
Molluscan species of minor commercial interest in Hellenic seas: Distribution, exploitation and...
42
Medit. Mar. Sci., 9/1, 2008, 77-118 77 Introduction Exploitation of mollusca as food and as ornaments in Hellas has a history dating back to the Paleolithic age (SHACKLE- TON & VAN ANDEL, 2007). However, the development of high density human populations near the coast has resulted in unprecedented and unsustainable levels of pressure nowadays. Unfortunately, sea- shells are collected in those areas least capable of withstanding exploitation. While Mediterranean Marine Science Volume 9/1, 2008, 77-118 Molluscan species of minor commercial interest in Hellenic seas: Distribution, exploitation and conservation status S. KATSANEVAKIS 1 , E. LEFKADITOU 1 , S. GALINOU-MITSOUDI 2 , D. KOUTSOUBAS 3 and A. ZENETOS 1 1 Hellenic Centre for Marine Research, Institute of Marine Biological Resources, P.O. Box 712, 19013, Anavissos, Attica, Hellas 2 Fisheries & Aquaculture Technology, Alexander Technological Educational Institute of Thessaloniki, 63200, Nea Moudania, Hellas 3 Department of Marine Sciences, School of Environment, University of the Aegean, 81100, Mytilini, Hellas e-mail: [email protected] Abstract Recognising the need for data gathering at taxon level besides the taxa of commercial interest and those listed as endangered in the Protocol of the Barcelona Convention, but still exploited by Man, the present study attempts to gather and review the available scientific information on molluscs of minor commercial importance in order to assist in the adequate management and protection of their popula- tions. Forty one species (18 gastropods, 13 bivalves, and 10 cephalopods) of minor commercial interest are treated in the present work with details on their biogeographic distribution, exploitation and conser- vation status in Hellas. Apart from a few species (e.g., Pinna nobilis, Lithophaga lithophaga, Donacilla cornea ), and these only at a local scale, there is no population assessment in Hellenic seas. The existing legislation for eleven of these species is not enforced in practice, and seems insufficient to guarantee their conservation. It is suggested that targeted collection data, networked nationally and internationally, should be promoted so this invaluable source of biodiversity information can be accessed for conserva- tion and planning purposes. Keywords: Mollusca exploitation; Conservation; Hellas. Review Article
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Molluscan species of minor commercial interest in Hellenic seas: Distribution, exploitation and...

Medit. Mar. Sci., 9/1, 2008, 77-118 77
Introduction
Exploitation of mollusca as food and asornaments in Hellas has a history datingback to the Paleolithic age (SHACKLE-TON & VAN ANDEL, 2007). However,
the development of high density humanpopulations near the coast has resulted inunprecedented and unsustainable levels ofpressure nowadays. Unfortunately, sea-shells are collected in those areas leastcapable of withstanding exploitation. While
Mediterranean Marine ScienceVolume 9/1, 2008, 77-118
Molluscan species of minor commercial interest in Hellenic seas: Distribution,exploitation and conservation status
S. KATSANEVAKIS1, E. LEFKADITOU1, S. GALINOU-MITSOUDI2, D. KOUTSOUBAS3
and A. ZENETOS1
1 Hellenic Centre for Marine Research, Institute of Marine Biological Resources, P.O. Box 712, 19013, Anavissos, Attica, Hellas2 Fisheries & Aquaculture Technology, Alexander Technological Educational Institute of Thessaloniki,63200, Nea Moudania, Hellas3 Department of Marine Sciences, School of Environment, University of the Aegean, 81100, Mytilini, Hellas
e-mail: [email protected]
Abstract
Recognising the need for data gathering at taxon level besides the taxa of commercial interest andthose listed as endangered in the Protocol of the Barcelona Convention, but still exploited by Man, thepresent study attempts to gather and review the available scientific information on molluscs of minorcommercial importance in order to assist in the adequate management and protection of their popula-tions. Forty one species (18 gastropods, 13 bivalves, and 10 cephalopods) of minor commercial interestare treated in the present work with details on their biogeographic distribution, exploitation and conser-vation status in Hellas. Apart from a few species (e.g., Pinna nobilis, Lithophaga lithophaga, Donacillacornea), and these only at a local scale, there is no population assessment in Hellenic seas. The existinglegislation for eleven of these species is not enforced in practice, and seems insufficient to guarantee theirconservation. It is suggested that targeted collection data, networked nationally and internationally,should be promoted so this invaluable source of biodiversity information can be accessed for conserva-tion and planning purposes.
Keywords: Mollusca exploitation; Conservation; Hellas.
Review Article

shelled mollusca are heavily exploited, somecephalopod species are impacted by uncon-trolled and unreported fishing, which mayhave a significant effect on their populations.
A total of 1160 mollusc species (1Solenogastra, 2 Caudofoveata, 771 Gas-tropoda, 308 Bivalvia, 19 Polyplacophora,12 Scaphopoda and 47 Cephalopoda) havebeen recorded so far in Hellenic seas(KOUTSOUBAS, 2004; CHINTIRO-GLOU et al., 2005; ZENETOS et al., 2005;LEFKADITOU, 2007; DELAMOTTE &VARDALA-THEODOROU, 2007). Atotal of 27 of these species (seventeenbivalves, four gastropods and sixcephalopods), comprising less than 2.5 %of the molluscan fauna of the Hellenicseas, have a commercial interest, particu-larly in fisheries and aquaculture sincethey are collected and/or cultivated forhuman consumption. The status of thefishing of these commercially importantmolluscs has been recently reviewed(KOUTSOUBAS et al., 2007; LEFKA-DITOU, 2007).
However, apart from those commer-cially important species, there are manyother molluscs, which, although not com-mercially exploited, are either systemati-cally or occasionally collected for humanconsumption and other uses (fishing bait,jewellery, decoration, private and publiccollections). Furthermore, several species,for example some common small-sizedcephalopods, constitute trawl fisheries by-catches which may have a marketable frac-tion. Bivalve and gastropod molluscs arealso frequently used to biomonitor heavymetal pollution in the marine environ-ment. In most cases, the distribution, thedegree of exploitation, and the state ofthese populations have not been assessedin Hellenic seas. Uncontrolled and unre-ported fishing or other human activities
have been destructive and have caused asignificant decline in the Mediterraneanpopulations of several species (e.g., Pinnanobilis) and have brought others to nearextinction (e.g., Patella ferruginea)(BOUDOURESQUE, 2004). In addition,basic knowledge on the biology and ecolo-gy of these species is either lacking or isscattered in the literature.
The red list species approach to con-servation requires detailed knowledgeconcerning individual taxa (distribution,declines, threatening processes, biologicalattributes, etc.). However, it addressesonly the tip of the biodiversity iceberg. Thefirst step in knowledge acquisitionrequired for the taxon-based approach tobiodiversity and conservation is the studyand documentation of at least the majorcomponents of the fauna. Hence, the pres-ent study attempts to gather and reviewthe available scientific information onmolluscs of minor commercial importancein order to assist in the adequate manage-ment and protection of their populations.
Materials and Methods
Forty one species (18 gastropods, 13bivalves, and 10 cephalopods) of minorcommercial interest were reviewed in thepresent work (Table 1). The distribution ofthe listed species in Hellenic seas was gen-erally based on ZENETOS (1986; 1996),KOUTSOUBAS (1992), KOUTSOUBASet al. (1992, 1997, 1999, 2000a,b),ZENETOS et al. (2004, 2007), ANTO-NIADOU et al. (2005), DELAMOTTE &VARDALA-THEODOROU (2007),LEFKADITOU (2007), EVAGELO-POULOS & KOUTSOUBAS (2008), greyliterature such as academic dissertations andHCMR or EU technical reports, and/oron unpublished/personal observations.
Medit. Mar. Sci., 9/1, 2008, 77-11878
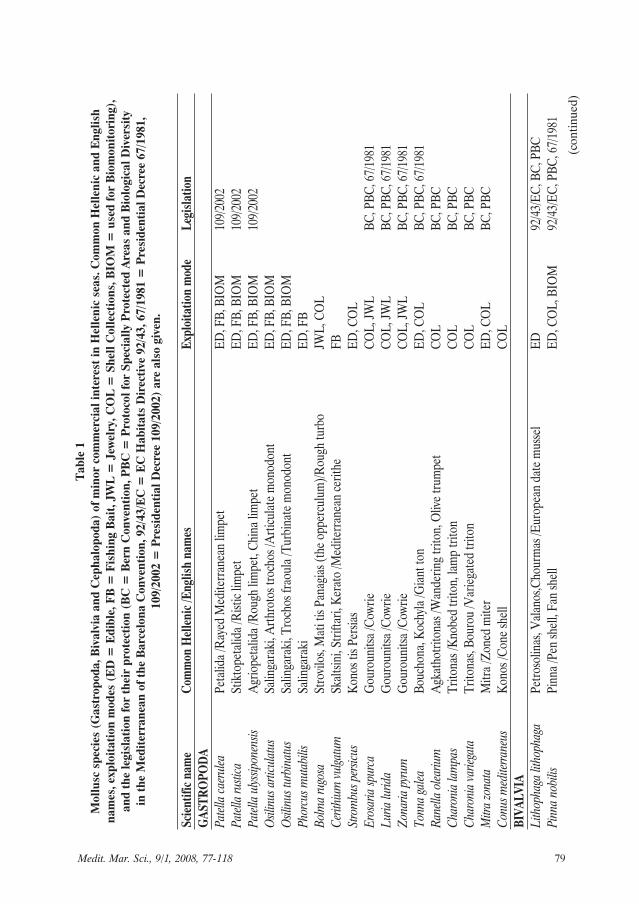
Medit. Mar. Sci., 9/1, 2008, 77-118 79
Tab
le 1
Mol
lusc
spe
cies
(G
astr
opod
a, B
ival
via
and
Cep
halo
poda
) of
min
or c
omm
erci
al in
tere
st in
Hel
leni
c se
as. C
omm
on H
elle
nic
and
Eng
lish
nam
es, e
xplo
itat
ion
mod
es (
ED
= E
dibl
e, F
B =
Fis
hing
Bai
t, JW
L =
Jew
elry
, CO
L =
She
ll C
olle
ctio
ns, B
IOM
= u
sed
for
Bio
mon
itor
ing)
,an
d th
e le
gisl
atio
n fo
r th
eir
prot
ecti
on (
BC
= B
ern
Con
vent
ion,
PB
C =
Pro
toco
l for
Spe
cial
ly P
rote
cted
Are
as a
nd B
iolo
gica
l Div
ersi
tyin
the
Med
iter
rane
an o
f the
Bar
celo
na C
onve
ntio
n, 9
2/43
/EC
= E
C H
abit
ats
Dir
ecti
ve 9
2/43
, 67/
1981
= P
resi
dent
ial D
ecre
e 67
/198
1,10
9/20
02 =
Pre
side
ntia
l Dec
ree
109/
2002
) ar
e al
so g
iven
.
Scie
ntifi
c na
me
Com
mon
Hel
leni
c /E
nglis
h na
mes
Expl
oita
tion
mod
eLe
gisl
atio
nG
ASTR
OPO
DA
Patel
la c
aeru
leaPe
talid
a /R
ayed
Med
iterr
anea
n lim
pet
ED, F
B, B
IOM
109/
2002
Patel
la ru
stica
Stik
tope
talid
a /R
istic
limpe
tED
, FB,
BIO
M10
9/20
02Pa
tella
ulys
sipon
ensis
Agr
iope
talid
a /R
ough
lim
pet,
Chin
a lim
pet
ED, F
B, B
IOM
109/
2002
Osil
inus
arti
cula
tus
Salin
gara
ki, A
rthro
tos t
roch
os /A
rticu
late
mon
odon
tED
, FB,
BIO
MO
silin
us tu
rbin
atus
Salin
gara
ki, T
roch
os fr
aoul
a /T
urbi
nate
mon
odon
tED
, FB,
BIO
MPh
orcu
s mut
abili
sSa
linga
raki
ED, F
BBo
lma
rugo
saSt
rovi
los,
Mat
i tis
Pana
gias
(the
opp
ercu
lum
)/Rou
gh tu
rbo
JWL,
CO
LC
erith
ium
vul
gatu
mSk
altsi
ni, S
trifta
ri, K
erat
o /M
edite
rran
ean
cerit
heFB
Stro
mbu
s per
sicus
Kon
os ti
s Per
sias
ED, C
OL
Eros
aria
spur
caG
ouro
units
a /C
owrie
COL,
JWL
BC, P
BC, 6
7/19
81Lu
ria lu
rida
Gou
roun
itsa
/Cow
rieCO
L, JW
LBC
, PBC
, 67/
1981
Zona
ria p
yrum
Gou
roun
itsa
/Cow
rieCO
L, JW
LBC
, PBC
, 67/
1981
Tonn
a ga
leaBo
ucho
na, K
ochy
la /G
iant
ton
ED, C
OL
BC, P
BC, 6
7/19
81Ra
nella
olea
rium
Agk
atho
trito
nas /
Wan
derin
g tri
ton,
Oliv
e tru
mpe
tCO
LBC
, PBC
Cha
roni
a la
mpa
sTr
itona
s /K
nobe
d tri
ton,
lam
p tri
ton
COL
BC, P
BCC
haro
nia
varie
gata
Trito
nas,
Bour
ou /V
arie
gate
d tri
ton
COL
BC, P
BCM
itra
zona
taM
itra
/Zon
ed m
iter
ED, C
OL
BC, P
BCC
onus
med
iterra
neus
Kon
os /C
one
shel
lCO
LBI
VALV
IALi
thop
haga
lith
opha
gaPe
troso
linas
, Val
anos
,Cho
urm
as /E
urop
ean
date
mus
sel
ED92
/43/
EC, B
C, P
BCPi
nna
nobi
lisPi
nna
/Pen
shel
l, Fa
n sh
ell
ED, C
OL,
BIO
M92
/43/
EC, P
BC, 6
7/19
81
(con
tinue
d)

Medit. Mar. Sci., 9/1, 2008, 77-11880
Tab
le 1
(co
ntin
ued)
Scie
ntifi
c na
me
Com
mon
Hel
leni
c /E
nglis
h na
mes
Expl
oita
tion
mod
eLe
gisl
atio
nBI
VALV
IAPi
ncta
da ra
diat
aM
arga
ritof
oro
strid
i /R
ayed
pea
rl oy
ster
ED, J
WL
, BIO
MPt
eria
hiru
ndo
Fter
ostri
do /E
urop
ean
wing
yste
rED
, CO
LC
hlam
ys v
aria
Kal
ogria
, Chl
amyd
a /V
arie
gate
d sc
allo
pED
, CO
LC
rass
adom
a m
ultis
triat
aG
ram
mot
o ch
teni
/Litt
le b
ay sc
allo
pED
, CO
LSp
ondy
lus g
aede
ropu
sG
aido
urop
odar
o, V
assil
iko
strid
i /Eu
rope
an th
orny
yste
rED
, CO
LC
rass
ostre
a gig
asPo
rtoga
liko
strid
i /G
iant
(or P
acifi
c) cu
pped
oys
ter
ED, B
IOM
Don
acill
a co
rnea
Am
mok
ochy
lo, M
ikri
ahiv
ada
/Ban
ded
wedg
e sh
ell
ED, B
IOM
Solen
mar
ginat
usSo
linas
, Am
mos
olin
as /E
urop
ean
razo
r cla
mED
, FB,
BIO
M10
9/20
02En
sis m
inor
Solin
as, A
mm
osol
inas
/Gia
nt ra
zor c
lam
ED, F
B10
9/20
02So
lecur
tus s
trigil
atus
Sam
ari,
Zam
pona
ki /S
andw
ichED
, FB,
CO
L10
9/20
02Ph
olas
dac
tylus
Dak
tilo,
Fte
ra a
ggel
on, F
olad
a /C
omm
on p
iddo
ckED
, FB
BC, P
BCC
EPH
ALO
POD
AAl
loteu
this
med
iaK
alam
arak
i /M
idsiz
e sq
uid
EDLo
ligo
forb
esi
Kal
amar
i /V
eine
d sq
uid
EDTo
daro
des s
agitt
atus
Thra
psal
o, K
atam
achi
/Eur
opea
n fly
ing
squi
dED
, FB
Toda
rops
is eb
lana
eTh
raps
alo
/Les
ser f
lying
squi
dED
, FB
Sepi
a ele
gans
Eleg
ant c
uttle
fish
EDSe
pia
orbi
gnya
naK
okki
noso
upia
/Pin
k cu
ttlef
ishED
Ross
ia m
acro
som
aM
ikro
soup
ia /S
tout
bod
tail
EDRo
ndele
tiola
min
orSo
upits
a /L
entil
bod
tail
EDSe
piett
a ow
enia
naSo
upits
a /C
omm
on b
odta
ilED
Scae
urgu
s uni
cirrh
usM
onok
ero
chta
podi
/Uni
horn
ed o
ctop
usED

A general map of Hellas with thenames of the gulfs and other locationsmentioned in the text is given in Figure 1.General information on the biology andecology of the species was gathered after athorough search of international andnational literature. Nomenclature for gas-tropods and bivalves followed the CheckList of European Marine MolluscaCLEMAM (http://www.somali.asso.fr/clemam/biotaxis.php), whereas that forcephalopods followed the classification ofliving coleoid species of the Mediter-ranean Sea by MANGOLD & vonBOLETZKY (1987).
Legislation concerning the conserva-tion of each species is reported whereavailable (Table 1). The existing legislationfor the conservation of marine species
includes the Presidential Decree 67/1981,the EC Habitats Directive 92/43, the BernConvention, and the Protocol for SpeciallyProtected Areas and Biological Diversity inthe Mediterranean of the Barcelona Con-vention (hereafter: Protocol of theBarcelona Convention). For speciesincluded in Annex IV of the Directive92/43/EC, all forms of deliberate capture orkilling of specimens in the wild, deliberatedisturbance, particularly during the periodof breeding, deliberate destruction or tak-ing of eggs from the wild, and deteriorationor destruction of breeding sites or restingplaces are strictly prohibited. For speciesincluded in Annex II of the Bern Conven-tion, the collection, keeping in captivity,killing, destruction or collecting of eggs,disturbance especially during the reproduc-
Medit. Mar. Sci., 9/1, 2008, 77-118 81
Fig. 1: A general map of Hellas, with indications of the main locations mentioned in the text.

tion period, possession or commerce of liveor dead specimens are strictly prohibited.For species included in Annex II of theProtocol of the Barcelona Convention, col-lection, capture, killing, commerce, trans-portation and disturbance especially duringreproduction should be prohibited or con-trolled. The Presidential Decree (PD)67/1981 prohibits killing, causing injury,capture, collection, commerce and trans-portation of all included species. Further-more, according to the PD 86/98 (regard-ing shell-fishing) as reformed by the PD227/2003, and the PD 109/2002 (regardingfishing baits), fishing, transportation andcommerce of any species of shelled mollus-ca not included in these PDs is prohibited.Thus, among the bivalves and gastropodsof this study, exploitation is prohibited forall species except Solecurtus strigilatus,Ensis siliqua, Solen marginatus, Pholasdactylus and Patella spp., which are includ-ed in PD 109/2002.
The species listed below are sortedfirst taxonomically (gastropods, bivalvesand cephalopods) and then phyllogeneti-cally. For each species, relevant informa-tion is categorized into four sections, i.e.General, with some diagnostic features ofthe species and useful information on itsautecology, Distribution where the globaland the Hellenic distribution of the speciesis given, Exploitation, where the mode ofexploitation and the uses of the species inthe past and at present are reported, andConservation Status, in which possible pop-ulation assessments of the species and rel-evant legislation for its conservation arepresented. In Exploitation, the real modeof exploitation is presented and not thataccording to existing legislation, as in mostcases legislation is violated and species forwhich fishing is prohibited may be exten-sively exploited. Con-generic species are
treated together.The selection of the forty-one species
listed in this paper was based on data avail-ability and the expert knowledge of theauthors. Several other species, not includ-ed in this list might also be considered asmolluscs of minor commercial interest, asfor example, Glycymeris glycymeris, Gly-cymeris bimaculata, Acanthocardia tubercu-lata, Glossus humanus, Phalium granula-tum, Cassidaria echinophora, which arecollected in great numbers by shell collec-tors. In this review, we included thosespecies that we subjectively considered asmost important.
Results
1. Gastropods
Family Patellidae: Patella caerulea (Lin-naeus, 1758), P. rustica (= lusitanica) (Lin-naeus, 1758), P. ulyssiponensis (= aspera)(Gmelin, 1791)
General: Three species have beenreported in Hellenic seas: Patella caerulea,P. rustica (= lusitanica), and P. ulyssipo-nensis (= aspera) (KOUTSOUBAS, 1992;DELAMOTTE & VARDALA-THEO-DOROU, 2007). Limpets (Patella sp.) arecommon inhabitants of the hard substratecommunities in the intertidal and uppersubtidal zones. They have a strong conicalshell with radiating ridges, which may haveepiphytes or epizooites growing on it. Lim-pets attach themselves to the substratumusing their strong muscular foot, whichenables them to withstand heavy waveaction. They are herbivorous and feedmostly at night grazing on algae found onrocks. They use a very hard-toothed radu-la to scrape rock surfaces, often limitingalgal growth on exposed and moderately
Medit. Mar. Sci., 9/1, 2008, 77-11882

exposed shores. They have a homingbehaviour and return to the same spotafter eating, using sensitive chemorecep-tors to find their way back; sometimes theyuse the same ‘home’ for their whole life.P. caerulea may reach a length of 70 mm,P. ulyssiponensis a length of 50 mm, whileP. rustica is relatively smaller, attaining alength of 45 mm.
Distribution: P. caerulea is presentthroughout the Mediterranean Sea and ina few places in the eastern Atlantic (Bay ofBiscay, Canary Islands, Moroccan coast).P. ulyssiponensis has the widest distribu-tion, being abundant on Atlantic shoresfrom Norway to Mauritania, in theMediterranean and Black Sea, and in theMacaronesian Islands. P. rustica occurs inthe Mediterranean Sea and on the Atlanticcoasts from Biarritz (France) to Maurita-nia, in Cape Verde, the Canary Islands,Selvagens, Desertas, and Madeira. P.caerulea is the most common limpet inHellenic seas. P. rustica, and P. ulyssipo-nensis are also common in most Helleniccoastal areas.
Exploitation: Limpets are widely col-lected for human consumption and as fish-ing bait. They are also used as bioindica-tors in heavy metal bioaccumulation stud-ies (e.g., CONTI & CECCHETTI, 2003).Their fishing in Hellenic seas is regulatedby the PD 109/2002.
Conservation Status: Although nopopulation assessment has been conduct-ed, all three species seem to be rathercommon.
Family Trochidae: Osilinus turbinatus(= Monodonta turbinata) (Von Born,1778), Osilinus articulatus (= Monodontaarticulata) (Lamarck, 1822), and Phorcusmutabilis (= Monodonta mutabilis) (Philip-pi, 1846)
General: Monodonts are commoninhabitants of the hard substrate commu-nities in the upper subtidal zone. Theheight of Osilinus turbinatus, O. articulatus,and Phorcus mutabilis may reach 30 mm,30 mm and 18 mm respectively. They areherbivorous and feed mostly at night graz-ing on algae found on rocks.
Distribution: All three species areabundant in the Mediterranean basin,while O. atriculatus and O. turbinatus arealso found in the eastern Atlantic. In Hel-lenic seas, O. turbinatus seems to be com-mon everywhere, while O. articulatus hasbeen reported in the Saronikos, Evvoikosand Pagasitikos Gulfs, along the Pelopon-nesian coasts, the Sporades, the N.Aegean and Ionian Sea, and P. mutabilisin the Saronikos, Korinthiakos andEvvoikos Gulfs, the N. Aegean, along thePeloponnesian coasts, Rodos and the Ion-ian Sea.
Exploitation: Small quantities of Mon-odonts (especially O. turbinatus) are col-lected for human consumption and spo-radically offered in small taverns in fishingvillages especially in the Dodekanisosislands (S Aegean Sea) and the ChalkidikiPeninsula (N Aegean Sea). They are alsoused as bioindicators in heavy metal bioac-cumulation studies (e.g., CONTI &CECCHETTI, 2003).
Conservation Status: Presumably, thethree species are abundant in Hellenicseas. There is no relevant legislation. Nopopulation assessment has been conduct-ed in Hellenic seas.
Bolma (= Astraea) rugosa (Linnaeus, 1767)
General: Bolma rugosa is an inhabitantof photophilic algae or coralligenousassemblages and Posidonia oceanicameadows (KOUTSOUBAS, 1992;
Medit. Mar. Sci., 9/1, 2008, 77-118 83

ANTONIADOU et al., 2005; personalobservations). Its shell may reach a heightof 60 mm and it has a thick, glossy, creamto orange, calcified operculum, which inHellas is called ‘Mati tis Panagias’ (i.e.,‘the Virgin Mary’s eye’).
Distribution: Bolma rugosa is found inthe Mediterranean and E Atlantic Ocean(around the Mediterranean basin),including Madeira, the Azores and theCanary Islands. B. rugosa is common andmay be found almost everywhere in Hel-lenic seas.
Exploitation: In Hellenic seas, B.rugosa is exploited (especially on theislands of the Kyklades plateau) for itsbeautifully coloured calcified operculum,which is utilized in jewellery making.
Conservation Status: It is not protect-ed by legislation. No population assess-ment has been conducted in Hellenicseas.
Cerithium vulgatum (Bruguière, 1792)
General: Approximately 300 speciesbelong to the family Cerithiidae, whichare small and medium sized shells foundin temperate and tropical seas. C. vulga-tum adults may reach a height of 5–7 cm.C. vulgatum may be found in a variety ofbiotopes, mainly on muddy substrate, onrocky and pebble bottoms, and in seagrassbeds. It prefers areas of low wave intensi-ty, such as lagoons and embayments. It isa herbivore, feeding mainly on smallalgae.
Distribution: C. vulgatum lives in theMediterranean Sea and E Atlantic, fromthe British Isles to the coast of Senegal. InHellenic seas, C. vulgatum is abundantalong most coastal areas.
Exploitation: Cerithium vulgatum, hasbeen found in a range of habitats, domi-
nating the archaeological record in theFranchthi Cave, an Upper Paleolithic findin the southern Peloponnisos, Hellas. Per-haps this small, gregarious snail was col-lected in such numbers not for food but forthe purpose of conversion into body orna-ments, as the perforations in most shellssuggest (SHACKLETON & VANANDEL, 2007). Nowadays, there is onlylimited exploitation of the species and theyare used as fishing bait.
Conservation Status: C. vulgatum isvery common. However, there is no popu-lation assessment in Hellenic seas.
Strombus (= Conomurex) persicus (= de-corus raybaudii) (Swainson, 1821)
General: Strombus persicus mayexceed a height of 75 mm, is gonochoristicwith sexual dimorphism in its radula(MUTLU, 2004). Larvae are plank-totrophic, ensuring long-distance disper-sal. All Strombus species are herbivorous,feeding on algae. In the Indo-Pacificregions, S. persicus lives on sandy bottomsand in coral sand (ABBOTT, 1960). In theMediterranean, adult S. persicus inhabitsandy or slightly gravelly bottoms and feedon seaweeds and detritus, while juveniles(shell length <2 cm) were found either liv-ing in colonies on small rocks covered byalgae or on sandy bottoms with detritus(MUTLU, 2004).
Distribution: S. persicus is a subtropicalspecies, initially restricted to the southArabian and part of the Iranian coast, thathas entered the Mediterranean presum-ably via shipping through the Suez Canaland has become locally invasive in thesouth-eastern Mediterranean (OLI-VERIO, 1995; STREFTARIS &ZENETOS, 2006). S. persicus was firstrecorded in the Mediterranean Sea in 1978
Medit. Mar. Sci., 9/1, 2008, 77-11884

in Iskenderum (southern Turkish coasts)(NICOLAY, 1986) and was successivelyfound in Israel, Hellas, Cyprus, Syria andLebanon (ZENETOS et al., 2004). In Hel-las, it has been established in Rodos, thesouth Peloponnisos coasts, Kriti, the Kyk-lades, the Argolikos, Evvoikos, andSaronikos gulfs (ZENETOS et al., 2004;YOUNG, 2007; ZENETOS et al., 2007;personal observations); it has not yet beenreported in the Ionian or the N AegeanSea (Fig. 2).
Exploitation: The species has been
exploited in Rodos Island over recentyears and sporadically offered for humanconsumption in fish restaurants.
Conservation Status: It is a successfulinvader in the Mediterranean Sea and itsspatial distribution keeps increasing(ZENETOS et al., 2004; YOUNG, 2007;ZENETOS et al., 2007). There is no rele-vant legislation for its conservation.Although there are detailed records of itsinvasion into the Mediterranean and Hel-lenic seas, no population assessment hasbeen conducted in Hellas.
Medit. Mar. Sci., 9/1, 2008, 77-118 85
Fig. 2: Known distribution of Strombus persicus in Hellenic seas.

Family Cypraeidae: Luria (= Cypraea)lurida (Linnaeus, 1758), Erosaria (=Cypraea) spurca (Linnaeus, 1758), Zonar-ia (= Cypraea) pyrum (Gmelin, 1791)
General: The shells are extremelysmooth and shiny and exhibit a great vari-ety of colouration, size, and sculpture. TheFrench call them ‘porcelains’ because oftheir texture. The mantle of the animalusually covers most of the shell surfacekeeping it in excellent condition through-out its life and without epibionts. Cowrieshave no operculum and a long, very nar-row, aperture which is lined with ‘teeth’.Luria lurida may exceed 65 mm in height.It lives in rocky areas of the subtidal zoneand is a nocturnal animal. During daylightit usually remains hidden in crevices or insponge colonies, while at night it is activesearching for its prey. L. lurida selectivelyfeeds on sponges having a strong prefer-ence for Aplysina aerophoba. It has alsobeen observed to prey on the spongesChondrosia reniformis, Chondrilla nucula,and Suberites domuncula, while the ascidi-an Ciona intestinalis is also preyed on occa-sionally, when of small size (DONEDU &MANUNZA, 1995). In the N Aegean, L.lurida specimens have been found incolonies of A. aerophoba and Petrosia fici-formis (KOUTSOUBAS, 1992). Althoughits longevity is not accurately known, L.lurida specimens have been kept alive inaquariums for more than 11 years.Erosaria spurca may reach 39 mm inheight. It is a nocturnal animal, remaininghidden during daylight hours, and lives inrocky subtidal areas and in seagrass beds.In Messina (Sicily) it is normally found onsubstrates such as shaded walls covered bychlorophyceans (Cladophora prolifera),pheophyceans (Padina pavonica, Colpo-menia sinuosa), encrusting demosponges
(Spirastrella cunctatrix, Cacospongia sp.),and colonial hydrozoans (Eudendrium sp.)(LIPPARINI & NEGRA, 2002). Zonariapyrum may exceed 50 mm in height. Speci-mens from the N Aegean were found onrocky substrate with many Axinella spp.sponges, at a depth of 25 m (KOU-TSOUBAS, 1992).
Distribution: The family Cypraeidae(cowries) includes approximately 190species, mostly distributed in tropical andsubtropical waters. Luria lurida is the mostcommon cowry of the Hellenic seas. Itsdistribution extends to the MediterraneanSea and E and W Atlantic Ocean, whileafter the opening of the Suez Canal it hasalso invaded the Red Sea and is consid-ered as an anti-lessepsian migrant(KOUTSOUBAS, 1992; DELAMOTTE& VARDALA-THEODOROU, 2007).L. lurida has been reported in theSaronikos, Evvoikos, Pagasitikos,Lakonikos, and Korinthiakos gulfs, theKyklades, the Dodekanissa, Kriti, the NAegean, and the Ionian Sea. Erosaria spur-ca is found in the Mediterranean, the RedSea and the Atlantic Ocean. It is less com-mon in Hellenic seas than L. lurida and ithas been reported in the Saronikos,Evvoikos, Messiniakos and Korinthiakosgulfs, the Kyklades, the Dodekanisos,Kriti, Rodos, Samos, and the N Aegean.Zonaria pyrum appears to be the leastabundant of the three species. It is foundin the Mediterranean Sea and E Atlantic.In Hellenic seas it has been reported in theEvvoikos Gulf, along the Peloponnisoscoasts, the Kyklades, the Dodekanisos,Samos, the N Aegean, Kriti, and the Ion-ian Sea.
Exploitation: All three species areamong the most beautiful shells in Hel-lenic seas and are very popular amongshell collectors. They are not edible and
Medit. Mar. Sci., 9/1, 2008, 77-11886

are exploited to be used in jewellery or tobe sold to shell collectors.
Conservation Status: All three speciesare under strict protection according to theBern Convention (Annex II), the Protocolof the Barcelona Convention (Annex II),and the Presidential Decree 67/1981.However, they are still illegally collectedand sold, and the relevant legislation has inpractice never been implemented. No pop-ulation assessment has been conducted inHellenic seas for any of the three species.
Tonna (= Dolium) galea (Linnaeus, 1758)
General: T. galea is one of the largestMediterranean gastropods with a heightthat may exceed 29 cm. It is gonochoristicand has no operculum. It lives insandy/muddy bottoms and seagrass beds,at depths from a few meters to 120 m. It isan active carnivore, mostly predating onholothurians but also on sea urchins andstarfish. T. galea is equipped with a partial-ly invaginable and highly extensible pro-boscis, which is capable of engulfing preythat rarely manages to escape. T. galea hasenlarged salivary glands whose secretioncontains 2.20–4.88 % sulfuric acid in addi-tion to hydrochloric acid (FÄNGE &LIDMAN, 1976) that can mediate the dis-solving of the prey integuments. Theacquisition of enlarged salivary glandscapable of secreting acid was first discov-ered in 1853 by Troschel who found thatafter the mollusc shell was crushed it pro-truded the proboscis over a distance of 13-16 cm and released a jet of clear fluidinducing active dissolution of a marblefloor (FÄNGE & LIDMAN, 1976). T.galea is a nocturnal species and is usuallyburied in the sand during daylight hoursfor protection. The common octopus is itsmain natural predator.
Distribution: The family Tonnidaeincludes approximately 30 species living intemperate and tropical seas. T. galea is acosmopolitan species. It has a wide distri-bution in Hellenic seas and has beenrecorded in the Saronikos, Korinthiakos,and Evvoikos gulfs, the N Aegean Sea (SEThermaikos Gulf, Chalkidiki - SE Toron-aios Gulf), the Kyklades, the Ionian Seaand the Cretan Sea.
Exploitation: T. galea is edible and issporadically collected in many Hellenicareas (Evvoikos Gulf, Gulf of Kavala,Thracian Sea) by divers for its flesh andalso for its shell or as a by-catch by fishinggear (e.g. trawlers, nets). In a fisheries’ sur-vey using bottom nets in the SE Ther-maikos Gulf during summer, 1 individualper 1500 m nets per day was caught in 25 %of the cases (GALINOU MITSOUDI,unpublished data).
Conservation Status: Although, nopopulation assessment has been conduct-ed in Hellenic seas, it is believed that itspopulations have greatly declined in thelast decades. It is a protected speciesaccording to Annex II of the Bern Con-vention, the Protocol of the BarcelonaConvention (Annex II), and the Presiden-tial Decree 67/1981; however, it is stillexploited.
Family Ranellidae: Charonia lampas (Lin-naeus, 1758), Charonia variegata (= trito-nis variegata) (Lamarck, 1816)
General: Charonia lampas and C. varie-gata (tritons) are among the largest gas-tropods of the Mediterranean Sea with aheight that may exceed 35 cm. They aregonochoristic gastropods and have a cor-neous operculum. Adult tritons are activepredators and mainly feed on starfish, seaurchins, sea cucumbers, and molluscs. Tri-
Medit. Mar. Sci., 9/1, 2008, 77-118 87

ton locates its prey by chemoreception ofcurrent-born prey substances. It grips itsprey with its muscular foot and uses itsradula to saw through the prey’s integu-ments. Once it has penetrated, its paralyz-ing saliva subdues the prey and the snailfeeds on it.
Distribution: C. variegata lives in rockyshores of temperate and tropical waters inthe Atlantic Ocean and the MediterraneanSea. It occurs mainly in the warm waters ofthe western Atlantic, in the Azores, theCanary and Cape Verde Islands (easternAtlantic) and in the eastern Mediter-ranean; in the western Mediterranean ithas been observed only sporadically(TEMPLADO, 1991). In Hellenic seas, C.variegata has been reported in theSaronikos, Korinthiakos, Lakonikos andEvvoikos gulfs, the Kyklades, Kriti,Dodekanisos, the N Aegean, and the Ion-ian Sea. C. lampas is found in the EAtlantic Ocean and the MediterraneanSea. In Hellenic seas, it has been reportedin the N Aegean Sea and the coasts of thePeloponnisos, Samos, and Kriti.
Exploitation: From ancient times, peo-ple of many different cultures, mainly fish-ermen, have removed the tip of the shellof the triton or drilled a hole in the tip andthen utilized the shell as a trumpet tocommunicate over long distances. Manypeople find triton shells striking andattractive and so they are collected andsold as part of the international shelltrade. This has contributed to the over-fishing and depletion of many triton pop-ulations worldwide.
Conservation Status: Nowadays, tri-tons are protected in many areas of theworld. In the Mediterranean, both C. lam-pas and C. variegata have been declared asendangered species and are strictly pro-tected according to the Bern Convention
(Annex II) and the Protocol of theBarcelona Convention (Annex II). Nopopulation assessment has been conduct-ed in Hellenic seas.
Ranella (= Argobuccinum) olearium (Lin-naeus, 1758)
General: Ranella olearium is an outershelf–upper bathyal species of the familyRanellidae. It is a large probosciferous gas-tropod with a height that may exceed 22 cm.Ranellinae are generalist predators, mostlyfeeding on echinoderms, ascidians, andpolychaetes; however, little is known aboutthe specific feeding habits of R. olearium.
Distribution: R. olearium is a cosmo-politan species and is mostly found in theNE Atlantic (SW British Isles to Maureta-nia, including the Azores, Canary Islands,Lusitanian and Meteor group seamounts),the Mediterranean Sea, S Africa (IndianOcean) and New Zealand, while it has alsobeen recorded occasionally in the W andSE Atlantic, the Caribbean Sea, the S Indi-an Ocean, and the SW Pacific(SCARABINO, 2003). In Hellenic seas, ithas been reported around the SporadesIslands, in the Evvoikos, on the Kymicoasts, and Chalkidiki.
Exploitation: It is a by-catch species oftrawl fishery. It is popular among shell col-lectors.
Conservation Status: The species isstrictly protected according to the BernConvention (Annex II) and the Protocol ofthe Barcelona Convention (Annex II).However, its populations are probablygreatly affected by trawling. In a study ofthe effects of commercial trawling on themalacological communities on the conti-nental shelf and slope of Portugal(MALAQUIAS et al., 2006), R. oleariumaccounted for 7.3 % of the total number of
Medit. Mar. Sci., 9/1, 2008, 77-11888

individuals caught in the fish trawls.Although most of the individuals caught bytrawlers are discarded, mortality mayoccur as a consequence of aggressive han-dling by fishers, thermal stress, or from set-tling on unsuitable substratum. The statusof R. olearium populations has not beenassessed in Hellenic seas or anywhere else.
Mitra zonata (Marryat, 1818)
General: Mitra zonata belongs to thefamily Mitridae and is well differentiatedfrom the other congeneric species due to itslarger size. It seems to be rather rare, andlives in sandy, coralligenous, or muddy bot-toms of deep waters. KOUTSOUBAS(1992) has reported the collection of threesamples that were on a red coral (Coralli-um rubrum) colony at a depth of 75 m. Itmay exceed a height of 85 mm. Species ofthe family Mitridae are carnivores usuallyfeeding on sipunculans and small gas-tropods; however, the specific diet of M.zonata is unknown.
Distribution: M. zonata occurs in a fewlocalities of the E Atlantic Ocean (e.g. theCanary Islands, the Azores, Madeira,Angola) and the Mediterranean Sea. InHellenic seas, it has been reported in theSaronikos and Evvoikos gulfs, Chalkidiki,Kymi (Evvoia), the Ionian Sea, and N Spo-rades Islands.
Exploitation: The startling contrast ofits beautiful colours and its rarity make M.zonata very appreciated and popularamong shell collectors. It is edible and iseaten in Croatia; it is not eaten in Hellas.
Conservation Status: M. zonata is understrict protection according to the BernConvention (Annex II) and the Protocol ofthe Barcelona Convention (Annex II). Nopopulation assessment has been conduct-ed in Hellenic seas.
Conus mediterraneus (= ventricosus)(Hwass in Bruguière, 1792)
General: Conus mediterraneus has adistinctively conical shape and can grow upto 7 cm. It has a brown, elliptical, corneousoperculum of length about 30 % of theaperture length. C. mediterraneus is apredatory, vermivorous species, beingactive mostly at night. It catches its preyusing singular, venomous, barbed, har-poon type radular teeth that are thrust intotheir prey with the use of their long andhighly maneuverable proboscis. In a studyof its feeding ecology in Tunisia, C.mediterraneus was found to eat fourteenspecies of polychaete which together witha gastropod of the Nereididae specieswere the most common food items(TAYLOR, 1987). Cone snails possess alarge, coiled venom gland, and use theircomposite venom to envenomate theirprey and to defend themselves from pred-ators and competitors. This venom con-tains a wide array of cysteine-rich, neu-roactive oligopeptides (named conopep-tides), which interfere with neuromusculartransmission (MASSILIA et al., 2001). C.mediterraneus crude venom extracts have aparalytic effect on polychaete worms andon molluscs (FAINZILBER &KLEIMAN, 1989).
Distribution: The family Conidae hasmany representatives in tropical and sub-tropical seas (approximately 400 species)but C. mediterraneus is the single Conusspecies living in the Mediterranean Sea; itis also found on the coasts of westernAfrica. It is a very common species of therocky upper subtidal zone in Hellenic seasand may be found everywhere, from the NAegean to the Libyan Sea, and from theIonian Sea to the Dodekanisos.
Exploitation: Its exploitation is very
Medit. Mar. Sci., 9/1, 2008, 77-118 89

limited, for private or public collections. Itis not edible.
Conservation Status: Presumably, thespecies is abundant. There is no relevantlegislation. No population assessment hasbeen conducted in Hellenic seas.
2. Bivalves
Lithophaga lithophaga (Linnaeus, 1758)
General: The European date musselLithophaga lithophaga is an endolithicspecies and an inhabitant of hard substrata(calcified) communities in the midlittoraland upper sublittoral zones (GALINOU-MITSOUDI & SINIS, 1994). The autoe-cology (ecology, population dynamic, biol-ogy) of L. lithophaga has been extensivelystudied in the Evvoikos Gulf (GALINOU-MITSOUDI & SINIS, 1994, 1995). Thepopulation density of this species, whichseems to be dependent on the substratumtype (limestones), the presence of otherorganisms (e.g. endolithic), predation andenvironmental conditions, is higher in theN. Evvoikos Gulf (36 inds/dm3). The age offirst reproduction is 2+ years and thereproductive activity is annual and contin-uous. The main reproductive period is dur-ing the summer and autumn at water tem-peratures > 14ÆC. L. lithophaga is a long-lived species (> 54 years) and its growthrate is one of the slowest among bivalves.The highest growth rate is observed at theend of spring and early summer. In theEvvoikos Gulf, the annual productivity ofL. lithophaga was 1659.6 g/m2 and theannual turnover ratio (P/B) was 0.845,which is among the lowest values reportedin the literature.
Distribution: L. lithophaga is endemicin the Mediterranean Sea. In Hellenicseas, the species is found in the Evvoikos
Gulf, the Ionian islands, the Ipeiros coast,the Korinthiakos Gulf, the Sea of Kythira,the N Sporades Islands, the Dodekanisos,Kriti, Lesvos island., Maliakos, Argolikosand Chalkidiki (Fig. 3) (ZENETOS 1986;1996; GALINOU-MITSOUDI & SINIS,1994, 1997; GEROVASILEIOU et al.,2007; personal observations).
Exploitation: In the past, L. lithophagawas extensively exploited in Hellenic seasand it was found in seafood markets andfish restaurants in many locations. Thespecies is still exploited by divers (illegally)as a delicious food resource, especially inEvvoia.
Conservation Status: The species isunder strict protection according to the92/43/EC Directive (Annex IV), the BernConvention (Annex II), and the Protocolof the Barcelona Convention (Annex II).The population of Evvoikos Gulf has beenassessed in the past (GALINOU-MITSOUDI & SINIS, 1994).
Pinna nobilis (Linnaeus, 1758)
General: The fan mussel Pinna nobilisoccurs at depths of between 0.5 and 60 m,typically in soft-bottom areas overgrownby seagrass (mostly Posidonia oceanicaand Cymodocea nodosa meadows) butalso in unvegetated sandy bottoms(KATSANEVAKIS, 2006; 2007a). It isthe largest Mediterranean bivalve and oneof the largest in the world, attaininglengths of up to 120 cm. P. nobilis is long-lived and may live over 20 years(GALINOU-MITSOUDI et al., 2006).Fan mussels live partially buried (usually>35 % of their length) by the anteriorportion of the shell and attached by theirbyssus threads to the substratum. Repro-duction of P. nobilis takes place by meansof external fertilization and its success
Medit. Mar. Sci., 9/1, 2008, 77-11890

depends on the proximity of other individ-uals spawning synchronously. When apopulation becomes sparse (as is the casefor most fan mussel populations in theMediterranean) failure of fertilizationbecomes a critical issue for the survival ofthe species. The planktonic period isshort, hence populations (especially inclosed embayments), which are relativelyisolated are not easily recolonised ifdepleted. Natural mortality is strikinglysize dependent and P. nobilis suffers highnatural mortality during the first year oflife; the probability of death by natural
causes quickly diminishes as the fan mus-sels grow in size (KATSANEVAKIS,2007b). Growth rates have a seasonal pat-tern, with an extended period of very slowgrowth between late autumn and earlyspring and a peak in growth rates duringlate spring - early summer, probably relat-ed to an optimum combination of temper-ature and food availability (KATSA-NEVAKIS, 2007b).
Distribution: P. nobilis is endemic tothe Mediterranean Sea. The global popu-lation of P. nobilis has been greatlyreduced over the past few decades as a
Medit. Mar. Sci., 9/1, 2008, 77-118 91
Fig. 3: Population density hotspots of Lithophaga lithophaga.

result of recreational and commercial fish-ing for food, the use of its shell for decora-tive purposes, and incidental mortality bytrawlers, bottom nets, or anchoring.Although P. nobilis has become rare inmany parts of the Mediterranean, impor-tant local populations still exist in Hellenicseas (Fig. 4), especially in the Korinthi-akos, Evvoikos, and Thermaikos gulfs, theislands of Chios and Lesvos (NE Aegean),NW Kriti, N Karpathos, as well as the Ion-ian Sea (ZENETOS, 1986; 1996; HAMESet al., 2001; GALINOU-MITSOUDI et al.,2006; KATSANEVAKIS, 2006; 2007a;
GEROVASILEIOU et al., 2007; personalobservations).
Exploitation: From ancient times untilearly in the 20th century, P. nobilis wasexploited for its byssus, from which anextremely fine and valuable fabric was pro-duced, called ‘sea silk’. P. nobilis has alsobeen extensively exploited for human con-sumption and use of its shell for decorativepurposes. ‘Semi-quality’ pearls may befound in some aged fan mussels. Nowadaysit has been declared an endangered speciesand its exploitation has been banned. How-ever, it is still illegally exploited and in cer-
Medit. Mar. Sci., 9/1, 2008, 77-11892
Fig. 4: Population density hotspots of Pinna nobilis.

tain areas fishing mortality greatly exceedsnatural mortality and is a critical determi-nant of the spatial distribution of thespecies. In Lake Vouliagmeni, mortalitydue to illegal fishing by free divers hascaused a very low population density of thespecies in shallow areas (KATSA-NEVAKIS, 2007b). P. nobilis has been usedas a bioindicator in heavy metal bioaccumu-lation studies. In Hellenic seas, it was select-ed as an indicator species to study chromi-um contamination in a gulf receiving tan-nery wastes (CATSIKI et al., 1994).
Conservation Status: The species isunder strict protection according to the92/43/EC Directive (Annex IV), the Proto-col of the Barcelona Convention (AnnexII), and the Presidential Decree 67/1981. Afew populations of P. nobilis have beenassessed in Hellenic seas, and some synop-tic results of reported population density orabundance are given in Table 2. In two ofthese areas (Vouliagmeni Lake and SoudaBay) total abundance of P. nobilis has beenestimated using Density Surface Modellingwith Distance Sampling (KATSA-NEVAKIS, 2007a), while in another threeareas population densities have been esti-mated (Table 2). Specific mtDNA regionswere genetically characterized in pinnidsfrom the Thessaloniki and Thermaikosgulfs, Vouliagmeni Lake and NE Chios.These results could contribute to screeninggenetic variability and population dynam-ics, as well as for conservation plans forspecies protection and/or aquacultureinvestigations (KATSARES et al., 2008).
Pinctada radiata (Leach, 1814)
General: Pinctada radiata is an epifau-nal suspension feeder of the subtidal zoneand a fouling species, living attached by itsbyssus to hard substrata. Usually it attains a
Medit. Mar. Sci., 9/1, 2008, 77-118 93
Tab
le 2
Est
imat
ed p
opul
atio
n de
nsit
y or
abu
ndan
ce o
f Pin
na n
obili
s in
Hel
leni
c si
tes.
Loca
tion
Stud
y Ar
eaYe
arD
ensi
ty(in
ds/m
2 )Ab
unda
nce
(inds
)R
efer
ence
(hec
tare
s)m
ean
max
estim
ate
95%
CI
Lake
Vou
liagm
eni (
Kor
inth
iako
s)15
0.4
2004
0.00
570.
1785
0042
00 –
1290
0K
ATS
AN
EVA
KIS
, 200
6La
ke V
oulia
gmen
i (K
orin
thia
kos)
150.
420
060.
0045
0.24
6770
5450
–84
00K
ATS
AN
EVA
KIS
, 200
7aSo
uda
Bay
(Krit
i)14
71.5
2007
0.00
890.
5313
0900
1006
00 –
1704
00K
ATS
AN
EVA
KIS
, unp
ublis
hed
data
NE
Chio
s Isl.
(Aeg
ean)
0.08
820
060.
060.
07G
ALI
NO
U-M
ITSO
UD
I, un
publ
ished
dat
aN
E K
efal
loni
a Is
l.(Io
nian
)20
000.
08H
AM
ES et
al .
, 200
1Ep
anom
i (Th
erm
aiko
s Gul
f)0.
007
2004
1.04
1.3
GA
LIN
OU
-MIT
SOU
DI e
t al .
, 200
6

length of 50-65 mm, but may exceed 100mm in length. It is a protandric hermaphro-dite species with sex inversion occurring inshells of 32-57 mm length. Gonad maturityis controlled by temperature (TLIGZOUARI & ZAOUALI, 1994).
Distribution: Pinctada radiata origi-nates from the Indo-Pacific and the RedSea. Following the opening of the SuezCanal, P. radiata has entered the far east-ern Mediterranean. It was also intention-ally imported for mariculture in manyareas of Hellas and Italy during the lastcentury. Up to date, it has been recordedas common in the eastern Mediterraneanwith sporadic occurrences in the westernbasin (ZENETOS et al., 2004). Besides itsadaptation to the subtropical environ-ment of the south-eastern Mediter-ranean, its tolerance to chemical contam-ination has enhanced its expansion inenclosed polluted ecosystems. In Hellenicseas (Fig. 5), it is mostly reported close toareas where it was originally introducedfor aquaculture (mainly in theDodekanisos islands, Argolikos, theSaronikos, Evvoia, and the island ofLesvos in the N Aegean Sea). Recently,the species was also recorded in the Kyk-lades Isles complex (in 2006) and in Kriti(in 2003) (ZENETOS et al., 2007).
Exploitation: Pearl shells (mother-of-pearl) have been used by the people ofthe Pacific and other regions as utensils,implements and ornamentation, while theoyster itself has been a basic food item.Pearl oyster meat is a delicacy in manywestern cultures. However, mother-of-pearl shells are of little commercial valuein the Mediterranean. Because of its toler-ance to chemical contamination and theability of oysters to accumulate metals toseveral orders of magnitude higher thanthe background medium, it has been used
as a bioindicator of heavy metal pollution(e.g., AL-MADFA, et al., 1998; GÖKSU etal., 2005).
Conservation Status: It is a successfulinvader into the Mediterranean Sea and itsspatial distribution keeps increasing(ZENETOS et al., 2004; ZENETOS et al.,2007). There is no relevant legislation forits conservation and there is no need forprotective measures. Although there aredetailed records of its invasion in theMediterranean and Hellenic seas, no pop-ulation assessment has been conducted inHellas.
Pteria hirundo (Linnaeus, 1758)
General: Pteria hirundo may reach 10cm in length. It has an aggregated distribu-tion and usually many individuals arefound attached by their robust byssus togorgonians, on rocky bottoms, or inclumps of many individuals on muddy bot-toms in the sublittoral and bathyal zones.
Distribution: P. hirundo is found in theMediterranean Sea and the E Atlanticfrom the British Isles south to the coastsof Portugal. In Hellenic seas, it has beenreported in the Saronikos, Patraikos, Pag-asitikos, and N Evvoikos gulfs, Astakos,the Kyklades, the N Sporades Islands, andChalkidiki (Toronaios and S Sithonia).
Exploitation: Its exploitation is verylimited, for private or public collections.
Conservation Status: No populationassessment has been conducted in Hel-lenic seas.
Family Pectinidae: Chlamys varia (Lin-naeus, 1758), Crassadoma (Chlamys) mul-tistriata (Poli, 1795)
General: Chlamys varia is a scallopwith a thin, narrow, elongated oval shell
Medit. Mar. Sci., 9/1, 2008, 77-11894

that may reach a length of 80 mm. Itscolour is very variable and may be white,yellow, orange, pink, red, purple, orbrown. Crassadoma multistriata looks likeC. varia but is shorter and with distinctivedecoration. It usually attains a length of15–30 mm. Both species live on sandy,muddy, or rocky bottoms of the sublittoralzone. Young individuals are either free-liv-ing or attached by byssus threads, whileadults are usually permanently cementedto hard surfaces by their right valve. Bothspecies are commonly found in Posidoniaoceanica beds. The spat settlement occurs
during late spring and summer (PEN~A etal., 1996).
Distribution: C. varia is found in theMediterranean Sea and the E AtlanticOcean from the British Isles south to Sene-gal, while C. multistriata is found in theMediterranean Sea and the E AtlanticOcean from the Faroe Islands to the coastsnear the Mediterranean. In Hellenic seas,C. varia is quite common and seems to bepresent everywhere on the Hellenic coasts.C. multistriata has been reported in the Sa-ronikos, Patraikos, Korinthiakos, Evvoikos,and Messiniakos gulfs, the Sporades, the
Medit. Mar. Sci., 9/1, 2008, 77-118 95
Fig. 5: Known distribution of Pinctada radiata in Hellenic seas.

Kyklades, the Dodekanisos, Kymi (Evvoia),Kavo Doro, and Chalkidiki.
Exploitation: Both species are edibleand in a few European locations (e.g., onthe French Atlantic coasts) C. varia is tar-geted by a commercial fishery. In Hellenicseas C. varia is locally exploited for humanconsumption. C. varia has been reportedto highly concentrate trace metals andradionuclides and has been used as a bio-monitor of marine pollution (BUSTA-MANTE et al., 2002; BUSTAMANTE &MIRAMAND, 2005). Because of theirbeautiful and variable colouration both C.varia and C. multistriata are popularamong shell collectors.
Conservation Status: No populationassessment has been conducted in Hel-lenic seas.
Spondylus gaederopus (Linnaeus, 1758)
General: Spondylous gaederopus mayexceed a height of 15 cm. It lives on rockybottoms of the sublittoral zone from 1 m toabout 50 m depth, permanently cementedto hard surfaces by its right valve. It prefersexposed positions on vertical rocks withslow but steadily moving currents (POPPE& GOTO, 1993). The species is oftenaccompanied by the sponge Crambecrambe that covers the upper valve andcamouflages its shell.
Distribution: S. gaederopus is found inthe Mediterranean and the Atlantic Oceannear Gibraltar, including Madeira, theAzores, the Canaries, and the Cape VerdeIslands. In Hellenic seas, it has beenrecorded in the Saronikos, Lakonikos,Messiniakos, Thermaikos, Evvoikos,Patraikos, and Korinthiakos gulfs, the Pelo-ponnisos, the Kyklades, the Dodekanisos,Kriti, Astakos, Kymi (Evvoia), Kefallonia,and the Chalkidiki peninsula.
Exploitation: Archaeological evidenceshows that people in Neolithic Europetraded the shells of S. gaederopus to makebangles, bracelets, belt-buckles and otherornaments as long as 5000 years ago. Theshells were mostly harvested from theAegean Sea and then transported intocontinental Europe. S. gaederopus orna-ments from the Neolithic age have beenfound in many caves and archeologicalsites in Hellas (IFANTIDIS, 2006). It isedible and has been collected for humanconsumption since antiquity. In someareas it is considered a delicacy and for thisreason was named the ‘royal oyster’. It isalso popular among shell collectors.
Conservation Status: No populationassessment has been conducted in Hel-lenic seas. POPPE & GOTTO (1993)pointed out that the species was rathercommon till the beginning of the 1980s,when (for unknown reasons) its popula-tions markedly declined or even disap-peared in some areas.
Crassostrea gigas (= angulata) (Thun-berg, 1793)
General: Crassostrea gigas is a largeoyster whose size may exceed 30 cm (andmay reach 40 cm in exceptional cases). Theshell surface is extremely rough, coarselysculptured with concentric ridges and 6–7thick and bold ribs that commonly give theshell edges an undulating appearance. Theshape of the shell varies depending on theenvironmental conditions. It prefers firmsubstrates of the sublittoral zone usually atdepths between 5 and 40 m and occasion-ally to depths of about 80 m. It may also befound on mud or mud-sand bottomsattached to debris, rocks, or other bivalveshells. It is an estuarine species and opti-mal salinity range is between 20 and 25ò.
Medit. Mar. Sci., 9/1, 2008, 77-11896

It is a protandrous hermaphrodite and thetiming of sex change is erratic and season-al. Spawning usually depends on a rise inwater temperatures above 20ÆC, givingapproximately 50-100 million eggs in a sin-gle spawning; larvae are planktonic (FAO,2008). The biology, ecology, and especiallythe aquaculture of the species have beenextensively studied (~1000 related pub-lished papers in the Science Citation Indexbetween 1970 and 2007).
Distribution: C. gigas originates in thecoastal and marine waters of Japan, NorthWest Pacific ocean. It has been introducedfor mariculture into North America, and isnow found from SE Alaska to Baja Cali-fornia. In Europe, it was first introduced inthe Atlantic (Portugal, Spain, France, Ire-land, U.K.), and may be found fromMorocco to Norway. It is also widespreadin the Mediterranean (France, Adriaticlagoons, Malta, Spain, Tunisia, Italy, Hel-las, Turkey). In Hellenic seas, it has beenreported in the Patraikos and Korinthi-akos gulfs (Zenetos et al, 2004). There isalso unconfirmed information that thespecies was imported for aquaculture intothe Thermaikos Gulf in 2007.
Exploitation: It is both fished and culti-vated for human consumption in manycountries. Through its potential for rapidgrowth and its wide ranging tolerance toenvironmental conditions, it has becomethe oyster of choice for cultivation in manyregions of the world. It is the dominantshellfish in a growing United States aqua-culture industry along the Pacific Coast.By 2003, global production of this specieshad expanded to 4.38 million tonnes, morethan any other species of fish, mollusc orcrustacean. Nearly 84 % of global C. gigasproduction was in China (FAO, 2008). C.gigas has also been used as a bioindicatorof heavy metal pollution (e.g., SHULKIN
et al., 2003). It is not a target species inHellenic seas, as it has low population den-sities and narrow distribution.
Conservation Status: It is classified asone of the Worst Invasive Species inEurope (EEA, 2007). Its dense aggrega-tions limit food and space for other benthicspecies (competition), it destroys habitatsand causes eutrophication of water. It alsocauses causes serious decline in macrofau-na and zooplankton, but enhances bacte-ria, microfauna and meiofauna. No popu-lation assessment has been conducted inHellenic seas.
Donacilla (= Mactra) cornea (Poli, 1791)
General: Donacilla cornea is an inhabi-tant of the midlittoral soft substratum andis found mostly in moderately exposedbeaches with medium to very coarse sand(MAVIDIS et al., 2006). It is a relativelysmall bivalve and its maximum length mayreach 25 mm. The individuals of D. corneaare distinctively patterned and coloured,exhibiting one of the most striking exam-ples of massive polymorphism in nature(OWEN & WHITELEY, 1988). D. corneais gonochoristic, its life expectancy is 3–4years, has an annual reproductive cycle, ismature at a length of approximately 10mm, and spawning (in the ThermaikosGulf) occurs between March and July witha peak in June (MAVIDIS & KOUKOU-RAS, 2001a). In a study of the midlittoralsoft substratum assemblages of the NorthAegean Sea (KOUKOURAS & RUSSO,1991), among the five different assem-blages that were described, one assem-blage was characterized by the very highdominance of D. cornea and the poly-chaete Ophelia bicornis. This assemblagewas dominant in the greatest part of theThermaikos and Strymonikos gulfs and
Medit. Mar. Sci., 9/1, 2008, 77-118 97

was located on coarse sandy substrates.Mean densities of D. cornea (± SE) were1993 (± 350) inds/m2 and 1255 (± 205)inds/m2 in the D. cornea–O. bicornis assem-blages of the Thermaikos and Strymonikosgulfs, respectively. On Korinos beach(Thermaikos Gulf) the abundance of D.cornea had two peaks, one at the mean sealevel (with a density that exceeded 12000inds/m2) and a second below the low tidelevel (MAVIDIS & KOUKOURAS,2001b).
Distribution: It is found in the Mediter-ranean, the Black Sea, and the E Atlanticin areas close to the Mediterranean basin.In Hellenic seas, D. cornea is very abun-dant in the N Aegean and is also found inthe Evvoikos, Saronikos, Lakonikos,Amvrakikos, and Patraikos gulfs, the Kyk-lades, the Ionian and the Cretan Seas.
Exploitation: The species is collectedby hand in very small quantities and usedoccasionally as fishing bait and ratherrarely for human consumption. There isample evidence that Donacilla corneaalong with Tapes decussatus and, Donaxtrunculus were among the marinemolluscan resources available toFranchthi's inhabitants around 13,000 BC(SHACKLETON & VAN ANDEL, 2007).D. cornea has been used as a bioindicatorof heavy metal pollution (e.g., REGOLI etal., 1991).
Conservation Status: It is very abun-dant in many midlittoral soft substratumareas. The populations of the Thermaikosand Strymonikos gulfs have been assessedin the past (KOUKOURAS & RUSSO,1991; MAVIDIS & KOUKOURAS,2001b). There is no relevant legislation.The regional IUCN status of CriticallyEndangered species was proposed for D.cornea in the Black Sea (MICU & MICU,2006).
Solen marginatus (Pulteney, 1799)
General: Solen marginatus is a large,shallow-water, elongate clam, which mayreach a length of 13 cm. It is an inhabitantof the midlittoral and upper sublittoralzone, living burried in the sand. It is a filterfeeder and lives in a vertical position in thesediment in more or less permanent tubesin which they can ascend or descend. Withits specially adopted piston-like foot,which can be inflated with blood, it is ableto burrow rapidly in its tube or even dig anew tube.
Distribution: S. marginatus is found inthe eastern Atlantic from Mauretania tothe North Sea, including the Mediter-ranean and the Black Sea. In Hellenicseas, S. marginatus has been reported fromthe coasts of the Peloponnisos (both in theIonian Sea and the Aegean Sea) and fromthe Amvrakikos, Thermaikos, Pagasitikosand Saronikos gulfs.
Exploitation: In Hellenic seas, it isexploited locally both for human consump-tion and as fishing bait. Its fishing is regu-lated by the PD 109/2002. S. marginatushas been used as a bioindicator of marinepollution by organochlorine compoundssuch as PCBs and DDTs (THOMPSON etal., 1999).
Conservation Status: No populationassessment has been conducted in Hel-lenic seas.
Ensis minor (Chenu, 1843)
General: Ensis minor is a large burrow-ing, shallow-water, elongate clam, whichmay reach a length of 20 cm. It is an inhab-itant of the midlittoral and upper sublit-toral zone, living buried in the sand mostlyat depths shallower than 4 m, but in theMediterranean may be found down to
Medit. Mar. Sci., 9/1, 2008, 77-11898

about 25 m (VAN URK, 1964). It is a fil-ter feeder and lives in a vertical position inthe sediment in more or less permanenttubes. In the Gulf of Trieste, gametogene-sis begins in September and the maximumreproductive activity occurs between Jan-uary and March (VALLI et al., 1983).
Distribution: E. minor is distributed inthe eastern Atlantic, from Norway to Por-tugal, including the Mediterranean Sea. InHellenic seas, E. minor has been reportedfrom Kriti and the Saronikos, Pagasitikosand Thermaikos gulfs.
Exploitation: In Hellenic seas, it isexploited locally both for human consump-tion and as fishing bait. In Italy it is exten-sively fished and is one of the most impor-tant fishery resources of the Gulf of Tri-este (DEL PIERO & DACAPRILE,1998). Its fishing in Hellenic seas is regu-lated by the PD 109/2002.
Conservation Status: No populationassessment has been conducted in Hel-lenic seas.
Solecurtus strigilatus (Linnaeus, 1758)
General: Solecurtus strigilatus mayexceed 10 cm in length. It lives buried inthe soft substrata of the sublittoral and,rarely, of the bathyal zones. It burrows rap-idly and to relatively great depths. Adultsmay live 50 cm below the sea floor andattempts to excavate them from their bur-rows release rapid escape reactions where-by the animals descend obliquely still fur-ther into the sediment (BROMLEY &ASGAARD, 1990). The location of S.strigilatus individuals on the seafloor ismarked by two conspicuous round orificesabout 1 cm in diameter and 2 cm apart,with siphons never observed to projectfrom these openings. When in danger ofpredation S. strigilatus will abort parts or
all of its siphons, which contract rhythmi-cally as they float away in the water col-umn; this is probably part of the escapestrategy of the animal, as the aborted partswill attract the predator and the bivalvewill have the chance to escape, while itssiphons will soon regenerate (BROMLEY& ASGAARD, 1990).
Distribution: S. strigilatus is found inthe Mediterranean Sea and on neighbour-ing E Atlantic coasts. In Hellenic seas, ithas been reported in the Saronikos, Ther-maikos, Evvoikos, Patraikos, Korinthi-akos, and Kavala gulfs, and the coasts ofthe Peloponnisos, Kefallonia, and Kriti.
Exploitation: Its exploitation is verylimited, for human consumption and forprivate or public collections. It is also usedas fishing bait, especially in the NorthEvvoikos Gulf. Its fishing in Hellenic seasis regulated by the PD 109/2002.
Conservation Status: No populationassessment has been conducted in Hel-lenic seas.
Pholas dactylus (Linnaeus, 1758)
General: Pholas dactylus grows up to15 cm long. It lives in midlittoral and shal-low sublittoral areas bored into soft rockssuch as, shale, peat, chalk, sandstone, stiffclay, or wood. It creates a conical burrow,which has a narrow entrance and a largerrounded chamber. Where abundant, itsborings can severely compromise thestructural stability of the shore, and canresult in increased rates of coastal erosion(PINN et al., 2005). P. dactylus gives off ablue, luminous secretion when irritated. Itsluminescence has been known sinceancient times (e.g., Pliny the Elder docu-mented the luminescence of P. dactyluseven whilst being eaten) and has a historicplace in the biochemistry of biolumines-
Medit. Mar. Sci., 9/1, 2008, 77-118 99

cence, since it was the first organism usedto describe the oxygen-requiring luciferin-luciferase reaction that is universal in bio-luminescence. It is a gonochoristic specieswith a spawning period during summerand a life span of 14 years (HILL, 2006).
Distribution: It is found in the Mediter-ranean, the Black Sea, the Red Sea, and inthe E Atlantic from the British Isles southto the coasts of Morocco. In Hellenic seas,it has been reported in the Thermaikos,Kalloni, Maliakos, Evvoikos, Saronikos,Amvrakikos, and Argolikos gulfs, on theIonian coasts of the Peloponnisos, andKefallonia.
Exploitation: It has been extensivelyfished for human consumption and as fish-ing bait.
Conservation Status: P. dactylus wasonce prevalent across the entire Mediter-ranean and on the Atlantic coast ofEurope, but it has disappeared from mostsites due to human collection for food andbait and/or as a result of marine pollution.The species is under strict protectionaccording to the Bern Convention (AnnexII), and the Protocol of the BarcelonaConvention (Annex II). Surprisingly, it isincluded as legally collected fishing bait inPD 109/2002 but the Bern Convention(adopted by the Hellenic Law 1335/1983)and the Barcelona Convention prevailover the PD. No population assessmenthas been conducted in Hellenic seas.
3. Cephalopods
Alloteuthis media (Linnaeus, 1758)
General: A. media is a small-sizedmember of the family Loliginidae, withfemales attaining a mantle length up to12 cm and males up to 9 cm. The durationof its life cycle is about one year
(ALIDROMITI, 2007). A. media is amongthe main cephalopod species of the conti-nental shelf community (GONZALEZ &S NCHEZ, 2002; LEFKADITOU et al.,2004) and is mostly found on sandy andmuddy grounds and also in brackish waters(UNSAL et al., 1999). It inhabits a widedepth range from the surface down to 600m, but is mostly found at depths <150 m(LEFKADITOU, 2007). It performs sea-sonal migrations between offshore (in win-ter) and inshore (in summer) areas(ROPER et al., 1984), as well as diel verti-cal migrations (ZUEV & NESIS, 1971).Maturation takes place over a wide rangeof mantle lengths. Spawning in the Me-diterranean occurs at depths of 10-100 mmostly in sandy areas or seagrass mead-ows. Spawning period lasts from March toOctober in the western part of theMediterranean and potential fecundity(PF) has been estimated to be about 950-1400 eggs (MANGOLD-WIRZ, 1963),whereas in the eastern part spawningoccurs all year round and PF is some 1000-4000 eggs, most of which are released dur-ing multiple spawning with female growthcontinuing between spawning events(LAPTIKHOVSKY et al., 2002). The dietof A. media consists of larvae and juvenilesof fishes, copepods and euphausiids(ZUEV & NESIS, 1971). A. media ispreyed on by demersal fish of medium andlarge size, mostly by gadids and some elas-mobranchs (ZUEV & NESIS, 1971;BELLO, 1997; VELASCO et al., 2001).
Distribution: A. media is common inthe Mediterranean Sea including the Seaof Marmara. In the E Atlantic its rangeextends from 60Æ to 21Æ N, but it is mainlyfound in the southern part of the NorthSea, the Irish Sea, the English Channel,the Bay of Biscay and off the south coast ofthe Iberian peninsula. It is widespread in
Medit. Mar. Sci., 9/1, 2008, 77-118100

Hellenic seas and has been reported as themost frequently caught cephalopodspecies on the shelf of the N. Aegean(LEFKADITOU, 2006), S. Aegean(LEFKADITOU et al., 2003b), and theIonian Sea (Table 3).
Exploitation: A. media is a by-catch ofbottom trawlers and beach-seiners that ismostly discarded by Hellenic trawlers(MACHIAS et al., 2001), whereas its dis-card percentage by Italian and Spanishtrawlers fishing in the western Mediter-ranean ranges from 10 to 50 % and is con-sidered accidental due to the fast sortingoperations on deck (SARTOR et al.,1998).
Conservation Status: No populationassessment has been conducted in Hel-lenic seas.
Loligo forbesi (Steenstrup, 1856)
General: Loligo forbesi is a large-sizedloliginid; males generally grow larger than
females and have higher growth rates(PIERCE et al., 1994a,b). Adult body sizeusually reaches 100–650 mm ML in males(weight range 155–3700 g) and 175–350mm ML in females (weight range200–1150 g), while maximum values of 937mm ML and 8308 g for males, and 462 mmML and 2184 g for females have beenreported from the Azores (MARTINS,1982). It has an annual life-cycle and issemelparous, exhibiting ‘intermittent ter-minal spawning’, in which the females lay1000 to 23000 eggs in batches and dieshortly after spawning (PIERCE et al.,1994b). L. forbesi migrates to shallowerwaters to spawn. A large number of preyspecies, including various polychaetes,molluscs, crustacean and fish have beenidentified in L. forbesi stomachs, with crus-taceans being relatively more important inthe diet of small squid while larger squidprey predominantly on fish (MARTINS,1982; PIERCE et al., 1994b). Amongcephalopod remains, those of L. forbesi
Medit. Mar. Sci., 9/1, 2008, 77-118 101
Ionian Sea ArgosaronikosSpecies 10-200 m 200-500 m 10-200 m 200-500 mAlloteuthis media 2948 82 453 95Loligo forbesi 2 38 2 112Todarodes sagittatus 0 13 0 14Todaropsis eblanae 26 124 0 17Sepia elegans 259 24 231 163Sepia orbignyana 46 5 777 238Sepietta oweniana 40 428 35 53Rondeletiola minor 7 296 8 34Rossia macrosoma 5 17 0 277*Scaeurgus unicirrhus 59 3 232 126
*exceptionally high value observed only in 1995
Table 3Indicative population density (in number of individuals per square km)of cephalopodspecies of minor importance,estimated during the MEDITS survey of 2005, on shelf
(10-200 m)and upper slope (200-500)of the Ionian Sea and the region of Argosaronikos.

have also been occasionally identified.Large demersal fish and some marinemammals have been reported to prey onL. forbesi (DALY et al., 2001; SANTOS etal., 2001).
Distribution: L. forbesi occurs in theMediterranean Sea and the NE Atlanticfrom 20 ÆN to 63 ÆN, where it is moreabundant in the northern part of its range.In the temperate part of its distributionalrange it is mainly found over the shelf, butin subtropical areas and in the Mediter-ranean in deeper waters, reaching depthsof 720 m in the Ionian Sea(LEFKADITOU et al., 2003a). It is wide-spread in Hellenic seas, being more com-mon in the neritic zone of regions withabrupt waters like the Ionian Sea and thearea of the Kyklades, Kriti and theDodekanisos islands, while it is scarcenorth of 40Æ ¡ in the Aegean Sea(KOUTSOUBAS et al., 1999; LEFKA-DITOU, 2006).
Exploitation: L. forbesi is largely fishedas a by-catch from bottom trawl fisheries.Fishery statistics do not distinguish L.forbesi from L. vulgaris, since they are ofsimilar appearance and equal commercialvalue. However, landings in Scottishwaters can generally be assumed to be L.forbesi (PIERCE et al., 1994a), while thosein the Mediterranean mainly consist of L.vulgaris (GUERRA et al., 1994). There isno estimation of the marketable propor-tion of L. forbesi in the landings of lolig-inids in Hellenic fisheries.
Conservation Status: Since the early1990s, L. forbesi has apparently disap-peared from much of the southern part ofits range in the eastern Atlantic (Franceand Portugal), while at the same time itsabundance in northern waters (Scotland)has increased. This shift in its distributionrange may be associated with the increase
in sea surface temperature after 1993 andsubsequent high levels (CHEN et al.,2004). No population assessment has beenconducted in Hellenic seas.
Todarodes sagittatus (Lamarck, 1798)
General: T. sagittatus is a large squid ofthe family Ommastrephidae. Its mantlelength attains 75 cm, but usually in trawlcatches it does not exceed 35 cm, as imma-ture individuals make up the vast majorityof specimens (BORGES & WALLACE,1993; QUETGLAS et al., 1998; LORDANet al., 2001; LEFKADITOU, unpublisheddata). This neritic and oceanic squidoccurs from surface to near-bottom waterlayers over the continental shelf and slopeup to 2500 m. It carries out vertical move-ments between the surface at night andnear the sea bottom during the day(MANGOLD-WIRZ, 1963; KORZUM etal., 1979). Estimates of its life span rangefrom 9 to 14 months. Its life span wasfound longer in higher latitudes and abouta month longer in females than in earliermaturing males. T. sagittatus is a fast grow-ing and early maturing squid, its matura-tion found earlier and subsequently itsgrowth rate lower in NW African waters(ARKHIPKIN et al., 1999) compared withthose of northernmost areas (LORDAN etal., 2001; QUETGLAS & MORALES-NIN, 2004). Mature individuals are pres-ent all year round but spawning appears topeak in autumn-winter (QUETGLAS etal., 1998; ARKHIPKIN et al., 1999).Potential fecundity estimates range from200000 to 950000 eggs (LAPTI-KHOVSKY & NIGMATULLIN, 1999;LORDAN et al., 2001). Its diet consists offish, crustaceans and cephalopods (indecreasing order of importance), with can-nibalism also identified (QUETGLAS et
Medit. Mar. Sci., 9/1, 2008, 77-118102

al., 1999; LORDAN et al., 2001). YoungerT. sagittatus in some areas have shownpreference for small pelagic prey in com-parison to specimens in more advancedadult stages (PIATKOWSKI et al., 1998),a fact that indicates their preying mostly inupper water layers. T. sagittatus is the mostabundant prey in the stomachs of sword-fish from the eastern Mediterranean sug-gesting a high abundance of the speciesparticularly during summer (PERISTE-RAKI et al., 2005). Apart from swordfish,other large pelagic fishes, sharks andcetaceans are important predators of thisspecies (ROPER et al., 1984).
Distribution: T. sagittatus is broadly dis-tributed throughout the north and north-east Atlantic, from the Arctic Ocean toabout 13ÆS and to approximately 40ÆW,including the North Sea and the Mediter-ranean Sea, where it is more abundant offthe Tunisian and Libyan coasts, in thestraits of Sicily and Messina, in the north-ern Tyrrhenian Sea and in the centralAdriatic (BELLO, 1985; JEREB &RAGONESE, 1994; RELINI et al.,1999).In Hellenic seas it has been reported inthe Aegean (N< 40Æ 30’) and the IonianSeas, being caught at depths between 120and 800 m, more frequently during sum-mer and autumn, but never exceeding 10individuals per hour of trawling(LEFKADITOU et al., 2003a).
Exploitation: A directed fishery by jig-ging machines exists for this species offNorway, with highly variable annual yields,which peaked in the early 1980s reaching10,000 metric tons. It usually constitutes ofa by-catch of trawl fisheries and occasion-ally of long-line and pelagic net fisheries,while it is also fished with hand-jigs by arti-sanal and sports fishermen, mainly duringsummer over deep waters. In someMediterranean countries it has a relatively
high commercial value. It is marketedfresh, frozen, salted or dried and is alsoused as bait in finfish fisheries.
Conservation Status: No populationassessment has been conducted in Hel-lenic seas. No official statistics are avail-able for T. sagittatus in any ICES area orthe Mediterranean Sea, because landingsof ommastrephid species (Illex coindetii,Todaropsis eblanae, Todarodes sagittatus,and Ommastrephes bartramii) are allpooled together (PEREIRA et al., 1998;JEREB, 2000). Thus, no populationassessment is possible based on commer-cial catches.
Todaropsis eblanae (Ball, 1841)
General: T. eblanae is a demersal mid-sized ommastrephid squid, with a maxi-mum recorded mantle length 29 cm forfemales and 23 cm for males (ROBIN etal., 2002). It is associated with sandy andmuddy bottoms and it mostly occurs on theshelf break and upper slope, not ascendingto the surface or approaching the shore. Itspopulations have been considered asindicative of the cephalopod community atthese depths, in the south-eastern Atlantic(ROELEVELD et al., 1992), the BalearicSea (QUETGLAS et al., 2000), the northTyrrhenian (SANCHEZ et al., 1998), thesouth-eastern Adriatic (KRSTULOVIC-SIFNER et al., 2005) and the eastern Ion-ian Sea (LEFKADITOU et al., 2001).Mature individuals are found all yearround, with maximum incidence of maturefemales from mid-spring to early autumn(GONZALEZ et al., 1994; RELINI et al.,1999; ROBIN et al., 2002). T. eblanae fol-lows the multiple pattern of spawning, itsPF decreasing with latitude and reaching275 000 for larger females off West Africa(LAPTIKHOVSKY & NIGMATULLIN,
Medit. Mar. Sci., 9/1, 2008, 77-118 103

1999) but not exceeding 28 000 in Scottishwaters (HASTIE et al., 1994). Teleosteanfish have been found to compose themajor part of its stomach contents, withcrustacean and cephalopod (including itsown species) percentages varying by geo-graphic area and specimen size (RASEROet al., 1996; LORDAN et al., 1998). Itsmost important predators are teleost fish,toothed whales and selachians.
Distribution: T. eblanae is distributed inthe Mediterranean Sea and E Atlantic,from 61Æ 15' N to 40Æ S, as well as in theSW Pacific and the SW Indian Oceans offAustralia. It is widespread in Hellenic seas,where it has been recorded from 100 to 850m of depth, being more abundant on theupper slope of regions with steep waters,like the Toroneos Gulf (LEFKADITOU,2006) and the Ionian Sea (Table 3).
Exploitation: T. eblanae is a by-catch inbottom trawl fisheries that is either dis-carded or landed with Illex coindetii. Noofficial statistics or research estimates existon the landed catch of T. eblanae fromHellenic waters. Studies of trawl fisherylandings have shown a marked seasonalityof the species catches, with a peak duringsummer in the northern Tyrhenian Sea(BELCARI et al., 1998) and almost equallevels of T. eblanae and I. coindetii landingsfrom northern Spanish Atlantic waters(GONZALEZ et al., 1994).
Conservation Status: Due to its poolingwith other ommastrephid species in fisherystatistics, it is not possible to assess catchlevels as an index of stock biomass. Indica-tive trends of density and biomass indicesfor this species on the fishing groundsacross the north coasts of the Mediter-ranean Sea might probably be feasiblethrough the database of the MEDITS(International Bottom Trawl Surrvey inthe Mediteranean) surveys, which have
been conducted during the summer overthe continental shelf and slope since 1994.
Sepia elegans (Blainville, 1827)
General: Sepia elegans is a small cuttle-fish; males attain a mantle length of 72 mmand females up to 89 mm, their maximumweight not exceeding 60 g (REID et al.2005). S. elegans is a sublittoral species ofsandy and sandy-muddy bottoms of thecontinental shelf and the upper slope,being among the dominant cephalopodspecies on the upper shelf (SANCHEZ etal. 1998; KRSTULOVIC-SIFNER et al.,2005; LEFKADITOU, 2007). In the Seaof Marmara, S. elegans has also beenrecorded in brackish waters (18-25 psu)(UNSAL et al., 1999), which indicates ahigh degree of salinity tolerance. Matureindividuals are present all year round, withmaximum incidence during summer andautumn in the north Aegean Sea (D’ONGHIA et al., 1996), while during springand summer in the western Mediterranean(MANGOLD-WIRZ, 1963). A seasonalmigration has been reported in some areaswith individuals spending their winter indeep waters (200–400 m) and then migrat-ing into shallower waters in spring andsummer, which however, has not beenobserved in other areas (RELINI et al.,1999). Feeding studies conducted in the E.Atlantic have shown the preference of S.elegans for small crustaceans, fish andpolychaetes (REID & JEREB, 2005). Itsremains have been frequently found in thestomach contents of Scyliorhinus caniculaand Raya clavata from the Aegean Sea(KABASAKAL, 2002).
Distribution: S. elegans is found in theeastern Atlantic from Ireland and westernScotland (~50ÆN) to the Sahara Bank andNamibia (21ÆS) and throughout the
Medit. Mar. Sci., 9/1, 2008, 77-118104

Mediterranean Sea including the brackishwaters of the Sea of Marmara (UNSAL etal., 1999) It is common in Hellenic seaswhere it has been reported at depths ofbetween 20 and 600 m.
Exploitation: S. elegans is a by-catchspecies of trawl fisheries; its catchesexhibiting a marketable proportion in Por-tugal, Spain, Italy and Hellas (SARTOR etal., 1998; MACHIAS et al., 2001;SENDAO et al., 2002) depending on thelanding port and sale perspectives of thespecies, which is usually marketed mixedwith S. orbignyana and small S. officinalis.In some areas S. elegans represents a sig-nificant percentage of the catch and con-stitutes a valuable resource locally (REIDet al., 2005).
Conservation Status: No populationassessment has been conducted in Hel-lenic seas or other geographical areas.
Sepia orbignyana (Ferussac, 1826)
General: Sepia orbignyana is a cuttle-fish of medium size, with maximum mantlelength recorded in the Mediterranean Sea9.3 cm for males and 9.6 cm for females(REID et al., 2005). Mature females arecaught from spring to autumn whilemature males are collected during all sea-sons (RELINI et al., 1999;LEFKADITOU, 2006), a fact that indi-cates earlier maturation of the males.Mature females carry about 400 oocytesand males about 100 spermatophores(REID et al., 2005). According to a studyof its feeding in the Tuscan waters, crus-taceans constitute its preferred prey, fol-lowed by fish, cephalopods and echino-derms (RELINI et al., 1999).
Distribution: Sepia orbignyana extendsfrom Irish waters south to Angola (17Æ S)in the eastern Atlantic and throughout the
Mediterranean Sea, existing also in thebrackish waters of the Sea of Marmara(REID et al., 2005). Its bathymetric rangeextends from a few meters down to middleslope, with its deepest record from a haulat 700-770 m in the southern Aegean Sea(MEDITS – International bottom trawlsurvey in the Mediterranean, unpublisheddata). However, S. orbignyana is more fre-quently caught on lower continental shelf,being an indicative species of cephalopodassemblages at these depths (SANCHEZet al., 1998; QUETGLAS et al., 2002;LEFKADITOU, 2006). Its remains havebeen noted among prey species of Rayaclavata from the Aegean Sea(KABASAKAL, 2002) and the demersalfish community in the Bay of Biscay(VELASCO et al., 2001).
Exploitation: Sepia orbignyana repre-sents a common trawl by-catch, regularlypresent in Italian and Spanish markets(RELINI et al., 1999), being marketablealso in Hellas (MACHIAS et al., 2001). Itis marketed frozen and fresh, usuallymixed with S. elegans and small individualsof the common cuttlefish Sepia officinalis,so that there are no separate statisticaldata for this species’ landings.
Conservation Status: Density and bio-mass data have been collected systemati-cally for Sepia orbignyana during the trawlsurveys conducted in the Mediterraneanwaters of European countries since thesummer of 1994, but are not sufficient forstock assessment of this short-livedspecies.
Rossia macrosoma (Delle Chiaje, 1830)
General: Rossia macrosoma is thelargest sepiolid species in the Mediter-ranean Sea; its mantle length attaining 5.4cm for males and 8.4 cm for females
Medit. Mar. Sci., 9/1, 2008, 77-118 105

(MANGOLD-WIRTZ, 1963;LEFKADITOU, 2006). Mature speci-mens of both sexes are found during allseasons, indicating an extended spawningperiod that probably peaks in spring andautumn (REID & JEREB, 2005). Poten-tial fecundity reaches 208 eggs, whilecounts of spermatophores inside a male’sSac of Needham do not exceed 150(LEFKADITOU, unpublished data). Noinformation is available on its feedinghabits. Remains of R. macrosoma havebeen identified among the stomach con-tents of demersal fishes and chon-drichthyans (BELLO, 1997; VELASCO etal., 2001).
Distribution: It is widespread in theMediterranean Sea and across the north-eastern Atlantic, extending also intowaters around Greenland (REID &JEREB, 2005). It has been reported atdepths ranging between 32 and 900 m,caught more frequently on upper slopes. Ithas been found in the Aegean Sea and theIonian Sea, at depths between 72 and 700m. Mature individuals of both sexes aredistributed mainly on upper slopes where-as younger individuals are mainly found onthe lower shelf (LEFKADITOU &KASPIRIS, 2004; ROSA et al., 2006).
Exploitation: It consistently exhibitsboth a marketable and discarded amountin Hellenic, Italian and Spanish trawl fish-eries (SARTOR et al., 1998; MACHIASet al., 2001). There are no official statisticsfor R. macrosoma commercial catches,which are usually landed mixed with othersepiolids and small eledonids.
Conservation Status: There are nostudies targeting assessment of thisspecies’ stocks. Data on the number ofindividuals and their total weight by haulhave been collected in the framework ofthe International bottom trawl survey in
the Mediterranean (MEDITS), conductedduring the summer over the continentalshelf and slope of European countriessince 1994.
Rondeletiola minor (Naef, 1912)
General: Rondeletiola minor is one ofthe smaller members of the family Sepioli-dae, attaining a mantle length up to 2.3 cm(REID & JEREB 2005) and recognized bythe roundish pair of light organs presentinside the mantle cavity on the ventral sideof the ink sac, easily seen from the ventralmantle opening. Mature individuals arefound throughout the year, with females ma-turing at 1.3-1.7 cm and males at 1.1-1.3 cm(SALMAN & KATAGAN, 1996; LEFKA-DITOU & KASPIRIS, 2004), a factexplaining the almost exclusive presence ofmature individuals among males collectedby selective bottom trawl nets. Potentialfecundity of females reaches 160 eggs(LEFKADITOU unpublished data) whilein males 280 spermatophores have beencounted on average (SALMAN &KATAGAN, 1996). R. minor shows anaggregation forming behaviour, beingamong the cephalopod species character-izing shelf-break areas (GONZALEZ &SANCHEZ, 2002; LEFKADITOU, 2006).R. minor remains have been reported inthe stomach content of Raya clavata(BELLO, 1997).
Distribution: Rondeletiola minor iswidespread in the Mediterranean Seaincluding the brackish waters of the Sea ofMarmara (UNSAL et al., 1999) and in theeastern Atlantic from the north coast ofthe Iberian Peninsula up to the waters offNamibia (30oS) (REID & JEREB 2005).It has been reported at depths rangingfrom 17 to 900 m, with its deepest recordsreported from the northeastern Ionian Sea
Medit. Mar. Sci., 9/1, 2008, 77-118106

(LEFKADITOU et al., 2003a). In theAegean Sea it has been caught down to770 m, with the greatest part of its popula-tion distributed over the continental slope(LEFKADITOU et al., 2003b, LEFKADI-TOU & KASPIRIS, 2004).
Exploitation: Rondeletiola minor is aby-catch species of trawl fisheries, knownto be consumed only in Italy (SARTOR etal., 1998).
Conservation Status: No populationassessment has been conducted for thisspecies across its distributional range.
Sepietta oweniana (Pfeffer, 1908)
General: Sepietta oweniana is the mostabundant species of the Sepiolidae family.Its mantle length reaches 5 cm in theNorth Sea but does not exceed 4 cm in theMediterranean Sea, with females general-ly maturing at around 2.5 cm and males ataround 2.0 cm. Mature individuals arepresent throughout the year with maxi-mum incidence of mature femalesobserved from spring to autumn (RELINIet al., 1999; LEFKADITOU &KASPIRIS, 2004). Fecunded eggs arereleased in one or several spawning eventsand potential fecundity ranges from 50 to240 eggs, while counts of male’s sper-matophores range between 50 and 824(SALMAN, 1998; LEFKADITOU, 2006).S. oweniana feeds primarily on crus-taceans, while cephalopod and fish pro-portions in their stomach contents varyconsiderably by geographic area and depthof capture (LEFKADITOU, 2006). S.oweniana has been found in the stomachsof Stenella coeruleoalba (SANTOS et al.,1995), Galeus melastomus (BERGSTROM& SUMMERS, 1983; BELLO, 1997), Chi-maera monstrosa (BELLO, 1997), Rayaclavata (KARABASAKAL, 2002) and
Aristaeomorpha foliacea (BELLO &PIPITONE, 2002).
Distribution: Sepietta oweniana is wide-spread in the Mediterranean Sea includingthe southwestern part of the Sea of Mar-mara (UNSAL et al., 1999) and in thenortheastern Atlantic from Norway toMauritania (15o N), while its occurrence inthe Indian Ocean is doubtful (REID &JEREB, 2005). It is present on soft bot-tom, of continental shelf and slope, butmore abundant on the upper slope, whereit has usually been found among the dom-inant species of cephalopod assemblages(SANCHEZ et al., 1998; BIAGI et al.,2002; GONZALEZ & SANCHEZ, 2002;LEFKADITOU, 2006). It is common inHellenic seas, where it has been recordedat depths of between 20 and 820 m, beingthe most abundant cephalopod species onthe eastern Ionian slope (Table 3).
Exploitation: Sepietta oweniana consti-tutes the major part of landed sepiolids,particularly appreciated in some marketsof the western Mediterranean, represent-ing a significant by-catch of trawl fisheries,especially in Norwegian lobster fishinggrounds (RELINI et al., 1999). Accordingto the results of a study of Hellenic trawlfisheries’ discards for 1995-1998, by-catch-es of this species were only discarded(MACHIAS et al., 2001).
Conservation Status: Studies on thestate of its exploitation and populationassessment are lacking.
Scaeurgus unicirrhus (Orbigny, 1840)
General: Scaeurgus unicirrhus is amedium-sized octopus, with mantle lengthreaching 12 cm for females (MANGOLD–WIRTZ, 1963) and 8 cm for males(LEFKADITOU, 2006). Mature malesare met throughout the year but mature
Medit. Mar. Sci., 9/1, 2008, 77-118 107

females have been caught in the Mediter-ranean Sea from early spring to earlyautumn (MANGOLD –WIRTZ, 1963;LEFKADITOU, unpublished data).Potential fecundity of females is about1000 eggs, while counts of spermatophoresin the males’ spermatophoric sac do notexceed 10. Prey spectrum of the speciesindicates its association with the bottom,made up mainly of carideans crustaceansand in smaller proportions of teleosts,cephalopods and polychaetes (RELINI etal., 1999).
Distribution: Scaeurgus unicirrhus is acosmopolitan species, scattered in tropicaland warm temperate waters, associatedwith sandy and coralline bottoms(ROPER et al., 1984). It has been report-ed at depths between 30 and 850 m, beingrelatively more abundant on lower shelfand upper slope (SANCHEZ et al., 1998;QUETGLAS et al., 2000; GONZALEZ &SANCHEZ, 2002; LEFKADITOU et al.,2003a). It is widespread in Hellenic seas,presenting a wider depth range in the Ion-ian Sea (43-850 m) in comparison to that(30-550 m) in the Aegean Sea, where itseems to be relatively more abundant(Table 3).
Exploitation: Scaeurgus unicirrhus is aby-catch of bottom trawlers that is mostlydiscarded by Hellenic trawlers(MACHIAS et al., 2001) and only occa-sionally marketed with musky octopuses(LEFKADITOU, unpublished data). Inthe western Mediterranean, its discardpercentage by Italian and Spanish trawlersranges from 0 to 50 % (SARTOR et al.,1998), but its economic value is lower thanthat of other commercial octopod species(RELINI et al., 1999).
Conservation Status: No populationassessment has been conducted in Hel-lenic seas or other geographic areas.
Conclusion
People who exploit marine resourceschoose among them according to techno-logical, economic, and social considera-tions. Prehistoric selectivity in theirexploitation has often been postulated.Many molluscs, although not commercial-ly important, have been exploited forhuman consumption or for other uses(fish bait, shell collections, the jewelleryindustry). Although for some speciesthere are indications of substantial popu-lation decline, there is a lack of scientificeffort towards a better understanding oftheir ecology and assessing the status oftheir populations. The situation is evenworse for species of no importance forhumans (not evaluated in this review). Sci-entific effort and financing targeted a fewcommercial species (KOUTSOUBAS etal., 2007; LEFKADITOU, 2007), whilemolluscs of minor or no commercialimportance are neglected. For most ofthese species there is no populationassessment in Hellenic seas with only afew exceptions on a local scale (e.g., Pinnanobilis, Lithophaga lithophaga, Donacillacornea). However, many of these speciesare important constituents of the Mediter-ranean ecosystems and as they are under avariety of pressures nowadays (marinepollution, alien species’ invasions, climatechange, destructive fishing methods, over-exploitation) there is a pressing need toassess the status of their populations andact towards their conservation. Existinglegislation is not enforced in practice andis extensively violated, thus the conserva-tion of the molluscan species is notensured. In addition, a good knowledge ofthe ecology and status of the aforemen-tioned species and appropriate conserva-tion plans are needed.
Medit. Mar. Sci., 9/1, 2008, 77-118108

An important goal in the next decadeis to network targeted collection datanationally and internationally so thisinvaluable source of biodiversity informa-tion can be accessed for conservation andplanning purposes.
Acknowledgements
We would like to thank Dimitris Pour-sanidis for the preparation of the mapspresented in this paper. Evi Vardala-Theodorou has provided useful informa-tion for some species.
References
ABBOTT, T.R., 1960. The genus Strom-bus in the Indo-Pacific. Indo-PacificMollusca, 1: 33–146.
ALIDROMITI, C., 2007. Study of age andgrowth of Alloteuthis media(Cephalopoda: Loliginidae) in theNorth Aegean Sea. Investigation ofgeographical and gender differencesin the statolith morphology and themantle growth. MSc dissertation,University of Athens, Athens, Hellas,68 pp (in Hellenic, English abstract).
AL-MADFA, H., ABDEL-MOATI,M.A.R. & AL-GIMALY, F.H., 1998.Pinctada radiata (Pearl Oyster): abioindicator for metal pollution moni-toring in the Qatari waters (ArabianGulf). Bulletin of Environmental Cont-amination and Toxicology, 60: 245-251.
ANTONIADOU C., KOUTSOUBAS D.& CHINTIROGLOU, C., 2005. Mol-luscan fauna from infralittoral hardsubstrate assemblages in the NorthAegean Sea. Belgian Journal of Zoolo-gy, 135 /2: 119-126.
ARKHIPKIN, A., LAPTIKHOVSKY, V.& GOLUB, A., 1999. Population
structure and growth of the squidTodarodes sagittatus (Cephalopoda:Ommastrephidae) in north-westAfrican waters. Journal of the MarineBiological Association of the UnitedKingdom, 79: 467-477.
BELCARI, P., SARTOR, P. & DERANIERI, S., 1998. I cefalopodi nellosbarcato commerciale con reti astrasciso del Mar Tirreno settentri-onale. Biologia Marina Mediterranea, 5:318-325.
BELLO G., 1985. Su una raccolta dicephalopodi pescati nel mesobatialedel golfo di Taranto. Bolletino Malaco-logico, 21/10-12: 275-280.
BELLO, G., 1997. Cephalopods from thestomach contents of demersal chon-drichthyans caught in the Adriatic Sea.Vie Milieu, 47: 221-227.
BELLO, G. & PIPITONE, C., 2002. Preda-tion on cephalopods by the giant redshrimp Aristaeomorpha Foliacea. Jour-nal of the Marine Biological Associationof the United Kingdom, 82: 213-218.
BERGSTROM, B. & SUMMERS, W.,1983. Sepietta oweniana. In: P.R. Boyle(ed.) Cephalopod life cycles. Volume1, Academic press: 75-91.
BIAGI, F., SARTOR, P., ARDIZZONE,G.D., BELCARI, P., BELLUSCIO,A. & SERENA, F., 2002. Analysis ofdemersal fish assemblages of the Tus-cany and Latium coasts (north-Western Mediterranean): communitystructure and biodiversity. ScientiaMarina, 66 (suppl. 2): 233-242.
BORGES, T.C. & WALLACE, J.C., 1993.Some aspects of the fishery biology ofthe Ommastrephid squid Todarodessagittatus (Lamarc, 1798) from theNortheast Atlantic. In: Recentadvances in Fisheries Biology, edited byT. Okutani, R.K. O’Dor & T.
Medit. Mar. Sci., 9/1, 2008, 77-118 109

Kubodera, Tokay University Press,Tokyo, pp. 25-36.
BOUDOURESQUE, C.F., 2004. Marinebiodiversity in the Mediterranean: sta-tus of species, populations and com-munities. Travaux scientifiques du Parcnational de Port-Cros, 20: 97-146.
BROMLEY, R.G. & ASGAARD, U.,1990. Solecurtus strigilatus: a jet-pro-pelled burrowing bivalve. p. 313-320.In: The Bivalvia – Proceedings of aMemorial Symposium in Honor of SirCharles Maurice Yonge, Edinburg,1986, B. MORTON, Hong KongUniversity Press, Hong Kong.
BROWN, K.M. & RICHARDSON, T.D.,1987. Foraging ecology of the south-ern oyster drill Thais haemastoma(Gray): constraints on prey choice.Journal of Experimental Marine Biologyand Ecology, 114: 123-141.
BROWN, K.M. & ALEXANDER Jr, J.E.,1994. Group foraging in a marine gas-tropod predator: Benefits and costs toindividuals. Marine Ecology ProgressSeries, 112: 97-105.
BUSTAMANTE, P & MIRAMAND, P.,2005. Subcellular and body distribu-tions of 17 trace elements in the varie-gated scallop Chlamys varia from theFrench coast of the Bay of Biscay.Science of the Total Environment, 337:59-73.
BUSTAMANTE, P., GERMAIN, P.,LECLERC, G. & MIRAMAND, P.,2002. Concentration and distributionof 210Po in the tissues of the scallopChlamys varia and the mussel Mytilusedulis from the coasts of Charente-Maritime (France). Marine PollutionBulletin, 44: 997-1002.
BUTLER, P.A., 1985. Synoptic review ofthe literature on the southern oysterdrill Thais haemastoma floridana.
NOAA Technical Report NMFS, 35: 1-9.CATSIKI, V.A., KATSILIERI, C. &
GIALAMAS, V., 1994. Chromiumdistribution in benthic species from agulf receiving tannery wastes (Gulf ofGeras - Lesbos Island, Greece).Science of the Total Environment, 145:173-185.
CHEN, C.S., PIERCE, G.J., WANG, J.,ROBIN, J-P., POULARD, J.C.,PEREIRA, J., ZUUR, A.F., BOYLE,P.R., BAILEY, N., BEARE, D.J.,JEREB, P., RAGONESE, S.,MANNINI, A., & ORSI RELINI, L.,2006. The apparent disappearance ofLoligo forbesi from the south of itsrange in the 1990s: trends in Loligo spp.abundance in the northeast Atlanticand possible environmental influences.Fisheries Research, 78: 44-54.
CHINTIROGLOU, C., ANTONIADOU,C., VAFIDIS, D. & KOUTSOUBAS,D., 2005. A review of the biodiversityof hard substrate invertebrate commu-nities in the Aegean Sea. Mediter-ranean Marine Science, 6/2: 51-62.
CONTI, M.E. & CECCHETTI, G., 2003.A biomonitoring study: trace metals inalgae and molluscs from Tyrrheniancoastal areas. Environmental Research,93: 99-112.
DALY, H. I., PIERCE, G. J., SANTOS,M. B., ROYER, J., CHO, S. K., G., S.,ROBIN, J.-P. & HENDERSON, S.,2001. Cephalopod consumption byfish in UK waters. Fisheries Research,52: 51-64.
DELAMOTTE, M. & VARDALA-THEODOROU, E., 2007. Shells fromGreek seas. Goulandris Natural Histo-ry Museum, Athens.
DEL PIERO, D. & DACAPRILE, R.,1998. The alternating recruitment pat-tern in Ensis minor, an exploited
Medit. Mar. Sci., 9/1, 2008, 77-118110

bivalve in the Gulf of Trieste, Italy.Hydrobiologia, 375/376: 67-72.
DONEDU, M. & MANUNZA, B., 1995.Mollusca Gastropoda found in associ-ation with Verongia aerophoba (Sch-midt). La Conchiglia 275: 4-7.
EEA, 2007. Halting the loss of biodiversityby 2010: proposal for a first set of indi-cators to monitor progress in Europe,EEA Technical report No 11/2007,182pp, http://reports.eea.europa.eu/technical_report_2007_11/en/Tech_report_11_2007_SEBI.pdf
EVAGELOPOULOS ∞. & D.KOUTSOUBAS, 2008. Communitystructure and dynamics of the mollus-can macrofauna at the marine-lagoon-al environmental transition in the Kal-loni solar saltworks (Lesvos Island, NEAegean Sea, Greece). Journal of Nat-ural History (in press).
FAINZILBER, M. & KLEIMAN, Y.,1989. Preliminary research on thevenom of Conus ventricosus Gmelin(Mollusca: Gastropoda) from theMediterranean Sea. Israel Journal ofZoology, 35: 96.
FÄNGE, R. & LIDMAN, U., 1976. Secre-tion of sulfuric acid in Cassidariaechinophora lamark (Mollusca: Meso-gastropoda). Comparative Biochem-istry and Physiology, 53: 101-103.
FAO, 2008. Cultured Aquatic SpeciesInformation Programme: Crassostreagigas (Thunberg, 1793). WWW publi-cation, http://www.fao.org/fishery/cul-turedspecies /Crassostrea_ gigas.
GALINOU-MITSOUDI, S. & SINIS, A.I.,1994. Reproductive cycle and fecundi-ty of date mussel Lithophaga lithopha-ga, (L.) (Bivalvia: Mytilidae). Journal ofMolluscan Studies, 60: 371-385.
GALINOU-MITSOUDI, S. & ∞. π.SINIS, 1995. Age and growth of
Lithophaga lithophaga, (L.) (Bivalvia:Mytilidae) based on annual growthlines in the shell. Journal of MolluscanStudies, 61: 435-453.
GALINOU-MITSOUDI, S. & SINIS,A.I., 1997. Population dynamics of thedate mussel Lithophaga lithophaga, (L.1758) (Bivalvia: Mytilidae), in theEvvoikos gulf (Greece). Helgolander,51: 137-154.
GALINOU-MITSOUDI, S., VLAHAVAS,G. & PAPOUTSI, O., 2006. Popula-tion study of the protected bivalvePinna nobilis (Linaeus, 1758) in Ther-maikos Gulf (North Aegean Sea).Journal of Biological Research, 5: 47-53.
GEROVASILEIOU, V., SINI, M.I.,POURSANIDIS, D., LEKKAS, V.,FILIOS, G. & KOUTSOUBAS, D.,2007. Biodiversity in marine areas offthe island of Lesvos (NE Aegean),potentially targets for the establish-ment of diving parks (PreliminaryResults). Proceedings of the 13th Pan-hellenic Ichtyhologists Conference,Mytilini: 567-570.
GÖKSU, M.Z.L., AKAR, M., EVIK, F.& FINDIK, Ö., 2005. Bioaccumula-tion of some heavy metals (Cd, Fe, Zn,Cu) in two bivalvia species (Pinctadaradiata Leach, 1814 and Brachidontespharaonis Fischer, 1870). Turkish Jour-nal of Veterinary and Animal Sciences,29: 89-93.
GONZALEZ, A.F., RASERO, M. &GUERRA, A., 1994. Preliminarystudy of Illex coindetii and Todaropsiseblanae (Cephalopoda: Ommas-trephidae) in northern SpanishAtlantic waters. Fisheries Research, 21:115-126.
GONZALEZ, M. & S NCHEZ, P., 2002.Cephalopod assemblages caught bytrawling along the Iberian Peninsula
Medit. Mar. Sci., 9/1, 2008, 77-118 111

Mediterranean coast. Scientia Marina,66 (suppl. 2): 199-208.
GUERRA, A., S NCHEZ, P., ROCHA,F., 1994. The Spanish fishery for Loli-go: recent trends. Fisheries Research,21: 217-230.
HAMES, S., TZANATOS, E. & WHITE,M., 2001. An investigation into the dis-tribution and abundance of Pinnanobilis in North East Kefallonia.Fiskardo’s Nautical & EnvironmentalClub: 10-11.
HASTIE, L.C., JOY, J.B., PIERCE, G.J.& YAU, C., 1994. Reproductive biolo-gy of Todaropsis eblanae (Cephalopo-da: Ommastrephidae) in Scottishwaters. Journal of the Marine BiologicalAssociation of the United Kingdom, 74:367-382.
HILL, J.M., 2006. Pholas dactylus, com-mon piddock. Marine Life Informa-tion Network: Biology and SensitivityKey Information Sub-programme [on-line]. Plymouth: Marine BiologicalAssociation of the UK. [cited14/09/2007]. Available from:<http://www.marlin.ac.uk/species/Pholasdactylus.htm>
IFANTIDIS, F., 2006. ‘Enigmatic’notched Spondylus ornaments fromthe Neolithic: New evidence from theAegean. The Archaeo+MalacologyGroup Newsletter, 9: 3-5.
JEREB P., 2002. Recent trends inMediterranean Cephalopods captureproduction. Report of the ICESWorking Group on Cephalopod Fish-eries and life-history, working docu-ment, 3: 73-82.
JEREB, P. & RAGONESE, S., 1994. Themediterranean teuthofauna: towards abiogeographical coverage by reg cen-cus. II: Strait of Sicily. Bolletino Mala-cologico, 30/5-9: 11-172.
KARABASAKAL, H., 2002. Cephalo-pods in the stomach contents of fourElasmobranch species from the north-ern Aegean Sea. Acta Adriatica, 43/1:17-24.
KATSANEVAKIS, S., 2006. Populationecology of the endangered fan musselPinna nobilis in a marine lake. Endan-gered Species Research, 1: 51-59.
KATSANEVAKIS, S., 2007a. Density sur-face modeling with line transect sam-pling as a tool for abundance estima-tion of marine benthic species: thePinna nobilis example in a marine lake.Marine Biology, 152: 77-85.
KATSANEVAKIS, S., 2007b. Growth andmortality of the fan mussel Pinnanobilis in Lake Vouliagmeni(Korinthiakos Gulf, Greece): a gener-alized additive modeling approach.Marine Biology, 152: 1319-1331.
KATSARES V., TSIORA, ∞.,GALINOU-MITSOUDI, S. &IMSIDRIDOU, A., 2008. Geneticstructure of the endangered speciesPinna nobilis L., 1758 (Mollusca:Bivalvia) inferred from mtDNAsequences. Biologia (in press).
KORSUN, Y., NESIS, K.N., NIGMA-TULLIN, C., OSTAPENKO, A.A. &PINCHUKOV, M.A., 1979. New dataon the distribution of squids, familyOmmastrephidae, in the world ocean.Oceanology, 19: 472-475.
KOUKOURAS, A. & RUSSO, A., 1991.Midlittoral soft substratum macrofau-nal assemblages in the North AegeanSea. P.S.Z.N.I.: Marine Ecology, 12:293–316.
KOUTSOUBAS, D., 1992. Contribution tothe study of the gastropod molluscs on thecontinental shelf of the N Aegean Sea.PhD Thesis, Aristotelion University ofThessaloniki, ISSN 1105-5049, 586 pp.
Medit. Mar. Sci., 9/1, 2008, 77-118112

KOUTSOUBAS, D., 2004 Biogeographyof the Gasteropod molluscs in Hel-lenic seas. Proceedings of the 7th Pan-Hellenic Symposium of the HellenicGeographic Society, Mytilini, pp. 444-449.
KOUTSOUBAS D., KOUKOURAS A.,KARAKASSIS I. & DOUNAS, C.,1992. Contribution to the knowledgeof Gastropoda and Bivalvia (Mollus-ca) of Crete Island. Bolletino Malaco-logico, 28/1-4: 69-82.
KOUTSOUBAS, D., KOUKOURAS, A.& VOULTSIADOU-KOUKOURA,E., 1997. Prosobranch mollusc faunaof the Aegean Sea: New information,Check List, Distribution. Israel Journalof Zoology, 43: 19-54.
KOUTSOUBAS D., ARVANITIDIS C.,VALAVANIS V., GEORGAKA-RAKOS S., KAPANTAGAKIS A.,MAGOULAS A. & G. KOTOULAS,1999. Cephalopod resources in theEastern Mediterranean with particularemphasis on Greek Seas: present andfuture perspectives. ICES LivingResources Committee, ICESCM19/K: 3, Working Group on Cephalo-pod Fisheries and Life History, Herak-lion, 25-26 March 1999, pp. 12-25.
KOUTSOUBAS D., TSELEPIDES A. &ELEFTHERIOU, A., 2000a. Deep seamolluscan fauna of the Cretan Sea(Eastern Mediterranean): faunal, eco-logical and zoogeographical remarks.Senckenbergiana maritim·, 30/3-6: 85-98.
KOUTSOUBAS D., ARVANITIDIS C.,DOUNAS C., & DRUMMOND, L.,2000b. Community structure anddynamics of the molluscan fauna in aMediterranean lagoon (Gialovalagoon, SW Greece). Belgian Journalof Zoology, 130: 135-142.
KOUTSOUBAS, D., GALINOU-
MITSOUDI, S., KATSANEVAKIS,S., LEONTARAKIS, P., METAXA-TOS, A. & ZENETOS, A., 2007. II.5.Bivalve and gastropod molluscs ofcommercial interest for human con-sumption in the Hellenic seas. p. 70-84. In: State of Hellenic Fisheries,C.Papaconstantinou, A. Zenetos, V.Vassilopoulou & G. Tserpes (Eds),Athens, HCMR publ.
KRSTULOVIC-SIFNER, S., LEFKADI-TOU, E., UNGARO, N., CERIOLA,L., OSMANI, K., KAVADAS, S. &VRGOC, N., 2005. Composition anddistribution of the cephalopod faunain the easterna Adriatic and easternIonian Sea. Israel Journal of Zoology,51: 315–330.
LAPTIKHOVSKY, V.V. & NIGMA-TULLIN, C.M., 1999. Egg size and fe-cundity in females of the subfamiliesTodaropsinae and Todarodinae (Cepha-lopoda: Ommastrephidae). Journal ofthe Marine Biological Association of theUnited Kingdom, 79: 569-570.
LAPTIKHOVSKY, V., SALMAN, A.,ÖNSOY, B. & KATAGAN, T., 2002.Systematic position and reproductionof squid of the genus Alloteuthis(Cephalopoda: Loliginidae) in theeastern Mediterranean. Journal of theMarine Biological Association of theUnited Kingdom, 82: 983-985.
LEFKADITOU, E., 2006. Taxonomy andbiology of Cephalopods in the NorthAegean Sea. PhD thesis, University ofPatras, Patras, Hellas, 298 pp + Annex-es (in Hellenic, English abstract).
LEFKADITOU, E., 2007. II.4. Review ofcephalopod fauna in Hellenic waters.p. 62-69. In: State of Hellenic Fisheries,C. Papaconstantinou, A. Zenetos, V.Vassilopoulou & G. Tserpes (Eds),Athens, HCMR publ.
Medit. Mar. Sci., 9/1, 2008, 77-118 113

LEFKADITOU, E. & KASPIRIS, P.,2005. Distribution and abundance ofsepiolids (Mollusca: Cephalopoda) offthe north-eastern Greek coasts. Bel-gian Journal of Zoology, 135: 199-204.
LEFKADITOU, ∂., MAIORANO, P. &.MYTILINEOU, CH., 2001. Cephalo-pod species captured by deep-waterexploratory trawling in the easternIonian Sea. NAFO Special Sympo-sium on Deep-Sea Fisheries,Varadero, Cuba, September 2001,NAFO SCR Doc.01/131, 8pp.
LEFKADITOU, E., MYTILINEOU, C.,MAIORANO, P. & D' ONGHIA, G.,2003a. Cephalopod species capturedby deep-water exploratory trawling inthe northeastern Ionian Sea. Journal ofNorthwestern Atlantic Fisheries Science,31: 431-440.
LEFKADITOU, E., PERISTERAKI, P.,BEKAS, P., TSERPES, G., POLITOU,C.-Y. & PETRAKIS, G., 2003b.Cephalopod distribution in the south-ern Aegean Sea. Mediterranean MarineScience, 4/1: 79-84.
LIPPARINI, G. & NEGRA, O., 2002.Increased polymorphism appearingrecently in a population of Erosariaspurca (Linné, 1758) (GastropodaProsobranchia: Cypraeidae) along aportion of the Sicilian coast. LaConchiglia, 302: 23-31.
LORDAN, C., BURNELL, G.M. &CROSS, T.F., 1998. The diet and eco-logical importance of Illex coindetiiand Todaropsis eblanae (Cephalopoda:Ommastrephidae) in Irish waters.South African Journal of MarineScience, 20: 153-163.
LORDAN, C., COLLINS, M.A., KEY,L.N. & BROWNE, E.D., 2001. Thebiology of the ommastrephid squid,Todarodes sagittatus, in the north-east
Atlantic. Journal of the Marine Biologi-cal Association of the United Kingdom,81: 299-306.
MACHIAS, A., VASSILOPOULOU, V.,VATSOS, D., BEKAS, P., KALLIA-NIOTIS A., PAPACONSTA-NTINOU, C. & TSIMENIDES, N.,2001. Bottom trawl discards in thenortheastern Mediterranean Sea.Fisheries Research, 53: 181-195.
MALAQUIAS, M.A.E., BENTES, L.,ERZINI, K. & BORGES, T.C., 2006.Molluscan diversity caught by trawlingfisheries: a case study in southern Por-tugal. Fisheries Management and Ecol-ogy, 13: 39-45.
MANGOLD-WIRZ, K., 1963. Biologiedes céphalopodes benthiques et nec-toniques de la Mer Catalane. VieMilieu, 13: 1-285.
MANGOLD K. & S.V. BOLETZKY,1987. Céphalopodes. p.633-714. InFiches FAO d’ identification des espècespour les besoins de la pêche (Révision 1)Méditerranée et mer Noire. Zone depêche 37. Vol. I. Végétaux et Invertébrés.Fischer W., Bauchot M.-L. & M. Sch-neider (Eds). Rome: FAO.
MARTINS, H.R., 1982. Biological studiesof the exploited stock of Loligo forbesi(Cephalopoda) in the Azores. Journalof the Marine Biological Association ofthe United Kingdom, 62: 799-808.
MASSILIA, G.R., SCHININA, M.E.,ASCENZI, P. & POLTICELLI, F.,2001. Contryphan-Vn: a novel peptidefrom the venom of the Mediterraneansnail Conus ventricosus. Biochemicaland Biophysical Research Communica-tions, 288: 908-913.
MAVIDIS, M. & KOUKOURAS A,2001a. Life cycle and production ofDonacilla cornea (BIVALVIA:MESODESMATIDAE). 22nd Pan-
Medit. Mar. Sci., 9/1, 2008, 77-118114

hellenic Conference of the HellenicSociety of Biological Sciences, Chios,24–27 May 2001, p. 104.
MAVIDIS, M. & KOUKOURAS A,2001b. Population dynamics ofDonacilla cornea (BIVALVIA:MESODESMATIDAE). 22nd Pan-hellenic Conference of the HellenicSociety of Biological Sciences, Chios,24–27 May 2001, p. 106.
MAVIDIS, M., CHARTOSIA, N.,KITSOS, M.-S., NIKOLAIDOU, A.,KOUKOURAS, A. & ELEFTHE-RIOU, A., 2006. Factors affecting thedistribution of the bivalve Donacillacornea on Mediterranean sandy beach-es. 10th International Congress on theZoogeography and Ecology of Greeceand Adjacent Regions, Patras, 26–30June 2006, p. 77.
MICU, D. & MICU, S., 2006. Recentrecords growth and proposed IUCNstatus of Donacilla cornea (Poli, 1795)from the Romanian Black Sea. Cerc-etari Marine I.N.C.D.M., 36: 117-132.
MUTLU, E., 2004. Sexual dimorphisms inradula of Conomurex persicus (Gastro-poda: Strombidae) in the Mediterra-nean Sea. Marine Biology, 145: 693-698.
NICOLAY, K., 1986. Nonstop spreadingof Mediterranean Strombus. LaConchiglia, 18/202-203: 29.
OLIVERIO, M., 1995. The status of theliving Mediterranean Strombus, or:what is a lessepsian migrant. NotiziarioCISMa, 16[1994]: 35-40.
OWEN, D.F. & WHITELEY, D., 1988.The beach clams of Thessaloniki:reflexive or apostatic Selection? Oikos,51: 253-255.
PEN~A J.B., CANALES, J., ADSUARA,J.M. & SOS, M.A., 1996. Study of sea-sonal settlements of five scallopspecies in the western Mediterranean.
Aquaculture International, 4: 253-261.PEREIRA, J.M.F., MORENO, A. &
CUNHA, M.M., 1998. Western euro-pean squid distribution: a review.ICES C.M. 1998/M:29, 20pp.
PERISTERAKI, P., TSERPES G. &LEFKADITOU E., 2005. Investiga-tion of prey-predator relationsthrough cephalopod remains inXiphias gladius stomachs. Journal ofFish Biology, 67/2: 549-554.
PIATOWSKI, U., HERNANDEZ-GARCIA, V. & CLARKE, M.R.,1998. On the biology of the Europeanflying squid Todarodes sagittatus(Lamarck, 1798) (Cephalopoda,Ommastrephidae) in the central-east-ern Atlantic. South African Journal ofMarine Science, 20: 375-383.
PIERCE, G.J., BOYLE, P.R., HASTIE,L.C. & SHANKS, A.M., 1994a. Distri-bution and abundance of the fishedpopulation of Loligo forbesi in UKwaters: analysis of fishery data. Fish-eries Research, 21: 193-216.
PIERCE, G.J., BOYLE, P.R., HASTIE,L.C. & KEY, L., 1994b. The life histo-ry of Loligo forbesi (Cephalopoda:Loliginidae) in Scottish waters. Fish-eries Research, 21: 17-41.
PINN, E.H., RICHARDSON, C.A.,THOMPSON, R.C. & HAWKINS,S.J., 2005. Burrow morphology, biom-etry, age and growth of piddocks (Mol-lusca: Bivalvia: Pholadidae) on thesouth coast of England. Marine Biolo-gy 147: 943-953.
POPPE, G.T. & GOTO, Y., 1993. Euro-pean Seashells. Vol. II (Scaphopoda,Bivalvia, Cephalopoda). VerlagChrista Hemmen, Wiesbaden, 221 pp.
QUETGLAS, A., ALEMANY, F.,CARBONELL, A., MERELLA, P. &S NCHEZ, P., 1998. Some aspects
Medit. Mar. Sci., 9/1, 2008, 77-118 115

of the biology of Todarodes sagittatus(Cephalopoda: Ommastrephidae)from the Balearic Sea (westernMediterranean). Scientia Marina,62/1-2: 73-82.
QUETGLAS, A., ALEMANY, F.,CARBONELL, A., MERELLA, P. &S NCHEZ, P., 1999. Diet of theEuropean flying squid Todarodes sagit-tatus (Cephalopoda: Ommastrephi-dae) in the Balearic Sea (westernMediterranean). Journal of the MarineBiological Association of the UnitedKingdom, 79: 479-486.
QUETGLAS, A., CARBONELL, A. &SANCHEZ, P., 2000. Demersal conti-nental shelf and upper slope cephalo-pod assemblages from the BalearicSea (north-western Mediterranean).Biological aspects of some deep-seaspecies. Estuarine, Coastal and ShelfScience, 50: 739-749.
QUETGLAS, A. & MORALES-NIN, B.,2004. Age and growth of the ommas-trephid squid Todarodes sagittatus fromthe western Mediterranean Sea. Jour-nal of the Marine Biological Associationof the United Kingdom, 84: 421-426.
RASERO, M., GONZALEZ, A.F.,CASTRO, B.G. & GUERRA, A.,1996. Predatory relationships of twosympatric squid, Todaropsis eblanaeand Illex coindetii (Cephalopoda:Ommastrephidae) in Galician waters.Journal of the Marine Biological Associ-ation of the United Kingdom, 76: 73-87.
REGOLI, F., ORLANDO, E., MAURI,M., NIGRO, M., COGNETTI, G.A.,1991. Heavy metal accumulation andcalcium content in the bivalve Donacil-la cornea. Marine Ecology ProgressSeries, 74: 219-224.
REID, A. & JEREB, P., 2005. FamilySepiolidae. In: Cephalopods of the
World. An annotated and illustrated cat-alogue of species known to date. Volume1. Chambered nautiluses and sepioids(Nautilidae, Sepiidae, Sepiolidae, Sepi-adariidae, Idiosepiidae and Spirulidae),P. Jereb & C.F.E. Roper (Eds). FAOSpecies Catalogue for Fisheries Pur-poses 4/1: 153-203.
REID, A., JEREB, P. & ROPER, C.F.E.,2005. Family Sepiidae. In:Cephalopods of the World. An annotat-ed and illustrated catalogue of speciesknown to date. Volume 1. Chamberednautiluses and sepioids (Nautilidae,Sepiidae, Sepiolidae, Sepiadariidae,Idiosepiidae and Spirulidae), P. Jereb& C.F.E. Roper (Eds). FAO SpeciesCatalogue for Fisheries Purposes 4/1:57-152.
RELINI, G., BERTRAND, J. &ZAMBONI, A. (Eds), 1999. Synthesisof the knowledge on bottom fisheryresources in the central Mediter-ranean (Italy and Corsica). BiologiaMarina Mediterranea, 6 (suppl.1).
RICHARDSON, T.D. & BROWN, K.M.,1990. Wave exposure and prey sizeselection in an intertidal predator.Journal of Experimental Marine Biologyand Ecology, 142: 105-120.
RILOV, G., BENAYAHU, Y. &GASITH, A., 2001. Low abundanceand skewed population structure ofthe whelk Stramonita haemastomaalong the Israeli Mediterranean coast.Marine Ecology Progress Series, 218:189-202.
ROBIN, J.P., DENIS, V., ROYER, J.,CHALLIER, L., 2002. Recruitment,growth and reproduction in Todarop-sis eblanae (Ball, 1841), in the areafished by French Atlantic trawlers.Bulletin of Marine Science, 71: 711-724.
ROELEVELD, M.A.C., LIPINSKI, M.R.,
Medit. Mar. Sci., 9/1, 2008, 77-118116

AUGUSTIN, C.J. & STEWARTB.A., 1992. The distribution and abun-dance of cephalopods on the continen-tal slope of the eastern South Atlantic.South African Journal of MarineScience, 12: 739-752.
ROPER, G.F.E., SWEENEY, M.J. &NAUEN, E., 1984. FAO species Cata-logue. Vol. 3 - Cephalopods of theworld. An annotated and illustratedcatalogue of species of interest to fish-eries. FAO Fisheries Synopsis No 125/ Vol.3: 277p. Rome: FAO.
ROSA, R., PEREIRA, J., COSTA, P.R. &NUNES, M.L., 2006. Distribution,abundance, biology and biochemistryof the stout bobtail squid Rossia macro-soma from the Portuguese coast.Marine Biology Research, 2: 14-23.
SALMAN, A., 1998. Reproductive biologyof Sepietta oweniana (Pfeffer, 1908) (Se-piolidae: Cephalopoda) in the AegeanSea. Scientia Marina, 62/4: 379-383.
SALMAN, A. & KATAGAN, T., 1996. Apreliminary study on reproductionbiology of Rondeletiola minor (Naef,1912) (Sepiolidae:Cephalopoda) inthe Aegean Sea. Su Urunleri Dergisi,13/3-4, 403-408 (in Turkish, abstract inEnglish).
S NCHEZ, P., BELCARI, P. &SARTOR, P., 1998. Composition andspatial distribution of cephalopods intwo north-western Mediterraneanareas. Payne A.I.L., Lipinski M.R.,Clarke M.R. & M.A.C. Roeleveld(Eds) Cephalopod Biodiversity, Ecol-ogy and Evolution, South African Jour-nal of Marine Science, 20: 17-24.
SANTOS, M.B., PIERCE, G.J.,WIJNSMA, G., ROSS, H.M. &REID, R.J., 1995. Diets of smallcetaceans stranded in Scotland 1993-1995. International Council for the
Exploration of the Sea, Copenhagen(Denmark), Marine Mammals Com-mittee, ICES Council Meeting Papers,8 pp.
SANTOS, M. B., CLARKE, M. R. &PIERCE, G. J., 2001. Assessing theimportance of cephalopods in thediets of marine mammals and othertop predators: problems and solutions.Fisheries Research, 52: 121 - 139.
SARTOR P., BELCARI P.,CARBONELL A., GONZALEZ M.,QUETGLAS A. & S NCHEZ P.,1998. The importance of cephalopodsto trawl fisheries in the westernMediterranean. In: Cephalopod Bio-diversity, Ecology and Evolution.A.I.L. Payne, M.R. Lipinski, M.R.Clarke & M.A.C. Roeleveld (Eds),South African Journal of MarineScience, 20: 67-72.
SCARABINO, F., 2003. Ranella olearium(Linnaeus, 1758) (Gastropoda: Ton-noidea): confirmation of its presencein Uruguayan waters. Communica-ciones de la Sociedad MalacolÔ’gica delUruguay, 8/78–79: 215-217.
SHACKLETON, J.C. & VAN ANDEL,TJH., 2007. Prehistoric shore environ-ments, shellfish availability and shell-fish Gathering at Franchthi, Greece.Geoarchaeology, 1: 127- 143.
SHULKIN, V.M., PRESLEY, B.J. &KAVUN, V.Ia., 2003. Metal concen-trations in mussel Crenomytilusgrayanus and oyster Crassostrea gigasin relation to contamination of ambi-ent sediments. Environment Interna-tional, 29: 493-502.
STREFTARIS, N. & ZENETOS, A.,2006. Alien marine species in theMediterranean – the 100 ‘Worst Inva-sives’ and their impact. MediterraneanMarine Science, 7: 87-117.
Medit. Mar. Sci., 9/1, 2008, 77-118 117

TAYLOR, J.D., 1987. Feeding ecology ofsome common intertidal neogas-tropods at Djerba, Tunisia. Vie etMilieu, 37: 13-20.
TEMPLADO, J., 1991. Las species delgenero Charonia (Mollusca: Gas-tropoda) en el Mediterraneo. p.133-140. In: Les espèces marines à pro-téger en Méditerranée. C.F.Boudouresque, M. Avon & V. Gravez,France, GIS Posidonie publ.
THOMPSON, S., BUDZINSKI, H.,GARRIGUES, P. & NARBONNE,J.F., 1999. Comparison of PCB andDDT distribution between water-col-umn and sediment-dwelling bivalves inArcachon Bay, France. Marine Pollu-tion Bulletin, 38: 655-662.
TLIG ZOUARI, S., & ZAOUALI, J.,1994. Reproduction de Pinctada radia-ta (Leach, 1814, Mollusque, Bivalve)dans les îles Kerkennah (Tunisie). VieMarine, 4: 41-45.
UNSAL I., UNSAL N., ERK, M.H. &KABASAKAL, H., 1999. Demersalcephalopods from the Sea of Marmarawith remarks on some ecological char-acteristics. Acta Adriatica, 40: 105-110.
VALLI, G., BRAIDA, R. & GASPARI,T., 1983. Considerations sur la repro-duction chez Ensis minor (Chenu)(Mollusca, Bivalvia). 28th CIESMCongress Proceedings, 239-240.
VAN URK, R.M., 1964. The Genus Ensisin Europe. Basteria, 28: 13-44.
VELASCO, F., OLASO, I. & S NCHEZ,F., 2001. The role of cephalopods asforage for the demersal fish communi-ty in the southern Bay of Biscay. Fish-
eries Research, 52: 65-77.YOUNG, L.A., 2007. Expansion of the
distribution of Strombus persicus: firstdocumentation from the SaronikosGulf (eastern Mediterranean).Xenophora, 116: 24-25.
ZENETOS, A., 1986. Systematics, ecologyand distribution of bivalvia (Mollusca)of the Patraikos Gulf. Ph.D. thesis,University of Athens, 268pp.
ZENETOS, A., (Ed) 1996. The marineBivalvia (Mollusca) of Greece. In: Hel-lenic Zoological Society & NCMRFauna Greciae VII, 319pp.
ZENETOS, A., GOFAS, S., RUSSO, G.& TEMPLADO, J., 2004. CIESMAtlas of exotic species in the Mediter-ranean. Vol. 3, Molluscs. CIESM Pub-lishers, Monaco.
ZENETOS, A., ARVANITIDIS, C.,THESSALOU-LEGAKI, M. &SIMBOURA, N., 2005. VI.4 Zooben-thos – Soft bottom fauna. In: State ofthe Hellenic Marine Environment. Pap-athanassiou, E. & Zenetos, A. (Eds),Athens, HCMR, 236-246.
ZENETOS, A., VASSILOPOULOU, V.,SALOMIDI, M. & POURSANIDIS,D., 2007. Additions to the marine alienfauna of Greek waters (2007 update).JMBA2, Biodiversity Records, pub-lished on-line.
ZUEV, G.V. & NESIS, K.N., 1971. Squid(Biology and Fishing). In SelectedEnglish translations of Kir N. NesisCephalopod Publications, vol. II(compiled by M.J. SWEENEY), 291p. Washington (U.S.A.): SmithsonianInstitution Libraries, 2003.
Medit. Mar. Sci., 9/1, 2008, 77-118118
Published on line: June 2008




















