
Moluscos límnicos invasores no Brasil : biologia, prevenção e controle
412
Organizadores: Maria Cristina Dreher Mansur Cintia Pinheiro dos Santos Daniel Pereira Isabel Cristina Padula Paz Manuel Luiz Leite Zurita Maria Teresa Raya Rodriguez Marinei Vilar Nehrke Paulo Eduardo Aydos Bergonci
Transcript of Moluscos límnicos invasores no Brasil : biologia, prevenção e controle

Organizadores:Maria Cristina Dreher Mansur
Cintia Pinheiro dos SantosDaniel Pereira
Isabel Cristina Padula PazManuel Luiz Leite Zurita
Maria Teresa Raya RodriguezMarinei Vilar Nehrke
Paulo Eduardo Aydos Bergonci

Patrocínio e Apoio:

Organizadores:Maria Cristina Dreher Mansur
Cintia Pinheiro dos SantosDaniel Pereira
Isabel Cristina Padula PazManuel Luiz Leite Zurita
Maria Teresa Raya RodriguezMarinei Vilar Nehrke
Paulo Eduardo Aydos Bergonci
Porto Alegre2012

Dados Internacionais de Catalogação na Publicação (CIP)
Bibliotecário Responsável
Ginamara Lima Jacques PintoCRB 10/1204
E24 Moluscos límnicos invasores no Brasil : biologia, prevenção e controle / organizadorMaria Cristina Dreher Mansur ... [et al.]. – Porto Alegre : Redes Editora, 2012.
412 p. ; 29,7 cm.
ISBN: 978-85-61638-46-7
1. Meio Ambiente – Brasil. 2. Moluscos Límnicos. 3. Moluscos Invasores no Brasil. 4. Mexilhão-Dourado.I. Mansur, Maria Cristina Dreher.
CDD 639.42
Organizadores:Maria Cristina Dreher Mansur
Cintia Pinheiro dos SantosDaniel Pereira
Isabel Cristina Padula PazManuel Luiz Leite Zurita
Maria Teresa Raya RodriguezMarinei Vilar Nehrke
Paulo Eduardo Aydos Bergonci
Projeto editorial:Guacira Gil e Salete Campos de Moraes
Redes Editora
Projeto gráfico, editoração e capas:Eleandro Moysés
Foto capa:Macroaglomerado de mexilhão-dourado, Lago Guaíba, RS. M.C.D. Mansur.
Impressão:Gráfica Evangraf
Tiragem: 1000 exemplares
Redes Editora Ltda.Av. Plínio Brasil Milano, 388, conj. 501 – 90520-000 – Porto Alegre/RS – Brasil
http://www.redeseditora.com.br/loja - E-mail: [email protected]: (51)8109.4238

PREFÁCIO ...................................................................................09
APRESENTAÇÃO ................................................................ 11
LISTA DE COLABORADORES ...........................13
SEÇÃO 1Moluscos invasores nos ecossistemas continentais do Brasil
CAPÍTULO 1 ............................................................................19Abordagem conceitual dos moluscos invasores nos ecossistemas límnicos brasileiros
CAPÍTULO 2 ............................................................................25Espécies de moluscos límnicos invasores no Brasil
CAPÍTULO 3 ............................................................................ 51Genética da bioinvasão do mexilhão-dourado
SEÇÃO 2Biologia de moluscos invasores
CAPÍTULO 4 ............................................................................ 61Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
CAPÍTULO 5 ............................................................................ 75Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
CAPÍTULO 6 ............................................................................95Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
Sumário
CAPÍTULO 7 .........................................................................111A gametogênese em Limnoperna fortunei (Dunker, 1857)
CAPÍTULO 8 .........................................................................119Dinâmica reprodutiva de Corbicula fluminea e Corbicula largillierti
CAPÍTULO 9 ........................................................................ 125Gastrópodes límnicos invasores: morfologia comparada
SEÇÃO 3Monitoramento de moluscos invasores
CAPÍTULO 10 ......................................................................139Como monitorar bivalves invasores no plâncton? Método da microscopia óptica
CAPÍTULO 11 ......................................................................143O método molecular de prospecção do mexilhão-dourado
CAPÍTULO 12 ......................................................................149Método de detecção e quantificação de larvas do mexilhão-dourado Limnoperna fortunei, usando PCR quantitativo em tempo real
CAPÍTULO 13 ......................................................................155Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
SEÇÃO 4Aspectos populacionais de moluscos invasores e relações com variáveis ambientais

CAPÍTULO 14 ......................................................................187Limnoperna fortunei na Bacia da Lagoa dos Patos e Lagoa Mirim
CAPÍTULO 15 ......................................................................193Limnoperna fortunei na bacia hidrográfica do baixo Rio Jacuí e Lago Guaíba, Rio Grande do Sul
CAPÍTULO 16 ......................................................................197Limnoperna fortunei na bacia do Rio Paraguai, Mato Grosso do Sul
CAPÍTULO 17 ......................................................................201Limnoperna fortunei associada a macrófitas aquáticas na bacia do Rio Paraguai, Mato Grosso
CAPÍTULO 18 ..................................................................... 207Experiência da procura e monitoramento no início da invasão de mexilhão-dourado no Rio Paraná e no reservatório de Itaipu
SEÇÃO 5Moluscos invasores e a comunidade bentônica
CAPÍTULO 19 ......................................................................221Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
CAPÍTULO 20 ..................................................................... 235Efeito do macrofouling sobre a comunidade de invertebrados aquáticos
CAPÍTULO 21 ......................................................................243Limnoperna fortunei na bacia hidrográficado baixo Rio Jacuí: relações com a comunidade de macroinvertebrados bentônicos
SEÇÃO 6Efeito da filtração de bivalves invasores sobre a comunidade planctônica
CAPÍTULO 22 ..................................................................... 249O impacto de Limnoperna fortunei sobre as cianobactérias
CAPÍTULO 23 ......................................................................255Os impactos do mexilhão-dourado sobre a comunidade planctônica
SEÇÃO 7Prevenção
CAPÍTULO 24 ..................................................................... 265Ações de Eletrobras–FURNAS na divulgação daCampanha de Controle do Mexilhão-Dourado
CAPÍTULO 25 ......................................................................271Ações desenvolvidas pelo Ministério do Meio Ambiente – 2001 a 2011
SEÇÃO 8Delineamento experimental: seleção de métodos de controle populacional
CAPÍTULO 26 ......................................................................279Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
SEÇÃO 9Controle populacional de bivalves invasores: métodos químicos
CAPÍTULO 27 ..................................................................... 297Controle Químico: conceitos básicos
CAPÍTULO 28 ..................................................................... 299Microencapsulados
CAPÍTULO 29 ......................................................................303Formas de Cloro
CAPÍTULO 30 ......................................................................307Sulfato de Cobre
CAPÍTULO 31 ......................................................................311Outros compostos
CAPÍTULO 32 ......................................................................317Seleção de materiais e revestimentos parao controle de incrustações do mexilhão-dourado em hidrelétricas.
CAPÍTULO 33 ......................................................................323Tintas anti-incrustantes no controle do mexilhão-dourado

SEÇÃO 10Controle populacional de bivalves invasores: métodos físicos
CAPÍTULO 34 ..........................................................................331Controle Físico: conceitos básicos
CAPÍTULO 35 ..........................................................................335Controle de Limnoperna fortunei com aplicação de radiação ultravioleta
CAPÍTULO 36 ..........................................................................339Controle de bivalves com a utilização do ultrassom
SEÇÃO 11Controle populacional de bivalves invasores: métodos biológicos
CAPÍTULO 37 ..........................................................................345Controle Biológico: conceitos básicos
CAPÍTULO 38 ..........................................................................351Controle Microbiano
CAPÍTULO 39 ..........................................................................357Predadores potenciais para o controle do mexilhão-dourado
SEÇÃO 12Modelos de análise de risco e previsão de cenários de dispersão de moluscos invasores
CAPÍTULO 40 ..........................................................................367Análise de risco de introdução de moluscos aquáticos invasores
CAPÍTULO 41 ..........................................................................373Modelos de previsão da distribuição do mexilhão-dourado no Brasil
CAPÍTULO 42 ..........................................................................377Previsão de cenários de distribuição de Limnoperna fortunei na Laguna dos Patos
SEÇÃO 13Manejo de Espécies Invasoras
CAPÍTULO 43 ..........................................................................383Manejo integrado de espécies invasoras de moluscos límnicos
REFERÊNCIAS .......................................................................389


9
Prefácio
Com vistas a incentivar a busca constante por inovações e fazer frente aos desafios tecnológicos do setor elétrico, foi regulamentado o Programa de Pesquisa e Desenvolvimento (P&D). Neste contexto, as em-presas concessionárias, permissionárias ou autorizadas de distribuição, transmissão e geração de energia elétrica devem aplicar anualmente um percentual mínimo de sua receita operacional líquida no Programa de Pesquisa e Desenvolvimento do Setor de Energia Elétrica (ANEEL). Esta publicação é produto final do Projeto “Estudos biológicos e ambientais como base à prevenção e ao controle do mexilhão-dourado, Limnoperna fortunei (Dunker, 1857), em rios e reservatórios brasileiros”, que foi financiado pelo Programa de P&D ANEEL/FURNAS, tendo como sede das pesquisas o Centro de Ecologia da Universidade Federal do Rio Grande do Sul (UFRGS) e a Fundação Luiz Englert como admi-nistradora junto à citada Universidade. Está subdividido em 13 seções, dentro das quais estão distribuídos 43 capítulos, redigidos por diversos especialistas de projeção no Brasil e no exterior além dos pesquisadores envolvidos diretamente no projeto. Sua importância é singular, devido ao somatório de pesquisas direcionadas à compreensão do comporta-mento e do ciclo de vida do mexilhão-dourado, como base aos diferentes métodos testados para a prevenção e ao controle do mesmo. A chegada do mexilhão-dourado L. fortunei marcou um dos mais alarmantes casos de bioinvasão das águas continentais brasileiras. Atra-vés do seu alto poder reprodutivo este invasor tem causado sérios pro-blemas de entupimento nos sistemas coletores de água, canalizações, re-frigeradores de indústrias, unidades geradoras de energia e sistemas de cultivo em tanques-rede, bem como impactos ambientais, devido a sua alta densidade populacional. Os impactos no ambiente poderão ser mais expressivos em longo prazo com trocas na estrutura da cadeia trófica, como já relatados na Argentina e no Brasil. As espécies invasoras foram consideradas como a terceira maior ameaça para a biodiversidade pela União Internacional para a Conservação da Natureza (IUCN) e o Go-verno brasileiro. Já o Ministério do Meio Ambiente – MMA considera espécies introduzidas a segunda maior causa de extinção de espécies. A presente obra recebe importante contribuição com base em aná-lises genéticas das populações invasoras da espécie L. fortunei no Brasil, que podem ser resumidas como: o mexilhão-dourado invadiu as águas

10
continentais brasileiras mais de uma vez; as populações que invadiram o Brasil vieram de diferentes locais; a migração natural não é vetor mais importante deste processo de bioinvasão. A capacidade de detecção precoce de espécies aquáticas invasoras durante o processo de sua dispersão é essencial para a definição de medi-das de controle ou erradicação. O monitoramento de bivalves invasores pode se dar por vários métodos. O mais óbvio e simples é a observação das conchas dos indivíduos adultos no ambiente. Embora esta seja a ma-neira mais fácil, a detecção é tardia e prejudica ações de controle ou eliminação dos moluscos. Para contornar este problema, são apresenta-dos, neste Guia, métodos clássicos de detecção morfológica das larvas, e técnicas mais sensíveis e específicas que se referem à detecção via PCR e quantificação de larvas em amostras de plâncton por PCR em tempo real. Na tentativa de controlar as espécies invasoras, são apresentados métodos químicos, físicos e biológicos, com comentários sobre as vanta-gens e desvantagens de cada um deles. O controle de organismos invasores (bioincrustações) no mundo está sendo impulsionado pela busca por maior eficiência com baixo cus-to, associado a um baixo impacto ambiental e operacional. Vale ressal-tar, que novas informações e desdobramentos dessa pesquisa financiada através do Programa da ANEEL de Pesquisa e Desenvolvimento, possibi-litarão a melhoria de procedimentos e a aplicação de ações preventivas e corretivas que aumentem a eficiência no controle do mexilhão-dourado, contribuindo efetivamente para uma performance adequada do setor elétrico, preservando e cuidando do meio ambiente.
Paulo S. Formagio

11
Apresentação
O ambiente de água doce, já tão disputado pelo homem e que em futuro muito próximo deverá apresentar os problemas mais críticos no nosso planeta devido à escassez de água, nas últimas décadas tem ser-vido de cenário para as invasões causadas por moluscos límnicos, com todas as consequências para o ecossistema e aos sistemas construídos pelo homem. Os impactos destas invasões têm sido complexos e incal-culáveis. No meio ambiente, a integridade das comunidades naturais é afetada com reflexos na biodiversidade, na cadeia trófica, na qualidade ambiental, com riscos à saúde humana e problemas sanitários imprevisí-veis. Nos sistemas de hidroelétricas, barramentos, refrigeração de indús-trias, canalizações, tratamento de água, pesca, turismo etc., os prejuízos econômicos ultrapassam a casa dos bilhões de dólares americanos. As invasões causadas por espécies exóticas são uma consequência da própria evolução do homem moderno. As companhias de comércio operam globalmente (Walker, 2005), contribuindo para a eliminação ou redução das barreiras naturais que sempre separaram e mantiveram a integridade dos ecossistemas (Silva et al., 2004). Os bivalves asiáticos do gênero Corbicula chegaram por primeiro na década de 1970 e o mexilhão-dourado na de 1990. Todas as áreas dos primeiros registros foram nas cercanias de portos, recaindo a suspeita de que tenham sido transportados na água de lastro de navios transoceâni-cos. Acompanhamos pessoalmente as invasões dos bivalves junto à ba-cia do Lago Guaíba que banha Porto Alegre, capital do Rio Grande do Sul, no extremo sul do Brasil. Como especialista em bivalves nativos, foi muito difícil explicar o que estávamos presenciando. As primeiras dúvidas surgiram com todas aquelas formas diferentes de Corbicula que foram aparecendo em sequência, uma dando lugar à outra. Seriam uma ou mais espécies? Nossa espécie nativa Cyanocyclas limosa, da mesma família, havia praticamente desaparecido. Depois o mexilhão-dourado. Em aproximadamente dois anos após os primeiros registros, a popula-ção alcançou densidades de 140.000 indivíduos e recrutas em mais de 1 milhão por m2. Ficamos pasmos com a rapidez da modificação do am-biente lacustre. Cada mês que íamos a campo havia uma surpresa, uma nova alteração, principalmente em se tratando da fauna e flora ripária e bentônica. Todas as estações de tratamento da água e sistemas de refri-geração de fábricas que utilizam água bruta no entorno do lago, tiveram problemas de entupimentos e partiram para soluções emergenciais – nem

12
sempre recomendadas ambientalmente. Foi também dificílimo, inicial-mente, convencer e sensibilizar os órgãos ambientais – para obtermos licenças de coleta – e os financiadores de pesquisa, sobre a necessidade de desenvolvermos pesquisas sobre o tema e ter projetos aprovados. Esta obra representa o esforço de uma equipe de biólogos especia-lizados em diferentes áreas, desde a Taxonomia, Ecologia, Biologia Mo-lecular e Controle Biológico, com apoio de engenheiros químicos, am-bientais, e barrageiros, no sentido de reunir dados disponíveis e integrar os conhecimentos. O ponto de partida para a organização de todos os es-tudos e subprojetos foi, inicialmente, a pesquisa básica, que nos permitiu aprofundar os conhecimentos sobre taxonomia e morfologia comparada das espécies de bivalves invasores, em todas as fases de desenvolvimento, dando-nos a certeza da identidade, do número de espécies e das respec-tivas áreas de invasão. Calcados neste pilar, e no aprendizado de novas técnicas de coleta e monitoramento foi possível, com mais segurança, direcionar as pesquisas para a compreensão das interferências e atuações das espécies invasoras no meio ambiente e realizar experimentos para adequar e adaptar técnicas direcionadas ao controle das diferentes espé-cies invasivas de moluscos no ambiente límnico brasileiro. E, por último, objetivando um controle com técnicas ambientalmente compatíveis, su-gerir um manejo integrado. A ideia do livro surgiu durante a elaboração de um projeto de Pesquisa e Desenvolvimento (P & D) sugerido pelo biol. Rodrigo De Filippo, da empresa FURNAS, por demanda da mesma. Este projeto foi idealizado em 2008/9, iniciado em outubro de 2009, com a finalização prevista para setembro de 2012 e o lançamento do livro em julho deste ano. Reunimos neste livro também o resultado de projetos desenvolvidos anteriormente, com recursos do GLOBALLAST que subsidiou a Força Tarefa Nacional de Combate ao Mexilhão-Dourado no período de 2003 a 2004; com o Auxílio para pesquisa CTHIDRO/CNPq durante o pe-ríodo de 2005 a 2008, sob a coordenação geral do Dr. Flávio da Costa Fernandes do IEAPM – Marinha do Brasil, além de recursos obtidos pelo edital Universal do CNPq e projetos de P & D financiados pela TRACTEBEL, AES Tietê, ENERPEIXE e CEEE. Parte das pesquisas que compõem vários capítulos deste livro, fo-ram desenvolvidas no Centro de Ecologia da Universidade Federal do Rio Grande do Sul, com recursos FURNAS/ANEEL que permitiram equipar e adaptar oito laboratórios para estudos do mexilhão-dourado. Também foram convidados pesquisadores de outras instituições brasileiras (IEAPM, IFRR, NUPÉLIA/UEM, FURG, UFMT, UERJ, ICMBio, IBA-MA, MMA, FURG, FIOCRUZ, UFP) e do exterior (UNRN e UNLP, na Argentina, e ICPIEE, no Japão), que contribuíram com textos de altíssi-ma qualidade para a obra apresentada. Quando este livro estiver publicado é possível que invasoras mais agressivas tenham chegado ao nosso país e novas técnicas mais eficientes tenham sido pesquisadas e testadas. Assim este livro não é definitivo, mas uma ponte para facilitar novos estudos e descobertas.
Maria Cristina Dreher Mansur

13
ColaboradoresARCHUBY, FERNANDOUniversidad Nacional de Río Negro (UNRN), Instituto de Paleobiología y Geología, Calle Isidro Lobos y Belgrano, 8332, General Roca, Río Negro, Argentina, e-mail: [email protected]
ARENZON, ALEXANDREUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
ARRUDA, JANINE OLIVEIRAUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
BARBOSA, FABIANA GONÇALVESUniversidade Federal do Rio Grande (FURG), CENOSYS, Av. Itália km 8, Carreiros, 96203-900Rio Grande, RS, Brasil, e-mail: [email protected]
BASEGIO, TANIA MARIA Universidade Federal do Rio Grande do Sul (UFRGS), Escola de Engenharia, Departamento de Materiais, Av. Osvaldo Aranha, 99, Centro, 90035-190 Porto Alegre, RS, Brasil, e-mail: [email protected]
BELZ, CARLOS EDUARDOUniversidade Federal do Paraná (UFPR), Centro de Estudos do Mar, CEM, Av. Beira Mar, s/n, Balneário Pontal do Sul, Pontal do Paraná, 83255-000 Pontal do Paraná, PR, Brasil, e-mail: [email protected]
BEMVENUTI, CARLOS EMÍLIOUniversidade Federal do Rio Grande (FURG), Instituto de Oceanografia, Laboratório de Ecologia de Invertebrados Bentônicos, Campus Carreiros, Base Oceanográfica, Av. Itália, Km 8, s/n, Cx. postal 474, 96201-900Rio Grande, RS, Brasil, e-mail: [email protected]
BERGMANN, CARLOS PÉREZUniversidade Federal do Rio Grande do Sul (UFRGS), Escola de Engenharia, Departamento de Materiais, Av. Oswaldo Aranha 99 sala 705, 90035-190 Porto Alegre, RS, Brasil, e-mail: [email protected]
BERGONCI, PAULO EDUARDO AYDOS Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil,e-mail: [email protected]
BERUTTI, FELIPE AMORIMUniversidade Federal do Rio Grande do Sul (UFRGS), Escola de Engenharia, Departamento de Materiais, Av. Osvaldo Aranha, 99 sala 705C, 90035-190 Porto Alegre, RS, Brasil, e-mail: [email protected]
BOEGER, WALTER ANTONIOUniversidade Federal do Paraná (UFPR), Departamento de Zoologia, Setor de Ciências Biológicas, Grupo Integrado de Aquicultura e Estudos Ambientais, Av. Francisco H. dos Santos, s/n, Jardim das Américas, Cx. postal 19073, 81531-980 Curitiba, PR, Brasil, e-mail: [email protected]
CALAZANS C., SÁVIO HENRIQUEInstituto de Estudos do Mar Almirante Paulo zoreira (IEAPM), Rua Kioto, 253, Praia dos Anjos, 28930-000 Arraial do Cabo, RJ, Brasil, e-mail: [email protected]
CALHEIROS, DÉBORA FERNANDESEmbrapa PantanalRua 21 de Setembro, 1880, Corumbá, MSe-mail: [email protected]
CALLIL, CLAUDIA TASSOUniversidade Federal do Mato Grosso (UFMT), Instituto de Biociências, Departamento de Biologia e Zoologia, Av. Fernando Correa da Costa, n° 2367, Bairro Boa Esperança, 78060-900 Cuiabá, MT, Brasil, e-mail: [email protected]
COLLING, LEONIR ANDRÉUniversidade Federal do Rio Grande (FURG), Instituto de Oceanografia, Laboratório de Ecologia de Invertebrados Bentônicos, Av. Itália, Km 08, Cx. postal 474, 96203-000 Rio Grande, RS, Brasil, email: [email protected]
COUTINHO, RICARDOInstituto de Estudos do Mar Almirante Paulo Moreira (IEAPM), Rua Kioto, 253, Praia dos Anjos, 28930-000 Arraial do Cabo, RJ, Brasil, e-mail: [email protected]

14
DANELON, OLGA MARIAInstituto de Estudos do Mar Almirante Paulo Moreira (IEAPM),Rua Kioto, 253. Praia dos Anjos, 28930-000 Arraial do Cabo, RJ, Brasil, e-mail: [email protected]
DARRIGRAN, GUSTAVOMuseo de La Plata, División Zoología Invertebrados (UNLP - FCNyM), Grupo de Investigación sobre Moluscos Invasores/Plagas (GIMIP), Paseo del Bosques /n, La Plata 1900, La Plata, Argentina, www.malacologia.com.ar , e-mail: [email protected]
DE FILIPPO, RODRIGORua Pitangueiras 315/73, Mirandópolis, 04052-020 São Paulo, SP, Brasil, e-mail: [email protected]
ENDO, NORIYUKIInstituto Central de Pesquisa da Indústria de Energia Elétrica (ICPIEE), Laboratório de Pesquisa de Ciências Ambientais, 1646 Abiko, Abiko-shi, Chiba-ken 270-1194, Japão, Endereço atual: Himeji Eco tech Co., Ltd., Himeji, 672-8023 Hyogo, Japão, e-mail: [email protected]
FACHINI, ALINEUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
FERNANDES, FLAVIO DA COSTAInstituto de Estudos do Mar Almirante Paulo Moreira (IEAPM),Rua Kioto, 253. Praia dos Anjos, 28930-000 Arraial do Cabo, RJ, Brasil, e-mail: [email protected]
FERNADES, LIVIA VIANA DE GODOYInstituto de Estudos do Mar Almirante Paulo Moreira (IEAPM), Rua Kioto, 253. Praia dos Anjos, 28930-000 Arraial do Cabo, RJ, e-mail: [email protected]
FERNANDEZ, MONICA AMMONLaboratório de Malacologia, Instituto Oswaldo Cruz (FIOCRUZ), Av. Brasil 4365, Manguinhos, 21040-900 Rio de Janeiro, RJ, Brasil, e-mail: [email protected]
FIGUEIREDO, GABRIELA CRISTINA SANTOS DEUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
FORMAGIO, PAULO SERGIOEstação de Hidrobiologia e Piscicultura de Furnas (ELETROBRAS/FURNAS). Rua Lavras 288, 37945-000 Furnas, MG, Brasil, e-mail: [email protected]
FREITAS, SUZANA MARIA FAGONDES DE Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Laboratório de Invertebrados Bentônicos I, Av. Bento Gonçalves, 9500, prédio 43435, sala 204, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
FUJITA, DANIELE SAYURIInstituto Federal de Educação, Ciência e Tecnologia de Roraima (IFECTR), Campus Amajari, Av. Tepequém, s/n Centro, 69343-000 Amajari, RR, Brasil, e-mail: [email protected]
GAZULHA, VANESSAUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970, Porto Alegre, RS, Brasil, e-mail: [email protected]
GOMES, ANA LÚCIA TEIXEIRAUniversidade Federal de Mato Grosso (UFMT), Instituto de Biociência, Laboratório de Ecologia Animal CCBS-III, Av. Fernando Corrêa da Costa, n° 2367, Bairro Boa Esperança, 78060-900 Cuiabá, MT, Brasil, e-mail: [email protected]
GONÇALVES, ISABELA CRISTINA BRITOUniversidade do Estado do Rio de Janeiro (UERJ), Instituto de Biologia Roberto Alcântara Gomes, Laboratório de Malacologia Límnica e Terrestre, Rua São Francisco Xavier, 524, PHLC 525-2, Maracanã,20550-900 Rio de Janeiro, RJ, Brasil, e-mail: [email protected]
GOULART, MARCILIA BARBOSA Estação de Hidrobiologia e Piscicultura de Furnas, Rua Lavras 288, Usina de Furnas, 37945-000 São José da Barra, MG, Brasil
HAGE-MAGALHÃES, LILIAN RIBEIROe-mail:[email protected]
KAPUSTA, SIMONE CATERINA Instituto Federal de Educação, Ciência e Tecnologia do Rio Grande do Sul (IFECTRGS), Campus Porto Alegre, Rua Cel. Vicente, 281, 90.030-040, Porto Alegre, RS, Brasil, e-mail: [email protected]
LOPES, MICHELLE DAS NEVES Centro Universitário (FACVEST, NDE), Ciências Biológicas, Av. Marechal Floriano, 947, Centro, 88501-103 Lages, SC, Brasil, e-mail: [email protected]
MANSUR, MARIA CRISTINA DREHERUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
MARÇAL, SANDRA FRANCISCAUniversidade Federal de Mato Grosso(UFMT), Programa de Pós Graduação em Ecologia e Conservação da Biodiversidade, Av. Fernando Correa da Costa, 2367, Bairro Boa Esperança, 78060-900 Cuiabá, MT, Brasil, e-mail: [email protected]

15
MASSOLI, EDSON VIANA JUNIORUniversidade de Varzea Grande (UNIVAG), Centro Universitário, GPA Ciências Agrárias e Biológicas, Av. Dom Orlando Chaves, 2655, Cristo Rei, 78118-900Várzea Grande, MT, Brasil, e-mail: [email protected]
MELO, ADRIANO SANCHESUniversidade Federal de Goiás (UFG), Instituto de Ciências Biológicas, Campus Samambaia, Cx. postal 131, 74001-970 Goiânia, GO, Brasil, e-mail: [email protected]
MIYAHIRA, IGOR CHRISTOUniversidade do Estado do Rio de Janeiro (UERJ), Instituto de Biologia Roberto Alcântara Gomes, Laboratório de Malacologia Límnica e Terrestre, Rua São Francisco Xavier 524, PHLC 525-2, Maracanã, 20550-900 Rio de Janeiro, RJ, Brasil, e-mail: [email protected]
NEHRKE, MARINEI VILAR Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
NOGATA, YASUYUKIInstituto Central de Pesquisa da Indústria de Energia Elétrica (ICPIEE), Laboratório de Pesquisa de Ciências Ambientais 1646 Abiko, Abiko-shi, 270-1194 Chiba, Japão,e-mail: [email protected]
OLIVEIRA, ARTHUR SCHRAMM DE Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
OLIVEIRA, MARCIA DIVINA DEEmpresa Brasileira de Pesquisa Agropecuária (EMBRAPA - Pantanal), Limnologia, Rua 21 de setembro, 1880, 79320-900Corumbá, MS, Brasil, e-mail: [email protected]
OSTRENSKY, ANTONIO Universidade Federal do Paraná (UFPR), Departamento de Zootecnia, Grupo Integrado de Aquicultura e Estudos Ambientais, setor de Ciências Agrárias, Rua dos Funcionários, 1540, Juvevê, 80035-050 Curitiba, PR, Brasil, e-mail: [email protected]
PATELLA, RAQUEL Universidade Federal do Paraná (UFPR), Departamento de Zoologia, Grupo Integrado de Aquiculturae Estudos Ambientais, setor de Ciências Biológicas, Av. Francisco H. dos Santos, s/n, Jardim das Américas, Cx. postal 19073, 81531-980 Curitiba, PR, Brasil,e-mail: [email protected]
PAZ, ISABEL CRISTINA PADULAUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
PEDROZO, CATARINA DA SILVAUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
PEREIRA, DANIEL Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
PIMPÃO, DANIEL MANSUR Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA), Rua 229, n. 95, Setor Leste Universitário, 74605-090 Goiânia, GO, Brasil, e-mail: [email protected]
PINILLOS, ANA CECILIA MAGARIÑO Universidade Federal do Mato Grosso (UFMT), Instituto de Biociências, Graduanda em Ciências Biológicas, Av. Fernando Corrêa da Costa, nº 2367, 78060-900 Cuiabá, MT, Brasil, e-mail: [email protected]
PINOTTI, RAPHAEL MATHIASUniversidade Federal do Rio Grande (FURG), Instituto de Oceanografia, Laboratório de Ecologia de Invertebrados Bentônicos, Av. Itália, km 8, Campus Carreiros, Cx. postal 474, 96201-900 Rio Grande, RS, Brasil, e-mail: [email protected]
POMBO, VIVIAN BECKMinistério do Meio Ambiente (MMA), Departamento de Conservação da Biodiversidade, Esplanada dos Ministérios, Bloco B, 70068-900 Brasília, DF, Brasil, e-mail:[email protected]
POSTIGLIONE, RAONI ROCHAUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
RAYA-RODRIGUEZ, MARIA TERESAUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
RESENDE, FERNANDO ANTONIO BLANCODepartamento de Equipamento Rotativo, Furnas Centrais Elétricas - Escritório Central, Rua Real Grandeza 219, Botafogo, 22281-900 Rio de Janeiro, RJ, Brasil.
SANTOS, CINTIA PINHEIRO DOS Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]

16
SANTOS, SILVIO CARLOS ALVES DOSAES Tietê, Departamento de Meio Ambiente, Usina Hidrelétrica de Promissão Rua Olavo Bilac, 620, Cx. Postal 64, 16400-970 Lins, SP, Brasil, e-mail: [email protected]
SANTOS, SONIA BARBOSAUniversidade do Estado do Rio de Janeiro (UERJ), Instituto de Biologia Roberto Alcântara Gomes, Laboratório de Malacologia Límnica e Terrestre, Rua São Francisco Xavier 524, PHLC 525-2, Maracanã 20550-900 Rio de Janeiro, RJ, Brasil, e-mail:[email protected]
SEBASTIANY, JULIANA BRASILIENSEInstituto Biológico do Meio Ambiente (IBIOMA), Rua Conselheiro Araújo, Centro, 80060-230 Curitiba, PR, Brasil, e-mail: [email protected]
SILVA, EDSON PEREIRA DA Universidade Federal Fluminense (UFF), Instituto de Biologia, Departamento de Biologia Marinha, Laboratório de Genética Marinha e Evolução, Outeiro São João Batista, s/nº, Valonguinho, Cx. postal 100.644, 24001-970 Niterói, RJ, Brasil, e-mail: [email protected]
SOARES, MÍRIAM DE FREITASFundação Estadual de Proteção Ambiental Henrique Luiz Roessler (FEPAM), Departamento de Laboratórios, Divisão de Química, Rua Aurélio Porto, 45, 90620-090 Porto Alegre, RS, Brasil, e-mail: [email protected]
SOARES, VINÍCIUS CORREA DA COSTAUniversidade Federal do Mato Grosso (UFMT), Instituto de Biociências, Departamento de Biologia e Zoologia, Laboratório de Ecologia Animal, Avenida Fernando Corrêa da Costa, n° 2367, Boa Esperança, 78060-900 Cuiabá, MT, Brasil, mail: [email protected]
SOUZA, ANDRESSA MORAES SOFIA DE Universidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, mail: [email protected]
SOUZA, ESTEVÃO CARINO FERNANDES DEInstituto Chico Mendes de Conservação da Biodiversidade (ICMBio), Cx. postal 7993, 70670-350 Brasília, DF, Brasil, e-mail: [email protected]
TAKEDA, ALICE MICHIYOUniversidade Estadual de Maringá (UEM), DBI/PEA/NUPELIA, Av. Colombo, 5790, Bloco H-90, 87020-900 Maringá, PR, Brasil, e-mail: [email protected]
THIENGO, SILVANA CARVALHOInstituto Oswaldo Cruz (FIOCRUZ), Laboratório de Malacologia, Av. Brasil 4365 Manguinhos, 21040-900 Rio de Janeiro, RJ, Brasil, e-mail: [email protected]
TSCHÁ, MARCEL KRUCHELSKIUniversidade Federal do Paraná (UFPR), Departamento de Zoologia, Grupo Integrado de Aquicultura e Estudos Ambientais, Setor de Ciências Biológicas, Av. Francisco H. dos Santos, s/n, Jardim das Américas, Cx. postal 19073, 81531-980 Curitiba, PR, Brasil, e-mail: [email protected]
UHDE, VERAUniversidade Federal de Mato Grosso (UFMT), Instituto de Biociências, Laboratório de Ecologia Aquática, Av. Fernando Corrêa da Costa, nº 2367, Bairro Boa Esperança, 78060-900 Cuiabá, MT, Brasil, e-mail: [email protected]
VANIN, ALINE SALVADORUniversidade Federal do Rio Grande do Sul (UFRGS), Instituto de Biociências, Centro de Ecologia, Av. Bento Gonçalves, 9500, setor 4, prédio 43411, Cx. postal 15007, 91501-970 Porto Alegre, RS, Brasil, e-mail: [email protected]
VICENZI, JULIANEUniversidade Federal do Rio Grande do Sul (UFRGS), Escola de Engenharia, Departamento de Materiais, Av. Osvaldo Aranha, 99, Centro, 90035-190 Porto Alegre, RS, Brasil, e-mail: [email protected]
VIEIRA, JOÃO PAES Universidade Federal do Rio Grande (FURG), Instituto de Oceanografia, Av. Itália, km 8, Vila Carreiros, 96201-900 Rio Grande, RS, Brasil. e-mail: [email protected]
VILLANOVA, DANIELA LUPINACCI Instituto Federal de Educação, Ciência e Tecnologia do Rio Grande do Sul (IFECTRGS), Campus Farroupilha. Av. São Vicente, 785, Centro, 95180-000 Farroupilha, RS, Brasil, e-mail: [email protected]
XIMENES, RENATA FREITAS Universidade do Estado do Rio de Janeiro (UERJ), Instituto de Biologia Roberto Alcântara Gomes, Laboratório de Malacologia Límnica e Terrestre, Rua São Francisco Xavier 524, PHLC 525-2, Maracanã, 20550-900 Rio de Janeiro, RJ, Brasile-mail: [email protected]
ZURITA, MANUEL LUIZ LEITE M & Z Gestão e Processos Ambientais, Rua Prof. Cristiano Fischer 181 /802, 91410-001 Porto Alegre, RS, Brasil, e-mail: [email protected]

Seção
1Moluscos invasores nos ecossistemas
continentais do Brasil


19
Quando abordamos o tema espécies in-vasoras, inúmeros conceitos disponíveis na li-teratura científica podem ser adotados, assim como uma ampla discussão pode ser estabe-lecida. No entanto, do ponto de vista prático e considerando o público alvo ao qual ele se destina, dentre os quais cabe citar o setor elé-trico e os órgãos ambientais, faz-se necessária a apresentação de alguns conceitos seguidos no exterior e no Brasil. As espécies introduzidas ou exóticas cor-respondem àquelas presentes em um ecossiste-ma de onde ela não é originária, ou seja, nativa. Várias designações de espécies introduzidas são utilizadas: espécies não indígenas; alienígenas; não nativas; estrangeiras; exóticas; transplan-tadas e alóctones (Espinola & Ferreira Junior, 2007). Algumas espécies exóticas têm gran-de capacidade de invasão e de colonização de ambientes devido às características biológicas, genéticas, fisiológicas e ecológicas que confe-
rem tolerância à maioria dos fatores ambientais (Machado & Oliveira, 2009). Ainda segundo os mesmos autores, “ao longo das últimas dé-cadas, o processo de globalização, associado à intensificação e à velocidade do deslocamento humano e de cargas pelos quatro cantos do mundo contribuiu, sobremaneira, para a que-bra de barreiras ecológicas, tendo como uma de suas consequências o aumento expressivo da introdução de espécies exóticas nas socie-dades”. Segundo Valéry et al. (2008a) a “inva-são biológica consiste de espécies que adqui-rem uma vantagem competitiva, seguida do desaparecimento de obstáculos naturais à sua proliferação, o que permite que ela se disperse rapidamente e conquiste novas áreas, nas quais se torna uma população dominante”. Ainda cabe citar o conceito de espécies engenheiras do ecossistema, estabelecido por Jones et al. (1994). O autor define como enge-nheiros de ecossistema os organismos que dire-ta ou indiretamente modificam a disponibilida-
CAPÍTULO 1
Abordagem conceitual dos moluscos invasores nos ecossistemas límnicos brasileiros
Flavio da Costa FernandesMaria Cristina Dreher Mansur
Daniel PereiraLivia Viana de Godoy Fernandes
Sávio Calazans CamposOlga Maria Danelon

Fernandes et al.
Abordagem conceitual dos moluscos invasores nos ecossistemas límnicos brasileiros
20
de de recursos para outras espécies, causando alterações bióticas e abióticas. Dessa forma, es-pécies introduzidas com características invasi-vas, que se enquadram no conceito de espécies engenheiras do ecossistema, causam impacto ainda maior sobre o funcionamento do ecossis-tema. Espinola & Ferreira Junior (2007) apre-sentaram uma ampla discussão sobre concei-tos, modelos e atributos de espécies invaso-ras. Os autores ressaltaram a necessidade de elaborar legislações com base em conceitos claros e de aplicação direta, extremamente desejáveis para subsidiar ações estratégicas que previnam invasões biológicas. Também apontam que a Portaria do Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA) Nº 145-N, de 29 de ou-tubro de 1998, apresenta diversos termos para denominar espécies introduzidas, levando a confusões técnicas e recomendam a utilização do termo espécie não indígena, não importan-do se a espécie é proveniente de outro país ou não. Considerando as dimensões continentais do Brasil, é muito comum a introdução de es-pécies nativas com potencial invasivo, de uma bacia hidrográfica em outra na qual não ocor-ria. No entanto, sem considerar a questão le-vantada pelos autores acima, Lopes & Villac (2009) uniformizam os conceitos sobre espécie exótica e espécie invasora no livro editado e publicado pelo Ministério do Meio Ambiente intitulado “Informe sobre espécies exóticas in-vasoras Marinhas no Brasil”. Esta nomenclatu-ra é seguida por este Ministério e seus órgãos no processo de tomadas de decisões quanto a esta problemática ambiental. As categorias que compõem esta nomenclatura seguem abaixo: Categorias genéricas
Exótica: espécie registrada fora de sua área de distribuição original. Nativa: espécie que vive em sua região de origem (em contraste à espécie exótica). Criptogênica: espécie de origem biogeo-gráfica desconhecida ou incerta – este termo deve ser empregado quando não existe uma evidência clara de que a espécie seja nativa ou exótica.
Categorias específicas para espécies exóticas:
Contida: quando a presença da espécie exótica foi detectada apenas em ambientes ar-tificiais controlados, isolados total ou parcial-mente do ambiente natural (aquário comercial, cultivo para fins científicos, tanque de água de lastro de navios etc.). Detectada: quando a presença da espé-cie exótica foi detectada no ambiente natural, porém sem aumento posterior de sua abun-dância e/ou de sua dispersão (considerando o horizonte de tempo das pesquisas ou levanta-mentos a respeito); ou, alternativamente, sem que tenham sido encontradas informações sub-sequentes sobre a situação populacional da es-pécie (registro isolado). Estabelecida: quando a espécie introdu-zida foi detectada de forma recorrente, com ciclo de vida completo na natureza e indícios de aumento populacional ao longo do tempo em uma região restrita ou ampla, porém sem apresentar impactos ecológicos ou socioeconô-micos aparentes. Invasora: quando a espécie estabeleci-da possui abundância ou dispersão geográfica que interferem na capacidade de sobrevivência de outras espécies em uma ampla região geo-gráfica ou mesmo em uma área específica, ou quando a espécie estabelecida causa impactos mensuráveis em atividades socioeconômicas ou na saúde humana.
Aspectos da bioinvasão
As características biológicas, químicas e físicas do ecossistema determinam a sua vul-nerabilidade à invasão. Por exemplo, um am-biente muito degradado estará mais suscetível à invasão do que ambientes ecologicamente equilibrados. Contudo, é importante ressaltar que é baixa a probabilidade de uma espécie exótica sobreviver e, além disso, se estabelecer em um ecossistema diferente do de sua origem. Somente espécies com características invasivas conseguem romper barreiras fisiológicas e eco-lógicas, estabelecendo-se em áreas distintas da original com sucesso reprodutivo, crescimento populacional e dispersão geográfica acelerada. Também é importante considerar que a inva-

CAPÍTULO 1
Seção 1
21
são biológica apresenta diferentes fases. Uma espécie inicialmente não considerada como in-vasora, e sim simplesmente exótica, pode apre-sentar comportamento invasivo com o passar do tempo. Darrigran & Damborenea (2009a), com base em autores consagrados, sintetizam as fases de uma invasão biológica: repouso ou espera, quando uma espécie chega no ambiente e se encontra dentro da capacidade de carga do mesmo, com crescimento populacional lento; pânico, quando a população se expande de for-ma acelerada, excedendo a capacidade de carga do ambiente invadido, e freio, quando ocorre a diminuição do crescimento exponencial da fase anterior até uma capacidade de carga menor a inicial, passando para uma etapa de equilíbrio oscilatório. Porém, uma vez estabelecida, dificil-mente a invasão será reversível num ecossistema. Naturalmente, há a dispersão de espécies onde anteriormente não eram nativas. O vetor natural pode ser alguma outra espécie, como peixes ou corrente marinha, etc. A capacidade do homem de transportar por longas extensões de terra, mar ou rio trouxe o aumento da pos-sibilidade de introdução de espécies por onde ele circule. Com o advento da navegação e, mais recentemente, com a globalização, a intro-dução de organismos aquáticos em ambientes não nativos se tornou caso frequente. Os navios transportam, não intencionalmente, organismos incrustados em sua estrutura, como casco e hé-lice. Mais importante que isso, os navios trans-portam água de lastro (esta confere estabilidade ao navio), a qual representa o principal vetor de introdução de espécies exóticas. Quando o na-vio não está transportando mercadoria, ele fica leve e precisa encher seus tanques de lastro com água, para garantir flutuabilidade e navegabili-dade. Ao deslastrar para carregar a mercadoria, o navio joga água de outra localidade neste por-to. Nesta água deslastrada, estão presentes va-rias formas de vida que podem encontrar boas condições para sobreviver e se estabelecer neste novo local. Quanto maior a carga de indivíduos introduzidos e a frequência com que isto aconte-ce, maior a probabilidade da introdução e invasão serem bem sucedidas. Portanto, com o aumento do transporte marítimo, devido ao crescente in-tercâmbio comercial entre os países, torna-se maior o número de casos de espécies invasoras no Brasil e no mundo (Silva et al., 2004).
Os problemas causados por espécies in-vasoras levam, no mínimo, ao desequilíbrio ecológico, principalmente, quando não encon-tram predador local. Por exemplo, o mexilhão--zebra (Dreissena polymorpha) tem causado prejuízos imensos por incrustação nos Grandes Lagos, nos EUA; a dispersão global de cólera (Vibrio cholerae) tem causado grave problema sanitário; o ctenóforo Mnemiopsis leidy, inva-sor no Mar Negro, causou redução drástica na pescaria de enchovas e espadas; na Austrália, a estrela-do-mar Asterias amurensis causou grande prejuízo ao cultivo de marisco; o bival-ve Isognomon bicolor invadiu a região interma-real do litoral brasileiro, impedindo a fixação de muitas espécies nativas (Silva et al., 2004; Darrigran & Damborenea, 2009a). Como parar a dispersão de espécies exó-ticas pelo mundo via água de lastro e bioin-crustação dos navios? Com certeza não iremos parar de usar os navios. A engenharia procu-ra alternativas para um navio que não preci-se de lastro para garantir o equilíbrio, mas o caminho é longo. As indústrias e institutos de pesquisa buscam por material ou tinta anti-in-crustante para as estruturas dos navios sujeitas à incrustação. Pesquisam, também, por produ-tos ou processos que possam tratar a água de lastro, visando à morte do maior número de indivíduos possível, presentes ali. Entretanto, qualquer alternativa de solução deverá prevenir o surgimento de novos problemas que impossi-bilitem seu uso. Por exemplo, o tratamento não pode causar poluição do meio ambiente, nem danificar as estruturas do navio, tão pouco ser economicamente inviável. As preocupações envolvendo introdução de espécies exóticas por água de lastro vêm crescendo em todo o mundo desde a década de 90. Ainda não há legislação internacional vigente com diretrizes para o controle e geren-ciamento da água de lastro, apenas algumas re-comendações da Organização Marítima Inter-nacional (IMO). Entretanto, em alguns países, já existe uma legislação nacional para controle da água de lastro. Desde 2005, entrou em vi-gor no Brasil, a Norma da Autoridade Marí-tima 20 (NORMAM 20), a qual exige que o navio em viagem internacional, que traga água de lastro, faça troca em alto mar, ou seja, numa profundidade superior a 200 m. O método pre-

Fernandes et al.
Abordagem conceitual dos moluscos invasores nos ecossistemas límnicos brasileiros
22
ventivo de troca de água em alto mar baseia-se no fato que o meio ambiente oceânico é inós-pito a organismos de água doce, estuarinos e a maioria dos organismos de águas costeiras e vice-versa. Ou seja, uma vez que os portos fi-cam em regiões costeiras ou de rios, a troca em alto mar levará águas dos portos para o oceano e do oceano para os portos, o que minimiza muito a transferência de espécies indesejáveis. A NORMAM 20 também exige que navios em cabotagem, oriundos de portos de água doce, façam também a troca no mar antes de deslas-trarem em outro porto de água doce (Silva et al., 2004; Fernandes et al., 2009). A introdução de espécies com potencial invasivo pode ocorrer por diferentes formas. Os moluscos bivalves invasores apresentam um desenvolvimento inicial em forma de larva que passa por várias fases até chegar ao juvenil que será recrutado no ambiente bentônico, onde se desenvolverá até a fase adulta (ver Capítulo 6). Em algumas espécies, as larvas se desenvolvem no plâncton; em outras, se desenvolvem dentro do corpo da mãe, sendo liberadas, na fase final
de larva ou como juvenil, no bentos. Na fase lar-val, os organismos podem ser sugados por bom-bas para o interior de tanques destinados a con-ter a água de lastro (Fig. 1). Também podem ser sugados para o interior de cisternas, de estações de tratamento de água, canais de irrigação e siste-mas de refrigeração de indústrias e usinas gerado-ras de energia elétrica. Outra forma de transporte seria através da dispersão de indivíduos adultos de espécies incrustantes, por meio de cascos de embarcações que circulam pelas hidrovias. A seguir estão alguns potenciais vetores que causam a introdução e a dispersão de mo-luscos límnicos invasores: a) Além da água de lastro, outra via de introdução é a prática de aquariofilia. b) Embarcações de pequeno a grande porte em trânsito pelas hidrovias ou mesmo via terrestre, pois alguns bivalves incrustados po-dem sobreviver por dias expostos ao ar. c) Águas contidas em tanques e cisternas das embarcações, contaminadas com larvas e juvenis de moluscos invasores, transportadas para outra bacia hidrográfica.
Figura 1. Navio deslastrando água contaminada, enquanto é carregado com mercadorias; ciclo de vida do mexilhão-dourado: fases planctônica e bentônica (M.C.D. Mansur).

CAPÍTULO 1
Seção 1
23
d) A areia retirada de rios, contaminada com moluscos invasores, transportada para ou-tros mananciais. e) Desvio de cursos naturais de corpos hídricos (transposição de bacias) contaminados com espécies invasoras. f) A aquicultura torna-se um potencial vetor, uma vez que a água contendo os alevinos ou os peixes pode conter larvas ou até mesmo organismos adultos de moluscos invasores. g) Peixes malacófagos são potenciais ve-tores de introdução, pois há disseminação do molusco pela área de abrangência do peixe, já
que alguns mexilhões podem passar através do tubo digestório dos peixes e saírem vivos nas fezes (ver Capítulo 39). h) A água dos recipientes que transpor-tam as iscas vivas para pesca pode conter larvas dos bivalves invasores. i) A utilização de moluscos invasores como isca constitui risco de dispersão em festi-vais de pesca. j) A transposição, por via terrestre, de barcos utilizados para pesca, lazer ou inspeção do reservatório de bacias contaminadas para bacias não contaminadas.


25
1. Introdução
Os moluscos são animais invertebrados que apresentam o corpo coberto pelo manto, geralmente protegido por uma concha (à exce-ção: lesmas e polvos). Os moluscos estão divi-didos em sete subgrupos ou Classes, de acor-do com certas características como a forma da concha e do pé. No ambiente de água doce te-mos moluscos de dois grupos apenas, os bival-ves e os gastrópodes. Os bivalves, geralmen-te chamados de mariscos, como o nome diz, apresentam a concha dividida em duas valvas articuladas. Os gastrópodes ou caracóis apre-sentam uma concha geralmente helicoidal. Nos ambiente de água doce do Brasil temos espécies invasoras tanto de bivalves como de gastrópodes. É senso comum que as espécies exóticas trazem prejuízos ao meio ambiente e à economia. Todavia, outras questões relativas ao processo evolutivo (Mooney & Cleland, 2001), à biologia da conservação (Coblentz, 1990; Olden et al., 2004) e à introdução de
novos parasitos que afetam a saúde humana e animal, incluindo as relações ecológicas das comunidades invadidas (Font, 2003; Torchin et al., 2003) são ainda pouco exploradas no Brasil.
Bivalves límnicos invasores 2. O mexilhão-dourado Limnoperna fortunei (Dunker, 1857)
2.1. Características da espécie
O mexilhão-dourado, L. fortunei (Fig. 1), é um bivalve pequeno (entre 2 e 3 cm) com características típicas da família dos mexilhões marinhos (Mytilidae), como: formato da con-cha mitiloide, modo de vida gregário, forman-do incrustações sobre substratos duros (Fig. 1B), onde se fixa com auxílio de fios de bisso. No entanto, é o único que vive na água doce (ver morfologia no Capítulo 4).
CAPÍTULO 2
Espécies de moluscos límnicos invasores
no Brasil
Sonia Barbosa dos SantosSilvana Carvalho Thiengo
Monica Ammon FernandezIgor Christo Miyahira
Isabela Cristina Brito GonçalvesRenata de Freitas Ximenes
Maria Cristina Dreher MansurDaniel Pereira

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
26
As incrustações formadas pelo mexi-lhão-dourado são volumosas. Vários indivídu-os que se sobrepõem fixando-se ao substrato e entre si, pelos fios de bisso. Os menores preen-chem os espaços entre os maiores, formando assim verdadeiros macroaglomerados compac-tos e resistentes à força da correnteza. Desenvolve-se preferencialmente em am-bientes lóticos e bem oxigenados; apresenta um comportamento invasivo (Morton, 1973, 1977; Darrigran & Damborenea, 2006a), e é transformador do meio ambiente, pois altera a composição do bentos e a paisagem ribeirinha (Mansur et al., 2004a, b).
2.2. Etimologia
O nome comum “dourado” deriva de sua cor predominantemente amarelada. O nome científico do gênero Limnoperna é compos-
to pelas palavras gregas que significam água doce para “Limno” e mexilhão para “perna”. O nome específico fortunei provavelmente re-fere-se à cor dourada da concha ou sua elevada abundância.
2.3. Impactos ambientais
O mexilhão-dourado (L. fortunei) e o mexilhão-zebra Dreissena polymorpha (Pallas, 1771) compartilham uma série de caracterís-ticas próprias das espécies invasivas e seme-lhanças quanto: ao tamanho; dispersão através de larva planctônica; serem dioicos, com ca-pacidade de adesão ao substrato duro através da formação de fios de bisso bastante fortes e resistentes; comportamento gregário com crescimento rápido alcançando altas densida-
Figura 1. Limnoperna fortunei (Dunker, 1857): 1A, vista externa do mexilhão-dou-rado (valvas esquerdas); 1B, macroaglomerados do mexilhão-dourado sobre tronco de madeira retira-dos do fundo do Lago Gua-íba, Porto Alegre, RS, Brasil (Fotos: M.C.D. Mansur).
des populacionais e a capacidade de causarem impactos ambientais consideráveis. Karatayev et al. (1997) observaram que D. polymorpha, após invadir um ambiente, torna-se o único in-vertebrado bentônico dominante, superando a biomassa dos outros componentes da comuni-dade. Martin & Darrigran (1994), Darrigran et al. (1998a), Darrigran (2002) e Darrigran & Damborenea (2005) descrevem exemplos do impacto ambiental negativo observado no ecossistema após o assentamento do L. fortunei nas margens da bacia do Rio de La Plata. Des-tacam a alteração da composição da malaco-fauna nativa, com o deslocamento das três es-pécies de gastrópodes mais comuns no litoral, que passam a ter uma ocorrência esporádica ou acidental com a presença do mexilhão-doura-do, enquanto outra nativa passa a aumentar sua população proporcionalmente à do invasor. Desde os primeiros registros da pre-
sença do mexilhão-dourado no Lago Guaíba (Mansur et al., 1999), coletas quantitativas revelaram que L. fortunei alterou a vegetação marginal (Mansur et al., 2003). No lago pre-dominam grandes sacos (baías) vegetadas por juncos da espécie Scirpus californicus (C.A. Mey.) Steud. Os substratos rochosos limitam--se aos pontais e algumas ilhas, portanto, são relativamente escassos. O mexilhão fixou-se inicialmente na base dos juncos mais afastados da margem, cujos rizomas e raízes de cabeleiras (Fig. 2A) ficam expostos devido à remoção do sedimento pelas ondas. A seguir, o mexilhão se aglomerou ocupando grandes superfícies so-bre os rizomas dos juncos (Fig. 2B), formando verdadeiros colchões (“mussel beds”) sobre o sedimento. Depois de um tempo, observou-se que os juncos apodreceram, restando apenas os

CAPÍTULO 2
Seção 1
27
aglomerados do mexilhão. A diminuição dos juncais foi acompanhada e documentada por fotos (Figs. 2C e 2D). Além dos substratos duros, como tron-cos e pedras, L. fortunei também se fixa so-bre vários animais bentônicos como bivalves, gastrópodes e crustáceos, prejudicando-os ou levando-os à morte precoce, interferindo des-te modo na biodiversidade bentônica. No caso dos bivalves, o mexilhão prefere fixar-se à re-gião posterior, que fica exposta (Figs. 2E e 2F). O aglomerado impede o movimento das valvas e também dificulta os processos de inalação, filtração, excreção e locomoção, levando o mo-lusco à morte. Aglomerados em gastrópodes podem impedir o fechamento do opérculo (Fig. 2G), tornando o molusco vulnerável à preda-ção. Houve um registro da fixação do L. for-tunei nas partes moles do bivalve nativo Leila blainvilliana Lea, 1834, no interior da área do sifão exalante (Mansur et al., 2003). No verão, o nível da água diminuiu, ex-
pondo os aglomerados, que apodrecem, exa-lando cheiro repugnante. Após a rarefação dos juncais, o mexilhão fixou-se em outras plantas marginais como o “sarandi” Cephalanthus gla-bratus (Spreng.) K. Schum que, devido ao peso dos aglomerados, acaba tombando e quebran-do (Fig. 2H). Conchas do mexilhão-dourado roladas e partidas, em grandes quantidades, modificam a paisagem (Fig. 2I).
2.4. Impactos econômicos
As aglomerações formadas pelo mexi-lhão-dourado causam a obstrução de encana-mentos, redução do diâmetro de tubulações, entupimentos de filtros, bombas, grades e trocadores de calor, principalmente em usinas geradoras de energia (Figs. 3A, 3B, 3C), em indústrias que utilizam água bruta para refri-geração e em estações de tratamento da água bruta para abastecimento (Figs. 3D, 3E, 3F). A parada destas unidades para a retirada dos
Figura 2. Danos ambientais cau-sados pelo Limnoperna fortunei no Lago Guaíba, RS, Brasil: 2A, assentamento sobre o rizoma do “junco” Scirpus californicus; 2B, “mussel beds” sobre Scirpus californicus no segundo ano da invasão; 2C, juncais em 2001, antes da invasão; 2D, modifica-ção da paisagem, após 2 anos de assentamento nos rizomas; 2E, aglomerado sufocando o bivalve nativo Diplodon sp.; 2F, aglome-rado sufocando o bivalve nativo Leila blainvilliana; 2G, no umbí-lico do gastrópode Pomacea ca-naliculata (Lamarck, 1822); 2H, aglomerados sobre ramos do “sarandi” Cephalanthus glabra-tus; 2I, conchas roladas, modifi-cando a paisagem das praias na orla (Fotos: M.C.D. Mansur).

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
28
aglomerados e consequente desentupimento gera despesas consideráveis. Em apenas um dia de limpeza são computadas perdas econômicas na ordem de milhões de reais. No caso do entupimento de uma estação de tratamento de água pelo mexilhão-dourado, são necessárias várias etapas de limpeza, con-siderando que cada estrutura ou subsistema exige um procedimento diferente. Exempli-ficamos como segue: a) parada do sistema, b) abertura da casa de captação para a retirada de uma bomba e a grade protetora, com um guindaste (Figs. 3E, 3F); c) raspagem de cada
estrutura para a retirada dos mexilhões e dos fios do bisso (Fig. 3D); d) lixa das superfícies para receber uma nova pintura; e) pintura; f) desobstrução dos encanamentos com maior diâmetro com ajuda de mergulhadores; g) ras-pagem das paredes internas dos encanamentos (em algumas estações foi construída uma rede secundária para revezamento nas limpezas); h) retirada dos mexilhões das paredes e da super-fície nos tanques de floculação, decantação e filtragem; i) montagem da unidade do sistema; j) tratamento especial da água para recuperar a potabilidade e o sabor da água, após este tra-tamento em função da presença do mexilhão--dourado. Tubulações menores devem ter cai-xas de inspeção. No caso de filtros e trocadores de calor, estes devem ser desmontados e raspa-dos. Antes de qualquer medida, as estruturas devem ser limpas. Colocar somente substâncias moluscicidas não resolve, pois os aglomerados continuam fixos pelos fios de bisso, por muito tempo. Prejuízos também foram sentidos em marinas e na navegação interna (Figs. 3G, 3H). Em embarcações o mexilhão se incrusta não somente no casco, mas também no in-terior dos motores, dos encanamentos, das bombas, dos sistemas de refrigeração com água bruta, nos lemes e nas hélices (Mansur et al., 2003), o que danifica as peças e di-minui o desempenho e a velocidade da nave. Em tanques-rede, muito utilizados atualmen-te na piscicultura, o mexilhão adere às telas e demais estruturas de suporte e flutuação dos mesmos. A obstrução dessas estruturas altera a oxigenação do tanque e o peso dos aglome-rados pode afundá-lo, facilitando a fuga dos alevinos (Fig. 4). Quando o mexilhão se incrusta sobre madeira, ferro ou concreto, pode fechar toda a superfície criando um ambiente anóxico sob a camada incrustante. Este ambiente é propício às bactérias anaeróbicas que produzem ácido sulfúrico durante seu metabolismo. Este ácido, muitíssimo corrosivo, acelera o apodrecimento e a corrosão do material da base. O mau cheiro e as conchas quebradas roladas nas margens durante o período de es-tiagem, afastam os turistas que normalmente frequentam os balneários e marinas nas orlas de veraneio.
Figura 3. Danos econômicos causados pelo Limnoperna fortu-nei: 3A, filtro autolimpante obstruído; 3B, detalhe da tampa; 3C, cilindros do interior do mesmo filtro; 3D, início do proces-so de limpeza de uma bomba captadora de água incrustada; 3E, bomba içada com guindaste pelo teto; 3F, grade protetora da bomba; 3G, trapiche de marina; 3H, casco de embarcação utilizada para pesca no Lago Guaíba e Lagoa dos Patos, sendo transportada por rodovia para outro manancial, oferecendo risco de contaminação. Fontes: A, B, C Gustavo Darrigran, www.malacologia.com.ar; D, E, F José Imada, CORSAN; G, Augusto Chagas: Biociências, 2004, 13(1); H, M.C.D. Mansur: Revista Brasileira de Zoologia 2003, 20(1).

CAPÍTULO 2
Seção 1
29
2.5. Danos à saúde
Por serem ativos filtradores, os molus-cos bivalves em geral, costumam atuar como bioacumuladores de uma série de substâncias tóxicas, inclusive metais pesados, em seus tecidos e na concha. Como os bivalves são muito apreciados pelos peixes, existe risco sanitário de bioacumulação de metais nos humanos que consomem peixes e em sua ali-mentação. Grande parte dos moluscos bivalves dul-cícolas hospeda trematódeos que completam seu ciclo em peixes, aves e mamíferos e, mui-to raramente, no homem. Estes parasitos são geralmente danosos aos moluscos. Instalam-se nas gônadas provocando a castração do ani-mal. Hiroko et al. (2004) estudaram o efeito de um trematódeo sobre o crescimento e a repro-dução do mexilhão-dourado no Japão e cons-tataram que a produção de gametas foi inibida pelo parasita. No entanto, até o momento, não foram registrados trematódeos parasitos do mexilhão-dourado, que completem seu ciclo no ser humano. Em moluscos estuarinos como Mytella guyanensis (Lamarck, 1819) (Bivalvia: Myti-lidae) e nos cultivos de mexilhões marinhos foram encontrados sítios de infecção por pro-tozoários do gênero Nematopsis Schneider, 1892 (Apicomplexa: Eugregarinida: Porospo-ridae) em várias áreas da costa brasileira do norte, nordeste e do sul, até Santa Catarina (Matos et al., 2001; Pinto & Boehs, 2008). No entanto, nada ainda foi registrado para o mexilhão-dourado.
2.6. Origem e dispersão.
L. fortunei é nativa do sudeste asiático, vi-vendo nos ambientes de água doce como, lagos, rios e regiões estuarinas, com baixa salinida-de (<13‰), da Coréia, China, Laos, Camboja, Vietnã, Indonésia e Tailândia (Ricciardi, 1998). Matsui et al. (2002), com base nos trabalho de Nishimura & Habe (1987), mencionam que essa espécie provavelmente chegou ao Japão em 1987, aderida às valvas de corbículas asiáticas que im-portam da China, como alimento. Foi introduzi-da em Taiwan (2000) e na América do Sul (1991), provavelmente via água de lastro (Pastorino et al., 1993; Darrigran & Pastorino, 1995).
2.7. Distribuição Geográfica na América do Sul
Na América do Sul o mexilhão-dourado foi primeiramente registrado no Rio de La Plata (Pastorino et al., 1993; Darrigran & Pastorino, 1995) e, subindo o Rio Paraná, dispersou-se em direção norte, alcançando em 10 anos, cinco pa-íses, na seguinte sequência: Argentina (em 1991), Uruguai (em 1994), Paraguai (em 1997), Brasil e Bolívia (em 1998) (Darrigran & Mansur, 2006, 2009). Estes autores (op. cit.) apresentam deta-lhada cronologia da invasão do mexilhão-dou-rado na América do Sul. Assim, neste capítulo, apresentamos uma síntese, com atualização dos registros a partir do ano de 2006 e, na Figura 5, um mapa com a distribuição atual de L. for-tunei, os primeiros registros, locais com danos econômicos e registros de exemplares, porém, em populações não consolidadas. No Rio Paraguai, o ponto mais à mon-tante do registro de ocorrência do mexilhão--dourado é ainda a cidade de Cáceres, onde foi encontrado em cisterna de embarcação num estaleiro, porém, a população ainda não se es-tabeleceu no meio ambiente. Registros consoli-dados referem a espécie para a baía da Gaíva e no curso inferior do Rio Cuiabá. Em 2001, foi registrado para o curso in-ferior do Rio Uruguai e na hidrelétrica de Salto Grande (Darrigran & Mansur, 2006, 2009). Em 2007, na foz do Rio Quaraí, na fronteira com o Brasil (Correio do Povo, 31/1/2007a). Existe ainda um registro para o Arroio Touro Passo (Correio do Povo, 13/03/2007b), nas
Figura 4. Rede de um tanque para a criação de esturjão no Rio Uruguai. Fonte: Gustavo Darrigran, www.malacologia.com.ar

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
30
proximidades da cidade de Uruguaiana, no Rio Grande do Sul, porém não soubemos da existên-cia de exemplares-testemunho para comprovar a ocorrência deste registro e do anterior, em Quaraí. Considerando o alto Rio Paraná, existem registros para a represa de Jaciretá (1998) e Itaipu (2001), e várias outras nos afluentes como o Igua-çu (2003), Paranapanema (2002), Tietê (2004) e no Paranaíba, até a jusante da represa de São Si-mão, onde permanece sem avançar, desde 2004 (Darrigran & Mansur, 2006, 2009). Registros recentes (novembro de 2011) para os reservató-rios de Porto Colômbia, Mascarenhas de Moraes e Marimbondo, marcam a presença da espécie em grande parte do Rio Grande, fronteira do es-tado de São Paulo com Minas Gerais. Uma invasão secundária, originária, pro-vavelmente, da água de lastro de navios argenti-nos, trouxe o mexilhão-dourado até o porto de Porto Alegre, junto ao Lago Guaíba, onde foi detectado pela primeira vez em 1998. Este lago integra as bacias do Atlântico Sul e Sudeste, sem comunicação direta com as bacias do Paraná ou Uruguai, no Rio Grande do Sul (Mansur et al., 2004a). Desse lago o mexilhão-dourado se dis-persou à montante da bacia do Rio Jacuí, e ao extremo sul do Estado, em direção à República Oriental do Uruguai, pelo lado Leste, onde hoje ocupa várias sub-bacias. Levantamentos efetu-ados no período de 2005 a 2008 na sub-bacia do Rio Jacuí, pertencente à bacia do Atlântico Sul e Sudeste, permitiram verificar a distribui-ção do mexilhão-dourado à montante deste rio até a desembocadura do Rio Pardo e no porto graneleiro de Estrela. Em novembro de 2009, foi registrada a presença do mexilhão-dourado na barragem Capingui, município de Marau, no alto Jacuí. Uma nova bacia, a das lagoas da Pla-nície Costeira do Rio Grande do Sul, situada a Nordeste do sistema Guaíba – Patos – Mirim, foi contaminada em 2009 (Freitas et al., 2009).
3. O berbigão asiático Corbicula fluminea (Müller, 1774) 3.1. Características da espécie
C. fluminea (Fig. 6A) é semelhante a um berbigão marinho, porém habita somente am-bientes de água doce. A concha é robusta com
um comprimento que varia entre 2 e 6 cm, tendo altura menor que o comprimento. O for-mato da concha é cordiforme em vista frontal. Possui superfície externa castanho escura, com algum brilho, ondulações comarginais espaça-das (1 mm cada) e posteriormente um prolon-gamento denominado “rostro” (ver morfologia no Capítulo 4). Produz fios de bisso diferentes do produzido pelo mexilhão-dourado. Nesta espécie o bisso se constitui num cordão muci-laginoso elástico presente apenas na fase de re-crutamento. Acima de 5 mm de comprimento, não produz mais o cordão mucoso. Este fio ou cordão mucoso auxilia na locomoção do ani-mal, aglutinando areia e evitando o arraste do molusco pela correnteza. Não forma aglomera-dos como o mexilhão-dourado; vive livremente no sedimento onde costuma enterrar-se parcial ou totalmente. Seu comportamento é gregário, formando densas populações (Fig. 6B). Apre-senta estratégias de sobrevivência e reprodução surpreendentes. Geralmente é considerada her-mafrodita, porém estudos recentes revelaram
Figura 5. Distribuição atual do mexilhão-dourado destacando a invasão cronológica e áreas com danos econômicos. Mapa: L. Lucatelli e M.C.D. Mansur.

CAPÍTULO 2
Seção 1
31
aspectos pouco comuns sobre a biologia re-produtiva da espécie como, poliploidia, esper-matozoide biflagelado e diploide, clonagem e androgênese (Komaru & Konishi, 1996, 1999; Komaru et al., 1997; Qiu et al., 2001). Assim como o mexilhão-dourado, é uma espécie com características invasivas e trans-formadoras do meio ambiente. Foi considerada praga nos Estados Unidos onde à espécie causou imensos problemas ambientais e econômicos. O comportamento é infaunal, ou seja, afunda--se no substrato, onde vive total ou semienter-rada. Prefere ambientes lóticos. Em ambientes lênticos ocupa áreas marginais bem oxigenadas, com declive suave e substrato preferencialmente arenoso. Segundo Ituarte (1994), grandes popu-lações de C. fluminea e C. largillierti (Philippi, 1844) conviviam ao longo de extensas superfí-cies arenosas na porção intertidal do estuário do Rio de La Plata. No entanto, segundo Darrigran (1991), as espécies competem entre si e depois de alguns anos, só se encontra C. fluminea.
3.2. Etimologia
O nome científico do gênero Corbicula e da família Corbiculidae tem sua origem na pa-Corbiculidae tem sua origem na pa-lavra latina corbis que significa cesto. A forma arredondada e inflada da concha, a cor de pa-lha e a ornamentação ondulada lembram uma pequena cestinha ou bolsinha. A terminação cola também de origem latina, se refere à produ-ção ou formação de algo, portanto, corbi + cola seria aquela ou aquele que produz a cestinha.
O nome popular só é citado nas enciclo-pédias portuguesas como amêijoa asiática. Em algumas regiões do Brasil é conhecida como berbigão asiático. Em meios acadêmicos, sim-plesmente por corbícula asiática ou simples-mente corbícula.
3.3. Impactos ambientais
Após a invasão de C. fluminea em várias bacias hidrográficas brasileiras, houve uma diminuição drástica das populações nativas de moluscos bentônicos, principalmente dos bivalves das famílias Mycetopodidae e Hyrii-dae. Este lamentável fato foi observado e re-latado após vários anos de trabalho de campo por Takeda et al. (2000) no alto Rio Paraná e por Beasley et al. (2003) no Pará. Em todas as bacias ocupadas por esta espécie, C. fluminea passou a apresentar, em poucos anos, densi-dades populacionais bem maiores do que as espécies nativas. Isom (1986) considera que densidades de C. fluminea acima de 200 in-divíduos por metro quadrado constituem um potencial para severos problemas ambientais e econômicos. Mansur & Garces (1988) en-contraram densidades de até 5295 ind/m2 em canal da região do Taim, no extremo sul do Rio Grande do Sul, em um canal que drena um açude junto ao curso inferior do Rio Caí no Rio Grande do Sul. Mansur et al. (1994) observaram uma população de 4173 ind/m2, com exemplares maiores que 7,7 mm de com-primento, aglomerando-se em até três níveis
Figura 6. Corbicula fluminea (Müller, 1774) o berbigão asiático: 6A, vista interna e externa das valvas de quatro exemplares em diferentes tamanhos (Foto: M.C.D. Mansur), escala 1 cm; 6B, concheiro natural indicando a alta densidade da espécie nas margens do Lago Guaíba em janeiro 2012 (Foto: G. Figueiredo), escala 10 cm.

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
32
de profundidade no substrato. Segundo Isom (1986), citando Prokopovich (1969), as corbí-culas, durante o processo de filtração, conso-lidam uma grande quantidade de partículas em suspensão, envolvendo-as em muco que é rejeitado como pseudofezes e sedimentado no fundo. Estas, junto com o acúmulo de conchas, alteram profundamente o ambiente bentônico. Sousa et al. (2008a, b), ainda mencionam que esta espécie, devido à grande abundância, pode interferir significativamente na reciclagem de nutrientes e na interação bêntico-pelágica e deste modo ser potencialmente responsáveis por importantes alterações ecológicas. Sousa et al. (2009) alertam, também, que seus efeitos modificadores do ambiente se equiparam ao de um invasor engenheiro de ecossistema e que esta espécie deveria ser considerada com muita seriedade nas iniciativas de conservação, mane-jo e restauração do ambiente.
3.4. Impactos econômicos
A espécie pode causar obstrução de en-canamentos, canais, sistemas de refrigeração de indústrias, usinas atômicas (nos EUA) e de hidrelétricas (Fig. 7). A entrada dos exempla-res nos sistemas dá-se na fase larval final como pedivéliger ou juvenil que, devido às pequenas dimensões e leveza, são sugados para dentro do sistema, passando pelos filtros protetores. Geralmente crescem dentro dos trocadores de calor, obstruindo as aberturas e provocando, consequentemente, um superaquecimento do sistema. Paralisação de usinas hidrelétricas e sistemas de abastecimento urbano de água por esta espécie, já foram registradas no Brasil. Na América do Norte, onde C. flumi-nea foi introduzida em torno do ano de 1922 (Counts, 1986), passou a despertar grande interesse devido aos problemas ambientais e econômicos ocasionados. Foram realizados vários simpósios, com publicação de inúmeros trabalhos. Calculam-se investimentos acima de um bilhão de dólares americanos anuais para reduzir a eficiência da espécie. Capital este, in-vestido em equipamentos, pesquisa, montagem de laboratórios e planos de controle da espécie (Isom, 1986). É importante ainda mencionar que a areia utilizada na construção, uma vez
contendo corbículas, forma um concreto de péssima qualidade. As conchas se descalcificam rapidamente, favorecendo infiltrações de água que danificam a estrutura, provocando racha-duras. As conchas fechadas contém ar em seu interior que pode migrar para a superfície du-rante a concretagem, fragilizando a estrutura (Isom, 1986).
3.5. Danos à saúde
Segundo Bendati (2000), o corbiculídeo nativo Neocorbicula limosa, por ser um filtra-dor, como os demais representantes da famí-lia Corbiculidae, atua como bioacumulador inclusive de metais pesados que podem entrar na cadeia alimentar, o que pode trazer alto risco para saúde humana. Os moluscos cons-tituem alimento para os peixes carnívoros e omnívoros. Cataldo et al. (2001) utilizaram C. fluminea como biomarcador de gradientes de poluição e constataram nos tecidos da mesma, o acúmulo significativo de cobre e cádmio em áreas mais poluídas do delta do Rio Paraná, Argentina. As corbículas também podem ser hospedeiros intermediários de trematódeos. Chung, et al. (2001) constataram em laborató-
Figura 7. Registro da obstrução pelo berbigão asiático Cor-bicula fluminea (Müller, 1774), na UHE Porto Colômbia, bacia do Rio Grande, alto Rio Paraná, MG, em 1998. 7A, em enca-namentos; 7B, em trocadores de calor (Foto: E.T Monteiro da Silva - FURNAS).

CAPÍTULO 2
Seção 1
33
rio a possibilidade de C. fluminea ser um se-gundo hospedeiro de Echinostoma cinetorchis Ando & Ozaki, 1923 (Trematoda: Echinos-tomatidae). Mamíferos carnívoros, roedores e inclusive o homem, além de aves aquáticas, ser-vem de hospedeiros definitivos desta espécie. Os hospedeiros definitivos tornam-se infecta-dos ao ingerirem os hospedeiros secundários do trematódeo, ainda cru ou mal cozido. O trematódeo adulto se instala no intestino del-gado podendo causar fortes dores abdominais, náusea, vômitos, diarreia e febre.
3.6. Origem e dispersão
É originária da Ásia, Coréia e sudeste da Rússia. Segundo Sousa et al. (2008a, b), é con-et al. (2008a, b), é con-siderada como a espécie invasora “NIS” (“Nui-sance Invasive Species”) de maior importância nos ecossistemas aquáticos. Nos últimos 80 anos, foi introduzida nas Américas, na África e na Europa, provavelmente via água de lastro, com registros para ilhas do Pacífico (Araujo et al., 1993). Invadiu os Estados Unidos (na dé-cada de 1920), primeiramente na parte oeste e em poucos anos ocupou todos os ambientes de água doce daquele país (Counts, 1986). Na América do Sul, os primeiros registros datam da década de 70, hoje ocupa as bacias sul-ame-ricanas, desde a Colômbia até o norte da Pata-gônia (Mansur et al., 2011a, 2012).
3.7. Distribuição geográfica
Apareceu na América do Sul na década de 70, nas proximidades do porto de Buenos Aires, Argentina (Ituarte, 1981), e em Porto Alegre, no sul do Brasil (Veitenheimer-Men-des, 1981). Em torno de 1985, Martinez (1987) identificou a corbícula como C. manilensis (Philippi, 1844), um sinônimo de C. fluminea, na Venezuela. Em 1994, no limite norte do Peru com o Equador e em 2000, em rios que correm para o Pacífico neste país (Mansur et al., 2004c). Desde o registro destes primeiros focos de invasão, a espécie vem se dispersando à montante e por todas as bacias do continente (Fig. 8). No Rio Uruguai, foi registrada para o curso inferior em 1986, para o curso médio em 1988/9 e para o superior nas nascentes junto ao Rio Pelotas, em 1996 (Rodrigues et al., 1998a,
b). Callil & Mansur (2002) registram a chega-da de C. fluminea no Pantanal Norte, próximo de Cuiabá em 1998. Um ano antes, Cazzani-ga (1997) relata sobre a chegada da espécie na Patagônia, Argentina. No alto Rio Paraná, em 1997 (Pereira, 1997). Na bacia Amazônica foi registrada desde 1997/8 para o baixo Rio To-cantins (Beasley et al., 2003); nas nascentes do Tocantins, em 1999 (Thiengo et al., 2005); no alto Rio Solimões (Lee et al., 2005); no Rio Negro, Amazônia Central, próximo de Ma-naus (Pimpão & Martins, 2008; Pimpão et al., 2008).
4. O berbigão asiático roxo Corbicula largillierti (Philippi, 1844)
4.1. Características da espécie Corbicula largillierti (Fig. 9) apresenta a concha mais frágil, menor e menos inflada que C. fluminea (não ultrapassa 2,5 cm de compri-mento), formato triangular, quase equilateral, altura menor que o comprimento, umbos bai-xos e arredondados, sem rostro, ondulações
Figura 8. Distribuição atual do berbigão asiático Corbicula flu-minea (Müller, 1774) na América do Sul. Mapa: G. Figueiredo e M.C.D. Mansur.

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
34
ou estrias comarginais na superfície externa, muito finas e justapostas (2 a 3 por mm, na parte mediana da concha), perióstraco de cor esverdeada, palha ou arroxeada, pouco brilho. Por dentro é roxo escuro ou cinza, sem brilho, linha palial evidente e sem sinus (ver morfo-logia no Capítulo 4). Facilmente confundida com a nativa Cyanocyclas (=Neocorbicula) li-mosa, quando vista externamente (Mansur et al., 2012) e com C. fluminea, principalmente quando esta não atingiu seu desenvolvimen-to completo. Produz fios de bisso diferentes do produzido pelo mexilhão-dourado. Nesta espécie o bisso se constitui num cordão muci-laginoso elástico, presente apenas na fase de recrutamento. Este fio, também chamado de cordão mucoso, auxilia na locomoção do ani-mal, aglutinando areia e evitando o arraste do molusco pela correnteza. Não forma aglomera-
dos como o mexilhão-dourado, vive livremente no sedimento onde costuma enterrar-se parcial ou totalmente. Seu comportamento também é invasivo, vive de forma gregária, formando densas populações. No entanto, quando com-partilha o mesmo ambiente que C. fluminea, com o passar do tempo sua população diminui gradativamente e pode desaparecer (Darrigran, 1991). Apresenta o comportamento infaunal.Habita ambientes bentônicos de água doce tan-to lóticos como lênticos, dando preferência por águas mais oxigenadas, declive suave do terre-no, substrato macio com a presença de areia fina. Geralmente é mais abundante em açudes e reservatórios do que C. fluminea.
4.2. Etimologia
Como a espécie acima o nome da família Corbiculidae e do gênero Corbicula vem dos termos latinos corbis e cola de origem latina, que se referem ao produtor de uma cesta. A palavra largillierti foi dada em homenagem ao colecionador Largilliert. É um Corbiculidae com aspecto de um berbigão de água doce de coloração mais es-verdeada por fora e roxo por dentro, razão do nome berbigão roxo. Em meios acadêmicos a espécie é conhecida por corbícula asiática roxa ou simplesmente corbícula roxa.
4.3. Impactos ambientais
Em grandes densidades C. largillierti provoca, assim como C. fluminea, alterações no sedimento dos mananciais e causa a dimi-nuição drástica das populações da fauna nati-va de moluscos bentônicos, principalmente do Corbiculidae nativo, Cyanocyclas (=Neocorbi-cula) limosa. Em todas as bacias ocupadas por C. largillierti, esta passou a apresentar, em pou-cos anos, densidades populacionais bem maio-res do que das espécies nativas.
4.4. Impactos econômicos
A espécie tem causado os mesmos pro-blemas de obstruções em sistemas de resfria-mento de termo e hidrelétricas (Fig. 9B), e de abastecimento de água, exigindo a parada des-tes sistemas e mão de obra especializada para
Figura 9. Corbicula largillierti (Philippi, 1884) o berbigão asi-ático roxo: 9A, Acima, vista externa da valva esquerda; em baixo, vista interna da valva direita (Foto: M.C.D. Mansur); 9B, Registro da obstrução dos trocadores de calor de uma unida-de hidroelétrica (Foto: W. Santiago).

CAPÍTULO 2
Seção 1
35
a limpeza, com consequentes perdas econômi-cas. Como C. fluminea, a entrada dos exem-plares nos sistemas dá-se na fase de véliger tardio ou pedivéliger que, devido às pequenas dimensões, passam pelos filtros protetores. Ge-ralmente crescem dentro dos trocadores de ca-lor, obstruindo as aberturas, provocando supe-raquecimento do sistema. Deve-se considerar que a areia contendo corbículas roxas, também fragiliza a concretagem.
4.5. Danos à saúde
As espécies do gênero Corbicula são bio-bio-acumuladoras de metais pesados, que podem entrar na cadeia alimentar através de peixes carnívoros e onívoros que, por sua vez, servem de alimento para populações humanas, o que implicaria em riscos para saúde. Não se conhe-cem trematódeos que se hospedem nesta espé-cie, nem da possibilidade de ser intermediário de alguma zoonose.
4.6. Lugar de origem e dispersão
Espécie de origem asiática, originalmen-te descrita do Rio Yangtse-Kiang da China. Sua distribuição compreende a China central e nor-te, assim como a península Coreana (Ituarte, 1994). Foi introduzida na América do Sul e no Brasil, provavelmente via água de lastro.
4.7. Distribuição Geográfica
C. largillierti encontra-se introduzida na América do Sul (Ituarte, 1981, 1982, 1984a e 1994; Olazarri, 1986; Mansur et al., 2004c, 2012; Callil & Mansur, 2002). Por guardar certas semelhanças com a Corbiculidae nativa, Cyanocyclas (=Neocorbicula) limosa, a inva-sora passou muito tempo despercebida no iní-cio de sua dispersão na América do Sul. Foi, e ainda hoje tem sido, também confundida com C. fluminea. Fatos estes, dificultam sobrema-neira o mapeamento de sua distribuição. Hoje ocupa duas grandes áreas na América do Sul (Fig. 10): o Nordeste e Leste do Brasil, desde o Ceará até o Rio de Janeiro e, mais ao Sul, a bacia do baixo e médio Paraná e o Rio Para-guai; o Rio Uruguai e o sistema das bacias da Laguna dos Patos e Mirim que drenam para o
Atlântico. Não foi ainda registrada para o alto Rio Paraná. Mais recentemente foi detectada pela primeira vez no Rio Tocantins, inician-do assim sua dispersão pela bacia Amazônica (Figueiredo et al., 2011; Mansur et al., 2011a, 2012).
5. O berbigão asiático Corbicula fluminalis (Müller, 1774)
5.1. Características da espécie
Corbicula fluminalis (Müller, 1774) (Fig. 11) apresenta concha robusta quase equilateral, inflada, altura igual ou maior que o compri-mento que varia de 1 a 3 cm, linhas comar-ginais semelhantes às de C. fluminea, porém menos espaçadas (uma a cada 0,7 mm) e bai-xas, charneira muito arqueada, umbos muito altos e praticamente centrados (quase na me-tade do comprimento), sem rostro, cor mais escura, tendendo ao roxo forte internamente e o perióstraco, do marrom ou negro. Conchas pequenas geralmente mais claras (detalhes da morfologia, ver no Capítulo 4).
Figura 10. Distribuição atual do berbigão asiático roxo Cor-bicula largillierti (Philippi, 1884) na América do Sul. Mapa: G. Figueiredo e M.C.D. Mansur.

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
36
Geralmente são encontradas em margens rasas, arenosas e nas proximidades de juncais. Mar-tins et al. (2004) revelaram que de 317 exem-plares do gênero Corbicula coletados entre ju-nho de 2002 a junho de 2003 no Lago Guaíba, RS, 185 corresponderam a C. fluminea, 98 a C. fluminalis e 34 a C. largillierti. Não foram documentados impactos am-bientais advindos da presença desta espécie. Até o momento observou-se que na presença de C. fluminea, as populações de C. fluminalis diminuem gradativamente em número. Porém, por ser uma invasora e ainda pouco conheci-da, não sabemos das suas qualidades invasi-vas nem do seu período de “repouso” no novo ambiente, pois se trata de uma introdução re-lativamente recente. Mansur et al. (2004c) a coletou em 1990 no baixo Rio Jacuí, RS e a identificou como Corbicula sp “b”. Martins et al. (2004) registram C. aff. fluminalis para o Lago Guaíba, Viamão, RS, com as datas de coleta no período de junho de 2002 a junho de 2003. Segundo Darrigran & Damborenea (2009b) o crescimento populacional de uma espécie invasora passa por uma série de fases. Quando a espécie invasora chega num novo ambiente, seu crescimento inicial é lento, com-preendendo a fase do “repouso” ou “espera”. Este período varia para cada espécie e pode até levar muitos anos. A seguir, na segunda fase, denominada de “pânico”, a população cresce e se expande rapidamente. Na terceira fase acon-tece um “freio” e uma redução no crescimento populacional com um declínio na densidade. É a fase do “equilíbrio oscilatório”. No entanto, dependendo das condições ambientais de cada local, a fase do “pânico” pode retornar.
5.4. Impactos econômicos
Não foram documentados impactos eco-nômicos desta espécie sobre sistemas construí-dos.
5.5. Danos à saúde
As espécies do gênero Corbicula são bio-bio-acumuladoras de metais pesados, que podem entrar na cadeia alimentar através de peixes carnívoros e omnívoros que, por sua vez, ser-vem de alimento para populações humanas,
Figura 11. Corbicula fluminalis (Müller, 1774) o berbigão asi-ático: A, vista externa da valva esquerda; B, vista interna da valva direita (Foto: M.C.D. Mansur).
5.2. Etimologia
O nome específico fluminalis, vem do latim “flumen” que significa rio. Corbicula fluminalis poderia ser definida como a espécie com a concha em forma de cestinha, que vive nos rios. Em algumas regiões como RJ e MG são chamadas de berbigões asiáticos. Em meios acadêmicos todas as espécies de corbículas são mais conhecidas como corbícula asiática ou so-corbícula asiática ou so-mente corbícula. Segundo Korniushin (2004), a espécie que durante muitos anos foi considerada por Morton (1982) – revisor da espécie – como C. fluminalis, seria, na realidade, uma espé-cie muito afim à estuarina Corbicula japoni-ca. Esta confusão de Morton (op. cit.) trouxe muitas incertezas na identificação da espécie. A morfologia da espécie conferia com os exem-plares coletados no baixo Rio Jacuí, bacia do Atlântico Sul, em 1990, porém o ambiente aqui apresenta salinidade zero. Por precaução, Man-sur et al. (2004c) a chamaram inicialmente de Corbicula sp “b”. Martins et al. (2004) a regis-tram como C. aff. fluminalis para o Lago Gua-íba, RS. Somente após a revisão de Korniushin (2004) foi possível determinar a espécie C. flu-minalis com segurança na América do Sul.
5.3. Impactos ambientais
As populações desta espécie encontradas no sul do Brasil são pequenas e pouco densas.

CAPÍTULO 2
Seção 1
37
o que implicaria em riscos para saúde. Não se conhecem trematódeos que se hospedem nesta espécie, nem da possibilidade de ser intermedi-ário de alguma zoonose.
5.6. Lugar de origem e dispersão nas áreas invadidas
Originalmente descrita do Rio Eufra-tes, na Mesopotâmia, Ásia menor (Araujo et al., 1993). Posteriormente, também foi re-gistrada para Ásia Central, Cáucaso, África (Korniushin, 2004) e China (Glaubrecht et al., 2007), sendo esta provavelmente a distribuição original da espécie. Foi introduzida na Europa, América do Sul e no Brasil, provavelmente via água de lastro.
5.7. Distribuição Geográfica
Registros da espécie se limitam ao com-plexo lagunar e fluvial do sistema Guaíba–Pa-tos-Mirim, no Rio Grande do Sul e no Uruguai e na bacia do Rio de La Plata, na Argentina (Fig. 12). Não há citações para outras bacias no Brasil (Mansur et al., 2011a, 2012). Não existem também registros publicados sobre a
presença de C. fluminalis no Rio de La Plata e contribuintes, nem no Rio Uruguai. Por co-municação pessoal do malacólogo Dr. Cristián Ituarte, do Museu Argentino de Ciências Natu-rais “Bernardino Rivadávia”, Buenos Aires, C. fluminalis ocorreu no Rio de La Plata, mas hoje não é mais encontrada.
6. O berbigão asiático rosa Corbicula sp.
6.1. Características da espécie
Corbicula sp. apresenta concha robusta quase equilateral, inflada (Fig.13), altura me-nor que o comprimento que chega a 4,3 cm, sem rostro, margem anterior levemente côncava na frente dos umbos, linhas comarginais mais delicadas, semelhantes às de C. largillierti (uma a cada 0,7 mm) e baixas, charneira reforçada, relativamente pouco arqueada, umbos baixos e arredondados, levemente voltados para a frente (prosógiros) e quase centrados (quase na meta-de do comprimento), internamente cor clara,
Figura 12. Mapa com a distribuição atual do berbigão asiático Corbicula fluminalis (Müller, 1774) na América do Sul. Mapa: G. Figueiredo e M.C.D. Mansur.
Figura 13. Corbicula sp. o berbigão asiático rosa: 13A, vista interna da valva direita; 13B, vista externa da valva esquerda (Foto: M.C.D. Mansur).

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
38
levemente lilás ou rosa e o perióstraco castanho com brilho. Muito semelhante a C. largillierti, porém bem mais robusta.
6.2. Etimologia
A espécie não foi determinada. Em meios acadêmicos do sul do Brasil é conhecida por corbícula ou berbigão rosa.
6.3. Impactos ambientais
As populações desta espécie encontradas no sul do Brasil são pequenas e pouco densas. Geralmente são encontradas em margens rasas, arenosas e nas proximidades de juncais.
6.4. Impactos econômicos
Não foram documentados impactos econô-micos desta espécie sobre sistemas construídos.
6.5. Danos à saúde
As espécies do gênero Corbicula são bio-bio-acumuladoras de metais pesados, que podem
entrar na cadeia alimentar através de peixes carnívoros e omnívoros que, por sua vez, ser-vem de alimento para populações humanas, o que implicaria em riscos para saúde. Não se conhecem trematódeos que se hospedem nesta espécie, nem da possibilidade de ser intermedi-ário de alguma zoonose.
6.6. Lugar de origem e dispersão
Provavelmente do sudeste asiático
6.7. Distribuição Geográfica
Registros da espécie (Fig. 14) se limitam ao complexo lagunar e fluvial do sistema Gua-íba-Patos, no Rio Grande do Sul, Brasil (Man-sur et al., 2011a).
Gastrópodes límnicos invasores 7. O caracol asiático ou caramujo-trombeta Melanoides tuberculata (Müller, 1774)
7.1. Características da espécie Melanoides tuberculata (Müller, 1774) é um Caenogastropoda da família Thiaridae, cujas conchas em geral são espessas e forte-mente esculturadas (Fig. 15). A concha de M. tuberculata é moderadamente grossa, alongada e espiralada, turriforme (em forma de torre), com 12 a 16 voltas nos animais adultos, alcan-çando em média 35 mm de comprimento e 12 mm de largura, mas alguns indivíduos podem ultrapassar 40 mm de comprimento e 14 mm de largura (Brandt, 1974). A descoloração do ápice e sua ausência, assim como a ausência das voltas iniciais é frequente, pois a concha sofre a ação do ácido carbônico presente na água. O umbílico é fechado. O perióstraco é amarron-zado ou amarelado, mas existe grande variação de cor de acordo com o ambiente. A abertura é relativamente pequena, oval-alongada. A con-cha é bem ornamentada, apresentando linhas espirais com tubérculos de tamanhos variados. Manchas coloridas (flâmulas) acastanhadas ou marrons estão presentes, assim como uma
Figura 14. Distribuição atual do berbigão asiático rosa Cor-bicula sp., na América do Sul. Mapa: G. Figueiredo e M.C.D. Mansur

CAPÍTULO 2
Seção 1
39
banda colorida na base da columela em alguns exemplares. Apresenta um opérculo córneo, preso ao pé, que fecha a abertura da concha (Figs. 15, 16). O animal possui uma probóscide alongada e o pé é ovalado (Fig. 16). O manto é amarelado, com manchas escuras; o bordo do mato é franjado (Fig. 16). Os animais vivem so-bre substratos lamosos, macios e turvos, onde
podem enterrar-se, mas também sobre substra-tos mais rígidos, formados por seixos de vários tipos e tamanhos (Fig. 17). Também são encon-trados na vegetação aquática, entre as raízes de plantas flutuantes e em substratos artificiais como garrafas pet, caixas de papel, etc. Apre-sentam comportamento gregário, formando grandes aglomerados populacionais (Fig. 18).
Figura 15. Melanoides tuberculata (Müller, 1774): A, aspecto geral da concha. À esquerda, vista ventral; à direita, vista dorsal. B, detalhe do opérculo córneo, paucispiral. C, detalhe da escultura da concha de exemplar jovem à microscopia de varredura (Fotos: A, P. M. Coelho; B, I.C. Miyahira; C, A.C.N. Moraes, Laboratório de Microscopia Eletrônica UERJ).
Figura 16. Melanoides tuberculata (Müller, 1774): A, aspecto do animal vivo, mostrando o pé ovalado com o opérculo preso em sua porção posterior, probóscide alongada, tentácu-los filiformes, com os olhos na base. B, detalhe da porção anterior, vista dorsal, mostrando as franjas do bordo do manto. C, detalhe da cabeça e pé, vista ventral, mostrando a pig-mentação do manto (Fotos: A, P.M. Coelho; B, S.B. Santos; C, I.C. Miyahira).

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
40
7.2. Etimologia
O nome do gênero é uma contração do nome genérico Melania com o sufixo latino –oi-des, do grego eidos (εἶδος), que significa forma, aparência, semelhança. Portanto, Melanoides significa de forma similar à Melania, que é ou-tro gênero do grupo Cerithioidea. Melania por sua vez deriva do grego melaina (μελαινα), que significa escuro, preto, dizendo respeito à cor escura do perióstraco desses gêneros. A palavra tuberculata significa tubérculos, se referindo à escultura em forma de tubérculos que cobre a concha do animal. Na literatura a combinação Melanoides tuberculatus é muito comum, pois o sufixo “oides” denota uma palavra mascu-
lina, de acordo com o Código Internacional de Nomenclatura Zoológica (ICZN, 2000). De acordo com o Artigo 30.1.4.4. do ICZN (2000), todo nome finalizado em -oides é mas-culino, exceto quando o autor ao criar o gênero estabeleceu o gênero feminino devido à com-binação específica original. Como Melanoides foi estabelecido por Olivier (1804) com a com-binação original Melanoides fasciolata, ou seja, um epíteto feminino, a combinação correta é Melanoides tuberculata (Müller, 1774).
7.3. Impactos ambientais
Atualmente, os problemas relacionados aos impactos no meio ambiente oriundos da
Figura 17. Melanoides tuberculata (Müller, 1774): Tipos de substratos ocupados. A, substrato lamoso, fino, águas turvas. B, substrato rígido, seixos de diversos tipos e tamanhos, águas límpidas. (Fotos: A, S.B. Santos; B, I.C. Miyahira).
Figura 18. Melanoides tuberculata (Müller, 1774): Agregado formado por centenas de indi-víduos em um riacho na Ilha Grande, Rio de Janeiro (Foto I.C. Miyahira).

CAPÍTULO 2
Seção 1
41
introdução de M. tuberculata são os principais observados no Brasil. É uma espécie bentônica, capaz de formar populações extremamente densas (Miyahira, 2010), que muitas vezes cobrem inteiramente o substrato (Fig. 15C), afetando assim diretamente as demais espécies de macroinvertebrados bentônicos (Moreno & Callisto, 2006). As interações de M. tubercu-lata com outras espécies são pouco estudadas. Freitas et al. (1987) sugerem uma diminuição das populações de Biomphalaria tenagophi-la (d’Orbigny, 1835) e de Pomacea haustrum (Reeve, 1856), em função do crescimento da população de M. tuberculata na Lagoa da Pam-pulha, Minas Gerais. Giovanelli et al. (2003) apresentam estudos de campo e laboratoriais sugerindo o deslocamento de B. tenagophila
por M. tuberculata. Por sua vez, Thiengo et al. (2005) relacionam o decréscimo das popula-ções de Aylacostoma tenuilabris (Reeve, 1860), em função do crescimento das populações de M. tuberculata na área da Usina Hidrelétrica de Serra da Mesa, Goiás. Efeitos negativos sobre o tamanho populacional de macroinvertebrados não-moluscos foram observados por Miyahi-ra (2010) em um estudo realizado na Vila do Abraão, Ilha Grande (Angra dos Reis, Rio de Janeiro). A capacidade de produzir material fino particulado e liberar partículas de blocos compactos é comprovada para M. tuberculata (Dudgeon, 1982). Além de ocupar espaço no substrato, altera os processos de sedimentação e turbação, modificando os processos físicos relacionados ao ambiente, afetando por fim as espécies que nele vivem. Melanoides tuberculata também disponi-biliza no ambiente na qual é introduzida um “novo” substrato duro (a concha), que pode ser colonizado por algas e outras espécies menores de moluscos, como ancilídeos e jovens bionfa-lárias, por exemplo, e invertebrados em geral. Algas filamentosas (Fig. 19) e nematódeos de vida livre também já foram observados aderi-dos às conchas de M. tuberculata (Miyahira, 2010). A dispersão deste tiarídeo exótico pode levar para outros lugares, inadvertidamente, essas outras espécies aderidas às suas conchas. Outro fator a ser considerado é o mau cheiro ocasionado pela morte de milhares de exemplares quando ocorre a descida do nível da água em reservatórios artificiais, por ocasi-ões de seca (Fig. 20).
7.4. Impactos econômicos Não existem relatos de impactos econô-micos extensos de M. tuberculata como aqueles ocasionados por C. fluminea e L. fortunei, es-pécies capazes de parar indústrias e hidroelé-tricas devido à obstrução de captações de água, filtros e turbinas. Podemos relacionar a introdução deste tiarídeos com impactos de menor monta. A in-trodução desta espécie em pisciculturas pode levar a gastos com controle de pragas, além de representar uma possibilidade de estabeleci-mento de algumas verminoses nos peixes, le-
Figura 19. Melanoides tuberculata (Müller, 1774) com filamen-tos de macroalgas aderidas à superfície da concha (Foto I.C. Miyahira).
Figura 20. Acúmulos concêntricos de conchas de moluscos mortos quando da descida da água em um açude no Nordes-te do Brasil. No detalhe, conchas de Melanoides tuberculata (Müller, 1774) e de Biomphalaria straminea (Dunker, 1848) (Fotos S.B. Santos).

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
42
vando a prejuízos. Agudo-Padrón (2010) rela-tou a dispersão de M. tuberculata associada ao comércio de camarão. A presença de impurezas no produto comercializado (camarão) pode acarretar na diminuição do valor final.
7.5. Impactos na saúde humana
Melanoides tuberculata é o primeiro hos-pedeiro intermediário de vários trematódeos de importância médica, listados na Tabela I. No Brasil, duas espécies de trematódeos foram registradas por Pinto & Melo (2010a, b) infec-tando M. tuberculata: Centrocestus formosanus (Nishigori, 1924) e Philophthalmus gralli Ma-this & Leger, 1910. Centrocestus formosanus, o helminto responsável pela centrocestíase, tem como segundo hospedeiro intermediário diversas espécies de peixes dulceaquícolas e como hospedeiros definitivos, aves e mamíferos (Scholz & Salgado-Maldonado, 2000; Aguilar--Aguilar et al., 2009). Espécies do gênero Phi-lophthalmus Looss, 1899 parasitam os olhos de aves, que são seus hospedeiros definitivos, podendo infectar ocasionalmente o homem e outros animais, através do contato direto com os olhos ou por via oral (Waikagul et al., 2006; Derraik, 2008). Também há o relato de um caso huma-no autóctone de paragonimíase no Brasil, em Salvador, Bahia, mas a participação de M. tu-berculata na transmissão do parasito não foi investigada (Lemos et al., 2007). A doença é
provocada pela infecção por Paragonimus spp., que é um parasito pulmonar (Keiser & Utzin-ger, 2009; Pinto, 2009). No Brasil, imigrantes asiáticos já foram encontrados infectados por Clonorchis sinensis (Cobbold, 1875), porém não há o registro de transmissão autóctone por este parasito no país (Leite et al., 1989). C. sinensis é parasito das vias biliares de seres humanos (Keiser & Utzin-Keiser & Utzin-ger, 2009; Pinto, 2009). O fato de M. tuberculata participar do ciclo de vida de várias espécies de trematóde-os justifica a necessidade de um maior conhe-cimento sobre a espécie, pois ela pode estar envolvida na introdução, manutenção e disse-minação de parasitos de interesse médico no Brasil. A Tabela II lista os casos mencionados na literatura.
7.6. Origem e dispersão.
Müller (1774) aponta como localidade-tipo a costa do Coromandel, no sudeste da Índia, mas existem divergências quanto à distribuição original da espécie. Pilsbry & Bequaert (1927) indicam como faixa original de distribuição o norte e o leste da África e o sul da Ásia, incluin-do a Indonésia. Benthem-Jutting (1956) apresen-tou uma distribuição englobando a África, sul da Ásia e norte da Austrália. Facon et al. (2003) modificaram um pouco esta distribuição in-cluindo toda a Oceania. Glaubrecht et al. (2009) tratam M. tuberculata como sendo espécie exó-
Tabela I. Trematódeos de importância médica que possuem Melanoides tuberculata (Müller, 1774) como primeiro hospedeiro intermediário.
Trematódeo Enfermidade Órgãos afetadosSegundo
hospedeiro intermediário
Hospedeiros definitivos Referências
Paragonimus westermani (Kerbert, 1878) paragonimíase pulmões Crustáceos Humanos
e outros animais 1, 4
Clonorchis sinensis (Cobbold, 1875)
clonorquíase fígado Peixes Humanos 4
Centrocestus formosanus (Nishigori, 1924) centrocestíase intestino delgado Peixes
Peixes, répteis, aves e mamíferos 2, 4, 5
Haplorchis pumilio(Looss, 1896) haplorquíase intestino delgado Peixes Aves e mamíferos 4, 5
Philophthalmus gralli Mathis and Leger, 1910 filofitalmíase olhos _
Aves;ocasionalmente
humanos e outros animais
4, 5
1- Guimarães et al., 2001; 2- Umadevi & Madhavi, 2006; 3- Díaz et al., 2008, 4- Derraik, 2008; 5- Pinto & Melo, 2010a, b.

CAPÍTULO 2
Seção 1
43
tica na Austrália e afirmam que só recentemen-te foi introduzida naquele país. Acreditamos que a distribuição apresentada por Pilsbry & Bequaert (1927) seja a mais próxima da origi-nal, por ser a mais antiga, antes da fase de ex-pansão desta espécie. Esta distribuição também é a que não apresenta conflitos com as demais, ou seja, Benthem-Jutting (1956) e Facon et al. (2003). Murray (1964) fez o primeiro registro de M. tuberculata nas Américas; a seguir, em 1967, a espécie foi registrada para o Brasil (Vaz et al., 1986a). Estes seriam os dois primeiros registros fora da área natural de ocorrência, considerando o proposto por Pilsbry & Be-quaert (1927). Na década de 70 temos outros registros para os Estados Unidos da América (Abbott, 1973; Dundee & Paine, 1977) e para o México e Panamá (Abbott, 1973). Durante a década de 80 a espécie passou a ser encontra-da nas ilhas do Caribe (Samadi et al., 1999) e foi efetuada a primeira tentativa de introdução com fins de controle biológico de planorbíde-os na mesma região (Pointier et al., 1989). A partir de então temos registros para a Europa (Stagl, 1993; Juricková, 2006) e para a Oceania (Duggan, 2002; Glaubrecht et al., 2009). Atualmente, esta espécie pode ser cha-mada de cosmopolita (Fig. 21), como suge-rem Ismail & Arif (1993) e Elkarmi & Ismail (2007), pois temos registros da mesma para to-dos os continentes a exceção da Antártica. Na distribuição original de Pilsbry & Be-
quaert (1927) a espécie estava limitada à região tropical. Aparentemente, as regiões mais frias impõem certos limites para uma maior expan-são de sua distribuição, pois a maioria dos re-gistros se encontra em regiões de clima mais ameno. Por exemplo, na Nova Zelândia os re-gistros são limitados à águas termais (Duggan, 2002; Derraik, 2008). No Brasil, a espécie ain-da não foi encontrada no Rio Grande do Sul, extremo sul do país (Thiengo et al., 2007). Os registros de M. tuberculata usual-mente tratam de uma região ou país específico (e.g. Abbott, 1973; Vaz et al., 1986a; Duggan, 2002; Santos et al., 2007). Uma compilação dos dados disponíveis na literatura mostrou que existem registros para 55 países, sendo 25 (45,5%) dentro da área natural de distribuição e 30 (54,54%) fora desta área, considerando--se a proposta de Pilsbry & Bequaert (1927) para a área nativa e ressaltando que esses au-tores apenas delimitaram uma área no mapa, não especificando os países. Percebe-se a gran-de expansão desta espécie, pois na maior parte dos registros da literatura foi classificada como “exótica”.
7.7. Meios de introdução
No Brasil, parece que a introdução de M. tuberculata foi acidental, provavelmente via comércio aquarista (Vaz et al., 1986a). O pri-meiro relato sobre o encontro da espécie data de 1967, quando o dono de uma indústria de
Tabela II. Trematódeos e tipos cercarianos encontrados em Melanoides tuberculata (Müller, 1774) no Brasil, com seus respecti-vos hospedeiros e locais de ocorrência.
Espécie Tipo cercariano Hospedeiro definitivo Ocorrência no Brasil Referências
Centrocestus formosanus “Pleurolophocercous cercaria”
PeixeRéptilAve
Mamífero
RJ: Guapimirim, Maricá,
Rio de Janeiro (Vargem Pequena, Vargem Grande,
Prainha) e Angra dos Reis (Ilha Grande).
Boaventura et al. (2002); Bógea et al. (2005);
Ximenes (2011)
DF: Brasília (Lago Paranoá)
Andrade et al. (2008)
MG: Belo Horizonte (Represa da Pampulha)
Pinto & Melo (2010a)
Philophthalmus gralli “Megalurous cercaria”
Ave;ocasionalmente
humanos e outros animais
MG: Belo Horizonte (Represa da Pampulha) Pinto & Melo (2010 b)

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
44
artefatos de conchas recebeu de um fornecedor diversas conchas coletadas em local indetermi-nado no município de Santos, São Paulo (Vaz et al., 1986a). Em 1971, seis exemplares foram obtidos em uma loja de comércio de peixes or-namentais em Atibaia, São Paulo, mas não foi possível descobrir a origem desses exemplares (Vaz et al., 1986a). A partir do encontro de exemplares no bairro do Macuco, Santos, em 1976, deflagrando-se a suspeita sobre a presen-ça de um molusco exótico em São Paulo, um levantamento efetuado pela SUCEN (Superin-tendência de Controle de Endemias), em 1984 e 1985 verificou sua ocorrência em dez municí-pios, o que levou Vaz et al. (1986a) a suporem que sua introdução no Brasil tenha sido ante-rior à 1967. Após o alerta sobre a presença de um caracol exótico no Brasil, diversos relatos começaram a surgir, o primeiro para o Lago Paranoá, Brasília, em 1984 (Vaz et al., 1987).
7.8. Formas de dispersão
No Brasil, a dispersão de M. tuberculata tem sido relacionada principalmente ao comér-cio aquarista (Vaz et al., 1986a). Os juvenis da espécie, logo após a liberação pela fêmea, são
de tamanho diminuto, medindo entre 2,6 e 3,4 mm de altura (Berry & Kadri, 1974), portanto, passando facilmente despercebidos aos comer-ciantes e aquariófilos, aderidos ao substrato ou às plantas aquáticas. Cowie & Robinson (2003) citaram o comércio de aquário como fonte de introdução intencional e acidental de diversas espécies. A introdução acidental deste tiarídeo também pode ser relacionada à piscicultura e ao comércio de conchas (Vaz et al., 1986a; Thien-go et al., 1998). Recentemente, Agudo-Padrón (2010) reportou a dispersão de M. tubercula-ta, Pomacea bridgesii (Reeve, 1856), P. lineata (Spix, 1827) e P. canaliculata (Lamarck, 1819), facilitada pela carcinicultura, através do trans-porte de camarões das áreas produtoras para outras regiões, sendo os moluscos transporta-dos junto ao camarão. A introdução e dispersão intencional de M. tuberculata foi reportada principalmente para as ilhas do Caribe. A espécie foi utiliza-da como controle biológico das espécies veto-ras da esquistossomose, caramujos do gênero Biomphalaria (Pointier & McCullough, 1989; Pointier et al., 1989, 1993). A espécie rapida-mente se estabeleceu nos locais onde foi intro-duzida. No Brasil, existem estudos utilizando
Figura 21. Distribuição conhecida de Melanoides tuberculata (Müller, 1774). Em azul, área de distribuição original; em vermelho, áreas invadidas, segundo dados da literatura. Em amarelo, a provável localidade-tipo (Mapa I.C. Miyahira).

CAPÍTULO 2
Seção 1
45
M. tuberculata como controle biológico, porém a espécie já se encontrava introduzida antes dos estudos (e.g. Freitas et al., 1987; Giovanelli et al., 2005). Não foram localizados registros for-mais da introdução intencional de M. tubercu-lata para controle biológico ou qualquer outro fim. Alguns atributos biológicos da espécie facilitam a sua dispersão e estabelecimento em um novo ambiente. Por se tratar de uma espécie na qual a partenogênese é comum, formando populações com predominância de fêmeas, em teoria, basta a introdução de uma única fêmea para o estabelecimento de uma nova população. A espécie também apresen-ta boa resistência à dessecação (Facon et al., 2004) e é capaz de suportar águas com tem-peraturas elevadas (Benthen-Jutting, 1956; Duggan, 2002), assim como águas estuarinas (Santos et al., 2007; Barroso & Matthews--Cascon, 2009) e até salgadas (Wingard et al., 2008). Estas características aumentam a probabilidade de sobrevivência dos indivídu-os durante o transporte, seja esse acidental ou intencional, aumentando ainda mais a probabilidade de sobrevivência ao transpor-te, facilitando a introdução em novas locali-dades e a reintrodução. Apesar da grande atenção dada à in-trodução passiva de M. tuberculata, aciden-tal ou intencional, esses moluscos também são capazes de se deslocar ativamente nos corpos d’água, inclusive contra a correnteza e escalando degraus naturais ou artificiais (Fig. 22), como observado por Miyahira et al. (2009).
Figura 22. Exemplares de Melanoides tuberculata (Müller, 1774) escalando ativamente um degrau artificial em um ria-cho na Ilha Grande, Rio de Janeiro (Foto I.C. Miyahira).
Figura 23. Distribuição geográfica de Melanoides tuberculata (Müller, 1774) no Brasil (Mapa P.M. Coelho).
7.9. Distribuição geográfica no Brasil
Melanoides tuberculata é encontrada em praticamente todas as regiões hidrográfi-cas brasileiras, exceto no extremo sul da Bacia do Atlântico Sul e Sudeste (inexistem registros para o Rio Grande do Sul) e para a maior parte da Bacia do Rio Amazonas (inexistem registros para os estados do Acre, Amazonas, Rondônia, Roraima e Amapá), porém essas ausências po-dem representar apenas um artefato da falta de estudos. Segundo dados da literatura (Fernan-dez et al., 2003, Thiengo et al., 2005, 2007; Souto et al., 2011) e em material depositado em algumas coleções científicas, M. tubercula-ta está presente em 201 municípios brasileiros (Fig. 23).
8. Helisoma duryi Wetherby, 1879) e Helisoma trivolvis (Say, 1817)
8.1. Características das espécies
Ambas as espécies, Helisoma duryi (We-therby, 1879) e Helisoma trivolvis (Say, 1817), pertencem à família Planorbidae, apresentando conchas planispirais, isto é, os giros crescem

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
46
em um mesmo plano, dando à concha aspecto de disco ou moeda (Fig. 24). A principal carac-terística é a concha planispiral com giros que crescem rapidamente em diâmetro, com a su-tura bem profunda em ambos os lados. O lado direito é côncavo e o esquerdo é pouco escava-do, tendendo a aplanado. A abertura da concha torna-se, às vezes, muito dilatada, o que nos permite diferenciá-los de outros Planorbidae, como Biomphalaria spp. Segundo Paraense (1975, 1976a, b) He-lisoma duryi (Fig. 25A, B) atinge 18 mm de diâmetro, 8,5 mm na largura da abertura e 5 giros, enquanto que Helisoma trivolvis (Figs. 25C, D) é um pouco maior, alcançando 25 mm de diâmetro, 9 mm na largura da abertura e 5,5 giros. Em campo, sua diferenciação é difí-cil, sendo necessária a dissecação para a corre-ta identificação.
8.2. Etimologia
O nome do gênero vem do grego helis, significando enrolado em espiral e soma, cor-po, ou seja, corpo enrolado em espiral. O epi-teto específico duryi foi concedido em home-nagem ao naturalista Charles Dury. O epíteto trivolvis significa três voltas.
8.3. Impactos ambientais, econômicos e na saúde humana
Embora não haja registro de qualquer impacto ambiental ou econômico, a utilização de H. duryi no controle de moluscos hospe-deiros de Schistosoma mansoni Sambon, 1907 e Schistosoma haematobium (Bilharz, 1852) foi registrada na década de 70. Os trabalhos de Milward-de-Andrade (1978a, b), Milward--de-Andrade & Souza (1979) e Milward-de--Andrade et al. (1979) mostraram as seguintes vantagens adaptativas de H. duryi sobre Biom-phalaria glabrata (Say, 1818), sob condições de laboratório: (1) ser mais profílera e possuir menor mortalidade; (2) ser resistente ao des-secamento ambiental; (3) suportar a falta de alimento; (4) ser refratária ao S. mansoni. Com base nesses fatos, Milward-de-Andrade (1979) conjecturou a capacidade de H. duryi colonizar os ecossistemas neotropicais, e de maneira dire-ta ou indireta, exercer importante papel no con-trole biológico da esquistossomose mansônica. Fernandez et al. (2010) assinalaram que, até o presente, H. duryi não constitui ameaça à malacofauna nativa, nem tem causado perdas econômicas ou danos à saúde humana. Entre-tanto, esses autores ressaltaram a importância da utilização de mecanismos que impeçam a expansão da distribuição dessa e de outras espécies exóticas para a preservação de nossa biodiversidade.
8.4. Origem e dispersão
Segundo Paraense (1981a) algumas espé-cies do gênero Helisoma Swainson, 1840 ex-pandiram-se de seu domínio Neártico original para a América do Sul a oeste dos Andes, até o Peru. A localidade-tipo de H. duryi é Ever-glades da Flórida, Estados Unidos da Améri-ca. Após estudo comparativo de amostras de
Figura 24. Helisoma duryi (Wetherby, 1879). Aspecto do ani-mal jovem, vivo (Foto P.M. Coelho).
Figura 25. Aspecto da morfologia da concha das espécies de Helisoma Swainson, 1840 presentes no Brasil. A (lado direito) e B (lado esquerdo) da concha de H. duryi (Wetherby, 1879). C (lado direito) e D (lado esquerdo) da concha de H. trivolvis (Say, 1817) (Fotos P.M. Coelho).

CAPÍTULO 2
Seção 1
47
populações pertencentes ao gênero Helisoma, Paraense (1976b) revelou as seguintes espé-cies nominais de H. trivolvis: Planorbis fove-alis Menke, 1830 (Jamaica), Planorbis affinis Adams, 1849 (Jamaica), Planorbis auriculatus Clessin, 1884 (Jamaica), Planorbis intermedius “Philippi” Dunker, 1850 (México), Planorbis ancylostomus Crosse & Fischer, 1879 (Méxi-co), Planorbis equatorius Cousin, 1887 (Guaya-quil, Equador) e Planorbis lentus Say, 1834 (New Orleans, Estados Unidos da América). Este autor refere-se, além das localidades-tipos destas espécies nominais, outras localidades de H. trivolvis: o Lago Xochimilco (México), Tembladera, Cajabamba, Puerto Chicama e Lima (Peru), e o Lago Miragoane (Haiti).
8.5. Meios de introdução
Helisoma duryi vem sendo introduzi-da em diversas áreas a leste dos Andes, segu-ramente por ação humana (Paraense, 1981a), tendo sido encontrada como população natu-ralizada no Brasil Central (Paraense, 1976a) ou colonizando ambientes artificiais como aquá-rios ou lagos artificiais (Thiengo et al., 1998; Fernandez et al., 2010). Os meios de introdução de H. trivolvis possivelmente são os mesmos de H. duryi, in-cluindo o transporte através de plantas aquáti-cas, provável meio de introdução da população encontrada pela equipe do Laboratório de Ma-lacologia do Instituto Oswaldo Cruz na cidade do Rio de Janeiro.
8.6. Formas de dispersão
Principalmente plantas aquáticas que são utilizadas para suprir o oxigênio necessário à criação e ao transporte de peixes de água doce, geralmente ornamentais. Nos Estados do Rio de Janeiro e Ceará, a ocorrência de populações de H. trivolvis e H. duryi foi observada em locais onde haviam peixes ornamentais destinados ao comércio ou como uma forma de controle bioló-gico das larvas de Aedes aegypti Linnaeus, 1758. Os meios de dispersão são por ação humana, onde as desovas ou pequenos filhotes são carrea-dos nas folhas ou talos das plantas aquáticas, ou ainda sobre conchas de outros moluscos, como os ampulariídeos Pomacea spp., geralmente co-
locados em aquários para manterem os vidros limpos, uma vez que são animais raspadores, que se alimentam do limo que se forma nas paredes. De acordo com Madsen & Frandsen (1989) populações de H. duryi são observadas em aquários na Europa, o que possivelmente vem ocorrendo no Brasil com as espécies de Helisoma.
8.7. Distribuição geográfica no Brasil
Helisoma duryi se encontra presente nas Bacias Hidrográficas do Atlântico Nordeste Oriental, do Atlântico Sul e Sudeste, Atlântico Leste e do Tocantins-Araguaia. Helisoma tri-volvis foi observado somente na Bacia Hidro-gráfica do Atlântico Sudeste, no Rio de Janeiro (Fig. 26).
9. Physa acuta Draparnaud, 1805
9.1. Características da espécie
Physa acuta é um gastrópode Pulmona-ta Basommatophora (olhos na base dos tentá-culos), da família Physidae, caracterizada pela concha sinistrógira (enrolamento da concha
Figura 26. Distribuição atual das espécies de Helisoma Swain-son, 1840 no Brasil (Mapa I.C. Miyahira).

Santos et al.
Espécies de moluscos límnicos invasores no Brasil
48
para a esquerda). A concha é oval-alongada, fina, lisa, moderadamente brilhante e translú-cida; cinco giros, espira curta e sutura ligeira-mente marcada. Abertura grande, sinistrógira, ocupando cerca de ¾ do total do comprimen-to da concha. Massa cefalopediosa uniforme, de cor cinza escuro, teto da cavidade palial profundamente pigmentado, visível através da concha. As conchas possuem em média 16 mm de comprimento e 9 mm de largura (Paraense & Pointier, 2003), mas podem alcançar mais de 20 mm em ambientes com muita matéria orgânica à disposição (Sonia B. Santos, obser-vação pessoal). Apresenta grande plasticidade fenotípica, fato que justifica a imensa lista com 155 sinônimos fornecida por Paraense (2011). A Figura 27A mostra um topotipo, onde se observa a concha com espira mais alongada e volta corporal mais estreita, diferindo dos exemplares de espira mais curta e volta corpo-ral mais abaulada, comuns no Brasil (Figs. 27B, 28). Habita diversos tipos de ambientes límni-cos, inclusive antrópicos, alcançando elevada densidade populacional, como registrado por
Figura 27. Physa acuta Draparnaud, 1805. Exemplares vivos. A, topotipo, mostrando a espira mais alongada e a volta cor-poral mais estreitada. B, morfotipo comum no Brasil, com a espira mais curta e volta corporal mais abaulada (Fotos: P.M. Coelho).
Figura 28. Physa acuta Draparnaud, 1805: Aspecto da morfo-logia da concha; à esquerda, lado dorsal; à direita, lado ven-tral. Exemplar CMIOC 4798 (Fotos: P.M. Coelho).
França et al. (2007), em reservatórios do baixo Tietê, onde a espécie alcançou 4350 ind./m-2 no período chuvoso. Toleram ambientes poluídos (Appleton, 1996; Fraga, 2002), sendo que na África do Sul podem atingir até 3.000 ind./m2
(Appleton, 1996).
9.2. Etimologia
O nome do gênero Physa é originário do termo grego physao, que significa saco, bolsa, enquanto o epíteto específico vem do termo em latim acutus, cujo significado é agudo, afiado. Physa acuta Draparnaud, 1805 poderia ser de-finida como uma espécie com a concha em for-ma de bolsa com uma das extremidades afiada, correspondendo ao ápice da concha (Te, 1980).
9.3. Impactos ambientais e econômicos
Não há registros.
9.4. Impactos na saúde humana
Não há registros. Todavia, existem evi-dências, através de infecções experimentais, de que P. acuta possa atuar como hospedeiro intermediário de Fasciola hepatica Linnaeus, 1758 (Barros et al., 2002) e de Echinostoma spp. (Morales et al., 1987).
9.5. Origem da espécie e dispersão
Physa acuta foi descrita por Draparnaud em 1805, baseada em espécimes coletados na bacia do Rio Garonne, na França, mas é prová-vel que P. acuta seja originária da América do Norte (Dillon et al., 2002). Muitos registros dessa espécie fora de sua localidade-tipo têm sido relatados em países da Europa, Ásia, África, além de registros na Aus-trália, Havaí e nos Estados Unidos (Paraense & Pointier, 2003; Madsen & Frandsen, 1989): Eu-ropa (Alemanha, Áustria, Bélgica, Bulgária, Escó-cia, Espanha, Grécia, Holanda, Hungria, Ingla-terra, Irlanda do Norte, Itália, Polônia, Portugal, República Tcheca), Ásia (Azerbaijão, Bangladesh, China, Geórgia, Índia, Irã, Iraque, Israel, Japão, Jordânia, Coréia e Macau), África (Argélia, Egi-to, Etiópia, Quênia, Ilhas Maurício, Rodésia, Madagascar, La Reunion, Zimbábue, Marrocos,

CAPÍTULO 2
Seção 1
49
Namíbia, Nigéria, África do Sul, Uganda, Sudão, Tunísia, Zaire), Austrália, Havaí e Estados Uni-dos (Massachusetts, Virgínia). Há ainda ocorrên-cia dessa espécie nas Américas Central e do Sul (Argentina, Barbados, Bolívia, Brasil, Chile, Co-lômbia, Cuba, Equador, El Salvador, Guatemala, Jamaica, Martinica, México, Nicarágua, Pana-má, Peru, Porto Rico, Santa Cruz, São Tomás, Trinidade, Uruguai e Venezuela).
9.6. Meios de introdução
É provável que P. acuta tenha sido intro-duzida de forma não intencional, por meio do comércio ou transporte de plantas aquáticas e atividades de aquariofilia.
9.7. Formas de dispersão
A aquariofilia e o comércio de plantas aquáticas e de peixes para piscicultura são considerados fontes de dispersão dessa espé-cie em nível global. É uma espécie bastante cosmopolita, capaz de se dispersar rapidamente e colonizar diversas áreas, encontrado em variados ambientes de água doce, como tanques pequenos, represas, lagos e córregos, geralmente de curso lento, eutrofizados ou não.
9.8. Distribuição geográfica no Brasil
Os registros de Physa acuta no Bra-sil (Fig. 29) estão, por ora, restritos às Bacias Hidrográficas do Atlântico Leste e Sudeste, segundo dados da literatura (Thiengo et al., 1998, 2004, 2005; Miyahira et al., 2010) e da Coleção Malacológica do Instituto Oswaldo Cruz (CMIOC). Fernandez (2011) relatou o encontro de populações de P. acuta no reserva-tório da Usina Hidrelétrica de Serra da Mesa, Bacia Hidrográfica Tocantins-Araguaia, em biótopos com ocorrência de Physa marmorata Guilding, 1828.
Figura 29. Distribuição atual de Physa acuta Draparnaud 1805 no Brasil (Mapa: P.M. Coelho).


51
Introdução
As bioinvasões são a chegada, estabele-cimento e subsequente difusão de espécies não nativas em comunidades naturais nas quais elas não existiam. A introdução destes organismos é, geralmente, mediada pela atividade humana e pode alcançar níveis extremos (Souza et al., 2009). A introdução de organismos exóticos pode ter consequências irreversíveis e devasta-doras para os ecossistemas naturais, afetando tanto a biodiversidade quanto as atividades econômicas. Abordagens ecológicas do proble-ma esbarram em dificuldades que o uso da Ge-nética, em alguns casos, pode resolver (Silva, 2004). Por exemplo, o desenvolvimento da téc-nica de PCR (Reação em cadeia da polimerase, do inglês Polymerase Chain Reaction) propor-cionou uma facilidade na identificação de ovos, larvas e jovens no plâncton (Pie et al., 2006; Boeger et al., 2007). Este tipo de abordagem é extremamente útil quando é preciso identificar as espécies invasoras nas fases mais iniciais do
desenvolvimento larval, como é o caso das bio-invasões mediadas pela água de lastro. Dados genéticos gerados por eletroforese de aloenzi-mas ou pelas mais diversas técnicas de DNA podem, também, ser usados para distinguir as espécies invasoras das nativas. Do mesmo modo, esta capacidade de identificação mole-cular das espécies, pode auxiliar o sistemata clássico no seu trabalho de pesquisar e definir diferenças morfológicas em grupos de espécies de classificação difícil. Outra questão que pode ser abordada a partir das técnicas e teoria da genética é a origem das populações invasoras. A resposta para esta questão pode ser inferida a partir do cálculo de índices de identidade gêni-ca entre populações de vários possíveis pontos de origem e a população invasora em questão. As identidades gênicas são calculadas par a par, neste caso, o par “população invasora–popula-ção do possível ponto de origem” que apresen-tar o maior valor de identidade gênica estará indicando, possivelmente, a origem geográfica da bioinvasão (Silva, 2001a). A Genética pode
CAPÍTULO 3
Genética da bioinvasão do mexilhão-dourado
Edson Pereira da Silva

Edson Pereira da Silva
Genética da bioinvasão do mexilhão-dourado
52
ser utilizada, também, no estudo da dinâmica dos processos de invasão e é, a este caso, que este capítulo se dedica: compreender aspectos da genética e evolução da bioinvasão do mexi-lhão-dourado no Brasil. A abordagem genética aplicada ao pro-blema da bioinvasão da espécie Limnoperna fortunei (Dunker, 1857) envolve a busca e o uso de marcadores moleculares (genes e seus produtos diretos) e sua interpretação usando a fundamentação teórica da genética de popula-ções. No próximo item esta abordagem é suma-riada.
A abordagem genética
A constituição genética dos indivíduos (genótipo) e o problema das leis governando a sua herança constituem o objeto de estudo da genética. A genética de populações, por sua vez, está preocupada com o estudo dos genóti-pos de grupos de indivíduos, as populações, e como esta constituição genética pode mudar ao longo das gerações. A mudança da composição genética das populações, ao longo das gera-ções, constitui o processo evolutivo e, por isto mesmo, estudar genética de populações é, tam-bém, estudar o processo evolutivo. Para que o processo evolutivo ocorra, a primeira condição é que haja variação gênica presente nas popu-lações. De outro modo, não é possível que haja
mudança ao longo do tempo. Desta forma, o trabalho de medir e caracterizar a variação gênica presente em populações naturais, bem como o entendimento dos mecanismos que determinam o seu padrão de distribuição nas populações, é condição fundamental para se estudar genética de populações (Silva, 2001b).Uma forma de medir e caracterizar a variação gênica presentes nas populações naturais é a utilização de marcadores moleculares que são locos gênicos que apresentam alguma variabi-lidade (Silva & Russo, 2000; Silva, 2004). A clássica técnica de eletroforese de aloenzimas, que teve grande impacto nos estudos populacio-nais na década de 1960, cada vez menos usada, e as várias técnicas de DNA, cada vez mais ro-tineiras, constituem as ferramentas moleculares de amostragem de variação gênica úteis para es-tudos a respeito da genética e evolução (Fig. 1). A eletroforese pode ser definida, de ma-neira geral, como a migração de partículas sob a ação de uma corrente elétrica. A técnica se baseia nas características físico-químicas das proteínas, que apresentam cargas diferentes, e no fato de que, pelos princípios da biologia mo-lecular, espera-se certa correspondência entre sequência de nucleotídeos no DNA e a sequên-cia de ácidos aminados na proteína por ele co-dificada, ou seja, entre o gene e o seu produto. Desta forma, proteínas diferentes apresentam cargas diferentes, que por sua vez irão manifes-
Figura 1. A união da genética com a teoria evolutiva permite o entendimento de como a composição genética das populações muda ao longo do tempo (genética de populações). A união dos princípios da biologia molecular com a genética permite estu-dar a variação fenotípica como variação em moléculas como o DNA e as aloenzimas (os marcadores moleculares).

CAPÍTULO 3
Seção 1
53
tar diferentes mobilidades eletroforéticas, que representam diferentes alelos. Desta forma, es-tudar variação de proteínas (aloenzimas) é es-tudar variação no DNA (Silva, 2009). A Figura 2 apresenta um esquema deste método. No caso da variação de DNA, são várias as formas pelas quais ela pode ser estudada. Por exemplo, segmentos de DNA amplificados por intermédio de reações em cadeia da poli-merase (PCR), que contém sequências repeti-tivas, constituem o tipo de variação conhecida como microssatélites (repetições constituídas de 2 a 10 pares de bases) ou minissatélites (mais de 10 pares de base de repetição). Outra forma de variação gênica é aquela que tem origem em mutações pontuais. Neste caso, o estudo pode ser feito pela utilização de enzimas de restri-ção que cortam o segmento em regiões por elas reconhecidas (Polimorfismo de tamanho dos fragmentos de restrição, RFLP do inglês restriction fragment lenght polymorphism), por processos desnaturativos que revelam muta-ções pontuais refletidas em polimorfismos de conformação das fitas simples de DNA (por
exemplo, a técnica de SSCP, do inglês single strand conformational polymorphism), ou ain-da, por sequenciamento total do segmento. Alternativamente, as facilidades da técnica de PCR podem ser utilizadas para amplificar seg-mentos aleatórios de DNA ao longo de todo o genoma dos organismos, como no caso da téc-nica de RAPDs (do inglês randomly amplified polimorphic DNAs) (Avise, 2004). Na Figura 3 estas técnicas estão esquematizadas de modo a dar uma ideia daquilo que foi resumido aqui. A escolha da técnica a ser utilizada de-pende fundamentalmente do problema a ser in-vestigado. Por exemplo, técnicas como RAPDs produzem uma quantidade muito grande de variação observável, o que é útil para estudos de paternidade e modos de reprodução, mas não são indicadas para estudo de filogenia (pa-rentesco entre espécies diferente), para os quais técnicas como RFLPs e sequenciamento são excelentes. Do mesmo modo, estudos de estru-turação populacional têm em microssatélites e aloenzimas uma boa alternativa de amostragem de variação gênica (Silva & Russo, 2000).
Figura 2. Representação esquemática do método de eletroforese de aloenzimas para amostragem da variação gênica em po-pulações naturais.

Edson Pereira da Silva
Genética da bioinvasão do mexilhão-dourado
54
Genética da bioinvasão
Por definição, espécies exóticas se acham em regiões biogeográficas diferentes daquelas nas quais evoluíram e se adaptaram, dessa for-ma, bioinvasoras têm que enfrentar pressões seletivas novas bem como novas situações de stress (Souza et al., 2009). Dentre os tipos de mudança evolutiva a que estão sujeitas as espé-cies invasoras podem ser citados: bottlenecks, efeito de pequeno número de genes e hibridiza-ção (Lee, 2002; Prentis et al., 2008). Em populações naturais pequenas, o aca-so tem um papel importante na determinação de quais genes estarão presentes na próxima geração, força evolutiva conhecida como deriva genética. Um caso extremo de deriva é a redu-ção drástica do tamanho populacional, que tem como consequência a redução dos níveis de va-riação gênica da população (bottleneck ou gar-
galo de garrafa, ver Figura 4). A chegada aciden-tal de um ou poucos indivíduos de uma espécie em um novo ambiente, como se dá no caso das bioinvasões, é um exemplo de bottleneck (co-nhecido, nesse caso, como efeito fundador). Modelos teóricos têm sido desenvolvidos para o estudo dessa dinâmica da bioinvasão. Garcia--Ramos & Rodriguez (2002) examinaram a in-teração entre o processo de adaptações locais e a densidade populacional na velocidade do pro-cesso de invasão. O modelo mostrou que as inte-rações entre os fatores genéticos e demográficos reduzem a velocidade de expansão das espécies, comparado com outros modelos que levam em conta apenas os fatores demográficos. A redu-ção da velocidade de bioinvasão é resultado de uma limitada capacidade das populações de se adaptarem a ambientes novos devido aos baixos níveis de variação gênica. Contudo, embora seja geralmente considerado que as mudanças gené-
Figura 3. Reação em cadeia da polimerase (PCR) e variação na forma de microssatélites (no exemplo, o motivo GATT tem uma, duas e três repetições), RFLP (no exemplo, um, dois e três fragmentos de restrição), SSCP (desnaturação das fitas duplas de DNA pela temperatura e polimorfismos de conformação das fitas simples) e RAPD (segmentos aleatórios amplificados por primers em sítios de homologia no DNA genômico).

CAPÍTULO 3
Seção 1
55
ticas não são detectáveis em tempos ecológicos, evidências de estudos com seleção artificial têm demonstrado que as populações podem sofrer mudanças rápidas de caracteres morfológicos, envolvendo em torno de apenas 100 gerações (Barton & Turelli, 1989). Uma alternativa às mudanças lentas en-volvidas no processo de adaptação é a hibridi-zação. Lee (2002) aponta que a hibridização, entre espécies ou entre populações da mesma espécie (dos bioinvasores com espécies nativas ou com outras espécies invasoras) pode reduzir a perda de variação gênica associada ao pro-cesso de bioinvasão (bottleneck), bem como produzir uma gama de novos genótipos impor-tantes à adaptação da espécie invasora ao novo ambiente. Os efeitos positivos da hibridização ao processo de bioinvasão incluem crescimento mais rápido, maior tamanho dos híbridos e um aumento da agressividade. A variação gênica, seja ela de origem re-combinacional ou mutacional, é extremamen-te importante para o processo de adaptação. Contudo, não se pode negligenciar o efeito de um pequeno número de genes na habilidade de colonização das espécies invasoras. Exemplos notáveis do efeito de um ou poucos genes no sucesso da colonização de ambientes novos tem sido demonstrado para espécies terrestres (Lee, 2002). Embora, não tenha sido encontrada, ainda, uma contraparte para o ambiente aquá-tico, genes dessa natureza não devem ser exclu-sivos de determinados grupos. Por fim, embora não seja uma estratégia adaptativa, mas uma contingência histórica, ou-
tro fator importante para o sucesso das espécies invasoras é o estado de depauperamento do am-biente invadido (Cohen & Carlton, 1988). Am-bientes poluídos, por exemplo, podem facilitar o crescimento de espécies invasoras que, nessas condições, provavelmente, encontram um am-biente de menor competição (Duffy et al., 2000).
O mexilhão-dourado
Para entender o padrão de invasão do mexilhão-dourado nas águas brasileiras, foram utilizados marcadores moleculares de aloenzi-mas para testar diferentes hipóteses sobre a sua dinâmica de invasão. Foram feitas amostragens (ver Figura 5) em três pontos no Brasil (Lago Guaíba, RS; Porto Esperança, MS e Usina de Itaipu, PR) e dois na Argentina (Balneário Ba-gliardi e Usina de Yaciretá). As populações invasoras de L. fortunei, tanto no Brasil quanto na Argentina, apresen-taram altos níveis de variação gênica (Fig. 6), que não estão de acordo com um evento de bottleneck, no qual as populações são estabele-cidas a partir de pequenos estoques com a con-sequente redução da variação gênica da espécie no novo ambiente. Altos níveis de variação gê-nica em espécies invasoras, embora surpreen-
Figura 4. Efeito bottleneck a que estão sujeitas populações in-vasoras, tendo como consequência a redução da quantidade de variação presente na população de origem.
Figura 5. Localização das cinco populações amostradas (Lago Guaíba, RS; Porto Esperança, MS; Usina de Itaipu, PR; Balneá-rio Bagliardi, ARG e Usina de Yaciretá, ARG).

Edson Pereira da Silva
Genética da bioinvasão do mexilhão-dourado
56
dentes, não são uma exceção. Holland (2000, 2001), por exemplo, estudando 6 populações nativas (2 do Brasil, incluindo Rio Grande, 1 da Venezuela e 3 do sul da África) e 6 popula-ções invasoras (Golfo do México) do mexilhão marrom Perna perna encontrou altos níveis de variação gênica para dois locos de microssaté-lites, além de pouca ou nenhuma diferença en-tre as populações nativas e invasoras. Holland concluiu, então, que as populações no Golfo do México foram o resultado de um único evento de bioinvasão, que ele denominou “evento de captura de pool genético”, provavelmente, de-corrente de descarga de água de lastro. No caso do mexilhão-dourado, no en-tanto, os níveis de variação gênica são elevados, mas existe grande diferença genética entre as cinco populações amostradas. Estes resultados
sugerem que as populações estudadas, prova-velmente, foram fundadas a partir de muitos eventos de invasão advindos de diferentes re-giões geográficas. Este fenômeno de mistura genética de populações oriundas de diferen-tes localidades é chamado de Efeito Wahlund (Fig. 7). Nestes casos, as populações invasoras apresentam altos níveis de variação gênica (re-sultado da mistura de populações) e grandes diferenças genéticas entre elas (resultado das diferentes regiões). Outra característica do Efeito Wahlund são os desvios significativos ao esperado pelo equilíbrio de Hardy-Weinberg. A hipótese do equilíbrio de Hardy-Wein-berg postula que a frequência de alelos em uma geração é exatamente a mesma da geração ante-rior, ou seja, a frequência de alelos permanece a mesma, geração após geração, quando ocorre acasalamento ao acaso. A constância da frequên-cia de alelos – e consequentemente da composi-ção genotípica da população – significa que, na ausência de forças evolutivas específicas (seleção natural, deriva genética, mutação e migração) para modificar as frequências dos alelos, o me-canismo da herança mendeliana, por si só, man-tém as frequências dos alelos constantes e, assim, preserva a variabilidade genética (Hartl, 2000; Silva, 2001a). Numa população que é o resul-tado da mistura de muitas populações diferen-tes, as frequências dos alelos são diferenciadas, portanto, o equilíbrio de Hardy-Weinberg só pode ser alcançado depois de várias gerações de acasalamento ao acaso. Antes que isto aconteça, muitas das combinações de alelos esperadas não serão encontradas na população misturada. No trabalho com Limnoperna foram encontrados muitos desvios significativos das proporções es-peradas pelo equilíbrio de Hardy-Weiberg para as três populações brasileiras e as duas argen-tinas, o que é uma clara indicação de que elas foram fundadas pela mistura de populações de diferentes origens geográficas. O fato de que as populações invasoras, tanto no Brasil como na Argentina, estão dife-renciadas geneticamente conduz a outra conclu-são importante: a migração natural não é vetor mais importante do processo de invasão da es-pécie L. fortunei em curso nas águas brasileiras. Contudo, Kimura et al. (2003) estudando po-pulações de L. fortunei oriundas da Argentina, Japão, Tailândia e China encontraram, também,
Figura 6. O elevado nível de variação gênica da espécie Limno-perna fortunei pode ser visualizado no gráfico que relaciona o número de locos polimórficos (P) com a heterozigosidade (H) em vários grupos de organismos. A variação gênica de L. fortunei (triangulo amarelo) está muito acima daquela obser-vada para bivalves.
Figura 7. Efeito de mistura de populações que produz o au-mento da quantidade de variação da população invasora em relação às populações originais (Pop 1-4).

CAPÍTULO 3
Seção 1
57
grande diferenciação genética entre as popula-ções do Japão e Tailândia, bem como diferen-ças relevantes entre Argentina e Japão/China. Estes resultados indicam alto potencial de di-ferenciação gênica para a espécie L. fortunei, pelo menos em nível macrogeográfico.
Conclusão
As três conclusões principais das análises genéticas com as populações invasoras da espé-cie L. fortunei no Brasil podem ser resumidas como se segue: 1) o mexilhão-dourado invadiu as águas continentais brasileiras mais de uma vez; 2) as populações que invadiram o Brasil vieram de diferentes locais;
3) a migração natural não é vetor mais importante deste processo de bioinvasão. O estudo da genética das espécies in-vasoras tem sido pouco explorado, contudo, seu conhecimento pode fornecer informações importantes sobre os padrões de bioinvasão. As invasões biológicas são, geralmente, cons-tituídas de rápidos eventos evolutivos, resul-tando em populações que são geneticamente dinâmicas, tanto no espaço, quanto no tempo. Espera-se ter demonstrado neste capítulo que a utilização da abordagem genética, a partir do uso de marcadores moleculares, se cons-titui numa alternativa importante no esforço para entender e controlar o processo de bioin-vasão nos ecossistemas aquáticos continentais do Brasil.


Seção
2Biologia de moluscos invasores


61
1. Introdução
O conhecimento da morfologia externa e interna funcional dos moluscos bivalves inva-sores é de extrema importância para a defini-ção das espécies, do entendimento de seu papel ecológico nos ecossistemas, bem como para a compreensão da ação dos métodos de controle populacional sobre seus organismos. No Brasil, Limnoperna fortunei (Dunker, 1857) pode ser confundida com algumas espé-cies de Dreissenidae que ocorrem em água doce ou salobra, respectivamente, Mytilopsis lopesi Alvarenga & Ricci, 1989 e Mytilopsis leuco-phaeta (Conrad, 1831). As espécies invasoras Corbiculidae (C. fluminea, C. largillierti, C. fluminalis e Corbi-cula sp.) são comumente confundidas com os corbiculídeos nativos como Cyanocyclas limosa e Cyanocyclas paranensis e outras espécies des-te gênero que não foram ainda bem definidas e que são abundantes nas regiões norte e nordes-te do Brasil.
Visando diferenciar as espécies invasoras entre si e das demais espécies nativas e invaso-ras de bivalves de água doce que ocorrem no território brasileiro, num primeiro momento descrevemos sua morfologia externa (concha) e interna, e posteriormente, num quadro compa-rativo, apresentamos os aspectos morfológicos dos principais grupos de bivalves límnicos bra-sileiros.
2. Morfologia externa e interna de Limnoperna fortunei
2.1. Morfologia externa A concha de L. fortunei (Fig. 1) é forma-da por duas valvas iguais de contorno triangu-lar com a base alongada; alcançando o compri-mento de 2,5 a 3 cm, excepcionalmente 3,5 a 6 cm; a margem anterior é arredondada e baixa com umbos (protuberância dorsal em cada val-va) subterminais, não agudos, porém pequenos e arredondados situados um pouco acima da
CAPÍTULO 4
Bivalves invasores límnicos: morfologia comparada de
Limnoperna fortunei e espécies de Corbicula spp.
Maria Cristina Dreher Mansur

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
62
extremidade anterior e carenados em ambos os lados da concha; esta carena umbonal se-gue pelas laterais das valvas de forma oblíqua, terminando junto à extremidade posterior; margem dorsal ascendente, reta; na metade do comprimento, forma uma elevação arredonda-da (o topo), depois desce oblíqua e levemente curvada até a extremidade posterior que é ar-redondada e se situa próximo da base; margem ventral muito variável em vista lateral, desde reta até acentuadamente arqueada para dentro ou côncava, porém, em vista frontal ela é con-vexa.
A forma da concha lembra a de muitos mexilhões marinhos de tamanho mediano a pe-queno, como os de costões rochosos à beira mar, do gênero Brachidontes e o sururu do mangue do gênero Mytela. Externamente, lembra também as espécies do gênero Mytilopsis, como por exem-plo, o M. lopesi presente nos rios do baixo Ama-zonas e o de água salobra M. leocophaeta nativo da América do Norte, porém, recentemente, re-gistrado para as proximidades do porto de Recife (Souza et al., 2005). A concha é formada por três camadas. A mais externa é o perióstraco, depois vem a ca-mada prismática calcária, composta por cristais microscópicos dispostos perpendicularmente à superfície e por dentro a camada nacarada, com-posta por lamelas horizontais. O perióstraco é orgânico, relativamente liso, fino, translúcido, le-vemente amarelado ou castanho claro e brilhan-te. Apesar da superfície externa ser relativamente lisa, aparecem débeis linhas de crescimento que acompanham o contorno das bordas, são as li-nhas comarginais. Nas bordas da concha o peri-óstraco é mais espesso e virado para o interior da concha (Morton, 1973). A cor da concha (Fig. 2) é geralmente marrom escura, às vezes esverdeada acima e castanho claro abaixo da carena umbo-nal. A cor da superfície externa depende da com-posição do nácar interno da concha que é de cor púrpura acima e branco abaixo da carena umbo-nal (Morton, 1973). Comumente a área púrpu-ra é cruzada por raios brancos que partem dos umbos em direção ao declive posterior. Exempla-res mais velhos são geralmente mais escuros e os menores mais claros. Frequentemente aparecem albinos que, devido à ausência da cor púrpura interna, apresentam-se amarelados ou dourados em vista externa. Na superfície interna da concha (Fig. 3), podemos observar impressões ou cicatrizes mus-culares diferentes (heteromiários). Na região an-terior junto à cavidade sub umbonal aparece a impressão do músculo retrator anterior do bisso e do pé, que é pequena; ventralmente na frente, aparece a impressão do músculo adutor anterior também pequena e elíptica; abaixo do topo, uma impressão pequena correspondente ao músculo retrator posterior do pé; a seguir as impressões do músculo retrator posterior do bisso, que se apresenta dividida em duas partes: a primeira que é bilobada (Fig. 3 - RPB1) seguida da segun-
Figura 1. Morfologia externa da concha de Limnoperna for-tunei. Carena umbonal (C); topo (T); umbo (U). Escala: 1 cm. (Foto: M.C.D. Mansur).
Figura 2. Vista externa da concha de Limnoperna fortunei, com as variações de coloração: Acima, exemplar albino; no centro, exemplar rajado com raios internos sem a cor púrpura do nácar; em baixo, exemplar escuro, com o nácar interno de cor púrpura (região anterior à esquerda e posterior à direita). (Foto: M.C.D. Mansur).

CAPÍTULO 4
Seção 2
63
da simples (Fig. 3 - RPB2); logo atrás se encontra a maior impressão, correspondente ao músculo adutor posterior. A linha palial une as impressões do adutor anterior e do posterior paralelamente à borda ventral. É a linha onde os músculos da borda do manto aderem à concha. O ligamento é estreito e reto, composto por proteínas elásticas, conecta as valvas dorsalmente atrás dos umbos. Isto significa que o mexilhão é opistodético. Microscopicamente o ligamento é formado por duas camadas de proteínas elásticas que estão cobertas pelo perióstraco. Este tipo de ligamento (com três camadas) é encontrado ape-nas nos mitilídeos onde se inclui o L. fortunei. Em Dreissena polymorpha o ligamento é também opistodético, porém formado por apenas uma camada complexa (Morton, 1973). Abaixo do li-gamento existe a charneira que é muito fina, reta e desprovida de dentes. Não existe apófise, nem septo, nem chanfro ou entalhe bissal.
Como os demais mitilídeos marinhos ou de água salobra, L. fortunei apresenta fios de bis-so que permitem uma adesão firme ao substrato. Fios de bisso também estão presentes no gênero Mytilopsis (Dreissenidae) e em outros como nos gêneros Eupera e Byssanodonta (Sphaeriidae) na-tivos da América do Sul.
2.2. Morfologia interna de L. fortunei
Para observar a morfologia interna do mexilhão-dourado é necessário separar as valvas, descolando-se os músculos e o manto da super-fície interna valvar. Para não destruir os órgãos internos, é necessário introduzir um bisturi de ponta fina logo abaixo do topo, mas de forma inclinada para afastar primeiramente os múscu-los da superfície interna valvar e depois liberar o manto de sua adesão junto à linha palial. As valvas ainda ficam unidas dorsalmente pelo liga-mento. Retirando-se as duas valvas, observa-se o corpo do mexilhão-dourado revestido pelo manto e vários músculos. Mais internamente, o corpo do mexilhão apresenta uma massa visceral na parte dorsal e a cavidade do manto na região ventral. A massa visceral envolve os músculos e os órgãos responsáveis circulação, reprodução, digestão e filtração. Uma vista esquemática dos principais órgãos da cavidade do manto consta na Figura 4. Uma descrição detalhada da morfo-logia interna e funcional de L. fortunei também pode ser encontrada em Morton (1973). Muitos aspectos internos também se assemelham aos do mexilhão marinho Mytilus edulis descrito con-forme Renner & Cunha (1986) e Fox (2006a).
Figura 3. Interior da valva direita do Limnoperna fortunei com detalhe das impressões musculares: ligamento (L), linha palial (LP), músculo adutor anterior (MAA), músculo adutor poste-rior (MAP), perióstraco (PER), retrator anterior do bisso e pé (RABP), retrator posterior do bisso (RPB1), retrator posterior do bisso (RPB2), retrator posterior do pé (RPP), topo (T), umbo (U). (Desenho: M.C.D. Mansur).
Figura 4: Vista esquemática dos princi-pais órgãos da cavidade do manto do mexilhão-dourado: bisso (BI), borda do manto (BM), boca (BO), cavidade palial (CP), cavidade suprabranquial (CSB), de-mibrânquia externa (DBE), demibrânquia interna (DBI), divertículos digestivos (DD), ligamento (L), manto (M), músculo adutor anterior (MAA), membrana branquial (MB) músculo adutor posterior (MAP), palpo la-bial (PL), pé (P), retrator anterior do bisso e pé (RABP), retrator posterior do bisso (RPB1), retrator posterior do bisso (RPB2), retrator posterior do pé (RPP), sifão exa-lante (SE), sifão inalante (SI), umbo (U), corrente inalante (←), corrente exalante e direção das fezes (↑), percurso e direção das pseudofezes no sifão inalante (→→) (Desenho: M.C.D. Mansur).

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
64
2.2.1. Musculatura
No corpo de L. fortunei pode-se reconhe-cer vários músculos com diferentes funções (Fig. 4) Os músculos adutores que se estendem de uma valva a outra são responsáveis pela adução valvar (fechamento das duas valvas) e fixação do corpo à concha. O músculo adutor anterior é pequeno e se situa na frente, ventralmente; o músculo adutor posterior é grande, próximo dos sifões posteriores. Dorsalmente, abaixo do topo, existe um músculo grande e forte dividido em dois componentes (músculo retrator poste-rior do bisso “1” e músculo retrator posterior do bisso “2”) que são responsáveis pela adesão e sustentação do bisso. Na frente destes dois mús-culos existe um menor que é o músculo retrator posterior do pé, responsável pelos movimen-tos de retração do pé. Sob a cavidade umbonal destaca-se o músculo retrator anterior do bisso e do pé. Os dois músculos adutores são unidos por uma linha de pequenos músculos, a linha palial (Fig. 3), que acompanha a borda ventral das valvas; esta linha é íntegra, sem sinus palial (reentrância da linha palial). Na realidade estes pequenos músculos são a inserção dos músculos radiais das bordas do manto que servem para o controle e movimentação das dobras e aberturas do manto e do sifão.
2.2.2. O manto
É um órgão comum a todos os moluscos (Fig. 4). De uma maneira geral ele reveste e protege o corpo e é responsável pela formação da concha. No caso dos bivalves, o manto também tem outras funções importantes. Ele forma a cavidade palial ou do manto que é cheia de água, onde acontece a condução e seleção de partículas e, no caso dos Mitilídeos, em cada lobo, o man-to, internamente, abriga parte das gônadas. Nos demais bivalves límnicos dos gêneros Dreissena, Mytilopsis, Congeria que pertencem à família Dreissenidae, e as espécies de corbiculídeos, es-ferídeos e unionídeos, as gônadas estão presentes somente na massa visceral e não no manto. O manto é formado por dois lobos, cada um cobrindo a superfície interna da respectiva valva. No mexilhão-dourado, os lobos estão fun-didos dorsalmente acima do sifão exalante, entre os sifões exalante e inalante (apenas acima do ina-
lante) e separados ventralmente junto à abertura do pé. Através desta abertura, o pé distende-se e os fios do bisso exteriorizam-se. Junto à borda livre de cada lobo do manto existem três pregas: a prega externa é responsá-vel pela formação das camadas da concha; entre a externa e a mediana existe uma glândula que produz o perióstraco; a dobra interna é sensitiva e apresenta franjas que se expandem e retraem junto ao sifão inalante.
2.2.3. Cavidade palial Entre os dois lobos livres do manto, está a cavidade palial ou do manto (Fig. 4) que se estende para os lados da massa visceral. Divide--se em duas câmaras pelo diafragma ou septo branquial situado na linha dorsal das brânquias e entre os sifões: a câmara ventral é grande e facilmente visualizada, bastando separar as valvas e os lobos do manto. Contém a abertura inalante, as brânquias, o pé, os palpos labiais e a boca, órgãos que atuam na condução e sele-ção de partículas alimentícias, rejeição, troca de gases, locomoção e fixação. A câmara dor-sal ou suprabranquial (Fig. 4) é muito estreita e praticamente limitada ao canal suprabranquial, os espaços do sifão exalante e, dorsalmente, o ânus. Para ser visualizada é necessário romper a base dorsal das brânquias.
2.2.3.1. Sifões
São curtos formando simples aberturas (Fig. 4) não portadoras de tentáculos ou papilas: o sifão exalante é afunilado, formado somente pela fusão das pregas internas da borda do man-to que selam a câmara dorsal da cavidade palial, logo acima do sifão inalante; o sifão inalante é aberto e, portanto, podemos também denomi-ná-lo de abertura inalante. Isto significa que os dois lobos do manto não se fundem ventralmen-te e que o sifão inalante não é separado da fres-ta por onde se exterioriza o pé/bisso. As bordas deste sifão são onduladas e se distendem para fora da concha quando o animal está em ati-vidade de filtração. Em Corbicula, Dreissena e Mytilopsis forma-se um curto sifão inalante que é fechado ventralmente com curtos tentáculos na borda e separado da fresta do pé bissado. O sifão varia de comprimento conforme o gênero.

CAPÍTULO 4
Seção 2
65
2.2.3.2. Membrana branquial
Presente na base dorsal da abertura ina-lante (Fig. 4) tem o formato de uma meia-lua e é também denominada de septo branquial por Morton (1973). A borda distal desta membra-na apresenta pequenos tentáculos simples e um maior no centro com ramificações ou com um conjunto de tentáculos terminais (Figs. 5A e 5B). Pela associação com algumas espécies ma-rinhas, Morton (1973) relaciona a função desta membrana com a de uma válvula, presente em algumas espécies marinhas. Com a presença dos tentáculos sugerimos ter também função sensitiva. Esses costumam curvar-se e direcio-nar sua extremidade distal para a abertura ina-lante, dando a impressão de funcionarem como um periscópio.
2.2.3.3. Brânquias Dentro da cavidade do manto ou cavi-dade palial e de cada lado do pé estão as brân-quias (Fig. 4). Cada brânquia compreende duas demibrânquias, uma externa e outra interna. Cada demibrânquia é formada por uma lamela descendente e uma ascendente (Fig. 6). As uni-dades de cada lamela são os filamentos. Cada filamento possui cílios frontais na parte mais externa e cílios laterais entre os mesmos e de encontro aos cílios do filamento vizinho. Os cílios laterais promovem as correntes respirató-rias e de seleção das partículas. Os cílios fron-tais movem as partículas alimentícias ao longo da superfície branquial em direção ao sulco oral ventral (Fig. 6B). As demibrânquias são muito alongadas e semelhantes entre si, considerando a forma
muito achatada e o número de filamentos, no entanto as externas são maiores que as internas (Fig. 6A). As brânquias são do tipo primitivo (filibranquiados), típico dos mitilídeos. Os fila-mentos apresentam-se justapostos, mas indepen-dentes, apenas unidos por fusão ciliar formando as pontes ciliares ou junções interfilamentares, assim como na junção das demibrânquias ao man-to e à massa visceral. Também existem junções entre os filamentos da lamela ascendente e da descendente. Estas são as junções interlamela-res que mantem os filamentos opostos unidos, mas não por cílios, e sim por conexões muito pequenas de tecido. Neste tipo de brânquia os filamentos podem ser separados e individuali-zados com muita facilidade, o que não acontece com os outros bivalves nativos e invasores que ocupam as águas continentais brasileiras. Em Dreissena os filamentos, entre si e as lamelas ascendente e descendente, estão fortemente fusionados por tecidos de conexão formando verdadeiras lamelas branquiais (eulamelibrân-quios) assim como os Corbiculidae, Sphaerii-dae e Unionoida. Os cílios das brânquias formam correntes para selecionar partículas. As partículas aceitas são direcionadas por cílios formando correntes de aceitação que seguem até os palpos labiais e destes em direção à boca. Em L. fortunei ocor-rem correntes de aceitação, na base dorsal das demibrânquias somando três de cada lado da cavidade palial e mais duas nas margens das demibrânquias (Fig. 6A). Nestas, forma-se um verdadeiro canal longitudinal chamado sulco oral, por onde passam as correntes de aceita-ção (Fig. 6B). Seriam, portanto, 5 correntes de aceitação de cada lado do animal, o que au-menta a eficiência de filtração nesta espécie em
Figura 5. Detalhe dos tentáculos da membrana branquial situada na base dorsal da abertura inalante do Limno-perna fortunei: 5A, tentáculo central com ramificações simples; 5B, tentácu-lo central com ramificações terminais. Membrana branquial (MB), tentáculo ramificado (TR), tentáculo simples (TS) (Fotos: M.C.D. Mansur).

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
66
comparação com os demais bivalves límnicos invasores e nativos. Nos demais bivalves límni-cos nativos, em Corbicula e Dreissena, só existe um canal oral na demibrânquia interna, além de uma ou mais correntes de aceitação na re-gião dorsal das demibrânquias. No animal vivo é possível visualizar o movimento dos cílios e a formação das corren-tes de aceitação na superfície das brânquias. Para tal, é necessário usar bastante aumento na lupa cobrindo a brânquia com um filme de água. O batimento dos cílios e movimento das correntes pode ser visto onde a luz se reflete ou colocando pó de carmim ou grafite na su-perfície da brânquia. As partículas são movidas rapidamente sobre as brânquias em direção ao sulco oral ventral na margem distal das brân-quias (Fig. 6). Neste sulco as partículas selecio-nadas e aceitas são transportadas pelos cílios para frente do animal, passando das brânquias em direção aos palpos labiais e à boca, situada na região anterior do animal.
manto onde, uma vez envolvidas em muco, for-mam as pseudofezes que são levadas ao exterior da cavidade palial através de correntes próprias.
2.2.3.5. Pé e bisso É delgado e afilado na extremidade distal (Figs. 4, 7). Na fase de recrutamento e juvenil, o pé pode se distender muito, quase alcançando o comprimento da concha. A medida que o animal cresce e assume sua vida de epibionte (animal que vive fixo sobre o substrato) graças a produção de fios de bisso, o pé torna-se reduzido a ponto de não poder mais locomover o animal, quando ar-rancado do substrato. Os fios de bisso apresen-tam um aspecto córneo e são bastante resisten-tes. São secretados pela glândula do bisso que se localiza na parte dorsal do interior do pé, junto à massa visceral. O produto desta glândula se ex-terioriza por um canal bissal que se abre na face ventral posterior do pé. O bisso na realidade é um conjunto de fios. Cada fio é produzido indi-
2.2.3.4. Palpos labiais.
Dois de cada lado do corpo (Figs. 4 e 7), são muito alongados e estreitos, conectando-se e envolvendo a parte anterior das brânquias. As su-perfícies justapostas de cada par são pregueadas. Estas pregas recebem as partículas vindas dos sulcos orais das duas demibrânquias e, através de uma série de correntes, promovem a condução dos alimentos aceitos em direção à boca. As par-tículas rejeitadas são conduzidas em direção ao
Figura 6. Brânquia esquerda de Limno-perna fortunei: 6A, Esquema de um corte frontal da metade esquerda do animal com as setas demarcando as correntes de seleção de partículas nas superfícies branquiais e os pontos negros demarcan-do o direcionamento das partículas via oral; 6B, Detalhe da extremidade distal de um filamento branquial com o perfil do sulco oral, onde as partículas selecio-nadas são direcionadas por movimentos ciliares aos palpos labiais e à boca: cílios (CI), cavidade palial (CP), câmara supra-branquial (CSB), demibrânquia externa (DBE), demibrânquia interna (DBI), dobras da borda do manto (DBM), eixo central (EC), gônadas (G), junção interlamelar (JI), lamela ascendente (LA), lamela des-cendente (LD), manto (M), massa visceral (MV), pé (P), sulco oral (SO). (Desenho e foto de M.C.D. Mansur).
vidualmente e fixado pelo molusco ao substrato através de um pequeno disco adesivo ou botão. Este bisso permite ao animal fixar-se ao substrato e também se aglomerar, formando incrustações volumosas, os macroaglomerados.
2.2.3.6. Função da cavidade do manto
A principal função da cavidade do manto é a seleção ou filtração de partículas alimentícias em suspensão que entram pelo sifão inalante.

CAPÍTULO 4
Seção 2
67
Estas partículas integrantes do plâncton são movimentadas por cílios que se encontram nas superfícies do manto, e dos demais órgãos contidos na cavidade palial. Na câmara ventral, as correntes ciliares sobre as brânquias e entre os palpos labiais são de aceitação e rejeição; as correntes ciliares do manto que reveste a ca-vidade palial, incluindo as da massa visceral e do pé são todas de rejeição e servem para man-ter a limpeza desta cavidade (Morton, 1973). Estas correntes conduzem as partículas rejeita-das para a região posterior e, bem próximo da borda interna posterior do sifão inalante, são conduzidas para cima, em direção a abertura exalante (Fig. 4). Durante o trajeto de rejeição, as partículas são envolvidas por muco produ-zido por células especiais do manto, formando então as pseudofezes. Bem junto à área dorsal da abertura inalante, as correntes de rejeição de ambos os lados do manto se unem formando um cordão único de pseudofezes que é expeli-do de forma quase contínua quando o animal não é perturbado. Este tipo de liberação das pseudofezes pela região dorsal da abertura ina-lante difere dos bivalves nativos e das corbícu-las onde as pseudofezes se concentram na base inferior da abertura inalante sendo expelidas por movimentos rápidos e intermitentes por adução valvar. Na câmara dorsal do L. fortu-nei, todas as correntes são de limpeza e saem junto com as fezes pelo sifão exalante. Assim fezes e pseudofezes são expelidas sob a forma de cordões paralelos na mesma direção dorsal. Estudos realizados Jørgensen (1990), para medir o índice ou taxa de filtração de partículas em suspensão em vários bivalves re-velaram que L. fortunei pode ser considerado como um dos bivalves filtradores mais eficien-tes conhecidos até o momento. Assim é possí-vel que o alto dinamismo da atividade ciliar relacionado ao tamanho proporcionalmente grande da cavidade palial, somado a ampla superfície ciliar disponível nas brânquias e palpos labiais e ao número proporcionalmente alto (dez) de correntes de aceitação (Fig. 6A), possa explicar a alta eficiência deste molusco na seleção de partículas, o que colocaria em risco a biodiversidade planctônica e a fauna que depende desta. Mais detalhes nos Capítu-los 22 e 23.
2.3. Órgãos da massa visceral
Contem os órgãos internos responsáveis pela digestão, circulação, excreção e reprodução (Fig. 7). De uma maneira geral seguem o padrão das espécies marinhas do gênero Mytilus ou Perna.
2.3.1. Sistema digestivo
Inicia pela boca, seguindo pelo esôfago, estômago, glândula digestiva, saco do estile-te, intestino e reto, percorrendo a parte mais dorsal do animal (Fig. 7). As partículas acei-tas pelas brânquias chegam aos palpos labiais, pelo canal oral ventral de cada demibrânquia. Entram pela boca em forma de fresta, percor-rem o esôfago curto e ciliado em direção ao estômago alongado e pequeno. Na realidade o esôfago e o estômago também são áreas de seleção e condução de partículas. O estômago compreende internamente uma rede complexa de áreas de seleção formadas por sulcos, pregas e duas tiflossoles ciliadas, cuja função é separar as partículas orgânicas alimentícias das mine-rais indigestas. As partículas orgânicas seguem em direção aos divertículos digestivos e as re-jeitadas são conduzidas por um sulco à região posterior do estômago que se comunica com o intestino onde se formam as fezes. Existem várias aberturas da glândula digestiva no estô-mago. Inicialmente o intestino é associado ao saco do estilete e separado deste apenas pelas duas tiflossoles que se prolongam para dentro do estômago. O saco secreta o estilete cristali-no que é um bastonete transparente, gelatinoso e longo, que se projeta para dentro do estôma-go em direção ao escudo gástrico, situado do lado esquerdo. O estilete cristalino é composto por enzimas digestivas (Fox, 2006a) e, girando, ajuda a mover um cordão de muco com par-tículas alimentícias que vem do esôfago para dentro do estômago. O intestino compreende três regiões com aproximadamente o mesmo comprimento: o intestino anterior associado ao saco do estilete sai do estômago e segue até a altura do músculo adutor posterior, formando a primeira parte; volta para a frente da massa visceral que seria a segunda porção, o intestino mediano, e sobe formando a parte final, o in-testino terminal, também denominado de reto, que atravessa o pericárdio, penetra e atravessa

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
68
o ventrículo cardíaco, contorna o músculo adu-tor posterior, terminando no ânus próximo da abertura exalante. Os demais órgãos da massa visceral, também seguem o padrão dos mexilhões ma-rinhos. Mais detalhes sobre as estruturas e função das mesmas em todo o sistema digesti-vo, estão descritas detalhadamente por Mor-ton (1973) que comenta que o intestino médio e o saco do estilete em L. fortunei seriam ba-sicamente o mesmo que do mexilhão azul do mar Mytilus edulis descrito por Giusti (1971).
2.3.2. Sistema circulatório Está localizado dorsalmente, próximo do final do ligamento e abaixo do topo da concha. O coração é composto por um ventrículo único situado dentro da cavidade pericárdica que cons-titui o espaço do celoma que nos bivalves é muito reduzido (Fig. 7). A cada lado do ventrículo te-mos uma aurícula formada por finíssima mem-brana, que recebe o sangue (hemolinfa) oxigena-do oriundo do vaso eferente branquial que drena as brânquias. O ventrículo envolve o reto (intesti-no posterior) que também penetra na cavidade do pericárdio. Na frente do ventrículo encontra-se o bulbo aórtico que recebe a aorta anterior.
2.3.3. Sistema reprodutor
Encontra-se na massa visceral e no me-sossoma (Fig. 7), porém a maior parte das gô-nadas está no interior do manto. O mesossoma é uma bolsa de contorno arredondado situado de cada lado da massa visceral, entre o pé e o
músculo adutor posterior. Os folículos femini-nos, quando maduros, são amarelados. Os ma-chos, de cor cinza claro. As gônadas de cada lado do animal conectam-se com ductos que le-vam ao gonóporo que apresenta papila genital. Esta se localiza na cavidade suprabranquial.
2.3.4. Sistema excretor Formado por dois nefrídios muito alon-gados situados na massa visceral, acima da base dorsal das brânquias. Cada nefrídio se conecta com a cavidade pericárdica e desemboca no canal suprabranquial através de um nefridióporo lade-ado por uma fina papila urinária que se localiza na base posterior da papila genital. A glândula pericárdica também está associada ao sistema ex-cretor. É lobulada e está conectada às aurículas.
2.3.5. Sistema nervoso
Formado por gânglios, conetivos e nervos conforme o padrão geral dos bivalves. Um par de gânglios cerebrais está localizado nos lados do esôfago, e estão conectados ao estatocisto; um par de gânglios pediosos muito próximos no plano mediano do pé e um par visceral na base ventral do músculo adutor posterior.
3. Morfologia externa e interna de Corbicula fluminea (Müller, 1774)
3.1. Morfologia externa
A concha de Corbicula fluminea (Figs. 8,
Figura 7: Vista esquemática dos prin-cipais órgãos da massa visceral do mexilhão-dourado: ânus (A), abertura pé-bisso (APB), aurícula (AU), bisso (BI), boca (BO), demibrânquia externa (DBE), demibrânquia interna (DBI), divertículo digestivo (DD), estômago (E), esôfago (ES), gônadas (G), intestino anterior (IA), intestino médio (IM), intestino terminal ou reto (IT), músculo adutor anterior (MAA), músculo adutor poste-rior (MAP), membrana branquial (MB), mesossoma (ME), pé (P), palpos labiais (PL), pericárdio (PR), rim (R), sifão exa-lante (SE), sifão inalante (SI), ventrículo (V) (Desenho: M.C.D. Mansur).

CAPÍTULO 4
Seção 2
69
9, 10) é formada por duas valvas iguais de con-torno triangular quase equilátero com a região posterior projetada formando o rostro. O com-primento varia de 2 cm a 4,8 cm, excepcional-mente mais de 5 cm. A forma da concha lembra a de um berbigão marinho. É bastante robusta e vista de frente tem o contorno de um coração. Todas as margens são regularmente arredon-dadas com exceção da posterior, que forma o rostro cuja aresta posterior situa-se bem abai-xo da impressão do músculo adutor posterior (Ituarte, 1994). Os umbos situam-se na região dorsal, quase na metade do comprimento, e são bem salientes inflados e afilados no bico (Fig. 10). A superfície externa é ripada por estrias ou ondulações comarginais (paralelas à margem – são como estrias que acompanham a orienta-ção da borda ou margem ventral) relativamente espaçadas, uma por milímetro (Mansur et al., 2009) e recoberta por uma película conheci-da pelo nome de perióstraco que é liso, fino, translúcido, levemente amarelado, esverdeado ou, castanho claro e brilhante. Em áreas de maior dinamismo da água, a espécie costuma apresentar menor altura, coloração escura, sem brilho e muito erodida na área umbonal, fican-do à mostra a camada subjacente da concha que é branca e calcária. Abaixo do perióstraco, a estrutura interna da concha é composta por várias camadas de prismas que se organizam de forma oblíqua e cruzada conferindo muita resistência à concha (Mansur & Meier-Brook, 2000). Este tipo de organização se denomina porcelanizada e é particular dos bivalves da ordem Veneroida onde se enquadram todos os representantes da família Corbiculidae e a maioria dos bivalves marinhos como berbigões, mariscos brancos, maçambiques e outros.
Dorsalmente e logo atrás dos umbos, as duas valvas estão unidas pelo ligamento que é curto e saliente (Figs. 9, 10). Este compreende uma formação protuberante, de aspecto cór-neo, composto de proteína elástica. Por dentro da valva (Fig. 10) e abaixo do ligamento, as valvas apresentam uma superfície de articulação, mais ou menos reforçada, de-nominada charneira. Esta também auxilia na união das valvas pela presença de dentes que funcionam como uma engrenagem. Os dentes cardinais são curtos, dispostos radialmente e centralizados logo abaixo dos umbos, em nú-mero de três em cada valva. Os dentes laterais são alongados para frente e para trás dos um-bos e apresentam as superfícies serreadas. A charneira direita apresenta dois dentes laterais anteriores e dois dentes laterais posteriores. À esquerda, apresenta apenas um dente lateral alongado anterior e um posterior. Por dentro, a cor é branca ou levemente amarelada e sem brilho no centro, com a borda abaixo da linha palial, arroxeada ou marrom, com certo bri-lho. Na superfície interna das valvas veem-se ainda duas demarcações arredondadas: a im-pressão do músculo adutor anterior e a impres-são do músculo adutor posterior e uma linha que une estas duas impressões paralela à borda ventral, ou seja, a linha palial. Nesta espécie, como também nas outras corbículas invasoras, esta linha é íntegra, ou seja, sem a reentrância denominada sinus que é comum nas espécies nativas da mesma família, como Cyanocyclas limosa (Ver Capítulos 5 e 6). Em áreas de maior correnteza da água ou batimento de ondas provocadas pelo vento, a espécie costuma apresentar-se muito erodida na área umbonal, menor altura, coloração es-cura e sem brilho, em relação às populações de águas mais calmas. Este fenômeno, estudado e ilustrado por Ituarte (1994), também ocorre com as demais espécies de Corbicula e mesmo com as nativas Cyanocyclas.
3.2. Morfologia interna
Para observar as partes moles da Corbi-cula é necessário descolar da concha os dois músculos adutores com um bisturi (Fig. 11). Como as valvas e os adutores são muito ro-bustos, é muito difícil abrir a concha sem que-
Figura 8. Concha de Corbicula fluminea (Müller 1774): vista externa da valva esquerda (em baixo) e vista interna da valva direita (acima) (Foto: M.C.D. Mansur).

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
70
brar o bisturi. Assim recomenda-se anestesiar os moluscos dentro de um pote pequeno com água natural, cobrindo-os o suficiente para que e água mal ultrapasse o nível do animal. Sobre a superfície da água, espalhar cristais de men-tol até fechar a superfície. Levar o pote à par-te inferior do refrigerador e esperar 48 horas. Quando as valvas estiverem entreabertas e o pé não reagir ao toque, é momento de observar o corpo do animal, cortando os adutores e afas-tando as valvas. As valvas ainda ficam unidas dorsalmente pelo ligamento. O corpo é coberto pelo manto de tecido bastante fino e delicado. Este órgão apresen-ta dois lobos muito achatados que produzem a concha pela superfície externa e pelas bordas livres. As bordas destes lobos estão unidas dor-salmente, acima e abaixo dos sifões exalante e inalante, e separadas anteroventralmente junto à abertura do pé. Através desta abertura, o pé se distende para cavar e enterrar o animal no sedimento. Junto à borda livre de cada lobo do man-to existem as três pregas comuns aos demais bivalves e com as mesmas funções sensitivas e de formação das camadas da concha. Segun-do Martins et al. (2006), a prega mediana apresenta papilas distribuídas desde a região posterior, que diminuem progressivamente de tamanho até finalizarem próximo da região anterior; o manto de exemplares fixados de C. fluminea apresenta uma coloração branca, de aspecto leitoso o que contrasta com o manto branco, translúcido das outras duas espécies, C. largillierti e C. fluminalis. Como na maioria dos bivalves, o corpo
da Corbicula esta dividido numa massa visceral compacta na parte dorsal e a cavidade do man-to na região ventral.
3.2.1. Órgãos da cavidade do manto
Afastando os lobos do manto e cortando as uniões acima e abaixo dos sifões exalante e inalante, podemos visualizar a cavidade do manto ou palial (Fig. 11). Nesta cavidade estão expostos: o pé central e nas laterais, as brân-quias e os palpos labiais. Na região posterior, os sifões curtos e fechados apresentam bordas ladeadas por tentáculos dispostos em duas fi-leiras: uma de tentáculos filtradores e a outra de tentáculos sensoriais, conforme Britton & Morton (1982); Mansur & Meier-Brook (2000) e Martins et al. (2006). Segundo os úl-timos autores C. fluminea apresenta um denso anel de pigmentos ao redor dos sifões inalante e exalante. O pé é volumoso, de perfil arredondado, mais espesso na borda ventral. Na fase adulta o pé não apresenta glândula bissogênica nem forma fios de bisso. Quando recruta ou muito jovem, o pé produz um fio mucoso e pegajo-so que serve para aderi-lo ao substrato ou em grãos de areia, para não ser arrastado pela cor-renteza. Não é firme como o fio de bisso do mexilhão-dourado, mas levemente elástico. Este fio logo desaparece e não é mais produ-zido quando o animal ultrapassa os 5 mm de comprimento. O pé da Corbicula está mais ha-bilitado para enterrar o animal no sedimento do que para a locomoção. Quando observamos os bivalves nativos Unionoida rastejarem para
Figura 9 e 10. 9, Vista externa esquemática da valva esquerda da corbícula asiática Corbicula fluminea (Müller 1774), (Modifi-cado de FOX, 2006b, por M.C.D. Mansur). 10, Vista interna esquemática da valva direita de Corbicula fluminea (Modificado de Fox, 2006b, por M.C.D. Mansur).

CAPÍTULO 4
Seção 2
71
águas mais profundas durante a seca, geral-mente a Corbicula caminha para trás em cír-culos quando tenta fugir do mesmo problema (Mansur & Meier-Brook, 2000). Cada brânquia (uma de cada lado) é for-mada por duas demibrâquias (Fig. 12). Ao con-trário do mexilhão-dourado, a demibrânquia interna é maior e mais ventral e a demibrâquia externa, menor e mais dorsal. As brânquias da C. fluminea, como a grande parte dos bivalves tem o filamento como unidade. No entanto, os filamentos em Corbicula são unidos lateralmente por tecidos e as lamelas descendente e ascendente por sua vez também conectadas por pontes de tecidos, formando assim brânquias verdadeiras ou eulamelibrânquias, bastante reforçadas. Se-eulamelibrânquias, bastante reforçadas. Se-gundo Martins et al. (2006) em C. fluminea a demibrânquia externa atinge a metade da altu-ra da interna. Em relação ao tipo de incubação predomina e endobranquial, ou seja, a grande parte dos embriões se concentra no marsúpio da
demibrânquia interna. Esporadicamente são en-contrados na externa, mas em geral em pequeno número. Além disto, as grandes superfícies cilia-das nas brânquias, também tem a função de filtrar as partículas alimentícias que são inala-das junto com a corrente branquial através da abertura inalante. Os cílios formam correntes de aceitação e rejeição (Fig. 12). As correntes de aceitação e que conduzem o alimento para a região frontal estão situadas na base dorsal das duas lamelas descendentes e no sulco ven-tral presente apenas na borda da demibrânquia interna. Estas correntes seguem em direção aos palpos labiais, onde são conduzidas à boca. Os palpos labiais, um par de cada lado da boca, situados logo atrás do músculo adutor anterior, são relativamente curtos e o contorno ventral arredondado. Apresentam superfícies internas com pregas que orientam e selecionam as partí-culas que vem das brânquias.
Figuras 11 e 12. 11, Órgãos da cavidade palial de Corbicula fluminea e parcialmente da massa visceral, após remoção do lobo esquerdo do manto: ânus (A), aurícula (AU), bico (B), borda do manto (BM), boca (BO), cavidade palial (CP), demibrânquia exter-na (DBE), demibrânquia interna (DBI), divertículo digestivo (DD), dente lateral posterior (DLP), gônadas (G), intestino terminal (IT), ligamento (L), lobo direito do manto (LDM), músculo adutor anterior (MAA), músculo adutor posterior (MAP), marsúpio (MAR), massa visceral (MV), nefrídio (N), pé (P), palpos labiais (PL), pericárdio (PR), sifão exalante (SE), sifão inalante (SI), umbo (U), ventrículo (V). (Modificado de FOX, 2006b, por M.C.D. Mansur). 12, Esquema de um corte frontal da metade esquerda do animal, com as setas demarcando as correntes de seleção de partículas nas superfícies branquiais e os pontos negros demar-cando o direcionamento das partículas via oral: cavidade palial (CP), câmara suprabranquial (CSB), demibrânquia externa (DBE), demibrânquia interna (DBI), dobras da borda do manto (DBM), eixo central (EC), septos interlamelares (SI), lamela ascendente (LA), lamela descendente (LD), manto (M), massa visceral (MV), pé (P), sulco oral (SO). (Desenho: M.C.D. Mansur).

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
72
3.2.2. Órgãos da massa visceral
O digestório, com um trato que se inicia pela boca, com dois palpos labiais de cada lado, o esôfago achatado, um estômago com muitas do-bras e aberturas que se comunicam com a glân-dula digestiva (Mansur & Meier-Brook, 2000), um saco do estilete cristalino e o intestino, que depois de formar várias voltas, atravessa o ven-trículo cardíaco finalizando junto ao sifão exa-lante; o sistema circulatório, com um ventrículo e duas aurículas laterais contidos no pericárdio; o sistema excretor, representado por dois nefrídios; o glandular, com duas glândulas pericárdicas; o nervoso, formado por gânglios pares, situados no pé, acima do músculo adutor posterior e atrás do músculo adutor anterior, conectados por nervos; o sistema reprodutor compreende duas gônadas, uma de cada lado do animal, formadas por fo-lículos masculinos, femininos e hermafroditas, que ocupam grande parte da massa visceral. Se-gundo Martins et al. (2006), as gônadas em C. fluminea apresentam uma coloração verde acin-zentada e, segundo Fox (2006b), correspondem aos ovários formados por folículos arborescentes; às vezes, parte das gônadas apresenta-se de cor branca brilhante que, segundo o mesmo autor, correspondem aos testículos. Os folículos escoam seu conteúdo por go-nodutos que se exteriorizam por um único poro de cada lado do animal. Os espermatozoides são biflagelados e quando maduros formam mórulas espermáticas que são liberadas ao meio ambien-te. A fecundação é interna, ocorrendo dentro dos tubos aquíferos situados nas brânquias, junto ao marsúpio. Como visto acima, as brânquias, além da troca gasosa, também servem para incubação dos embriões junto a áreas especiais situadas nas demibrânquias internas, denominadas marsúpios (ocasionalmente podem ser encontrados embriões fora do marsúpio, nas demibrânquias externas). Nos guias para estudos morfológicos e de dissecação de Corbicula fluminea oferecidos por Britton & Morton (1982) e Fox (2006b) podemos encontrar mais detalhes sobre a anatomia desta espécie. Estudos morfológicos foram desenvolvi-dos por Mansur & Meier-Brook (2000), visando o relacionamento filogenético das famílias Corbi-culidae e Sphaeriidae, mostram dados compara-tivos da concha e partes moles, distintivos entre a invasora C. fluminea e a nativa Cyanocyclas limo-sa. Martins (2004) e Martins et al. (2004, 2006)
trouxeram grande contribuição para a distinção das três espécies do gênero Corbicula encontra-das na bacia do Lago Guaíba no Rio Grande do Sul (C. fluminea, C. largillierti e C. fluminalis), com base na morfologia da concha e das partes moles, tais como: aberturas branquiais, brân-quias e localização dos marsúpios.
4. Morfologia externa e interna de Corbicula largillierti (Philippi, 1844)
4.1. Morfologia externa
A concha C. largillierti (Fig. 13) é mais frágil e menor que as outras corbículas citadas (não passa de 2,5 cm de comprimento), umbos baixos e arredondados, sem rostro, ondula-ções comarginais na superfície externa, mui-to finas e justapostas (duas a três por mm na parte mediana da concha), perióstraco de cor esverdeada palha ou arroxeada, pouco brilho. Por dentro, na parte central a cor é roxa escura ou tendendo ao roxo ou cinza, sem brilho, a borda também arroxeada, levemente brilhante abaixo da linha palial que não apresenta sinus palial. Facilmente confundida com a nativa Cyanocyclas, quando vista externamente. Comparando a concha, Callil & Mansur (2002) observaram diferenças significativas na relação entre peso e comprimento das espécies C. fluminea e C. largillierti coletadas na parte norte do Pantanal do Mato Grosso. A primeira apresenta-se relativamente maior em termos de comprimento e peso que a segunda.
Figura 13. Concha de Corbicula largillierti: vista externa da valva esquerda (abaixo) e vista interna da valva direita (acima) (Foto: M.C.D. Mansur).

CAPÍTULO 4
Seção 2
73
4.2. Morfologia interna
Considerando os dois sifões de C. lar-gillierti, segundo Martins (2004) e Martins et al. (2006), apenas o exalante apresenta o anel externo com pigmentos, havendo uma concen-tração maior na região do diafragma que di-vide os sifões. Em C. largillierti, os tentáculos no sifão inalante apresentam uma organização aparente em três fileiras desencontradas, sendo a mais externa também de tentáculos menores, assim como observado nas outras duas espé-cies. Massa visceral: Em C. largillierti a epi-derme de um modo geral é bastante translú-cida e fina, permitindo visualizar as gônadas que também ocupam grande parte da massa visceral. C. largillierti apresenta a incubação endobranquial, com as larvas ocupando toda extensão das demibrânquias internas; as exter-nas são muito curtas não alcançando a metade da altura das internas (Martins et al., 2006). Também invasiva, mas pouco sabemos sobre a biologia e o comportamento desta espécie, uma vez que raros investimentos de pesquisa foram direcionados a mesma na América do Sul. Esta espécie não ocorre na América do Norte.
5. Morfologia externa e interna de Corbicula fluminalis (Müller, 1774)
5.1. Morfologia externa
A concha de Corbicula fluminalis (Mül-ler, 1774) (Fig. 14) é robusta, quase equilateral, inflada, altura igual ou maior que o compri-mento (<3 cm), linhas comarginais semelhan-tes às de C. fluminea, porém menos espaçadas (uma a cada 0,7 mm) e baixas, charneira muito arqueada, umbos muito altos e praticamente centrados (quase na metade do comprimento), sem rostro, cor mais escura, tendendo ao roxo forte internamente e o perióstraco do castanho esverdeado ao negro.
5.2. Morfologia interna Em espécimes de C. fluminalis, os ten-táculos do sifão inalante estão organizados em
três ou quatro fileiras, sendo uma fileira ex-terna de tentáculos menores e as fileiras mais internas com tentáculos maiores, entremeados por tentáculos de tamanhos intermediários. Na base dos sifões não ocorre um segundo anel externo pigmentado, como descrito para as outras espécies, C. fluminea e C. largillierti. Porém, assim como nas outras duas espécies, apresentam um anel interno de pigmentos lo-calizado junto à base dos tentáculos do sifão inalante. Em C. fluminalis, segundo Martins et al. (2006), a epiderme é bastante translúcida e fina, permitindo igualmente visualizar as gô-nadas que também ocupam grande parte da massa visceral como em C. largillierti. Quanto às brânquias, os mesmos autores (op. cit.) cons-tataram que C. fluminalis apresenta incuba-ção branquial nas quatro brânquias, portanto tetrabranquial, e as demibrânquias externas atingem a metade da altura das demibrânquias internas. A altura das demibrânquias externas coincide com as das C. fluminea. Korniushin (2004) também revisou a es-pécie quanto aos aspectos da concha, da ana-tomia interna, tipo de incubação branquial e caracteres reprodutivos, redefinindo a espécie com base no estudo de exemplares oriundos da Ásia Central, África e China. Após estes estu-dos, o mesmo autor (op. cit.) considerou que a espécie de Corbicula não incubadora de larvas, identificada anteriormente por Morton (1982) como C. fluminalis, apresenta caracteres anatô-micos e reprodutivos diferentes e foi por ele re-considerada como C. cf japonica (Prime, 1864).
Figura 14. Vista externa e interna da concha de Corbicula flu-minalis (Müller, 1774). (Foto: M.C.D. Mansur).

M.C.D. Mansur
Bivalves invasores límnicos: morfologia comparada de Limnoperna fortunei e espécies de Corbicula spp.
74
6. Q
uad
ro c
om
para
tivo
da
morf
olo
gia
do L
. fo
rtunei
e o
utr
as
esp
éci
es d
e b
ivalv
es lím
nic
os
bra
sile
iros
nat
ivos
e in
vaso
res.
Car
acte
res
Fam
ília
Gên
eros
e s
pp.
Form
a co
ncha
Cha
rnei
raB
isso
na
fase
ad
ulta
Tip
o de
B
rânq
uia
Cor
rent
es
acei
taçã
o na
s br
ânqu
ias
Tip
o de
la
rva
Cic
lo la
rval
PéA
bert
ura
inal
ante
Lim
nope
rna
fort
unei
mit
iloid
eSi
mpl
es li
sa
sem
den
tes
sim
filib
rânq
uia
10ve
licon
cha
No
plan
cton
redu
zido
aber
ta
Dre
isse
nida
eM
ytilo
psis
lo
pesi
mit
iloid
eC
om s
epto
e
apóf
ise
sim
eula
mel
ibrâ
nqui
a?
velic
onha
Incu
bado
no
mar
súpi
o at
é jo
vem
redu
zido
fech
ada
Cor
bicu
lidae
Cor
bicu
latr
iang
ular
Den
tes
card
inai
s e
late
rais
ser
read
osnã
oeu
lam
elib
rânq
uia
4ve
licon
cha
No
mar
súpi
o at
é ve
liger
dese
nvol
vido
fech
ada
Cor
bicu
lidae
Cya
nocy
clas
tria
ngul
arD
ente
s ca
rdin
ais
e la
tera
is s
erre
ados
não
eula
mel
ibrâ
nqui
a4
velic
onch
aN
o m
arsú
pio
até
jove
mde
senv
olvi
dofe
chad
a
Spha
eriid
ae
Pisi
diu
Spha
eriu
m
Mus
culiu
m
pequ
ena
oval
ada
Den
tes
card
inai
s e
late
rais
sim
ples
não
eula
mel
ibrâ
nqui
a 4
velic
onch
aN
o m
arsú
pio
até
jove
m
dese
nvol
vido
fech
ada
Spha
eriid
ae
Eup
era
Byss
anod
onta
pequ
ena
oval
ada
Den
tes
card
inai
s e
late
rais
sim
ples
sim
eula
mel
ibrâ
nqui
a 4
velic
onch
aE
m s
acos
em
brio
nári
os
no m
arsú
pio
até
jove
m
dese
nvol
vido
fech
ada
Hyr
iidae
Dip
lodo
n C
asta
lia
Trip
lodo
nPa
xyod
on
Pris
odon
vari
ável
Esq
uiso
dont
enã
oeu
lam
elib
rânq
uia
6gl
oquí
dio
Mar
súpi
o at
é gl
oquí
dio
e pa
rasi
to p
eixe
dese
nvol
vido
gera
lmen
te
aber
ta
Myc
etop
odid
aeA
nodo
ntit
esM
ycet
opod
aM
ycet
opod
ella
L
eila
H
aasi
caFo
ssul
aM
onoc
oncy
laea
Ba
rtle
ttia
vari
ável
Sem
ou
com
de
ntes
car
dina
isnã
oeu
lam
elib
rânq
uia
6la
sídi
oM
arsú
pio
até
lasí
dio
e pa
rasi
to p
eixe
vari
ável
gera
lmen
te
aber
ta

75
Com a introdução de espécies de molus-cos límnicos invasores de origem asiática no Brasil, a busca por métodos de controle po-pulacional, visando à minimização dos danos econômicos, tem sido alvo de investimentos prioritários pelos setores de geração de energia hidrelétrica e termoelétrica, bem como de cap-tação e tratamento de água para abastecimento público. Os programas de controle populacio-nal de espécies de moluscos invasores envolvem desde métodos simples, como remoção mecâ-nica, até métodos mais complexos de contro-le físico (UV e Ultrassom), químico (cloração, ozonização, aplicação de sulfato de cobre, etc.) e biológico (bactérias malacopatogênicas). A eficácia do controle populacional dos moluscos invasores requer o conhecimento pré-vio das características biológicas e ecológicas da espécie-alvo. E o primeiro passo é reconhe-cer a identidade taxonômica da espécie. Mesmo sendo as espécies invasoras muito peculiares e distintas das espécies nativas, a identificação taxonômica errônea é muito comum. Deve-se
considerar que nem sempre o controle é reali-zado por biólogos, pois diferentes profissionais atuantes na área de meio ambiente coordenam atividades em plantas industriais e nos sistemas naturais nos quais estas estão inseridas. Além disso, é cada vez mais preocupante o estado de conservação das espécies nativas de bivalves límnicos, que vêm sofrendo redução po-pulacional em decorrência de diferentes ativida-des humanas impactantes aos ecossistemas aquá-ticos continentais, são elas: destruição de habitats marginais, assoreamento, alteração da qualidade da água, eutrofização, barramentos, alterações populacionais e das rotas migratórias da ictiofau-na, bem como a competição por recursos e habi-tats com espécies de moluscos límnicos invasores. Com base nessas considerações, tomou-se a iniciativa de elaborar uma chave dicotômica para a identificação e diferenciação dos bivalves invasores introduzidos no Brasil dos bivalves na-tivos encontrados no país. A determinação específica de espécies de bivalves de água doce não é uma tarefa fácil.
CAPÍTULO 5
Identificação e diferenciação dos bivalves límnicos invasores
dos demais bivalves nativos do Brasil
Daniel Pereira Maria Cristina Dreher Mansur
Daniel Mansur Pimpão

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
76
Muitas espécies foram descritas para o Brasil e o grande número de sinônimos é um fato marcante, sendo necessária a revisão taxo-nômica deste grupo.
Estruturas e orientação da concha para estudo
As principais estruturas da concha uti-lizadas na diferenciação de gêneros e espécies de moluscos nativos e invasores encontram-se discriminadas na Tabela I e na Figura 1. A orientação da concha para a tomada de medidas biométricas para fins de diferen-ciação estatística dos gêneros e espécies é re-alizada da seguinte forma: toma-se a concha (Fig. 6A), orientando-se os umbos para cima e a lúnula para frente; o ligamento e o escudo devem ficar voltados para o observador; assim, a valva direita será correspondente ao lado di-reito do observador e vice-versa; portanto, a região anterior estará voltada para frente e a
posterior, voltada para o observador; em caso de dúvidas, observar o ligamento que é sem-pre posterior; para tomar as medidas de uma concha é necessário visualizar duas linhas bá-sicas, x e y; após marcar um ponto central em cada músculo adutor, unindo-se estes pontos se obtêm a linha x; bipartindo o umbo, a linha y passa pelo bico, perpendicularmente a linha x, e finaliza na borda ventral. A largura da concha (z) é tomada com as duas valvas fecha-das (Fig. 6B). Para a diferenciação estatística de espé-cies e gêneros com base em medidas da concha a análise multivariada de discriminantes canô-nicas (ADC) é um dos procedimentos mais uti-lizados (ver Capítulo 6). Pereira et al. (2011a) diferenciou 11 espécies do gênero Anodontites com base no comprimento, altura, largura e posição do umbo. Pimpão et al. (2012) também comparou a morfometria de conchas larvais de espécies amazônicas de Hyriidae por meio do uso da ADC.
Tabela I. Termos malacológicos citados na chave dicotômica.
Termo Definição
Apófise (a) Projeção aderida ao septo, junto ao compartimento mais interno da cavidade umbonal, onde se fixa e se sustenta o músculo retrator do pé.
Borda da concha Extremidade marginal das valvas.
Bico (b) Extremidade, ápice da prodissoconcha e umbo. Pode ter direcionamento dife-rente do umbo (Fig. 1B, D e F).
Bisso (bi)Filamento orgânico ou conjunto de fios fabricados pela glândula do bisso no interior do pé do animal e serve para fixá-lo ao substrato. Presente em Limno-perna fortunei (Fig. 2A), Mytilopsis e Eupera spp.
Borda prismática (bp) Extremidade marginal interna das valvas, geralmente de aspecto visivelmente diferente do restante da superfície interna das valvas em Mycetopodidae.
Capacete Prodissoconcha claramente separada e projetada formando uma tampa ou ca-pacete.
Carena (ca)Saliência externa que parte do umbo em direção a região póstero-inferior. Linha longitudinal que marca a troca de inclinação da superfície externa da concha (Fig. 1A).
Cavidade Subumbonal (csu) Reentrância interna dorsal, sob os umbos (Fig. 1B).
Charneira (c)Região de articulação das valvas, situada dorsalmente sob os umbos. Apresen-ta-se sob diversos aspectos: lisa, estreita espessa, provida de dentes, fossetas ou lamelas cujas disposições variam muito (Fig. 1B, D e F).
Concha oval Semelhante a um ovo com dois eixos de simetria.
Concha ovoide Semelhante a um ovo com um eixo de simetria.
Concha romboide Semelhante a uma elipse com bordas paralelas como um polígono paralelogra-mo (com quatro lados iguais e paralelos).
Concha triangular Semelhante a um triângulo.
Dentes cardinais (dc) Dentes situados na charneira, logo abaixo dos umbos (Fig. 1D e F).
Dente pseudocardinal anterior (dpa) e posterior (dpp)
Dentes da charneira de formato irregular, de difícil individualização e distinção entre eles, em alguns casos. Característicos de Hyriidae (Fig. 1B).
Dentes laterais (dl) Dentes alongados em forma de lamela, situados na frente e atrás dos cardinais (Fig. 1B, D e F).

CAPÍTULO 5
Seção 2
77
Tabela I. Termos malacológicos citados na chave dicotômica.
Termo Definição
Equilateral Quando as valvas são simétricas em relação a um plano vertical que passa pelos umbos e perpendicular à charneira.
Escultura umbonal (eu) Ornamentação externa da concha em forma de proeminências. Pode ser radial (Fig. 1A), concêntrica e reticular.
Escudo Depressão externa achatada na região posterior delimitada pela carena (Fig. 6B).
Expansão alada Projeção em forma de asa situada nas extremidades anterior/dorsal ou poste-rior/dorsal (Fig. 4A).
Fosseta Cavidade na charneira para o encaixe de dente da valva oposta.
Heterodonte
Tipo de charneira que apresenta dentes cardinais medianos em número máxi-mo de três, com fossetas correspondentes em cada valva e ainda apresentam antero e posteriormente aos cardinais, dentes laterais alongados (Presente em Corbiculidae e Sphaeriidae) (Fig. 1D e F).
Impressão dos músculos anteriores (iaa) e posteriores (iap)
Cicatrizes deixadas na superfície interna da concha, onde os músculos do ani-mal se fixam na mesma (Fig. 1B, D e F).
Impressão dos músculos adutores Duas impressões grandes de contorno ovalado; uma situada na região anterior e a outra na região posterior da concha (Fig. 1B, D e F).
Impressão dos músculos dorsais Impressões pequenas na cavidade umbonal.
Inequilateral Quando as valvas são assimétricas em relação a um plano vertical que passa pelo umbo e perpendicular à charneira.
Ligamento (l)
Estrutura orgânica que une as duas valvas dorsalmente e força a abertura das valvas quando do relaxamento dos músculos adutores. Firme, mas elástica quando o animal está vivo; quebradiça depois do animal morto, com o resse-camento da concha. Geralmente externo (Fig. 1A, B, D e F).
Linhas ou estrias comarginais (ec) Marcas ou linhas na superfície externa da concha que delimitam a margem de um estágio de crescimento. Paralelas à margem da valva (Fig. 1A e E).
Linha palial (lp) Linha paralela aos bordos da margem ventral da concha, deixada pela aderência do manto. Liga as duas impressões dos músculos adutores (Fig. 1B, D, F e 3A).
Lúnula Depressão externa pequena na região anterior logo após os umbos (Fig. 6B).
Nodosidade Saliência em forma de nódulo na superfície externa da concha (Fig. 4B).
Pé em forma de machado Pé típico de bivalves Unionoida, em forma de lâmina de machado (Fig. 5A).
Pé com extremidade intumescida Pé alongado e com a extremidade intumescida, funcionando como uma ânco-ra; não laminar distinto dos demais bivalves (Fig. 5B).
Perióstraco Camada orgânica, fina, que reveste a superfície externa da concha. Protege as camadas calcárias da dissolução pela água.
Rostro Projeção da região posterior da valva, que a torna inequilateral (Fig. 4C).
Prodissoconcha (pc) Concha embrionária dos bivalves, secretada pela larva e geralmente mantida nos indivíduos adultos (Fig. 1E).
Septo (s) Projeção que divide a cavidade umbonal de cada valva em dois compartimen-tos. Nele se fixa e se sustenta o músculo adutor anterior. (Fig. 2B).
Sinus palial ou seio palial (sp)Reentrância de linha palial em forma de “U” ou “V” invertido, junto a impres-são do músculo adutor posterior (Presente em Leila blainvilliana (Fig. 3A, B) e Neocorbicula).
Umbo (u)Elevação na região dorsal de cada valva (Fig. 1A, D, E e F). Quanto a sua loca-lização podem ser denominados frontais, terminais e subterminais. É a região mais antiga da concha, geralmente gasta em Unionoida.
Diversidade de bivalves límnicos no Brasil: espécies invasoras e nativas
São conhecidos 22 gêneros de bivalves lí-mnicos para o Brasil, sendo que dois destes es-tão representados apenas por espécies invasoras
neste país, são eles: Limnoperna (Mytilidae) e Corbicula (Corbiculidae). A Tabela II apresenta um resumo da classificação destes gêneros. A seguir, é apresentada a primeira chave para determinação e diferenciação das espécies de bivalves límnicos invasores dos demais bi-valves nativos citados para o Brasil.

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
78Figura 1. Estruturas da concha de bivalves límnicos: 1A, vista externa da valva esquerda e 1B, interna da valva direita de Castalia sp; 1C, vista interna da valva direita de Anodontites patagonicus; 1D, vista interna da valva direita de Corbicula fluminea; 1E, vista exter-na da valva esquerda de Pisidium sp. e vista interna das valvas esquerda e direita de Pisidium sp. Prodissoconcha (pc); umbo (u); bico (b); escultura umbonal (eu); carena (ca); declive dorsal (dd); estrias ou linhas comarginais (ec); ligamento (l); cavidade subumbonal (csu); charneira (c); dente cardinal (dc); dente pseudocardinal anterior (dpa); dente pseudocardinal posterior (dpp); dente lateral (dl); impressão do músculo adutor anterior (iaa); impressão do músculo adutor posterior (iap); linha palial (lp); borda prismática (bp). (Desenho: M.C.D. Mansur).
A relação das espécies de moluscos cita-das para o Brasil encontra-se discriminada na Tabela III. São conhecidas 114 espécies de mo-luscos bivalves límnicos com registros para o Brasil, sendo que destas, apenas 5 espécies (4% das espécies de bivalves límnicos citados para o país) são invasoras (Fig. 7). No entanto, o im-pacto deste pequeno número de espécies inva-
soras aliado aos impactos ambientais de origem antrópica tem contribuído para a diminuição das populações das espécies nativas. Dentre os impactos decorrentes das espécies invasoras podemos citar a competição por espaço, re-cursos alimentares, bem como pela formação de macroaglomerados bioincrustantes, quando considerado o mexilhão-dourado.

CAPÍTULO 5
Seção 2
79
Figura 2. Concha mitiloide: 2A, fios de bisso (bi) em Limnoperna fortunei; 2B, septo (s) e apófise (ap) em Mytilopsis. Escala: 0,5 cm. (Foto e desenho: M.C.D. Mansur).
Figura 3. Representação da linha palial (lp) e do sinus palial (sp) em conchas do gênero Leila (Unionoida, Mycetopodidae): 3A, concha com a linha palial demarcada e 3B, não demarcada. A seta indica o sinus palial. Escala: 1 cm. (Fotos: M.C.D. Mansur).
Figura 4. Estruturas da concha: 4A, expansão alada em Hyriidae; 4B, nodosidades sobre o perióstraco de Hyriidae; 4C, rostro em Corbiculidae. Escala: 1 cm. (Fotos: M.C.D. Mansur e D. Pimpão).
Figura 6. Orientação da concha de um bivalve: 6A, borda dorsal, ventral, anterior e posterior; 6B, vista da largura da concha (z). Posição do umbo (pu) determinada pela distância entre o bico do umbo e o limite anterior perpendicular à linha Y (altura) (Desenhos: M.C.D. Mansur).
Figura 5. Pé de bivalves límnicos: 4A, pé típico dos bivalves límnicos, em forma de machado representado por um indivíduo do gênero Leila; 4B, pé de com extremidade intumescida, típico do gênero Mycetopoda (Fotos: M.C.D. Mansur).

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
80
Tabela II. Gêneros de bivalves límnicos nativos e invasores citados para o Brasil.
Gêneros OrigemClasse BIVALVIA Linné, 1758
Subclasse PTERIOMORPHIA Beurlen, 1944 Ordem MYTILOIDA Férrusac, 1822
Superfamília MYTILOIDEA Rafinesque, 1815Família MYTILIDAE Rafinesque, 1815
Subfamília Mytilinae Rafinesque, 1815Gênero Limnoperna Rochebrune, 1882 Invasor
Subclasse PALAEOHETERODONTA Newell, 1965Ordem UNIONOIDA Stoliczka, 1871
Superfamília UNIONOIDEA Fleming, 1828Família HYRIIDAE Fleming, 1828
Subfamília Hyriinae Swainson, 1840Gênero Diplodon Spix, 1827 NativoGênero Castalia Lamarck, 1819 NativoGênero Prisodon Schumacher, 1817 NativoGênero Paxyodon Schumacher, 1817 NativoGênero Triplodon Spix, 1827 Nativo
Superfamília ETHERIOIDEAFamília MYCETOPODIDAE Gray, 1840
Subfamília Anodontitinae Modell, 1942Gênero Anodontites Bruguière, 1792 Nativo
Subgênero Anodontites s.s. NativoSubgênero Lamproscapha Nativo
Subfamília Mycetopodinae Adams & Adams, 1856Gênero Mycetopoda Orbigny, 1835 Nativo
Subfamília Monocondylaeinae Modell, 1942Gênero Monocondylaea Orbigny, 1835 NativoGênero Fossula Lea, 1870 NativoGênero Haasica Strand, 1932 NativoGênero Tamsiella Haas, 1931 Nativo
Subfamília Leilinae Morretes, 1949Gênero Leila Gray, 1840 Nativo
Subfamília BartlettiinaeGênero Bartlettia H. Adams, 1866 Nativo
Subclasse HETERODONTA Neumayr, 1884Ordem VENEROIDA Adams & Adams, 1858
Subordem ARCTICINA Newell, 1965Superfamília SPHAERIOIDEA Rafinesque, 1820
Família SPHAERIIDAE Rafinesque, 1820Subfamília Sphaeriinae Baker, 1927
Gênero Musculium Link, 1807 NativoGênero Sphaerium (Scopoli, 1777) NativoGênero Pisidium C. Pfeiffer, 1821 Nativo
Subfamília Euperinae Heard, 1965Gênero Eupera Bourguignat, 1854 Nativo
Superfamília CORBICULOIDEAFamília CORBICULIDAE Gray, 1847
Gênero Corbicula Megerle Von Mühlfeld, 1811 InvasorGênero Cyanocyclas Blainville, 1818 NativoGênero Polymesoda Rafinesque, 1828 Nativo
Superfamília DREISSENOIDEAFamília DREISSENIDAE
Gênero Mytilopsis Conrad, 1857 Nativo
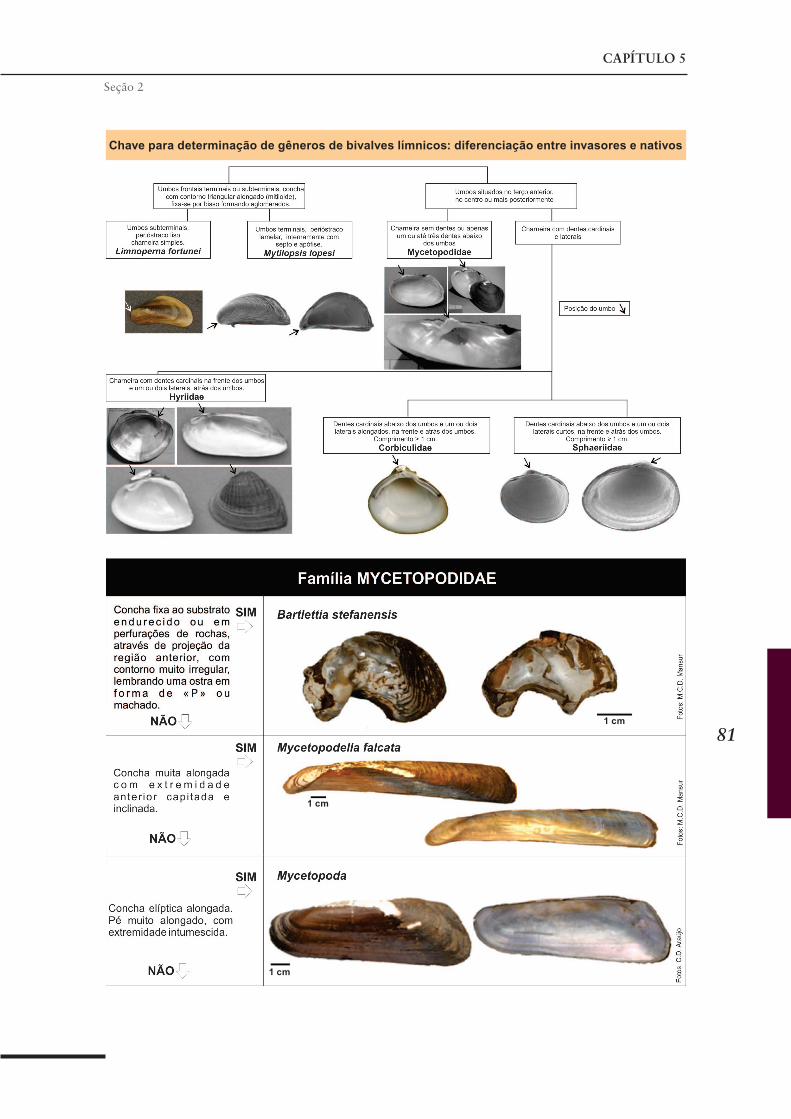
CAPÍTULO 5
Seção 2
811 cm
1 cm
1 cm
Chave para determinação de gêneros de bivalves límnicos: diferenciação entre invasores e nativos

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
82
1 cm
1 cm
1 cm 1 cm
1 cm
1 cm 1 cm

CAPÍTULO 5
Seção 2
83
1 cm
1 cm
1 cm
1 cm
1 cm
1 cm
0,5 cm

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
84
0,5 cm
3 mm
1 mm
2 mm
1 mm
0,5 cm

CAPÍTULO 5
Seção 2
85
Dentre as famílias mais representativas em número de espécies podemos citar Hyriidae (52 espécies; 45% das espécies de bivalves lím-nicos), Mycetopodidae (32; 28%) e Sphaeriidae (20; 17%), sendo todas estas espécies represen-tantes da malacofauna nativa (Fig. 8). Dentre os gêneros mais representativos em número de espécies podemos citar Diplo-don (37 espécies; 32% das espécies de bivalves límnicos), Anodontites (14; 12%), Castalia (11; 10%), Eupera e Pisidium (9; 8%), e Monocon-dylaea (7; 6%), sendo todas estas espécies re-presentantes da malacofauna nativa (Fig. 9). O Brasil apresenta oito grandes bacias hidrográficas dos rios: 1, Amazonas; 2, Tocan-tins/Araguaia; 3, do Atlântico Norte/Nordeste; 4, São Francisco; 5, do Atlântico Leste; 6, Pa-raná/Paraguai; 7, Uruguai; e 8, bacia do Atlân-tico Sul/Sudeste (Fig. 10). O maior número de espécies é verificado nas bacias: Paraná/Para-guai (51 espécies; 22% das espécies de bivalves límnicos citados para o Brasil), Atlântico Sul/Sudeste (43; 18%), Uruguai (42; 18%) e Ama-zonas (40; 17%) (Fig. 11). No entanto, esses números podem não representar a diversidade real destas bacias, já que as bacias Amazonas, Atlântico Norte/Nordeste, Atlântico Leste, To-
cantins/Araguaia e São Francisco carecem de coletas e inventários malacofaunísticos. Das 114 espécies citadas para o Brasil, 45 carecem de revisão taxonômica (35% das espécies citadas). As maiores dificuldades são verificadas quanto à identidade taxonômica das espécies do gênero Diplodon que apresen-tam ampla variação morfológica. A falta de
Figura 7. Número e percentual de espécies de moluscos bival-ves límnicos nativos e invasores citadas para o Brasil.
Chave para determinação das espécies do gênero Corbicula
Adaptado de Mansur et al., 2011; Fotos: C.D. de Araújo.

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
86
Figura 9. Número e percentual de espécies de moluscos bivalves límnicos conhecidos para os gêneros citados para o Brasil.
Figura 8. Número e percentual de espécies de moluscos bivalves límnicos conhecidos para as famílias citadas para o Brasil.

CAPÍTULO 5
Seção 2
87
conhecimento da real identidade taxonômica das espécies é um agravante no que diz respei-to à conservação, sendo que algumas espécies constam em listas vermelhas. Apenas 1% das espécies citadas para o Brasil está enquadrada na categoria Criticamente Em Perigo (CEP) na lista vermelha brasileira de espécies ameaça-das; 11%, na categoria Em Perigo (EP) e 9% em Vulnerável (VU) (Fig. 12). As demais espécies não constam em listas, mas isso não significa que não estejam ameaçadas, pois as listas não foram revisadas desde suas publicações. Além disso, muitas destas espécies têm sofrido ame-aças consideráveis, destacando-se a construção de novas barragens, assoreamento, desmata-mento, destruição de habitats, poluição e a ampliação da dispersão das espécies de bival-ves invasoras. Desta forma, 46 espécies (40,3% das espécies de bivalves límnicos citadas para o Brasil) são sugeridas como candidatas à ava-liação criteriosa em nova revisão da lista de es-pécies ameaçadas da fauna brasileira (Tab. III).
As demais espécies carecem de dados e, assim como as espécies candidatas, deverão ser alvo de pesquisas quanto aos seus níveis populacio-nais, preferências ambientais e distribuição ge-ográfica atual no país. Considerando dados históricos da ma-lacofauna límnica do estado de São Paulo (Ihering, 1893; Vaz et al., 1983, 1985, 1986b, 1987, 1992; Teles et al., 1991; França et al., 2007; Suriani et al., 2007) e o diagnóstico apresentado no Capítulo 19 para a bacia do Rio Tietê, além de registros de coleções de museus, é possível estabelecer uma cronolo-gia, a qual ilustra a diminuição da riqueza de moluscos bivalves límnicos ao longo dos anos na bacia do Rio Tietê (Fig. 13). Cabe ressaltar que a construção das barragens situadas nes-ta bacia ocorreu no período da ditadura mili-tar. Naquela época, o licenciamento ambien-tal de empreendimentos hidrelétricos era um processo novo no país, carecendo de estudos criteriosos sobre a biota aquática e do estabe-
Figura 10. Bacias hidrográ-ficas do Brasil. Fonte: ANA (http://www.aneel.gov.br/area.cfm?id_area=104).
Figura 11. Número e per-centual de espécies de moluscos bivalves límni-cos citados nas bacias hi-drográficas do Brasil.
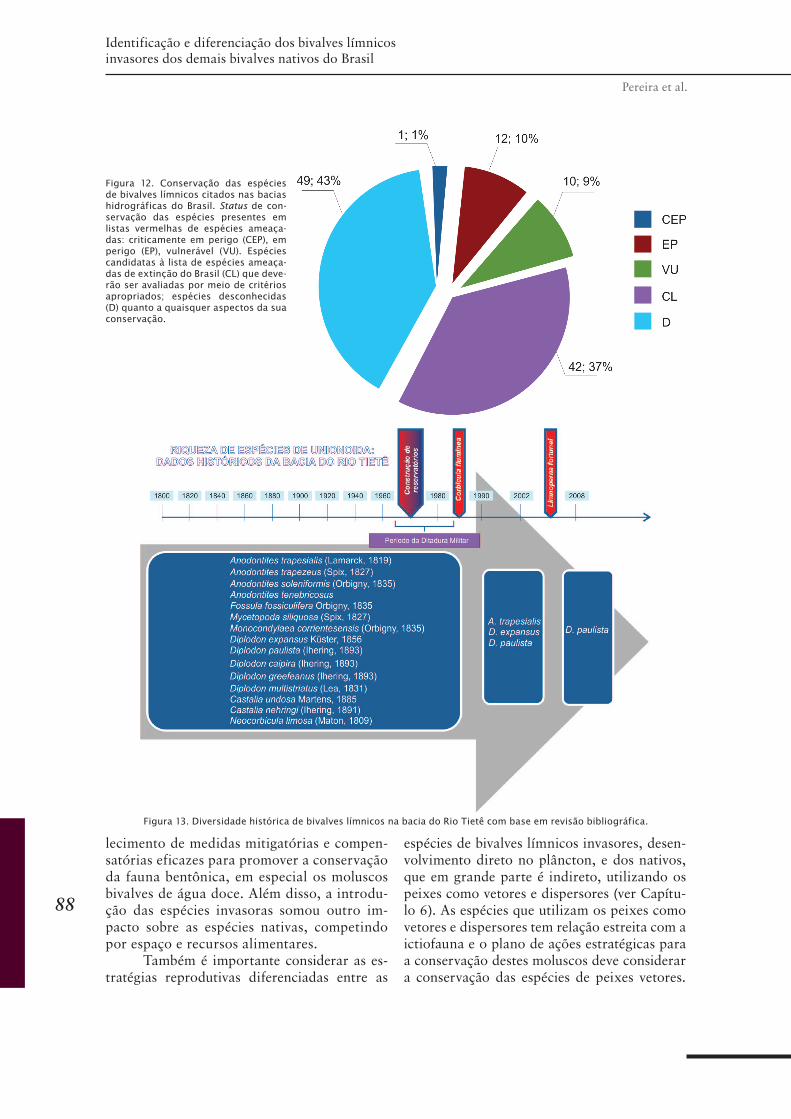
Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
88
Figura 12. Conservação das espécies de bivalves límnicos citados nas bacias hidrográficas do Brasil. Status de con-servação das espécies presentes em listas vermelhas de espécies ameaça-das: criticamente em perigo (CEP), em perigo (EP), vulnerável (VU). Espécies candidatas à lista de espécies ameaça-das de extinção do Brasil (CL) que deve-rão ser avaliadas por meio de critérios apropriados; espécies desconhecidas (D) quanto a quaisquer aspectos da sua conservação.
Figura 13. Diversidade histórica de bivalves límnicos na bacia do Rio Tietê com base em revisão bibliográfica.
lecimento de medidas mitigatórias e compen-satórias eficazes para promover a conservação da fauna bentônica, em especial os moluscos bivalves de água doce. Além disso, a introdu-ção das espécies invasoras somou outro im-pacto sobre as espécies nativas, competindo por espaço e recursos alimentares. Também é importante considerar as es-tratégias reprodutivas diferenciadas entre as
espécies de bivalves límnicos invasores, desen-volvimento direto no plâncton, e dos nativos, que em grande parte é indireto, utilizando os peixes como vetores e dispersores (ver Capítu-lo 6). As espécies que utilizam os peixes como vetores e dispersores tem relação estreita com a ictiofauna e o plano de ações estratégicas para a conservação destes moluscos deve considerar a conservação das espécies de peixes vetores.

CAPÍTULO 5
Seção 2
89
No entanto, a maioria dos vetores é desconhe-cida, o que demonstra a carência de dados que possam dar suporte à conservação de muitas espécies de moluscos bivalves. A construção de barragens, além de provocar uma alteração am-biental ao transformar um rio em lago, muitas vezes afeta a estrutura da ictiofauna (composi-ção e abundância de peixes), além de suas rotas
migratórias. Portanto, é de extrema importân-cia a pesquisa científica sobre a relação larva gloquídio e ou lasídio com o peixe vetor e dis-persor, visando descrever detalhadamente o ci-clo parasitário no peixe e o desenvolvimento da larva do bivalve até sua fase adulta bentônica, bem como a importância das rotas migratórias de peixes na dispersão desses moluscos.
Tabela III. Espécies de bivalves límnicos citados para as bacias hidrográficas do Brasil com base principalmente nas seguintes obras: Haas (1969); Ihering (1890, 1893, 1910); Ituarte & Mansur (1993); Mansur et al. (1987, 1988, 1991, 1994, 2008a, 2011a); Mansur & Meier-Brook (2000); Mansur & Pereira (2006); Mansur & Silva (1990); Mansur & Valer (1992); Ortmann (1921); Pereira et al. (2000a); Pimpão et al. (2008, 2012); Pimpão & Mansur (2009), Simone (2006), Scarabino & Mansur (2007), além do exame de coleções científicas. Status (S) de conservação das espécies: vulnerável (VU), em perigo (EP), criticamente em perigo (CEP), conforme Amaral et al (2008). Candidata à lista de espécies ameaçadas de extinção do Brasil (CL), espécies as quais são desco-nhecidas (D) quanto a quaisquer aspectos quanto a sua conservação *carece de revisão taxonômica.
Família/Gênero Espécie Bacias Hidrográficas Ameaças S
Mytilidae
Lymnoperna Rochebrune, 1882
Limnoperna fortunei(Dunker, 1857)
Paraná/Paraguai, Uruguai e Atlântico Sul/Sudeste. - D
Mycetopodidae
Anodontites Bruguière, 1792
Anodontites (A.) crispatus Bruguière, 1792*
Amazonas, Paraná/Paraguai, Atlântico Norte/Nordeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
CL
Anodontites (A.) elongatus (Swainson, 1823)
Amazonas, Paraná/Paraguai, Uruguai.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
VU
Anodontites (L.) ensiformis (Spix, 1827)
Amazonas, Tocantins/Araguaia, Paraná/Paraguai, Uruguai.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
VU
Anodontites (A.) ferrarisi (Orbigny, 1835)
Uruguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
EP
Anodontites (A.) iheringi (Clessin, 1882)*
Atlântico Sul/Sudeste.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
EP
Anodontites (A.) lucidus (Orbigny, 1835)
Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
CL
Anodontites (A.) moricandi (Lea, 1860)*
São Francisco.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Anodontites (A.) obtusus (Lamarck, 1819)*
São Francisco, Atlântico Norte/Nordeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
CL
Anodontites (A.) patagonicus
(Lamarck, 1819)*
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
D
Anodontites (A.) schombergianus
(Sowerby, 1870)*
Amazonas e Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
CL
Anodontites (A.)
soleniformis (Lamarck, 1819)*
Amazonas, São Francisco, Paraná/Paraguai.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
VU
Anodontites (A.) tenebricosus (Lea, 1834)*
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
VU
Anodontites (A.)
trapesialis (Lea, 1860)
Amazonas, Tocantins/Araguaia, Atlântico Norte/Nordeste, São Francisco, Atlântico Leste, Paraná/
Paraguai, Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal e poluição.
VU

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
90
Tabela III. Continua...
Família/Gênero Espécie Bacias Hidrográficas Ameaças S
Mycetopodidae
Anodontites (A.) trapezeus (Spix, 1827)
Amazonas, Tocantins/Araguaia, São Francisco, Atlântico Leste, Paraná/
Paraguai, Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal e poluição.
EP
Mycetopoda Orbigny, 1835
Mycetopoda legumen (Martens, 1888)
Uruguai, Atlântico Sul/Sudeste.
Erosão marginal, poluição, barramento e barreiras à piracema.
VU
Mycetopoda siliquosa (Spix, 1827)
Amazonas, Tocantins/Araguaia, Atlântico Norte/Nordeste, São Francisco, Atlântico Leste, Paraná/
Paraguai, Uruguai, Atlântico Sul/Sudeste.
Erosão marginal, poluição, barramento e barreiras à piracema. VU
Mycetopoda soleniformis Orbigny, 1835
Paraná/Paraguai, Uruguai.Erosão marginal, poluição,
barramento e barreiras à piracema. CL
Mycetopodella Marshall, 1927
Mycetopodella falcata (Higgins, 1868)
Amazonas. Erosão marginal, poluição, barramento e barreiras à piracema. CL
Monocondylaea Orbigny 1835
Monocondylaea corrientesensis Orbigny, 1834
Paraná/Paraguai, Uruguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Monocondylaea costulata Moricand, 1858
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
CL
Monocondylaea
franciscana (Moricand, 1837)
São Francisco.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Monocondylaea jaspidea Hupé, 1857
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
CL
Monocondylaea minuana Orbigny, 1835
Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
D
Monocondylaea
paraguayana Orbigny, 1835
Paraná/Paraguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
VU
Monocondylaea parchappi Orbigny, 1835
Paraná/Paraguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Fossula Lea, 1870
Fossula fossiculifera (Orbigny, 1835)
São Francisco, Atlântico Leste, Paraná/
Paraguai, Uruguai.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
EP
Tamsiella Haas, 1931
Tamsiella amazonica Bonetto, 1972*
Amazonas.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Tamsiella schroeteriana (Lea, 1852)*
Amazonas.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Haasica Strand, 1931
Haasica balzani (Ihering, 1893)
Paraná/Paraguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Leila Gray, 1838
Leila blainvilliana (Lea, 1834)
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema e espécies invasoras.
EP
Leila esula (Orbigny, 1835)
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema.
VU
Bartlettia H. Adams 1866
Bartlettia stefanensis Moricand, 1856
Paraná/Paraguai. Assoreamento, erosão marginal, poluição e barramento. VU
Hyriidae -

CAPÍTULO 5
Seção 2
91
Tabela III. Continua...
Família/Gênero Espécie Bacias Hidrográficas Ameaças S
Hyriidae -
Diplodon Spix, 1827
Diplodon (D.) aethipos (Lea, 1860)
Uruguai, Atlântico Sul/Sudeste.
- D
Diplodon (D.) berthae Ortman, 1921
Uruguai, Atlântico Sul/Sudeste.
- D
Diplodon (D.) besckeanus (Dunker, 1848)
Atlântico Leste. Assoreamento, erosão marginal e poluição. CL
Diplodon (D.) caipira (Ihering1893)*
Paraná/Paraguai. - EP
Diplodon (D.) delodontus (Lamarck, 1919)*
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
- D
Diplodon (D.) dunkerianus (Lea, 1956)*
Atlântico Leste. Assoreamento, erosão marginal e poluição.
EP
Diplodon (D.) ellipticus (Wagner, 1827)*
São Francisco, Atlântico Leste, Atlântico Sul/
Sudeste (?)- D
Diplodon (D.) expansus (Küster, 1856)*
Paraná/Paraguai, Atlântico Leste, Atlântico Sul/Sudeste (?).
Assoreamento, erosão marginal e poluição.
VU
Diplodon (D.) rhombeus (Spix, 1827)*
São Francisco, Paraná/Paraguai.
- D
Diplodon (D.) granosus (Bruguière, 1792)*
Atlântico Leste, Atlântico Sul/Sudeste. - D
Diplodon (D.) imitator Ortman, 1921*
Atlântico Sul/Sudeste. - D
Diplodon (D.) martensi (Ihering, 1893)*
Atlântico Leste, Uruguai, Atlântico Sul/Sudeste. - D
Diplodon (D.) multistratus(Lea, 1834)*
Atlântico Leste, Atlântico Sul/Sudeste. - D
Diplodon (D.) obsolescens (Baker, 1913)
Amazonas. - D
Diplodon (D.)
parallelopipedon (Lea, 1834)
Paraná/Paraguai, Uruguai. - D
Diplodon (D.) parodizi Bonetto, 1960
Paraná/Paraguai, Uruguai. - D
Diplodon (D.) paulista (Ihering, 1893)*
Paraná/Paraguai. - D
Diplodon (D.) piceus (Lea, 1860)
Uruguai. - D
Diplodon (D.) rhuacoicus (Orbigny, 1835)*
Uruguai, Atlântico Sul/Sudeste.
- D
Diplodon (D.) suavidicus (Lea, 1856)
Amazonas. - D
Diplodon (D.) vicarius Ortmann, 1821*
Paraná/Paraguai. - D
Diplodon (D.) wynami (Lea, 1860)*
Uruguai, Atlântico Sul/Sudeste.
- D
Diplodon (R.) burroughianus
(Lea, 1834)Paraná/Paraguai, Uruguai. Assoreamento, erosão marginal,
poluição e barramento. CL
Diplodon (R.) charruanus Orbigny, 1835*
Uruguai, Atlântico Sul/Sudeste. - D
Diplodon (R.) deceptus Simpson, 1914 sensu
Ortmann, 1921 Atlântico Sul/Sudeste. - D

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
92
Tabela III. Continua...
Família/Gênero Espécie Bacias Hidrográficas Ameaças S
Hyriidae -
Diplodon (R.) funebralis (Lea, 1860)*
São Francisco, Paraná/Paraguai.
Assoreamento, erosão marginal e poluição, barramento. CL
Diplodon (R.) koseritzi (Clessin, 1882)
Atlântico Sul/Sudeste.Assoreamento, erosão marginal
e poluição, barramento e espécies invasoras.
CEP
Diplodon (R.) hildae Ortman, 1921
Atlântico Sul/Sudeste. - -
Diplodon (R.) hylaeus (Orbigny, 1835)
Amazonas, Tocantins/Araguaia e Paraná/
Paraguai (?).
Assoreamento, erosão marginal e poluição, barramento.
CL
Diplodon (R.) iheringi (Simpson, 1900)
Atlântico Sul/Sudeste. Assoreamento, erosão marginal e poluição, barramento. EP
Diplodon (R.) peraeformis (Lea, 1860)
Uruguai. Assoreamento, erosão marginal e poluição, barramento. CL
Diplodon fontainianus (Orbigny, 1835)*
Paraná/Paraguai. Assoreamento, erosão marginal, poluição, barramento.
EP
Diplodon greeffeanus Ihering, 1893*
Paraná/Paraguai.
Diplodon pfeifferi (Dunker, 1848)
Atlântico Leste.Assoreamento, erosão marginal,
poluição, barramento.EP
Diplodon paranensis (Lea, 1834)*
Paraná/Paraguai. Assoreamento, erosão marginal, poluição, barramento. CL
Diplodon rotundus Wagner, 1827*
Tocantins/Araguaia, São Francisco e Paraná/Paraguai.
Assoreamento, erosão marginal, poluição, barramento. EP
Diplodon uruguayensis (Lea, 1860)*
Uruguai. Assoreamento, erosão marginal, poluição, barramento.
CL
Castalia Lamarck, 1819
Castalia ambigua Orbigny, 1835*
Amazonas e Paraná/Paraguai.
Assoreamento, erosão marginal, poluição, barramento, barreiras
à piracema e explotação.CL
Castalia duprei (Récluz, 1842)
Tocantins/Araguaia.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia inflata Orbigny, 1835
Amazonas, Paraná/Paraguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia martensi (Ihering, 1891)
Uruguai, Atlântico Sul/Sudeste.
Assoreamento, erosão marginal, poluição, barramento e barreiras à piracema e espécies invasoras.
CL
Castalia nehiringi (Ihering, 1893)
Paraná/Paraguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia orbignyi (Deville & Hupé, 1850)
Amazonas.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia psammoica (Orbigny, 1835)
Uruguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia quadrata (Sowerby, 1869)*
Amazonas.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia schombergiana Sowerby, 1869*
Amazonas.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia sulcata (Krauss, 1849)*
Amazonas.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
CL
Castalia undosa Martens, 1885
Paraná/Paraguai.Assoreamento, erosão marginal,
poluição, barramento e barreiras à piracema.
EP
Paxyodon Schumacher, 1817
Paxyodon syrmathophorus
(Meuschen, 1781)
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento, barreiras
à piracema e explotação.CL

CAPÍTULO 5
Seção 2
93
Tabela III. Continua...
Família/Gênero Espécie Bacias Hidrográficas Ameaças S
Hyriidae -
Prisodon Schumacher, 1817
Prisodon obliquus Schumacher, 1817*
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento, barreiras
à piracema e explotação.CL
Triplodon Spix, 1827
Triplodon corrugatus (Lamarck, 1819)
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento, barreiras
à piracema e explotação.CL
Triplodon chodo
Mansur & Pimpão, 2008
Amazonas. Poluição e espécies invasoras. CL
Dreissenidae
MytilopsisConrrad, 1858
Mytilopsis lopesiAlvarenga & Ricci, 1989
Amazonas, Tocantins/Araguaia.
Assoreamento, erosão marginal, poluição, barramento
e espécies invasoras.CL
Corbiculidae -
Cyanocyclas Blainville, 1818
Cyanocyclas amazonica (Prime, 1870)*
Amazonas. - D
Cyanocyclas brasiliana (Deshayes, 1854)*
Amazonas. - D
Cyanocyclas limosa (Maton, 1809)*
Paraná/Paraguai, Uruguai. Assoreamento, erosão marginal, poluição e espécies invasoras.
CL
Cyanocyclas paranensis (d’Orbigny, 1835)*
Paraná/Paraguai. Assoreamento, erosão marginal, poluição e espécies invasoras. CL
Corbicula Megerle Von Mühlfeld, 1811
Corbicula fluminalis (Müller, 1774)
Atlântico Sul/Sudeste. - D
Corbicula fluminea (Müller, 1774)
Amazonas, Tocantins/Araguaia, Atlântico Norte/Nordeste, São Francisco, Atlântico Leste, Paraná/
Paraguai, Uruguai, Atlântico Sul/Sudeste.
- D
Corbicula largillierti
(Philippi, 1844)
Tocantins/Araguaia, Atlântico Norte/
Nordeste, São Francisco, Atlântico Leste, Paraná/
Paraguai, Uruguai, Atlântico Sul/Sudeste.
- D
Corbicula sp.* Atlântico Sul/Sudeste. - D
Polymesoda Rafinesque, 1828
Polymesoda sp.*Amazonas, Atlântico
Norte/Nordeste. - D
Sphaeridae - D
Eupera Bourguignat, 1854
Eupera klappenbachi Mansur &
Veitenheimer, 1975Atlântico Sul/Sudeste. Poluição, drenagem de banhados
e remoção de macrófitas. D
Eupera platensis Doello-Jurado, 1921
Uruguai, Atlântico Sul/Sudeste.
Poluição, drenagem de banhados e remoção de macrófitas.
D
Eupera elliptica Ituarte & Dreher-
Mansur, 1993Paraná/Paraguai. Poluição, drenagem de banhados
e remoção de macrófitas. D
Eupera doellojuradoi Klappenbach, 1962
Uruguai. Poluição, drenagem de banhados e remoção de macrófitas. D
Eupera bahiensis (Spix & Wagner, 1827)
Atlântico Leste.Poluição, drenagem de banhados
e remoção de macrófitas.D
Eupera tumida (Clessin, 1879)
Amazonas, São Francisco, Atlântico Leste, Paraná/Paraguai.
Poluição, drenagem de banhados e remoção de macrófitas.
D

Pereira et al.
Identificação e diferenciação dos bivalves límnicos invasores dos demais bivalves nativos do Brasil
94
Tabela III. Continua...
Família/Gênero Espécie Bacias Hidrográficas Ameaças S
Sphaeridae - D
Eupera simoni (Jousseaume, 1889)
Amazonas, Paraná/Paraguai. Poluição, drenagem de banhados e remoção de macrófitas.
D
Eupera guaraniana Ituarte, 1994
Uruguai, Atlântico Sul/Sudeste.
Poluição, drenagem de banhados e remoção de macrófitas.
D
Eupera iguazuensis Ituarte, 1989
Paraná/Paraguai. Poluição, drenagem de banhados e remoção de macrófitas. D
Sphaerium (Scopoli, 1777)
Sphaerium cambaraense Mansur, Meier-Brook
& Ituarte, 2008
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
Poluição e drenagem de banhados. D
Musculium Link, 1807
Musculium argentinum (D’Orbigny, 1835)
Uruguai, Atlântico Sul/Sudeste. Poluição e drenagem de banhados. D
Pisidium C. Pfeiffer, 1821
Pisidium vile Pilsbry, 1897
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste. Poluição e drenagem de banhados. D
Pisidium dorbignyi Clessin, 1879
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste. Poluição e drenagem de banhados. D
Pisidium sterkianum Pilsbry, 1897
Amazonas, Paraná/Paraguai, Uruguai,
Atlântico Sul/Sudeste.Poluição e drenagem de banhados. D
Pisidium punctiferum (Guppy, 1867)*
Amazonas, Tocantins/Araguaia, São Francisco,
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
Poluição e drenagem de banhados. D
Pisidium taraguyense Ituarte, 2000
Paraná/Paraguai, Uruguai, Atlântico Sul/Sudeste.
Poluição e drenagem de banhados. D
Pisidium forense Meier Brook, 1967
Paraná/Paraguai, Atlântico Sul/Sudeste. Poluição e drenagem de banhados. D
Pisidium globulus* Atlântico Sul/Sudeste. Poluição e drenagem de banhados. D
Pisidium bejumaeH.B. Baker, 1930*
Amazonas. Poluição e drenagem de banhados. D
Pisidium boliviense Sturany, 1900*
Amazonas. Poluição e drenagem de banhados. D
Recomendações
Abaixo são listadas algumas ações prioritá-rias necessárias para minimizar os efeitos das espé-cies invasoras sobre as espécies nativas, assim como ações complementares visando à conservação dos bivalves nativos por meio da indução de projetos de pesquisa (básica e aplicada) e da apropriação dos dados gerados pelos gestores que atuam em órgãos ambientais, responsáveis pela conservação da biodi-versidade e licenciamento ambiental no país. Os temas prioritários de pesquisa são: im-pacto das espécies de bivalves límnicos invasores sobre os bivalves nativos; estudo morfológico das espécies nativas e invasoras visando à elaboração de chaves e catálogos; mapeamento das espécies nativas de bivalves citadas para o Brasil, espe-cialmente nas bacias subamostradas; descrição do ciclo de desenvolvimento dos bivalves nativos e reconhecimento das espécies de peixes vetores
e dispersores das larvas destes moluscos; relação das rotas migratórias de peixes vetores com a dis-persão de bivalves límnicos; criação de espécies de bivalves límnicos para repovoamento de áreas com declínio das populações das mesmas; avaliação do impacto da construção de reservatórios em série (cascata) sobre a distribuição da malacofauna e a busca de estratégias alternativas para a conserva-ção de espécies de bivalves típicos de água corren-te, visando subsidiar o estabelecimento de medi-das compensatórias e mitigadoras; reconstrução de habitats em represas e nos seus tributários para a conservação da malacofauna; monitoramen-to populacional das espécies invasoras e nativas; identificação de áreas para a implantação de uni-dades de conservação; estabelecimento de estraté-gias de conservação; revisão das listas de espécies ameaçadas estaduais e a nacional, visando rever a categorização das espécies enquadradas e a catego-rização de espécies que não constam nas listas.

95
Moluscos bivalves invasores de origem asiática que ocorrem no ambiente límnico brasileiro são, primordialmente, Limnoperna fortunei (Dunker, 1857), Corbicula fluminea (Müller, 1774) e Corbicula largillierti (Philippi, 1844). No extremo sul do Brasil temos ainda duas espécies de Corbicula que se encontram em áreas muito restritas das bacias que dre-nam a Laguna dos Patos e Mirim: C. fluminalis (Müller, 1774) e C. sp., ainda não identificada. As espécies invasoras devem ser monitoradas constantemente para o conhecimento de suas densidades, interferências ambientais e rotas de dispersão, principalmente em locais onde a água bruta é utilizada para a geração de energia elétrica, resfriamento de indústrias ou abaste-cimento e irrigação. Uma das maneiras de monitorar seria a análise direta do zooplâncton, já que os mo-luscos bivalves desenvolvem-se através de fa-ses larvais que podem frequentar o plâncton, principalmente em se tratando de invasoras. As larvas no plâncton acompanham a corrente da
água, com pouca autonomia de movimentação. Geralmente ficam no fundo, apresentando mo-vimentos circulares de migração vertical na co-luna d’água. É nesta fase planctônica que as es-pécies são facilmente levadas ou tragadas pelas correntes para dentro de canais, encanamentos, cisternas, bombas, tanques de alevinos, tanques de água de lastro, sistemas de refrigeração, etc. As larvas são muito pequenas, variando de 80 a 350 μm no comprimento da concha larval dependendo da espécie (Ortmann, 1921; Bonetto, 1960; Bonetto, 1963; Bonetto & Ez-curra, 1963, 1965; Parodiz & Bonetto, 1963; Mansur & Campos-Velho, 2000; Vale et al., 2005). O pequeno tamanho das larvas facilita sua dispersão e transporte dentro de tubulações e bombas, além de dificultar a localização a olho nu em ações de controle. Para a amostragem do zooplâncton sugere-se seguir os passos para o monitoramento que constam no Capítulo 10. Para o reconhecimento das espécies inva-soras na fase larval, é necessário diferenciá-las entre si e também das larvas de alguns bivalves
CAPÍTULO 6
Morfologia e ciclo larval comparados de bivalves
límnicos invasores e nativos
Maria Cristina Dreher MansurDaniel Mansur Pimpão
Paulo Eduardo Aydos BergonciCintia Pinheiro dos Santos
Gabriela Cristina Santos de Figueiredo

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
96
nativos que compartilham o mesmo ambiente. Apesar desses bivalves serem facilmente dife-renciáveis na fase adulta, seu desenvolvimento e a forma larval são muito semelhantes, princi-palmente nas últimas fases larvais. Apresentam-se neste capítulo, além das técnicas básicas para a observação das larvas ao microscópio, maneiras de reconhecê-las através da morfologia externa e da morfome-tria das conchas larvais. Objetiva-se também auxiliar na compreensão da diversidade de es-tratégias de desenvolvimento e proteção das crias no ambiente dulcícola, descrevendo-se, ao final do presente capítulo, as fases e os ciclos de desenvolvimento dos principais grupos de bi-valves límnicos que ocorrem no Brasil, mesmo aqueles sem larva planctônica.
1. Tipos de larvas planctônicas de bivalves límnicos
As larvas dos bivalves que mais frequen-tam o plâncton são as do tipo véliger, cujo cor-po é protegido por uma concha que pode ser univalve ou bivalve. Na Figura 1, apresenta-se uma chave pictórica para facilitar a diferencia-ção das conchas das principais formas larvais que ocorrem no plâncton, tanto de espécies in-vasoras como de nativas. Assim, reconhecem-se inicialmente dois tipos básicos: o lasídio, com uma concha única em forma de oito, próprio dos bivalves nativos da família Mycetopodidae; e as larvas com conchas bivalves, que podem ser do tipo glo-quídio, que apresenta uma aresta ventral, típica dos bivalves nativos da família Hyriidae ou do tipo veliconcha, com a borda ventral arredon-dada, comum às invasoras.
1.1. Veliconcha de Limnoperna fortunei
A veliconcha de L. fortunei (Fig. 2) apre-senta a charneira reta, sem deflexão central e sem projeções nas extremidades terminais; a superfície externa das valvas subdividida em prodissoconcha I, relativamente pequena, ocu-pando aproximadamente 2/3 da altura da ve-liconcha, seguida da prodissoconcha II, que ocupa o 1/3 restante. A superfície da prodisso-concha I apresenta uma escultura central tipo casca de ovo, sem estrias radiais; muito próximo
da linha dorsal, ocorre a demarcação da glându-la conchífera, representada apenas por poucos e pequenos poros centrais. A prodissoconcha II é bem demarcada por linhas comarginais. A prodissoconcha I é formada na fase larval “D”, apenas pela produção da glândula conchífera que deixa uma demarcação em for-ma de pequenos furos na região central pró-xima da charneira. Quando o manto larval se completa, a concha passa a ser produzida ape-nas pelas bordas do manto adquirindo então as demarcações comarginais estriadas que acom-panham o crescimento da concha e formam a prodissoconcha II. Quando a veliconcha se completa, temos a larva na fase de véliger que envolve todo o corpo protegendo-o, quando
Figura 1. Chave pictórica para a identificação de larvas de bivalves límnicos que frequentam o plâncton.

CAPÍTULO 6
Seção 2
97
fechada. Quando entreabre, expõe o véu, um órgão ciliado, responsável pela locomoção do animal. Os movimentos da larva são geralmen-te circulares e verticais (Ver mais detalhes no item 5.1, deste capítulo).
1.2. Veliconchas de Corbicula fluminea e Corbicula largillierti
A veliconcha de C. fluminea (Fig. 3A) pos-sui as duas extremidades da linha da charneira projetadas e o centro levemente deflexionado. A prodissoconcha I é relativamente grande, ocu-pando 5/6 da altura da veliconcha enquanto a prodissoconcha II apresenta-se muito reduzida, o equivalente ao 1/6 restante. A concha possui escultura radial na região central, situada logo abaixo da demarcação sob forma estrelada da glândula conchífera. As larvas de C. largillierti (Fig. 3B) apresentam a charneira reta e conchas compostas apenas da prodissoconcha I, cor-respondendo ao estágio larval “D”. Na região central não se observaram estrias radiais bem definidas como demarcações da glândula con-chífera, nem se encontraram estágios larvais mais avançados que este, como na espécie acima (mais detalhes no item 5.2).
1.3. O lasídio
Lasídio (Fig. 4) é uma larva do tipo vé-liger transformada para assumir uma fase pa-rasitária temporária em peixes. É próprio da família Mycetopodidae e apresenta caracterís-ticas como: concha larval única em forma de
capacete com uma reentrância de cada lado, o corpo composto por um órgão ciliado anterior em forma de língua ou bilobado, um corpo cen-tral com lobos ventrais e uma cauda posterior também bilobada com ganchos terminais. Para frente do animal, projeta-se um órgão adesivo anterior transparente que, dependendo da espé-cie, pode ser em forma de fita, larga na base afi-lando distalmente (gênero Monocondylaea, Fig. 4), em forma de flagelo (Leila blainvilliana) ou de flor de cartucheira com um microgancho em uma das extremidades, conhecido para duas es-pécies do gênero Anodontites e uma de Myceto-poda (Fig. 17). Não se conhecem os lasídios das demais espécies e gêneros desta família, como: Mycetopodella, Fossula, Haasica e Bartlettia, to-dos monotípicos (com uma espécie por gênero) e Tamsiella com duas espécies. O órgão adesivo anterior é uma estrutura que capacita a larva a se fixar externamente na pele, escamas e nada-deiras ou nas brânquias dos peixes. Os lasídios,
Figura 2. Vista lateral da veliconcha do bivalve invasor Myti-lidae, Limnoperna fortunei, sob microscopia eletrônica de varredura. Charneira (C); demarcação da glândula conchífera (GC); prodissoconcha I (PI); prodissoconcha II (PII). (Prepara-ção e foto: G. Figueiredo e M.C.D. Mansur).
Figuras 3. Vista lateral das veliconchas dos bivalves invasores Corbiculidae, sob microscopia eletrônica de varredura: 3A, Corbicula fluminea; 3B, Corbicula largillierti. Charneira (C); escultura radial (ER); demarcação da glândula conchífera (GC); prodissoconcha I (PI); prodissoconcha II (PII). (Preparação e foto: G. Figueiredo e M.C.D. Mansur).

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
98
assim como os gloquídios, são ectoparasitos de peixes, no entanto crescem muito e emitem haustórios que penetram nos tecidos dos peixes de onde retiram nutrientes.
1.4. Gloquídio
Gloquídio é um tipo de véliger habilitado ao ectoparasitismo temporário de peixes (Figs. 5 a 8), próprio dos representantes da família Hyriidae. Recebe um nome especial por apre-sentar características distintas como: duas val-vas com uma aresta na borda ventral, um gan-cho ventral em cada valva, um flagelo adesivo (ausente em Castalia), cílios sensitivos, cirros, um músculo adutor central, um véu muito ru-dimentar e células fagocitárias forrando inter-
Figura 4. Lasídio de Monocondylaea paraguayana d’Orbigny, 1835 em vista dorsal, com os lobos posteriores recurvados e escondidos sob a concha. Órgão adesivo anterior em forma de fita (AO); lobos anteriores (LA); concha larval (CL). (Fonte: Bonetto & Ezcurra (1963), autorizado).
Figura 5. Gloquídio de Diplodon martensi (Ihering, 1893), pa-rasito de peixe. Organização interna em vista ventral. Célu-las fagocitárias (CF); cirro sensitivo (CR); filamento adesivo (F); músculo adutor (MA); órgão ciliado (OC). Fonte: Mansur (1999) Revista Brasileira de Zoologia 16(2).
Figura 6. Larvas (gloquídios) de bivalves de água doce do gênero Diplodon. 6A, Diplodon obsolescens, vista interna das valvas com gancho; 6B, Diplodon hylaeus, vista interna de uma valva, sem gancho. Aresta ventral (AR), gancho (G). (Pre-paração e foto: D. Pimpão e M.C.D. Mansur).
namente as valvas (Fig. 5). Com o flagelo e os ganchos, o gloquídio torna-se habilitado para se fixar às brânquias, nadadeiras ou escamas dos peixes, que, por sua vez, são responsáveis pela dispersão destes moluscos. Geralmente a época de liberação dos gloquídios por parte dos adultos está relacionada à piracema, movi-mento de migração dos peixes à montante dos rios. Os gloquídios permanecem como parasi-tos dos peixes por um curto período de 20 a 30 dias, conforme Mansur & Silva (1999). No entanto, dentre os Hyriidae do gê-nero Diplodon, temos exceções. As espécies do subgênero Diplodon (Rhipidodonta) possuem gloquídios sem ganchos (Fig. 6B) e sem o fi-lamento adesivo. Segundo Bonetto & Ezcurra (1965), a presença de gancho está relacionada à condição parasitária. Para observar os gloquídios é necessário utilizar o microscópio óptico ou de varredura. Suas valvas apresentam um contorno subtrian-gular com pequenas variações entre os gêneros. Assim, é necessário em primeiro lugar enqua-drar as valvas em três padrões: equilátero nas es-pécies do gênero Castalia (= Castalina e Callo-
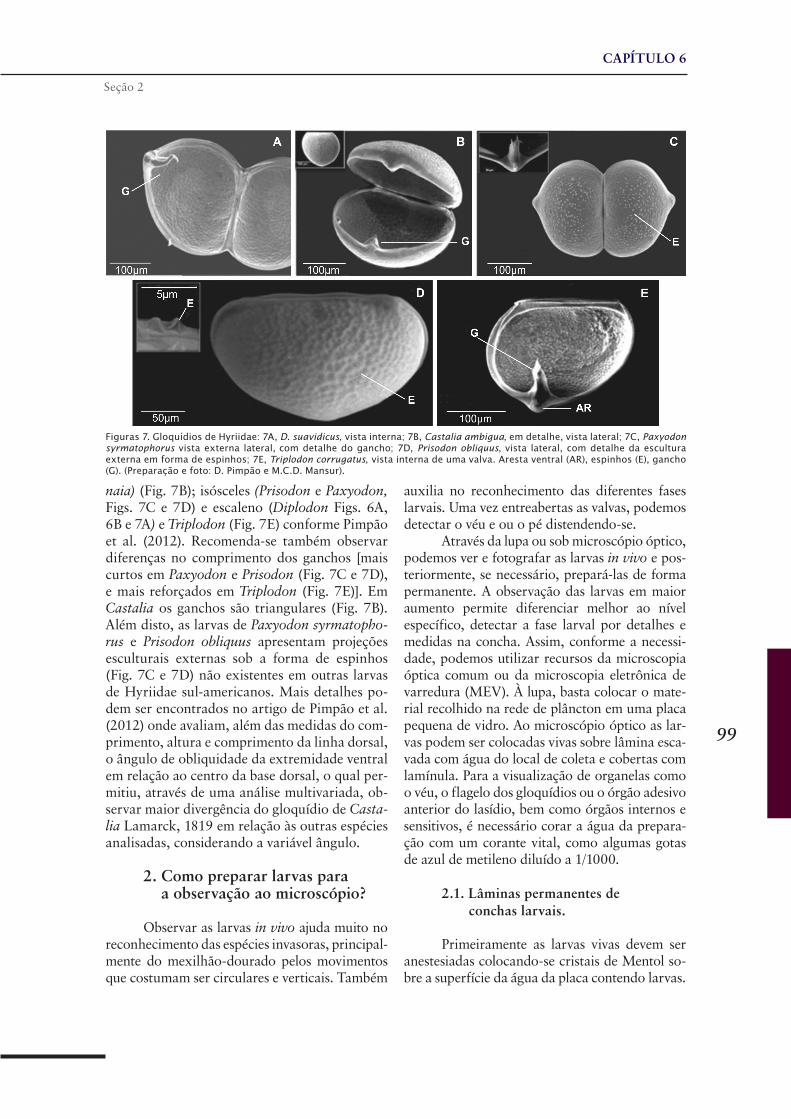
CAPÍTULO 6
Seção 2
99
naia) (Fig. 7B); isósceles (Prisodon e Paxyodon, Figs. 7C e 7D) e escaleno (Diplodon Figs. 6A, 6B e 7A) e Triplodon (Fig. 7E) conforme Pimpão et al. (2012). Recomenda-se também observar diferenças no comprimento dos ganchos [mais curtos em Paxyodon e Prisodon (Fig. 7C e 7D), e mais reforçados em Triplodon (Fig. 7E)]. Em Castalia os ganchos são triangulares (Fig. 7B). Além disto, as larvas de Paxyodon syrmatopho-rus e Prisodon obliquus apresentam projeções esculturais externas sob a forma de espinhos (Fig. 7C e 7D) não existentes em outras larvas de Hyriidae sul-americanos. Mais detalhes po-dem ser encontrados no artigo de Pimpão et al. (2012) onde avaliam, além das medidas do com-primento, altura e comprimento da linha dorsal, o ângulo de obliquidade da extremidade ventral em relação ao centro da base dorsal, o qual per-mitiu, através de uma análise multivariada, ob-servar maior divergência do gloquídio de Casta-lia Lamarck, 1819 em relação às outras espécies analisadas, considerando a variável ângulo.
2. Como preparar larvas para a observação ao microscópio? Observar as larvas in vivo ajuda muito no reconhecimento das espécies invasoras, principal-mente do mexilhão-dourado pelos movimentos que costumam ser circulares e verticais. Também
auxilia no reconhecimento das diferentes fases larvais. Uma vez entreabertas as valvas, podemos detectar o véu e ou o pé distendendo-se. Através da lupa ou sob microscópio óptico, podemos ver e fotografar as larvas in vivo e pos-teriormente, se necessário, prepará-las de forma permanente. A observação das larvas em maior aumento permite diferenciar melhor ao nível específico, detectar a fase larval por detalhes e medidas na concha. Assim, conforme a necessi-dade, podemos utilizar recursos da microscopia óptica comum ou da microscopia eletrônica de varredura (MEV). À lupa, basta colocar o mate-rial recolhido na rede de plâncton em uma placa pequena de vidro. Ao microscópio óptico as lar-vas podem ser colocadas vivas sobre lâmina esca-vada com água do local de coleta e cobertas com lamínula. Para a visualização de organelas como o véu, o flagelo dos gloquídios ou o órgão adesivo anterior do lasídio, bem como órgãos internos e sensitivos, é necessário corar a água da prepara-ção com um corante vital, como algumas gotas de azul de metileno diluído a 1/1000.
2.1. Lâminas permanentes de conchas larvais. Primeiramente as larvas vivas devem ser anestesiadas colocando-se cristais de Mentol so-bre a superfície da água da placa contendo larvas.
Figuras 7. Gloquídios de Hyriidae: 7A, D. suavidicus, vista interna; 7B, Castalia ambigua, em detalhe, vista lateral; 7C, Paxyodon syrmatophorus vista externa lateral, com detalhe do gancho; 7D, Prisodon obliquus, vista lateral, com detalhe da escultura externa em forma de espinhos; 7E, Triplodon corrugatus, vista interna de uma valva. Aresta ventral (AR), espinhos (E), gancho (G). (Preparação e foto: D. Pimpão e M.C.D. Mansur).

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
100
A água tem que ser mineral ou natural do local de coleta. Após a anestesia, que leva algumas ho-ras (acompanhar à lupa), transferir as larvas para uma proveta com auxílio de uma pipeta de Pas-teur, desprezando-se o excesso de água. Acres-centar 8 gotas de água sanitária ou hipoclorito de sódio comercial (5% NaOCl) para 10 ml de água deionizada durante aproximadamente 5 minutos. O hipoclorito remove os tecidos. Acompanhar o processo de limpeza através da observação dire-ta na lupa. Lavar o material renovando a água a cada minuto (tempo suficiente para as larvas decantarem ao fundo do tubo). Trocar o líquido apenas acima do nível das larvas. Desidratar rapi-damente por uma série crescente de álcool etílico de 50, 60, 70, 80, 90, 96% e, ao final, dois banhos de álcool isopropílico. Pipetar as veliconchas em pequenas quantidades sobre lâmina, montar com algum bálsamo de selagem histológica e cobrir com lamínula. Utilizar um anel ou moldura de papel entre lâmina e lamínula para o peso desta não quebrar as conchas. Mais detalhes em Man-sur & Campos-Velho (1990). Através da microscopia óptica, obser-vam-se vários caracteres importantes na dife-renciação interespecífica das conchas gloqui-dianas, como: a forma da charneira (área de articulação das valvas) reta ou com deflexão, borda ventral com aresta ou arredondada, po-sição da aresta em relação ao centro da char-neira, borda com reforço ou simples, presença ou não de ganchos e os diferentes tipos destes. Nas veliconchas das invasoras, a técnica permi-te observar em L. fortunei a charneira reta e as extremidades arredondadas, enquanto que em C. fluminea e C. largillierti a charneira é de-flexionada na região central e as extremidades são levemente projetadas. 2.2. Observação de veliconchas sob o microscópio eletrônico de varredura Proceder à anestesia e remoção dos teci-dos como descrito acima. Depois, macerar em água deionizada durante dois dias, trocando o líquido acima do nível das larvas a cada minu-to. Limpo o material, este deve ser pipetado so-bre papel filtro para secar. Com um pincel fino selecionar, sob à lupa, as valvas mais íntegras e fixá-las sobre o suporte “stub” já preparado com cola adesiva dupla face de cobre, devida-
mente numerada. Maiores detalhes em Mansur & Campos-Velho (1990). Sob microscopia eletrônica de varredura é possível observar mais caracteres que refor-çam as diferenças entre as espécies invasoras, L. fortunei (Fig. 2), C. fluminea e C. largillierti (Figs. 3A e 3B), principalmente na superfície externa da veliconcha que pode apresentar es-culturas ou não, vestígios da glândula conchífe-ra e demarcações que dividem as valvas em áre-as de proporções diferentes que correspondem a prodissoconcha I e prodissoconcha II.
3. Como avaliar a morfometria larval? Com base em apenas três medidas da concha larval (Fig. 8 e Quadro 1) referentes ao comprimento, altura e comprimento da char-neira, é possível diferenciar as espécies entre si e as invasoras das nativas. Por meio de lâminas permanentes, con-chas larvais são posicionadas em máxima ho-rizontalidade com as bordas no foco e mensu-radas conforme demonstrado na Figura 8. A charneira representa a base e ponto de referên-cia. Assim, o comprimento máximo da concha é mensurado pelo paralelo e a altura pela per-pendicular à linha da charneira. Para a discriminação das formas das con-chas larvais a partir de variáveis mensuráveis (Quadro 1), referimos a análise multivariada de fatores discriminantes canônicos, pois nesta
Figura 8. Orientação da concha larval, para a obtenção de me-didas (gloquídio de Triplodon corrugatus). Altura perpendicu-lar à linha da charneira (A), comprimento máximo paralelo ao da charneira (C), comprimento da charneira (CC). (Preparação e foto: D. Pimpão e M.C.D. Mansur).

CAPÍTULO 6
Seção 2
101
análise a ideia principal é retirar a influência do tamanho na discriminação entre as formas, re-sultando na identificação da variável ou variáveis responsáveis pela separação das mesmas (Perez--Neto, 1995). Este método foi aplicado recente-mente por Pimpão et al. (2012) para a discrimina-ção morfométrica de conchas larvais de espécies amazônicas de Hyriidae. Para a realização desta análise, recomenda-se a transformação dos dados originais em logaritmos (Ln ou Log10) a fim de diminuir a variabilidade entre os dados.
4. Morfometria larval comparada
A estatística descritiva de variáveis mor-fométricas das conchas larvais de 10 espécies de moluscos bivalves, sete nativas e três invasoras, pode ser verificada no Quadro 1. Constata-se que Diplodon obsolescens apresenta os maiores valores, ao passo que, L. fortunei, os menores. Através da morfometria larval, é possível diferenciar Corbicula fluminea de L. fortunei através da conformação e medidas do com-
primento da charneira. Comparando as três espécies invasoras, L fortunei apresenta pro-porcionalmente as menores medidas de com-primento, de altura e a charneira mais curta. C. largillierti possui charneira mais alongada das três espécies, mesmo considerando o fato do material examinado não estar em estágio larval tão avançado como as demais, ou seja, ainda em estágio de larva “D”. A análise discriminante canônica (ADC), apresentada graficamente por meio da Figura 9A, diferencia a forma larval de L. fortunei das demais espécies de bivalves com desenvol-vimento não planctônico. Uma segunda ADC realizada sem a presença de L. fortunei elucida mais claramente as diferenças entre as formas larvais das demais espécies, dada agora pela variável comprimento da charneira (Fig. 9B). Nesta segunda análise, as espécies de Diplo-don e Prisodon (D. obsolescens, D. hylaeus, D. suavidicus e P. obliquus) são discriminadas das demais, bem como as outras duas espécies inva-soras (C. fluminea e C. largillierti).
Quadro 1. Medidas de comprimento, altura e comprimento da charneira de larvas gloquídios (Hyriidae) e das invasoras Corbi-cula fluminea e C. largillierti (Corbiculidae) e Limnoperna fortunei (Mytilidae).

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
102
5. Descrição das fases larvais das espécies invasoras e nativas
O embrião dos bivalves, até atingir a fase juvenil passa, obrigatoriamente, por vários está-gios de desenvolvimento, a começar pelo ovo, depois blástula, gástrula, trocófora, larva “D” (quando se completa a veliconcha inicial corres-pondente a prodissoconcha I), véliger, com pre-sença de um véu ciliado bastante evidente e o início da prodissoconcha II, e o pedivéliger, que
depois da absorção do véu e maior desenvolvi-mento do pé, torna-se pós-larva ou juvenil.
5.1. Fases larvais de L. fortunei
L. fortunei libera os gametas para o meio ambiente e os ovos, fertilizados externamen-te, se desenvolvem numa larva planctônica. Os ovos fecundados (zigotos) são muito pequenos, (aproximadamente 80 μm) e o desenvolvimento inicial do embrião, a partir da divisão dos pri-meiros blastômeros, passa à gástrula, depois à fase de trocófora que leva ao todo de 5 a 6 ho-ras. A trocófora se transforma em véliger através do desenvolvimento do véu que é uma organela ciliada destinada à locomoção e filtração (Choi & Kim, 1985; Choi & Shin, 1985). Estes au-tores reconheceram três estágios de véliger que denominaram de larva “D”: primária, média e larva “D” propriamente dita, com os respectivos tamanhos de: 140.0 μm. 167.6 μm e 210.0 μm de comprimento por 97.3 μm, 137.6 μm e 178.2 μm em altura. O último estágio larval no qual o véu e o pé, já bem desenvolvidos, estão pre-sentes, trata-se de um juvenil em estágio inicial ou uma larva umbonada, pronta para assentar e colar ao substrato. Ressalta-se aqui que a deno-minação das fases larvais varia muito conforme o autor. As fases larvais de uma população de L. fortunei do sul do Brasil, foram descritas por Santos et al. (2005) como apresentando cinco estágios sem valvas e quatro valvados (Fig. 10). O primeiro reconhecido como uma mórula ci-liada (Fig. 10A), que se transforma numa trocó-fora (comprimentos respectivos de 80 μm a 125 μm) com quatro estágios distintos, trocófora de 1 à 4 (Figs. 10B a 10E). Na última fase de trocó-à 4 (Figs. 10B a 10E). Na última fase de trocó- 4 (Figs. 10B a 10E). Na última fase de trocó-fora (Fig. 10E) aparecem os primórdios da con-cha. Os estágios valvados incluem: a larva “D” (Fig. 10F, comprimento de 100 μm a 130 μm), o véliger de charneira reta (Fig. 10G, compri-mento de 140 μm a 180 μm), véliger umbonado (Fig. 10H, comprimento de 190 μm a 230 μm) e o pedivéliger (Fig. 10I, comprimento de 230 μm a 270 μm). Nesta fase, o pé se desenvolve e o véu é lentamente absorvido. Quando pós--larva ou plantígrada (Fig. 10J, comprimento aproximado de 300 μm), o pé secreta, através da glândula bissogênica, fios de bisso que per-mitem sua fixação no substrato.
Figura 9. Análise discriminante canônica entre as variáveis mensuradas de 10 espécies de larvas de bivalves: 9A, análise com Limnoperna fortunei; 9B, análise sem L. fortunei. Diplo-don obsolescens (1); D. hylaeus (2); D. suavidicus (3); Prisodon obliquus (4); Paxyodon syrmatophorus (5); Triplodon corrugatus (6); Castalia ambigua (7); Corbicula fluminea (8); C. largillierti (9); L. fortunei (10).

CAPÍTULO 6
Seção 2
103
Considerando o período de duração do ciclo planctônico da larva como livre natante, Choi & Shin (1985) referem aproximadamente um mês, no reservatório próximo de Seul, na Coréia do Sul.
5.2. Fases larvais de Corbicula spp.
Nos ambientes límnicos do Brasil po-demos encontrar quatro espécies invasoras de origem asiática do gênero Corbicula: C flumi-nea, C., largillierti, C. fluminalis e C. sp. (ain-da não identificada). Conforme Skuza et al. (2009), as espécies do gênero Corbicula são únicas em termos de reprodução, por serem tanto hermafroditas quanto dioicas e, segundo alguns autores, capazes de autofecundação e ou clonagem (ver Capítulo 8). Quanto ao desenvolvimento larval, as espécies de Corbi-cula spp. podem ser ovíparas, ovovivíparas e vivíparas. As espécies de água doce geralmente
são vivíparas com a incubação das larvas até a fase de pós-larva, no saco marsupial das demi-brânquias internas (endobranquial – caso mais comum) ou nas quatro brânquias (tetrabran-quial – casual). Martins (2004) e Martins et al. (2006) observaram em C. fluminea e C. lar-gillierti a incubação como endobranquial (Fig. 11), e em C. fluminalis como tetrabranquial (Ver Capítulo 4). O desenvolvimento larval foi descrito para a espécie C. fluminea mais detalhadamen-te por Kraemer & Galloway (1986) e King et al. (1986). Na Figura 12, esquematizam-se as fases larvais que são incubadas nos marsúpios até a fase de liberação ao meio ambiente. No marsúpio se desenvolvem: o embrião, passan-do pelos estágios larvais desde o ovo, blástula (175 μm), gástrula (de 175 – 180 μm), trocófora (180 – 190 μm), trocófora com primórdios da concha (±190 μm), larva véliger com presença de um véu ciliado bastante evidente (200 – 220 μm) e larva pedivéliger (225 – 230 μm), que depois de abortar o véu se torna pós-larva ou juvenil, sendo então liberada do marsúpio para o exterior. Este juvenil mede em torno de 220 a 270 μm de comprimento, não é livre natante, mas por ser muito leve, pode ser encontrado também no plâncton. Apresenta um pé adap-tado a rastejar em ambientes de correnteza, onde pode se locomover sem ser arrastado pe-las águas. Isto acontece graças a uma glândula que existe no pé e que produz um fio mucoso e elástico. Este não é um bisso, mas permite ade-são ao substrato à medida que o animal rasteja. Este fio também auxilia o animal a assumir sua vida bentônica infaunal (Kraemer & Galloway, 1986).
Figura 10. Fases larvais do mexilhão-dourado (L. fortunei). A, Mórula ciliada (80 µm); B a D, trocófora; E, trocófora com os primór-dios da concha; F, larva “D”; G, véliger de charneira reta; H, véliger umbonado ; I, pedivéliger e J, pós-larva. (Desenhos M.C.D. Mansur).
Figura 11. Brânquia direita de Corbicula largillierti com a de-mibrânquia externa rebatida e a interna contendo embriões incubados. Demibrânquia externa (DBE), demibrânquia inter-na (DBI), embriões (E), marsúpio (M). (Foto: M.C.D. Mansur).

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
104
Nas espécies C. fluminea e de C. lar-gillierti, tanto as larvas quanto a sequência de desenvolvimento no marsúpio é muito se-melhante (Figs. 13 e 14). Os embriões estão unidos por uma massa gelatinosa, formando uma estrutura compacta, no formato cilíndri-co e ocupam os tubos aquíferos das brânquias, principalmente nas fases iniciais. Nas fases de véliger em diante, ficam soltos e podem nadar no interior da brânquia. Quando exempla-res adultos com larvas incubadas entram em stress ou quando, após coleta, são colocados em aquário, facilmente liberam as larvas em estágios de véliger que nadam livremente. Não sabemos sobre suas condições de sobrevivência e se conseguem completar o desenvolvimento. C. fluminalis e C. sp. não têm seu desenvolvi-mento larval conhecido em nosso meio.
5.3. Fases larvais da espécie nativa Cyanocyclas limosa
Considerando os Corbiculidae nativos, só existem estudos realizados para a espécie Cyanocyclas limosa (Ituarte, 1984b, 1994; Focht & Veitenheimer, 2001) até recentemen-te conhecida por Neocorbicula limosa. Ituarte (1984b) observou que os embriões são incuba-dos nas demibrânquias internas por longos pe-ríodos e se nutrem do vitelo contido no ovo.
Em geral, são liberados aos poucos, a partir de 1,1 mm de comprimento, não na fase de pós-larva, como em Corbicula fluminea, mas como um juvenil em adiantado estado de desenvolvimento, uma miniatura do indivíduo adulto (Fig. 15B). Seria um caso de euviviparia (incubação prolongada, segundo Glaubrecht et al., 2006). Entretanto, muitos ainda per-manecem nas brânquias internas do indivíduo “mãe”. O número (assim como o tamanho) de embriões incubados é variável, não ultrapas-sando, em geral, 25 indivíduos por demibrân-quia. Raramente encontram-se 45 embriões nas duas demibrânquias e o desenvolvimento destes embriões não é sincronizado como em C. fluminea. Distinguem-se geralmente três ou quatro gerações com tamanhos diferentes, que estão relacionadas às distintas fases reprodu-tivas do indivíduo materno (Ituarte, 1984b). Em estudos realizados com uma população do Lago Guaíba, Focht & Veitenheimer-Mendes (2001) confirmaram e ilustram o tipo de incu-bação dos juvenis nas demibrânquias internas.
5.4. Fases larvais de Hyriidae
Os bivalves nativos Hyriidae no Brasil, reúnem os gêneros Diplodon (± 37 espécies), Triplodon (2 spp.), Paxyodon (1 sp.) e Prisodon (1 sp.) e Castalia (± 13 spp). Todas as larvas das
Figura 12. Esquema das fases larvais de Corbicula fluminea (Müller 1774) desde a formação do zigoto pelos gametas masculino e feminino (a e b) até as fases incubadas no marsúpio (de C até I), incluindo a fase de pedivéliger ou pós-larva que é liberada ao meio ambiente a partir da fase J (pós-larva J ou juvenil K). – momento da liberação da larva no ambiente; óvulo (A); esper-matozoide (B); embrião em clivagem com os primeiros blastômeros (C); gástrula (D); trocófora (E); trocófora com primórdios da concha (F); véliger de charneira reta (G); pedivéliger (H); pedivéliger abortando o véu (I); pós-larva com o manto e o pé extro-vertidos (J); juvenil (K). Fonte: Kraemer & Galloway (1986); King et al. (1986), adaptado por M.C.D. Mansur.
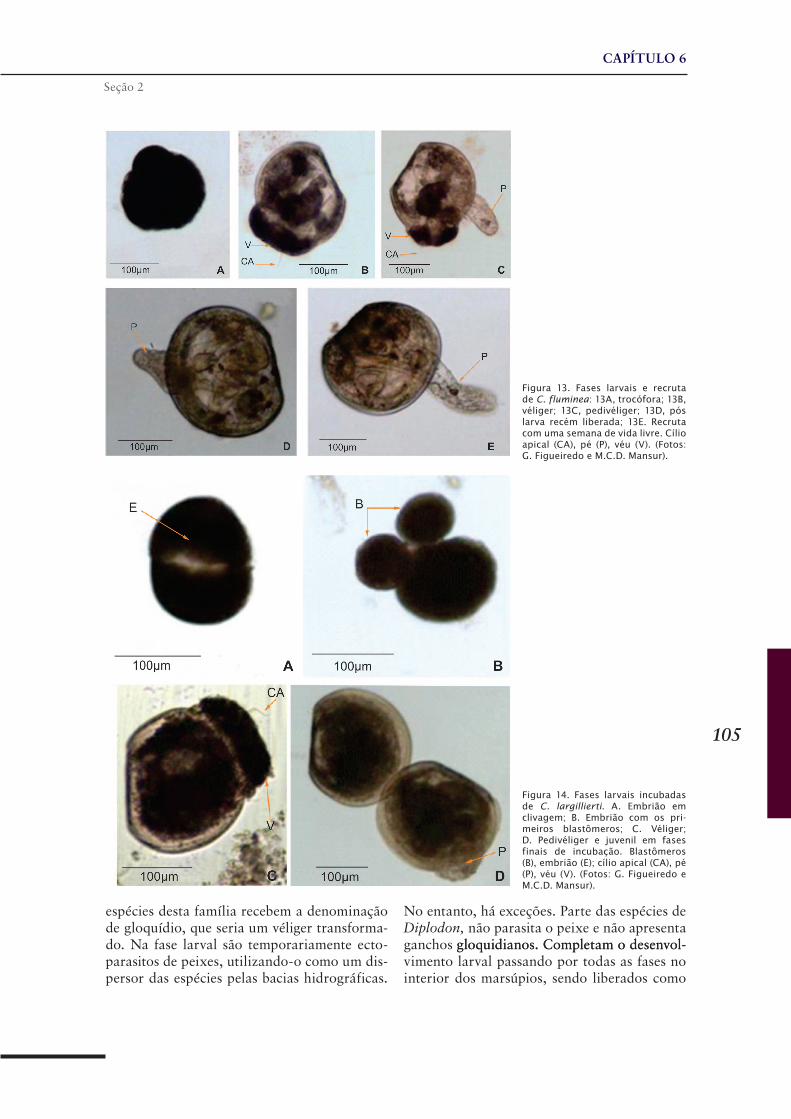
CAPÍTULO 6
Seção 2
105
Figura 13. Fases larvais e recruta de C. fluminea: 13A, trocófora; 13B, véliger; 13C, pedivéliger; 13D, pós larva recém liberada; 13E. Recruta com uma semana de vida livre. Cílio apical (CA), pé (P), véu (V). (Fotos: G. Figueiredo e M.C.D. Mansur).
Figura 14. Fases larvais incubadas de C. largillierti. A. Embrião em clivagem; B. Embrião com os pri-meiros blastômeros; C. Véliger; D. Pedivéliger e juvenil em fases finais de incubação. Blastômeros (B), embrião (E); cílio apical (CA), pé (P), véu (V). (Fotos: G. Figueiredo e M.C.D. Mansur).
espécies desta família recebem a denominação de gloquídio, que seria um véliger transforma-do. Na fase larval são temporariamente ecto-parasitos de peixes, utilizando-o como um dis-persor das espécies pelas bacias hidrográficas.
No entanto, há exceções. Parte das espécies de Diplodon, não parasita o peixe e não apresenta ganchos gloquidianos. Completam o desenvol-gloquidianos. Completam o desenvol-vimento larval passando por todas as fases no interior dos marsúpios, sendo liberados como
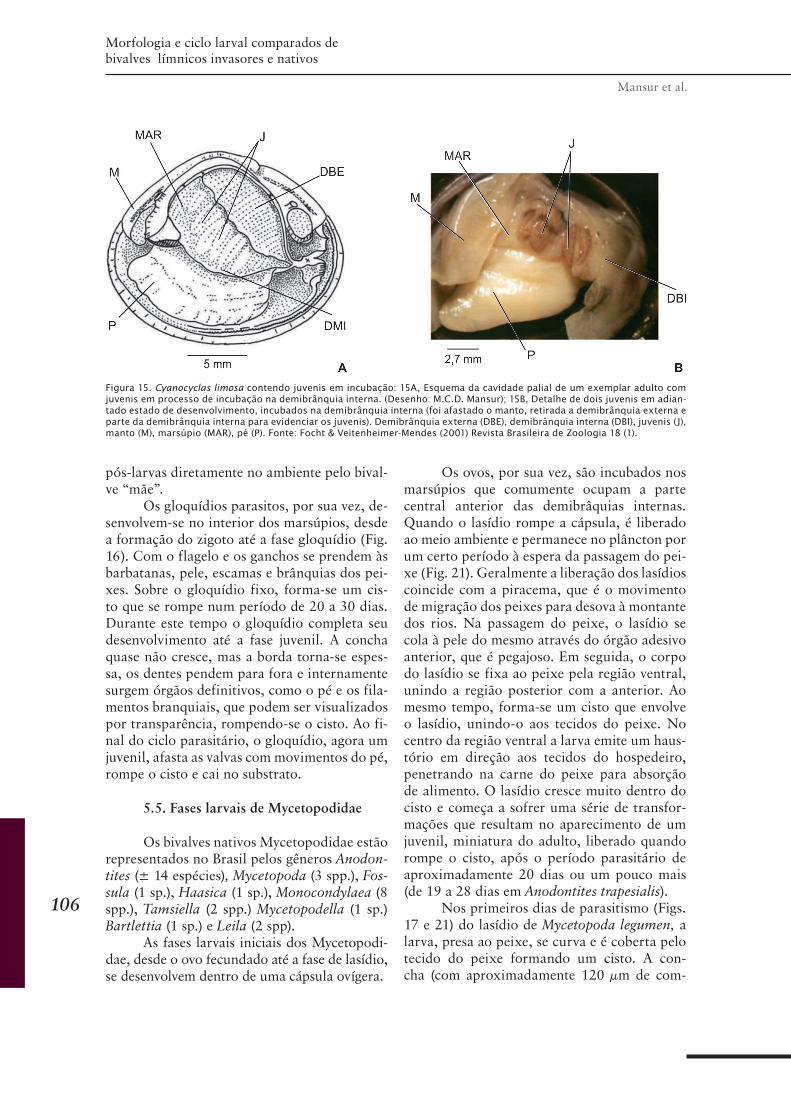
Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
106
Figura 15. Cyanocyclas limosa contendo juvenis em incubação: 15A, Esquema da cavidade palial de um exemplar adulto com juvenis em processo de incubação na demibrânquia interna. (Desenho: M.C.D. Mansur); 15B, Detalhe de dois juvenis em adian-tado estado de desenvolvimento, incubados na demibrânquia interna (foi afastado o manto, retirada a demibrânquia externa e parte da demibrânquia interna para evidenciar os juvenis). Demibrânquia externa (DBE), demibrânquia interna (DBI), juvenis (J), manto (M), marsúpio (MAR), pé (P). Fonte: Focht & Veitenheimer-Mendes (2001) Revista Brasileira de Zoologia 18 (1).
pós-larvas diretamente no ambiente pelo bival-ve “mãe”. Os gloquídios parasitos, por sua vez, de-senvolvem-se no interior dos marsúpios, desde a formação do zigoto até a fase gloquídio (Fig. 16). Com o flagelo e os ganchos se prendem às barbatanas, pele, escamas e brânquias dos pei-xes. Sobre o gloquídio fixo, forma-se um cis-to que se rompe num período de 20 a 30 dias. Durante este tempo o gloquídio completa seu desenvolvimento até a fase juvenil. A concha quase não cresce, mas a borda torna-se espes-sa, os dentes pendem para fora e internamente surgem órgãos definitivos, como o pé e os fila-mentos branquiais, que podem ser visualizados por transparência, rompendo-se o cisto. Ao fi-nal do ciclo parasitário, o gloquídio, agora um juvenil, afasta as valvas com movimentos do pé, rompe o cisto e cai no substrato.
5.5. Fases larvais de Mycetopodidae
Os bivalves nativos Mycetopodidae estão representados no Brasil pelos gêneros Anodon-tites (± 14 espécies), Mycetopoda (3 spp.), Fos-sula (1 sp.), Haasica (1 sp.), Monocondylaea (8 spp.), Tamsiella (2 spp.) Mycetopodella (1 sp.) Bartlettia (1 sp.) e Leila (2 spp). As fases larvais iniciais dos Mycetopodi-dae, desde o ovo fecundado até a fase de lasídio, se desenvolvem dentro de uma cápsula ovígera.
Os ovos, por sua vez, são incubados nos marsúpios que comumente ocupam a parte central anterior das demibrâquias internas. Quando o lasídio rompe a cápsula, é liberado ao meio ambiente e permanece no plâncton por um certo período à espera da passagem do pei-xe (Fig. 21). Geralmente a liberação dos lasídios coincide com a piracema, que é o movimento de migração dos peixes para desova à montante dos rios. Na passagem do peixe, o lasídio se cola à pele do mesmo através do órgão adesivo anterior, que é pegajoso. Em seguida, o corpo do lasídio se fixa ao peixe pela região ventral, unindo a região posterior com a anterior. Ao mesmo tempo, forma-se um cisto que envolve o lasídio, unindo-o aos tecidos do peixe. No centro da região ventral a larva emite um haus-tório em direção aos tecidos do hospedeiro, penetrando na carne do peixe para absorção de alimento. O lasídio cresce muito dentro do cisto e começa a sofrer uma série de transfor-mações que resultam no aparecimento de um juvenil, miniatura do adulto, liberado quando rompe o cisto, após o período parasitário de aproximadamente 20 dias ou um pouco mais (de 19 a 28 dias em Anodontites trapesialis). Nos primeiros dias de parasitismo (Figs. 17 e 21) do lasídio de Mycetopoda legumen, a larva, presa ao peixe, se curva e é coberta pelo tecido do peixe formando um cisto. A con-cha (com aproximadamente 120 μm de com-

CAPÍTULO 6
Seção 2
107
Figura 16. Gloquídio de Diplodon martensi (Ihering, 1893): 16A, vista frontal da larva parasitária de peixes, como encontrado no plâncton; 16B, gloquídio no 5º dia de vida parasitária como ectopa-rasito de peixe; 16C, gloquídio no 30º dia de vida parasitária; 16D, pós-larva no 1º dia de vida livre, em vista ventral com as valvas cer-radas; 16E, juvenil em vista lateral com o pé distendido. Concha lar-val (CL); coração (CO); dente glo-quidiano (DG); filamentos bran-quiais (FB); músculo adutor central (MA); músculo adutor anterior (MAA); músculo adutor posterior (MAP); pé (P); quisto larval (QL); raio da aleta do peixe (R). Fonte: Mansur (1999), Revista Brasileira de Zoologia, 16 (2).
Figura 17. Lasídio de Mycetopoda legumen (Mycetopodidae): 17A, vista dorsal de indivíduo com o órgão adesivo anterior completo; 17B, detalhe do lasídio em vista la-teral, com a concha o corpo, e par-te do órgão adesivo anterior; 17C, lasídio no 5º dia, encistado na pele do peixe; 17D, lasídio no 10º dia de parasitismo; 17E, lasídio no 17º dia, encistado; 17F, lasídio no 18º dia, encistado, e 17G, no 21º dia, juvenil afastando as valvas com o pé. Concha larval (CL); filamentos branquiais (FB); haustório (H); lobo anterior (LA); lobo posterior (LP); lobos ventrais (LV), órgão adesivo anterior (AO), pé (P); quisto larval (QL). Fonte: Veitenheimer-Mendes & Mansur (1978), Revista Brasileira de Biologia 38 (3), adaptado.
primento) se prolonga pelos lados em direção ventral, cobrindo as extremidades do corpo da larva; os 4 lobos ventrais da larva unem-se e projetam-se e no centro o haustório se prolon-ga e faz contato com os tecidos do peixe. Por
transparência pode-se ainda observar o corpo do lasídio com os minúsculos ganchinhos do lobo posterior, dentro da concha. No décimo dia de parasitismo a larva pode atingir 346 μm de comprimento por 213 μm de altura e toma a

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
108
forma de um rim. Na região ventral, a concha se solda deixando apenas a fresta para uma pe-quena porção projetada dos 4 lobos ventrais e no centro o haustório. No décimo quarto dia, o tamanho da larva passa a 545 μm de com-primento por 407 μm de altura. No décimo sétimo dia, a larva, agora com o comprimento que varia de 647 a 814 μm de comprimento por 462 a 582 μm de largura, apresenta na região dorsal, anterior e posterior, um marcado sul-co que irá definir a futura divisão da concha larval – até agora única – em duas valvas. No décimo oitavo dia, por transparência, é possí-vel visualizar o pé e os filamentos branquiais. No vigésimo primeiro dia, a larva, agora com 837 μm de comprimento por 647 μm de altura, mostra movimentos ativos do pé, que promo-vem o afastamento das valvas e o rompimento do cisto. Liberado, passa a ser um juvenil que irá se desenvolver no bentos até a fase adulta. Muitos juvenis desta família apresentam no pé um sulco ventral e produzem um fio de bisso temporário que os auxilia a se fixar junto à ve-getação marginal, para não serem arrastados pela correnteza.
6. Ciclos larvais
Conforme a família ou grupo dos bival-ves, a sequência de estágios larvais pode acon-tecer de diferentes modalidades que dependem do grau de proteção que é conferido ao em-brião ou de estratégias necessárias para a dis-persão da espécie. Assim, as larvas podem se desenvolver livremente no plâncton, ao sabor da corrente; totalmente incubados e protegidos
dentro da concha “mãe”, em espaços denomi-nados de marsúpios, geralmente situados nas demibrâquias internas (Fig. 18); parcialmente incubados nos marsúpios, quando só as últimas fases são planctônicas; ou, ainda, parcialmente incubados, completando seu desenvolvimento como parasitos temporários de peixes. Além disto, as larvas podem se desenvolver envoltas ou não por cápsulas ovígeras, mesmo dentro dos marsúpios. A proteção dos embriões cons-titui quase uma regra para os bivalves nativos de água doce. O desenvolvimento larval dos bivalves límnicos invasores e nativos que ocorrem no Brasil, pode ser sumarizado em 5 modalidades, conforme os esquemas a seguir (Figs. 19 a 23):
6.1. Ciclo completo planctônico
Todo o ciclo larval, desde a fecundação, ocorre no plâncton (Fig. 19). Na fase de pedi-véliger passa a frequentar o bentos como um jovem recruta. É o caso do mexilhão-dourado, bem como da maioria dos seus parentes mari-nhos (Mytilidae). Na Coréia, L. fortunei libera os gametas para o meio ambiente durante um período de 15 a 20 dias entre julho e agosto (Choi & Kim, 1985; Choi & Shin, 1985).
6.2. Ciclo parcialmente incubado no marsúpio e planctônico
As primeiras fases larvais (desde ovo até véliger) desenvolvem-se protegidas dentro do marsúpio. São liberadas no meio ambiente, num estádio de véliger ou pedivéliger (Fig. 20).
Figura 18. Órgãos da cavidade pa-lial de adulto do gênero Corbicula. Em cor amarela destaca-se a de-mibrânquia interna que contém o marsúpio, local de incubação das larvas. (Desenho: M.C.D. Mansur).

CAPÍTULO 6
Seção 2
109
É o caso das corbículas invasoras como C. fluminea, C. largillierti e C. fluminalis, nas quais todas as fases larvais, até pedivéliger, costumam acontecer dentro dos marsúpios. Porém, eventualmente, por razões ainda desco-nhecidas, podem liberar larvas ainda nas fases de véliger ou pedivéliger.
6.3. Ciclo parcialmente incubado no marsúpio e parasitário de peixe As primeiras fases larvais desde ovo até chegar a um tipo de véliger transformado (lasídio em Mycetopodidae, Fig. 21A; gloquídio em Hyrii-dae, Fig. 21B) permanecem incubados nos mar-súpios. São então liberados no plâncton e, em poucas horas, aderem às escamas, nadadeiras ou brânquias de peixes hospedeiros. Nestes se encistam e completam seu desenvolvimento como ectoparasitas até a fase juvenil. Rompem o cisto, passando a viver no bentos. É o caso dos bivalves de conchas nacaradas Mycetopo-didae e da maior parte das espécies da família Hyriidae.
6.4. Ciclo completo incubado no marsúpio
Todo o desenvolvimento larval ocorre protegido pela mãe dentro dos marsúpios. São liberados na fase juvenil, como uma mi-niatura do adulto. É o caso dos Sphaeriidae (Eupera, Byssanodonta, Pisidium, Musculium e Sphaerium), dos Corbiculidae nativos (Cya-nocyclas) cujos juvenis são liberados em está-dio adiantado de desenvolvimento (Focht & Veitenheimer, 2001, Fig. 22A) e dos Hyriidae do gênero Diplodon (Rhipidodonta), que não utilizam o peixe como hospedeiro e dispersor (Fig. 22B).
6.5. Ciclo completo incubado na cavidade palial
É o caso do nosso falso mexilhão-doura-do, Mytilopsis lopesi (Dreissenidae), endêmico do curso inferior do Rio Amazonas, excluindo a área do delta, cujos embriões se desenvolvem fora das brânquias, mas no interior da cavidade palial, aderidos ao manto. Os jovens são libera-dos em adiantado estádio de desenvolvimento (Mansur et al., 2011b, Fig. 23).
Figura 19. Ciclo larval planctônico desde a fecundação até o recrutamento.
Figura 20. Ciclo larval parcialmente incubado no marsúpio.
Figura 21. Ciclo larval parcialmente incubado no marsúpio e parasitário de peixe. 21A, Mycetopodidae (lasídio) e 21B, Hyriidae (gloquídio).
Figura 22. Ciclo completo incubado no marsúpio até juvenil. 22A, Cyanocyclas em estágio adiantado de desenvolvimento; 22B, Diplodon (Rhipidodonta).
Figura 23. Ciclo completo incubado na cavidade palial e a li-beração pelo sifão inalante de juvenis em estágio avançado de desenvolvimento. (D
esenh
os:
M.C
.D. M
ansu
r).

Mansur et al.
Morfologia e ciclo larval comparados de bivalves límnicos invasores e nativos
110
Mytilopsis lopesi Alvarenga & Ricci, 1989 apresenta aspecto mitiloide, vivendo fixo por fios de bisso em substratos duros, for-mando incrustações como pequenas manchas. Por este motivo tem sido confundido com o mexilhão-dourado L. fortunei. Apesar da apa-rente semelhança, são de famílias e até ordens de Bivalvia distintas. Segundo Cox (1969), os representantes da família Dreissenidae, (com os gêneros Dreissena, Mytilopsis, Congeria)
apresentam a larva livre natante planctotrófi-ca, passando por todas as fases larvais, desde trocófora à véliger, como a maioria das espécies marinhas. No entanto, nosso “falso mexilhão” Mytilopsis lopesi, incuba as larvas até adianta-do estado de desenvolvimento, não apresentan-do larva planctônica, motivo de não ter sido considerado na morfologia larval comparada. É também um caso de euviviparia, mas fora das brânquias.

111
A intensa capacidade reprodutiva do me-xilhão-dourado, Limnoperna fortunei (Dunker, 1857), tem facilitado a rápida disseminação e altas densidades desde a sua introdução aciden-tal na América do Sul, acerca de duas décadas (Pastorino et al., 1993; Mansur et al., 1999). Dentre os requisitos necessários para uma espé-cie se tornar um bioinvasor de sucesso, Ricciardi & Rasmussen (1998) elencam que a espécie deve apresentar um ciclo de vida curto; rápido cres-cimento, com maturidade sexual precoce e alta fecundidade, características típicas de espécies estrategistas-r, como o mexilhão-dourado, clas-sificado assim por Morton (1977). Informações detalhadas relacionadas a aspectos populacionais, em especial, às flutu-ações reprodutivas sazonais (Cataldo & Bolto-vskoy, 2000), à estrutura etária, à proporção entre os sexos (sex-ratio), dentre outras carac-terísticas, são essenciais, não apenas na investi-gação de mecanismos básicos de reprodução e desenvolvimento, mas também na previsão de
sua propagação, sendo assim fundamentais para a produção de estratégias adequadas de controle e mitigação dos impactos. Neste capítulo pre-tendemos relatar os processos de maturação e as mudanças cíclicas que ocorrem nas estruturas reprodutivas formadoras de gametas.
Dioicos, porém hermafroditas, como assim?
Bivalves, preferencialmente os de água doce, apresentam uma ampla plasticidade e variabilidade de estratégias quando se trata de produção de gametas. Há muito se relata e se descreve um gradiente compreendendo diferen-tes categorias sexuais, desde espécies exclusiva-mente dioicas, que apresentam sexos separados, até aquelas que são invariavelmente hermafrodi-tas funcionais (Coe, 1943). Em um mesmo gêne-ro é possível observar em espécies simpátricas, que coexistem sob condições ambientais idên-ticas, padrões gametogênicos completamente
CAPÍTULO 7
A gametogênese em Limnoperna fortunei
(Dunker, 1857)
Claudia T. CallilAna Lúcia Teixeira Gomes
Ana Cecília Pinillos Vinicius Soares

Callil et al.
A gametogênese em Limnoperna fortunei (Dunker, 1857)
112
distintos. Um exemplo desta plasticidade ocorre com duas espécies de bivalves nativos do gênero Anodontites: enquanto A. elongatus (Sawaison, 1829) é exclusivamente dioico, o seu parente e vizinho A. trapesialis (Lamarck, 1819) pode ser considerado um hermafrodita funcional convic-to, apresentando folículos femininos e masculi-nos ativos simultaneamente (Callil, 2003; Callil & Mansur, 2007; Callil, 2009). O mexilhão-dourado é dioico (Morton, 1982) (Fig. 1), apresenta sexos separados e é classificado por Choi & Shin (1985) como uma espécie gonocorística. Machos e fêmeas geral-mente estão presentes em iguais proporções, apesar de ocasionalmente ocorrer um ou outro caso de hermafroditismo (Tab. I). De todos os estudos disponíveis sobre reprodução e sex--ratio, apenas em dois locais foram observados indivíduos hermafroditas: no Rio da Prata, na Argentina (Darrigran et al., 1998b; 1999) e no Rio Paraguai em Corumbá, MS (Uliana & Callil, 2006). Apesar da baixa porcentagem
de hermafroditismo em mexilhão-dourado, quando ocorre, os gametas femininos e mas-culinos são produzidos em folículos distintos (Fig. 2). Para os Mytilidae, descritos geralmen-te como dioicos, o hermafroditismo esporádi-co parece ser possível (Villalobos et al., 2010), porém, raro. Entre 4.309 espécimes de Peru-mytilus purpuratus (Lamarck, 1819) analisados houve apenas um registro de hermafroditis-mo (0,023%) (Villalon apud Villalobos et al., 2010) e o primeiro registro para o mexilhão Aulacomya ater (Molina, 1782) foi feito por Vi-nuesa (1977). Em Corbicula fluminea (Müller, 1774) a incidência de hermafroditismo é me-nos rara do que em mexilhão-dourado, porém ainda considerada como ocasional (Massoli & Callil, 2012). Situação semelhante ocorre em Dreissena polymorpha (Pallas, 1771) espécie dioica, a qual é relatada cerca de 4 a 8% de hermafroditismo (Nichols, 1993). Seria opor-tuno investigar as causas do hermafroditismo em espécies dioicas de comportamento invasor,
Tabela I. Frequência de ocorrência de machos, fêmeas e hermafroditas de Limnoperna fortunei.
Fonte Local Ano Machos Fêmeas Não Ident. Hermafroditas
Morton, 1982 Sudeste da Ásia 1971/74 34,30 65,70 0,00 0,00Darrigran et al., 1999 Rio da Prata, AR 1992/94 45,00 40,00 15,00 0,00Darrigran et al., 2003 Rio da Prata, AR 1992/94 61,47 38,28 0,00 0,25Uliana & Callil, 2006 Rio Paraguai, MS 2004/06 35,00 56,11 8,89 0,00Marcelo & Callil, 2006 Praia do Veludo, RS 2002 45,10 54,90 0,00 0,00Pestana et al., 2008 Res. Itaipu, PR 2003/05 55,80 26,90 15,40 0,00Uliana et al., 2006 Corumbá, MS 2006 42,00 46,60 10,60 0,60Gomes & Callil, 2010 Rio Paraná, PR 2006/08 37,00 40,00 23,00 0,00Pinillos & Callil, 2011a Rio Baía, PR 2006/08 41,76 55,58 2,35 0,00Pinillos & Callil, 2011b Ilha da Pintada, RS 2010 36,00 50,00 14,00 0,00Gomes & Callil, 2011 Cais do Porto, RS 2010 37,00 56,20 6,00 0,00
Figura 1. Diferenciação sexual em indivíduos de mexilhão-dourado, Limnoperna fortunei ativos reprodutivamente (A). Vista superficial externa em microscópio estereoscópico, do manto de um indivíduo feminino em início de maturação. As estruturas na ponta das setas são os folículos em desenvolvimento (B). Fotos: A, A.L.T. Gomes e B, C.T. Callil.

CAPÍTULO 7
Seção 2
113
como L. fortunei, D. polymorfa e C. fluminea, mesmo porque, apesar de apresentarem esta se-melhança no comportamento reprodutivo, essas espécies pertencem a grupos filogenéticos distin-tos. Tal comportamento, mesmo que em baixas proporções, provavelmente seja acessado como uma estratégia de estabelecimento populacional no início de instalação dos bioinvasores.
Estruturas reprodutivas No mexilhão-dourado, assim como em outros Mytilideos, a produção de gametas ocorre em estruturas acinosas que se distribuem naturalmente pelo manto e ocasionalmente na massa visceral (Fig. 1) (Morton, 1973, 1982). Não há dimorfismo sexual evidente, po-rém após a abertura da concha, apesar da mor-fologia externa do manto ser semelhante entre os machos e as fêmeas, a presença de ovócitos ou esperma pode ser identificada a olho nu, por padrões de textura e coloração distintos, comuns nos Mytilidae. Fêmeas maduras apre-sentam uma coloração que varia em tons de castanho claro a acobreado quando no pico de maturação, enquanto que os machos, nesse momento, apresentam o manto branco leitoso (Fig. 1). Esta diferenciação entre machos e fê-meas é ainda mais evidente no marisco Perna perna (Linnaeus, 1758), um parente marinho do mexilhão-dourado, em que nos machos a coloração é branca-leitosa e nas fêmeas verme-lha-alaranjada (Lunetta, 1969). Incontestavelmente, informações mais confiáveis e úteis sobre a morfologia são ob-tidas apenas a partir de preparações histoló-gicas das gônadas (Seed & Suchanek, 1992). Embora trabalhoso (provavelmente a razão principal para o seu uso limitado), este método oferece informações detalhadas sobre os teci-dos e possibilita acompanhar pormenores do ciclo reprodutivo completo, incluindo o tempo real de desova. A identificação e descrição dos elementos morfológicos e gametogênicos uti-lizados para o monitoramento do ciclo sexual estão representados na Figura 2. Nas fêmeas, a gônada é composta de vesículas, geralmen-te denominadas de ácinos (Bielefeld, 1991) ou de folículos ovarianos (Haag & Garton, 1992), por ser o local aonde os ovócitos irão se desen-volver (Fig. 3). Nos machos, as vesículas onde
os espermatozoides se desenvolvem têm sido denominadas de lobos espermatogênicos (Gar-ton & Haag, 1993) (Fig. 4). Os ovócitos e espermatozoides apresen-tam produção cíclica, culminando em um ou mais momentos de desova, comumente asso-ciada a fatores ambientais. Esses processos co-ordenados (maturação e desova) dos gametas são importantes para uma reprodução bem sucedida. Para estudar a gametogênese é usual categorizar e caracterizar as sucessivas altera-ções que os folículos e lobos passam durante a gametogênese e, para isso, apresentamos uma descrição simplificada, organizada em uma es-cala, compreendendo cinco fases de desenvol-vimento gametogênico para ambos os sexos: IM – Início de Maturação, EM – Em Matu-ração, MA – Maturo, EL – Eliminando e RP – Repouso (Figs. 3 e 4). Estes estádios foram adaptados a partir de sistemas de classificação proposto por Callil (2003) e validadas com base em Damborenea & Penchaszadeh (2006). A aplicação de estádios de desenvolvi-mento foi por muito tempo utilizado, mesmo considerando a subjetividade do método, uma vez que, apesar de serem elencados critérios morfo-histológicos para a classificação, o trei-namento e ‘olhar’ do pesquisador podem ser tendenciosos. Daí a importância de avaliações quantitativas (Callil & Mansur, 2007), minimi-zando a subjetividade categórica e transforman-do medidas como diâmetro, área ou perímetro de ovócitos, folículos e lobos em variáveis con-tínuas, mais robustas analiticamente. Pensando nisso, Pinillos & Callil, (2011a, b) validaram quantitativamente os estádios de maturação, a partir de aproximadamente 15 mil folículos mensurados provenientes de amostras mensais de 15 indivíduos oriundos do Rio Baía, PR. A variância de dados biométricos dos elementos gametogênicos (diâmetro, área e perímetro de ovócitos, folículos femininos e lobos esperma-togênicos) foi considerada como variável-res-posta e os estádios como a variável-preditora. O diâmetro (20-300 μm); área (36-4525,3 μm) e perímetro (85,4-876,5 μm) diferem significa-tivamente (p<0,001) quando considerados os diferentes estádios (IM, EM, MA e EL). O está-dio repouso (RP) não foi considerado devido ao fato da população, no local estudado, apresen-tar desenvolvimento gametogênicos contínuo.

Callil et al.
A gametogênese em Limnoperna fortunei (Dunker, 1857)
114
Figura 2. Morfologia microscópica das estruturas gametogênicas utilizadas como elementos de classificação dos estádios de desenvolvimento folicular. A) manto feminino, B) manto masculino, C) folículos femininos, D) detalhes dos folículos femininos, E) folículos masculinos, F) detalhes dos folículos masculinos, G) hermafrodita com manto masculino apresentando folículo feminino, H) hermafrodita com manto feminino apresentando folículo masculino.
Manto: a epiderme do manto é caracteriza-da externamente por um epitélio simples de células cúbicas (Ee) e internamente por uma camada simples de células colunares (Ei). Na porção mediana observa-se um estroma de preenchimento (Esp), que fica quase que imperceptível durante perío-dos de intensa atividade gametogênica; o manto está completamente ocupado por folículos e lobos gametogênicos.
Folículos e gametas femininos: estruturas acinosas, delimitadas pelo epitélio folicu-lar (Ef) composto por células germinativas primordiais femininas (cgpf), que vão dar origem às ovogônias (ovg). Estas se de-senvolvem em ovócitos (ovc), onde são evidentes o núcleo (n) e nucléolo (nc). É possível observar grânulos de vitelo (v) nos ovócitos maduros e em eliminação.
Lobos e gametas masculinos: estruturas acinosas (Lm), delimitadas pelo epité-lio folicular (efm), composto por células germinativas primordiais (cgp), que dão origem às espermatogônias (esg) e vão se diferenciar na série espermática radial (SER): espermatócitos primários (ep1) e secundários (ep2); espermátides (epm) e espermatozoides (epz), que são elimina-dos através de ductos (dc).
Hermafroditismo: ocorrência esporádica de um ou mais folículos masculinos den-tre os femininos, podendo ocorrer o in-verso. Darrigran et al. (1999) observaram produção de ovócitos e espermatozoides simultaneamente em um mesmo folículo.
Ciclo reprodutivo
Os primeiros estudos que investigaram a produção de gametas em mexilhão-dourado são oriundos dos locais de origem desta espé-cie. Na China, durante a fase de estabelecimen-to, a espécie apresentou dois picos reprodutivos anuais Morton (1982), e em Kyoto, no Japão, a atividade reprodutiva foi descrita como ocor-rendo apenas uma vez ao ano (Iwasaki & Uryu, 1998). Segundo Morton (1982), duas desovas
anuais são comuns em bivalves que ocupam águas rasas. Durante o início do processo de estabe-lecimento do mexilhão-dourado na América do Sul, a produção de gametas foi contínua com proliferação permanente de elementos ga-metogênicos. Cinco picos de eliminação anu-ais foram inicialmente descritos no Balneario Bagliardi, Rio da Prata, Argentina (Darrigran et al., 1999). Boltovskoy & Cataldo (1999) in-feriram uma desova contínua com base na den-

CAPÍTULO 7
Seção 2
115sidade de larvas no ambiente, porém, diferen-ças entre dados histológicos e abundância de larvas no plâncton podem ser explicadas como resultante da deriva de larvas de aglomerados de indivíduos isolados em uma área de estudo contígua (Haag & Garton, 1992). No Brasil, durante a fase de estabeleci-mento da população na Praia do Veludo, Lago Guaíba, RS, a população eliminou gametas durante todo o período estudado (Marcelo & Callil, 2006). Em experimentos desenvol-vidos no reservatório de Itaipu, Rio Iguaçu, Silva (2006) descreve três picos anuais e ainda no Paraná, no Rio Baía, o mexilhão-dourado
também apresenta proliferação contínua com dois picos pronunciados de eliminação. Neste mesmo estudo, porém no Rio Paraná, local re-lativamente próximo, a gametogênese não foi contínua, apresentando por três meses conse-cutivos, um período de inatividade gonadal – Repouso (Gomes & Callil, 2010; Callil et al., 2012). Os locais estudados no Rio Paraná e no Rio Baía são relativamente próximos, porém dis-tintos quanto às características ambientais. Esse padrão de interrupção do processo gametogêni-co, entrando em estado de Repouso, também foi detectado no Rio Paraguai, em Corumbá, MS, região central do Brasil (Uliana et al., 2008).
Figura 3. Sucessão de estádios de desenvolvimento de gametas femininos de Limnoperna fortunei.
IM – Início da Maturação: manto com epiderme externa evidente e estroma de preenchimento abundante. Os folículos femininos são pequenos e esparsos e apresentam o epitélio folicular espessado pela presença de elementos proliferati-vos – ovogônias; é possível observar raros ovócitos em início de desenvolvimento.
EM – Em Maturação: manto com presen-ça de folículos femininos em expansão. Internamente os ovócitos de tamanhos variados estão em plena proliferação. Epitélio folicular está se estendendo em detrimento do aumento do volume dos ovócitos. Processo de vitelogênese ativo.
MA – Maturo: manto completamente ocu-pado por folículos repletos de gametas femininos. O intenso processo de vitelo-gênese promove aumento expressivo no tamanho dos ovócitos, tornando-os de forma poliédrica. Núcleos e nucléolos aparentes.
EE – Em Eliminação: observa-se o início do esvaziamento do manto e dos folículos. Os folículos femininos apresentam espa-ços entre os ovócitos que estão sendo eliminados através dos ductos. Os ductos são constituídos de um epitélio simples de células cuboides, densamente ciliadas internamente.

Callil et al.
A gametogênese em Limnoperna fortunei (Dunker, 1857)
116
Tabela II. Revisão dos estudos relacionados ao ciclo reprodutivo de mexilhão-dourado na América do Sul durante o período de instalação e estabelecimento do mexilhão-dourado, Limnoperna fortunei.
Fonte Local Períodoamostral Início Final Abordagem Variação
dos dados Reprodução Repouso
Morton, 1982 Hong Kong, CH
3 anos 1971 1974 Gametas 10,0-60,0μm mai/jun; nov/dez
ausente
Darrigran et al., 1999
Rio de La Plata, AR 30 meses jul/92 nov/94 Gametas 41,19-64,37μm contínuo ausente
Cataldo & Bolto-vskoy, 2000
Rio de La Plata, AR
17 meses set/97 jan/99 Larvas0-28.000 ind.m-3 ago/abri mai/jul
Darrigran et al., 2002
Rio Paraná, AR
14 meses abr/99 mai/01 Larvas 0-259,3 ind.m-3 out/mar mai/ago
Uliana & Callil, 2006
Rio Paraguai, MS 18 meses ago/04 mar/06 Gametas
5,27-127,52 μm out/fev jun/ago
Marcelo & Callil, 2006 Rio Jacuí, RS 12 meses jan/02 dez/02 Gametas 31,0-65,5 μm contínuo ausente
Pestana et al., 2008
Rio Paraná, PR
13 meses jan/05 fev/06 Larvas 948,5 ind.m-3 jan/jun jul/ago
Pestana et al., 2008
Itaipu, R. Iguaçu, PR 13 meses jan/05 fev/06 Larvas 2.755 ind.m-3 set/fev jul/ago
Boltovskoy et al., 2009a
Paraná-Prata, AR 10 anos 1997 2006 Larvas 450-7.000
ind.m-3 verão> 17ºC inverno < 17ºC
Nakano et al., 2010
Lago Takenuma, JP
18 meses jun/07 nov/08 Larvas 0-200 ind.m-3 jul/set (17-28ºC)
out/mai
Nakano et al., 2010
Lago Ohshio, JP 18 meses jun/07 nov/08 Larvas
0-10.000 ind.m-3
jul/set (17-28ºC) out/mai
Oliveira et al., 2011
Rio Paraguai, MS 47 meses jan/04 nov/07 Larvas 1.629 ±
4.571set/abr
(28-35ºC) ausente
Oliveira et al., 2011
Rio Miranda, MS
47 meses jan/04 nov/07 Larvas 990 ± 2.292 ago/set fev/mar
Morton (1982) já afirmava que seria muito difícil relacionar algum padrão de deso-va, não apenas para L. fortunei, como também para D. polymorpha e C. fluminea, porque estas espécies geralmente são oportunistas, podendo apresentar diversas estratégias para ocupar um gradiente ambiental. A literatura disponível ilustra a afirmação de Morton (1982) e eviden-cia que durante o processo de estabelecimento da espécie em um novo ambiente, ainda não existem padrões definidos, uma vez que há uma grande variação na gametogênese e mo-mentos de desova e, consequentemente, nas informações relacionadas às densidades larvais e ao período em que as larvas ocorrem em ele-vadas densidades.
Influências ambientais na gametogênese
Muito se discute sobre a influência de fatores ambientais como agentes determinan-
tes de respostas reprodutivas. A temperatura, disponibilidade de alimentos e os efeitos de adensamento entre mexilhões vizinhos (macro-fouling) têm sido descritas como variáveis críti-cas que influenciam no processo de produção e eliminação de gametas. No caso de organismos de ampla distribuição, a identificação de fatores que interferem diretamente em determinados processos biológicos pode ser bem complicada. Desde o início da invasão, durante o processo de instalação de L. fortunei, a partir de uma detalhada revisão da pouca literatura disponí-vel até então, quase que exclusivamente rela-cionada à distribuição desta espécie, Ricciardi (1998) já sugeriu alguns limites de tolerância às variáveis físicas e químicas da água. Todos os trabalhos disponíveis sobre a gametogênese do mexilhão-dourado associam quase que exclusivamente à temperatura como fator determinante do desenvolvimento game-togênico (Darrigran et al., 1999; Maroñas et al., 2003; Darrigran et al., 2003; Damborenea

CAPÍTULO 7
Seção 2
117Figura 4. Sucessão de estádios de desenvolvimento de gametas masculinos de Limnoperna fortunei.
& Penchaszadeh, 2006) e larval (Darrigran, et al., 2002; Cataldo et al., 2005a; Bolovskoy et al., 2009; Pestana et al., 2008; Oliveira et al., 2011). É de se esperar que a variação cícli-ca da temperatura não esteja diretamente re-lacionada ao ciclo gametogênico, mas sim, as oscilações repentinas de temperatura (Morton, 1982), que seriam o gatilho que dispara a deso-va (Ferreira & Magalhães, 2002). Entretanto, é quase consenso a ideia de que o efeito siner-gético de um conjunto de variáveis ambientais atua sincrônica e simultaneamente no ciclo re-produtivo. Morton (1982) já havia sinalizado que existe uma multiplicidade de fatores que determinam mudanças associadas à qualidade
de água e absolutamente um único fator pode não ser o determinante exclusivo pelo início e taxa de gametogênese. Em locais onde as variações de tempe-ratura são inexpressivas, as características da água podem ser reguladas pela dinâmica hí-drica, típica das planícies de inundação sazo-nais, como o Pantanal e a Planície Aluvial do Rio Paraná. Nestas áreas úmidas, o pulso de inundação periódico (Junk et al., 1989) pode ser considerado o espelho que resume o siner-gismo de fatores que regulam a dinâmica sexu-al em moluscos límnicos (Callil, 2003; Callil & Mansur, 2007; Callil, 2009; Oliveira et al., 2011; Callil et al., 2012).
IM – Início da Maturação: no manto, o estroma de preenchimento apresenta inúmeras células pequenas e isoladas, ocupando a maior parte do manto. Lobos gametogênicos reduzidos e escassos. No epitélio folicular são visíveis espermato-gônias, sempre periféricas, e espermató-citos primários desordenados.
EM – Em Maturação: expansão dos lobos pelo manto, que ainda apresenta-se ra-zoavelmente ocupado pelo estroma. Nos lobos, camadas de espermatogônias des-prendem-se do epitélio e começam a se diferenciar em espermatócitos primários. Apesar das células entrarem em meiose, ainda não é evidente a definição da série espermática radial.
MA – Maturação: os lobos aumentam mui-to em número e tamanho e preenchem completamente o manto. A série esper-mática radial torna-se evidente e bem organizada; os espermatozoides, aos mi-lhares, aparecem sempre associados ao lúmen folicular.
EE – Em Eliminação: lobos ainda ocupam todo o manto, porém, com lúmen eviden-te. Série espermática radial frouxamente desorganizada. O lúmen está mais amplo que no estádio anterior e há a presença – quase que exclusiva – de espermatozoi-des, que também podem ser visualizados ocupando toda a extensão dos ductos.

Callil et al.
A gametogênese em Limnoperna fortunei (Dunker, 1857)
118
Sabemos que, para detectar fatores li-mitantes, experimentos rigorosos em campo e laboratório devem ser desenvolvidos a fim de testar, inclusive, a variabilidade da tolerância. Estudos relacionados à temperatura como fa-tor regulador do desenvolvimento larval foram conduzidos por Cataldo et al. (2005a), compro-vando que há uma relação direta entre o in-cremento das taxas de desenvolvimento com o aumento de temperatura. Oliveira et al. (2011) demonstram que a temperatura, profundidade e velocidade da água são os fatores que afetam estágios imaturos da população de mexilhão--dourado no Rio Paraguai, em Corumbá, e dis-cutem que o efeito combinado de várias con-dições causadas por diferentes fatores podem explicar as baixas densidades em alguns locais do Pantanal. Além dos fatores abióticos, devemos le-var em consideração as interferências bióticas como qualidade e disponibilidade de recursos alimentares e interações inter e intraespecífi-cas. O efeito da alimentação x sucesso repro-dutivo tem sido atribuído por Borcherding (1991), Ram et al. (1996) e Claxton & Mackie (1998), sendo que a qualidade de alimento pode influenciar inclusive o tamanho dos ovó-citos (Wacker & Von Elert, 2003; Wilhelm & Adrian, 2007). Morton (1982) ainda conside-rou a possibilidade de que a inalação de esper-matozoides de indivíduos da mesma espécie induz à desova simultânea. Esta estratégia é
vantajosa para indivíduos que formam aglome-rados, maximizando o sucesso de fecundação, principalmente durante o processo de instala-ção e estabelecimento da espécie em um novo ambiente. Apesar de seus impactos ecológicos e econômicos o mexilhão-dourado pode ser usa-do como um novo recurso e de fácil obtenção para estudos de mecanismos de reprodução.
Fecundação
A proximidade de indivíduos de sexos opostos dentro dos agrupamentos, macrofou-ling, facilita o sucesso de fecundação externa. A fecundação desta espécie é externa e todas as fases de desenvolvimento larval (mórula, trocó-fora, larva D, véliger, pedivéliger e plantígrada) são planctônicas, até que a fase de recruta irá colonizar novos locais (Boltovskoy et al., 1999; Santos et al., 2005). Liu et al. (apud Morton, 1982) afirmaram que L. fortunei incubam os ovos dentro das quatro demibrânquias, porém, o próprio Morton não confirma esta informação e ainda questiona este tipo de comportamento, uma vez que não é usual dentre os Mytilidae, os quais não apresentam estruturas adequadas para a função de incubação nos ctenídeos. Nós temos processado histologicamente várias cen-tenas de indivíduos e em momento algum foram observados casos de incubação, corroborando os questionamentos de Morton (1982).

119
1. Introdução
Espécies asiáticas de Corbicula ocorrem na América do Sul desde o final da década de 60 e início da de 70, no entanto, pouco ainda se sabe sobre suas estratégias reprodutivas ao lon-go de gradientes climáticos diferenciados nos mananciais que ocuparam e seguem invadindo.Até há pouco tempo, os dados na literatura eram extremamente contraditórios quanto à reprodução das corbículas de origem asiática. C. fluminea (Müller, 1774) foi considerada dioica com protandria, isto é, as gônadas de um mesmo indivíduo seriam primeiramente masculinas e mais tarde se tornariam femini-nas (Britton & Morton, 1982). Na América do Norte, Kraemer et al. (1986) observaram que C. fluminea é monoica com hermafroditismo simultânea, ou seja, os folículos reprodutivos são masculinos e femininos no mesmo indiví-duo. Mais tarde, Morton (1986) descreveu a espécie C. fluminea da Ásia como dioica com uma alta percentagem de hermafroditas em
ambientes lênticos, porém hermafrodita com igual percentagem de fêmeas e machos no am-biente lótico. Além disso, várias características incomuns da biologia reprodutiva, tais como poliploidia, esperma biflagelados diploides, androgênese e clonalidade foram também ob-servadas (Komaru & Konishi, 1996, 1999; Ko-maru et al., 1997; Qiu et al., 2001). Kraemer & Galloway (1986) e Krae-mer et al. (1986) constataram autofecunda-ção e fecundação cruzada em C. fluminea, e a presença de um espermatozoide biflagelado. Estudos mais recentes surpreenderam com as descobertas de Park & Chung (2004), que dis-cordam quanto à autofecundação. Estes auto-res observaram que: C. fluminea se reproduz por partenogênese, sem autofertilização ou fecundação cruzada por óvulos ou espermato-zoides; que a espécie é um hermafrodita fun-cional, com as gônadas formadas por folículos masculinos, femininos ou hermafroditas, no mesmo indivíduo; que no folículo hermafro-dita, os óvulos são circundados por inúme-
CAPÍTULO 8
Dinâmica reprodutiva de Corbicula fluminea e Corbicula largillierti
Maria Cristina Dreher MansurAline Salvador Vanin
Paulo Eduardo Aydos BergonciArthur Schramm de Oliveira

Mansur et al.
Dinâmica reprodutiva de Corbicula fluminea e Corbicula largillierti
120
ros espermatozoides que atuariam apenas no estímulo para a clivagem inicial dos óvulos maduros para a partenogênese. Constataram também triploidia, o que impediria uma fe-cundação cruzada normal. Através dos estudos genéticos desenvol-vidos por Lee et al. (2005) sobre as popula-ções de Corbicula do continente americano (incluindo populações do Rio Paraná), foi constatada que grande parte da população seria triploide. Duas linhagens minoritárias, ocorrendo em regiões limitadas, foram ainda detectadas com um padrão diferenciado em relação aos mitocôndrios e ribossomos. No entanto, todas as populações apresentam o es-permatozoide biflagelado, que seria o marca-dor morfológico da clonalidade da espécie. Para Fox (2004), os espermatozoides que são liberados para o ambiente via aber-tura exalante, formam, ainda nos folículos, grandes esferas denominadas esferas ou mó-rulas espermáticas ou aglomerados espermáti-cos (“sperm clusters”) segundo Park & Chung (2004). Cada esfera é formada por centenas de espermatozoides. Esta esfera, uma vez liberada no ambiente, entra através do sifão inalante na cavidade palial de um outro indivíduo. Ali, a mórula se desmonta em espermatozoides indi-viduais que atravessam as brânquias penetran-do na câmara suprabranquial onde os óvulos presentes são fertilizados. Estes óvulos, agora embrionados, são incubados dentro dos tubos aquíferos das demibrânquias, na área do mar-súpio, permanecendo ali até a fase de pedivé-liger quando são liberados ao ambiente pela abertura exalante. Britton & Morton (1982) comentam que o gênero Corbicula pode estar sujeito às pressões ambientais, tanto de espécies consi-deradas “r” estrategistas quanto as “k” estra-tegistas, o que explicaria seu grande sucesso como invasora nos mananciais norte-america-nos. Sob esta perspectiva, populações locali-zadas em ambientes de águas lênticas, como reservatórios, por exemplo, tendem a investir no crescimento corporal, podendo atrasar o período reprodutivo e até mesmo exibir fe-cundidade reduzida. Em habitat nativo, onde estão sujeitas a condições variáveis e menos estáveis, estes moluscos podem rapidamente se ajustar como “r” estrategistas.
2. Períodos reprodutivos
Segundo Fox (2004), a liberação das lar-vas acontece duas vezes por ano: na primavera e no outono. Na América do Sul, estudos revelaram que C. fluminea apresenta um evento repro-dutivo por ano: Cataldo & Boltovskoy (1999) relataram para populações do Rio Paraná, um período reprodutivo entre os meses de outu-bro e novembro; Ituarte (1985) observou na população do Rio da Prata, um pico no mês de setembro. Massoli-Junior (2006) e Massoli--Junior & Callil (2012), ao analisarem aspectos populacionais e reprodutivos de uma popula-ção da praia da Vereda, Rio Cuiabá, Município de Santo Antônio do Leverger, MT, constata-ram que o início do recrutamento ocorreu no começo da estiagem (junho de 2005), o declí-nio populacional no começo da cheia (dezem-bro de 2004 e 2005) e um pico de atividade reprodutiva no período da seca (setembro de 2005), com elevada proporção de indivíduos maturos e em maturação e com os maiores di-âmetros de folículos e ovócitos. Observaram, ainda, a predominância de folículos femininos em relação aos masculinos e hermafroditas. Sobre aspectos reprodutivos da espécie C. largillierti (Philippi, 1844) encontram-se dados apenas em Ituarte (1984a) que estudou a dinâ-mica populacional desta espécie no Rio da Prata, Argentina. O autor a descreveu como hermafro-dita funcional com os tecidos masculinos e femi-ninos não claramente segregados na massa visce-ral, e uma fração maior do feminino. Descreveu também um ciclo reprodutivo bianual, com a li-beração de larvas pedivéliger na primavera e no verão e um tempo de vida aproximado de 24 a 30 meses. Porém, encontrou ao longo de todo o ano, indivíduos com óvulos maduros, e conside-rou que a temperatura seria o fator regulador do sincronismo para a liberação das larvas. Obser-vou também que as gônadas se desenvolvem cedo nos indivíduos e que 26% da população apresen-taram indivíduos com embriões nas brânquias, a partir dos 11 a 12 mm de comprimento. A in-cubação dos embriões ocorre nas demibrânquias internas e a liberação dos mesmos se deu quando atingiram a fase de larva véliger tardio (225 a 230 μm de comprimento) ou de pedivéliger (235 a 240 μm de comprimento).

CAPÍTULO 8
Seção 2
121
3. Aspectos da dinâmica reprodutiva de C. largillierti e C. fluminea em um reservatório tropical brasileiro. Estudos recentes sobre duas populações (C. fluminea e C. largillierti) amostradas men-salmente (de fevereiro de 2010 a abril de 2011) no reservatório de Peixe-Angical, município de Peixe, rio Tocantins, avaliaram a ocorrência de larvas incubadas (Mansur et al., 2011c). Exem-plares foram coletados na margem do reserva-tório, anestesiados com mentol e fixados em álcool 70%. As valvas foram entreabertas para a retirada das brânquias e estas examinadas quanto à presença e ausência de embriões (Fig. 1). As larvas foram separadas, homogeneizadas em câmara de contagem (100 quadrículas/5 mm) (Fig. 2) sob a lupa. Quando em pequeno número (<100) a contagem foi absoluta, acima deste valor foi por sorteio de 10 quadrículas e multiplicadas por 10. A comparação entre o numero médio de indivíduos incubados por espécie foi realizada por meio do teste de Mann-Whitney (p =0,05). O mesmo teste foi aplicado para a comparação entre as estações do ano (seca e chuvosa) para cada espécie. A temperatura na água do reservatório variou de 26,4ºC a 34ºC, com as mais elevadas de novembro a fevereiro. Os períodos de seca na área estudada vão de maio a setembro e os de cheia, de outubro a abril. Os resultados revelaram que durante o período de fevereiro de 2010 a abril de 2011, aproximadamente 1/3 dos indivíduos adultos apresentaram larvas incubadas nas brânquias. Dentre os 909 espécimes de C. largillierti, 231 apresentaram larvas incubadas nas brânquias, ou seja, 25,41% da população analisada es-tavam em atividade reprodutiva. De maneira semelhante, C. fluminea apresentou 334 in-divíduos incubados dentre os 1217 indivíduos analisados, isto é, 27,44% do total estavam em processo reprodutivo, com larvas incubadas nas brânquias. O exame do conteúdo larval junto às brânquias revelou que as populações das espé-cies C. largillierti e C. fluminea apresentaram produção larval praticamente durante todo o ano (Figs. 3 e 4), tanto durante o período de
seca (maio a setembro) quanto no chuvoso (ou-tubro a abril) (Fig. 4). Com base na Figura 5 também se observa que C. largillierti apresenta uma diferença de cerca de 10% entre os perío-dos de seca e chuva, provavelmente relacionada à falta de exemplares desta espécie nos meses de outubro e dezembro de 2010 e janeiro de 2011 (Fig. 3). Em fevereiro (16/02/2011) constatou-se, em três exemplares de C. largillierti, ovos em clivagem inicial, em grande quantidade, e ra-ras larvas em estágio de pós-larva ainda retidas nos marsúpios, demonstrando a continuidade do ciclo (Fig. 6). A incubação de larvas nas demibrânquias interna e externa dos bivalves analisados revelou que tanto para C. largillierti quanto para C. fluminea, a incubação ocorre em sua maior parte nas demibrânquias internas e eventualmente aparecem embriões esparsos nas externas, porém em baixa quantidade (Fig. 7). Comparando apenas a ocorrência de embri-ões nas demibrânquias externas, C. largillierti apresentou proporcionalmente maior ocorrên-cia de larvas do que C. fluminea. A observação dos estágios larvais revelou que o desenvolvi-mento dos embriões e a liberação das larvas é sincronizado nas duas espécies.
Figura 1. Brânquia direita de Corbicula largillierti com a demib-rânquia externa rebatida e a interna contendo embriões encu-bados. Legenda: DBE – demibrânquia externa, DBI – demibrân-quia interna, E – embriões, M – marsúpio. Foto: M.C.D. Mansur
Figura 2. Câmara de contagem (100 quadrículas/ 5 mm). Foto: M.C.D. Mansur

Mansur et al.
Dinâmica reprodutiva de Corbicula fluminea e Corbicula largillierti
122
Figura 3. Variação temporal do percentual de indivíduos de Corbicula largillierti incubados no reservatório da UHE Peixe-Angical.
Figura 4. Variação temporal do percentual de indivíduos de Corbicula fluminea incubados no reservatório da UHE Peixe-Angical.
Figura 5. Percentual de indivíduos adultos incubados e não incubados das duas espécies de corbículas por estação chuvosa e seca.
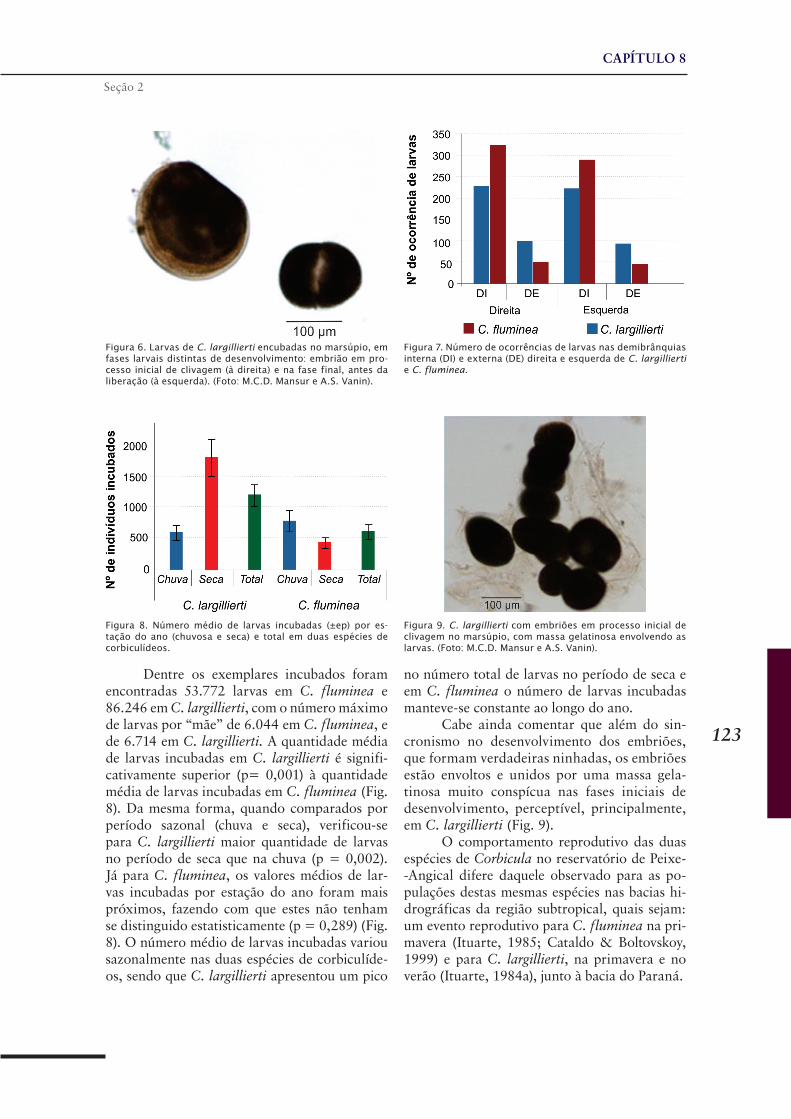
CAPÍTULO 8
Seção 2
123
Dentre os exemplares incubados foram encontradas 53.772 larvas em C. fluminea e 86.246 em C. largillierti, com o número máximo de larvas por “mãe” de 6.044 em C. fluminea, e de 6.714 em C. largillierti. A quantidade média de larvas incubadas em C. largillierti é signifi-cativamente superior (p= 0,001) à quantidade média de larvas incubadas em C. fluminea (Fig. 8). Da mesma forma, quando comparados por período sazonal (chuva e seca), verificou-se para C. largillierti maior quantidade de larvas no período de seca que na chuva (p = 0,002). Já para C. fluminea, os valores médios de lar-vas incubadas por estação do ano foram mais próximos, fazendo com que estes não tenham se distinguido estatisticamente (p = 0,289) (Fig. 8). O número médio de larvas incubadas variou sazonalmente nas duas espécies de corbiculíde-os, sendo que C. largillierti apresentou um pico
Figura 6. Larvas de C. largillierti encubadas no marsúpio, em fases larvais distintas de desenvolvimento: embrião em pro-cesso inicial de clivagem (à direita) e na fase final, antes da liberação (à esquerda). (Foto: M.C.D. Mansur e A.S. Vanin).
Figura 7. Número de ocorrências de larvas nas demibrânquias interna (DI) e externa (DE) direita e esquerda de C. largillierti e C. fluminea.
Figura 8. Número médio de larvas incubadas (±ep) por es-tação do ano (chuvosa e seca) e total em duas espécies de corbiculídeos.
no número total de larvas no período de seca e em C. fluminea o número de larvas incubadas manteve-se constante ao longo do ano. Cabe ainda comentar que além do sin-cronismo no desenvolvimento dos embriões, que formam verdadeiras ninhadas, os embriões estão envoltos e unidos por uma massa gela-tinosa muito conspícua nas fases iniciais de desenvolvimento, perceptível, principalmente, em C. largillierti (Fig. 9). O comportamento reprodutivo das duas espécies de Corbicula no reservatório de Peixe--Angical difere daquele observado para as po-pulações destas mesmas espécies nas bacias hi-drográficas da região subtropical, quais sejam: um evento reprodutivo para C. fluminea na pri-mavera (Ituarte, 1985; Cataldo & Boltovskoy, 1999) e para C. largillierti, na primavera e no verão (Ituarte, 1984a), junto à bacia do Paraná.
Figura 9. C. largillierti com embriões em processo inicial de clivagem no marsúpio, com massa gelatinosa envolvendo as larvas. (Foto: M.C.D. Mansur e A.S. Vanin).

Mansur et al.
Dinâmica reprodutiva de Corbicula fluminea e Corbicula largillierti
124
No Pantanal Norte, onde as temperatu-ras são sempre altas com ciclos de cheia e seca, acontece um evento reprodutivo no início da seca para C. fluminea (Massoli-Junior, 2006). No reservatório do Rio Tocantins, ambas as es-pécies C. fluminea e C. largillierti apresentam em torno de 26% dos indivíduos com presença de larvas incubadas durante todo o ano. O nú-mero médio de larvas incubadas em C. largillier-ti foi significativamente maior na estação seca. Segundo McMahon & Williams (1986)
temperaturas altas, próximas de 36ºC, seria o limite letal de populações naturais de C. flumi-nea nos EUA e temperaturas em torno de 30ºC impediriam a formação do cordão mucoso no juvenil, o que dificultaria o recrutamento dos animais junto ao substrato. No entanto, mesmo sob temperaturas extremas na água do reser-vatório, que podem chegar a 34ºC no período de novembro a fevereiro, não foram observados declínios na densidade larval nem populacional das duas espécies (Mansur et al., 2011c).

125
1. Introdução
A morfologia da concha é um dos mais tradicionais critérios para a identificação siste-mática dos moluscos. Mesmo considerando a convergência morfológica, que pode ser obser-vada em grupos sem relacionamento filogenéti-co próximo, ainda assim os detalhes da concha, sua ornamentação e escultura, são importan-tes auxiliares nesta tarefa. A Classe Gastropo-da é tradicionalmente dividida nas Subclasses Prosobranchia, Opistobranchia e Pulmonata (Hyman, 1967; Ruppert et al., 2005). Essa classificação admite como critério as sucessivas modificações da cavidade palial (= cavidade do manto) e das estruturas nela contidas. Assim, Prosobranchia significa que a cavidade palial e as estruturas associadas (entre elas brânquias e osfrádio) estão situadas na porção anterior da massa visceral, acima da cabeça do gastrópode (as brânquias estão situadas em posição anterior ao pericárdio); nos Opistobranchia, a cavidade palial e as brânquias, em consequência de um
processo chamado detorção da massa visceral, ocupam a posição posterior; nos Pulmonata, a cavidade palial ocupa a posição anterior, assim como nos prosobrânquios, mas as brânquias es-tão ausentes. Nesse caso, a troca gasosa passa a ser efetuada por uma extensa rede vascular no teto da cavidade palial. Em alguns grupos a cavidade palial é reduzida e em outros desa-parece por completo, como em algumas lesmas terrestres. Entre o molusco ancestral, o padrão prosobrânquio típico, o padrão opistobrânquio típico e o padrão pulmonado, existem diversas “etapas intermediárias” que suportam a hipó-tese gradista de evolução no sentido Proso-branchia-Opistobranchia-Pulmonata (Hyman, 1967; Brusca & Brusca, 2007). A Subclasse Pulmonata engloba gastró-podes límnicos e terrestres, com conchas espi-raladas, que podem estar reduzidas ou mesmo ausentes, geralmente sem opérculo, hermafro-ditas, com uma aurícula anterior ao ventrículo, sem brânquias e com a abertura da cavidade do manto modificada pela fusão com a região
CAPÍTULO 9
Gastrópodes límnicos invasores: morfologia comparada
Monica Ammon FernandezSonia Barbosa dos Santos
Igor Christo MiyahiraIsabela Cristina Brito Gonçalves
Renata de Freitas XimenesSilvana Carvalho Thiengo
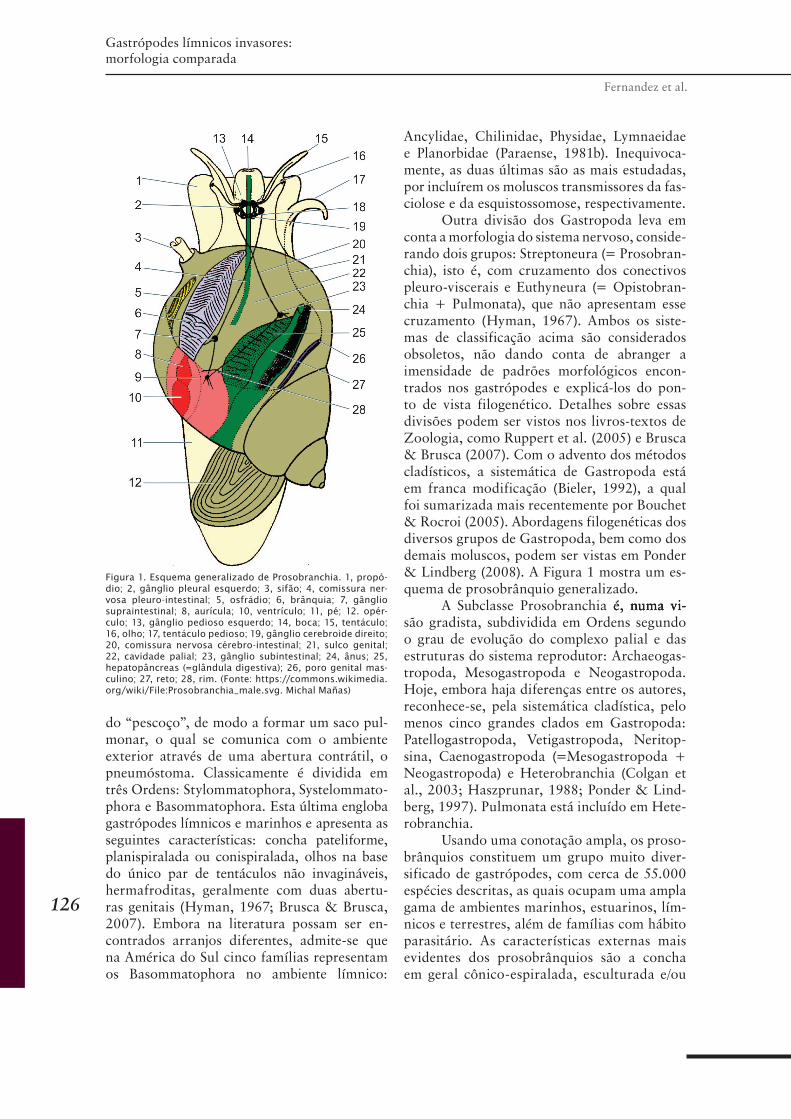
Fernandez et al.
Gastrópodes límnicos invasores: morfologia comparada
126
do “pescoço”, de modo a formar um saco pul-monar, o qual se comunica com o ambiente exterior através de uma abertura contrátil, o pneumóstoma. Classicamente é dividida em três Ordens: Stylommatophora, Systelommato-phora e Basommatophora. Esta última engloba gastrópodes límnicos e marinhos e apresenta as seguintes características: concha pateliforme, planispiralada ou conispiralada, olhos na base do único par de tentáculos não invagináveis, hermafroditas, geralmente com duas abertu-ras genitais (Hyman, 1967; Brusca & Brusca, 2007). Embora na literatura possam ser en-contrados arranjos diferentes, admite-se que na América do Sul cinco famílias representam os Basommatophora no ambiente límnico:
Ancylidae, Chilinidae, Physidae, Lymnaeidae e Planorbidae (Paraense, 1981b). Inequivoca-mente, as duas últimas são as mais estudadas, por incluírem os moluscos transmissores da fas-ciolose e da esquistossomose, respectivamente. Outra divisão dos Gastropoda leva em conta a morfologia do sistema nervoso, conside-rando dois grupos: Streptoneura (= Prosobran-chia), isto é, com cruzamento dos conectivos pleuro-viscerais e Euthyneura (= Opistobran-chia + Pulmonata), que não apresentam esse cruzamento (Hyman, 1967). Ambos os siste-mas de classificação acima são considerados obsoletos, não dando conta de abranger a imensidade de padrões morfológicos encon-trados nos gastrópodes e explicá-los do pon-to de vista filogenético. Detalhes sobre essas divisões podem ser vistos nos livros-textos de Zoologia, como Ruppert et al. (2005) e Brusca & Brusca (2007). Com o advento dos métodos cladísticos, a sistemática de Gastropoda está em franca modificação (Bieler, 1992), a qual foi sumarizada mais recentemente por Bouchet & Rocroi (2005). Abordagens filogenéticas dos diversos grupos de Gastropoda, bem como dos demais moluscos, podem ser vistas em Ponder & Lindberg (2008). A Figura 1 mostra um es-quema de prosobrânquio generalizado. A Subclasse Prosobranchia é, numa vi-é, numa vi- numa vi-são gradista, subdividida em Ordens segundo o grau de evolução do complexo palial e das estruturas do sistema reprodutor: Archaeogas-tropoda, Mesogastropoda e Neogastropoda. Hoje, embora haja diferenças entre os autores, reconhece-se, pela sistemática cladística, pelo menos cinco grandes clados em Gastropoda: Patellogastropoda, Vetigastropoda, Neritop-sina, Caenogastropoda (=Mesogastropoda + Neogastropoda) e Heterobranchia (Colgan et al., 2003; Haszprunar, 1988; Ponder & Lind-berg, 1997). Pulmonata está incluído em Hete-robranchia. Usando uma conotação ampla, os proso-brânquios constituem um grupo muito diver-sificado de gastrópodes, com cerca de 55.000 espécies descritas, as quais ocupam uma ampla gama de ambientes marinhos, estuarinos, lím-nicos e terrestres, além de famílias com hábito parasitário. As características externas mais evidentes dos prosobrânquios são a concha em geral cônico-espiralada, esculturada e/ou
Figura 1. Esquema generalizado de Prosobranchia. 1, propó-dio; 2, gânglio pleural esquerdo; 3, sifão; 4, comissura ner-vosa pleuro-intestinal; 5, osfrádio; 6, brânquia; 7, gânglio supraintestinal; 8, aurícula; 10, ventrículo; 11, pé; 12. opér-culo; 13, gânglio pedioso esquerdo; 14, boca; 15, tentáculo; 16, olho; 17, tentáculo pedioso; 19, gânglio cerebroide direito; 20, comissura nervosa cérebro-intestinal; 21, sulco genital; 22, cavidade palial; 23, gânglio subintestinal; 24, ânus; 25, hepatopâncreas (=glândula digestiva); 26, poro genital mas-culino; 27, reto; 28, rim. (Fonte: https://commons.wikimedia.org/wiki/File:Prosobranchia_male.svg. Michal Mañas)

CAPÍTULO 9
Seção 2
127
ornamentada, e a presença de um opérculo córneo ou calcário, preso à região dorsal do pé, que fecha a abertura da concha. Trata-se de um grupo predominantemente marinho, mas com diversas incursões na água doce (por exemplo, Ampullariidae, Hydrobiidae, Thia-ridae e Pleuroceridae) e no ambiente terrestre (por exemplo, Helicinidae e Cyclophoridae). A Figura 2 mostra alguns representantes do grupo em nossos sistemas límnicos.
2. Morfologia comparada de Melanoides tuberculata (Müller, 1774)
Melanoides tuberculata é um Caenogas-tropoda do clado Cerithioidea, considerado monofilético (Colgan et al., 2000; Simone, 2001; Lydeard et al., 2002) e um clado basal em Caenogastropoda (Strong et al., 2011). Na sistemática gradista está inserido na Ordem Mesogastropoda (Hyman, 1967). A morfolo-gia da concha turriforme, com pequena aber-tura, canal sifonal anterior e uma ou mais linhas espirais com tubérculos, espinhos ou nódulos, é considerada quase que exclusiva dos Cerithioidea (Sälgeback & Savazzi, 2006), representando diversas adaptações funcio-nais. Segundo Strong et al. (2011), o clado Ce-rithioidea é caracterizado pelos machos afáli-cos, pelos gonodutos abertos e reprodução via espermatóforos, caracteres considerados ple-siomórficos, além da complexidade do trato genital feminino. Melanoides tuberculata, incluída em Thia-ridae, pode ser confundida com outras espécies de moluscos límnicos, tanto nativas do Brasil, como as dos gêneros Aylacostoma Spix in Wag-ner, 1827 (Thiaridae) (Fig. 4F) e Doryssa H. & A.
Adams, 1854 (Pleuroceridae) (Fig. 2C), como as exóticas, como por exemplo Tarebia grani-fera (Lamarck, 1822) (Thiaridae). Esta última espécie ainda não foi registrada para o Brasil, mas está presente na América do Norte, Amé-rica Central e Caribe (Pointier et al., 1998). Na América do Sul, temos registros apenas na Ve-nezuela (Pointier et al., 1994; Amarista et al., 2001). Segundo Simone (2006), existem 33 espécies nominais de Aylacostoma no Brasil, as quais ocorrem em todas as regiões de nos-so país. No gênero Doryssa são reconhecidas 37 espécies nominais, as quais se distribuem principalmente no norte do Brasil, na região Amazônica (Simone, 2006). Ambos os gêne-ros guardam muitas semelhanças, sendo difícil separá-los somente pelos caracteres da concha (Morrison, 1954). Os Rissooidea límnicos no Brasil são de pequeno porte, com conchas na escala dos milí-metros, raramente alcançando 7 mm. Podem ser confundidos com jovens de M. tuberculata, com até cinco ou seis voltas, especialmente os gêne-ros com concha cônico-alongada, como Heleobia Simpson, 1865 e Idiopyrgus Pilsbry, 1911. Um exame sob binocular é capaz de diferenciá-los, pois em geral os Rissooidea límnicos apresen-tam conchas lisas, com voltas arredondadas e raramente o ápice é perdido. Nos ambientes de água doce do Brasil este grupo é representado por pequenos caramujos pertencentes aos gê-neros Littoridina Eydoux & Souleyet, 1852 (= Heleobia); Potamolithus Pilsbry, 1896; Lyrodes Döring, 1884; Sioliella Haas, 1949 e Idiopyrgus, segundo Simone (2006). Todavia, os Rissooidea límnicos no Brasil possuem a concha lisa (Fig. 2A), enquanto nos tiarídeos a concha é bastante esculturada (Fig. 2D).
Figura 2. Conchas de repre-sentantes de prosobrânquios em ambientes límnicos no Brasil: A, Hydrobiidae (Ris-sooidea); B, Ampullariidae, C, Pleuroceridae D, Thiari-dae. Escala, 1 cm. (Fotos: A, S. Almeida; B e D, P.M. Coe-lho; C, L.R. Simone).

Fernandez et al.
Gastrópodes límnicos invasores: morfologia comparada
128
2.1. Morfologia externa
No Brasil existem poucos estudos sobre a morfologia de M. tuberculata. Simone (2001, 2006) apresenta comparações da morfologia da concha e das partes moles com outros Cerithioi-dea. Descreve a concha como sendo de tamanho médio (cerca de 3 cm), cuja escultura apresenta aspecto reticulado. O perióstraco é escuro, es-pesso, e o óstraco bege, com manchas marrons em disposição axial. Simone (2001) também afir-ma que a presença de uma escultura radial bem marcada distingue M. tuberculata das espécies de Aylacostoma. Gonçalves et al. (2010) acom-panharam uma população de M. tuberculata du-rante três anos na Ilha Grande, Angra dos Reis, RJ, e observaram a existência de três morfotipos (Fig. 3), baseados nos padrões de coloração. Lago et al. (2009) analisaram a conquiliometria de 11 populações de M. tuberculata oriundas do estado do Rio de Janeiro, provenientes de dois diferentes biótopos (rio e vala), encontrando diferença sig-nificativa apenas no diâmetro da abertura entre conchas dos dois biótopos. Apesar das semelhanças com espécies nativas de Thiaridae e de Pleuroceridae, alguns detalhes permitem a sua diferenciação. As con-chas de Pleuroceridae e de Thiaridae nativos, usualmente são grossas e apresentam forte es-culturação, composta por nódulos e costelas (Fig. 2 C-D, Fig. 4F). É difícil estabelecer um quadro comparativo, pois existe uma ampla va-riação de forma nas espécies dos gêneros, que se refletem nas proporções da concha e na colo-ração, como se observa em M. tuberculata (Fig. 4 A-E). Talvez a característica mais conspícua seja a espessura da concha, muito grossa e re-sistente em Aylacostoma e em Doryssa, quando comparadas à Melanoides Oliver, 1804, cuja concha é mais delicada, apesar de espessa. A concha de Aylacostoma é alongada, mas existe uma tendência ao abaulamento da volta corporal em muitas espécies, com for-mação de uma quilha ou carena (Fig. 4F); em Melanoides a concha nunca forma carena e o perfil alongado cresce suavemente, sem abaula-mento pronunciado da volta corporal, que em geral não apresenta quilha (Fig. 4 A-E). Outra diferença é a presença de um curto entalhe si-fonal na margem inferior da abertura da con-cha de Aylacostoma, aspecto não observado em M. tuberculata.
Figura 3. Variação morfológica em M. tuberculata (Müller, 1774) da Ilha Grande, Rio de Janeiro, mostrando as variações na ornamentação colorida da concha. A, ausência de flâmulas acastanhadas e banda columelar; B, presença de flâmulas e ausência de banda columelar; C, presença de banda colume-lar e flâmulas. Escala = 1 cm (Fotos: I.C. Miyahira).
Figura 4. Morfologia de conchas de Melanoides tuberculata (Müller, 1774) (A, B, C, D, E) em comparação com concha de Aylacostoma sp. (F). A, Vila do Abraão, Ilha Grande, RJ; B, Praia Vermelha, Ilha Grande, RJ; C, Vala da Fiocruz, Rio de Janeiro, RJ; D, Maricá, Rio de Janeiro, RJ; E, Jijoca de Jericoacoara, CE; F, Mocambinho, MG. Todas as escalas = 1 cm.

CAPÍTULO 9
Seção 2
129
Em Doryssa encontramos desde conchas lisas até fortemente esculturadas, com perda do ápice em algumas espécies. Em geral inexiste entalhe sifonal, assim como em M. tuberculata; a volta corporal é abaulada em algumas espé-cies. Ou seja, existe uma superposição de ca-racterísticas morfológicas que dificultam uma caracterização exata, as quais exigem uma re-visão das espécies atribuídas à Aylacostoma e Doryssa. Melanoides tuberculata possui pé e ca-beça com coloração escura (Simone, 2001) ou com manchas amareladas (Brandt, 1974). A pro-bóscide é ampla, achatada dorso-ventralmente, com a margem anterior bilobada. Os tentácu-los possuem comprimento semelhante ao da probóscide. Os olhos são escuros, próximos à base dos tentáculos e sem omatóforos (Simone, 2001). O bordo do manto é franjado (Benthem--Jutting, 1956; Brandt, 1974, Simone, 2001).
2.2. Morfologia interna
Uma descrição detalhada da morfologia interna de fêmeas de M. tuberculata foi apre-sentada por Simone (2001). A disposição dos principais órgãos observados externamente pode ser vista na Figura 5.
Manto
Apresenta coloração amarelo-creme pá-lido, deixando perceber por transparência os órgãos da massa visceral. O bordo do manto é esbranquiçado e espessado, com lobos franja-dos, em geral de forma regular, mas podem ser irregulares, como figurado por Simone (2001).
Cavidade palial
A abertura da cavidade palial é ampla. No teto da cavidade palial (Fig. 6) ob-servamos o osfrádio, a brânquia e o oviduto. O osfrádio é alongado, semelhante a um sulco, com aproximadamente metade a um terço do comprimento da brânquia, situado posterior-mente ao início da mesma.
Sistema digestório
O sistema digestório é completo, apresen-tando massa bucal, esôfago, estômago, glândula
digestiva, intestino e ânus. O esôfago não apre-senta glândulas acessórias. O reto é largo e am-plo. O ânus é sifonado (Simone, 2001). A rádula (Fig. 7) é curta e possui um formato semelhante ao de um “S”. Os dentes raquidianos apresentam sete cúspides cada; os dentes laterais com cinco cúspides cada, sen-do a segunda mais larga; os dentes marginais possuem cinco cúspides cada (Simone, 2001). A rádula é do tipo tenioglossa, cuja fórmula radular é 2.1.1.1.2 (Benthem-Jutting, 1956). Brandt (1974) fornece a fórmula radular com a indicação do número de dentículos em cada dente: C 4-1-4, L 1-1-4, M1 9, M2 10.
Sistema circulatório
O sistema circulatório é similar ao de outros tiarídeos. Apresenta um coração gran-de, situado posteriormente na cavidade palial, depois das brânquias. A aurícula é maior que o ventrículo (Simone, 2001, Fig. 8).
Figura 5. Fêmea de Melanoides tuberculata (Müller, 1774), re-tirada da concha: bm, bordo do manto franjado; g, gônada imersa na glândula digestiva; i, intestino; m, marsúpio; mv, massa visceral. Escala= 1 cm. (Foto: R.F. Ximenes).
Figura 6. Desenho esquemático do teto da cavidade palial de Melanoides tuberculata (Müller, 1774). (Desenho: R.F. Ximenes).

Fernandez et al.
Gastrópodes límnicos invasores: morfologia comparada
130
Sistema excretor
O sistema excretor apresenta rim gran-de, de cor creme, medindo aproximadamente de um terço à metade do comprimento da vol-ta corporal, situado no limite posterior-direito da cavidade palial. Possui dois lobos, ambos relativamente ocos, com uma série de septos transversais. O nefróstoma está localizado na extremidade posterior direita da membrana entre o rim e a cavidade palial, próximo ao reto (Simone, 2001, Fig. 8). Difere das espécies nativas de Thiaridae já conhecidas, do gênero Aylacostoma, que também apresentam um rim bilobado, mas um dos lobos é sólido, glandular (Simone, 2001).
Sistema reprodutivo
Melanoides tuberculata é uma espécie dioica, como a maioria dos prosobrânquios, apesar da partenogênese ser o meio de repro-dução mais comum. As fêmeas possuem um sistema reprodutivo composto por ovário, ovi-duto palial e marsúpio (Berry & Kadri, 1974). O ovário apresenta uma coloração amarelada (Fig. 9A) e se encontra parcialmente inserido na glândula digestiva (Berry & Kadri, 1974; Heller & Farstay, 1989). O oviduto palial é to-talmente fechado, sendo um tubo estreito com paredes espessas, com abertura próxima ao ânus, sem glândulas acessórias (Berry & Ka-dri, 1974; Simone, 2001). O marsúpio é uma
Figura 7. Rádula de Melanoides tuberculata, exemplar oriundo da Vila do Abraão, Ilha Grande, Angra dos Reis, Rio de Janeiro. A, Panorama geral da rádula, escala = 100 µm. B, Detalhe do dente raquidiano (cen-tral), escala = 10 µm. C, Detalhe do dente lateral, escala = 10 µm. D, Detalhe dos dentes marginais. Escala= 20 µm. (Fotos: A.C.N. Moraes, Laboratório de Microsco-pia Eletrônica UERJ).
Figura 8. Detalhe do complexo renopericárdico: à esquerda, Melanoides tuberculata (Müller, 1774); à direita, Aylacostoma exoplicata Simone, 2001. (aa), aorta anterior; (au), aurícula; (cp), cavidade palial; (in), intestino; (mr), membrana renal; (nj), ne-fróstoma; (ov), oviduto; (pe), pericárdio; (ra), lobo renal anterior; (rp), lobo renal posterior; (se), saco do estilo; (ve), ventrículo. Modificado de Simone (2001, figs. 176 e 120, respectivamente).

CAPÍTULO 9
Seção 2
131
câmara espaçosa, localizado na região antero-dorsal do corpo, dorsal ao esôfago, abrindo-se para o exterior através de um pequeno poro do lado direito, acima da massa pediosa, próximo ao músculo columelar (Simone, 2001). Exter-namente, o marsúpio forma uma protuberância atrás dos tentáculos e da massa bucal. O mar-súpio internamente é dividido por partições de tecido muscular em compartimentos irregu-lares. Normalmente encontra-se no marsúpio de M. tuberculata juvenis em todas as fases de desenvolvimento, de ovos até juvenis de cinco a seis voltas (Berry & Kadri, 1974; Dudgeon, 1986; Simone, 2001; Ben-Ami & Hodgson, 2005). Os machos são raros e apresentam tes-tículo grande e vermelho-acastanhado no ani-mal vivo (Fig. 9B), com tamanho aproximado ao da glândula digestiva (Heller & Farstay, 1989). O testículo é lobulado e se estende ao longo das voltas corporais, embutido na glân-dula digestiva. Os lobos testiculares confluem em um ducto que conduz à próstata. A partir da próstata se estende um longo vaso deferente (Livshits & Fishelson, 1983).
3. Helisoma duryi (Wetherby, 1879) e Helisoma trivolvis (Say, 1817)
3.1. Morfologia comparada de Helisoma spp.
Helisoma duryi (Fig. 10A) e Helisoma trivolvis (Fig. 10B) podem ser confundidos com ou-tros planorbídeos nativos, como por exem-plo, Biomphalaria tenagophila (d’Orbigny, 1835), Biomphalaria occidentalis Paraense, 1981 e Biomphalaria tenagophila guaibensis Paraense, 1984, os quais geralmente apresen-tam conchas planispirais altas. A concha das espécies mencionadas e de Helisoma spp. são apresentadas na Figura 11. A diferenciação dessas espécies é importante tanto sob o pon-to de vista biológico, quanto parasitológico, uma vez que B. tenagophila é uma das três es-pécies transmissoras naturais de Schistosoma mansoni Sambon, 1907, trematódeo que causa a esquistossomose, uma das principais ende-mias do país, pois ainda acomete milhares de pessoas.
Figura 9. Melanoides tuberculata (Müller, 1774). Comparação entre a coloração das gônadas de fêmeas (A) e machos (B). Fotos: R.F. Ximenes.
Figura 10. Exemplares vivos de Helisoma. A, H. duryi (Wetherby, 1879); B, H. trivolvis (Say, 1817). Escala: 5 mm. Fotos: P.M. Coelho.
As espécies de Biomphalaria (Fig. 11 C-E) possuem concha mais frágil e espessura da borda da concha mais delicada, quando com-parada às de Helisoma spp., embora a observa-ção do complexo genital seja necessária para a diferenciação específica. A relação tamanho da

Fernandez et al.
Gastrópodes límnicos invasores: morfologia comparada
132
Tabela I. Diferenças entre as conchas de Helisoma trivolvis (Wetherby, 1879); e Helisoma duryi (Say, 1817).
Helisoma trivolvis Helisoma duryi
Diâmetro da concha Até cerca de 25 mm
Até cerca de 18 mm
Largura na abertura 9 mm 8,5 mm
Número de giros 5,5 5
Fonte: Paraense, 1975; Paraense, 1976a, b.
concha e crescimento dos giros pode também ser um indicativo, uma vez que em Helisoma os giros crescem rapidamente em diâmetro. Seme-lhanças entre Biomphalaria sp. e Helisoma sp. encontram-se também na morfologia interna, como por exemplo, no tubo renal (em forma de J), mandíbula (com uma peça transversal su-perior mais larga e duas peças laterais inferio-res verticais muito mais estreitas), rádula (em fita aproximadamente retangular), hemolinfa (vermelha, devido à presença de hemoglobina), pseudobrânquia (foliácea, aproximadamente triangular, com a face lateral percorrida longi-tudinalmente pela extremidade da crista retal) e posição de alguns órgãos (glândulas salivares passando por fora do anel ganglionar perieso-fagiano e intestino anterior cruzando a super-fície ventral da glândula do albúmen), como descrito por Paraense (1975).
3.2. Morfologia externa
Há algumas décadas, pelo menos duas espécies, H. duryi e H. trivolvis, originárias dos Estados Unidos da América, têm sido registra-das em algumas ilhas do Caribe e em países da América do Sul, inclusive no Brasil. As conchas destas duas espécies são muito semelhantes, o que torna muito difícil a identificação apenas pela conquiliologia (Tab. I). Ambas as espécies apresentam concha pla-
Figura 11. Conchas de espécies nativas e exóticas de Planorbidae. Para cada espécie é apresentada, respectivamente, o lado direito, a visão frontal e o lado esquerdo. A, Helisoma duryi (Wetherby, 1879); B, Helisoma trivolvis (Say, 1817); C, Biomphalaria tenagophila (d’Orbigny, 1835); D, Biomphalaria occidentalis Paraense, 1981; E; Biomphalaria tenagophila guaibensis Paraense, 1984; Escala: 5 mm. (Fotos: P.M. Coelho).

CAPÍTULO 9
Seção 2
133
nispiral, que aumentam rapidamente em diâme-tro, carenados (com vinco) no lado esquerdo e sub-carenados no lado direito, separados por suturas profundas; abertura da concha dilatada, quando comparada às espécies do gênero Biomphalaria. Em Helisoma, geralmente as estrias de crescimen-to são muito visíveis nos giros, uma característica do gênero, de acordo com Paraense (1975). Tanto B. occidentalis quanto B. tenago-phila apresentam concha planispiral com apro-ximadamente 7 giros ou voltas que crescem lentamente em diâmetro, carenados, mais acen-tuadamente no lado esquerdo em B. tenagophila e em ambos os lados em B. occidentalis; sutu-ras profundas e abertura variando de deltoide a cordiforme (Fig. 12). Ao descrever B. tenago-phila guaibensis, Paraense (1984) comentou que a concha dos espécimes do Guaíba, RS, não se distingue das de B. tenagophila. Estas três espé-cies apresentam a concha mais alta quando com-parada às demais espécies de Biomphalaria Sul--americanas, de acordo com Paraense (1981b). Durante muito tempo B. occidentalis, com ocorrência principalmente no Brasil Cen-tral, foi identificada como B. tenagophila. O estudo da morfologia interna, entretanto, re-
velou diferenças, entre elas, a ausência de bolsa vaginal em B. occidentalis e a forma e tamanho do prepúcio, maior nessa espécie (Fig. 13). É interessante ressaltar que apesar da grande se-melhança, B. occidentalis não é transmissora da esquistossomose.
3.3. Morfologia interna
Os planorbídeos são hermafroditas, ca-pazes de realizar a autofecundação, quando ne-cessário. Em linhas gerais o sistema reprodutor é composto por ovoteste (onde são produzidos óvulos e espermatozoides), vesícula seminal, ovispermiduto, oviduto, vagina, espermiduto, próstata, espermateca, canal deferente, bainha do pênis, prepúcio e pênis. O complexo penia-no de Helisoma (Fig. 13) difere na forma e ta-manho dos órgãos, quando comparado com as espécies de Biomphalaria (Fig. 14). As principais características que permi-tem a rápida distinção entre H. duryi e H. tri-volvis estão no comprimento do canal da esper-mateca, bem mais longo em H. trivolvis, e no prepúcio e bainha do pênis, em forma de pera em H. duryi (Fig. 13, Tab. II).
Figura 12. Desenhos das conchas de espécies nativas e exóticas de Planorbidae. Para cada espécie estão ilustrados, respecti-vamente, o lado esquerdo, a visão frontal e o lado direito. A, Helisoma duryi (Wetherby, 1879); B, H. trivolvis (Say, 1817); C, B. tenagophila (d’Orbigny, 1835); B. occidentalis Paraense, 1981; E, B. tenagophila guaibensis Paraense, 1984. Extraídos de Para-ense (1975, 1981b, 1984, 2003).

Fernandez et al.
Gastrópodes límnicos invasores: morfologia comparada
134Figura 14. Sistema reprodutor de espécies de Biomphalaria. A, Biomphalaria tenagophila (d’Orbigny, 1835); B, Biomphalaria occidentalis Paraense, 1981; C, Biomphalaria tenagophila guaibensis Paraense, 1984; bo, bolsa do oviduto; bp, bainha do pênis; bv, bolsa vaginal; cc, canal coletor do ovoteste; cd, canal deferente; cop, canal do órgão prepucial; des, duto da espermateca; ed, espermiduto; eg, encruzilhada genital; es, espermateca; gn, glândula nidamental; mr, músculo retrator do complexo pe-niano; mp, músculo protrator do complexo peniano; odd, ovispermiduto distal; odp, ovispermiduto proximal; ot, ovoteste; ov, oviduto; pp, prepúcio; pr, próstata; ut, útero; va, vagina; vs, vesícula seminal. Extraídos de Paraense (1975, 1981b, 1984, 2003).
Tabela II. Diferenças na morfologia interna de Helisoma trivolvis (Say, 1817) e Helisoma duryi (Wetherby, 1879).
Helisoma trivolvis Helisoma duryi
Próstata Com cerca de 6 a 14 divertículos arborescentes Com cerca de 4 a 8 divertículos longos, quase todos arborescentes
Bainha do pênis Aproximadamente cilíndrica Piriforme
Prepúcio Aproximadamente em forma de ânfora Com parede projetada lateralmente pelo órgão prepu-cial
Espermateca Muito longa, canal geralmente mais de 1,5 ve-zes mais longo que o corpo e notavelmente ex-pandido na base
Ovoide ou claviforme, canal um pouco mais longo que o corpo e alargando-se na direção da base.
Fonte: Paraense, 1975; Paraense, 1976a, b.
Figura 13. Sistema reprodutor. A, Helisoma duryi (Wetherby, 1879); B, Helisoma trivolvis (Say, 1817). bp, bainha do pênis; bo, bolsa do oviduto; bv, bolsa vaginal; cc, canal coletor do ovoteste; cd, canal deferente; cop, canal do órgão prepucial; des, duto da espermateca; ed, espermiduto; eg, encruzilhada genital; es, espermateca; gn, glândula nidamental; mr, músculo retrator do complexo peniano; mp, músculo protrator do complexo peniano; odp, ovispermiduto proximal; odd, ovispermiduto distal; ot, ovoteste; ov, oviduto; pp, prepúcio; pr, próstata; ut, útero; va, vagina; vs, vesícula seminal. Extraídos de Paraense (1975, 1981b, 1984, 2003).

CAPÍTULO 9
Seção 2
135
4. Morfologia comparada de Physa acuta Draparnaud, 1805, Physa marmorata Guilding, 1828 e Lymnaea columella Say, 1817
4.1. Morfologia externa Physa acuta Draparnaud, 1805 e Physa marmorata Guilding, 1828 estão entre as es-pécies incluídas na família Physidae no Brasil, enquanto Lymnaea columella Say, 1817 é a espécie mais amplamente distribuída de Lym-naeidae. De acordo com Simone (2006) ou-tras duas espécies ocorrem no país, Lymnaea rupestris Paraense, 1982 e Lymnaea viatrix d’Orbigny, 1835. A diferenciação entre fisídeos e limneídeos é importante, uma vez que L. columella é a princi-pal espécie vetora da Fasciola hepatica Linnaeus, 1758 no Brasil. Os limneídeos têm a concha cô-nica, com abertura dextrógira (voltada para di-reita), ao contrário dos fisídeos que apresentam concha sinistrógira (voltada para esquerda) (Figs. 15, 16). Os olhos estão situados na base interna dos tentáculos, os quais são achatados e triangu-lares nos limneídeos e alongados e filiformes nos fisídeos. Outra diferença é a forma do pé, cordi-forme em L. columella e alongado em Physa spp. (Fig. 17). Ambas as famílias estão amplamente distribuídas nas regiões tropicais do Continente Sul-americano, habitando águas lênticas, porém os fisídeos são mais tolerantes aos ambientes po-luídos (Paraense, 1981b).
4.2. Morfologia interna de Physa acuta Draparnaud, 1805 e Physa marmorata Guilding, 1828 As principais diferenças entre P. acuta e P. marmorata encontram-se no sistema re-produtor (Fig. 18). De acordo com Paraense (1986b, 1987) o ovoteste, ovispermiduto, ve-sícula seminal, oviduto, glândula nidamental, útero, vagina, espermoduto, próstata e vaso deferente de ambas as espécies são iguais. A principal diferença encontra-se na presença de uma glândula prepudial sobre o prepúcio em P. acuta, a qual ocupa cerca de um terço do com-primento deste órgão, não encontrada em P. marmorata. Além desta, outras diferenças po-dem ser observadas nestas espécies (Tab. III).
Figura 15. Conchas de espécies de Physa e Lymnaea. A, Physa marmorata Guilding, 1828; B, Physa acuta Draparnaud, 1805; C, Lymnaea columella Say, 1817. Escala 5 mm. (Fotos: P.M. Coelho).
Figura 16. Desenhos das conchas: A, Physa acuta Draparnaud, 1805; B, Physa marmorata Guilding, 1828; C, Lymnaea columella Say, 1817. Extraídos de Paraense (1986a, b) e Paraense & Pointier (2003).
Figura 17: Diferença entre os tentáculos e forma do pé: A, ten-táculos triangulares e pé cordiforme em Lymnaea columella Say, 1817; B, tentáculos filifomes e pé alongado em Physa acuta Draparnaud, 1805. Escala 5 mm. (Fotos: P.M. Coelho).

Fernandez et al.
Gastrópodes límnicos invasores: morfologia comparada
136 Agradecimentos Ao Dr. Luiz Ricardo Simone, do Museu de Zoologia da Universidade de São Paulo pela ces-são de algumas fotos e figuras que ilustram esse
Tabela III. Diferenças anatômicas entre Physa acuta Draparnaud, 1805 e Physa marmorata Guilding, 1828.
Physa acuta Physa marmorata
Espermateca Oval ou piriforme Oblongo, com uma moderada cons-trição no meio e um tanto curvo
Canal da espermateca Duto uniformemente estreito, com a base ex-pandida, um pouco mais longa que o corpo
Duto uniformemente estreito, um pouco mais longo que o corpo
Bainha do pênis Mesma largura do pênis e termina em um bulbo um tanto mais estreito que a porção proximal
Bainha do pênis apresenta uma ex-pansão terminal bulbosa cuja extre-midade se insere na porção caudal do prepúcio.
Prepúcio Muito mais largo que o bulbo da bainha do pênis, com ombro pouco saliente
Com ombro bem definido, muito mais largo que a porção estreita da bainha do pênis
Canal peniano Com abertura subterminal Com abertura lateral, mais ou menos na junção dos terços médio e inferior
Proporção bainha do pênis/prepúcio Cerca de 1,49 (1,25 – 1,83 ). Cerca de 2.08 (1,45 - 2,75).
Fonte: Paraense, 1986b; Paraense, 1987.
Figura 18. Desenhos esquemáticos do sistema reprodutor: A, Physa marmorata Guilding, 1828; B, Physa acuta Draparnaud, 1805. (bbp), bulbo da bainha do pênis; (bp), bainha do pênis; (bv), bolsa vaginal; (cc), canal coletor do ovoteste; (cd), canal deferente; (des), duto da espermateca; (dga), duto da glândula de albúmen; (ed), espermiduto; (es), espermateca; (gn), glândula nidamental; (gp), glândula prepucial; (mr/mrc), músculo retrator do complexo peniano; (mp), músculo protrator do complexo peniano; (mpc), músculo protrator do complexo peniano; (odd); ovispermiduto distal; (odp), ovispermiduto proximal; (ot), ovo-teste; (ov), oviduto; (pp), prepúcio; (pp), próstata; (ut), útero; (va), vagina; (vs), vesícula seminal. Extraídos de Paraense (1975, 1981b, 1984, 2003).
capitulo. Ao Eduardo Prado e ao Pablo Menezes Coelho, do Laboratório de Referência em Mala-cologia, Fiocruz, RJ, pelo trabalho iconográfico. Ao Laboratório de Microscopia Eletrônica da UERJ pela imagem de varredura da rádula.

Seção
3Monitoramento de moluscos invasores


139
Os ecossistemas são formados por dife-rentes organismos. Em ambientes aquáticos en-contramos o plâncton, quaisquer organismos encontrados na coluna d’água que são desloca-dos de acordo com as massas de água (Esteves, 1988). O plâncton pode ser classificado de di-versas formas: em função de suas dimensões, do biótopo, distribuição vertical, duração da vida planctônica e de sua nutrição (Omori & Ikeda, 1984). O plâncton de forma geral se di-vide em fitoplâncton e zooplâncton. O fitoplâncton constitui a porção ve-getal do plâncton, aquela capaz de sintetizar matéria orgânica através da fotossíntese. Este é composto basicamente por algas, desempe-nhando um papel importante por estar na base da cadeia alimentar. Já o zooplâncton é cons-tituído por organismos heterotróficos (seres incapazes de produzir seu próprio alimento e se nutrem de substâncias orgânicas produzidas por outros seres), pertencentes ao reino animal, como pequenos invertebrados e formas larvais de alguns organismos. Muitos organismos do
zooplâncton realizam migrações diárias, deter-minadas pela dinâmica do ambiente. A comunidade planctônica apresenta um caráter muito dinâmico, com elevadas taxas de reprodução e perda, respondendo rapidamente às alterações físicas e químicas do meio aquá-tico e estabelecendo complexas relações intra e interespecíficas na competição e utilização do espaço e dos recursos (Valiela, 1995). Variações no regime meteorológico, ca-racterísticas geomorfológicas regionais e os im-pactos antropogênicos nas áreas costeiras, es-tabelecem, em conjunto, o regime hidrográfico particular de cada região e, consequentemente, as características taxonômicas e a dinâmica espaço-temporal de suas comunidades planctô-nicas (Brandini et al., 1997). As espécies de bivalves invasores en-contrados no ambiente límnico brasileiro são principalmente Limnoperna fortunei (Dunker, 1857), Corbicula fluminea (Müller, 1774) e Corbicula largillierti (Philippi, 1844). As espé-cies invasoras possuem uma grande capacidade
CAPÍTULO 10
Como monitorar bivalves invasores no plâncton?
Método da microscopia óptica
Cintia Pinheiro dos SantosMarinei Vilar Nehrke
Maria Cristina Dreher MansurVanessa Gazulha

Santos et al.
Como monitorar bivalves invasores no plâncton? Método da microscopia óptica
140
reprodutiva, geralmente causando problemas ambientais e econômicos (ver Capítulo 2). De-vido a estes motivos, estas espécies invasoras devem ser monitoradas constantemente consi-derando que as mesmas apresentam fase larval total ou parcialmente planctônica (Detalhes de diferenciação larval das espécies invasoras, ver Capítulo 6). O êxito de qualquer monitoramento so-bre o plâncton depende de um desenho amos-tral adequado de acordo com o tipo de infor-mação que se deseja obter. Fatores importantes a serem considerados são as variações espaciais horizontais e verticais, as variações sazonais, o tamanho dos organismos, e se o estudo é qua-litativo ou quantitativo. Estes fatores, entre ou-tros, irão determinar os métodos de amostra-gem e análise mais adequados. Os métodos para a coleta de plâncton po-dem agrupar-se em duas grandes categorias, as garrafas e as redes (González de Infante, 1988). As garrafas, como por exemplo, as garrafas de Van Dorn, são utilizadas para coletar um certo volume de água que deve ser concentrado por filtração ou sedimentação para reunir o plânc-ton em um volume pequeno de água. Tem como vantagem a captura das formas pequenas do plâncton, porém, seu volume reduzido (de 2 a 6L) é uma desvantagem quando a população é pequena. As redes são os instrumentos de coleta de plâncton mais antigos e de uso mais generali-zado. As redes podem apresentar diferentes mo-delos. O tecido filtrante das redes de plâncton é de nylon e pode apresentar diferentes aberturas de malha (μm). As redes com aberturas de malha mais reduzidas têm maior tendência a acumular os organismos e partículas suspensas, dificul-tando a filtração. Já as redes com maior aber-tura de malha tendem a perder os organismos forçadamente permanecendo em rede só os de maior tamanho. Na base da rede deve haver um copo coletor, as laterais do copo deverão ser for-radas com a mesma tela, assim, os organismos coletados permanecerão neste copo até o térmi-no da filtração. As coletas podem ser feitas por arrastos, em que a rede é passada várias vezes no ambiente; ou a água também pode ser recolhida através de um balde; ou através de uma bomba e passada pela rede em quantidades conhecidas. A vantagem das redes é o grande volume de água que pode ser coletado.
No caso das larvas planctônicas de bival-ves invasores, o método mais adequado para a coleta é a rede de plâncton, devido especial-mente à baixa densidade destes organismos no ambiente aquático. É importante obter amos-tras com réplicas (no mínimo, em triplicata) e coletar em diferentes locais do corpo d’água, especialmente, em lagos e reservatórios gran-des, onde possa haver grande heterogeneidade espacial. Após a coleta é necessária a preservação do material amostrado, a qual pode ser realiza-da por método físico via congelamento e poste-rior liofilização, e químico pelo uso de formol e etanol. A utilização do formol requer mais tem-po para provocar a morte dos organismos, as-sim eles podem se contrair e expelir o conteúdo dos intestinos. Portanto, recomenda-se a adi-ção de anestésico antes da fixação. Além disso, o pH ácido da solução, com o tempo, pode cau-sar danos aos animais (Haney & Hall, 1973). A imersão em etanol é uma técnica simples e eficiente, onde o material coletado é imedia-tamente imerso em etanol 95%. Esse fixador causa morte imediata e evita alguns problemas, como distorção de corpos (Hall, 1964). Para a obtenção de larvas das espécies invasoras citadas anteriormente, é necessário efetuar coletas no plâncton através de rede de nylon, comumente utilizado em serigrafia, com abertura de malha em torno de 30 μm (micrômetros) ou 150 fios. No laboratório, o conteúdo do copo coletor deve ser examinado sob estereomicroscópio para quantificação, uma vez que as larvas de bivalves possuem tamanho diminuto. Já para a identificação das fases larvais, é necessário um microscópio com grande resolução óptica (Coelho, 2004). (Ver Capítulo 6).
Metodologia para coleta e contagem de larvas
1.1. Coleta
A coleta de larvas é realizada através da filtração de 1 m3 de água (Fig. 1), com rede de plâncton com malha de 30 μm e com o auxílio de uma bomba de sucção (Fig. 1C), o tempo de filtração varia conforme a vazão

CAPÍTULO 10
Seção 3
141
do equipamento (Fig. 1A). Esta quantidade elevada de água (aprox. 1000L) que se costu-ma coletar para quantificação das larvas de bivalves está relacionada ao fato de que estas normalmente ocorrem em baixa densidade no plâncton. Ao término da filtração, as larvas concentradas são retiradas do tubo coletor, o volume aproximado é de 100 ml (Fig. 1D). A amostra é acondicionada em pote plástico (Fig. 1E), logo deve-se adicionar 100 ml de álcool 96% ao volume da amostra a fim de preservar a mesma, totalizando 250 ml (Fig. 1F).
1.2. Quantificação da densidade larval
A análise das amostras no laboratório envolve duas etapas: a preparação da amostra e a quantificação.
1.2.1. Preparação da amostra a ser analisada
* Troca-se o recipiente do campo por um Becker de 400 ml; * Espera-se 15 minutos para a amostra decantar;
* Com auxílio de uma pipeta, retira-se a água da superfície do Becker até chegar ao valor estipulado de 100 ml. Este será o volume a ser analisado.
1.2.2. Quantificação da amostra
Para a quantificação das amostras pre-paradas anteriormente em laboratório (Fig. 2), estas são homogeneizadas e retira-se uma alíquota de 2 ml e coloca-se sobre uma pla-ca de contagem quadriculada (Fig. 2A). Esta amostra (Fig. 2C) é quantificada com o auxí-lio do microscópio estereoscópico (Fig. 2B). Cada larva encontrada (Figs. 2D e 2E) nas alíquotas quantificadas é pipetada e retirada da placa de contagem, procedimento adotado para que não haja recontagem de larvas, evi-tando, assim, que a amostra seja superestima-da.
1.3. Estimativa da Densidade Larval
A densidade larval (ind.m-3) de cada es-pécie de bivalve (p. ex. L. fortunei, C. fluminea, C. largillierti) é obtida utilizando-se a seguinte fórmula (APHA, 2005):
Figura 1. Etapas da coleta. 1A: filtração do plâncton; 1B: detalhe da rede de plâncton; 1C: detalhe da bomba de sucção; 1D: retirada das larvas concentradas no tubo coletor; 1E: acondicionamento da amostra e 1F: preservação da amostra coletada em campo.

Santos et al.
Como monitorar bivalves invasores no plâncton? Método da microscopia óptica
142
onde, C= número de indivíduos contados V’= volume da amostra concentrada (mL) V’’= volume da alíquota (mL) V’’’= volume da amostra coletada (m3)
Por exemplo, se em campo foram coletados 1000L (=1 m³) de água e concentrados em frascos de 100 mL, e em laboratório, foram triadas alí-quotas referentes à 10 mL da amostra e quantifica-dos 200 indivíduos (larvas) no total, o cálculo da densidade larval (ind.m-3) será da seguinte forma:
Figura 2. Etapas em laboratório. 2A, placa contagem contendo 2 ml de amostra; 2B, microscópio estereoscópico utilizado para quantificação; 2C, visão geral da amostra sob microscópio estereoscópico; 2D e 2E, larvas fotografadas sob microscópio óptico encontradas nas amostras coletadas durante monitoramento; 2D, Limnoperna fortunei no estágio véliger (220 µm) e 2E, larva véliger de Corbicula fluminea (220 µm).
Desta forma, o resultado do cálculo indica que a densidade larval na amostra em questão é de 2000 ind/m³. É importante ressaltar que para uma avaliação eficiente da densidade larval recomenda--se a triagem de alíquotas referentes a, no mínimo, 10% do total da amostra. Quando o número de in-divíduos contados for inferior a 100, recomenda-se a triagem de mais alíquotas até a contagem mínima de 100 larvas, ou ainda, a triagem total da amostra.

143
A capacidade de detecção precoce de es-pécies aquáticas invasoras durante o processo de sua dispersão é essencial para a definição de medidas de controle ou mitigação. O mo-nitoramento de bivalves invasores pode se dar por vários métodos. O mais óbvio e simples é a observação das conchas dos indivíduos adul-tos no ambiente. Embora esta seja a maneira mais fácil, a detecção é tardia e prejudica ações mitigadoras de controle ou eliminação dos mo-luscos. A presença de indivíduos adultos quase sempre já é um indício de que as populações se encontram em adiantado processo de coloniza-ção do ambiente. Além disso, a observação de moluscos fixados em substratos em ambientes de águas mais profundas ou turvas, sejam na-turais ou artificiais, costuma ser pouco efetiva. No caso de espécies que apresentam fases larvais planctônicas, o monitoramento também pode ser realizado pela prospecção de larvas desses moluscos bivalves a partir da cole-ta de amostras do zooplâncton, seguida da ob-servação em microscópio estereoscópico. Essa
metodologia permite a detecção precoce dessas espécies antes do estabelecimento de grandes e extensas populações, porém, apresenta desvan-tagens, pois exige experiência e conhecimento taxonômico das larvas da espécie-alvo. Além disso, o processamento microscópico é lento e realizado geralmente a partir de subamostra-gens, o que pode resultar em falsos negativos. Adicionalmente, a identificação de larvas de moluscos bivalves é geralmente possível ape-nas através das fases larvais mais tardias ou de microscopia óptica ou eletrônica de varredura (Kimura & Sekiguchi, 1994; Ozawa & Seki-guchi, 2002). Na maioria das vezes, a prospec-ção sob lupa de larvas de espécies de bivalves exige o conhecimento prévio sobre as espécies de moluscos coexistentes na área de interesse e da morfologia geral de cada uma das suas fa-ses larvais. Muitas larvas de bivalves possuem extrema semelhança morfológica induzindo o identificador a erros, como frequentemen-te ocorre com as espécies Corbicula fluminea (Müller, 1774) e Limnoperna fortunei (Dunker,
CAPÍTULO 11
O método molecular de prospecção do
mexilhão-dourado
Marcel K. TscháRaquel Patella
Antonio OstrenskyWalter A. Boeger

Tschá et al.
O método molecular de prospecção do mexilhão-dourado
144
1857) em amostras de zooplâncton de ambien-tes aquáticos continentais sul-americanos. O desenvolvimento de técnicas para de-tecção molecular de organismos planctônicos, todavia, tornou possível o monitoramento de larvas de espécies de bivalves (Claxton & Bou-ling, 1998; Toro, 1998). Com o uso desse mé-todo, larvas podem ser detectadas com grande sensibilidade e especificidade, em um curto espaço de tempo, sem necessariamente exigir conhecimentos específicos em biologia larval do técnico responsável. O método proposto por Pie et al. (2006), utiliza marcadores moleculares para detectar o DNA de L. fortunei em amostras de plâncton. Esse protocolo tornou-se uma ferramenta indis-pensável, de agilidade e eficiência superior quan-do comparado aos métodos tradicionais para a prospecção precoce do mexilhão-dourado. O protocolo de prospecção molecular já foi extensamente testado, tendo apresentado resultados bastante satisfatórios quanto à sua especificidade (Boeger et al., 2007). A sensibili-dade do método é, também, um de seus pontos mais fortes. O limiar de detecção é cerca de 600 vezes maior do que a quantidade de DNA extraída de uma única larva (Pie et al., 2006). Assim, em teoria, o método é capaz de indicar a presença de apenas uma larva em aproxima-damente 200 L de água filtrada com redes de zooplâncton. Na prática, o protocolo tem se mostrado eficiente na detecção de larva em bai-xa concentração no ambiente, inclusive quan-do larvas não são detectadas através de análises microscópicas e quando adultos não são detec-tados em amostras ambientais (Boeger et al., 2007; Darrigran et al., 2009). O protocolo molecular é descrito nesse capítulo de forma objetiva, apresentando deta-lhes de procedimentos e soluções para proble-mas de processamento. Alternativamente, esse protocolo pode ser utilizado para realizar a identificação de animais jovens ou adultos, sem-pre que houver dúvidas sobre sua identidade.
Coleta e fixação de material biológico As amostras de zooplâncton devem ser coletadas dos locais onde se objetiva prospec-tar as fases larvais do mexilhão-dourado, como
por exemplo, tanques de estações de tratamento de água, reservatório de usinas hidrelétricas ou no próprio curso natural de um rio. Esta cole-ta é feita com a utilização de redes de arrasto de plâncton, com abertura de malha de aproxima-damente 53 μm, em arrastos de cerca de 5 min, preferencialmente em arrasto diagonal (do fundo para a superfície). Devido à esperada natureza em manchas na distribuição do plâncton, múltiplas amostras devem ser obtidas de cada ambiente. Quando se capta água diretamente do meio (ex. sistema de resfriamento de usinas hidroelétricas), recomenda-se a filtração de cerca de 4.000 L de água. Em campo, o material coletado por este arrasto deve ser rapidamente concentrado por se-dimentação (alguns minutos são suficientes para que os organismos da amostra se concentrem no fundo do recipiente) e remoção do excesso de água. Em seguida, etanol 92-95% deve ser adi-cionado em uma proporção aproximada de 1:2 (volume de amostra : volume de etanol). Cada amostra deve então ser devidamente etiquetada, com anotações do modo de arrasto (ou bombea-mento e filtragem), local, data, horário e respon-sável pela coleta (dentre outras, conforme inte-resse específico). A adequada etiquetagem dos recipientes é um procedimento imprescindível. Informações adicionais quanto ao estado físico do zooplâncton ou aparente presença de ciano-bactérias são de grande importância para se de-terminar procedimentos subsequentes da análise. Sugere-se também que, a partir do mate-rial coletado sejam geradas subamostras antes do início das atividades em laboratório, propiciando assim, contraprovas das análises (recomendações adicionais são encontradas no item “Sugestões e resolução de problemas” no final do capítulo). A amostra coletada deve ser então enca-minhada ao laboratório, onde será pré-proces-sada com auxílio de uma bomba a vácuo (Fig. 2). O pré-processamento visa separar as par-tículas do plâncton da água, possibilitando a extração mais eficiente do DNA. Esse proce-dimento tem se mostrado bastante eficiente na prática, embora não seja imprescindível. Amos-tras de animais adultos podem ser obtidas pre-servando um pedaço de tecido (ex. músculo, manto, gônada) em etanol 92% ou em outro líquido utilizado nos procedimentos usuais de laboratórios de genética (ex. DMSO, tampões).

CAPÍTULO 11
Seção 3
145
Figura 1. Síntese da metodologia molecular utilizada na pros-pecção das larvas de L. fortunei.
Procedimentos laboratoriais
Uma síntese do processamento das amos-tras é apresentada na Figura 1. A amostra de zoo-plâncton coletada é pré-processada através de um conjunto de peneiras em série, que removem parte de organismos indesejáveis na análise molecular (Quadro I e Fig. 2). Neste conjunto são utilizadas duas peneiras com diâmetro de abertura de malha de 300 μm e 100 μm, considerando os tamanhos registrados para as larvas de L. fortunei (Kimura e Sekiguchi, 1994). Dessa maneira as formas de zo-oplâncton menores que 100 μm, incluindo os está-μm, incluindo os está-m, incluindo os está-gios larvais iniciais de mexilhão-dourado, passam pelas peneiras e são retidas em um filtro de papel instalado abaixo no sistema. Tanto o material re-tido no filtro de papel, como nas demais malhas do sistema de filtragem, é raspado e o DNA total da amostra é extraído e purificado utilizando-se procedimentos usuais de biologia molecular.
Figura 2. Sistema de filtragem acoplado à bomba a vácuo para pré-processamento do zooplâncton. Esquema explodi-do à direita
– funil simples;– tubo PVC 2” de 15 cm sem rosca;– malha de abertura de 300 μm;– união roscável 2”;– tubo PVC 2” de 8 cm sem rosca;– malha de abertura de 100 μm;– união roscável 2”;– tubo PVC 2” de 8 cm com rosca;– papel filtro;– união roscável 2”;– tubo PVC 2” de 15 cm com rosca;– bucha de redução 2”;– adaptador autoajustável com rosca, anel para caixa d’água e junta de vedação;– frasco de 2 L em acrílico com tampa de pressão;– adaptador autoajustável com rosca e junta de vedação;– adaptador interno para mangueira.
Quadro I. A bomba a vácuo de pré-processamento do zooplâncton
O sistema descrito abaixo é recomendado para o pré-processamento das amostras de zooplâncton. Este sistema é composto por uma série de malhas com aberturas distintas e um filtro de papel, utilizado para retenção de material a ser processado a vácuo. O equipamento é montado com material facilmente encontrado no mercado, incluindo tubos de PVC, recipiente de acrílico e mangueiras. Um diagrama explodido do mecanismo é apresentado abaixo.
Todas as peças devem ser conectadas uma a outra, na mesma ordem descrita acima. As duas últimas peças devem ser fixadas ao frasco por um orifício artesanalmente aberto na lateral do recipiente, as demais peças devem ser presas na tampa do frasco.

Tschá et al.
O método molecular de prospecção do mexilhão-dourado
146
O procedimento descrito representa a aplicação do marcador molecular específico para prospecção do DNA do mexilhão-doura-do. O método é bastante simples e intuitivo e baseia-se na Reação em Cadeia da Polimerase do DNA (cuja sigla mais popular é PCR – Poly-merase Chain Reaction). Iniciadores da PCR específicos para L. fortunei foram desenvolvidos a partir de um fragmento de DNA mitocondrial, a subunida-de I do gene do Citrocromo Oxidase (COI). Na reação de PCR descrita a seguir, mesmo na presença de DNA de outras espécies, os ini-ciadores específicos são capazes de amplificar apenas o fragmento de COI de L. fortunei. Para evitar falsos negativos, devido a problemas de integridade do DNA total amostrado (decor-rente, por exemplo, da má fixação no campo) um par de iniciadores universais (fragmentos do gene nuclear 18S) é utilizado junto aos ini-ciadores/marcadores específicos. A reação do marcador é, portanto, conhecida como um PCR multiplex (ou duplex) que é o nome dado a reações que envolvam a amplificação de mais de um fragmento de DNA. Ambos marcadores moleculares utilizados estão listados na Tabela I com suas respectivas sequências. Veja proce-dimento no Quadro III:
O procedimento de extração utilizado no Laboratório de Ecologia Molecular é apresentado pas-so a passo no Quadro II.
Quadro II. Extração do DNA
1. Homogeneizar do filtrado com 98 μL de tampão de digestão (Tris 50 nM, pH 8.0) e 2 μL de proteinase K (20 ng/mL) a 50ºC durante 3 h;
2. Adicionar 200 μL de EZ-DNA® (Genomic DNA Isolation Kit – Biological Industries) e incubar a 50°C durante 30 min;
3. Centrifugar a 10.400 RPM por 10 min;
4. Passar o sobrenadante para novo tubo – este procedimento elimina os fragmentos insolúveis;
5. Neste novo tubo, adicionar etanol absoluto e centrifugar a 7.300 RPM por 5 min para precipitação do DNA;
6. Remover o etanol sobrenadante através de pipetagem – atenção para não remover o pellet que deve estar no fundo do tubo;
7. Repetir os procedimentos 5 e 6 com aplicação de etanol 96% para a lavagem do DNA;
8. Secar a amostra em temperatura ambiente por cerca de 10 min;
9. Dissolver o DNA em 50 μL de NaOH 8 mM.
A reação multiplex da PCR (Reação em Cadeia da Polimerase) é realizada em um volume total de 25 μL seguindo os seguintes passos:1. Adicionar água destilada ultrapura para completar o volume de 24 μL/tubo;2. Preparar uma solução estoque (em gelo) contendo volume suficiente para o número de amostras e controles (positivo e negativo) contendo: 1,5 mM de MgCl2, 0,5 mM de dNTPs, reação de buffer 1X, BSA 1:100, 0,625 unidades enzima Taq polimerase, 0,8 mM de cada indicador universal e 4 mM de cada indicador específico para L. fortunei.3. Agitar em vórtex.4. Distribuir 24 μL em cada tubo de amostra e controles;5. Adicionar 1 μL da amostra contendo o DNA extraído na diluição desejada.6. Programar o termociclador e proceder a PCR da seguinte maneira: a. desnaturação inicial a 94ºC por 4 min; b. 32 ciclos de cada um dos seguintes: b.1. desnaturação a 94ºC por 30 s. b.2. anelamento a 58,2ºC por 30 s. b.3. extensão a 70ºC por 1 min. c. Extensão final a 70ºC, desta vez por 4 min;7. Submeter o produto da PCR ao gel de agarose 2% e à eletroforese com voltagem aproximada de 80V;8. Corar o gel de agarose por 20 min em solução de brometo de etídio ou GelRed®;9. Visualizar sob luz ultravioleta.
Quadro III. Amplificação do DNA

CAPÍTULO 11
Seção 3
147
Tabela I. Sequências dos marcadores moleculares universais (indicadores) e específicos empregados na prospecção molecular do mexilhão-dourado.
Nome do primer Sequência (5’- 3’) Região do DNA Tipo do primer
COIR1 TCCAACCAGTCCCTACTCCACCCTCTA COI Específico
COIF1 TTTAGAGTTAGCACGTCCTGGTAGGTT COI Específico
7F GCCCTATCAACTTACGATGGTA 18S Universal
1100R GATCGTCTTCGAACCTCTG 18S Universal
Figura 3. Exemplo de gel de agarose visualizado em luz UV. L: Marcador de peso molecular. C+: controle positivo. 1 e 2: amos-tras positivas para L. fortunei. 3, 4, 5 e 6: amostras com integridade do DNA inadequada para análise. 7 e 8: amostras negativas para L. fortunei. C-: controle negativo.
Em todas as reações, um controle posi-tivo (DNA de L. fortunei adulto) deve ser inclu-ído como referência para a posição das bandas de cada fragmento e o correto funcionamento dos reagentes. Um controle negativo (reação sem amostra de DNA) é, também, indispensá-vel na reação, pois controla eventuais contami-nações nas reações. O padrão de bandas formado nos géis de agarose pelas amostras em análise é comparado com o padrão de bandas produzido pelo con-trole positivo (Fig. 3, coluna C+). A coincidên-cia na presença de banda específica (produzida pelos marcadores/iniciadores específicos) indica a presença de L. fortunei na amostra (Fig. 3, co-lunas 1 e 2). As amostras consideradas positivas para o bivalve devem apresentar também as ban-das universais (Fig. 3, colunas 1 e 2). Amostras
que apresentam apenas bandas resultantes dos iniciadores universais (controle da qualidade do DNA das amostras) devem ser consideradas negativas para o mexilhão-dourado. Devido aos eventuais problemas relacionados à concentra-ção do DNA de L. fortunei, todas as amostras devem ser submetidas às reações do marcador em pelo menos duas diluições (1:1 e 1:100), pre-ferencialmente três (1:1, 1:10, 1:100). Os resultados das reações devem ser confirmados periodicamente. Uma das formas de realizar essa avaliação do procedimento é sequenciar amostras positivas para bandas específicas escolhidas aleatoriamente, com subsequente comparação de sua composição nucleotídica com sequencias de L. fortunei dis-poníveis em bancos de dados via internet (ex. Genbank, BOLD).

Tschá et al.
O método molecular de prospecção do mexilhão-dourado
148
1. Quais são os procedimentos adequados para se evitar dano ao DNA ou mesmo perda de material durante a extração? 1.1. Manter a amostra de zooplâncton devidamente fixada e refrigerada – entre 2 e 8ºC – até a aplicação do protocolo de extração. 1.2. Fracionamento da amostra ou coleta de mais de uma amostra por ponto de coleta. Assim, caso ocorram problemas durante o procedimento de extração do DNA, as informações ge néticas não serão perdidas,
2. Nenhuma banda visível nas amostras e controle positivo com bandas? 2.1. DNA degradado. 2.2. Observar se o extrato recebido apresenta excesso de cianobactérias e/ou resíduos de solo, fatores que podem inibir as reações moleculares. 2.3. Verificar como o material foi armazenado durante a coleta e transporte. A má fixação e a extensa exposição da amostra do zooplâncton ao calor podem danificar o material coletado. A verificação da concentração e pureza de cada amostra extraída e um teste de amplificação com iniciadores universais de fragmentos menores (em torno de 200pb) confirmará a frag mentação do material genético da amostra. 2.4. Reagentes mal conservados ou mal pipetados durante a aplicação do protocolo molecular podem prejudicar a amplificação do DNA da amostra.
3. Apenas a banda do marcador específico aparece nas amostras? 3.1. Os iniciadores específicos podem estar consumindo os reagentes para a amplificação do fragmento universal. 3.2. Considerar resultado como positivo. 3.3. Verificar a qualidade da solução do conjunto de iniciadores universais.
Quadro IV. Sugestões e resolução de problemas

149
Introdução
O mexilhão-dourado Limnoperna fortunei (Dunker, 1857) é uma espécie bivalve de água--doce invasora identificada como contribuinte em vários problemas ecológicos e econômicos (Darrigran et al., 1998a, Ricciardi, 1998, Pen-chaszadeh et al., 2000, Magara et al., 2001, Boltovskoy et al., 2006, Pie et al., 2006). Em 2006, essa espécie foi identificada como uma espécie exótica invasora no Japão, devido a seus efeitos adversos nos ecossistemas. Durante o ciclo de vida do L. fortunei, a etapa planctô-nica é uma fase importante em que a distribui-ção da espécie é facilitada. A detecção e identi-ficação precisa da larva do mexilhão-dourado geralmente são realizadas com o auxílio de um microscópio óptico. Esse método consome muito tempo e requer experiência com taxo-nomia. Portanto, o desenvolvimento de um método mais direto seria benéfico para estudos sobre a ecologia e o gerenciamento dessa espé-cie. Pie et al. (2006) desenvolveram um método
sensível para a detecção de larvas do mexilhão--dourado que emprega uma abordagem base-ada no PCR (polymerase chain reaction, ou seja, reação em cadeia da polimerase) usando um conjunto específico de primers: LIMNO.COIR1 e LIMNO.COIF1. Assim, Boeger et al. (2007) demonstram que esse método permitiu a detecção de 1 a 5 larvas. No entanto, o mé-todo de detecção baseado em PCR não pode ser utilizado para quantificar com precisão as amostras de larvas. O método de detecção e quantificação de organismos planctônicos usando a reação em cadeia da polimerase quantitativa em tem-po real (qPCR) foi reconhecido como uma fer-ramenta eficiente, por Darling e Blum (2007). O qPCR permite detectar com precisão e quan-tificar um DNA específico em uma mistura de solução de DNA complexa, mesmo que a quan-tidade do DNA alvo (organismo) esteja em uma concentração muito baixa. Na última década, as aplicações de qPCR passaram a ser ferra-mentas largamente utilizadas na detecção e
CAPÍTULO 12
Método de detecção e quantificação de larvas do mexilhão-dourado Limnoperna fortunei,
usando PCR quantitativo em tempo real
Noriyuki EndoYasuyuki Nogata

Endo & Nogata
Método de detecção e quantificação de larvasdo mexilhão-dourado Limnoperna fortunei, usando PCR quantitativo em tempo real
150
quantificação de um DNA (organismo) especí-fico em misturas complexas, e foram aplicadas não apenas na detecção e quantificação das lar-vas de mexilhão-dourado (Endo et al., 2009), mas também para larvas do abalone Haliotis kamtschatkana (Vadopalas et al., 2006), para 13 espécies de cirripédios (Endo et al., 2010), para o molusco asiático Corbula amurensis (Smith et al., 2012), entre outros. O qPCR mo-nitora a amplificação de DNA em “tempo real” e detecta o DNA de espécies alvo. A quantidade de DNA molde inicial é quantificada baseada no fator de amplificação. O fluxograma de uma detecção de qPCR e o método de quantificação de larvas de mexi-lhão-dourado é exibido na Figura 1. As amos-tras de plâncton obtidas em campo são subme-tidas à extração de DNA e à análise de qPCR usando um conjunto de primers específicos para o mexilhão-dourado. O conjunto de pri-mers foi desenvolvido no gene mitocondrial ci-tocromo c oxidase subunidade 1 (CO1) do me-xilhão-dourado para a amplificação de qPCR. A amplificação é altamente específica para a espécie e amplifica apenas 138pb do fragmento de DNA do mexilhão-dourado, mesmo quan-do há DNA de outras espécies. A amplificação do fragmento de DNA por qPCR utilizando o conjunto de primers indica a existência de lar-vas de mexilhão-dourado na amostra analisa-da, sendo o número de larvas calculado pelo fator de amplificação. O método qPCR permite a detecção e quantificação por meio de um pro-cesso de análise simples, que economiza tem-po. Além disso, com o método qPCR é possível detectar e quantificar larvas com precisão, pois não é preciso experiência prévia em identifica-ção de larvas de mexilhão-dourado. Este capítulo explica o procedimento de pré-tratamento de amostras para análise da qPCR. Desta forma, será demonstrado como ocorre o processo de detecção e quantificação de larvas de mexilhão-dourado usando a qPCR.
Método de amostragem e preparação de amostras para análise de qPCR
No método padrão para coleta de amos-tras de plâncton do campo se utiliza uma rede de plâncton (NXX13, 300 mm de diâmetro).
Geralmente, se coletam amostras a par-tir de 3 puxadas verticais da rede, a aproxima-damente 10 m de profundidade em um ponto de amostragem. O fluxômetro é útil para o cálculo da densidade larval. Para uma análi-se mais precisa, é importante reduzir a lama que contamina a amostra. Os ácidos húmicos e outras substâncias húmicas da lama formam complexos com o DNA extraído que não são separados facilmente e interferem na extração do DNA. Além disso, essas substâncias inibem a reação da Taq polimerase, que é a reação enzimática-chave do PCR (Burgmann et al., 2001).
Figura 1. Fluxograma de uma detecção de qPCR e método de quantificação das larvas de mexilhão-dourado. Os zooplânctons maiores e outras substâncias nas amostras são removidos atra-vés de malhas de nylon e a solução resultante é submetida à ex-tração de DNA. Posteriormente, o DNA extraído é usado como modelo para a análise quantitativa de PCR em tempo real (qPCR).

CAPÍTULO 12
Seção 3
151
As amostras são imediatamente fixadas em etanol 99,5% e armazenadas a 4°C até se-°C até se-C até se-rem submetidas à extração de DNA. Esse pro-cesso evita que ocorra a degradação do DNA na amostra pela DNase, sendo particularmente importante para uma detecção e quantificação precisa quando o método qPCR é usado.
Extração do DNA molde a partir de amostras de plâncton obtidas em campo
Os zooplânctons maiores e outras subs-tâncias, como areia, folhas caídas e demais resíduos contidos nas amostras são removidos com uma malha de nylon de 420 μm. O plânc-ton que passa pela malha, incluindo as larvas do mexilhão-dourado, é coletado utilizando malhas de nylon de 74 μm. Um método de isolamento de DNA base-ado em filtro é indicado para análise de qPCR. Em geral, a eficiência da extração de DNA é re-duzida em proporção ao aumento da quantida-de de amostras. A baixa eficiência na extração de DNA é uma das causas para a diminuição da precisão das análises de detecção e quantifica-ção. Assim, quantidades grandes de amostras de plâncton devem ser divididas em duas a dez partes iguais (peso úmido aproximado de 5 a 25 mg, consulte o manual de instruções do kit de extração do DNA), colocadas em tubos de coleta de amostras de 1,5 ou 2 ml e submetidas separadamente à extração de DNA. O etanol é removido do tubo de coleta de amostra utilizando um evaporador centrífugo e o plâncton é homogeneizado em um tubo de co-leta com 3 a 5 esferas de zircônio (1 a 3 mm de diâmetro) utilizando um homogeneizador “bead beater” em velocidade máxima por 5 minutos. Em seguida, o DNA total é extraído do plâncton homogeneizado com o método de isolamento de DNA baseado em filtro usando os kits de extra-ção Quick Gene-Mini80 e Quick Gene (Kurabo Industries, LTD., Osaka, Japão). Se as amostras estiverem contaminadas com grandes quantidades de lama, a eficiência da extração de DNA pode ser aprimorada com o uso de reagente especializado na extração de DNA do solo (por exemplo, o kit de isolamen-to de DNA do solo da Norgen Biotek Corpo-ration, Ontário, Canadá). A solução de DNA
resultante deve ser armazenada a uma tempera-tura entre -20 e -70°C até ser submetida à aná-°C até ser submetida à aná-C até ser submetida à aná-lise de qPCR.
Amplificação de qPCR
A análise de qPCR é conduzida pelo mé-todo de intercalação SYBR® Green I usando um SYBR® Premix Ex TaqTM. A fluorescência do corante SYBR® Green I aumenta entre 100 e 200 vezes quando vinculado ao sulco menor de um DNA de cadeia dupla e o sinal fluorescente corresponde à quantidade de DNA de cadeia dupla na solução da reação. A fluorescência é monitorada pelo detector do sistema PCR em tempo real ao fim da etapa de prolongamento da reação de PCR, tendo e a eficiência da reação é calculada. A amplificação de qPCR é realizada com um conjunto de primers específico para o mexilhão-dourado (Limf-F: 5’-TCTCTTCAT-TTAGCTGGTGCGTC-3’ e Limf-R: 5’-CAA-AACAGATGTAACTGCAAGAGAC-3’, Endo et al., 2009) usando como um sistema Smart Cycler II (TaKaRa Bio Inc.). A reação de PCR em tempo real é conduzida em um volume total de 25 μl de solução de reação contendo 12,5 μl de SYBR® Premix Ex TaqTM, 2 μl de DNA molde extraído da amostra de plâncton, 1 μl de cada primer específico (0,2 μM cada) e 8,5 μl de água para PCR. Para a verificação da conta-minação dos reagentes, sempre deve ser usado com as amostras o controle negativo da reação com a água para PCR em vez do DNA molde. A reação de amplificação é conduzida a 95°C por 10 segundos, seguida por 40 ciclos de 95°C por 5 segundos e 60°C por 30 segundos. Após a amplificação, o valor do ciclo limite (Ct) é calculado automaticamente a partir do cruza-mento entre o traçado de amplificação e a linha limite (Fig. 2). O valor de Ct obtido é traçado nas curvas padrão e a quantidade de larvas é calculada (conforme será descrito adiante).
Verificação de especificidade da amplificação de qPCR
A especificidade das reações de amplifica-ção é examinada através de curvas de desnatu-ração geradas após a amplificação da qPCR. A análise das curvas de desnaturação é usada para distinguir amplificações alvo de produtos não

Endo & Nogata
Método de detecção e quantificação de larvasdo mexilhão-dourado Limnoperna fortunei, usando PCR quantitativo em tempo real
152
específicos, como dímeros de primer e/ ou “mis-primed”. A análise da curva de desnaturação é realizada com uma transição de temperatura linear de 60°C a 95°C após a amplificação da qPCR. A fluorescência é medida continuamente e a temperatura de desnaturação dos produtos do qPCR é determinada pela diminuição de fluo-rescência causada pela degeneração. O resultado da análise da curva de desnaturação da amplifi-cação da qPCR do mexilhão-dourado é exibido na Figura 3. A temperatura de desnaturação do produto da qPCR obtido com um conjunto de primers específico para o mexilhão-dourado é de aproximadamente 79°C. Os produtos que apresentam menor temperatura de desnaturação são provavelmente produtos da amplificação de dímeros de primer ou “misprimed”. A especifi-cidade da qPCR pode ser aprimorada através da otimização das condições da reação de acordo com o aparelho e os reagentes. Em alguns casos a causa é a degradação dos primers, ou a con-centração muito alta de DNA (incluindo DNA alvo e não alvo) na solução de reação. Uma boa prática é realizar a análise da curva de desnatu-ração após cada reação de qPCR para controlar a qualidade da identificação e da detecção.
Análise dos dados
A quantidade de larvas de mexilhão--dourado na solução de DNA é estimada tra-çando-se os valores de Ct na curva padrão. Quando a amostra de plâncton é subdividida
em várias partes, a quantidade de larvas na so-lução de DNA é multiplicada pelo número de divisões. A partir da quantidade de água fil-trada na amostragem de plâncton obtida em campo é possível calcular a densidade larval. A curva padrão é obtida a partir da amplificação de 2 μl de DNA molde procedente da qPCR de uma série de larvas (por exemplo, 1, 2, 5, 10, 20 e 50 indivíduos, Figura 4). A amplifica-ção para curvas padrão é conduzida conforme descrito acima. As eficiências de amplificação (E) foram calculadas a partir da inclinação das curvas padrão de acordo com a equação E = 10(-1/inclinação)-1. Um E de 1,0 corresponde a uma eficiência de amplificação de 100%. Os dados quantitativos de alta qualidade podem ser ob-tidos na faixa E que vai de 0,9 a 1,1 e quando houver um coeficiente de correlação alto (R2 > 0,98). Ao usar um reagente de lotes diferen-tes, uma nova curva padrão deve ser preparada para uma análise mais precisa. A análise de qPCR permite estimar não apenas a quantidade de larvas de mexilhão--dourado, mas também a quantidade de DNA de mexilhões dourados encontrados em amos-tras de plâncton. A quantidade de DNA de me-xilhões dourados é calculada traçando-se os valores de Ct na curva padrão obtida de dilui-ções seriais do DNA molde (por exemplo, 0,5, 2,5, 5, 10, 25 e 50 ng / μl) extraído de um me-xilhão-dourado adulto. O conteúdo do DNA na amostra está estreitamente relacionado ao número de larvas, sendo assim, a dinâmica
Figura 2. Exemplo de uma amplificação experimental realizada em um sistema Smart Cycler® II (TaKaRa Bio Inc.). A amplificação de qPCR foi conduzida em um conjunto de primers específico para mexilhões dourados e 2 µl de DNA molde extraídos de uma série de larvas do mexilhão (1, 2, 5, 10, 20 e 50 indivíduos). O sinal de fluorescência é monitorado continuamente e o traçado de amplificação foi obtido a partir da unidade de fluorescência relativa. Assim, a linha limite foi determinada automaticamente para a posição ideal, e os valores de Ct foram calculados a partir do cruzamento do traçado de amplificação e a linha limite.
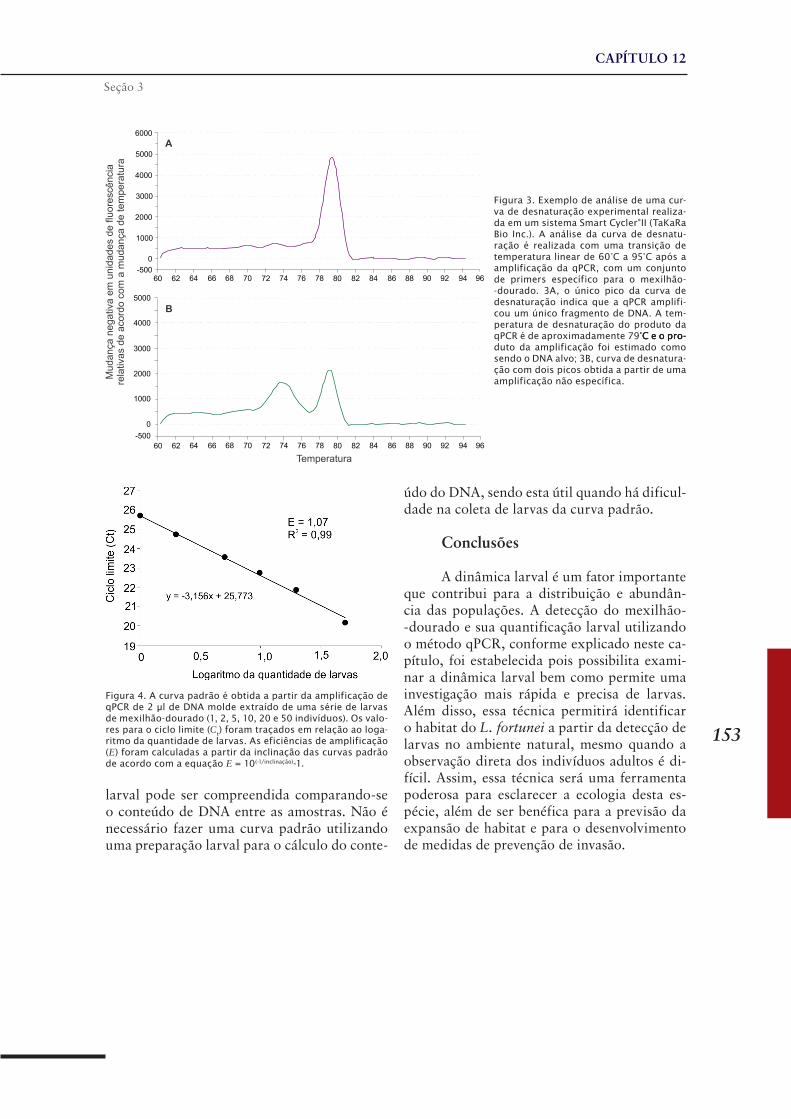
CAPÍTULO 12
Seção 3
153
Figura 3. Exemplo de análise de uma cur-va de desnaturação experimental realiza-da em um sistema Smart Cycler®II (TaKaRa Bio Inc.). A análise da curva de desnatu-ração é realizada com uma transição de temperatura linear de 60°C a 95°C após a amplificação da qPCR, com um conjunto de primers específico para o mexilhão--dourado. 3A, o único pico da curva de desnaturação indica que a qPCR amplifi-cou um único fragmento de DNA. A tem-peratura de desnaturação do produto da qPCR é de aproximadamente 79°C e o pro-°C e o pro-C e o pro-duto da amplificação foi estimado como sendo o DNA alvo; 3B, curva de desnatura-ção com dois picos obtida a partir de uma amplificação não específica.
Figura 4. A curva padrão é obtida a partir da amplificação de qPCR de 2 µl de DNA molde extraído de uma série de larvas de mexilhão-dourado (1, 2, 5, 10, 20 e 50 indivíduos). Os valo-res para o ciclo limite (C
t) foram traçados em relação ao loga-
ritmo da quantidade de larvas. As eficiências de amplificação (E) foram calculadas a partir da inclinação das curvas padrão de acordo com a equação E = 10(-1/inclinação)-1.
larval pode ser compreendida comparando-se o conteúdo de DNA entre as amostras. Não é necessário fazer uma curva padrão utilizando uma preparação larval para o cálculo do conte-
údo do DNA, sendo esta útil quando há dificul-dade na coleta de larvas da curva padrão.
Conclusões
A dinâmica larval é um fator importante que contribui para a distribuição e abundân-cia das populações. A detecção do mexilhão--dourado e sua quantificação larval utilizando o método qPCR, conforme explicado neste ca-pítulo, foi estabelecida pois possibilita exami-nar a dinâmica larval bem como permite uma investigação mais rápida e precisa de larvas. Além disso, essa técnica permitirá identificar o habitat do L. fortunei a partir da detecção de larvas no ambiente natural, mesmo quando a observação direta dos indivíduos adultos é di-fícil. Assim, essa técnica será uma ferramenta poderosa para esclarecer a ecologia desta es-pécie, além de ser benéfica para a previsão da expansão de habitat e para o desenvolvimento de medidas de prevenção de invasão.


155
Macroinvertebrados bentônicos são ani-mais que habitam o substrato de lagos, rios, arroios, estuários e mares, podendo viver den-tro ou sobre o substrato, por meio da constru-ção de tubos ou casas, fixos sobre rochas ou materiais orgânicos, nos quais vivem durante parte ou todo o ciclo de vida. Por convenção, segundo Hauer & Resh (1996), esses consistem na fauna de invertebrados que é retida numa malha com diâmetro de abertura de 500 μm; no entanto, alguns estádios de vida em que os organismos apresentam menor tamanho não são retidos nesta malha, o que justifica o uso de malhas de aberturas inferiores (250-300 μm). A comunidade de macroinvertebrados bentônicos límnicos é composta por uma va-riedade de grupos taxonômicos, incluindo mo-luscos, insetos (adultos e imaturos), crustáceos, anelídeos etc. Um dos grupos mais representa-tivos em número de espécies depois dos insetos são os moluscos de água doce. Somente dentre os bivalves são conhecidos cerca de 114 espé-cies para o Brasil (ver Capítulo 5). Apesar do
pequeno número de espécies de moluscos in-vasores – 5 de bivalves e 4 de gastrópodes –, o impacto destas invasoras pode ser devasta-dor pra os demais componentes da macrofauna bentônica. Até a década de 1990, a bioincrustação em ecossistemas límnicos da América do Sul era pouco representativa, salvo aquelas forma-das pelas espécies do filo Porifera. No entanto, com a introdução do mexilhão-dourado, Lim-noperna fortunei (Dunker, 1857), em diversas bacias hidrográficas brasileiras, um novo cená-rio se estabeleceu para macrofauna bentônica límnica. O mexilhão-dourado produz fios de bisso que possibilitam a incrustação do mes-mo sobre diferentes tipos de substratos duros formando macroaglomerados. Darrigran & Armengol (1998) e Darrigran et al. (1998a) ve-rificaram intensas modificações na estrutura da fauna bentônica na bacia do Rio La Plata (Argentina) em decorrência da colonização de substratos duros pelo mexilhão-dourado. O monitoramento de moluscos invasores
CAPÍTULO 13
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
Daniel PereiraJanine Oliveira Arruda
Paulo Eduardo Aydos Bergonci Arthur Schramm de Oliveira
Raoni PostiglioneMaria Cristina Dreher Mansur

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
156
numa bacia hidrográfica deve considerar não somente a quantificação da população destas invasoras, mas também a avaliação da estrutu-ra da comunidade bentônica, com o objetivo de verificar alterações e impactos sobre esta fauna tão importante para a manutenção dos ecossis-temas aquáticos e seus níveis tróficos. O plano de amostragem deve conside-rar distintas áreas da bacia, desde tributários de altitudes elevadas até as terras baixas, nas quais estações de coleta deverão ser definidas com base nas zonas do rio, seus trechos de cor-redeiras e poções, bem como os respectivos mi-cro-habitats (rocha contínua, seixos, cascalho, areia, silte/argila, folhiço) típicos destes trechos (Figs. 1 e 2). Na bacia do arroio Velhaco (Fig. 2), o diagnóstico da distribuição espacial da malacofauna nativa e invasora considerou dis-tintas unidades de paisagem da bacia e Pereira
em fase de licenciamento devem ser diagnosti-cados quanto à presença de espécies invasoras, com o objetivo de subsidiar o manejo da fauna bentônica após a operação, bem como a mini-mização de impactos gerados por bioincrus-tação em decorrência de uma planta geradora industrial projetada de forma alheia à presença de organismos bioincrustantes.
1. Equipamentos e métodos de amostragem
Diferentes equipamentos e métodos de amostragem da fauna bentônica e, em especial, da malacofauna límnica, foram desenvolvidos para situações específicas encontradas em di-ferentes ambientes e micro-habitats. A relação comentada destes diferentes equipamentos e métodos de amostragem é descrita a seguir.
et al. (2011a) constataram que este gradiente ambiental foi determinante na composição de espécies ao longo do rio. No caso de represas, deve-se avaliar a localização de estações de amostragem no sen-tido da corrente do canal principal do rio, bem como de transeções perpendiculares à margem com no mínimo três pontos equidistantes que deverão ser escolhidos com base na batimetria e morfometria do reservatório (Fig. 3). O plano de amostragem de uma represa deve prever a localização de estações de amostragem desde o início do reservatório, aonde a água ainda é corrente, até o barramento, além dos seus prin-
cipais tributários. A Figura 4 exibe um plano de amostragem utilizado para o diagnóstico da malacofauna bentônica da UHE Ibitinga no Rio Tietê. Critérios semelhantes devem ser aplicados a sistemas lacustres, lagunares e es-tuarinos, desta vez não considerando corrente, mas sim a direção dos ventos que influencia nos processos deposicionais dos corpos d’água.O monitoramento deve ser realizado de for-ma integrada nas bacias, com base na parce-ria entre diversas instituições. Não apenas os reservatórios construídos devem ser monito-rados quanto à presença de moluscos e outros invertebrados invasores, mas também aqueles
Figura 1. Microhabitats de rios e riachos. Desenho adaptado de FISRWG (1998) por Daniel Pereira.

CAPÍTULO 13
Seção 3
157
1.1. Amostradores
A escolha do amostrador adequado re-quer um planejamento prévio, no qual se deve considerar o tipo de ambiente (rio, riacho, re-presa, lago, lagoa, laguna, açude, etc.). Tam-bém é importante considerar a profundidade do local, o tipo de substrato e o habitat a ser amostrado. Amostradores com grande superfí-cie amostral resultam em amostras grandes que demandam elevado tempo de processamento. Amostradores com superfície amostral peque-na resultam em amostras menores que conse-quentemente demandam menor tempo de pro-cessamento; no entanto, selecionam o tamanho dos organismos a serem amostrados. Além do mais, a variabilidade das unidades amostrais aumenta em razão do efeito de borda. O mais
adequado é escolher um tamanho de amos-trador que possibilite a amostragem de popu-lações dos diferentes grupos taxonômicos que compõem a comunidade. O delineamento de um plano amostral deve considerar aspectos da morfologia do corpo d’água como: tipologia e declividade de margem; áreas de poção e corredeira no caso de rios; diferentes substratos; áreas de maior e menor velocidade; diferentes profundidades; a localização de tributários; proximidade a lo-cais propícios a contaminação por invasoras (portos, piers, ancoradouros). O ideal é que em cada micro-habitat de um determinado sistema sejam obtidos, no mí-nimo, três unidades de amostra. No entanto, o mais aconselhável e realizar uma amostragem piloto preliminar com a finalidade de avaliar a suficiência amostral (ver Brandimarte et al., 2004). A periodicidade deve ser no mínimo sa-zonal, mas quanto se trata do monitoramento de espécies invasoras e seu impacto sobre a fauna bentônica é recomendado amostragens mensais.
1.1.1. Pegadores ou busca-fundos Coletam uma área conhecida de substra-to por meio de sua penetração no mesmo, em função de seu peso; o fechamento é provocado por mecanismos ativados por molas, gravida-de, braços em alavanca ou cordas (Brandimarte et al., 2004).
Figura 2. Unidades de paisa-gem de uma bacia e estações de coleta em um diagnóstico das espécies límnicas nativas e invasoras da malacofauna. Fonte: Pereira et al. (2011), Bio-temas 24.
Figura 3. Estações de coleta situadas em um trecho do rio Tietê e seus tributários para o diagnóstico da distribuição de espécies invasoras e nativas da malacofauna. Mapa adaptado de Tundisi et al. (2008), Estudos Avançados 22(63) por Daniel Pereira.

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
158
1.1.1.1. Busca-fundo de Petersen: feito de ferro ou aço inoxidável; possui duas garras ou mandíbulas como se fossem valvas da concha de um molusco bivalve; o busca-fundo pode apresentar diferentes tamanhos, variando em área de 0,06 a 0,09 m2 (Fig. 5); seu peso apro-ximado é de 13,7 kg, podendo chegar até 31,8 kg, quando pesos auxiliares são adicionados nos lados; para usá-la, as mandíbulas abertas são lentamente colocadas no fundo da estação de coleta, evitando a perturbação dos materiais
mais leves; para soltar a trava que mantém as mandíbulas abertas, libera-se a tensão exercida na corda; quando o busca-fundo é levantado, o sistema de alavanca fecha as mandíbulas, mor-dendo o substrato e coletando a amostra; é am-plamente usado para amostrar fundo compacto constituído de areia, cascalho e argila em áreas de correnteza e águas profundas. (APHA, 1992). Hongzhu et al. (1999) coletaram exem-plares de Nemertea, Nematoda, Annelida, Mollusca (classes Bivalvia e Gastropoda) e Ar-thropoda no estudo preliminar de zoobentos do Lago Poyang (China), utilizando um busca--fundo de Peterson de 0,0625 m2. Wanganeo et al. (2011), no estudo dos macroinvertebrados bentônicos em dois reservatórios artificiais de Bhoj (município de Bhopal, Índia) utilizaram o mesmo amostrador, porém com 0,027 m2
de área, e capturaram exemplares dos grupos Mollusca (incluindo exemplares do gênero Li-mnoperna), Arthropoda, Annelida e Crustacea.
1.1.1.2. Busca-fundo de Van Veen: os longos braços do amostrador tendem a estabi-lizá-lo sem perturbar a água na interface água/substrato; este é basicamente uma versão me-lhorada do busca-fundo de Petersen (Fig. 6), sendo usado nos substratos lama, cascalho, sei-xos e areia; este amostrador é pesado e deve ser lançado por meio de uma plataforma de barco ou navio, por elevadores mecânicos ou hidráu-licos; utilizado em amostragens em mar aberto e em grande lagos (APHA, 1992). No estudo de Abílio et al. (2007) sobre os macroinvertebrados aquáticos da bacia hidrográ-fica do Rio Taperoá, bioma caatinga (Brasil), foi utilizado um busca-fundo Van Veen de 0,04 m2 de área, cujos grupos amostrados foram Arthro-poda, Mollusca (classes Bivalvia e Gastropoda), Crustacea e Annelida. Ourives (2010), utilizando o mesmo amostrador, porém com 0,12 m2, em seu estudo da distribuição espacial da macrofauna bêntica no estuário do Rio Cachoeira (município de Ilhéus, Bahia, Brasil) coletou exemplares de Mollusca (classes Bivalvia e Gastropoda), Polycha-eta, Nemertea, Echinodermata e Crustacea.
1.1.1.3. Busca-fundo de Ekman: feito de latão ou aço inoxidável com bitola de 12 a 20 cm, pesando aproximadamente 3,2 kg (Fig. 7); a parte que parece uma caixa, e que segura a
Figura 4. Proposta de um plano de amostragem da malaco-fauna bentônica e macroinvertebrados associados em reser-vatórios. Pontilhado amarelo demarcam transecções centrais paralelas à margem e perpendiculares à margem; as bolinhas amarelas representam pontos de amostragem. Em zoom são verificados habitats importantes para amostragem como ro-chas, a vegetação flutuantes e o sedimento de fundo. Dese-nho: A.S. de Oliveira.
Figura 5. Busca-fundo de Petersen de dois distintos modelos: A: aço inox leve para águas mais calmas; B: aço inox pesada para rios correntosos. Fotos: D. Pereira e J.O. Arruda.

CAPÍTULO 13
Seção 3
159
amostra, possui mandíbulas na parte inferior, operadas por molas, que são armadas manual-mente (tomar cuidado ao erguer as mandíbulas e engatilhá-las, pois estas podem se desarmar e causar danos às mãos do operador); no topo do busca-fundo estão duas tampas sobrepos-tas curvadas, que são mantidas parcialmente abertas pela água que passa através do compar-timento de amostragem durante a descida do amostrador; quando este está sendo retirado da água, as tampas são mantidas fechadas pela pressão da água; o acionamento do fechamento das mandíbulas é feito através de um mensagei-ro; as dimensões mais utilizadas são: 15 x 15 cm (0,0225 m2), 23 x 23 cm (0,0529 m2) e 30 x 30 cm (0,09 m2); o amostrador não é apropria-do para fundos rochosos, arenosos e ou em am-bientes com crescimento moderado de macró-fitas, porque pequenos seixos, areia ou talos de macrófitas dificultam o fechamento adequado das mandíbulas; é utilizado exclusivamente em amostragens de substrato lodoso e com lama, em água com pouca correnteza (APHA, 1992). Na investigação sobre a comunidade de macroinvertebrados do sistema hidrológico do Taim (Rio Grande do Sul, Brasil), Würdig et al. (2007) utilizaram um amostrador busca-fundo Ekman, coletando exemplares de Oligochaeta, Turbellaria, Hirudinea, Gastropoda, Bivalvia (incluindo exemplares de bivalves invasores da família Corbiculidae), Crustacea e Insecta. Si-raj et al. (2010) coletaram Mollusca (gastrópo-des da família Lymnaeidae e bivalves diversos, incluindo da família Corbiculidae), Annelida e Arthropoda no estudo ecológico dos macro-bentos no charco de Shallabugh (Índia) usando o busca-fundo Ekman de 0,0015 m2. 1.1.1.4. Busca-fundo de Ponar: similar ao busca-fundo de Petersen em tamanho, peso, sistema de alavanca, mecanismo de fechamen-to e compartimento de amostra (Fig. 8); possui placas laterais e uma tela na parte superior do compartimento da amostra para evitar perda desta durante o processo de fechamento; pode ser usado em todos os tipos de substrato, ex-ceto em rochas, sendo mais eficiente quando usado em areia, cascalho ou em pequenas pe-dras com lama; geralmente utilizado em rios de profundidade média a profunda, em lagos e reservatórios (APHA, 1992).
Figura 6. Busca-fundo de Van Veen. Acima, da esquerda para a direita: vista ventral com as mandíbulas fechadas, ventral com as mandíbulas abertas; abaixo, vista dorsal e lateral. Fo-tos: A.S. de Oliveira.
Figura 7. Busca-fundo de Ekman, com as mandíbulas fechadas (esquerda) e abertas (direita). Fotos: A.S. de Oliveira.
Figura 8. Busca-fundo de Ponar, com as mandíbulas fechadas (esquerda) e abertas (direita). Fotos: A.S. de Oliveira e M.V. Nehrke.

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
160
Ramsey & White (2007) coletaram exem-plares de Arthropoda, Annelida e Mollusca (bi-valves, incluindo a família Corbiculidae) utili-zando busca-fundo Ponar de 0,0522 m2 de área amostral no Lago Kentucky (EUA). Taurusman (2010) fez uso do mesmo amostrador, porém com 0,023 m2 de área amostral, em seu estudo sobre a comunidade de macroinvertebrados da baía de Jakarta (Índia), coletando exemplares de Polychaeta, Bivalvia e Crustacea.
1.1.2. Dragas Usada em amostragens quali-quantati-vas. A draga é usada para capturar membros da epifauna, mas devido a sua limitada penetração no sedimento, não é adequada em amostragens de animais que se enterram (Holme & McIn-tyre, 1971).
1.1.2.1. Draga de arrasto modelo adap-tado de Holme e McIntyre (1971): consiste basicamente em duas armações principais de ferro no formato de um retângulo; uma arma-ção molda a abertura do amostrador; a haste de ferro do retângulo é perfurada e apresen-ta duas alças em forma de meia-lua nas late-rais de menor comprimento; na parte superior das alças se encontram argolas para amarrar os cabos ou cordas nas quais será suspenso o amostrador; outro retângulo sem alças forma o fundo do amostrador; ambos os retângulos são conectados por hastes de ferro aparafusadas; anteriormente à montagem da estrutura metá-lica, os sacos, o externo, de pano, e o interno, com a malha amostral adequada, são presos aos retângulos por anilhas plásticas; as laterais de maior comprimento do retângulo da abertura devem ser denteadas, com a finalidade de pene-trar no substrato (Fig. 9); a draga, amarrada a um cabo, deve ser colocada na água na posição horizontal e descida lentamente para que não mude de posição, até que fique deitada sobre o fundo; feito isso, o barco deve percorrer uma distância “x” ou deve-se anotar o tempo de ar-raste; em todas as estações de coleta deve-se usar o mesmo procedimento; por meio de um guincho, o amostrador é puxado até o barco onde o mesmo é virado sobre uma bandeja para que a lama coletada seja escoada e posterior-mente peneirada.
1.1.3. Extratores Os extratores são usados em amostra-gens quali-quantitativas. Estes variam desde tubos puxados manualmente até modelos im-pulsionados por explosão com flutuação auto-mática (APHA, 1992).
1.1.3.1. Corers (do inglês, “extrator”): são artefatos cilíndricos, simples ou múltiplos, que coletam uma coluna de substrato por operação manual, gravidade ou envio de mensageiros (Fig. 10); possuem, frequentemente, mecanismo de fechamento na extremidade superior que impede a perda de material (Brandimarte et al., 2004). São usados para amostrar sedimentos profundos. São mais eficientes que o amostrador Surber, quando usados em com-binação com peneiras para sedimentos finos em uma área amostral pequena, de 0,0013 a 0,0026 m2. Para o uso eficiente como amos-tradores de superfície, é necessária uma densa população de animais.
1.1.4. Amostradores por Sucção São cilindros que são forçados contra o substrato e recebem sucção por bomba compressora (de ar ou água); consequentemente,
Figura 9. Draga de arrasto (modelo adaptado de Holme, 1971). Estrutura metálica em ferro (f). Foto: D. Pereira.
Figura 10. Amostrador Corer. Foto: A.S. de Oliveira.

CAPÍTULO 13
Seção 3
161
ocorre subida de água, de sedimentos e de organismos que são selecionados usando rede in situ ou são totalmente recolhidos e selecio-nados a posteriori (Brandimarte et al., 2004).
1.1.4.1. Amostradores por Sucção de Ki-kuchi et al. (2006): o corpo consiste de uma caixa quadrada de metal (placas de ferro galva-nizadas) de 30 cm de altura por 30 cm de lar-gura e 30 cm de profundidade; são adicionados mais 10 cm em cada largura para a adição de peso (Fig. 11); a parte de baixo da caixa é aber-ta e encontra-se em contato com o sedimento de fundo; as laterais da caixa são fechadas; na parte superior há um tubo de aço inoxidável de 5 cm de diâmetro e 25 cm de comprimento que penetra no meio da caixa; na extremidade supe-rior deste tubo, do lado de fora da caixa, é aco-plado uma mangueira para aspirar o sedimento; na extremidade posterior do tubo, que se encon-tra dentro da caixa, encontra-se um filtro com malha de 1 cm de diâmetro para impedir que grandes objetos sejam aspirados e consequente-mente danifiquem o equipamento; em cada uma das quatro extremidades da região superior da caixa há uma alça que contém cordas que são amarradas e se unem em uma braçadeira e esta ao cabo de aço do guincho; lastros são adiciona-dos sobre a superfície da caixa e nas laterais para dar estabilidade ao amostrador. Brooks & Boulton (1991) utilizaram amostrador de sucção que encobriu 0,05 m2 do substrato a 10 cm de profundidade nas estações de coleta do Rio Finniss (Austrália), onde exem-plares de Nematoda, Annelida, Mollusca (classe Gastropoda), Crustacea e Arthropoda foram amostrados. Kikuchi et al. (2006) coletaram Annelida, Mollusca (classes Bivalvia e Gastro-poda), Crustacea, Arthropoda e Platyhelmin-thes utilizando um amostrador de sucção de 30
cm de comprimento por 30 cm de largura e 30 cm de profundidade no Rio Tocantins (Brasil).
1.1.5. Delimitadores
Constam de aparelhos que delimitam uma área sobre o substrato para amostragem. Estes podem estar associados a redes ou não. A área do substrato é perturbada, em geral, manualmente, com consequente liberação dos animais, que são capturados pela rede. Exem-plo: amostradores Surber e Hess. Mais detalhes sobre sua estrutura e funcionamento são apre-sentados por Klemm et al. (1990). Tendo área delimitada, seus resultados são expressos em densidade por metro quadrado.
1.1.5.1. Quadrados ou cilindros: podem ser feitos de diferentes materiais como PVC, fer-ro, aço inoxidável e madeira, bem como podem apresentar diferentes tamanhos (Fig. 12). Este amostrador delimita uma área sobre um substra-to, onde os organismos são removidos por raspa-gem, com o auxílio de um pincel ou, no caso de sedimento fino, com o auxílio de uma espátula ou, simplesmente, catados manualmente.
Figura 11. Amostrador de suc-ção: 11A, visão tridimensional do amostrador; 11B, disposição dos equipamentos usados pelo amostrador de sucção, com o amostrador em si, uma bomba se sucção e uma rede. Dese-nhos: J.O. Arruda adaptado de Kikuchi et al. (2006), Acta Lim-nologica Brasiliensia, 18(1).
Figura 12. Delimitadores no formato quadrado (ao fundo) e ci-líndrico (à frente). Foto: D. Pereira e J.O. Arruda.

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
162
Pereira et al. (2001) coletaram moluscos límnicos no bentos marginal do arroio Capi-vara, município de Triunfo (Brasil), utilizando quadrats de 0,22 m2. Kikuchi & Uieda (2005), amostrando áreas no córrego Itaúna no mu-nicípio de Itatinga (Brasil), delimitadas por quadrante de 30 cm x 30 cm (0,09 m2), com profundidade máxima de 10 cm, coletaram Pla-tyhelminthes, Nematoda, Arthropoda, Mollus-ca (classes Bivalvia e Gastropoda) e Crustacea.
1.1.5.2. Surber: consiste em dois quadra-dos delimitadores de ferro (30 x 30 cm; 0,09 m2), unidos por uma extremidade articulável, sendo que um deles é posicionado horizontal-mente sobre o substrato enquanto que o outro fica na posição vertical (Fig. 13); o conjunto desses dois quadrados forma uma espécie de “L” dobrável; em cada lateral do “L” é preso um tecido em forma de triângulo; no quadrado vertical é preso um saco de malha de 250 μm
e 60 cm de comprimento; o amostrador é posi-cionado sob o substrato de forma que a abertu-ra do saco junto ao quadrado vertical seja posi-cionada contra a correnteza; o coletor revolve o substrato delimitado pela área do quadrado ho-rizontal e a correnteza leva os organismos para dentro do saco; os triângulos laterais de pano impossibilitam que os organismos fujam pela lateral, fazendo com que a corrente de água os direcione para dentro do saco de malha. Silva, L. et al. (2005), utilizando amostra-dor Surber de 0,225 mm de abertura de malha e 0,1 m2 de área no Rio das Almas e no córrego do Inferno (Minas Gerais, Brasil), coletaram exem-plares de Arthropoda, Mollusca (classe Bival-via), Annelida e Platyhelminthes. Kikuchi et al. (2006) coletaram Annelida, Mollusca (classes Bivalvia e Gastropoda), Crustacea e Arthropoda utilizando Surber em estações de coleta no Rio Tocantins (Brasil). Milesi et al. (2009) usaram o mesmo amostrador com 250 μm de abertura de malha e 0,1 m2 de área amostral em riachos da região norte do Rio Grande do Sul (Brasil), cole-tando organismos dos grupos Platyhelminthes, Annelida, Mollusca (classes Bivalvia e Gastro-poda), Crustacea e Arthropoda.
1.1.5.3. Hess: consiste em um cilindro de 50 cm de altura com 30 cm de diâmetro (0,15 m2); na parede do cilindro há duas aberturas re-tangulares (28 cm de largura por 40 cm de altu-ra; 0,112 m2), uma na região oposta da outra; em uma delas encontra-se acoplado uma malha de 250 μm, fechando esta abertura (região anterior); no retângulo oposto (região posterior) há aco-plado um saco no formato triangular de malha de 250 μm no qual, em sua extremidade distal, é acoplado um copo rosqueável similar aos uti-lizados em rede de plâncton; o cilindro também possui duas alças laterais situadas na sua parte su-perior; o cilindro é colocado na posição vertical sobre o substrato; apoiado pelas alças este é gira-do para a esquerda e para a direita até penetrar no substrato, sendo que a região anterior deve ficar posicionada no sentido contra a correnteza para que a corrente de água passe por essa abertura e saia pela posterior, onde se localiza o saco trian-gular (Fig. 14); o coletor revolve com as mãos o substrato delimitado pelo fundo do cilindro; a corrente de água que passa pelo equipamento carrega os detritos e os invertebrados para den-
Figura 13. Amostrador de Suber. Foto: D. Pereira e J.O. Arruda.
Figura 14. Amostrador de Hess. Foto: D. Pereira e J.O. Arruda.

CAPÍTULO 13
Seção 3
163
tro do saco triangular; depois de todo o material retido no saco ter sido deslocado para dentro do copo rosqueável, este é desenroscado e o material nele contido é acondicionado em potes ou sacos plásticos contendo álcool 80%. Metzeling et al. (2003) coletaram exem-plares de Mollusca (classes Gastropoda e Bi-valvia, incluindo exemplares da família Corbi-culidae), Arthropoda, Nemertea, Nematoda, Annelida, Crustacea e Platyhelminthes com um amostrador Hess de 0,1 m2 de área e 0,25 mm de abertura de malha em vários rios dos estados de Victoria e South Wales (Austrália). Kikuchi et al. (2006) coletaram Annelida, Mollusca (classes Bivalvia e Gastropoda), Crustacea e Arthropoda utilizando Hess em estações de coleta no Rio Tocantins (Brasil). Alonso & Ca-margo (2010), utilizando amostrador Hess com malha de 250 μm de abertura e 0,01815 cm2 de área amostral, coletaram no Rio Henares em Guadalajara (Espanha) espécimes de Crusta-cea, Arthropoda, Mollusca (classes Bivalvia e Gastropoda) e Platyhelmintes.
1.1.6. Conchas
1.1.6.1. Concha de captura (Carvalho et al., 2005): a concha de captura é constituída de um recipiente de alumínio de fundo crivado preso na ponta de um cabo (Fig. 15). A concha é empregada para coletar moluscos aderidos à vegetação submersa, emergente e flutuante, permitindo o escoamento da água e a retenção dos moluscos. Também pode ser utilizada para coletar moluscos junto ao sedimento de fundo. No estudo do gastrópode Melanoides tuberculatus no município de Guapimirim (RJ) realizado por Giovanelli et al. (2005), foi utili-zada uma concha metálica com malha de 4 mm de abertura, que era movida um passo a cada conchada aplicada, cobrindo uma distância de 30 m em 20 minutos para cada estação de cole-ta. Já no estudo de Núñez (2010), uma concha de captura com 15 cm de diâmetro e abertura da malha de 1,4 mm foi utilizada para coleta do caramujo Physa acuta no município de La Plata (Argentina).
1.1.6.2. Concha raspadora de superfí-cie: constituída de um recipiente de alumínio em forma de meia-lua com o fundo reto e cri-
vado, preso na ponta de um cabo (Fig. 16); a extremidade reta da meia lua é denteada; uti-lizada para raspar superfícies incrustadas por bivalves em paredões de concreto; a área amos-trada é delimitando, sendo a largura a mesma do amostrador e a distância perpendicular à largura do amostrador percorrida no processo de raspagem o comprimento da área amostral; a multiplicação do valor da distância raspa-da pela largura do amostrador fornece a área amostrada; a partir da área é possível calcular a densidade populacional na parede incrustada.
Figura 15. Coletores de moluscos ou conchas de captura. Foto: Ministério da Saúde (2007).
Figura 16. Concha raspadora de superfície. Foto: D. Pereira e J.O. Arruda.

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
164
1.2. Substratos artificiais
Os substratos artificiais são artefatos que procuram imitar certas características do ambiente a ser amostrado (Rosenberg & Resh, 1982), como disponibilizar material para colo-nização por organismos bentônicos. Esse ma-terial pode ser encontrado tanto no ambiente a ser amostrado quanto fora dele. Os substratos artificiais são ferramentas amplamente utiliza-das no monitoramento de macroinvertebrados límnicos e marinhos, especialmente de espécies que se fixam ou formam incrustações sobre su-perfícies duras. Os substratos artificiais padro-nizam a amostragem, reduzem a variabilidade e o tempo de processamento de amostras (Gi-bbons et al., 1993). Podem ser seletivos para certos organismos; não fornecem informação sobre o substrato local; requerem tempo de colonização relativamente longo para que seja alcançado o equilíbrio da comunidade, o que o torna inadequado para estudos de curta dura-ção, sujeito a perda ou a alteração de eficiência, em razão de vandalismo, sedimentação, secas e enchentes, possibilidade de perda de orga-nismos durante o recolhimento dos substratos (Brandimarte et al., 2004). O período mínimo de submersão de um substrato artificial para a avaliação da colo-nização da fauna bentônica em ecossistemas límnicos é de 75 dias, de acordo com Henri-ques-de-Oliveira (2002). Para a colonização de substratos pelo mexilhão-dourado recomenda--se o tempo mínimo de três e cinco meses para a visualização de incrustações por indivíduos adultos, o que varia em função da disponibi-
lidade de densidades elevadas de larvas no plâncton, bem como de outros fatores como a competição com outros organismos incrus-tantes (porífera, perifíton, etc.), a qualidade e velocidade da água. Quando se objetiva apenas avaliar o recrutamento do mexilhão-dourado, colonizações resultantes de um mês de expo-sição são suficientes. A magnitude da incrusta-ção pode ser avaliada por meio da quantifica-ção da densidade de indivíduos incrustados em substratos submersos em diferentes períodos de exposição (1 mês, 2 meses, 3 meses, ..., 12 meses), conforme a Figura 17, por meio de di-ferentes substratos que poderão ser escolhidos pelo coordenador do programa de monitora-mento ou pelo pesquisador ,de acordo com o objetivo proposto e a natureza dos dados que se deseja coletar. 1.2.1. Estruturas tridimensionais de amianto (Morton, 1977): são placas no formato de um quadrado que são encaixadas e que, em um primeiro momento, uma encontra-se perpendicular a outra, formando um eixo X e um eixo Y e, num segundo momento, adiciona-se uma outra placa que forma o eixo Z (Fig. 18). Esta estrutura tridimensional apresenta 24 superfícies que somadas fornecem a área amostral. Morton (1977), no estudo da dinâmica populacional de L. fortunei em um reserva-tório em Hong Kong, montaram duas séries de armações, penduradas a 0,3 m de profun-didade em intervalos de 0 m, 0,3 m, 0,6 m, 0,9 m e 1,2 m de distância entre si e ligadas por uma linha de nylon a um peso de concreto
Figura 17. Delineamento amostral de monitoramento do mexilhão dourado nas fases bentônicas (recruta e adulto) com substra-tos em forma de “x” de madeira submersos no delta do Jacuí, Porto Alegre, RS, durante projeto coordenado pelo IEAPM (anos de 2006-2008), com financiamento do CT-HIDRO. Tempos de exposição dos substratos: 1 mês, densidade mensal (M), 2 meses (2), 3 meses (3), 4 meses (4), 5 meses (5), ..., 12 meses (12). As profundidades não diferem estatisticamente quanto a profundidade; sendo assim, em cada conjunto se considera três réplicas. Desenho: D. Pereira.

CAPÍTULO 13
Seção 3
165
deitado sobre o fundo e a boias. Cada série de armações foi mensalmente inspecionada por um período de 28 meses para verificar a colonização pelo mexilhão-dourado. Uma vez colonizado, os exemplares de mexilhão foram medidos em intervalos mensais para acompa-nhar seu crescimento de mm até o maior com-primento. A cada mês, seis exemplares adultos de mexilhão-dourado foram coletados sobre pedras dentro do reservatório para a inspeção de seus ctenídeos (também conhecidos como brânquias), visando busca de ovos encubados e larvas. No final do experimento as arma-ções foram levadas ao laboratório e o número total de exemplares assentados em cada uma das superfícies e fendas de cada armação foi registrado e medido. A temperatura e o oxigê-nio dissolvido foram registrados, no local do experimento a 0 m, 0,6 m e 1,2 m de profun-didade. Estes dados têm sido correlacionados à época de reprodução e o assentamento de larvas de mexilhão, objetivando estimar valo-res ótimos sobre quando esses processos ocor-rerão.
1.2.2. Estruturas tridimensionais de PVC (Boltovskoy & Cataldo, 1999): ar-mações compostas por três placas retangulares de PVC dispostas entre si para definir três pla-nos perpendiculares (Fig. 19); esta é presa ao ancoradouro por dois furos opostos; este de-senho garante que as superfícies orientadas em todas as direções fiquem disponíveis para colo-nização e que todo o aparato fique orientado da mesma forma, com a base da frente para a jusante; o material aderido à superfície da ar-mação é removido e filtrado em um malha de
25 μm de abertura, com o intuito de eliminar as partículas de argila e sílica; os mexilhões maiores que 2-3 mm de comprimento retidos são manualmente separados; o remanescente da amostra é reamostrando utilizando um se-parador de plâncton Folsom até se obter uma fração contendo ao menos 150 indivíduos juve-nis menores que 2 mm. Boltovskoy & Cataldo (1999) estuda-ram a dinâmica populacional do mexilhão--dourado no baixo Rio Paraná (Argentina) utilizando amostradores tridimensionais de PVC. Os autores fizeram amostragens men-sais ao longo de 11 meses, em um ano onde houve extrema influência do fenômeno cli-mático El Niño. Os autores concluíram que a reprodução de L. fortunei foi contínua entre os meses de setembro e março, sendo que no período de temperaturas mais baixas houve um declínio acentuado da reprodução na po-pulação estudada.
Figura 18. Estrutura tridimensional de amianto proposto por Morton (1977). Desenho: J.O. Arruda e R. Postiglione. Adaptado de Morton (1977), Malacologia 16(1).
Figura 19. Amostrador tridimensional de PVC: 19A, estrutura formada por três placas de PVC perpendiculares; 19B, esque-ma de um separador de plâncton de Folsom. Desenhos: Raoni Postiglione. Adaptado de Boltovskoy & Cataldo, Biofouling (1999), 14(3) por R. Postiglione.

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
166
1.2.3. Estruturas tridimensionais de PVC (Sylvester et al., 2007a): armação cons-tituída de uma gaiola formada por três placas de PVC mutuamente perpendiculares, forman-do quatro seções; três delas são recobertas por telas protetoras de plástico com aberturas de malha de 40, 15 e 5 mm cada uma (Fig. 20); a armação é colocada dentro da água na área amostral e presa um metro abaixo da superfície d’água por um peso de 20 kg.
da última réplica; após o período de exposição adotado, os substratos colonizados são primei-ramente lavados sobre rede de plâncton 30 μm com auxílio de água destilada e de um pincel para a remoção de recrutas e, posteriormente, de espátulas para a remoção de adultos; os fil-trados são fixados em álcool 70%; a contagem total é realizada no laboratório e quando ele-vadas densidades são verificadas é realizada a quantificação por subamostragem; para tal, o material fixado é novamente filtrado por uma rede de 150 μm; o material retido é examinado ao microscópio estereoscópico para a remoção e quantificação total dos adultos; o restante do filtrado contendo os recrutas é removido para um copo de béquer de 25 ml, com o auxílio de um pincel e água destilada; o material contido no béquer é avolumado com álcool 70%; o vo-lume é agitado (25 vezes); com uma seringa é retirada uma alíquota (2 ml) e removida para uma placa quadriculada (área total de 23 cm2 e quadrícula de 1 cm2); a placa é agitada sobre a bancada por meio de movimentos circulares (25 vezes nos sentidos horário e anti-horário); sob o microscópio estereoscópico o material contido na placa é quantificado; com base na área amostral do substrato artificial (0,0722 m2) também são calculadas as densidades mé-dias expressas em m2. Oliveira et al. (2006) usaram amostra-dor artificial de concreto para o estudo do mexilhão-dourado no Rio Paraguai, Porto Es-perança (Brasil) e, posteriormente, adotaram o substrato de madeira na forma de X. Os au-tores encontraram uma densidade média de 523,8 e 238 ind.m-2 nos meses de agosto de 2001 e fevereiro de 2002, respectivamente. Mansur et al. (2009) utilizaram o substrato de madeira no formato de um X no estudo da colonização do mexilhão-dourado no delta do Rio Jacuí e no cais do porto em Porto Alegre (RS, Brasil). Foram utilizadas três réplicas do substrato artificial, expostas por três meses, nas profundidades de 1, 1,5 e 2 m. Os autores não encontraram diferença significativa quanto às diferentes profundidades testadas e nem em relação à posição do mexilhão no substrato ar-tificial. Entretanto, encontraram diferença na densidade média dos adultos entre as estações de coleta estudadas. As densidades médias de adultos encontradas por Oliveira et al. (2006)
Figura 20. Amostrador tridimensional de PVC. Adaptado de Sylvester et al. (2007), Hydrobiologia 589 por R. Postiglione.
Sylvester et al. (2007a) utilizaram a arma-ção de PVC revestida por tela plástica de malha de 5 mm para coletar L. fortunei no delta do baixo Rio Paraná (Argentina). Os demais gru-pos animais coletados junto com o mexilhão--dourado foram Oligochaeta, Chironomidae, Hirudinea, Rotifera, Copepoda, Turbellaria, Nematoda, Insecta, Naupii, Tardigrada, Cla-docera, Tanaidacea, Ostracoda, Hydracarina e outros moluscos da classe Gastropoda.
1.2.4. Estruturas tridimensionais de ma-deira no formato de “X” (Fontes et al., 2002; Mansur et al., 2009): o substrato consiste em um “x” de madeira com alças metálicas pelas quais o amostrador é fixado a um píer (Fig. 21), paredão de concreto ou plataforma flutuante por meio de cordas; geralmente são fixadas três réplicas conectadas por corda a 0,5 m de dis-tância uma da outra (em canais entre 5 e 6 m de profundidade não se verifica diferenças signifi-cativas entre as réplicas, de acordo com Mansur et al., 2009); para que o substrato permaneça na posição vertical na coluna d’água, um peso (rocha, tijolo ou metal) é fixado na alça inferior

CAPÍTULO 13
Seção 3
167
foram inferiores a observadas por Mansur et al. (2009) utilizando o mesmo substrato.
1.2.5. Substrato cerâmico – tijolo (Santos et al., 2008): os tijolos de cerâmica são retan-gulares e perfurados medindo aproximadamen-te 14 cm de comprimento x 10 cm de largura (0,014 m2); os tijolos são inseridos em suportes de ferro na forma de estantes, que por sua vez são cravados no sedimento; permanecem sus-pensos a uma altura de no mínimo 20 cm do fundo (Fig. 22); também podem ser fixados sus-pendendo-os por cordas em um píer, plataforma flutuante, paredão de hidrelétrica ou porto; a quantificação dos indivíduos fixados ao substra-to é realizada nas duas fases bentônicas de de-senvolvimento do mexilhão (recruta< 0,5 mm; adulto>0,5 mm); o microscópio estereoscópico com ocular milimetrada é utilizado na quanti-ficação; a área avaliada do tijolo é computada para que se possa calcular a densidade (ind.m-2).
Na investigação do mexilhão-dourado no delta do Rio Jacuí, Lago Guaíba e laguna dos Patos no Rio Grande do Sul, Mansur et al. (2003) utilizaram tijolos de cerâmica de 14 X 10,5 x 9,5 cm, vazados por seis perfurações de 3 cm e duas de 1 cm de diâmetro para coleta quantitativa. Estes foram colocados quinzenal-mente e mensalmente, na periferia e dentro de juncais [Scirpus californicus (C. A. Mey) Steud.], para a avaliação da incidência e do tempo de fixação do L. fortunei. Além desses, outros ti-jolos foram mantidos por três, seis meses e um ano no local de amostragem. Cada tijolo era numerado e ancorado ao fundo, através de uma haste de ferro com 2 cm de comprimento e 0,5 cm de diâmetro, transpassada por uma abertu-ra de menor diâmetro do tijolo e enterrada no substrato a uma profundidade de aproximada-mente 50 cm. Como a haste era longa, parte dela encontrava-se emersa, facilitando a locali-zação do ponto de coleta. Esta permaneceu ao
Figura 21. Substrato artificial de madeira em forma de X: 21A, esquema dos substra-tos suspensos por cordas em diferentes profundidades, ancorados no sedimento por um tijolo cerâmico; 21B, terminolo-gia do substrato cerâmico; 21C, substra-to sem incrustação (área: 0,072m2); 21D, substrato com incrustação; 21E, visão da superfície superior do substrato com in-crustações. Fonte: Mansur et al., 2009, Biotemas 22(1).
Figura 22. Substrato artificial cerâmico (tijo-lo) para amostragem do mexilhão-dourado: 22A, substratos fixados em suportes de ferro na forma de estante; 22B, substrato incrustado pelo mexilhão-dourado. La-teral canaliculada incrustada (p) e furo incrustado (f). Fonte: Santos et al., 2008, Revista Brasileira de Zoologia 25(3).

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
168
longo de todo o período de amostragem, sendo substituídos periodicamente apenas os tijolos. Santos et al. (2008) utilizaram tijolos para o acompanhamento do crescimento do mexilhão-dourado na praia do Veludo em Porto Alegre (RS, Brasil). As autoras utiliza-ram seis conjuntos de quatro tijolos cada na área amostral, que foram substituídos a cada seis meses e retirados ao final de um ano. Na contagem e medida do comprimento máximo dos indivíduos adultos, foram consideradas as quatro superfícies maiores de cada tijolo além de distância de 1 cm dentro de cada perfuração dos tijolos. Os animais foram medidos usando paquímetro digital de 0,01 mm de precisão. As autoras encontraram uma diferença significati-va do tamanho dos indivíduos entre as estações primavera-verão e outono-inverno.
1.2.6. Substrato cerâmico – tijoleta (Ber-gonci et al., 2009): o substrato (Fig. 23A, B) se constitui de uma tijoleta de cerâmica medindo 21 cm de comprimento, 6 cm de largura e 1 cm de espessura; os substratos são perfurados a 2 cm da extremidade com furadeira elétrica; são fixados por meio de anilhas plásticas em estru-turas metálicas denominadas bastidores (Fig. 23C); os bastidores são suspensos por cordas junto a um píer, plataforma flutuante, paredão de hidrelétrica ou porto; a quantificação dos indivíduos fixados ao substrato é realizada nas duas fases bentônicas de desenvolvimento do mexilhão (recruta<0,5 mm; adulto>0,5 mm); o microscópio estereoscópico com ocular mi-limetrada é utilizado na quantificação, com o auxílio de uma malha transparente quadricu-lada e segmentada (Fig. 24), sobreposta às su-
perfícies do substrato; em cada segmento são sorteadas cinco quadrículas (Qij) de 1 cm2, nas quais se quantificam os indivíduos recrutas e adultos; por meio da equação 1 (Dsn = (ΣQij/5)) são calculadas as densidades de indivíduos (ind.cm-2) nos três segmentos (Dsn; n = I, II ou III), de cada superfície; quando não são detectados indivíduos nas quadrículas de um segmento, procede-se à quantificação total dos indivídu-os no segmento, dividindo-se o valor resultan-te da quantificação pela área do segmento (42 cm); em seguida são obtidas as densidades das superfícies (Dln; n = lisa, I ou canaliculada, II) por meio da equação 2 (Dln = µDsn); a densidade média (D, ind.cm-2) de indivíduos no substrato cerâmico é obtida por meio da soma das densidades de ambas superfícies por meio da equação 3 (D = ΣDln). Os bastidores foram adaptados de mode-los desenvolvidos por Caprari (2006) com a fi-nalidade de fixar corpos de prova utilizados na avaliação de revestimentos anti-incrustantes. Bergonci et al. (2009) utilizaram o subs-trato artificial tijoleta para amostragem do mexi-lhão-dourado no cais do porto e no canal do Rio Jacuí em Porto Alegre (RS, Brasil). Os autores
Figura 23. Substrato cerâmico (tijoleta): 23A, superfície canelada; 23B, superfície lisa; 23C, bastidor de ferro com substra-tos cerâmicos (tijoletas) fixados, modifi-cado de Caprari (2006). Fonte: P.E.A. Ber-gonci et al., 2009, Biotemas 22(3). Fotos: D.Pereira e P.E.A. Bergonci.
Figura 24. Malha quadriculada para sobreposição no substra-to artificial e orientação da quantificação. Segmentos (Sn) na tijoleta: SI, SII e SIII. Fonte: Bergonci et al., 2009, Biotemas 22(3).

CAPÍTULO 13
Seção 3
169
compararam as densidades médias de recrutas e adultos nas duas estações de coleta, bem como nas distintas superfícies da tijoleta (lisa e canali-culada) e seus segmentos. Concluíram que houve diferença estatística nas densidades de recrutas e adultos nas estações de coleta, além de terem encontrado diferença entre os segmentos e as su-perfícies da tijoleta na estação do cais do porto.
1.2.7. Garrafa PET preenchida com o retículo celulósico da fruta da bucha (Lufa ci-lindra L.) (Volkmer-Ribeiro et al., 2004): cada amostrador consiste em duas garrafas verdes tipo PET descartáveis de dois litros cada, cujo fundo de cada garrafa é cortado com o auxí-lio de uma tesoura e desprezado; a cor verde é para diminuir a depredação causada pelo ser humano, uma vez que uma garrafa verde sub-mersa é mais difícil se ser vista do que uma gar-rafa transparente (Fig. 25); o restante das gar-rafas é lixado, tanto a superfície interna como a externa; as tampas são mantidas, fechando as aberturas de ambas as garrafas; as garrafas são cortadas longitudinalmente em seis tiras de aproximadamente 2 cm de largura, começando pela base, com o auxílio de uma tesoura; em cada tira são feitos pequenos cortes transver-sais a cada 2 mm, deixando-os com aparência de um pente; dentro das garrafas se adiciona um enchimento natural, constituído por uma rede de celulose que abriga as sementes da planta Luffa cylindra L.; o enchimento é adap-tado para o tamanho das garrafas que o abri-garão; uma garrafa é inserida dentro da outra, com as aberturas tampadas dispostas em lados opostos e com o enchimento dentro delas; as garrafas são então envoltas por uma pequena rede de nylon, cuja abertura é fechada sob a tampa de uma das garrafas PET; uma peça de metal pesando aproximadamente 300 g é fixa-da próximo à tampa de uma das garrafas; um fio de nylon de aproximadamente 3 m é fixado à tampa da outra garrafa; o amostrador é fi-xado, utilizando a corda de nylon, à vegetação adjacente ou a um tronco nas margens da esta-ção de coleta e, em seguida, arremessado para dentro da água; para a retirada do amostrador da água, desprende-se o fio de nylon amarrado à margem do rio e usa-o como guia para deter-minar o local exato onde a garrafa se alojou, to-mando o cuidado para não deslocá-la; um saco
plástico é mergulhado no fundo do rio e então o amostrador é ensacado, sendo o saco amar-rado e só então emerso da água; o saco plásti-co contendo o amostrador é colocado em um balde de plástico, aberto, adicionado formalina 5% para fixar o material, novamente fechado e transportado para o laboratório; o conteúdo de cada saco, junto com o amostrador, é evertido sobre uma peneira forrada com algodão e lava-do sobe água corrente; em seguida as garrafas, a rede de nylon e a rede de celulose são secas para posterior exame da fauna associada; os macroinvertebrados são identificados e conta-bilizados com o uso de estereomicroscópio. Volkmer-Ribeiro et al. (2004) utilizaram uma garrafa PET preenchida pela planta Luffa cylindra L. para amostragem de macroinverte-brados bentônicos no Rio Caí e no arroio Bom Jardim em Triunfo (RS, Brasil). Foram captu-rados exemplares de Turbellaria, Oligochaeta, Mollusca (classes Gastropoda e Bivalvia, in-cluindo o mexilhão-dourado), Crustacea, In-secta, Bryozoa e Porifera. Devido à efetividade na captura dos grupos mais representativos de macroinvertebrados bentônicos, os autores re-comendaram o uso deste amostrador no moni-toramento desta comunidade.
1.2.8. Garrafa PET com preenchimento por folhas de taboa (Typha dominguensis Pers.) (Queiroz et al., 2007): o coletor de substrato artificial desenvolvido baseia-se no estudo de Volkmer-Ribeiro et al. (2004); são utilizadas garrafas tipo PET com oito furos de 2 cm de diâmetro cada e 5 cm de distância entre eles, sendo quatro furos na parte superior e quatro
Figura 25. Esquema do amostrador de garrafa PET preenchido com retículo celulósico da fruta da bucha. Desenho: R. Rosa. Fonte: Volkmer-Ribeiro et al., 2004, Brazilian Journal of Bio-logy 64(3A).

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
170
na parte inferior da garrafa; além dos furos, é feito nas garrafas cortes longitudinais de 17 cm de comprimento por 1,5 cm de largura, unindo os furos superiores e inferiores de cada garra-fa; estes cortes têm a função de permitir que se coloque 60 g de folhas de taboa (Typha do-minguensis) dentro de cada garrafa (Fig. 26) e, também, para que estas folhas se mantenham em contato com a água; as garrafas são fixa-das em varas de bambu com 3 m de compri-mento utilizando uma abraçadeira de plástico, mantendo-se uma distância de 20 cm entre a garrafa e a ponta da vara de bambu inserida no fundo dos viveiros, de forma a manter a garra-fa exatamente sobre o sedimento do fundo dos viveiros; no topo de cada vara de bambu são colocadas marcações, como bandeiras colori-das, para diferenciar o tempo de permanência de cada garrafa na estação de coleta; após o tempo de amostragem, a garrafa é retirada da água e as amostras são acondicionadas em sa-cos plásticos. Queiroz et al. (2007) coletaram princi-palmente representantes de Gastropoda (famí-lia Thiaridae), Oligochaeta (Glossiphonidae) e Diptera (Chironomidae), utilizando amostra-dores artificiais de garrafa PET preenchidos por folhas de taboa em tanques de piscicultura no município de Itupeva (SP, Brasil). Os autores concluíram que em 40 dias ocorreu a coloniza-ção dos amostradores, sendo que após este pe-ríodo constatou-se uma diminuição no número de organismos no substrato.
1.2.9. Garrafa PET com tela de nylon (Pe-reira et al., 2010, adaptado de Volkmer-Ribeiro et al., 2004): consiste numa garrafa PET com uma tela de nylon (10 x 10 cm) presa na parede interna do amostrador; a superfície do amostrador é per-furada com a finalidade de permitir o fluxo da água; após a colonização o fundo e o gargalo da garrafa é removido com o auxílio de uma tesoura (Fig. 27); a malha de nylon (superfície amostral 1) é removida e fixada em álcool 70%; a quantifica-ção total de macroinvertebrados é realizada sob o microscópio estereoscópico, sendo os resultados expressos em densidade média (ind.m-2); a malha de nylon é um material inerte; após a retirada da malha, a lateral da garrafa é aberta, originando um retângulo de aproximadamente 576 cm2 (su-perfície amostral 2), o que varia de acordo com o fabricante do recipiente; o material fixado na superfície externa é quantificado sob o microscó-pio estereoscópico, os resultados podem ser ex-pressos em densidade de organismos (ind.m2) ou em percentual de cobertura (%), especialmente no caso de esponjas incrustantes. Pereira et al. (2010) amostraram molus-cos bivalves (mexilhão-dourado) e gastrópo-des, bem como Chironomidae e Oligochaeta no delta do Rio Jacuí e no cais do porto de Porto Alegre (RS, Brasil) utilizando o substrato artificial acima descrito.
1.3. Substratos naturais
Os substratos naturais são muitos variá-
Figura 26. Amostrador de garrafa PET preenchida com por folhas de taboa: 26A, amostrador artificial feito com garrafa PET e vara de bambu; 26B, detalhamento do amostrador com preenchido internamente com folhas de taboa. Fonte: Queiroz et al., 2007, Circular Técnica EMBRAPA 16.

CAPÍTULO 13
Seção 3
171
veis quanto ao tamanho e a forma. As macrófitas apresentam diferentes formas biológicas e ocu-pam diferentes habitats. As rochas apresentam diferentes tipos de texturas, porosidade, tama-nho e composição mineral. Desta forma, a coleta quantitativa nestes substratos é uma tarefa difícil. Alguns procedimentos são descritos a seguir:
1.3.1. Raízes e talos de água pé: o agua-pé-de-baraço Eichhornia azurea (Sw.) Kun-th forma colônias fixas (Fig. 28), sendo que sua raiz mais profunda se adere ao substrato. Logo, essa porção da raiz serve de substra-to para os bentos. Para a coleta dos bentos, puxa-se para fora da água, pela folhagem flu-tuante, um dos talos da macrófita. Continua--se puxando, com cuidado para não romper o talo e sem deixar que ele retroceda para dentro d’água, até que as várias cabeleiras de raízes mais novas saiam da água. Na sequên-cia, são deslocadas as raízes fixas no substra-to do fundo, o que se observa quando, além do barro, a raiz apresenta alguma quantidade de lodo e, o talo e a raiz passam a apresentar cor marrom enegrecida. Essa secção do talo, com uma ou mais cabeleiras de raízes, deve ser cortada do talo subjacente e colocada em saco plástico para análise dos bentos. Uma vez a raiz retirada da água, esta não deve ser novamente mergulhada no rio. A cabeleira se-lecionada opera como uma “draga”, trazendo junto a si uma porção do substrato com a fau-na dela contida. Por isso procura-se exercer tração contínua no talo de modo a evitar que as raízes entrem novamente na água e assim se perca o material retido nas cabeleiras. No la-boratório, cada amostra retirada do saco plás-tico é lavada em uma bacia com água comum,
através de imersões sucessivas até que as cabe-leiras das raízes fiquem limpas do sedimento e do lodo. O lodo, água e sedimento que se encontram no saco plástico também são verti-dos nesta mesma bacia. O lodo resultando da lavagem é deixado para decantar. A água so-brenadante é escorrida utilizando um pano de algodão para reter eventuais animais. O lodo, então, é passado aos poucos em água corrente através do mesmo pano para retenção da fau-na nele contida. Em seguida o pano é fechado em trouxa e mergulhado dentro de um frasco contendo álcool 70% para posterior contagem dos organismos. Estes são contados a olho nu e também através de estereomicroscópio com aumento de até 10x. As raízes lavadas são se-paradas dos caules, restando destes apenas os nós. Raízes e nós de uma mesma amostra são colocados em uma bandeja forrada com jor-nal para retirada do excesso de umidade. Esta bandeja é colocada em uma estufa com circu-lação forçada de ar para secagem. No caso de haver disponível uma estufa comum ao invés de uma com circulação forçada de ar, o jornal da bandeja deverá ser trocado sempre que ficar encharcado ou muito úmido e as raízes e caules deverão ser reviradas para acelerar a perda de água e evitar que a amostra mofe. Se possível, a amostra deverá ficar exposta ao sol ou em um ambiente que possua umidade baixa. Quando o jornal estiver pouco úmido, praticamente seco, a bandeja é então levada a uma estufa a 70ºC para secagem. O processo de secagem em uma estufa comum, secagem prévia da amostra com o uso de jornal, leva em torno de uma semana. Após a secagem das raízes e caule, a amostra é pesada em uma balança digital analítica de 0,01 g de precisão. Com o peso do substrato seco e a contagem do número de indivíduos da amostra, relaciona-se o percentual (para cada 100 g de raiz) ao número de indivíduos de cada um dos grupos faunísticos encontrados na amostra. O mesmo procedimento de triagem e secagem pode ser realizado a partir dos talos do aguapé. Os resultados devem ser expressos separadamente para raízes e talos.
1.3.2. Rochas: podem ser coletadas manualmente na beira de rios e lagos, ou em áreas mais profundas por meio de mergulho (Fig. 29); devem-se coletar rochas de tamanhos
Figura 27. Esquema do amostrador de garrafa PET com tela de nylon. Superfície amostral 1 (a), malha de nylon (área: 100cm2) fixada na lateral interior da garrafa; superfície amostral 2 (b), retângulo (área: 576cm2) formado pelo corte da lateral exte-rior da garrafa PET nas linhas tracejadas (lt); orifícios (o) com diâmetro de 1cm. Fonte: Pereira et al., 2010, Biotemas, 23(1).

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
172
similares; os organismos aderidos são removi-dos com pincel umedecido em água destilada e por raspagem com espátula; os resultados po-dem ser expressos em densidade de organismos (ind.m-2), quando for possível quantificar a área da rocha (sendo que esta não é uma tarefa fácil) ou como abundância de organismos em cada ré-plica do substrato. Um método apropriado para a quantificação de organismos em rochas encon-tra-se descrito em Fischer & Gaupp (2004).
2. Processamento de amostras em campo e no laboratório
2.1. Peneiramento das amostras de sedimento
As amostras de sedimento coletadas com busca fundo, dragas e delimitadores (sem rede) são peneiradas em uma série de peneiras com malhas diversas sobrepostas sobre uma mesa (Fig. 30) e lavadas com água corrente que pode ser jorrada por uma mangueira conectada a uma bomba a bateria.
2.2. Acondicionamento das amostras
As amostras são acondicionadas em potes plásticos em diferentes tamanhos (Fig. 30A). As etiquetas de identificação devem ser inseridas no interior do frasco junto à amostra e externamente ao frasco. A etiqueta interna é feita com papel vegetal e caneta nanquim e posteriormente inserida num tubo de eppen-dorf (Fig. 30B) que é colocado dentro do fras-co, evitando se assim que o papel rasgue ou se degrade. A etiqueta externa é feita com fita crepe larga com boa propriedade adesiva e a anotação deve ser feita a lápis ou nanquim para que no caso de vazamento do álcool este não remova a anotação.
Figura 28. O aguapé-de-baraço Eichhornia azurea (Sw.) Kunth enraizado no subs-trato na margem de um corpo d´água. Fonte: Volkmer-Ribeiro et al. 2004, Bra-zilian Journal of Biology 64(3A).
Figura 29. Macroinvertebrados aderidos a rochas: 29A, manchas de cor bege são poríferos; 29B, Limnoperna fortunei incrustado em rocha com fauna acompanhante de gastrópodes do gênero Potamolithus sp. Fotos: A, D. Pereira; B, M.C.D. Mansur.
Figura 30. Conjunto de peneiras com malhas de 1 mm; 0,5mm e 0,25 mm para a lavagem de sedimento e triagem prévia de organismos bentônicos. Foto: D. Pereira.
Figura 31. Materiais utilizados no acondicionamento das amos-tras: 31A, potes plásticos de diferentes tamanhos, picetas con-tendo água e álcool 70%, bandeja de plástico e alcoômetro; 31B, tubos plásticos tipo eppendorf (os dois à esquerda) e potes de acrílico (os dois à direita) que recebem. as etiquetas para identi-ficação da amostra. Fotos: D. Pereira e J.O. Arruda.

CAPÍTULO 13
Seção 3
173
Tab
ela
I. M
éto
dos
e eq
uip
am
en
tos
de
am
ost
rag
em
de
molu
scos
inva
sore
s e
mac
roin
vert
eb
rad
os
ass
oci
ad
os.
Mo
dif
icad
o d
e Bra
nd
imart
e et
al. (
20
04), b
ase
ad
o e
m o
bse
rvaç
ões
de
cam
po
e n
a lite
ratu
ra a
cim
a ci
tad
a.
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
BU
SCA
-FU
ND
OS
Pete
rsen
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
se u
sado
apr
opri
adam
en-
te;
perm
ite
amos
trag
em
quan
tita
tiva
; -
amos
tra
a m
aior
ia d
os t
i-po
s de
sub
stra
to;
- ide
al p
ara
amos
trar
loca
is
prof
undo
s.
- m
ater
ial d
e al
to c
usto
;-
mod
elo
padr
ão é
mui
to p
esad
o, r
eque
rend
o ba
rco
gran
de c
om g
uinc
ho;
- se
des
cido
rap
idam
ente
, po
de c
ausa
r on
das
de c
hoqu
e qu
e pe
rtur
bam
o f
undo
; -
man
díbu
las
não
pene
tram
pr
ofun
dam
en-
te,
port
anto
, or
gani
smos
em
pro
fund
idad
es
mai
ores
não
são
am
ostr
ados
; - m
andí
bula
s fac
ilmen
te b
loqu
eada
s por
det
ri-
tos,
cau
sand
o pe
rda
de m
ater
ial;
-
quan
do o
bstr
uído
, é n
eces
sári
o la
nçar
nov
a-m
ente
o a
mos
trad
or, o
que
pod
e re
quer
er u
m
mai
or t
empo
de
cole
ta, a
lém
de
caus
ar m
últi
-pl
as p
ertu
rbaç
ões
no s
ubst
rato
;- n
ão p
ode
ser
empr
egad
o em
fund
os la
jead
os
ou c
om r
ocha
s m
uito
gra
ndes
;-
os m
odel
os m
aior
es n
eces
sita
m d
e gu
inch
o pa
ra r
etor
nar
a su
perf
ície
com
as
amos
tras
;-
se a
bran
gem
áre
a m
uito
gra
nde,
com
o re
-su
ltad
o po
de h
aver
aum
ento
do
tem
po g
asto
na
tri
agem
.
Rio
s, r
iach
os e
rep
resa
s co
m
fund
os
de
lam
a,
arei
a e
casc
alho
.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados
Van
Veen
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
mec
anis
mo
de
fech
a-m
ento
da
s m
andí
bula
s m
elho
r qu
e do
bus
ca-f
un-
do P
eter
sen;
-
amos
tra
a m
aior
ia d
os
tipo
s de
sub
stra
to;
- id
eal
para
am
ostr
ar l
o-ca
is p
rofu
ndos
.
- m
ater
ial d
e al
to c
usto
;-
os m
aior
es s
ão m
uito
pes
ados
, re
quer
endo
ba
rco
gran
de c
om g
uinc
ho;
- m
andí
bula
s po
dem
ser
blo
quea
das
por
pe-
dras
e g
alho
s;
- nã
o é
efic
ient
e pa
ra o
rgan
ism
os e
scav
ado-
res;
- nã
o po
de s
er e
mpr
egad
o em
fun
dos
laje
a-do
s ou
com
roc
has
mui
to g
rand
es;
- ne
cess
ita
de g
uinc
ho p
ara
reto
rnar
a s
uper
-fí
cie
com
as
amos
tras
;-
se a
bran
gem
áre
a m
uito
gra
nde,
com
o re
sul-
tado
pod
e ha
ver
aum
ento
do
tem
po g
asto
na
tria
gem
.
Rio
s, r
iach
os e
rep
resa
s co
m
fund
os
de
lam
a,
arei
a e
casc
alho
.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
174
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
BU
SCA
-FU
ND
OS
Ekm
an
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
fáci
l op
eraç
ão
man
ual
em
água
ra
sa,
pode
ndo
ser
empu
rrad
a co
ntra
o
subs
trat
o;
- ab
ertu
ras
supe
rior
es
redu
zem
a
form
ação
de
on
das
de
choq
ue
ou
e pe
rtur
baçã
o do
sub
stra
to;
- am
ostr
a su
bstr
atos
de
la
ma
ou a
reia
;-
idea
l pa
ra a
mos
trar
lo-
cais
pro
fund
os.
- m
ater
ial d
e al
to c
usto
;- p
elo
fato
de
sere
m le
ves,
as
man
díbu
las
não
pene
tram
em
sub
stra
tos
duro
s;
- m
andí
bula
s fr
eque
ntem
ente
nã
o fe
cham
co
mpl
etam
ente
por
blo
quei
o ou
fal
ha n
o m
ecan
ism
o de
fec
ham
ento
; -
inef
icie
nte
onde
a v
eloc
idad
e da
águ
a é
re-
lati
vam
ente
alt
a;-
não
pode
ser
em
preg
ado
em f
undo
s la
jea-
dos
ou c
om r
ocha
s m
uito
gra
ndes
; -
se a
bran
gem
áre
a m
uito
gra
nde,
com
o re
-su
ltad
o po
de h
aver
aum
ento
do
tem
po g
asto
na
tri
agem
.
Lago
s, la
goas
, lag
unas
e
açud
es c
om f
undo
s de
la
ma
e ar
eia.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados
Pona
r
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
mel
hor
pene
traç
ão
que
outr
os b
usca
-fun
dos
(mes
mo
a ve
rsão
peq
uena
te
m b
oa p
enet
raçã
o, c
on-
side
rado
seu
tam
anho
);
- pl
acas
late
rais
e t
elas
re-
duze
m fo
rmaç
ão d
e on
das
de c
hoqu
e;
- m
elho
r am
ostr
ador
qu
anti
tati
vo p
ara
ambi
en-
tes
de á
gua
doce
; -
vers
ão
pequ
ena
pode
se
r op
erad
a m
anua
lmen
te
sem
bar
co o
u gu
inch
o;-
amos
tra
a m
aior
ia d
os
tipo
s de
sub
stra
to;
- id
eal
para
am
ostr
ar l
o-ca
is p
rofu
ndos
.
- m
ater
ial d
e al
to c
usto
;-
quan
do e
m g
rand
es d
imen
sões
são
mui
to
pesa
das,
req
uere
ndo
um b
arco
de
port
e co
m-
patí
vel c
om a
inst
alaç
ão d
e um
gui
ncho
; -
man
díbu
las
pode
m
perm
anec
er
aber
tas
quan
do
obst
ruíd
as
por
pedr
as
e de
trit
os,
caus
ando
per
da d
e am
ostr
a;
- a
vers
ão p
eque
na n
ão é
efi
cien
te e
m á
guas
rá
pida
s;-
se a
bran
gem
áre
a m
uito
gra
nde,
com
o re
-su
ltad
o po
de h
aver
aum
ento
do
tem
po g
asto
na
tri
agem
.
Rio
s, r
iach
os e
rep
resa
s co
m
fund
os
de
lam
a,
arei
a e
casc
alho
.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados

CAPÍTULO 13
Seção 3
175
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
DR
AG
A
Dra
ga d
e ar
rast
o m
odel
o ad
apta
do
de H
olm
e (1
971)
- pe
rmit
e a
cole
ta d
e or
-ga
nism
os
mai
ores
qu
e nã
o sã
o ca
ptur
ados
pel
os
dem
ais
equi
pam
ento
s;-
perm
ite
uma
amos
tra-
gem
m
ais
repr
esen
tati
va
da
com
posi
ção
de
es-
péci
es
do
sedi
men
to
de
fund
o.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- dif
ícil
oper
ação
, poi
s a
drag
a po
de n
ão d
es-
cer
na p
osiç
ão c
orre
ta e
sub
ir s
em m
ater
ial;
- ne
cess
ita
de g
uinc
ho p
ara
reto
rnar
a s
uper
-fí
cie
com
as
amos
tras
;-
cole
ta a
pena
s or
gani
smos
ent
erra
dos
su-
perf
icia
lmen
te n
o se
dim
ento
e n
ão c
aptu
ra
orga
nism
os e
scav
ador
es;
- po
de f
icar
pre
sa a
o fu
ndo
quan
do p
rese
nte
roch
as, g
alho
s ou
tro
ncos
.
Lago
s, la
goas
e la
guna
s.A
bund
ânci
a C
orbi
cula
larg
illie
rti
Cor
bicu
la fl
umin
eaC
orbi
cula
flum
inal
isC
orbi
cula
sp.
Lim
nope
rna
fotu
nei
Mel
anoi
des
tube
rcul
atus
Mac
roin
vert
ebra
dos
asso
ciad
os
CO
RE
RS
Cor
ers
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
boa
pene
traç
ão e
m s
edi-
men
tos
mol
es;
- pe
quen
o vo
lum
e da
uni
-da
de
amos
tral
pe
rmit
e an
ális
e de
núm
ero
mai
or
de r
éplic
as e
m c
urto
in-
terv
alo
de t
empo
; -
no
caso
do
s m
anua
is,
pode
m s
er u
sado
s em
am
-bi
ente
s ra
sos;
-
pequ
ena
pert
urba
ção
da
inte
rfac
e ág
ua/s
ubst
rato
.
- ta
man
ho
pequ
eno
da
unid
ade
amos
tral
po
de s
er p
robl
ema
em s
itua
ções
em
que
os
orga
nism
os s
ão e
scas
sos
ou c
om t
aman
ho
mai
or q
ue a
áre
a am
ostr
al;
- nã
o re
tém
are
ia;
- pod
e oc
orre
r a
perd
a de
org
anis
mos
peq
ue-
nos
e ep
ibên
toni
cos
em f
unçã
o de
ond
as d
e ch
oque
;-
a va
riab
ilida
de d
as u
nida
des
amos
trai
s au
-m
enta
em
raz
ão d
o ef
eito
de
bord
a.
Lago
s, la
goas
, lag
unas
e
açud
es.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.M
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados
SUC
ÇÃ
O
Am
ostr
ador
es
por
sucç
ão
- fr
eque
ntem
ente
pe
r-m
item
es
tim
ativ
as
po-
pula
cion
ais
com
m
aior
ex
atid
ão q
ue p
egad
ores
e
core
rs.
- op
eraç
ão r
eque
r eq
uipa
men
tos
caro
s (p
or
exem
plo,
com
pres
sor
de a
r) e
, as
veze
s, m
er-
gulh
ador
em
loca
is p
rofu
ndos
.
Rio
s de
gra
nde
port
e,
com
el
evad
a pr
ofun
-di
dade
e s
ubst
rato
ro-
chos
o do
tip
o la
jead
o,
casc
alho
, se
ixos
, ar
eia
e lo
do.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.M
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
176
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
DE
LIM
ITA
DO
RE
S
Qua
drad
o ou
ci
lindr
o
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
faci
lmen
te t
rans
port
ado;
- id
eal
para
a c
olet
a de
bi-
valv
es e
nter
rado
s em
sed
i-m
ento
s;-
idea
l pa
ra c
olet
as e
m l
o-ca
is r
asos
.
- se
a c
olet
a fo
r fe
ita
em u
ma
área
com
águ
a co
rren
te,
o m
ater
ial
cole
tado
pod
e se
r de
s-lo
cado
, per
dend
o se
ass
im p
arte
da
amos
tra;
- nã
o pe
rmit
e co
leta
em
loca
is p
rofu
ndos
.
Ria
chos
ras
os,
mar
gens
de
lago
s, la
goas
e d
e la
-gu
nas.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
usM
acro
inve
rteb
rado
s as
soci
ados
Surb
er
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
faci
lmen
te t
rans
port
ado;
- id
eal p
ara
cole
tas
em lo
-ca
is r
asos
.
- a
efic
iênc
ia d
a am
ostr
agem
dep
ende
de
corr
ente
za,
não
send
o po
ssív
el a
mos
trar
em
ág
ua p
arad
a;-
difi
culd
ade
de d
ispo
siçã
o em
alg
uns
tipo
s de
sub
stra
to;
- ef
iciê
ncia
bai
xa e
m f
luxo
len
ta p
orqu
e os
or
gani
smos
não
são
arr
asta
dos
para
o i
nte-
rior
da
rede
; -
amos
trag
em li
mit
ada
a pr
ofun
dida
des
infe
-ri
ores
a 3
0 cm
.-o
s or
gani
smos
com
alt
a m
obili
dade
pod
em
esca
par
quan
do o
sub
stra
to é
per
turb
ado;
-o
rgan
ism
os m
enos
móv
eis
pode
m s
er s
upe-
rest
imad
os e
m r
elaç
ão a
os c
om m
aior
mob
i-lid
ade.
Rio
s e
riac
hos,
pri
nci-
palm
ente
em
cor
rede
i-ra
s.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.M
elan
oide
s tu
berc
ulat
usPh
ysa
acut
aH
elis
oma
spp.
Mac
roin
vert
ebra
dos
asso
ciad
os
Hes
s
- en
cont
rado
no
mer
cado
em
vár
ios
tam
anho
s;-
pode
ser
usa
do e
m le
itos
co
bert
os p
or v
eget
ação
;-
idea
l par
a co
leta
s em
lo-
cais
ras
os.
-os
orga
nism
os c
om a
lta
mob
ilida
de n
ão e
scap
am
quan
do
o su
bstr
ato
é pe
rtur
bado
(po
r ex
empl
o ca
rang
uejo
s da
fam
ília
Ae-
glid
ae).
- a
efic
iênc
ia d
a am
ostr
agem
dep
ende
de
corr
ente
za,
não
send
o po
ssív
el a
mos
trar
em
ág
ua p
arad
a;-
difi
culd
ade
de d
ispo
siçã
o em
alg
uns
tipo
s de
sub
stra
to;
- am
ostr
agem
lim
itad
a a
prof
undi
dade
s in
fe-
rior
es a
80
cm;
essa
pro
fund
idad
e po
de s
er
vari
ável
de
acor
do c
om a
alt
ura
do c
ilind
ro
do a
mos
trad
or.
Rio
s e
riac
hos,
pri
nci-
palm
ente
em
cor
rede
i-ra
s.
Den
sida
de (
m2 )
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.M
elan
oide
s tu
berc
ulat
usPh
ysa
acut
aH
elis
oma
spp.
Mac
roin
vert
ebra
dos
asso
ciad
os

CAPÍTULO 13
Seção 3
177
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
CO
NC
HA
S
Con
cha
cole
tora
de
mol
usco
s
- m
ater
ial d
e ba
ixo
cust
o;-
fáci
l man
usei
o;-
adeq
uado
par
a am
bien
-te
s lê
ntic
os, s
em b
atim
en-
to d
e on
das;
- ad
equa
do
para
co
le-
tas
qual
itat
ivas
em
áre
as
com
veg
etaç
ão s
ubm
ersa
, em
erge
nte
e fl
utua
nte,
al
ém
de
sedi
men
to
de
fund
o.
- nã
o é
adeq
uado
par
a co
leta
s qu
anti
tati
vas;
- se
o co
leto
r in
clin
ar d
entr
o d’
água
, a a
mos
-tr
a po
de s
er p
erdi
da.
Rio
s, r
iach
os,
repr
esas
, la
gos,
lag
oas,
lag
unas
e
açud
es.
Abu
ndân
cia
Cor
bicu
la la
rgill
iert
iC
orbi
cula
flum
inea
Cor
bicu
la fl
umin
alis
Cor
bicu
la s
p.M
elan
oide
s tu
berc
ulat
usPh
ysa
acut
aH
elis
oma
dury
iH
elis
oma
triv
olvi
sM
acro
inve
rteb
rado
s as
soci
ados
Con
cha
rasp
ador
a de
sup
erfíc
ie
- m
ater
ial d
e al
to c
usto
;-
fáci
l man
usei
o;-
adeq
uado
par
a am
bien
-te
s lê
ntic
os,
com
ou
sem
ba
tim
ento
de
onda
s;-
idea
l pa
ra c
olet
as q
uan-
tita
tiva
de
m
olus
cos
in-
crus
tant
es e
m p
ared
ões
de
conc
reto
em
hid
relé
tric
as,
port
os, e
tc.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- nã
o é
adeq
uado
par
a su
perf
ície
s ir
regu
la-
res;
- um
a co
leta
efi
cien
te r
eque
r um
a ra
spag
em
cont
ínua
e u
nifo
rme.
Am
bien
tes
cons
truí
dos
nas
mar
gens
de
ri
os,
repr
esas
, lag
os, l
agoa
s e
lagu
nas.
Den
sida
de (
m2 )
Lim
nope
rna
fotu
nei
Mac
roin
vert
ebra
dos
asso
ciad
osM
acro
inve
rteb
rado
s as
soci
ados
SUB
STR
AT
OS
AR
TIF
ICIA
IS
Est
rutu
ras
trid
imen
sion
ais
de
amia
nto
(Mor
ton
et a
l., 1
977)
- m
ater
ial d
e ba
ixo
cust
o;-
não
exig
e es
trut
uras
adi
-ci
onai
s co
mo
bast
idor
es,
pode
ndo
ser
fixa
do
por
mei
o de
cor
das;
- ade
quad
o pa
ra a
mbi
ente
s lê
ntic
os,
com
ou
sem
bat
i-m
ento
de
onda
s e
lóti
cos
em
dife
rent
es
níve
is
de
corr
ente
za;
- ad
equa
do
para
lo
cais
pr
ofun
dos,
com
o re
pres
as;
- fá
cil
obse
rvaç
ão d
as d
is-
tint
as f
ases
de
vida
(be
ntô-
nica
s),
níve
is d
e co
loni
za-
ção,
e a
glom
erad
os;
- ad
equa
do
para
el
evad
a pr
ofun
dida
de.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- o
amia
nto
é um
mat
eria
l can
cerí
geno
;-
difí
cil
man
usei
o no
mic
rosc
ópio
est
ereo
s-có
pico
;- a
ras
page
m d
os a
glom
erad
os p
ara
cole
ta d
e in
diví
duos
adu
ltos
pod
e da
nifi
car
indi
vídu
os
recr
utas
e jo
vens
;-
os s
ubst
rato
s de
vem
ser
lav
ados
com
águ
a de
stila
da e
pin
cel p
ara
que
não
ocor
ra a
per
-da
de
indi
vídu
os jo
vens
e r
ecru
tas.
Rio
s,
repr
esas
, la
gos,
la
goas
e la
guna
s.D
ensi
dade
(m
2 )L
imno
pern
a fo
tune
i M
acro
inve
rteb
rado
s as
soci
ados

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
178
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
SUB
STR
AT
OS
AR
TIF
ICIA
IS
Est
rutu
ras
trid
imen
sion
ais
de
PVC
(B
olto
vsko
y &
Cat
aldo
, 199
9)
- m
ater
ial d
e ba
ixo
cust
o;-
não
exig
e es
trut
uras
adi
-ci
onai
s co
mo
bast
idor
es,
pode
ndo
ser
fixa
do
por
mei
o de
cor
das;
- ade
quad
o pa
ra a
mbi
ente
s lê
ntic
os,
com
ou
sem
bat
i-m
ento
de
onda
s e
lóti
cos
em
dife
rent
es
níve
is
de
corr
ente
za;
- ad
equa
do
para
lo
cais
pr
ofun
dos,
com
o re
pres
as;
- fá
cil
obse
rvaç
ão d
as d
is-
tint
as f
ases
de
vida
(be
ntô-
nica
s),
níve
is d
e co
loni
za-
ção,
e a
glom
erad
os;
- ad
equa
do
para
el
evad
a pr
ofun
dida
de.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- di
fíci
l m
anus
eio
no m
icro
scóp
io e
ster
eos-
cópi
co;
- a r
aspa
gem
dos
agl
omer
ados
par
a co
leta
de
indi
vídu
os a
dult
os p
ode
dani
fica
r in
diví
duos
re
crut
as e
jove
ns;
- os
sub
stra
tos
deve
m s
er l
avad
os c
om á
gua
dest
ilada
e p
ince
l par
a qu
e nã
o oc
orra
a p
er-
da d
e in
diví
duos
jove
ns e
rec
ruta
s.
Rio
s,
repr
esas
, la
gos,
la
goas
e la
guna
s.D
ensi
dade
(m
2 )L
imno
pern
a fo
tune
i M
acro
inve
rteb
rado
s as
soci
ados
Est
rutu
ras
trid
imen
sion
ais
de P
VC
(Sy
lves
ter
et a
l., 2
007)
- m
ater
ial d
e m
édio
cus
to;
- nã
o ex
ige
estr
utur
as a
di-
cion
ais
com
o ba
stid
ores
, po
dend
o se
r fi
xado
po
r m
eio
de c
orda
s;- a
dequ
ado
para
am
bien
tes
lênt
icos
, co
m o
u se
m b
ati-
men
to d
e on
das
e ló
tico
s em
di
fere
ntes
ní
veis
de
co
rren
teza
;-
adeq
uado
pa
ra
loca
is
prof
undo
s, c
omo
repr
esas
;-
adeq
uado
pa
ra
elev
ada
prof
undi
dade
.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- di
fíci
l m
anus
eio
no m
icro
scóp
io e
ster
eos-
cópi
co;
- as
dif
eren
tes
mal
has
sele
cion
am t
aman
hos
dos
orga
nism
os i
ncru
stan
tes
ou c
olon
izad
o-re
s, d
ific
ulta
ndo
a in
terp
reta
ção
dos
resu
lta-
dos.
Rio
s,
repr
esas
, la
gos,
la
goas
e la
guna
s.A
bund
ânci
aL
imno
pern
a fo
tune
i M
acro
inve
rteb
rado
s as
soci
ados

CAPÍTULO 13
Seção 3
179
Tab
ela
I.C
on
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
SUB
STR
AT
OS
AR
TIF
ICIA
IS
Est
rutu
ras
trid
imen
sion
ais
de m
adei
ra n
o fo
rmat
o de
“X
” (F
onte
s et
al.,
20
02;
Man
sur
et
al.,
2009
)
- m
ater
ial d
e m
édio
cus
to;
- nã
o ex
ige
estr
utur
as a
di-
cion
ais
com
o ba
stid
ores
, po
dend
o se
r fi
xado
po
r m
eio
de c
orda
s;- a
dequ
ado
para
am
bien
tes
lênt
icos
, co
m o
u se
m b
ati-
men
to d
e on
das
e ló
tico
s em
di
fere
ntes
ní
veis
de
co
rren
teza
; -
adeq
uado
pa
ra
loca
is
prof
undo
s, c
omo
repr
esas
;-
fáci
l ob
serv
ação
das
dis
-ti
ntas
fas
es d
e vi
da (
bent
ô-ni
cas)
, ní
veis
de
colo
niza
-çã
o, e
agl
omer
ados
;-
adeq
uado
pa
ra
elev
ada
prof
undi
dade
.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- di
fíci
l m
anus
eio
no m
icro
scóp
io e
ster
eos-
cópi
co;
- a r
aspa
gem
dos
agl
omer
ados
par
a co
leta
de
indi
vídu
os a
dult
os p
ode
dani
fica
r in
diví
duos
re
crut
as e
jove
ns;
- os
sub
stra
tos
deve
m s
er l
avad
os c
om á
gua
dest
ilada
e p
ince
l par
a qu
e nã
o oc
orra
a p
er-
da d
e in
diví
duos
jove
ns e
rec
ruta
s.
Rio
s,
repr
esas
, la
gos,
la
goas
e la
guna
s.D
ensi
dade
(m
2 )L
imno
pern
a fo
tune
i M
acro
inve
rteb
rado
s as
soci
ados
Est
rutu
ras
de
cerâ
mic
a –
tijo
lo
(San
tos
et a
l.,
2008
)
- fa
cilm
ente
en
cont
rado
no
mer
cado
;-
mat
eria
l de
baix
o cu
sto;
- fá
cil
man
usei
o no
m
i-cr
oscó
pio
este
reos
cópi
co;
- ad
equa
do p
ara
o m
oni-
tora
men
to
em
ambi
ente
s lê
ntic
os,
com
re
duzi
do
bati
men
to d
e on
da o
u ló
ti-
cos,
com
bai
xa c
orre
ntez
a;-
fáci
l ob
serv
ação
das
dis
-ti
ntas
fas
es d
e vi
da (
bent
ô-ni
cas)
, ní
veis
de
colo
niza
-çã
o, e
agl
omer
ados
;-
ótim
a co
nser
vaçã
o do
m
ater
ial b
ioló
gico
dur
ante
o
proc
essa
men
to e
m c
am-
po e
labo
rató
rio;
- po
de s
er f
acilm
ente
fix
a-do
por
cor
das.
- qu
ando
uti
lizad
as e
stan
tes
de f
erro
, o
cus-
to a
umen
ta,
além
de
difi
cult
ar a
fix
ação
no
ambi
ente
;-
não
é in
dica
do p
ara
ambi
ente
s ló
tico
s, c
om
elev
ada
corr
ente
za;
- m
ater
ial f
rági
l e q
uebr
adiç
o;-
viab
iliza
ele
vado
ass
enta
men
to,
devi
do à
s ca
ract
erís
tica
s te
xtur
ais
do m
ater
ial,
pode
n-do
não
rep
rese
ntar
os
níve
is p
opul
acio
nais
em
sub
stra
tos
natu
rais
(ex
ceto
alg
uns
tipo
s de
sub
stra
tos
roch
osos
);
- o m
ater
ial d
ever
ser
des
idra
tado
a 6
0o C e
m
forn
o de
Pas
teur
até
24
hora
s.
Rep
resa
s, l
agos
, la
goas
e
lagu
nas.
Den
sida
de (
m2 )
Lim
nope
rna
fotu
nei
Mac
roin
vert
ebra
dos
asso
ciad
os

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
180
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
SUB
STR
AT
OS
AR
TIF
ICIA
IS
Est
rutu
ras
de
cerâ
mic
a –
tijo
leta
(B
ergo
nci e
t al
., 20
09)
- fa
cilm
ente
en
cont
rado
no
mer
cado
;-
mat
eria
l de
baix
o cu
sto;
- fá
cil
man
usei
o no
m
i-cr
oscó
pio
este
reos
cópi
co;
- ad
equa
do p
ara
o m
oni-
tora
men
to
em
ambi
ente
s lê
ntic
os,
com
re
duzi
do
bati
men
to d
e on
da o
u ló
ti-
cos,
com
bai
xa c
orre
ntez
a;-
fáci
l ob
serv
ação
das
dis
-ti
ntas
fas
es d
e vi
da (
bent
ô-ni
cas)
, ní
veis
de
colo
niza
-çã
o, e
agl
omer
ados
;-
ótim
a co
nser
vaçã
o do
m
ater
ial b
ioló
gico
dur
ante
o
proc
essa
men
to e
m c
am-
po e
labo
rató
rio;
-
os b
asti
dore
s po
dem
ser
re
apro
veit
ados
.
- re
quer
est
rutu
ras
de f
erro
ade
quad
as p
ara
fixa
ção
(bas
tido
res)
de
elev
ado
cust
o;
- os
bas
tido
res
não
são
enco
ntra
dos
no m
er-
cado
, dev
endo
ser
man
ufat
urad
os;
- os
bas
tido
res
nece
ssit
am d
e m
anut
ençã
o de
vido
à c
orro
são;
- m
ater
ial (
bast
idor
es +
sub
stra
tos)
pes
ado;
- nã
o é
indi
cado
par
a am
bien
tes
lóti
cos,
com
el
evad
a co
rren
teza
;-
mat
eria
l frá
gil e
que
brad
iço;
- vi
abili
za e
leva
do a
ssen
tam
ento
, de
vido
às
cara
cter
ísti
cas
text
urai
s do
mat
eria
l, po
den-
do n
ão r
epre
sent
ar o
s ní
veis
pop
ulac
iona
is
em s
ubst
rato
s na
tura
is (
exce
to a
lgun
s ti
pos
de s
ubst
rato
s ro
chos
os);
- o
mat
eria
l dev
er s
er d
esid
rata
do a
60o C
em
fo
rno
de P
aste
ur a
té 2
4 ho
ras.
Rep
resa
s, l
agos
, la
goas
e
lagu
nas.
Den
sida
de (
m2 )
Lim
nope
rna
fotu
nei
Mac
roin
vert
ebra
dos
asso
ciad
os
Gar
rafa
PE
T –
com
pr
eenc
him
ento
de
mat
eria
l cel
ulós
ico
da b
ucha
de
Luff
a cy
lindr
ica
(Vol
kmer
-Rib
eiro
et
al.,
200
4)
- m
ater
ial d
e ba
ixo
cust
o;-
adeq
uado
par
a am
bien
tes
lóti
cos
com
bai
xa p
rodu
-çã
o pe
rifí
tica
, ve
loci
dade
re
duzi
da e
teo
r de
sól
idos
se
dim
entá
veis
red
uzid
os;
- nã
o ex
ige
estr
utur
as a
di-
cion
ais
com
o ba
stid
ores
, po
dend
o se
r fi
xado
po
r co
rdas
;-
o m
ater
ial
cole
tado
é
lava
do
em
pene
ira
sobr
e ág
ua c
orre
nte
para
pos
te-
rior
aná
lise
sob
mic
rosc
ó-pi
o es
tere
oscó
pico
.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- fo
rmaç
ão d
e bi
ofilm
es (
alga
s e
prot
ozoá
-ri
os) o
u ac
umul
ação
de
sedi
men
tos
na s
uper
-fí
cie
exte
rna
da g
arra
fa;
- ac
umul
ação
de
sedi
men
to in
tern
amen
te n
a te
la;
- a
colo
niza
ção
dos
dem
ais
mac
roin
vert
ebra
-do
s é
mai
s fa
vore
cida
que
a d
os m
olus
cos
inva
sore
s;-
rom
pim
ento
das
gar
rafa
s, e
m l
ocai
s co
m
bati
men
to d
e on
das,
ou
com
ele
vada
cor
ren-
teza
;-
o cr
esci
men
to p
opul
acio
nal
no s
ubst
rato
é
pouc
o re
pres
enta
tivo
dev
ido
aos
inte
rfer
en-
tes
cita
dos;
- nã
o é
adeq
uado
par
a a
obse
rvaç
ão d
e ag
lo-
mer
ados
e d
os n
ívei
s de
col
oniz
ação
;- n
ão r
epre
sent
am o
s nív
eis p
opul
acio
nais
em
su
bstr
atos
na
tura
is,
difi
cult
ando
qu
alqu
er
tipo
de
com
para
ção.
Rio
s,
riac
hos,
aç
udes
, re
pres
as, l
agos
, lag
oas
e la
guna
s.
Abu
ndân
cia
Lim
nope
rna
fotu
nei
Mac
roin
vert
ebra
dos
asso
ciad
os

CAPÍTULO 13
Seção 3
181
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
SUB
STR
AT
OS
AR
TIF
ICIA
IS
Gar
rafa
PE
T –
com
pr
eenc
him
ento
po
r fo
lhas
de
tabo
a (T
ypha
do
min
guen
sis
Pers
.) (
Que
iroz
et
al.,
2007
)
- m
ater
ial d
e ba
ixo
cust
o;-
adeq
uado
par
a am
bien
tes
lóti
cos
com
bai
xa p
rodu
-çã
o pe
rifí
tica
, ve
loci
dade
re
duzi
da e
teo
r de
sól
idos
se
dim
entá
veis
red
uzid
os;
- nã
o ex
ige
estr
utur
as a
di-
cion
ais
com
o ba
stid
ores
, po
dend
o se
r fi
xado
po
r co
rdas
;-
o m
ater
ial
cole
tado
é
lava
do
em
pene
ira
sobr
e ág
ua c
orre
nte
para
pos
te-
rior
aná
lise
sob
mic
rosc
ó-pi
o es
tere
oscó
pico
.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- fo
rmaç
ão d
e bi
ofilm
es (
alga
s e
prot
ozoá
-ri
os) o
u ac
umul
ação
de
sedi
men
tos
na s
uper
-fí
cie
exte
rna
da g
arra
fa;
- ac
umul
ação
de
sedi
men
to in
tern
amen
te n
a te
la;
- a
colo
niza
ção
dos
dem
ais
mac
roin
vert
ebra
-do
s é
mai
s fa
vore
cida
que
a d
os m
olus
cos
inva
sore
s;-
rom
pim
ento
das
gar
rafa
s, e
m l
ocai
s co
m
bati
men
to d
e on
das,
ou
com
ele
vada
cor
ren-
teza
;-
o cr
esci
men
to p
opul
acio
nal
no s
ubst
rato
é
pouc
o re
pres
enta
tivo
dev
ido
aos
inte
rfer
en-
tes
cita
dos;
- nã
o é
adeq
uado
par
a a
obse
rvaç
ão d
e ag
lo-
mer
ados
e d
os n
ívei
s de
col
oniz
ação
;- n
ão r
epre
sent
am o
s nív
eis p
opul
acio
nais
em
su
bstr
atos
na
tura
is,
difi
cult
ando
qu
alqu
er
tipo
de
com
para
ção.
Rio
s,
riac
hos,
aç
udes
, re
pres
as, l
agos
, lag
oas
e la
guna
s.
Abu
ndân
cia
Lim
nope
rna
fotu
nei
Mac
roin
vert
ebra
dos
asso
ciad
os
Gar
rafa
PE
T –
co
m t
ela
de n
ylon
(P
erei
ra e
t al
., 20
10)
- m
ater
ial d
e ba
ixo
cust
o;-
adeq
uado
par
a am
bien
tes
lóti
cos
com
bai
xa p
rodu
-çã
o pe
rifí
tica
, ve
loci
dade
re
duzi
da e
teo
r de
sól
idos
se
dim
entá
veis
red
uzid
os;
- nã
o ex
ige
estr
utur
as a
di-
cion
ais
com
o ba
stid
ores
, po
dend
o se
r fi
xado
po
r co
rdas
;-
fáci
l m
anus
eio
das
tela
s so
b o
mic
rosc
ópio
est
ere-
oscó
pico
.
- nã
o é
com
erci
aliz
ado,
dev
endo
ser
man
u-fa
tura
do;
- fo
rmaç
ão d
e bi
ofilm
es (
alga
s e
prot
ozoá
-ri
os) o
u ac
umul
ação
de
sedi
men
tos
na s
uper
-fí
cie
exte
rna
da g
arra
fa;
- ac
umul
ação
de
sedi
men
to in
tern
amen
te n
a te
la;
- ro
mpi
men
to d
as g
arra
fas,
em
loc
ais
com
ba
tim
ento
de
onda
s, o
u co
m e
leva
da c
orre
n-te
za;
- o
cres
cim
ento
pop
ulac
iona
l no
sub
stra
to é
po
uco
repr
esen
tati
vo d
evid
o ao
s in
terf
eren
-te
s ci
tado
s;-
não
é ad
equa
do p
ara
a ob
serv
ação
de
aglo
-m
erad
os e
dos
nív
eis
de c
olon
izaç
ão;
- não
rep
rese
ntam
os n
ívei
s pop
ulac
iona
is e
m
subs
trat
os
natu
rais
, di
ficu
ltan
do
qual
quer
ti
po d
e co
mpa
raçã
o.
Rio
s,
riac
hos,
aç
udes
, re
pres
as, l
agos
, lag
oas
e la
guna
s.
Abu
ndân
cia
Lim
nope
rna
fotu
nei
Mac
roin
vert
ebra
dos
asso
ciad
os

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
182
Tab
ela
I.
Con
tinua...
Mét
odo
de
amos
trag
emV
anta
gens
Des
vant
agen
sE
coss
iste
ma/
Hab
itat
/ M
icro
habi
tat
Exp
ress
ão d
e R
esul
tado
sE
spéc
ie a
ser
mon
itor
ada
SUB
STR
AT
OS
NA
TU
RA
IS
Raí
zes
e ta
los
de
água
pé
- o
mét
odo
pode
ser
apl
i-ca
do e
ada
ptad
o a
outr
as
mac
rófi
tas;
-per
mit
e am
ostr
ar c
om f
a-ci
lidad
e em
loc
ais
onde
os
rios
são
mui
to e
ncai
xado
s,
prof
undo
s e
de f
undo
ro-
chos
o, s
endo
invi
ável
a u
ti-
lizaç
ão d
e am
ostr
ador
es;
- a
faun
a qu
e oc
orre
nes
te
subs
trat
o é
mai
s ri
ca q
ue
em m
uito
s su
bstr
atos
art
i-fi
ciai
s.
- em
mui
tos
loca
is n
ão s
ão v
erif
icad
os e
xem
-pl
ares
do
agua
pé;
- nu
m p
rogr
ama
de m
onit
oram
ento
não
são
en
cont
rada
s m
uita
s ve
zes
a m
esm
a es
péci
e de
mac
rófi
ta e
m t
odos
os
trec
hos
e es
taçõ
es
de c
olet
a de
um
a ba
cia
hidr
ográ
fica
, o
que
difi
cult
a a
aval
iaçã
o in
tegr
ada
de r
esul
tado
s;-
a fa
una
que
ocor
re n
este
sub
stra
to é
dis
tin-
ta d
aque
la v
erif
icad
a no
sub
stra
to d
e fu
ndo,
se
ndo
cons
ider
ada
uma
amos
trag
em c
ompl
e-m
enta
r e
não
alte
rnat
iva.
Rio
s,
riac
hos,
aç
udes
, re
pres
as, l
agos
, lag
oas
e la
guna
s.
Abu
ndân
cia
ou
dens
idad
e.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
usPh
ysa
acut
aH
elis
oma
dury
iH
elis
oma
triv
olvi
s
Roc
has
- am
ostr
agem
fác
il e
rápi
-da
;- d
ifer
ente
s ta
man
hos
de r
ocha
são
enc
ontr
a-do
s no
s di
stin
tos
trec
hos
de u
ma
baci
a.-
a ár
ea d
as r
ocha
s é
de d
ifíc
il qu
anti
fica
ção.
Rio
s,
riac
hos,
aç
udes
, re
pres
as, l
agos
, lag
oas
e la
guna
s.
Abu
ndân
cia
ou
dens
idad
e.L
imno
pern
a fo
tune
iM
elan
oide
s tu
berc
ulat
us
2.3. Anestesia
Muitos organismos devem ser anestesia-dos antes de conservados, pois caso não sejam, retraem o corpo, inviabilizando a posterior de-terminação específica ou, em alguns casos, até em nível de gênero ou família. Alguns procedi-mentos são citados a seguir: para moluscos, co-locar os indivíduos em um recipiente com água e mentol, refrigerar em geladeira, procurando não agitar o recipiente ou movê-lo de lugar para que os organismos possam relaxar e atingir o estágio de letargia; para hidroides, o anestésico mais usado é o mentol, sendo também usado o cloreto e o sulfato de magnésio, dentre outros; nos ectoprotozoa, usar uma solução de 1% de stovaine ou eucaine, que é adicionado gota a gota a intervalos de 10 a 15 minutos entre as primeiras duas doses e 5 minutos para as su-cessivas ou, como anestésico alternativo, usar mentol; para os hirudíneos, usar uma solução de álcool a 5-10%, suco de limão ou água ga-sosa; para os crustáceos, usar uma solução de formol a 1-2% (Mugnai et al., 2010).
2.4. Conservação das amostras
Apesar no uso consagrado de álcool 70% para a conservação de amostras biológicas, no caso de amostras de sedimento para avaliação da fauna bentônica, recomenda-se a utilização de álcool 80-85% uma vez que as amostras de sedimento apresentam elevado teor de umidade e matéria orgânica. O uso de formol é extrema-mente inapropriado já que causa a descalcifica-ção das conchas dos moluscos, inviabilizando assim a sua identificação.
2.5. Processamento de amostras em laboratório
O processamento das amostras ocorre no laboratório. Para a triagem de uma amostra, o sedimento é retirado do pote plástico e co-locado por meio de uma colher de plástico so-bre uma placa de acrílico (Fig. 32) escavada no formato de um U, que é levada ao estereomi-croscópio para triagem dos organismos. Se há uma grande quantidade de amostra, coloca-se

CAPÍTULO 13
Seção 3
183
apenas uma parte do sedimento na placa, sen-do o restante adicionado à medida que a tria-gem do material anterior tenha sido finalizada. Na placa escavada e com o auxílio de pinças e/ ou agulha de aço inoxidável, analisa-se o sedi-mento. Os organismos encontrados são pinça-dos usando pinça de aço maleável (Fig. 33) e colocados em uma placa de acrílico com poços escavados e preenchidos por álcool 70%. Pos-teriormente, esses organismos, se forem peque-nos, são transferidos para tubos plásticos com tampa – tipo Eppendorfs – e preenchidos com álcool 70%; se forem maiores, são transferidos para potes de acrílico que também são preen-chidos por álcool 70%. São utilizadas pinças maleáveis, pincel e/ou pera de plástico para a transferência dos organismos para os recipien-tes anteriormente mencionados. Tanto o tubo como o pote, deve conter uma etiqueta, feita de papel vegetal e escrita a caneta nanquim, com informações sobre a amostra (data, cole-tor e local de coleta). Após a identificação do organismo, é adicionada outra etiqueta ao tubo ou pote com o nome do táxon. Como o pote de acrílico e o tubo plástico secam em contato com álcool 70%, abrindo pequenas fissuras e assim deixando vazar seu conteúdo, estes são acondicionados em frascos de vidro (Fig. 28) com tampa metálica preenchidos por álcool 70%. Assim, mesmo que apareçam fissuras, o organismo dentro do tubo ou frasco não se de-teriorará com a evaporação do álcool.
2.6. Identificação dos táxons
Ser bentólogo não é uma tarefa fácil, pois exige conhecimento sobre a taxonomia de vários grupos de invertebrados. Assim como uma única pessoa não consegue se especializar em todos os grupos taxonômicos que estão pre-sentes na comunidade de macroinvertebrados, por outro lado nem sempre se têm à disposição os especialistas dos grupos taxonômicos que se necessita. Além disso, um programa de mo-nitoramento bem executado e planejado pode gerar um número de amostras muito elevado, dificultando ainda mais o processamento e de-terminação, pela falta de profissionais treina-dos.
Figura 32. Placas de acrílico escavada em formato de U (es-querda) e com oito poços escavados (direita). Foto: J.O. Ar-ruda.
Figura 33. Materiais utilizados na triagem das amostras. Da esquerda para a direita: pinças de diferentes tamanhos, agu-lha, pincel, pera de plástico e colher de plástico. Foto: J.O. Arruda.
Figura 34. Frascos de vidro preenchido por álcool 70% e con-tendo os tubos plásticos (esquerda) e potes de acrílicos (direi-ta) com os organismos identificados das amostras coletadas. Foto: J.O. Arruda.

Pereira et al.
Como monitorar moluscos límnicos invasores bentônicos e macroinvertebrados associados?
184
No caso do monitoramento de espécies invasoras, a correta determinação específica das mesmas é primordial. Não só dos molus-cos límnicos, alvo de discussão neste livro, mas também das demais espécies de macroinverte-brados invasores que ocupem os mesmos ni-chos. Num programa de monitoramento onde se objetiva avaliar o impacto dos moluscos e outros invertebrados invasores sobre a fauna
bentônica de forma geral, os demais macroin-vertebrados devem ser identificados, no míni-mo, em nível de família. Quando espécies ame-açadas e endêmicas estão presentes na bacia monitorada, o monitoramento deve considerar a determinação específica das mesmas, pois a presença de espécies invasoras pode acarretar no declínio das populações de espécies amea-çadas e endêmicas.

Seção
4Aspectos populacionais de moluscos invasores e
relações com variáveis ambientais


187
Com uma área aproximada de 10.227 km2 (265 km de extensão por até 60 km de largura), a Lagoa dos Patos se encontra orien-tada no sentido NE-SO na planície costeira do extremo sul do Brasil, sendo considerada a maior laguna estrangulada do mundo (Kjerfve, 1986). A conexão deste corpo lagunar com o oceano atlântico ocorre em sua porção mais ao sul através de um único e estreito canal (0,5 a 3 km de largura), onde águas de origem marinha se misturam com aquelas mais doces originan-do uma região estuarina com cerca de 971 km2
(Asmus, 1997). A Lagoa Mirim, por sua vez, apresenta uma área aproximada de 3.749 km2 (185 km de extensão por 20 km de largura, em média) e uma extensa bacia hidrográfica trans-fronteiriça (62.250 km2 de área total), onde prevalece o regime de águas compartilhadas entre o Brasil (76%) e o Uruguai (FAO, 1972). Conectando a Lagoa dos Patos (na altura da cidade de Pelotas) ao extremo nordeste da Lagoa Mirim, encontra-se o Canal São Gon-çalo, um canal de água doce com 61 Km de
extensão que é influenciado pelo deságue dos rios tributários Piratini, Tigre e Arroio Pelotas. Além destes rios, o Canal também recebe os fluxos dos corpos lagunares (Patos e Mirim), que dependem do desnível entre ambas somada à ação dos ventos (Kotzian & Marques, 2004). O fluxo de enchente ou vazante neste canal é controlado por uma barragem eclusa que im-pede a entrada de água salgada proveniente da região estuarina da Lagoa dos Patos ao interior da Lagoa Mirim, em períodos de forte estia-gem (Burns et al., 2006a). As características hidrográficas na La-goa dos Patos, em especial em sua região es-tuarina, são fortemente influenciadas pelos ciclos hidrológicos (pluvial e fluvial) na bacia de drenagem do Sistema Patos – Mirim, onde aproximadamente 85 % do volume de água é proveniente dos rios Guaíba, Camaquã e Canal São Gonçalo, que deságuam nas regiões nor-te, central e sul desta laguna, respectivamente (Garcia, 1997). Estudos pioneiros sobre a presença do
CAPÍTULO 14
Limnoperna fortunei na Bacia da Lagoa dos Patos
e Lagoa Mirim
Leonir André CollingRaphael Mathias PinottiCarlos Emílio Bemvenuti

Colling et al.
Limnoperna fortunei na Bacia da Lagoa dos Patos e Lagoa Mirim
188
bivalve exótico de água doce Limnoperna for-tunei (Dunker, 1857) no Sistema Patos – Mirim evidenciam o início de sua invasão e distribui-ção a partir do Lago Guaíba em 1998 (Mansur et al., 1999). A partir destes primeiros registros de ocorrência na porção norte (região límnica) da laguna, a expansão da distribuição desta es-pécie exótica foi observada mais ao sul (região pré-límnica), tendo sido registrada a presença de L. fortunei em Arambaré no ano 2000 e em São Lourenço do Sul em 2001 (Mansur et al., 2003). Com a progressiva colonização do bival-ve em direção ao sul da Lagoa dos Patos, exem-plares da espécie foram registrados no Arroio Pelotas e no Canal São Gonçalo, podendo as-sim ter dado início à expansão de sua distri-buição no segundo maior corpo lagunar cos-teiro do Rio Grande do Sul e Uruguai, a Lagoa Mirim (Brugnoli et al., 2005; Capítoli et al., 2008). O Arroio Pelotas foi considerado um local-chave na dispersão da espécie, uma vez que nesta área foram identificados importantes aspectos ecológicos que facilitariam sua persis-tência local e aumento de sua distribuição atra-vés de potenciais eventos reprodutivos (Capí-toli & Bemvenuti, 2004). Dentre os principais fatores observados estão a presença de diversos tipos substrato para sua fixação (marinas, tra-piches e embarcações) e o predomínio de água doce na área, impedindo a ocorrência de even-tos de mortalidade devido à elevada salinidade, assim como a registrada na porção estuarina da Lagoa dos Patos em 2004, durante um período de estiagem. O Canal São Gonçalo também se mos-trou um importante vetor na distribuição do bivalve L. fortunei para a Lagoa Mirim. A construção da barragem eclusa, que entrou em funcionamento em 1977, dividiu este Canal em dois setores característicos, os quais apresen-tam condições límnicas à montante (em dire-ção à Lagoa Mirim) e predomínio de condições estuarinas à jusante (em direção à Lagoa dos Patos), ambos importantes no processo de ex-pansão geográfica desta espécie. Quando efe-tuado o primeiro registro da espécie no setor estuarino do Canal São Gonçalo (2002), a presença do bivalve ainda não havia sido regis-trada no setor límnico do Canal, na Mirim e
nem nos seus tributários (Capítoli et al., 2008). Entretanto, um prolongado período de predo-mínio de águas doces entre a eclusa e a Lagoa dos Patos (durante os anos de 2002-2003), alia-do à permanente ocorrência de condições lím-à permanente ocorrência de condições lím- permanente ocorrência de condições lím-nicas no São Gonçalo (à montante da eclusa), provavelmente possibilitaram a ocorrência – e consequente expansão – do bivalve para o inte-rior da Lagoa Mirim, via Canal São Gonçalo, como registrado no ano de 2005 (Burns et al., 2006b). Pescadores do município de Arroio Grande, moradores do distrito de Santa Isabel (porção sul do Canal São Gonçalo) relatam o deslocamento de embarcações artesanais até a Lagoa dos Patos, apontando esta laguna como a possível origem do bivalve neste setor da La-goa Mirim (Pieve et al., 2009), assim como dis-cutido em estudos anteriores (Capítoli & Be-mvenuti, 2004; Brugnoli et al., 2005). Ainda, a conexão de corpos de água entre a Lagoa Mi-rim e a Reserva Ecológica do Taim (localizada na porção sudeste desta lagoa), provavelmente, favoreceu a expansão na distribuição de L. for-tunei nas áreas de banhado e lagos interiores, como na Lagoa Nicola (Burns et al., 2006a). No Uruguai, a presença deste bivalve in-vasor havia sido reportada para quatro das seis maiores Bacias Hidrográficas do país, as bacias dos rios Prata, Santa Lucía, Negro e Uruguai, sem que se houvesse detectado sua presença nas bacias do Atlântico e Laguna Merín (Brug-noli et al., 2005). No entanto, os primeiros re-gistros de L. fortunei nesta última bacia (setor uruguaio da Lagoa Mirim) foram incluídos no trabalho de revisão de Langone (2005), repor-tando sua presença nas localidades de Balne-ario Merín e na usina de captação de água de Río Branco, às margens do Rio Jaguarão. Pos-teriormente, sua presença também seria regis-trada na desembocadura do Rio Tacuarí, um dos rios uruguaios afluentes da Lagoa Mirim (Brugnoli et al., 2009). São recorrentes os relatos de prejuízos a atividades humanas proporcionados por bival-ves invasores. Dentre os principais registros são citados o bloqueio de sistemas de captação de água, entupimento de filtros e incrustação de superfícies submersas (Mackie, 1991; Smit et al., 1992; Schloesser, 1995; Darrigran, 2002; Mansur et al., 2003). No estuário da Lagoa dos

CAPÍTULO 14
Seção 4
189
Patos, uma região com atividades econômicas voltadas ao ambiente costeiro e numerosa frota pesqueira artesanal, os problemas causados por sua incrustação são relatados em comunidades de pescadores, onde os organismos são encon-trados fixos em calões de pesca, trapiches e em-barcações, além dos registros de macrofouling em tubulações de resfriamento de indústrias e refinarias nesta laguna (Capítoli & Bemvenuti, 2004; Capítoli et al., 2008). Eventos em escala global como o ENSO – El Niño, que ocorrem em intervalos interanu-ais pouco previsíveis e ocasionam um aumento de precipitação no sul do Brasil (Ropelewski & Halpert, 1987; Grimm et al., 2000), são asso-ciados a períodos prolongados de predomínio de água doce no estuário da Lagoa dos Patos. Por outro lado, sob influência de fenômenos ENSO – La Niña, a região sul do Brasil e o Uruguai sofrem anomalias negativas de pre-cipitação – seca (Grimm et al., 2000), ocasio-nando menores descargas de água doce nesta laguna. Por ser uma espécie de água doce, L. fortunei não tolera ambientes de águas salobras
ou marinhas, sendo assim estabelecidos limites fisiológicos que inibem sua presença e dispersão na região estuarina da Lagoa dos Patos. Neste sentido, a baixa tolerância da es-pécie à salinização oferece um fator contro-lador natural à sua distribuição no ambiente estuarino, em diferentes cenários hidrológicos (Capítoli et al., 2008). O evento com influência mais evidente sobre a biota estuarina foi o El Niño 2002-2003, sendo observado um prolon-gado cenário de água doce e expansão de L. fortunei na região (Fig. 1). Após a diminuição da vazão fluvial na bacia de drenagem, houve um predomínio de águas salobras e salinida-des elevadas entre os anos 2004-2006, ocasio-nando a mortalidade daquelas populações do bivalve amplamente distribuídas e impedindo novas colonizações nos anos seguintes (Fig. 1). Outro evento El Niño ocorrido no verão 2009-2010 provocou o predomínio de condi-ções límnicas na região estuarina da laguna, sendo novamente registrada uma colonização do bivalve (Fig. 1), observada através de experi-mentos de incrustação nos quais se utilizou pla-
Figura 1. Valores diários (linha cinza) e média móvel trimestral (linha preta) da salinidade na região estuarina da Lagoa dos Patos, no período entre janeiro/2001 e setembro/2011 (Banco de Dados PELD – Programas Ecológicos de Longa Duração, Site 8). SOI = Southern Index Oscillation (Bom, 2011).

Colling et al.
Limnoperna fortunei na Bacia da Lagoa dos Patos e Lagoa Mirim
190
cas de madeira como substrato para sua fixação (Santos, C.L. 2011). A partir da elevação dos valores de salinidade, novas colonizações de L. fortunei não foram encontradas, evidenciando a variação deste parâmetro ambiental como o principal fator regulador da sobrevivência e distribuição das populações desta espécie no extremo sul da Lagoa dos Patos.
Uma síntese da distribuição de L. fortu-nei na Bacia Patos-Mirim pode ser visualizada na Figura 2. A ampla distribuição de popula-ções deste bivalve encontra-se atualmente es-tabelecida em regiões límnicas, tanto lênticas como lóticas, não encontrando barreiras fisio-lógicas para sua sobrevivência e reprodução. Situação particular ocorre na região estuarina
Figura 2. Áreas de ocorrência do bivalve invasor Limnoperna fortunei, até o presente: Lagoa dos Patos (LP) e sua região estuari-na (ELP); Canal São Gonçalo e seus tributários; e Lagoa Mirim (LM), tanto no estado do Rio Grande do Sul, quanto em território uruguaio (UY).

CAPÍTULO 14
Seção 4
191
da Lagoa dos Patos, onde a ocupação temporá-ria de L. fortunei é dependente dos regimes de adoçamento ou salinização, associados a dife-rentes eventos ENSO. Neste ambiente estuarino, populações permanentes podem ser observadas por toda a região sul da laguna quando em regime de condições límnicas (BS: baixa salinidade – El Niño; Figura 2), ficando restritas a ambientes
de água doce quando a região estuarina se en-contra sob alta salinidade (AS: alta salinidade – La Niña; Fig. 2). A correlação entre eventos de colonização deste bivalve relacionados a pe-ríodos de água doce fornece subsídios para o es-tabelecimento de relações positivas entre estes dois processos, com provável aumento da distri-buição de L. fortunei pelo estuário da Lagoa dos Patos durante os próximos eventos El Niño.


193
No Rio Grande do Sul são delimitadas três grandes regiões hidrográficas, reconheci-das pelas direções principais de escoamento dos rios. Uma delas é a Região Hidrográfica da Bacia do Lago Guaíba, tendo como principal canal de escoamento o Rio Jacuí. Esta região apresenta uma área de 84.763,5 km2, sendo a mais densamente povoada do estado e de maior concentração de atividades comerciais e indus-triais. Nesta região hidrográfica são reconheci-das oito bacias, identificadas por um grande rio ou lago formador, como é o caso do Rio Jacuí, que forma duas bacias, uma denominada de ba-cia do alto Jacuí (porção inicial) e outra de ba-cia do Pardo-Baixo Jacuí (porção final). Den-tre os rios da Bacia do Cai, Sinos e Gravataí, que juntamente com o Jacuí desembocam no Lago Guaíba e assim formam o Delta do Jacuí (área de 47,18 km2), o Rio Jacuí responde por 84,6% do volume total de agua que desemboca no Lago Guaíba (Menegat & Kirchhein, 1998). Na bacia do Guaíba até o início da década de 1990, conhecia-se apenas a invasão de bivalves
asiáticos do gênero Corbicula, que foram intro-duzidos em meados da década de 1970 (Veite-nheimer-Mendes, 1981). No início de 1999 foi constatada a presença do mexilhão-dourado nas margens do Lago Guaíba (Mansur et al., 1999). Como os corbiculídeos, esta também é uma espécie de bivalve invasora e é oriunda do sudeste asiático. De acordo com Mansur et al. (2003), a introdução do mexilhão-dourado na bacia do Lago Guaíba aconteceu de maneira não intencional, através da água de lastro de navios, provavelmente, oriundos da Argenti-na, onde a espécie foi registrada pela primeira vez, mais especificamente, no Rio da Prata, em 1991, próximo da cidade de La Plata (Pastori-no et al., 1993). L. fortunei distingue-se dos demais bi-valves nativos do sul do Brasil (Unionoida e Ve-neroida) que vivem enterrados nos substratos arenosos, a exceção das espécies bissadas de Eupera Bourguinat, 1854 que não formam ma-croaglomerações, por apresentar a produção de fios de bisso, o qual possibilita sua fixação
CAPÍTULO 15
Limnoperna fortunei na bacia hidrográfica do baixo
Rio Jacuí e Lago Guaíba, Rio Grande do Sul
Paulo Eduardo Aydos BergonciCintia Pinheiro dos Santos
Marinei Vilar NehrkeDaniel Pereira
Maria Cristina Dreher Mansur

Bergonci et al.
Limnoperna fortunei na bacia hidrográfica do baixo Rio Jacuí e Lago Guaíba, Rio Grande do Sul
194
e consequente formação de macroaglomerados compactados sobre substratos duros (Mansur & Pereira, 2006). Esta característica dificulta o monitoramento da espécie em ambiente na-tural, em função da variabilidade de substratos ao qual ela adere e da inaplicabilidade de méto-do tradicionais de amostragem (dragas, cores, quadrats e puçás, entre outros). Desta forma, se faz necessária a utilização de métodos como os substratos artificiais, uma vez que estes pa-dronizam a amostragem, reduzem a variabili-dade e o tempo de processamento das amostras (Gibbons et al., 1993). Sob esta perspectiva, diversos tipos de substratos artificiais são utilizados para moni-torar populações de L. fortunei, como estrutu-ras tridimensionais de abestos (Morton, 1977), adaptações em PVC (Boltovskoy & Cataldo, 1999), em madeira no formato de “X” (Fontes et al., 2002; Mansur et al., 2009), garrafas PET (Faria et al., 2006; Pereira et al., 2010) e subs-tratos cerâmicos (Mansur et al., 2003; Santos, 2004; Terra et al., 2007; Santos et al., 2008; Bergonci et al., 2009). Os métodos empregados pela equipe coordenada pela Dra. Maria Cristina Dreher Mansur constam em Mansur et al. (2003), San-
tos (2004), Santos et al. (2008), Bergonci et al. (2009), Mansur et al. (2009) e Pereira et al. (2010). Serão apresentados alguns dos resulta-dos obtidos no monitoramento com substrato artificial de madeira (Mansur et al., 2009). A área do monitoramento (Fig. 1) situa-se no del-ta do Rio Jacuí, na unidade fisiográfica da De-pressão Central do Estado do Rio Grande do Sul, na região metropolitana de Porto Alegre. O Rio Jacuí deságua no Lago Guaíba por meio de um delta formado por uma série de distri-butários, dentre os quais se destaca o Canal do Jacuí (profundidade média de 8 m). No Cais do Porto, junto à área urbana de Porto Alegre, a paisagem marginal foi significativamente alte-rada por meio de um aterro. Atualmente, neste local se verificam margens íngremes (paredes concretadas) e águas profundas (5 m). As esta-ções de coleta situam-se no delta do Rio Jacuí: na margem direita do Canal do Jacuí (CJ) e na Ilha da Pintada e Cais do Porto (PO), no centro da cidade. Foram submersos doze conjuntos, com três réplicas, de um substrato artificial de ma-deira em forma de xis (Fig. 2) nas estações de coleta acima descritas. O período de submersão foi de outubro de 2006 a setembro de 2007.
Figura 1. Estações de coleta onde foram realizados os monitoramentos. CJ = canal do Rio Jacuí; PO = Cais do Porto. Mapa: R.A. Ramos (MCN/FZB-RS) em Pereira et al. (2010).
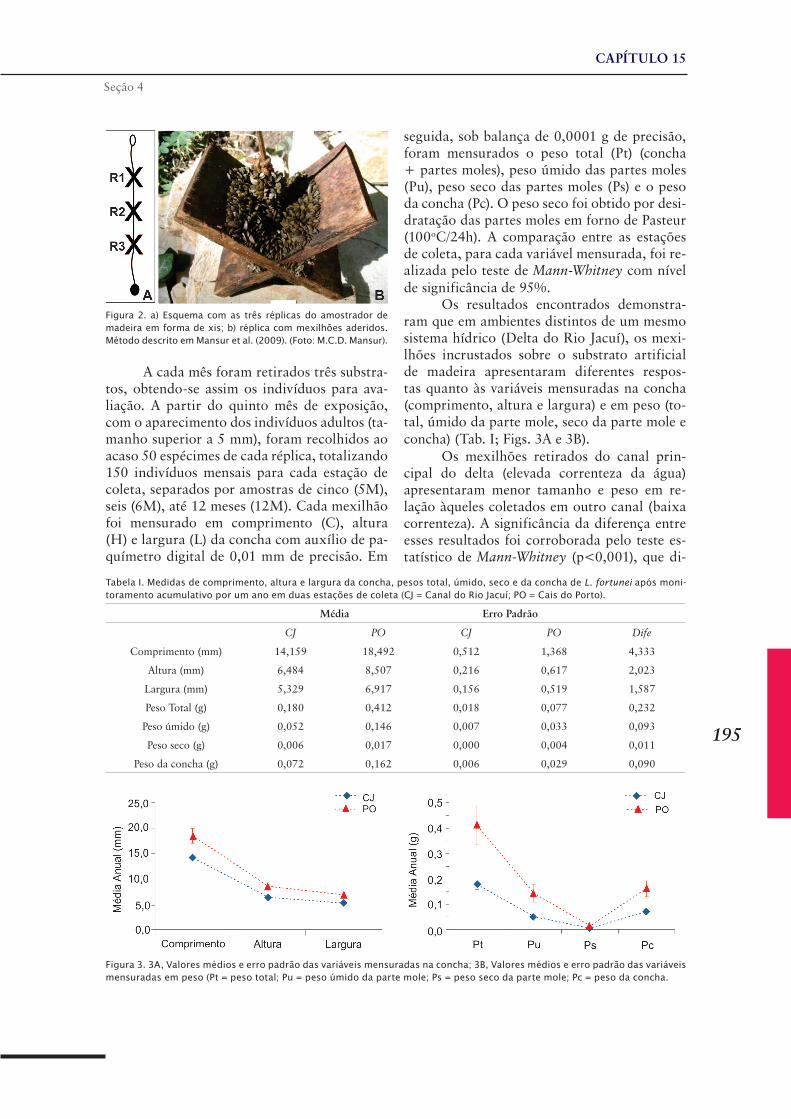
CAPÍTULO 15
Seção 4
195
A cada mês foram retirados três substra-tos, obtendo-se assim os indivíduos para ava-liação. A partir do quinto mês de exposição, com o aparecimento dos indivíduos adultos (ta-manho superior a 5 mm), foram recolhidos ao acaso 50 espécimes de cada réplica, totalizando 150 indivíduos mensais para cada estação de coleta, separados por amostras de cinco (5M), seis (6M), até 12 meses (12M). Cada mexilhão foi mensurado em comprimento (C), altura (H) e largura (L) da concha com auxílio de pa-químetro digital de 0,01 mm de precisão. Em
seguida, sob balança de 0,0001 g de precisão, foram mensurados o peso total (Pt) (concha + partes moles), peso úmido das partes moles (Pu), peso seco das partes moles (Ps) e o peso da concha (Pc). O peso seco foi obtido por desi-dratação das partes moles em forno de Pasteur (100oC/24h). A comparação entre as estações de coleta, para cada variável mensurada, foi re-alizada pelo teste de Mann-Whitney com nível de significância de 95%. Os resultados encontrados demonstra-ram que em ambientes distintos de um mesmo sistema hídrico (Delta do Rio Jacuí), os mexi-lhões incrustados sobre o substrato artificial de madeira apresentaram diferentes respos-tas quanto às variáveis mensuradas na concha (comprimento, altura e largura) e em peso (to-tal, úmido da parte mole, seco da parte mole e concha) (Tab. I; Figs. 3A e 3B). Os mexilhões retirados do canal prin-cipal do delta (elevada correnteza da água) apresentaram menor tamanho e peso em re-lação àqueles coletados em outro canal (baixa correnteza). A significância da diferença entre esses resultados foi corroborada pelo teste es-tatístico de Mann-Whitney (p<0,001), que di-
Figura 2. a) Esquema com as três réplicas do amostrador de madeira em forma de xis; b) réplica com mexilhões aderidos. Método descrito em Mansur et al. (2009). (Foto: M.C.D. Mansur).
Tabela I. Medidas de comprimento, altura e largura da concha, pesos total, úmido, seco e da concha de L. fortunei após moni-toramento acumulativo por um ano em duas estações de coleta (CJ = Canal do Rio Jacuí; PO = Cais do Porto).
Média Erro Padrão
CJ PO CJ PO Dife
Comprimento (mm) 14,159 18,492 0,512 1,368 4,333
Altura (mm) 6,484 8,507 0,216 0,617 2,023
Largura (mm) 5,329 6,917 0,156 0,519 1,587
Peso Total (g) 0,180 0,412 0,018 0,077 0,232
Peso úmido (g) 0,052 0,146 0,007 0,033 0,093
Peso seco (g) 0,006 0,017 0,000 0,004 0,011
Peso da concha (g) 0,072 0,162 0,006 0,029 0,090
Figura 3. 3A, Valores médios e erro padrão das variáveis mensuradas na concha; 3B, Valores médios e erro padrão das variáveis mensuradas em peso (Pt = peso total; Pu = peso úmido da parte mole; Ps = peso seco da parte mole; Pc = peso da concha.
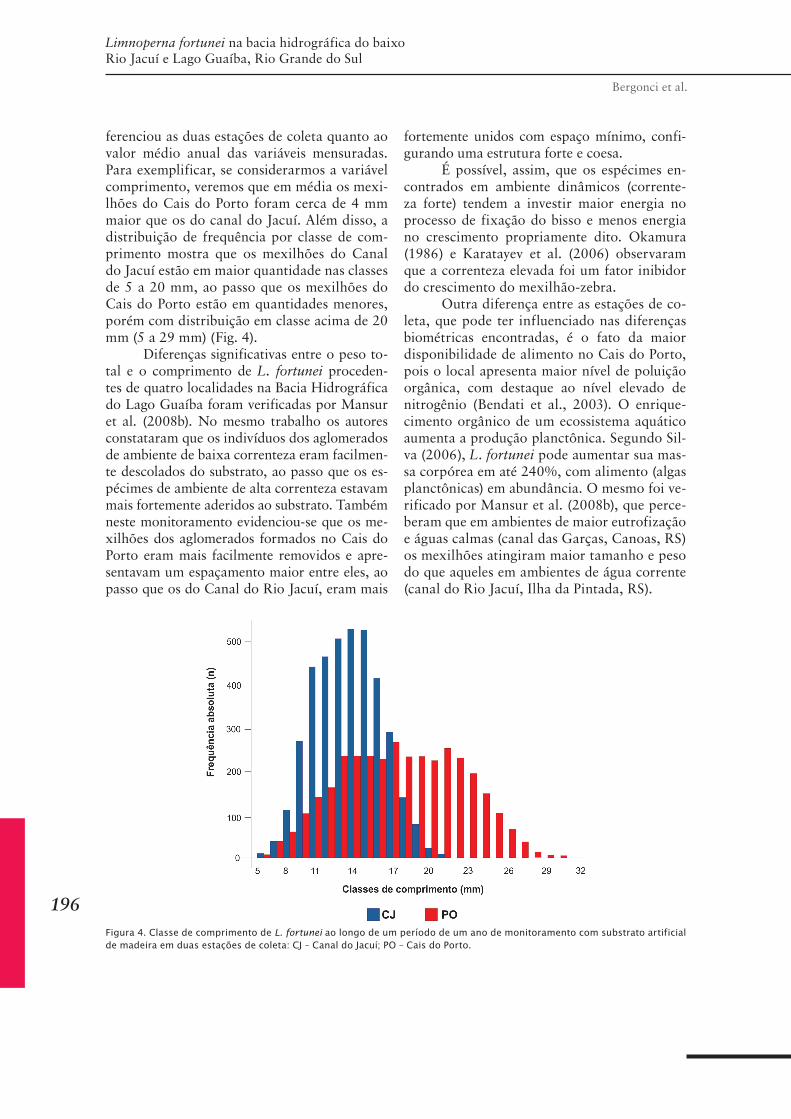
Bergonci et al.
Limnoperna fortunei na bacia hidrográfica do baixo Rio Jacuí e Lago Guaíba, Rio Grande do Sul
196
ferenciou as duas estações de coleta quanto ao valor médio anual das variáveis mensuradas. Para exemplificar, se considerarmos a variável comprimento, veremos que em média os mexi-lhões do Cais do Porto foram cerca de 4 mm maior que os do canal do Jacuí. Além disso, a distribuição de frequência por classe de com-primento mostra que os mexilhões do Canal do Jacuí estão em maior quantidade nas classes de 5 a 20 mm, ao passo que os mexilhões do Cais do Porto estão em quantidades menores, porém com distribuição em classe acima de 20 mm (5 a 29 mm) (Fig. 4). Diferenças significativas entre o peso to-tal e o comprimento de L. fortunei proceden-tes de quatro localidades na Bacia Hidrográfica do Lago Guaíba foram verificadas por Mansur et al. (2008b). No mesmo trabalho os autores constataram que os indivíduos dos aglomerados de ambiente de baixa correnteza eram facilmen-te descolados do substrato, ao passo que os es-pécimes de ambiente de alta correnteza estavam mais fortemente aderidos ao substrato. Também neste monitoramento evidenciou-se que os me-xilhões dos aglomerados formados no Cais do Porto eram mais facilmente removidos e apre-sentavam um espaçamento maior entre eles, ao passo que os do Canal do Rio Jacuí, eram mais
Figura 4. Classe de comprimento de L. fortunei ao longo de um período de um ano de monitoramento com substrato artificial de madeira em duas estações de coleta: CJ – Canal do Jacuí; PO – Cais do Porto.
fortemente unidos com espaço mínimo, confi-gurando uma estrutura forte e coesa. É possível, assim, que os espécimes en-contrados em ambiente dinâmicos (corrente-za forte) tendem a investir maior energia no processo de fixação do bisso e menos energia no crescimento propriamente dito. Okamura (1986) e Karatayev et al. (2006) observaram que a correnteza elevada foi um fator inibidor do crescimento do mexilhão-zebra. Outra diferença entre as estações de co-leta, que pode ter influenciado nas diferenças biométricas encontradas, é o fato da maior disponibilidade de alimento no Cais do Porto, pois o local apresenta maior nível de poluição orgânica, com destaque ao nível elevado de nitrogênio (Bendati et al., 2003). O enrique-cimento orgânico de um ecossistema aquático aumenta a produção planctônica. Segundo Sil-va (2006), L. fortunei pode aumentar sua mas-sa corpórea em até 240%, com alimento (algas planctônicas) em abundância. O mesmo foi ve-rificado por Mansur et al. (2008b), que perce-beram que em ambientes de maior eutrofização e águas calmas (canal das Garças, Canoas, RS) os mexilhões atingiram maior tamanho e peso do que aqueles em ambientes de água corrente (canal do Rio Jacuí, Ilha da Pintada, RS).

197
Introdução Informação sobre limite de tolerância da espécie a variáveis ambientais são essenciais para prever novas invasões e dimensionar os impactos que a espécie pode causar. Relatos de sobrevivência, crescimento e reprodução em condições estressantes revelam os limites de tolerância da espécie. Os estudos de Oli-veira e colaboradores na planície do Pantanal ampliaram alguns dos limites de tolerância do molusco invasor Limnoperna fortunei (Dunker, 1857). A interação rio-planície, numa das maiores áreas alagadas do mundo, provoca al-terações nas características da água e apresenta valores extremos de variáveis limnológicas que causam a mortalidade da espécie anualmente, representando um controle populacional natu-ral. É difícil apontar a variável mais importan-te como controle, porque potenciais estressores são variáveis no tempo e no espaço e tendem a coincidir ou sobrepor-se no tempo. Quanto mais áreas forem colonizadas por L. fortunei
mais informação teremos sobre seus limites ambientais, embora sua ampla tolerância a fa-tores ambientais torne difícil identificar quais variáveis são realmente importantes no seu es-tabelecimento num novo ambiente.
Área de ocorrência e densidades do mexilhão-dourado na Bacia do Alto Paraguai Na bacia do Alto Rio Paraguai, o primei-ro registro de L. fortunei foi em 1998, no Rio Paraguai (18º49’18”S e 57º39’13”W), próximo a Corumbá (MS). A área de ocorrência de L. fortunei é mostrada na Figura 1. Foram encon-tradas conchas entre 10 e 20 mm, indicando que a espécie já estava no ambiente há aproxi-madamente um ano. A área de ocorrência atual de L. fortunei no Pantanal está restrita ao canal principal do Rio Paraguai, lagoas marginais co-nectadas e nas partes inferiores dos tributários, como os rios Cuiabá, Miranda e Apa. O limite superior no Rio Paraguai não está bem estabe-
CAPÍTULO 16
Limnoperna fortunei na bacia do Rio Paraguai,
Mato Grosso do Sul
Marcia Divina de OliveiraDébora Fernandes Calheiros

Oliveira & Calheiros
Limnoperna fortunei na bacia do Rio Paraguai, Mato Grosso do Sul
198
lecido. Callil, C.T. (comunicação pessoal) en-controu exemplares do mexilhão-dourado no porto de Cáceres (norte do Mato Grosso), mas a espécie não se estabeleceu. Com relação aos tributários do Rio Para-guai, L. fortunei ocorre no Rio Miranda desde 2003, e avançou aproximadamente 100 km rio
acima depois do primeiro registro (Albuquer-que, S., comunicação pessoal). No Rio Apa a espécie ocorre desde 2000 sem registro exato da área de ocorrência, e na foz do Rio Cuiabá, 2008, também sem registro exato da área colo-nizada (Oliveira et al., 2011). A introdução de L. fortunei na bacia do
Figura 1. Localização da bacia do Alto Paraguai na América do Sul, drenagem da Bacia do Alto Paraguai e área de ocorrência de L. fortunei. Fonte: modificado de Oliveira et al. (2010a).

CAPÍTULO 16
Seção 4
199
Alto Rio Paraguai foi facilitada pelo tráfego de barcaças na hidrovia Paraguai-Paraná, as quais podem transportar os organismos no casco. A dispersão entre Rio Paraguai e afluentes tem sido mais lenta devido ao menor tráfego de em-barcações (Oliveira, M. et al., 2010a, 2011). Depois de 2 a 3 anos da presença de L. fortunei no Rio Paraguai, a densidade média de L. fortunei, medida em áreas com densa coloni-zação, foi de 1.772 ind./m3 para larvas, 88.853 juvenis/m2 e aproximadamente 35.000 adultos/m2. No Rio Miranda, as densidades hoje são semelhantes às do Rio Paraguai, com maior densidade nos afloramentos rochosos. O moni-toramento da densidade da população no Rio Paraguai e tributários não tem sido feito desde 2007.
Atividade reprodutiva e fatores limitantes ao estabelecimento do mexilhão-dourado
A presença de larvas e juvenis indica re-produção quase contínua por cerca de 7-8 me-ses no Rio Paraguai, semelhante a outros luga-res na América do Sul (Cataldo & Boltovskoy, 2000; Santos et al., 2005). Tanto a temperatura da água quanto o pulso de inundação influenciam significativa-mente a atividade reprodutiva e a densidade de L. fortunei no Pantanal, embora de maneiras diferentes. Enquanto a temperatura parece atu-ar diretamente no ciclo reprodutivo, a sazona-lidade do pulso de inundação altera as variá-veis químicas, como nos eventos de depleção de oxigênio, e causa mortalidade dos mexilhões adultos (Oliveira et al., 2011), afetando direta-mente a densidade e reprodução. No Rio Paraguai L. fortunei está expos-ta a temperaturas relativamente elevadas (até 34ºC) em comparação ao seu ambiente nativo na China (média mensal máxima até 27,8ºC) (Ho et al., 2003) e outros ambientes na Améri-ca do Sul, onde a temperatura máxima mensal da água é geralmente abaixo de 30ºC (Bolto-vskoy & Cataldo, 1999). Cataldo & Boltovskoy (2000) sugerem que em temperaturas abaixo de 16,7ºC L. fortunei para de reproduzir. No en-tanto, observamos que uma queda na tempera-tura da água no início do inverno de 5 a 8ºC, mais que valores absolutos, podem estimular a
parada reprodutiva de L. fortunei no Pantanal, uma vez que as temperaturas mínimas no Rio Paraguai ficam acima de 17ºC (Oliveira et al., 2011). De acordo com Mackie & Claudi (2010), as variáveis que mais afetam o potencial de so-brevivência dos moluscos são os níveis de cál-cio, alcalinidade, pH e dureza total, as chama-das variáveis essenciais. Baseado nos ambientes onde ocorre L. fortunei, Ricciardi (1998) sugeriu que o pH ideal para infestações de L. fortunei é acima de 6,4. O pH do Rio Paraguai é em torno de 6,5, mas pH em torno de 6,0 é bastante comum e pode chegar a 5,0 ou menos, durante a fase de inundação. Embora mortalidades tenham sido observadas anualmente durante a inundação, consideramos que em pH 6.0 é possível ter po-pulação estabelecida, ainda que em densidades menores àquelas obtidas em lugares com pH mais alto (Oliveira et al., 2011). Oliveira et al. (2011) consideram que águas com cálcio acima de 1,0 mg/L têm poten-cial para receber o mexilhão-dourado, embora a espécie possa ocorrer em baixas densidades. Durante as inundações o ambiente pode ficar limitante para o mexilhão-dourado, que não sobrevive quando concentrações de oxigênio dissolvido de 0.0 mg/l, pH em torno de 5.0, índice de saturação da Calcita (IScalcita) de –4,0, CO2 livre por volta de 100mg/L, e temperatu-ra maior que 30ºC ocorrem simultaneamente (Oliveira, M. et. al., 2010b). Essas condições foram observadas em eventos de depleção de oxigênio dissolvido denominado localmente como ‘decoada’ na Baía do Castelo (lago co-nectado ao Rio Paraguai), onde a espécie foi eliminada no ano de 2007. Apesar de elimina-da, é provável que a espécie volte a colonizar o ambiente devido ao contato das águas da baía com as do Rio Paraguai nos anos subsequentes, porque outros lagos rio acima funcionam como reservatórios de larvas, e não apresentam com frequência o fenômeno de ‘decoada’. Eventos anuais de ‘decoada’ no Rio Miranda (Passo do Lontra), provavelmente foram responsáveis por baixas densidades de larvas e juvenis observa-das neste rio (Oliveira et al., 2011). A predação por peixes e a flutuação do nível d’água tam-bém são fatores controladores da densidade da espécie (Oliveira et al., 2006).

Oliveira & Calheiros
Limnoperna fortunei na bacia do Rio Paraguai, Mato Grosso do Sul
200
L. fortunei tem sido considerada uma pra-ga no Brasil por causar danos nas estações de captação de água, sistemas de refrigeração de hidrelétricas, e sistemas de cultivo em tanques--rede, bem como impactos ambientais, devido a sua alta densidade populacional. Incrustações de L. fortunei são observadas nas embarcações que navegam no Rio Paraguai, mas não representam grandes desafios quanto ao controle, e nem perdas econômicas, quando comparados ao sistema de captação de água e hidrelétricas localizadas no Rio Paraná. Os impactos no ambiente poderão ser mais expressivos em longo prazo com trocas na estrutura da cadeia trófica, como relatado em ambientes da Argentina (Darrigran & Damborenea, 2011). Apesar da mortalidade anual e oscila-ção na densidade populacional, o que temos observado é que L. fortunei tem resistido, re-colonizando os ambientes após um período de
depleção de oxigênio. Nos anos mais secos o fenômeno de ‘decoada’ é de baixa intensidade e favorece o crescimento da espécie por não ha-ver mortalidades na população. Eventos de depleção de oxigênio somen-te ocorrem na área de inundação do Rio Para-guai, incluindo o trecho final dos tributários como Cuiabá e Miranda. Quando L. fortunei atravessa essa barreira não encontra fatores limitantes ao seu desenvolvimento, se houver meios de introdução, certamente a espécie irá se estabelecer na parte alta dos rios da bacia. Um indicativo desta invasão é a expansão das espécies também de moluscos invasores Cor-bicula fluminea (Müller, 1774), Corbicula lar-gillierti (Philippi, 1844) e Melanoides tubercu-latus na bacia do Rio Miranda (Oliveira, M. et al., 2010c), cuja ocorrência é ampla, incluindo a cabeceira do Rio Miranda e a maioria dos tributários.

201
Introdução
O bioma Pantanal é caracterizado pela inundação sazonal, que determina processos ecossistêmicos específicos, com a ocorrência de plantas e animais adaptados à mudança anual de encolhimento e expansão de habitats, em virtude do regime hidrológico sazonal (Junk et al., 1989; Alho, 2011). Dessa forma, ocorrem áreas sujeitas a intenso alagamento, periódico ou permanente, áreas com baixo índice de ala-gamento, e áreas nunca alagadas (Scremin-Dias et al., 2011). Nas últimas décadas, a perda de habitats e de biodiversidade têm se intensificado nesse sistema, decorrentes do desmatamento, prin-cipalmente no Cerrado de Planalto, onde nas-cem os rios que alimentam o Pantanal (Silva et al., 2010). Associado à conversão de habitats naturais em pastagens, está a degradação dos recursos hídricos pela contaminação por agro-tóxicos e herbicidas, resíduos sólidos urbanos e também pela introdução de espécies exóticas.
As espécies invasoras foram considera-das como a terceira maior ameaça para a bio-diversidade pela União Internacional para a Conservação da Natureza (IUCN) e Governo brasileiro (Alho et al., 2011). Já o Ministério do Meio Ambiente – MMA, considera espécies in-troduzidas a segunda maior causa de extinção de espécies (Coradin, 2006). A introdução de espécies exóticas nos ecossistemas aquáticos e terrestres do Pantanal e seu impacto sobre as populações, comunida-des e funcionamento do ecossistema devem ser tema prioritário para pesquisas na região. Dentre as espécies exóticas aquáticas, o molusco bivalve trazido da China Limnoperna fortunei (Dunker, 1957) é o mais conhecido atualmente. O mexilhão-dourado foi registra-do no Rio Paraguai a partir de 1999 (Oliveira et al., 2000). A combinação da maturidade sexual precoce, alta fecundidade, ampla tolerância a condições diversas permitem que L. fortunei seja um invasor de sucesso em novos ambientes (Darrigran, 2002). A colonização pode ocorrer
CAPÍTULO 17
Limnoperna fortunei associada a macrófitas aquáticas
na bacia do Rio Paraguai, Mato Grosso
Sandra F. Marçal Claudia T. Callil

Marçal & Callil
Limnoperna fortunei associada a macrófitas aquáticas na bacia do Rio Paraguai, Mato Grosso
202
em estruturas não naturais ou naturais, como troncos de árvores, raízes, rochas e plantas aquáticas (Boltovskoy et al., 2006). No Panta-nal, a ausência de substratos duros fez com que se fixasse em macrófitas aquáticas (Callil et al., 2006). Isso aumentou as preocupações com o crescimento populacional deste bivalve nos rios e lagoas do Pantanal Matogrossense, pois se acredita que em alta densidade, o agregamen-to dos indivíduos possa alterar a composição e abundância da fauna de invertebrados nativos (Darrigran & Pastorino, 2000). Assim, alguns trabalhos foram desenvolvidos a fim de enten-der a dinâmica populacional e possíveis efeitos na estrutura da comunidade de invertebrados associados às raízes da macrófita flutuante Ei-chhornia crassipes Mart. (Solms) (Callil et al., 2006, Marçal & Callil, 2008).
Locais e método de amostragem
A área de estudo abrangeu um trecho do alto Rio Paraguai, entre o Parque Nacional do Pantanal e a Reserva Particular do Patrimônio Natural Acurizal, MT. Nesse trecho, 15 lagoas marginais foram visitadas na estação seca, em setembro de 2005. Foram feitas amostragens de invertebra-dos associados a raízes da macrófita flutuante E. crassipes. Para isso, tréplicas da vegetação flutuante de estandes marginais monoespecífi-cos foram coletadas com auxílio de uma tesou-ra de poda, utilizando-se um quadrado 25 x 25 cm. A coleta das réplicas foi realizada em três pontos distantes entre si em cada Lagoa e teve como objetivo aumentar a variabilidade das amostras nas unidades amostrais (lagoas).
Figura 1. Área de ocorrência de Limno-perna fortunei, referente aos estudos desenvolvidos com macrófitas em la-gos rasos adjacentes ao Rio Paraguai (Imagem de satélite: Google Earth; mapa: Marçal & Callil, 2008. Acta li-minologica Brasiliensia 20(4):359-371).

CAPÍTULO 17
Seção 4
203
A biomassa da planta foi obtida também a partir de amostras em tréplica a partir da ae-ração forçada em estufa a 60ºC (cerca de 72 horas) e pesagem em balança analítica. Para estudo dos invertebrados, a massa de raízes das plantas foram lavadas em peneiras de 2 mm, 1 mm e 0,25 mm, pré-triada e a parte resultante retida na última peneira foi fixada em álcool 70%. Os invertebrados foram triados e identificados sob microscópio estereoscópico. A densidade de L. fortunei nas plantas foi obtida para cada lagoa. Para estudo da es-trutura populacional de L. fortunei, os exem-plares coletados foram medidos em seu maior comprimento com auxílio de paquímetro digi-tal. A estrutura etária da população foi ava-liada utilizando um gráfico de distribuição de frequências por classes de comprimento.
Análise de dados A abundância da comunidade de ma-croinvertebrados obtidos para cada amostra fo-ram somados e expressos em número de indiví-duos por 100 g de peso seco de raiz (ind.100–1 PS) e indivíduos por metro quadrado (ind.m-2).Avaliamos a relação da abundância da comuni-dade de macroinvertebrados com a densidade de L. fortunei utilizando correlação e análise multivariada (Marçal & Callil, 2008). Os dados de densidade, comprimento médio da concha e classe modal da população de mexilhão-dourado foram apenas usados para interpretações da interferência na estrutu-ra da comunidade de macroinvertebrados asso-ciados a E. crassipes.
Características populacionais de L. fortunei em lagoas do Rio Paraguai A densidade de L. fortunei obtida no es-tudo é similar a já registrada para a região do Pantanal. Houve uma grande variação nesses valores de densidade (21-3616 ind.m-2), e, em grande parte, estes foram inferiores aos valores observados em afloramentos rochosos no iní-cio da colonização dessa espécie no Rio Para-guai (1000 ind.m-2 em 1999) (Oliveira, 2003). Nas 15 lagoas visitadas (Fig. 1), a densi-
dade média foi de 1327 ind.m-2. As lagoas que apresentaram as maiores abundâncias e densi-dades foram: Turco com 576 indivíduos (3616 ind.m-2); Canafisto com 501 (3008 ind.m-2); Três Bocas com 477 (2917 ind.m-2). Essa den-sidade de mexilhão em E. crassipes é superior a registrada entre 2001 e 2002 por Oliveira et al. (2006) em substratos artificiais de concreto (88,9 ind.m-2) e madeira (523,8 ind.m-2) e em 2004 por Callil et al. (2006) em Eichhornia azurea (107.75 ind.m-1), ambos no Rio Paraguai (Tab. I). A maior densidade observada em subs-tratos rochosos em relação à apresentada para macrófitas demonstra que os primeiros ofere-cem maior estabilidade e maior possibilidade de agregamento aos mexilhões. Assim como observado para o mexilhão-zebra Dreissena polymorpha (Pallas, 1771) nos Grandes Lagos dos Estados Unidos (Diggins et al., 2004), as macrófitas parecem prover um substrato alter-nativo para o mexilhão-dourado no Pantanal, podendo atuar na dispersão desse bivalve Rio Paraguai abaixo por deriva. Quanto ao comprimento médio das val-vas nas lagoas, observamos variação entre 1,8 – 9,26 mm (Lagoa Caracarazinho e Lagoa da Cláudia, respectivamente). A distribuição de frequência de L. fortunei por classes de compri-mento demostrou classe modal no intervalo de 1,31-2,61 mm e 2,62-3,92 mm, com exceção da Lagoa da Cláudia, que apresentou comprimen-to modal entre 14,40-15,70 mm (Tab. I). Estes valores são compatíveis com aqueles descritos por Marcelo et al. (2004) e Callil et al. (2006) ambos no Rio Paraguai, os quais relatam que as amplitudes de comprimento variam entre 1 a 28 mm e de 5 a 13 mm, respectivamente. O aumento da população de L. fortunei no Rio Paraguai e em suas lagoas adjacentes têm se mostrado diferente dos ambientes co-lonizados no Rio da Prata, onde o incremento na população desse bivalve se deu de maneira acelerada, com rápida dispersão e registro de altas densidades em pouco tempo de coloniza-ção, cerca de 150.000 ind.m-2 em três anos de invasão (Darrigran & Pastorino, 2000). Esse crescimento explosivo foi também registrado no Lago Guaíba, RS, aonde L. fortunei chegou a 143.500 ind.m-2 (Mansur et al., 2004c). A baixa densidade de L. fortunei obser-

Marçal & Callil
Limnoperna fortunei associada a macrófitas aquáticas na bacia do Rio Paraguai, Mato Grosso
204
vada no Rio Paraguai em relação aos registros da região Sul do Brasil, pode estar relacionada às peculiaridades hidrológicas dessa planície de inundação, a qual é caracterizada pela flutua-ção sazonal do nível da água (Junk et al., 1989), o que torna os substratos colonizados mais ins-táveis. A grande escassez de substratos duros e, quando existentes (pedras, afloramentos de rochas e troncos de árvores), ficam expostos à dessecação no período de águas baixas. Já no período de águas altas, os estandes de E. cras-sipes e outras macrófitas podem ser arrastados. Nesse período, ocorre também a decomposição da matéria orgânica carreada pela enchente, que ao decompor-se diminui a concentração de oxigênio na água, controlando a reprodu-ção (Uliana & Callil, 2006) e o crescimento das populações de L. fortunei (Oliveira et al., 2006).
Fauna de macroinvertebrados associados à macrófita flutuante E. crassipes Nas 15 lagoas amostradas (Fig. 1) foram coletados um total de 86.943 indivíduos e 64 taxa de invertebrados, distribuídos nos filos Nematoda, Annelida, Mollusca e Arthropoda (Tab. I, Fig. 2). Desse total, Hydrobiidae foi o táxon mais abundante com 17966 indivíduos (21%), seguido de Ostracoda com 17872 (20,6%), Hydracarina 11353 (13,05%) e Eupera sp. (Sphaeridae) com 8322 (9,57%). Apresentaram também abundância expressiva os táxons Chi-ronomidae (7%), Trichoptera (5,4%), além de L. fortunei e Planorbidae que juntos somaram 8,83% do total de indivíduos (Tab. I, Fig. 2). A estrutura da comunidade de macroin-vertebrados foi testada em relação à variação na abundância de L. fortunei nas macrófitas. O resultado demostrou que o mexilhão-doura-do não afetou a estrutura da comunidade de macroinvertebrados amostrados nas plantas. Porém, observamos por meio de correlação uma pressão negativa sobre Cladocera frente à abundância de L. fortunei. Acreditamos que a diluição das amostras pode ter enfraquecido o resultado, devido ao grande número de lagoas amostradas e o número de amostras coletadas em cada uma. Porém, a densidade de L. fortu-
nei obtida foi muito baixa quando comparada aos valores registrados para o sul do Brasil com valores próximos a 100.000 ind.m-2 (Darrigran & Pastorino, 2000). A ocorrência de altas densidades do me-xilhão-dourado na América do Sul tem levado a uma diminuição da abundância e riqueza de espécies de invertebrados (Darrigran et al., 1998a; Ricciardi, 1998; Orensanz et al., 2002; Conde et al., 2002; Mansur et al., 2003; Scara-bino, 2004; Brugnoli et al., 2005). Os resultados aqui apresentados não são suficientes para demonstrar que o pulso de inundação pode controlar a densidade de mexilhão-dourado no Rio Paraguai. Porém, o tempo decorrido após a introdução da espécie e as baixas densidades registradas, nos faz in-ferir que esse processo possa ser determinante no controle populacional. Entretanto, a inun-dação sazonal no Pantanal não pode conter a dispersão da espécie, que pode ocorrer devido ao transbordamento lateral da água para os campos no período de cheia e maior densidade de organismos nas lagoas laterais ao Rio Para-guai, que atuam como fontes dispersoras. Considerando os resultados desse traba-lho, fica evidente que no Pantanal a flutuação anual do nível da água e a escassez de subs-tratos estáveis podem ser fatores limitantes para o crescimento populacional de L. fortunei (Oliveira et al., 2006), o que torna pouco pro-vável a ocorrência de densidades equivalentes às encontradas na Argentina (Darrigran & Pas-torino, 2000) e sul do Brasil (Mansur et al., 2004c). No entanto, somente após a realização de estudos mais detalhados sobre os fatores que interferem no crescimento populacional dessa espécie invasora, bem como a construção de um conhecimento aprofundado das comuni-dades associadas a E. crassipes, que considerem as variações temporais e relações tróficas esta-belecidas, será possível avaliar de forma efetiva o efeito da introdução de L. fortunei na estru-tura da fauna de macroinvertebrados.
Observações e recomendações
O equilíbrio de um sistema natural de-pende da ação combinada e equilibrada entre a estrutura do ambiente, que reflete o quão ínte-

CAPÍTULO 17
Seção 4
205
Figura 2. Abundância relativa (%) dos taxa de invertebrados associados a E. crassipes nas 15 lagoas marginais ao Rio Paraguai, Mato Grosso amostradas em setembro de 2005. (Lagoas = L1 a L15).
gro este se encontra, com seu funcionamento, determinando, assim, não apenas a estabili-dade, mas também a amplitude de resiliência. A ação de uma espécie invasora com um alto poder de interferência nestes atributos, integri-dade e estabilidade, deve receber atenção con-tínua (Fig. 3). Novos estudos são necessários, considerando o monitoramento dos fatores ambientais e dos padrões de biodiversidade, para que se entenda a evolução do mexilhão--dourado nos sistemas aquáticos do Pantanal. A estratégia mais eficiente para o contro-le da dispersão do mexilhão-dourado e outras espécies no Pantanal é a sensibilização para a problemática que os bioinvasores podem causar para a fauna aquática. Ações em diferentes ní-veis são necessárias como medidas mitigadoras, iniciando pelos pescadores e ribeirinhos que desconhecem os efeitos negativos de sua práti-ca, quando transportam de um lugar para ou-tro, água, iscas, e embarcações, até os gestores e autoridades ambientais que, no estado de Mato Grosso, tem negligenciado a intensidade da problemática. Propomos aqui o planejamento estratégico considerando cinco níveis para o de-senvolvimento de ações específicas integradas, como medidas de monitoramento e controle da dispersão de bioinvasores (Fig. 4). Os efeitos dos impactos na bacia do Rio Paraguai são evidentes quando causados pela degradação do solo e poluição, porém são in-conspícuos os efeitos causados nos sistemas aquáticos causados pela introdução de espécies exóticas. Alho (2011) e Alho et al. (2011) apon-tam a organização e continuidade das pesquisas na forma de um amplo programa de conserva-ção. Este deverá identificar, avaliar e mitigar os
Figura 3. Modelo conceitual básico da ação de uma ou mais es-pécies invasoras frente aos principais atributos do ecossistema.
Figura 4. Níveis para o desenvolvimento de ações estratégicas para a gestão adequada relacionada à interferência de espé-cies invasoras.
impactos ambientais no Pantanal e dessa forma restaurar, proteger e preservar habitats naturais dos impactos causados pela má gestão dos re-cursos naturais, causada, dentre outros fatores, pela fragilidade das organizações ambientais para aplicar a legislação. Dessa forma, a apro-vação de projetos de desenvolvimento na região do Pantanal deve perpassar pela utilização sus-tentável dos recursos biológicos. A investigação científica poderá incitar a discussão de avanços, problemas e prioridades dentro um plano de ação para conservar a biodiversidade regional.

Marçal & Callil
Limnoperna fortunei associada a macrófitas aquáticas na bacia do Rio Paraguai, Mato Grosso
206
Tabela I. Coordenadas geográficas, biomassa de E. crassipes (raiz e total), densidade de invertebrados e de L. fortunei o comprimento médio de L. fortunei nas 15 lagoas marginais ao Rio Paraguai, Mato Grosso amostradas em setembro de 2005. L1 a L15 = Lagoas, PS = Peso seco.
LagoasCoordenadas Biomassa de
E. crassipes (PS)Densidade de Invertebrados L. fortunei
Comp. Médio
Lat/Long Raiz (g.m-2)
Planta (g.m-2)
ind/100g PS ind.m-2 ind.m-2 (mm)
Acurizal S17°49’24,8’’ W57°33’48,9’’ 609.76 1274.51 6399 39019 21 0
Caracará S17°52’50,9’’ W57°28’24,8’’ 881.07 1600.53 2884 25408 283 2,82
Joãozinho S17°52’13,2’’ W57°28’51,8’’ 1248.16 2109.01 2348 29301 1792 3,93
Comprida S17°52’36,9’’ W57°30’39,3’’ 853.76 1509.76 6111 52171 533 3,7
Piuval S17°53’37,6’’ W57°29’57,4’’ 707.09 1865.39 4124 29157 827 6,38
Desprezo S17°53’53,4’’ W57°29’00,8’’ 997.76 1633.76 1930 19259 384 3,74
Turco S17°48’05,9’’ W57°15’35,9’’ 658.83 1535.79 5388 35499 3616 3,14
Bigueirinho S17°48’07,7’’ W57°34 05,5’’ 1041.65 1956.48 2091 21776 725 2,44
Canafisto S17°47’33,8’’ W57°33’18,9’’ 749.60 1677.92 2888 21648 3008 1,96
Caracarazinho S17°50’23,2’’ W57°29’12,1’’ 941.92 1839.79 2099 19771 219 1,8
Três Bocas S17°51’21,1’’ W57°28’57°,1’’ 1162.08 1973.33 1460 16971 2917 3,08
Claúdia S17°51’20,1’’ W57°30’11,2’’ 1071.79 1936.80 5501 58955 443 9,26
Sandrinha S17°51’02,9’’ W57°29’45,0’’ 1220.80 1851.41 2446 29856 2539 3,18
Inês S17°49’54,9’’ W57°31’46,4’’ 1772.36 3738.45 1838 32568 2426 3,14
Figueira S17°48’32,5’’ W57°32’35,3’’ 991.68 1845.12 2920 28960 1483 2,43

207
A procura de L. fortunei, seja da larva ou do adulto, em um ambiente desconhecido e nunca estudado, é uma tarefa árdua para muitos pesquisadores, quando solicitados por órgãos governamentais ou privados. Tendo em vista a necessidade de uma análise preliminar do ambiente, muitas vezes ainda sem financia-mento, é preciso verificar com cuidado qual método poderá ser utilizado naquele ambiente para detectar esses bivalves. Dever-se-á tentar não incorrer no erro metodológico. Mesmo as-sim, com todos os cuidados utilizados pode-se não encontrar o mexilhão-dourado, devido a muitos fatores ainda não pesquisados sobre a biologia dessa espécie (Takeda et al., no prelo). Em janeiro de 2001, no reservatório de Itaipu ainda não havia sido detectado nenhum mexilhão-dourado, então foi solicitada a veri-ficação da sua existência ao longo do canal à jusante da barragem desse reservatório. O reservatório de Itaipu é muito utiliza-do por pescadores, tanto amadores como pro-fissionais, bem como por turistas que utilizam os campings espalhados em suas margens. A
primeira ocorrência de L. fortunei foi constata-da na estação ICLI, onde se localiza o Iate Clu-be do Lago Itaipu, com embarcações de turistas advindas de diversos lugares. O estudo desenvolvido ao longo de re-servatório, com início em maio de 2001, foi o primeiro no Brasil que utilizou uma abordagem experimental dentro do reservatório, em uma extensão tão ampla. Os experimentos condu-zidos mostraram-se efetivos, possibilitando de-terminar a colonização do mexilhão-dourado ao longo do reservatório de Itaipu, desde a zona lacustre até a de transição do reservatório.
1. Fase I
As estações de coletas foram identificadas pela sigla E1 a E20 correspondendo, respectiva-mente, às estações 1 a 20 desde Encarnación, à jusante, até Ciudad del Este, à montante (Fig. 1A e B). Para realizarem-se as amostragens de campo foi necessário levar em consideração as seguintes condições: o tempo disponível, a segurança da equipe de trabalho e a verba disponível.
CAPÍTULO 18
Experiência da procura e monitoramento no início da invasãode mexilhão-dourado no Rio Paraná
e no reservatório de Itaipu
Alice Michiyo Takeda Daniele Sayuri Fujita

Takeda & Fujita
Experiência da procura e monitoramento no início da invasão de mexilhão-dourado no Rio Paraná e no reservatório de Itaipu
208
1.1. Morfologia e profundidades das estações de coleta à jusante do reservatório de Itaipu.
As estações foram distribuídas em um trecho de 344 km onde o Rio Paraná corre di-retamente sobre o basalto da Formação Serra Geral. Esse trecho do rio flui através de uma garganta, provavelmente de origem estrutural, alcançando os maiores valores de velocidade e profundidade em locais onde seu canal se apresenta mais estreito. Consequentemente, é nesse trecho que o rio tem os maiores valores de potência de fluxo (power stream), exceto no trecho das antigas Sete Quedas. Em algumas estações a velocidade atinge cerca de 2,46 m/s, até 68 m de profundidade, para valores de po-tência de fluxo em torno de 94,00 W/m-2.
Muito embora o rio apresente um gra-diente hidráulico variando entre 0,024 e 0,054 m/km, o gradiente do fundo, controlado por tectonismo, apresentou valores totalmente anômalos, inclusive com situações de inversão de gradiente. Isso provoca um fluxo extrema-mente turbulento com formação de vórtices, correntes ascendentes e refluxos. Os dados gerais referentes à profundida-de, velocidade de fluxo, gradiente hidráulico, largura do canal e potência de fluxo estão apre-sentados na Figura 2.
1.2. Amostragem de larvas de Limnoperna fortunei (Dunker, 1857)
As amostragens de campo foram realiza-
Figura 1. A, Mapa e localização das estações de coletas de E1 a E20.
A

CAPÍTULO 18
Seção 4
209
das, mensalmente, na Fase I – de janeiro a março de 2001, à jusante do reservatório de Itaipu (E1 a E20). Na Fase II coletou-se no reservatório de Itaipu – de abril de 2001 a agosto de 2002. Na Fase I do projeto realizaram-se as co-letas no Rio Paraná, onde as fortes correntezas predominaram durante quase todo curso do rio, com alguns trechos apresentando afloração de rochas, onde se formavam redemoinhos. Durante a subida do rio, uma caminho-nete da Itaipu/ Paraguai esperava a equipe no local pré-determinado para o dia. No barco tipo chatão, motor 70, era carregado o material de coleta, como bomba de sucção, mangueiras, potes, frascos de álcool, rede de plâncton etc. A equipe de coleta formada por dois téc-nicos (Itaipu – Brasil) para auxiliar as coletas, parar o barco no meio do canal jogando uma âncora de ferro e na filtração de água, um téc-
nico do Itaipu – Paraguai para tomar as medi-das de dados físicos e químicos da água e duas pesquisadoras da Universidade Estadual de Maringá para tomar os dados biológicos. Nessas condições, optou-se, para cada estação, coletar 100 litros de água com auxílio de moto-bomba, pois em alguns trechos esse volume de água foi o máximo de tempo dis-ponível para o barco ficar parado no meio do canal sem afundar. Na região central, coleta-ram-se em três profundidades: superfície, meio e fundo (comprimento máximo da mangueira – 40 m), 60 litros de água com balde, além de amostragem qualitativa com rede (5 minutos). Na margem, também foram coletados 100 li-tros na superfície, juntamente com a coleta de balde. Portanto, em cada estação foi coletado um total de 520 litros de água, além de amostra para análise qualitativa de rede.
Figura 1.B, Perfis de profundidade das estações.
B

Takeda & Fujita
Experiência da procura e monitoramento no início da invasão de mexilhão-dourado no Rio Paraná e no reservatório de Itaipu
210
Em cada campanha, com 20 estações, fo-ram amostrados no total, 10.400 litros obtidos com bomba e filtrada em rede de plâncton de 25 μm. Além disso, realizou-se amostragem quali-tativa de aproximadamente 10 minutos de água filtrada pela mesma rede. Nessa fase não foi de-tectada nenhuma larva nesse trecho do canal. Em três meses de coleta não foi observa-da nenhuma larva de L. fortunei. Devido à for-te correnteza desse trecho, seria pouco provável à subida de larvas do reservatório de Yacyretá para o reservatório de Itaipu pelo estreito canal do Rio Paraná por meio natural. Provavelmen-te, o fator antrópico ajudou a subida dos inva-sores diretamente para região próxima ou no reservatório de Itaipu.
2. Fase II
A partir de abril de 2001, após o primei-ro registro de um espécime adulto na Usina de Itaipu (Zanella & Marenda, 2002), passou-se a coletar amostras de água, do substrato artifi-cial e de zoobentos do reservatório.
2.1. Amostragem e distribuição das larvas de Limnoperna fortunei
As coletas de água, para verificar a ocor-rência de larvas de L. fortunei no reservatório de Itaipu, foram realizadas em quatorze esta-ções do corpo do reservatório (Fig. 3).
R1 = em frente à Usina, mais ou menos próximo a Unidade 11 (25o23’40,6’’S; 54o34’50,6’’WO).
R2 = na praia de Santa Teresinha (25o21’46,2’’S; 54o30’24,9’’WO).R3 = no município de São Miguel do Igua-çu, em frente à foz do Rio Ocoí (25o14’07,6’’S; 54o27’12,8’’WO).R4 = no município de Missal (25o02’27,0’’S; 54o26’03,0’’WO).R5 = no município de Missal – perto da mar-gem com pedreira, onde foi visto o mexilhão (25o04’09,6’’S; 54o22’51,3’’WO)R6 = no município de Santa Helena (24o46’21,8’’S; 54o21’35,3’’WO).R7 = no município de Santa Helena (Sartori), próxima a tanque rede – onde já foi visto o me-xilhão (24o50’02,0’’S; 54o19’24,1’’WO).R8 = no município de Pato Bragado (Porto Britânia), no meio do canal (24o38’55,0’’S; 54o19’16,0’’WO).R9 = no município Entre Rios, no meio do ca-nal (24o40’55,5’’S; 54o20’03,6’’WO).R10 = no município de Marechal Cândido Ron-don, no canal do reservatório (24o29’38,7’’S; 54o19’38,6’’WO).R11 = no município de Arroio Guaçu (24o22’51,6’’S; 54o15’38,6’’WO).R12 = no município de Guaíra, no meio dos pa-liteiros (24o06’05,8’’S; 54o19’19,2’’WO).R13 = no município de Guaíra, fora do reserva-tório, depois da Ponte MT-PR (24o03’15,9’’S; 54o15’22,4’’WO).R14 = em frente do Iate Clube do Lago de Itai-pu (25o26’12,6’’S; 54o30’45,4’’WO).
Em cada estação, foram obtidas amos-tras em três profundidades, utilizando-se uma
Figura 2. Perfil batimétrico, velocidade de fluxo, gradiente hidráulico, largura do canal e potencia do fluxo das estações de coleta à jusante do reservatório. E = estação de coleta.

CAPÍTULO 18
Seção 4
211
moto-bomba para filtrar 200 litros de água por amostra, em uma rede de plâncton de 25 μm de abertura de malha. O material coletado em campo foi acondicionado em frasco de polieti-leno devidamente etiquetado e fixado em álco-ol 70º. As amostras foram analisadas no Labo-º. As amostras foram analisadas no Labo-. As amostras foram analisadas no Labo-ratório de Zoobentos do Nupelia. Todas as larvas de Bivalvia encontradas nas amostras foram retiradas e analisadas (me-didas e morfologia externa) em microscópio óptico. Para o reconhecimento de larvas de L. fortunei foi necessário ter muito cuidado na identificação, principalmente onde havia mui-tos indivíduos de Corbicula, como foi o caso do reservatório de Itaipu. A detecção das larvas de L. fortunei no reservatório de Itaipu, apesar de intensas co-letas de água, foi observada apenas em feve-reiro de 2002 (Fig. 4), sugerindo, assim, que a detecção das larvas no início da colonização,
pelo método tradicional, é mais difícil devido a pouca quantidade de adultos que estão se re-produzindo. Observou-se um pico na densidade de larvas em maio e junho de 2002, decrescendo nos meses subsequentes (Fig. 4). O decrésci-mo dessas larvas pode indicar um período de baixa reprodução, porém, destaca-se que essa pesquisa enfocou o início da colonização do mexilhão-dourado no reservatório, quando a densidade ainda era muito baixa em relação aos resultados observados na Argentina por Cataldo & Boltovskoy (2000), que encontra-ram até 33.706 larvas m-3 no Rio de La Plata. Dessa forma, nada pode ser concluído a respei-to da atividade reprodutiva dessa espécie no reservatório de Itaipu. Cataldo & Boltovskoy (2000) sugerem a temperatura de 16-17ºC como limite para a reprodução do mexilhão-dourado. Tendo em vista que as temperaturas da água do reservató-rio não decrescem tanto durante certo período (de um mês ou mais), pode-se esperar que, com a adaptação dessa espécie, a reprodução tenha sido mais intensa do que na Argentina.
2.2. Amostragem e distribuição de Limnoperna fortunei com substrato artificial
Os substratos artificiais são mais fáceis de serem controlados, servem para coletar os invertebrados incrustantes. Os substratos artificiais foram insta-lados em janeiro de 2001, em sete regiões do reservatório de Itaipu e sem réplicas. A partir de abril de 2001, instalaram-se mais substratos artificiais ao longo do reservatório de Itaipu,
Figura 3. Mapa do reservatório de Itaipu com os pontos de coleta de larvas de L. fortunei.
Figura 4. Média das densidades de larvas registradas no re-servatório de Itaipu ao longo de dois anos de coleta.

Takeda & Fujita
Experiência da procura e monitoramento no início da invasão de mexilhão-dourado no Rio Paraná e no reservatório de Itaipu
212
com réplicas em cada região, uma vez que o monitoramento passou a ser no reservatório, abandonando-se as coletas à jusante do mesmo. Necessitou-se de constante reposição dos 37 substratos artificiais instalados em maio de 2001, ao longo e à jusante do reservatório, devido ao desaparecimento constante dos mes-mos. As coletas à jusante do reservatório foram abandonadas devido a necessidade de reposi-ção total a cada mês. O substrato artificial foi feito com uma boia fixada no leito do reservatório (Fig. 5), onde duas placas de madeira em X foram fixa-das (Fig. 6). Um substrato foi instalado a 1,5 m (S1) e outro a 3,0 m (S2) da superfície da água. As amostras dos substratos foram raspadas, mensalmente, utilizando-se um quadrado con-feccionado com borracha e fixados imediata-mente em álcool 70º. Todo material foi triado sob microscópio estereoscópio e os organismos contados. Os experimentos com substratos artifi-ciais foram realizados no corpo do reservatório (Fig. 7) nas seguintes estações:
UsinaU1 – (substrato de madeira)U2 – (substrato de aço)U3 – (substrato de cimento)
RefúgioR1 – (aço) 25o24’14,8’’S – 54o34’45,9’’WOR2 – (cimento) 25o24’06,7’’S – 54o34’48,5’’WOR3 – (madeira) 25o23’40,6’’S – 54o34’50,6’’WO
Iate Clube – ICLII1 – 25o26’12,4’’S – 54o30’45,4’’WOI2 – 25o26’34,2’’S – 54O29’59,1’’WOI3 – 25o25’02,6’’S – 54o32’06,6’’WO
Santa Teresinha do IguaçuT1 – 25o22’03,4’’S – 54o30’17,2’’WOT2 – 25o22’00,3’’S – 54o30’27,8’’WOT3 – 25o21’55,3’’S – 54o30’53,2’’WO
São MiguelL1 – 25o13’41,0’’ S – 54o 14’27,4’’WOL2 – 25o14’31,7’’ S – 54o23’47,2’’WOL3 – 25o13’15,1’’ S – 54o26’33,8’’WO
Santa HelenaH1 – 24o48’34,1’’S – 54o20’01,4’’WOH2 – 24o47’29,0’’S – 54o20’57,9’’WOH3 – 24o46’43.2’’S – 54o21’11,8’’WO
Entre RiosE1 – 24o40’01,7’’S – 54o16’50,7’’WOE2 – 24o39’50,8’’S – 54o18’09,4’’WOE3 – 24o40’00,1’’S – 54o18’32,7’’WO
Pato Bragado (Porto Britânia)B1 – 24o38’44,5’’S – 54o18’42,6’’WOB2 – 24o38’39,5’’S – 54o19’11,4’’WOB3 – 24o38’35,8’’S – 54o19’22,8’’WO
Porto MendesM1 – 24o26’44,5’’S – 54o18’37,7’’WOM2 – 24o27’15,0’’S – 54o18’59,3’’WOM3 – 24o29’16,1’’S – 54o19’46,2’’WO
Arroio-GuaçuA1 – 24o23’04,5’’S – 54o15’24,3’’WOA2 – 24o22’57,6’’S – 54o15’28,4’’WOA3 – 24o22’51,2’’S – 54o15’26,9’’WO
GuaíraG1 – 24o06’19,9’’S – 54o18’54,3’’WOG2 – 24o05’12,9’’S – 54o17’49,7’’WOG3 – 24o06’19,9’’S – 54o18’54,3’’WO
Figura 5. Boia do substrato artificial instalada no reservatório de Itaipu.
Figura 6. Substratos artificiais que ficam pendurados na boia.

CAPÍTULO 18
Seção 4
213
Lago de Itaipu, com a máxima densidade de incrustação de 92.656 ind.m-2 nos substratos desta mesma estação (Figs. 9 e 10). O aumento exponencial de Limnoperna nos substratos artificiais coincidiu com a dimi-nuição de densidade das larvas de Chironomi-dae ao longo do tempo (Fig. 11). A proliferação de L. fortunei deve afetar outras comunidades do reservatório de Itaipu, mostrados aqui com o experimento realizado com substrato artificial. Para Oligochaeta (F(2, 1300)=59,708; p<0,000), Cladocera (F(2, 1300)=12,371; p<0,000) e Ostracoda (F(2, 1300)=30,103; p<0,000) obser-vou-se uma preferência pelo estrato boia, en-quanto Limnoperna (F(2, 1300)=13,501; p<0,000) e Trichoptera (F(2, 1300)=5,9603; p<0,00265) fo-ram registrados em maiores densidades nos estratos S1 e S2 (Fig. 12). Ephemeroptera (F(2, 1300)=3,3303; p<0,003609) apresentou maiores abundâncias nos estratos superiores (boia e S1). Para as larvas de Chironomidae não foram observadas prefe-rências por um dos estratos (boia, S1 e S2).
Figura 7. Mapa do reservatório de Itaipu com a localização de substrato artificial.
Figura 8. Média e erro padrão da densidade de Limnoperna fortunei nos substratos artificiais ao longo dos meses.
Figura 9. Substrato artificial ainda não colonizado por mexi-lhão-dourado.
Figura 10. Substrato artificial no final do experimento, com bastante mexilhão-dourado.
Nos substratos artificiais instalados no reservatório de Itaipu desde maio de 2001, ob-servou-se a rápida colonização e dispersão de L. fortunei ao longo do mesmo. A densidade de L. fortunei mostrou aumento progressivo des-de sua primeira ocorrência em agosto de 2001 (Fig. 8), na estação em frente ao Iate Clube do

Takeda & Fujita
Experiência da procura e monitoramento no início da invasão de mexilhão-dourado no Rio Paraná e no reservatório de Itaipu
214
Os substratos artificiais mostraram ser muito eficientes para monitorar os mexilhões dourados.
2.3. Amostragem e distribuição de Limnoperna fortunei em zoobentos
As amostragens bentônicas foram reali-zadas nas oito estações, abrangendo o braço do tributário até a proximidade do antigo canal do Rio Paraná, estabelecendo-se diversos pontos, especialmente próximos às margens até perto do canal antigo do Rio Paraná. As oito estações selecionadas nos muni-cípios (Fig. 13) foram: Guaíra (Taturí) (4 pontos) Mercedes (Arroio Guaçu) (6 pontos) Marechal Cândido Rondon (Porto Mendes) (7 pontos) Entre Rios do Oeste (rio São Francisco Verdadeiro) (7 pontos) Santa Helena (rio São Francisco Falso) (17 pontos) Missal (rio São Vicente) (12 pontos)
São Miguel (rio Ocoí) (19 pontos) Refúgio Biológico de Bela Vista (12 pontos)
As amostragens foram feitas com pegador de fundo tipo Petersen modificado (0,0345 m2) e acondicionadas em galões (Fig. 14). Em cada ponto de coleta, foram realizadas quatro amos-tragens, sendo três para análise biológica e uma para análise sedimentológica, totalizando men-salmente 252 amostras para a análise biológica. Logo após a coleta, o material biológico foi transportado para margem e lavado em jogo de peneiras com malhas de abertura 2,0, 1,0 e 0,5 mm (Fig. 15). Os organismos encontrados nas malhas de 2,0 e 1,0 mm foram retirados e fi-xados imediatamente em álcool 70º. O material retido na malha de 0,5 mm foi acondicionado em potes de polietileno em álcool 70º, para pos-º, para pos-, para pos-terior triagem sob microscópio estereoscópio, no Laboratório de Zoobentos do Nupelia.
Figura 11. Distribuição temporal dos valores médios da den-sidade (ind.m-2) de invertebrados.
Figura 12. Distribuição dos valores médios da densidade (ind.m-2) de invertebrados nos diferentes estratos.
Figura 13. Mapa do reservatório de Itaipu com os pontos de coleta de zoobentos.

CAPÍTULO 18
Seção 4
215
Local Coordenadas S Coordenadas WO
GR1 24º06’38,6’’ 54º18’48,0’’
GR2 24º06’30,6’’ 54º18’42,0’’
GR3 24º06’23,2’’ 54º18’33,6’’
GR4 24º06’19,3’’ 54º19’01,6’’
AG1 24º22’32,9’’ 54º15’51,5’’
AG2 24º22’57,7’’ 54º15’33,9’’
AG3 24º22’50,7’’ 54º15’23,0’’
AG4 24º23’00,7’’ 54º15’20,6’’
AG5 24º23’02,0’’ 54º15’22,4’’
AG6 24º23’03,7’’ 54º15’26,8’’
PM1 24º29’30,4’’ 54º20’08,8’’
PM2 24º27’17,1’’ 54º18’36,3’’
PM3 24º28’47,8’’ 54º19’01,2’’
PM4 24º29’37,4’’ 54º19’02,2’’
PM5 24º29’50,7’’ 54º19’09,5’’
PM6 24º30’12,3’’ 54º19’17,8’’
PM7 24º29’38,8’’ 54º18’49,1’’
ER1 24o40’55,5” 54o20’36”
ER2 24º39’39,8’’ 54º18’05,9’’
ER3 24º39’49,4’’ 54º17’57,1’’
ER4 24º40’03,3’’ 54º17’49,9’’
ER5 24º40’11,2’’ 54º16’56,3’’
ER6 24º40’06,8’’ 54º16’50,6’’
ER7 24º40’00,9’’ 54º16’38,9’’
SH1 24º45’40,9’’ 54º22’17,5’’
SH2 24º45’59,5’’ 54º20’54,3’’
SH3 24º46’50,9’’ 54º21’18,1’’
SH4 24º46’41,7’’ 54º20’17,8’’
SH5 24º47’20,5’’ 54º20’43,1’’
SH6 24º47’27,2’’ 54º20’53,7’’
SH7 24º47’28,2’’ 54º21’03,4’’
SH9 24º47’59,9’’ 54º20’42,4’’
SH8 24º47’54,1’’ 54º20’32,2’’
SH10 24º48’02,0’’ 54º20’52,4’’
SH11 24º48’39,9’’ 54º20’17,3’’
SH12 24º48’38,5’’ 54º20’03,6’’
SH13 24º48’27,6’’ 54º19’46,4’’
SH14 24º49’22,8’’ 54º20’14,4’’
SH15 24º50’14,1’’ 54º20’28,8’’
SH16 24º50’00,0’’ 54º20’10,7’’
SH17 24º50’12,4’’ 54º20’06,1’’
MS1 25º02’03,2’’ 54º26’54,4’’
Local Coordenadas S Coordenadas WO
MS2 25º02’40,7’’ 54º26’15,6’’
MS3 25º02’49,6’’ 54º25’33,6’’
MS4 25º01’19,9’’ 54º25’28,7’’
MS5 25º01’55,1’’ 54º24’46,6’’
MS6 25º01’32,1’’ 54º23’36,2’’
MS7 25º02’12,1’’ 54º23’44,3’’
MS8 25º02’50,8’’ 54º23’54,6’’
MS9 25º03’24,3’’ 54º24’23,3’’
MS10 25º04’18,0’’ 54º23’12,4’’
MS11 25º03’53,0’’ 54º22’02,0’’
MS12 25º03’38,3’’ 54º22’13,0’’
SM1 25º14’07,6’’ 54º27’12,8’’
SM2 25º13’26,0’’ 54º27’33,6’’
SM3 25º12’32,4’’ 54º25’57,5’’
SM4 25º12’23,4’’ 54º23’59,7’’
SM5 25º14’05,8’’ 54º26’41,6’’
SM6 25º14’47,0’’ 54º25’01,7’’
SM7 25º13’28,0’’ 54º23’27,9’’
SM8 25º14’56,0’’ 54º23’46,5’’
SM9 25º15’27,1’’ 54º23’36,6’’
SM10 25º15’08,1’’ 54º22’44,7’’
SM11 25º13’36,9’’ 54º22’08,3’’
SM12 25º13’21,5’’ 54º21’08,5’’
SM13 25º12’39,7’’ 54º19’57,1’’
SM14 25º13’13,0’ 54º19’27,5’
SM15 25º12’38,2’’ 54º18’29,1’’
SM16 25º12’20,4’’ 54º16’36,9’’
SM17 25º13’32,5’’ 54º15’38,3’’
SM18 25º13’32,5’’ 54º14’20,4’’
SM19 25º13’57,0’’ 54º13’44,1’’
RF1 25º24’19,1’’ 54º33’51,9’’
RF2 25º24’42,9’’ 54º32’02,7’’
RF3 25º24’58,2’’ 54º31’09,9’’
RF4 25º25’42,8’’ 54º30’39,4’’
RF5 25º26’24,1’’ 54º29’54,3’’
RF6 25º26’56,2’’ 54º29’56,9’’
RF7 25º26’46,4’’ 54º30’21,9’’
RF8 25º26’19,0’’ 54º30’45,3’’
RF9 25º25’32,1’’ 54º31’17,5’’
RF10 25º26’10,1’’ 54º33’27,5’’
RF11 25º26’35,0’’ 54º32’34,3’’
RF12 25º26’42,4’’ 54º33’03,8’’
Tabela I. Coordenadas dos pontos de cada estação de coleta. GR = Guaíra; AG = Arroio-Guaçu; PM = Porto Mendes; ER = Entre Rios; SH = Santa Helena; MS = Missal; SM = São Miguel; RF = Refúgio.

Takeda & Fujita
Experiência da procura e monitoramento no início da invasão de mexilhão-dourado no Rio Paraná e no reservatório de Itaipu
216
No reservatório de Itaipu, a coleta de zoobentos mostrou-se muito eficiente para de-tecção de mexilhão-dourado e, principalmen-te, para analisar melhor as condições e tipo de substratos utilizados pelos adultos. A maior densidade média de mexilhão-dourado foi en-contrada no clasto de argila, presente nas regi-ões marginais do reservatório (Fig. 16).
3. Considerações finais
Para se estabelecer um modelo de moni-toramento de mexilhão-dourado é necessária uma trama complexa de conhecimentos. Esses bivalves estão sujeitos a um número de diferen-tes agentes estressores. Esses agentes podem
Figuras 14, 15. 14, Pegador tipo Petersen (modificado); 15, Coleta e lavagem do material bentônico.
ser resultados de ações humanas ou simples-mente devido a eventos naturais locais, dife-rentes do país de origem da espécie. Portanto, o mexilhão-dourado não está adaptado a essas situações, como a conhecida “dequada”, prin-cipalmente a depleção de oxigênio dissolvido na água, no Pantanal, muitas vezes citadas pela Oliveira et al. (2006), podendo ser um dos fa-tores limitante para sua proliferação. O estresse pode levar a alterações nas concentrações de neurotransmissores ou pro-dução de hormônios, afetando o organismo nos níveis celulares e histológicos. A esse nível, ocorrem mudanças metabólicas, tais como, a osmorregulação e funções imunológicas, po-dendo atingir o organismo como um todo. Por
14 15
Figura 16. Média mensal das densidades de mexilhão-dou-rado em todos os pontos de coleta ao longo do reservató-rio de Itaipu.

CAPÍTULO 18
Seção 4
217
sua vez, os organismos respondem através de mudanças na dinâmica do crescimento e repro-dução, culminando em alterações que atingem toda a população e, consequentemente, poden-do levar a uma extinção total da invasora não bem sucedida. O mexilhão-dourado parece ter superado, por enquanto, todos os obstáculos naturais e antrópicos mantendo-se nos am-
bientes já colonizados como nos reservatórios, a planície aluvial do Rio Paraná, o Rio Ivinhe-ma, entre outros. Os métodos utilizados para análise da malacofauna do reservatório Itaipu, dos anos 2001 a 2002, resultaram em dados inéditos so-bre o mexilhão-dourado, antes nunca pesqui-sado no Brasil, com a instalação dos substratos artificiais ao longo do reservatório e coletas de zoobentos que mostraram a sua incrível rapidez no avanço da colonização (Fig. 17). A pesquisa com as larvas de mexilhão mostrou alguns pe-ríodos de picos de reprodução no reservatório de Itaipu e a pesquisa de zoobentos mostrou onde realmente os indivíduos de L. fortunei vi-vem no reservatório. A proliferação de L. fortunei deve afetar outras comunidades do reservatório de Itaipu, especialmente as espécies de Bivalvia nativas, sendo aconselhável dar continuidade as pesqui-sas para a obtenção de mais dados biológicos para poder sugerir um controle natural.
4. Agradecimentos
À Itaipu Binacional, pelo financiamento do projeto; ao NUPELIA, pelo apoio logístico; à pesquisadora Inês Ezcurra de Drago (INALI – Argentina), pelos ensinamentos sobre as larvas de mexilhão-dourado; ao Dr. José C. Stevaux, pelo cálculo do gradiente hidráulico e potência do fluxo das estações de coleta à jusante do re-servatório; ao motorista e técnico Celsão /UEM; aos funcionários da Itaipu Binacional, pelo apoio e amizade durante as longas e pesadas jornadas de coletas (mas sempre alegres!) e aos auxiliares contratados pelo projeto, via UEM.
Figura 17. Mapa com a localização dos substratos e registros mensais (maio/2001 a agosto/2002) no reservatório de Itaipu desde a primeira colonização de Limnoperna fortunei.


Seção
5Moluscos invasores e a comunidade bentônica


221
Ao analisarem a distribuição da fauna bentônica das represas Ibitinga, Bariri e Bar-ra Bonita do médio Rio Tietê, no período de novembro de 2002 a agosto de 2003, Suriani et al. (2007) verificaram que as espécies de mo-luscos invasores, Melanoides tuberculata (Mül-ler, 1774) e Corbicula fluminea (Müller, 1774), foram dominantes. França et al. (2007) reali-zaram no mesmo período, um levantamento, semelhante às autoras acima, para as represas do baixo Tietê, com resultados semelhantes em relação à distribuição e abundância dos mes-mos moluscos invasores. Ambos as publicações alertaram também sobre a eutrofização dos re-servatórios do trecho baixo e médio do Tietê, com altas concentrações de nutrientes (total e dissolvido). Em ambos os trabalhos, as autoras não registraram a presença da espécie invasora Li-mnoperna fortunei (Dunker, 1857), conhecida popularmente como o mexilhão-dourado. Ave-lar et al. (2004), registra pela primeira vez o mexilhão-dourado no estado de São Paulo em
novembro de 2002, em uma localidade do Rio Paranapanema, na confluência com o Rio Pa-raná, próxima à localidade de Rosana (22º31’S - 53º00’W). Darrigran & Mansur (2006) men-cionaram que no final de 2004, a espécie foi encontrada no Rio Tietê, na represa de Barra Bonita, já próximo à capital do estado de São Paulo. A exata chegada do mexilhão-dourado em Ibitinga não tem registro, porém estima--se ter sido em período anterior a esta data, pois Barra Bonita situa-se mais à montante no mesmo rio. Pareschi et al. (2008) calcularam a velocidade de invasão do mexilhão-dourado ao longo do alto Rio Paraná, desde a chegada ao reservatório de Itaipu, em 2001, até 2005, ano estimado da chegada a Ibitinga/Bariri. Os autores acreditam que a velocidade de invasão tenha sido na ordem de 264 km nestes 4 anos. Tundisi et al. (2008) alertaram sobre a vulnera-bilidade da bacia do Rio Tietê com presença do mexilhão-dourado, devido aos inúmeros danos que esta invasora tem produzido aos sistemas aquáticos, alterando canais, tubulações e estru-
CAPÍTULO 19
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré,
Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
Daniel PereiraPaulo Eduardo Aydos Bergonci
Cintia Pinheiro dos SantosVanessa Gazulha
Maria Cristina Dreher MansurCarlos Perez Bergmann
Juliane Vicenzi Silvio Santos

Pereira et al.
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
222
turas, com prejuízos para a sua manutenção, especialmente em represas. Os reservatórios no Brasil, especialmen-te no Estado de São Paulo, são importantes ecossistemas que foram construídos nos úl-timos 40 anos. Alguns destes reservatórios, principalmente os do sistema em cascata do Médio e Baixo Rio Tietê, encontram-se em estado eutrófico. Florações de cianobactérias são frequentemente relacionadas a estas con-dições eutróficas, o que compromete os usos múltiplos destes reservatórios, implicando em riscos à saúde humana e à biota aquática (Mi-nillo, 2005). Os maiores problemas estão rela-cionados à produção de toxinas. As toxinas das cianobactérias podem causar intoxicações em animais e seres humanos, e até mesmo a morte pela ingestão de água contaminada (Carmicha-el et al., 2001). Diversos estudos demonstram que estas toxinas afetam as comunidades aquá-ticas (Lampert, 1982; Demott, 1999). Durante projeto de P & D financiado pela AES Tietê e executado pela Fundação Luiz Englert, um diagnóstico da distribuição espacial do mexilhão-dourado e dos moluscos límnicos associados (nativos e invasores), em diferentes substratos, revelou algumas relações com o fitoplâncton, cianobactérias, microcisti-nas e a qualidade da água na sub-bacia do mé-dio Rio Tietê/Jacaré, especialmente nos reser-vatórios de Ibitinga e Bariri, no período de 12 a 14 de maio de 2008, em oito estações de coleta (Tab. I). A área pertence à sub-bacia do médio Rio Tietê/Jacaré, situado no centro do Estado de São Paulo (entre 49º32’ – 47º30’ longitude
e 21º37’ – 22º51’ de latitude) com uma área de drenagem de 11.749 km2. Este trecho engloba três rios principais, o Rio Tietê (em um total de 150 km da usina hidrelétrica (UHE) Barra Bonita até a UHE da Ibitinga), o Rio Jacaré--Guaçu e o Rio Jacaré Pepira. O relevo é variá-vel com a altitude máxima de 800 m. A área do Tietê/Jacaré localiza-se na Depressão Periférica do Estado de São Paulo, onde se encontram os aquíferos Bauru/Serra e Geral/Botucatu. Na re-gião predominam os solos de areias quartzosas profundas a moderadas e em menores quanti-dades o latossolo roxo eutrófico. O clima da região, segundo classificação de Koppen, situa--se entre clima tropical úmido (de outubro a março) e inverno seco (de abril a setembro). A disponibilidade hídrica anual está na ordem de 4.400 mm, com um período de precipita-ção mais intenso de outubro a março (verão) e um período de menor precipitação de abril a setembro (inverno) (Tundisi et al., 2008). Segundo CETESB (apud Suriani et al., 2007) a área apresenta “intensa atividade industrial com forte pressão antrópica”. Para um diagnóstico desta natureza dife-rentes técnicas são empregadas (Tab. II): coleta de três rochas em cada estação de coleta para a quantificação da abundância média do me-xilhão-dourado; três lançamentos (réplicas) de draga de Petersen (área: 10 x 10 cm) para coleta do mexilhão-dourado e demais moluscos ben-tônicos presentes no sedimento, sendo os resul-tados expressos em densidade média (ind./m2), considerando, em cada estação de coleta. Os moluscos associados à vegetação de macrófitas aquáticas são coletados por meio do exame de
Tabela I. Coordenadas geográficas das estações de coleta (E1 a E8) situadas na sub-bacia do médio Rio Tietê/Jacaré (São Paulo, Brasil).
Estações de coleta Coordenadas geográficas
E1 Canal lateral de deságue da eclusa a jusante da UHE Ibitinga (lótico) 21°45’24,67’’ S 48°59´38,05’’ W
E2 UHE Ibitinga próximo ao barramento (lêntico) 21°45’ 51,30’’ S 48°59’19,57’’ W
E3 Trecho inferior do rio Jacaré Guaçu (lótico) 21°49’28,55’’ S 48°49’53,13’’ W
E4 Trecho inferior do rio Jacaré Guaçu sob influência do lago da UHE Ibitinga (lêntico) 21°50’31,51’’ S 48°57’01,73’’ W
E5 Trecho inferior do rio Jacaré Pepira sob influência do reservatório (lêntico) 21°53’38,17’’ S 48°48’51,37’’ W
E6 Início do reservatório de Ibitinga (lêntico 22°01’01,26’’ S 48°52’54,14’’ W
E7 Rio Tietê à montante do reservatório de Ibitinga e jusante de Bariri (lótico)
22°08’43,16’’ S 48°45’04,70’’ W
E8 UHE Bariri próximo ao barramento (lêntico) 22°04’15,63’’ S 48°44’49,49’’ W

CAPÍTULO 19
Seção 5
223
raízes de macrófitas de acordo com o método descrito por Volkmer-Ribeiro et al. (1984); co-leta de larvas do mexilhão-dourado por meio de três réplicas de água filtrada (1000L), em rede de 30 μm, com auxílio de moto bomba. Amos-tragem do fitoplâncton através da passagem de frasco na superfície da água. As amostras (três réplicas) são acondicionadas em recipientes de 100mL e fixadas com formol a 4%. As análi-ses qualitativa e quantitativa são realizadas em Câmara de Sedgewick-Rafter sob microscópio óptico, com aumento de 200x. Os resultados são expressos em densidade, atingindo eficiên-cia amostral de 80%, de acordo com a seguinte fórmula (Pappas & Stormer, 1996):
Eficiência = [(nº de indivíduos - nº de táxons) / nº de indivíduos] x 100
Os táxons fitoplanctônicos considerados como dominantes são aqueles que ocorreram em densidades acima de 50% da densidade
total da alíquota analisada, enquanto os abun-dantes foram os que ocorreram acima do valor médio obtido ao dividir-se a densidade total da subamostra pelo número de táxons registrados na mesma (Lobo & Leighton, 1986). Além dos métodos de amostragem da bio-ta aquática, a avaliação da qualidade da água também se faz necessária. Para tal são obtidos dados de temperatura do ar e da água, bem como do oxigênio dissolvido e pH. Amostras de água também são coletadas e preservadas para a análise de sólidos suspensos totais, turbidez, condutividade, fósforo total, nitrogênio amo-niacal e total. Os métodos de coleta, preserva-ção e análise de água estão descritos em APHA (2004). Para a coleta de água para a determi-nação de microcistinas é necessário no mínimo réplicas através da passagem de frasco na super-fície da água. A determinação das concentrações de microcistinas se faz através do método ELISA (kit Beacon), que determinada à concentração total de microcistinas (intra e extracelular).
Figura 1. Estações de coleta (E1 a E8) si-tuadas na sub-bacia do médio Rio Tietê/Jacaré (São Paulo, Brasil): E1, canal lateral de deságue da eclusa a jusante da UHE Ibi-tinga (lótico); E2, reservatório UHE Ibitinga próximo ao barramento (lêntico); E3, tre-cho inferior do Rio Jacaré Guaçu (lótico); E4, trecho inferior do Rio Jacaré-Guaçu sob influência do lago da UHE Ibitinga (lêntico); E5, trecho inferior do Rio Jacaré Pepira sob influência do reservatório (lên-tico); E6, no início do reservatório de Ibi-tinga (lêntico), E7, Rio Tietê à montante do reservatório de Ibitinga e jusante de Bariri (lótico) e, E8, reservatório de Bariri próxi-mo ao barramento (lêntico). Mapa adapta-do de Tundisi et al. (2008) por D. Pereira.
Tabela II. Plano de amostragem utilizado nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil. Macrófitas aquáticas ausentes (ma); substrato rochoso (sr) impediu dragagem.
Plano de amostragem Estações de coleta Unidade
E1 E2 E3 E4 E5 E6 E7 E8Amostragem de sedimento (draga de Petersen)* sr sr X x x x x x ind.m-2
Amostragem de rochas x x X x x x x x ind.rochaExame de macrófita aquática (E. crassipes) ** ma x Ma x x x ma x ind./100gExame de macrófita aquática (Poacea) ** ma ma X ma ma ma x ma ind./100gAmostragem de larvas de L. fortunei*** x x X x x x x x ind.m-3
Amostragem de fitoplâncton/cianobactérias x x X x x x x x cél/mLAmostragem de água/cianotoxinas x x X x x x x x μg.L-1
Métodos de coleta e análise: *Mansur et al. (2003) e Takeda et al. (2005); **Volkmer-Ribeiro et al. (1984); ***Santos et al. (2005).

Pereira et al.
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
224
Para a análise estatística dos dados ob-tidos são realizados diferentes procedimen-tos: análise de correlação (Spearman’s rho), com finalidade de verificar relações espaciais entre o mexilhão-dourado e demais variáveis (moluscos límnicos, fitoplâncton e qualidade da água); análise de variância (ANOVA, Dun-can a posteriori, α=0,05), com a finalidade de verificar variação espacial; análises de corres-pondência canônica (CCA), seguida de teste de Monte Carlo através do programa PC-ORD (Mccune & Mefford, 1997), com a finalidade de detectar padrões espaciais e relações com as variáveis ambientais. No diagnóstico em questão se observa a total dominância das invasoras L fortunei (Fig. 2), seguida de Melanoides tuberculata, e Corbi-cula fluminea. Suriani et al. (2007) constatou a dominância do gastrópode invasor Melanoides tuberculata em elevadas densidades, anterior-mente à introdução de L. fortunei. Atualmente este gastrópode mantém densidades elevadas similares às verificadas pelas autoras (op. cit.), nos anos de 2002 e 2003, porém as densidades do mexilhão-dourado são mais elevadas. Corbicula fluminea predomina com maiores densidades nas estações E5 e E7 (Fig. 2), em densidades bem inferiores a de L. for-tunei e M. tuberculata. Suriani et al. (2007) também registraram a presença de C. flumi-nea em densidades relativamente baixas. Esta espécie requer substrato arenoso e águas com elevada saturação de oxigênio (Mansur et al., 1994). Segundo Boltovskoy et al. (1997) este C. fluminea é sensível à poluição orgânica. Com base nos dados logaritmizados de densidade média de moluscos encontrados no sedimento de fundo foram verificados alguns padrões de variação espacial da malacofauna. As estações de coleta não diferem estatisticamente (ANO-VA, Duncan a posteriori, α=0,05) quanto às densidades do táxon Pomacea sp.; as estações E5 e E7 (maiores densidades) diferem das es-tações E3, E6 e E8 (menores) quanto ao táxon Melanoides tuberculata; as estações de coleta E5 e E7 (maiores densidades) diferem das de-mais estações de coleta (menores) quanto ao táxon C. fluminea; as estações de coleta E4 e E8 (maiores densidades) diferem entre si e das demais das estações (menores) quanto ao táxon L. fortunei. O diagnóstico diverge dos resulta-
dos obtidos por Suriani et al. (2007) que regis-tra Aylacostoma tenuilabris (Bernardi, 1856), Biomphalaria glabrata (Say, 1818), Biomphala-ria intermedia (Paraense & Deslandes, 1962), Diplodon expansus (Küster, 1853), Physa cubensis (Pfeiffer, 1839) e Pomacea canalicula-ta (Lamarck, 1822). Não se verifica correlação significativa entre o mexilhão-dourado e os de-mais moluscos no sedimento. Os táxons de moluscos encontrados nos talos e raízes de macrófitas flutuantes do gê-nero Eichhornia na bacia do médio Rio Tietê/
Figura 2. Densidade populacional média de moluscos límnicos nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil A, no sedimento; B, densidade em raízes de macrófitas aquáticas. M. tuberculata (Mt), Pomacea sp. (Psp), Cochiliopidae (Co), Physa sp. (Ph), Biomphalaria sp. (Bi), Drepanotrema sp (Dr.), Antillorbis sp. (An), O. matheroni (Om), Eupera sp. (Eu), Pomacea sp. (Psp), C. fluminea (Cf) e L. fortunei (Li).

CAPÍTULO 19
Seção 5
225
Jacaré (Fig. 2B) são típicos deste tipo de subs-trato (Volkmer-Ribeiro et al., 1984; Pereira et al., 2000a, b; Pfeifer & Lopes-Pitoni, 2003; Abílio, et al., 2006; Arruda et al., 2009). Os talos (estolões) e as raízes do aguapé oferecem substrato, abrigo e disponibilizam alimento (perifíton) aos moluscos raspadores, no caso os gastrópodes. Já os bivalves filtradores, que vivem na macrófita, devem apresentar os fios de bisso para sua fixação, como é o caso de Eupera sp. e L. fortunei. Corbicula fluminea é um filtrador que se enterra no sedimento e, por não apresentar fios de bisso na fase adulta, não teria condições de utilizar a macrófita como substrato. A dominância da espécie bissada L. for-tunei no aguapé é notória sobre as demais, em especial à montante dos reservatórios (E2, E4, E5 e E8). L. fortunei têm sido registrado numa grande variedade de espécies e formas biológicas de macrófitas aquáticas (Mansur et al., 2003, 2004b). Em alguns casos, até colonizando espécies arbóreas marginais aos rios e lagos que não teriam capacidade de abrigar fauna de macroinvertebrados, como é o caso de dos galhos de sarandi, no Lago Guaíba (RS), oferecendo um novo micro-habitat para moluscos e crustáceos (Mansur et al., 2008c). É importante considerar que a invasora M. tuberculata dominava a região (Suriani et al., 2007), até a chegada do mexilhão-dourado, que no momento é o dominante na taxocenose. Com base nos dados logaritmizados de densidade média, em raízes de macrófitas aquá-ticas, os seguintes padrões de variação espacial da malacofauna são verificados: as estações de coleta não diferem estatisticamente (ANOVA, Duncan a posteriori, α=0,05) quanto aos tá-xons Melanoides tuberculata, Physa sp., Biom-phalaria sp.; Drepanotrema sp., Antillorbis sp., Eupera sp. e Cochiliopidae. No entanto, a es-tação E2 difere das demais por apresentar as densidades mais elevadas de L. fortunei em raiz de macrófita aquática. A estação de coleta E4 (maior densidade) difere das estações E3, E6, E7 e E8 (menor) quanto à densidade Poma-cea sp. É verificada correlação positiva entre o mexilhão-dourado e o gastrópode Physa sp. (r=0,413; p=0,023; Spearman’s rho). A relação entre a população desta invasora e demais com-ponentes da macrofauna é pouco conhecida e
merece ser elucidada em pesquisa destinada ao tema. Silva et al. (2007) também não verificou qualquer correlação entre as densidades do me-xilhão-dourado e dos demais moluscos límni-cos associados no aguapé (Eichornia crassipes), em lagos do Rio Paraguai. A densidade larval média é de 1,79±0,74 ind.m-3, variando de zero a 5,33 (Fig. 3). Os maiores valores de densidade larval (ind.m-3) ocorrem à jusante da UHE Ibitinga (E1) e à ju-sante da UHE Bariri (E7). De todos os locais amostrados, na estação de coleta situada no Rio Jacaré Pepira (E5) as larvas não são detec-tadas, porém foi registrada a presença de indi-víduos adultos neste local. As estações E1 e E7 diferem estatistica-mente (ANOVA, Duncan a posteriori, α =0,05) das demais estações, quanto à densidade média de larvas logaritmizada. A densidade larval ob-servada no período amostrado é relativamente baixa quando comparada de uma maneira ge-ral com resultados obtidos por Santos (2004) e Santos et al. (2008) para o Lago Guaíba no Rio Grande do Sul e Pestana et al. (2008), no Rio Paraná, junto à foz do Rio Iguaçu. Segundo Morton (1982), a espécie apresenta pulsos re-produtivos que coincidem mais ou menos com as temperaturas da água, mais altas e mais bai-xas. De acordo com Darrigran et al. (2002), as baixas temperaturas inibem a liberação de lar-vas do mexilhão-dourado. Boltovskoy & Catal-do (1999) observaram que a densidade larval no plâncton junto ao baixo Rio Paraná acon-tece de setembro a março e que os pulsos não
Figura 3. Densidade média, erro padrão, mínimo e máximo (ind.m-3) de larvas de Limnoperna fortunei no plâncton (moni-toramento), nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.

Pereira et al.
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
226
são bem evidentes no verão, mas as temperatu-ras mais baixas tornam a atividade reprodutiva descontínua. Na região amostrada, na época do diagnóstico, não foi possível um monitoramento larval periódico para a verificação dos pulsos de densidade larval, mas as datas de coleta coinci-dem com um período de temperaturas de outo-no (mês de maio), mais amenas, o que poderia justificar a baixa densidade encontrada. Outros fatores ambientais que poderiam influenciar na densidade do mexilhão seriam o tipo de substrato e a correnteza. O mexilhão--dourado prefere substratos duros e águas bem oxigenadas, onde costuma se avolumar (Morton, 1982). A maior ocorrência de larvas nos locais com maior movimentação das águas (jusantes dos barramentos Ibitinga e Bariri) confere com os dados observados por Bergonci et al. (2009) durante um ano de coletas mensais, que referem a maior densidade larval encontrada na margem do canal principal do Rio Jacuí em relação ao cais do porto de Porto Alegre, Delta do Jacuí, Rio Grande do Sul. Os ambientes juntos às es-tações E3 (Jacaré-Guaçu), ambiente lótico, e E5 (Jacaré Pepira), apresentaram-se compostos por sedimentos lodosos e pouco consistentes, não apresentando condições ideais para a fixação e sobrevivência do molusco e consequentemente acarretando baixa produção larval. A comunidade fitoplanctônica está repre-sentada por 43 táxons, identificados em nível
específico, na sua maioria. Estes táxons encon-tram-se distribuídos em 6 classes: Chlorophyce-ae, Zygnematophyceae, Cyanophyceae, Dino-phyceae, Euglenophyceae e Bacillariophyceae (Tab. V). Os grupos fitoplanctônicos que apresen-tam maior riqueza específica, em ordem decres-cente, são Cyanophyta (Cyanophyceae), Hete-rokontophyta (Bacillariophyceae) e Chlorophyta (Chlorophyceae) (Fig. 4). Os grupos Dynophyta (Dinophyceae) e Euglenophyta (Euglenophyce-ae) têm baixa representatividade em termos de riqueza, ocorrendo apenas na E4, com 01 espé-cie cada. De um modo geral, a comunidade fito-planctônica demonstra baixa riqueza específica nas diferentes estações amostradas. As cianobactérias apresentam as maio-res densidades em todas as estações de coleta da sub-bacia do médio Rio Tietê/Jacaré (Figs. 5 e 6). A E1 destaca-se por apresentar densi-dades muito elevadas em relação às demais (3738,9 cél.ml-1). As estações de coleta E2, E4 e E8 apresentam densidades intermediárias. As densidades mais baixas são registradas nas estações 5 (0,2 cél.ml-1) e 6 (1,3 cél.ml-1), sen-do a abundância relativa das cianobactérias próxima de 100 % na maioria das estações de coleta (Fig. 7). Nas estações E5 e E6, foi re-gistrada maior distribuição de densidades entre os grupos Cyanophyta, Heterokontophyta e Chlorophyta, ainda assim, com o predomínio
Figura 4. Riqueza específica dos grupos fitoplanctônicos (n° espécies) nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.

CAPÍTULO 19
Seção 5
227
Tabela III. Composição de espécies do fitoplâncton/cianobactérias nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.
E1 E2 E3 E4 E5 E6 E7 E8CHLOROPHYTA Chlorophycea Chlorella sp. x Kirchneriella lunaris x Monoraphidium sp. x x Oedogonium sp. x Pediastrum duplex x x x Scenedesmus obliquus x Scenedesmus quadricauda x Sphaerocystis sp. x x x x x x Volvox aureus xZygnematophyceae Cosmarium contractum x Cosmarium punctulatum x x Closterium sp. x Eudorina elegans x x Micrasterias sp. x Staurastrum gracile x x x x x Staurodesmus sp. xCYANOPHYTA Cyanophyceae Anabaena cf. crassa x x x x x Aphanizomenon cf. gracile x x x x Aphanocapsa cf. incerta x x x x Aphanothece minutissima x x Chroococcus sp. x x cf. Cylindrospermopsis raciborskii x cf. Gomphosphaerium x cf. Heteroleibleinia x Lyngbya sp. x x x x x Microcystis aeruginosa x x x x x x x Microcystis cf. botrys x x x Microcystis cf. novacekii x x x x Microcystis cf. protocystis x x x x x Oscillatoria sp. x x x x Planktothrix sp. x Synechococcus cf. elongatus x x x x x x xDINOPHYTA Dinophyceae Dinoflagelado 1 xEUGLENOPHYTA Euglenophyceae Phacus sp. xHETEROKONTOPHYTA Bacillariophyceae Asterionella formosa x x x x Aulacoseira granulata x x x x Cyclotella sp. x Gomphonema sp. x x x x Melosira varians x x x x x x Navicula sp. x x x x x Nitzschia palea x x x x x x x Pinnularia gibba x x Pleurosira laevis x x Surirella tenera x x x Synedra sp. x x x Tabellaria sp. x x Diatomácea 1 x
Pereira et al.
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
228
do grupo das cianobactérias. No estado de São Paulo, desde o final da década de 1990, tem sido constatada a ocorrência frequente de florações de cianobactérias nos reservatórios do Rio Tie-tê, acompanhadas de consideráveis mudanças na composição e estrutura das comunidades planc-tônicas (Calijuri et al., 2002; Matsumura-Tundi-si et al., 2006). Estudos recentes revelam a do-minância das espécies dos gêneros Microcystis e Anabaena nos primeiros reservatórios do médio
Tietê e aquelas dos gêneros Cylindrospermopsis e Raphidiopsis nos reservatórios do baixo Rio Tietê (Takenaka, 2007). Outro estudo, no mé-dio e baixo Tietê, registrou que as cianofíceas ocorreram em maior frequência e abundância em relação às demais classes fitoplanctônicas, com a presença de espécies tóxicas dos gêneros Microcystis, Anabaena, Cylindrospermopsis, Ra-phidiopsis e Planktothrix em todas as amostras analisadas (Minillo, 2005).
Figura 5. Densidade absoluta (cél.mL-1) dos grupos fitoplanctônicos nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.
Figura 6. Abundância relativa (%) dos grupos fitoplanctônicos nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.

CAPÍTULO 19
Seção 5
229
A análise quali-quantitativa da comuni-dade fitoplanctônica permite identificar as es-pécies dominantes e abundantes ocorrentes nos reservatórios de Ibitinga e Bariri, nas diferentes estações de coleta (Fig. 8). As espécies domi-nantes registradas são Microcystis aeruginosa, Oscillatoria sp. e Synechococcus cf. elongatus, todas pertencentes às cianobactérias. A espé-cie M. aeruginosa é a única espécie dominante nas estações de coleta E1, E2, E6, E7 e E8. A espécie Oscillatoria sp. foi dominante nas esta-ções de coleta E3 e E4, sendo que nesta última, M. aeruginosa é registrada como abundante. E a estação de coleta E5, teve a presença de S. cf. elongatus como espécie dominante. A análise da variação espacial da comu-nidade fitoplanctônica permite identificar a ocorrência predominante das cianobactérias nos Reservatórios de Ibitinga e Bariri, tanto em riqueza, quanto em densidade. A dominância das cianobactérias demonstrou o grau de eu-trofização do reservatório e a possibilidade da ocorrência de florações produtoras de toxinas. A ocorrência de florações está mais relacionada aos períodos de verão e primavera, ocorrendo em menores densidades nos períodos de outo-no e inverno. Com base na análise de correlação bi-variada (Spearman’s rho) são verificadas as seguintes correlações significativas entre o mexilhão-dourado e o fitoplâncton: densidade
populacional nas macrófitas (recrutas e adul-tos) versus Microcystis (r=0,522; p=0,026); versus Chlorophyta (r=0,839; p<0,0001); versus Cyanophyta (r=0,812; p<0,0001); ver-sus Dynophyta (r=0,536; p=0,022); densida-de populacional no bentos (sedimento) versus Chlorophyta (r=0,490; p=0,024), versus Cya-nophyta (r=0,570; p=0,007); Heterokonto-phyta (r=0,734; p<0,0001); densidade popu-lacional larvas no plâncton versus Cyanophyta (r=0,436; p=0,033); versus Heterokontophyta (r=0,483; p=0,017). As relações entre o fitoplâncton e o me-xilhão-dourado são pouco conhecidas. Este bivalve filtrador, como qualquer outro bivalve, utiliza o material em suspensão na água (par-ticulado orgânico, plâncton, etc.), em sua ali-mentação. No entanto pouco se sabe sobre a composição de sua dieta e sobre o seu papel como consumidor na cadeia trófica. Em con-dições de laboratório, Silva (2006) observou que em contato com alimentação adequada (Chlorella sp., Chlorophyta; Spirulina sp., Cya-nophyta; e Scenedesmus sp., Chlorophyta), L. fortunei pode aumentar sua massa corpórea em até 240% em 75 dias, enquanto que, espécimes sem alimento podem resistir até 63 dias sem incremento de peso. Provavelmente, as diferen-tes fases de vida do mexilhão selecionem com-ponentes diferenciados do plâncton, quanto ao tamanho e composição nutricional, em sua ali-
Figura 7. Espécies do fitoplâncton dominantes/abundantes (abundância relativa %) nas estações de coleta distribuídas na sub--bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.

Pereira et al.
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
230
mentação, de acordo com a disponibilidade no ambiente e nos micro-habitats. A Tabela III apresenta o enquadramento da água em classes de qualidade conforme CO-NAMA (2005), resolução nº 357. Os valores verificados de fósforo total no presente estudo são comparáveis aos valores apresentados por Thomaz (2006), para o trecho do Rio Tietê en-tre a UHE Ibitinga e Bariri. De modo geral, a qualidade da água é pior à montante de Ibitin-ga, Estações de E4 a E8. A concentração de microcistinas na água dos reservatórios estudados varia de 0 a 1,8 μg.L-1 (Fig. 8) e podem ser consideradas baixas, visto que a concentração de cianobactérias no período amostrado também foi baixa, não sen-do registrado nenhum evento de floração. As concentrações de microcistinas nas florações de cianobactérias nos reservatórios do Tietê po-dem chegar em torno de 138 μg.g-1 de material algal liofilizado, e seus extratos brutos podem ser tóxicos a cladóceros e camundongos (Ro-cha, 2008). Estudo recente no baixo e médio Tietê, detectou microcistinas na maioria dos reservatórios, com concentrações elevadas que variaram de 2,17 a 150 μg.L-1 (Minillo, 2005). A toxicidade das florações do Tietê para a bio-ta vem sendo observada através de bioensaios com camundongos e cladóceros (Sotero-Santos et al., 2006; Takenaka et al., 2007). A toxicida-de destas florações para o mexilhão-dourado é ainda desconhecida. A cianobactéria M. aeru-ginosa foi a espécie dominante nas estações de coleta em que se detectou microcistinas. Esta
espécie é de ampla distribuição, e reconheci-damente produtora de microcistinas, sendo assim a principal responsável pelas toxinas na água dos reservatórios, no período amostrado. É uma cianobactéria comum nos reservatórios tropicais eutróficos e frequentemente responsá-vel pelas florações, sendo também uma indica-dora de baixa qualidade da água. A estação de coleta 3 teve a cianobac-téria S. cf. elongatus como dominante. Esta espécie é reconhecida como não produtora de toxinas, o que justifica o resultado obtido para esta estação (Fig. 8). Já a estação de co-leta E5, foi caracterizada pela dominância de Oscillatoria sp. Esta cianobactéria não pro-duz microcistinas, corroborando o resultado para esta estação de coleta. A Oscillatoria é produtora de anatoxina-a, uma neurotoxina, que não foi avaliada no presente estudo. Estudos no Tietê demonstraram a ocorrência de diversas cianotoxinas, principalmente para as neurotoxinas, com a presença de diferentes variantes no grupo das toxinas paralíticas: saxitoxinas, neosaxitoxinas e goniautoxinas (Takenaka, 2007). Desta forma, salienta-se a importância de avaliar a concentração de neurotoxinas em pesquisas futuras nestes reservatórios, devido à ocorrência do gênero como dominante. Com base na análise de correlação bi-variada (Spearman’s rho) foram verificadas as seguintes correlações positivas entre o mexi-lhão-dourado e as variáveis da qualidade da água (Tab. IV): densidade populacional nas
Figura 8. Concentração de microcistinas (µg.L-1) nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil.

CAPÍTULO 19
Seção 5
231
macrófitas (recrutas e adultos) versus condu-tividade (r=0,436; p=0,048); versus nitrato (r=-0,471; p=0,031); versus sólidos suspensos totais (r=-0,543; p=0,011); versus turbidez (r=0,564; p=0,008); densidade populacio-nal no bentos (recrutas e adultos) versus con-dutividade (r=0,524; p=0,026); versus pH (r=0,635; p=0,05); versus turbidez (r=-0,728; p=0,01); versus temperatura da água (r=0,532; p=0023); larvas no plâncton versus turbidez (r=-0,447; p=0,029); versus nitrogênio amo-niacal (r=-0,525; p=0,008). O teor de sólidos totais suspensos é um fator determinante no processo de filtração dos bivalves límnicos, auxiliando na incorporação de metais presentes no meio aquático (Callil & Junk, 1999). Em parte, as correlações verifica-das entre as densidades do mexilhão-dourado no plâncton, macrófitas e bentos (sedimento) corroboram estudo anterior (Mansur et al., 2008c) referente ao monitoramento (variação espacial e temporal) do mexilhão-dourado e variáveis ambientais no delta do Rio Jacuí, Rio Grande do Sul, que demonstrou: correla-ção positiva e significativa para os mexilhões incrustados em substrato artificial versus con-dutividade; correlação negativa e significativa para as larvas de mexilhões no plâncton ver-sus nitrogênio amoniacal. Cabe ressaltar que a presente análise é de ordem espacial. O mo-nitoramento continuado desta bacia poderia explicar melhor a relação entre a dinâmica po-pulacional do mexilhão-dourado e as variáveis ambientais. Por meio da CCA (Fig. 9), os eixos 1 (50,7%) e 2 (16,9%) respondem 67,5% da va-riância dos dados de densidade larval do me-xilhão-dourado, fitoplâncton, cianobactérias e cianotoxinas ao longo das estações de coleta na bacia do Rio Tietê. Esta ordenação demonstra um claro gradiente espacial que distingue as es-tações de coleta 1 e 2 das demais, assim como dentre as demais, pode ser verificado a diferen-ciação das estações 3 e 4, das estações de coleta 5, 6, 7 e 8. As estações 5 a 8 representam espécies fitoplanctônicas de ocorrência exclusiva nes-tes locais, com baixa densidade e representa-tividade. Estas estações localizam-se mais à montante recebendo, portanto, menor carga de nutrientes. O suprimento de nutrientes, espe-
cialmente o fósforo, é fundamental, pois regula o crescimento do fitoplâncton. Nas Estações de Coleta 5 e 6, o mexilhão-dourado está presente no bentos, tanto sobre substratos duros, quanto no aguapé. No entanto, a densidade de larvas foi muito baixa. A qualidade da água é com-prometida pela baixa concentração de oxigênio dissolvido, principalmente na estação 6. As es-tações 7 e 8 têm qualidade de água semelhante, especialmente baixa concentração de oxigênio dissolvido e elevada condutividade. Na E7 a densidade larval é elevada. As estações de cole-ta 1 e 2 apresentam baixa riqueza fitoplanctô-nica e densidades mais elevadas, com a domi-nância da cianobactéria M. aeruginosa. Na E1, as diatomáceas tiveram sua maior representati-vidade em termos de densidade. Nas Estações de coleta 3 e 4 ocorre dominância da ciano-bactéria Osillatoria sp. Na E4, as clorofíceas têm sua maior representatividade em termos de densidade. O Teste de Monte Carlo demonstra correlação alta (Eixo 1, r=0,99 e p=0,01; Eixo 2, r=0,94 e p=0,01) e significativa entre matri-zes de densidades de organismos e variáveis de qualidade da água (variação espacial). As áreas que apresentam maior densida-de larval, de recrutas e adultos estão próximas e à montante das represas. O mexilhão-dourado demonstra alta capacidade invasiva e dominân-cia na área. Comparando os dados de densida-de levantada em períodos anteriores observa-se que o gastrópode invasor Melanoides tubercu-lata que predominava em abundância na área, aparentemente ainda não sofreu redução em sua população, porém é superado em muitas vezes pela do mexilhão-dourado. No diagnós-tico apresentado, as amostragens quantitativas não revelam espécies de bivalves nativos. Um único exemplar de Diplodon paulista se encon-trava nas margens do reservatório. O bivalve invasor C. fluminea, por preferir um substrato diferente ao do mexilhão-dourado, qual seja, o sedimento arenoso do fundo no qual se en-terra, provavelmente não esteja ainda sofrendo com a presença competitiva do mexilhão-dou-rado. A relação entre as densidades do mexi-lhão (larvas e adultos) e os grandes grupos de táxons que compõem o fitoplâncton, indicam possíveis e estreitas relações tróficas entre estes organismos, que atuam nos processos ecológi-cos do meio aquático. Sugere-se para pesquisas
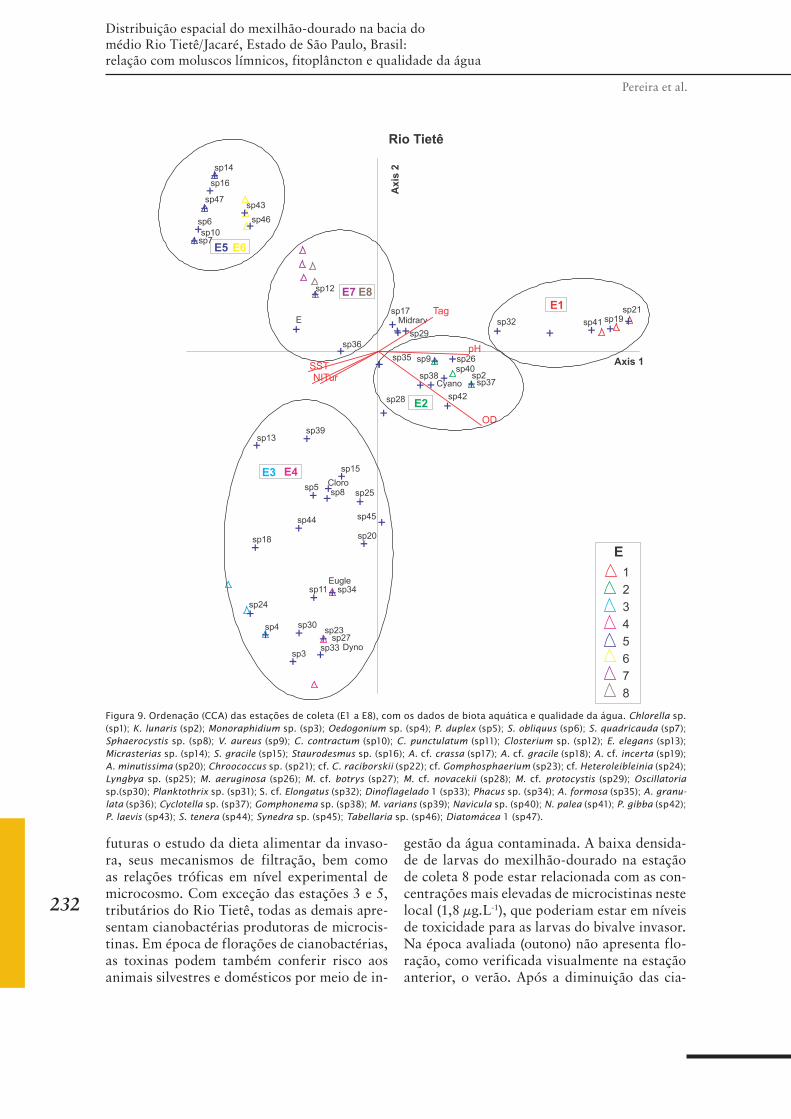
Pereira et al.
Distribuição espacial do mexilhão-dourado na bacia do médio Rio Tietê/Jacaré, Estado de São Paulo, Brasil: relação com moluscos límnicos, fitoplâncton e qualidade da água
232
futuras o estudo da dieta alimentar da invaso-ra, seus mecanismos de filtração, bem como as relações tróficas em nível experimental de microcosmo. Com exceção das estações 3 e 5, tributários do Rio Tietê, todas as demais apre-sentam cianobactérias produtoras de microcis-tinas. Em época de florações de cianobactérias, as toxinas podem também conferir risco aos animais silvestres e domésticos por meio de in-
Figura 9. Ordenação (CCA) das estações de coleta (E1 a E8), com os dados de biota aquática e qualidade da água. Chlorella sp. (sp1); K. lunaris (sp2); Monoraphidium sp. (sp3); Oedogonium sp. (sp4); P. duplex (sp5); S. obliquus (sp6); S. quadricauda (sp7); Sphaerocystis sp. (sp8); V. aureus (sp9); C. contractum (sp10); C. punctulatum (sp11); Closterium sp. (sp12); E. elegans (sp13); Micrasterias sp. (sp14); S. gracile (sp15); Staurodesmus sp. (sp16); A. cf. crassa (sp17); A. cf. gracile (sp18); A. cf. incerta (sp19); A. minutissima (sp20); Chroococcus sp. (sp21); cf. C. raciborskii (sp22); cf. Gomphosphaerium (sp23); cf. Heteroleibleinia (sp24); Lyngbya sp. (sp25); M. aeruginosa (sp26); M. cf. botrys (sp27); M. cf. novacekii (sp28); M. cf. protocystis (sp29); Oscillatoria sp.(sp30); Planktothrix sp. (sp31); S. cf. Elongatus (sp32); Dinoflagelado 1 (sp33); Phacus sp. (sp34); A. formosa (sp35); A. granu-lata (sp36); Cyclotella sp. (sp37); Gomphonema sp. (sp38); M. varians (sp39); Navicula sp. (sp40); N. palea (sp41); P. gibba (sp42); P. laevis (sp43); S. tenera (sp44); Synedra sp. (sp45); Tabellaria sp. (sp46); Diatomácea 1 (sp47).
gestão da água contaminada. A baixa densida-de de larvas do mexilhão-dourado na estação de coleta 8 pode estar relacionada com as con-centrações mais elevadas de microcistinas neste local (1,8 μg.L-1), que poderiam estar em níveis de toxicidade para as larvas do bivalve invasor. Na época avaliada (outono) não apresenta flo-ração, como verificada visualmente na estação anterior, o verão. Após a diminuição das cia-

CAPÍTULO 19
Seção 5
233
nobactérias, os mexilhões recrutaram todos os substratos naturais da área. As macrófitas con-sistem num estoque de mexilhões recrutas com potencial de incrustação. Fica recomendado o Tabela IV. Qualidade da água nas estações de coleta distribuídas na sub-bacia do médio Rio Tietê/Jacaré, estado de São Paulo, Brasil. Classes de qualidade da água (C1, C2 e C3) conforme resolução 357 do CONAMA.
Variáveis Estações de Coleta
E1 E2 E3 E4 E5 E6 E7 E8
Temperatura do ar (°C) 22 18 20 20 25 18 23 23
Temperatura da água (°C) 22 22 22 20 22 21 22 22
Oxigênio Dissolvido6.14 (C1)
6.18 (C1)
6.47 (C1)
4.12 (C3)
3.8 (C3)
3.66 (C3)
4.68 (C3)
3.88 (C3)
pH 7.19 (C1)
7.11 (C1)
7.45 (C1)
6.87 (C1)
6.67 (C1)
7.32 (C1)
7.22 (C1)
7.25 (C1)
Sólidos Suspensos Totais (mg/L) 1 1 1 24 2 5 7 11
Condutividade 140 156 147 49 35 168 143 173
Turbidez (UTN) 1.58 (C1)
1.75 (C1)
2.00 (C1)
11.58 (C1)
6.08 (C1)
3.66 (C1)
2.22 (C1)
2.96 (C1)
Fósforo total (mg/L)0.022 (C1)
0.022 (C2)
0.01 (C1)
0.126 (C2)
0.01 (C1)
0.033 (C1)
0.038 (C1)
0.048 (C3)
Nitrogênio Amoniacal (mg/L) 0.1 0.2 0.1 0.2 0.63 0.1 0.1 0.1
Nitrogênio Total (mg/L) 2.0 (C1)
2.0 (C1)
2.0(C1)
4.0 (C1)
2.0 (C1)
2.6 (C1)
2.5 (C1)
2.5 (C1)
Enquadramento (C1) (C2) (C1) (C3) (C3) (C3) (C3) (C3)
monitoramento permanente do mexilhão-dou-rado e de cianobactérias no trecho estudado, com a finalidade de subsidiar o manejo das po-pulações do invasor e das cianobactérias.


235
Ao longo dos vinte anos de invasão de Limnoperna fortunei (Dunker, 1857) na Amé-rica do Sul, muitos esforços têm sido emprega-dos para monitorar a dispersão (Pastorino et al., 1993; Mansur et al., 2003; Darrigran & Damborenea, 2005; Darrigran et al., 2007; Oliveira, 2009) e os impactos causados nos di-ferentes ecossistemas invadidos (Boltovskoy et al., 2006; Sylvester et al., 2007b; Sardiña et al., 2008; Marçal & Callil, 2008 e Darrigran & Damborenea, 2011). A magnitude dos impactos do mexi-lhão-dourado no ambiente está relacionada com o tamanho populacional e a suscetibilidade do sistema à invasão (Hicks, 2004). As densida-des são altas no início da invasão e, com o tem-po, vão reduzindo até alcançar um equilíbrio estável (Mansur et al., 2003). A atividade de filtração exercida por L. fortunei afeta direta-mente atributos físicos da coluna d’água como fluxo (Boltovskoy et al., 2006), transparência e penetração de luz (Sylvester et al., 2005; Bol-tovskoy et al., 2009b). O sedimento recebe um
aporte de matéria orgânica proveniente da ati-vidade de filtração e deposição de partículas (Ricciardi et al., 1997; Boltovskoy et al., 2006, 2009b) e os diferentes substratos ocupados por L. fortunei tornam-se mais heterogêneos, atra-vés da criação de novos micro-habitats inters-ticiais entre as valvas, possibilitando proteção contra predadores e maximização da utilização do espaço (Sylvester et al., 2007b; Sardiña et al., 2008). As modificações físicas observadas na coluna d’água, no sedimento e nos substratos afetam indiretamente as comunidades biológi-cas (Darrigran & Damborenea, 2011). Cataldo & Boltovskoy (2000) relatam que a comunida-de de algas pode ser afetada de forma positi-va ou negativa. A maior penetração de luz na coluna d’água pode promover um aumento na densidade de populações fitoplanctônicas e no crescimento de plantas submersas (Boltovskoy et al., 2009b). Por outro lado, a atividade de filtração de densas populações de L. fortunei pode também diminuir a quantidade de fito-
CAPÍTULO 20
Efeito do macrofouling sobre a comunidade
de invertebrados aquáticos
Vera UhdeEdson V. Massoli Jr.
Claudia T. Callil

Uhde et al.
Efeito do macrofouling sobre a comunidade de invertebrados aquáticos
236
plâncton na coluna d’água que pode ser con-vertido em biomassa ou ainda ser depositado no substrato por meio das fezes e pseudofezes (Fig. 1). A introdução de L. fortunei no Rio Pa-raguai ocorreu em 1999, através da navegação fluvial, que age como dispersor de adultos e larvas planctônicas em rios e lagos no Pantanal (Mansur et al., 2004c; Oliveira et al., 2006). Por conta da pouca disponibilidade de subs-tratos duros no Pantanal, o mexilhão-dourado utiliza raízes e estolões de macrófitas aquáticas para se fixar (Marçal & Callil, 2008). A arqui-tetura e o habitat formado pelas raízes das ma-crófitas aquáticas favorecem o crescimento de perifíton (Toft et al., 2003; Poi de Neiff & Neiff, 2006) e aumentam a complexidade estrutural na região litorânea de lagoas, que abriga os in-vertebrados nativos e o bioinvasor L. fortunei (Oliveira et al., 2006). O interesse em entender o efeito da pre-sença de L. fortunei na estrutura da comuni-dade de invertebrados, associado a macrófitas aquáticas flutuantes no Pantanal, motivou o desenvolvimento do estudo de Marçal & Callil (2008) em lagos rasos adjacentes ao Rio Para-guai, MT. Desta maneira, discutiremos os re-sultados de Marçal & Callil (2008) juntamente com Darrigran et al. (1998a) referentes ao efeito
de L. fortunei sobre a comunidade de inverte-brados aquáticos. Traçaremos um paralelo com os dados disponíveis na literatura para Dreisse-na polymorpha, uma vez que, as informações disponíveis para o mexilhão-zebra podem ser utilizadas para prever impactos causados pelo mexilhão-dourado. Embora os mecanismos impostos por cada espécie sejam similares, o resultado final dentro da comunidade pode ser bem diferente, dependendo das restrições do ecossistema e das espécies que compõe a comu-nidade (Boltovskoy et al., 2006).
Interferência do macrofouling na comunidade de invertebrados aquáticos
Nematoda
Nematódeos de vida livre são muito abundantes e diversos nos sistemas aquáticos, além de estarem presentes em todos os tipos de habitats límnicos (Abebe et al., 2008). Em locais com presença do mexilhão-dourado este grupo é beneficiado e aumenta sua densidade, como observado por Darrigran et al. (1998a) e Marçal & Callil (2008) (Fig. 2A). O mesmo efeito positivo sobre os nematódeos também pode ser observado em locais invadidos pelo
Figura 1. Fluxograma ilustran-do os efeitos de L. fortunei sobre as comunidades aquá-ticas na coluna d’água e no sedimento.

CAPÍTULO 20
Seção 5
237
mexilhão-zebra (Dreissena polymorpha) (Zai-ko et al., 2009) (Fig. 2A). Este aumento signi-ficativo na densidade média dos nematódeos pode estar relacionado à maior quantidade de matéria orgânica e bactérias associadas às fezes e pseudofezes depositadas pelos mexilhões so-bre a superfície do sedimento (Dermott & Ke-rec, 1997), além de outros fatores específicos inerentes a cada local invadido (Radziejewska et al., 2009).
Annelida
Oligochaeta
A Classe Oligochaeta, juntamente com as larvas de Chironomidae, são os principais componentes da fauna de invertebrados, em diferentes tipos de habitats que compõe as co-munidades bentônicas em ambientes lênticos e lóticos (Harman, 1982). No Rio de La Plata, Argentina, a den-sidade média de Oligochaetas aumentou sig-nificativamente com a presença de L. fortunei (Darrigran et al., 1998a), enquanto que em la-goas marginais ao Rio Paraguai nenhum efeito foi observado, a densidade média de Oligocha-etas entre locais invadidos não apresentou uma variação expressiva (Marçal & Callil, 2008) (Fig. 2B). Estudos realizados com Dreissena polymorpha demonstram relações positivas (Atalah et al., 2010; Ricciardi et al., 1997) e neutras (Zaiko et al., 2009) do mexilhão-zebra com a densidade de Oligochaeta (Fig. 2B). O efeito positivo do mexilhão-dourado sobre a densidade de Oligochaetas é atribuído à biodeposição de nutrientes através das fezes e pseudofezes (Ricciardi et al., 1997). A ativida-de de filtração remove grandes quantidades de matéria orgânica particulada da coluna d’água e as depositam no sedimento, aumentando a fon-te de alimentação para Oligochaetas (Sephton et al., 1980; Gonzalez & Downing, 1999). Outro fator que contribui para favorecimento dos Oligochaetas é a baixa disponibilidade de oxigênio nos espaços intersticiais das conchas, devido à decomposição bacteriana dos biode-pósitos (Darrigran & Damborenea, 2011). Estudos que demonstraram relações neutras entre a presença do invasor e a densi-dade de Oligochaetas atribuem esse resultado
ao comportamento específico de cada espécie (Karatayev et al., 2010). Influências antropo-gênicas também podem afetar a densidade de Oligochaetas. Nalepa et al. (2003) argumen-tam que os efeitos do mexilhão-zebra nos Oli-gochaetas são insignificantes em comparação com os efeitos do nível de fósforo na água.
Hirudinea
Hirudinea é uma classe representante do filo Annelida, formada predominantemente por invertebrados de água doce, que podem ser encontrados em todo o mundo (Klemm, 1972). Nos locais invadidos pelo mexilhão-dourado e mexilhão-zebra a densidade média dos hirudí-neos aumenta (Fig. 2C). Esse efeito está cor-relacionado com o aumento da abundância de invertebrados nas agregações de mexilhões, o que gera, consequentemente, o aumento da dis-ponibilidade alimentar das espécies carnívoras, como os hirudineos (Zaiko et al., 2009).
Mollusca
Gastropoda
Os gastrópodes correspondem à maior e mais bem sucedida classe dentro do filo Mollus-ca, principalmente pela grande variedade de habitats que ocupam e papéis funcionais que desempenham (Meier-Brook, 1984). Em locais invadidos por mexilhão-dourado foram obser-vados diferentes efeitos sobre a densidade da classe Gastropoda. Darrigran et al. (1998a), em estudo realizado no Rio de La Plata, observou uma relação positiva entre mexilhão-dourado e gastrópodes. Já Marçal & Callil (2008) no Pan-tanal, demonstraram uma relação negativa. Em estudos realizados com o mexilhão-zebra, tam-bém foram registrados efeitos positivos (Hor-vath et al., 1999; Mörtl & Rothhaupt, 2003) e negativos (Zaiko et al., 2009) (Fig. 2D). Os diferentes efeitos nas densidades, tanto na presença de mexilhão-dourado como para mexilhão-zebra, podem estar relaciona-dos com os papéis funcionais de cada espécie de gastrópode. Physella sp. (Pulmonata; Phy-sidae) alimenta-se principalmente como um raspador, sendo beneficiada com a presença do mexilhão, que estimula o crescimento de

Uhde et al.
Efeito do macrofouling sobre a comunidade de invertebrados aquáticos
238
algas através de claridade da água. Darrigran et al. (1998a) também registraram que algumas espécies endêmicas, como Gundlachia concen-trica (Ancylidae) e Chilina fluminea (Chilini-dae), que eram abundantes antes da invasão, diminuiram em números ao longo do tempo, enquanto Heleobia piscium (Cochiolidae) au-mentou em proporção direta com a densidade de L. fortunei.
Bivalvia
Em relação aos demais bivalves, nas la-goas marginais ao Rio Paraguai invadidas por L. Fortunei, a densidade apresentou um discre-to aumento (Fig. 2E). Em locais invadidos por Dreissena polymorpha, podemos observar que ocorreu tanto efeito positivo como negativo sobre outros bivalves (Fig. 2E). A redução na abundância de bivalves tem sido geralmente re-gistrada (Ward & Ricciardi, 2007; Zaiko et al., 2009) na presença de mexilhões-zebra. Segun-do Karatayev et al. (2007a) os unionoides liga-dos com mexilhão-zebra podem ter dificulda-de de enterrar e movimentar a concha através do sedimento ou, ainda, abrirem suas conchas para a respiração, alimentação e reprodução. A ligação dos mexilhões pode impedir os Unio-noides de abrir suas conchas para a respiração, alimentação e reprodução.
Crustacea
Os microcrustáceos planctônicos são re-presentados neste trabalho pelos grupos Am-phipoda, Cladocera, Copepoda e Isopoda. A presença de L. fortunei provocou apenas uma leve redução na densidade destes microcrustá-ceos (Fig. 3A), enquanto que, na presença de Dreissena polymorpha, ocorreu sempre um efeito positivo, com aumento da densidade (Mörtl & Rothhaupt, 2003; Zaiko et al., 2009; Atalah et al., 2010) (Fig. 3A). A maior complexidade estrutural do ha-bitat, gerada pela presença dos mexilhões, pro-porciona aos microcrustáceos maior chance de fugir da predação, já que fazem parte da dieta de vários outros organismos e, desta forma, au-mentam suas densidades (Beekey et al., 2004). Gonzalez & Downing (1999) argumentam que Gammarus roeseli (Amphipoda) também é be-
neficiado pela biodeposição de D. polymorpha, portanto, o aumento da matéria orgânica tam-bém afeta a densidade destes organismos.
Insecta
A classe Insecta ou Hexapoda, que con-tém mais de 750.000 espécies descritas é o maior grupo de animais no mundo. Embora sejam es-sencialmente animais terrestres, os insetos tam-bém ocupam habitats aquáticos, principalmente na fase imatura (Ruppert & Barnes, 1996).
Chironomidae
As larvas da família Chironomidae pos-suem elevada diversidade e densidade em ecos-sistemas de água doce, participando signifi-cativamente na composição da biota aquática (Callil & Cruz, 2009). Em locais invadidos pelo mexilhão-dourado a densidade de larvas de Chironomidae não variou (Darrigran et al., 1998a) ou até sofreu efeito negativo (Marçal & Callil, 2008) (Fig. 3B). Enquanto que na pre-sença do mexilhão-zebra o efeito na densidade de larvas de Chironomidae foi sempre positivo (Fig. 3B). O aumento na densidade de larvas de Chironomidae pode ser determinado pela me-nor capacidade de forageamento dos peixes, devido à maior complexidade estrutural do local (Beekey et al., 2004). Assim como os mi-crocrustáceos que possuem maior chance de fugir dos predadores, as larvas de Chironomi-dae, como Chironomus, também encontram re-fúgio entre as conchas e acabam beneficiadas. Para Botts et al. (1996) e Mörtl & Rothhaupt (2003) o aumento da densidade pode estar re-lacionado à maior fonte de alimentação gerada pela biodeposição das fezes e pseudofezes do mexilhão-zebra.
Coleoptera Os coleópteros aquáticos ocupam uma ampla variedade de ambientes aquíferos, in-cluindo lagos, riachos e estuários (Benetti et al., 1998). Nas lagoas marginais ao Rio Para-guai, invadidas por L. fortunei, a densidade de coleópteros sofreu efeito negativo, diminuin-do significativamente (Marçal & Callil, 2008)

CAPÍTULO 20
Seção 5
239
Figura 2. Variação na densidade média de invertebrados aquáticos em locais invadidos e não invadidos por Limnoperna fortunei e Dreissena polymorpha. A = Nematoda, B = Oligochaeta, C = Hirudinea, D = Gastropoda, E = Bivalvia. Os números representam as fontes dos dados: 1 = Darrigran et al. (1998), 2 = Marçal & Callil (2008), 3 = Atalah et al. (2010), 4 = Horvath et al. (1999), 5 = Mörtl & Rothhaupt (2003), 6 = Zaiko et al. (2009), 7 = Ricciardi et al. (1997). Os valores de densidade foram logaritmizados para permitir a comparação entre as fontes de dados.

Uhde et al.
Efeito do macrofouling sobre a comunidade de invertebrados aquáticos
240
(Fig. 3C). Ao contrário, a ordem Coleoptera parece ser beneficiada com a invasão de D. polymorpha, tanto Horvath et al. (1999) quan-to Zaiko et al. (2009) registraram a presença de coleópteros somente depois da presença do mexilhão-zebra (Fig. 3C).
Ephemeroptera As ninfas de Ephemeroptera habitam principalmente rios de regiões tropicais e subtropicais (Tundisi & Tundisi, 2008), e são comumente utilizadas como indicadores am-bientais, devido à sua exigência por águas bem oxigenadas (Callil & Cruz, 2009). Sua densida-de média não varia com a presença ou ausência do bioinvasor L. fortunei (Fig. 3D). Enquanto que na presença de D. polymorpha, larvas de Ephemeroptera sofrem efeito positivo, aumen-tando sua densidade média (Fig. 3D). Ninfas de Ephemeroptera são raspadoras de superfície (Tundisi & Tundisi, 2008), desta maneira, podem ser beneficiadas com a presen-ça do mexilhão-zebra, que altera a disponibili-dade de nutrientes nos habitats, por deposição de fezes e pseudofezes (Mörtl & Rothhaupt, 2003). A depuração dos nutrientes de mexi-lhão-zebra também pode conseguir estimular o crescimento de perifíton e assim atrair raspa-dores (Ricciardi et al., 1997).
Trichoptera Larvas de Trichoptera, assim como Ephe-meroptera, são utilizadas como indicadoras am-bientais, devido as suas exigências por águas bem oxigenadas. Uma característica marcante deste grupo é a capacidade de construção de abrigos fi-xos, casas e tubos (Callil & Cruz, 2009). Na pre-sença de L. fortunei diminuiu sua densidade mé-dia, sofrendo efeito negativo (Fig. 3E). Enquanto que na presença de D. polymorpha o efeito foi sempre positivo, aumentando a densidade média de larvas de Trichoptera (Fig. 3E). A densidade de larvas de Trichoptera pode variar dependendo da influência espe-cífica de cada espécie. Grandes larvas, como Polycentropodidae, são territoriais e evitam receber água previamente filtrada por seus vi-zinhos, dessa forma, competem com mexilhões por posições ótimas no substrato (Ricciardi
et al., 1997). Por outro lado, pequenas larvas podem ser beneficiadas ao explorar as cor-rentes de filtração do mexilhão-zebra. Larvas Brachycentrus orientam-se para a abertura an-terior e ficam expostas à corrente de filtração do mexilhão-zebra; nesta posição, as larvas são capazes de fazer a filtração de particulas de ali-mentos recebidos (Ricciardi et al., 1997).
Considerações Finais
A análise da literatura disponível sobre a relação entre L. fortunei e a comunidade de invertebrados aquáticos mostra que os efeitos podem ser positivos ou negativos dependendo do grupo taxonômico considerado. A compa-ração entre os dados de L. fortunei em dife-rentes regiões da América do Sul e Dreissena polymorpha na América do Norte e Europa, demonstra padrões distintos de efeitos sobre a comunidade de invertebrados aquáticos. Apesar da irregularidade dos efeitos, ob-servamos um padrão de favorecimento para os grupos Nematoda, Annelida e Chironomidae atribuído pela maioria dos autores ao incre-mento de matéria orgânica oriunda das fezes e pseudofezes, bem como o aumento da comple-xidade estrutural dos substratos causado pela presença do macrofouling. Tal padrão de favo-recimento não é observado para o estudo reali-zado por Marçal & Callil (2008) em lagoas no Pantanal. Os efeitos divergentes observados para L. fortunei no Pantanal podem estar associados a fatores como o tempo de invasão, que é me-nor em comparação as demais regiões conside-radas. Outro diferencial é a disponibilidade sa-zonal das macrófitas aquáticas, que constituem o principal substrato para colonização. Devido a essas restrições, a população de mexilhão tem suas densidades limitadas, provavelmente mi-nimizando os impactos sobre as comunidades nativas. Contudo, consideramos que a ação de L. fortunei sobre as comunidades de invertebra-dos podem ocorrer de maneira muito variável, dependendo do comportamento e nicho trófi-co dos organismos que compõe a comunida-de, bem como a restrições do ambiente para o desenvolvimento da população do invasor. Destacamos ainda a necessidade da realização

CAPÍTULO 20
Seção 5
241
de monitoramento contínuo da invasão de L. fortunei na bacia do Alto Paraguai, bem como estudos com delineamentos amostrais que per-
Figura 3. Variação na densidade média de invertebrados aquáticos em locais invadidos e não invadidos por Limnoperna fortunei e Dreissena polymorpha. A = Crustacea, B = Chironomidae, C =Coleoptera, D = Ephemeroptera, E = Trichoptera. Os números representam as fontes dos dados: 1 = Darrigran et al. (1998), 2 = Marçal & Callil (2008), 3 = Atalah et al. (2010), 4 = Horvath et al. (1999), 5 = Mörtl & Rothhaupt l (2003), 6 = Zaiko et al. (2009), 7 = Ricciardi et al. (1997). Os valores de densidade foram logaritmizados para permitir a comparação entre as fontes de dados.
mitam identificar os efeitos específicos desse invasor sobre diferentes nichos tróficos dos sis-temas.


243
A espécie Limnoperna fortunei (Dunker, 1857), mexilhão-dourado, introduzida na América do Sul em 1991 (Pastorino et al., 1993), pode ser considerada como “engenheira do ecossistema”, pois através de suas conchas, dispostas na forma de aglomerados, cria novos habitats, altera o ambiente natural, provocan-do o deslocamento de algumas espécies nati-vas e favorecendo o estabelecimento de outras (Gutiérrez et al., 2003; Darrigran et al., 2003). Além dos conhecidos prejuízos à economia, por ser de hábito epifaunal (Darrigran & Ez-curra de Drago, 2000a), estes organismos pro-vocam mudanças na biodiversidade local, com-petindo por alimento e por espaço, sufocando bivalves nativos e servindo de substrato para al-gumas espécies de invertebrados (Darrigran et al., 1998a; Mansur et al., 2008c). No entanto, poucos são os estudos da sua influência sobre a macrofauna bentônica acompanhante (Fig. 1). Para avaliar a estrutura da macrofauna bentônica associada aos aglomerados de mexi-lhão-dourado no Rio Jacuí foram realizadas 12
campanhas amostrais, com frequência quinze-nal de abril a agosto de 2007 e mais duas cam-panhas com intervalos de tempo maiores, uma coleta em outubro de 2007 e outra em janeiro de 2008. As amostragens foram efetuadas em uma boia de sinalização de navegação fluvial, localizada nas proximidades do lançamento da água de resfriamento da Usina Termelétrica Charqueadas – UTCH, no município de Char-queadas, RS, Brasil. Para comparações com a fauna dos aglomerados, foram obtidas amos-tras dos sedimentos no entorno da boia. Este trabalho foi financiado com recursos do P&D da TRACTEBEL ENERGIA/SUEZ/ANEEL. Em cada campanha foram obtidas 3 amostras de aglomerados de mexilhão-dou-rado, com um amostrador do tipo raspador. Cada amostra foi colocada em uma bandeja e delimitada a área amostral de 100 cm2 (10 cm x 10 cm). As amostras foram fixadas com formol 10% e encaminhadas para processamento. Os sedimentos no entorno foram amostrados com o busca fundo do tipo “Petersen” modificado
CAPÍTULO 21
Limnoperna fortunei na bacia hidrográfica do baixo Rio Jacuí: relações com a comunidade
de macroinvertebrados bentônicos
Simone Caterina Kapusta Suzana Maria Fagondes de Freitas

Kapusta & Freitas
Limnoperna fortunei na bacia hidrográfica do baixo Rio Jacuí: relações com a comunidade de macroinvertebrados bentônicos
244
com o esforço de três pegadas. As amostras ob-tidas foram lavadas em campo, acondicionadas em sacos plásticos, etiquetadas, fixadas com formol 10% e processadas em laboratório. Para a análise dos resultados, conside-rou-se a densidade média de indivíduos de L. fortunei e da macrofauna bentônica, por metro quadrado (ind/m2), por mês amostrado. Para a macrofauna associada a L. fortunei também foi considerada a abundância relativa (AR), sen-do os táxons classificados como: raros: <1%; ocasionais: ≥1% e <10%; comuns: ≥10% e <30%; abundante: ≥30% e <50% e dominan-te: ≥50%. Os táxons também foram categoriza-dos por meio da frequência de ocorrência (FO), como acidental: FO <25%; acessório: ≥25% e <40%; constante/frequente: ≥40% e <60% e muito frequente: FO ≥60%. Análises de Va-riância (ANOVA) unifatoriais foram efetuadas para testar a significância da densidade de or-ganismos, transformada por log (x+1), entre
riação temporal da densidade de L. fortunei, no estuário do Rio de La Plata, Argentina, en-tre os anos de 1991 a 2001, registraram pico de 150.000 ind/m2, em março de 1995. Belz (2006) estudando a densidade, recrutamento e o crescimento de L. fortunei, no reservató-rio da Usina Hidrelétrica de Itaipu (Paraná), entre dezembro de 2004 a dezembro de 2005, registrou as maiores densidades nos meses de temperatura mais elevada (dezembro a março), e um declínio da mesma à medida que se apro-ximam os meses mais frios. O autor observou um crescente incremento na densidade de me-xilhões, a partir do mês de agosto. No presente estudo, ocorreu uma tendência de aumento de densidade em agosto, com uma diminuição em outubro, quando se esperava uma maior densi-dade. Este declínio na densidade do mexilhão--dourado, no mês de outubro, provavelmente ocorreu devido às condições adversas de pluvio-sidade e temperatura, registradas neste período.
Figura 1. Densidade média de L. fortunei e da macrofauna bentônica (ind/m2) e desvio padrão, por mês amostrado (março a outubro de 2007, janeiro de 2008), coletados no Rio Jacuí, RS.
as campanhas amostrais. Quando a ANOVA foi significativa, utilizou-se o teste de Tukey, de comparação de médias, para identificar as diferenças. Para estas análises foi utilizado o programa Statistica® 5.0. As densidades de mexilhão-dourado fo-ram mais elevadas nos meses de maior tempera-tura (janeiro e março), conforme já evidencia-do por outros estudos (Darrigran et al., 2003; Belz, 2006). A densidade em março foi signifi-cativamente mais elevada do que a encontrada de abril a outubro. A densidade nos meses de junho, julho e outubro foram significativamen-te menores que as registradas em março, abril e janeiro. Darrigran et al. (2003) analisando a va-
A densidade média da macrofauna ben-tônica em março foi significativamente mais elevada do que a encontrada em maio e julho. A densidade em maio foi significativamente me-nor do que a de março, agosto e janeiro (Fig.1). A maior riqueza taxonômica da ma-crofauna bentônica, em termos absolutos, foi observada nos aglomerados de L. fortunei, to-talizando 23 famílias, enquanto que nos sedi-mentos foram registradas 15 famílias (Tab. I). A dominância das famílias foi diferen-ciada entre os substratos coletados no Rio Ja-cuí, conforme pode ser visto nas Figuras 2 e 3. Darrigran et al. (1998a) documentaram mu-danças da fauna de substratos no Rio de La Pla-ta ocupados pelo mexilhão-dourado, com um

CAPÍTULO 21
Seção 5
245
aumento significativo na riqueza e abundância de invertebrados não-moluscos (Oligochaeta, Hirudinea, Chironomidae, dentre outros) e a redução em diversidade e número de espécies de moluscos nativos. Naididae foi classificada como dominan-te nos aglomerados de mexilhão, no Rio Jacuí, Chironomidae e Oribatidae como comuns, Ba-etidae, Leptohyphidae e Cochliopidae como ocasionais e as demais famílias como raras (Tab. I). Chironomidae, Baetidae, Leptohyphidae, Naididae e Oribatidae foram as famílias com
a maior frequência de ocorrência nas amostras (FO ≥60%), sendo classificadas como “muito frequentes”. Mansur et al. (2008c) observaram a ocorrência de invertebrados associados aos macroaglomerados de L. fortunei sobre galhos de sarandi na Praia do Veludo, Lago Guaíba, RS, e registraram as famílias Cochliopidae como constante nas amostras e Lithoglyphidae e Hyallelidae como acessórias, as demais foram consideradas acidentais. As diferenças na estrutura da comunida-de associada ao mexilhão quando comparada
Tabela I. Presença (X) e ausência (-) de famílias da macrofauna bentônica, associadas aos aglomerados de L. fortunei e nos sedimentos, Rio Jacuí, RS.
Figura 2. Abundância relativa das famílias da macrofauna bentônica, acompanhante de L. fortunei.

Kapusta & Freitas
Limnoperna fortunei na bacia hidrográfica do baixo Rio Jacuí: relações com a comunidade de macroinvertebrados bentônicos
246
com a comunidade associada ao sedimento de fundo (Fig.3), caracterizam-se pela maior di-versidade de táxons da macrofauna associada ao micro-habitat criado pela população de L. fortunei, bem como pela maior oferta de perifí-ton, evidenciada principalmente nos meses mais quentes. De acordo com Mansur et al. (2008c) os macroaglomerados provavelmente oferecem diversos recursos alimentares por meio da pro-dução de pseudofezes, substrato para o perifí-ton, acúmulo de detritos e micro-organismos. Torgan et al. (2009) consideram que a riqueza de espécies de diatomáceas encontrada sobre as conchas de L. fortunei demonstra que este molusco pode ser considerado como um ha-bitat favorável à presença de várias espécies, e
isso se deve provavelmente pela localização dos aglomerados próximos à superfície da água, na zona eufótica, pela disponibilização de nutrien-tes inorgânicos dissolvidos no próprio meio e pelo suporte à fixação das diatomáceas perifíti-cas. Os resultados encontrados no presente estudo estão corroborando os estudos acerca da presença de L. fortunei nos ecossistemas aquá-ticos, concluindo que estes organismos, são ca-pazes de modificar a estrutura da comunidade bentônica e, por conseguinte alterar a cadeia trófica (Martin & Darrigran, 1994; Darrigran et al., 1998a; Darrigran, 2002; Pelichotti et al., 2002).
Figura 3. Abundância relativa das famílias da macrofauna bentônica, registradas nos sedimentos.

Seção
6Efeito da filtração de bivalves invasores sobre a comunidade planctônica


249
Desde a invasão de Limnoperna fortunei (Dunker, 1857) na América do Sul, as pesqui-sas enfocaram principalmente aspectos relacio-nados à sua distribuição espacial e temporal, ao ciclo reprodutivo e ao desenvolvimento larval. Os efeitos da filtração do mexilhão-dourado na disponibilidade de nutrientes e na estrutura das comunidades planctônicas ainda são pouco conhecidos. O mexilhão-dourado é bastante se-melhante ao bivalve invasor na Europa e Améri-ca do Norte Dreissena polymorpha, conhecido como mexilhão-zebra. Desta forma, é bastante provável que os impactos ecológicos que o me-xilhão-zebra promoveu na Europa e América do Norte também ocorram na América do Sul com o mexilhão-dourado. O mexilhão-zebra tem a capacidade de alterar a composição e a abundância das comu-nidades planctônicas através da filtração seleti-va das partículas em suspensão. Alguns estudos sugerem que a filtração seletiva de D. polymor-pha em ecossistemas com a presença de ciano-
bactérias poderia promover a ocorrência de florações tóxicas (Vanderploeg et al., 2001). Entretanto, os estudos a respeito dos efeitos do mexilhão-zebra sobre as densidades de cia-nobactérias e a ocorrência de florações tóxicas são contraditórios. Alguns estudos observaram o declínio das densidades de cianobactérias (Bastviken et al., 1998, Dionisio-Pires & Van Donk, 2002), enquanto outros demonstraram efeito contrário, onde a filtração do mexilhão--zebra promoveu o aumento das densidades de cianobactérias (Vanderploeg et al., 2001, Ni-cholls et al., 2002). A dominância de cianobactérias em ecos-sistemas de água doce é um problema mundial, especialmente naqueles sujeitos à eutrofização, devido à formação de florações e produção de toxinas, conhecidas como cianotoxinas. O tipo mais comum de intoxicação envolvendo cia-nobactérias é ocasionado pelas hepatotoxinas (microcistinas e nodularinas), que agem sobre o citoesqueleto das células provocando hemor-
CAPÍTULO 22
O impacto de Limnoperna fortunei sobre as cianobactérias
Vanessa Gazulha

Gazulha, V.
O impacto de Limnoperna fortunei sobre as cianobactérias
250
ragias. A exposição crônica a doses não letais de hepatotoxinas pode causar tumores no fíga-do, e consequentemente, trazer sérios danos a seres humanos e animais (Falconer, 1991; Car-michael, 1992). O mexilhão-dourado é invasor na Amé-rica do Sul há mais de quinze anos, ocorrendo sempre em densidades massivas que podem su-perar 140.000 ind m-2 (Darrigran & Mansur, 2006). As taxas de filtração registradas para L. fortunei são mais elevadas do que aquelas registradas para outros bivalves invasores de água doce, como D. polymorpha, D. bugensis e Corbicula fluminea. As elevadas taxas de fil-tração do mexilhão-dourado associadas às suas densidades massivas indicam seu grande poten-cial para alterar a estrutura trófica dos ecossis-temas invadidos. Desta forma, pretende-se abordar o comportamento alimentar e as taxas de fil-tração do mexilhão-dourado na presença de cianobactérias tóxicas e não-tóxicas (Mi-crocystis aeruginosa e Planktothrix sp.) e fito-plâncton não-tóxico (Nitzschia palea e Mono-raphidium sp.). O presente capítulo apresenta resultados referentes à primeira avaliação dos efeitos de cianobactérias tóxicas na alimen-tação e sobrevivência do mexilhão-dourado, assim como as primeiras estimativas das ta-xas de filtração das larvas deste bivalve inva-sor baseado em Gazulha (2010) e Gazulha et al. (2012a, b).
Taxas de filtração, ingestão e produção de pseudofezes
Quando se trata de avaliar as taxas de filtração de bivalves é importante deixar claro as variações existentes, pois é fundamental no momento de comparar resultados de diferentes estudos. As taxas de filtração (FR) ou taxa de clareamento (CR) foram estimada considerando as partículas capturadas pelo mexilhão. A taxa de ingestão (IR) é igual à taxa de filtração menos a taxa de produção de pseudofezes (PPR). As pseudofezes são as partículas filtradas aglomeradas com muco que são expelidas periodicamente pelo sifão inalante, isto é, partículas filtradas, mas não ingeridas. FR é igual à IR somente se não forem produzidas pseudofezes. As taxas de filtração
do mexilhão-dourado foram estimadas baseadas na fórmula contida em Coughlan (1969).
Análise das pseudofezes e fezes
Foi desenvolvido um método para esti-mar precisamente a produção de pseudofezes pelo mexilhão dourado, conforme descrito a seguir. Durante o curso dos experimentos, o comportamento de cada exemplar de L. fortunei foi monitorado sob microscópio estereoscópico acoplado à câmera digital. As pseudofezes e fe-zes foram capturadas no momento da liberação com a utilização de pipetas capilares e fixadas com Lugol a 1%. As pseudofezes foram desinte-gradas em equipamento de ultrassom Bandelin Sonorex RK100H durante 10 min para separar as células do muco sem danificá-las. As partí-culas alimentares foram estimadas em câmara de contagem de Sedgewick-Rafter para acessar a taxa de produção de pseudofezes (PPR).
Padrão de liberação das pseudofezes e fezes
O padrão de liberação das pseudofe-zes foi diferenciado entre os tratamentos. As pseudofezes, no tratamento fitoplâncton não--tóxico (diatomácea Nitzschia), foram liberadas constantemente, durante toda a duração do experimento e sempre sob a forma de grandes aglomerados globulares ou sob a forma de cor-dões (Fig. 1). No tratamento cianobactéria (Mi-crocystis tóxica e não-tóxica), as pseudofezes foram liberadas com menor frequência, sempre sob a forma de pequenos aglomerados globu-lares (Fig. 2). O padrão de liberação das fezes foi o mesmo em todos os tratamentos. As fezes apresentaram aspecto semelhante a “tiras acha-tadas” (formato do intestino de L. fortunei) de coloração marrom (Fig. 3).
L. fortunei adulto na presença de M. aeruginosa tóxica e diatomácea não-tóxica N. palea - Short Term Grazing Experiment (1h)
FR do mexilhão-dourado variou de 95,6 a 817,5 mL ind-1 h-1, e o valor médio foi de 519,3 mL ind-1 h-1 (Fig. 4). FR em re-lação à massa corporal variou de 2,4 a 24,5

CAPÍTULO 22
Seção 6
251
mL mgDW-1 h-1, e o valor médio foi de 10,6 mL mgDW-1 h-1(Fig. 4). FRs de L. fortunei foram significativamente maiores na presen-ça da Nitzschia não-tóxica do que nas outras combinações alimentares (p<0,05, ANOVA). Apesar das elevadas FRs sobre a diatomácea não-tóxica, o mexilhão-dourado rejeitou mais células de Nitzschia como pseudofezes (p<0,05, ANOVA) e ingeriu mais células de Microcystis (p<0,05, ANOVA).
L. fortunei adulto na presença de M. aeruginosa tóxica e diatomácea não-tóxica N. palea - Long Term Grazing Experiment (120h)
IR na presença de Microcystis tóxica variou de 31,8 a 54,6 mL ind-1 h-1 e de Microcystis não--tóxica variou de 36,3 a 62,5 mL ind-1 h-1 (Fig. 5). Não houve diminuição significativa da IR de L. for-tunei sob exposição à Microcystis tóxica ao longo
Figura 1. Pseudofezes de L. fortunei no tratamento fitoplâncton não-tóxico (Nitzschia). Da esquerda para a direita: a) pseudo-fezes sob a forma de grandes aglomerados globulares, b) pseudofezes sob a forma de cordões, c) aglomerado de células de Nitzschia sob a forma de pseudofezes. Sifão inalante (SI), sifão exalante (SE), pseudofezes (Ps).
Figura 2. Pseudofezes de L. fortunei no tratamento cianobactéria tóxica (Microcystis). Da esquerda para a direita: a) pseudofe-zes sob a forma de pequenos aglomerados globulares, b) aglomerado de células de Microcystis sob a forma de pseudofezes. Sifão inalante (SI), sifão exalante (SE), pseudofezes (Ps).
Figura 3. Fezes de L. fortunei. Da esquerda para a direita: a) liberação das fezes sob a forma de “tiras achatadas”, b) fezes. Sifão exalante (SE), fezes (F).

Gazulha, V.
O impacto de Limnoperna fortunei sobre as cianobactérias
252
do experimento de filtração (p<0,05, ANOVA), indicando que o mexilhão-dourado ingeriu células tóxicas durante os 5 dias de experimento. Os mexilhões foram capazes de se alimentar e sobreviver na presença da cianobactéria tóxica, sugerindo que L. fortunei apresenta mecanismos de sobrevivência diante das cianotoxinas, além de evidenciar o seu potencial como vetor para a trans-ferência das cianotoxinas para os níveis tróficos superiores, aumentando o risco de bioacumulação nos ecossistemas aquáticos.
L. fortunei adulto na presença de ciano-bactérias solitárias (Microcystis), coloniais (Microcystis) e filamentosas (Planktothrix)
FRs na presença das cianobactérias uni-celulares, coloniais e filamentosas foram seme-
lhantes, no entanto, houve uma grande diferen-ça nas IRs e PPRs (p<0,05, ANOVA; Fig. 6). As células solitárias foram amplamente aceitas como alimento, enquanto que as colônias e fi-lamentos foram expulsos abundantemente nas pseudofezes. L. fortunei apresentou capacidade de selecionar partículas alimentares, e desta forma, promover a dominância de determi-nadas espécies do plâncton nos ecossistemas aquáticos. A presença do mexilhão-dourado pode-ria levar a uma redução de células solitárias de cianobactérias (Microcystis), e favorecer o au-mento em densidade de cianobactéiras coloniais (Microcystis) e filamentosas (Planktothrix), e de diatomáceas (Nitzschia). O mexilhão-dourado apresenta potencial para remover células tóxi-cas de cianobactérias (Microcystis), entretanto,
Figura 4. Taxa de Filtração (FR), Taxa de Ingestão (IR) e Taxa de Produção de Pseudofezes (PPR) de Limnoperna fortunei (mL ind-1 h-1 e mL mgDW-1 h-1). Nitzschia não-tóxica (N), Microcystis tóxica (NPLJ-4) e não-tóxica (NPCD-1), e mistura de Nitzschia+Microcystis tóxica (Mix N+NPLJ-4) (símbolo= Média, barra= Erro Padrão).
Figura 5. Taxa de Ingestão (IR) de Limnoperna fortunei (mL ind-1 h-1). Microcystis tóxica (NPLJ-4) e não-tóxica (NPCD-1) (símbolo= Média, barra= Erro Padrão).

CAPÍTULO 22
Seção 6
253
este potencial ficaria reduzido em eventos de floração, onde as formas coloniais, preferen-cialmente rejeitadas por L. fortunei, são predo-minantes. Neste caso, a presença do bivalve no ambiente poderia ainda potencializar a ocor-rência da floração via rejeição das cianobacté-rias coloniais nas pseudofezes.
Taxas de filtração das larvas de L. fortunei na presença de Microcystis tóxica e não-tóxica e Monoraphidium não-tóxico
FR da larva de L. fortunei variou de 9,9 a 44,5 μL larva-1 h-1, com o valor médio de 24,7 μL larva-1 h-1 (Fig. 7). As maiores FRs foram
registradas na presença de Monoraphidium não-tóxico, com o valor médio de 28,2 μL larva-1 h-1. As larvas ingeriram Monoraphidium, assim como Microcystis não-tóxica e tóxica a taxas similares, indicando que não houve efeito negativo das toxinas de cianobactérias na filtração das larvas durante os 4 dias de experimento (p>0,05, ANOVA).
Sobrevivência das larvas de L. fortunei na presença de cianobactérias tóxicas e fitoplâncton não-tóxico
Elevada sobrevivência das larvas foi regis-trada na presença da alga verde Monoraphidium
Figura 6. Taxa de Filtração (FR), Taxa de Ingestão (IR), e Taxa de Produção de Pseudofezes (PPR) de L. fortunei (mL ind-1 h-1 e mL mgDW-1 h-1) na presença de cianobactéria filamentosa, colonial e unicelular (símbolo= Média, barra= Erro Padrão). Floração natural da filamentosa Planktothrix (NB F), M. aeruginosa colonial (NPCD-1 C), M. aeruginosa unicelular (NPCD-1 S).
Figura 7. Taxa de Filtração (FR) das larvas de Limnoperna fortunei. Monoraphidium não-tóxico (MONO), Microcystis tóxica (NPLJ-4) Microcystis não-tóxica (NPCD-1) (símbolo= Média, barra= Erro Padrão).

Gazulha, V.
O impacto de Limnoperna fortunei sobre as cianobactérias
254
e de seston natural durante todo o experimento (p<0,05, ANOVA; Fig. 8). Após quatro dias de exposição, a sobrevivência das larvas diminuiu na presença de Microcystis tóxica, seguida pelo extrato de Microcystis tóxica+seston natural, e Microcystis não-tóxica (p <0,05, ANOVA). As larvas demonstraram maior sensibilida-de à natureza da partícula em comparação com os adultos de L. fortunei. As cianobactérias não tiveram efeito negativo nas taxas de filtração das larvas, indicando a incapacidade destas de de-tectar e evitar as partículas tóxicas. Entretanto, ocorreu um aumento significativo da mortalida-de larval na presença das cianobactérias tóxicas e não-tóxicas em longo prazo (96h). Estes resulta-dos demonstram que além da toxicidade da par-tícula alimentar, a qualidade desta pode ser um fator importante na seletividade alimentar das larvas. O predomínio das cianobactérias no am-biente poderia ter como consequência o aumento da mortalidade das larvas de L. fortunei.
Figura 8. Sobrevivência (%) das larvas de L. fortunei na presença de Monoraphidium não-tóxico (MONO), Microcystis não-tóxica (NPCD-1), Microcystis tóxica (NPLJ-4), Seston Natural (NS), extrato de Microcystis tóxica (NPLJ-4 + NS) (símbolo= Média, barra= Erro Padrão).
Considerações finais
No presente estudo, foram abordados apectos relacionados à seletividade alimentar de L. fortunei na presença de cianobactérias tó-xicas e não-tóxicas, e fitoplâncton não-tóxico. Algumas questões foram esclarecidas, entre-tanto ainda existe uma lacuna no entendimen-to dos mecanismos de seleção de partículas do mexilhão-dourado, especialmente no que diz respeito à qualidade do alimento, como a seleção de partículas orgânicas ou inorgâni-cas, a capacidade de ingestão e a eficiência de digestão. Além disso, estudos sobre os efeitos da filtração do mexilhão-dourado na estrutu-ra trófica dos ecossistemas são raros. A conti-nuidade de estudos desta natureza é de grande importância no sentido de se compreender a interação existente entre esta espécie invasora, o plâncton e o bentos nos ecossistemas invadi-dos.

255
Em bivalves filtradores, como o mexi-lhão-dourado, a captação das partículas em suspensão na água ocorre através de um sifão inalante. Os cílios das brânquias geram cor-rentes que impulsionam a água pelo sifão até chegar à cavidade do manto e brânquias. As partículas sólidas são removidas pelos cílios das brânquias e, aquelas selecionadas para ingestão, são dirigidas até a boca, enquanto que as partículas indesejáveis são envoltas em muco e eliminadas como pseudofezes por um canal adjacente ao sifão inalante (Morton, 1973). Embora a ocorrência da seleção de par-tículas alimentares pelos bivalves seja reconhe-cida, ainda existem importantes questões sobre os critérios de seleção utilizados (Baker et al., 2000). Segundo Bernard (1974), a seletividade pode ser dividida em duas categorias princi-pais: 1) a separação de partículas inorgânicas das partículas orgânicas, que são geralmente preferidas em relação às primeiras, e 2) a pos-às primeiras, e 2) a pos- primeiras, e 2) a pos-sível seleção entre partículas orgânicas, onde
se incluem organismos planctônicos das mais variadas formas, tamanhos e estruturas. Denomina-se plâncton o conjunto de or-ganismos diminutos (μm a poucos mm) que têm pouco poder de locomoção e vivem livremente na coluna de água, sendo muitas vezes arrastados pelas correntezas. Algumas espécies são fotossintetizantes, constituindo o fitoplâncton (também chamado de algas), enquanto outras são heterotróficas, constituindo o zooplâncton. Encontram-se na base das cadeias alimentares aquáticas, servindo como alimento a diversos organismos, incluindo bivalves. Experimentos abordando a seletividade de bivalves sobre a comunidade fitoplanctônica são os mais comuns, utilizando principalmente algumas espécies de algas cultivadas em labo-ratório (Berg et al., 1996; Baker et al., 1998; Gazulha et al., 2012a, b). Poucos são os estudos sobre a filtração do plâncton natural e, espe-cialmente, sobre o zooplâncton. Em laborató-rio, Horgan & Mills (1997), Bastviken et al. (1998), Dionisio-Pires et al. (2004) e Naddafi
CAPÍTULO 23
Os impactos do mexilhão-dourado sobre
a comunidade planctônica
Aline FachiniVanessa Gazulha
Catarina da S. Pedrozo

Fachini et al.
Os impactos do mexilhão-dourado sobre a comunidade planctônica
256
et al. (2007) realizaram estudos sobre a filtra-ção do mexilhão-zebra (Dreissena polymorpha) somente com a comunidade fitoplanctônica na-tural. Já Rojas Molina et al. (2010) estudaram a dieta natural do mexilhão-dourado, incluin-do fito e zooplâncton, analisando o conteúdo estomacal de exemplares coletados na planície de inundação do Rio Paraná. Assim como es-tes, diversos outros trabalhos tentam elucidar a relação existente entre a chegada dos bivalves invasores e as alterações observadas na densi-dade e composição da comunidade planctônica dos ambientes invadidos (Nicholls & Hopkins, 1993; Caraco et al., 1997; Pace et al., 1998; Smith et al., 1998; Jack & Thorp, 2000; Thorp & Casper, 2002; Rojas Molina & Paggi, 2008; David et al., 2009).
Para a realização dos experimentos em laboratório, exemplares de L. fortunei e orga-nismos planctônicos foram coletados no Lago Guaíba em Porto Alegre, RS. Os experimen-tos consistiam, basicamente, em manter cada mexilhão filtrando ativamente durante 1h em uma suspensão contendo os organismos planc-tônicos (104 org. L-1), conforme visualizado na Figura 1. Ao final dos experimentos, os organismos planctônicos presentes no recipiente controle, no recipiente experimental e nas pseudofezes coletadas foram identificados, contabilizados e medidos. A partir disto, estimaram-se quais espécies foram filtradas, ingeridas ou rejeitadas pelos mexilhões durante os experimentos, se-gundo o diagrama descrito na Figura 2.
Figura 1. Recipiente controle, contendo somente plâncton (à esquerda) e recipien-te experimental, contendo plâncton e um mexilhão (à direita). A pipeta foi utilizada para a coleta das pseudofezes.
Para entendermos os efeitos da filtração do mexilhão-dourado sobre o plâncton é im-portante conhecer quais espécies estão mais sus-cetíveis à captura, ingestão ou rejeição. Tanto o plâncton consumido quanto o que é rejeitado nas pseudofezes podem constituir significantes perdas energéticas para a região pelágica (Hor-gan & Mills, 1997) em função das altas taxas de filtração registradas para L. fortunei (Sylvester et al., 2005). A maioria dos estudos foca na fil-tração de pequenas partículas fitoplanctônicas, mas ainda existem questões sobre quais tama-nhos e tipos de plâncton podem ser retirados da água pelos mexilhões (Horgan & Mills, 1997). Alguns trabalhos (Bastviken et al., 1998; Baker et al., 1998) salientam a importância de se ana-lisar as pseudofezes para uma melhor observa-ção dos possíveis padrões de seletividade, já que a filtração dos mexilhões pode se dar em uma ampla faixa de partículas, mas parte delas ser
rejeitada na cavidade do manto, brânquias e pal-pos labiais e deslocada da coluna d’água para o sedimento. Dessa forma, a ausência da partícula na suspensão não significa necessariamente que ela foi utilizada como alimento. O mecanismo de seleção alimentar em filtradores é controlado por diversos fatores físi-cos, químicos e biológicos, e muitos estudos têm revelado que diferenças no tamanho, na forma e na concentração de partículas, assim como suas moléculas de superfície, metabólitos e car-gas eletrostáticas podem influenciar na escolha (Espinosa et al., 2008). No estudo realizado por Fachini et al. (2011), cujos resultados principais serão apresentados a seguir, foram testadas as capacidades seletivas do mexilhão-dourado so-bre a forma e o tamanho dos organismos de uma comunidade planctônica natural, a fim de sub-sidiar a avaliação dos possíveis efeitos sobre sua estrutura.

CAPÍTULO 23
Seção 6
257
Uma vez que o objetivo principal dos experimentos realizados era identificar quais formas e tamanhos planctônicos foram prefe-rencialmente filtrados, ingeridos ou rejeitados pelos mexilhões, as espécies registradas nestas três situações foram, então, agrupadas arbitra-riamente de acordo com suas formas segundo a Tabela I. Imagens de alguns dos organismos podem ser visualizadas nas Figuras 7 e 8. Estes mesmos organismos fito e zoo-planctônicos foram, após, categorizados quan-to ao tamanho, gerando a distribuição ilustrada na Tabela II. Deve-se lembrar de que somente organismos maiores do que 30 μm estão pre-sentes em função da coleta por filtração em rede. Para uma melhor compreensão, os resul-
tados obtidos foram transformados em taxas de filtração, de ingestão e de produção de pseu-dofezes de acordo com fórmula proposta por Coughlan (1969) e em índices de seletividade alimentar de acordo com fórmula proposta por Strauss (1979). As taxas de filtração do mexilhão-dou-rado sobre a comunidade planctônica natural durante os experimentos variaram entre 36,8 e 86,4 mL mexilhão-1 h-1, com média de 61,9 mL mexilhão-1 h-1. Estas taxas ficaram abaixo de algumas taxas já registradas para L. fortunei em estudos de laboratório (entre 125 e 350 mL mexilhão-1 h-1, de acordo com Sylvester et al., 2005; e entre 95,6 e 817,5 mL mexilhão-1 h-1, de acordo com Gazulha et al., 2012a, b), mas o fato de se utilizar uma comunidade planctônica
Figura 2. Estimativa dos organismos planctônicos filtrados, ingeridos e rejeitados pelo mexilhão-dourado através dos experi-mentos.
Tabela I. Classificação dos organismos planctônicos quanto à forma.
FITOPLÂNCTON ZOOPLÂNCTON
Categoria Categoria
Gênero e/ou Família/Ordem Gênero e/ou Família/Ordem
Diatomáceas unicelulares (60-550 µm)Pennales, Centrales
Rotíferos Brachionidae (90-400 µm)Brachionus, Keratella, Kellicottia, Platyonus
Espinhosas (30-95 µm)Desmodesmus, Kirchneriella, Monactinus, Pediastrum, Scene-desmus, Staurastrum, Tetraedron
Outros Rotíferos (100-300 µm)Conochilus, Bdelloidea, Lecane, Lepadella, Trichocerca
Alongadas (350-1050 µm)Closterium, Closteriopsis, Euglena
Cladóceros (190-990 µm)Alona, Bosmina, Chydorus, Ceriodaphnia, Diaphanosoma
Filamentosas 1 (150-1200 µm) Aulacoseira, Mougeotia, Planktothryx
Copépodos (400-1850 µm)Calanoida, Cyclopoida
Filamentosas 2 (850-19000 µm)Spyrogyra
Náuplios de Copépodos (100-400 µm)Calanoida, Cyclopoida
Coloniais mucilaginosas (100-700 µm)Aphanocapsa, Microcystis, Volvox, Westella
Protozoários testáceos (50-300 µm)Tecamoebas, Ciliophora

Fachini et al.
Os impactos do mexilhão-dourado sobre a comunidade planctônica
258
natural e não somente espécies de algas isola-das deve ser considerado. Segundo Vanderplo-eg et al. (2009), diferentes taxas de filtração tem sido observadas para o mexilhão-zebra se alimentando do seston natural, e estas taxas seriam consideravelmente mais baixas do que aquelas observadas em culturas de algas de la-boratório. Portanto, deve-se ter cautela ao ex-trapolar as taxas obtidas em laboratório, geral-mente sob condições ótimas, para situações no ambiente natural. Como podemos observar na Figura 3, uma parcela equivalente dos organismos planc-tônicos filtrados acabou sendo ingerida e rejei-tada pelos mexilhões durante os experimen-tos, demonstrando que grandes quantidades de plâncton podem ser deslocadas da coluna d’água para o sedimento sob a forma de pseu-dofezes (Fig. 4). Embora se tenha observado que os organismos planctônicos sejam libera-dos ainda vivos nestas pseudofezes, a viabilida-de fica certamente comprometida em meio ao emaranhado de muco. Como hipótese principal do estudo re-alizado, tínhamos que L. fortunei utilizaria não só fitoplâncton, mas também zooplâncton como alimento, e que este bivalve selecionaria os organismos baseado nas formas e tamanhos, de maneira que os menores e com menos proje-ções seriam preferidos como alimento. Nossos resultados, a exemplo do registrado por Rojas Molina et al. (2010), mostraram que o mexi-lhão-dourado foi capaz de se alimentar tanto
de fito quanto de zooplâncton e que a seletivi-dade foi positiva para organismos de pequeno a moderado tamanho e limitada capacidade de escape, independente de possuir espinhos, pelo menos na escala observada. Isto indica que o movimento, além do tamanho, foi mais impor-tante do que a forma para a seletividade ali-mentar do mexilhão-dourado. A Figura 5 nos mostra como foi a seleção alimentar do mexilhão-dourado em função da forma dos organismos fito e zooplanctônicos. O índice apresentado faz uma proporção entre os itens (categorias) alimentares disponíveis à filtração do mexilhão com os itens ingeridos
Tabela II. Classificação dos organismos planctônicos quanto ao tamanho.
Fitoplâncton Diatomáceas unicelulares Filamentosas 1 Filamentosas 2 Espinhosas Alongadas Coloniais mucilaginosas
< 100 μm x x
100 a 300 μm x x x
300 a 600 μm x x x x
600 a 1000 μm x x x x
> 1000 μm x x x
Zooplâncton Cladóceros Copépodos Náuplios Copepoda Rotíferos Rotíferos
Brachionidae Protozoários testáceos
< 100 μm x x x
100 a 300 μm x x x x x
300 a 600 μm x x
600 a 1000 μm x x
> 1000 μm x
Figura 3. Taxas de filtração (TF), produção de pseudofezes (TPP) e ingestão (TI) do mexilhão-dourado sobre a comunida-de planctônica natural (barras = média ± EP, p < 0,001).

CAPÍTULO 23
Seção 6
259
pelo bivalve, indicando se o item foi seleciona-do de forma positiva, negativa ou indiferente. Observa-se que entre o zooplâncton, os grupos de Rotíferos foram consumidos preferencial-mente em relação ao grupo dos Copépodos pelo mexilhão. Já entre o fitoplâncton, o grupo das Espinhosas e Filamentosas 1 teve preferên-cia em relação ao grupo das Filamentosas 2. Quanto ao tamanho dos organismos, observa-se na Figura 6 que o mexilhão apresen-tou seletividade negativa para as três maiores categorias do zooplâncton, enquanto que para
o fitoplâncton somente a última categoria não foi preferida. Esta diferença se deve, provavel-mente, ao fato de os organismos zooplanctô-nicos maiores terem uma capacidade natatória que os permite escapar da corrente inalante do mexilhão, sendo, desta forma, inacessíveis ao consumo do bivalve. Assim, pelos resultados deste estudo, observou-se que as categorias de forma mais ingeridas de fitoplâncton (Espinhosas e Fila-mentosas 1) e de zooplâncton (Outros rotíferos e Rotíferos Brachionidae) também pertenciam
Figura 4. Limnoperna fortunei liberando pseu-dofezes: 4A, pseudofe-zes sendo liberadas (SI = Sifão Inalante; Ps = Pseu-dofezes); 4B, Pseudofezes contendo algas diatomá-ceas; 4C, Pseudofezes contendo algas filamen-tosas; 4D, Pseudofezes contendo um microcrus-táceo Cladócero.
Figura 5. Seletividade alimentar do mexilhão-dourado sobre as diferentes formas de organismos planctônicos. Índices positi-vos = preferência; índices negativos = rejeição ou inacessibilidade. O tamanho das barras indica a intensidade da seleção (* p<0,05).

Fachini et al.
Os impactos do mexilhão-dourado sobre a comunidade planctônica
260
às menores categorias de tamanho. As projeções e espinhos exibidos por alguns destes organis-mos, nesta escala, parecem não ter tido efeito contra a filtração dos mexilhões e o tamanho dos organismos planctônicos parece ter sido o determinante para a ingestão. As formas Colo-niais, Alongadas e, especialmente, Filamentosas 2, que se enquadravam nas maiores categorias de tamanho, foram filtradas e ingeridas em me-nores taxas. Outro resultado interessante observado foi o pequeno consumo do grupo Diatomáceas, cujos organismos enquadraram-se nas menores categorias de tamanho planctônico e estiveram bastante abundantes no plâncton disponibiliza-do à filtração do mexilhão durante os experi-mentos. Baixas taxas de filtração de diatomáce-as também foram registradas em outros estudos não só para L. fortunei, mas também para ou-tros bivalves (Cucci et al., 1985; Shummay et al., 1985; Baker et al., 1998; Bastviken et al., 1998; Gazulha et al., 2012a, b), indicando que, neste caso, algum fator além das dimensões da alga também influenciou no seu baixo consumo. Mensurar impactos do mexilhão-dourado sobre a comunidade planctônica no ambiente na-tural é difícil em função do grande número de variáveis bióticas e abióticas envolvidas e do po-tencial que este bivalve tem como espécie-enge-nheira de modificar tanto a estrutura biológica quanto física do ambiente. Mas alguns estudos no ambiente natural (Rojas Molina & Paggi, 2008) indicam a presença do mexilhão-dourado como a grande responsável pelas alterações verificadas
na comunidade planctônica, demonstrando que o investimento em pesquisas sobre o comporta-mento alimentar do mexilhão-dourado pode ser útil em fornecer subsídios para o conhecimento e avaliação dos impactos deste bivalve invasor. Nesse sentido, os resultados do estudo aqui apre-sentado indicam que o L. fortunei, através de sua seletividade alimentar, pode promover o decrés-cimo na densidade de organismos planctônicos de menor tamanho tanto do fito quanto do zo-oplâncton, enquanto rejeita o consumo daqueles organismos maiores ou capazes de evitar a cor-rente inalante do bivalve. A estrutura da comuni-dade planctônica pode sofrer alterações devido a esta ingestão seletiva do mexilhão-dourado e os impactos podem ser diferenciados em cada am-biente, dependendo, entre outros fatores, da di-versidade de organismos planctônicos que com-põem a comunidade. Esta alteração no plâncton, associada ao incremento de recursos alimentares no bentos proporcionado pelos mexilhões, acaba refletindo diretamente sobre o comportamento alimentar e a estrutura das demais comunidades biológicas do sistema. As informações sobre as interações do L. fortunei com as comunidades biológicas do Lago Guaíba e, especialmente, com sua co-munidade planctônica, ainda são insuficientes para que se tenha uma dimensão dos impactos causados pelo mexilhão-dourado neste local. Mas a grande densidade de mexilhões que co-lonizam este sistema torna cada vez mais neces-sários o monitoramento, a pesquisa e o manejo constantes neste ambiente.
Figura 6. Seletividade alimentar do mexilhão-dourado sobre os diferentes tamanhos de organismos planctônicos (em µm). Índices positivos = preferência; índices negativos = rejeição ou inacessibilidade. O tamanho das barras indica a intensidade da seleção (*p<0,05).

CAPÍTULO 23
Seção 6
261
Figura 7. Grupos de Fitoplâncton: 1-6) Diatomáceas; 7-14) Espinhosas; 15-18) Alongadas; 19-23) Filamentosas 1; 24) Filamento-sas 2; 25-27) Coloniais mucilaginosas. Grupos de Zooplâncton: 28-30) Copépodos.
Figura 8. Grupos de Zooplâncton: 31) Náuplio; 32-36) Cladóceros; 37-43) Rotíferos; 44-50) Rotíferos Brachionidae; 51-60) Pro-tozoários testáceos.


Seção
7Prevenção


265
Em 2001, o biólogo Hélio Martins Fon-tes Junior, da Itaipu Binacional, apresentou a uma plateia de pesquisadores e outros convi-dados, a notícia de que o mexilhão-dourado havia sido encontrado nas paredes internas da tomada d’água da Usina. Em dez anos, o pe-queno molusco havia percorrido mais de mil quilômetros desde a foz do Rio da Prata até Itaipu. O tema foi discutido internamente e o departamento de meio ambiente de FURNAS foi convidado a participar da vistoria da capta-ção de água para o sistema de refrigeração da subestação de Itaipu, operada por FURNAS. A empresa tinha uma preocupação inicial: asse-gurar que a infestação não prejudicasse a re-frigeração dos gigantescos transformadores de voltagem da subestação que transmite a energia de Itaipu para todo o país. Descobriu-se que não havia risco algum devido aos procedimen-tos operacionais adotados, que não favoreciam a fixação dos mexilhões, seja nos equipamen-tos da subestação, seja no sistema de captação.
A partir daí, FURNAS constituiu um grupo de trabalho multidisciplinar, então com-posto por biólogos e engenheiros, que parti-ciparam de um treinamento promovido pela bióloga da CEMIG, Maria Edith Rolla, em Belo Horizonte, e também no laboratório da Divisão de Ecossistemas Aquáticos da Itaipu Binacional, coordenado pela zootecnista Car-la Canzi. Nas duas ocasiões foram discutidas técnicas de monitoramento e identificação de larvas e adultos. Algumas semanas depois, seus representantes participaram de um evento para discutir o combate ao mexilhão-dourado na América do Sul. Essas atividades iniciais foram marcan-tes para a definição de uma estratégia de atu-ação para a empresa. Até aquele momento, as usinas hidrelétricas operadas pela empresa es-tavam distantes das áreas infestadas. Em 2003, o Limnoperna fortunei (Dunker, 1857) já havia chegado ao Pantanal e ao reservatório de Ilha Solteira, formado pelos rios Grande e Paranaí-ba, que formam o Rio Paraná. O grupo sentiu
CAPÍTULO 24
Ações de Eletrobras–FURNAS na divulgação da Campanha de
Controle do Mexilhão-Dourado
Rodrigo De FilippoPaulo Sérgio Formagio
Fernando Antonio Blanco ResendeMarcilia Barbosa Goulart

De Filippo et al.
Ações de Eletrobras–FURNAS na divulgação da Campanha de Controle do Mexilhão-Dourado
266
a necessidade de fazer uma mobilização interna e apresentar o problema para as unidades com maior risco de introdução do bivalve. Quatro usinas hidrelétricas foram de-finidas como prioritárias para essa atividade inicial: UHE Itumbiara, situada nos estados de Minas Gerais e Goiás, no Rio Paranaíba; UHEs Marimbondo e Porto Colômbia, nos estados de Minas Gerais e São Paulo, no Rio Grande; e APM Manso, no estado do Mato Grosso, no Rio Manso. Foi elaborada uma apresentação para os funcionários das áreas de operação e manutenção, e o treinamento para coleta de larvas com rede de plâncton e instalação de ti-jolos e outros materiais submersos próximos da barragem, para a detecção de incrustação dos moluscos adultos. A resposta dos técnicos envolvidos foi altamente positiva. O grupo de trabalho come-çou a receber sugestões de atividades e convites para fazer a apresentação em eventos internos e externos. Ao mesmo tempo, FURNAS se fez repre-sentar na Força Tarefa Nacional para Controle do L. fortunei (mexilhão-dourado), constituída pelo Ministério do Meio Ambiente, por meio da Portaria no 494, de 22 de dezembro de 2003. Percebeu-se ali que o setor elétrico, a Marinha e demais envolvidos, articulam-se na busca por uma ação de controle ao L. fortunei. Merecem destaque as campanhas de divulgação promo-vidas pela CESP, coordenadas pelo agrônomo André Luis Mustafá, e as pesquisas financiadas pela CEMIG. A CESP gentilmente recebeu o grupo de FURNAS na UHE Sérgio Motta, para mostrar os procedimentos de manutenção adotados após a infestação do mexilhão-dourado. É importante destacar aqui a integração entre as empresas do setor elétrico. E deve ser destacado o papel de Itaipu Binacional, que foi a primeira empresa brasileira a discutir o tema, em 1997, antes mesmo da chegada do mexilhão no seu reservatório. No escritório central da empresa, o gru-po de trabalho viu que era necessário fazer algo complementar, ao que já se desenvolvia no setor. O setor de comunicação foi acionado, e o debate se ampliou com a participação de jornalistas e designers. As usinas sob conces-são de FURNAS estavam distantes de Itaipu,
de modo que foi decidido o investimento em ações preventivas. Foram avaliadas as prováveis rotas de embarcações de pesca esportiva e co-mercial, e quatro usinas foram definidas como de maior risco de infestação: Porto Colômbia e Marimbondo, no Rio Grande; Itumbiara, no Rio Paranaíba; e Manso, no Rio Manso. Essas usinas foram visitadas e o alerta foi ouvido com atenção. Todos os profissionais envolvidos se en-gajaram de forma entusiasmada na campanha. Sugestões brotavam de todos os setores visita-dos. Mesmo distantes do escritório central, as unidades regionais se organizaram e multipli-caram a informação. Os convites para eventos foram inúmeros. Era necessário produzir mate-rial e tornar a campanha institucional. Surgiram o folder, a cartilha com perso-nagens exclusivos, seguida da logomarca como personagem principal, impressa em bonés, camisetas, cadernos, canetas, cartazes e um número telefônico denominado “Disque Me-xilhão”. O conselho diretor da empresa criou formalmente o Grupo de Controle do Mexi-lhão-dourado, com recursos anuais da ordem de R$250.000,00. Estava constituída a cam-panha “Mexilhão-Dourado: Não dê carona a esse Bicho!”, cuja página pode ser acessada no endereço: http://www.furnas.com.br/meioam-biente_mexilhao2.asp. O material e as fotos logo despertaram o interesse da Editora Moderna, que há cinco anos introduziu o tema no conteúdo de seu li-vro Projeto Araribá Ciências, para turmas do 7º ano do Ensino Fundamental. Vale a pena detalhar um pouco a criação do Disque Mexilhão, pois é um exemplo do compromisso dos profissionais de FURNAS. A empresa dispõe de um serviço similar dedicado a receber avisos de queimada nas proximidades das linhas de transmissão sob sua concessão. Foi feita uma apresentação ao grupo de aten-dentes, seguida de uma discussão sobre como eles deveriam proceder. Havia um conjunto de perguntas e respostas já definidas, mas os atendentes pediram autorização para fazerem algumas adaptações. O resultado foi um fluxo-grama objetivo que definia se poderia se tratar de mexilhão-dourado, de Corbicula, ou de gas-trópodes. O primeiro evento oficial da campanha

CAPÍTULO 24
Seção 7
267
ocorreu em 2005, na UHE Porto Colômbia, em Planura, MG, que teve a participação do biólogo Sílvio Carlos Santos, da AES-Tietê, que relatou como a empresa estava lidando com o problema nos reservatórios do Rio Tietê. Em seguida foi lançada a campanha idealizada pelos técnicos de FURNAS, no encontro dos pescadores artesanais profissionais, promovido pela Colônia de Pescadores Z 10 na mesma ci-dade de Planura, onde foram distribuídos aos presentes camisetas, bonés, folders e cartilha alusivos à campanha, e realizada uma palestra. Daí em diante foram vários os eventos dos quais o grupo participou, desde seminários acadêmicos a festas populares locais e também palestra em escolas. Outra medida bem sucedida foi a parceria com empresas de co-a parceria com empresas de co-mercialização de material de pesca, para as quais foram disponibilizados fôlderes e carti-lhas, acrescentadas ao material adquirido pelos compradores. A receptividade era grande e teve alcance surpreendente, como foi o caso de um pesca-dor, Sr. Francisco, que ligou de Pedrinhas Pau-lista, na divisa de São Paulo com o Paraná, para o Disque Mexilhão. Ele disse que recebera o folheto juntamente com a mercadoria que com-prara numa casa de pesca de Minas Gerais. Fez questão de elogiar o material e avisava que o L. fortunei já havia invadido o Rio Paranapanema. Outro caso pitoresco, foi a apresentação para multiplicadores das cidades de Planura e Colômbia dois dias antes do Carnaval de 2006 (Fig. 1). A Banda Eva estava hospedada no mes-mo hotel da equipe de FURNAS. Depois de uma conversa com o empresário da banda – e da entrega dos kits de bonés, camisetas e ca-dernos – a cantora cumpriu sua promessa e, du-rante o show, gritou pela turma do mexilhão--dourado. Dois eventos foram os mais marcantes da campanha: a participação na expedição pelo Rio Paranaíba, articulada pelas prefeituras dos municípios do comitê de bacia que se formava; e no 26º Festival de Pesca de Cáceres. A expedição foi realizada em 2008. O objetivo principal era fazer um movimento po-lítico entre as prefeituras para incentivar a ade-são ao comitê de bacia hidrográfica que estava sendo constituído. A intenção era percorrer o Rio Paranaíba, desde a nascente até sua junção
com o Rio Grande, no reservatório de Ilha Sol-teira, com parada em algumas cidades para di-vulgação do comitê. A equipe de FURNAS deu suporte às medições de qualidade da água rea-lizadas pelas empresas SANEAGO, de Goiás, e COPASA, de Minas Gerais. Houve também o apoio de profissionais acadêmicos e alunos da Universidade Luterana do Brasil (ULBRA) da cidade de Itumbiara, com quem FURNAS já mantinha estreita relação. FURNAS mobilizou duas embarcações para fazerem todo o trajeto, três automóveis e dezoito funcionários. A estratégia era montar a base nos pontos de desembarque para fazer a desinfecção de todos os barcos, juntamente com a distribuição de cartilhas, folhetos e ade-sivos. Todos os funcionários usavam a camise-ta e o boné da campanha, e foram comprados cinco borrifadores (Fig. 2). Entre os membros da equipe havia um jornalista responsável pe-los contatos com a imprensa. Ao mesmo tempo eram feitos contatos com representantes de es-colas e das secretarias municipais de educação, para divulgar o tema e ceder material gráfico.
Figura 1. Multiplicadora de UHE Porto Colômbia, no estado de São Paulo, durante campanha de divulgação realizada na cidade.

De Filippo et al.
Ações de Eletrobras–FURNAS na divulgação da Campanha de Controle do Mexilhão-Dourado
268
O material da campanha foi distribuído para várias escolas dos três estados. O contato com professores e secretários de educação foi muito proveitoso (Fig. 3). A cartilha e o folheto criaram oportunidade para os educadores tra-balharem um tema específico em sala de aula, diretamente relacionado com o seu ambiente. Foram realizadas duas entrevistas em rá-dio, três entrevistas para TV e distribuídos relea-ses para jornais. A campanha de FURNAS foi di-fundida em pelo menos um site na Internet: http://www.paragonbrasil.com.br/news.php?id=988. Cerca de cinco mil cartilhas e folhetos foram distribuídos nos stands montados em cada parada (Fig. 4). Havia carência absoluta de informação sobre o mexilhão-dourado em alguns locais, de modo que o tema ficou per-manentemente vinculado à campanha realiza-da pela Empresa. A participação de FURNAS no 16º Festival de Pesca de Cáceres, no Mato Grosso, originou--se da preocupação com a circulação de embar-cações por todo o estado, devido aos inúmeros festivais de pesca organizados pelos municípios. Em 2006, o grupo saiu de Minas Gerais e Rio de Janeiro para divulgar a campanha da empresa no 26º Festival de Pesca de Cáceres (Fig. 5). A razão dessa participação era a pro-ximidade do local com o Aproveitamento Múl-tiplo de Manso (APM Manso) sob concessão da empresa. Para esse festival foi confecciona-do um adesivo para as embarcações. A presen-ça de FURNAS foi tão marcante que o stand da empresa foi visitado por todos (Fig. 6). As apresentações eram quase que contínuas. Pro-fessores locais solicitavam material para traba-lharem o tema em suas escolas. Após o evento, a Secretaria de Meio Am-biente do Mato Grosso convidou a empresa para participar do grupo de trabalho que de-veria criar uma estratégia de combate ao mexi-lhão-dourado no estado. O risco da dispersão do mexilhão era grande, devido aos inúme-ros eventos de pesca que ocorrem no estado. FURNAS colaborou ativamente com o grupo, do qual participavam as pesquisadoras Maria Cristina Dreher Mansur e Claudio Tasso Callil. Dali surgiu a proposta do projeto de pesquisa que está sintetizado neste livro. Um segundo desdobramento da parti-cipação no município de Cáceres foi a efetiva estruturação de um programa de prevenção do
Figura 2. Desinfecção de embarcação durante a expedição do Rio Paranaíba, em 2008.
Figura 3. Entrega de cartilhas para professoras do Ensino Fun-damental com cartilhas, em Inaciolândia, no estado de Goiás.
Figura 4. Movimentação no stand de FURNAS durante a expe-dição do Rio Paranaíba, em 2008.
Figura 5. Abertura de uma etapa de pesca do festival de Cá-ceres, no estado do Mato Grosso.

CAPÍTULO 24
Seção 7
269
mexilhão-dourado no estado do Mato Grosso. Em setembro 2007, o grupo foi convidado a proferir uma palestra para os membros da Se-cretaria de Meio Ambiente do Mato Grosso do Sul, acerca das ações tomadas por FURNAS para impedir a colonização do mexilhão-dou-rado, pois esse molusco estava sendo levado em direção à região de Bonito, no Mato Grosso do Sul, através do Rio Miranda. Em recente viagem ao Rio Teles Pires, na divisa com Goiás, foi documentado o adesivo referente ao festival de pesca de Paranaíba, Fest Praia, ocorrido em 2009, num barco que foi so-corrido devido a uma pane no motor (Fig. 7). Os livros de administração exortam as capacidades de liderança e gerencial como base para o sucesso de uma atividade ou carreira. A campanha desenvolvida por FURNAS parece ter rompido esse paradigma. Quem era o líder? Fato é que não havia liderança porque ela não era necessária. Todos sabiam exatamente o que fazer, e estavam livres para tomarem decisões. O que movia o grupo e que o levava a agregar mais gente em torno dele foram compromisso, entusiasmo, e capacidade de persuasão. Foram esses os principais ingredientes que multiplicaram a informação e fizeram da cam-panha uma referência para muitos. Um grupo heterogêneo e disperso por vários setores da empresa conseguiu fazer uma campanha de educação ambiental em cinco grandes estados brasileiros: São Paulo, Minas Gerais, Goiás, Mato Grosso e Mato Grosso do Sul. O que deveria exigir um enorme esforço de coordenação e controle fluiu naturalmente, evento após evento. Parte dos frutos já foi colhida, o mexilhão não foi introduzido nos reservatórios da empre-sa. A outra parte, os resultados de dois anos de pesquisa, está sendo apresentada agora. O grupo sempre acreditou que essa for-ma de divulgação adotada pela empresa foi a melhor maneira de difundir a existência do mexilhão-dourado, sua biologia e os danos que ele causa ao meio ambiente, pois se conseguiu que um número grande de pessoas tomasse co-nhecimento de sua existência, o que pode ter contribuído para retardar a introdução do mo-lusco no Rio Grande por quase quatro anos. Todo o trabalho da equipe de FURNAS culminou com a assinatura de um contrato de projeto de pesquisa e desenvolvimento com a
Figura 6. Animadores no stand de FURNAS em Cáceres, no estado do Mato Grosso.
Fundação Luiz Englert/UFRGS, denominado “Estudos biológicos e ambientais como base à prevenção e ao controle do mexilhão-dourado L. fortunei (Dunker, 1857), em rios e reser-vatórios brasileiros” objetivando desenvolver metodologias para o controle biológico do me-xilhão-dourado, contribuindo assim para um controle efetivo desse molusco que tem causa-do prejuízo ao meio ambiente e as hidrelétri-cas e estações de tratamento de água nos locais onde ele está ocorrendo.
Figura 7. Adesivo do Fest Praia, no município de Paranaíta, no estado do Mato Grosso.


271
O Ministério do Meio Ambiente – MMA, considerando os crescentes impactos negativos gerados pelas espécies exóticas invasoras e as dificuldades de controle e monitoramento e, considerando ainda, as fragilidades existentes para o enfrentamento da introdução de novas espécies, a vulnerabilidade natural de um país de proporções continentais e os elevados custos decorrentes da dispersão de espécies exóticas invasoras no País, decidiu tratar o tema como uma de suas prioridades. A primeira reunião sobre o tema ocor-reu em 2001, quando o MMA em parceria com a Embrapa, realizou, em Brasília, a “Reu-nião de Trabalho sobre Espécies Exóticas In-vasoras”. O evento contou com a participação dos países da América do Sul (Argentina, Bolí-via, Brasil, Chile, Colômbia, Equador, Guiana Francesa, Paraguai, Peru, Suriname, Uruguai e Venezuela) e com a colaboração do Governo dos Estados Unidos da América e do Progra-ma Global para Espécies Exóticas Invasoras – GISP. Entre as recomendações aprovadas
destacam-se: prevenção e controle dos im-pactos de espécies exóticas invasoras sobre os ecossistemas naturais e sobre a rica biodiver-sidade da região; elaboração de diagnósticos nacionais, pesquisa, capacitação técnica, for-talecimento institucional, sensibilização pú-blica, coordenação de ações e harmonização de legislações; atenção urgente ao problema de introdução de espécies exóticas invasoras nas diferentes bacias hidrográficas da região e ecossistemas transfronteiriços; e coordenação e cooperação entre os setores agrícolas, flo-restais, pesqueiros e ambientais nacionais no tratamento dessa questão. Atendendo algumas das recomendações decorrentes desta reunião internacional, o MMA vem desenvolvendo ações voltadas à pre-venção das introduções; ao monitoramento; ao controle/manejo; e à erradicação. Estas ações dizem respeito à revisão e desenvolvimento de normativas relacionadas à matéria; realização de inventários das espécies exóticas ocorrentes nos diversos ecossistemas brasileiros, inclusive
CAPÍTULO 25
Ações desenvolvidas pelo Ministério do Meio Ambiente
2001 a 2011
Vivian Beck Pombo

Pombo, V. B.
Ações desenvolvidas pelo Ministério do Meio Ambiente - 2001 a 2011
272
no âmbito de bacias hidrográficas; discussões relativas à elaboração de listas oficiais de es-pécies exóticas invasoras em âmbito estadual e nacional; e estímulo à abertura de linhas de fi-nanciamento para pesquisa no Fundo Nacional de Meio Ambiente. Parceria Fundo Nacional do Meio Ambiente – FNMA e Projeto de Con-servação e Utilização Sustentável da Diversida-de Biológica Brasileira – PROBio. Ainda em 2001, o MMA lançou o Edital FNMA/PROBio 04/2001. A partir desse Edital foram selecionados nove projetos relacionados a espécies invasoras presentes no país que ser-viram de base para o início dos trabalhos no Departamento de Conservação da Biodiversi-dade, que deu continuidade ao desenvolvimen-to das ações, a exemplo das descritas a seguir.
Primeiro Informe Nacional sobre Espécies Exóticas Invasoras
Em 2003, o MMA, considerando suas responsabilidades frente ao tema e diante da constatação da precariedade de referências bibliográficas e de pesquisas no Brasil relacio-nadas à questão, decidiu realizar o I Informe Nacional sobre Espécies Exóticas Invasoras, com dois diagnósticos, um sobre as “Espécies Exóticas Invasoras Atuais e Potenciais” e ou-tro sobre a estrutura existente no país para tratar o problema. Os levantamentos abor-daram as espécies que afetam os ambientes terrestres, marinho e de águas continentais, bem como aquelas que afetam os sistemas de produção e a saúde humana. Como resultado dessa iniciativa, foram listadas 543 espécies na condição de exóticas invasoras aos dife-rentes ecossistemas brasileiros, sendo: 176 de organismos que afetam o ambiente terrestre; 66 que afetam o ambiente marinho; 49 que afetam as águas continentais; 155 que afetam os sistemas de produção; e 97 que afetam a saúde humana. Tal levantamento foi realizado com re-cursos financeiros do Projeto de Conservação e Utilização Sustentável da Diversidade Bioló-gica Brasileira – PROBio, que firmou convê-nios com a The Nature Conservancy/Instituto Hórus, com a Fundação de Estudos e Pesqui-sas Aquáticas – Fundespa, com a Fundação Arthur Bernardes – Funarbe, com a Empresa
Brasileira de Pesquisa Agropecuária – Embra-pa e com a Fundação para o Desenvolvimento Científico e Tecnológico em Saúde – Fiotec foram executados cinco subprojetos relaciona-dos, respectivamente, às espécies que afetam o ambiente terrestre, o ambiente marinho, as águas continentais, os sistemas de produção (agricultura, pecuária e silvicultura) e a saúde humana. Os relatórios apresentados após o levan-tamento estão em fase de revisão e sistematiza-ção visando à publicação de livros específicos sobre cada um dos temas abordados no levan-tamento, porém podem ser consultados em sua versão original no endereço: http://www.mma.gov.br/sitio/index.php?ido=publicacao.publicacoesPorSecretaria&idEstrutura=14.
Força Tarefa Nacional para o Controle do Mexilhão-Dourado
Por meio da Portaria nº 494, de 22 de de-zembro de 2003, a Ministra do Meio Ambiente criou, em caráter emergencial, a Força Tarefa Nacional para o Controle do Mexilhão-dou-rado – FTN, visando o controle do mexilhão--dourado. Ao final das atividades do Grupo de Trabalho relativo à Força Tarefa Nacional foi elaborado o “Plano de Ação Emergencial para o Controle do Mexilhão-Dourado”, com de-talhes sobre componentes de fiscalização, ca-pacitação, monitoramento e comunicação que deveriam ser contemplados com ações para o controle das populações já estabelecidas e ade-quação da legislação para prevenção de novas introduções. O Relatório da Força Tarefa lista as principais conclusões, ou seja: i) o controle e contenção da dispersão do mexilhão-dourado devem-se constituir em tarefas de caráter per-manente; ii) deve haver restrição nas atividades de tráfego hidroviário; iii) devem ser amplia-das as ações de fiscalização de embarcações de pequeno porte de pesca e recreio transporta-das por via rodoviária; iv) maior controle no transporte de fauna e flora; e v) desaconselha a transposição de águas e aconselha um maior rigor na fiscalização em relação às navegações internacional e de cabotagem. Após a finalização dos trabalhos da FTN, o MMA entendeu que as informações

CAPÍTULO 25
Seção 7
273
existentes até aquele momento, bem como as informações técnicas sobre o controle do mo-lusco, não eram suficientes para subsidiar a tomada de decisões. A FTN considerou ainda a necessidade de serem disponibilizadas infor-mações científicas sobre a biologia, a ecologia e o comportamento da espécie Limnoperna for-tunei (Dunker, 1857). Assim, entendeu neces-sário o desenvolvimento de estudos e pesquisas voltados à identificação de formas de controle da infestação das águas interiores brasileiras pelo mexilhão-dourado, o desenvolvimento de tecnologias para mitigação dos seus impactos sobre os múltiplos usos da água, a exemplo da geração de energia elétrica, captação de água para diversos fins, navegação, pesca e aquicul-tura. Para a execução dos estudos fomenta-dos pelo CNPq, foi organizada uma rede de pesquisa, com a contratação de nove (9) ins-tituições, atuantes na temática, a seguir dis-criminadas: a Pontifícia Universidade Católica do Rio Grande do Sul – PUCRS; a Universida-de Estadual de Maringá – UEM; o Instituto de Estudos do Mar Almirante Paulo Morei-ra – IEAPM; a Empresa Brasileira de Pesquisa Agropecuária – EMBRAPA Pantanal; a Uni-versidade Federal de Mato Grosso – UFMT; a Universidade Federal do Paraná – UFPR; a Pontifícia Universidade Católica do Rio de Janeiro – PUCRJ; a Universidade Federal de São Carlos – UFSC; o Centro de Pesquisa e Gestão de Recursos Pesqueiros Continentais – CEPTA IBAMA. A coordenação ficou a cargo do IEAPM. O estudo acima referido encontra-se registrado no CNPq sob o Processo de nº 507675/ 2004.
GT CONAMA para Revisão da Portaria IBAMA nº 145/98
Desde 2005, discute-se, em um Grupo de Trabalho – GT criado no âmbito do CONA-MA, a elaboração de instrumento normativo, que estabeleça critérios para a introdução, rein-trodução e a translocação de organismos vivos em ambientes aquáticos. Esse GT, designado pela Câmara Técnica de Biodiversidade, Fauna e Recursos Pesquei-ros tem a incumbência de elaborar uma minuta de Resolução para ser apresentada à plenária
da Câmara Técnica do CONAMA, para deli-beração. Tal Resolução, se aprovada, deverá substituir a Portaria IBAMA nº 145, de 29 de outubro de 1998, em vigor.
I Simpósio Brasileiro sobre Espécies Exóticas Invasoras
No ano de 2005, o MMA e o Instituto Brasileiro do Meio Ambiente e dos Recursos Renováveis – IBAMA, em parceria com a Fio-cruz, a Embrapa, o Instituto Oceanográfico da USP – IOUSP, Instituto Hórus e a Universidade de Viçosa – UFV, realizaram o I Simpósio Bra-sileiro sobre Espécies Exóticas Invasoras, no período de 4 a 7 de outubro, em Brasília, DF. O Simpósio contou com a presença de sete países: África do Sul, Argentina, Brasil, Estados Unidos, Jamaica, Nova Zelândia e Portugal. En-tre os atores brasileiros, tivemos representação das cinco regiões geopolíticas. A abertura do evento foi presidida pela então ministra Marina Silva, que em seu discurso propôs a criação de uma Câmara Técnica Permanente sobre Espé-cies Exóticas Invasoras, no âmbito da Comissão Nacional de Biodiversidade – CONABio. Todo material apresentado no Simpósio e autorizado pelos autores está disponível na página do MMA no endereço www.mma.gov.br/invasoras.
Publicação Espécies Exóticas Invasoras: Situação Brasileira
Por ocasião da realização da VIII Confe-rência das Partes – COP8, da Convenção sobre Diversidade Biológica, em 2006, o MMA ela-borou e publicou um livreto com 24p, deno-minado “Espécies Exóticas Invasoras: Situação Brasileira”, com breves informações das ações até então desenvolvidas por este órgão central da Política de Meio Ambiente no Brasil. Esta publicação (Brasil, 2006) encon-tra-se disponível para download no endereço http://www.mma.gov.br/estruturas/174/_publi-cacao/174_publicacao17092009113400.pdf
Resolução CONAMA nº 394/07
Em atendimento as recomendações da FTN, o MMA participa diretamente da ela-

Pombo, V. B.
Ações desenvolvidas pelo Ministério do Meio Ambiente - 2001 a 2011
274
boração de instrumentos normativos para pre-venir a introdução, não somente do mexilhão--dourado, como também, de outras espécies exóticas invasoras. Nesse contexto, é importante ressaltar a Resolução CONAMA nº 394, de 6 de novem-bro de 2007, que “estabelece os critérios para a determinação de espécies silvestres a serem criadas e comercializadas como animais de es-timação”. A prática de adoção de animais de es-timação exóticos tem sido uma das causas de introdução e dispersão de espécies exóticas potencialmente invasoras, em razão da soltu-ra desses em ambientes naturais, quando seus proprietários desistem de sua guarda e os des-cartam.
Câmara Técnica Permanente sobre Espécies Exóticas Invasoras – CTPEEI
A CTPEEI é um fórum específico para o debate do tema. Criada pela Deliberação CONA-CONA-Bio nº 49, de 30 de agosto de 2006, sofreu alteração em sua composição pela Deliberação nº 55, de 21 de outubro de 2008. A CTPEEI foi instalada em 2008 e desde então, foram realizadas cinco reuniões, cuja memória e documentos avaliados estão disponíveis no endereço http://www.mma.gov.br/index.php?ido=conteudo.monta&idEstrutura=15&idConteudo=7434.
Estratégia Nacional sobre Espécies Exóticas Invasoras
Em decorrência dos trabalhos da CTPEEI, foi elaborada e aprovada, em outubro de 2009, no âmbito da Comissão Nacional de Biodiversidade, a Estratégia Nacional sobre Es-pécies Exóticas Invasoras, conforme Resolução CONABIO nº 5/2009. A Estratégia Nacional se constitui no pri-meiro documento aprovado no âmbito do Go-verno Federal que pode orientar as diferentes esferas do governo no trato das questões relati-vas às espécies exóticas invasoras. Obviamente, legislações específicas serão necessárias para prevenir ou diminuir a introdução e a translo-cação de exóticas invasoras no país. Ainda, a Estratégia Nacional representa, um importante
instrumento para a internalização e implemen-tação no país do artigo 8 (h) da Convenção so-bre Diversidade Biológica. Da mesma forma, a Estratégia se traduz em uma efetiva ferramenta que o país dispõe para a consecução das deter-minações das Decisões V/8, VI/23 e IX/4, das Conferências das Partes, da CDB.
Informe sobre as Espécies Exóticas Invasoras Marinhas no Brasil
Em dezembro de 2009 foi lançado o pri-meiro de uma série de cinco livros que aborda-rão a situação das espécies exóticas invasoras nos diferentes ambientes (marinho, de águas continentais e terrestres), sistemas de produção e saúde humana. O primeiro livro está contri-buindo com as informações científicas básicas sobre as espécies exóticas marinhas e deverá ser decisivo como subsídio para a tomada de decisões e planejamento das ações necessárias para o enfrentamento das invasões biológicas no país. Esta publicação encontra-se disponível para download no endereço: h t t p: / /w w w.mma.gov.br/estruturas/174/_publicacao/174_publicacao16032010050723.pdf
Estratégia Nacional de Comunicação e Educação Ambiental - ENCEA
Finalmente, e considerando que as espé-cies exóticas invasoras representam a segunda maior causa de perda de biodiversidade em ní-vel mundial, a primeira no âmbito das Unida-des de Conservação – UCs e, considerando ain-da que essas UCs representam redutos chaves para a preservação da diversidade biológica nos diferentes biomas brasileiros, o Departamento de Conservação da Biodiversidade, da Secreta-ria de Biodiversidade e Florestas, em conjun-to com outras secretarias do MMA, elaborou um documento que está sendo chamado “Es-tratégia Nacional de Comunicação e Educação Ambiental no Âmbito do Sistema Nacional de Unidades de Conservação – ENCEA”. Tal do-cumento deverá definir os princípios, as dire-trizes, os objetivos e os métodos para o forta-lecimento e a implementação de atividades de

CAPÍTULO 25
Seção 7
275
comunicação e de educação ambiental em UCs, Corredores Ecológicos, Mosaicos de UCs, Re-servas da Biosfera, e seus entornos, incluindo o tema “espécies exóticas invasoras e as invasões biológicas”, finalizada e publicada em dezem-bro de 2010, disponível nos endereços: www.mma.gov.br/ea; encea.blogspot.com e www.icmbio.gov.br.
Espécies Exóticas Invasoras de Águas Continentais no Brasil
Em fase de finalização, o MMA vem trabalhando no livro que irá apresentar a com-pilação dos resultados apresentados em 2005, no relatório final do I Informe Nacional sobre Espécies Exóticas Invasoras relativo às águas
continentais, com atualizações e acréscimo de informações. A publicação será composta de 11 capí-tulos, trazendo dados sobre as espécies exóticas já presentes no país, suas situações populacio-nais, características, origem, distribuição geo-gráfica e outras, dos seguintes grupos taxonô-micos: microrganismos; cnidários; crustáceos e microcrustáceos; moluscos; peixes; anfíbios e répteis e macrófitas. Este livro também será um marco, uma vez que temos carência de informações cientí-ficas compiladas sobre as espécies exóticas de águas continentais. Sendo, portanto, um impor-tante documento para subsidiar as decisões dos gestores e usuários dos recursos hídricos e de espécies exóticas para produção em larga escala.


Seção
8Delineamento experimental: seleção de métodos de controle populacional


279
Quando um cientista elabora uma hi-pótese de trabalho é necessário que estabeleça critérios que validem de forma irrefutável se a mesma é válida ou não, e é neste ponto que o delineamento experimental entra no processo, visto que experimentos mal projetados ou mal conduzidos podem levar a conclusões errôneas. O delineamento experimental é a base onde está apoiado todo o sistema científico, e consiste no planejamento e condução de ex-perimentos de modo que seja possível o reco-lhimento de dados que possam ser analisados, usando métodos estatísticos apropriados e que conduzam a conclusões válidas e objetivas. O desenvolvimento e adaptação de tec-nologias que promovam o controle populacio-nal de espécies invasoras, com a minimização de impacto de subprodutos e metabólitos tóxi-cos, requerem métodos científicos adequados. Para tal, a correta abordagem experimental re-quer um planejamento otimizado visando obter respostas rápidas e precisas quanto à eficácia de métodos de controle, dosagens adequadas e suas implicações ambientais.
As espécies de moluscos invasores lím-nicos que têm sido alvo de preocupação quan-to aos impactos econômicos são Limnoperna fortunei (Dunker, 1857), Corbicula largillierti (Philippi, 1844) e Corbicula fluminea (Müller, 1774). Portanto, a maior parte do conhecimen-to experimental resulta de projetos de pesquisa e desenvolvimento no controle destes bivalves. Desta forma muitos dos exemplos citados neste capítulo se referem a estas espécies, mas podem ser utilizados como referência no planejamento de experimentos que visem testar métodos de controle para outras espécies invasoras.
1. Tipos de experimentos
A experimentação do uso de substâncias químicas, radiações, ondas sonoras ou até mes-mo, inimigos naturais, no controle de molus-cos invasores, pode ser realizada de diferentes formas com diferentes métodos e tempos de execução. O objetivo, o custo e viabilidade são as características que costumam limitar os ex-perimentos (Zagatto & Bertoletti, 2006).
CAPÍTULO 26
Planejamento experimental para a seleção de métodos de controle
populacional de moluscos invasores
Daniel PereiraSávio Henrique Calazans CamposLivia Viana de Godoy Fernandes
Isabel Cristina Padula PazCintia Pinheiro dos SantosFlavio da Costa Fernandes
Maria Teresa Raya RodriguezAlexandre Arenzon
Maria Cristina Dreher Mansur

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
280
Gambetta (2011) define muito bem as fases de uma pesquisa experimental desde as fases de laboratório até a sua aplicação na in-dústria, como segue: * Escala de bancada. Os experimentos conduzidos em laboratório são geralmente re-ferenciados como “em escala de bancada” e se caracterizam pela escala reduzida de uma si-tuação hipoteticamente real. Em linhas gerais uma boa fase de pesquisa em bancada gera re-sultados sólidos e confiáveis que permitem afir-mar que a tecnologia é promissora. * Escala piloto. Os estudos em escala piloto são a solução adotada para sanar os li-mitantes encontrados em escala de bancada. O principal entrave à utilização dessas plantas--piloto são os custos envolvidos no desenvolvi-mento, montagem e operação de tais unidades. Esses custos, entretanto, são ordens de gran-deza acima dos investimentos na montagem de uma unidade de bancada, mas muito inferio-res aos de uma tentativa frustrada de construir uma planta industrial com base em premissas equivocadas ou em informações obtidas em es-cala de bancada. * Escala industrial. A partir do proje-to da planta-piloto e das informações obtidas ao longo de sua operação, pode-se iniciar um projeto de planta industrial, levando em conta as características técnicas e econômicas, que permitirão construir uma planta industrial competitiva. O marco final de uma pesquisa científica bem sucedida é exatamente quando essa nova tecnologia é adotada em uma planta industrial de forma que seja competitiva frente a outros processos e ou produtos e se complete o ciclo da inovação. Dessa forma, recupera-se o investimento realizado na pesquisa e no de-senvolvimento, garantindo a sustentabilidade do empreendimento em termos econômicos, ambientais e sociais.
2. Fases de desenvolvimento do organismo alvo
O planejamento de experimentos com moluscos invasores límnicos implica em uma primeira pergunta: qual fase de desenvolvi-mento do animal deve ser exposta ao agente controlador em meu experimento? Os molus-cos límnicos invasores (bivalves e gastrópodes)
apresentam uma fase planctônica, na forma larval, e, duas fases bentônicas: recruta e adul-ta. A escolha das fases adequadas para a experi-mentação deve considerar uma série de fatores, dentre eles, a disponibilidade da mesma no am-biente aquático para coleta. A Tabela I discri-mina as vantagens e desvantagens de cada fase em experimentos de seleção de agentes contro-ladores.
3. Manutenção dos organismos-alvo em laboratório
Escolhida a fase a ser testada em um ex-perimento, um novo desafio surge: como man-ter o organismo vivo em meu laboratório? Até o momento não foi possível reprodu-zir o mexilhão-dourado em laboratório, assim como as espécies do gênero Corbicula. Desta forma, os organismos-teste não são reproduzi-dos, mas apenas mantidos em laboratório sob condições adequadas à sua sobrevivência. O sucesso da manutenção de organismos-alvo em laboratório depende de várias etapas desde sua coleta, transporte e manutenção para a sobre-vida, até o seu uso.
3.1. Coleta do organismo-alvo
A coleta de larvas de L. fortunei é feita diretamente no plâncton e as amostras devem ser transportadas sob refrigeração (ver Capí-sob refrigeração (ver Capí-tulos 10 e 11). Alguns cuidados devem ser tomados durante o transporte de indivíduos adultos: no caso de espécies do gênero Corbi-cula, o ideal é coletar os organismos e enrolar em algodão ou gaze umedecida e acondicioná--los em potes plásticos, que são posteriormen-te colocados em isopor com gelo em escamas. Desta forma se reduz o metabolismo do ani-mal, que não realizará a filtração. E não ha-vendo água não se observa efeitos da degrada-ção da qualidade da mesma sobre os bivalves. Esta forma de acondicionamento é apropriada para larvas de espécies do gênero Corbicu-la, que tem curta permanência no plâncton. Quando o animal é desembalado e colocado em bandeja com água reconstituída (ver item 4.2.3 deste capítulo) acrescida de alimento com base em algas e ração de peixe, as larvas são liberadas logo em seguida na bandeja e po-

CAPÍTULO 26
Seção 8
281
dem ser facilmente removidas com pipeta de Pasteur. Pouco se conhece sobre a reprodução destes organismos sendo difícil reproduzi-los em laboratório. Quanto ao mexilhão-dourado (L. fortu-nei), com base no conhecimento de sua capaci-dade de resistência ao dessecamento (Montalto & Drago, 2003), o transporte sem a imersão em água oferece uma condição adequada sem per-
das consideráveis (Campos, 2009). Transporte similar ao relatado para os bivalves do gênero Corbicula também podem ser realizados desde que os aglomerados sejam bem lavados e des-membrados, cuidando-se para não danificar o pé do animal no momento da desagregação. O transporte em água é possível, mas requer aten-ção à degradação da qualidade da mesma que depende do tempo de deslocamento.
Tabela I. Vantagens e desvantagens do uso de diferentes fases de desenvolvimento de moluscos invasores em experimentos visando à seleção de métodos de controle populacional.
Fases Vantagens Desvantagens
Larvas
A coleta é facilmente realizada com rede de plâncton.
É a fase inicial do desenvolvimento, desta forma o controle das larvas inibe a incrustação ou acumulação.
Poderá requerer concentrações ou doses letais menos elevadas.
A presença de larvas no plâncton pode ser determinada pela sazonalidade.
Indivíduos muito pequenos dificultando a avaliação da letalidade.Quando grandes volumes de água são tratados, a sele-ção e quantificação de organismos são mais difíceis.
A utilização de amostras concentradas de plâncton nos experimentos impossibilita a utilização de um número padrão de indivíduos gerando muita variabilidade nos dados.
Difícil manutenção em laboratório o que implica em coleta de organismos a cada experimento.
Recrutas
Organismos recrutados em substratos artificiais tem tamanho mais uniforme do que indivíduos recrutados em substratos naturais.
É uma das fases de maior mobilidade, sendo interessante compreender a ação dos métodos de controle nesta fase.
É possível padronizar o número de indivíduos nos testes com facilidade.
Manutenção em laboratório é possível.
A presença de recrutas no macrofauna bentônica pode ser determinada pela sazonalidade.
O recrutamento de organismos incrustantes ocorre em substratos artificiais num tempo mínimo de exposição de três meses.
Indivíduos muito pequenos dificultando a avaliação da letalidade.
Adultos
A coleta é facilmente realizada manualmente ou por meio de amostradores e substratos artificiais.
Indivíduos grandes facilitam a avaliação da letalidade.
Organismos recrutados em substratos artificiais tem tamanho mais uniforme do que indivíduos recrutados em substratos naturais.
O controle dos adultos inibe a produção larval.
Os organismos podem ser coletados durante todo o ano.
É possível padronizar o número de indivíduos nos testes com facilidade.
Manutenção em laboratório é possível.
Requer maiores cuidados com manutenção (alimenta-ção, renovação do meio, etc).
Poderá requerer concentrações ou doses letais mais elevadas.

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
282
3.2. Condições de manutenção
A manutenção do mexilhão-dourado em laboratório depende do controle da qualidade da água, a qual está diretamente relacionada a alguns parâmetros tais como temperatura (24-26ºC), pH (6,5-7,5), oxigenação (Saturado), acúmulo de metabólitos (troca de água).
3.3. Alimentação
As relações ideais para sustentar uma população de mexilhões em função de volu-mes de água não são definitivas. Em seu estu-do, Campos (2009) utiliza caixa de água para manutenção e aclimatação dos organismos em laboratório, alimentando-os com um composto à base de Saccharomyces cerevisae fornecidas duas vezes ao dia e complementada com as mi-croalgas Ankistrodesmus sp. e Pseudokirchne-riella subcapitata
3.4. Cuidados de biossegurança: o que fazer para o organismo não fugir do laboratório?
Quanto à biossegurança, deve-se ado-tar medidas para a manipulação adequada de agentes biológicos, químicos e físicos para prevenir acidentes e reduzir riscos inerentes às atividades desenvolvidas. Tratando-se de um organismo invasor torna-se necessário adotar práticas para a sua contenção dentro do labo-ratório, principalmente em áreas que tenham ambientes não invadidos e propícios para que o mexilhão possa se estabelecer (rios, lagos e lagoas). A adoção de boas práticas laborato-riais é essencial para que se evite a dispersão de larvas, juvenis ou mesmo adultos na drenagem urbana. Algumas práticas recomendadas para manter um nível adequado de biossegurança e evitar a dispersão deste organismo para outros corpos hídricos são listadas a seguir.
* Capacitação das pessoas que manipu-lam os moluscos invasores; * Controle do acesso ao laboratório de experimentos, cultivo/manutenção; * Limpeza adequada dos estabelecimen-tos e equipamentos do laboratório com agentes
(substância oxidante) minimizando a possibili-dade de transporte de larvas e juvenis; * Uso de EPIs, vidrarias e equipamentos minimizando a possibilidade de dispersão do organismo para fora da área controlada do la-boratório; * Tratar a água descartada do cultivo, que deve ser renovada a toda semana, tratar descartes de resíduos, como conchas e animais mortos, com substâncias oxidantes; * Controlar e tratar efluentes mantendo um sistema sanitário diferenciado.
4. Planejando um experimento
O planejamento do experimento come-ça com a seleção adequada dos indivíduos da espécie-alvo, sua adequada manutenção em sistema de cultivo, tipo de meio de cultivo, de-senho experimental (delineamento), escolha dos testes estatísticos a análise de dados, e, finalmente, a validação dos métodos com base em testes ecotoxicológicos, visando avaliar os possíveis danos ambientais potenciais do tipo de método de controle populacional sobre a biota aquática ou até mesmo sobre operadores do sistema de controle. A desconsideração de qualquer um destes passos pode invalidar os dados obtidos em um experimento mal plane-jado.
Figura 1. Sistema de tanques para manutenção de mexilhões em laboratório (Características: 500 Litros; 24-26ºC; pH: 6,5-7,5; Oxigênio saturado por aeração constante; Circulação de água constante; Filtração em sistema de filtro biológico e substituição semanal de um terço do volume dos tanques) (Campos, 2009).

CAPÍTULO 26
Seção 8
283
4.1. Seleção e preparação dos organismos alvos
Para experimentos com larvas do mexi-lhão-dourado, são geralmente escolhidas lar-vas véliger de charneira reta e véliger umbona-da de acordo com Santos et al. (2005). Para os experimentos com adultos, dentre os critérios de seleção, devem ser considerados além do tamanho (Fig. 2), características vitais como produção de bisso, filtração e locomoção. Os organismos que não produzem bisso ou que apresentam suas conchas abertas sem atividade filtrante devem ser descartados. No caso de or-ganismos adultos, é aconselhável a limpeza das conchas com escova de dente para que se remo-vam algas perifíticas com potencial de desen-volvimento dentro da unidade experimental.
4.2. Meio de Cultivo
No caso dos organismos aquáticos o meio de cultivo é o tipo de água na qual o mesmo vai permanecer durante todo o experimento. A qua-lidade desta água implica em diferentes interfe-rências nos resultados finais do experimento. Portanto a escolha de um determinado meio de cultivo deve considerar os objetivos do experi-mento, o tempo de duração do mesmo, a viabili-dade técnica e possíveis implicações da qualidade do meio sobre o desenvolvimento e manutenção do organismo alvo e na qualidade dos resultados.
4.2.1. Água bruta
A utilização da água bruta em experi-mentos tem como objetivo avaliar a interferên-cia da qualidade da água do local sobre o mé-todo de controle (físico, químico ou biológico). A água bruta pode ser um meio utilizado em testes de bancada e estação piloto. Santos, C. P. (2011) avaliou a ação da luz ultravioleta sobre larvas do mexilhão-dourado em água bruta em experimento utilizando estação-piloto, com o objetivo de avaliar a relação da turbidez com a eficiência do tratamento. Soares et al. (2009) utilizaram água bruta para avaliar a toxicidade do sulfato de cobre sobre indivíduos adultos do mexilhão-dourado em testes de bancada. Os autores escolheram utilizar água bruta, devido à presença de sólidos suspensos. A presença de
sólidos na água é um fator determinante para a incorporação de metais por organismos filtra-dores (Callil & Junk, 1999).
4.2.2. Água deionizada
A utilização da água deionizada em ex-perimentos busca eliminar a interferência da qualidade da água sobre o método de controle (físico, químico ou biológico). A água deioni-zada pode ser utilizada em testes de bancada e estação-piloto, onde o tempo de residência do fluido nos canos e muito pequeno e o experi-mento de curto período de duração. Santos, C. P. (2011) avaliou a ação da luz ultravioleta sobre larvas do mexilhão-dourado em água deioniza-da, em experimento utilizando estação-piloto, com o objetivo de eliminar turbidez e aumen-tar eficiência do tratamento. A desvantagem da água deionizada é que pode causar desequilíbrio osmótico nos organismos-alvo mais sensíveis, principalmente em experimentos com maior duração. No entanto, Santos, C. P. (2011) não detectou diferenças significativas quanto à mor-talidade entre grupos controle com água bruta e água deionizada em seus experimentos.
4.2.3. Água reconstituída
O uso de água reconstituída (Fig. 3) em ensaios com organismos aquáticos é padrão em ensaios ecotoxicológicos, pois suas caracterís-ticas físico-químicas são mais estáveis devido ao tamponamento. Águas podem se reconstitu-ídas a partir de águas brutas ou águas deioni-zadas. Desta forma as características químicas das águas são ajustadas para aquelas desejadas no experimento. No caso da reconstituição da água a partir da água deionizada, é garantida a ausência de matéria orgânica, que pode degra-
Figura 2. Seleção de tamanho de L. fortunei para experimen-tos em laboratório (Fernandes & Coutinho, 2008).

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
284
dar-se e alterar a qualidade da água. Pereira et al. (2011b) utilizaram água reconstituída para a realização de testes visando à seleção de agen-tes microbianos no controle de L. fortunei.
4.3. Delineamento
4.3.1. Número de indivíduos
O número de indivíduos usados em ex-perimentos necessita ser considerado em fun-ção da variabilidade e representatividade ob-
tida dentro das análises estatísticas a serem adotadas. Estudos de experimentos desenha-dos para cálculos de concentração letal (CL50) com o mexilhão-dourado geralmente utilizam um número de 10 ou 20 indivíduos em todas as réplicas e de 3 a 5 replicações das concentra-ções (Patrício et al., 2003; Soares et al., 2009; Campos, 2009). Pereira et al. (2011b) utiliza-ram 10 indivíduos e 5 replicações para cada tratamento. Um caminho a ser adotado antes da definição do delineamento do experimento é realizar experimentos pilotos com 10, 20 ou 30 indivíduos e 3 a 5 replicações das concen-trações e escolher o modelo mais otimizado do experimento com menor variação dos dados.
4.3.2. Repetição
O experimento deve ser repetido no mí-nimo três vezes para validação dos resultados, confirmando que os mesmos são devido ao agente testado, e não fruto do acaso.
4.3.3. Controle negativo e positivo
É fundamental que todo planejamento de experimento inclua um controle (testemunha) que seja mantido sobre as mesmas condições dos organismos-alvo, apenas sem a inserção do composto/organismo a ser avaliado. Esse tipo de controle também é chamado controle negativo. Em ensaios onde já são conhecidos compostos ou organismos que não apresentem efeitos sobre o organismo-alvo (efeito neutro), esses são utilizados nas mesmas condições de todas as réplicas do ensaio, sendo denominado controle positivo.
4.3.4. Aleatorização
Impedir que a heterogeneidade de fato-res ambientais que ocorrem de forma diferen-cial no espaço possam influenciar os resultados dos ensaios.
4.3.5. Condições ambientais da sala de experimentos
Por se tratar de experimentos com orga-nismos aquáticos não é necessário controle da umidade da sala. A temperatura da sala deve permanecer entre 21 e 22ºC. Não é necessária
Figura 3. Procedimento para a preparação de água reconsti-tuída.

CAPÍTULO 26
Seção 8
285
a iluminação (fotoperíodo), já que as condições de aplicação em campo, onde as incrustações ocorrem muitas vezes em tubulações e equipa-mentos, não apresentam luminosidade.
4.3.6. Exemplos de experimentos
São descritos a seguir alguns modelos de experimentos realizados para selecionar méto-dos de controle de moluscos invasores.
4.3.6.1. Controle biológico: seleção de agentes microbianos Um exemplo de experimento, em escala de bancada, para a avaliação da ação de agen-tes microbianos e ou produtos formulados so-bre indivíduos adultos do mexilhão-dourado é descrita a seguir (Fig. 4).
4.3.6.2 Controle físico Dentre os métodos de controle físico são descritos abaixo, dois exemplos de experimen-tos, um com ondas ultrassônicas e outro com luz ultravioleta.
4.3.6.2.1. Ultrassom Um exemplo de experimento, em escala de bancada, para a avaliação da ação de ondas
ultrassônicas sobre larvas de L. fortunei e C. fluminea é descrita a seguir (Fig. 5).
4.3.6.2.2. Luz ultravioleta (UV) Um exemplo de experimento, em escala piloto, para avaliação da ação germicida de luz ultravioleta sobre larvas do mexilhão-dourado é descrita a seguir (Fig. 6) de acordo com San-tos, C. P. (2011).
4.3.6.3. Controle químico: substâncias moluscidas Um exemplo de experimento, em esca-la de bancada, para a avaliação da ação de um moluscida (sulfato de cobre) sobre indivíduos adultos de L. fortunei é descrito a seguir (Fig. 7), de acordo com Soares et al. (2009). O exem-plo descrito pode ser adaptado para avaliação de outras substâncias químicas.
5. Avaliação
A avaliação dos resultados dos experi-mentos requer, primeiramente, o estabeleci-mento de critérios de avaliação da mortalidade, bem como cálculos e procedimentos estatísti-cos adequados para consolidar o experimento através da correta interpretação dos resultados,
Figura 4. Procedimento para a preparação de experimento para seleção de agentes microbianos.

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
286
Figura 5. Procedimento para a preparação de experimento para avaliar a atuação de ondas ultrassônicas no controle da popu-lação larval de L. fortunei e C. fluminea.
bem como fornecer subsídios para a tomada de decisão referente à aplicação da tecnologia em campo.
5.1. Critérios de letalidade
Testes de toxicidade realizados com me-xilhões, geralmente causam uma grande varia-bilidade devido à capacidade de isolar-se do meio externo (Cataldo et al., 2002a; Rajago-pal et al., 2003), necessitando de uma atenção maior no momento da definição da condição de vida do mexilhão, após a sua exposição a um biocida. Torna-se necessário, neste perío-do, a observação das reações vitais do mexilhão o qual, muito vezes, permanece em estado de defesa, mantendo as valvas fechadas. Sendo as-sim, alguns estudos inserem um período (geral-mente 48h), após a exposição, para confirmar as reações vitais dos organismos sobreviventes. Dentro deste período consideram a morte do organismo, ao confirmar sua inércia, falta de reação a estímulos (como um toque com bastão de vidro) e, também, apresentar valvas abertas
e o manto exposto (Darrigran & Damborenea, 2001; Campos, 2009; Soares et al., 2009). Santos, C. P. (2011) avalia a mortalidade de larvas do mexilhão-dourado com base nos cri-térios de mobilidade, adução valvar e ausência de batimentos do estilete cristalino (Tab. II). Soares et al. (2009) definem alguns critérios, com base em Morton (1973), para uma avaliação quali--quantitativa do comportamento de L. fortunei exposto ao sulfato de cobre. Com esses critérios os autores buscaram não apenas avaliar a mor-talidade, mas compreender a ação da substância sobre o comportamento do molusco (Tab. II).
5.2. Confirmação da ação letal de microrganismos
Após a constatação da morte de um or-ganismo-alvo é necessário confirmar se este foi morto realmente pelo microrganismo inocula-do. Uma maneira clássica é o reisolamento do microrganismo após a inoculação do organis-mo-alvo em um meio de cultura apropriado para o crescimento do microrganismo testado

CAPÍTULO 26
Seção 8
287
5.3.1. Mortalidade absoluta (MA)
Número absoluto de indivíduos mortos no experimento para cada tratamento (Alves, 1998).
5.3.2. Mortalidade corrigida (MC)
Considera a mortalidade ocorrida no tratamento-testemunha (sem aplicação do agente estressor) no cálculo. A correção da mortalidade pode ser feita pelo uso da fórmula de Abbott, como segue abaixo, (Alves, 1998). Importante destacar que experimentos com mortalidade maior de 10% no tratamento controle devem ser excluídos e repetidos.
5.3.3. Relação dose-resposta ou concentração-resposta
Descreve a mudança em efeito sobre um organismo causado por diferentes níveis de ex-posição (ou concentrações) a um estressor (que pode ser químico, físico ou biológico), após certo tempo de exposição (Crump et al., 1976). O termo dose pode ser utilizado quando se sabe exatamente a quantidade de substância a que os organismos foram submetidos, quando não se conhece a quantidade de agente estres-sor que atingiu diretamente o organismo-alvo, o termo correto é concentração (Alves, 1998). Em experimentos com substâncias químicas, ondas ultrassônicas e radiações ultravioleta, é possível obter a dosagem. Já aqueles realizados com agentes microbianos, apenas a concentra-ção é obtida. O estudo de dose-resposta e desenvol-vimento de modelos de dose-resposta é pri-mordial para determinar as doses “seguras” e “letais” das substâncias as quais os organismos são submetidos. Graficamente, os dados de dose-respos-ta são demonstrados em uma curva de dose--resposta como um gráfico X-Y relacionando a magnitude do agente de controle (ex.: concen-tração de bactérias/ml, dose radiação ultravio-leta) à resposta dos organismos (no caso, me-xilhão-dourado). A resposta é avaliada como
Figura 6. Procedimento para a preparação de experimento para avaliar a atuação de radiação ultravioleta no controle da população larval de L. fortunei e C. fluminea.
(Fig. 9). Esta confirmação também pode ser feita por métodos moleculares e histológicos (Molloy & Mayer, 2007).
5.3. Interpretação de resultados
Em ensaios de avaliação de produtos e/ou microrganismos para o controle de pragas se utilizam alguns cálculos padrões e procedimen-tos estatísticos que permitem a inferência sobre a efetividade dos mesmos sobre a praga-alvo.

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
288
Figura 7. Procedimento para a preparação de experimento para avaliar a atuação de substâncias químicas no controle da po-pulação adulta de L. fortunei.
Figura 8. Mexilhões mortos após infecção por microrganismo (isolado Bb7B) e reisolamento em placa com meio de cultura. Fotos: A.S. de Oliveira e D. Pereira.

CAPÍTULO 26
Seção 8
289
Tabela II. Indicadores comportamentais e de letalidade para L. fortunei em experimentos: larvas de acordo com Santos, C. P. (2011) e adultos de acordo com Soares et al. (2009), com base em Morton (1973).
Indicadores Definição
Larvas
M Presença de mobilidade (M).
AV Adução valvar após estímulo mecânico.
AB Ausência dos batimentos do estilete cristalino após estímulo mecânico.
Adultos
M Número de indivíduos em deslocamento (M) pela superfície de vidro do aquário.
IM Número de indivíduos sésseis (IM) no aquário.
A Número de indivíduos com valvas abertas (A).
F Número de indivíduos com valvas fechadas (F).
DA A densidade de indivíduos aglomerados (DA) foi obtida pelo somatório de indivíduos aglomerados dividido pelo número de aglomerados.
PE Número de indivíduos com o pé exposto (PE).
PR Número de indivíduos com o retraído (PR).
SE Os sifões expostos (SE) indicam filtração ativa; os sifões são considerados expostos quando ultrapassam a borda posterior da concha.
SR Os sifões retraídos (SR) indicam filtração não ativa; os sifões são considerados não expostos quando não ultrapassam a borda posterior da concha.
RTP A reação ao toque positiva (RTP) é indicada pela movimentação das valvas ou pela retração do pé ou sifões, por meio do contato com a extremidade de um palito de madeira.
RTN A reação ao toque negativa (RTN) é indicada quando as valvas não se fecham ou o pé e sifões não se retraem por meio do contato com a extremidade de um palito de madeira.
V
Os indivíduos vivos (V) retraem o pé e sifões, fechando as valvas, quando tocados com a extremidade de um palito de madeira; quando as valvas estão fechadas, via dissecação se observa a movimentação das brânquias ou retração do pé, quando tocados com a extremidade de uma agulha histológica; os indiví-
duos vivos apresentam uma cor amarela ou marrom intensa.
MO
Os indivíduos mortos (MO) não retraem o pé e sifões, nem fecham as valvas, quando tocados com a extremidade de um palito de madeira; quando as valvas estão fechadas, via dissecação não se observa movimentação das brânquias ou retração do pé, quando tocados com a extremidade de uma agulha
histológica ou de um bisturi; a cor dos indivíduos mortos é esbranquiçada.
mortalidade, e pode ser expressa em números absolutos ou como porcentual de indivíduos mortos. A dose é geralmente plotada no eixo X, enquanto a mortalidade é plotada no eixo Y. O primeiro ponto no gráfico onde a resposta é acima de zero (ou acima da resposta do trata-mento controle), é denominado dose limiar. A mortalidade também varia em função do tempo. Sendo assim, estudos preliminares devem testar diferentes tempos de exposição dependen-do do objetivo do experimento e dos agentes de controle testados. No caso do controle microbia-no, a ação tóxica da bactéria evolui com o passar
do tempo, sendo necessário de 8 a 10 dias para se verificar uma ação efetiva (Fig. 11).
5.3.4. Concentração Letal Média (CL50) e Tempo Letal Médio (TL50)
Análise estatística de curvas de dose resposta pode ser realizada por métodos de regressão como Probit e Logit. Além desta aná-lise, são calculados as regressões lineares e os coeficientes de determinação, com base nos da-dos de mortalidade e concentrações do agente estressor. Regressão do tipo Probit é a mais uti-lizada para determinação da Concentração Le-

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
290
tal Média (CL50) e Tempo Letal Médio (TL50) a partir das curvas de dose resposta. Os cálculos de concentração letal (CL) determinam a quantidade necessária do agen-te estressor para que ocorra a mortalidade de 50% (CL50) ou 95% (CL95) da população en-saiada. Os cálculos de tempo letal (TL) deter-minam o tempo necessário de exposição do organismo-alvo ao agente estressor para que ocorra a mortalidade de 50% (TL50) ou 95% (TL95) da população ensaiada. Devem ser con-siderados os limites de confiança e a significân-cia do teste. A análise conjunta dos parâmetros des-critivos de mortalidade absoluta e corrigida, juntamente com os dados obtidos via regres-são de Probit, permite a visualização de quais tratamentos e concentrações são mais efetivos no controle de determinada praga-alvo (Tab. III).
5.3.5. Análise de dados
Soares et al. (2009) utilizaram diferentes abordagens na análise de dados de mortalidade e de critérios de letalidade do mexilhão-dou-rado exposto ao sulfato de cobre. Por meio da análise de variância (ANOVA, Dunnet a pos-teriori), com nível de significância de 95%, compararam tratamentos e controles quanto à mortalidade. Utilizaram análise multivariada para avaliar os dados obtidos pelas observações do comportamento (com base nos critérios dis-criminados na Tab. II) do mexilhão-dourado frente à exposição ao controlador químico em duas etapas: a primeira, análise de agrupamen-to (matriz de correlação de Pearson entre vari-áveis; agrupamento pelo método do centroide) (Fig. 12) e por meio da Análise de Componen-tes Principais (ACP) (Fig. 13), com a finalidade de ordenar e agrupar as variáveis que melhor explicam o comportamento; a segunda, por meio de análise de discriminante (AD) (Fig. 14), com a finalidade de discriminar os dife-rentes tratamentos quanto aos padrões de com-portamento. Os autores acima verificaram, por meio da ACP (Fig. 12), que os componentes 1 e 2 responderam por 55,96% da variância dos in-dicadores comportamentais e de letalidade. O componente 1 (35,05%) apresentou forte
Figura 9. Mortalidade de Limnoperna fortunei exposto a Ba-cillus thuringiensis sv. Israelensis IIRAC 22 em experimento de bancada: relação dose-resposta
Figura 10. Mortalidade de Limnoperna fortunei exposto a Ba-cillus thuringiensis sv. Israelensis IIRAC 22 em experimento de bancada: relação dose-resposta ao longo do tempo.
correlação positiva com o número de indiví-duos com sifões expostos – SE (0,935); com o número de indivíduos com valvas abertas – A (0,926); com a densidade de indivíduos aglo-merados – DA (0,604) e o número de indivídu-os com reação ao toque positiva – RTP (0,573). Também apresentou forte correlação negativa com o número de indivíduos com sifões não ex-postos – SNE (-0,935); o número de indivíduos com valvas fechadas – F(-0,926); com o núme-ro de indivíduos com reação ao toque negativa – RTN (-0,641) e com concentrações de sulfa-to de cobre utilizadas – C (-0,600). O compo-nente 2 (20,91%) apresentou forte correlação positiva com o número de indivíduos mortos – MO (0,949); com a temperatura do aquário –T (0,865) e com o tempo de exposição – TE (0,844). Este componente também apresentou forte correlação negativa com o número de in-

CAPÍTULO 26
Seção 8
291
divíduos vivos – V (-0,949). A ACP (Fig. 12) ordenou os indicadores comportamento e le-talidade de L. fortunei em concordância com os grupos (G1 a G5) formados por meio da análise de agrupamento hierárquico (Fig. 13) dos mesmos indicadores: G1 (F, SNE, RTN e
os mexilhões apresentaram pouca mobilidade durante os experimentos. O G5 refere-se os organismos vivos. A avaliação comportamen-tal do mexilhão durante os experimentos per-mitiu definir parâmetros de comportamento para o entendimento do efeito moluscida da
Tabela III. Mortalidade corrigida de L. fortunei e concentração letal (CL50
) de duas variedades de B. thurin-giensis. Experimentos de bancada realizados por Pereira et al. (2011b).
Tratamento Mortalidade corrigida (%) CL50
B. thuringiensis sv. israelensis IRAC22(107 cél./ml)
72 7,6 x 106 cél/mL
B. thuringiensis sv. kurtaski HD-1(107 cél./ml)
62,5 8,65 x 106 cél/mL
C); G2 (MO, T e TE); G3 (número de indi-víduos com o pé retraído – PNE, número de indivíduos com o pé exposto – PE, número de indivíduos sésseis – IMV e o número de in-divíduos em deslocamento – MV); G4 (RTP, DA, A e SE) e G5 (V). O G1 reuniu as variáveis que indicaram filtração inativa e altas concen-trações de sulfato de cobre. Ao contrário do G1, o G4 reuniu variáveis que indicaram fil-tração ativa e a formação de aglomerados. O G2 reuniu variáveis que indicaram a mortali-dade ao longo do tempo associada ao aumen-to da temperatura nos aquários. O G3 reuniu variáveis relacionadas à mobilidade, que não contribuíram para a descrição dos padrões de comportamento dos organismos testes, pois
substância testada. Os organismos quando ex-postos reagiram imediatamente fechando as valvas. Após alguns minutos abriam as valvas e iniciavam novamente a atividade de filtra-ção de forma pouco intensa, como pode ser observado por meio do nível de exposição dos sifões. No entanto, com o passar do tempo de exposição, o efeito das concentrações sobre os moluscos inibiu a atividade de filtração e a mobilidade dos mexilhões. A análise mul-tivariada demonstrou claramente a distinção entre grupos de filtração ativa e inativa. Após a exposição ao cobre os mexilhões diminuí-ram a atividade de filtração, a mobilidade e a reação ao toque, demonstrando um estado letárgico. Outros indivíduos mantiveram as
Figura 11. Análise multivariada dos indicadores comportamentais e de letalidade de Limnoperna fortunei: agrupamento hierárquico (método do centroide) com base em matriz de correlação (Pearson) entre os indica-dores comportamentais e de letali-dade (grupos 1 e 2: G1 e G2) (Fonte: Soares et al., 2009).

Pereira et al.
Planejamento experimental para a seleção de métodos de controle populacional de moluscos invasores
292
Figura 12. Análise multivariada dos indicadores comportamentais e de letalidade de Limnoperna fortunei. As variáveis agrupadas no dendo-grama ilustrado na figura anterior apresentam a mesma relação na or-denação obtida por análise de com-ponentes principais (ACP). (grupos 1 a 4 do dendograma, G1 a G4) dos indicadores de letalidade. (Fonte: So-ares et al., 2009).
Figura 13. Ordenação (análise de dis-criminante canônica) dos tratamen-tos e controles quanto aos indicado-res de comportamento e letalidade de Limnoperna fortunei. (Fonte: So-ares et al., 2009).
valvas fechadas. Esse comportamento indicou provavelmente um mecanismo de reação do bivalve diante da sinalização da presença de uma substância de elevada toxicidade. Por meio da AD (Fig. 14), as funções 1 e 2 responderam 94,8% da variância dos trata-mentos quanto aos indicadores comportamen-
tais e de letalidade. A função 1 (67,8%) apre-sentou forte correlação positiva com a variável discriminante RTN (0,464) e negativa com A (-0,600). A função 2 (27,0%) apresentou forte correlação positiva com a variável discrimi-nante RTP (0,488) e negativa com TE (-0,331). Com base nesta análise foi possível diferenciar

CAPÍTULO 26
Seção 8
293
as observações referentes ao controle (sem adi-ção de sulfato de cobre) do tratamento 4,872 mg L-1, e estes dos demais tratamentos (9,135 mg L-1 e 15,225 mg L-1). A avaliação propos-ta por meio de indicadores de comportamento e letalidade do mexilhão-dourado, durante os testes de toxicidade consistiu num comple-mento eficiente da avaliação da toxicidade do sulfato de cobre ao mexilhão-dourado e pode ser adaptada em experimentos com outras substâncias que causam efeitos diferenciados sobre a fisiologia e comportamento das es-pécies alvos. Também pode ser adaptada aos experimentos realizados com outras espécies de bivalves. Critérios específicos a espécies de gastrópodes devem ser desenvolvidos ou e/ou adaptados.
5.4. Avaliação ecotoxicológica
Duas abordagens ecotoxicológicas fi-nalizam a avaliação e seleção de métodos de controle populacional de moluscos invasores: uma, a avaliação da toxicidade do meio (água) no qual é aplicado o tratamento; outra, a ava-liação ecotoxicológica da substância moluscida ou do produto formulado.
5.4.1. Avaliação ecotoxicológica da água tratada
Visa verificar se a presença de subpro-dutos ou metabólitos no meio tratado confe-re toxicidade a organismos representantes da biota aquática. Santos, C. P. (2011) avaliou a toxicidade de água submetida à radiação ultra-violeta, para o controle de larvas do mexilhão--dourado, por meio de ensaios ecotoxicológicos crônicos com organismos-teste de três níveis tróficos (Pimephales promelas, Ceriodaphnia dubia e Pseudokirchneriella subcapitata).
5.4.2. Avaliação ecotoxicológica de substâncias moluscidas ou produto formulado
Visa verificar se a substância moluscida ou produtos formulados apresentam toxici-dade às espécies não-alvo. Várias espécies de organismos-teste não-alvo podem ser utiliza-das aqui, incluindo espécies exóticas e nativas. A maioria das espécies utilizadas como orga-nismos-teste no Brasil hoje, é exótica, mas os métodos para utilizá-las são padronizados pela ABNT, o que garante uma maior confiabilida-de e repetibilidade dos resultados. Seria interessante padronizar testes eco-toxicológicos com espécies de bivalves nativos, mas muitas dificuldades podem ser encontra-das neste sentido. Um fato a se considerar é que os bivalves nativos, em sua maioria, não são facilmente criados em laboratório e seus aspectos reprodutivos são muito pouco conhe-cidos. No caso do mexilhão-dourado, a escolha de um organismo-teste não alvo filogenetica-mente relacionado se torna impossível, uma vez que as demais espécies da família Mytilidae são todas marinhas. Quanto aos corbiculídeos, a padronização de testes com espécies do gênero Cyanocyclas pode ser uma alternativa, mas re-quer estudos prévios sobre a criação e reprodu-ção desta espécie em laboratório. A busca de in-dicadores representantes das famílias Hyriidae e Mycetopodiade também se torna uma tarefa difícil, pois muitas das espécies utilizam pei-xes em seu desenvolvimento larval parasitário. Além disso, muitas espécies destas duas últimas famílias apresentam populações reduzidas ou enquadradas em alguma categoria de ameaça de extinção. Uma alternativa seria padronizar tes-tes com espécies da família Sphaeriidae que são mais abundantes, não apresentam ciclo parasitá-rio e apresentam ampla distribuição no Brasil.


Seção
9Controle populacionalde bivalves invasores:
métodos químicos


297
As pesquisas e o uso prático de agentes químicos constituem-se nas primeiras tentati-vas para o controle populacional de bivalves in-vasores. As razões para tanto estão associadas à facilidade de sua aplicação e a experiências já conhecidas da ação desses agentes químicos como agentes desinfetantes e bactericidas no tratamento de águas para os mais diversos usos pelo homem. Desta forma, é natural que encontremos na literatura avaliações para o controle de bi-valves com substâncias químicas já amplamente utilizadas para o controle de outras espécies.Via de regra, a aplicação de agentes químicos apresenta aspectos operacionais extremamente simples, pois o agente está disponível no merca-do a custos relativamente baixos, a distribuição do produto no corpo d’água muitas vezes pode ser direta ou, quando muito, exigir a simples dissolução e diluição antes dessa distribuição e, às vezes, não requer equipamentos especiais para a distribuição no corpo d’água ou, quando muito, simples dosadores.
Entretanto, o uso desses agentes quími-cos vem encontrando restrições na medida em que pesquisas são desenvolvidas sobre os efei-tos negativos sobre outras espécies e sobre o meio ambiente como um todo. Estas pesquisas, sobre efeitos indesejá-veis dos agentes químicos em águas superfi-ciais, têm promovido alterações nas legislações ambientais, limitando o lançamento de pro-dutos químicos em águas superficiais. Assim, as doses dos agentes químicos permitidas pela legislação, podem estar aquém das doses neces-sárias para o controle eficaz da população de bivalves. Cabe salientar que os agentes químicos usualmente utilizados no controle populacio-nal de bivalves são classificados como substân-cias tóxicas. Em termos legais, devemos sempre con-sultar os órgãos de regulação e fiscalização do meio ambiente e solicitar a devida autorização ou licença ambiental para a aplicação de agen-tes químicos nos corpos d’água, sob pena de
CAPÍTULO 27
Controle Químico: conceitos básicos
Maria Teresa Raya Rodriguez

Rodriguez M. T. R.
Controle Químico: conceitos básicos
298
estarmos cometendo um ato ilícito enquadrado como crime ambiental. Várias famílias de agentes químicos têm sido empregadas para o controle populacional pretendido. Entre eles podemos citar substân-cias oxidantes e sais, tais como, cloro e seus de-rivados (dióxido de cloro, cloroaminas e ácido hipoclorídrico), compostos de bromo, ozônio, peróxido de hidrogênio, permanganato de po-tássio, nitrato de amônia e sulfato de cobre. As tintas anti-incrustantes têm sido tra-dicionalmente aplicadas para a proteção de cascos de embarcações marítimas com a fina-lidade de inibir a fixação e o desenvolvimento de organismos, pela liberação de um biocida misturado à tinta. Estes biocidas como o TBTO (óxido de estanho tributil, um óxido de organoestânicos), têm sido utilizados com sucesso para a preven-ção da incrustação de bivalves. Entretanto, es-tes compostos químicos são substâncias tóxicas que levaram alguns países a proibir seu uso. Atualmente, as pesquisas vêm sendo de-senvolvidas com o intuito de produzir tintas anti-incrustantes à base de silicone, substância não tóxica, que impede ou diminui considera-velmente a força de fixação para os bivalves. Tintas a base de zinco também parecem muito promissoras. Assim, as pesquisas com tintas anti-in-crustantes devem considerar, além da eficácia como proteção à incrustação em superfícies, a minimização dos impactos ambientais para os ambientes aquáticos. Tão importante quanto à escolha do agente químico a ser utilizado, é a forma como o mesmo deve ser distribuído no corpo d’água. Esta distribuição está associada à ação preten-dida. Basicamente temos duas situações:
a) Ação Corretiva- são usadas depois dos bivalves invasores terem se estabelecido no cor-po d’água.
b) Ação Preventiva- são utilizadas para que os bivalves invasores não se estabeleçam no corpo d’água que se pretende proteger.
Exemplo de ação corretiva é a aplicação do agente químico nos momentos em que os bivalves adultos estão mais vulneráveis, situa-ção que ocorre logo após o período de desova; os mais jovens são mais susceptíveis à ação do agente químico. Em se tratando de ação preventiva, te-mos três estratégias a serem consideradas na aplicação do agente químico, a saber:
* Ação intermitente – aplicado a interva-los de tempo (por exemplo, a cada 6 horas) com o intuito de destruir os bivalves nas primeiras fases de vida, situação em que se encontram mais vulneráveis.
* Ação semicontínua – aplicado a inter-valos regulares de tempo, por exemplo, de 15 minutos, com o intuito de prejudicar que os bi-valves realizem a filtragem fechando a concha. Ao reabri-la, após 15 a 30 minutos, ficam sub-metidos à nova carga do agente químico.
* Ação contínua – aplicada ininterrupta-mente com o propósito de proteger sistemas de alta segurança, onde a ocorrência de qualquer tipo de incrustação não pode ser tolerada. Visa causar a mortalidade de espécies adultas, prejudicar a reprodução e destruir as espécies jovens.
Devemos em nossas avaliações, com o uso de agentes químicos, para o controle po-pulacional de bivalves, levar em consideração a relação causa (ação do agente químico) e efeito (sobre os organismos). A causa sempre será função da dose a que o organismo ficou submetido. A dose, por sua vez, é função da concentração do agente químico e do tempo de exposição do organismo. Para sistemas fechados (estáticos) o cálculo da dose é relativamente simples porque a concen-tração do agente químico e o tempo de exposição do organismo são dados fáceis de serem obtidos. Para sistemas abertos (com fluxo), entretanto, o cálculo da dose tornar-se mais complexo, pois exige o conhecimento da dinâmica do processo.

299
Introdução
O desenvolvimento de técnicas inovado-ras, produtos, métodos e substâncias alterna-tivas, para o controle de organismos invasores (bioincrustações) no mundo estão sendo impul-sionando pela busca por maior eficiência com baixo custo, associado a um baixo impacto am-biental e operacional. Neste contexto, biocidas microencapsulados foram propostos em estu-dos realizados pela universidade de Cambrid-ge-UK e atualmente são comercializados. Microcápsulas são utilizadas em diferentes atividades tecnológicas (tintas, têxteis, eletrônicas etc.) sendo mais empregadas na indústria alimen-tícia e farmacológica. As razões para o encapsu-lamento são incontáveis, e para o uso no controle de mexilhões, ele serve para direcionar a entre-ga de substâncias, isolar e controlar a dissolução de biocidas evitando a degradação do princípio ativo. As microcápsulas propostas para o contro-le de mexilhões são produzidas como pequenas partículas (3-800 μm) que contêm compostos ativos (20-30%) revestidos por uma cobertura
ou proteção, geralmente um invólucro nutritivo ou atrativo (alginatos, proteínas, lipídeos etc.) (Fig. 1). Os microencapsulados testados contra o mexilhão-dourado no Brasil foram partículas microscópicas com tamanhos variados (entre 10-500 μm) feitas de uma mistura de goma, óleo e cera que recobrem biocidas do tipo cloreto de potássio e ou compostos de aminas (Fig. 2) (Campos, 2009) tendo sido produzidos de forma experimental e atualmente, a sua produção está estabilizada e sendo oferecida de forma comercial (BioBullet.com) (Quadro 1). O revestimento externo de ácido graxo e o núcleo tóxico para mexilhões são inofensivos para os seres humanos. A toxicidade para outros animais aquáticos dependerá das concentrações ministradas, as quais geralmente estão bem acima das utilizadas contra os mexilhões. As substâncias que compõem os microencapsulados utilizados são aprovadas para a utilização em tratamento de água potável na Europa (cloreto de potássio e compostos de amônia quartenária). Os métodos químicos comuns (substân-
CAPÍTULO 28
Microencapsulados
Sávio Henrique Calazans CamposFlavio da Costa Fernandes

Calazans C. & Fernandes
Microencapsulados
300
cias dissolvidas na água) provocam a reação de defesa natural dos mexilhões, os quais fecham suas conchas por até 3 semanas resistindo às dosagens do biocida (Cataldo et al., 2002, Mansur et al., 2003). Esta capacidade de prote-ção dos mexilhões sobre os biocidas dissolvidos amplificam o risco de dano ambiental, princi-palmente sobre os organismos não-alvo, além do alto custo com dosagens contínuas para a remoção dos organismos. Desta forma, os mi-croencapsulados tornam-se uma alternativa às substâncias dissolvidas, as quais são ampla-mente utilizadas com um alto custo ambiental e baixa eficiência contra mexilhões (bivalves filtradores). Ao considerarmos que os mexilhões são filtradores típicos (retiram seu alimento do material suspenso na água), o uso de biocidas microencapsulados surge como uma solução sedutora, na qual possibilita diminuir o volu-me do biocida utilizado. Desta forma, ao filtrar as partículas suspensas na água, os mexilhões concentram no estômago a dose específica do biocida que seria dispersado (Aldridge et al., 2006). Esta estratégia de dosagem aumenta a aceitabilidade do mexilhão diminuindo a sua reação de defesa (fechamento das conchas) intensificando o efeito do biocida ministrado (Costa, 2008). Inicialmente, a invenção dos microen-capsulados manteve-se focada no controle do mexilhão-zebra (Dreissena polymorpha), o qual rapidamente se tornou uma das maiores pestes ecológicas e ambientais (Costa et al., 2008). Atualmente a proposta dos desenvolvedores do produto é a de integrar uma estratégia de tratamento com um plano de controle de do-
Quadro 1. Usos propostos pelo fabricante para o uso dos microencapsulados disponibilizados:
Atividade Aplicações
Controle para usinas hidrelétricas Manutenção de limpeza em sistemas de refrigeração e fluxo de água;
Tratamento de águas industriais Tratamento ao longo de tubulações, sistemas de transporte e armazenamento de águas;
Controle para fábricas de papel Manutenção em sistemas de águas evitando entupimentos;
Transporte Marítimo e Petroleiro Remoção e ou controle de bioincrustação e água de lastro;
Agropecuária Limpeza de canais e dispersores e Irrigação;
Fazendas de cultivo de camarões Remoção de animais filtradores;
Incremento de Cultivos Enriquecimento da alimentação com dosagens de vitaminas;
Fonte: www.biobullet.com
Figura 1. A ideia de encapsulamento de biocida para o con-trole de mexilhões. A - representação de biocida encapsula-do; B - fotografia de endoscópio, biocida microencapsulado sendo transportado pelas brânquias de um mexilhão-zebra vivo (créditos: Dr. Paul Elliot, Universidade de Cambridge-UK). Microcápsulas similares foram utilizadas em testes com o Mexilhão-Dourado.
Figura 2. Foto de microcápsulas de KCl utilizadas em expe-rimentos com Limnoperna fortunei (adaptado de Campos, 2009).

CAPÍTULO 28
Seção 9
301
sagens e de monitoramento para o controle da infestação, no qual o uso de microcápsulas com biocidas tem se mostrado ser uma alternativa para controlar o mexilhão-zebra, com eficiên-cia e menor dano ambiental em instalações de tratamento de água (Aldridge et al., 2006). Estudos realizados com o mexilhão-ze-bra na Europa verificaram que a concentração de biocida necessária para matar 50% dos or-ganismos em testes decresceu de 210 para 107 mg/L ao encapsular o biocida (Costa, 2008) onde 1g/L do microencapsulado correspondia 300 mg/L de KCl. Testes com uma formula-ção de amônia quaternária DB-45 encapsula-da (poliDADMAC), em bioensaios semiestáti-cos, ofereceu clara evidência do seu potencial. Onde, o composto foi mais efetivo contra o mexilhão-zebra do que sua condição original não encapsulada (Costa, 2008). Nestes bioen-saios, o produto particulado reduziu em três vezes a quantidade de princípio ativo requeri-do para causar 90% de mortalidade em 12h de tratamento.
Resultados com Mexilhão-Dourado
Em estudo com o mexilhão-dourado no Brasil, Calazans C. et al. (prelo 2012) re-alizaram testes de toxicidade para a determi-nação da concentração letal dos organismos expostos (CL50%) em sistemas semiestáticos (com troca de água) para avaliar o potencial das microcápsulas (Campos, 2009). Neste es-tudo, verificou-se os efeitos da mortalidade do L. fortunei exposto à cloreto de potássio
e compostos quaternários de amônia dissol-vidos (poliDADMAC) e microencapsulados. Dentre as substâncias estudadas, o KCl foi o menos tóxico, em ambas as formas, dissolvi-da (CL50=3.445 mg.L-1) e microencapsuladas (CL50=2.737 mg.L-1), sendo que a forma mi-croencapsulada foi mais eficiente, com uma CL50 25% menor e uma menor variabilidade da resposta dos mexilhões. Ao termos em con-ta que a quantidade de substância ativa (KCl) do produto é de 30% do peso total do micro-encapsulado, percebemos o efeito potenciali-zador do encapsulamento. Desta forma, o KCl microencapsulado apresentou uma concentra-ção letal (CL50) de 821 mg.L-1 o que represen-tou ser 4 vezes maior que a toxicidade com-parada com o KCl dissolvido. Os compostos quaternários de amônia DB-45 exibiram uma CL50 em 48h de 315 mg.L-1. No entanto, ao considerarmos que somente 20% é o biocida, a toxicidade real estimada passa para (CL50) 63 mg.L-1. Estes valores indicam toxicidade 8.6 vezes maior do que o KCl microencapsulado, mesmo apresentando uma variabilidade maior das mortalidades entre as concentrações de 25 a 200 mg.L-1. Ao considerarmos o intuito de remover os organismos, os resultados apresen-taram a menor concentração de efeito obser-vado (CEO=100% mortalidade) em torno de 500 mg.L-1 do produto (Fig. 3 e Tab. I).
Figura 3. A- Cloreto de potássio dissolvido; B- KCl microencapsulado; C-polyDADMAC microencapsulado (Reproduzido de Ca-lazans C. et al., prelo 2012).

Calazans C. & Fernandes
Microencapsulados
302
Os microencapsulados também foram avaliados em um sistema de fluxo contínuo (Campos, 2009). Considerando-os como testes piloto, tais resultados assinalam sobre o com-portamento destes compostos em um sistema dinâmico. Os experimentos com KCl apresen-taram uma relação dose-dependente atingindo 15% de mortalidade em 6h. Enquanto que o composto de amina causaram pelo menos 40% de mortalidade em três experimentos também de 6h com concentração de 90 mg.L-1. O uso deste produto para tratamento em sistemas condutores de água necessita de uma dinâmica de dosagem contínua o que deve ser associada ao uso de um dispositivo misturador/dosador (Calazans C. et al., prelo 2012).
Conclusão Testes realizados com o mexilhão-zebra e também com o mexilhão-dourado demons-tram que o uso de microencapsulado provou ser eficaz, alcançando 100% de mortalidade em laboratório com uma baixa variabilidade de resposta em relação às substâncias quími-cas dissolvidas (Calazans C. et al., prelo 2012). Tais resultados estimulam a expectativa em um produto altamente eficiente. Os benefícios pro-porcionados por biocidas encapsulados podem oferecer vantagens econômicas e operacionais, bem como benefícios ambientais em função de considerar os hábitos dos mexilhões. Esta técnica oferece como vantagem; A proteção do principio ativo até o seu momento de reação; A possibilidade da dosagem específica necessária para o controle do mexilhão acompanhada de uma diminuição das emissões no efluente; A
possibilidade selecionar os diferentes filtrado-res (organismos alvo e não alvo) através da di-ferenciação do tamanho das partículas as quais eles se alimentam. Em contrapartida, esta téc-nica oferece ação somente sobre os organismos adultos os quais conseguem ingerir as cápsulas; Em tratamentos de águas de resfriamento é ne-cessário muito material, o que o torna pouco competitivo em relação aos biocidas estabeleci-dos no mercado (ex. cloro) (Polman & Janssen--Mommen, 2011). Desta forma, ainda existe a necessidade de melhorar a tecnologia para am-pliar a desempenho da formulação do molusci-cida em condições reais de densidade de mexi-lhões, qualidade de água e hidrodinamismos, além de tornar esta técnica ainda mais accessí-vel comercialmente. Em um estudo solicitado pelo governo espanhol, o método de controle dos microencapsulados não foi classificado como a melhor técnica disponível justamente em função das necessidades de aprimoramento das capacidades de retenção das micropartí-culas. Por outro lado, a Agência Ambiental do Reino Unido já deliberou a descarga (de até 10 mg/l), dos compostos de amônia quaternária (princípio ativo com maior eficiência das mi-crocápsulas) na água potável, como também em efluentes, neste último, estuda-se a associação de dosagens de bentonita para torná-los inofen-sivos (Polman & Janssen-Mommen, 2011). No Brasil ainda não há nenhuma deliberação para uso deste produto em larga escala. Certamente a continuidade da busca, no uso e no desenvol-vimento de produtos inovadores irá promover a conquista de produtos altamente efetivos para o controle de bioincrustação em sistemas de águas.
Tabela I. Valores de concentrações letais CL50-48h, das concentrações de efeito não-observado (CENO) e me-nor concentração com efeito observável (CEO), previstos com a regressão Probit, com intervalos de confiança de 95%, para cada produto químico testado contra Limnoperna fortunei em teste de toxicidade semiestático. (Reproduzido de Calazans C. et al., prelo 2012).
ProdutoCL50-48h (mg.L-1) -95.0%CL +95.0%CL N¥ [n] CENO CEO
poliDADMAC-m‡ 315,3 287,1 343,6 8 [13] 25 500
KCl-m‡ 2.737,0 2.650,2 2.823,9 6 [10] 250 6000
KCl dissolvido 3.444,9 3.367,2 3.522,5 2 [9] 10 2000
KCl-m-ft* - - - 3 [4] 90 1000
poliDADMAC-m-ft* - - - 3 [1] 90 90m = microencapsulado; CL = Concentração Letal, CENO = Concentração de efeito não observado; CEO = Con-centração de efeito Observado.* No tratamento em tubos com fluxo contínuo o número de testes e a mortalidade não foram suficientes para prever a LC-50.¥ ‘N’ significa a partir do número de experimentos independentes e ‹n› para o número de concentrações testadas.‡ O LC50 estimado é baseado no peso total da microcápsula, que são 30% e 20% de produto ativo para PolyDADMAC e KCl, respectivamente.

303
Há diversas substâncias químicas, orgâ-nicas ou inorgânicas, que possuem o átomo de cloro em sua composição. Isto confere a essas substâncias propriedades oxidantes, permitin-do seu uso como desinfetante em geral, para tratamento de água e como alvejante. Exemplos de compostos clorados para desinfecção são: cloro molecular (Cl2), hipoclorito de cálcio, hipoclorito de sódio (NaClO), dicloroisocia-nurato de sódio (dicloro), tricloroisocianurato de sódio (tricloro) e dióxido de cloro (ClO2), entre outros. Alguns desses produtos foram estudados para uso como biocida, neste caso, moluscocida. A seguir podemos acompanhar um pouco dessa história. O NaClO (água sanitária), o dicloro e tricloro liberam ácido hipocloroso (HClO) quando em contato com a água. Neste caso, o dicloro e o tricloro, além do HClO, formam isocianurato de sódio (Fig. 1). O HClO é o princípio ativo. Ele que é capaz de interagir quimicamente com a matéria orgânica existen-te na água, oxidando e consumindo a matéria
orgânica e matando os organismos. Sua eficiên-cia como biocida é dependente do pH da água a ser tratada. Quanto a isso, o dicloro e o triclo-ro são capazes de manter sua eficiência numa faixa de pH um pouco maior do que o NaClO (Fernandes, 2009). Em meio aquoso, o NaClO se dissocia gerando um equilíbrio de um ácido fraco, como pode se observar nas equações apresentadas na Figura 1. O dicloro e o tricloro funcionam como armazenadores de HClO, liberando-o lenta-mente. Cada molécula de dicloro libera duas moléculas de HClO. O tricloro, três. Os cloros orgânicos são muito utilizados para tratamento de piscina, pois mantêm o nível de cloro baixo e constante, até o próximo tratamento, geral-mente, 24h depois. Caso seja usado o NaClO na piscina, ou em qualquer outro meio aquoso, o HClO será liberado totalmente, atingindo al-tas concentrações inicialmente e, com o passar do tempo, decaindo até zero. O HClO é volátil e facilmente perdido para a atmosfera. Outra
CAPÍTULO 29
Formas de Cloro
Livia Viana de Godoy FernandesSávio Henrique Calazans Campos
Flavio da Costa Fernandes

Fernandes et al.
Formas de Cloro
304
característica vantajosa do dicloro, é que sua solução a 1% possui um pH de 6,0 – 8,0, en-quanto o pH do NaClO varia de 11,0 – 12,0 (cáustico) e o tricloro apresenta, 2,7-2,9 (ácido) (Macêdo, 2004). O dicloro é menos corrosivo ao meio (tubulações e equipamentos). A grande vantagem do uso do NaClO está no custo, o que faz seu uso mais frequente na prática. O ClO2 é um gás esverdeado. O princípio ativo, neste caso, é a própria molécula de ClO2, pelo seu efeito oxidante. Sua eficiência como biocida é pouco afetada pelo pH do meio. Por se apresentar no estado gasoso à temperatura ambiente, deve ser produzido no local do seu uso. Há no mercado a opção do ClO2 estabi-lizado em forma líquida, que com a adição do ativador libera a molécula de ClO2 no local e na hora do uso (Macêdo, 2004, Almeida, 2007). Uma substância química para ser usada como biocida deve apresentar o mínimo de toxicidade ao ecossistema, assim como os sub-produtos oriundos da sua utilização. Reações de cloração da água envolvendo o HClO (uso do NaClO, dicloro e tricloro) geram subprodu-tos tóxicos aos humanos e ao meio ambiente, representados, principalmente, pelos trialome-tanos (THM’s):
clorato, podem ser nocivos para a saúde huma-na (WHO, 2008). A Legislação Brasileira possui a Resolu-ção nº 357, de março de 2005 do CONAMA para definição da concentração limite de certas substâncias em efluentes. Esta Legislação re-gulamenta o limite de 1 mg/L (ou 1000 μg/L) de clorofórmio, tetracloreto de carbono, triclo-roetano e diclorometano para efluentes (todos esses também subprodutos tóxicos da cloração da água). Porém, não contempla concentração limite de cloro livre, assim como para clorito, clorato e ClO2. Um conceito importante de se ter em mente é o de cloro livre. As concentrações dos produtos clorados, geralmente, são apresenta-das em função do cloro livre e não do produ-to em si. A quantificação de cloro livre estará em função da molécula do cloro (Cl2), ou seja, relaciona quanto de HClO está disponível no meio. Isto significa que o HClO pode ser libe-rado por diferentes equilíbrios químicos, seja a partir do ClO- ou das moléculas de cloros orgânicos, como dicloro e tricloro, que servem como armazenadores. Os métodos de determi-nação de cloro livre fornecem a concentração do produto em função de cloro livre e não do produto em si. O método mais importante para quantificação de cloro livre é o método colori-métrico DPD (Clesceri et al., 1995). Vamos ver agora alguns trabalhos sobre o uso dessas substâncias cloradas como molus-cocidas. Campos (2009) determinou a CL50 (concentração letal para 50% dos organismos testados) do NaClO, do dicloro e do ClO2 (forma líquida estabilizada da empresa Bera-ca, Dióx50®) para o L. fortunei (Tab. I). Nesta Tabela, também estão expostos resultados não publicados anteriormente, de experimentos
Figura 1. Equilíbrio químico formado com a reação do dicloroisocianurato de sódio com água.
O dicloro e o tricloro são pouco tóxicos, assim como seus subprodutos, os isocianuratos (Clasen & Edmondson, 2006; Hammond et al., 1986). O ClO2, assim como seus subprodu-tos gerados com a cloração da água, clorito e

CAPÍTULO 29
Seção 9
305
cada molécula de dicloro libera duas de HClO, nesta concentração de 0,0004 mol/L de diclo-ro, 0,0008 mol/L de HClO seriam geradas no meio. A CL 50 do NaClO encontrada foi 0,53 mg/L. O que corresponde a 0,000007 mol/L de NaClO, que equivale à mesma quantidade de HClO. Ou seja, quando usamos NaClO como fonte de HClO, precisamos de uma quantida-de cerca de 100 vezes menor de HClO como biocida do que quando este HClO está acopla-do ao dicloro. Pode-se supor que a caracterís-tica do dicloro de liberar o HClO aos poucos, como um armazenador, não é tão eficiente e interessante como moluscocida, uma vez que o mexilhão ganha tempo e se fecha. Neste caso, o NaClO age de forma mais agressiva por “ata-car” com todas suas moléculas de HClO livres no meio. O mexilhão sofre dano mesmo antes de se fechar. Resultados encontrados por outros auto-res também indicam que o mexilhão é sensível a baixas concentrações de hipoclorito. Cataldo et al. (2002a) encontraram uma CL50 = 5,5 mg/L e Morton (1977) controlavam a rein-festação do mexilhão-dourado em instalações de hidroelétricas com 0,5 mg/L de cloro ati-vo. Porém, inicialmente, altas concentrações de cloro (200 mg/L) foram utilizadas durante vários dias, até semanas e depois doses redu-zidas (1,0 mg/L) foram usadas para remoção da população do molusco. Portanto, como se observa também em outros trabalhos (Doher-ty et al., 1986; Belanger et al., 1985; O’Neill, 1997 – citado em Cataldo, et al., 2002a), os
Tabela I. CL 50 de diferentes biocidas clorados.
Biocida Dicloro NaClO ClO2 Tricloro
CL50 (mg/L) 108,96 0,53 13,99 105,69
com o tricloro. O experimento com tricloro foi feito apenas uma vez e por isso deve ser ob-servado com mais cautela do que os outros. A metodologia utilizada nos testes de ecotoxici-dade pode ser encontrada em Campos (2009). Estes testes são semiestáticos, com período de exposição dos mexilhões aos biocidas de 48h, com 3 réplicas e 20 organismos em cada uma. Os cálculos de CL50 foram realizados com o auxílio de softwares específicos, PROBITOS 1.5 e Trimmed Spearman-Karber (TSK) 1.5. As CL50 estão em função da concentração do produto (não de cloro livre). Na Figura 2, temos um exemplo de como ocorre a mortalidade do mexilhão-dourado frente ao biocida. Neste caso, o biocida usado foi o dicloro, porém comportamento similar foi observado com os demais biocidas clora-dos. Nota-se que há uma grande variabilidade da mortalidade que pode ser visualizada pelos grandes desvios-padrão. Em determinada fai-xa de concentração, essa variabilidade é ainda maior. No caso do dicloro, essa faixa crítica de concentração foi cerca de 10 a 150 mg/L. Abai-xo dessa faixa a mortalidade não chega a 10% e, acima, mantém-se nos 100%. Essa variabi-lidade encontrada, principalmente nessa fai-xa crítica, é devida à capacidade do mexilhão de se fechar em ambiente adverso. Ao perce-ber que há uma ameaça química dissolvida na água, o mexilhão fecha a concha e para de fil-trar. Desta forma, o mexilhão se isola por um tempo do ambiente externo. Após determinado tempo, o mexilhão se abre e volta a filtrar. Se o ambiente estiver adequado, ele se restabelece. Se não, ele morre pela ação do biocida ou por inanição, por parar de filtrar. Justamente por essa capacidade de se fechar e se isolar do am-biente externo é que ocorre tanta variabilidade de mortalidade. Ao transformar as CL50 que estão em mg/L em mol/L, podemos comparar os bio-cidas em função do número de moléculas de HClO. Por exemplo, a CL 50 do dicloro é 109 mg/L, isto equivale a 0,0004 mol/L. Como
Figura 2. Mortalidade do L. fortunei em diferentes concentra-ções de dicloro (Campos, 2009).

Fernandes et al.
Formas de Cloro
306
tratamentos de curto período de tempo de con-tato com cloro não afetam os moluscos, pro-vavelmente pela capacidade dos mexilhões em detectar o elemento tóxico na água e fechar fortemente suas valvas. Rajagopal et al. (2003) concluíram que com uma dose constante de 1 mg/L de hipoclorito por 588 a 1104 h é possí-vel obter 100% de mortalidade dos adultos de Dreissena polimorpha, Mytilopsis leucophaeta e Mytilus edulis. Cataldo et al. (2002a) também encontraram muita resistência para combater o mexilhão-dourado em aplicações industriais, com uma concentração de 5 mg/L a 15ºC, sen-do aplicada por mais de um mês para matar a metade dos animais expostos, obtendo con-tudo, uma mortalidade maior com o aumento da temperatura, demonstrando que a tempera-tura influencia no efeito letal do hipoclorito. Os tratamentos de longa duração ressaltam a possibilidade de que os mexilhões estão sendo controlados por inanição causada pelo fecha-mento das conchas e não por ação direta do biocida, que em baixas concentrações já teriam eficiência para a prevenção do assentamento de larvas. O caminho trilhado até aqui mostra a di-ferença do efeito dos biocidas clorados frente à mortalidade do mexilhão-dourado para cálcu-lo de CL50. O passo seguinte seria a busca por concentrações ideais dos biocidas para serem usados nas tubulações para se minimizar as bioincrustações. Na prática, concentrações bai-xas e frequentes, contínuas ou não, devem ser avaliadas. Mäder Netto (2011) fez experimen-tos com o dicloro em tubulações de hidrelétri-cas e mediu seu efeito sobre a bioincrustação
do mexilhão-dourado (além da corrosão da tu-bulação) ao longo de 12 meses. A concentração usada no experimento foi de 1 ppm (dentro da tubulação) durante 8 h/dia. No décimo mês, a densidade da bioincrustação foi suficiente para prejudicar o perfeito funcionamento de filtros e trocadores de calor de usinas hidrelétricas. Ainda assim este resultado foi bom comparado ao experimento controle: após 12 meses, a bio-incrustação no experimento controle era mais de cinco vezes maior do que no experimento com o uso do dicloro. É evidente que os assuntos envolvendo a cloração da água são extensos e complexos. Estudos nessa área ainda são deficientes e devem ser desenvolvidos, pois são muitas as questões não respondidas. No presente capítulo, abordou-se de forma geral e simples este assunto. Para estudos mais aprofundados, recomenda-se a leitura das referências citadas seria um bom começo. Resumidamente, a escolha da substância usada para minimizar a bioincrustação deve levar em conta as seguintes questões: * a especificidade do efeito biocida sobre o organismo alvo e sua baixa toxicidade ao am-biente; * o tempo de contato do biocida com a massa de água. Por exemplo, o NaClO é melhor para tempos curtos de reação, como é o caso do controle de bioincrustação em tubulações, e o dicloro é melhor para tempos longos de reação, como no tratamento de águas represadas; * o pH da água a ser tratada; * o efeito oxidante do biocida frente ao tipo de material que constitui a tubulação ou a superfície que vai ser tratada.

307
O Sulfato de Cobre (II) ou Sulfato Cúpri-co (CuSO4) é um sal obtido pela reação entre minerais oxidados de cobre e ácido sulfúrico. Este sal existe sob algumas formas, que se dife-rem por seu grau de hidratação. A forma pen-tahidratada (CuSO4.5H2O) apresenta-se na forma de um sal azul brilhante (Fig. 1). O sulfato de cobre é um produto comu-mente utilizado como fungicida, bactericida, inseticida, moluscida e herbicida em atividades agrícolas, mas seus efeitos sobre a biota aquá-tica podem causar drásticas reduções nas po-pulações de macroinvertebrados (EPA, 1986; Doudoroff & Katz, 1953). A presença de cobre em concentrações superiores a 5 mg.l-1 torna a água para consumo humano impalatável (Branco, 1986). O cobre disponível do meio aquático é facilmente incorporado pelos moluscos herbí-voros, detritívoros, filtradores (Bendati, 2000; Pip, 1990; Villar et al., 1999), mas quando al-tas concentrações estão disponíveis, o metal pode causar a morte destes organismos. Na-
turalmente, o mexilhão-dourado, assim como outros bivalves límnicos, acumula o metal co-bre nos seus tecidos (Tab. I). Segundo Sylves-ter et al. (2005), L. fortunei tem capacidade de filtração muito elevada, quando comparado aos demais bivalves límnicos, provável razão da elevada concentração de cobre nos tecidos desta invasora. Os bivalves límnicos, inclusive L. fortunei, são utilizados como monitores am-bientais. Mas quando metais estão presentes em elevadas concentrações no meio aquático, conferem efeito tóxico sobre estes organismos, podendo causar a morte. O sulfato de cobre foi amplamente utili-zado como moluscida no controle de planorbí-deos (Reddy et al., 2004), ampularídeos (Wada, 2004) e outros gastrópodes terrestres (Amusan et al., 2002) e, também, para o mexilhão-zebra (Dreissena polymorpha) (Waller et al., 1993). Para o controle de mexilhão-dourado no Rio Grande do Sul (Brasil), especialmente na bacia do Lago Guaíba, o sulfato de cobre foi utilizado por captadoras de água (Colares
CAPÍTULO 30
Sulfato de Cobre
Daniel PereiraMíriam de Freitas Soares

Pereira & Soares
Sulfato de Cobre
308
et al., 2002a). No entanto, nenhum estudo foi realizado no sentido de dimensionar o impacto deste tratamento ao ecossistema, considerando a formação de subprodutos. Soares et al. (2009) verificaram que a Concentração Efetiva para causar a mortalida-de de 50% (CE50) e 95% (CE95) da população do mexilhão-dourado, em experimentos de laboratório (limite de confiança 95%) realiza-do com a adição de sulfato de cobre (mg L-1) em água bruta foi de 8,48 (6,69, limite infe-rior; 10,42, superior) e 19,08 (15,58 e 25,43) respectivamente. Os autores verificaram que 90,8% da mortalidade da população podem ser explicada pelas concentrações de sulfato de cobre r2 = 0,908) (Fig. 2). Também veri-ficaram que os organismos, quando expostos ao sulfato de cobre, reagiram imediatamente fechando as valvas. Após alguns minutos abri-ram as mesmas e iniciaram novamente a ati-vidade de filtração de forma pouco intensa, como pode ser observado por meio do nível de exposição dos sifões. No entanto, com o pas-sar do tempo de exposição, o efeito das con-centrações sobre os moluscos inibiram a ativi-dade de filtração, mobilidade dos mexilhões e a reação ao toque, demonstrando um estado de letargia. Em concentrações mais elevadas os indivíduos mantiveram as valvas fechadas. Esse comportamento indicou provavelmente um mecanismo de reação do bivalve diante da sinalização da presença de uma substância de elevada toxicidade. A Tabela II apresenta a comparação en-tre os valores das concentrações letais (CL50) destinadas ao controle de L. fortunei e outros moluscos límnicos nativos e invasores. Agentes genotóxicos (como os compos-tos de cobre) podem provocar mutações nas células, com formação de tumores, que po-dem causar efeitos letais ou subletais em um organismo (Arnaiz, 1997). Villela et al. (2006)
verificaram danos celulares significativos na hemolinfa de L. fortunei após 48h de exposi-ção ao sulfato de cobre, em concentrações igual ou superior a 3,75 mg L–1. Também verifica-ram alterações nos valores do índice de danos no DNA, em concentrações igual ou superior a 7,5 mg L–1. Soares et al. (2009) verificaram mortalidade de organismos expostos ao sulfato de cobre em concentrações superiores a 6,69 mg L-1, concentração muito próxima à faixa de concentrações na qual se verifica efeitos muta-gênicos nesta espécie quando exposta ao sulfa-to de cobre. A concentração efetiva de sulfato de co-bre de 8,48 mg L-1 (equivalente a 1,23 mg L-1 de cobre), necessária para causar a mortalidade de 50% da população do mexilhão-dourado, em testes de bancada (Soares et al., 2009), foi su-perior ao padrão permitido para o lançamento de efluentes líquidos (0,5 mg L-1 de cobre) em águas superficiais segundo resolução nº 128 do CONSEMA (2006) e ao padrão para águas de classe 3 (0,013 mg L-1 de cobre) segundo reso-lução nº 357 do CONAMA (2005). As concentrações letais verificadas por Soares et al. (2009) foram superiores àquelas indicadas por Colares et al. (2002b) no con-trole do mexilhão-dourado em captadoras de água para abastecimento da cidade de Porto Alegre (0,5 a 2,0 mg L-1 de sulfato de cobre) pelo Departamento Municipal de Águas e Es-gotos. Mas cabe ressaltar que testes de bancada e de campo diferem. O cálculo da concentração de um mo-luscida a ser aplicado em uma captadora deve
Figura 1. Sulfato de cobre pentahidratado comercial
Figura 2. Equações de regressão e coeficientes de determina-ção obtidos por diagrama de dispersão dos valores de mor-talidade (N) do mexilhão-dourado, L. fortunei, sob diferentes concentrações de sulfato de cobre (48 h de exposição).

CAPÍTULO 30
Seção 9
309
considerar a vazão e a diluição da substância moluscida (Souza, 1995). Além disso, o modo de aplicação do controlador químico (intermi-tente, contínuo ou semicontínuo) deve ser con-siderado no dimensionamento do tratamento (Darrigran & Damborenea, 2006b). A aplicação do sulfato de cobre como controlador químico em sistemas abertos, sem
Tabela I. Concentrações de cobre em tecidos de bivalves límnicos.
Espécie Concentração (mg kg-1) Autor
Limnoperna fortunei (Dunker, 1857) 27,560±2,406 (20,360-40,700) Soares et al. (2009)
Limnoperna fortunei 12,05 e 42,0 Villar et al. (1999)
Cyanocyclas limosa (Maton, 1811) 7,03 e 28,39 Bendati, 2000
Mytella falcata (Orbigny, 1846) 7,30±1,80 Carvalho et al. (2000)
Anodonta grandis Say, 1829 45,30±3,10 Pip (1990)
Tabela II. Concentração letal (CL50
) de sulfato de cobre para causar a mortalidade 50% da população em diferentes espécies de moluscos límnicos e respectivos tempos de exposição (TE).
Espécie CL50 (mg L-1) TE Autor
Limnoperna fortunei (Dunker, 1857) 8,48 48 h Soares et al. (2009)
Corbicula manilensis (Müller, 1774) 0.1 84 h Orme e Kegley (2006)
Biomphalaria alexandrina (Ehrenberg, 1831) 0.005 8 h Orme e Kegley (2006)
Lymnaea natalensis Krauss, 1848 0.8 24 h Orme e Kegley (2006)
Pomacea canaliculata (Lamarck, 1822) 0.003 8 h Orme e Kegley (2006)
Melanoides tuberculata (Müller, 1774) 0.1 a 0.2 24 h Orme e Kegley (2006)
Mytilus galloprovincialis (Lamarck, 1819) 2.5 48 h Barosa et al. (2003)
posterior tratamento da água para a remoção de resíduos de cobre e sem a devida remoção e acon-dicionamento dos mexilhões mortos em aterros sanitários, causaria a contaminação dos corpos hídricos e de toda a sua biodiversidade por meio da cadeia trófica. As dosagens são elevadas e po-dem resultar em resíduos superiores aos padrões estabelecidos pela legislação ambiental.


311
A busca pelo melhor produto para o con-trole do Limnoperna fortunei (Dunker, 1857) estende-se sobre outras substâncias de uso co-mum, na tentativa de mitigar os efeitos da in-crustação sobre os sistemas e instalações. Desta forma, uma variedade de substâncias está sen-do testada e avaliada dentro de diferentes abor-dagens e estratégias de controle. Ainda não se obteve a substância perfeita para controle do mexilhão e, certamente, esta não seria a expec-tativa ou pretensão. No entanto, encontrar e ou desenvolver o uso de substâncias, considerando suas principais características, oferece a possi-bilidade de traçar a melhor estratégia de con-trole para as diferentes condições, sistemas e instalações. Estratégias estas que visem reduzir os riscos laborais e ambientais nos tratamentos, como também diminuir os custos com as dosa-gens dos produtos. Nos capítulos anteriores, acompanhamos os estudos desenvolvidos nesta área para determi-nados produtos específicos. Neste capítulo, iremos abordar algumas outras substâncias com potencial
biocida que também foram testadas para controle do mexilhão-dourado. A tabela a seguir contém os nomes dos produtos que também foram avaliados para controle do L. fortunei. Os diferentes produtos possuem suas peculiaridades e seus efeitos sobre o mexilhão--dourado atuando de diferentes formas, sendo eles coagulantes, disruptores osmótico e altera-dores de pH (Tab. 1). Os resultados e os efei-tos encontrados para estas substâncias estão descritos separadamente de acordo com cada produto. Em relação aos sais, biologicamente os organismos necessitam controlar a quan-tidade de água e de minerais em suas células. Este controle, chamado de osmorregulação, é afetado pela concentração de sais do meio ex-terno e possuem restrições para o controle de mudanças extremas das concentrações. Sendo assim, a aplicação de um choque osmótico em organismos de água doce induz à morte celular acarretando a morte do organismo. Esta técnica pode auxiliar em determinadas circunstâncias, porém, existe o inconveniente de seu uso
CAPÍTULO 31
Outros compostos
Sávio Henrique Calazans CamposLivia Viana de Godoy Fernandes
Flavio da Costa Fernandes

Calazans C. et al.
Outros compostos
312
em instalações em virtude do aumento da corrosão (Polman & Janssen-Mommen, 2011), sem contar que em determinados ambientes o descarte de água salgada pode representar um impacto ambiental severo.
Cloreto de potássio (KCl)
O cloreto de potássio é sabidamente um moluscocida que oferece uma seletividade de-sejada sobre mexilhões em relação aos outros organismos do ecossistema. No entanto, para obterem os mesmos efeitos, diferentes estudos encontraram variações na ordem de grandeza (CL50 de 138 mg.L-1 a 5.500 mg.L-1) para a con-centração do biocida contra o mexilhão-zebra (Fisher et al., 1991; Waller et al., 1993; Wildrid-ge et al., 1998). Campos (2009) verifica o efeito
desta substância em sua forma dissolvida contra o mexilhão-dourado encontrando uma CL50 estimada em 293,38 mg.L-1, e concentrações acima de 4.000 mg.L-1 apresentaram 100% de mortalidade. Uma característica destacada por Campos (2009) foi o efeito letárgico provoca-do pelo KCl: os mexilhões permaneciam com a concha aberta, desfalecidos, com movimentos lentos e até mesmo fechando as valvas e deixan-do parte do corpo para fora (Fig. 1), podendo se restabelecer após a exposição.
Cloreto de sódio (NaCl)
Cloreto de Sódio – NaCl: O cloreto de sódio é um sal muito abundante, encontrado em sua forma dissociada nos oceanos, não sen-do prejudicial para os organismos marinhos.
Tabela 1. Produtos testados contra o mexilhão-dourado.
Produtos Características* Uso* Como efluente*
Cloreto de Potássio (KCl) Sal sólido branco cristalino nas condições normais;
Uso medicinal para reposição de sais como também como injeção letal; aplicações cien-tíficas; processamento de comida no uso doméstico; Fertilizante; Provoca choque osmótico celular;
Causa sérias alterações em ambientes de água doce.
Cloreto de Sódio (NaCl) Sal sólido branco cristalino nas condições normais; Po-pular sal de cozinha.
Uso doméstico; Uso Indus-trial na produção de hidró-xido de sódio, cloro, hidro-gênio e de forma indireta na produção de acido clorídrico por eletrólise de sua solução aquosa.
Causa sérias alterações em ambientes de água doce.
Menadiona Bissulfito de nico-tinamida (MNB) - Kavist®
Coloração creme cristalino, moderadamente solúvel em água.
Forma sintética do precursor da menadiona, precursores da vitamina K3 (envolvida no processo de coagulação do sangue) muito usada como suplemento alimentar de animais, por ser uma forma solúvel em água.
Moderadamente tóxico aos sistemas aquáticos.
MXD-100 Composto de amônia qua-ternária, extrato de plantas e taninos. Capacidade de combinar-se com proteínas e polissacarídeos.
Produto ativo usado em ou-tras finalidades como para proteger materiais contra o ataque de insetos, fungos ou de bactérias em residências
Biodegradável. Sem efeitos conhecidos sobre o ambien-te aquático do local de des-carte.
Hidróxido de Sódio (NaOH) Composto sólido, branco, e cáustico. Provoca queima-duras graves. Popular soda cáustica
Comumente usado em solu-ções bem diluídas para desin-fecção em geral, doméstica, industrial etc. Sua solução possui pH básico, aumento do pH; Consumo de matéria orgânica.
Altera o pH do corpo recep-tor podendo gerar mortanda-de de organismos em contato direto.
*Dados obtidos junto aos fabricantes e em ficha técnica dos produtos (FISPQ e MSDS).

CAPÍTULO 31
Seção 9
313
Tabela II - Valores normativos que caracterizam os tipos de água.
Tipo de água g/L
água doce NaCl < 0,5
água salobra 0,5 > NaCl < 30
água salgada NaCl > 30
Tabela III - Condições de sobrevivência relacionada à salinidade.
Condição Sobrevivência Salinidade g/L
muito favorável > 75% 0 < sal. < 5,1
relativamente favorável < 75% 5,1 < sal. < 11,6
área desfavorável < 50% 11,6 < sal. < 33
* Tabela adaptada de Barbosa & Melo, 2009.
Figura 1 - Organismo vivo, com reação letárgica. Manto para fora da concha, sendo lentamente retraído.
Este sal participa de forma direta no fun-cionamento fisiológico de organismos adapta-dos ao meio, seja água doce ou salgada. Em laboratório, foi encontrado que em 24h de ex-posição a concentrações acima de 8 g/L afetava a condição de sobrevivência do L. fortunei e que em 240h de exposição consegue-se mortalida-de de 100% utilizando concentrações acima de 4 g/L (Angonesi & Bemvenuti, 2008). Estudos de toxicidade aguda para este sal apresentaram uma concentração letal (50% população) em 48h de 5,64 g/L e uma concentração de 20 g/L para atingir 100% de mortalidade em todas as réplicas em 48h (Calazans et al., 2007). Em estudos que verificavam a resistência do L. fortunei em ambientes estuarinos de Hong Kong verificou-se que esta espécie possui baixa tolerância à salinidade, suportando uma varia-ção de 0-12 g/L (Deaton et al., 1989). Na Lagoa dos Patos (RS, Brasil), Angonesi & Bemvenuti (2008) verificam que populações do molusco não resistem a 4 g/L ao longo de 10 dias de ex-posição. Valores que coincidiram com obser-vações realizadas no estuário do Rio da Prata, onde as colônias foram registradas em áreas nas quais a salinidade média não é superior a 3 g/L (Darrigran & Pastorino, 1995; Darrigran, 2002; Brugnoli et al., 2005). No Brasil, temos a classificação de salinidade de corpos hídricos de acordo com CONAMA 357/2005 (Tab. II). Na Lagoa dos Patos um estudo estimou a capacidade de sobrevivência do mexilhão-dou-rado em diferentes concentrações salinas natu-rais, estimando as condições de sobrevivência do mexilhão-dourado em diferentes salinida-des (Barbosa & Melo, 2009) (Tab. III).
Estudos verificaram que o mexilhão-zebra necessita de quantidades mínimas de determi-nados sais (Na, K, Mg e Cl) para sobreviver, não resistindo por muito tempo em água deio-nizada, onde o tempo letal de 50% (TL50) en-contrado foi menor que 4h. No entanto, outros bivalves de água doce toleram água deioniza-da durante semanas, sem mortalidade (Krogh, 1939; Murphy & Dietz, 1976; McCorkle & Dietz, 1980; Scheide & Dietz, 1982; Dietz et al., 1994), inclusive o L. fortunei (Dealton et al., 1989).
Menadiona bissulfito de nicotinamida (MNB)
Fernandes & Coutinho (2008) testaram os efeitos da menadiona bissulfito de nicotina-mida – MNB contra o mexilhão-dourado mo-tivados pelo fabricante Vanetta S.p.A. a qual comercializa o produto como Kavist®. Os re-sultados obtidos mostraram que a MNB gera um efeito de mortalidade muito variável e sem relação dose-dependência (maior concentração = maior mortalidade), onde a CL50 estimada (0,88 mg/L) não corresponde ao valor real na mortalidade encontrada. Este produto também apresentou maior toxicidade sobre os micro-crustáceos Daphnia similis do que sobre o me-xilhão-dourado, desta forma, causando maior efeito sobre os organismos do plâncton do que sobre o organismo-alvo. Uma peculiaridade do produto é que os mexilhões tratados com MNB aparentavam estar mortos, com conchas abertas e sem reação ao toque. Porém, em sua maioria, conseguiam se restabelecer ao serem colocados em água sem o produto (48h após

Calazans C. et al.
Outros compostos
314
a exposição) (Fig. 2). Resultados de morbidez também são relatados por Oduho et al. (1993) em aves, quando em altas doses de MNB como suplemento alimentar.
2011). Alguns autores estudaram a eficiência do MXD-100 como anti-incrustante e obtive-ram bons resultados (Mata et al., 2008; Bonafé et al., 2010). Estudos realizados pela Fundação Centro Tecnológico de Minas Gerais – CETEC definiram CL50 do MXD-100 para o mexi-lhão-dourado como 0,051 mg/L (exposição de 48h ao produto), sem causar toxicidade sobre os microcrustáceos Daphnia similis. Também verificaram que as associações aumentaram a sobrevivência dos mexilhões (Tab. V) (Fernan-des & Coutinho, 2008). Mata et al. (2007) ob-tiveram resultados de 100% de letalidade para larvas de L. fortunei nas concentrações acima de 0,5 mg/l de MXD-100, após 24h de expo-sição. Mäder Netto (2011) conseguiu um con-trole de 99% da bioincrustação do mexilhão--dourado em tubulações de hidrelétricas (por 12 meses, tempo de duração do experimento). O MXD-100 também foi testado nas Usinas de Nova Ponte, Irapé e Queimados da CEMIG, Porto Primavera e Ilha Solteira da
Tabela V – CL50
em ppm* dos produtos testados com Daphnia similis e Limnoperna fortunei.
Substâncias Testadas Daphnia similis Limnoperna fortunei
MXD-100 0,12 0,051
MXD-100 + Kavist 0,28 7,1
MXD-100 + Cloreto de Potássio 5,40 13,40
*partes por mil.
Tabela IV - CL50
em ppm do MNB e associações sobre o L. fortunei e D. simillis.
Substâncias Testadas Daphnia similis Limnoperna fortunei *
MNB 0,88 11
MNB + MXD-100 0,28 7,1
MNB + Cloreto de Potássio 144 874
*partes por mil.
Figura 2 - Efeito “falso-morto” causado pela substância MNB. [Adaptado de Fernandes & Coutinho (2008), onde: 2A, orga-nismo exposto ao produto e 2B, organismo recuperado, após exposição].
Testes combinando MNB com outros produtos foram utilizados pelo laboratório do CETEC – MG comparando os efeitos sobre o mexilhão-dourado e organismos não alvos (D. similis). Os resultados são apresentados na ta-bela abaixo e mostram que todas as suas combi-nações diminuíram os efeitos de toxicidades de outras substâncias sobre o mexilhão (Fernan-des & Coutinho, 2008).
MXD-100
O MXD-100 possui muitas característi-cas que fazem do seu uso como biocida uma ótima opção, dentre elas: fácil manuseio, esto-cagem e aplicação. Testes realizados em labo-ratórios demonstraram que o MXD-100 é bio-degradável (Ecolyzer, 2007 apud Mäder Netto, 2011), não corrosivo a metais (CEFET/MG, 2006), de baixa toxicidade a seres humanos (Ecolyzer 2007 apud Mäder Netto, 2011) e a biota aquática (CETEC/MG, 2007). Uma des-vantagem do seu uso é o custo (Mäder Netto,
CESP, Ibitinga da AES Tietê, Governador José Richa da COPEL, Manso de FURNAS e Itaipu, utilizam ou já testaram o MXD-100 em seus sistemas, considerando como satisfatório os re-sultados obtidos, contudo sem uma avaliação detalhada sobre a eficiência e ou comparação com outras metodologias (Mäder Netto, 2011).
Hidróxido de sódio (NaOH)
O hidróxido de sódio está sendo con-siderada uma ótima opção para controle de mexilhão-dourado em tubulações de hidrelé-

CAPÍTULO 31
Seção 9
315
trica, demonstrando eficiência na diminuição da deposição de lama ferruginosa, redução da corrosão química e biológica (LACTEC, 2004; Segundo et al., 1993). A injeção de NaOH (pH de 8 a 8,5) faz com que os grupos carboxílicos dos ácidos húmicos e compostos orgânicos se-jam ionizados, evitando assim a complexação do ferro e, consequentemente, minimizando a deposição de lama ferruginosa (“fouling”) nos sistemas de resfriamento (LACTEC, 2004; Se-gundo et al., 1993). Em estudo realizado na usina de Salto Caxias, sob condições reais de uso, Mäder Netto (2011) consegue controle de 99% da bioincrustação, em 12 meses de experimento, adicionando ininterruptamente NaOH na tubulação, de forma a manter um pH 9. Em termos gerais, as avaliações de dife-rentes substâncias auxiliam no desenvolvimento de estratégias de controle do mexilhão-doura-do oferecendo novas alternativas de tratamen-to, ao ponto que destaca novos produtos e suas melhores formas de utilização. Em relação às sustâncias citadas neste capítulo podemos des-tacar que o uso do KCl pode ser estratégico,
tendo em vista seu efeito anestésico, podendo ser útil para um controle associado com baixas concentrações de compostos oxidativos (clo-ros), demandando apenas avaliações sobre as possíveis combinações e viabilidade econômi-ca. O MXD-100 e o NaOH apresentaram bons resultados para o controle do mexilhão-doura-do. Entretanto, segundo Mäder Netto (2011), o NaOH foi mais eficiente no controle da depo-sição da lama ferruginosa, além de apresentar um menor custo. Considerando assim o NaOH uma boa opção em relação a outros produtos, principalmente em relação ao seu custo. Atualmente, com as substâncias testadas é possível manter os sistemas de resfriamento, tratados tanto com MXD-100 ou com NaOH, operando normalmente em cursos d’água inva-didos por L. fortunei, sem acontecer redução na vazão do sistema de resfriamento e aumen-tos de temperatura ocasionados pela bioincrus-tação. A continuidade na avaliação de substân-cias certamente irá oferecer ainda mais opções de tratamento que possam se adequar em dife-rentes sistemas e ou condições de regulamenta-ção ambiental.


317
A bioincrustação marinha tem sido enfren-tada por alguns milhares de anos, desde as culturas mais antigas com o uso de cera, alcatrão e asfalto, bem como no período dos Fenícios e Cartagineses com a utilização de piche e, possivelmente, cobre como cobertura (Yebra et al., 2004; Lopes et al., 2004). A evolução apontou para o uso de folhas de cobre aos cascos de madeira, porém com o uso de embarcações de ferro, esta alternativa ficou im-possibilitada devido à ocorrência de corrosão gal-vânica. Até meados de 2000, o método mais utili-zado de combate à bioincrustação foi a utilização de tintas contendo TBT (tributil-estanho). Porém, com a proibição de uso destes compostos organo--estanho pela “International Maritime Organiza-tion” – IMO – Agência das Nações Unidas, foi necessário encontrar novas alternativas ambien-talmente viáveis e sustentáveis. Algumas alterna-tivas desenvolvidas foram: revestimentos a base de tintas com cobre, resinas, de teflon, utilização de biocidas naturais e substância 5-ciano-4-(4- clo-rofenilamino)tieno[2,3-b] piridina como agentes anti-incrustantes (Meador, 2000).
Diversas pesquisas têm sido realizadas no intuito de investigar materiais e revestimentos que proporcionem soluções via seleção de ma-terial e engenharia de superfícies para o contro-le de incrustações do mexilhão-dourado. Faria et al. (2006) testaram as características anti-in-crustantes de materiais de diferentes naturezas submersos no Rio Paraguai: i) cerâmico: vidro; ii) metais: aço inoxidável, alumínio, aço carbo-no, zinco, cobre, latão e iii) polímeros: polieti-leno, polipropileno, poliuretano, PET, PVC e teflon. Experimentos semelhantes foram reali-zados com CPs revestidos com tintas a base de óxidos de cobre e zinco na Usina Hidrelétrica de Jaciretá, no Rio Paraná (Caprari & Lecot, 2001). Caprari (2006) apresenta uma revisão sobre fundamentos e métodos experimentais com tintas anti-incrustantes para o controle de incrustações do mexilhão-dourado. Outra maneira de prevenir e eliminar os efeitos nocivos de moluscos é desenvolven-do materiais que controlam os fenômenos que acontecem na biointerface, envolvendo intera-
CAPÍTULO 32
Seleção de materiais e revestimentos para o controle de incrustações do
mexilhão-dourado em hidrelétricas
Carlos Pérez BergmannJuliane Vicenzi
Tania Maria BasegioDaniela Lupinacci Villanova
Daniel PereiraPaulo Bergonci
Cintia Pinheiro dos Santos Maria Cristina Dreher Mansur

Bergmann et al.
Seleção de materiais e revestimentos para o controle de incrustações do mexilhão-dourado em hidrelétricas
318
ções entre superfícies de material e o meio bio-lógico. Eventualmente parte deste esforço leva ao desenvolvimento de processos que transfor-mem as superfícies mais resistentes à deteriora-ção (fouling) devido a proteínas, células, bacté-rias ou outros organismos. No caso do mexilhão-dourado, evitar a incrustação deste molusco em materiais sig-nifica inibir sua fixação em superfícies através da modificação das mesmas. Em usinas hidre-létricas o material base de equipamentos, tu-bulações, grades, comportas é, geralmente o aço. Este material devido suas características de superfície e composição química não evi-ta a incrustação do mexilhão-dourado. Logo, entende-se que a modificação superficial seja uma alternativa eficaz para inibir tal compor-tamento do molusco, bem como viável econo-micamente para as Usinas. Este capítulo tem por objetivo apresen-tar resultados de pesquisas laboratoriais e in loco sobre a utilização de diferentes materiais e revestimentos visando o controle de incrus-tações do mexilhão-dourado. Alguns desses resultados já foram obtidos por meio de pro-jeto P&D ANEEL/AES TIETÊ e publicados em Bergmann et al. (2010a e b). Outros foram obtidos por meio de P&D ANEEL/CEEE e são inéditos.
Testes laboratoriais: investigação de tintas com pigmentos nanoestruturados de óxidos de cobre, zinco e cobre+zinco
Em testes laboratoriais foram avaliadas 18 estações divididas em 3 grupos de revesti-mentos nanoestruturados a base de: óxido de cobre, óxido de zinco e óxidos de cobre e zin-co, considerando-se metodologia de Mansur et al. (2009). Os CPs utilizados foram placas me-tálicas de aço com revestimento de tintas ela-boradas com óxidos nanoestruturados de zinco e de cobre obtidos a partir da síntese por com-bustão. Estes óxidos foram misturados em di-ferentes concentrações (2,5%, 5%, 7,5%, 10% e 12,5%) a uma tinta esmalte base para metais e a seguir a mesma foi aplicada sobre as placas metálicas. As placas metálicas foram dispostas no fundo da estação de testes e para avaliar a eficiência dos óxidos nanoestruturados foram
dispostos 30 mexilhões dourado jovens. Os organismos permaneceram nas caixas pelo período de três meses. Amostras de água fo-ram recolhidas para a análise de metais (cobre e zinco), além disso, as variáveis temperatura, pH, oxigênio dissolvido e condutividade foram mensuradas. Durante o período de realização dos testes seguiu-se o protocolo de avaliação adaptado de Soares et al. (2008). Os resultados (Fig. 1) mostraram que para a tinta com pigmentos nanoestruturado de cobre apenas na concentração de 7,5% ocorreu uma maior mortalidade dos mexilhões-dourado e constatou-se a fixação de mexilhões junto às placas. Já para a tinta com acréscimos de ZnO foi observado que a partir de 2,5% de adição deste óxido houve a mortalidade de 90% dos indivíduos. Acréscimos maiores do óxido oca-sionaram 100% da mortalidade do mexilhão--dourado. Constatou-se uma tendência logarít-mica na mortalidade de mexilhões em função do acréscimo de ZnO nanoestruturado na tinta a base esmalte, com uma significância elevada (R2 = 0,99). Pode-se considerar então a efici-ência deste pigmento nanoestruturado devido a maior mortalidade em relação ao CuO, bem como em função da tendência logarítmica apre-sentada. O uso sinérgico dos dois óxidos não ocasionou mudança significativa de comporta-mento em relação ao acréscimo de apenas ZnO. Notou-se que as demais variáveis da água avaliadas não foram fatores determinantes na mortalidade do mexilhão-dourado, pois se man-tiveram em níveis aceitáveis para a sobrevivência do mexilhão-dourado: i) temperatura: de 24,3 a 28,3ºC; ii) pH: 4,8 a 5,9; iii) OD: 7,8% a 9,7%. Apenas a condutividade elétrica da água apre-sentou variações significativas em relação às es-
Figura 1. Correlação entre mortalidade do mexilhão-dourado e revestimentos com diferentes concentrações de óxidos na-nométricos de cobre, zinco e cobre + zinco.

CAPÍTULO 32
Seção 9
319
tações experimentais com tintas á base de ZnO e CuO + ZnO nanoestruturados, destacando-se uma tendência de acréscimo da condutividade com o tempo de exposição das placas metálicas com tintas na água. Esse aumento sugere que es-teja ocorrendo a solubilidade dos metais. Talvez este seja um dos fatores determinantes para a mortalidade do mexilhão-dourado. Para verifi-car esta observação, avaliou-se a lixiviação dos metais durante o experimento. Para as estações experimentais com corpos-de-prova revestidos com tintas e pig-mento nanoestruturado de CuO, praticamente não houve lixiviação de metais, e nota-se uma aleatoriedade na mortalidade deste bioinvasor. Infere-se assim, que provavelmente o metal Cu não colaborou para a mortalidade do mexilhão. Para as estações experimentais com cor-pos-de-prova revestidos com tintas e pigmento nanoestruturado de ZnO, praticamente não houve lixiviação do metal Cu e a lixiviação do Zn apresentou um máximo de 1,53 mg/L para o revestimento com 7,5% de ZnO nanoestrutu-rado para 62 dias de ensaio. Com o auxilio da Figura 2, nota-se a tendência exponencial da mortalidade em função do tempo transcorrido de ensaio. Concluiu-se a partir da análise dos resultados que a lixiviação de Zn na ordem de até 0,5 mg/L, apresenta-se suficiente para oca-sionar a mortalidade do mexilhão-dourado. Nas estações experimentais com corpos--de-prova revestidos com tintas e pigmentos nanoestruturados da mistura de CuO+ZnO, praticamente não houve lixiviação do metal Cu e a lixiviação do Zn apresentou picos de concentração de Zn em função da quantidade de dias de exposição dos CPs, bem como da quantidade adicionada de CuO+ZnO. Para qualquer quantidade de CuO+ZnO nanoestru-
turados adicionados em 35 dias de exposição ocorreu o máximo de lixiviação do metal Zn. Na Figura 2 nota-se uma tendência exponen-cial da mortalidade em função do tempo trans-corrido de ensaio, com o máximo em 7 dias para as concentrações investigadas. Concluiu--se ainda que a lixiviação de Zn na ordem de 0,5 mg/L, apresentou-se suficiente para ocasio-nar a mortalidade do mexilhão-dourado. Comparando-se os valores lixiviados de Cu e Zn em função dos valores permitidos de dissolução destes metais na água, segundo reso-luções ambientais, pode-se notar que não foram atingidos os valores limites, isto é estão abaixo dos valores estabelecidos por lei. Observa-se que os valores de cobre dissolvido na água são infe-riores ao padrão permitido para lançamentos de efluentes líquidos (0,5 mg/L de cobre) em águas superficiais segundo resolução n. 128 do CON-SEMA (2006) e ao padrão para águas de classe 3 (0,013 mg/L de cobre) segundo resolução n. 357 do CONAMA (2005). Quanto aos valores de zinco dissolvido na água, estes são inferiores ao padrão permitido para lançamentos de efluentes líquidos (2,0 mg/L de zinco) em águas superfi-ciais segundo resolução n. 128 do CONSEMA (2006) e superiores ao padrão para águas de classe 3 (0,09 mg/L de zinco) segundo resolução n. 357 do CONAMA (2005).
Testes in loco: avaliação de revestimentos com óxidos nanométricos e convencional em tinta a base de resina epóxi poliamina
A fim de testar e validar os materiais e reves-timentos anti-incrustantes ao mexilhão-dourado em testes in loco foi realizado um experimen-
Figura 2. Número de mexilhões mortos nas estações experimentais de CuO (2A), ZnO (2B) e CuO + ZnO (2C) em função do tempo de experimento e da quantidade de óxido adicionado à tinta a base esmalte.

Bergmann et al.
Seleção de materiais e revestimentos para o controle de incrustações do mexilhão-dourado em hidrelétricas
320
to com um ano de duração, no reservatório da Usina Hidrelétrica de Ibitinga (UHE Ibitinga - AES Tietê), na bacia do médio Rio Tietê/Jacaré, município de Ibitinga, estado de São Paulo, na localização (21º45’26.05”S e 49º59’30.11”W e altitude de 398 m) do experimento junto ao paredão do reservatório. A escolha do local de submersão se deu a partir da verificação de grandes incrustações na referida UHE. Os CPs (CP) utilizados foram placas metálicas retangulares (21 x 6 x 0,3 cm) com área total de 252 cm2. Foram utilizados CPs sem e com revestimento (Tab. I). Os revesti-mentos aplicados às superfícies dos CPs cons-tituem uma mistura de óxidos (nanométricos ou convencionais) em diferentes concentrações e uma tinta comercial a base de resina epóxi poliamina (REP), livre de solventes orgânicos, com produtos de funções anticorrosivas, gran-de resistência química, que reage com a água e com os óxidos do substrato, proporcionan-do excelente aderência e proteção, com alta retenção nas arestas. Os CPs foram fixados aleatoriamente em estruturas metálicas deno-minadas bastidores (Caprari & Lecot, 2001; Caprari, 2006) e submersos no reservatório da UHE Ibitinga. Ficaram submersos durante os meses de outubro (2007) a outubro (2008). Foi considerada a incrustação de toda a super-fície do CP para fins de quantificação da den-sidade (ind.cm-2). Após a retirada dos CPs da água, estes foram secos em forno de Pasteur a 60ºC e os aglomerados formados por adultos quantificados. Aplicou-se a análise de variância ANO-VA com nível de significância de 95% segui-da de múltiplas comparações pelo método de Duncan (p = 0,05), com o auxílio do pacote estatístico SPSS 11.5. Os valores densidades de mexilhões nos CPs foram submetidos à transformação logarítmica (Log (x+1)) a fim de que fossem minimizados os desvios da nor-malidade. Avaliou-se a presença larval na área de estudo para evitar que este fosse um parâmetro que causasse erro na avaliação dos resultados. Para tanto, utilizaram-se as metodologias de i) amostragem: Santos et al. (2005), ii) recru-tamento em substrato cerâmico: Soares et al. (2008) e iii) amostragem e quantificação dos mexilhões: Bergonci et al. (2009).
Os valores médios, erro padrão, míni-mo e máximo da densidade de mexilhões ade-ridos aos CPs estão discriminados na Tabela II. De acordo com a avaliação ANOVA houve a diferenciação entre alguns dos CPs testados (Fcalc = 4,108; p = 0,000). Segundo o teste de Duncan realizado a posteriori, houve a for-mação de cinco grupos para os valores da den-sidade de mexilhões incrustados sobre os CPs, conforme a Figura 3.Tabela I. Materiais e revestimentos testados (Fonte: Berg-mann et al., 2010a).
Material Revestimento
1 Aço galvanizado (AG) Ausente
2 Aço carbono (AC) Ausente
2.2 Tinta comercial a base de Resi-na Epóxi Poliamina (REP) + óxido de cobre nanométrico (nan) 5,0%
2.3 REP + óxido de cobre nan 10,0%
2.4 REP + óxido de cobre convencional (con) 5,0%
2.5 REP + óxido de cobre con 10,0%
2.6 REP + óxido de zinco nan 5,0%
2.7 REP + óxido de zinco nan 5,0%
2.8 REP + óxido de zinco nan 10,0%
2.9 REP + óxido de cobre con 2,5% + óxido de zinco con 2,5%
2.10 REP + óxido de cobre nan 2,5% + óxido de zinco nan 2,5%
2.11 REP + oxido de cobre nan 5,0% + óxido de zinco nan 5,0%
2.12 REP + óxido de cobre nan 38,0%
2.13 REP + óxido de cobre nan 15,0%
2.14 REP + óxido de cobre nan 0,0%
2.15 REP + óxido de cobre con 38,0%
2.16 REP + óxido de cobre con 15,0%
2.17 REP + óxido de cobre con 0,0%
3 Aço inox (AI) Ausente
4 Aço cobreado (PC) Ausente
5 Acrílico (ACR) Ausente

CAPÍTULO 32
Seção 9
321
O grupo 1, constituído por aço carbono revestido com tinta a base de resina epóxi po-liamina óxido de zinco convencional 5% e o aço carbono revestido com tinta a base de re-sina epóxi poliamina com óxido de cobre con-vencional 10% foram os que se demonstraram mais eficientes no controle das incrustações de L. fortunei. Os demais grupos, em sua maioria, apresentaram uma considerável sobreposição das densidades nos CPs e se revelaram pouco eficientes. Segundo Faria et al. (2006), com base em experimentos de laboratório e campo, os materiais que formam óxidos, como o zin-co e o aço cobreado, impedem a fixação do mexilhão-dourado e proporcionam alta mor-talidade dos organismos frente a esses óxidos, principalmente em placas de cobre. Esta in-formação corrobora com os resultados obti-
Tabela II. Densidade média (ind.cm-2), erro padrão e valores mínimo e máximo, em ordem crescente, de espécimes de L. fortunei aderidos aos materiais e revestimentos testados (Fonte: Bergmann et al., 2010a).
Nº Material Média Erro Padrão Mínimo Máximo N
1 AC* REP** + ZnO con 5% 0,173 0,047 0,111 0,266 3
2 AC REP + CuO con*** 10% 0,351 0,139 0,079 0,540 3
3 AC REP + ZnO nan**** 10% 0,992 0,362 0,456 1,683 3
4 PC 1,027 0,269 0,060 3,040 12
5 AC REP + CuO nan 10% 1,176 0,410 0,492 1,909 3
6 AC REP + CuO con 5% 1,214 0,527 0,321 2,488 4
7 AC REP + CuO con 2,5% + ZnO con 2,5% 1,332 0,434 0,464 1,786 3
8 AC REP + ZnO con 10% 1,470 0,527 0,417 2,032 3
9 AC REP + CuO nan 5% 1,608 0,711 0,187 2,365 3
10 AC REP + CuO nan 0% 1,646 0,427 0,563 2,909 5
11 AC REP + CuO nan 15% 1,684 0,581 0,583 2,560 3
12 AC REP + CuO con 0% 1,775 0,495 0,833 3,694 5
13 AC REP + CuO nan 38% 1,809 0,420 0,837 2,698 4
14 AC REP + CuO nan 2,5% + ZnO nan 2,5% 1,848 1,213 0,278 4,234 3
15 AG 2,093 0,361 0,512 4,659 12
16 AC REP + CuO con 38% 2,713 1,214 1,119 9,968 7
17 AC REP + CuO con 15% 3,163 0,614 1,694 5,210 6
18 AC 3,706 0,739 0,782 6,635 10
19 AI 3,986 0,821 1,611 8,913 11
20 ACR 4,006 0,489 1,020 6,802 10
21 AC REP + CuO nan 5% + ZnO nan 5% 4,646 3,409 0,282 11,365 3
dos no teste in loco nos CPs em aço, revestidos com pigmento a base de óxido de zinco con 5% e óxido de cobre con 10%. De acordo com Caprari & Lecot (2001), com base em experi-mentos de campo, os revestimentos com base em partículas de óxidos de zinco se demons-traram promissores para o controle da fixação do mexilhão-dourado. Resultado semelhante foi observado nos testes laboratoriais descri-tos neste capítulo. Conforme Caprari (2006), a solubilida-de do óxido cuproso (Cu2O), um dos óxidos mais amplamente utilizados em revestimentos anti-incrustantes, é baixa em água doce. Pou-co se sabe sobre os mecanismos de solubili-zação deste óxido em ambientes límnicos, já que seu emprego se restringe ao controle de incrustações pelo mexilhão-dourado. Na água do mar, onde a solubilidade deste óxido é alta,

Bergmann et al.
Seleção de materiais e revestimentos para o controle de incrustações do mexilhão-dourado em hidrelétricas
322
Figura 3. Densidade média de L. fortunei nos respectivos CPs tes-tados no reservatório da UHE Ibitinga, São Paulo. Grupos forma-dos por meio da ANOVA (Duncan a posteriori): G1 (p = 0,397); G2 (p = 0,065); G3 (p = 0,132); G4 (p = 0,061); G5 (p = 0,072).
Figura 4. Equipamento incrustados pelo mexilhão-dourado na UHE Ibitinga, município de Ibitinga, estado de São Paulo. A: cesto de inox; B: tampa e C: parte superior da junção dos cestos do filtro de Hi-flow (Fonte: Bergmann et al., 2010a).
Figura 5. Tampas dos filtros de Hi-flow após a retirada. Perma-nência das mesmas durante 11 meses de operação. A: cobre, destacando a presença de um indivíduo de mexilhão-dourado e B: de zinco, não se constatou a presença deste bioinvasor.
tem demonstrado eficiência sobre todos os or-ganismos incrustantes, a exceção das algas. O aço inoxidável, um dos CPs que apresentaram maior quantidade de mexi-lhões aderidos neste estudo, corrobora com os experimentos realizados por Faria et al. (2006), que constataram que este não é um bom material anti-incrustante, devido à alta força média de desligamento e maior por-centagem de organismos aderidos. Quando comparados ao substrato cerâmico submerso no mesmo período na UHE, o aço carbono revestido com tinta a base de resina epóxi poliamina apresentou elevados percentuais de redução da bioincrustação.
Testes in loco: avaliação de revestimentos de Cu e Zn em partes de equipamentos
Aplicou-se, via aspersão térmica, a de-posição de cobre e zinco metálicos em tam-pas de dois filtros Hi-flow contaminados (Fig. 4) na UHE Bariri, situada logo a mon-tante da UHE Ibitinga. Neste experimento foram escolhidos estes revestimentos devido ao curto espaço de tempo para a parada do equipamento, evitando o processo demorado de pintura que exige várias de mãos de tinta. Por outro lado a aspersão térmica é um pro-cedimento rápido, com duração de no máxi-mo duas horas. Além disso, a deposição de metal tem maior vida útil do que o revesti-mento por tintas anti-incrustantes. O perío-do deste experimento foi de 11 meses. Os resultados mostraram que cobre e zinco metálicos são eficientes quanto à in-crustação de L. fortunei, tendo em vista que as tampas do filtro Hi-Flow permaneceram por 11 meses em operação. Pode-se observar que apenas na tampa revestida com cobre houve a ocorrência de um indivíduo incrus-tado, visível a olho nu, conforme a Figura 5. Também se fez a coleta do material da super-fície das tampas e analisou-se o mesmo, onde não foi verificada a presença de indivíduos recrutas. Pode-se concluir que o uso de revesti-mentos de Cu e Zn depositados por arco spray apresentou elevada eficácia em relação aos materiais sem proteção.

323
Antes da invasão do mexilhão-dourado, a incrustação em regiões neotropicais era res-trita às águas salgadas ou estuarinas. Hoje, de-vido a sua fecundidade elevada e à ausência de inimigos naturais eficientes, é possível encon-trar organismos incrustantes que obstruem as tubulações e os sistemas de água para as cida-des, indústrias e hidrelétricas (Darrigran et al., 2002). Cascos de pequenas embarcações são sem dúvida um dos principais mecanismos de dispersão do mexilhão-dourado atualmente nas regiões sul e sudeste do Brasil. A capacida-de dessa espécie de se fixar e ser transportada por esses vetores é grande se considerarmos a reduzida velocidade que essas embarcações atingem. Além disso, tintas anti-incrustantes utilizadas em sistemas de água doce não pos-suem a mesma eficácia daquelas utilizadas em ambientes marinhos onde as formas incrustan-tes são bem mais diversas e as tintas bem mais tóxicas. Torna-se, por isso, imprescindível que tintas anti-incrustantes que controlem de for-
ma eficiente a fixação do mexilhão-dourado nos cascos de embarcações sejam utilizadas para que se reduza sua dispersão nos ecossiste-mas aquáticos Brasileiros pelas embarcações. Além das embarcações, a presença do mexilhão-dourado em estruturas metálicas, em concreto no interior de usinas e em estações de tratamento de água tem causado sérios proble-mas e altos custos de manutenção e limpeza. Com o propósito de identificar qual re-vestimento anti-incrustante existente no mer-cado brasileiro permite o maior controle da fi-xação de Limnoperna fortunei (Dunker, 1857) em embarcações e estruturas submersas com um menor custo ambiental foram desenvolvi-das uma série de testes no Instituto de Estudos do Mar Almirante Paulo Moreira. Para tal, primeiramente foram realiza-dos contatos com diferentes grupos de pes-quisa visando obter resultados de testes anti--incrustantes disponíveis na forma de relatório ou publicações científicas. Após análise desses trabalhos, as tintas que apresentaram os me-
CAPÍTULO 33
Tintas anti-incrustantes no controle do mexilhão-dourado
Ricardo CoutinhoEstevão Carino Fernandes de Souza
Lilian Haje

Coutinho et al.
Tintas anti-incrustantes no controle do mexilhão-dourado
324
lhores resultados foram utilizadas nos testes anti-incrustantes em conjunto com outras for-mulações. Além disso, inicialmente foi realizado um levantamento dos fabricantes de revesti-mentos industriais no Brasil para que fossem identificadas as empresas e produtos que pode-riam ser utilizados na pesquisa. Oitenta empre-sas foram encontradas. Destas, 17 trabalham com produtos anti-incrustantes. Estas empre-sas foram convidadas a participar da pesquisa através de carta consulta e e-mail, mas somente 8 se interessaram, fornecendo 11 produtos para serem testados. Os testes em laboratório foram reali-zados com placas de madeira preparadas no Laboratório de Engenharia e Modificação de Superfícies – LEMS do CETEC – Fundação Centro Tecnológico de Minas Gerais. As más-caras foram feitas com fita crepe e retiradas após a cura completa de cada revestimento. Os revestimentos foram aplicados com trin-chas seguindo a recomendação de cada um dos fabricantes. Indivíduos jovens do mexilhão--dourado foram coletados na Bacia do Lago Guaíba, Rio Grande do Sul e mantidos em tanques no laboratório de processos incrustan-tes do IEAPM, em Arraial do Cabo, RJ, para serem utilizados nos experimentos. Os corpos de prova (placas de madeira) foram pintados em padrão “tabuleiro de xadrez” com um dos revestimentos anti-incrustantes em quadrados 3 x 3 cm intercalados com quadrados controle
(sem pintura). Em seguida, foram lavados em água corrente e submersos em bandejas, para liberação residual dos revestimentos, durante um período de 120 horas antes do início do experimento. Os mexilhões foram colocados em uma bacia com aeração em água desclorada durante um período de 4 horas. Nos ensaios, 3 indivíduos foram colocados em cubas plásticas contendo 2 litros de água desclorada sobre o centro de uma placa de madeira quadrada de 15 cm de lado. Dez réplicas foram feitas para cada revestimento avaliado. O experimento foi realizado durante 15 horas, a partir das 16 ho-ras, em função do comportamento mais ativo dos mexilhões no período noturno. A posição e fixação dos mexilhões foram observadas ao fim do experimento e o número de indivídu-os presentes nas regiões “tratamento” e “con-trole” das placas foram comparados para cada revestimento analisado com teste t de Student (Zar, 1999). Os indivíduos foram observados nas 24 horas seguintes ao fim do experimento para observar um possível efeito letal dos reves-timentos avaliados em médio prazo. A toxicidade dos revestimentos utiliza-dos nos bioensaios em laboratório foi analisada no Laboratório de Ecotoxicologia do CETEC, em Belo Horizonte – MG. Em todos os bioensaios realizados a quantidade de mexilhões presentes e/ou fixa-dos na região “tratamento” das placas avalia-das foi maior que na região “controle” (Tab. I) para todos os 11 revestimentos analisados.
Tabela I. Comparação (teste t de Student) entre o número de indivíduos de Limnoperna fortunei presentes nas regiões “tratamento” (n=8) e “controle” (n=8) das placas utilizadas nos bioensaios de laboratório para cada revestimento avaliado.
Tratamento Controle
Revestimentos avaliados X EP X EP t P
B 30 - 0 - - -
A 19 0,98 11 0,98 5,76 <0,01
E 20,12 1,56 9,87 1,56 4,64 <0,01
I 20,62 0,68 9,37 0,68 11,7 <0,01
K 18,75 1,57 8,25 1,57 4,74 <0,01
G 21,75 1,25 8,25 1,25 7,64 <0,01
C 22,25 1,40 7,75 1,40 7,33 <0,01
F 24,62 1,05 5,37 1,05 12,95 <0,01
J 25,62 1,10 4,37 1,10 13,65 <0,01
D 27 0,91 3 0,91 18,72 <0,01
H 27,50 0,57 2,63 0,65 28,77 <0,01
* todos os indivíduos permaneceram imóveis até o fim do experimento.

CAPÍTULO 33
Seção 9
325
Nos testes realizados no Laboratório de Ecotoxicologia do CETEC, 3 revestimentos foram classificados como tendo “alta toxicida-de”, 2 como “média” e 4 como “baixa” (Tab. II). O revestimento C, feito a base de silicone, não foi avaliado por ser um anti-incrustante mecânico, não químico. O revestimento I, não foi avaliado por problemas operacionais. Nas placas em que foi testado o reves-timento B, todos os mexilhões permaneceram imóveis em todo o período de teste sugerindo um efeito letal muito forte do revestimento, o que poderia ser comprovado pelo teste de to-xicidade (Tab. II). Nos demais revestimentos, apesar do número de indivíduos ter sido maior na região “tratamento” do que na “controle”, não houve fixação dos mexilhões com forma-ção de bisso, mostrando uma eficácia dos pro-dutos para impedir a incrustação do substra-to pelos mexilhões. Nesse contexto, os cinco revestimentos selecionados para os testes de campo por apresentarem uma combinação en-tre eficiência na inibição da fixação do mexi-lhão-dourado nos bioensaios em laboratório e baixa toxicidade foram E, F, H, J e K. Após os ensaios de laboratório, os cin-co revestimentos anti-incrustantes que apre-sentaram uma combinação entre eficiência na inibição da fixação do mexilhão-dourado nos bioensaios em laboratório e baixa toxicidade foram testados no campo. Os testes foram rea-lizados em 4 locais de ocorrência do mexilhão--dourado: na bacia do Lago Guaíba – RS, Porto Rico – PR, Santa Clara D’Oeste – SP e Corum-bá – MS, entre janeiro e maio de 2007. Estru-turas foram confeccionadas para a colocação das placas nas áreas testadas. Cada estrutura era composta por um cano rígido de PVC de 1 ¼ com 2 metros de comprimento, preso por cabos pelas extremidades a uma boia que era presa num píer. As placas de madeira de 20 x 25 cm de lado e 15 mm de espessura foram transpassadas pelos canos e travadas por fios de cobre para evitar o contato entre elas. Em cada estrutura haviam 5 placas pintadas com cada um dos cinco revestimentos testados e 2 placas controle, uma para o revestimento (pin-tada apenas com o primer) e outra para o pri-mer. Em cada local de estudo foram colocadas 4 estruturas. Após o período de estudo, todo o material perifítico das placas foi removido com
pincel para frascos de 250 ml e fixados em ál-cool a 70%, totalizando 28 amostras por local estudado. O material foi avaliado em microscó-pio invertido com objetiva de 40x em lâmina de sedimentação no laboratório de Microscopia do IEAPM, pela Dra. Maria Helena Baeta Neves, para verificar a presença do mexilhão-dourado nas amostras e contagem de pós-larvas. Dentre as localidades avaliadas nos bio-ensaios de campo, as placas das estruturas de Corumbá não apresentaram nenhuma fixação de organismos devido ao fenômeno da deco-ada. Já em Santa Clara D’Oeste, Porto Rico e Guaíba, a fixação do mexilhão-dourado variou bastante entre os locais e entre os revestimen-tos avaliados. Em laboratório, foram encontra-dos 36 taxa do fitobentos no Rio Paraná (Porto Rico), 28 no Lago Guaíba (Porto Alegre) e 45 no Rio Grande (São Paulo). No bioensaio realizado em Porto Alegre, no Lago Guaíba, houve diferença na fixação do mexilhão-dourado entre as placas dos di-ferentes revestimentos testados (ANOVA, F = 19,62; g.l. = 6; p<0,001). E e H não diferiram (p>0,05) do primer e do controle na fixação dos mexilhões. Os revestimentos de melhor de-sempenho por rank de eficiência foram J, K e F, embora não tenha havido diferença significati-va na fixação de pós-larvas. Em Santa Clara D’Oeste, no Rio Grande, não houve diferença na fixação do mexilhão--dourado entre as placas dos diferentes revesti-mentos testados e do primer (p>0,05 para to-das as comparações), mas todos diferiram das placas controle (ANOVA, F = 11,09; g.l. = 6; p<0,001). Todos os revestimentos avaliados fo-ram eficientes e apresentaram baixos níveis de
Tabela II. Toxicidade dos revestimentos avaliados com a Con-centração Letal para 50% da amostra (CL
50) de Daphnia similis.
Revestimentos avaliados CL50 (%) Toxicidade
B 1,47 Alta
G 10,21 Alta
A 13,63 Alta
H 33,32 Média
D 33,77 Media
K 49,94 Baixa
E 52,95 Baixa
F 72,95 Baixa
J 75,48 Baixa
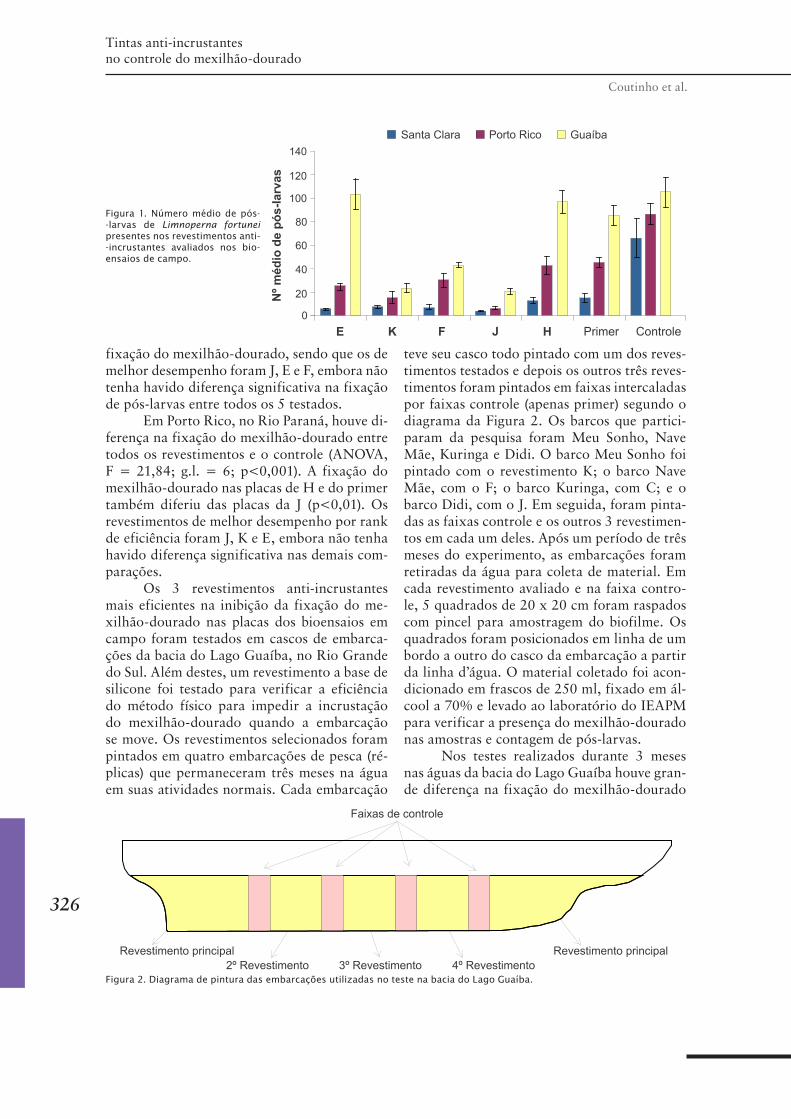
Coutinho et al.
Tintas anti-incrustantes no controle do mexilhão-dourado
326
fixação do mexilhão-dourado, sendo que os de melhor desempenho foram J, E e F, embora não tenha havido diferença significativa na fixação de pós-larvas entre todos os 5 testados. Em Porto Rico, no Rio Paraná, houve di-ferença na fixação do mexilhão-dourado entre todos os revestimentos e o controle (ANOVA, F = 21,84; g.l. = 6; p<0,001). A fixação do mexilhão-dourado nas placas de H e do primer também diferiu das placas da J (p<0,01). Os revestimentos de melhor desempenho por rank de eficiência foram J, K e E, embora não tenha havido diferença significativa nas demais com-parações. Os 3 revestimentos anti-incrustantes mais eficientes na inibição da fixação do me-xilhão-dourado nas placas dos bioensaios em campo foram testados em cascos de embarca-ções da bacia do Lago Guaíba, no Rio Grande do Sul. Além destes, um revestimento a base de silicone foi testado para verificar a eficiência do método físico para impedir a incrustação do mexilhão-dourado quando a embarcação se move. Os revestimentos selecionados foram pintados em quatro embarcações de pesca (ré-plicas) que permaneceram três meses na água em suas atividades normais. Cada embarcação
teve seu casco todo pintado com um dos reves-timentos testados e depois os outros três reves-timentos foram pintados em faixas intercaladas por faixas controle (apenas primer) segundo o diagrama da Figura 2. Os barcos que partici-param da pesquisa foram Meu Sonho, Nave Mãe, Kuringa e Didi. O barco Meu Sonho foi pintado com o revestimento K; o barco Nave Mãe, com o F; o barco Kuringa, com C; e o barco Didi, com o J. Em seguida, foram pinta-das as faixas controle e os outros 3 revestimen-tos em cada um deles. Após um período de três meses do experimento, as embarcações foram retiradas da água para coleta de material. Em cada revestimento avaliado e na faixa contro-le, 5 quadrados de 20 x 20 cm foram raspados com pincel para amostragem do biofilme. Os quadrados foram posicionados em linha de um bordo a outro do casco da embarcação a partir da linha d’água. O material coletado foi acon-dicionado em frascos de 250 ml, fixado em ál-cool a 70% e levado ao laboratório do IEAPM para verificar a presença do mexilhão-dourado nas amostras e contagem de pós-larvas. Nos testes realizados durante 3 meses nas águas da bacia do Lago Guaíba houve gran-de diferença na fixação do mexilhão-dourado
Figura 1. Número médio de pós--larvas de Limnoperna fortunei presentes nos revestimentos anti--incrustantes avaliados nos bio-ensaios de campo.
Figura 2. Diagrama de pintura das embarcações utilizadas no teste na bacia do Lago Guaíba.

CAPÍTULO 33
Seção 9
327
entre as embarcações e entre os revestimentos avaliados. No barco Didi não houve diferença na fixação do mexilhão-dourado entre os diferen-tes revestimentos testados e o controle (p>0,05 para todas as comparações) com exceção da J e K (p<0,01 para ambas as comparações) (ANOVA, F = 6,66; g.l. = 4; p<0,01). A não existência de diferença significativa na fixação de pós-larvas segundo o teste de Tukey deve--se provavelmente à heterogeneidade de vari-âncias (teste de Cochran, C = 0,88; g.l. = 4; p<0,001). Os revestimentos de melhor desem-penho por rank de eficiência foram J, K e C. No barco Kuringa também não houve di-ferença na fixação do mexilhão-dourado entre os diferentes revestimentos testados e o con-trole (p>0,05 para todas as comparações) com exceção da J (p<0,01) (ANOVA, F = 6,68; g.l. = 4; p<0,01). A não existência de diferença significativa na fixação de pós-larvas segundo o teste de Tukey deve-se provavelmente à hete-rogeneidade de variâncias (teste de Cochran, C = 0,68; g.l. = 4; p<0,001). Os revestimentos de melhor desempenho por rank de eficiência foram J, K, C e F, embora não tenha havido diferença significativa na fixação de pós-larvas. No barco Meu Sonho não houve diferen-ça na fixação do mexilhão-dourado entre os diferentes revestimentos testados e o controle (ANOVA, F = 2,78; g.l. = 4; p>0,05). A não existência de diferença significativa na fixa-ção de pós-larvas segundo a ANOVA, apesar de se verificar visualmente (Fig. 2) uma menor fixação do mexilhão nos revestimentos F e K deve-se provavelmente à heterogeneidade de variâncias (teste de Cochran, C = 0,56; g.l. = 4; p<0,001). No barco Meu Sonho, os reves-
timentos de melhor desempenho por rank de eficiência K, F e J. No barco Nave Mãe não houve diferen-ça na fixação do mexilhão-dourado entre os diferentes revestimentos testados e o controle (ANOVA, F = 0,27; g.l. = 4; p>0,05). Os re-vestimentos de melhor desempenho por rank de eficiência foram F e K. Analisando o desempenho dos 4 reves-timentos nas 4 embarcações avaliadas, o mais eficiente para evitar a fixação do mexilhão--dourado foi o K.
Conclusões Dentre os 11 revestimentos existentes no mercado brasileiro e testados ao longo do estu-do, aquele que apresentou o melhor desempe-nho para controlar a fixação de L. fortunei em embarcações e estruturas submersas com um menor custo ambiental, foi a tinta Coper Bot-tom 60 do fabricante Sherwin-Willians Tintas Sumaré (código K). Entretanto, devido aos re-sultados observados ao longo do estudo, outros dois revestimentos mostraram-se adequados para o controle do mexilhão-dourado: a tinta Aemme Long Life Plus (código J) e Revran AF LCL 870 (código F) dos fabricantes Promar e Renner Herrman, respectivamente. Esses 3 re-vestimentos mostraram baixa toxicidade e alta eficiência para evitar a incrustação do mexi-lhão-dourado em experimentos de laboratório e no ambiente natural. Um programa de pintura dos cascos das embarcações deveria ser implementado com urgência em áreas críticas de dispersão, como o Mato Grosso, visando o controle desse invasor e evitar que ele se espalhe para novas bacias.
Figura 3. Número médio de pós--larvas de Limnoperna fortunei presentes nos revestimentos anti--incrustantes avaliados em quatro embarcações na bacia do Lago Guaíba.


Seção
10Controle populacionalde bivalves invasores:
métodos físicos


331
A aplicação de métodos físicos para o controle populacional de bivalves invasores possui a vantagem de apresentar menor im-pacto ambiental quando comparada a métodos químicos, por não apresentar contaminantes residuais nos ambientes aquáticos. Entretanto, apresenta como restrição os custos advindos da instalação de sistemas e equipamentos para realizar o controle. Os tipos de métodos físicos com poten-cial de aplicação são bastante variados. Podem estar centrados na propriedade de alguns ma-teriais em dificultar a incrustação dos bivalves, na retenção das espécies adultas em sistemas de filtros, na destruição das espécies jovens por dosagem de radiações, em processos de remo-ção mecânica de espécies incrustantes, entre outros. A escolha do método mais conveniente a ser aplicado depende, entre outros aspectos, do volume e das características físicas e quími-cas da água onde o controle será realizado, das características e dimensões das superfícies que
se deseja proteger de incrustações e dos custos para a instalação dos sistemas e equipamentos necessários para aplicação do método. De forma geral, os métodos físicos podem ser classificados em preventivos e corretivos. Os métodos preventivos visam inibir o desenvolvimento populacional dos bivalves ou inibir sua incrustação em sistemas e equipa-mentos que se deseja proteger. Os métodos corretivos visam permitir a re-moção de bivalves já incrustados em superfícies. Dentre os métodos preventivos merecem destaque aqueles a seguir apresentados.
Sistema de filtração com filtro de areia ou em equipamentos específicos A aplicação de filtros de areia pode ser viável quando a vazão de água bruta a ser tra-tada é baixa, da ordem de até 10 m3 /h por metro quadrado de área filtrante. Nesse caso o material retido pode ser separado por lavagem
CAPÍTULO 34
Controle Físico: conceitos básicos
Manuel Luiz Leite Zurita

Zurita, M.L.L
Controle Físico:conceitos básicos
332
em contracorrente ou por raspagem da super-fície filtrante. Os filtros industriais podem ser aplicados para operar até centenas de m3 /h de água bruta. Cabe salientar que os filtros não retêm os bivalves na forma larval.
Aplicação de radiações ultravioleta
A radiação UV é comumente aplicada para esterilizar ambientes hospitalares e desin-fetar a água potável. A eficiência deste método está direta-mente ligada às características físicas da água bruta. Se esta água apresenta alta turbidez e/ou material sólido em suspensão a eficiência deste método é sensivelmente reduzida. Os bivalves são destruídos pela radia-ção ultravioleta quando a luz penetra através da célula e é absorvida pelo ácido nucleico. A absorção da luz ultravioleta pelo ácido nucleico provoca alterações da informação genética que incapacita a reprodução da célula. As avaliações até o momento realizadas para o controle populacional de bivalves inva-sores indicam que este método pode apresentar bons resultados para a mortalidade de bivalves na forma larval e, dependendo da dose de UV a que os bivalves venham a ser submetidos, até mesmo para espécies adultas. Devemos ressaltar que, apesar das avalia-ções de laboratório usualmente relacionar tempo de exposição à radiação UV, devemos em nossas avaliações sempre relacionar os efeitos observa-dos com a dose que o bivalve ficou submetido. Desta forma, os resultados registrados podem ser comparados com os resultados de outras avalia-ções realizadas por outros pesquisadores. A Dose é a energia UV incidente por uni-dade de área em uma superfície, normalmente expressa na unidade mJ/cm2 ou J/m2. A dose de UV recebida por um organismo aquático em um reator considera os efeitos da intensidade da ra-diação UV (absorção, refração e reflexão) sobre a água e a superfície do reator e a eficácia germici-da dos comprimentos de onda UV transmitidos.
Aplicação de Ultrassom
Este método utiliza ondas sonoras aplica-das à água bruta que geram vibrações conheci-das como cavitação. Essas vibrações provocam
efeitos danosos sobre membranas e valvas dos bivalves jovens e na forma larval, provocando rachaduras. A concha, uma vez danificada, não se desenvolve. Os estudos indicam que, dependendo da frequência da radiação acústica e da dose aplicada, os efeitos podem ser sentidos também por espécies adultas. Para avaliações com a aplicação de ul-trassom valem as mesmas considerações apre-sentadas para as radiações UV em relação à im-portância de relacionar os efeitos observados com a dose incidente.
Proteção catódica
O método de proteção catódica de su-perfícies metálicas contra a corrosão, através da aplicação de corrente elétrica contínua, é um processo utilizado de forma rotineira. Estudos iniciais indicam a possibilidade das superfícies submetidas a esse tipo de pro-teção poderem também inibir a deposição de bivalves invasores nessas superfícies. Atualmente estão sendo desenvolvidas avaliações com aplicação de correntes elétricas com intensidade de até 10 mA/ft2.
Limpeza mecânica Dentre os métodos corretivos merece destaque a limpeza mecânica. Este método é aplicado quando uma su-perfície plana ou uma tubulação recebe a in-crustação por bivalves invasores, impedindo ou dificultando as funções desses equipamentos. Os procedimentos a serem utilizados para realizar essa limpeza dependem das ca-racterísticas da superfície incrustada, das suas dimensões e das condições de acesso às áreas incrustadas.
Os principais métodos são:
a) Dutos de grande diâmetro Para este tipo de duto pode-se aplicar um raspador com o diâmetro ligeiramente inferior ao diâmetro interno do duto, que se desloca pelo interior do duto retirando o material depo-sitado. O material incrustante pode ser retirado do interior do duto pela própria ação do elemen-to raspador ou com o auxílio de fluxo de água.

CAPÍTULO 34
Seção 10
333
b) Superfície imersa em água Este método exige o emprego de mergu-lhadores. Estas pessoas retiram o material in-crustado com o uso de bombas de alta pressão e de instrumentos raspadores.
c) Bomba de alta pressão Este método pode apresentar eficácia no processo de retirada de material incrus-
tante em acessórios de tubulações e dutos de pequeno comprimento. O uso de raspa-dores manuais facilita a retirada do material incrustado. Também podemos aplicar subs-tâncias abrasivas para facilitar o trabalho. Os bivalves invasores retirados das superfícies devem ser imediatamente retirados do local de trabalho, tendo em vista o forte odor que podem exalar.


335
O mexilhão-dourado, Limnoperna fortu-nei (Dunker, 1857), é uma espécie de molusco bivalve invasor. Esse organismo vem se multipli-cando e se dispersando à montante das bacias hidrográficas do Sul, sudeste e centro-oeste do Brasil (Darrigran & Ezcurra-de-Drago, 2000b; Mansur et al., 2004a; Darrigran & Dambore-nea, 2009c). As razões para este crescimento descontrolado estão associadas a sua alta capaci-dade reprodutiva (Morton, 1982), a falta de pre-dadores eficazes (Darrigran & Pastorino, 2003) e a sua capacidade de se dispersar através da in-crustação nos cascos de embarcações (Mansur et al., 2003). A larva do mexilhão-dourado, em todas suas fases, vive livremente no plâncton, se des-locando a favor da corrente (Darrigran & Dam-borenea, 2009c). No momento da captação de água bruta por bombas e turbinas, as larvas po-dem ser sugadas para o interior das tubulações, motores, sistemas de resfriamento, irrigação e de tratamento de água bruta, onde continuam se desenvolvendo. Quando pós-larva ou recruta
(comprimento aproximado de 300 μm), secreta fios de bisso que permitem sua fixação e aglome-ração no substrato. Em sistemas que utilizam es-sas águas, como estações de tratamento de água e unidades hidroelétricas, o mexilhão-dourado vem causando problemas de obstrução em di-versos equipamentos, tais como: tubulações, fil-tros, bombas captadoras de água e trocadores de calor (Figs. 1A e 1B). Ao longo do tempo, torna-se necessária a desativação temporária de parte destes sistemas produtivos para a remoção do molusco e manutenção de maquinários, acar-retando em prejuízos operacionais e financeiros (Darrigran & Ezcurra-de-Drago, 2000b). Devi-do a estes problemas existe necessidade de bus-car técnicas que possam diminuir a quantidade de indivíduos e minimizar os problemas provo-cados pelo mexilhão-dourado. Dentre as metodologias desenvolvidas no controle de espécies invasoras, na busca de tec-nologias limpas e efetivas inclui-se a utilização de desinfecção com radiação ultravioleta (UV). A radiação UV é uma técnica amplamente uti-
CAPÍTULO 35
Controle de Limnoperna fortunei com aplicação de radiação ultravioleta
Cintia Pinheiro dos SantosMaria Teresa Raya Rodriguez
Maria Cristina Dreher MansurMarinei Vilar Nehrke
Manuel Luiz Leite Zurita

Santos et al.
Controle de Limnoperna fortunei com aplicação de radiação ultravioleta
336
lizada para desinfecção de água, com ação bac-tericida e germicida (Wolfe, 1990). Sua ação é letal para diversos organismos, pois atinge di-retamente o material genético (ácidos nucleicos DNA/RNA) das células (Fig. 2). A luz UV pene-tra na célula e é absorvida pelo ácido nucleico, provocando alterações da informação genética e incapacitando a célula para reprodução. Caso a replicação ocorra, as novas células serão mutan-tes e os descendentes incapazes de se duplicar (Friedberg et al., 1995). Diversos estudos comprovaram a inativa-ção de microrganismos, por desinfecção fotoca-talítica, utilizando irradiação UV, com diferentes dosagens e comprovaram sua eficiência (Tab. I). Objetivou-se através deste trabalho o de-
Figura 1. Problemas provocados pelas incrustações do mexilhão-dourado. 1A, entupimento em termelétrica; 1B, incrustação em bomba de captação de água. Fotos: A, R.C.M. Tissot; B, J. Imada.
Figura 2. Efeito da radiação UV sobre a cadeia de DNA, fazen-do com que o filamento do DNA não tenha mais capacidade de encaixe. Modificado de Herring, 2006.
senvolvimento de método para o controle da biomassa na fase larval de L. fortunei com a apli-cação da radiação ultravioleta. Os experimentos foram realizados no la-boratório em unidade piloto (Fig. 3), esta essen-cialmente constituída por: * Dois tanques, em forma cilíndrica, con-feccionados em aço inox, capacidade para 220 litros, fundo cônico, drenos de fundo e tendo externamente um indicador de nível com trena para determinar as vazões utilizadas nas avalia-ções; * Uma bomba centrífuga de ½ CV pro-vida de manômetro para recirculação da água com sistema de fluxo contínuo e válvulas para controle de vazão; * Dois conjuntos de unidades de ultravio-leta, modelo UVNat 7501, com corpo de mate-rial termoplástico, lâmpadas com potência de 75 W e área externa de 910 cm2.
Figura 3. Unidade piloto, onde foram realizados os testes de ex-posição de larvas de Limnoperna fortunei à radiação ultravioleta, Centro de Ecologia, Universidade Federal do Rio Grande do Sul.

CAPÍTULO 35
Seção 10
337
Para o cálculo da dosagem de radiação foram levantadas as características dos reatores de radiação UV e realizados os cálculos de do-sagem com a associação da vazão de água pre-sente no reator e o tempo de exposição. A ação germicida das radiações ultravioleta emitidas pela lâmpada, segundo dados do fabricante do reator, é da ordem de 40% da potência da lâm-pada, ou seja, 30 W. Desta forma, consideran-do que a área externa da lâmpada é de 910 cm2,
a ação germicida aplicada foi de 33 mW/cm2. Para o cálculo do tempo de retenção em segundos levou-se em consideração o volume do líquido na câmara e a vazão, sendo representada pela equação 4,6 L/ Vazão (L/h) x 3600 s/h. A Dosagem de UV para um reator é ex-pressa em mWs/cm2 e seu cálculo contempla o produto da ação germicida (mW/cm2) e o Tem-po de Retenção (s). Desta forma, para a uni-dade piloto instalada, a dosagem em mWs/cm2 pode ser expressa pelas equações: – 547000/Vazão (L/h) para um reator; – 1094 000/Vazão (L/h) para dois reatores. As amostras de larvas foram coletadas no Cais do Porto, delta do Jacuí, município de Porto Alegre, Rio Grande do Sul, Brasil (30°01’31,08’’S; 51°13’38,26’’W), obtidas com auxílio de uma bomba de sucção, através da fil-tragem de 1000L de água, em rede de plâncton com abertura de malha de 30 μm. Amostras de larvas véliger foram quantificadas com o auxí-éliger foram quantificadas com o auxí-liger foram quantificadas com o auxí-í-lio de microscópio estereoscópico. O volume de água utilizado em cada tes-te foi de 145L [1L (CT) + 144L (água bruta ou deionizada)]. As réplicas da concentração teste (CT) foram expostas à radiação ultravio-leta em diversas dosagens no intervalo de 200 a 800 mWs/cm2. Para obtenção das dosagens--teste trabalhou-se com as vazões de 1400, 2000, 2400, 3500 e 4200 L/h o que representa, respectivamente, o tempo de retenção de apro-ximadamente 6, 4, 2,5 e 2 minutos. Realiza-ram-se os experimentos de exposição com dois tipos de água (deionizada e bruta), seis réplicas em cada dosagem e o controle, totalizando 72 amostras. Como controle do experimento, as amostras CT circulavam no sistema na estação piloto com a as mesmas condições dos experi-mentos, mas sem a exposição à radiação UV. Após a exposição à radiação UV nos diferentes tempos e vazões, a água resultante foi refiltrada em rede de plâncton de 30 μm, concentrada a 100 mL e levada ao microscó-ó-pio estereoscópico. Os critérios definidos para a avaliação da letalidade neste trabalho foram a falta de movimentos, a adução valvar e a ausên-cia dos batimentos do estilete cristalino, após estímulo mecânico. Esta avaliação foi realizada logo após o término do experimento. Os resultados encontrados são apresen-tados na Tabela II.
Tabela I. Doses de radiação para inativação de diversos or-ganismos
BACTÉRIAS Dose de UV (mWcm2)
Bacillus anthracis (esporos de Antrax) 46,2
Clostridium tetani 23,1
Sarcina lutea 26,4
Shigella dysenteriae 4,2
Salmonella typhimurium 15,2
Vibrio cholerae 6,5
Escherichia coli 6,6
VÍRUS
Hepatite infecciosa 8,0
Rotavírus As-11 9,9
FUNGOS
Aspergillus niger 330
Rhizopus nigricans 220
PROTOZOÁRIOS E HELMINTOS
Giardia lamblia (Cistos) 100
Entamoeba hystolytica 84
ALGAS
Chlorella vulgaris 22
Microcystis aeruginosa 75
BIVALVES
Dreissena polymorpha (larvas) 12,5
Fonte: Alam et al. (2001), Chalker-Scott et al. (1994), Gon-çalves et al. (2003) e Wright et al. (1997).
Tabela II. Cálculo da dosagem de radiação ultravioleta; mor-talidade larval percentual média em água deionizada (AD) e água bruta (AB) das larvas de Limnoperna fortunei resultante em cada experimento na unidade piloto.
Vazão(L/h)
Tempo(min)
Dosagem(mWs/cm2)
AD(%)
Desv.pad
AB(%)
Desv.pad
1400 06:05 781 99,5 0,56 90,9 5,53
2000 04:15 547 95,1 2,74 68,0 57,17
2400 03:38 456 90,0 2,98 79,1 7,15
3500 02:37 313 88,9 5,92 29,3 3,62
4200 02:05 260 67,7 4,56 53,1 21,37

Santos et al.
Controle de Limnoperna fortunei com aplicação de radiação ultravioleta
338
A maior taxa de mortalidade larval ocorreu a uma dosagem de 781 mWs/cm2. As mortalidades percentuais das larvas de mexi-lhão-dourado, para dosagens em torno de 781 mWs/cm2 tanto com água deionizada (99%) como com água bruta (90%) (Fig. 4), corro-boram com o trabalho de Wright et al. (1997) cuja eficiência da radiação ultravioleta no con-trole de D. polymorpha, em laboratório, atin-giu percentuais semelhantes aos 90%. Uma vez que a exposição de radiação ultravioleta provoca 100% de redução do assentamento de cracas e outras formas de larvas em tubos transparentes por onde circula a água salgada, é pertinente supor que, nos casos de água doce, a eficiência seja maior, pois há uma quantida-de menor de minerais que absorvem a radiação (Chalker-Scott et al., 1994). Por outro lado, as flutuações nos percentuais de mortalidade para água bruta nas dosagens de 260 e 313 mWs/cm2
podem estar associadas às diferenças da quali-dade da água a cada dia de experimento, parâ-metros que poderiam influenciar nas condições dos testes, segundo Gonçalves et al. (2003). O valor de DL50 (limite de confiança: 95%) obtido por análise de regressão foi de
Figura 4. Variação na mortalidade média em água deionizada e água bruta.
324 mWs/cm2, que corresponde à dosagem letal necessária para causar a mortalidade de 50% das larvas de mexilhão-dourado nos tes-tes na unidade piloto. Esta dosagem foi supe-rior às dosagens utilizadas para a inativação de bactérias para desinfecção da água (Lobo et al., 2009). Sendo assim, a dosagem de ra-diação ultravioleta aplicada para eliminar as larvas do mexilhão-dourado, quando aplica-da em tubulações de água potável, poderia também atuar no tratamento da qualidade da água e diminuir os insumos no tratamento tradicional. Os resultados do trabalho demonstraram que a radiação UV apresenta grande eficiência no controle das larvas de mexilhão-dourado, sendo que a mortalidade instantânea das larvas aproximou-se dos 100% nas condições do teste com a dosagem de 781 mWs/cm2 e apresentan-do DL50 de 324 mWs/cm2 . Para um controle de larvas em unidades de maior porte, recomenda-se a implantação de sis-temas que permitam uma maior flexibilidade na potência dos equipamentos emissores de UV para que seu uso possa manter a eficiência pretendida com diferentes qualidades de água bruta.

339
As ondas ultrassônicas são vibrações me-cânicas em um líquido, sólido ou gás. Durante o século XIX foi demonstrado a sensibilidade do ouvido humano, capaz de detectar sons cujas frequências de ondas estejam entre 16 Hz e 21 kHz, aproximadamente (Fig. 1). No início do século XX, foi possível produzir e detectar ondas sonoras com frequência acima do limite audível pelo homem, dando origem ao termo ultrassom (Okuno et al., 1986, Luque-Garcia & Castro, 2003). Os estudos sobre a utilização do ultras-som como método de controle de moluscos teve seu início na década de 70 na União Soviética, para controle do mexilhão-zebra (Dreissena polymorpha) reduzindo o assentamento do mo-lusco no caso particular dos sistemas de refri-geração (Donskoy & Ludyanskiy, 1995), porém é uma técnica muito mais antiga. O ultrassom foi descoberto por Curie em 1880, ao estudar o efeito piezelétrico. Já em 1894, Thornycroft & Barnaby observaram que na propulsão de mís-seis lançados pelo destróier, uma fonte de vi-
bração era gerada causando implosão de bolhas e ou cavidades na água (Suslick, 1989; 1994). Essa vibração ficou conhecida como cavitação. O ultrassom por possuir propriedades de degradação da matéria orgânica, ação mi-crobiocida e também dispersante (Gonze et al., 1999), contribui formando compostos de bai-xo peso molecular. Classicamente, a literatura demonstra que a onda ultrassônica contínua, primariamente aquece o tecido, determinando os efeitos térmicos. Já a onda ultrassônica pul-sada é utilizada para atuar no nível celular e/ou molecular, principalmente através da alteração da permeabilidade da membrana, gradientes de concentração iônica e atividade bioquími-ca celular (Mendonça et al., 2006; Ter Harr, 2007). O ultrassom também possui um meca-nismo químico, responsável pela formação de radicais livres e peróxido de hidrogênio (En-tezari et al., 2004). Diversos autores relatam a eficácia do ultrassom na redução de mesófilos em água (Jyoti & Pandit, 2004; Seymour et al., 2002), na redução de fungos filamentosos
CAPÍTULO 36
Controle de bivalves com a utilização do ultrassom
Cintia Pinheiro dos SantosJuliane Vicenzi
Felipe Amorim BeruttiMaria Cristina Dreher Mansur
Carlos Pérez BergmannMaria Teresa Raya Rodriguez
Marinei Vilar NehrkeManuel Luiz Leite Zurita

Santos et al.
Controle de bivalves com a utilização do ultrassom
340
e leveduras em água, frutas e outros vegetais (Idrissi et al., 1996; Radel et al., 2000; Nasci-mento et al., 2005). Em espécies de larvas de moluscos, Ese-erco (1992) descreve que a energia acústica na escala de 39 a 45 kHz fragmenta a larva do me-xilhão-zebra num intervalo de alguns segundos, em fluxo de água. Este experimento também foi eficiente para alguns adultos no intervalo de 19 a 24 horas. Donskoy & Ludyanskiy (1995), realizaram estudos, em laboratório, utilizando ultrassom para o controle do mexilhão-zebra. Os experimentos demonstraram que sons de baixa frequência em torno de 200 Hz foram efetivos no controle dos aglomerados de me-xilhão-zebra. Os melhores resultados encon-trados estão no uso integrado do som com a vibração. Esta aplicação inviabilizou e destruiu as larvas do mexilhão-zebra. Estudos realiza-dos pela Ontário Hydro demonstraram que a vibração, aplicada na prevenção da fixação dos mexilhões juvenis e adultos sobre estruturas da usina, foi efetiva (Kowalewski et al., 1993). Experimentos laboratoriais utilizando o ultrassom como forma de controle do mexi-lhão-zebra, Dreissena polymorpha, também fo-ram realizados com sucesso por Breitig (1961).
No estágio larval deste molusco, este au-tor obteve sucesso com 3 segundos, 1 minuto e 3 minutos de ultrassom sob 22 kHz e 800 kHz. O ultrassom exerce efeitos de pressão mecânica sobre as valvas das pequenas larvas, provocando rachaduras. A concha, uma vez danificada, não se desenvolve. Porém existe a necessidade de avaliar os demais componentes do plâncton, para verificar se estes não sofrem prejuízos. O Centro de Ecologia e o Laboratório de Materiais Cerâmicos da Escola de Engenharia da Universidade Federal do Rio Grande do Sul desenvolveram estudos sobre o controle físico de larvas de moluscos invasores com ondas ul-trassônicas. Foram realizados testes em banca-da, conforme metodologia que segue. Os testes foram realizados com amostras de zooplânc-ton e de larvas de Corbicula fluminea (Müller, 1774).
Teste em Bancada
Os testes em bancada foram realizados colocando-se as larvas, coletadas no Lago Gua-íba (Porto Alegre/RS), num volume determina-do de água destilada e submetendo-as às on-das ultrassônicas, utilizando um equipamento gerador de ultrassom que permite variar a fre-quência e a potência aplicada de ondas de ul-trassom (Fig. 2), permitindo, por conseguinte, variar a dose aplicada de ultrassom. O conceito de dose aplicada de ultrassom para uma dada frequência, utilizado neste estudo, é apresenta-do pela relação:
Dose Aplicada (Kws/L) = Potência (Kw)x tempo de permanência (s)/ Volume (L)
Para efeitos de comparação, observou--se o mesmo tempo de exposição ao ultrassom na análise de ensaios conduzidos com frequên-cias diferentes (20, 100, 200, 400 e 800 kHz). Observou-se que nas faixas de frequências tes-tadas há uma diminuição acentuada do efeito da exposição ao ultrassom na mortalidade das larvas com o aumento da frequência. Este re-sultado mostrou que a frequência de 20 kHz é mais eficiente na mortalidade larval que as ou-tras frequências nas condições ensaiadas. Esses resultados corroboram os estudos de Holm et
Figura 1. Região de frequência do som.
Figura 2. Detalhes do experimento de bancada

CAPÍTULO 36
Seção 10
341
al. (2008) com zooplâncton, que recomendam a utilização do ultrassom para o tratamento da água de lastro. A Figura 3 apresenta os resulta-dos obtidos do percentual de mortalidade para o tempo máximo de 240 segundos em todas as frequências estudadas. A dose aplicada para esta situação foi da ordem de 44 Kws/L. Para as larvas de Corbicula fluminea a dose aplicada de ultrassom de 44 Kws/L tam-bém apresentou 100% de mortalidade. A via-bilidade dos organismos foi avaliada através da observação dos mesmos em microscópio este-reoscópico. Os testes realizados indicam que a mor-
Figura 3. Mortalidade de larvas de Limnoperna fortunei em função da frequência do ultrassom para um tempo de 240 segun-dos, equivalente a uma dose aplicada de 44 Kws/L.
talidade de larvas de L. fortunei tende a 100% quando sobre elas aplicamos doses de ultras-som superiores a 10 kws/L para a frequência de 20 kHz, conforme indica a Figura 4. Os testes realizados com a técnica de aplicação de ultrassom indicam condições fa-voráveis como metodologia para o controle de larvas de espécies invasoras, em especial em dutos transportadores de águas contaminadas pela presença dessas larvas. A evolução e o aprimoramento dessa técnica, utilizando uni-dades piloto, permitirá definir os parâmetros para o desenvolvimento de projetos em escala industrial.
Figura 4. Mortalidade registrada em relação à dose aplicada de ultrassom para frequência de 20 kHz.


Seção
11Controle populacionalde bivalves invasores:
métodos biológicos


345
O controle biológico, por definição, é o uso de organismos vivos para reduzir a den-sidade populacional ou impacto de um orga-nismo-praga específico, tornando este menos abundante ou menos danoso (Eilenberg et al., 2001). Também é definido como a redução dos efeitos adversos de uma espécie pela atividade de outra (Van Lenteren, 2008). Este processo ocorre normalmente na natureza, e é um dos principais mecanismos de manutenção do equi-líbrio das densidades populacionais, que tam-bém são reguladas pela competição, escassez de alimento e fatores ambientais. Os asiáticos foram pioneiros no uso de inimigos naturais para controle de insetos-pra-ga. As formigas e joaninhas foram os primeiros agentes de controle biológico a serem utiliza-dos. Na Figura 1, uma linha do tempo ilustra a evolução do uso de agentes de controle biológi-co no mundo. Os inimigos naturais, quando empre-gados de forma estratégica no controle de organismos-praga, recebem a denominação de
Agente de Controle Biológico (ACB). Existem três categorias de ACBs: 1) Predadores: organismos que usam ou-tros como alimento (Raffel et al., 2008). No controle populacional de moluscos podemos citar o uso de Pomacea haustrum Reeve, 1856, na predação de desovas de Biomphalaria glabra-ta Say, 1818, Biomphalaria straminea (Dunker, 1848) e Biomphalaria tenagophila d’Orbigny, 1835 (Guimarães et al., 1983a), em experimen-tos de laboratório. Redução de 50,5%, 24,5% e 22,5% das desovas das respectivas espécies ocorreram devido à predação pelo ampularí-deo. Outro exemplo investigado por Guima-rães et al. (1983b) foi o uso da sanguessuga Helobdella triserialis lineata Blanchard, 1949, como predador de B. glabrata (Guimarães et al., 1983b). A ação deste anelídeo hirudíneo foi eficaz na predação de indivíduos recém eclodi-dos, jovens e adultos B. glabrata de até 10 mm, em condições de laboratório. 2) Patógenos: organismos que causam doenças em outros organismos (Chaube & Pu-
CAPÍTULO 37
Controle Biológico: conceitos básicos
Isabel Cristina Padula PazDaniel Pereira

Paz & Pereira
Controle Biológico: conceitos básicos
346
dhir, 2005). Genthner et al. (1997) testaram a linhagem 2771 de Bacillus alvei, em adultos de D. polymorpha (mexilhão-zebra). Os moluscos tratados com a bactéria apresentaram vacuoliza-ção no epitélio do tubo digestivo em 24 horas. Molloy & Mayer (2007) testaram 10 linhagens de Pseudomonas fluorescens para controle de Dreissena polymorpha e D. rostriformis bugen-sis. Os pesquisadores verificaram que a linha-gem Pf-CL145A de P. fluorescens causou mais de 90% de mortalidade em D. polymorpha. 3) Parasitas: qualquer organismo que viva
dentro ou sobre outro organismo e que depen-da totalmente ou em parte de seus nutrientes, comumente exibindo algum grau de adaptação estrutural, e causando algum grau de dano ao seu hospedeiro (Price, 1977). Nematódeos têm sido testados no controle de lesmas terrestres (Kaya, 2000-2001). O nematódeo Phasma-rhabditis hermaphrodita (A. Schneider, 1859) é utilizado como agente de controle biológico comercial de lesmas da família Limacidae, na Inglaterra (Morley & Morritt, 2006). Como uma estratégia de manejo de or-ganismos-praga, o controle biológico se ocupa da ação de parasitas, patógenos ou predadores, na redução das populações abaixo dos limiares de dano econômico. A Figura 2 apresenta um modelo de variação temporal de populações de espécies invasoras antes e após a introdu-ção de um ACB: na fase inicial, denominada fase de repouso, a presença da espécie invasora muitas vezes não é notada, sendo constatada, geralmente, somente na fase de pânico, quan-do a espécie se reproduz desenfreadamente, ocupando vários compartimentos e nichos do ecossistema. Após esta fase, inimigos naturais e fatores limitantes começam atuar sobre a po-pulação invasora, desencadeando uma nova di-nâmica populacional; a partir daí ocorre certo equilíbrio, embora a invasora continue causan-do impactos de ordem econômica e ambiental. Nesta fase, a aplicação de métodos de controle biológico visa diminuir a população invasora abaixo do limiar de dano econômico, mas é quase impossível uma redução em nível da fase inicial de colonização. O uso de ACBs no manejo de organis-mos-praga pode ser didaticamente dividido em quatro tipos (Fig. 3). A Tabela I sumariza as principais vanta-gens e desvantagens do uso de agentes de con-trole biológico (ACB) e agentes químicos (AQ). A fraca correlação entre resultados de controle de pragas obtidos em laboratório e em campo, quando usados ACB, geralmente é fru-to de problemas que ocorrem antes e durante a aplicação no ambiente alvo. A ausência de cuidado com as condições ambientais e a falta da elaboração de estraté-gias de inoculação adequadas ao hábito da praga surgem como os principais limitantes ao sucesso na inoculação de ACB. Além disso,
Figura 1. Evolução histórica do uso de inimigos naturais no controle biológico de pragas (adaptado de Daane & Mills, 2009).

CAPÍTULO 37
Seção 11
347
após a liberação ocorrem diversas interações na natureza que podem interferir no processo de introdução e estabelecimento do ACB em uma determinada área. Por esse motivo, é primaz que na elabo-ração de um programa de controle biológico sejam realizados ensaios em vários níveis ex-
perimentais (bancada, piloto e campo), e que todas as exigências para o estabelecimento do ACB sejam respeitadas, para que o controle biológico não perca força e o sucesso seja al-cançado. Cabe ressaltar que praticamente todos os estudos com controle biológico, advêm da
Figura 2. Dinâmica populacional da espécie-praga antes e após a intro-dução de um agente de biocontrole (adaptado de Darrigran & Dambore-nea, 2009b). Agente de Controle Bio-lógico (ACB).
Figura 3. Tipos de controle biológico, conforme Eilenberg et al. (2001). Figura gentilmente cedida por Roderick, extraído de Roderick & Navajas (2003).

Paz & Pereira
Controle Biológico: conceitos básicos
348
Tabela I. Vantagens e desvantagens do uso de agentes de controle biológico (ACB) e químico (AQ). Adaptado de Alves et al. (1999).
Agente Vantagens Desvantagens
ACB
1. Automultiplicação e dispersão do ACB
2. Especificidade e seletividade ao organis-mo alvo
3. Permite uso associado com outras técni-cas de controle de pragas
4. Permite melhoramento genético conven-cional e via transgenia, no caso de agentes microbianos
5. Riscos menores de poluição e de toxicida-de aos organismos não alvos
6. Baixo risco de desenvolvimento de resis-tência nas populações de pragas.
1. Espectro de ação limitado ao organismo alvo ou aos or-ganismos relacionados
2. Requer mais tempo para o controle efetivo da praga
3. Requer condições ambientais adequadas para introdu-ção e estabelecimento na área de aplicação
4. Necessita estratégias de aplicação compatíveis com habi-tat e características da praga alvo
5. Necessidade de armazenamento adequado à espécie de ACB, por se tratar de organismos vivos
AQ
1. Amplo espectro de ação
2. Ação rápida e eficaz
3. Não dependente de fatores ambientais
4. Necessidade de poucos cuidados com ar-mazenamento
1. Necessidade de reaplicação a cada ressurgência da praga
2. Baixa seletividade
3. Alto risco de poluição ambiental e de impacto sobre or-ganismos não alvo, muitas vezes seus resíduos persistem em diferentes níveis da cadeia trófica
4. Alto risco de seleção de populações resistentes
agronomia, devido à importância das pragas e doenças nos sistemas agrícolas, e da necessida-de da busca de alternativas aos métodos quími-cos amplamente utilizados atualmente nesses sistemas, os quais são onerosos tanto ao bolso do produtor, quanto ao ambiente. Dentro da grande área “Controle Bio-lógico” existe um gradiente no número de estudos de acordo com o tipo de pragas-alvo, como demonstrado na Figura 4, montada após o levantamento de artigos publicados em duas bases de dados, Web of Science e Scopus. Nesse levantamento foi possível verificar que o me-nor número de artigos publicados se refere ao controle biológico de moluscos, e a maior parte dos mesmos objetivam o controle de molus-cos vetores de doenças a humanos, seguido de moluscos-praga agrícolas (Daoust et al., 2010; Eskelson et al., 2011). Em ambientes aquáticos, o caso de uso de controle biológico de maior sucesso se refe-re ao uso da bactéria Bacillus thuringiensis no controle de larvas de mosquitos. Essa bactéria é aplicada em diversos locais no mundo e seu uso no Brasil é preconizado pelo Ministério da Saúde (Alves & Lopes, 2008), visto os mos-
quitos serem vetores de diversas doenças. No Brasil, em 2002, a aplicação de B. thuringiensis sv. israelensis para controle de larvas de Aedes aegypti, juntamente com outras políticas, leva-ram a redução de 73,3 % dos casos de dengue (Alves & Lopes, 2008). Apesar de estratégias de controle mi-crobiano de moluscos incrustadores do gênero
Figura 4. Gradiente e distribuição de publicações referentes ao controle biológico de diferentes grupos de pragas.

CAPÍTULO 37
Seção 11
349
Dreissena já terem sido testadas em laboratório, com o uso de algumas espécies de Bacillus (Singer et al., 1997), Aeromonas e Shewanella (Gu & Mitchell, 2001), o único registro de aplicação em campo de ACB para o controle de moluscos incrustadores é com o uso da bactéria Pseudomonas fluorescens CL145A para contro-le de Dreissena polymorpha e D. rostriformis bugensis, a qual já foi patenteada nos Estados Unidos e Canadá (Molloy & Mayer, 2007). O projeto “Monitoramento e controle do mexilhão-dourado Limnoperna fortunei
(Dunker, 1857), em rios e reservatórios brasi-leiros” financiado por FURNAS e coordenado por Maria Tereza Raya Rodriguez é pioneiro na busca de agentes de controle microbiano para o manejo do mexilhão-dourado. Não foram encontrados registros de uso de predadores e parasitas como agentes de bio-controle do mexilhão-dourado, L. fortunei, en-tretanto, existe referência sobre a predação do mexilhão-dourado (Silveira, 2010) e mexilhão--zebra por peixes e aves (Crosier & Molloy, 2007).


351
O controle microbiano é uma subdivisão do controle biológico, que se baseia na capa-cidade de microrganismos causarem doenças em organismos-pragas e/ou invasores. Fungos e bactérias são os inimigos naturais mais utili-zados em programas de controle populacional que usam agentes microbianos, mas nematoi-des e vírus também são utilizados. O uso de microrganismos patogênicos é amplamente difundido na agricultura, no controle de microrganismos causadores de do-enças de plantas, nematoides e, principalmen-te, de insetos (Alves, 1998; Melo & Azevedo, 2000a, b; Alves & Lopes, 2008), sendo em al-guns patossistemas a única opção disponível. Atualmente, na área de saúde pública, o uso de bactérias patogênicas no controle da população de larvas de mosquitos vetores de doenças é uma realidade, sendo seu uso preconizado pelo Ministério da Saúde (Alves & Lopes, 2008). Há quase cinco mil anos doenças de in-setos, como abelha e bicho-da-seda, já eram conhecidas pelos egípcios e chineses. A primei-ra aplicação de um vírus para controle de uma
praga florestal foi feita pelo inglês J. Evellyn há mais de trezentos anos (Alves & Lopes, 2008). Apesar da grande importância dos mo-luscos como espécies-praga, estudos visando o controle microbiano dos mesmos, sejam os de importância médica, agrícola ou ambiental são pouco comuns, sendo que a maioria das pes-quisas busca predadores e competidores dos moluscos-praga (Brasil, 2007). Mesmo após Madsen (1990) ter suposto que os micropara-sitas seriam as formas mais efetivas de controle de moluscos, e hoje, passados quase 25 anos, estudos usando microrganismos para controle de moluscos ainda são escassos. Publicações referentes ao controle mi-crobiano de moluscos em nível de bancada de laboratório se restringem a experimentos com bactérias (Genthner et al., 1997; Singer et al., 1997; Oliveira et al., 2004; Halima et al., 2006; Ali et al., 2010; Pereira et al., 2011b), com poucos estudos usando microrganismos formulados e disponíveis comercialmente (Os-man & Mohamed, 1991; Mayer, 2009; Osman et al., 2011), como sumarizado nas Tabelas I e II.
CAPÍTULO 38
Controle Microbiano
Isabel Cristina Padula PazDaniel Pereira
Andressa Moraes Sofia de Souza

Paz et al.
Controle Microbiano
352
Bioensaios visando à seleção de agentes microbianos para o controle do mexilhão-dou-rado, Limnoperna fortunei (Dunker, 1857), foram realizadas por Pereira et al. (2011b). Os autores testaram uma cepa de Bacillus thurin-
giensis sv. israelensis e uma de Bacillus thurin-giensis sv. kurstaki. Os melhores resultados fo-ram obtidos por meio da primeira cepa (72% de mortalidade corrigida na concentração de 107 UFC.ml-1; CL50 7,66 x 106).
Tabela I. Agentes de controle biológico (ACB) avaliados por diversos autores no controle de espécies de bivalves invasores ou vetores de doenças.
Espécie Alvo ACB Modo de ação
Mortalidade corrigida
(%)
LC50(UFC.ml-1)
CT Autor
Dreissena polymorpha
Bacillus alvei 2771
Vacuolização no epitélio do tubo digestivo em 24 horas
- - - Genthner et al. (1997)
Biomphalaria glabrata
Bacilus alvei 2771 C
- - 6,3x102♠ - Singer et al. (1997)
Biomphalaria glabrata
Bacillus brevis SS86-4 C
- - 6,03 x 102♠ - Singer et al. (1997)
Biomphalaria glabrata
Bacillus circulans 42G1 C
- - 5,13x102♠ - Singer et al. (1997)
Biomphalaria glabrata
Brevibacillus laterosporus NRS 1111
- 100♣94♦
0,05 mg.ml-1 Oliveira et al. (2004)
Dreissena polymorpha
Bacilus alvei 2771 C
- - 3,01 x 102♥9,33 x 103♠5,25 x 104•
Singer et al. (1997)
Dreissena polymorpha
Bacilus brevis SS86-4 C
- - 7,94x10♠ - Singer et al. (1997)
Dreissena polymorpha
Bacilus circulans 42G1 C
- - 9,33 x 10♥1 x 107•
- Singer et al. (1997)
Dreissena polymorpha
Bacillus circulans 42G1 P
- - 2 x 108* - Singer et al. (1997)
Dreissena polymorpha
Brevibacillus laterosporus 1647 P
- - SA♥♠2x108*
- Singer et al. (1997)
Dreissena polymorpha
Aeromonas media - 100 - 107 UFC.ml-1 Gu e Mitchell (2001)
Dreissena polymorpha
Aeromonas veronii - 100 - 107 UFC.ml-1 Gu e Mitchell (2001)
Dreissena polymorpha
Aeromonas salmonicida - 54,5 - 107 UFC.ml-1 Gu e Mitchell (2001)
Dreissena polymorpha
Shewanella putrefaciens - 100 - 107 UFC.ml-1 Gu e Mitchell (2001)
Dreissena polymorpha
Pseudomonas fluorescensPf-CL145A
Destruição do sistema digestivo
>90 - - Molloy & Mayer (2007)
Dreissena rostriformis
bugensis
Pseudomonas fluorescensPf-CL145A
Destruição do sistema digestivo
>90 - - Molloy & Mayer (2007)
Limnoperna fortunei
Bacillus thuringiensis sv. israelensis IIRAC 22
- 72 7,66 x 106 107 UFC.ml-1 Pereira et al. (2011)
Limnoperna fortunei
Bacillus thuringiensis sv. kurstaki HD-1
- 62,5 8,65 x 106 107 UFC.ml-1 Pereira et al. (2011)
Oncomelania hupensis
Bacillus thuringiensis 66
- 47,5 - - Halima et al. (2006)
Oncomelania hupensis
B. thuringiensis sv. israelensis 977 e NRRL HD-522
(Proteína solúvel)
- 70 - 0,0009 mg.ml-1
Ali et al. (2010)
SA (sem atividade); ♣ 0 a 3 dias após a eclosão; ♦ 12-15 dias após a eclosão; ♥ 8-12 mm de comprimento; ♠2-5 mm de compri-mento; • véliger estágio D; * véliger pré-estágio D.

CAPÍTULO 38
Seção 11
353
Espécies de bactérias dos gêneros Bacillus e Pseudomonas representam a grande maioria dos agentes microbianos testados no contro-le de moluscos-praga, assim como ocorre no controle de insetos-praga e fitopatógenos (Paz, 2009; Lanna Filho et al., 2010). Isso se deve à ampla gama de metabólitos produzidos pe-las espécies destes gêneros de bactérias, o que permite que as mesmas possuam atividade so-bre diversos hospedeiros. Além de haver um grande conhecimento adquirido sobre os me-canismos de ação envolvidos nos processos de
representantes de diferentes níveis tróficos do meio aquático (algas, invertebrados e peixes), assim como em mamíferos e outros organismos pertinentes. Após os testes, na quinta etapa, o produto comercial é formulado, podendo ser na forma líquida ou sólida (pastilhas solúveis em água, granulado, pó, etc.). Na sexta etapa o microrganismo, transformado em um produto formulado, é reavaliado quanto à ecotoxicida-à ecotoxicida- ecotoxicida-de por meio de organismos não-alvo. São reco-mendados testes com organismos representantes de diferentes níveis tróficos do meio aquático
Tabela II. Agentes de controle biológico (ACB) utilizados comercialmente no controle de espécies de invertebrados invasores ou vetores de doenças.
Espécie Alvo ACB Mortalidade (%)
CL50(mg.ml-1) Produto Autor
Biomphalaria alexandrina Bacillus thuringiensis - 0,5 Thuricide 16000 U.I. Osman & Mohamed (1991)
Biomphalaria alexandrina
Bacillus thuringiensis sv. kurstaki - 392,31 Dipel 2x Osman et al. (2011)
Biomphalaria truncatus Bacillus thuringiensis - 0,4 Thuricide 16000 U.I. Osman & Mohamed (1991)
Dreissena polymorpha
Pseudomonas fluorescens 92,7 - Zequanox Mayer (2009)
patogênese e morte dos hospedeiros por esses microrganismos, geralmente o uso dos mesmos em altas densidades é de baixo impacto ao am-biente e ao homem. Com relação ao controle de bivalves in-crustadores, as pesquisas são ainda mais escas-sas e restringem-se basicamente ao controle do mexilhão-zebra Dreissena polymorpha e do mexilhão quaga Dreissena rostriformis bugensis na América do Norte (Genthner et al., 1997; Singer et al., 1997; Molloy, 1998; Oliveira et al., 2004; Halima et al., 2006; Ali et al., 2010), e em escala de laboratório, do mexilhão-doura-do, L. fortunei (Pereira et al., 2011b). O processo de seleção de agentes micro-bianos para o controle efetivo de uma praga-al-vo é muito longo. As fases envolvidas desde o isolamento do agente até sua certificação e per-missão de uso em campo são descritas a seguir e ilustradas na Figura 1. A primeira etapa envol-ve a coleta de diferentes tipos de substratos que abrigam microbiota diversificada com poten-cial para a seleção de agentes microbianos de controle. Os substratos amostrados geralmente são procedentes do mesmo habitat da espécie praga. Os microrganismos destes substratos são isolados e purificados, dando a origem a uma
bacterioteca e/ou micoteca. Após esta etapa, os microrganismos são caracterizados com base em critérios taxonômicos, por meio da obser-vação macroscópica da morfologia de colônias cultivadas em meio de cultura a base de Agar e nutrientes, em técnicas de microscopia óptica (coloração de Gram) e provas bioquímicas (Fig. 2). Também são utilizados testes moleculares para a determinação taxonômica mais segura dos isolados. Uma terceira etapa compreen-de os bioensaios de bancada e campo, com os organismos-alvo, visando à seleção de agentes de controle eficazes. Geralmente é necessária a avaliação de um grande número de isolados até a obtenção de um que acumule características de interesse, como mortalidade da espécie-alvo e baixa ecotoxicidade (Mayer, 2009). No tra-balho de Molloy & Mayer (2007), cerca de 700 isolados bacterianos oriundos de solo foram avaliados até a seleção de Pseudomonas fluo-rescens Pf-CL145A, efetiva contra os mexilhões zebra e quaga, e inócuo a organismos não-alvo. Na quarta etapa o microrganismo, no caso da ausência de informações sobre sua inocuidade, é avaliado por meio de testes ecotoxicológicos, em relação a seus efeitos sobre organismos não--alvo. São recomendados testes com organismos

Paz et al.
Controle Microbiano
354
(algas, invertebrados e peixes), assim como em mamíferos e outros organismos pertinentes, o que depende do modo de ação e método de apli-cação do agente de biocontrole. Por último, o produto comercial recebe o registro dos órgãos competentes, após ter sido constatada baixa ou média toxicidade aos organismos não-alvo, re-cebendo assim permissão para comercialização. A partir daí, o produto pode ser aplicado em campo pelos profissionais que realizarão o con-trole populacional da espécie-alvo. Atualmente, os testes ecotoxicológicos são realizados com base no produto formulado (testes ecotoxicológicos da FASE II), mas se recomenda a avaliação prévia da toxicidade dos isolados se-
lecionados anteriormente a etapa de formulação do produto (testes ecotoxicológicos da FASE I). Desta forma, são evitados custos adicionais na formulação de produtos que podem ser rejeitados na etapa de registro. No caso de produtos formu-lados com base em micro-organismos conhecidos quanto à reduzida toxicidade, os testes ecotoxico-lógicos da FASE I podem ser dispensados. Embora os agentes de controle biológi-co sejam geralmente patógenos naturais das espécies-alvo, a aplicação desses organismos de maneira antrópica gera, também, preocupação com a segurança ambiental, pois a utilização desses patógenos no ambiente tem por objetivo causar danos somente à espécie-alvo (Oliveira-
Figura 1. Processo de seleção de agentes microbianos para o controle efetivo de uma praga-alvo.

CAPÍTULO 38
Seção 11
355
Figura 2. Isolamento e caracterização de microrganismos isolados de diferentes substratos: 2A e 2B, Isolamento; 2C, bacté-ria purificada em cultura; 2D, coleção de bactérias (bacterioteca); 2E, cartela de tipos de células bacterianas; 2F, cartela de tipologia de colônias; 2G, Identificação bacteriana usando aspectos fisiológicos e nutricionais, pelo uso de placas de BIOLOG (Foto: V. Duarte); 2H, gel de agarose mostrando bandas relativas a um fragmento do gene rDNA 16S bacteriano usado para sequenciamento e identificação de bactérias.
-Filho, 2008). A fase crítica, do ponto de vista ambiental, no desenvolvimento de um produto a base de agentes microbianos, é a certificação de que o micro-organismo não apresente efei-tos danosos sobre organismos não-alvo. A cer-
tificação é concedida ou não após a avaliação de estudos ecotoxicológicos pelos órgãos regu-lamentadores. Os estudos ecotoxicológicos do produto formulados são realizados em quatro fases (Fig. 3) (Melo & Azevedo, 2000b).

Paz et al.
Controle Microbiano
356
No Brasil, o registro de produtos a base de agentes de controle biológico são avaliados sobre três aspectos: eficiência no controle do organismo-alvo realizado pelo Ministério da Agricultura (MAPA); efeitos sobre a saúde hu-mana, realizado pela Agência Nacional de Vi-gilância Sanitária (ANVISA) do Ministério da Saúde; e, quanto à periculosidade ambiental do produto, avaliado pelo Instituto Brasileiro de Meio Ambiente e dos Recursos Naturais (IBA-MA), do Ministério do Meio Ambiente. Os moluscicidas com base em agentes de biocontrole apresentam uma alternativa ao con-trole de moluscos-praga, já que os métodos tradi-cionais de controle químico liberam uma série de subprodutos tóxicos ao meio ambiente e apresen-tam amplo espectro de ação.
Figura 3. Fases dos estudos eco-toxicológicos utilizados na avalia-ção necessária para o registro do produto comercial.
O amplo conhecimento adquirido pela comunidade científica, principalmente no âm-bito agrícola e de saúde pública, sobre a efeti-vidade e modo de ação de agentes microbianos no controle de insetos, nematoides e fitopatóge-nos, assim como sobre os níveis de toxicidade destes agentes a organismos não-alvo, servem de suporte para o fortalecimento de linhas de pesquisa direcionadas ao controle biológico de moluscos. Baseado nos dados apresentados na Tabe-la I, cinco espécies do gênero Bacillus, uma de
Brevibacillus, três de Aeromonas e uma de Pseu-domonas apresentam potencial no controle po-pulacional de moluscos. Bacillus thuringiensis, um dos agentes de biocontrole mais usados no mundo, apresenta potencial para o controle do mexilhão-dourado, L. fortunei e de O. hulepense, molusco transmissor da esquitossomose. Dentre as espécies de Aeromonas deve-se considerar: A. salmonicida é um conhecido patógeno de peixes (Sharp & Secombes, 1993), sendo assim seu uso poderia afetar a ictiofauna; A. veronii também é patógeno de peixe (Rahman et al., 2002), além de patógeno humano (Gröbner et al., 2007). Já, quanto à espécie A. media, não foram encontrados registros de patogenicidade. Pseudomonas fluorescens, agora descrita como patógena ao mexilhão-zebra, é uma bactéria
cosmopolita que apresenta atividade biocon-troladora de fitopatógenos (Gnanamangai & Ponmurugan, 2012). Para a consolidação do uso de agentes microbianos no controle de moluscos límnicos é necessário o conhecimento do modo de ação dos patógenos sobre as pragas-alvo; da toxici-dade das dosagens recomendadas sobre organis-mos aquáticos não-alvo; a prospecção de outras espécies e isolados bacterianos e fúngicos; além do aprimoramento dos protocolos de avaliação ecotoxicológica de produtos formulados.

357
Introdução
Na América do Sul, a partir da última década, começou a ser registrada a presença do molusco invasor Limnoperna fortunei (Dunker, 1857) (Bivalvia; Mytilidae) conhecido como mexilhão-dourado. Este bivalve encontrou nas bacias hidrográficas sul-americanas condições favoráveis para dispersão e crescimento, colo-nizando diversos sistemas de drenagem (Darri-gran, 2002; Mansur et al., 1999, 2003). O primeiro registro da ocorrência de L. fortunei na Lagoa Mirim foi em 2005 (Lango-ne, 2005). No mesmo ano foi registrada a pre-sença da espécie no Canal São Gonçalo, bacia de drenagem da Lagoa Mirim, e na estação eco-lógica do Taim (Burns et al., 2006a, b). Assim como o bivalve corbícula [Corbicula fluminea (Müller, 1774)], que foi registrada no Guaíba em 1978 (Mansur et al., 2004c), e no Taim em 1991 (Mansur et al., 1991), L. fortunei foi re-gistrado no Lago Guaíba em 1998 e levou cerca de 10 anos para transpor a Barragem Eclusa da
Lagoa dos Patos, que separa a Lagoa dos Patos da Lagoa Mirim, e invadir e se expandir por toda a região.
O cenário das alterações tróficas
Entre os inúmeros impactos causados pelo mexilhão-dourado, as alterações na cadeia trófica parecem ser de particular importância, uma vez que a atividade de filtragem destes or-ganismos leva a mudanças nas taxas de sedi-mentação, ciclagem de nutrientes, abundância e composição do seston (Cataldo et al., 2005b; Sylvester et al., 2005). Por outro lado, os maio-res níveis tróficos parecem se beneficiar dos recursos alimentares adicionais, representados pelas larvas planctônicas e adultos bentônicos do mexilhão-dourado (Montalto et al., 1999; Ferriz et al., 2000; Penchaszadeh et al., 2000; García & Protogino, 2005; Boltovskoy et al., 2006; Sylvester et al., 2007b). É relatado por Darrigran (2002) que a elevada capacidade re-produtiva e a ausência de predadores naturais
CAPÍTULO 39
Predadores potenciais para o controle do mexilhão-dourado
Michelle Lopes João Vieira

Lopes & Vieira
Predadores potenciais para o controle do mexilhão-dourado
358
favorecem a formação de grandes aglomerados populacionais. Desta forma, esse molusco cau-sa alterações na abundância relativa de muitos grupos do zooplâncton e pode gerar modifica-ções potenciais na trama trófica (Brugnoli et al., 2005). Uma vez que, altas densidades de L. fortunei certamente representam uma abun-dante oferta de alimento para os peixes, em particular para aqueles que apresentam elevada plasticidade alimentar (Silva, G. et al., 2005), é esperado que este recurso seja incorporado na trama trófica do sistema. Embora a erradicação de espécies aquá-ticas invasoras seja uma tarefa difícil, espera--se que o consumo intenso por certas espécies possa ter um papel importante no controle do mexilhão-dourado (Oliveira, C. et al., 2010, citando Vitule et al., 2009). Em ambientes neo-tropicais, L. fortunei foi prontamente incorpo-rada como item alimentar de diversas espécies de peixes demersais1, em particular por aque-las que apresentam uma elevada plasticidade alimentar (Montalto et al., 1999; Ferriz et al., 2000; Penchaszadeh et al., 2000; García & Protogino, 2005; Boltovskoy et al., 2006; Syl-vester et al., 2007b). No sistema Patos-Mirim, por exemplo, o mexilhão-dourado foi registra-do como um dos itens de maior importância, e que foi rapidamente incorporado na dieta do pintado Pimelodus pintado (Baptista & Zibetti, 2006; Lopes, 2010), que é um peixe omnívoro e que possui uma estratégia alimentar genera-lista-oportunista2.
Predadores potenciais
A diversidade de peixes do sistema Patos--Mirim é alta (Malabarba, 1989, Vieira et al., 2010). Dentre as mais de cinquenta (50) espé-cies de peixes registradas para a Lagoa Mirim e Canal São Gonçalo (Garcia et al., 2006; Burns, 2010; Ceni, 2010), trinta e uma (31) se desta-cam por sua abundância ou presença sistemáti-ca, tanto na zona rasa como na zona profunda (Tab. I). Um estudo com dezenove (19) destas es-pécies, coletadas na Lagoa Mirim e na região profunda (3-6 m) do Canal São Gonçalo – um elemento de ligação entre as Lagoas dos Patos e Mirim –, demonstrou a presença de mexi-lhão-dourado em 12 das 19 espécies, as quais
apresentaram uma variação de frequência de ocorrência (FO) entre 1,4% nos exemplares de cascuda (Loricariichthys anus) até 96% em exemplares de joaninha (Crenicichla punctata) (Tab. I). Dentre os Characiformes, duas espécies se destacaram como predadores de mexilhão--dourado. Em primeiro, encontra-se o grande lambari omnívoro oportunista Astyanax aff. fasciatus, que pode atingir até 160 mm CT (comprimento total), e que apresentou cerca de 70% de seus estômagos com mexilhão-doura-do. Em segundo, a traíra Hoplias aff. malaba-ricus que apresentou 20% dos seus estômagos com mexilhão-dourado, e que representa o mais importante recurso pesqueiro na região. Diversas outras espécies de Characiformes, embora amplamente distribuídas na região, não apresentaram mexilhões-dourado adultos no trato digestório. No entanto, não é possível descartar a hipótese do uso de larvas de me-xilhão-dourado como alimento pelos juvenis e adultos da maioria dos pequenos lambaris. Os Clupeiformes, amplamente distribuídos na La-goa Mirim e Canal São Gonçalo têm uma ali-mentação zooplantófaga3, e assim como os pe-quenos Characiformes provavelmente também ingerem as larvas de mexilhão-dourado como alimento. Dentre os Siluriformes e os Perciformes, que compartilham modificações morfológicas específicas à vida demersal, alimentando-se no fundo, tanto em zonas rasas como profundas, encontram-se a grande maioria das espécies de grande porte exploradas pela pesca da região. Garcia & Montalto (2006) observaram que os peixes que predam eficientemente esses molus-cos invasores, geralmente têm bocas adaptadas para sucção ou possuem dentes incisivos fortes ou molariformes, podendo ainda ter os dentes faringeanos com diferentes graus de desen-volvimento. Os altos índices de frequência de ocorrência (Tab. I) de mexilhão-dourado ob-servados para espécies como joaninha (Creni-cichla punctata), pintado (Pimelodus pintado), Cará (Geophagus brasiliensis), e de raros exem-plares de corvina (Micropogonias furnieri) que ainda são capazes de cruzar a barragem eclusa do Canal São Gonçalo, demonstraram a ampla capacidade, por parte destes peixes, de modifi-carem suas dietas, diante das alterações do am-
1 Demersais: animais aquáticos que, apesar de terem capacidade de natação ativa, vivem a maior parte do tempo em associação com o substrato.2 Generalista-oportunista. Generalista: comportamento alimentar de alguns organismos que consomem uma grande variedade de itens em pequenas quantidades. Oportunista: organismos que consomem os itens mais disponíveis no ambiente.3 Zooplantófaga: alimentação baseada em zooplâncton, diminutos animais flutuantes sendo muitas vezes arrastados pelo fluxo de água.

CAPÍTULO 39
Seção 11
359
Tabela I. Lista de espécies de peixes (Ordem, Família e Espécie) mais frequentes (quadrado hachurado) ou presentes (X) nas zonas rasas (RASA) e profundas (PROF) do Canal São Gonçalo e Lagoa Mirim, RS. ALI FO = frequência de ocorrência de mexilhão-dourado nos estôma-gos analisados (N/A = espécies cujos estômagos não foram analisados); PESCA = (*) se importante na pesca da região; SG = São Gonçalo e MI = Lagoa Mirim; COMP. MAX = comprimento máximo observado nas amostras. Fonte: Burns, 2010; Ceni, 2010 e Lopes, 2010.
AU PES RASA PROF COMP.
Espécie FO% CA SG MI SG MI MAX
Characiformes
Characidade Astyanax aff. fasciatus (Cuvier, 1819) 72,2 * X 146-165
Characidade Cheirodon ibicuhiensis Eigenmann, 1915 N/A X X 53-91
Characidade Cyanocharax alburnos (Hensel, 1870) N/A X X 74-94
Characidade Cheirodon interruptus (Jenyns, 1842) N/A X X 40-61
Characidade Astyanax eigenmanniorum (Cope, 1894) N/A X X X 130-130
Characidade Charax stenopterus (Cope, 1894) 0,0 X X X X 125-126
Characidade Oligosarcus robustus Menezes, 1969 0,0 * X X X X 234-274
Characidade Oligosarcus jenynsili (Günther, 1864) 0,0 * X X X X 240-331
Erythrinidae Hoplias aff. malabaricus (Bloch, 1794) 20,0 * X X X 550-640
Curimatidae Cyphocharax voga Hensel, 1870 2,1 X X X X 274-292
Siluriformes
Pimelodidae Parapimelodus nigribarbis (Boulenger, 1889) 0,0 X X 246-306
Pimelodidae Pimelodus pintado Azpelicueta, Lundberg & Loureiro, 2008
52,4 * X X 446-556
Loricariidae Loricariichthys anus (Valenciennes, 1836) 1,4 * X 412-615
Loricariidae Hypostomus commersoni Valenciennes, 1836 2,4 * X X X 615-631
Loricariidae Rineloricaria microlepidogaster (Regan, 1904) 55,6 X X X 260-265
Loricariidae Rineloricaria longicauda Reis, 1983 0,0 X X 185-225
Loricariidae Rineloricaria strigilata (Hensel, 1868) 13,8 X X 250-298
Heptapteridae Pimelodella australis Eigenmann, 1917 0,0 X X X 162-180
Aspredinidae Pseudobunocephalus iheringii (Boulenger, 1891) N/A X X X 73-106
Auchenipteridae Trachelyopterus lucenai Bertoletti, Pezzi da Silva & Pereira, 1995
0,0 X X 224-261
Heptapteridae Rhamdia aff. quelen Quoy & Gaimard, 1824 31,0 * X X X 700-922
Clupeiformes
Clupeidae Platanichthys platana (Regan, 1917) N/A X X 106-117
Clupeidae Lycengraulis grossidens (Agassiz, 1829) N/A X X X 261-286

Lopes & Vieira
Predadores potenciais para o controle do mexilhão-dourado
360
4 Vágil: organismos que se locomovem livres no ambiente.
Tabela I. Continua...
AU PES RASA PROF COMP.
Espécie FO% CA SG MI SG MI MAX
Perciformes
Sciaenidae Micropogonias furnieri (Desmarest, 1823) 41,7 * X 118-258
Cichlidae Geophagus brasiliensis (Quoy & Gaimard, 1824) 42,1 * X X X X 235-240
Cichlidae Crenicichia punctata Hen-sel, 1870 96,3 * X X 280-360
Atheriniformes
Atherinopsidae Odontesthes aff. perugiae (Evermann & Kendall, 1906)
* 220-250
Atherinopsidae Odontesthes argentinensis (Valenciennes, 1835) * 102-255
Atherinopsidae Odontesthes bonariensis (Valenciennes, 1835) * 145-250
Atherinopsidae Odontesthes humensis de Buen, 1953 * 220-250
Atherinopsidae Odontesthes mirinensis Bemvenuti, 1995 * 200-240
biente, revelando-se potenciais predadores de mexilhão-dourado. Este fato já foi previamente observado para P. pintado, que rapidamente incorporou a espécie invasora como um novo recurso alimentar no Canal São Gonçalo (Bap-tista & Zibetti, 2006; Lopes, 2010). Dentre a Família Loricariidae observou--se um contraste. Peixes com hábito bentônico e detritívoros, como as espécies de cascuda Lo-ricariichthys anus e Hypostomus commersoni, apresentaram baixas frequências de ocorrência de mexilhão-dourado, em contraste com cascu-das da espécie Rineloricaria strigilata e Rinelori-caria miocrolepdogaster que apresentaram 13% e 55% de frequência de ocorrência, respectiva-mente (Tab. I). No entanto, a ingestão do mexi-lhão-dourado não implica necessariamente uma estratégia bem sucedida para o predador, como evidenciado pela alta incidência de conchas não digeridas e intactas no trato digestório. A pre-sença de moluscos intactos em várias espécies de peixes sugere que os moluscos não podem ser digeridos e, provavelmente, passam pelo interior do peixe ainda vivos (Oliveira, C. et al., 2010).
Predação seletiva
O comprimento total de concha dos espé-cimes de mexilhão-dourado capturados no am-biente variou entre 4 mm e 32 mm com uma dis-tribuição de tamanho unimodal com moda na
classe de 10 mm. (Fig. 1A). Os comprimentos de concha de L. fortunei encontrados nos tratos di-gestórios variaram entre 3 e 15 mm nas espécies analisadas (Fig. 1A). A diferença na distribuição de tamanho dos indivíduos nos tratos digestó-rios e no ambiente natural, por si só sugere uma predação seletiva sobre os indivíduos menores. Independente do tamanho e do compor-tamento de forrageamento do predador, apenas os indivíduos de L. fortunei menores que 14 mm em média, são predados. A incidência de indivíduos de L. fortunei menores que 14 mm na dieta das espécies detritívoras raspadoras de fundo, como as cascudas R. microlepdogaster e R. strigilata, as quais não apresentam caracterís-ticas adaptadas à predação de moluscos, confir-ma a hipótese levantada por Lopes (2010). Tal hipótese sugere que, os indivíduos de mexilhão--dourado com até 14 mm CT apresentam um comportamento vágil4 (Uryu et al., 1996), mo-vendo-se sobre o fundo lamoso do Canal São Gonçalo, estando mais disponíveis à predação dos peixes, do que os indivíduos maiores que 14 mm que tendem a se aglomerar ou se esconder em reentrâncias do substrato. Os indivíduos vá-geis, mais disponíveis, seriam “coletados” aci-dentalmente pelos detritívoros. Evidências mostram que L. fortunei é predado em seus diferentes estágios de desen-volvimento, por várias espécies de peixes da região neotropical (Montalto et al., 1999), mas

CAPÍTULO 39
Seção 11
361
os mexilhões pequenos (<15 mm) são mais fre-quentes e abundantes no trato digestório destes peixes. As médias de comprimento de concha de L. fortunei encontrados nos tratos digestó-rios dos peixes analisados no sistema Patos-Mi-rim, apresentaram valores inferiores a 14 mm, independente do comprimento total dos preda-dores (Fig. 1), confirmando os dados já descri-tos para o pintado (P. pintado) neste sistema (Lopes, 2010). Observações feitas em predado-res de grande porte como o armado (Pterodo-ras granulosus, 170-550 mm CT), encontraram resultados semelhantes, onde L. fortunei foi encontrado em uma faixa de tamanho de 8-17 mm de comprimento médio (Cantanhêde et al., 2008), apesar de o mexilhão ao longo do seu desenvolvimento crescer até mais de 30 mm (Boltovskoy & Cataldo, 1999; Maroñas et al., 2003, Mansur et al., 2008b).
Figura 1. A) Frequência de tamanho de L. fortunei coletados no ambiente (AMBIENTE) e no conteúdo estomacal (ALIMEN-TAÇÃO) dos peixes (vide lista Figura 1B) no Canal São Gonça-lo, RS. B) Distribuição do comprimento de concha (cm) de L. fortunei nos diferentes predadores coletados no Canal São Gonçalo, RS. Os códigos dos peixes são: Rineloricaria stri-gilata RINSTR (12,2-21,0 cm CT), Pimelodus pintado PIMPIN (6,4-33,3 cm CT), Rhamdia aff. quelen RHAQUE (16,0-45,4 cm CT), Hoplias malabaricus HOPMAL (14,6-38,2 cm CT), Rinelo-ricaria microlepdogaster RINMIC (15,5-28,0 cm CT), Astyanax fasciatus ASTFAS (12,0-16,5 cm CT), Hypostomus commersoni HYPCOM (12,5-50,2 cm CT) e Crenicichla punctata CREPUN (12,8-26,0 cm CT).
Aspectos do Controle Biológico
Desde a invasão de L. fortunei na Améri-ca do Sul, mudanças na dieta das espécies tem sido observadas, principalmente em peixes om-nívoros oportunistas, que passaram de uma die-ta de baixa qualidade baseada em plantas, para uma dieta rica e energética dominada por esses moluscos (Montalto et al., 1999; Ferriz et al., 2000, Garcia & Protogino, 2005; Boltovskoy et al., 2006). Isto se confirma com as espécies observadas neste estudo, as quais apresentam hábitos generalistas e omnívoros, consumindo de forma oportunista os itens disponíveis em maior quantidade no ambiente (Fig. 2). As características morfofuncionais do apa-rato trófico dos peixes condicionam o gradien-te de digestão de L. fortunei (Montalto et al., 1999), como verificado para os predadores de mexilhão-dourado no estuário do Rio da Prata, que apresentaram em sua maioria, aparatos bu-cais adaptados, desde pequenas placas mandibu-lares a dentes incisivos e dentição faringeana em vários graus de desenvolvimento (Garcia & Pro-togino, 2005). A grande quantidade de conchas trituradas presentes nos tratos digestórios do lambari (A. fasciatus), do cará (G. brasiliensis), da joaninha (C. punctata) e da corvina (M. furnie-ri), refletem a presença de aparatos bucais desen-volvidos nestas espécies (Bemvenuti & Moresco, 2005; Hartz et al., 1996), os quais permitiram quebrar facilmente a frágil concha de L. fortunei. Semelhanças funcionais do mexilhão--dourado com o mexilhão-zebra (Dreissena polymorpha) sugerem fortemente que altas densidades de L. fortunei podem ativamente transferir grandes quantidades de matéria or-gânica da coluna d’água aos domínios bentôni-cos, através da filtração e da formação de fezes e pseudofezes (Karatayev et al., 2007b), que por sua vez aumenta a densidade de invertebra-dos bentônicos (Botts et al., 1996; Darrigran, 2002). Estudos realizados no Rio Paraná e no Rio da Prata na Argentina, e mais recentemente no reservatório de Itaipu, revelam que algumas das espécies comercialmente mais importantes, como o armado (Pterodoras granulosus) e a pia-va (Leporinus obtusidens), se alimentam prefe-rencialmente de L. fortunei em frequências de até 100% (Penchaszadeh et al., 2000; Cataldo et al., 2002b; Oliveira, C. et al., 2010).

Lopes & Vieira
Predadores potenciais para o controle do mexilhão-dourado
362
Figura 3. Potenciais predadores de mexilhão-dourado no sistema Patos-Mirim, RS. A) Crenicichla punctata (JOANINHA), B) Geophagus brasiliensis (CARÁ), C) Hoplias malabaricus (TRAÍRA), D) Astyanax fascinatus (LAMBARI), E) Pimelodus pintado (PINTADO), F) Rineloricaria strigilata, G) Rhamdia aff. quelen ( JUNDIÁ), H) Rineloricaria microlepdogaster (CASCUDA).
Figura 2. Distribuição do IIR (relati-vizado a 100%) dos itens encontra-dos no trato digestório dos peixes coletados no Canal São Gonçalo, RS. Os códigos dos peixes são: Charax stenopterus (CHASTE), Oligosarcus robustus (OLIROB), Parapimelodus nigribarbis(PARNIG), Pimelodella australis (PIMAUS), Rineloricaria longicauda (RINLON), Trachelyopterus lucenai (TRALUC), Cyphocharax voga (CYPVOG), Hypostomus commersoni (HYPCOM), Rineloricaria strigilata (RINSTR), Rhamdia aff. quelen (RHAQUE), Hoplias malabaricus (HOPMAL), Micropogonias furnieri (MICFUR), Pimelodus pintado (PIMPIN), Geophagus brasiliensis (GEOBRA), Rineloricaria microlepdogaster (RINMIC), Astyanax fasciatus (ASTFAS), Crenicichla punctata (CREPUN).

CAPÍTULO 39
Seção 11
363
Os efeitos de L. fortunei sobre as comu-nidades de peixes não estão restritos às espécies que consomem o molusco diretamente (Fig. 3), em particular aqueles que podem separar e es-magar as conchas, mas também se estendem às espécies que podem se beneficiar deste novo recurso indiretamente, incluindo muito dos grandes exemplares, e comercialmente valio-sas espécies que se alimentam de outros peixes que por sua vez predaram o mexilhão-dourado (Penchaszadeh et al., 2000; Boltovskoy et al., 2006). Desta forma, tanto as espécies piscí-voras, como a traíra (H. malabaricus) e o jun-diá (Rhamdia. aff. quelem), quanto espécies omnívoras generalistas-oportunistas, como o lambari (A. fasciatus) e o pintado (P. pintado) podem ser afetadas de forma positiva pela pre-sença de L. fortunei. No entanto, impactos ne-gativos sobre a ictiofauna também podem ser
esperados, como os observadas neste estudo em alguns exemplares de cascudas das espécies R. microlepdogaster e R. strigilata, que apresen-taram deformidades na região anal, causadas por conchas não digeridas de L. fortunei, tor-nando estas espécies suscetíveis a patógenos e infecções. É importante ressaltar que independen-te do tamanho e do comportamento de forra-geamento do predador, apenas os indivíduos de L. fortunei menores que 14 mm em média, são predados. Neste contexto, futuras investi-gações a respeito destas complexas interações podem contribuir na avaliação dos impactos do mexilhão-dourado sobre os peixes e a trama trófica da Lagoa Mirim e Canal São Gonçalo, porém não existem evidências de campo, nem na bibliografia, que os peixes possam controlar esta espécie invasora.


12Seção
Modelos de análise de risco e previsão de
cenários de dispersão de moluscos invasores


367
Os moluscos representam um grupo, dentro de uma longa lista de organismos inva-sores que vêm causando impactos ambientais, sociais e econômicos em todo o mundo (Di Castri, 1989; Carlton & Geller, 1993; Jones et al., 1997; Carlton, 1999; Mack et al., 2000). A introdução destas espécies, na maioria dos ca-sos, tem uma forte correlação com a atividade humana e, em especial, com a expansão e glo-balização do comércio. A navegação marítima, por exemplo, já resultou no translado de um grande número de espécies de plantas e ani-mais de um lugar a outro do planeta, de forma intencional ou acidental (Di Castri, 1989). Des-de o século XV, ela tem sido um meio de trans-porte não só de pessoas, mas também de ou-tros mamíferos, pássaros, plantas e organismos acompanhantes como artrópodes, moluscos e outros invertebrados (Carlton, 1999). Assim, chegaram às Américas o gado, o rato domés-tico e tantos outros animais e plantas exóticos que hoje são comuns em nosso ambiente. Os impactos ambientais causados pela
introdução de espécies invasoras são, na maio-ria das vezes, de difícil percepção, principal-mente pelos seus efeitos tardios. Este fato pode levar à errônea e perigosa ideia de que esses or-ganismos já se integraram ao ecossistema onde se instalaram. Esta crença fatalista induz ao pensamento de que as invasões biológicas são um fato consumado e que já seria tarde para fazer algo. Isto, somado aos benefícios econô-micos que algumas espécies invasoras têm ge-rado a determinados setores que as exploram, se traduziu em uma falta de interesse em tra-balhos de prevenção, controle ou erradicação. Exceções parecem estar associadas aos casos nos quais estas espécies causam prejuízos eco-nômicos significativos a outros setores, como é o caso do molusco invasor Limnoperna fortu-nei (Dunker, 1857) que tem causado grandes problemas ao setor energético brasileiro (Dar-rigran et al., 2007). Em todos os casos, os esforços de erra-dicação são dispendiosos e muitas vezes inefi-cazes, especialmente quando se trata de espé-
CAPÍTULO 40
Análise de risco de introdução de moluscos
aquáticos invasores
Carlos Eduardo Belz Juliana Brasiliense Sebastiany

Belz & Sebastiany
Análise de risco de introdução de moluscos aquáticos invasores
368
cies aquáticas (Cowie, 2004). Na água, vários problemas se somam, como a facilidade de dis-persão de ovos e larvas pela correnteza, a difi-culdade para visualizar a espécie e sua área de ocorrência e as dificuldades de coleta. Assim, a melhor opção é a prevenção. Por mais compli-cado e demorado que possa parecer, prevenir é o melhor caminho. Isto pode significar o impe-dimento da introdução de uma espécie em uma região, ou pelo menos, o adiamento deste even-to, o que gera economia de gastos e um tempo maior para busca de soluções. Porém, para que um trabalho de prevenção seja eficaz é neces-sário um bom conhecimento sobre a espécie e sobre o ambiente, além de ferramentas capazes de analisar estas informações e permitir sua in-terpretação. Um melhor entendimento do processo de bioinvasão pode levar a muitos benefícios, tais como: uma previsão mais apurada dos ní-veis e direção da dispersão, uma concentração de esforços em áreas mais críticas e um maior conhecimento dos mecanismos de dispersão e da biologia das espécies invasoras, o que traz informações relevantes também, para futuras invasões (Johnson & Padilla, 1996). Neste sen-tido, vários trabalhos têm sido publicados com o objetivo de desenvolver um sistema eficaz de prevenção, gerando um novo campo de pesqui-sas, o da ecologia das bioinvasões (Keller et al., 2007; Lockwood et al., 2007).
Conceitos
Dentro do entendimento dos processos ecológicos envolvidos com as bioinvasões, dois temas de grande importância são a predição e a análise de risco. Estes dois termos têm tido distintas interpretações por parte dos pesqui-sadores, o que acaba gerando uma série de dúvidas conceituais. A predição é “a geração de dados sobre a probabilidade de ocorrência de um evento de bioinvasão”. Três critérios são considerados de suma importância para o desenvolvimento de sistemas de predição: 1) serem abertos para revisões e adições de no-vos dados; 2) terem uma estrutura lógica que permita a inclusão de fatores independentes, importantes no processo de invasão; e 3) per-mitir sua replicabilidade. Já o risco é “o pro-duto da probabilidade de ocorrência de algum
evento e suas consequências” (NRC, 2002). Se-gundo Guilam (1996), a análise de risco pode ser definida como a identificação de danos potenciais e suas consequências, usando dados anteriores, observação sistemática, análise estatística e experimentação ou intuição. Estes dados anteriores são obtidos com os estudos de predição. A análise pode ter desde uma abordagem mais orientada no sentido da quantificação do risco, tentando excluir fatores subjetivos ou julgamentos individuais, até uma aborda-gem mais subjetiva, levando em consideração experiências e observações empíricas. Porém, o ideal é a sobreposição destes dois enfoques, criando uma interação proveitosa entre as ciên-cias quantitativa e qualitativa, já que o ambien-te não se comporta de forma linear como as ciências exatas.
Métodos para a predição e a análise de risco
Não é por acaso que existem poucos trabalhos de pesquisa na área da prevenção de bioinvasões, já que é um campo ainda reple-to de incertezas e com um grande número de variáveis a serem analisadas onde, em muitos casos, é difícil estabelecer as correlações entre estas variáveis e seus possíveis resultados. As dificuldades em estabelecer uma aná-lise estatística confiável somam-se ao fato de haver uma responsabilidade e um risco muito grande para o pesquisador ao tentar extrapolar estes dados para a realidade. Estas incertezas também geram uma falta de credibilidade nos resultados alcançados. Uma metodologia muito importante de pesquisa para trabalhos de predição e análi-se de risco é a análise dos vetores de disper-são. Carlton & Ruiz (2004) a consideram a peça chave na prevenção de novas invasões e sugerem um modelo conceitual para carac-terizar estes vetores. Este modelo envolve a identificação e análise dos seguintes pontos: “causa” (acidental ou intencional); “rotas” (os caminhos pelo qual a espécie é transpor-tada – corredores de bioinvasão); “vetores” (as formas pelas quais a espécie é transpor-tada); “tempo associado ao vetor” (como os vetores operam no tempo em termos de fre-

CAPÍTULO 40
Seção 12
369
quência, duração e período); “biota associa-da ao vetor” (descrição da biota associada ao vetor em termos de diversidade, densida-de e condição) e “força do vetor” (número de invasões estabelecidas por meio do ve-tor em uma região geográfica). Esta análise pode ser complexa e trabalhosa, ainda mais quando se trata de ambientes tão distintos como os encontrados na América do Sul. Po-rém, trata-se de um caminho promissor no sentido de evitar novas introduções. No Brasil, são escassos os estudos sobre vetores de dispersão de moluscos invasores, estando a maioria das pesquisas voltadas para descrição geográfica da área de ocorrência e as-pectos da biologia populacional. Um método de grande importância para entender como esses vetores interagem no ambiente é a modelagem. Porém, ela tam-bém apresenta grandes limitações. A tradição histórica da modelagem na Ecologia é marca-da pela perseguição de um ideal representado pelos modelos da Física. Não é de se surpreen-der, dada a relativa simplicidade dos modelos físicos, seu histórico mais antigo, e seu incom-parável poder de predição e generalização. Po-rém, na Ecologia, a história tem sido um pouco diferente. Sistemas ecológicos não são como os sistemas estudados pelos físicos. Os mode-los lineares, por não se aplicarem a muitas das situações não lineares e complexas observadas na natureza, têm falhado em demonstrar dinâ-micas realistas e em confirmar a existência de regras gerais. Neste sentido, historicamente, o realismo biológico foi deixado de lado em troca da conveniência matemática (Giacomini, 2007). Perspectivas melhores têm sido alcança-das nas últimas décadas com o uso de novos conceitos em modelagem como as Redes Neu-rais Artificiais (Özesmi et al., 2006), o Sistema de Informações Geográficas (SIG) (Haltuch et al., 2000), a análise de paisagens e os Mode-los Baseados no Indivíduo (MBI) (Railsback, 2001), dando um caráter mais sistêmico aos modelos.
Estudo de caso
Neste tópico é apresentado um estudo de caso que sugere uma metodologia de análise
de risco para introdução do molusco invasor L. fortunei em reservatórios de usinas hidrelétri-cas. O estudo exemplifica a criação de um ín-dice de vulnerabilidade ao mexilhão-dourado, chamado aqui de IVMD. O IVMD, criado para avaliação da vul-nerabilidade de um reservatório suscetível a re-ceber e desenvolver uma população desta espé-cie, tem uma estrutura múltipla, integrada por seis blocos. Os primeiros quatro blocos têm um ca-ráter básico e quantificam uma condição ou circunstância de um determinado reservatório com relação à vulnerabilidade acima citada. O quinto bloco é, na realidade, um parâmetro de ajuste, que incorpora ao IVMD o risco direto de contaminação (RD) e o sexto bloco é com-posto por uma série de variáveis excludentes (VE) que podem apresentar valores extremos que atuem como fatores determinantes de pri-meira ordem, ao estabelecer a vulnerabilidade de um reservatório diante do mexilhão-doura-do. Esta lista de parâmetros que formam estes blocos não é fixa, sendo que novos pa-râmetros podem ser integrados, dependendo do nível de conhecimento da espécie e das ca-racterísticas específicas de cada região onde o método for empregado. A seguir são descritos mais detalhada-mente os componentes que podem fazer parte destes seis blocos.
Bloco 1 – Características Ecológicas do Reservatório (CER) Este bloco refere-se aos aspectos limno-lógicos do reservatório, que podem apresen-tar um papel de maior ou menor importância diante da pressão colonizadora e expansiva do mexilhão-dourado. Neste sentido, podem ser selecionadas as seguintes variáveis: * Estabilidade do volume de água: que está em função do Tempo de Residência da Água e da Oscilação do Nível da Água; * Regime térmico; * Índice de Estado Trófico; * Concentração de cálcio; * pH da água; * Morfologia do reservatório; * Presença de espécies aquáticas exóticas; * Presença de moluscos invasores.

Belz & Sebastiany
Análise de risco de introdução de moluscos aquáticos invasores
370
Bloco 2 – Vetores de Dispersão (VD) Refere-se a aspectos vinculados aos ve-tores de dispersão que atuam no reservatório, tais como a prática da pesca, a navegação re-creativa, o transporte de areia, a piscicultura e o transporte fluvial (Belz, 2006). Este bloco pode ser quantificado da seguinte forma: * Acessibilidade ao corpo d’água; * Navegabilidade; * Intensidade da pesca recreativa; * Intensidade do transporte fluvial de pessoas e mercadorias; * Presença de hidrovias e eclusas; * Intensidade de comercializaçãode areia; * Intensidade de comercialização de matrizes e alevinos de peixes.
Bloco 3 – Condições Geográficas da Bacia Hidrográfica (CGBH)Estas variáveis informam da possibilidade de dispersão que pode ter o molusco até um deter-minado reservatório, em função de suas con-dições geográficas como localização, conexão com a bacia hidrográfica, proximidade de pon-tos já contaminados, etc. e pode ser representa-do pelas seguintes variáveis: * Conectividade; * Tipo de reservatório: artificial ou natural; * Distância a focos de contaminação pelo mexilhão-dourado.
Bloco 4 – Efeitos Socioeconômicos e Ecológicos (ESE) Este grupo de variáveis pretende medir o grau de transcendência social, econômica e ecoló-gica que pode ser alcançada pela presença do me-xilhão-dourado num determinado reservatório. Estão relacionadas aos usos não recreativos e seu valor de conservação natural, seja pelos habitats representados ou pelas espécies presentes (biodi-versidade) e pode incluir as seguintes variáveis: * Usos da água; * Número de captações de água; * Efeitos ecológicos: que pode ser quantificado com base em indicadores ambientais.
Bloco 5 – Risco Direto de Contaminação (RD) Trata-se do risco extremo que um reser-vatório apresenta em função de se encontrar
à jusante e conectado com outros reservató-rios ou rios onde o mexilhão-dourado já está presente. Neste caso, a aplicação do IVMD ao reservatório inferior deveria resultar num máximo valor de vulnerabilidade, independen-temente dos valores alcançados pelos outros componentes do índice. Com o objetivo de que a incorporação de tal parâmetro ao índice não o distorça, naqueles casos em que não seja aplicável, sua quantifica-ção pode ser planejada como um termo inde-pendente, de acordo com a seguinte expressão: RD = K(100-SBB) Onde: K é um coeficiente entre 0 e 1, de acordo com suposições predefinidas; SBB é o somatório dos quatro blocos bá-sicos que compõem o IVMD; 100 é um valor constante que corresponde ao valor máximo alcançável pelo somatório.
Bloco 6 – Variáveis Excludentes As variáveis que podem configurar este bloco são as seguintes: * Temperatura da água (VT); * Concentração de cálcio (VCc); * pH da água (VpH); * Salinidade (VSal). O bloco de variáveis excludentes (VE) formula-se como um produto de todas as vari-áveis que nele intervém: VE = VT x VCc x VpH x VSal A incorporação das variáveis excludentes ao IVMD deve ser de forma condicionante ao resultado final do índice, com independência do valor do resto das variáveis. No caso, por exem-plo, de um reservatório apresentar temperatura acima da tolerável pela espécie, o valor de VT seria zero, consequentemente, VE também seria zero, o que traria um IVMD nulo para o reser-vatório estudado, indicando que este ambiente não é vulnerável à introdução da espécie.
Organização e interpretação
Dada a natureza de cada variável, bem como suas diferentes unidades e intervalos de

CAPÍTULO 40
Seção 12
371
medida que influem no IVMD, pode-se optar por distribuir todos os valores possíveis de tais variáveis em relação a uma escala comum. Ou seja, uma padronização que incorpora as pon-derações e facilita o planejamento do índice, além de resultar num valor global. No caso de RD (risco direto) e VE (variáveis excludentes) estes podem sofrer uma padronização distinta, já que atuam de forma independente das de-mais variáveis. Segue uma exemplificação de como as variáveis que compõem o IVMD podem ser pa-dronizadas:
Variáveis (Blocos 1 a 4) Para as variáveis que compõem estes blocos, se pode adotar uma padronização de escala de 0 a 5 como a exemplificada na (Tab. I) para a variável “Tempo de Residência da Água”, onde quanto maior o tempo de resi-dência em dias, maior o risco de permanência de larvas da espécie no reservatório, conse-quentemente, maior o risco de contaminação. Então, por exemplo, para um tempo de resi-dência maior que 295 dias, o peso seria o má-ximo (cinco).
Risco direto de contaminação
Este parâmetro pode ser quantificado admitindo-se quatro valores possíveis ao coe-ficiente que o determina: K=1, casos em que existe, com toda a certeza, mexilhão-dourado em reservatórios à montante e que estão co-nectados com o objeto de estudo; K=0,8, exis-te uma suspeita fundada de que pode haver a espécie em reservatórios à montante e que, estão conectados com o que é objeto de estu-do; K=0,5, existem indícios da existência de mexilhão-dourado em reservatórios à montan-te e que, estão conectados com o que é objeto de estudo; K=0, sabe-se com certeza que não existe a espécie em reservatórios à montante ou nos que estão conectados ao que é objeto de estudo.
Variáveis excludentes
De acordo com as referências sobre tole-rância da espécie diante das variáveis selecio-nadas, podem ser estabelecidas escalas, como a exemplificada na (Tab. II), onde um reservató-rio com valores extremos de temperatura não permitiria a sobrevivência de larvas e adultos da espécie, gerando um valor de VT=0 e con-sequentemente, anulando os efeitos de todas as outras variáveis.
Tabela I. Exemplo de padronização para a variável Tempo de Residência da Água (TRA). Valor máximo (VMáx) e valor mínimo (VMín).
Variável VMáx VMínPesos
1 2 3 4 5
TRA (Dias) 1 5 <5 5 a 100 100 a 195 195 a 290 >295
Tabela II. Valores limites (mínimos e máximos) de temperatu-ra da água (T) para a sobrevivência de Limnoperna fortunei.
T(ºC) Valor VT
<-2 ou >40 0
<5 ou >35 0,8
5 a 35 1,0
O IVMD foi planejado primeiramente como um somatório dos 4 blocos básicos, de-finidos cada um deles com um peso próprio (CER + VD + CGBH + ESEE). Sobre tal so-matório, aplica-se, com caráter aditivo, o parâ-metro que mede o risco direto de contamina-ção (RD) e, com caráter multiplicativo, o grupo de variáveis excludentes (VE). Para cada grupo de variáveis ainda pode ser estipulado um peso, de acordo com a rele-vância de cada bloco e para que o índice varie de 0 (nula vulnerabilidade) a 100 pontos (má-xima vulnerabilidade). A forma como estas variáveis podem in-teragir para gerar o índice depende do grau de complexidade que se deseje estabelecer. Como sugestão, segue uma simplificação que pode ser aplicada aos dados, gerando a seguinte equa-ção:
Uma vez organizado o IVMD, passa-se para a etapa de interpretação dos dados iden-tificados. Tal interpretação pode e deve ser abordada a partir de dois enfoques. O primeiro refere-se ao valor obtido dos blocos 1 a 3, que resume a vulnerabilidade de cada reservatório,
IVMD=[(CER+VD+CGBH+ESEE)+RD]xVE

Belz & Sebastiany
Análise de risco de introdução de moluscos aquáticos invasores
372
incluindo suas aptidões para receber o mexilhão--dourado e o segundo enfoque é referente ao po-tencial de gravidade dos efeitos de sua presença (Bloco 4). Desta forma é possível reconhecer, além do valor global do índice, qual ou quais são os aspectos e componentes de maior importância no resultado final, como na (Fig. 1).
Conclusões
Embora seja um assunto discutido há muitos anos e ultimamente esteja fazendo parte do discurso de ambientalistas no mundo todo, o que se vê, na prática, é que pouca importância tem se dado ao assunto bioinvasões. Muitas pes-quisas têm direcionado seu foco para a biologia das espécies e o monitoramento de sua disper-são e, pouca ênfase tem sido dada aos estudos de prevenção. O trabalho de Carlton & Ruiz (2004), que discute o conceito e aplicação da ciência dos vetores, e o de Arim et al. (2006), que discute a teoria da invasão, indicam que o caminho mais sensato a seguir passa por uma análise profunda dos processos que envolvem a chegada e a dispersão destas espécies em novos ambientes. Somente desta forma seremos capa-zes de intervir no processo e minimizar os im-pactos causados pelas invasões biológicas. Muitas visões pessimistas cercam o tema prevenção. Realmente, aprofundamentos neste tema exigem muito esforço e poucas vezes ob-têm algum resultado conclusivo. Mas isso é um fato que evidencia o quanto ainda se desconhe-ce do assunto.
Figura 1. Relação entre a gravidade dos efeitos (ESEE) e a vul-nerabilidade intrínseca (CER+VD+CGBH).
A identificação e ação direta sobre pos-síveis vetores de dispersão, somada à proposi-ção de um modelo de análise de risco, podem criar subsídios para adoção imediata de ações concretas de prevenção. Porém, isso depende, também, da atuação política, o que torna ne-cessário levar este conhecimento às instâncias governamentais. Por outro lado, a intervenção direta so-bre os vetores de dispersão, principalmente sobre os antrópicos, exige cuidado para que atividades econômicas envolvidas não sejam penalizadas pela falta de conhecimento mais aprofundado do processo de dispersão da es-pécie e da sua biologia. O caminho a ser seguido deve ser o da multidisciplinaridade e da união de esforços, criando uma frente única de discussão do tema e criando mecanismos para controlar o proble-ma crescente das bioinvasões.

373
Invasões normalmente são registradas quando as espécies já estão estabelecidas. Ava-liar o potencial que um ambiente tem de rece-ber uma dada espécie tem a vantagem de que a invasão pode ser avaliada antes da introdução, sendo possível estabelecer medidas preventivas, áreas prioritárias para monitoramento, manejo e conservação da biodiversidade. Cenários de dispersão normalmente são desenvolvidos comparando-se as caracterís-ticas do ambiente onde a espécie ocorre e as características de ambientes potencialmente re-ceptores. Sendo assim, a modelagem de nicho ecológico, a qual permite identificar ambientes onde a espécie pode sobreviver, caso seja in-troduzida, tem sido uma ferramenta útil (Peter-son, 2003, 2006). Dois exemplos desse tipo de modelagem são a previsão da expansão dos moluscos aquá-ticos invasores D. polymorpha na América do Norte (Peterson & Vieglais, 2001; Drake & Bossenbroek, 2004) e L. fortunei em escala global (Kluza & McNyset, 2005). Nesse tipo
de modelagem foram utilizados dados climato-lógicos, geológicos ou topográficos como ca-madas ambientais, analisados em algoritmos matemáticos como GARP e BIOCLIM. Eles mostraram bons resultados, embora com nível de detalhamento pequeno. Para menor escala, como rios dentro de uma bacia hidrográfica, variáveis limnológicas podem dar melhores resultados. A dificulda-de é a disponibilidade de dados. Latini (2006) foi pioneiro na modelagem de nicho ecológico utilizando variáveis limnológicas para prever a distribuição de peixes na bacia do Rio Doce, até então esse tipo de modelagem era feito com dados climatológicos, geológicos e topográfi-cos. No caso dos moluscos aquáticos, a con-centração de cálcio e o pH da água, são consi-deradas variáveis essenciais para o desenvolvi-mento (Mackie & Claudi, 2010). Estas variáveis foram utilizadas para prever a distribuição potencial e a densidade de D. polymorpha na América do Norte sem uso de modelagem, uti-
CAPÍTULO 41
Modelos de previsão da distribuição do
mexilhão-dourado no Brasil
Marcia Divina de Oliveira

Oliveira M. D.
Modelos de previsão da distribuição do mexilhão-dourado no Brasil
374
lizando-se principalmente análises estatísticas e sobreposição de mapas (Ramcharan et al., 1992; Neary & Leach, 1992; Mellina & Ras-mussen, 1994). Nos tópicos seguintes será mostrada a experiência na previsão da distribuição poten-cial do molusco aquático invasor L. fortunei usando variáveis limnológicas em diferentes es-calas espaciais: bacia hidrográfica do Rio Para-guai, rios Brasileiros e rios Norte Americanos, de acordo com Oliveira, M. et al. (2010a, d).
Modelos
Oliveira, M. et al. (2010a) avaliaram a distribuição potencial de L. fortunei na bacia do Rio Paraguai através de análise explorató-ria baseada na distribuição espacial da concen-tração de cálcio e do índice de saturação do carbonato de cálcio (IScalcita), e aplicando dois modelos de nicho ecológico: GARP (Genetic Algorithm for Rule-set Prediction) e Maxent (Maximum Entropy Method). O modelo GARP descreve condições ambientais onde a espécie seria capaz de man-ter populações viáveis, se introduzida. Como entrada para o modelo GARP usou-se dois conjuntos de pontos, um com dados da ocor-rência da espécie (coordenadas geográficas) e o outro, camadas ambientais, que representa os parâmetros ambientais que podem limitar a so-brevivência da espécie no novo ambiente (Sto-ckwell & Noble, 1992; Peterson & Cohoon, 1999; Stockwell & Peters, 1999). O algoritmo Maxent estima a distribui-ção geográfica das espécies procurando a pro-babilidade de distribuição de máxima entropia (isto é, mais espalhada ou mais próxima da distribuição uniforme), sujeita a um conjunto de restrições que representam a informação incompleta sobre a distribuição que se dese-ja (Phillips et al., 2006). Ambos os modelos, GARP e Maxent, são baseados em registros de presença, tendo o mesmo formato de dados.
Variáveis
Um ponto crítico da modelagem é deci-ítico da modelagem é deci-tico da modelagem é deci-dir quais variáveis, ou combinações dessas va-riáveis, deve-se utilizar, além da sazonalidade no caso das variáveis limnológicas, a qual pre-
cisa ser considerada. Guisan & Zimmermann (2000) discutem métodos para ajudar a definir as variáveis, incluindo análises estatísticas. Para estimar a distribuição potencial de L. fortunei, Oliveira, M. et al. (2010a, d) fi-zeram suas previsões utilizando somente vari-áveis limnológicas como temperatura da água, oxigênio dissolvido, condutividade elétrica, pH, concentração de cálcio, sólidos suspensos totais e índice de saturação da calcita (IScalcita). Embora o limite de tolerância de L. for-tunei a variáveis ambientais não esteja bem estabelecido, os autores consideraram os re-querimentos mínimos de pH e cálcio para o desenvolvimento de uma população viável L. fortunei como 6.0 e 1.0 mg l-1 , respectivamen-te. O índice de saturação do carbonato de cál-cio inclui variáveis como temperatura da água, pH, alcalinidade, condutividade e concentra-ção de cálcio, cujo limite inferior foi estabele-cido em –4.0. Sólidos suspensos totais, veloci-dade da água e clorofila a não foram incluídas nos modelos por não estar disponível para muitos ambientes, mas serviram de base para a discussão das áreas potenciais. A concentração de sólidos suspensos combinados com baixa disponibilidade de ali-mento (clorofila em torno de 1.0 μg/L) e alta velocidade da água (em torno de 0.9 m/s), comuns em ambientes lóticos, podem ser li-mitantes para o desenvolvimento das larvas, como sugerido por Oliveira et al. (2011) em seus estudos no Rio Miranda. Estudos sobre o efeito dessas variáveis para L. fortunei não são conclusivos. A escolha das variáveis deve ser feita de acordo com o ambiente. Por exemplo, no Pan-tanal onde eventos de depleção de oxigênio dissolvido provocam mortalidade da popula-ção de L. fortunei na maioria dos anos (Calhei-ros & Hamilton, 1998; Hamilton et al., 1997), variáveis como oxigênio dissolvido, pH e CO2 livre precisam ser consideradas no estabeleci-mento da espécie (Oliveira, M. et al., 2010a).
Exemplos da aplicação de modelos de nicho ecológico
Bacia do alto Rio Paraguai: Segundo Oli-veira, M. et al. (2010a), considerando somente a concentração de cálcio, a maioria dos rios da

CAPÍTULO 41
Seção 12
375
bacia do Alto Paraguai tem médio a alto poten-cial para o desenvolvimento de L. fortunei, e somente alguns rios com cálcio menor que 1.0 mg l-1 pode ter baixo risco, mas ainda alguma chance de suportar populações de L. fortunei considerando a tolerância a águas diluídas rela-tada por Deaton et al. (1989). A previsão base-ada na IScalcita forneceu resultados comparáveis àqueles quando cálcio é utilizado. Águas com cálcio maior que 1.0 mg l-1 e IScalcita acima de –4 tem potencial para o estabelecimento de L. for-tunei com maior probabilidade de população abundante em águas acima desses limites. Regiões indicadas pelos modelos GARP e Maxent como tendo alto risco de invasão nos rios Paraguai, Cuiabá, Miranda/Aqui-dauana e Apa foram coincidentes com áreas onde o cálcio é maior que 1.0 mg l-1, enquanto regiões com menor chance de invasão foram aquelas onde o cálcio é abaixo de 1.0 mg/L, corroborando a ideia de que estes modelos podem dar bons resultados usando variáveis limnológicas. Previsões a partir de modelos empíricos (cálcio e IScalcita) foram consistentes, pois os locais onde a espécie já está presente apresentaram média a alta probabilidade da espécie se estabelecer. A concordância entre modelos usan-do cálcio e IScalcita e os modelos matemáticos (GARP and Maxent) sugerem que ambos os modelos empíricos podem ser utilizados para prever a expansão de L. fortunei como uma al-ternativa aos modelos matemáticos mais com-plexos, os quais exigem mais conhecimento das ferramentas de SIG e domínio de programas computacionais. A vantagem dos modelos em-píricos de cálcio e IScalcita é que eles são fáceis de usar, embora o cálculo do IScalcita requeira vários dados (Oliveira, M. et al., 2010a). Os modelos empíricos são particular-mente eficientes para previsões em pequenas escalas. Neary & Leach (1992) previram a expansão de D. polymorpha em Ontario (Ca-nadá) usando cálcio e pH. A atual distribuição desta espécie depois de aproximadamente 16 anos (Zoltak, 2008) coincide com as áreas de alto risco indicadas nesta previsão. Modelos que simultaneamente consideram Ca e pH são similares para o modelo de IScalcita desde que estas são as variáveis mais importantes no con-trole da IScalcita.
Outros rios Brasileiros: segundo aná-lise de Oliveira, M. et al. (2010d), a maioria dos rios brasileiros tem concentração de cálcio maior que 1 mg/L e IScalcita maior que –4, valo-res mínimos encontrados no Rio Paraguai onde L. fortunei mantem populações ativas, com me-lhores condições para calcificação e crescimen-to se comparadas ao Rio Paraguai. De acordo com os autores em alguns tributários do Rio Amazonas, como Rio Negro, porções do Rio Tapajós e Xingu, e o Rio Araguaia L. fortunei terá menos chance de se estabelecer, pois são rios de águas mais ácidas e com menor concen-tração de cálcio. Oliveira, M. et al. (2010d) previram im-portantes áreas localizadas no estuário do Rio Amazonas bem como áreas nas partes superio-res de tributários do Rio Amazonas como Ta-pajós, Teles Pires, Araguaia e outros rios com condições favoráveis ao desenvolvimento de L. fortunei. Estas mesmas áreas foram previstas como pouco favoráveis para L. fortunei pelo modelo global de Kluza e McNyset (2005). Este é o diferencial dos modelos baseados em dados limnológicos, pois estes fornecem infor-mação detalhada e específica para o sistema aquático, onde se pode analisar cada sistema de rio separadamente. O sistema de rios da bacia Amazônica tem grande potencial de receber a espécie por água de lastro devido aos portos de Manaus e Belém, e sua dispersão pode ser feita pela na-vegação fluvial de grande e pequeno porte, o meio mais comum de transporte na região. Por isso, é importante o uso de ferramentas que ajudem a melhorar a previsão de estabeleci-mento da espécie na região Amazônica, caso ela alcance o Rio Amazonas. O Rio São Francisco possui águas com características muito favoráveis ao estabele-cimento de L. fortunei (Oliveira, M. et al., 2010d). Embora Boltovskoy et al. (2006) con-sideraram que a navegação seja restrita e não intensa comercialmente, outros meios de intro-dução, como a pesca esportiva e profissional, criação de peixes em tanques-rede, movimen-tação de barcos, dentre outros meios, podem favorecer a entrada da espécie, devido à proxi-à proxi- proxi-midade com a bacia do Rio Paraná. Rios norte-americanos: enquanto cál-cio e IScalcita são bons indicadores para prever a

Oliveira M. D.
Modelos de previsão da distribuição do mexilhão-dourado no Brasil
376
expansão de L. fortunei nas águas brasileiras, eles não são para os rios norte-americanos (Oli-veira, M. et al., 2010d). Esses rios tem concen-tração de cálcio, pH e IScalcita muito acima do nível requerido, e então os dados limnológicos usados na modelagem dos rios brasileiros não se aplicaram para os rios norte-americanos. De acordo com Oliveira, M. et al. (2010d) a princi-pal variável que pode limitar o estabelecimento de L. fortunei nos rios americanos analisados é a temperatura da água, e talvez turbidez e ali-mento, sendo os limites dessas duas variáveis desconhecidos para L. fortunei. Tolerância térmica depende do tempo de exposição bem como dos valores máximos e mínimos. A temperatura de inverno na Améri-ca do Norte varia entre 0 e 5ºC no trecho supe-ºC no trecho supe-C no trecho supe-rior do Rio Mississippi e seu tributário, o Rio Missouri. Esta baixa temperatura pode durar por 3 meses nestes rios e impedir o desenvolvi-mento de L. fortunei, caso ela seja introduzida. Já no Rio Colorado, mesmo durante o inver-no, a temperatura da água fica acima de 8ºC, e pode ser favorável ao desenvolvimento de L. fortunei, caso a espécie alcance os Estados Uni-dos. Turbidez e alimento devem ser conside-rados no caso dos rios norte-americanos, visto que o efeito negativo de alta turbidez inorgâ-nica foi observado para o mexilhão-zebra em grandes rios, como Mississippi, quando com-binado com alta temperatura (Alexander et al., 1994; Allen et al., 1999). A concentração de se-dimentos suspensos são maiores nos rios norte--americanos comparados aos rios do Pantanal (Oliveira, M. et al., 2010d), onde a combinação de turbidez e escassez de alimento foi conside-rado um fator importante no desenvolvimento das larvas por Oliveira et al. (2011). Outro fator importante no estuário do Rio Mississippi, que se assemelha ao Rio Pa-raguai, é o fenômeno de depleção de oxigê-nio, o qual também provoca mortalidade de Dreissena polymorpha anualmente (Mihuc et al., 1999). Acima do delta a concentração de oxigênio dissolvido é superior a 1.0 mg l-1. O
mesmo pode ser observado no Rio Grande. A distribuição potencial de L. fortunei sobrepõe à distribuição atual de corbiculídeos e dreisse-nídeos na América do Norte, e se estende para áreas que são mais quentes, com menor cálcio e sujeitas a eventos de depleção de oxigênio. Des-ta forma, os mesmos modelos de distribuição potencial usados para L. fortunei na América do Sul não podem ser aplicados na América do Norte. Modelos globais como o de Kluza & McNyset (2005), usando dados climatoló-gicos também não são capazes de identificar máximos e mínimos de temperatura, e princi-palmente a duração das temperaturas mínimas para a sobrevivência de L. fortunei.
Considerações finais
As ferramentas para modelagem têm sido aprimoradas tornando mais confiáveis e mais realísticos os resultados gerados pela modela-gem. Os métodos de previsão incluem métodos estatísticos, algoritmos matemáticos, ou, sim-plesmente comparação de habitats usando ca-racterísticas ambientais que sejam importantes para o estabelecimento de uma espécie. Cabe ao usuário a difícil tarefa de decidir qual a me-todologia é mais adequada para a sua previsão, considerando, principalmente, sua escala espa-cial e a base de dados disponível. Há muitas incertezas na modelagem de nicho ecológico, como acuracidade, tamanho da amostra, vari-áveis utilizadas. O grau de incerteza aumenta com o tamanho da escala geográfica. A acura-cidade dos modelos ainda é algo questionável. O resultado final de qualquer previsão deve ser avaliado por especialistas nas áreas afins, que são os principais validadores desses métodos de previsão por ter conhecimento da biologia da espécie. A modelagem de nicho eco-lógico tem mostrado bons resultados, mas é pre-ciso estar atento, pois o nicho ecológico da es-pécie é muito mais complexo do que os modelos possam retratar, e não considera as interações bióticas, como predação, competição, habilida-de da espécie de dispersar, dentre outros.

377
Limnoperna fortunei (Dunker, 1857), popularmente conhecido por mexilhão-doura-do, é um dos casos mais emblemáticos de bioin-vasão aquática no Brasil. Este molusco bivalve de água doce, nativo de rios do sudeste asiá-tico, foi registrado em 1998, tanto na região do Lago Guaíba, Rio Grande do Sul (Mansur et al., 1999) como da região do Pantanal no estado do Mato Grosso (Oliveira et al., 2006). A partir da invasão, L. fortunei disseminou--se por diversos corpos hídricos e atualmente se encontra presente em ambientes continen-tais (lagos, reservatórios e rios) (Mansur et al., 2003; Oliveira et al., 2006) e estuarinos (Darrigran & Pastorino, 1995; Brugnoli et al., 2005; Capítoli et al., 2008) do sul da América do Sul L. fortunei tem causado danos ambien-tais e econômicos (Darrigran & Pastorino, 1995; Mansur et al., 2004b). Esses danos são geralmente elevados devido ao seu hábito epi-zoico, capacidade de formar macroaglomera-ções e as frequentes explosões populacionais
no ambiente invadido (Darrigran & Pastorino 1995; Mansur et al., 2004b). Considerando que a erradicação da espécie é extremamente difícil (Oliveira et al., 2006), medidas de con-trole de sua propagação são importantes. Em estuários, a presença e distribuição de L. fortunei podem ser limitadas pela salini-dade (Darrigran & Pastorino, 1995; Brugno-li et al., 2005; Capítoli & Bemvenuti, 2004). No estuário do Rio da Prata, Argentina, Dar-rigran & Pastorino (1995) constataram que a distribuição de L. fortunei é limitada por altos valores de salinidade e que a espécie resiste a salinidades de até 3 ups (unidade padrão de sa-linidade). Em outro estudo, na costa Uruguaia do Rio da Prata, o papel da salinidade como o principal fator abiótico limitante da distribui-ção de L. fortunei foi descrito por Brugnoli et al. (2005). Segundo estes autores, L. fortunei ocorre nas áreas correspondentes às iso-halinas de valores 2-3 ups em Punta Espinillo e nas lo-calidades de Colônia e Montevidéu com salini-dades de 0 e 1,6 ups respectivamente
CAPÍTULO 42
Previsão de cenários de distribuição de Limnoperna fortunei na Laguna dos Patos
Fabiana Gonçalves BarbosaAdriano Sanches Melo

Barbosa & Melo
Previsão de cenários de distribuição de Limnoperna fortunei na Laguna dos Patos
378
Distribuição potencial de L. fortunei em relação a variações de salinidade na Laguna dos Patos
A Laguna dos Patos localiza-se na planí-cie costeira do estado do Rio Grande do Sul, Brasil. Ela representa a maior área lagunar da América do Sul e a maior laguna costeira do tipo “estrangulada” do mundo (Kjerve, 1986). Ela pode ser dividida em três regiões ecológicas distintas: i) a região límnica, que compreende a célula do setor norte da Laguna dos Patos; ii) a região pré-límnica, que se estende desde 31°05’S até o limite da região estuarina; e iii) a região estuarina, na porção sul da laguna, delimitada ao norte por uma linha imaginá-ria traçada entre a Ponta da Feitoria (31°41’S e 52°02’O) à Ponta dos Lençóis (31°11’S e 52°04’O) (Capítoli & Bemvenuti, 2004) (Fig. 1). Na região estuarina da Laguna dos Patos o aporte de água salgada proveniente do Ocea-no Atlântico é essencialmente regido pelo nível da água na Laguna dos Patos e intensidade dos ventos atuantes (Möller et al., 2001). Neste es-
tuário, a salinidade segue um padrão sazonal: i) valores baixos no outono e inverno, devido às descargas de água doce e fortes ventos de nor-deste e ii) valores altos na primavera e verão, em períodos dominados por ventos de sudeste e baixo nível da água que favorecerem a sali-nização da região (Niencheski & Baumgarten, 1997) (Fig. 2). O modelo preditivo ajustado por Barbosa & Melo (2009) para predizer a sobrevivência de L. fortunei em relação a variações de salinidade na Laguna dos Patos demonstrou que a sobrevi-vência da espécie foi afetada negativamente pela salinidade, com alta sobrevivência em águas com salinidade de até 5 ups e decréscimo em salini-dades mais altas. A partir do modelo ajustado, estes autores criaram cenários de distribuição potencial das áreas favoráveis à sobrevivência de L. fortunei na Laguna dos Patos em função da salinidade para as quatro estações climáticas do ano (Fig. 3). Os cenários obtidos mostraram duas situações relevantes: i) durante o período de baixas salinidades (inverno), as regiões límni-ca, pré-límnica e estuarina da Laguna dos Patos
Figura 1. Laguna dos Patos e sua divisão ecológica. Fonte: modificada de Barbosa & Melo (2009).

CAPÍTULO 42
Seção 12
379
mostram-se favoráveis à sobrevivência da espé-cie e ii) no período de altas salinidades (verão), a região estuarina apresenta-se desfavorável à sobrevivência do molusco (Fig. 3). Adicionalmente, estes cenários preditos apresentaram uma distribuição de sobrevivên-cia de L. fortunei perante variações de salini-dade na Laguna dos Patos bastante precisa, especialmente em comparação aos registros históricos da distribuição do mexilhão na La-guna dos Patos. Atualmente, L. fortunei possui populações persistentes nas regiões límnica e pré-límnica da Laguna dos Patos (Mansur et al., 2004b; Capítoli et al., 2008). Estudos sobre a distribuição de L. fortunei em distintos am-bientes do estuário da Laguna dos Patos (Capí-toli & Bemvenuti, 2004; Burns et al., 2006b; Piedras et al., 2007) sugerem que as coloniza-ções e/ou persistências ocorrem concomitante-mente à predominância de baixas salinidades. A salinização é um fator natural contro-lador da disseminação de L. fortunei durante períodos de salinização do baixo Arroio Pelo-tas, localizado no município de Pelotas (Pie-dras et al., 2007). Entretanto, a espécie man-tém estoques populacionais persistentes em locais com salinidade inferior a 5 ups. No Ca-nal de São Gonçalo, que conecta o estuário da
Laguna dos Patos com a Lagoa Mirim, foram encontradas populações estabelecidas de L. fortunei na região límnica, embora populações na região estuarina deste canal não consigam se estabelecer devido aos eventos de estiagem e salinização (Burns et al., 2006b). No estuá-rio da Laguna dos Patos, durante o período de 2002-2003, o mexilhão-dourado colonizou o setor norte do estuário, assim como a região estuarina próxima a desembocadura do Canal do Rio Grande (Capítoli & Bemvenuti, 2004). Posteriormente, Capítoli & Bemvenuti (2004) registraram a mortalidade destas populações anteriormente estabelecidas no corpo do es-tuário, e isto aconteceu após uma progressiva salinização que ocorreu no verão de 2004. A Laguna dos Patos, juntamente com o seu estuário, possui uma grande importância ecológica, econômica e social para o estado do Rio Grande do Sul, Brasil (Asmus & Ta-gliani, 1998). Atualmente, as populações per-sistentes de L. fortunei nas regiões límnica e pré-límnica da Laguna dos Patos e na Lagoa Mirim (Mansur et al., 2004b; Capítoli et al., 2008) têm causado grandes danos ambientais e econômicos (Mansur et al., 2004b). Duran-te condições de vazante, ou seja, descarga de água doce em direção à região sul da laguna,
Figura 2. Salinidades nas regi-ões ecológicas da Laguna dos Patos para as quatro estações climáticas registradas de 2000 a 2004. As caixas representam os percentis 25 e 75%. A linha hori-zontal dentro da caixa represen-ta a mediana e o ponto a média. As barras de erro representam o mínimo e o máximo. Fonte: modi-ficada de Barbosa & Melo (2009).

Barbosa & Melo
Previsão de cenários de distribuição de Limnoperna fortunei na Laguna dos Patos
380
pode haver transporte de larvas de L. fortunei destas regiões para o estuário da Laguna dos Patos e consequente colonização no setor norte do estuário, uma vez que essa parte do estuário é favorável à sobrevivência da espécie durante o inverno e relativamente favorável nas outras
Figura 3. Mapas de distribuição predita das áreas favoráveis à sobrevivência de Limnoperna fortunei na Laguna dos Patos (Rio Grande do Sul, Brasil) em função da salinidade para cada estação climática. Onde: i) área muito favorável – sobrevi-vência >75% (0< salinidade< 5,1); ii) área relativamente favorável – 75% > sobrevi-vência > 50% (5,1<salinidade<11,6); e iii) área desfavorável - 50% > sobrevivência (11,6< salinidade< 33). Fonte: modificada de Barbosa & Melo (2009).
estações climáticas do ano (Fig. 3). Neste sen-tido, os cenários preditos de sobrevivência da espécie (Barbosa & Melo, 2009) (Fig. 3) po-dem fornecer subsídios para ações de controle à distribuição e persistência de L. fortunei na Laguna dos Patos.

13Seção
Manejo de espéciesinvasoras


383
Para poder estabelecer as estratégias mais adequadas para o manejo das bioinvasões, deve-se delinear um programa de manejo, em que devem estar claros dois critérios: (1) o cri-tério utilizado para definir uma espécie inva-sora; (2) o critério de sucesso para medir um programa de manejo de espécies invasoras. (1) O primeiro critério, segundo Pereyra (ms.), permitirá definir as prioridades no ma-nejo da bioinvasão, concentrando os recursos nas espécies mais problemáticas para a região. Esse autor assinala que, segundo Valéry et al. (2008b), os dois conceitos mais difundidos para definir as espécies invasoras são o bioge-ográfico e o do impacto ocasionado; também Valéry et al. (2008b) propõem sua própria defi-nição, baseando-se no processo de invasão pro-priamente dito. Cabe destacar que cada uma das definições consideradas apresenta virtudes e carências. O critério biogeográfico é o mais prático, permitindo, em princípio, melhor ma-nejo das espécies em trânsito a partir de seu lugar de origem (Hulme, 2006). O conceito
embasado no processo de invasão dá lugar ao manejo de uma espécie que, por seu compor-tamento dominante diante de certos fatos ou impactos ambientais, afeta a estabilidade de uma comunidade, ainda que essa seja nativa. Por último, e coincidindo com Davis (2006), as espécies invasoras, sob o ponto de vista do impacto que causam, são um transtorno para a biodiversidade, e algumas delas constituem um importante problema socioeconômico. Neste capítulo, entende-se espécie inva-sora como o organismo introduzido em con-sequência da atividade humana em área ou ecossistema em que não é nativo; que coloni-za ambientes naturais e não naturais, conver-tendo-se, assim, em um agente de mudança e ameaça à biodiversidade nativa (Darrigran & Damborenea, 2009a) e de potenciais danos econômicos (Darrigran, 2009). (2) Para o caso do segundo critério, de-ve-se evitar limitar a medição do sucesso do programa de manejo de espécies não nativas à diminuição ou à eliminação da espécie invaso-
1 Tradução de Maria Regina Lucena Borges Osório.
CAPÍTULO 43
Manejo integrado de espécies invasoras1
Gustavo DarrigranFernando Archuby
Maria Cristina Dreher Mansur

Darrigran et al.
Manejo integrado de espécies invasoras
384
ra em estudo. Essa forma de pensar desvia-se do objetivo final desses programas, que não é somente a eliminação das espécies não nativas, mas também a restauração da biodiversidade do ecossistema. Embora seja natural que a distribuição das espécies mude ao longo do tempo, o ho-mem tem incrementado essa distribuição não só em escala espacial, cruzando barreiras in-transponíveis aos organismos (montanhas, oceanos, desertos, etc.), mas também tem acelerado sua taxa de dispersão (Darrigran & Guimarey, no prelo). Além disso, nas últimas décadas, incrementou-se ainda mais o número de bioinvasões, fundamentalmente por meio de dois processos: 1) Incremento do comércio, devido à “globalização”. Esse processo provocou um au-mento na quantidade dos corredores de invasão (rotas aquáticas, terrestres ou aéreas; comerciais e/ou turísticas) e o acesso do que se denomina propágulo (p. ex., um exemplar fecundado, ou uma população, um quisto ou uma fase de resis-tência de uma espécie não nativa), com a capaci-dade de gerar a população inicial em uma região na qual essa espécie não é nativa. 2) Incremento de ambientes naturais im-pactados. Já na década de 1950, Elton (1958) afirmava que, em um ambiente não impactado, era difícil a introdução de propágulos. Nesse caso, a mudança global é encarregada de trans-formar os ambientes em mais “frágeis” e, por-tanto, mais suscetíveis a permitir o ingresso das bioinvasões. Como exemplo do que foi mencionado nos parágrafos anteriores, Falk-Petersen et al. (2006) consideram o caso da Baía de San Fran-ía de San Fran-a de San Fran-cisco (Califórnia). Esses autores examinaram dados obtidos no estuário da referida Baía, analisando, como variável temporal, o decurso de 140 anos. Ao longo desse período, foram monitorados os organismos aquáticos (plantas, protistas, invertebrados e vertebrados) e foi ge-rada uma Curva de Invasão Cumulativa (Fig. 1). Essa curva não apresenta um incremento li-near, salvo uma taxa acelerada de introdução e estabelecimento de espécies não nativas. Nessa figura, pode-se observar que aproximadamen-te 50% das espécies não nativas introduziram--se nos últimos 35 anos. A invasão de espécies não nativas aumentou em uma média de:
* uma (1) espécie não nativa, a cada 55 semanas no período entre 1850 e 1960; * uma (1) espécie não nativa, a cada 14 semanas no período entre 1961 e 1995.
Esse exemplo, que sustenta a expansão das bioinvasões nas últimas décadas, somado à tendência desse fenômeno para ocasionar perda da biodiversidade (homogeneização global do ambiente) e problemas econômicos associados, viu-se acompanhado de um incremento no nú-mero de publicações científicas, o que, por sua vez, assinala um aumento no número de pes-quisas realizadas. Esse fato positivo ocasiona um problema: a multiplicidade de termos técni-cos, muitos sendo sinônimos que perturbam a explicação dos processos de bioinvasão. Hicks (2004), com a finalidade de chegar à solução desse problema, propõe uma simplificação es-trutural para conseguir explicar o processo de bioinvasão. Esse autor sugere sua referência em duas fases, uma representada pela capacidade de invadir (“invasividade”) da espécie em estu-do e a outra considerando o ambiente e sua sus-cetibilidade a ser invadido (“invasibilidade”). Durante o processo de invasão, uma es-pécie passa por uma série de fases ou etapas (Fig. 2). Em cada uma dessas fases, existe uma drástica mudança de situação da espécie, de acordo com o tipo de ambiente onde se en-contre. A estratégia de manejo de uma espécie
Figura 1. Curva de Invasão Cumulativa (quantidade de bioin-vasões ao longo do tempo) no estuário da Baía de San Francis-co. Modificada de Falk-Petersen et al. (2006).

CAPÍTULO 43
Seção 13
385
invasora é altamente dependente da etapa em que esta se encontra no processo de invasão. Em geral, é nesse item que o critério de mane-jo aplicado nos diferentes casos de bioinvasões apresenta uma falha. Essa falha relaciona-se com as políticas de manejo, que não conside-ram que em cada fase do processo de invasão exista uma forte variação da situação particu-lar da espécie. Portanto, em cada etapa da inva-são, os esforços de manejo devem ser distintos. Pode-se chegar ao extremo em que esse manejo será inútil, se não forem consideradas previa-mente as próprias condições de cada etapa de invasão. Deve-se observar, por exemplo, que há etapas em que é ótimo investir recursos na prevenção, enquanto outras, no controle, e as-sim sucessivamente. Com base nisso e seguindo Hicks (2004), pode-se fazer um gráfico desse fato (Fig. 3), contrastando uma curva de custos (econômicos, esforços), com uma curva que re-presente a probabilidade de erradicação de uma espécie não nativa. Desse gráfico, depreende-se que a combinação de custo mínimo de erradi-cação e a máxima probabilidade de sucesso é observada no início do processo, quando, em geral, por falhas de monitoramento, não é evi-dente o recente assentamento da espécie. Ao contrário, quando a espécie se torna evidente, isto é, quando passou o tempo de menor custo e a probabilidade de sucesso é quase nula, é que se começa a realizar o manejo da espécie inva-sora. Cabe ressaltar que, independentemen-te do tempo, geralmente se começa a realizar o manejo nos casos em que a bioinvasão afeta o homem, seja direta ou indiretamente, e não quando o impacto que ocasiona é apenas em detrimento da biodiversidade nativa não utili-zada como recurso pelo homem. O que foi mencionado no parágrafo an-terior leva a se inferir que, uma vez detectada a bioinvasão, já ocorreu seu impacto sobre o ecossistema em estudo. Caso se chegue, então, à erradicação pouco provável da espécie nessa fase, deve-se considerar também em que situa-ção se encontra esse ecossistema. Segundo Bull & Courchamp (2009), ainda que a eliminação seja de uma única espécie não nativa já assenta-da em um ecossistema, não conduz necessaria-mente à restauração da biodiversidade afetada. Em alguns casos, inclusive, tem o potencial de
provocar efeitos opostos, exercendo impacto sobre o ecossistema e sua biota nativa. Desse modo, pode-se afirmar que a erradicação de espécies não nativas não leva necessária e au-tomaticamente à restauração ou à recuperação das condições ambientais originais da biodiver-sidade afetada. Os estudos de pré-erradicação são essenciais para se estimar a probabilidade dessas incidências. Entender quais são as es-pécies que estão presentes em um ecossistema,
Figura 2. Etapas no processo de invasão de uma espécie exó-tica. Modificada de Lockwood et al. (2007).
Figura 3. Relação entre o custo da erradicação de uma espécie invasora e a probabilidade de erradicação ao longo do tempo.

Darrigran et al.
Manejo integrado de espécies invasoras
386
como interagem e onde se localizam na rede alimentar, situará o encarregado de realizar o manejo da bioinvasão em uma boa posição para tomar decisões relativas aos possíveis efeitos sobre o ecossistema. Além disso, permitirá não somente o sucesso na eliminação de espécies não desejadas, mas também a maior probabi-lidade de alcançar o objetivo principal que um programa de manejo de espécies invasoras deve ter: restauração da biodiversidade nativa. Portanto, a implementação de um ma-nejo integrado e sustentável de uma bioinvasão deve ser completada com a geração de conhe-cimento sobre a mesma. Dessa forma, em pri-meira instância, se poderá realizar a explicação ou descrição, predição e risco de invasão e, em segunda instância, se poderá chegar ao objeti-vo final do programa de manejo de uma bioin-vasão: a restauração. Entende-se por explica-ção da invasão o estudo descritivo do evento, acompanhado de estudos experimentais, dinâ-mica populacional, análises estatísticas, etc. [p. ex., Darrigran & Damborenea (eds.) 2006a]. A predição da invasão refere-se à avaliação da probabilidade de que ocorra um evento de in-
vasão (Belz et al., no prelo). Finalmente, o ris-co de invasão é o produto da probabilidade de que ocorra um evento e de suas consequências (Belz, 2009). Tendo por base que conservar a biodiver-sidade é um ponto crucial para a vida na Ter-ra, que, por sua vez, está sendo devastada pela ação do homem, por meio da destruição do habitat, em primeiro lugar, e da introdução de espécies não nativas, em segundo lugar (Hayes, 2003), as áreas de trabalho para a conservação da biodiversidade podem servir de eixo articu-lador para se estabelecer um padrão de manejo de bioinvasões. Portanto, nesse nível de situa-ção global diante das bioinvasões, devem ser estabelecidas com precisão as áreas de traba-lho, que também devem estar integradas com políticas ambientais regionais de ação (Darri-gran & Darrigran, 2001). Com fundamento no que foi exposto até agora, o manejo integrado de espécies invaso-ras depende de três componentes (Fig. 4), o pri-meiro eixo sendo o critério adotado no concei-to de espécie invasora e a forma de medição do sucesso do programa de manejo desenvolvido. O segundo eixo é o marco de conhecimentos prévios ou conhecimentos de base com que se propõe o manejo, enquanto o terceiro eixo é representado pela geração do conhecimento, que ajusta e sustenta, interagindo com os ou-tros dois eixos. Não obstante, nesse diagrama de fluxo falta um quarto eixo que teria o po-der de tornar o programa de manejo de bio-invasões sustentável no tempo. Cowie (2004) também estabelece três componentes básicos para um programa de conservação da biodiver-sidade (Fig. 5). O primeiro deles é o conheci-mento científico gerado a partir das Ciências Naturais, o qual deve chegar às organizações ambientais locais e regionais com a finalidade de realizar um manejo (o segundo componen-te) adequado. Ainda que se cumpram esses dois pontos anteriores, também assinala, de modo contundente, que, sem o apoio do setor da po-lítica socioambiental, somente se teria suces-so em pequenas áreas e durante curto tempo, tanto quanto durasse a gestão ou o grupo de pesquisa que a realizasse. Esse terceiro compo-nente é o mais complexo dos três, já que é en-
Figura 4. Diagrama de três componentes no manejo integra-do de espécies invasoras.
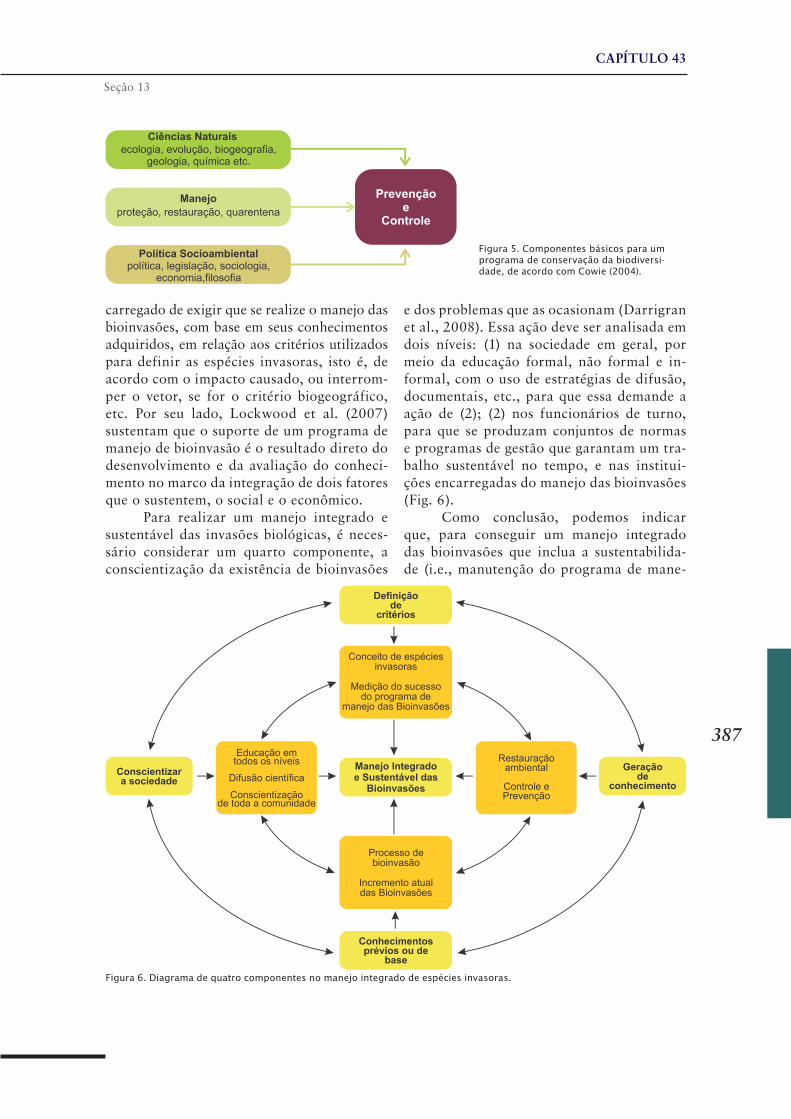
CAPÍTULO 43
Seção 13
387
carregado de exigir que se realize o manejo das bioinvasões, com base em seus conhecimentos adquiridos, em relação aos critérios utilizados para definir as espécies invasoras, isto é, de acordo com o impacto causado, ou interrom-per o vetor, se for o critério biogeográfico, etc. Por seu lado, Lockwood et al. (2007) sustentam que o suporte de um programa de manejo de bioinvasão é o resultado direto do desenvolvimento e da avaliação do conheci-mento no marco da integração de dois fatores que o sustentem, o social e o econômico. Para realizar um manejo integrado e sustentável das invasões biológicas, é neces-sário considerar um quarto componente, a conscientização da existência de bioinvasões
e dos problemas que as ocasionam (Darrigran et al., 2008). Essa ação deve ser analisada em dois níveis: (1) na sociedade em geral, por meio da educação formal, não formal e in-formal, com o uso de estratégias de difusão, documentais, etc., para que essa demande a ação de (2); (2) nos funcionários de turno, para que se produzam conjuntos de normas e programas de gestão que garantam um tra-balho sustentável no tempo, e nas institui-ções encarregadas do manejo das bioinvasões (Fig. 6). Como conclusão, podemos indicar que, para conseguir um manejo integrado das bioinvasões que inclua a sustentabilida-de (i.e., manutenção do programa de mane-
Figura 5. Componentes básicos para um programa de conservação da biodiversi-dade, de acordo com Cowie (2004).
Figura 6. Diagrama de quatro componentes no manejo integrado de espécies invasoras.

Darrigran et al.
Manejo integrado de espécies invasoras
388
jo no tempo), é necessário contar com uma sociedade atenta a esse problema, que exija dos funcionários a continuidade das ações no tempo. Isso se consegue graças à incorpora-ção do quarto componente, “conscientizar a sociedade”, que mantém ativos os componen-tes e que consegue retroalimentar-se com os mesmos, por meio da “educação em todos os níveis”, “a difusão científica” e a “conscienti-zação da comunidade em seu conjunto”.
Agradecimentos Ao Lic. Prof. Alfredo Vilches pelos concei-tos proporcionados na realização das Figuras 4 e 6. À Lic. Mirta Lagreca, do pessoal de apoio da Comisión de Investigaciones Científicas (CIC), da Província de Buenos Aires, Argentina, pela ajuda técnica oferecida. Para a elaboração deste capítu-lo, contou-se com o auxílio da FCNyM (UNLP, Argentina); PIP nº 1017 (CONICET, Argentina).

389
Referências
Abbott, R.T. 1973. Spread of Melanoides tuberculata. The Nautilus 87(1):29.Abebe, E.; Decramer, W.; De Lay, P. 2008. Global diversity of nematodes (Nematoda) in freshwater. Hydrobiologia 595:67-78.Abílio, F.J.P.; Fonseca-Gessner, A.A., Leite, R.L., Ruffo, T.L.M. 2006. Gastrópodes e outros invertebrados do sedimento e as-sociados à macrófita Eichhornia crassipes de um açude hiper-trófico do semi-árido paraibano. BioTerra 1(supl.):165-178.Abílio, F.J.P.; Ruffo, T.L.M.; Souza, A.H.F.F.; Junior, E.T.O.; Meireles, B.N.; Santana, A.C.D. 2007. Macroinvertebra-dos bentônicos como bioindicadores de qualidade ambien-tal de corpos aquáticos da caatinga. Oecologia Brasiliensis 11(3):397-409. Agudo-Padrón, A. I. 2010. O camarão de cultivo produzido no nordeste do Brasil como vetor de introdução e dispersão do caramujo invasor Melanoides tuberculatus (Müller, 1774) (Thiaridae) no estado de Santa Catarina. Informativo SBMa 41(171):10-12.Aguilar-Aguilar, R.; Martínez-Aquino, A.; Pérez-Rodríguez, R.; Pérez-Ponce-De-León, G. 2009. Digenea, Heterophyidae, Centrocestus formosanus (Nishigori, 1924) metacercariae: Distribution extension for Mexico, new state record, and geo-graphic distribution map. Check List 5(2):357–359. Alam, Z.B.; Otaki, M.; Furumai,H.; Ohgaki, S. 2001. Direct and indirect inativation of Microcystis aeruginosa by UV-radi-ation. Water Research 35 (4):1008-1014.Aldridge, D.C.; Elliott, P.; Moggridge, G.D. 2006. Microen-capsulated BioBullets for the control of biofouling zebra mus-sels. Environmental Science & Technology 40:975-979.
Alexander, J.E. JR; Thorp, J.H.; Fell, R.D. 1994. Turbidity and temperature effects on oxygen consumption in the zebra mussel (Dreissena polymorpha). Canadian Journal of Fisheries and Aquatic Sciences 51:179-184.Alho, C.J.R. 2011. Concluding remarks: overall impacts on biodiversity and future perspectives for conservation in the Pantanal biome. Brazilian Journal of Biology 71(1):337-341.Alho, C.J.R.; Mamede, S.; Bitencourt, K.; Benites, M. 2011. Introduced species in the Pantanal: implications for conserva-tion. Brazilian Journal of Biology 71(1):321-325.Ali, B.A.; Salem, H.H.; Wang, X.M.; Huang, T.H.; Xie, Q.D.; Zhang, X.Y. 2010. Effect of Bacillus thuringiensis var. israe-lensis endotoxin on the intermediate snail host of Schitosoma japonicum. Current Research in Bacteriology 3(1):37-41.Allen, Y.; Thompson, B.A.; Ramcharan, C.W. 1999. Growth and mortality rates of the zebra mussel, Dreissena polymorpha, in the Lower Mississippi River. Canadian Journal of Fisheries and Aquatic Sciences 56: 748-759.Almeida, C.M.M. 2007. Desinfecção com dióxido de cloro. Boletim SPQ 105: 21-29.Alonso, A.; Camargo, J. A. 2010. Comparison of two methods for estimating the abundance, diversity and habitat preference of fluvial macroinvertebrates in contrasting habitats. Limnolo-gica 40: 23-29. Alves, S.B. 1998. Controle microbiano de insetos. FEALQ, Pi-racicaba. 1163 p.Alves, S.B.; Lopes, R.B. 2008. Controle Microbiano de Pragas na América Latina: avanços e desafios. FEALQ: Piracicaba. 414 p.

390
Amaral, A.C.Z.; Volkmer-Ribeiro, C.; Mansur, M.C.D.; San-tos, S.B.; Avelar, W.E.P.; Mattews-Cascon, H.; Leite, F.P.P.; Melo, G.A.S.; Coelho, P.A.; Buckup, G.B.; Buckup, L.; Ven-tura, C.R.R.; Tiago, C.G. 2008. A Situação de Ameaça dos Invertebrados Aquáticos no Brasil. p. 156-351. In: Machado, A.B.M.; Drummond, G.M.; Paglia, A.P. (Orgs.). Livro Verme-lho da Fauna Brasileira Ameaçada de Extinção. Ministério do Meio Ambiente – MMA, Brasília, v. 1.Amarista, M.; Niquil, N.; Balzana, C.B.; Pointier, J.P. 2001. Inter-Inter-specific competition between freshwater snails of medical impor-fic competition between freshwater snails of medical impor-c competition between freshwater snails of medical impor-tance: a Venezuelan example. Comptes Rendus de l’Académie des Sciences, Series III, Sciences de la Vie 324:143-148.Amusan, A.A.S.; Anyaele, O.O.; Lasisi, A.A. 2002. Effects of copper and lead on growth, feeding and mortality of terrestrial gastropod Limicolaria flammea (Müller, 1774). African Jour-nal of Biomedical Research 5:47-50.Andrade, C.P.; Pinto, H.A.; Coscarelli, D.; Vidigal, T.H.D.A.; Melo, A.L. 2008. Infecção natural de Melanoides tuberculatus (Caenogastropoda: Thiaridae) por larvas de trematódeos dige-néicos em Brasília, Distrito Federal. In: Congresso Brasileiro de Zoologia, 27, 2008, Curitiba. Resumos... p. 1483.Angonesi, L.G.; Bemvenuti, C.E. 2008. Tolerance to salinities shocks of the invasive mussel. Control 98(1):66-69.APHA. 1992. “Standard Methods for the Examination of Wa-ter and Wastewater”. Ed: Greenberg, A. E.; Clescer, L. S.; Ea-ton, A. D.; Franson, M. A. H. 18th ed. American Public Health Association, Washington. 887p. APHA. 2004. Standards Methods for the Examination of Water and Wastewater. American Public Health Association, Washington, New York. 1368p. APHA. 2005. American Public Health Association; American Water Works Association; Water environment Federation. Standard Methods for the Examination of Water & Wastewa-ter. 21ª ed. Washington.Appleton, C.C. 1996. Freshwater mollusks of Southern Africa. University of Natal Press, Pietermaritzburg. 64 p.Araujo, R.; Moreno, D.; Ramos, M.A. 1993. The asiatic clam Corbicula fluminea (Müller, 1774) (Bivalvia, Corbiculidae) in Europe. American Malacological Bulletin 10(1):39-49.Arim, M.; Abades, S. R.; Neill, P.E.; Lima, M.; Marquet, P.A. 2006. Spread dynamics of invasive species. PNAS 103(2): 374-378.Arnaiz, R.R. 1997. Las toxinas ambientales y sus efectos ge-néticos. La Ciência para Todos, Fondo de Cultura Econômica, México. 95p.Arruda, J.O.; Pereira, D.; Bergonci, P.E.A.; Santos, C.P.; Man-sur, M.C.D. 2009. Novos registros de Omalonyx matheroni (Pontiez; Michaud, 1835) (Mollusca, Gastropoda, Succinei-dae) para os Estados de São Paulo e Paraná, Brasil. Biotemas 22(2):181-184.Asmus, M.L. 1997. Coastal plain and Patos Lagoon. p. 09-12. In: Seeliger, U.; Odebrecht, C.; Castello, J.P. (Eds). Subtropi-cal Convergence Environments: the coast and sea in the Sou-thwestern Atlantic. Springer-Verlag, Berlin, 308p.Asmus, M.L.; Tagliani, P.R.A. 1998. Considerações sobre o manejo ambiental. p. 225-229. In: Seeliger, U.; Odebrecht, C.; Castello, P. (Eds). Os ecossistemas costeiro e marinho do extremo Sul do Brasil. Ecoscientia, Rio Grande, Brasil, 326p.Atalah, J.; Kelly-Quinn, M.; Irvine, K.; Crowe, T. P. 2010. Im-pacts of invasion by Dreissena polymorpha (Pallas, 1771) on the performance of macroinvertebrate assessment tools for eu-trophication pressure in lakes. Hydrobiologia 654:237–251.Avelar, W.E.P.; Martins, S.L.; Vianna, M.P. 2004. A new occur-A new occur-rence of Limnoperna fortunei (Dunker 1856) (Bivalvia, Myti-lidae) in the State of São Paulo, Brazilian Journal of Biology 64(4):739-742.
Avise, J.C. 2004. Molecular Markers, Natural History and Evolution. Sinauer Associates, Inc., EUA.Baker, S.M.; Levinton, J.S.; Kurdziel, J.P.; Shumway, S.E. 1998. Selective feeding and biodeposition by zebra mussels and their relation to changes in phytoplankton composition and seston load. Journal of Shellfish Research 17:1207-1213.Baker, S.M.; Levinton, J.S.; Ward, J.E. 2000. Particle transport in the zebra mussel Dreissena Polymorpha (Pallas). Biological Bulletin 199:116-125.Baptista, C.T.; Zibetti, V. 2006. Mexilhão dourado (Limno-perna fortunei) como recurso alimentar de peixes do Arroio Pelotas, Pelotas, RS: Resultados Preliminares XV Congresso de IC.Barbosa, F.G.; Melo, A.S. 2009. Modelo preditivo de sobrevi-vência do mexilhão dourado (Limnoperna fortunei) em rela-ção a variações de salinidade na Laguna dos Patos, RS, Brasil. Biota Neotropica 9(3):407-412. Barosa, J.; Ferreira, A.; Fonseca, B.; Souza, I. 2003. Teste de toxicidade do sulfato de cobre para o mexilhão mediterrâneo Mytilus galloprovincialis (Lamarck), em água do mar. Univer-sidade de Algarve, Faro, 13p.Barros, J.S.L.; Pile, E.A.; Vasconcellos, M.C.; Santos, J.A.A.; Lessa, C. 2002. Experimental infection of Physa cubensis Pfeiffer, 1839 and Lymnaea columella with Fasciola hepatica Linnaeus, 1758 miracidiae. Brazilian Journal of Veterinary Re-search and Animal Science 39 (3):121-123.Barroso, C.X.; Matthews-Cascon, H. 2009. Distribuição es-pacial e temporal da malacofauna no estuário do Rio Cea-rá, Ceará, Brasil. Pan-American Journal of Aquatic Sciences 4(1):79-86.Barton, N.; Turelli, M. 1989. Evolutionary quantitative genet-ics: How little do we know? Ann. Rev. Genet. 23:237-370.Bastviken, D.T.E.; Caraco, N.F.; Cole, J.J. 1998. Experimen-tal measurements of zebra mussel (Dreissena polymorpha) im-pacts on phytoplankton community composition. Freshwater Biology 39:375-386.Beasley, C.; Tagliaro, C.; Figueiredo, W.B. 2003. The occur-rence of the Asian clam Corbicula fluminea in the lower Ama-zon Basin. Acta Amazônica 33(2):317-323.Beekey, M.A.; McCabe, D.J.; Marsden. 2004. Zebra mussel colonization of soft sediments facilitates invertebrate commu-nities. Freshwater Biology 49(5):535545.Belanger, S.E.; Farris, J.L.; Cherry, D.S.; JR. Cairns, J. 1985. Sediment preference of the freshwater Asiatic clam, Corbicula fluminea. The Nautilus 99(2-3) 66-73.Belz, C.E. 2006. Análise de risco de bioinvasão por Limnoper-na fortunei (Dunker, 1857): um modelo para a bacia do Rio Iguaçu, Paraná. Tese de Doutorado, Pós-Graduação em Zoolo-gia, Universidade Federal do Paraná, Brasil, 102pp.Belz, C.E. 2009. Análise de risco aplicada às bioinvasões. p. 229-245. In: Darrigran, G.; Damborenea, C. (Eds). Introdu-ção a Biologia das Invasões. O Mexilhão Dourado na Améri-ca do Sul: biologia, dispersão, impacto, prevenção e controle. Cubo Editora, São Carlos, 246p.Belz, C.E.; Darrigran. G.; Mäder Netto, O.; Ribeiro Junior, P. (no prelo). Analysis of Dispersion Vectors in Inland Waters: The Case of the Invading Bivalves in South America. Journal of Shellfish Research.Bemvenuti, M.A.; Moresco, A. 2005. Peixes: Áreas de banhados e lagoas costeiras do extremo sul do Brasil. Editora ABRH. 63 p. Ben-Ami, F.; Hodgson, A.N. 2005. Ovoviviparity and the structure of the brood pouch in Melanoides tuberculata. (Gas-(Gas-tropoda: Prosobranchia: Thiaridae). Journal of Morphology 263: 322–329.Bendati, M.M. 2000. Avaliação da concentração de metais pe-sados em moluscos no Lago Guaíba. ECOS 17:1-9

391
Bendati, M.M.; Schwarzbach, M.S.R.; Maizonave, C.R.M.; Bringhenti, M.L.; Almeida, L.B. 2003. Avaliação da qualidade da água do Lago Guaíba: subsídios para a gestão da bacia hi-drográfica. Ecos 7:1-34.Benetti, C.J.; Fiorentin, G.L.; Régil Cueto, J.A.; Pacho Miguel, R.R. 1998. Coleopterofauna aquática na Floresta Nacional de Francisco de Paula, RS, Brasil. Acta Biologica Leopoldensia 20:91-101.Benthem-Jutting, W.S.S. 1956. Systematic studies on the non-marine Mollusca of the Indo- Australian archipelago. V – Crit-ical revision of the Javanese freshwater gastropods. Treubia 23:259-477.Berg, D.J.; Fischer, S.W.; Landrum, P.F. 1996. Clearance and processing of algal particles by zebra mussels (Dreissena poly-morpha). Journal of Great Lakes Research 22:779-788.
Bergmann, C.P.; Mansur, M.C.D.; Bergonci, P.E.A.; Pereira, D.; Santos, C.P.; Basegio, T. ; Vicenzi, J.; Santos, S.C.A. 2010a. Seleção de materiais e revestimentos para auxiliar no controle da incrustação do mexilhão dourado na Usina Hidrelétrica de Ibitinga (SP, Brasil). Matéria 15:21-30.
Bergmann, C.P.; Mansur, M.C.D.; Pereira, D.; Bergonci, P.E.A.; Santos, C.P.; Basegio, T. ; Vicenzi, J.; Santos, S.C.A. 2010b. Avaliação de materiais e revestimentos para o contro-le de incrustações do mexilhão dourado Limnoperna fortunei (Dunker, 1857). Biotemas 23:87-100.Bergonci, P.E.A.; Mansur, M.C.D.; Pereira, D.; Santos, C. P. dos. 2009. Population sampling of the golden mussel Lim-noperna fortunei (Dunker, 1857) based on artificial ceramic substrate. Biotemas 22(3): 85-94.Bernard, F.R. 1974. Particle sorting and labial palp function in the pacific oyster Crassostrea gigas (Thunberg, 1795). Biologi-cal Bulletin 146(1):1-10.Berry, A.J.; Kadri, A.H. 1974. Reproduction in the Malayan freshwater cerithiacean gastropod Melanoides tuberculata. Journal of Zoology 172: 369-381.Bielefeld, U. 1991. Histological observation of goands and di-gestive gland in starving Dreissena polymorpha. Malacologia 33:31-42.Bieler, R. 1992. Gastropods phylogeny and systematics. Annu-al Review on Ecology and Systematics 23: 311-338.Biobullet to combat invasive Zebra Mussel. Microencapsula-Microencapsula-tion report. publicado em 09/2009 em: microencapsulation.org/2009/09/biobullets-combat-invasive-zebra-mussel/; acessa-do 09/11/11.Biobullets. Res.: Dr. David Aldridge, MD. Disponível em: <http://www.biobullets.com/>. Acessado em 09/11/11.Boaventura, M.F.; Fernandez, M.A.; Thiengo, S.C.; Silva, R.E.; Melo, A.L. 2002. Formas larvais de Trematoda prove-nientes de gastrópodes límnicos da microrregião Rio de Janei-ro, sudeste do Brasil. Lundiana 3:45-49.Boeger, W.A.; Pie, M.R.; Falleiros, R.M., Ostrensky, A.; Darri-gran, G., Mansur, M.C.D.; Beltz, C.E. 2007. Testing a molecu-Testing a molecu-lar protocol to monitor the presence of golden mussel larvae (Limnoperna fortunei) in plankton samples. Journal of Plank-ton Research 29(11):1015–1019Bogéa, T.; Cordeiro, F.M.; Gouveia, J.S. 2005. Melanoides tuberculatus (Gastropoda:Thiariidae) as intermediate host of Heterophyidae (Trematoda:Digenea) in Rio de Janeiro metro-politan area, Brazil. Revista do Instituto de Medicina Tropical de São Paulo 47:87-90.Boltovskoy, D.; Cataldo, D.H. 1999. Population dynamics of Limnoperna fortunei, an invasive fouling mollusc, in the lower Paraná River (Argentina). Biofouling 14(3):255-263. Boltovkoy, D.; Correa, N.; Cataldo, D.; Stripeikis, J.; Tudino, M. 1997. Environmental stress on Corbicula fluminea (Bival-
via) in the Paraná River delta (Argentina): complex pollution-related disruption of population structures. Archive für Hy-drobiologie 138(4): 483-507.Boltovskoy, D.; Gibbons, M.J.; Hutchings, L.; Binet, D. 1999. General biological features of the South Atlantic. In:Boltovskoy, D. (Eds.). Zooplankton of the South Atlantic.Backhuys Publishers, Leiden.pp. i-xvi+1706.(2 volumes).Boltovskoy, D.; Correa, N.; Cataldo, D.; Sylvester, F. 2006. Dispersion and ecological impact of the invasive freshwater bivalve Limnoperna fortunei in the Río de la Plata watershed and beyond. Biological Invasions 8(4):947-963.Boltovskoy, D.; Sylvester, F.; Otaegui, A.; Leites, V.; Cataldo, D. 2009a. Austral Ecology 34:719–730.Boltovskoy, D.; Karatayev, A.; Burlakova, L.; Cataldo, D.; Karatayev, V.; Sylvester, F.; Mariñelarena, A. 2009b. Signifi-cant ecosystem-wide effects of the swiftly spreading invasive freshwater bivalve Limnoperna fortunei. Hydrobiologia 636:271–284.Bom, 2011. Australian Government Bureau of Meteorology. Southern Oscillation Index (SOI). Disponível em: <http://www.bom.gov.au/climate/current/soi2.shtml>.Bonafé, R.A.; Dias, C.A.; Mata, F.A.R.; Campos, M.C.S. 2010. Uso do antiincrustante MXD-100 para controle do me-xilhão dourado. IV Seminário Brasileiro de Meio Ambiente e Responsabilidade Social do Setor Elétrico.Bonetto, A.A. 1960. Contribución al conocimiento de las glo-chidias del gênero Diplodon y su aplicación a los estúdios sis-temáticos. Actas y Trabajos del Primer Congr. Sud-americano de Zoologia 2:43-59.Bonetto, A.A.; Ezcurra, I.D. 1963. Notas malacologicas. I:(1) El desarrollo del glochidium de Diplodon delodontus delodon-tus (Lam.); (2) El glochidium de Prisodon (Triplodon) corru-gatus Lam.; (3) El lasidium de Monocondylea paraguayana d’Orb. Buenos Aires, Physis 24 (67):17-21.Bonetto, A.A.; Ezcurra, I. D. 1965. Estudio comparado de las formas larvales de Mutelidae Ortmann y su significación siste-mática y zoogeográfica (Moll. Pelecypoda). Anais do Segundo Congr. Latino-Americano de Zoologia 2:55-71.Borcherding, J. 1991. The annual reproductive cycle of the freshwater mussel Dreissena polymorpha Pallas in lakes.Oeco-logia 87:208-218.Botts, P.S.; Patterson, B.A.; Schloesser, D.W. 1996. Zebra mus-sel effects on benthic invertebrates: physical or biotic? Journal of the North American Benthological Society 15:179–184.Bouchet, P.; Rocroi, J.P. 2005. Classification and nomenclator of gastropod families. Malacologia 47:1–397.Branco, S.M. 1986. Hidrobiologia Aplicada à Engenharia Sa-nitária. 3 ed. CETESB, São Paulo, 616p. Brandimarte, A.L.; Shimizu, G.Y.; Anaya, M.; Kuhlmann, M.L. 2004. Amostragem de invertebrados bentônicos. In: Bi-cudo, C.E. de M.; Bicudo, D. de C. (Org.). Amostragem em Limnologia. São Carlos, Rima, p. 213-230. Brandini, F.P.; Lopes, R.M.; Gutseit, K.S.; Spach, H.L.; Sassi, R. 1997. Planctolologia na plataforma continental do Bra-sil. Diagnose e revisão bibliográfica. Rio de Janeiro, MMA--CIRM-FEMAR, 196p.Brandt, R.A. 1974. The non-marine aquatic Mollusca of Thai-land. Archiv für Molluskenkunde 105(1-4):1-423. Brasil. Ministério da Saúde. 2007. Vigilância e controle de moluscos de importância epidemiológica – diretrizes técnicas: Programa de Vigilância e Controle da Esquistossomose (PCE). Ministério da Saúde, Secretaria de Vigilância em Saúde, De-partamento de Vigilância Epidemiológica.– 2. ed. – Brasília : Editora do Ministério da Saúde, 178 p.Breitig, G. 1961. Dreissena polymorpha Pallas Schädling in Kraft-und Industrieanlagen Norddeutschlands und Versuche

392
zuihrer Bekämpfung mit Ultraschal. Verh Internat. Verein. Limnol. 14: 690-694.Britton, J.C.; Morton, B. 1982. A dissection guide, field and laboratory manual for the introduced bivalve Corbicula flu-minea. In: Burch, J.B.; White-Rudolph, J. (eds.). Malacologi-cal Review, (Supplement 3), 82p.Brooks, S.S.; Boulton, A.J. 1991. Recolonization dynamics of benthic macroinvertebrates after artificial and natural distur-bances in an Australian Temporary system. Australian Journal of Marine & Freshwater Researcher 42:295-308. Brugnoli, E.; Clemente, J.; Boccardi, L.; Borthagaray, A.; Scarabino, F. 2005. Golden mussel Limnoperna fortunei (Bivalvia:Mytilidae) distribution in the main hydrographycal basins of Uruguay: updates and predictions. Anais da Acade-mia Brasileira de Ciencias 77(2):235-244.Brugnoli, E.; Lanfranconi, A.; Muniz, P. 2009. El mejillón do-rado, 15 años de invasión en Uruguay: preguntas y respues-tas. Disponível em: <http://limno.fcien. edu.uy/pdf/informes/Lfortunei-Brugnolietal09.pdf>.Brusca, R.C.; Brusca, G.J. 2007. Invertebrados. 968 pp. Rio de Janeiro: Guanabara Koogan.Bull, L.S.; Courchamp, F. 2009. Management of interacting invasives: ecosystem approaches. Em: Clout, M.N; Williams, P.A. (Eds.) 2009. Invasive Species Management. A Handbook of Principles and Techniques. Oxford University Press, New York. 308p.Burgmann, H.; Pesaro, M.; Widmer, F.; Zeyer, J. 2001. A strategy for optimizing quality and quantity of DNA extracted from soil. Journal of Microbiological Methods 45:7-20.Burns, M.D.M. 2010. Consequências da barragem eclusa do canal São Gonçalo para a ictiofauna do sistema Patos - Mirim, Pós-graduação em Oceanografia biológica. Universidade Fede-ral do Rio Grande, Rio Grande, p. 143.Burns, M.D.M.; Garcia, A.M.; Bemvenuti, M.A.; Vieira, J.P.; Marques, D.M.L.M.; Moresco, A.; Condini, M.V.L. 2006a. Bivalvia, Mytilidae, Limnoperna fortunei: Distribution exten-sion. Check list 2(2): 41-43.Burns, M.D.M.; Geraldi, R.M.; Garcia, A.M.; Benvenu-ti, C.E.; Capítoli, R.R.; Vieira, J.P. 2006b. Primeiro registro de ocorrência do mexilhão dourado Limnoperna fortunei (Dunker, 1857) na bacia de drenagem da Lagoa Mirim, RS, Brasil. Biociências 14(1): 83-84.Calazans, C.S.H.; Fernandes, F.C.; Fernandes, L.V.G.; Santos, O.C.; Souza, T.O.J; Patrício, F.C.; Fróes, C.L. 2007. Morta-Morta-lidade do mexilhão dourado exposto à KCl, NaCl e ao KA-VISTÒ. Painel apresentado no II Encontro Sul-Americano de Integração de Ações para o Controle do Mexilhão Dourado – SULMEX.Calazans, C.S.H.; Fernandes, F.D.; Aldridge, D.; Rebelo, M.D. 2012. Assessment of dissolved and microencapsulated biocides toxicity for the control of golden mussel Limnoperna fortunei infestation. Magazine Review 1-11.Calheiros, D.F.; Hamilton, S.K. 1998. Limnological conditions associated with natural fish kills in the Pantanal Wetland of Brazil. Verhandlungen des Internationalen Verein Limnologie26: 2189–2193.Calijuri, M.C.; Dos-Santos, A.C.A.; Jati, S. 2002. Tempo-Tempo-ral changes in the phytoplankton community structure in a tropical and eutrophic reservoir (Barra Bonita, SP – Brazil). J. Plankton Res., 4:617-634.Callil, C. T. 2003. Base de dados direcionada a elaboração de um programa de monitoramento de águas continentais utili-zando moluscos bivalves. 227f. Tese – Pós-graduação em bio-ciências – Pontifícia Universidade Católica do Rio Grande do Sul – PUCRS, Porto Alegre. Callil, C.T. 2009. Influence of environmental factors on re-
production of two sympatric species of Anodontites (Myceto-podidae: Unionoida) in the center of south America. Abstracts Book of the International Symposium of Freshwater Mollusk Conservation Society. April, 19-24. Baltimore, Maryland.Callil, C.T.; Cruz, R.F. 2009. Macroinvertebrados bentônicos como indicadores de alterações ambientais. p. 84-94. In: Fi-gueiredo, D.M; Salomão, F.X.T. (Eds). Bacia do Rio Cuiabá: uma abordagem socioambiental. EdUFMT, Cuiabá, Brasil, 216p.Callil, C.T.; Junk, W.J. 1999. Concentração e incorporação de mercúrio por Moluscos Bivalves (Anodontites trapesialis La-marck, 1819 e Castalia ambigua Lamark, 1819) do Pantanal de Poconé - MT, Brasil. Biociências 7(1):3-28.Callil, C.T.; Mansur, M.C.D. 2002. Corbiculidae in the Panta-nal: history of invasion in southeast and central South America and biometrical data. Amazoniana 17(1-2):153-167. Callil, C.T.; Mansur, M.C.D. 2007. Gametogênese e dinâmica da reprodução de Anodontites trapesialis (Lamarck) (Unionoi-da, Mycetopodidae) no Lago Baía do Poço, planície de inun-dação do Rio Cuiabá, Mato Grosso, Brasil. Revista Brasileira de Zoologia 24(3):825-840.Callil, C.T.; Mansur, M.C.D.; Marcelo, S.M. 2006. Bivalves invasores no Pantanal In: Malacologia Brasileira – Palestras do XVIII EBRAM, Sociedade Brasileira de Malacologia Rio de Janeiro, p. 87-100.Callil, C.T.; Gomes, A.L.T.; Pinillos, A.C.; Takeda, A.M. 2012. Variation in gametogenesis and spawning of Limnoperna for-tunei (Bivalvia) in the alluvial floodplain of the upper Paraná River (Brazil). International Review of Hidrobiology. In press.Campos, S.H.C. 2009. Avaliação de biocidas dissolvidos e mi-croencapsulados para o controle do mexilhão dourado Lim-noperna fortunei (Dunker, 1857). Universidade Federal do Rio de Janeiro (UFRJ), Mestrado em Ciências Biológicas (Biofísi-ca), Rio de Janeiro, 63p.Cantanhêde, G.; Hahn, N.S.; Gubiani, E.A.; Fugi, R. 2008. Invasive molluscs in the diet of Pterodoras granulosus (Valen-ciennes, 1821) (Pisces, Doradidae) in the Upper Paraná River floodplain, Brazil. Ecology of Freshwater Fish 17: 47–53.Capítoli, R.R.; Bemvenuti, C.E. 2004. Distribuição do mexi-Distribuição do mexi-lhão dourado Limnoperna fortunei (Dunker, 1857) na área estuarina da Lagoa dos Patos e Canal São Gonçalo. Anais do VI Simpósio de Ecossistemas Brasileiros. Publicações ACIESP 110(1): 98-107.Capítoli, R.R.; Colling, L.A.; Bemvenuti, C.E. 2008. Cenários de distribuição do mexilhão dourado Limnoperna fortunei (Mollusca - Bivalvia) sob distintas condições de salinidade no complexo Lagunar Patos-Mirim, RS - Brasil. Atlântica 30:35-44.Caprari, J. 2006. Pinturas antiincrustantes. In: Darrigran, G.; Damborenea, C. (Eds.). Bio-invasión del mejillón dorado en el Continente Americano. 1. ed., capítulo 12, Buenos Aires, Argentina, Universidade Nacional de La Plata, Edulp, p. 181-211.Caprari, J.; Lecot, C. 2001. El control de bivalvos invasores Limnoperna fortunei (Dunker, 1857) en la central hidroe-lectrica Yaciretá mediante pinturas. In: Actas do Seminario Internacional sobre Gestión Ambiental e Hidroelectricidad. Yaciretá, p.1-13.Caraco, N.F.; Cole, J.J., Raymond, P.A.; Strayer, D.; Pace, M.L; Findlay, S.E.; Fischer, D.T. 1997. Zebra mussel invasion in a large, turbid river: phytoplankton response to increased grazing. Ecology 78:588-602.Carlton, J.T. 1999. The scale and ecological consequences of biological invasions in the World’s oceans. p.195-212. In: Sandlund, O.T.; Schei, P.J.; Viken, A. (Eds). Invasive Species and Biodiversity Management. Kluwer Academic Publishers, Netherlands, 435p.

393
Carlton, J.T.; Geller, J.B. 1993. Ecological roulette: biological invasions and the global transport of nonindigenous marine organisms. Science 261: 78-82.Carlton, J.T.; Ruiz, G.M. 2004. Vector science and integrated vector management in bioinvasion ecology: conceptual frame-works. In: Money, H.A.; McNeely, J.; Neville, L.E.; Schei, P.J.; Waage J.K. (Eds). Invasive Alien Species: A New Synthesis. Is-land Press, Covelo California.Carmichael, W. W. 1992. Cyanobacteria Secondary Me-tabolites - the Cyanotoxins. Journal of Applied Bacteriology 72:445-459.Carmichael, W.W.; Azevedo, S.; An, J.S.; Molica, R.J.R.; Jo-chimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesh-am, G.K. 2001. Human fatalities from cyanobacteria: chemi-cal and biological evidence for cyanotoxins. Environmental Health Perspectives 109: 663-668.Carvalho, G.P.; Cavalcante, P.R.S.; Castro, A.C.L.; Rojas, M.O.A.I. 2000. Preliminary assessment of heavy metal levels in Mytella falcata (Bivalvia Mytilidae) from Bacanga River Es-tuary, São Luiz, State of Maranhão, Northeastern of Brazil. Revista Brasileira de Biologia 60(1):11-16.Carvalho, O.; Jannotti-Passos, L.K.; Mendonça, C.L.F.G.; Cardoso, P.C.M.; Caldeira, R.L. 2005. Moluscos de impor-tância médica no Brasil. Centro de Pesquisas René Rachou – Fiocruz, Série Esquistossomose no 7. 52p.Cataldo, D.; Boltovskoy, D. 1999. Population dynamics of Corbicula fluminea (Bivalvia) in the Paraná River Delta, Ar-gentina. Hydrobiologia 380:153-163.Cataldo, D.; Boltovskoy, D. 2000. Yearly reproductive activity of Limnoperna fortunei (Bivalvia) as inferred from the occur-rence of its larvae in the plankton of the lower Paraná River and the Rio de la Plata estuary (Argentina). Aquatic Ecology 34:307–317.Cataldo, D.; Boltovskoy, D.; Stripeikis, J.; Pose, M. 2001. Condition index and growth rates of field caged Corbicula fluminea (Bivalvia) as biomarkers of pollution gradients in the Paraná river delta (Argentina). Aquatic Ecosystem Health and Management 4:187-201.Cataldo, D.; Boltovskoy, D.; Pose, M. 2002a. Control del mo-lusco incrustante Limnoperna fortunei mediante el agregado de moluscicidas al agua. In: Tercera jornada sobre conserva-ción de la fauna íctica en el Rio Uruguay, Paysandu, Uruguay.Cataldo, D.; Boltovskoy, D.; Marini, V.; Correa, N. 2002b. Limitantes de Limnoperna fortunei na bacia do Prata: a preda-ção por peixes. Tercera Jornada sobre Consevación de la fauna íctica en el Rio Uruguay. Paysandú, Uruguay, 25 a 26 de abril.Cataldo, D.; Boltovskoy, D.; Hermosa, J.; Canzi, C. 2005a. Temperature-dependent rates of larval development in Lim-noperna fortune (Bivalvia: Mytilidae). Journal of Molluscan Studies 71(1):41-46.Cataldo, D.; O’Farrel, I.; Paolucci, E.; Sylvester, F.; Boltovs-koy, D. 2005b. Efectos de Limnoperna fortunei sobre el fito-plancton y los nutrientes. Tercer Congreso Argentino de Lim-nología, Chascomús, 30 October 2 November.Cazzaniga, N.J. 1997. Asiatic clam, Corbicula fluminea, rea-ching Patagonia (Argentina). Journal of Freshwater Ecology 12:629-630.CEFET (Centro Federal de Educação Tecnológica de Minas Gerais). 2006. Avaliação de corrosão ASTM Copper Strip Corrosion. Standards: Método ASTM D130/IP154.Ceni, G. 2010. Distribuição da ictiofauna por zonas de pro-fundidade na Lagoa Mirim, RS, Brasil: um estudo de caso. Pós-graduação em Biologia de Ambientes Aquáticos Continen-tais. Universidade Federal do Rio Grande, Rio Grande.CETEC (Fundação Centro Tecnológico de Minas Gerais). 2007. Ensaio de ecotoxicidade aguda com Daphnia similis,
conforme Associação Brasileira de Normas Técnicas ABNT (1993). Certificado número 401980 de 12.07.2007.Chalker-Scott, L.; Scalia, J.; Titus, J. 1994. Influence of wide-range ultraviolet radiation upon behavoir and mortality of Dreissena polymorpha. In: Proceedings of The Fourth Inter-national Zebra Mussel Conference. Wisconsin Sea Grant insti-tute, University of Wisconsin, Madison, 161-177. Chaube, H.S.; Pundhir, V.S. 2005. Crop diseases and their management. PHI Learning Pvt. Ltd. 703p.Choi, S.S.; Kim, J.S. 1985. Studies on the metamorphosis and the growth of larvae in Limnoperna fortunei. Korean Journal of Limnology 1:13-18.Choi, S.S.; Shin, C.N. 1985. Studies on the early development and larvae of Limnoperna fortunei. Korean Journal of Mala-cology 1:5-12. (in Korean).Chung, P.R.; Jung, Y.; Park, Y. K.; Hwang, M.G.; Soh, C.T.I. 2001. Corbicula fluminea (Bivalvia: Corbiculidae): a possible second molluscan intermediate host of Echinostoma cinetor-chis (Trematoda: Echinostomatidae) in Korea. Korean Journal of Parasitology 39 (4):329-332.Clasen, T.; Edmondson, P. 2006. Sodium dichloroisocyanurate (NaDCC) tablets as an alternative to sodium hypochlorite for the routine treatment of drinking water at the household leve International. Journal of Hygiene and Environmental Health, 209, 173-181.Claxton, W.T.; Boulding, E.G. 1998. A new molecular techni-que for identifying field collections of zebra mussel (Dreissena polymorpha) and quagga mussle (Dreissena bugensis) veliger larvae applied to eastern Lake Erie, Lake Ontario, and Lake Simcoe. Canadian Journal of Zoology 76: 194-198.Claxton, W.T.; Mackie, G.L. 1998. Seasonal and depth varia-tions in gametogenesis and spawning of Dreissena polymorpha and Dreissena bugensis in Eastern Lake Erie.Canadian Journal of Zoology, 76:,2010–2019.Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. (Eds). 1995. Stan-dard Methods for the Examination of Water and Wastewater. American Public Health Association, Washington, DC, USA, 20th Edn.Coblentz, B.E. 1990. Exotic organisms: a dilemma for Conser-vation Biology. Conservation Biology 4:261-265.Coe, W.R. 1943. Development of the Primary Gonad and Dif-ferentiation of Sexuality in Teredo navalis and other Pelecypod Mollusks.Biological Bulletin 84:178-186.Coelho, R.M.P. 2004. Métodos de coleta, preservação, con-tagem e determinação de biomassa em zooplâncton de águas continentais. p. 149-166. In: Bicudo, C.E.M; Bicudo, D.C. Amostragem em Limnologia. Ed. RiMa, 371p. Cohen, A.N.; Carlton, J.T. 1998. Accelerating invasion rate in a highly invaded estuary. Science 279:555-558.Colares, E.R.C.; Bendati, M.M.A.; Suminsky, M. 2002a. Im-pactos da presença do mexilhão dourado em sistemas de abas-tecimento de água. ECOS 22:30-35.Colares, E.R.C.; Suminsky, M.; Bendati, M.M.A. 2002b. Diagnóstico e controle do mexilhão dourado, Limnoperna fortunei, em sistemas de tratamento de água em Porto Alegre (RS, Brasil). ABES, Porto Alegre, 4p.Colgan, D.J.; Ponder, W.F.; Eggler, P.E. 2000. Gastropod evo-lutionary rates and phylogenetic relationships assesses using partial 28S rDNA and historie H3 sequences”. Zoological Scripta 29: 29-63.Colgan, D.J.; Ponder, W.F.; Beacham, E.; Macaranas, J.M. 2003. Gastropod phylogeny based on six segments from four genes representing coding and non-coding and mitochondrial or nuclear DNA. Molluscan Research 23: 123-148.CONAMA. Resolução no 357, de 17 de março de 2005. Dis-põe sobre a classificação dos corpos de água e diretrizes am-

394
bientais para o seu enquadramento, bem como estabelece as condições e padrões de lançamento de efluentes, e dá outras providências. Diário Oficial da República Federativa do Brasil, Brasília, DF, 23p.Conde, D.; Paradiso, M.; Gorga, J.; Brugnoli, E.; De Leon, L.; Mandiá, M. 2002. Problemática de la calidad de água em el sistema de grandes embalses Del Rio Negro (Uruguay). Revista CIER 39: 51-68. CONSEMA. 2006. Resolução nº 128, de 24 de novembro de 2006. Disponível em: <http://www.mp.rs.gov.br/areas/am-biente/arquivos/boletins/bola_leg08_06/iig128.pdf> Acesso em: 19/agosto/ 2008.Coradin, L. 2006. Espécies exóticas invasoras: situação bra-sileira. Brasília: Ministério do Meio Ambiente, Secretaria de Biodiversidade e Florestas, 24p.Correio do Povo. 2007a. Barra do Quaraí – Mexilhão doura-do chega à região. Porto Alegre, 31 de janeiro de 2007, p.16.Correio do Povo. 2007b. Uruguaiana – Mexilhões dourados no arroio Toro Passo. Porto Alegre, 13 de março de 2007.Costa, R. 2008. Improved solutions for zebra mussel control. Ph D. Thesis. University of Cambridge.Costa, R.; Elliott, P.; Saraiva, P.M.; Aldridge, D.; Moggridge, G.D. 2008. Development of sustainable solutions for zebra mussel control through chemical product engineering, Chinese Journal of Chemical Engineering 16(3):435-440.Coughlan, J. 1969. Estimation of filtering rate from clearance of suspensions. Marine Biology 2:356-359.Counts, C.L. III. 1986. The Zoogeography and History of the invasion of the United States by Corbicula fluminea (Bivalvia: Corbiculidae). Proceedings of the Second International Cor-bicula Symposium. American Malacological Bulletin, Special edition (2):7-39.Cowie, R. H. 2004. Disappearing snails and alien invasions: the biodiversity/conservation interface in the pacific. Journal of Conchology. Especial Publication 3: 23-37.Cowie, R.H.; Robinson, D.G. 2003. Pathways of introduction of nonindigenous land and freshwater snails and slugs, pp. 93-122. In: Ruiz, G.M.; Carlton, J.T. Invasive species: vectors and management strategies. Island Press, Washington, 484p.Cox, L.R. 1969. General features of Bivalvia. Part N. Mol-lusca. p. N3-Nl29. In: Moore, R.C. (Ed.). Treatise on inver-tebrate paleontology. Vol. 1. Geological Society of America, Boulder, Colorado.Crosier, D.M.; Molloy, D.P. 2007. Management and Control. Dis-ponível em < http://el.erdc.usace.army.mil/zebra/zmis/zmishelp/management_and_control.htm>. Acesso em 25 agosto 2011.Crump, K. S.; Hoel, D.G.; Langley, C.H.; Peto, R. 1976. “Fun-damental Carcinogenic Processes and Their Implications for Low Dose Risk Assessment”. Cancer Research 36: 2973–2979.Cucci, T.L.; Shummay, S.E.; Newell, R.C.; Selvin, R.; Guil-lard, R.R.L.; Yentsch, C.M. 1985. Flow cytometry – a new method for characterization of differential ingestion, digestion and egestion by suspension feeders. Marine Ecology Progress Series 24:201-204. Daane, K.; Mills, N.J. 2009. History of Biological Control. Disponível em: <http://nature.berkeley.edu/biocon/BC%20Class%20Notes/6-11%20BC%20History. pdf>.Acesso em: 13 outubro de 2011.Damborenea, C.; Penchaszadeh, P.E. 2006. Biología repro-ductiva de Limnopernafortunei. p.69-82. In: Darrigran, G.; Damborenea, C. (Eds). Bio-invasión del mejillón dorado en el continente americano. Edulp, La Plata, Argentina, 219p.Daoust, S.P.; Mader, B.J.; Maure, F.; Mclaughlin, J.D.; Tho-mas, F.; Rau, M.E. 2010. Experimental evidence of size/age-biased infection of Biomphalaria glabrata (Pulmonata: Pla-norbidae) by an incompatible parasite species: Consequences
for biological control. Infection, Genetics and Evolution 10: 1008-1012.Darrigran, G. 1991. Competencia entre dos especies de pe-lecipodos invasores, Corbicula fluminea (Muller, 1774) y C. largillierti (Philippi, 1844) en el litoral argentino del estuario del Rio de la Plata. Biología Acuática 15 (2):214-215. Darrigran, G. 2002. Potential impacto of filterfeeding invad-ers on temperate inland freshwater environments. Biological Invasions 4:145-156. Darrigran, J. 2009. Governabilidade dos recursos hídricos e as bioinvasões. p. 1-29. Em: Darrigran G.; Damborenea. C. (Eds) Introdução a Biologia das Invasões. O mexilhão doura-do na América do Sul: biologia, dispersão, impacto, prevenção e controle. Cubo Editora. São Carlos, 246p.Darrigran, G.; Armengol, M.F.L. 1998. Distribucion, estruc-tura y composicion de la malacofauna presentes en substrato duro de la costa argentina del Rio de La Plata. Gayana Zoo-logía 62(1):79-89. Darrigran, G.; Damborenea, C. 2001. Concentraciones letales de un biocida para adultos del molusco invasor Limnoperna fortunei (Mytilidae). ACTAS Seminario Internacional sobre Gestión Ambiental e Hidroelectricidad, 19 al 22 de septiem-bre de 2001 – Complejo Hidroeléctrico de Salto Grande 119.Darrigran, G.; Damborenea, C. 2005. A South American bioin-vasion case history: Limnoperna fortunei (Dunker, 1857), the golden mussel. American Malacological Bulletin 20:105–112.Darrigran, G.; Damborenea, C. 2006a. (Eds.). Bio-invasión del mejillón dorado en el continente americano. EDULP, La Plata, Argentina. 218 p.Darrigran, G.; Damborenea, C. 2006b. Aspectos generales vinculados a la prevención y control. p. 154- 167. In: Darri-gran, G.; Damborenea, C. (Eds.) Bio-invasión del mejillón do-rado en el continente americano. Edulp, La Plata, 2006. 220 p.Darrigran, G.; Damborenea, C. 2009a. Bioinvasões. p.1-29. In: Darrigran, G.; Damborenea, C. (Eds). Introdução à Bbio-logia das Invasões. O mexilhão dourado na América do Sul: biologia, dispersão, impacto, prevenção e controle. Editora Cubo Multimídia, São Carlos, 245p.Darrigran, G.; Damborenea, C. 2009b. Introdução à Biologia das Invasões. O mexilhão dourado na América do Sul: biolo-gia, dispersão, impacto, prevenção e controle. Cubo Multimí-dia, São Carlos, 260 p.Darrigran, G.; Damborenea, C. 2009c. Aspectos gerais vincu-lados à prevenção e controle. p.155-167. In: Darrigran, G.; Damborenea, C. (Eds). Introdução à Biologia das Invasões. O mexilhão dourado na América do Sul: biologia, dispersão, im-pacto, prevenção e controle. Cubo Editora, São Carlos, 246p.Darrigran, G.; Damborenea, C. 2011. Ecosystem Engineering Impact of Limnoperna fortunei in South America. Zoological Science 28(1):1-7.Darrigran, G.; Darrigran, J. 2001. El mejillón dorado: una obstinada espécie invasora. Ciencia Hoy 61:20-23. Buenos Aires.Darrigran, G.; Ezcurra-de-Drago, I. 2000a. Invasion of the exotic freshwater mussel Limnoperna fortunei (Dunker, 1857) (Bivalvia: Mytilidae) in South America. The Nautilus, 114(2):69-73.Darrigran, G.; Ezcurra-de-Drago, I. 2000b. Distribución de Limnoperna fortunei (Dunker, 1857) (Mytilidae), en La Cuenca Del Plata, Region Neotropical. Medio Ambiente 13(2):75-79.Darrigran, G.; Guimarey, P. (no prelo). Bio Invasiones en el Sur del Cono Sur. Una aproximación de las invasiones de los moluscos acuáticos.In: Santos, S. B. dos; Pimenta, A.D.; Fer-nandez, M.A.; Thiengo, S.C. (Org.). Tópicos em Malacologia – Ecos do XIX Encontro Brasileiro de Malacologia - Fiocruz, ICICT Rio de Janeiro, Brasil (2008).

395
Darrigran, G.; Mansur, M.C.D. 2006. Distribución, abundan-2006. Distribución, abundan-cia y dispersión, pp. 93-110. In: Darrigran, G.; Damborenea, C. (Eds.). Bio-invasión del mejillón dorado en el continente americano. Edulp, La Plata. 220p. Darrigran, G.; Mansur, M.C.D. 2009. Introdução e dispersão do Limnoperna fortunei. p.89-109 In:Darrigran, G.; Dambo-renea, C. (Eds). Introdução à Biologia das Invasões. O mexi-lhão dourado na América do Sul: biologia, dispersão, impacto, prevenção e controle. Cubo Editora. São Carlos, 246p.Darrigran, G.; Pastorino, G. 1995. The recent introduction of a freshwater asiatic bivalve, Limnoperna fortunei (Mytilidae) into South America. The Veliger 38(2):171-175.Darrigran, G.; Pastorino, G. 2000. Distribution of the golden mussel Limnoperna Fortunei (Dunker, 1857), after 10 years of American invasion. In: World Congress of Malacology, Vi-enna. Darrigran, G.; Pastorino, G. 2003. The golden mussel, Lim-noperna fortunei (Dunker, 1857) (Bivalvia, Mytilidae) in the neotropical region: a 10 year story of invasion. Tentacle 11: 8-9.Darrigran, G., Martin, S.M., Gullo, B., Armendariz, L. 1998a. Macroinvertebrates associated with Limnoperna fortunei (Dunker, 1857) (Bivalvia, Mytilidae) in Río de la Plata, Argen-tina. Hydrobiologia 367:223-230.Darrigran G.; Damborenea C.; Penchaszadeh, P. 1998b. A case of hermaphroditism in the freshwater invading bivalve Limnoperna fortunei (Dunker, 1857) (Mytilidae) from its first American habi-tat (Río de la Plata, Argentina).Iberus 16(2):99-104.Darrigran, G.; Penchaszadeh, P.; Damborenea, C. 1999. The reproductive cycle of Limnoperna fortune (Dunker, 1857) (Mytilidae) from a Neotropical Temperate Locality. Jounal of Shellfish Research, 18(2):361-365.Darrigran, G.; Penchaszadeh, P.; Damborenea, C.; Greco, N. 2002. Abundance and distribution of the golden mussel (Limnoperna fortunei) larvae in a Hydroeletric Power Plant in South America. In: Proceeding of the 11th International Aquatic Nuisance Species and Zebra Mussel Conference. Al-exandria, Virginia, USA, 312-316.Darrigran, G.; Damborenea, C.; Penchaszadeh, P.; Taraborelli, C. 2003. Adjustments of Limnoperna fortune (Bivalvia: Myti-lidae) after ten years of invasion in Americas. Journal of Shell-fish Research 22(1):141-146.Darrigran, G.; Damborenea, C.; Greco, N. 2007. Freshwater invasive bivalves in man-made environments: A case study of larvae biology of Limnoperna fortunei in a Hydroelectric Pow-er Plant in South America. AMBIO 36(7):575-579.Darrigran, G.; Vilches, A.; Legarralde, T. 2008. Desinterés del Pasado, Decisiones del Futuro: Educación para prevenir las in-vasiones biológicas. Revista Educación en Biología 11(1):39-52.Darrigran, G.; Boeger, W.; Damborenea, C.; Moroñas, M. 2009. Evaluation of sampling and analysis techniques for eraly detection of Limnoperna fortunei (Mytilidae) in limit areas of its distribution. Brazilian Journal of Biology 69(3): 979-980.David, K.A.; Davis, B.M.; Hunter, R.D. 2009. Lake St. Clair zooplankton: evidence for post-Dreissena changes. Journal of Freshwater Ecology 24 (2):199-209.Davis, M.A. 2006. Invasion Biology 1958-2005: the Pursuit of Science and Conservation. p.35-64. In: Cadotte, M.W.; Mc-Mahon, S.M.; Fukami, T. (Eds). Conceptual Ecology and In-vasions Biology: Reciprocal Approaches to Nature. Springer, London, UK.Deaton, L.E.; Derby, J.G.S.; Subhedar, N.; Greenberg, M.J. 1989. Osmoregulation and salinity tolerance in two species of bivalve mollusk: Limnoperna fortunei and Mytilopsis leuco-phaeta. Journal of Experimental Marine Biology and Ecology 133: 67-79.
Demott, W.R. 1999. Foraging strategies and growth inhibition in five daphnids feeding on mixtures of a toxic cyanobacteria and a green alga. Freshwater Biology 42: 263-274.Dermott, R.; Kerec, D. 1997. Changes to the deepwater benthos of eastern Lake Erie since the invasion of Dreissena: 1979–1993. Canadian Journal of Fisheries and Aquatic Scien-ces 54:922–930.Derraik, J.G.B. 2008. The potential significance to human he-alth associated with the establishment of the snail Melanoides tuberculata in New Zeland. The New Zeland Medical Journal 121(1021):25-32.Di Castri, F. 1989. History of biological invasions with special emphasis on the old world. p.130. In: Drake, J.A.; Mooney, H.A.; Castri, F. di; Groves, R.H.; Kruger, F.J.; Rejmanek, M.; Williamson, M. (Eds). Biological invasions: a global perspecti-ve. John Wiley, New York.Díaz, M. T; Hernandez, L. E.; Bashirullah, A. K. 2008. Studies on the life cycle of Haplorchis pumilio (Looss, 1896) (Trema-toda: Heterophyidae) in Venezuela. Revista Científica 18: 35-42.Dietz, T.H.; Lessard, L.J.W.; Silverman, H. 1994. Osmoregu-Osmoregu-lation in Dreissena polymorpha: The importance of Na, Cl, K and particularly Mg. Biological Bulletin 187:76-83.Diggins, T.P.; Weimer, M.; Stewart, K.M.; Baier, R.E.; Meyer, A.E., Forsberg, R.F.; Goehle, M.A. 2004. Ephiphytic refugi-um: are two species of invading freshwater bivalves partition-ing spatial resources? Biological Invasions 6: 83-88. Dillon, R.T. Jr.; Wethington, A.R.; Rhett, J.M.; Smith, T.P. 2002. Populations of the European freshwater pulmonate Phy-sa acuta are not reproductively isolated from American Phy-sa heterostropha or Physa integra. Invertebrate Biology 121: 226−234.Dionisio-Pires, L.M.; Van Donk, E. 2002. Comparing grazing by Dreissena polymorpha on phytoplankton in the presence of toxic and non-toxic cyanobacteria. Freshwater Biology 47:1855-1865.Dionisio-Pires, L.M.; Jonker, R.R.; Van Donk, E.; Laanbroek, H.J. 2004. Selective grazing by adults and larvae of the zebra mussel (Dreissena polymorpha): application of flow cytometry to natural seston. Freshwater Biology 49:116–126.Doherty, F.G.; Cherry, D.S.; Cairns J. JR. 1986. Control of the freshwater fouling bivalve Corbicula fluminea by haloge-nation. Archives of Environmental Contamination and Toxi-cology 15:535–542.Donskoy, D.M.; Ludyanskiy, M. 1995. Low Frequency Sound as a Control Measure for Zebra mussel Fouling. Proceedings of The Fifth International Zebra mussel and Others Aquatic Nuisance Organisms Conference p. 103-108. Toronto, Canada. Doudoroff, P.; Katz, M.1953. Critical review of literature on the toxicity of industrial wastes and their components to fish. Sewage and Industrial Wastes 25: 802-839.Drake, J.M.; Bossenbroek, J.M.; 2004. The potential distribu-tion of zebra mussels in the United States. BioScience 54(10): 931-941.Dudgeon, D. 1982. An investigation into some physical and biotic effects of flooding on reservoir mud previously subjec-ted to a period of aerial exposure. Hydrobiologia 97:27-35.Dudgeon, D. 1986. The life cycle, population dynamics and productivity of Melanoides tuberculata (Müller, 1774) (Gas-tropoda: Prosobranchia: Thiaridae) in Hong Kong. Journal of Zoology 208: 37-53.Duffy, M.A.; Perry, L.J. C.; Kearns, M.; Weider, L.J.; Hairston, N.G. 2000. Paleogenetic evidence for a past invasion of Onon-daga Lake, New York, by exotic Daphinia curvirostris using mtDNA from dormant eggs. Limnology and Oceanography 45:1409-1414.

396
Duggan, I.C. 2002. First record of a wild population of the tropical snail Melanoides tuberculata in New Zealand natural waters. New Zealand Journal of Marine and Freshwater Re-search 36:85-89.Dundee, D.S.; Paine, A. 1977. Ecology of the snail Melanoides tuberculata (Müller), intermediate host of human liver fluke (Opisthorchis sinensis) in New Orleans, Lousiana. The Nau-tilus 91 (1):17-20.Eilenberg, J.; Hajek, A.; Lomer, C. 2001. Suggestions for uni-fying the terminology in biological control. BioControl 46: 387-400.Elkarmi, A.Z.; Ismail, N.S. 2007. Growth models and shell morphometrics of tow populations of Melanoides tuberculata (Thiaridae) living in hot springs and freshwater pools. Journal of Limnology 66 (2):90-96.Elton, C.S. 1958. The Ecology of Invasions by animals and plants. Catalogue 6041/U. Methuen and CO LTD. UK, 181p.Endo, N.; Sato, K.; Nogata, Y. 2009. Molecular based method for the detection and quantification of larvae of the golden mussel Limnoperna fortunei using real-time PCR. Plankton & Benthos Research 4:125-128.Endo, N.; Sato, K.; Matsumura, K.; Yoshimura, E.; Odaka, Y.; Nogata, Y. 2010. Species-specific detection and quantifica-tion of common barnacle larvae from the Japanese coast using quantitative real-time PCR. Biofouling 13:901- 911.Entezari, M.H.; Nazary, S.H.; Khodaparast, M.H.H. 2004. The direct effect of ultrasound on the extraction of ultrasound on the extraction of date syrup and and its micro-organisms. Ultrasonics Sonochemistry 11(6): 379-384. EPA. 1986. Guidance for registration of pesticide products containing copper sulfate. Office of Pesticide Programs, Whashington. G-31.Eseerco (Empire State Eletric Energy Research Corporation). 1992. Evaluation of Ultrasound for Zebra Mussel Mitigation. Research Report 91: 17.Eskelson, M.J.; Chapman, E.G.; Archbold, D.D.; Obrycki, J.J.; Haewood, J.D. 2011. Molecular identification of preda-tion by carabid beetles on exotic and native slugs in a straw-berry agroecosystem. Biological Control 56: 245-253.Espinola, L.A.; Ferreira Julio Junior, H. 2007. Invader species: concepts, models and attributes. INCI 32(9):580-585.Espinosa, E.P.; Allam, B.; Ford, S.E. 2008. Particle selection in the ribbed mussel Geukensia demissa and the Eastern oyster Crassostrea virginica: effect of microalgae growth stage. Estu-arine, Coastal and Shelf Science 79:1-6. Esteves, F.A. 1988. Fundamentos de Limnologia. Ed. Interci-ência, 575p. Fachini, A.; Gazulha, V.; Pedrozo, C.S. 2011. Filtração do bi-valve invasor Limnoperna fortunei (Dunker, 1857), o mexi-lhão dourado, sobre a comunidade planctônica natural e na presença de cianobactéria tóxica. Dissertação de Mestrado, Programa de Pós-Graduação em Ecologia, Universidade Fede-ral do Rio Grande do Sul, 68p.Facon, B.; Pointier, J.P.; Glaubrecht, M.O.; Poux, C.; Jarne, P.; David, P. 2003. A molecular phylogeography approach to bio-A molecular phylogeography approach to bio-logical invasions of the New World by parthenogenetic thiarid snails. Molecular Ecology 12:3027-3039.Facon, B.; Machline, E.; Pointier, J. P.; David, P. 2004. Varia-tion in desiccation tolerance in freshwater snails and its conse-quences for invasion ability. Biological Invasions 6:283-293.Falconer, I.R. 1991. Tumor promotion and liver-injury caused by oral consumption of cyanobacteria. Environmental Toxi-cology and Water Quality 6:177-184.Falk-Petersen, J.; Bohn, T.; Sandlund, O.T. 2006. On the numerous concepts in invasion biology. Biological Invasions 8:1409-1424.
FAO, 1972. Desarrollo de la Cuenca Hidrografica de la Lagu-na Merín. 86p.Faria, E.A.; Branco, J.R.T.; Campos, M.C.S.; Oliveira, M.D.; Rolla, M.E. 2006. Estudo das características antiicrustantes de materiais. Revista da Escola de Minas 59(2):233-238.Fernandes, F.C.; Coutinho, R. 2008. Programa para controle do mexilhão dourado, Limnoperna fortunei (Dunker, 1857), nas águas juridiscionais brasileiras (CT-HIDRO/CNPq). Rio de Janeiro: Instituto de Estudos do Mar Almirante Paulo Mo-reira (IEAPM), p. 381.Fernandes, F.C.; Campos, S.H.C; Plastina, A. 2009. Estrutura de prevenção e controle. p. 397-423. In: MMA (Ministério do Meio Ambiente). Informe sobre as espécies exóticas invasoras marinhas do Brasil. Série Biodiversidade, 33. MMA, Brasília – DF, Brasil, 439p.Fernandes, L.V.G. 2009. Estudo da formação de subprodutos halogenados em águas e fluvias tratadas com dicloroisocianu-rato e hipoclorito de sódio para controle do mexilhão doura-do (Limnoperna fortunei). Dissertação de Mestrado, Pós-Gra-duação Química, Universidade Federal do Rio, Brasil, 77p.Fernandez, M.A. 2011. Variação espaço-temporal da malacofauna límnica em dois reservatórios no rio Tocantins, com ênfase na transmissão da esquistossomose, no período entre junho de 2004 e outubro de 2010. Tese de Doutora-. Tese de Doutora-do, Pós-graduação em Biologia Parasitária, Instituto Oswaldo Cruz – Fundação Oswaldo Cruz, 188p.Fernandez, M.A.; Thiengo, S.C.; Simone, L.R.L. 2003. Distri-bution of the introduced freshwater snail Melanoides tubercu-latus (Gastropoda: Thiaridae) in Brazil. Nautilus 117:78-82.Fernandez, M.A.; Thiengo, S.C.; Bezerra, F.S.M.; Alencar, L.M.S. 2010. Current distribution of the exotic freshwater snail Helisoma duryi (Gastropoda: Planorbidae) in Brazil. Nautilus 124 (1):44-50.Ferreira, J.F; Magalhães, A.R.M. 2002. Cultivo de mexilhões. Disponível em <http:www. http://bgnaescola.files.wordpress.com/2010/09/cultivo_mexilhoes.pdf>. Acesso em:29 outubro 2011. Ferriz, R.A.; Villar, C.A.; Colautti, D.; Bonetto, C. 2000. Ali-Ali-mentación de Pterodoras granulosus (Valenciennes) (Pisces, Doradidae) en la baja cuenca del Plata. Revista del Museo Ar-gentino de Ciencias Naturales 2: 151-156.Figueiredo, G.; Mansur, M.C.D; Pimpão, D.; Bergonci, P.E.A. ; Santos, C.P.; Rafaelli, L.; Oliveira, A.; Rodriguez, M. 2011. Morfometria e Morfologia larval de bivalves nativos e invaso-res como base ao monitoramento das áreas límnicas no Brasil (Bivalvia, Mytilidae, Corbiculidae, Hyriidae). In: XXII Encon-tro Brasileiro de Malacologia, Fortaleza, Livro de Resumos: 174-176.Fischer, C.; Gaupp, R. 2004. Multi-scale surface area quanti-fication – a systematic method to evaluate the reactive surface area of rocks. Chemie der Erde / Geochemistry 64:241-256.Fisher, S.W.; Stromberg, P.; Bruner, K.A.; Boulet, L.D. 1991. Molluscicidal activity of potassium to the zebra mussel, Dreis-sena polymorpha: toxicity and mode of action. Aquatic Toxi-cology 20:219–234.Focht, T.; Veitenheimer-Mendes, I.L. 2001. Distribuição sazo-nal e reprodução de Neocorbicula limosa (Maton) (Bivalvia, Corbiculidae) no Lago Guaíba, Rio Grande do Sul, Brasil. Re-vista Brasileira de Zoologia 18(1):35-4.Font, W.F. 2003. The global spread of parasites: what do Ha-waiian streams tell us. BioScience 53:1061-1067.Fontes, H.M.; Takeda, A.M.; Fugita, D.S. 2002. Experience in Itaipu reservoir (Brazil) with artificial substrate (in situ): is it best way of monitoring an invading species like Limnoperna fortunei (Bivalvia, Mytilidae)? Proceedings of International Symposium on Reservoir Management in Tropical and Sub-Tropical Regions, Foz do Iguaçu, Brasil, p.673-679.

397
Fox, R. 2004. Invertebrate anatomy online. Corbicula flu-minea, Asian Clam. In: Invertebrate Zoology, Laboratory exercises. Home page, Biology 310. 16 p. (www.lander.edu/rsfox/310corbiculalab.html). Fox, R. 2006a. Invertebrate anatomy online. Mytilus edulus, Blue mussel. In: Invertebrate Zoology, Laboratory exercises. Cap 12. Mollusca p. 367.Home Page: http://lanwebs.lander.edu/faculty/rsfox/invertebrates/mytilus.html (último acesso em 22-3-2012).Fox, R. 2006b. Invertebrate anatomy online. Corbicula flu-minea, Asian Clam.In: Invertebrate Zoology, Laboratory ex-ercises. Cap 12. Mollusca p. 367. Home Page: http://lanwebs.lander.edu/faculty/rsfox/invertebrates/corbicula.html (último acesso em 22-3-2012).Fraga, S.A.M. 2002. Efeitos do impacto por esgoto doméstico na decomposição e fauna associada, em três rios do Parque Estadual da Pedra Branca, Rio de Janeiro, RJ. Tese de Douto-rado, Pós-graduação em Conservação e Manejo da Vida Silves-tre, Universidade Federal de Minas Gerais, 94 p.França, R.S., Suriani, A.L.; Rocha, O. 2007. Composição das espécies de moluscos bentônicos nos reservatórios do baixo Rio Tietê (São Paulo, Brasil) com uma avaliação do impacto causado pelas espécies exóticas invasoras. Revista Brasileira de Zoologia 24(1):41-51.Freitas, J.R.; Bedê, L.C.; Marco JR, P.; Rocha, L.A.; Santos, M.B.L. 1987. Population dynamics of aquatic snails in Pampu-lha reservoir. Memórias do Instituto Oswaldo Cruz 82 (Suppl. IV):299-305.Freitas, S.M.F.; Kapusta, S.C.; Würdig, N.L. 2009. Invertebra-dos bentônicos das lagoas da planície Costeira do Rio Grande do Sul. p.120-141. In: Würdig, N.L.; Freitas, S.M.F. et al., (Org.) Ecossistemas e Biodiversidade do Litoral Norte do RS. Porto Alegre, Nova Prova, 288p.Friedberg, E.C.; Walker, G.C.; Side, W. 1995. DNA and muta-genesis. ASM Press, Washington, 264p.Gambetta, R. 2011. Plantas piloto: o caminho entre a pesquisa na bancada e a produção em escala industrial. Disponível em: <www.embrapa.br/imprensa/artigos/2011/plantas-piloto>. Acesso em: 09/04/2012.Garcia, A.M.; Bemvenuti, M.A.; Vieira, J.P.; Motta Marques, D.M.; Burns, M.D.M.; Moresco, A.; Condini, M.V.L. 2006. Checklist comparison and dominance patterns of the fish fau-na at Taim Wetland, South Brazil. Neotropical. Ichthyology 4(2): 261-268.Garcia, C.A.E. 1997. Hydrographic Characteristics. p. 18-20. In: Seeliger, U.; Odebrecht, C.; Castello, J.P. (Eds). Subtropical Convergence Environments: the coast and sea in the South-western Atlantic. Springer-Verlag, Berlin, 308p.Garcia, M.; Montalto L., 2006. Los peces depredadores de Limnoperna fortunei en los ambientes colonizados. p. 111-127. In: Darrigran, G.; Damborenea, C. (Eds.). Bio-invasion del mejillón dorado en el continente americano. La Plata, Edulp.Garcia, M.L.; Protogino, L.C. 2005. Invasive freshwater mol-Invasive freshwater mol-luscs are consumed by native fishes in South America. Journal of Applied Ichthyology 21: 34–38Garcia-Ramos, G.; Rodriguez, D. 2002. Evolutionary speed of species invasions. Evolution 56(4):661-668.Garton, D.; Haag, W. 1993. Seasonal reproductive cycles and settlement patterns of Dreissena polymorpha in western Lake Erie. In: Nalepa, T.F.; Schloesser, D.W. (Eds.), Zebra mussels: biology, impact and control. Lewis Publishers, Boca Raton, Florida. 111-128p.Gazulha, V. 2010. O mexilhão dourado Limnoperna fortunei (Dunker, 1857) na presença de cianobactérias: taxas de fil-tração, comportamento alimentar e sobrevivência. Programa de Pós-Graduação em Recursos Hídricos e Saneamento Am-
biental da Universidade Federal do Rio Grande do Sul. Porto Alegre, 104p. [Tese de Doutorado].Gazulha, V.; Mansur, M.C.D.; Cybis, L.F.; Azevedo, S.M.F.O. 2012a. Feeding behavior of the invasive bivalve Limnoperna fortunei (Dunker, 1857) under exposure to toxic cyanobac-teria Microcystis aeruginosa. Brazilian Journal of Biology 72(1):41-49.Gazulha, V.; Mansur, M.C.D.; Cybis,L.F.; Azevedo, S.M.F.O. 2012b. Grazing impacts of the invasive bivalve Limnoperna fortunei (Dunker, 1857) on single-celled, colonial, and fila-mentous cyanobacteria. Brazilian Journal of Biology 72(1):33-39.Genthner, F.J.; Winstead, J.T.; Gillet, J.E.; Van Fleet, A.L.; Viel, J.J.; Genovese, E.E.; Singer, S. 1997. Effects of a mol-luscicidal strain of Bacillus alvei on digestive tubules of zebra mussels, Dreissena polymorpha. Journal of Invertebrate Patho-logy 69:289-291.Giacomini, H.C. 2007. Sete motivações teóricas para o uso da modelagem baseada no indivíduo em ecologia. Acta Amazôni-ca 37(3): 431-446.Gibbons, W.N., Munn, M.D.; Paine, M.D. 1993. Guidelines for monitoring benthos in freshwater environments. Report prepared for Environment Canada, North Vancouver, BC. EVS Consultants, North Vancouver, British Coumbia. 81p. Giovanelli, A.; Vieira, M.V.; Silva, C.L.P.A.C. 2003. Apparent competition through facilitation between Melanoides tubercu-latus and Biomphalaria tenagophila and the control of schis-tosomiasis. Memórias do Instituto Oswaldo Cruz 98 (3):429-431.Giovanelli, A.; Silva, C.L.P.A.C.; Leal, G.B.E.; Baptista, D.F. 2005. Habitat preference of freshwater snails in relation to environmental factors and the presence of the competitor snail Melanoides tuberculatus (Müller, 1774). Memórias do Insti-tuto Oswaldo Cruz 100(2):169-176. Giusti, F. 1971. The fine structure of the style sac and intestine in Mytilus galloprovincialis Lam. Proceedings of the Malaco-logical Society of London 39:95-104.Glaubrecht, M.; Fehér, Z.; Rintelen, T. von. 2006. Brooding in Corbicula madagascarensis (Bivalvia, Corbiculidae) and the re-peated evolution of viviparity in corbiculids. Zoologica Scripta 35(6):641-654.Glaubrecht, M.; Fehér, Z.; Köhler, F. 2007. Inventorizing an invader:Annotated Type Catalogue of Corbiculidae Gray, 1847 (Bivalvia, Heterodonta, Veneroida), including Old World lim-nic Corbicula in the Natural History Museum Berlin. Malaco-logia 49 (2):243-272.Glaubrecht, M.; Brinkmann, N.; Pöppe, J. 2009. Diversity and disparity ‘down under’:Systematics, biogeography and repro-ductive modes of the ‘marsupial’ freshwater Thiaridae (Cae-nogastropoda, Cerithioidea) in Australia. Zoosystematics and Evolution 85 (2):199-275.Gnanamangai, B.M.; Ponmurugan, P. 2012.Evaluation of va-rious fungicides and microbial based biocontrol agents against bird’s eye spot disease of tea plants Crop Protection 32:111-118.Gomes, A.L.; Callil, C.T. 2010. Monitoramento da gameto-gênese de Limnoperna fortunei (Dunker 1857) no Rio Paraná – PR. CNPq/PIBIC/UFMT, Cuiabá, Brasil, 20p.Gomes, A.L.; Callil, C.T. 2011. Monitoramento da gametogê-nese de Limnoperna fortunei (Dunker 1857) no Cais do Porto – RS. FAPEMAT/PIBIC/UFMT, Cuiabá, 26p.Gonçalves, R.F.; Couracci Filho, B.; Chernicharo, C.A.L.; LapollI, F.R.; Aisse, M.M.; Piveli, R P.; Sant’Ana, T.D. 2003. Desinfecção por radiação ultravioleta. p.209-275. In: Gonçal-ves, R. F. (Coordenador). Desinfecção de Efluentes Sanitários. PROSAB3, ABES, RIMA, Rio de Janeiro, Brasil, 438p.

398
Gonçalves, I.C.B.; Miyahira, I.C.; Santos, S.B. 2010. Morfo-logia da concha do molusco exótico Melanoides tuberculatus na Vila do Abraão, Ilha Grande, Angra dos Reis, RJ, Brasil. In: Livro de Resumos do XXVIII Congresso Brasileiro de Zoolo-gia, Belém, p. 1412.González de Infante, A. 1988. El plankton de las aguas conti-nentals. Secretaria General de La Organización de lós Estados Americanos (OEA). Programa Regional de Desarollo Científi-co y Tecnológico, Washington, D.C., 130p.Gonzalez, M.J.; Downing, A. 1999. Mechanisms underlying amphipod responses to zebra mussel (Dreissena polymorpha) invasion and implications for fish-amphipod interactions. Canadian Journal of Fisheries and Aquatic Sciences 56:679–685.Gonze, E.; Fourel, L.; Gonthier, Y.; Boldo, P.; Bernis, A. 1999. Wastewater pretreatmente with ultrasonic irradiation to re-duce toxicity. Chemical Engineering Journal 73(2):93-100.Grimm, A.M.; Barros, V.R.; Doyle, M.E. 2000. Climate vari-ability in Southern South America associated with El Niño and La Niña events. Journal of Climate 13:35-58.Gröbner, S.; Bissinger, A.L.; Raible, A.; Heeg, P.; Autenrieth, I.B.; Schmidt, S.M. 2007. Severe diarrhoea caused by Aeromo-nas veronii biovar sóbria in a patient with metastasised GIST. Polish Journal of Microbiology 56(4):277-279.Gu, J.D.; Mitchell, R. 2001. Antagonism of bacterial extra-cellular metabolites to freshwater-fouling invertebrate zebra mussels, Dreissena polymorpha. Journal of Microbiology 39: 133-138.Guilam, M.C.R. 1996. O conceito de risco. Sua utilização pela Epidemiologia, Engenharia e Ciências Sociais. Disponível em <www.ensp.fiocruz.br/projetos/esterisco/maryfim1.htm>. Acesso em: 09 janeiro 2012.Guimarães, C.T. et al. 1983a. Controle biológico: Pomacea haustrum Reeve, 1856 (Mollusca, Pilidae) sobre planorbíneos, em laboratório. Revista de Saúde Pública 17(2):138-147.Guimarães, C.T. et al. 1983b. Controle biológico: Helobdella triserialis lineata Blanchard, 1849 (Hirudinea: Glossiphoni-dae) sobre Biomphalaria glabrata Say, 1818 (Mollusca: Planor-bidae), em laboratório. Revista de Saúde Pública 17(6):481-492.Guimarães, C.T.; Souza, C.P.; Soares, D.M. 2001. Possible competitive displacement of planorbids by Melanoides tuber-culatus in Minas Gerais, Brazil. Memórias do Instituto Oswal-do Cruz 96 (Suppl.):173-176.Guisan, A.; Zimmermann, N.E. 2000. Predictive habitat distri-Predictive habitat distri-bution models in ecology. Ecological Modeling 135:147-186.Gutiérrez, J.; Jones, C.; Strayer, D., Iribarne, O. 2003. Mol-lusks as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101:79-90. Haag, W.R.; Garton, D.W. 1992. Synchronous spawning in a recently established population of the European zebra mussel, Dreissena polymorpha. Hydrobiologia 234:103-110.Haas, F. 1969. Superfamilia Unionacea. Das Tierreich. Eine Zusammenstellung und Kennzeichunung der rezenten Tierfor-men. Berlim, W. de Gruyter, v. 88: 663p.Halima, H.S.; Bahy, A.A.; Tian, H.H.; Qing, D.X. 2006. Mo-lecular characterization of novel Bacillus thuringiensis isolate with molluscicidal activity against the intermediate host of schitosomes. Biotechnology 5(4):413-420.Hall, D.J. 1964. An experimental approach to the dynamics of a natural population of Daphnia galeata mendotae. Ecology 45(1): 94-112. Haltuch, M.A.; Berkman, P.A.; Garton, D.W. 2000. Geogra-phic information system (GIS) analysis of ecosystem invasion: exotic mussels in Lake Erie. Limnology and Oceanography 45(8):1778-1787.
Hamilton, S.K.; Sippel, S.J.; Calheiros, D.F.; Melack, J.M. 1997. An anoxic event and other biogeochemical effects of the Pantanal wetland on the Paraguay River. Limnology and Oceanography 42(2):257-272.Hammond, B.G.; Barbee, S.J.; Inoue, T.; Ishida, N.; Levins-kas, G.J.; Stevens, M.W.; Wheeler, A.G.; Cascieri, T. 1986, A review of toxicology studies on cyanurate and its chlorinated derivatives. Environmental Health Perspectives 69, 287–292.Haney, J.F.; Hall, D.J. 1973. Sugar coated Daphnia: a preserva-tion technique for cladocera limn. Oceanography18:331-333.Harman, W.J. 1982. The aquatic Oligochaeta (Aeolosoma-tidae, Opistocystidae, Naididae) of Central America. South-western Naturalist 27(3):287-298.Hartl, D.L. 2000. A primer of population genetics. Sinauer Associates, Massachusetts.Hartz, S.M.; Silveira C.M.: Barbieri, G. 1996. Alimentação das espécies de Astianax Baird; Girard 1854, ocorrentes na Lagoa Caconde, RS, Brasil. (Teleostei; Characidae). Revista Unimar 18(2):269-281.Haszprunar, G. 1988. On the origin and evolution of major gastropod groups, with special reference to the Streptoneura. Journal of Molluscan Studies 54 (4): 367–441.Hauer, R.; Resh, V.H. 1996. Macroinvertebrates. In: Hawer, R.; Cambert, G. Methods in stream ecology. Academic Press, New York, p.453-474.Hayes, K.R. 2003. Biosecurity and the Role of Risk Assess-ment. En: Ruiz; Carlton (Eds). Invasive Species: Vectors and Management Strategies. Island Press, Washington, EEUU, 518p.Heller, J.; Farstay, V. 1989. A field method to separate males and females of the freshwater snail Melanoides tuberculata. Journal of Molluscan Studies 55: 427-429.Henriques de Oliveira, C. 2002. Macroinvertebrados asso-ciados à Typha domingensis Pers (Typhaceae) em duas lagoas no litoral norte fluminense e sua utilização em programas de biomonitoramento. Dissertação de Mestrado, Universidade Federal do Rio de Janeiro, Brasil, 92p.Herring, H. 2006. File: DNA UV mutation.gif. Disponível em: http://upload.wikimedia.org/wikipedia/commons/2/2f/DNA_UV_mutation.gif. Acesso em 01 de março de 2012.Hicks, G. 2004. Turning the Tide: Is aquatic bioinvaders research heading in the right direction? Aquatic Invaders 15(1):9–20.Hiroko, T.; Takuya, D.; Takaki, K. 2004. Effect of gasterostome trematode infection on growth and reproduction in Limnoper-na fortunei (Bivalvia: Mytilidae). Memoirs of Osaka Kyoiku University, Natural Science and Applied Science 53 (1):1-7.Ho, K.C.; Chow, Y.L.; Yau, J.T.S. 2003. Chemical and micro-biological qualities of the East River (Dongjiang) water, with particular reference to drinking water supply in Hong Kong. Chemosphere 52: 1441-1450. Holland, B.S. 2000. Genetics of marine bioinvasion. Hydro-biologia 420:63-71.Holland, B.S. 2001. Invasion without a bottleneck: microsa-tellite variation in natural and invasive populations of the bro-wn mussel Perna perna (L). Marine Biotechnology. 3:407-415.Holm, E.R., Stamper, D.M.; Brizzolara, R.A.; Barnes, L.; Dea-mer, N.; Burkholder, J.M. 2008. Sonication of bacteria, phyto-plankton and zooplankton: Application to treatment of ballast water. Marine Pollution Bulletin 56 (6): 1201-1208.Holme, N.A.; McIntyre, A.D. 1971. Methods for the study of Marine Benthos – IBP Handbook No 16. Oxford and Edin-burgh: Blackwell Scientific Publications, 334p. Hongzhu, W.; Zhicai, X.; Xiaoping, W.; Yanling, L. 1999. A preliminary study of zoobenthos in the Poyang lake, the largest freshwater lake of China, and its adjoining reaches of Changji-ang river. Acta Hidrobiologica Sinica 23 suppl:132-138.

399
Horgan, M.J.; Mills, E.L. 1997. Clearance rates and filtering activity of zebra mussel (Dreissena polymorpha): implications for freshwater lakes. Canadian Journal of Fisheries and Aquat-ic Sciences 54 (2):249-255.Horvath, T.G.; Martin, K.M.; LambertI, G.A. 1999. Effect of Zebra Mussels, Dreissena polymorpha, on Macroinvertebrates in a Lakeoutlet Stream. The American Midland Naturalist 142(2):340-347.Hulme, P.E. 2006. Beyond Control: Wider Implications for the Management of Biological Invasions. Journal of Applied Ecology 43:835-847. Hyman, L. H. 1967. The Invertebrates: Mollusca I. Mac Graw-Hill Book Company, New York, United States, 792 p.Idrissi, F.Z.; Agut, M.; Larrondo, J.; Calvo, M.A. 1996. Effect of ultrasound on fungal cells. Cytobios 88: 119-122.Ihering, H. 1890. Revision der von Spix in Brasilien gesammelt-en Najaden. Archiv für Naturgeschichte 11(2):117-170, est 9.Ihering, H. 1893. Najaden von S. Paulo und die geographische Verbreitung der süsswasser Faunen von Südamerika. Archive für Naturgeschichte 1(1/3):45-140. Ihering, H.1910. Über brasilianische Najaden. Senckenber-gische Naturforschende Gesellschaft 32:111-140. ICZN (International Code of Zoological Nomenclature). 2000. International Commission on Zoological Nomencla-ture. International Trust for Zoological Nomenclature, Lon-International Trust for Zoological Nomenclature, Lon-don. Disponível em <http://www.nhm.ac.uk/hosted-sites/iczn/code/>. Acesso em 31/01/2012.Ismail, N.S.; Arif, M.A.S. 1993. Population dynamics of Mela-noides tuberculata (Thiaridae) snails in a desert spring, United Arab Emirates and infection with larval trematodes. Hydro-biologia 257:57-64.Isom, B.G. 1986. Historical review of Asian clam (Corbicula) invasion and biofouling of water and industries in the Americas. American Malacological Bulletin, Special edition (2) Proceed-ings of the Second International Corbicula Symposium:1-5. Ituarte, C.F. 1981. Primera noticia acerca de la introducción de pelecípodos asiáticos en la área rioplatense (Mollusca, Cor-biculidae). Neotropica, 27 (7):79-82. Ituarte, C.F. 1982. Contribución a la biología de la familia Corbiculidae (Mollusca, Pelecypoda) en el Río de la Plata. Tesis número 408, Facultad de Ciencias Naturales y Museo, Universidad de La Plata.Ituarte, C.F. 1984a. Aspectos biologicos de las poblaciones de Corbicula largillierti (Philippi, 1844) (Mollusca Pelecypoda) en el Río de La Plata. Revista del Museu de La Plata (nueva serie) 13(143): 231-247.Ituarte, C.F. 1984b. El fenomeno de incubacion branquial em Neocorbicula limosa (Maton, 1809) (Mollusca Palecypoda). Neotropica 30(83):43-54.Ituarte, C.F. 1985. Growth dynamics in a natural population of Corbicula fluminea (Bivalvia, Sphaeriacea) at Punta Atalaya, Río de La Plata, Argentina. Studies on Neotropical Fauna and Environment 20: 217-225.Ituarte, C.F. 1994. Corbicula and Neocorbicula (Bivalvia: Cor-biculidae) in the Paraná, Uruguay, and Río de La Plata Basins. The Nautilus 107(4):129-136.Ituarte, C.F; Mansur, M.C.D. 1993. Eupera elliptica n. sp. una nueva especie en el Rio Iguazú, Misiones, Argentina. Neo-tropica 39(1):11-16.Iwasaki, K.; Uryu, Y. 1998. Life cycle of a freshwater myti-lid mussel, Limnoperna fortunei, in Uji River, Kyoto.Japanese Journal of Malacology 57(2):105–113.Jack, J.D.; Thorp, J.H. 2000. Effects of the benthic suspension feeder Dreissena polymorpha on zooplankton in a large river. Freshwater Biology 44:569-579.
Johnson, L. E.; Padilla, D. K. 1996. Geographic spread of exotic species: ecological lessons and opportunities from the invasion of zebra mussel Dreissena polymorpha. Biological Conservation 78: 23-33.Jones, C.G.; Lawton, J.H.; Shachak, M. 1994. Organisms as ecosystems engeneers. Oikos 69:373-386.Jones, C.G.; Lawton, J.H.; Shachak, M. 1997. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78(7): 1946-1957.Jørgensen, C.B., 1990. Bivalve filter feeding: hydrodynamics, Bioenergetics, Physiology and Ecology. Olsen; Olsen, 140 p.Junk, W.J.; Bayley, P.B.; Sparks, R.E. 1989. The flood pulse concept in river-floodplain systems. In: Dodge, D.P. (Ed.). In-ternational Large River Simposium (Lars). Canadian Special Publication of the Fisheries Aquatic Science 106:110-127.Juricková, L. 2006. Subulina octona (Bruguière, 1798) – a new greenhouse species for the Czech Republic (Mollusca: Gastro-poda: Subulinidae). Malacologica Bohemoslovaca 5(1):1-2.Jyoti, K.K.; Pandit, A.B. 2004. Effect of cavitation on chemical disinfection efficiency. Water Research 38(9):2249-2258. Karatayev, A.Y.; Burlakova, L.E.; Padilla, D.K. 1997. The effects of Dreissena polymorpha (PALLAS) invasion on aquatic communities in Eastern Europe. Journal of Shellfish Research 16:187-203.Karatayev, A.Y.; Burlakova, L.E.; Padilla, D.K. 2006. Growth rate and longevity of Dreissena polymorpha (Pallas): A review and recommendations for future study. Journal of Shellfish Re-search 25:23-32.Karatayev, A.Y.; Boltovskoy, D.; Padilla, D.K.; Burlakova, L.E. 2007a. The invasive bivalves Dreissena polymorpha and Lim-noperna fortunei: parallels, contrasts, potential spread and in-vasion impacts. The Journal of Shellfish Research 26:205–213.Karatayev, A.Y.; Padilla, D.K.; Minchin, D.; Boltovskoy D.; Burlakova L.E. 2007b. Changes in global economies and trade: the potential spread of exotic freshwater bivalves. Bio-logical Invasions 9:161–180.Karatayev, A.Y.; Burlakova, E.L.; Karatayev, V. A.; Boltovskoy, D. 2010. Limnoperna fortunei x Dreissena polymorpha: Popu-lation density and benthic community impacts of two invasive freshwater bibalves. Journal of Shellfish Research 29:985–984. Keiser, J.; Utzinger, J. 2009. Emerging foodborne trematodia-sis. Clinical Microbiology Reviews 22 (3):466-483. Keller, R. P.; Drake, J. M.; Lodge, D. M. 2007. Fecundity as a basis for risk assessment of nonindigenous freshwater mol-lusks. Conservation Biology 21(1): 191-200.Kikuchi, R.M.; Uieda, V.S. 2005. Composição e distribuição dos macroinvertebrados em diferentes substratos de fundo de um riacho no município de Itatinga, São Paulo, Brasil. Ento-mologia y Vectores 12(2):193-231. Kikuchi, R.M.; Fonseca-Gessner, A.A.; Shimizu, G.Y. 2006. Suction sampler for collection of benthic macroinvertebrates in several continental aquatic environments: a comparative study with the Hess and Surber samplers. Acta Limnologica Brasilensia 18(1):29-37. Kimura, T.; Sekiguchi, H. 1994. Larval and postlarval Shell morphology of two mytilid species Musculista senhousia (Ben-son) and Limnoperna fortunei kikuchii Habe. Venus 53: 307-318.Kimura, T.; Tabe, M.; Darrigran, G.; Panha, S. 2003. Genetic comparative study of Limnoperna fortunei (Bivalvia:Mytilidae) from two native and two invaded localities. p. 173. In: XVII Encontro Brasileiro de Malacologia. Rio de Janeiro, Brasil.King, C.A.; Langdon, C.J.; Counts III, C. 1986. Spawn-ing and early development of Corbicula fluminea (Bivalvia:Corbiculidae) in laboratory culture. American Mala-cological Bulletin 4(1):81-88.

400
Kjerfve, B. 1986. Comparative oceanography of coastal Lago-ons. p.63-81. In: Wolf, D.A. (Ed). Estuarine Variability. Aca-demic Press, Orlando, 509p.Klemm, D.J. 1972. The leeches (Anelida, Hirudinea) of Michi-gan. Michigan Academy 4(4):405–444.Klemm, D.J.; Lewis, P.A.; Fulk, F. 1990. Macroinvertebrate field and laboratory for evaluating the biological integrity of surface waters. Cincinnati: Usepa, 256 p. Kluza, D.A.; Mcnyset, M. 2005. Ecological niche modeling of aquatic invasive species. Aquat Invaders 16(1): 1-7.Komaru, A.; Konishi, K. 1996. Ultrastructure of bifiagellate sperm in the freshwater clam Corbicula leana (Prime). Inverte-brate Reproduction and Development 29:193–197. Komaru, A.; Konishi, K. 1999. Non-reductional spermatozoa in three shell color types of the freshwater clam Corbicula flu-minea in Taiwan. Zoological Science 16:105–108.Komaru, A.; Konishi, K.; Nakayama, I.; Kobayashi, T.; Sakai, H.; Kawamura, K. 1997. Hermaphroditic freshwater clams in the genus Corbicula produce non-reductional spermatozoa with somatic DNA content. Biological Bulletin 193:320–323. Korniushin, A. 2004. A revision of some Asian and African freshwater clams assigned to Corbicula fluminalis (Müller, 1774) (Mollusca: Bivalvia: Corbiculidae), with a review of anatomical characters and reproductive features based on mu-seum collections. Hidrobiologia 529:251-270.Kotzian, H.B.; Marques, D.M. 2004. Lagoa Mirim e a con-venção Ramsar: um modelo para ação transfronteiriça na con-servação de recursos hídricos. Revista de Gestão de Água da América Latina 1(2): 101-111.Kowalewski, J.J.; Patrick, P.H.; Christie, A.E. 1993. Effect of acoustic Energy on the Zebra Mussel (Dreissena polimorpha). In: Zebra Mussel: biology, impacts, and Control. Lewis Pub-lishers, Boca Raton. p.657-666.Kraemer, L.R.; Galloway, M. 1986. Larval Development of Corbicula fluminea (Müller) (Bivalvia: Corbiculacea): an ap-praisal of its heterochrony. American Malacogical Bulletin 4(1):61-79.Kraemer, L.R.; Swainson, C.; Galloway, M.; Kraemer, R. 1986. Bilogical basis of behavior in Corbicula fluminea, II. Functional morphology of reproduction and development and review of evidence for self-fertilizatin. In: Britton, J.C.; Pre-zant, R.S. (eds.). International Corbicula Symposium, Ameri-can Malacological Bulletin, Special Edition, Hattiesburg, 2(2): 193-201.Krogh, A. 1939. Osmotic regulation in aquatic animals. Cam-Cam-bridge Univ. Press, Cambridge.LACTEC – Instituto de Tecnologia para o Desenvolvimento. 2004. In: Estudos para controle da deposição de lama ferru-ginosa (“fouling”) nos sistemas de resfriamento das usinas de Nova Ponte e Emborcação da Companhia Energética de Mi-nas Gerais - CEMIG. Relatório Final. Curitiba, Paraná.Lago, D.V.; Fernandez, M.A.; Thiengo, S. 2009. Análise con-quiliológica de Melanoides tuberculatus (Gastropoda: Thia-ridae) obtidos em diferentes biótopos no Estado do Rio de Janeiro. Livros de Resumos do XXI EBRAM. 176p.Lampert, W. 1982. Further studies on the inhibitory effect of the toxic blue-green Microcystis aeruginosa on the filtering rate of zooplankton. Archiv für Hydrobiologie 95: 207-220.Langone, J.A. 2005. Notas sobre el mejillón dorado Limnoper-na fortunei (Dunker, 1857) (Bivalvia, Mytilidae) en Uruguay. Publicacion Extra del Museo Nacional de Historia Natural y Antropologia 1: 1-18. Disponível em <http://mnhn.gub.uy/ innovaportal/file/3717/1/PE1(enlinea).pdf>.Lanna Filho, R. Ferro, H.M.; Pinho, R.S.C. 2010. Controle biológico mediado por Bacillus subtilis. Revista Tropica – Ciencias Agrarias e Biologicas 4(2):1-12.
Latini, R. 2006. Aprimoramento de um método baseado em modelagem de nicho ecológico para prever a distribuição de espécies aquáticas em rios usando variáveis físico-químicas. Dissertação de Mestrado, Pós-Graduação em Ecologia, Mane-jo e Conservação da Vida Silvestre, Universidade Federal de Minas Gerais, Brasil. 79p.Lee, C.E. 2002. Evolutionary genetics of invasive species. Trends in Ecology & Evolution 17(8):386-391.Lee, T.; Siripattrawan, S.; Ituarte, C.F.; Ó Foighil, D. 2005. Invasion of clonal clams: Corbicula lineages in the New World. American Malacological Bulletin 20:113–122.Leite, O.H.M.; Higaki, Y.; Serpentini, S.L.P.; Carvalho, S.A.; Neto, V.A.; Torres, D.M.A.; Dias, R.M.D.S.; Chieff, J.P.P. 1989. Infecção por Clonorchis sinensis em imigrantes asiáticos no Brasil. Tratamento com praziquantel. Revista do Instituto de Medicina Tropical de São Paulo 31 (6):416-422.Lemos, A.C.M.; Coelho, J.C.; Matos, E.D.; Montal, G.; Aguiar, F.; Badaró, R. 2007. Paragonimiasis:first case reported in Brazil. Brazilian Journal of Infectious Diseases 11:153-156.Livshits, G.; Fishelson, L. 1983. Biology and reproduction of the freshwater snail Melanoides tuberculata (Gastropoda: Pro-sobranchia) in Israel. Israel Journal of Zoology 32:21-35.Lobo, E.A.; Leighton, G. 1986. Estruturas de las fitocenosis planctonicas de los sistemas de desembocaduras de rios y este-ros de la zona central de Chile. Revista de Biologia Marinha 22(1):143-170.Lobo, M. G.; Costa, B. P.; Wisbeck, E. 2009. Avaliação da de-sinfecção de água por reator utilizando radiação ultravioleta. Revista de Ciências Ambientais 3(1): 21-36.Lockwood, J.; M. Hoopes; Marchetti, M. 2007. Invasion Eco-logy. Blackwell Publishing. 304p.Lopes, C.C.; Lopes, R.S.C.; Cardoso, J.N. 2004. Síntese de glicero-fosfatos com atividade antiincrustante em embarca-ções –- UFRJ, Instituto de Pesquisa do Mar Almirante Paulo Moreira – IEAPM, Marinha do Brasil, Arraial do Cabo – RJ.Lopes, M.N. 2010. Estudos ecológicos sobre a espécie in-vasora Limnoperna fortunei (Dunker, 1857) em zonas pro-fundas do Canal São Gonçalo, Lagoa Mirim-RS-Brasil. Pós-graduação em Biologia de Ambientes Aquáticos Continentais. Universidade Federal do Rio Grande, Rio Grande. Lopes, R.M; Villac, M.C. 2009. Métodos. In: Lopes, R.M. (Org.). Informe sobre as espécies exóticas invasoras marinhas no Brasil. Brasília: MMA/SBF, 2009, p. 19-28.Lunetta, J.E. 1969. Fisiologia da Reprodução de mexilhões (Mytilus perna L. Mollusca Lamellibranchia). Boletim de Zoo-logia e Biologia Marinha 26:33-111.Luque-Garcia, J.L.; Castro, M.D.L. 2003. Ultrasound: a powerful tool for leaching. Trends in Analytical Chemistry 22(1): 41-47.Lydeard, C.; Holznagel, W.E.; Glaubrecht, M.; Ponder, W.F. 2002. Molecular phylogeny of circum-global, diverse gastro-pod superfamily (Cerithioidea: Mollusca: Caenogastropoda) - pushing the deepest phylogenetic limits of mitochondrial LSU rDNA sequences”. Molecular Phylogenetics and Evolution 22(3): 399–406.Macêdo, J.A.B. 2004. Águas e Águas. CRQ-MG, Belo Hori-Águas e Águas. CRQ-MG, Belo Hori-. CRQ-MG, Belo Hori-zonte, 977p.Machado, C.J.S.; Oliveira, A.E.S. 2009. Espécies exóticas in-vasoras: problema nacional ainda pouco conhecido. Ciência e Cultura 61(1):22-23. Mack, R. N.; Simberloff, D.; Lonsdale, W. M.; Evans, H.; Clout, M.; Bazzaz, F. A. 2000. Biotic Invasions: Causes, Epi-Biotic Invasions: Causes, Epi-demiology, Global Consequences and Control. Ecological Ap-plications 10(3): 689-710.Mackie, G.L. 1991. Biology of the exotic zebra mussel, Dreis-sena polymorpha, in relation to native bivalves and its poten-tial impact in Lake St. Clair. Hydrobiologia 219: 251-268.

401
Mackie, G.L.; Claudi, R. 2010. Monitoring and Control of Macrofouling Mollusks in Fresh Water Systems. 2nd. Edition. Taylor & Francis, Boca Raton, New York. 508p.Mäder Netto, O.S. 2011. Controle da incrustação de organis-mos invasores em materiais de sistemas de resfriamento de usinas hidrelétricas Dissertação de Mestrado, Pós-Graduação em Enge-nharia, Universidade Federal do Paraná, Brasil, 112p.Madsen, H. 1990. Biological methods for the control of freshwa-ter snails. Parasitology Today 6(7): 237-241.Madsen, H.; Frandsen, F. 1989. The spread of freshwater snails including those of medical and veterinary importance. Acta Tro-pica 46:139-146.Magara, Y.; Matsui. Y.; Goto, Y.; Yuasa, A. 2001. Invasion of the non-indigenous nuisance mussel, Limnoperna fortunei, into wa-ter supply facilities in Japan. Journal of Water Supply: Research and Technology-AQUA 50:113-124.Malabarba, L.R. 1989. Histórico sistemático e lista comentada das espécies de peixes de água doce do sistema da Laguna dos Patos, Rio Grande do Sul, Brasil. Comunicação do Museu de Ci-ências da PUCRS (Ser. Zoologia) 2:107-179.Mansur, M.C.D. 1999. Gloquídio de Diplodon martensi (Ihe-ring) (Mollusca, Bivalvia, Hyriidae) e seu ciclo parasitário. Curiti-ba, Revista Brasileira de Zoologia 16(2):185-194.Mansur, M.C.D.; Campos-Velho, N.M.R. 1990. Técnicas para o estudo dos gloquídios de Hyriidae (Mollusca, Bivalvia, Unio-noida). São Leopoldo, Acta Biologica Leopoldensia 12(1):5-18.Mansur, M.C.D.; Campos-Velho, N.M.R. 2000. The glo-The glo-chidium of Castalia martensi (Ihering, 1891) (Bivalvia, Unionoida:Hyriidae). Heldia 3(1):6-10. Mansur, M.C.D.; Garces, L.M.M.P. 1988. Ocorrência e densida-Ocorrência e densida-de de Corbicula fluminea (Müller, 1774) e Neocorbicula limosa (Matton, 1811) na Estação Ecológica do TAIM e áreas adjacen-tes, RS. Brasil. (Mollusca, Bivalvia, Corbiculidae). Iheringia, Série Zoologia (68):99-115.Mansur, M.C.D.; Meier-Brook, C. 2000. Morphology of Eupera (Bourguignat, 1854) and Byssanodonta Orbigny, 1846 and the phylogenetic affinities within Sphaeriidae and Corbiculidae (Bi-valvia - Veneroida). Archiv für Molluskenkunde der Senckenber-gischen Naturforschenden Gesellschaft 128(1):1-59.Mansur, M.C.D.; Pereira, D. 2006. Bivalves límnicos da bacia do Rio dos Sinos, Rio Grande do Sul, Brasil (Bivalvia, Unionoida, Veneroida e Mytiloida). Revista Brasileira de Zoologia 23(4):1123-1147.Mansur, M.C.D.; Silva, M.G.O. 1990. Morfologia e microana-tomia comparada de Bartlettia stefanensis (Moricand, 1856) e Anodontites tenebricosus (Lea, 1834) (Bivalvia, Unionoida, Mu-teloidea). Amazoniana 11(2): 147-166. Mansur, M.C.D.; Silva, M.G.O. 1999. Description of glochidia of five species of freshwater mussels (Hyriidae: Unionoidea) from South America. Philadelphia, Malacologia 41(2):1-9.Mansur, M.C.D.; Valer, R.M. 1992. Moluscos bivalves dos rios Uraricoera e Rio Branco. Amazoniana 12(1):85-100. Mansur, M.C.D.; Schulz, C.; Garces, L.M.M.P. 1987. Moluscos bivalves de água doce: identificação dos gêneros do sul e leste do Brasil. Acta Biologica Leopoldensia 9(2): 181-202.Mansur, M.C.D.; Veitenheimer-Mendes, I.L.; Almeidacaon, J.E.M. 1988. Mollusca, Bivalvia de um trecho do curso inferior do Rio Jacuí, Rio Grande do Sul, Brasil. Iheringia, Série Zoologia 67:87-108.Mansur, M.C.D.; Schulz, C.; Silva, M.G.O.; Campos-Velho, N.M.R. 1991. Moluscos Bivalves Límnicos da Estação Ecológica do Taim e áreas adjacentes. Rio Grande do Sul, Brasil. Iheringia, Série Zoologia 71:43-58.Mansur, M.C.D.; Valer, R.M.; Aires, N.C.M. 1994. Distribuição e preferências ambientais dos moluscos bivalves do açude do Par-que de Proteção Ambiental COPESUL, município de Triunfo, RS - Brasil. Biociências 2(1):27-45.
Mansur, M.C.D.; Richinitti, L.M.Z.; Santos, C.P. 1999. Lim-noperna fortunei (Dunker, 1857) molusco bivalve invasor na Bacia do Guaíba, Rio Grande do Sul, Brasil. Biociências 7(2):147-149. Mansur, M.C.D.; Santos, C.P.; Darrigran, G.; Heydrich, I.; Callil, C.T.; Cardoso, F.R. 2003. Primeiros dados quali-quan-titativos do mexilhão dourado, Limnoperna fortunei (Dunker, 1857), no Delta do Jacuí, no Lago Guaíba e na Laguna dos Patos, Rio Grande do Sul, Brasil e alguns aspectos de sua inva-são no novo ambiente. Revista Brasileira de Zoologia 20(1): 75-84.Mansur, M.C.D.; Quevedo, C.B.; Santos, C.P.; Callil, C.T. 2004a. Prováveis vias da introdução de Limnoperna fortu-nei (Dunker, 1857) (Mollusca, Bivalvia, Mytilidae) na Bacia da Laguna dos Patos, Rio Grande do Sul e novos registros de invasão no Brasil pelas Bacias do Paraná e Paraguai. 4, p. 33-38. In: Silva, J.S.V.; Souza, R.C.C. L. (Org.) Água de Lastro e Bioinvasão. Interciências, Rio de Janeiro, 224p.Mansur, M.C.D.; Cardoso, F.R.; Ribeiro, L.A. Santos, C.P.; Thormann, B.M.; Fernandes, F.C.; Richinitti, L.M.Z. 2004b. Distribuição e conseqüências após cinco anos da invasão do mexilhão dourado, Limnoperna fortunei (Dunker, 1857), no estado do Rio Grande do Sul, Brasil (Mollusca, Bivalvia, Myti-lidae). Biociências 12(2):165-172. Mansur, M.C.D.; Callil, C.T.; Cardoso, F.R.; Ibarra, J.A.A. 2004c. Uma retrospectiva e mapeamento da invasão de espé-cies de Corbicula (Mollusca, Bivalvia, Veneroida, Corbiculi-dae) oriundas do sudeste asiático, na América do Sul. 5, p.39-58. In: Silva, J.S.V.; Souza, R.C.C.L. (Orgs.) Água de Lastro e Bioinvasão. Interciências, Rio de Janeiro, 224p.Mansur, M.C.D.; Meier-Brook, C.; Ituarte, C. 2008a. A new species of Sphaerium Scopoli, 1777, from southern Brazil (Bi-valvia: Sphaeriidae). The Nautilus 121:228-235.Mansur, M.C.D.; Figueiró, H.; Santos, C.P.; Glock, L.; Ber-gonci, P.E.A., Pereira, D. 2008b. Variação espacial do compri-mento e do peso úmido total de Limnoperna fortunei (Dunker, 1857) no delta do Rio Jacuí e Lago Guaíba (RS, Brasil). Biote-mas 21(4): 49-54. Mansur, M.C.D.; Santos, C.P.; Richinitti L.M.Z.; Pereira, D.; Batista, C.B.; Silveira, M.B.; Alberto, R.M.F.; Silva, M.C.P. 2008c. Ocorrência de moluscos límnicos e crustáceos em ma-Ocorrência de moluscos límnicos e crustáceos em ma-croaglomerados do mexilhão dourado, Limnoperna fortunei (Dunker, 1857) sobre sarandi no Lago Guaíba (RS, Brasil). Biotemas 21(4):179-182.Mansur, M.C.D.; Pereira, D.; Santos, C.P.; Bergonci, P.E.A.; Thormann, B.M.; Takeda, A.M. 2009. Colonização de subs-trato artificial de madeira pelo mexilhão dourado, Limnoper-na fortunei (Dunker, 1857) (Bivalvia, Mytiloida, Mytilidae), no Delta do Rio Jacuí (RS, Brasil). Biotemas 22(1):75-80.Mansur, M.C.D.; Santos, C.P.; Nehrke, M.V. 2011a. Corbicu-lidae na América do Sul, espécies nativas e invasoras, dispersão e a situação das pesquisas no Brasil (Mollusca: Bivalvia). p. 348-362. In: Fernandez, M.A.; Santos, S.B.; Pimenta, A.D.; Thiengo, S.C. (Eds.). Tópicos em Malacologia – Ecos do XIX EBRAM, Sociedade Brasileira de Malacologia, Rio de Janeiro, 378p.Mansur, M.C.D; Santos, C.P; Pimpão, D.M. 2011b. Morfolo-gia do adulto e da larva, incubação e distribuição de Mytilopsis lopesi Alvarenga; Ricci, um Dreissenidae euvivíparo do bai-xo Amazonas (Bivalvia, Veneroida, Dreissenidae). p. 327. In: XXII Encontro Brasileiro de Malacologia. Livro de Resumos, Fortaleza, 04 a 08 de setembro de 2011. 611p. Mansur, M.C.D.; Santos, C.P.; Rafaelli, L.; Nehrke, M.V.; Bueno-e-Silva, M.; Bergonci, P. E. A.; Pereira, D.; Oliveira, A.S.; Bergmann, C.; Raya-Rodriguez, M.T. 2011c. Repro-Repro-ductive behavior of Corbicula fluminea and C. largillierti in a tropical reservoir, Brazil (Bivalvia, Corbiculidae). p. 41. In: 2nd World Conference on Biological Invasions and Ecosystem

402
Functioning, Biolief. Livro de Resumos. Session 9 – Population biology of invasive species. Mar del Plata, Argentina, Novem-ber 21-24 2011. 91p.Mansur, M.C.D.; Santos, C.P.; Pereira, D.; Bergonci, P.E.A.; Callil, C.T. 2012. Cap. 7 Moluscos Límnicos – Bivalves. p. 131-179. In: Informe sobre as Espécies Exóticas Invasoras de Águas Continentais no Brasil. Brasília, Ministério do Meio Ambiente – MMA. (em impressão).Marçal, S.F.; Callil, C.T. 2008. Structure of invertebrates com-munity associated with Eichhornia crassipes Mart. (Solms-Lau-bach) after the introduction of Limnoperna fortunei (Dunker, 1857) (Bivalvia, Mytilidae) in the Upper Paraguay River, MT, Brazil. Acta Limnologica Brasiliensia 20(4):359-371. Marcelo, M.S.; Callil, C.T. 2006. Gametogênese de Limno-perna fortunei (Dunker, 1857) no Lago Guaíba, Porto Alegre - RS. TCC. Graduação em ciências biológicas – Universidade Federal de Mato Grosso – UFMT. Cuiabá. 2006. 30p.Marcelo, M.S.; Callil, C.T.; Sampaio, M.B.; Mello, F.S. 2004. Ocorrência, densidade e aspectos populacionais do Mexilhão dourado, Limnoperna fortunei (Dunker, 1857) no Pantanal de Mato Grosso. In: Resumos. 56 Reunião Anual da SBPC, Cuiabá.Marcelo, M.S.; Uliana, E.R.; Callil, C.T.; Mansur, M.C.D.; Oliveira, M.D.; Mezzomo, B.P.; Massolii JR, E.V. 2005. As-pectos da reprodução de Limnoperna fortunei (Dunker, 1857) (Bivalve, Mytiliidae). In: Pimenta, A.;Santos, S. (Eds) XIX En-contro Brasileiro de Malacologia, 2005, Rio de Janeiro. Anais do XIX Encontro Brasileiro de Malacologia. p. 219.Maroñas, M.E.; Darrigran, G.A.; Sendra, E.D.; Breckon, G. 2003. Shell growth of the golden mussel, Limnoperna fortunei (Dunker, 1857) (Mytilidae), in the Rıo de la Plata, Argentina. Hydrobiologia 495:41-45.Martin, S.; Darrigran, G. 1994. Limnoperna fortunei (Dunker, 1857) en el Balneario Bagliardi, Rio de la Plata. Alteración en la composición de la malacofauna litoral. Tankay 1:164-166. Martinez, M.A.U.; Davolos, M.R.; Jafelicci, M. 2000. O efei-to do ultra-som em reações químicas. Química Nova 23 (2): 251-256.Martinez, R.E. 1987. Corbicula manilensis molusco introdu-cido en Venezuela. Acta Científica Venezuelana 38:384-385.Martins, D.S. 2004. Aspectos comparativos da morfologia e comportamento reprodutivo de três espécies de Corbicula Mühlfeld, no Lago Guaíba, Rio Grande do Sul, Brasil. UFRGS. Instituto de Biociências. Programa de Pós-Graduação em Bio-logia Animal. (Dissertação de mestrado). Porto Alegre. 2004, 107p.Martins, D.S.; Veitenheimer-Mendes, I.L.; Faccioni-Heuser, M.C. 2004. Corbicula (Bivalvia,Corbiculidae) em simpa-tria no Lago Guaíba, Rio Grande do Sul, Brasil. Biociências 12(2):129-138.Martins, D.S.; Veitenheimer-Mendes, I.L.; Faccioni-Heuser, M.C. 2006. Aspectos morfológicos e de incubação em três es-pécies de Corbicula Mühfeld, no Lago Guaíba, Rio Grande do Sul, Brasil (Bivalvia, Corbiculidae). Biota Neotropica 6(2):1-11.Massoli-Junior, E.V. 2006. Aspectos populacionais de Corbi-cula fluminea (Bivalvia: Veneroida) no Rio Cuiabá, Praia da Vereda, Santo Antônio do Leverger, MT. Departamento de Biologia e Zoologia, Instituto de Biociências, Universidade Fe-deral do Mato Grosso (Monografia Licenciatura em Ciências Biológicas). 26p.Massoli-Junior, E.V.; Callil, C.T. 2012. Influência da variação fluviométrica na densidade etária e atividade gametogênica Corbicula fluminea (Bivalvia: Veneroida) em trecho do Rio Cuiabá, Santo Antônio do Leverger, MT. Cap. 7. Moluscos Límnicos, p. 206-221. In: Informe sobre as Espécies Exóticas Invasoras de Águas Continentais no Brasil. Brasília, Ministé-rio do Meio Ambiente – MMA.
Mata, F.A.R.; Patricio, F.C.; Campos, M.C.S.; Fróes, C.L. 2007. Avaliação da mortalidade de larvas de mexilhão dou-rado, Limnoperna fortunei, (Dunker 1857), expostas em di-ferentes concentrações do produto MXD-100. II Encontro Sul-Americano de Integração de Ações para Controle do Me-xilhão Dourado.Mata, F.A.R.; Dias, C.A.; Patrício, F.C.; Rolla, M.E., Carvalho, M.D.; Freitas, L.C. 2008. Avaliação da eficiência da substância MXD-100 na prevenção de incrustação e corrosão, por lama ferruginosa, nos trocadores de calor tipo placa, da usina de Nova Ponte (CEMIG/MG). III Seminário Brasileiro de Meio Ambiente e Responsabilidade Social no Setor Elétrico.Matos, E.; Matos, R; Santos, M.N.S.; Giese, E.G. 2001. Ação de protozoários parasitas em moluscos da região ama-zônica: Nematopsis sp. XVII Encontro Brasileiro de Malaco-logia, Resumos, Recife, p.78-79.Matsui, Y.; Nagoya, K.; Funahashi, G.; Goto, Y.; Yuasa, A.; Yamamoto, H.; Ohkawa, K.; Magara, Y. 2002. Effectiveness of Antifouling Coatings and Water Flor in Controling Attach-ment of the Nuisance Mussel Limnoperna fortunei. Biofoul-ing 18 (2):137-148.Matsumura-Tundisi, T., Luzia, A.P.; Tundisi, J.G. 2006. Estado trófico dos reservatórios em cascata do Médio e Baixo Tietê (SP) e manejo para o controle da eutrofização, pp. 141-160. In: J. G. Tundisi, T. Matsumura-Tundisi; Galli, C.S. (eds.). Eutrofização na América do Sul: causas, conseqüências e tec-nologias para gerenciamento e controle. EUTROSUL – Rede Sul-Americana de eutrofização (IIE, IIEGA), 532p.Mayer, D. 2009. Daphnia magna acute toxicity tests of Pseu-domonas fluorescens CL145A. Disponível em: http://marrone-bioinnovations.com/pdf/zeq/AcuteToxicity-Daphniamagna.pdf.Mccorkle, S.; Dietz, T.H. 1980. Sodium transport in the freshwater asiatic clam, Corbicula fluminea. Biological Bulle-tin 159:325-336.Mccune, B.; Mefford, M.J. 1997. PC-ORD. Multivariate Analysis of Ecological Data – Version 3. MJM Software De-sign. Gleneden Beach. 273p.McMahon, R.F.; Williams, C.J. 1986. Growth, life cycle, uper thermal limit and downstream colonisation rates in a natural population of the freshwater bivalve mollusc, Corbicula flu-minea (Müller) receiving thermal effluents. In: Britton, J.C.; Prezant, R.S. (eds.). Proceeding of the second International Corbicula Symposium, American Malacological Bulletin, Spe-cial Edition 2(2): 231-239.Meador, J.P. 2000. Predicting the fate and effects of tributyl-tin in marine systems. Rev. Environmental Contamination and Toxicology 166:1-48.Meier-Brook, C. 1984. A preliminary biogeography of freshwater pulmonate gastropods. p. 23-37. In: Solem, A.; Bruggen, C. (Eds). World-Wide Snails: Biogeographical Stu-dies on non-marine Mollusca. Brill; Backhuys, Leiden.Mellina, E.; Rasmussen, J.B. 1994. Patterns in the Distribution and Abundance of Zebra Mussel (Dreissena polymorpha) in Rivers and lakes in Relation to Substrate and Other Physico-chemical Factors. Canadian Journal of Fisheries and Aquatic Science 51: 1024-1036.Melo, I.S.; Azevedo, J.L. 2000a. Controle Biológico. v. 2. Ja-guariúna: Embrapa Meio Ambiente. 387 p.Melo, I.S.; Azevedo, J.L., 2000b. Controle Biológico. v. 3. Ja-guariúna: Embrapa Meio Ambiente. 308 p.Mendonça, A.C., Ferreira, A.S.; Barbieri, C.H.; Thomazine, J.A.; Mazzer, N. 2006. Effects of low-power pulsed ultrasound on second-intention healing of total skin injuries in rats. Acta Ortopédica Brasileira 14(3):152-157.Menegat, R.; Kirchhein, R.E. 1998. Lagos rios e arroios: as doces águas da superfície. In: Menegat, R. (coord.). Atlas Am-biental de Porto Alegre. UFRGS, Porto Alegre, p. 35-42.

403
Metzeling, L.; Chessman, B.; Hardwick, R.; Wong, V. 2003. Rapid assessment of rivers using macroinvertebrates: the role of experience, and rapid comparisons with quantitative meth-ods. Hydrobiologia 510:39-52. Mihuc, T.B.; Battle, J.M.; Mihuc, J.R.; Bryan, C.F. 1999. Zebra mussel (Dreissena polymorpha) seasonal colonization patterns in a subtropical floodplain river. Hydrobiologia 392:121–128.Milesi, S.V.; Restello, R.M.; Hepp, L.U. 2009. Distribution of benthic macroinvertebrates in Subtropical streams (Rio Grande do Sul, Brazil). Biological Limnology 21(4):419-429. Milward-de-Andrade, R. 1978a. Fecundidade de Helisoma duryi e Biomphalaria glabrata, criadas em laboratório. Revista Brasileira de Malariologia e Doenças Tropicais 30:5-44.Milward-de-Andrade, R. 1978b. Competição entre Helisoma duryi e Biomphalaria glabrata, em condições de laboratório. Revista Brasileira de Biologia 38 (4):787-800.Milward-de-Andrade, R. 1979. Natalidade comparada de Helisoma duryi e Biomphalaria glabrata, em laboratório (Mollusca, Planorbidae). Revista Brasileira de Malariologia e Doenças Tropicais 31:35-58.Milward-de-Andrade, R.; Souza, C.P. 1979. Infecção experi-mental de Biomphalaria glabrata com Schistosoma mansoni, na presença de Helisoma duryi. Revista Brasileira de Mala-riologia e Doenças Tropicais 31 (Suppl.):185-186.Milward-de-Andrade, R.; Oliveira, S.; Ramos, A.A. 1979. Observação preliminar sobre liberação de cercarias de S. mansoni por Biomphalaria glabrata associadas à Helisoma duryi. Ciência e Cultura 31 (Suppl.):532.Minillo, A. 2005. Análise da distribuição, densidade e toxici-dade de florações de cianobactérias em reservatórios do médio e baixo Rio Tietê (SP) e relação com as características limnoló-gicas do sistema. Tese de Doutorado. Escola de Engenharia de São Carlos, Universidade de São Paulo, São Carlos.Ministério da Saúde. 2007. Vigilância e Controle de Moluscos de Importância Epidemiológica. 2a edição. Série A: Normas e Manuais Técnicos. 177p.Miyahira, I.C. 2010. Dinâmica populacional de Melanoides tuberculatus (Müller, 1774) em um riacho impactado da Vila do Abraão, Ilha Grande, Angra dos Reis, Rio de Janeiro, Brasil. UERJ. Dissertação (Ecologia e Evolução). 110p.Miyahira, I.C.; Lacerda, L.E.M.; Santos, S.B. 2009. Dis-persão ativa de Melanoides tuberculatus (Müller, 1774) e sua influência no processo de invasão. Informativo SBMa 40(167):3-5.Miyahira, I.C.; Lacerda, L.E.M.; Santos, S.B. 2010. How many species are introduced every day? Some insights from a tropical insular stream in Brasil. Tentacle 18:30-32.Möller, O.O.; Casting, P.; Salomon, J.C.; Lazure, P. 2001. The influence of local and non local forcing effects on the subtidal circulation of Patos Lagoon. Estuaries 24:297-311.Molloy, D.P. 1998. The potential for using biological control technologies in the management of Dreissena spp. Journal of Shellfish Research 17(1):177-183.Molloy, D.P.; Mayer, D.A. 2007. Overview of a Novel Green Technology: Biological Control of Zebra and Quagga Mus-sels with Pseudomonas fluorescens. Disponível em: www.aquaticnuisance.org/wordpress/wp-content/uploads/2009/01/Dreissena-Novel-Green-Technology-for-Dreissena-Control-4--Malloy.pdf.Montalto, L.; Drago, I.E. 2003. Tolerance to desiccation of an invasive mussel, Limnoperna fortunei (Dunker, 1857) (Bival-via, Mytilidae), under experimental conditions. Hydrobiolo-gia 498:161-167.Montalto, L; Oliveros, O. B.; Ezcurra de Drago, I.; Demonte, L.D. 1999. Peces del Rio Parana Medio predadores de una
especie invasora: Limnoperna fortunei (Bivalvia, Mytilidae). Revista de la Facultad de Bioquimica y Ciencias Biológicas de la Universidad Nacional del Litoral 3: 85-101.Mooney, H.A.; Cleland, E.E. 2001. The evolutionary impact of invasive species. Proceedings of the National Academy of Sciences 98:5446-5451.Morales, G.; Pino, L.; Angulo, N. 1987. Resistencia de Physa cubensis a la infección con Fasciola hepatica. Revista de La Facultad de Ciências Veterinárias 34(1/4):43-55.Moreno, P.; Callisto, M. 2006. Benthic macroinvertebrates in the watershed of an urban reservoir in southeastern Brazil. Hy-drobiologia 560:311-321.Morley, N.; Morritt, D. 2006. The effects of the slug biologi-cal control agent, Phasmarhabditis hermaphrodita (Nemato-da), on non-target aquatic molluscs. Journal of Invertebrate Pathology 92:112 - 114.Morrison, J.P.E. 1954. The relationships of old and new world melanians. Proceedings of the United States National Museum 103(3325):357-394.Mörtl, M.; Rothhaupt, K. 2003. Effects of adult Dreissena polymorpha on settling juveniles and associated macroinverte-brates. Hydrobiologia 88(6):561-569.Morton, B. 1973. Some aspects of the biology and functional morphology of the organs of feeding and digestion of Limno-perna fortunei (Dunker, 1857) (Bivalvia: Mytilacea). Malaco-logia 12(2):265-281.Morton, B. 1977. The population dynamics of Limnoperna fortunei (Dunker, 1857) (Bivalvia: Mytilacea) in Plover Cove Reservoir, Hong Kong. Malacologia 16(1):165–182.Morton, B. 1982. The reproductive cycle of Limnoperna fortunei (Dunker, 1857) (Bivalvia: Mytilidae) fouling Hong Kong’s raw water supply system. Oceanologia et Limnologia Sinica 13(4):319-324.Morton, B. 1986. Corbicula in Asia – an updated synthesis. In: Britton, J. C. (Ed.) Proceedings of the Second International Corbicula Symposium, American Malacological Bulletin, Special Edition, 2: 113124.Mugnai, R.; Nessimian, J. L.; Baptista, D. F. 2010. Manual de identificação de macroinvertebrados aquáticos do Estado do Rio de Janeiro. Rio de Janeiro, Technical Books, 176p.Müller, O.F. 1774. Vermium terrestrium et fluviatiles, seu ani-malium infusorium, helminthicorum et testaceorum, non ma-rinorum, succincta historia. Vol. 2. Heineck; Faber, Havniae et Lipsiae, 532p.Murphy, W.A.; Dietz, T.H. 1976. The effects of salt depletion on blood and tissue ion concentrations in the freshwater mus-sel, Ligumia subrostrata (Say). Journal of Comparative Physi-ology 108:233-242.Murray, H.D. 1964. Tarebia granifera and Melanoides tubercu-lata in Texas. American Malacological Union Annual Report 1964:15-16.Naddafi, R.; Pettersson, K.; Eklöv, P. 2007. The effect of sea-sonal variation in selective feeding by zebra mussels (Dreis-sena polymorpha) on phytoplankton community composition. Freshwater Biology 52: 823-842.Nakano, D.; Kobayashi, T.; Sakaguchi, I. 2010. Differences in larval dynamics of golden mussel Limnoperna fortunei be-tween dam reservoirs with and without an aeration system. Landscape and Ecological Engineering 6:53–60.Nalepa, T.F.; Fanslow, D.L.; Lansing, M.B.; Lang, G.A. 2003. Trends in the benthic macroinvertebrate community of Sagi-naw Bay, Lake Huron, 1987–96: responses to phosphorus abatement and the zebra mussel, Dreissena polymorpha. Jour-nal of Great Lakes Research 29:14–33.Nascimento, L.C.; Lima, L.C.O.; Piccoli, R.H.; Veiga, S.M.O.M.; Fiorini, J.E. 2005. Uso de derivados clorados, ozô-Uso de derivados clorados, ozô-

404
nio e ultra-som na sanificação de água e alimentos - Revisão. Higiene Alimentar 19(136): 48-57.Neary, B.P.; Leach, J.H. 1992. Mapping the potential spread of the zebra mussel (Dreissena polymorpha) in Ontario. Cana-dian Journal of Fisheries and Aquatic Sciences 49: 406–415.Nicholls, K.H.; Hopkins, G.J. 1993. Recent changes in Lake Erie (North Shore) phytoplankton: cumulative impacts of phosphorus loading reductions and the zebra mussel introduc-tion. Journal of Great Lakes Research 19:637-647.Nicholls, K.H.; Heintsch, L.; Carney, E. 2002. Univariate step-trend and multivariate assessments of the apparent effects of P loading reductions and zebra mussels on the phytoplankton of the Bay of Quinte, Lake Ontario. Journal of Great Lakes Research 28:15-31.Nichols, S.J. 1993. Spawning of zebra mussels (Dreissena polymorpha) and rearing of veligers under laboratory condi-tions. In: Nalepa, T.F.; Schloesser, D.W. (Eds.). Zebra mussels: biology, impacts, and control. Lewis Publishers, Boca Raton, Florida, 315p. Niencheski, L.F.H.; Baumgarten, M.G.Z. 1997. Environmen-tal chemistry. p.20-24. In: Seeliger, U.; Odebrecht, C.; Cas-tello, J.P. (Eds). Subtropical Convergence Environments: the coast and sea in Southwestern Atlantic. Springer-Verlag, Berlin, 308p. Nishimura, T.; Habe, T. 1987. Chinese freshwater mussels mi-xed in the imported Corbicula clam. Chiribotan (em japonês) 18:110-111.NRC (NATIONAL RESEARCH COUNCIL). 2002. Predicting Invasions of Nonindigenous Plants and Plant Pest. National Academy Press. Washington, D.C.Núñez, V. 2010. Differences on allocation of available resour-ces, in growth, reproduction, and survival, in an exotic gastro-pod of Physidae compared to an endemic one. Iheringia, Série Zoologia 100(3).O’Neill, C.R. Jr. 1997. Economic impact o zebra mussels – re-sults of the 1995 National zebra mussel information clearing-house study. Great Lakes Research Review. 3(1). 35-42.Oduho, G.W.; Chung, T.K.; Bakers, D.H. 1993. Menadiona nicotinamide bisulfite is a bioactive source of vitamin K and niacin activity for chicks. Journal of Nutrition 123:737-743. Disponível em: <http: jn.nutrition.org> acesso em: 19 janeiro de 2011.Okamura, B. 1986. Group living and the effects of spatial posi-tion in aggregations of Mytilus edulis. Oecologia 69(3):341-347.Okuno, E.; Caldas, I.L.; Chow, C. 1986. Física para ciências biológicas e biomédicas. São Paulo, Harba, 490p.Olazarri, J. 1986. Las almejas del género Corbicula en el río Uruguay. Resúmenes de Comunicaciones, Seminario “El Rio Uruguay y sus Recursos” CARU-INAPE-INIDEP, Entre Rios, Argentina, 2p.Olden, J.D.; Poff, N.L.; Douglas, M.R.; Douglas, M.E.; Faus-ch, K.D. 2004. Ecological and evolutionary consequences of biotic homogenization. Trends in Ecology and Evolution 19:18–24.Oliveira, C.R.C. de; Fugi, R.; Brancalhão, K.P.; Agostinho, A.A. 2010. Fish as Potential Controllers of Invasive Mollusks in a Neotropical Reservoir. Natureza & Conservação 8(2): 140-144.Oliveira, E.J.; Rabinovitch, L.; Monnerat, R.G.; Passos, L.K.J.; Zahner, V. 2004. Molecular characterization of Brevibacllus laterosporus and its potential use in biological control. Applied and Environmental Microbiology 70(11): 6657-6664.Oliveira, M.D. 2003. Ocorrência e impactos do Mexilhão dourado Limnoperna fortunei (Dunker, 1857) no Pantanal Mato-Grossense. Circular Técnica n. 38, EMBRAPA, MS. Co-rumbá. 6p.
Oliveira, M.D. 2009. Fatores atores reguladores e distribuição potencial do mexilhão dourado (Limnoperna fortunei Dunker 1857) na bacia do alto Rio Paraguai e outros rios brasileiro. Tese de Doutorado. Programa de Pós-Graduação em Ecologia, Conservação e Manejo de Vida Silvestre. Universidade Federal de Minas Gerais. 100p.Oliveira, M.D.; Takeda, A.M.; Barbosa, D.S.; Calheiros, D.F. 2000. Ocorrência da espécie exótica Mexilhão Dourado (Bi-valvia, Mytilidae) no Rio Paraguai, Pantanal, Brasil. In: Simpó-sio sobre Recursos Naturais e Sócio-Econômicos do Pantanal. Manejo e Conservação 3., Corumbá. Resumos... Corumbá: Embrapa Pantanal. Oliveira, M.D.; Takeda, A.M.; Barros, L.F.; Barbosa, S.D.; Re-zende, E.K. 2006. Invasion by Limnoperna fortunei (Dunker, 1857) (Bivalvia, Mytilidae) of the Pantanal wetland, Brazil. Biological Invasions 8(1):97-104.Oliveira, M.D.; Hamilton, S.K.; Calheiros, D.F.; Jacobi, C.M.; Latini, R.O. 2010a. Modeling the potential distribution of the invasive golden mussel Limnoperna fortunei in the Upper Paraguay River system using limnological variables. Brazilian Journal of Biology 70(3): 831-840. Oliveira, M.D.; Hamilton, S.K.; Calheiros, D.F.; Jacobi, C.M. 2010b. Oxygen Depletion Events Control the Invasive Golden Mussel (Limnoperna fortunei) in a Tropical Floodplain. Wet-lands, 30(4): 705-716.Oliveira, M.D.; Mansur, M.C.D.; Barbosa, D.S., Calhei-ros, D.F.; Callil .T. 2010c. Área de ocorrência de moluscos invasores na bacia do Rio Miranda, Pantanal Sul, MS. In: V Simpósio Sobre Recursos Naturais e Socioeconômicos do Pan-tanal. Resumos, Corumbá, MS. Oliveira, M.D.; Hamilton, S.K.; Jacobi, C.M. 2010d. Fore-Fore-casting the expansion of the invasive golden mussel Limnoper-na fortunei in Brazilian and North American rivers based on its occurrence in the Paraguay River and Pantanal wetland of Brazil. Aquatic Invasions 5(1):59-73.Oliveira, M.D.; Calheiros, D.F.; Jacobi, C.M.; Hamilton, S.K. 2011. Abiotic factors controlling the establishment and abun-Abiotic factors controlling the establishment and abun-dance of the invasive golden mussel Limnoperna fortunei. Bio-logical Invasions 13: 717-729. Oliveira-Filho, E.C. 2008. Avaliação da periculosidade am-biental de bioinseticidas como uma nova perspective para a ecotoxicologia no Brasil. Journal of Brazilian Society of Eco-toxicology 3(1): 1-7.Omori, M.; Ikeda, T. 1984. Methods in marine zooplankton ecology. John Wiley; Sons, New York: 332p.Orensanz, J.M.; Bortolus, A.; Casas, G.; Darrigran, G.; Elías, R.; Lópes-Gappa, J.J.; Obenat, S.; Pascual, M.; Pastorino, G.; Penchaszdeh, P.; Piriz, M.L.; Scarabino, F.; Schwindt, E.; Spivak, E.D.; Villarino, E.A. 2002. No longer the pris-tines confines of the World ocean: a survey of exotic marine species in the southwestern Atlantic. Biological Invasions 4: 115-143.Orme, S.; Kegley, S. 2006. PAN pesticide database. Disponível em: <http://www.pesticideinfo.org/Detail_Chemical.jsp?Rec_Id=PC33703> Acesso em: 19/agosto/ 2008.Ortmann, A.E. 1921. South American naiades; a contribution to the knowledge of the freshwater mussels of South America. Memories of Carnegie Museum 8(3): 451-684. Osman, G.Y.; Mohamed, A.M. 1991. Bio-efficacy of bacterial insecticide, Bacillus thuringiensis Berl. as biological control agent against snails vectors of Schistosomiasis in Egypt. Anzei-ger für Schädlingskunde 64(7):136-139.Osman, G.Y.; Mohamed, A.M.; Kader, A.A.; Mohamed, A.A. 2011. Biological studies on Biomphalaria alexandrina snails treated with Furcraea selloa marginata plant (family: Agava-ceae) and Bacillus thuringiensis kurstaki (Dipel-2x). Journal of Applied Pharmaceutical Science. 01(10): 47-55.

405
Ourives, T.M.S. 2010. Distribuição especial da macrofauna bêntica no estuário do Rio Cachoeira, Ilhéus, Bahia. Disser-tação de mestrado do programa de Pós-Graduação em Sis-temas Aquáticos Tropicais, área de concentração: Ecologia. Universidade Estadual de Santa Cruz. 41p. Disponível em: http://www.uesc.br/cursos/pos_graduacao/mestrado/ppsat/dissertacoes/00thailla_ourives.pdf.Ozawa, H.; Sekiguchi, H. 2002. Shell morphology of larvae and post-larvae of the two tropical Modiolus species, Modiolus auriculatus and Modiolus philippinarum (Bivalvia: Mytilidae). Venus 61: 25-39.Özesmi, S.L.; Tan, C.O.; Özesmi, U. 2006. Methodological is-Methodological is-sues in building, training, and testing artificial neural networks in ecological applications. Ecological Modelling 195: 83-93.Pace, M.L.; Findlay, S.E.G.; Fischer, D. 1998. Effects of an invasive bivalve on the zooplankton community of the Hudson River. Freshwater Biology 39:103-116.Pappas, J.L.; Stoermer, E.F. 1996. Quantitative method for determining a representative algal sample count. Journal of Phycology 32: 693-696.Paraense, W.L. 1975. Estado atual da sistemática dos planor-bídeos brasileiros. Arquivos do Museu Nacional 55:105-111.Paraense, W.L. 1976a. A natural population of Helisoma duryi in Brazil. Malacologia 15(2):369-376.Paraense, W.L. 1976b. Helisoma trivolvis and some of its syn-onyms in the Neotropical region (Mollusca: Planorbidae). Re-vista Brasileira de Biologia 36(1):187-204.Paraense, W.L. 1981a. Gastropoda. p. 201-207. In: Hulbert, S.H.; Rodrigues, G.; Santos, N.D. (Eds). Aquatic Biota of Tropical South America, Part. 2: Anarthopoda. San Diego State University, San Diego, California, 298p.Paraense, W.L. 1981b. Biomphalaria occidentalis sp. n. from South America (Mollusca Basommatophora Pulmonata). Me-mórias do Instituto Oswaldo Cruz 76(2):199-211.Paraense, W.L. 1984. Biomphalaria tenagophila guaibensis ssp. n. from Southern Brazil and Uruguay (Pulmonata: Planorbi-dae). I. Morphology. Memórias do Instituto Oswaldo Cruz 79(4): 465-469.Paraense, W.L. 1986a. Lymnaea columella: two new Brazilian localities in the states of Amazonas and Bahia. Memórias do Instituto Oswaldo Cruz 81: 121-123.Paraense, W.L. 1986b. Physa marmorata Guilding, 1828 (Pul-monata: Physidae). Memórias do Instituto Oswaldo Cruz 81:459-469.Paraense, W.L. 1987. Physa cubensis Pfeiffer, 1839 (Pulmo-nata: Physidae). Memórias do Instituto Oswaldo Cruz 82:15-20.Paraense, W.L. 2003. A bird’s eye survey of Central American planorbid molluscs. Memórias do Instituto Oswaldo Cruz 98: 51-67.Paraense, W.L. 2011. Sinonímia entre Physa acuta e Physa cubensis: morfologia e genética. In: Fernandez, M.A.; Santos, S.B.; Thiengo, S.C; Pimenta, A.D. (Eds). Tópicos em Mala-cologia. Ecos do XIX Encontro Brasileiro de Malacologia, Sociedade Brasileira de Malacologia; Technical Books, Rio de Janeiro, (no prelo).Paraense, W.L.; Pointier, J.P. 2003. Physa acuta Draparnaud, 1805 (Gastropoda: Physidae): a study of topotypic specimens. Memórias do Instituto Oswaldo Cruz 98:513-517.Pareschi, D.C.; Matsumura-Tundisi, T.; Medeiros, G.R.; Lu-zia, A.P.; Tundisi, J.G. 2008. First occurrence of Limnoper-na fortunei (Dunker, 1857) in the Rio Tietê watershed (São Paulo State, Brazil). Brazilian Journal of Biology 68(4, Sup-pl.):1107-1114.Park, G.M; Chung, E.Y. 2004. Histological studies on herma-froditism, gametogenesis and cyclic changes in the structures
of marsupial gills of the introduced Asiatic clam, Corbicula fluminea, and the Korean clam, Corbicula leana. Journal of Shellfish Research 23(1):179-184.Parodiz, J. J.; Bonetto, A.A. 1963. Taxonomy and Zoogeo-graphic relationships of the South American naiades (Pelecy-poda: Unionacea and Mutelacea). Malacologia 1(2):179-213.Pastorino, G.; Darrigran, G.; Martin, S.M.; Lunaschi, L. 1993. Limnoperna fortunei (Dunker, 1857) (Mytilidae), nuevo bival-vo invasor en aguas del Río de la Plata. Neotropica 39(101-102): 34.Patrício, F.C.; Fróes, C.L.; Campos, M.C.S. 2003. Avaliação da Toxicidade de Substâncias químicas potencialmente molus-cicidas para controle de Limnoperna fortunei. Relatório técni-co ANNAEL/ CEMIG/CETEC.Paz, I.C.P. 2009. Bactérias endofíticas de eucalipto e potencial uso no controle de doenças e promoção de crescimento de mu-das em viveiros florestais. Tese de doutorado, Pós-graduação em Fitotecnia, Universidade Federal do Rio Grande do Sul, Brasil, 111p.Pelichotti, P., Damborenea, C.; Darrigran, G. 2002. Variacio-nes espaciales de la composición y estructura de la macrofauna sociada a Limnoperna fortunei (Bivalvia Mytilidae) en la cuen-ca del plata. V Congreso Latinoamericano de Malacología – CLAMA. São Paulo Brasil.Penchaszadeh, P. E.; Darrigran,G.; Angulo,C.; Averbuj, A.; Brögger, M.; DogliottI A.; Pírez, N. 2000. Predation of the in-vasive freshwater mussel Limnoperna fortunei (Dunker, 1857) (Mytilidae) by the fish Leporinus obtusidens Valenciennes, 1846 (Anostomidae) in the Río de la Plata, Argentina. Journal of Shellfish Research 19: 229–231.Pereira, D.; Veitenheimer-Mendes, I.L.; Mansur, M.C.D.; Sil-va, M.C.P. 2000a. Malacofauna límnica do sistema de irriga-ção do arroio Capivara e áreas adjacentes, Triunfo, RS, Brasil. Biociências 8(1):137-157.Pereira, D.; Konrad, H.G.; Paloski, N.I. 2000b. Gastrópodos límnicos da bacia do Rio Camaquã, RS, Brasil. Acta Biologica Leopoldensia 22(1):55-66.Pereira, D.; Veitenheimer-Mendes, I.L.; Consoni, J.M.; Kon-rad, H.G. 2001. Composição e abundância de espécies de mo-luscos do bentos marginal da microbacia do arroio Capivara, Triunfo, RS, Brasil. Biociências 9(1):3-20.Pereira, D.; Mansur, M.C.D.; Volkmer-Ribeiro, C.; Oliveira, M.D.; Santos, C.P.; Bergonci, P.E.A. 2010. Colonização de substrato artificial por macroinvertebrados límnicos, no delta do Rio Jacuí (RS, Brasil). Biotemas 23(1):101-110.Pereira, D.; Arruda, J.O.; Menegat, R.; Porto, M.L.; Schwar-zbold, A.; Hartz, S.M. 2011a. Guildas tróficas, composição e distribuição de espécies de moluscos límnicos no gradiente flu-vial de um riacho subtropical brasileiro. Biotemas 24:21-36.Pereira, D.; Paz, I.C.P.; Souza, A.M.S.; Liedke, R. B. S.; Suzuki, M. T.; Azevedo, J. L.; Mansur, M. C. D.; Raya-Rodriguez, M.T. 2011b. Atividade moluscicida de Bacillus thuringiensis sobre o mexilhão dourado, Limnoperna fortune. In: XII Sim-pósio Brasileiro de Controle Biológico, São Paulo, CO 02.06.Pereira, P.A.C. 1997. Primeiro registro de Corbicula fluminea (Müller, 1774) (Corbiculidae), um molusco asiático, no Rio Tibagi (Primeiro de Maio, PR). In: Encontro Brasileiro de Ma-lacologia 15, Florianópolis. Livro de Resumos, p. 38.Pereyra, P. 2011. Invasão de Limnoperna fortunei (Dunker, 1857). Estudio del impacto causado en el ambiente natural y humano. Trabajo de Tesis de Doctorado (ms). Facultad de Ciencias Naturales y Museo Universidad Nacional de La Pla-ta.Perez-Neto, J. L. 1995. Introdução a análises morfométricas. In: Tópicos em tratamento de dados biológicos (Perez-Neto, J.L.; Valentin, F.A.S.; Fernandez, F. (Eds.). Oecologia Brasi-liensis 2: 57-89.

406
Pestana, D.; Pie, M.R.; Ostrensky, A.; Boeger, W. A.; Andre-oli, C.; Franceschi, F.; Lagos, P. 2008. Seasonal variation in larval density of Limnoperna fortune (Bivalvia, Mytilidae) in the Iguaçu and Paraná Rivers, in the Region of Foz do Igua-çu, Paraná, Southern Brazil. Brazilian Archives of Biology and Technology 51(3):607-612.Peterson, A.T. 2003. Predicting the geography of species’ inva-sions via ecological niche modeling. Quarterly Review of Biol-ogy 78(4):419-433.Peterson, A.T. 2006. Uses and requirements of ecological niche models and related distributional models. Biodiversity Infor-matics 3:59-72.Peterson, A.T.; Cohoon, K.P. 1999. Sensitivity of distributional prediction algorithms to geographic data completeness. Eco-logical Modeling 117:159–164.Peterson, A.T.; Vieglais, D.A. 2001. Predicting species in-vasions using ecological niche modeling: new approaches from bioinformatics attack a pressing problem. Bioscience 51(5):363-371.Pfeifer, N.T.S.; Lopes Pitoni, V.L.2003. Análise qualitativa es-tacional da fauna de moluscos Límnicos no Delta do Jacuí, Rio Grande do Sul, Brasil. Biociências 11(2):145-158.Phillips, S.J.; Robert, P.; Anderson, C.; Robert, E. 2006. Maxi-mum entropy modeling of species geographic distributions. Ecological Modelling 190: 231–259.Pie, M.R.; Boeger, W.A; Patella, L.; Falleiros, R.M. 2006. A fast and accurate molecular method for the detection of larvae of the golden mussel Limnoperna fortunei (Mollusca: Mytillidae) in plankton samples. Journal of Molluscan Studies 72: 218-219.Piedras, S.R.; Bager, A.; Corrêa, F. 2007. Ocorrência de Lim-noperna fortunei (Dunker, 1857) durante um período de sali-nização do baixo Arroio Pelotas, Pelotas, Rio Grande do Sul. Boletim do Instituto de Pesca 33:121-125.Pieve, S.M.N.; Kubo, R.R.; Coelho-de-Souza, G. 2009. Pesca-dores artesanais da Lagoa Mirim: Etnoecologia e Resiliência. Ministério do Desenvolvimento Agrário, Brasília, 244p.Pilsbry, H.A.; Bequaert, J. 1927. Aquatic Mollusks of Belgian Congo. Bulletin of the American Museum of Natural History 53:59-602.Pimpão, D.M.; Mansur, M.C.D. 2009. Chave pictórica para identificação dos bivalves do baixo Rio Aripuanã, Amazonas, Brasil (Sphaeriidae, Hyriidae e Mycetopodidae) Biota Neotro-pica 9(3):377-384.Pimpão, D.M.; Martins, D.S. 2008. Ocorrência do molusco asiático Corbicula fluminea (Müller, 1774) (Bivalvia, Corbicu-lidae) no baixo Rio Negro, Amazônia central. Acta Amazonica 38 (3):589-592.Pimpão, D.M.; Rocha, M.S.; Fettuccia, D.C. 2008. Freshwater mussels of Catalão and Negro River, state of Amazon, Brazil. Check List 4(4):395-400.Pimpão, D.M.; Mansur, M.C.D.; Bergonci, P.E.A.; Beasley, C.R. 2012. Comparative morphometry and morphology of glochidial shells of Amazonian Hyriidae (Mollusca: Bivalvia: Unionida). American Malacological Bulletin 30(1):73-84.Pinillos, A.C.M; Callil, C.T. 2011a. Estudo da reprodução de Limnoperna fortunei (Dunker, 1857) no Rio Baía – Paraná, Brasil. TCC. Graduação em Ciências Biológicas – Universida-de Federal de Mato Grosso – UFMT Cuiabá, Brasil 24p.Pinillos, A.C.M; Callil, C.T. 2011b. Gametogênese de Lim-noperna fortunei na Ilha da Pintada – RS. FAPEMAT/PIBIC/UFMT, Cuiabá, 23p.Pinto, H.A. 2009. Infecção natural de Melanoides tuberculata (Mollusca: Thiaridae) por Centrocestus formosanus (Tremato-da: Heterophyidae) e por Philophthalmus gralli (Trematoda: Philophthalmidae) no Brasil. Dissertação de Mestrado, Uni-versidade Federal de Minas Gerais. 99p.
Pinto, H.A.; Melo, A.L. 2010a. Melanoides tuberculata (Mollusca:Thiaridae) as na intermediate host of Centrocestus formosanus (Trematoda: Heterophyidae) in Brazil. Revista do Instituto de Medicina Tropical de São Paulo 52(4):207-210.Pinto, H.A.; Melo, A.L. 2010b. Melanoides tuberculata as In-termediate host of Philophthalmus gralli in Brazil. Revista do Instituto de Medicina Tropical de São Paulo 52(6):323-327.Pinto, T.R.; Boehs, G. 2008. Nematopsis sp. (Apicomplexa:Eugregarinida) em Mytella guyanensis (La-marck, 1819) (Bivalvia: Mytilidae) da região estuarina do Rio Cachoeira, Ilhéus, Bahia, Brasil. Brazilian Journal of Veteri-Brazilian Journal of Veteri-nary Research and Animal Science 45:95-100.Pip, E.1990. Copper, Lead and Cadmium concentrations in a sample of Lake Winnipeg Anodonta grandis. The Nautilus 103(4):140-142.Poi de Neiff, A.; Neiff, J.J. 2006. Riqueza de especies y simila-ridad entre invertebrados que viven en macrófitas de la plani-cie de inundación del río Paraná. Interciencia 31(3):220–225.Pointier, J.P.; Mccullough, F. 1989. Biological control of the snail hosts of Schistosoma mansoni in the Caribbean area using Thiara spp. Acta Tropica 46 (1989):147-155.Pointier, J.P; Guyard, A.; Mosser, A. 1989. Biological control of Biomphalaria glabrata and B. straminea by the competitor Thiara tuberculata in a transmission site of schistomiasis in Martinique, French West Indies. Annals of Tropical Medicine and Parasitology 83 (3):263-269.Pointier, J.P.; Théron, A.; Borel, G. 1993. Ecology of the introduced snail Melanoides tuberculata (Gastropoda:Thiaridae) in relation to Biomphalaria glabrata in the marshy forest zone of Guadeloupe, French West Indies. Journal of Molluscan Studies 59:421-428.Pointier, J.P.; Incani, R.N.; Balzan, C.; Chrosciechowski, P.; Prypchan, S. 1994. Invasion of the rivers of the Littoral Cen-tral Region of Venezuela by Thiara granifera and Melanoides tuberculata (Mollusca: Prosobranchia: Thiaridae) and the absence of Biomphalaria glabrata, snail host of Schistosoma mansoni. The Nautilus 17: 124-128.Pointier, J.P.; Samadi, S.; Jarne, P; Delay, B. 1998. Introduction and spread of Thiara granifera (Lamarck, 1822) in Martinique, French West Indies. Biodiversity and Conservation 7: 1277-1290.Polman, H.J.G.; Janssen-Mommen, J.P.M. 2011. Consultancy services to assist and advise Confederacion Hidrográfica del Ebro for control and mitigation of the Zebra mussel within the EU regulations. Ministerio de Medio Ambiente y Medio Rural y Mariño (España), Kema,115.Ponder, W.F.; Lindberg, D.R. 1997. Towards a phylogeny of gastropod molluscs: an analysis using morphological charac-ters. Zoological Journal of the Linnean Society 119: 83-265.Ponder, W.F.; Lindberg, D.R. (eds). 2008. Phylogeny and Evolution of the Mollusca. University of California Press, California. 469p.Prentis, P.J.; Wilson, J.R.U; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. 2008. Adaptive evolution in invasive spe-cies. Trends in Plant Science 16(6):288-294.Price, P.W. 1977. General concepts on the evolutionary biology of parasites. Evolution 31(2): 405-420.Prokopovich, N.P. 1969. Land subsidence along Delta-Mendo-ta Canal, California. Journal of International Society for Rock Mechanics 1:134-144.Qiu, A.; Shi, A.; Komaru, A. 2001. Yellow and brown shell color morphs of Corbicula fluminea (Bivalvia:Cobiculidae) from Sichuan province, China, are triploids and tetraploids. Journal of Shellfish Research 20:323–328.Queiroz, J.F.; Silveira, M.P.; Sitton, M.; Marigo, A.L.S.; Zam-bon, G.V.; Silva, J.R.; Carvalho, M.P.; Ribacinko, D.B. 2007. Coletor de macroinvertebrados bentônicos com substrato arti-ficial para monitoramento da qualidade de água em viveiros de produção de tilápia. Circular Técnica 16 EMBRAPA. p. 1-5.

407
Radel, S.; Mcloughlin, A.J.; Gherardini, L.; Doblhoff-Dier, O.; Benes, E. 2000. Viability of yeast cells in well controlled propagating and standing ultrasonic plane waves. Ultrasonics 38(1-8): 633-637.Radziejewska, T.; Fenske, C.; Wawrzyniak, W.; Riel, P.; Wo-zniczka, A.; Gruszka, P. 2009. The zebra mussel (Dreissena polymorpha) and the benthic community in a coastal Baltic lagoon: another example of enhancement? Marine Ecology 30:138–150.Raffel, T. R., Martin, L. B. et al. 2008. Parasites as predators: Unifying natural enemy ecology. Trends in Ecology and Evo-lution 23:610-618.Rahman, M.; Colque-Navaroo, P.; Kühn, I.; Huys, G.; Swin-gs, J.; Möllby, R. 2002. Identification and characterization of pathogenic Aeromonas veronii biovar sobria associated with epizootic ulcerative syndrome in fish in Bangladesh. Applied and Environmental Microbiology, 68(2):650-655.Railsback, S.F. 2001. Concepts from complex adaptive systems as a framework for individual-based modelling. Ecological Modelling 139: 47-62.Rajagopal, S.; Venugopalan, V. P.; Van Der Veld, G.; Jenner, H. A. 2003. Response of fouling brown mussel, Perna perna (L) to chlorine. Archives of Environmental Contamination and Toxicology 44:369-376.Ram, J.L.; Fong, P.P.; Garton, D.W. 1996. Physiological aspects of zebra mussel reproduction: Maturation, spawning and fer-tilization. American Zoologist 36:326-338.Ramcharan, C.W.; Padilla, D.K.; Dodson, S.I. 1992. Models to predict potential occurrence and density of zebra mussel, Dreissena polymorpha. Canadian Journal of Fisheries and Aquatic Science 49: 2611-2620.Ramsey, J.B.; White, D.S. 2007. Spatial distribution of ben-thic macroinvertebrates in a Sidearm Embayment of Kentucky Lake. Journal of Kentucky Academy Science 68(1):50-58. Reddy, A.; Ponder, E.L.; Fried, B. 2004. Effects of copper sul-fate toxicity on cercariae and metacercariae of Echinostoma caproni and Echinostoma trivolvis and on the survival of Biom-phalaria glabrata snails. Journal of Parasitology 90(6):13-32-1337.Renner, M.; Cunha, A.X. DA. 1986. Bivalvia, p 178-195 In: Kükenthal, W.; Renner, M. Guia de Trabalhos Práticos de Zo-ologia. 19 th Ed. Livraria Medina Coimbra, Traduzido e adap-tado por A. Xavier da Cunha. 472p.Ricciardi, A. 1998. Global range expansion of the Asian mus-sel Limnoperna fortunei (Mytilidae): another fouling threat to freshwater systems. Biofouling 13(2):97-106.Ricciardi, A.; Rasmussen, J.B. 1998. Predicting the identity and impact of future biological invaders: a priority for aquat-ic resource management. Canadian Journal of Fisheries and Aquatic Sciences 55:1759-1765.Ricciardi, A.; Whoriskey, F.G.; Rasmussen, J.B. 1997. The role of the zebra mussel (Dreissena polymorpha) in structuring macroinvertebrate communities on hard substrata. Canadian Journal of Fisheries and Aquatic Sciences 54:2596–2608.Rios Vivos. 2001. Rios Vivos. Disponível em: <http://www.riosvivos.org.br>. Rocha, O. 2008. Efeitos da toxicidade das cianobactérias em ecossistemas de água doce. Holos Environment 8 (2): palestra 4.Roderick, G.K.; Navajas, M. 2003. Genes in new environ-ments: genetics and evolution in biological control. Nature Reviews Genetics 4: 889-899.Rodrigues, P.A.; Richinitti, L.M.Z.; Mansur, M.C.D. 1998a. Moluscos bivalves do Rio Uruguai. In:Jornada de Iniciação Científica – Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Catálogo de Resumos, p. 95.Rodrigues, P.A.; Cunha, F.B.; Mansur, M.C.D.; Richinitti,
L.M.Z. 1998b. Moluscos bivalves do Rio Uruguai. 1 fase: Tre-cho superior do Rio. In: Salão de Iniciação Científica 10 e Feira de Iniciação Científica 7, Univ. Federal do Rio Grande do Sul, Porto Alegre, Livro de Resumos (Sessão 24 – Zoologia/Biodiversidade), p. 267-268.Rojas Molina, F.; Paggi, S.J. 2008. Zooplankton in the Paraná River floodplain (South America) before and after the invasion of Limnoperna fortunei (Bivalvia). Wetlands 28(3):695-702.Rojas Molina, F.; Paggi, J.C.; Devercelli, M. 2010. Zooplank-tophagy in the natural diet and selectivity of the invasive mol-lusk Limnoperna fortunei. Biological Invasions 12(6):1647-1659.Ropelewski, C.F.; Halpert, M.S. 1987. Global and regional scale precipitation patterns associated with El Niño/Southern Oscillation. Monthly Weather Review 115(8): 1606-1626.Rosenberg, D.M.; Resh, V.H. 1982. The use of artificial subs-trates in the study of freshwater benthic macroinvertebrates. In: Cairns Jr., J. (Ed.). Artificial substrates. Ann Arbor: Ann Arbor Science/ Butterworth Group, p. 175-235. Ruppert, E.E.; Barnes, D.R. 1996. Zoologia dos Invertebra-dos. Rocca, São Paulo, 1029p.Ruppert, E.E.; Fox; R.S.; Barnes, R.D. 2005. Zoologia dos invertebrados: uma abordagem funcional-evolutiva. Ed. Roca.Sälgeback, J.; Savazzi, E. 2006. Constructional morphology of cerithiform gastropods. Paleontological Research 10 (3): 233–259.Samadi, S.; Mavarez, J.; Pontier, J.P.; Delay, B.; Jarne, P. 1999. Microsatellite and morphological analysis of population struc-ture in parthenogenetic freshwater snail Melanoides tubercula-ta: inisights into the creation of variability. Molecular Ecology 8:1141-1153.Santos, C.L.T. 2011. Colonização de organismos incrustantes em substrato artificial na região estuarina da Lagoa dos Patos, sob a influência do fenômeno El Niño. Dissertação de Mestra-do, Pós-Graduação em Oceanografia Biológica, Universidade Federal do Rio Grande, FURG, Brasil, 42p.Santos, C.P. 2004. Avaliação da densidade e crescimento popu-lacional do mexilhão dourado Limnoperna fortunei (Dunker, 1857) em suas diferentes fases de vida no Lago Guaíba, muni-cípio de Porto Alegre, RS, como subsídios ao controle do bi-valve invasor. Dissertação de Mestrado, Universidade Federal do Rio Grande do Sul, Brasil, 93p.Santos, C.P. 2011. Desenvolvimento de metodologia para controle e mitigação dos efeitos causados pela incrustação do mexilhão dourado em tubulações de captação de água bruta. Tese de Doutorado, Programa de Pós-Graduação em Ecologia, Porto Alegre, 83p.Santos, C.P.; Würdig, N.L.; Mansur, M.C.D. 2005. Fases larvais do mexilhão dourado Limnoperna fortunei (Dunker) (Mollusca, Bivalvia, Mytilidae) na Bacia do Guaíba, Rio Gran-de do Sul, Brasil. Revista Brasileira de Zoologia 22(3):702-708.Santos, C.P.; Mansur, M.C.D.; Würdig, N.L. 2008. Varia-ções no comprimento dos indivíduos de uma população do mexilhão dourado, Limnoperna fortunei (Mollusca: Bivalvia: Mytilidae), ao longo do ano, na praia do Veludo, Lago Guaí-ba, Rio Grande do Sul, Brasil. Revista Brasileira de Zoologia 25(3):389-396. Santos, S.B.; Miyahira, I.C.; Lacerda, L.E.M. 2007. First re-First re-cord of Melanoides tuberculatus (Müller, 1774) and Biom-phalaria tenagophila (d’Orbigny, 1835) on Ilha Grande, Rio de Janeiro, Brazil. Biota Neotropica 7:361-364.Sardiña, P.; Cataldo, D.; Boltovskoy, D. 2008. The effects of the invasive mussel, Limnoperna fortunei, on associated fauna in South American freshwaters: importance of physical struc-ture and food supply. Fundamental and Applied Limnology 173:135-144.

408
Scarabino, F. 2004. Lista sistematica de los Gastropoda ma-rinos y estuarinos vivientes de Uruguay. Comunicaciones la Sociedad Malacológica del Uruguay 8: 305-346.Scarabino, F.; Mansur, M.C.D. 2007. Lista Sistemática de los bivalvia dulciaquícolas viventes de Uruguay. Comunicaciones de la Sociedad Malacológica del Uruguay 9:89-99.Scheide, J.I.; Dietz, T.H. 1982. The effects of independent sodium and chloride depletion on ion balance in freshwater mussels. Canadian Journal of Zoology 60:1676-1682.Schloesser, D.W. 1995. Introduced species: zebra mussel in North America. p. 337-356. In: Nieremberg, W.A. (Ed). En-cyclopedia of Environmental Biology. Academic Press, San Diego, 2114p.Scholz, T.; Salgado-Maldonado, G. 2000. The introduction and dispersal of Centrocestus formosanus (Nishigori, 1924) (Diginea:Heterophyidae) in Mexico:a review. American Mid-land Naturalist 143 (1):185-200.Scremin-Dias, E.; Lorenz-Lemke, A.P.; Oliveira, A.K.M. 2011. The floristic heterogeneity of the Pantanal and the occurrence of species with different adaptive strategies to water stress. Brazilian Journal of Biology 71(1): 275-282.Seed, R.; Suchanek, T.H. 1992. Population and community ecol-ogy of Mytilus. In: The mussel Mytilus: ecology, physiology, genet-ics and culture p.87–169. Elsevier, Amsterdam. (E. Gosling, ed.).Segundo, S.M.A.; Kosel, O.; Vianna, L.F.L.; Esmanhoto, E.; Neves, E.F.A. 1993. Metodologia desenvolvida para remoção de “fouling” no sistema de resfriamento das unidades gera-doras da usina Governador Parigot de Souza – US/GPS. XII SNPTEE.Sephton, T.W.; Paterson, C.G.; Fernando. C.H. 1980. Spa-tial interrelationships of bivalves and nonbivalve benthos in a small reservoir in New Brunswick, Canada. Canadian Journal of Zoology 58:852-859.Seymour, I.J.; Burfoot, D.; Smith, R.L.; Cox, L.A.; Lockwood, A. 2002. Ultrasound decontamination of minimally processed fruits and vegetables. International Journal of Food Science and Technology 37(5): 547-557.Sharp, G.J.E.; Secombes, C.J. 1993. The role of reactive oxy-gen species in the killing of the bacterial fish pathogen Ae-romonas salmonicida by rainbow trout macrophages. Fish & Shellfish Immunology, 3(2): 119-129.Shummay, S.E.; Cucci, T.L.; Newell, R. C.; Yentsch, C.M. 1985. Particle selection, ingestion and absorption in filter feeding bivalves. Journal of Experimental Marine Biology and Ecology 91:77-92.Silva, D.P. 2006. Aspectos Bioecológicos do Mexilhão Doura-do (Limnoperna fortunei) (Bivalvia Mytilidae) (Dunker, 1857). Tese de doutorado. Programa de pós-graduação em Engenharia Florestal, Universidade Federal do Paraná, Brasil, 123p.Silva, E.P. 2001a. Genética da bioinvasão. Pesquisa Naval 14:237-247.Silva, E.P. 2001b. A short history of evolutionary theory. His-tória, Ciências, Saúde-Manguinhos 8(3):671-687.Silva, E.P. 2004. Marcadores moleculares no rastreamento da bioinvasão. p. 191-203. In: Silva, J.S.V.; Souza, R.C.C.L. (Eds). Água de Lastro e Bioinvasão. Editora Interciência, Rio de Janeiro.Silva, E.P. 2009. Genética Marinha. p. 49-69. In: Pereira, R.C.; Soares-Gomes, A. (Eds). Biologia Marinha. 2ª ed. Editora In-terciência, Rio de Janeiro.Silva, E.P.; Russo, C. 2000. Techniques and statistical data analysis in molecular population genetics. Hydrobiologia 420:119-135.Silva, G.C.;Hahn, N.S.; Fugi, R.; Gubiani, E. 2005. Alimenta-ção de peixes – Participação de bivalves invasores na dieta de Pterodoras granulosus. PELD – Programa de Pesquisas Ecoló-gicas de Longa Duração.
Silva, J.S.V.; Fernandes, F.C.; Souza, R.C.L.; Larsen, K.T.S., Danelon, O.M. 2004. Água de lastro e bioinvasão. p.1-10. In: Silva, J.S.V.; Souza, R.C.L. (Orgs). Água de lastro e bioinvasão. Editora Interciência, Rio de Janeiro, 224p.Silva, J.S.V.; Abdon, M.M.; Moraes, J.A. 2010. Desmatamento na bacia do Alto Paraguai no Brasil. In: Anais do III Simpósio de Geotecnologias no Pantanal. Embrapa Informática Agrope-cuária/INPE. Cáceres, MT. p. 458-467.Silva, L.C.F.; Vieira, L.C.G.; Costa, D.A.; Lima-Filho, G.F.; Vital, M.V.C.; Carvalho, R.A.; Silveira, A.V.T.; Oliveira, L.C. 2005. Qualitative and quantitative benthic macroinvertebrate samplers in Cerrado stream: a comparative approach. Acta Limnologica Brasiliensia 17(2):123-128. Silveira, C.M. 2010. Controle Populacional de Moluscos In-vasores e outros Invertebrados: 4. Peixes como Agentes de Controle. In: Pereira, D.; Pinheiro, C.S.; Arenzon, A.; Rodri-guez, M.T.R., Paz, I.P.(org.). 2010. Livro de Resumos do 1º Workshop de Invertebrados Límnicos Invasores na América do Sul: Prevenção, Monitoramento e Controle. Centro de Ecologia (CENECO), Universidade Federal do Rio Grande do Sul, Porto Alegre, 45p.Simone, L.R.L. 2001. Phylogenetic analyses of Cerithioidea (Mollusca, Caenogastropoda) based on comparative morpho-logy. Arquivos de Zoologia 36 (2): 147-263.Simone, L.R.L. 2006. Land and Freshwater Molluscs of Bra-zil. São Paulo, EGB/Fapesp, 390p.Singer, S.; Van Fleet, A.L.; Viel, J.J.; Genevese, E.E. 1997. Bio-logical control of the zebra mussel Dreissena polymorpha and the snail Biomphalaria glabrata using Gramicidin S and D and moluscicidal strains of Bacillus. Journal of Industrial Microbi-ology and Biotechnology 18:226-231.Siraj, S.; Yousuf, A.R.; Bhat, F.A.; Parveen, M. 2010. The eco-logy of macrozoobenthos in Shallabugh wetland of Kashmir Himalaya, India. Journal of Ecology and the Natural Environ-ment 2(5):84-91. Skuza, L.; £abêcka, A. M.; Domaga£a. J. 2009. Cytogenetic and Morphological Characterization of Corbicula fluminalis (O. F. Müller, 1774) (Bivalvia: Veneroida: Corbiculidae):Taxonomic Status Assessment of a Freshwater Clam. Folia Biologica 57 (3-4):177-185. Smit, H.; De Vaate, A.B.; Fioole, A. 1992. Shell growth of the zebra mussel Dreissena polymorpha (Pallas) in relation to select-ed physicochemical parameters in the Lower Rhine and some associated lakes. Archiv für Hydrobiologie 124(3): 257-280.Smith, K.; Wood, S.A.; Mountfort, D.O.; Cary, S.C. 2012. Devel-opment of a real-time PCR assay for the detection of the invasive clam, Corbula amurensis, in environmental samples. Journal of Experimental Marine Biology and Ecology 412:52–57.Smith, T.E.; Stevenson, R.J.; Caraco, N.F.; Cole, J.J. 1998. Changes in phytoplankton community structure during the ze-bra mussel (Dreissena polymorpha) invasion of the Hudson Riv-er (New York). Journal of Plankton Research 20 (8):1567-1579.Soares, M.F.; Pires, M.J.R.; Mansur, M.C.D., Pereira, D.; Santos, C.P. 2008. Desenvolvimento e adequação de tecno-logias ambientais para o controle populacional do mexilhão dourado (Limnoperna fortunei). FEEVALE/PUCRS/CNPq/CT-HIDRO/ANA. http://cadastrocthidro.ana.gov.br/arquivos/relatorio_cnpq.pdf. Acessado em julho de 2009.Soares, M.F.; Pereira, D.; Santos, C.P.; Mansur, M.C.D.; Pi-res, M.; Breintebach, L.O.; Grespan, C. 2009. Toxicidade do sulfato de cobre ao mexilhão dourado, Limnoperna fortunei (Dunker, 1857), em água bruta. Journal of Brazilian Society of Ecotoxicology 4(1-3):37-48.Sotero-Santos, R.B., Souza e Silva, C.R., Verani, N.F., Nonaka, K.O.; Rocha, O. 2006. Toxicity of a cyanobacteria Bloom in Barra Bonita Reservoir (Middle Tietê River, São Paulo, Brazil). Ecotoxicology and Environmental Safety 64:163-170.

409
Sousa, R.; Rufino, M.; Gaspar, M.; Antunes, C.; Guilhermino, L. 2008a. Abiotic impacts on spatial and temporal distribution of Corbicula fluminea (Müller, 1774) in the River Minho Estu-ary, Portugal. Aquatic Conservation: Marine and Freshwater Ecosystems 18:98 –110. Sousa, R.; Antunes, C.; Guilhermino, L. 2008b. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: an overview. Annales de Limnologie. In-ternational Journal of Limnology 44 (2):85–94.Sousa, R.; Gutiérrez, J.L.; Aldridge, D.C. 2009. Non-indige-nous invasive bivalves as ecosystem engineers. Biological Inva-sions 11:2367–2385.Souto, L.S.; Brito, M.F.G.; Rosa, L.C. 2011. Melanoides tu-berculatus (Müller, 1774): a new threat to the conservation of aquatic native species in Sergipe, Brazil. Scientia Plena 7 (4):1-6.Souza, C.P. 1995. Molluscicide control of snail vectors of schisto-somiasis. Memórias do Instituto Oswaldo Cruz 90(2):165-168.Souza, J.R.B. de; Rocha, C.M.C. da; Lima, M.P.R. 2005. Ocorrência do bivalve exótico Mytilopsis leucophaeta (Con-rad) (Mollusca, Bivalvia) no Brasil. Revista Brasileira de Zoo-logia, 22(4):1206 – 1206.Souza, R.C.C.L.; Calazans, S.H.; Silva, E.P. 2009. Impacto das espécies invasoras no ambiente aquático. Ciência e Cultura 61(1):35-41.Stagl, V. 1993. Die Bruttasche von Melanoides tuberculata (O. F. Müller) (Gastropoda:Thiaridae). Annals Naturhistorisches Museum Wien 94/95:187-192.Stockwell, D.R.B.; Noble, I.R. 1992. Induction of sets of rules from animal distribution data: A robust and informative meth-od of analysis. Mathematic and Computer in Simulation 33: 385-390.Stockwell, D.R.B.; Peterson, D.P. 1999. The GARP modeling system: Problems and solutions to automated spatial predic-tion. International Journal of Geographic Information Sys-tems 1:143-158.Strauss, R.E. 1979. Reliability estimates for Ivlev’s electivity index, the forage ratio, and proposed linear index of food selection. Transactions of the American Fisheries Society 108:344-352.Strong, E.E.; Colgan, D.J.; Healy, J.M.; Lydeard, C.; Ponder, W.F.; Glaubrecht, M. 2011. Phylogeny of the gastropod super-family Cerithioidea using morphology and molecules. Zoolo-gical Journal of the Linnean Society 162 43–89.Suriani, A.L.; França, R.S.; Rocha, O. 2007. A malacofauna bentônica das represas do médio Rio Tietê (São Paulo, Brasil) e uma avaliação ecológica das espécies exóticas invasoras, Me-lanoides tuberculata (Müller) e Corbicula fluminea (Müller). Revista Brasileira de Zoologia 24(1):21-32.Suslick, K.S. 1989. The Chemical Effects of Ultrasound. Scien-tific American 2(260): 80-86Suslick, K.S. 1994. Yearbook of Science and the Future. Ency-clopedia Britannia, 139. Sylvester, F.; Dorado, J.; Boltovskoy, D.; Juárez, A.; Cataldo, D. 2005. Filtration rates of the invasive pest bivalve Limnoper-na fortunei as a function of size and temperature. Hydrobio-logia 534:71–80.Sylvester, F.; Boltovskoy, D.; Cataldo, D.H. 2007a. The invasi-ve bivalve Limnoperna fortunei enhances benthic invertebrates densities in South America floodplain rivers. Hydrobiologia 589:15-27. Sylvester, F.; Boltovskoy, D.; Cataldo, D. 2007b. Fast response of freshwater consumers to a new trophic resource: predation on the recently introduced Asian bivalve Limnoperna fortu-nei in the lower Paraná River, South America. Austral Ecology 32:403–415.
Takeda, A.M.; Higuti, J.; Fujita, D.S.; Bubena, M.R. 2000. Pro-Pro-liferação de uma espécie de bivalve invasora, Corbicula flumi-nea, na área alagável do Alto Rio Paraná (Brasil). In: Seminário Brasileiro sobre Água de Lastro 1, Arraial do Cabo, p.11.Takeda, A.M.; Mansur, M.C.D.; Fujita, D.S. 2005. Ocorrência de moluscos bivalves em diferentes reservatórios. p.161-167. In: Rodrigues L., Thomaz, S. M.; Agostinho, A. A.; Gomes; L.C. (Org.). Biocenoses em Reservatórios. 1 ed. São Carlos, RIMA. Takeda, A.M.; Fujita, D.S.; Mansur, M.C.D.; Callil, C.T.; Oli-veira, M.D. (no prelo). Amostragens de larvas, bentos e experi-mentos in loco: vantagens e desvantagens para monitoramento e detecção de Limnoperna fortunei (Dunker, 1857) (Mollusca, Mytilidae)” e “Mexilhão-dourado na Bacia do Paraná, com ênfase ao Estado do Paraná” Tópicos em Malacologia – Ecos do XIX Ebram, organizado por Sonia Barbosa dos Santos, Alexandre Dias Pimenta, Silvana Carvalho Thiengo e Monica Ammon Fernandez.Takenaka, R.A. 2007. Avaliação da Toxicidade de Microcystis aeruginosa e de florações naturais de cianobactérias de reserva-tórios do Rio Tietê, SP. Tese de Doutorado. Escola de Engenha-ria de São Carlos, Universidade de São Paulo, São Carlos. 330p.Takenaka, R.A.; Dellamano-Oliveira, M.J.; Rocha, O. 2007. Toxicidade de extratos de florações de cianobactérias de reser-vatórios do Rio Tietê, SP, aos dafinídeos Ceriodaphnia dubia e Ceriodaphnia silvestrii (Cladocera, Crustacea). Journal of the Brazilian Society of Ecotoxicology 2:147-156.Taurusman, A.A. 2010. Community structure of macrozoo-benthic feeding guilds in responses to eutrophication in Jakar-ta Bay. Biodiversitas 11(3):133-138. Te, G.A. 1980. New classification system for the family Phy-sidae (Pulmonata:Basommatophora). Archiv fur Mollusken-kunde 110:179-184.Teles, H.M.S.; Vaz J.F.; Glasser C.M.; Ciaravolo, R.M.C. 1991. Levantamento planorbídico do Estado de São Paulo, Brasil. 2ª Região Administrativa. Revista de Saúde Pública 23(3):179-83.Ter Harr, G. 2007. Therapeutic applications of ultrasound. Prog-ress in Biophysics and Molecular Biology 93 (1-3): 111-129.Terra, N.R.; Lemieszek, M.B.; Lemos, C.T.; Leite, E.H. 2007. Presença de Limnoperna fortunei (Dunker, 1857) nas bacias hidrográficas do Uruguai e Lago Guaíba, Rio Grande do Sul, Brasil. FEPAM em Revista 1(1):12-19.Thiengo, S.C.; Fernandez, M.A.; Boaventura; M.F.; Stortti; M.A. 1998. A survey of freshwater gastropods in the micror-A survey of freshwater gastropods in the micror-região serrana of the state of Rio de Janeiro, Brazil. Memórias do Instituto Oswaldo Cruz 93 (Suppl 1):233-234.Thiengo, S.C.; Mattos, A.C.; Boaventura, M.F.; Fernandez, M.A. 2004. Freshwater snails and schistosomiasis mansoni in the state of Rio de Janeiro, Brazil: IV - Sul Fluminense me-soregion. Memórias do Instituto Oswaldo Cruz 99:275-280.Thiengo, S.C.; Santos, S.B.; Fernandez, M.A. 2005. Malaco-fauna límnica da área de influência do Lago da usina hidre-létrica de Serra da Mesa, Goiás, Brasil. I. Estudo qualitativo. Revista Brasileira de Zoologia 22 (4):867-874.Thiengo, S.C.; Fernandez, M.A.; Mattos, A.C.; Barbosa, A.F. 2007. Dispersão do molusco introduzido Melanoides tuber-culatus (Muller, 1774) (Gastropoda; Thiaridae) no Brasil. p. 101-106. In: Santos, S.B.; Pimenta, A.D.; Thiengo. S.A.R.C.; Fernandez, M.A.; Absalão, R.S. (Org.). Tópicos em Malacolo-gia - Ecos do XVIII Encontro Brasileiro de Malacologia. So-ciedade Brasileira de Malacologia, Rio de Janeiro, xiv +365p.Thomaz, S.M. 2006. Efeitos de fatores limnológicos sobre a colonização e estrutura de assembléias de macrófitas aquáticas. In: Mariath, J.E.A.; Santos, R.P. (Org.). Os avanços da botâni-ca no início do século XXI: morfologia, fisiologia, taxonomia, ecologia e genética. Porto Alegre: Sociedade Botânica do Bra-sil, p. 124-139.

410
Thorp, J.H.; Casper, A.F. 2002. Potential effects on zooplank-Potential effects on zooplank-ton from species shifts in planktivorous mussels: a field experi-ment in the St Lawrence River. Freshwater Biology 47:107-119.Toft, J.D.; Simenstead, A.D.; Cordell, J. R.; Grimaldo, L. F. 2003. The effects of introduced water hyacinth on habitat structure invertebrates assemblages, and fish diets. Estuaries 26(3):746-758.Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; Mckenzie, V.J.; Kuris, A.M. 2003. Introduced species and their missing parasi-tes. Nature 421:628-630.Torgan, L.C.; Salomoni, S.E.; Bicca, A.B. 2009. Diatomáceas so-Diatomáceas so-bre Limnoperna fortunei (Dunker), molusco introduzido no Lago Guaíba, Sul do Brasil. Revista Brasileira de Botânica 32(1): 23-31.Toro, J.E. 1998. Molecular identification of four species of mussels from southern Chile by PCR-based nuclear markers: the potential use in studies involving planktonic surveys. Jour-nal of Shellfish Research 17: 1203-1205.Tundisi, J. G; Tundisi, T. M. 2008. Limnologia. Oficina de Textos, São Paulo. 632p.Tundisi, J.G.; Matsumura-Tundisi, T.; Pareschi, D.C.; Luzia, A.P.; Von Haeling, P.H.; Frollini, E.H. 2008. A bacia hidrográ-fica do Tietê/Jacaré: estudo de caso em pesquisa e gerencia-mento. Estudos Avançados 22(63):159-172. Uliana, E.R.; Callil, C.T. 2006. Avaliação do ciclo sexual do mexilhão dourado, Limnoperna fortunei (Dunker, 1857), no Rio Paraguai, Porto Esperança – MS. TCC. Graduação em Ciências Biológicas– Universidade Federal de Mato Grosso – UFMT Cuiabá, Brasil. 33p. Uliana, E.R.; Callil, C.T.; Oliveira, M.D. 2006. Avaliação temporal da reprodução do Mexilhão dourado, Limnoperna fortunei (Dunker, 1857) no Rio Paraguai, Corumbá, MS. In: IV Encontro Nacional sobre Água de Lastro, 2006, Arraial do Cabo. Anais do IV Encontro Nacional sobre Água de Lastro.Uliana, E.R.; Callil, C.T.; Mansur, M.C.D.; Oliveira, M.D.; Takeda, A.M.; Santos, C.P.; Pinillos, A.C.M.; Neis, E. 2008. Estudo da reprodução de Limnoperna fortunei (Dunker, 1857) em três estados brasileiros. In: V Seminário Brasileiro sobre Água de Lastro, 2008, Arraial do Cabo - RJ. Anais do V En-contro Nacional sobre Água de Lastro, 2008.Umadevi, K.; Madhavi, R. 2006. The life cycle of Haplorchis pumilio (Trematoda:Heterophyidae) from the Indian region. Journal of Helminthology 80:327–332.Uryu, Y.; Iwasaky, K.; Hinque, M. 1996. Laboratory experiments on behaviour and movement of a freshwater mussel, Limnoperna fortunei (DUNKER) Journal of a Mollusan Studies 62: 327-341.Vadopalas, B.; Bouma, J.V.; Jackels, C.R.; Friedman, C.S. 2006. Application of real-time PCR for simultaneous identifi-cation and quantification of larval abalone. Journal of Experi-mental Marine Biology and Ecology 334:219–228.Vale, R.S.; Beasley, C.R.; Tagliaro, C.H.; Mansur, M.C.D. 2005. The glochidium and marsupium of Castalia ambigua ambigua Lamarck, 1819, from northern Brazil. American Ma-lacological Bulletin 20:43-48.Valéry, L.; Fritz, H.; Lefeuvre, J.-C.; Simberloff, D. 2008a. Ecosystem-level consequences of invasions by native species as a way to investigate relationships between evenness and ecosystem function. Biological Invasions 11(3):609-617.Valéry, L.; Fritz, H.; Lefreuve, J.; Simberloff, D. 2008b. In se-arch of a real definition of the biological invasion phenome-non itself. Biological Invasions 10:1345-1351.Valiela, I. 1995. Marine Ecological Processes. 2ª ed. Springer-Verlag, New York. 686p.Van Lenteren, J.C. 2008. IOBC Internet Book of Biological Control Version 5. Disponível em: http://www.iobc-global.org/publications_iobc_internet_book_of_biological_control.html. Acesso em: 25 agosto 2011.
Vanderploeg, H.A.; Liebig, J.R.; Carmichael, W.W.; Agy, M.A.; Johengen, T.H.; Fahnenstiel, G.L.; Nalepa, T.F. 2001. Zebra mussel (Dreissena polymorpha) selective filtration promoted toxic Microcystis blooms in Saginaw Bay (Lake Huron) and Lake Erie. Canadian Journal of Fisheries and Aquatic Sciences 58:1208-1221.Vanderploeg, H.A.; Johengen, T.H.; Liebig, J.R. 2009. Feed-back between zebra mussel selective feeding and algal compo-sition affects mussel condition: did the regime changer pay a price for its success? Freshwater Biology 54:47–63.Vaz, J.F.; Elmor, M.R.D.; Gonçalves, L.M.C.; Ishihata, G.K. 1983. Resultados do levantamento planorbídico da área de Presidente Prudente. Estado de São Paulo. Revista do Instituto de Medicina Tropical de São Paulo 25:120-126.Vaz, J.F.; Teles, H.M.S.; Takaku, L. 1985. Levantamento pla-norbídico do Estado de São Paulo. 7ª Região Administrativa. Ciência e Cultura 37(12):2057-2062.Vaz, J.F.; Teles, H.M.S.; Correa, M.A.; Leite, S.P.S. 1986a. Ocor-rência no Brasil de Thiara (Melanoides) tuberculata (O.F. Müller, 1774) (Gastropoda, Prosobranchia), primeiro hospedeiro inter-mediário de Clonorchis sinensis (Cobbold, 1875) (Trematoda, Plathyhelmintes). Revista de Saúde Pública 20(4):318-322.Vaz, J.F.; Teles, H.M.S.; Fabbro, A.L.D.; Rosa W.S. 1986b. Le-vantamento planorbídico do Estado de São Paulo. 6ª Região Administrativa. Revista de Saúde Pública 20(5): 352-361.Vaz, J.F.; Mantegazza, E.; Teles, H.M.S.; Leite, S.P.S.; Morais, L.V.C. 1987. Levantamento planorbídico do Estado de São Paulo. 4ª Região Administrativa. Revista de Saúde Pública 21(5):371-379.Vaz, J.F.; Elmor, M.R.D.; Gonçalves, L.M.C. 1992. Levanta-mento planorbídico do Estado de São Paulo. 8ª Região Admi-nistrativa (Grande Área de São José do Rio Preto). Revista do Instituto de Medicina Tropical de Saõ Paulo 34(6): 527-34.Veitenheimer-Mendes, I.L. 1981. Corbicula manilensis, (Phi-lippi, 1844) molusco asiático, na bacia do Jacuí e do Guaíba, Rio Grande do Sul, Brasil (Bivalvia, Corbiculidae). Iheringia, Série Zoologia 60:63-74.Veitenheimer-Mendes, I.L.; Mansur, M.C.D. 1978. Myce-topoda legumen (Martens, 1988): lasídio e desenvolvimento parasitário (Bivalvia, Mycetopodidae). Rio de Janeiro, Revista Brasileira de Biologia 38(3):531-536.Vieira, J.P.; Garcia, A.M.; Moraes, L., 2010. A assembleia de peixes, in: Seelinger, U.; Odebrecht, C. (Eds.), O Estuário da Lagoa dos Patos. Um século de transformações. FURG, Rio Grande, pp. 79-88. Villalobos, D.M.; Paz, A.O.; González, M.T. 2010. Hermaph-Hermaph-roditism in Marine Mussel Perumytilus purpuratus (Lamarck, 1819), (Mollusca: Mytilidae). International Journal of Mor-phology 28(2):569-573.Villar, C.; Stripekis, J.; D’Huicque, L.; Tudino, M. 1999. Cd, Cu, and Zn concentration in sediments and the invasive bi-valves Limnoperna fortunei and Corbicula fluminea at the Rio de La Plata basin, Argentina. Hydrobiologia 416:41-49.Villela, I.V.; Oliveira, I. M.; Silva, J.; Henriques, J.A.P. 2006. DNA Damage and Repair in Haemolymph Cells of Golden Mussel (Limnoperna fortunei) Exposed to Environmental Contaminants. Mutation Research 697:78-86.Vinuesa, J.H. 1977. Sobre un caso de hermafroditismo en la cholga Aulacomya ater (Molina). Physis (37):63-65.Vitule J.R.S.; Freire C.A.; Simberlo, D. 2009. Introduction of non - native freshwater fish can certainly be bad. Fish and Fisheries 10: 98-108.Volkmer-Ribeiro, C.; Mothes de Morais, B.; Rosa Barbosa, R.; Mansur, M.C.D.; Veitenheimer-Mendes, I. L. 1984. Um estudo do bentos em raízes de Eichhornia azurea (Sw.) Kunt, do curso inferior de um rio Subtropical Sul-americano. Revista Brasileira de Biologia 44(2):125-132.

411
Volkmer-Ribeiro, C.; Guadagnin, D.L.; de Rosa-Barbosa, R.; Silva, M.M.; Drügg-Hanh, S.; Lopes-Pitoni, V.L.; Gas-tal, H.A.O.; Barros, M.P.; Demaman, L.V. 2004. A polyeth-A polyeth-ylenetherephthalate (PET) device for sampling freshwater benthic macroinvertebrates. Brazilian Journal of Biology 64(3A):531-541. Wacker, A.; Von Elert, E. 2003. Food quality controls repro-duction of the zebra mussel (Dreissena polymorpha). Oecolo-gia 135:332-338.Wada, T. 2004. Strategies for controlling the Apple Snail Pomacea canaliculata (Lamarck) (Gastropoda: Ampullaridae) in Japanese direct-Sown Paddy Fields. JARQ 38(2):75-80.Waikagul, J.; Dekumyoy, P., Yoonuan, T.; Praevanit, R. 2006. Conjunctiva philophthalmosis: a case report in Thailand. American Journal of Tropical Medicine and Hygiene 74:848-849.Waller, D. L.; Rach, J. J.; Cope, W.G.; Marking, L. L.; Fischer, S. W.; Dabrowska, H. 1993. Toxicity of candidate mollusci-cides to zebra mussels (Dreissena polymorpha) and selected nontarget organisms. Journal of Great Lakes Research. 19(4):695-702.Wanganeo, A.; Kumar, P.; Wanganeo, R.; Sonaullah, F. 2011. Variation in benthic population in two basins of Bhoj wetland, Bhopal. International Journal of Environmental Sciences 1(7):2004-2017. Ward, J.M.; Ricciardi, A. 2007. Impacts of Dreissena invasions in benthic macroinvertebrate communities: a meta-analysis. Diversity and Distributions 13:155–165.WHO. 2008. Guidelines for drinking-water quality. Third edi-tion incorporating the first and second addenda. Volume 1. Recommendations. Geneva, World Health Organization.Wildridge, P.J.; Werner, R.G.; Doherty, F.G.; Neuhauser, E.F., 1998. Acute toxicity of potassium to the adult zebra mussel Dreissena polymorpha. Archives of Environmental Contami-nation and Toxicology 34, 265-270.Wilhelm, S.; Adrian, R. 2007. Long-term response of Dreisse-na polymorpha larvae to physical and biological forcing in a shallow lake. Oecologia 151:104-114.Wingard, G.L.; Murray, J.B.; Schill, W.B.; Phillips, E.C. 2008. Red-rimmed melania (Melanoides tuberculatus) – A snail in Biscayne National Park, Florida - Harmful invader or just a nuisance? USGS Fact Sheet 2008-3006:1-6.
Wolfe, R.L. 1990. Ultraviolet disinfection of potable water. Current technology and research needs. Environ. Science Te-chnology 24(6): 768-773. Wright, D.A.; Magee, J. A.; Setzler-Hamilyon, E. M.; Chalker-Scott, L.; Morgan, G. L. 1997. Use of high energy monochromatic UV light to kill dreissenid larvae. p. 467-476. In: D’Itri, F.M. (Eds.) Zebra Mussel and Aquatic Nui-sance Species. Proceedings, International Zebra Mussel and Other Aquatic Nuisance Species Conference. CRC PRESS, Boca Raton, 638p.Würdig, N.L.; Cenzano, C.S.S.; Motta-Marques, D. 2007. Macroinvertebrate communities structure in different invi-ronments on the Taim Hydrological System in the state of Rio Grande do Sul, Brazil. Acta Limnologica Brasiliensia 19(4):427-438.Ximenes, R.F. 2011. Tamanho, parasitismo e maturidade re-produtiva em Melanoides tuberculatus (Müller, 1774) (Gas-tropoda, Thiaridae) da Vila do Abraão, Ilha Grande, Angra dos Reis, RJ. Monografia de Bacharelado, Curso de Ciências Biológicas, Universidade do Estado do Rio de Janeiro, Brasil, 59p.Yebra, D.M.; Kill, S.; Dam-Johansen, K. 2004. Antifouling technology: past, present and future steps towards efficient and environmentally friendly antifouling coatings. Progress in Organic Coatings 50: 75-104.Zagatto, P.A; Bertoletti, E. 2006. Ecotoxicologia aquática – Princípios e aplicações. Rima, São Paulo, 478 p.Zaiko, A.; Daunys, D.; Olenin, S. 2009. Habitat engineering by the invasive zebra mussel Dreissena polymorpha (Pallas) in a boreal coastal lagoon: impact on biodiversity. Helgoland Ma-rine Research 63(1):85–94.Zanella, O.; Marenda, L.D. 2002. Ocorrência de Limnoperna fortunei na Central Hidrelétrica de Itaipu, p. 41. In: V Con-gresso Latinoamericano de Malacologia. São Paulo, Instituto Butantan/ Instituto de Biociências – USP. Zar, J.H. 1999. Biostatistical Analysis. Prentice-Hall, 4th Edi-tion, 718p.Zoltak, J. 2008. Ontario zebra mussel distribution. Ontario Federation of Anglers and Hunters. <http://www.glu.org/sites/default/files/ZebraMussel_100dpi.pdf>. Acesso em: 26 julho 2011.





















