculture reshaped the demographic structure of
prehistoric populations? New evidence from the Levant. American
Journal of Physical Anthropology 124, 315–29.
Fino, N. (2004). Evidence for settlement organization at ‘Ain Jammam.
Pp. 105–11 in H. D. Bienert, H. G. K. Gebel and R. Neef (eds),
Central Settlements in Neolithic Jordan. Studies in Early Near
Eastern Production, Subsistence, and Environment 5. Berlin: ex
oriente.
Fujii, S. (2009) Wadi Abu Tulayha: a preliminary report of the 2008
summer final field season of the Jafr Basin Prehistoric Project,
Phase 2. ADAJ 53, 173–209.
Garrard, A., Baird, D., Colledge, S., Martin, L. and Wright, K. (1994)
Prehistoric environment and settlement in the Azraq Basin: an
interim report on the 1987 and 1988 excavation season. Levant 26,
73–109.
Gebel, H. G. K. (2004) Central to what? The centrality issue of the
LPPNB Mega-site phenomenon in Jordan. Pp. 1–19 in H. D.
Bienert, H. G. K. Gebel and R. Neef (eds), Central Settlements in
Neolithic Jordan. Studies in Early Near Eastern Production,
Subsistence, and Environment 5. Berlin: ex oriente.
— (2010) Commodification and the formation of Early Neolithic social
identity. The issues as seen from the southern Jordanian
Highlands. Pp. 35–80 in M. Benz (ed.), The Principle of Sharing.
Segregation and Construction of Social Identities at the Transition
from Foraging to Farming. Studies in Early Near Eastern
Production, Subsistence, and Environment 14. Berlin: ex
oriente.
— (2013) Basta radiocarbon dates. PPND: The Platform for Neolithic
Radiocarbon Dates. http://www.exoriente.org/associated_projects/
ppnd_site.php?s512
— and Bienert, H. D. (1997) Ba’ja hidden in the Petra mountains.
Preliminary report on the 1997 excavations. Pp. 221–62 in H. G.
K. Gebel, Z. Kafafi and G. O. Rollefson (eds), The Prehistory of
Jordan II. Perspectives from 1997. Studies in Early Near Eastern
Production, Subsistence, and Environment 4. Berlin: ex oriente.
Goring-Morris, A. N. and Belfer-Cohen, A. (2008) A roof over one’s
head: developments in Near Eastern residential architecture across
the Epipaleolithic-Neolithic transition. Pp. 239–86 in J.-P.
Bocquet-Appel and O. Bar-Yosef (eds), The Neolithic
Demographic Transition and its Consequences. Springer.
Gorsdorf, J. (2000) 14C ratings of the es-Sifiya settlement (Area C). Pp.
15–19 in H. D. Bienert and B. Muller-Neuhof (eds), At the
Crossroads: Essays on the Archaeology, History, and Current
Affairs of the Middle East. Amman: German Protestant Institute
of Archaeology in Amman.
Hecker, H. M. (1982) Domestication revisited: its implications for
faunal analysis. Journal of Field Archeology 9, 217–36.
Helmer, D. and Rocheteau, M. (1994) Atlas du squelette appendicu-
laire des principaux genres Holocenes de petits ruminants du nord
de la Mediterranee et de procheorient (Capra, Ovis, Rupicapra,
Capreolus, Gazella). Riches D’osteologie Animale Pour
L’archeologie. Serie B: Mammiferes 4, 3–21.
Hesse, B. (1984) These are our goats: the origins of herding in west
central Iran. Pp. 243–64 in J. Clutton-Broack and C. Grigson
(eds), Animals and Archaeology: 3. Early Herders and their Flocks.
Oxford: BAR International Series.
Horwitz, L. K. (2003) Temporal and spatial variation in Neolithic
caprine exploitation strategies: a case study from the site of
Yiftah’el (Israel). Paleorient 29, 19–58.
— and Ducos, P. (1998) An investigation into the origins of domestic
sheep in the southern Levant. Pp. 80–94 in H. Buitenhuis,
L. Bartosiewicz and A. M. Choyke (eds), Archaeozoology of the
Near East III Proceedings of the Third International Symposium on
the Archaeozoology of Southwestern Asia and Adjacent Areas.
Groningen: ARC Publication 18.
— and Ducos, P. (2005) Counting cattle: trends in Neolithic Bos
frequencies from the southern Levant. Revue de Paleobiologie,
Geneve 10, 209–24.
—, Tchernov, E., Ducos, P., Becker, C., von den Dreisch, A., Martin,
L. and Garrard, A. (1999) Animal domestication in the southern
Levant. Paleorient 25, 63–80.
Hunt, C. O., Elrishi, H. A., Gilbertson D. D., Grattan, J., McLaren, S.,
Pyatt, F. B., Rushworth, G. and Barker, G. W. (2004) Early
Holocene environments in the Wadi Faynan, Jordan. The
Holocene 14, 921–30.
Khalaily, H., Milevski, Y., Getzov, N., Hershkovitz, I., Barzilai, O.,
Yarosevich, A., Shlomi, V., Najjar, A., Zidan, O., Smithline,
H. and Liran, R. (2008) Recent excavations at the Neolithic site of
Yiftahel (Khalet Khalladyiah), Lower Galilee. Neo-Lithics 2/08,
3–11.
Kohler-Rollefson, I., Quintero, L. and Rollefson, G. (1993) A brief
note on the fauna from Neolithic ‘Ain Ghazal. Paleorient 19, 95–
97.
Kratochvil, Z. (1969) Species criteria on the distal section of the tibia in
Ovis ammon F. aries L. and Capra aegagrus F. hircus L. Acta
Veterinaria (Brno) 38, 483–90.
Kuijt, I. (2008) Demography and storage systems during the southern
Levantine Neolithic demographic transition. Pp 287–313 in J.-P.
Bocquet-Appel and O. Bar-Yosef (eds), The Neolithic
Demographic Transition and its Consequences. New York:
Springer.
— and Goring-Morris, N. (2002) Foraging, farming, and social
complexity in the Pre-Pottery Neolithic of the southern Levant:
a review and synthesis. Journal of World Prehistory 16, 361–440.
Lemonnier, P. (1993) Pigs as ordinary wealth: technical logic, exchange,
and leadership in New Guinea. Pp. 126–56 in P. Lemonnier (ed.),
Technological Choice: Transformations in Material Culture since
the Mesolithic. London: Routledge.
Levy, T. E., Adams, R. B., Witten, A. J., Anderson, J., Arbel, Y.,
Kuah, S., Moreno, J., Lo, A. and Wagonner, M. (2001) Early
metallurgy, interaction, and social change: the Jabal Hamrat
Fidan (Jordan) research design and 1998 archaeological survey:
preliminary report. Annual of the Department of Antiquities of
Jordan 45, 1–31.
Makarewicz More than meat
260 Levant 2013 VOL 45 NO 2

Mahasneh, H. M. (1997) The 1995 season at the Neolithic site of es-
Sifiya, Wadi Mujib, Jordan. Pp. 203–14 in H. G. K. Gebel,
Z. Kafafi and G. O. Rollfeson (eds), The Prehistory of Jordan,
II. Perspectives from 1997. Berlin: ex oriente.
Makarewicz, C. (2009) Complex caprine harvesting practices and
diversified hunting strategies: integrated animal exploitation
systems at Late Pre-Pottery Neolithic B ‘Ain Jammam.
Anthropozoologica 44/1, 1–24.
— (2011) Xiongnu pastoral systems: integrating economies of
subsistence and scale. Pp. 181–92 in U. Brosseder and B. K.
Miller (eds), Xiongnu Archaeology — Multidisciplinary
Perspectives on the First Steppe Empire in Central Asia. Bonn:
Bonn Contributions to Asian Archaeology 5.
— (in press) Assorted strategies of production and risk reduction in
Late PPNB caprine herding systems: zooarchaeological and
isotopic case studies from el-Hemmeh and Ba’ja. In G.
Rollefson and B. Finlayson (eds), Jordan’s Prehistory: Past and
Future Research. Amman: Department of Antiquities, Jordan.
— (forthcoming) Pig domestication processes and caprine husbandry at
Neolithic Wadi Shu’eib, Jordan: a zooarchaeological approach.
— and Austin, A. E. (2006) Late PPNB architecture at el-Hemmeh:
results from the third excavation season, 2006. Neolithics 10, 18–
23.
— and Tuross, N. (2006) Foddering by Mongolian pastoralists is
recorded in the stable carbon (d13C) and nitrogen (d15N) isotopes
of caprine dentinal collagen. Journal of Archaeological Science 33,
862–70.
— and Tuross, N. (2009) Variation in goat diet through the Later Pre-
Pottery Neolithic: diachronic shifts in human approaches to
caprine management at Basta. Pp. 287–304 in J. Shea and D.
Lieberman (eds), Transitions in Prehistory: Papers in Honor of
Ofer Bar-Yosef. Cambridge: American School of Prehistoric
Research.
— and Tuross, N. (2012) Finding fodder and tracking transhumance:
isotopic detection of early goat domestication processes in the
Near East. Current Anthropology 53, 495–505.
—, Goodale, N., Rassmann, P., White, C., Miller, H., Haroun, J.,
Carlson, E., Pantos, A., Kroot, M., Kadowaki, S., Casson, A.,
Williams, J. T., Austin, A. E. and Fabre, B. (2006) El-Hemmeh: a
multi-period Pre-Pottery Neolithic site in the Wadi el-Hasa,
Jordan. Eurasian Prehistory 4/1–2, 183–220.
Maltby, J.M. (1985) Patterns in faunal assemblage variability. Pp. 33–
74 in G. Barker and C. Gamble (eds), Beyond Domestication in
Prehistoric Europe: Investigations in Subsistence Archaeology and
Social Complexity. London: Academic Press.
Meadow, R. (1999) The use of size index scaling techniques for research
on archaeozoological collections from the Middle East. Pp. 285–
300 in C. Becker, H. Manhart, J. Peters and J. Schibler (eds),
Historia animalium ex ossibus: Beitrage zur Palaoanatomie,
Archaologie, Agyptologie, Ethnologie, und Geschichte der
Tiermedizin. Rahden: Verlag Marie Leidorf.
Milevski, I., Khalaily, H., Getzov, N. and Hershkovitz, I. (2008) The
plastered skulls and other PPNB finds from Yif-tahel, Lower
Galilee (Israel). Paleorient 34/2, 37–46.
Munson, P. J. (2000). Age-correlated differential destruction of bone
and its effects on archaeological mortality profiles of domestic
sheep and goats. Journal of Archaeological Science 27, 391–407.
Neef, R. (2004) Vegetation and plant husbandry. Pp. 187–218 in H. J.
Nissen, M. Muheisen and H. G. K. Gebel (eds), Basta I: The
Human Ecology. Berlin: ex oriente.
Nissen, H. J., Muheisen, M., Gebel, H.-G., Becker, C., Neef, R.,
Pachur, H.-J., Qadi, N. and Shultz, M. (1987) Report on the first
two seasons of excavations at Basta (1986–1987). Annual of the
Department of Antiquities of Jordan 31, 79–119.
Noddle, B. A. (1978) Some minor skeletal differences in sheep. Pp. 133–
41 in D.R. Brothwell, K.D. Thomas and J. Clutton-Brock (eds),
Research Problems in Zooarchaeology. Institute of Archaeology
Occasional Publication 3: London.
Payne, S. (1969) A metrical distinction between sheep and goat
metacarpals. Pp. 295–306 in P. J. Ucko and G. W. Dimbleby (eds),
The Domestication and Exploitation of Plants and Animals.
London: Duckworth.
— (1973) Kill-off patterns in sheep and goat: the mandible from Asvan
Kale. Anatolian Studies 23, 281–306.
— (1975) Partial recovery and sample bias. Pp. 7–17 in A. T. Clason
(ed.), Archaeozoological Studies. Amsterdam: Elsevier.
— (1985) Morphological distinctions between the mandibular teeth of
young sheep, Ovis, and goats, Capra. Journal of Archaeological
Science 12, 139–47.
Petersen, J. (2004) Khirbet Hammam (WHS 149): a Late Pre-Pottery
Neolithic B settlement in the Wadi el-Hasa, Jordan. Bulletin of the
American Schools of Oriental Research 334, 1–17.
Prummel, W. and Frisch, H. (1986). A guide for the distinction of
species, sex and body side in bones of sheep and goat. Journal of
Archaeological Science 13, 5567–77.
Redding, R. (1981) Decision-making in subsistence herding of sheep
and goats in the Middle East. Unpublished PhD thesis, University
of Michigan.
Rollefson, G. O. (2005) Stone tools from ‘Ayn Jamman, near Ras en-
Naqb, Jordan. Neolithics 5, 17–23.
— (2010). Blood loss: realignments in community social structures in
the LPPNB of highland Jordan. Pp. 183–202 in M. Benz (ed.), The
Principle of Sharing. Segregation, and Construction of Social
Identities at the Transition from Foraging to Farming. Berlin: ex
oriente.
Russell, N. (2012) Social Zooarchaeology: Humans and Animals in
Prehistory. Cambridge: Cambridge University Press.
Simmons, A., Rollefson, G. O., Kafafi, Z., Mandel, R. D., al-Nahar,
M., Cooper, J., Kohler-Rollefson, I. and Durand, K. (2001) Wadi
Shu’eib, a large Neolithic community in Central Jordan: final
report of test investigations. Bulletin of the American School of
Oriental Research 321, 1–39.
Stein, G. (1987) Regional economic integration in early state societies:
third millennium BC pastoral production at Gritille, southeast
Turkey. Paleorient 13, 101–11.
Twiss, K. (2003) They were what they ate: food and the construction of
social identity in the Neolithic of southern Jordan. Unpublished
PhD thesis, University of California, Berkeley.
— (2007) The zooarchaeology of Tel Tif-dan (Wadi Fidan 001),
southern Jordan. Paleorient 33, 127–45.
Uerpmann, H.-P. and Uerpmann, M. (1994) Animal bone finds from
Excavation 520 at Qala’at-Bahrain. Pp. 417–44 in F. Hojlund and
H. H. Andersen (eds), Qala’at al-Bahrain: The Northern City Wall
and the Islamic Fortress. Aarhus: Aarhus University Press.
Vigne, J.-D. and Helmer, D. (2007) Was milk a secondary product in
the Old World Neolithisation process? Its role in the domestica-
tion of cattle, sheep, and goats. Anthropozoologica 42, 9–40.
von den Driesch, A. and Wodtke, U. (1997) The fauna of ‘Ain Ghazal,
a major PPN and Early PN settlement in Central Jordan. Pp. 511–
56 in H. G. K. Gebel, Z. Kafafi and G. O. Rollefson (eds), The
Prehistory of Jordan II, Perspectives from 1997. Studies in Early
Near Eastern Production, Subsistence, and Environment 4. Berlin:
ex oriente.
von den Driesch, A., Cartajena, I. and Manhart, H. (2004) The Late
PPNB site of Ba’ja, Jordan: the faunal remains (1997 seasons). Pp.
271–88 in H. D. Bienert, H. G. K. Gebel and R. Neef (eds),
Central Settlements in Neolithic Jordan. Berlin: ex oriente.
Wasse, A. (2000) The development of goat and sheep herding during
the Levantine Neolithic. Unpublished PhD thesis, University
College London.
— (2002) Final results of analysis of the sheep and goat bone from ‘Ain
Ghazal, Jordan. Levant 34, 59–82.
White, C. and Wolff, N. (2012) Beyond the house: cultivation practices
in the Late PPNB. Pp. 279–300 in C. Foster and B. Parker (eds),
Household Archaeology in the Middle East and Beyond: Theory,
Method and Practice. Winona Lake, IN: Eisenbrauns.
Zeder, M. (2001) A metrical analysis of a collection of modern goats
(Capra hircus aegagrus and C. h. hircus) from Iran and Iraq:
implications for the study of domestication. Journal of
Archaeological Science 28, 61–79.
— (2006) Reconciling rates of long bone fusion and tooth eruption in
sheep (Ovis) and goat (Capra). Pp. 87–118 in D. Ruscillo (ed.),
Ageing and Sexing Animals from Archaeological Sites. Oxford:
Oxbow.
— and Lapham, H. A. (2010) Assessing the reliability of criteria used to
identify postcranial bones in sheep, Ovis, and goats, Capra.
Journal of Archaeological Science 37, 2887–905.
— and Pilaa, S. (2011) Assessing the reliability of criteria used to
identify manidibles and mandibular teeth in sheep, Ovis, and goats
Capra. Journal of Archaeological Science 37, 225–42.
Makarewicz More than meat
Levant 2013 VOL 45 NO 2 261




















