
Temperature Sensitivity of Cardiac Function in Pelagic Fishes
Upload
independentCategory
view
4download
0
P.S.Z.N. I: Marine Ecology, 17 (1-3): 447471 (1996) 0 1996 Blackwell Wissenschafts-Verlag, Berlin ISSN 0173-9565
Accepted June 16,1995
Pelagic-Benthic Coupling in the Nordic Seas: The Role of Episodic Events PAUL WASSMANN, INGER ANDREASSEN, MARIT REIGSTAD & DAG SLAGSTAD*
Norwegian College of Fishery Science, University of Tromse, N-9037 Trorns~, Norway. *The Foundation of Scientific and Industrial Research (SINTEF), Division of Automatic Control, The Norwegian Institute of Technology, N-7034 Trondheim-NTH, Norway.
With 10 figures
Key words: Plankton dynamics, advection, sedimentation, Nordic Seas.
Abstract. The consequences of the following episodic phenomena for the pelagic-benthic coupling in the Nordic Seas are illustrated: (1) Advection of water masses between fjords and shelf environments, (2) freshwater run-off and vertical stability, (3) dynamics of the marginal ice zone in the central and northern Barents Sea and the Polar Ocean, (4) drift patterns of sinking particles along the North Norwegian coast, ( 5 ) advection of zooplankton into subarctic fjords and the southern Barents Sea, zooplankton overwintering and composition, and (6) transport of organic particulate matter from the Barents Sea shelf. It is shown that physical processes in the north-eastern North Atlantic and Polar Ocean can be strongly variable on time scales of days to decades. They have a significant influence on the dynamics of pelagic-benthic coupling. The physical oceanography influences the vertical and horizontal particle flux not only directly (mixing, advection, up- and down-welling), but also indirectly through its impact on the biota (for example radiation, wind, ice cover, freshwater run-off and overwintering, advection and retention of zooplankton). Understanding pelagic-benthic coupling at high latitudes depends even more on a best possible understanding of the physical oceanography and the time scales involved than elsewhere.
Problem
The increasing interest in system-ecological research from the late 1960s onwards resulted in various research directions in marine ecology which previously were only touched upon marginally. Among these directions is pelagic-benthic coupling, the study of the relationship between processes in the water column and those of the nepheloid layer, the benthos, and the sediment surface. The main process connecting plankton and benthos inside the frame of the carbon cycle is the vertical flux of organic matter. Presently, spatial and temporal patterns in the flux of sinking organic matter are central to the understanding of elemental dynamics and foodwebs in the ocean (SMETACEK et al., 1984; LEGENDRE, 1990; WASSMANN et al., 1991; LOHRENZ et al., 1992). Sedimentation removes suspended biomass from the water column and concurrently represents an input of energy and matter to the benthos. It links, therefore, the production of the water column with that of the sediment (GREBMEIER & BARRY, 1991; WASSMANN, 1991; GRAF, 1992). Pelagic-benthic coupling in
U. S. Copyright Clearance Center Code Statement: 0173-9565/96/1701 - 0447 $ 1 l.H)/O

448 WASSMANN, ANDREASSEN, REIGSTAD & SLAGSTAD
general terms is understood as the connection between the pelagial and the benthal and vice versa. Here, we use the term exclusively to denote the processes connecting the pelagial with the benthal.
In the early 1980s, it was widely accepted that the sedimentation rate in a given region could be ascertained from knowledge of primary production: the higher the production, the greater the proportion settling out (e.g.. BERGER et al., 1989; WASSMANN, 1990). The concept of primary production, however, underwent a differentiation: we now distinguish "new" from "regenerated" production (EPPLEY & PETERSON, 1979), and the "new" production sets the upper limit to particle flux out of a given water column (PLAIT er al., 1988). It seems reasonable to assume that major flux pulses follow periods dominated by new production, e.g., the spring bloom. Attention is currently being focused on the processes that control the vertical flux of particles from the mixed layer as well as the formation and export of aggregates (e.g., ALLDREDGE & SILVER, 1988; K~BRBOE & HANSEN, 1993; PASSOW et al., 1994) and faecal pellets (e.g., BATHMANN et al., 1990a; Non et al., 1991).
At low latitudes and non-upwelling systems the annual phyto- and zooplankton cycles vary within narrow limits and pelagic systems are potentially capable of retaining and recycling most autochthonous organic material, although some losses due to sinking particles occur. With increasing latitude, however, the variability in the physical environment as well as the relationship between phyto- and zooplank- ton tightens, giving rise to less predictability and a tendency towards an increased episodic nature of the pelagic-benthic coupling. Locally, this is reflected in decoup- ling between primary and secondary producers, giving rise to extensive sedimenta- tion of ungrazed phytoplankton in shallow areas of the boreal zone (e.g., LAWS et al., 1988; WASSMANN, 1991). Some of the world's highest sediment carbon (Ro- MANKEVICH, 1984) and benthic biomass concentrations (ZENKEVITCH, 1963) are found on Arctic shelfs; this possibly reflects the tight pelagic-benthic coupling characteristic of shallow Arctic environments (HBPNER-PETERSEN, 1989; GREB-
The current understanding of pelagic-benthic coupling is too rudimentary to understand the involved dynamics as revealed by daily and seasonal changes in vertical flux. This is particularly true for subarctic and arctic areas, with their large seasonal variation in physical forcing. Here, the pelagic-benthic coupling and the vertical flux is strongly dominated by episodic events on daily, weekly, seasonal, and yearly time scales. As the physical forcing becomes more and more dominating and the response of the planktonic food web more unpredictable with increasing latitude, the episodic nature of the vertical flux gives rise to increasing ambiguity with regard to predicting seasonal and annual rates. This is particularly true if only average rates are available and if the physical oceanography and climatic forcing is inadequately known. This contribution gathers and presents examples from the literature or from ongoing research at the University of Trams@ with the aim to exemplify various episodic, yet important processes for pelagic-benthic coupling in the Nordic Seas. The selection is based on present evidence and our current understanding of this coupling. It does not claim to be comprehensive. The presen- tation also aims to add to the quantitative and qualitative understanding of pelagic- benthic coupling in Nordic Seas.
MEIER & BARRY, 1991).

Pelagic-benthic coupling in the Nordic Seas and episodic events 449
Scenarios and environments
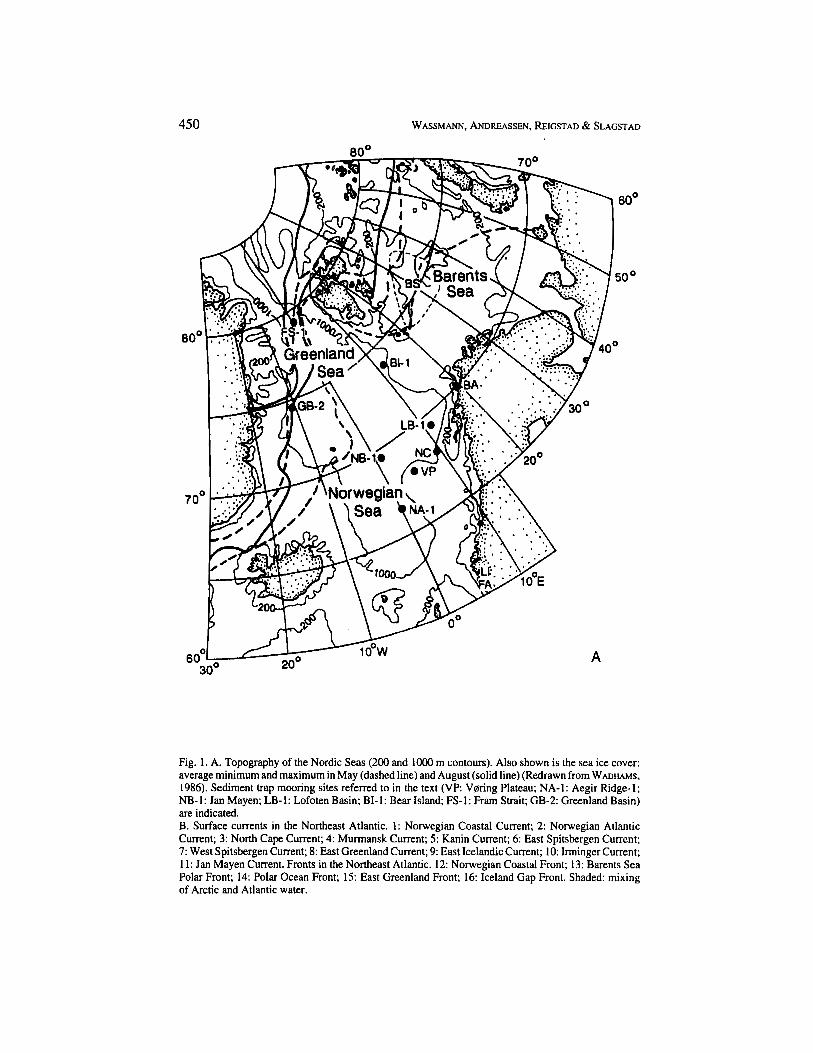
Figure 1 illustrates the areas in the Nordic Seas for which examples will be presented. The coasts are characterised by narrow and broad fjords; shallow banks and the shelf break dominate the outer coastal zone in this area (Fig. 1 A). Rivers discharge into the inner part of North Norwegian fjords, while glaciers are discharged into some of the fjords of Spitsbergen and all fjords in eastern Greenland. Sills are not prominent in most of the fjords in the area. This gives rise to trenches penetrating from the fjords to the shelf break, separating the shelf into shallow coastal banks. Thus, a relatively narrow coastal zone fringes the deep Norwegian Sea to the east and west. To the north, shallow shelf areas like the Barents Sea adjoin the deep basin and the coastal zone. The Barents Sea is permanently ice-free in the south and seasonally ice-covered in the centre and the east. Above 80" N, more or less permanent ice-cover (multi-year ice) is found (Fig. 1 A).
While the North Atlantic Current transports variable amounts of warm, saline water into the Arctic along the Norwegian coast, giving rise to branches onto the shelf and along the shelfbreak, Polar water penetrates from the Polar Ocean into the North Atlantic along the east coast of Greenland and the northern Barents Sea (Fig. 1 B). The transport of Arctic water gives rise to a band of ice-covered areas from the Fram Strait, along the coast of Greenland towards Iceland (Fig. 1 A). The depth of winter mixing varies regionally, with maxima of several thousands of meters south off Iceland and in the central Norwegian Sea where North Atlantic Deep and Bottom Water are formed. Extensive fronts between Arctic and Atlantic water exist (Fig. 1 B). For overviews on the physical oceanography in this area, see JOHANNESSEN (1986). Further, there is a large latitudinal gradient of incident light. Light available for primary production is further limited in the Nordic Seas due to a seasonally variable ice-cover in the west, north and north-east (Fig. 1 A).
In contrast to the circular flow of the North Pacific gyre, warm Atlantic water is transported northward beyond 80"N, whereas both cold Arctic surface water and deep bottom water are exported southwards into lower latitudes (Fig. 1 B). While the North Pacific could be characterised as a more closed and circular system, the North Atlantic is an open, asymmetric system, with horizontal advection being very important for structuring pelagic systems and, hence, also for pelagic-benthic coupling.
Pelagic-benthic coupling in fjords: multi-pulse sys- tems
Compared to the much better investigated West Norwegian fjords (see WASSMANN, 1991), North Norwegian fjords are characterised by weak or no stratification during winter and spring, deep or lacking sills, highly seasonal freshwater run-off, and a less significant influence of the Norwegian Coastal Current. Important sources of seasonal and annual variability with regard to the pelagic-benthic coupling are water column stability and turbulence as well as advection. A third source of variability, zooplankton biomass and composition, will be discussed later.
It is widely accepted that the seasonal development of a pycnocline, in combina- tion with increasing solar radiation in spring, is a prerequisite for vernal spring
450 WASSMANN, ANDREASSW, REIGSTAD & SLAGSTAD
80°
7 0'
6d A
Fig. 1. A. Topography of the Nordic Seas (200 and lo00 m contours). Also shown is the sea ice cover: average minimum and maximum in May (dashed line) and August (solid line) (Redrawn from WADHAMS, 1986). Sediment trap mooring sites referred to in the text (VP: Voring Plateau; NA-1: Aegir Ridge-1; NB-1: Jan Mayen; LB-1: Lofoten Basin; BI-1: Bear Island; FS-1: Fram Strait; GB-2: Greenland Basin) are indicated. B. Surface currents in the Northeast Atlantic. 1: Norwegian Coastal Current; 2: Norwegian Atlantic Current; 3: North Cape Current; 4: Murmansk Current; 5: Kanin Current; 6: East Spitsbergen Current; 7: West Spitsbergen Current; 8: East Greenland Current; 9: East Icelandic Current; 1 0 Irminger Current; 11: Jan Mayen Current. Fronts in the Northeast Atlantic. 12: Norwegian Coastal Front; 13: Barents Sea Polar Front; 1 4 Polar Ocean Front; 15: East Greenland Front; 16: Iceland Gap Front. Shaded: mixing of Arctic and Atlantic water.

Pelagic-benthic coupling in the Nordic Seas and episodic events 45 1
BOO
7 0'
blooms (SVERDRUP, 1953; SMETACEK & PASSOW, 1990). However, the spring bloom in North Norwegian fjords takes place in almost non-stratified waters (EL LERTSEN, 1993). This bas also been observed in other boreal shelf and open ocean environments (TOWNSEND et al., 1992), suggesting that the phenomenon is the rule rather than an exception in many boreal and polar environments. Deep penetration of light and the absence of wind-driven vertical mixing appear to support cell growth that overcomes the vertical excursion rates of the neutrally stable water column, giving rise to a bloom (TOWNSEND et al., 1992). Wind strength and direction during early spring, along with the light regime, play a crucial role for the onset, development, and fate of the spring bloom in North Norwegian fjords. Strong, topographically enforced winds of short duration (mostly down-fjord), frequently observed during spring in the innermost fjords, can mix dividing cells

452 WASSMANN, ANDREASSEN. B I G S T A D & SLAGSTAD
below the compensation depth and may delay spring bloom development. Also, phytoplankton is easily transported out of the fjords along with wind-induced surface currents. This can give rise to a sequence of minor, "unsuccessful" bloom events, spreading the spring bloom development over a longer time. A major pycnocline does not develop before the freshwater run-off starts. This usually takes place in late Maylearly June, but the interannual variation in time as well as run-off pattern is considerable. For example, mild winters and rain during early spring influence the onset of the spring bloom by increasing vertical stability, giving rise to blooms that develop earlier and exhaust nutrients more rapidly compared with a "normal" year.
Information from boreal fjordic environments in western Norway revealed that advection plays an important role for the carbon flux of fjords (MATTHEWS & HEIMDAL, 1980; LEWIS & THOMAS, 1986; LINDAHL, 1987; WASSMANN, 1991). Evidence for advection of phytoplankton over long distances within fjords and between fjords and the shelf has also been provided by several investigations from western Norway (e.g., BRAARUD et al., 1958). Unfortunately, no such information is available for the coastal environments of northern Norway. Variability in the general wind regime along the coast is probably the main forcing for the renewal of the intermediate layer (SVENDSEN, 1981,1991). Advection of nutrients may have large implications for the new production in open fjords. Advection of meso-zoo- plankton can exceed local growth on an annual basis, and primary production and especially zooplankton production can vary considerably between years due to hydrographic variability (MAITHEWS & HEIMDAL, 1980; LINDAHL, 1987; AKSNES et al., 1989). An "ecology of advection" (MATTHEWS, 1986) obviously plays an important, if not the key role for the production in open fjords and on the shelf. Hence, pelagic-benthic coupling is determined here by stochastic processes, and characterised by strong seasonal and interannual variability.
Fig. 2 A illustrates this for the subarctic fjord Balsfjord, close to the city of Tromsa in northern Norway. The first sedimentation maximum reflects a vernal phytoplankton bloom based on winter-accumulated nutrients (I 1). The first maxi- mum comes to an end during an exchange of high salinity water with less dense water (I 2). This water probably introduced new nutrients from the coastal area outside Balsfjord and induced a vernal bloom. This bloom gave rise to a second maximum in vertical flux of POC. Finally, a third water mass with increased salinity and probably depleted of nutrients came into the fjord (I 3). Sedimentation of POC following the last exchange of water decreased and was low for the rest of the period.
The hydrographic situation in 1982 was different (Fig. 2 B). CTD profiles revealed no exchange of water masses. Salinity decreased throughout the period March to June. Sedimentation of POC was generally lower than 100 mg + m-2 . d-1, with a slight increase from mid-April to midJune. These two scenarios suggest that vertical flux of POC in Balsfjord is subject to considerable interannual variation and that the exchange of water masses during spring increases the export of organic
'matter to the sediment. Depending on the frequency and duration of water ex- change, several maxima of POC sedimentation may take place. If no exchange takes place, grazing and remineralisation in the water column seems to be sufficient to retard the loss of suspended biomass by sinking. The timing of events like water

Pelagic-benthic coupling in the Nordic Seas and episodic events 453
A I1 12 13 - w I 6004 r o - c
30
60 - E
tl
+ 500
400
300
N
90 2 120 4
I
H 2oo 100 150
180
- March+ April __t_ May - June -I
0
30
60
300 90
B
N
- March + April - May ___c_ June
salinity isopleth 0 mg POC m-2.d-1
Fig. 2. Sedimentation of particulate organic carbon (POC) at 60 and 50 m depth (columns, mg . C . m-2 . d-I) in Balsfiord, northern Norway in 1992 and 1982 (A and B, respectively). Superimposed areisolines of salinity (33.5 PSU in 1992 and 33.0 PSU in 1982), illustrating the inflow (A) and presence (B) of saline water. Data from REIGSTAD (1994) and L u m et al. (1989). respectively.
exchange also determines if a bloom is exported to the shelf and does not sink
For the time being, no precise information is available as to what extent North Norwegian fjords are systems of net export or import of nutrients and biomass. However, large seasonal and interannual variation in the pelagic-benthic coupling must be assumed since climatic forcing plays an important role. North Norwegian fjords are therefore multi-pulse systems where several phytoplankton blooms can take place and either settle to the bottom or be exported, depending on seasonal and annual variability in the physical oceanography.
locally.
Pelagic-benthic coupling in fjords and on the shelf: advection, retention, sources, and sinks
Primary production, suspended biomass, and sedimentation estimates are strongly influenced by advection of water masses on time scales down to hours and days. Advection can exchange water masses with variable phytoplankton assemblages as well as quantities and qualities of settling organic matter. This results in large
WASSMANN, ANDREASSEN, REIGSTAD & SLAGSTAD 454
February March
-
0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 ~ x-poaition (in 100km) x-powition (in 10Okm)
horizontal distances between particle formation and deposition at the sediment surface. For example, particles such as diatoms and faecal pellets, which continu- ously sink from the sea surface at a speed of 10-60 m . d-l and are exposed to a low, unidirectional average horizontal current speed of 5 cm 3 s-l, will be transported between about 4-21 km before they reach a sediment trap or the sediment surface at 50 m depth. During summer, when settling rates of organic matter are usually much lower (typically 1 m . d-1 or less), and under the same conditions, most particles will derive from areas several hundred km away from the deposition area. What are the implications for pelagic-benthic coupling?
Due to the considerable differences in sinking rates and horizontal transport, only two categories of particles will settle inside North Norwegian fjords, which are up to about 100 km long. Fast-sinking phytoplankton cells, most probably aggregates, entering the aphotic zone derive from surface areas which are only a few km or less away from the sampling site. Particles such as meso-zooplankton faecal pellets will also sediment within a fjord. When settling rates are much lower, most particles sampled by the trap will originate from areas tens to several hundred km away from the sampling site, i.e., they derive from outside the investigation area. This means that during episodes of large-scale advection, common in Norwegian fjords under the influence of northerly winds along the coast (SVENDSEN 1981, 1991), the majority of vertically sinking particles derive from areas outside the fjords (REIG- STAD, 1994). Even during summer, despite of the usually calm hydrographic con- ditions, most of the particles, except for faecal pellets, derive from elsewhere.
Investigating advective movements of particles in fjords is difficult (and often purely speculative) since the physical oceanography is often poorly understood and concomitant current measurements are usually not available. However, 3-D models describing the current field on the North Norwegian shelf exist and represent an

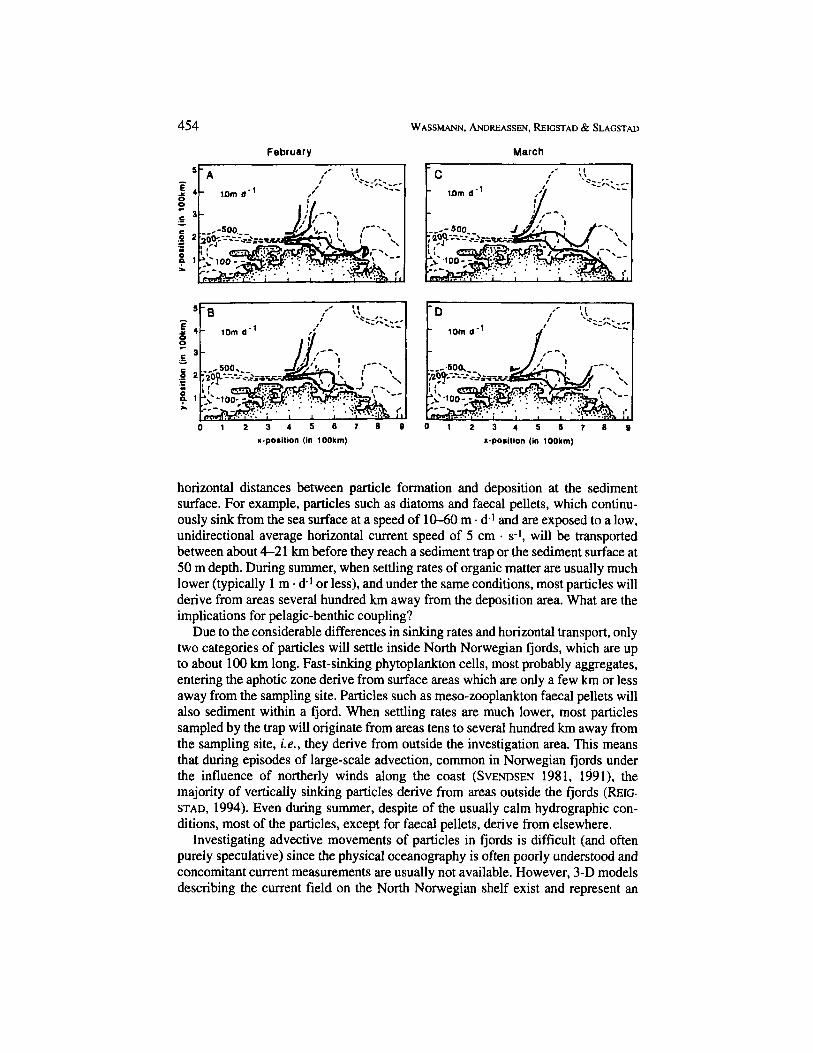
Pelagic-benthic coupling in the Nordic Seas and episodic events 455 Fig. 3 . Drift patterns of inert particles released at 100 and 200 m depth on the shelf and 500 and 1,ooO m depth off the shelf break off the fjord Ma- langen, close to the city of Trorns~, during February, March, and April. Topography is also shown (stip- pled lines: 100, 200 and 500 m equidistance). The particles were sinking at 1 m (upper panel) and 10 m . d-I (lower panel). Drift patterns were determined in a simulated current field in the open North Norwe- gian coastal zone based on a numerical model with 6.6 km grid size. (D. SLAGSTAD, unpubl. res.).
April
E - -
0 1 2 3 4 5 6 7 8 9
x-posltion (in 100krn)
important tool for investigating large-scale advective transports (WASSMANN & SLAGSTAD, 1995). This model was used to simulate the drift patterns of sinking particles.
In one simulation experiment, inert particles sinking at 1 and 10 m . d-1 were discharged in the surface layer off the fjord Malangen close to Tromsa in an average year current field during the months February, March, and April (Fig. 3). One of the most important variables determining current direction and speed is the prevail- ing, large-scale wind field. The model was run with an average year wind field. The particles began to sink at the 100 and 200 m isobaths on the shelf and at the 500 and 1000 m isobaths off the shelf. The simulations were run separately for each month and the drift trajectories plotted (shorter periods if the particles did reach the sediment surface). Particles starting at the 100 and 200 m isobaths drifted along the North Norwegian shelf shallower than 300 m depth. The particles sinking at 1 m . d-1 drifted 300400 km per month. They often came close to the coastline, but were probably not deposited. The particles sinking at 10 m . d-1 drifted about 200- 300 km in February, 300-400 km in March, and about 200 km in April, indicating monthly differences in the current field. They sedimented either close to the coast- line in February or further off-shore in March and April, indicating that deposition sites might vary over time according to the current field.
None of the sinking particles discharged off the North Norwegian shelf break reached the bottom within one month (Fig. 3). The particles discharged closest to the shelf break drift along with the current for about 200 km per month, following the shelf break close to the 500 m isobath. Only initially do the particles discharged further off the shelf break drift similarly to those in February: in March and April, however, they accumulated in a retention area. This indicates that the drift pattern of sinking particles in the western Norwegian Sea can be dramatically different over
456 WASSMA", h R E A S S E N , RElGSTAD & SLAGSTAD
. A I \ - temperature anomailee
1.6- Ice Index - - -
1.0-
0.5-
0 - 0- a
0.5-
1 .o-
1.6-
-20
-15
-10
- 5
P - 0 - -75
- - lo
.-15
-20
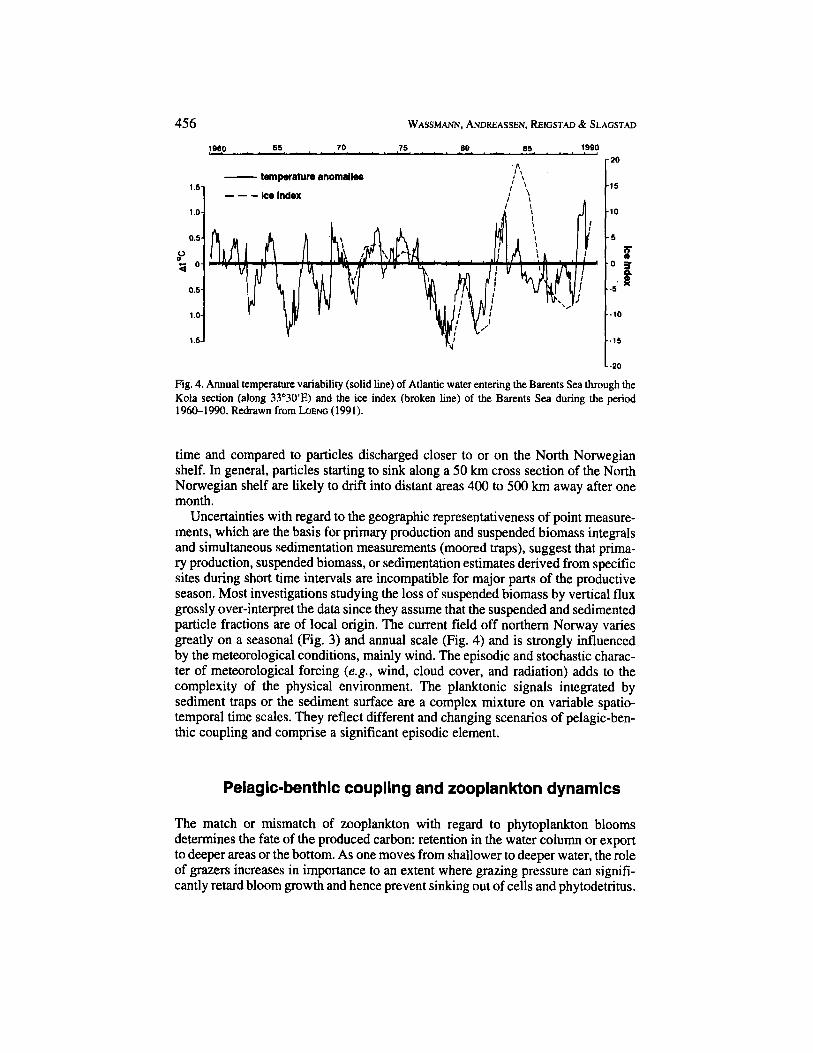
Fig. 4. Annual temperature variability (solid line) of Atlantic water entering the Barents Sea through the Kola section (along 33'30'E) and the ice index (broken line) of the Barents Sea during the period 1960-1990. Redrawn from h E N G (1991).
time and compared to particles discharged closer to or on the North Norwegian shelf. In general, particles starting to sink along a 50 km cross section of the North Norwegian shelf are likely to drift into distant areas 400 to 500 km away after one month.
Uncertainties with regard to the geographic representativeness of point measure- ments, which are the basis for primary production and suspended biomass integrals and simultaneous sedimentation measurements (moored traps), suggest that prima- ry production, suspended biomass, or sedimentation estimates derived from specific sites during short time intervals are incompatible for major parts of the productive season. Most investigations studying the loss of suspended biomass by vertical flux grossly over-interpret the data since they assume that the suspended and sedimented particle fractions are of local origin. The current field off northern Norway varies greatly on a seasonal (Fig. 3) and annual scale (Fig. 4) and is strongly influenced by the meteorological conditions, mainly wind. The episodic and stochastic charac- ter of meteorological forcing (e.g., wind, cloud cover, and radiation) adds to the complexity of the physical environment. The planktonic signals integrated by sediment traps or the sediment surface are a complex mixture on variable spatio- temporal time scales. They reflect different and changing scenarios of pelagic-ben- thic coupling and comprise a significant episodic element.
Pelagic-benthic coupling and zooplankton dynamics
The match or mismatch of zooplankton with regard to phytoplankton blooms determines the fate of the produced carbon: retention in the water column or export to deeper areas or the bottom. As one moves from shallower to deeper water, the role of grazers increases in importance to an extent where grazing pressure can signifi- cantly retard bloom growth and hence prevent sinking out of cells and phytodetritus.

Pelagic-benthic coupling in the Nordic Seas and episodic events 457
Whether sedimentation occurs subsequent to heavy grazing in the form of herbivore faeces or whether it is retarded altogether depends on the type of grazer dominating this trophic level. The pelagic-benthic coupling is mainly influenced by zooplankton by (a) advection of adult populations into and out of certain areas, (b) overwintering, and (c) the taxonomic composition of the zooplankton community.
a. Advection of zooplankton
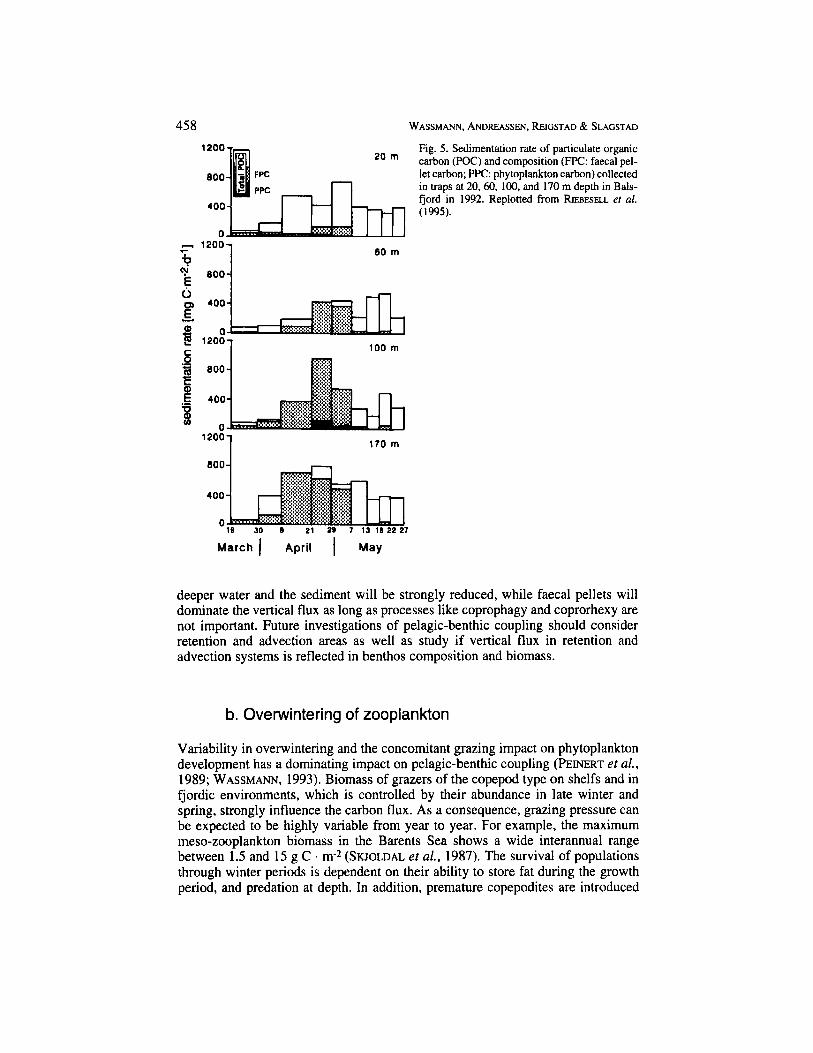
Rapid and extensive exchange of water masses between fjords and shelf waters are the rule rather than the exception in the coastal zone adjacent to the Nordic Seas. Not only nutrients, phytoplankton, and detritus follow along with the exchanged water masses, but also zooplankton populations. Meso- and macro-zooplankton advected into a fjordic environment may stay in the area as long as they keep close to the bottom and avoid being flushed out by intermediate or surface water exchange. However, when bottom water is exchanged, the zooplankton also follows. In 1992 several episodes of bottom water exchange were recorded in Balsfjord (Fig. 2), for details, see REICSTAD (1994). One of these events took place from about April 12 to May 10, when water masses with increased salinity and temperature entered and disappeared again from Balsfjord. The water exchange was clearly evident in the phytoplankton biomass, but a new zooplankton community, dominated by krill, was also probably flushed into and out of the fjord. Krill is a species which is easily advected into fjords (AKSNES ef al., 1989). The krill community produced a strong sedimentation signal in form of large, rapidly sinking faecal pellets that were caught in the traps (Fig. 5). The maximum in POC sedimentation during spring was strongly dominated by krill faeces, in particular at depths were krill is usually present (RIEBESELL et al., 1995). The advection of two new water masses terminated the dominance of krill faeces in the vertical flux after May 10 (Fig. 5). Episodically advected populations of meso- and macro-zooplankton obviously influence the pelagic-benthic coupling in fjords and probably other areas (WASSMANN etal., 1991). The impact of larger zooplankters on vertical flux adds to the interannual variability of pelagic-benthic coupling in coastal environments.
The temporal variation of the influx of Atlantic water into the Barents Sea (Fig. 4) implies that variable amounts of nutrients, phytoplankton, and detritus are transported along with the water into the southern and central Barents Sea. Depend- ing on its vertical position in the water column, this is also true for zooplankton. The advected zooplankton populations add to the local overwintering populations in the Barents Sea and significantly impact pelagic-benthic coupling due to variable grazing pressure (see below). Careful investigations of the Norwegian Sea and Barents Sea current fields revealed that retention areas with cyclonic and anti-cy- clonic circulation are found inside areas dominated by large-scale advection (G. PEDERSEN & D. SLAGSTAD, unpubl. res.). This implies that zooplankton accu- mulates in certain areas to densities significantly higher compared to the adjacent areas dominated by advecting populations. Preliminary investigations of meso-zoo- plankton distribution across advection and retention areas revealed that this is actually the case (G. PEDERSEN, pers. comm.). High zooplankton concentrations in retention areas will obviously affect pelagic-benthic coupling, probably increasing supply of organic matter to the sediment surface here. Phytoplankton export to
45 8 WASSMANN, ANDREASSEN, REIGSTAD & SLAGSTAD
Fig. 5. Sedimentation rate of particulate organic carbon (POC) and composition (FF'C: faecal pel- let carbon; PPC: phytoplankton carbon) collected in traps at 20.60, 100, and 170 m depth in Bals- fjord in 1992. Replotted from RIEBESELL er al. (1995).
2o
i ro m 12001
'18 30 B 21 29 7 13 1 8 2 2 27
March I April I May
deeper water and the sediment will be strongly reduced, while faecal pellets will dominate the vertical flux as long as processes like coprophagy and coprorhexy are not important. Future investigations of pelagic-benthic coupling should consider retention and advection areas as well as study if vertical flux in retention and advection systems is reflected in benthos composition and biomass.
b. Overwintering of zooplankton
Variability in overwintering and the concomitant grazing impact on phytoplankton development has a dominating impact on pelagic-benthic coupling (PEINERT et al., 1989; WASSMANN, 1993). Biomass of grazers of the copepod type on shelfs and in fjordic environments, which is controlled by their abundance in late winter and spring, strongly influence the carbon flux. As a consequence, grazing pressure can be expected to be highly variable from year to year. For example, the maximum meso-zooplankton biomass in the Barents Sea shows a wide interannual range between 1.5 and 15 g C . m-2 (SKJOLDAL et al., 1987). The survival of populations through winter periods is dependent on their ability to store fat during the growth period, and predation at depth. In addition, premature copepodites are introduced

Pelagic-benthic coupling in the Nordic Seas and episodic events 459
into the southern and central part of the Barents Sea by inflowing Atlantic water (see above). How will differences in the number of CV copepodites during spring influence pelagic-benthic coupling?
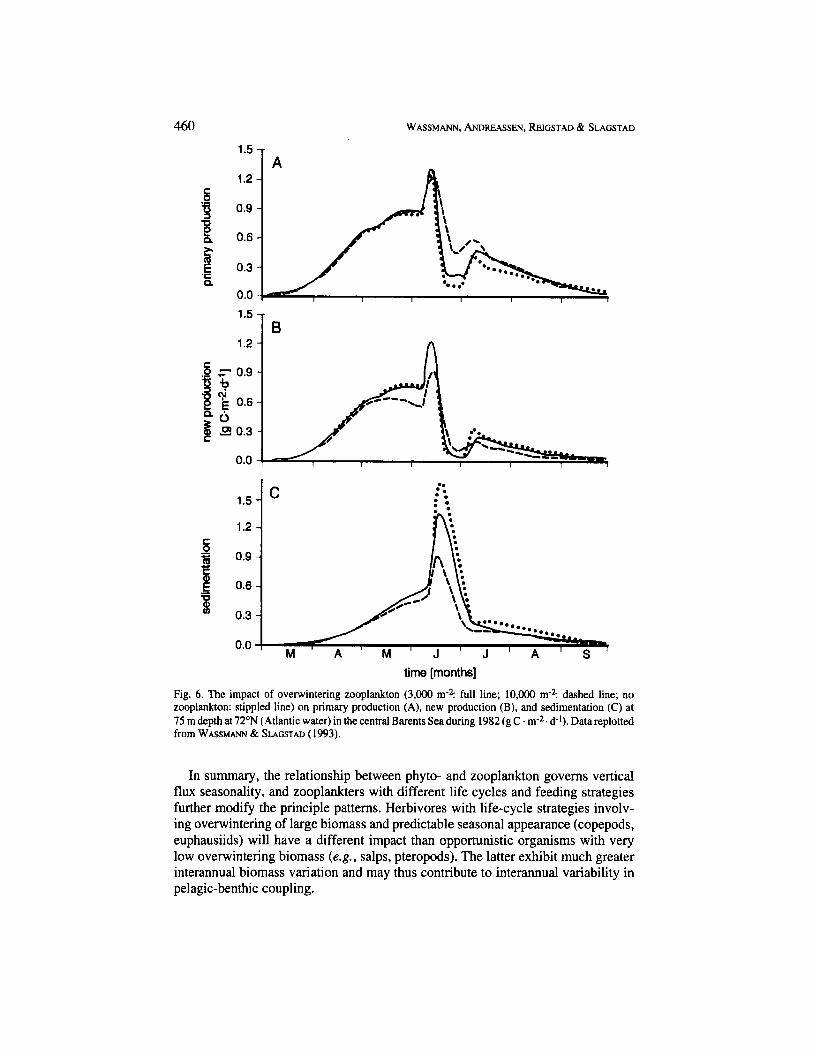
Fig. 6 is based on a simulation model (WASSMANN & SLAGSTAD, 1993) and shows how variable amounts of overwintering meso-zooplankton influence the time variation and magnitude of primary and export production in the southern Barents Sea. Due to increased remineralisation in the euphotic zone, increased numbers of meso-zooplankton increased primary production slightly from 76 to 85 g C . m2 . a-1. However, export production is significantly influenced by zooplankton grazing. 10,OOO individuals of overwintering meso-zooplankton per m* decreased sedimentation from 50 to 38 g C . m-2 . a'. Not only does the total export production decrease at higher grazing pressure, but the time variation of export production is also influenced by increased grazing as the amplitude of the daily sedimentation rate decreased (Fig. 6). The abundance of meso-zooplankton during spring has, thus, a pronounced influence on sedimentation, especially during late spring/early summer. The simulation model for the Barents Sea is very sensitive to changes in the number of overwintering and advected copepods, representing one of the most important variables in the Barents Sea ecosystem. Future investigations of pelagic-benthic coupling will have to focus specifically on the overwintering, advection, and grazing impact of zooplankton and the episodic nature of these processes.
c. Taxonomic composition of the zooplankton community
Field data (BATHMANN et al., 1987; PEINERT et al., 1987; NOJI, 1989) and laboratory experiments (LAMPITT et al., 1990) suggest that copepod grazing can counteract sedimentation by keeping matter in suspension. Being widespread and relatively evenly distributed horizontally, copepods cannot easily leave their feeding grounds. In this context, coprorhexy and coprophagy are important mechanisms (NOJI, 1991).
Grazing by large, roving crustaceans such as euphausiids produces faeces that sink too rapidly for easy recycling at their production depths. Thus they inherently deplete their own and others feeding grounds. Other herbivores such as pteropods and certain gelatinous plankters (salps, appendicularians) are less predictable in their appearance, but may be very abundant in some years and may significantly influence sedimentation (BATHMANN et aZ., 1991). They all feed efficiently and non-selectively on small particles, with salp faeces, for example, contributing significantly to the vertical flux (BATHMA", 1988; NOJI, 1989). Abandoned appen- dicularian houses (ALLDREDGE, 1986) may play a role in accelerating losses by scavenging small and slower-sinking particles. Years of very high salp abundance have been observed non-periodically in Norwegian waters (BRATTSTROM, 1972). This implies that flux patterns differ from normal years depending on the season in which salps appear: if large salp biomass appeared early in spring, most of the phytoplankton biomass could leave the upper layers due to rapidly sinking faeces (WASSMANN et al., 1991). This would leave copepods little opportunity to increase their population size. Such a scenario would result in a strong spring pulse in sedimentation.
460
t 'C i! 8 n
0.
WASSMANN, ANDREASSEN, REICSTAD & SLAGSTALI
1.5 -
1.2 -
0.9 -
A
'5 1 B 1.2
1.5
1.2 C 0 g 0.9 E
0.6
0.3
0.0
time [months] Fig. 6. The impact of overwintering zooplankton (3,000 m-? full line; 10,000 m2: dashed line; no zooplankton: stippled line) on primary production (A), new production (B), and sedimentation (C) at 75 m depth at 72"N (Atlantic water) in the central Barents Sea during 1982 (g C . m-2. d-1). Data replotted from WASSMANN & SLAGSTAD (1993).
In summary, the relationship between phyto- and zooplankton governs vertical flux seasonality, and zooplankters with different life cycles and feeding strategies further modify the principle patterns. Herbivores with life-cycle strategies involv- ing overwintering of large biomass and predictable seasonal appearance (copepods, euphausiids) will have a different impact than opportunistic organisms with very low overwintering biomass (e.g., salps, pteropods). The latter exhibit much greater interannual biomass variation and may thus contribute to interannual variability in pelagic-benthic coupling.

Pelagic-benthic coupling in the Nordic Seas and episodic events 46 1
Is the vertical flux of organic matter in the Norwegian Sea determined by zooplankton?
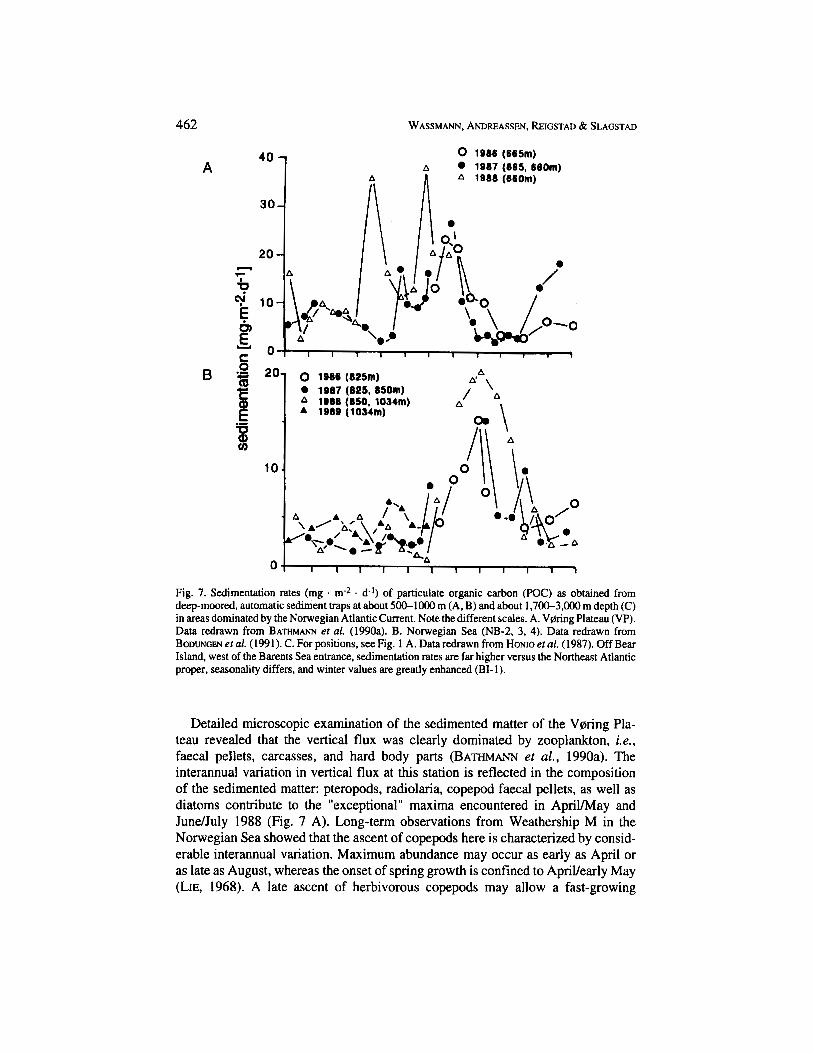
Extensive seasonal studies of the hydrography and the plankton dynamics in the Norwegian Sea are only available from Weathership M. However, seasonal investi- gations of sedimentation have been conducted with deep-moored automatic sedi- ment traps for several years all over the Norwegian Sea (Fig. 1 A). The evaluation of the processes influencing the pelagic-benthic coupling in the Norwegian Sea remains speculative since only one part of the necessary data is available. Further- more, comparing vertical fluxes from oceanic environments of the Nordic Seas with those of the fjords and shelf is difficult because the former are derived from trap deployments below 500 m: the fluxes here are greatly reduced due to remineralisa- tion of matter in the upper hundreds of meters. Some of the seasonal POC sedimen- tation rates from the Norwegian Sea are presented in Fig. 7.
The seasonal sedimentation of POC at about 660 m depth on the Vprring Plateau (VP) in the Norwegian Sea off the Norwegian Shelf at 67"N (Fig. 1 A) generally shows seasonal maxima during summer (June to August), with low sedimentation rates in spring and autumn (Fig. 7 A). Three years of trap deployments, however, showed significant interannual variations. The single peaks recorded in Novem- bermecember 1987 and spring and summer 1988 are unexplained. Despite the scatter in the data, the "regular" seasonal maximum of vertical flux clearly occurs on the Vprring Plateau in summer and is hence not related to the spring bloom, as is typical for shallow regions (e.g. , Fig. 2).
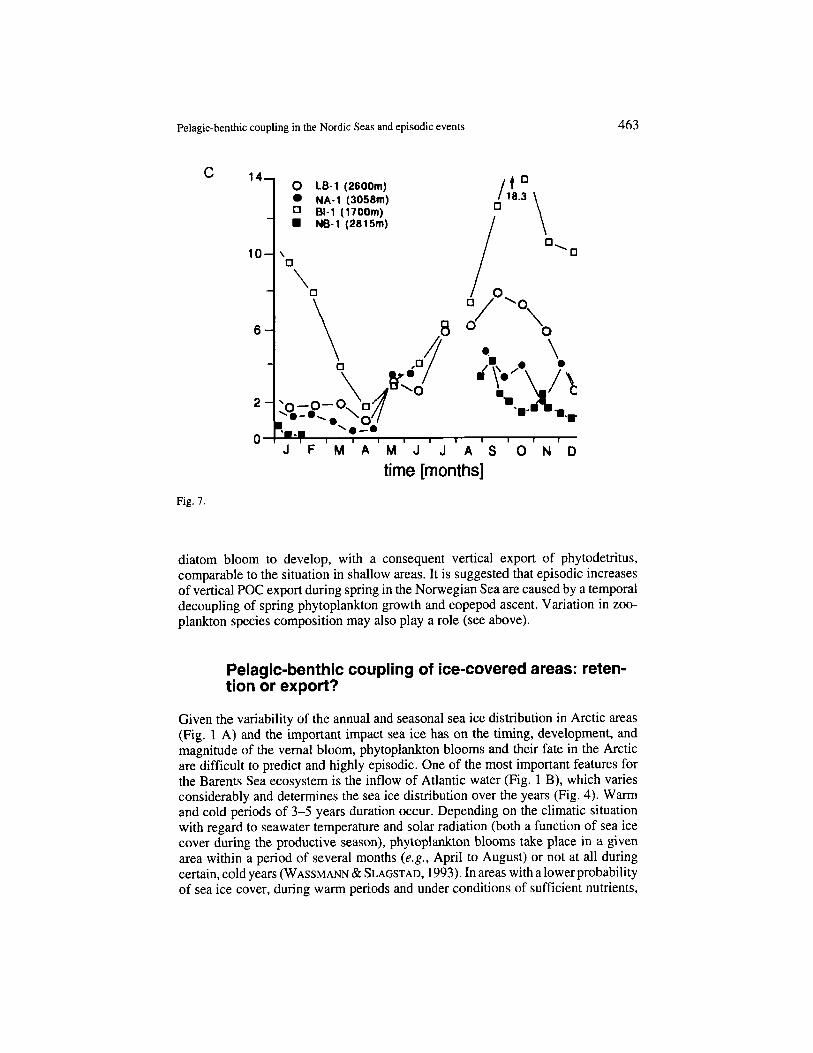
The same trend was also found in the Lofoten Basin in the central Norwegian Sea (LB-1) at 70"N (Fig. 1 B). The annual maximum was in late summer/early autumn (August-September) (Fig. 7 B). The considerable interannual variation encountered at the Vprring Plateau was not recorded here. The same trends were also recorded at depth as reflected by the data presented in Fig. 7 C, with the exception of the data off Bear Island, which will be commented upon later. POC sedimenta- tion maxima were recorded in late summer and autumn. What processes in the Norwegian Sea retard vertical flux of organic matter compared with the seasonal variation in new production, and what is the reason for interannual variability or the lack thereof?
As mentioned above, evaluations of the processes governing the vertical flux must be speculative. We suggest that variation in regeneration efficiency in the upper layers determines the seasonal delay and interannual variability of export production. Herbivorous copepods ascending from their overwintering depths in the open ocean can control phytoplankton production early in the growth season (BATH- MA" et al., 1990b). The large copepod faecal pellets are apparently prevented from sinking by processes such as coprophagy and coprorhexy LAM PI^ et al., 1990; No11 et al., 1991). These copepod activities effectively recycle matter in the upper layers, controlling the vertical flux (PEINERT et al., 1989; WASSMANN, 1993). An export of phytoplankton and phytodetritus from the upper layers during spring and early summer, as observed in shallow areas and fjords, therefore does not take place. When the recycling activities of the zooplankton population cease in late summedearly autumn and hibernation starts, the remaining detritus from the annual new production sinks, increasing the vertical flux of POC. This general pattern is valid for the entire Norwegian Sea (Fig. 7).
WASSMA", ANDREASSEN, k3GSTAD & SLAGSTAD 462
A
B
Fig. 7. Sedimentation rates (mg m-* . d-1) of particulate organic carbon (POC) as obtained from deep-moored, automatic sediment traps at about 500-1000 m (A, B) and about 1.700-3.000 m depth (C) in areas dominated by the Norwegian Atlantic Current. Note the different scales. A. V0ring Plateau (VP). Data redrawn from BATHMA" et al. (1990a). B. Norwegian Sea (NB-2, 3, 4). Data redrawn from BODUNGEN er al. (1991). C . For positions. see Fig. 1 A. Data redrawn from Ho~lo el al. (1987). Off Bear Island, west of the Barents Sea entrance, sedimentation rates are far higher versus the Northeast Atlantic proper, seasonality differs, and winter values are greatly enhanced (BI- 1).
Detailed microscopic examination of the sedimented matter of the Voring Pla- teau revealed that the vertical flux was clearly dominated by zooplankton, i.e., faecal pellets, carcasses, and hard body parts (BATHMANN et al., 1990a). The interannual variation in vertical flux at this station is reflected in the composition of the sedimented matter: pteropods, radiolaria, copepod faecal pellets, as well as diatoms contribute to the "exceptional" maxima encountered in ApriVMay and JundJuly 1988 (Fig. 7 A). Long-term observations from Weathership M in the Norwegian Sea showed that the ascent of copepods here is characterized by consid- erable interannual variation. Maximum abundance may occur as early as April or as late as August, whereas the onset of spring growth is confined to ApriVearly May (LIE, 1968). A late ascent of herbivorous copepods may allow a fast-growing
Pelagic-benthic coupling in the Nordic Seas and episodic events 463
C t o
0 \ 0 LB-1 (2600m)
0 81-1 (1700m) I NB-1 (2815m)
NA-1 (3058m)
I
14- 0 LB-1 (2600m)
- 0 81-1 (1700m) NB-1 (2815m)
0 NA-1 (3058m)
10- \"
6 -
\
1 1 1 I I I
J F M A M J J A S O N D time [months]
Fig. 7.
diatom bloom to develop, with a consequent vertical export of phytodetritus, comparable to the situation in shallow areas. It is suggested that episodic increases of vertical POC export during spring in the Norwegian Sea are caused by a temporal decoupling of spring phytoplankton growth and copepod ascent. Variation in zoo- plankton species composition may also play a role (see above).
Pelagic-benthic coupling of ice-covered areas: reten- tion or export?
Given the variability of the annual and seasonal sea ice distribution in Arctic areas (Fig. 1 A) and the important impact sea ice has on the timing, development, and magnitude of the vernal bloom, phytoplankton blooms and their fate in the Arctic are difficult to predict and highly episodic. One of the most important features for the Barents Sea ecosystem is the inflow of Atlantic water (Fig. 1 B), which varies considerably and determines the sea ice distribution over the years (Fig. 4). Warm and cold periods of 3-5 years duration occur. Depending on the climatic situation with regard to seawater temperature and solar radiation (both a function of sea ice cover during the productive season), phytoplankton blooms take place in a given area within a period of several months (e.g., April to August) or not at all during certain, cold years (WASSMANN & SLAGSTAD, 1993). In areas with a lower probability of sea ice cover, during warm periods and under conditions of sufficient nutrients,
464 WASSMA", ANDREASSEN, REIGSTAD & SLAGSTAD
phytoplankton blooms will take place annually during specific periods. However, the carbon flux in stratified, marginal ice zones are generally dominated by extensive, short-term blooms, aggregation, and sedimentation.
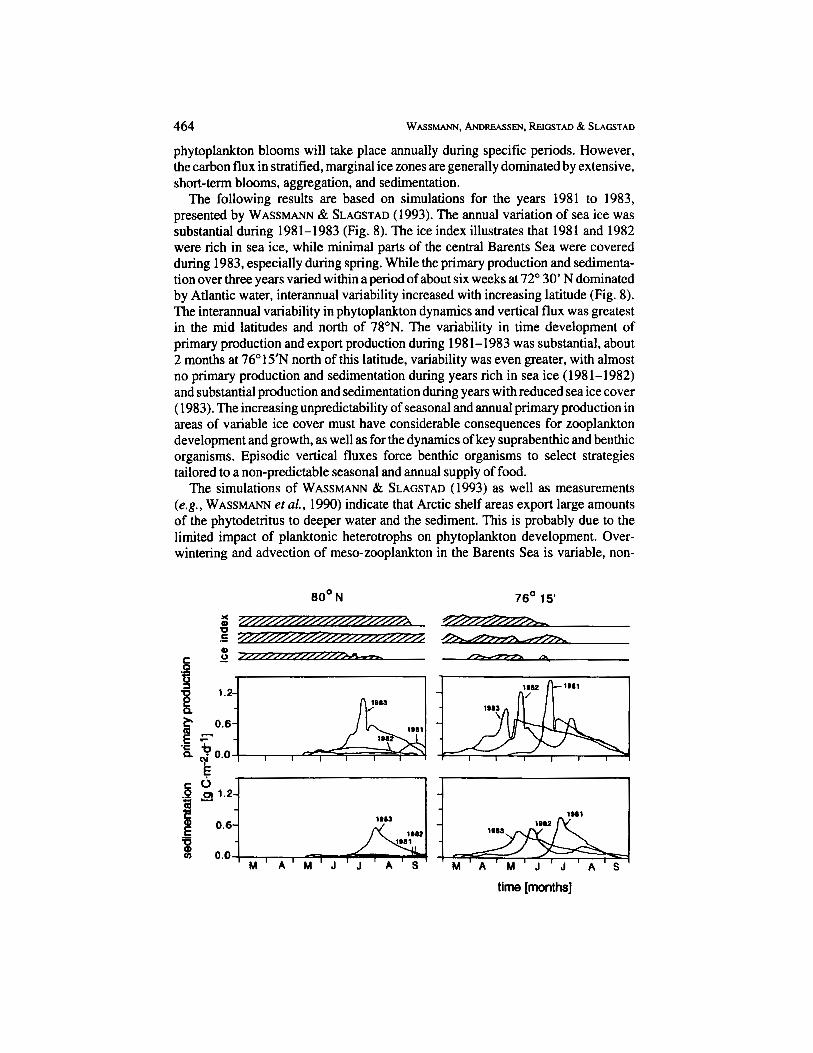
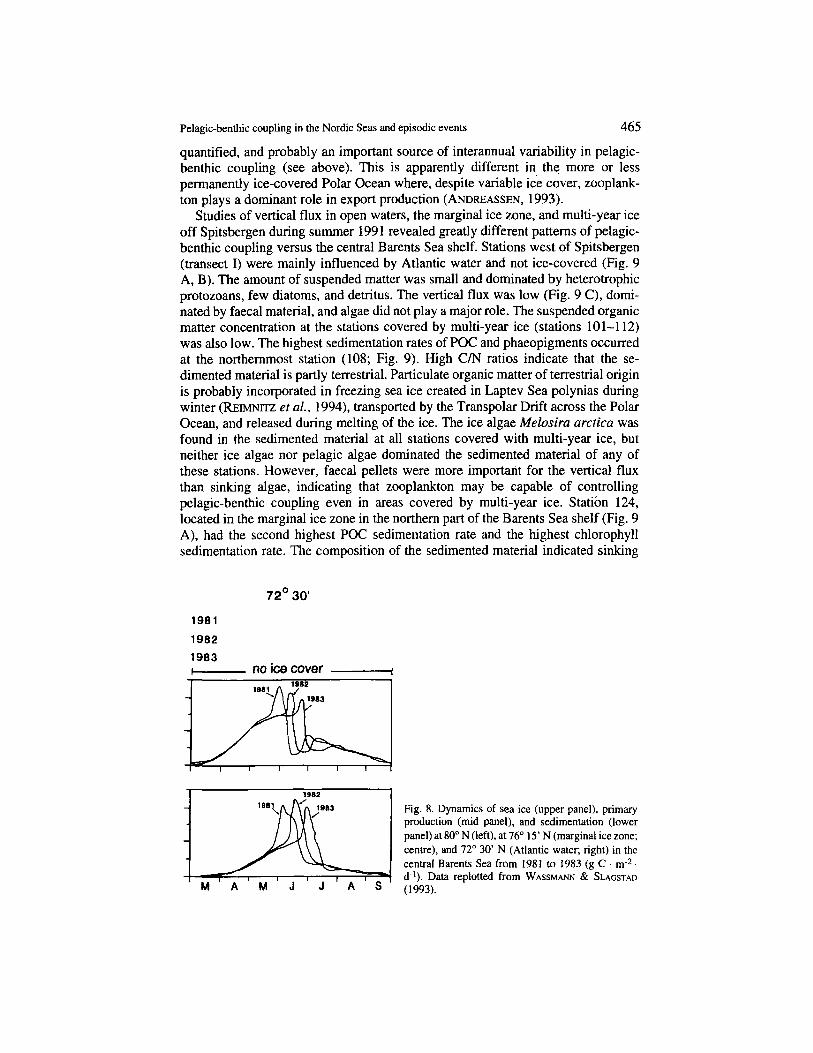
The following results are based on simulations for the years 1981 to 1983, presented by WASSMA" & SLAGSTAD (1993). The annual variation of sea ice was substantial during 1981-1983 (Fig. 8). The ice index illustrates that 1981 and 1982 were rich in sea ice. while minimal parts of the central Barents Sea were covered during 1983, especially during spring. While the primary production and sedimenta- tion over three years varied within a period of about six weeks at 72" 30' N dominated by Atlantic water, interannual variability increased with increasing latitude (Fig. 8). The interannual variability in phytoplankton dynamics and vertical flux was greatest in the mid latitudes and north of 78"N. The variability in time development of primary production and export production during 1981-1983 was substantial, about 2 months at 76" 15" north of this latitude, variability was even greater, with almost no primary production and sedimentation during years rich in sea ice (1981-1982) and substantial production and sedimentation during years with reduced sea ice cover ( 1983). The increasing unpredictability of seasonal and annual primary production in areas of variable ice cover must have considerable consequences for zooplankton development and growth, as well as for the dynamics of key suprabenthic and benthic organisms. Episodic vertical fluxes force benthic organisms to select strategies tailored to a non-predictable seasonal and annual supply of food.
The simulations of WASSMANN & SLAGSTAD (1993) as well as measurements (e.g., WASSMANN et al., 1990) indicate that Arctic shelf areas export large amounts of the phytodetritus to deeper water and the sediment. This is probably due to the limited impact of planktonic heterotrophs on phytoplankton development. Over- wintering and advection of meso-zooplankton in the Barents Sea is variable, non-
la83
A S
time [months]
Pelagic-benthic coupling in the Nordic Seas and episodic events 465
quantified, and probably an important source of interannual variability in pelagic- benthic coupling (see above). This is apparently different in the more or less permanently ice-covered Polar Ocean where, despite variable ice cover, zooplank- ton plays a dominant role in export production (ANDREASSEN, 1993).
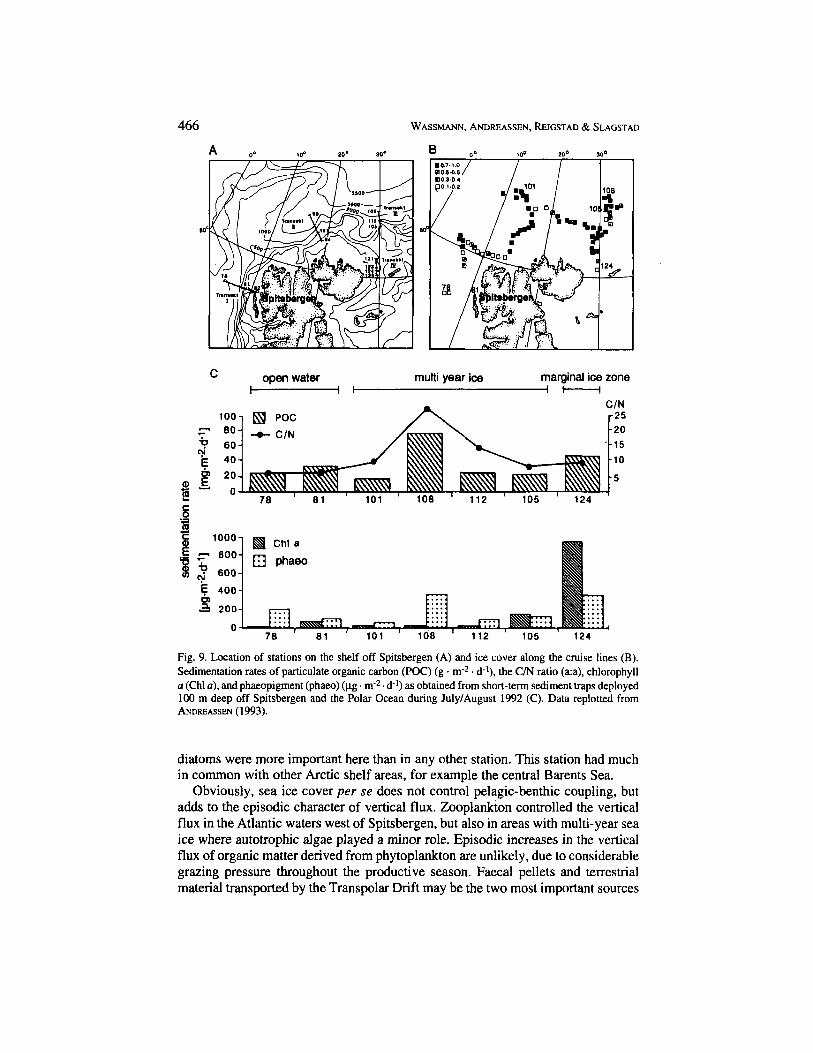
Studies of vertical flux in open waters, the marginal ice zone, and multi-year ice off Spitsbergen during summer 1991 revealed greatly different patterns of pelagic- benthic coupling versus the central Barents Sea shelf. Stations west of Spitsbergen (transect I) were mainly influenced by Atlantic water and not ice-covered (Fig. 9 A, B). The amount of suspended matter was small and dominated by heterotrophic protozoans, few diatoms, and detritus. The vertical flux was low (Fig. 9 C), domi- nated by faecal material, and algae did not play a major role. The suspended organic matter concentration at the stations covered by multi-year ice (stations 101-1 12) was also low. The highest sedimentation rates of POC and phaeopigments occurred at the northernmost station (108; Fig. 9). High C/N ratios indicate that the se- dimented material is partly terrestrial. Particulate organic matter of terrestrial origin is probably incorporated in freezing sea ice created in Laptev Sea polynias during winter (REIMNITZ et al., 1994), transported by the Transpolar Drift across the Polar Ocean, and released during melting of the ice. The ice algae Melosiru urctica was found in the sedimented material at all stations covered with multi-year ice, but neither ice algae nor pelagic algae dominated the sedimented material of any of these stations. However, faecal pellets were more important for the vertical flux than sinking algae, indicating that zooplankton may be capable of controlling pelagic-benthic coupling even in areas covered by multi-year ice. Station 124, located in the marginal ice zone in the northern part of the Barents Sea shelf (Fig. 9 A), had the second highest POC sedimentation rate and the highest chlorophyll sedimentation rate. The composition of the sedimented material indicated sinking
72' 30'
1981
1982
1983 1- no ice cover
Fig. 8. Dynamics of sea ice (upper panel), primary production (mid panel), and sedimentation (lower panel) at 80' N (left), at 76" 15' N (marginal ice zone; centre), and 72' 30' N (Atlantic water; right) in the central Barents Sea from 1981 to 1983 (g C . m2 d-l). Data replotted from WASSMA" & SLAGSTAD (1993).
466 WASSMANN, ANDREASSEN, REIGSTAD & SLAGSTAD
loD 20' 30'
m07.1.0 B0.5.0.0 I
8C
C open water multi year ice marginal ice zone - I I W
" 78 ' 81 ' 101 ' 108 ' 112 ' 105 ' 124
Fig. 9. Location of stations on the shelf off Spitsbergen (A) and ice cover along the cruise lines (B). Sedimentation rates of particulate organic carbon (POC) (g . m-2. d-*), the C/N ratio (a:a), chlorophyll u (Chl a), and phaeopigment (phaeo) (pg . m-2. d-I) as obtained from short-term sediment traps deployed 100 m deep off Spitsbergen and the Polar Ocean during July/August 1992 (C). Data replotted from ANDREASSEN (1993).
diatoms were more important here than in any other station. This station had much in common with other Arctic shelf areas, for example the central Barents Sea.
Obviously, sea ice cover per se does not control pelagic-benthic coupling, but adds to the episodic character of vertical flux. Zooplankton controlled the vertical flux in the Atlantic waters west of Spitsbergen, but also in areas with multi-year sea ice where autotrophic algae played a minor role. Episodic increases in the vertical flux of organic matter derived from phytoplankton are unlikely, due to considerable grazing pressure throughout the productive season. Faecal pellets and terrestrial material transported by the Transpolar Drift may be the two most important sources

Pelagic-benthic coupling in the Nordic Seas and episodic events 467
Fig. 10. Possible pathways of advective transport of tine sediment surface particles along with dense bottom water ("winter outbursts") from the Barents Sea shelf into the Nonvegian Sea and the Polar Ocean.
for sedimenting material in the Polar Ocean; the latter probably variable, depending on incorporation into the ice, ice drift, and melting. In the marginal ice zone of Arctic shelfs, however, zooplankton is unable to control the bloom, and ungrazed algae dominate the vertical flux, probably due to reduced overwintering.
Particle transport from the Barents Sea shelf
The general picture encountered in shallow coastal waters with a strong physical impact at the bed from waves, currents, and fishery is that the primary sedimentation flux - measured with a sediment trap near the bed - can be obscured by a much larger gross flux of resuspended particles caught in the process of redeposition (FLODERUS & PHIL, 1990). This difficulty must be kept in mind when considering pelagic-ben- thic coupling. We need more information about resuspension in order to estimate both sedimentation and horizontal transport. Currently, no easy and reliable tech- niques are available for this.

468 WASSMANN, ANDREASSEN, REIGSTAD 8t SLAGSTAD
Along the slopes of coastal areas and the shelf break, non-quantified amounts of organic matter, ultimately derived from primary production, are transported in the benthic nepheloid layer to the sediment surface and benthos of basins and the deep sea (GRAF, 1989). This implies local decoupling of pelagic and benthic production on seasonal time scales. This will be demonstrated by an example from the Barents Sea shelf.
Dense bottom water accumulating in the deeper parts of the Barents Sea can give rise to pulses of cold water which flow through the various troughs into the Arctic Ocean and the Norwegian Sea (QUADFASEL et al., 1988). The formation of this water results from winter cooling of surface water and is thus a function of climatic variability (MIDTTUN & LOENG, 1987; JAENG, 1991). The down-slope flow of dense bottom water at the steep shelf break between the Barents and Norwegian Seas may trigger a large-scale bottom flow over a wider shelf area. This flow, in turn, may flush out newly deposited, unconsolidated sediment from the shelf and troughs, causing a "winter outburst" transport of particulate matter (HONJO, 1990). These "outbursts" are reflected by high winter sedimentation rates of particulate carbon in the Northeastern Norwegian Sea off the Barents Sea shelf (BI) (HONJO ef al., 1988) which are far higher than for the central Norwegian Sea (HONJO, 1990; BODUNGEN et al., 1991) (see BI-1 in Fig. 7 C).
The example implies that the Barents Sea supplies the deeper parts of the Norwegian Sea with material, giving rise to additional food for the benthos, and signals in the sediment record. This is probably also true for the Arctic Ocean (Fig. 10). Due to the extensive transport of water masses to and from the highly productive Barents Sea shelf, not all of the annually produced organic matter is mineralised in the area of production. Some of the particulate matter will be advected from the shelf through the Bear Island, Victoria, and St. Anna troughs, in particular during periods of bottom water exchange. Once again, pelagic-benthic coupling is different in time and space, depending on the season, year, and area.
Conclusion
The pelagic-benthic coupling is ultimately governed by hydrodynamic processes. In contrast to the subarctic North Pacific, the Northeast Atlantic can be viewed as a huge advective flow-through system. Long-term changes in the location and strength of major current systems influence the North Atlantic as a whole (PARSONS & LALLI, 1988). TAYLOR (1978) and COLEBROOK (1985) observed that zooplankton is less abundant when the influence of the North Atlantic Current on the Northeast Atlantic is weak. Similar variability has also been observed in the Barents Sea (SKJOLDAL et al., 1987). The reasons for this variability remain poorly understood, but we speculate that zooplankton production and pelagic-benthic coupling in the Nordic Seas may change significantly on decadal scales as well.
We suggest that the Nordic Seas are strongly controlled by episodic events on various time scales. Future oceanographic studies in the Nordic Seas should care- fully consider the episodic nature of these ecosystems. Since the Nordic Seas experience large climatic and biological variations, no "normal" pattern of pelagic- benthic coupling can be derived from a few annual studies, let alone from sporadic investigations.

Pelagic-benthic coupling in the Nordic Seas and episodic events 469
Acknowledgements
The investigation was supported by the Norwegian Fisheries Research Council (NFFR) and is a contribution from the research programme North Norwegian Coastal Ecology (MARE NOR).
References
AKSNES, D. L., J. AURE, S. KAARTVEDT, T. MAGNESEN & J. RICHARD, 1989: Significance of advection for the carrying capacities of fjord populations. Mar. Ecol. Prog. Ser., 50: 263-274.
ALLDREDGE, A. L. 1986: Aggregate dynamics: Biological processes, which form, alter and destroy aggregates in the ocean. In: A. L. ALLDREDGE & E. 0. HARTWIG (Eds.), Aggregate dynamics in the sea: Workshop report. American Institute of Biological Sciences, Washington D.C.: 109-1 30.
- -, M. W. SILVER, 1988: Characteristics, dynamics and significance of marine snow. Prog. Oceanogr.,
ANDREASSEN, I. 1993: Suspendert organisk materide og vertikal fluks p i Svalbards nordlige sokkelom-
Mass occurrence of Sulpufusiformis in the spring of 1984 off Ireland: implication on processes. Mar. Biol., 97: 127-135.
- -. T. T. NOJI, M. Voss & R. PEINERT, 1987: Copepod fecal pellets: abundance, sedimentation and content at a permanent station in the Norwegian Sea in May/June 1986. Mar. Ecol. Prog Ser., 38: 45-5 1.
- -, T. NOJI & B. v. BODUNGEN, 1990b: Copepod grazing potential in late winter in the Norwegian Sea - a factor in the control of spring phytoplankton growth? Mar. Ecol. Prog. Ser., 60: 225-233.
- -, T. NOJI & B. v. BODLINGEN, 1991: Sedimentation of pteropods in the Norwegian Sea in autumn. Deep-sea Res., 38 (10): 1341-1360.
- - , R. PEINERT & B. v. BODUNGEN, 1990a: Pelagic origin and fate of sedimenting particles in the Norwegian Sea. Prog. Oceanogr., 24: 117-125.
BERGER, W., V. SMETACEK & G. WEER, 1989: Ocean productivity and paleoproductivity. In: W. BERGER, V. SMETACEK & G. WEFER (Eds.). Productivity of the ocean: present and past, John Wiley & Sons, New York: 1-34.
BODUNGEN, B. v., U. BATHMANN, M. Voss & M. WUNSCH, 1991: Vertical particle flux in the Norwegian Sea - resuspension and interannual variability. In: P. WASSMANN, A.3. HEISKANEN & LINDAHL (Eds.), Sediment trap studies in the Nordic countries 2. Proceedings. NurmiPrint OY, Nurmijkvi: 116-136.
BRAARUD), T., R. K. GAARDER & 0. NORDLI, 1958: Seasonal changes in the phytoplankton at various points of the Norwegian west coast. Fiskeridir. Skr. Ser. Havunders., 13: 1-77.
BRATTSTR~M, H., 1972: On Sulpafusiformis CUVIER (Thalaciacae) in Norwegian coastal and off-shore waters. Sarsia, 48: 71-90.
COLEBROOK, J. M., 1985: Continuous plankton records: overwintering and annual fluctuations in the abundance of zooplankton. Mar. Biol., 84: 261-265.
EILERTSEN, H.-C.. 1993: Spring blooms and stratification. Nature, 363: 24. EPPLEY, R. & B. J. PETERSON, 1979: Particulate organic flux and planktonic new production in the deep
FLODERLIS, S . & L. PHIL, 1990: Resuspension in the Kattegat: impact of variation in wind climate and
GRAF, G., 1989: Benthic-pelagic coupling in a deep-sea benthic community. Nature, 34: 437439. - - , 1992: Pelagic-benthic coupling: a benthic perspective. Oceanogr. Mar. Biol. Annu. Rev., 30:
GREBMEIER, 1. M. & J. P. BARRY, 1991: The influence of oceanographic processes on pelagic-benthic
HONJO, S., 1990: Particle fluxes and modem sedimentation in polaroceans. In: W. 0. SMITH (Ed.). Polar
- _ , S. J. MANCANIN, A. KAROWE, A. & B. L. WOODWARD, 1987: Particle fluxes, North-Eastem Nordic
- - , S. J. MANGANINI, G. & WEFER, 1988: Annual particle flux and a winter outburst of sedimentation
20: 41-82.
rAde. Unpubl. cand. scient. thesis, University of Tromsci; 103 pp.
ocean. Nature, 282: 677-680.
fishery. Estarine Coastal Shelf Sci., 31: 487-498.
149-1 90.
coupling in polar regions: A benthic perspective. J. Mar. Syst., 2: 495-5 18.
Oceanography. Academic Press, New York: 687-739.
Seas: 1983-1986. Woods Hole Oceanogr. Inst. Tech. Rep. WHOI-87-17; 85 pp.
in the northern Norwegian Sea. Deep-sea Res., 35: 1223-1234.

470 WASSMA", ANDREASSEN, REIGSTAD & SLAGSTAD
HBPNER-hmRSEN, G., 1989: Benthos, an important compartment in northern aquatic ecosystems. In: L. REY & V. ALEXANDER (Eds.), Proceedings of the sixth conference of the ComitB Arctique International. E. J. Brill, Leiden: 162-178.
JOHANNESSEN, 0. M., 1986: Brief overview of the physical oceanography in the Nordic Seas. In: B. G. HURDLE (Ed.), The Nordic Seas. Springer-Verlag. New York: 103-127.
KI0R!3OE, H. & J.-L. HANSEN, 1993: Phytoplankton aggregate formation: observations of patterns and mechanisms of cell sticking and the significance of exopolymeric material. J. Plankt. Res., 15:
LAMPITT, R. S . , T. NOJI & B. v. BODUNGEN, 1990: What happens to zooplankton fecal pellets? Implications for material flux. Mar. Biol., 104: 15-23.
LAWS, E. A., P. K. BIENFANG, A. D. Z ~ A N N & L. D. CONQUEST, 1988: Phytoplankton population dynamics and the fate of production during the spring bloom in Auke Bay, Alaska. Limnol. Oceanogr., 33: 57-65.
LEGENDRE, L., 1990: The significance of microalgal blooms for fisheries and for the export of particulate organic carbon in the ocean. J. Plankt. Res., 12: 681699.
LEWIS. A. G. & A. C. THOMAS, 1986: Tidal transport of planktonic copepods across the sill of a British Columbia fjord. J. Plankt. Res., 8: 1079-1089.
LIE, U., 1968: Variation in the quantity of zooplankton and propagation of C. finmarchicus at station M in the Norwegian Sea 1959-66. Fiskeridir. Skr. Ser. Havunders., 14 121-128.
LMDAHL, 0.. 1987: Plankton community dynamics in relation to water exchange in the Gulmdjord, Sweden. Ph. D. thesis, Univ. Stockholm; 138 pp.
LOENG, H., 1991: Features of the physical oceanographic conditions of the Barents Sea. In: E. SAKSHAUG, C. C. E. HOPIUNS & N. A. ~RITSLAND (Eds.), Proceedings of the Pro Mare Symposium on Polar Marine Ecology. Trondheim, 12-16 May 1990. Polar Res., 10: 5-18.
LOHRENZ, S. E., G. A. KNAL~ER, V. L. ASPER, M. TUEL, A. F. MICHAELS, & A. H. KNAP, 1992: Seasonal variability in primary production and particles flux in the northwestern Sargasso Sea: U.S. JGOFS Bermuda Atlantic Time-series Study. Deep-sea Res., 39: 1373-1391.
L m , S., J. P. TAASEN, C. C. E. HOPKINS & V. SMETACEK. 1989: Phytoplankton dynamics and sedimentation processes during spring and summer in Balsfjord, Northern Norway. Polar Biol., 10: 113-124.
MATWEWS, J. B. L.. 1986: A framework for discussion of marine zooplankton production in relation to freshwater runoff. In: S. SKRESLEIT (Ed.), The role of freshwater outflow in coastal marine ecosystems. Plenum Press, New York: 107-1 15.
- - & B. HEIMDAL, 1980: Pelagic productivity and food chains in fjord systems. In: H. J. FREELAND, D. M. FARMER, C. D. LEVINGS (Eds.), Fjord Oceanography. Plenum Press, New York 377-398.
MIDITUN, L. & H. LOENG, 1987: Climatic variations in the Barents Sea. In: H. ~ E N G (Ed.), The effect of oceanographic conditions on distribution and population dynamics of commercial fish stocks in the Barents Sea. Proc. 3rd Soviet-Norwegian Symp., Inst. Mar. Res., Bergen: 13-27.
NOJI. T. T.. 1989: The influence of zooplankton on sedimentation in the Norwegian Sea. Ber. Sonder- forschungsbereich 313, Univ. Kiel, 17: 1-183.
_ - , 1991: The influence of macrozooplankton on vertical particulate flux. Sarsia, 76: 1-9. _ - , K. W. ESTEP, F. MACINNRE & F. NORRBIN, 1991: Image analysis of fecal matter grazed upon by
three species of copepods. Evidence for coprorhexy, coprophagy and coprochaly. J. Mar. Assoc. U.K., 71: 465-480.
PARSON, T. R. & C. M. LALLI, 1988: Comparative oceanic ecology of the plankton communities of the subarctic Atlantic and Pacifjc oceans. Oceanogr. Mar. Biol. Annu. Rev., 26: 317-359.
PASSOW, U., A. ALLDREDGE & B. LOGAN, 1994: The role of particulate carbohydrate exudates in the flocculation of diatom blooms. Deep-sea Res., 41: 335-357.
F'EINERT, R., U. BATHMA", B. v. BODUNGEN & T. T. NOJI. 1987: The impact of grazing on spring phytoplankton growth and sedimentation in the Norwegian Current. In: E. T. DEGENS, E. I. IZDAR & S. HONJO (Eds.), Particle flux in the ocean. Mitt. Geol.-Palaont. Inst., Univ. Hamburg, SCO- PE/lJNEP Sonderband 62: 149-164.
_ - , B. v. BODUNGEN & V. SMETACEK, 1989: Food web structure and loss rates. In: W. BERGER, V. SMETACEK & G. WWEFER (Eds.), Productivity of the ocean: present and past. John Wiley & Sons, New York: 34-48.
PLAT, T.. W. G. HARRISON, M. L. LEWIS, W. K. W. Lr, S. SATHYENDRANATH, R. SMITH & A. F. VEZINA, 1988: Biological production and the oceans: the case for a consensus. Mar. Ecol. Prog. Ser., 52:
993- 10 18.
77-88.

Pelagic-benthic coupling in the Nordic Seas and episodic events 47 1
QUADFASEL, D., B. RUDELS & K. KURZ, 1988: Outflow of dense water from a Svalbard fjord into the Fram Strait. Deep-sea Res., 35: 1143-1 150.
REIGSTAD, M. 1994: Viroppblomstringsdynaikk og sedimentering i to nordnorske fjorder, Balsfjorden og Malangen, i 1992. Unpubl. cand. scient. thesis, University of Tromso, Norway. 79 pp.
REIMNIIZ, E., D. DETHLEFF & D. NURNBERG, 1994: Contrasts in Arctic shelf sea-ice regimes and some implication: Beaufort Sea versus Laptev Sea. Mar. Geol., 119: 215-225.
RIEBESELL, U., M. REIGSTAD, P. WASSMANN, T. NOJI, & U. PASSOW, 1995: On the trophic fate of fhaeocystis pouchetii (HARIOT): VI. Significance of fhaeocysris-derived mucus for vertical flux. Neth. J. Sea Res., 33 (2): 193-203.
ROMANKEVICH, YE. A,, 1984: Geochemistry of organic matter in the ocean. Springer Verlag, Berlin; 340 pp.
SKJOLDAL, H. R., A. HASSEL, F. REY & H. LOENG, 1987: Spring phytoplankton development and zooplankton reproduction in the central Barents Sea in the period 1979-1984. In: H. LOENG (Ed.), The effect of oceanographic conditions on distribution and population dynamics of commercial fish stocks in the Barents Sea. Roc. 3rd Soviet-Norwegian Symp., Inst. Mar. Res., Bergen: 59-89.
SMETACEK, V., B. v. BODUNGEN, B. KNOPPERS, R. PEINERT, F. POLLEHNE, P. STEGMA” & B. ZEITZSCHEL, 1984: Seasonal stages characterising the annual cycle of an inshore pelagic system. Rapp. P.-v. Reun. Cons. Int. Explor. Mer, 183: 126-135.
- - & U. PASSOW, 1990: Spring bloom initiation and SVERDRUP’S critical depth model. Limnol. Oceanogr., 35: 228-234.
SVERDRUP, H. U., 1953: On conditions for the vernal blooming of phytoplankton. J. Cons. Int. Explor.,
SVENDSEN, H.. 1981: Wind-induced variations of circulation and water level in coupled fjord-coast systems. In: R. S m & M. Mow (Eds.), The Norwegian Coastal Current. Univ. Bergen: 229-262.
- - , 1991: Preliminary results from a hydrographical investigation of Porsangenfjord, Altafjord and adjacent coastal waters, June-August 1990. Report, Geophysical Institute, Univ. Bergen, 31 pp.
TAYLOR, A. H., 1978: Long-term changes in North Atlantic Current system and their biological implication. Proc. R. Soc., Edinburgh, 76: 223-243.
TOWNSEND, D. W., M. D. KELLER, M. E. SIERACKI & S . G. ACKELSON, 1992: Spring phytoplankton blooms in the absence of vertical water column stratification. Nature, 360: 59-62.
WADHAMS, P., 1986: The ice cover. In: B. G. H, URDLE(E~.), The Nordic Seas. Springer Verlag, Berlin:
WASSMANN, P., 1990: Relationship between primary and export production in the boreal, coastal zone of the North Atlantic. Limnol. Oceanogr., 35: 464-471.
- - , 1991: Dynamics of primary production and sedimentation in shallow fjords and polls of western Norway. Oceanogr. Mar. Biol. Annu. Rev., 29: 87-154.
- - , 1993: The role of planktonic heterotrophs for the export of organic matter in eutrophicated aquatic environments. Mar. Pollut. Bull., 26: 636-643.
- -, R. PEINERT & V. SMETACEK, 1991: Patterns of production and sedimentation in the boreal and polar Northeast Atlantic. In: E. SAKSHAUG, C. C. E. HOPKINS & N. A. ~RITSLAND (Eds.), Proceedings of the Pro Mare Symposium on Polar Marine Ecology, Trondheim, 12-16 May 1990. Polar Res., 10:
- - & D. SLAGSTAD, 1993: Seasonal and interannual dynamics of carbon flux in the Barents Sea: a model approach. Polar Res., 13: 363-372.
- - & D. SLAGSTAD, 1995: Drift patterns of sinking particles in the north Norwegian coastal zone. Sediment trap studies in the Nordic countries 3. Proceedings. Nurmi Print OY, Nurmijmi: 52-69.
- -, M. VERNET, G. MITCHELL & F. REY, 1990: Mass sedimentation of Phaeocysfis poucherii in the Barents Sea. Mar. Ecol. Prog. Ser., 66: 183-195.
ZENKEVITCH, L. A., 1963: Biology of the seas of the USSR. George Allen & Unwin LTD., London;
18: 287-295.
,
21-84.
209-228.
955 pp.
Copyright © 2022 FDOKUMEN




















