
Molecular epidemiology of norovirus associated with gastroenteritis and emergence of norovirus GII.4...
9
Molecular epidemiology of norovirus associated with gastroenteritis and emergence of norovirus GII.4 variant 2012 in Japanese pediatric patients Aksara Thongprachum a,b , Wisoot Chan-it a , Pattara Khamrin c , Patchreenart Saparpakorn d , Shoko Okitsu a,b , Sayaka Takanashi a , Masashi Mizuguchi a , Satoshi Hayakawa b , Niwat Maneekarn c , Hiroshi Ushijima a,b,⇑ a Department of Developmental Medical Sciences, School of International Health, Graduate School of Medicine, The University of Tokyo, Tokyo, Japan b Division of Microbiology, Department of Pathology and Microbiology, Nihon University School of Medicine, Tokyo, Japan c Department of Microbiology, Faculty of Medicine, Chiang Mai University, Chiang Mai, Thailand d Department of Chemistry, Faculty of Science, Kasetsart University, Bangkok, Thailand article info Article history: Received 11 October 2013 Received in revised form 17 January 2014 Accepted 19 January 2014 Available online 4 February 2014 Keywords: Norovirus Epidemiology Gastroenteritis Japanese Variant abstract In late 2012, an outbreak of acute gastroenteritis due to norovirus variant Sydney_2012 occurred and have been reported from many counties. In this study, we described surveillance study of the incidence of norovirus infections among Japanese pediatric patients in association with gastroenteritis and inves- tigated the antigenic change of the new variant Sydney_2012 circulated in Japanese populations. A total of 2381 fecal specimens collected from children with acute gastroenteritis in Hokkaido, Tokyo, Shizuoka, Kyoto, Osaka, and Saga from 2009 to 2013 were examined for norovirus and further analyzed molecu- larly. A high proportion (39.3%) of norovirus positive samples and several genotypes were detected. Noro- virus GII.4 dominated over other genotypes (71.4%). The Den_Haag_2006b (43.2%) was detected as the predominant variant and co-circulated with New_Orleans_2009 (17.8%) until March 2012. Subsequently, they were displaced by Sydney_2012. The Sydney_2012 variant has been responsible for the majority of norovirus infections in 2012–2013 (85.7%). Although Sydney_2012 variant has a common ancestor with New_Orleans_2009 variant, analysis of P2 sub-domain showed a high level of diversity in comparison with other variants in four amino acid changes at the antigenic sites. The change in particular residue 393 of new variant may affect HBGA recognition. Analysis of noroviruses circulating in the past 4 years revealed a change of predominant variant of norovirus GII.4 in each epidemic season. The change of amino acid in putative epitopes may have led the virus escape from the existing herd immunity and explain the increase of new variant outbreaks. Ó 2014 Elsevier B.V. All rights reserved. 1. Introduction Norovirus is a major etiologic agent responsible for epidemic gastroenteritis worldwide. It is a single-stranded, positive sense RNA virus that belongs to the Caliciviridae family. The norovirus genome is 7.5 kb long and contains three open reading frames; ORF1 encodes the non-structural proteins, ORF2 encodes the VP1 major capsid protein, and ORF3 encodes the VP2 minor capsid pro- tein (Prasad et al., 1999). Based on genetic difference in the capsid gene, noroviruses are genetically highly diverse and divided into five genogroups (GI-V). They are further subdivided into several different genotypes, at least nine genotypes of GI, 21 genotypes of GII, 2 genotypes of GIII, and only one genotype of GIV and GV have been identified (Zheng et al., 2010; Kroneman et al., 2013). Genotype II.4 (GII.4) norovirus has been shown to spread rapidly and is the most commonly detected worldwide, particularly in association with outbreaks. Re- cent molecular surveillances indicate that GII.4 norovirus has evolved a series of genetic variants over the last 20 years, some of which persist and replace the previous circulating variants (Siebenga et al., 2010). Norovirus epidemic patterns in human pop- ulations include epidemic outbreaks of disease every 2–3 years, punctuated by the emergence of an antigenically distinct GII.4 strain that appears to escape human herd immunity to the previ- ous circulating strains (Debbink et al., 2012; Lindesmith et al., http://dx.doi.org/10.1016/j.meegid.2014.01.030 1567-1348/Ó 2014 Elsevier B.V. All rights reserved. ⇑ Corresponding author at: Department of Developmental Medical Sciences, School of International Health, Graduate School of Medicine, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan. Tel.: +81 3 5841 3509; fax: +81 3 5841 3629. E-mail address: [email protected] (H. Ushijima). Infection, Genetics and Evolution 23 (2014) 65–73 Contents lists available at ScienceDirect Infection, Genetics and Evolution journal homepage: www.elsevier.com/locate/meegid
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Molecular epidemiology of norovirus associated with gastroenteritis and emergence of norovirus GII.4...

Infection, Genetics and Evolution 23 (2014) 65–73
Contents lists available at ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsevier .com/locate /meegid
Molecular epidemiology of norovirus associated with gastroenteritis andemergence of norovirus GII.4 variant 2012 in Japanese pediatric patients
http://dx.doi.org/10.1016/j.meegid.2014.01.0301567-1348/� 2014 Elsevier B.V. All rights reserved.
⇑ Corresponding author at: Department of Developmental Medical Sciences,School of International Health, Graduate School of Medicine, The University ofTokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan. Tel.: +81 3 5841 3509; fax:+81 3 5841 3629.
E-mail address: [email protected] (H. Ushijima).
Aksara Thongprachum a,b, Wisoot Chan-it a, Pattara Khamrin c, Patchreenart Saparpakorn d,Shoko Okitsu a,b, Sayaka Takanashi a, Masashi Mizuguchi a, Satoshi Hayakawa b, Niwat Maneekarn c,Hiroshi Ushijima a,b,⇑a Department of Developmental Medical Sciences, School of International Health, Graduate School of Medicine, The University of Tokyo, Tokyo, Japanb Division of Microbiology, Department of Pathology and Microbiology, Nihon University School of Medicine, Tokyo, Japanc Department of Microbiology, Faculty of Medicine, Chiang Mai University, Chiang Mai, Thailandd Department of Chemistry, Faculty of Science, Kasetsart University, Bangkok, Thailand
a r t i c l e i n f o a b s t r a c t
Article history:Received 11 October 2013Received in revised form 17 January 2014Accepted 19 January 2014Available online 4 February 2014
Keywords:NorovirusEpidemiologyGastroenteritisJapaneseVariant
In late 2012, an outbreak of acute gastroenteritis due to norovirus variant Sydney_2012 occurred andhave been reported from many counties. In this study, we described surveillance study of the incidenceof norovirus infections among Japanese pediatric patients in association with gastroenteritis and inves-tigated the antigenic change of the new variant Sydney_2012 circulated in Japanese populations. A totalof 2381 fecal specimens collected from children with acute gastroenteritis in Hokkaido, Tokyo, Shizuoka,Kyoto, Osaka, and Saga from 2009 to 2013 were examined for norovirus and further analyzed molecu-larly. A high proportion (39.3%) of norovirus positive samples and several genotypes were detected. Noro-virus GII.4 dominated over other genotypes (71.4%). The Den_Haag_2006b (43.2%) was detected as thepredominant variant and co-circulated with New_Orleans_2009 (17.8%) until March 2012. Subsequently,they were displaced by Sydney_2012. The Sydney_2012 variant has been responsible for the majority ofnorovirus infections in 2012–2013 (85.7%). Although Sydney_2012 variant has a common ancestor withNew_Orleans_2009 variant, analysis of P2 sub-domain showed a high level of diversity in comparisonwith other variants in four amino acid changes at the antigenic sites. The change in particular residue393 of new variant may affect HBGA recognition. Analysis of noroviruses circulating in the past 4 yearsrevealed a change of predominant variant of norovirus GII.4 in each epidemic season. The change ofamino acid in putative epitopes may have led the virus escape from the existing herd immunity andexplain the increase of new variant outbreaks.
� 2014 Elsevier B.V. All rights reserved.
1. Introduction
Norovirus is a major etiologic agent responsible for epidemicgastroenteritis worldwide. It is a single-stranded, positive senseRNA virus that belongs to the Caliciviridae family. The norovirusgenome is �7.5 kb long and contains three open reading frames;ORF1 encodes the non-structural proteins, ORF2 encodes the VP1major capsid protein, and ORF3 encodes the VP2 minor capsid pro-tein (Prasad et al., 1999).
Based on genetic difference in the capsid gene, noroviruses aregenetically highly diverse and divided into five genogroups (GI-V).They are further subdivided into several different genotypes, atleast nine genotypes of GI, 21 genotypes of GII, 2 genotypes of GIII,and only one genotype of GIV and GV have been identified (Zhenget al., 2010; Kroneman et al., 2013). Genotype II.4 (GII.4) norovirushas been shown to spread rapidly and is the most commonlydetected worldwide, particularly in association with outbreaks. Re-cent molecular surveillances indicate that GII.4 norovirus hasevolved a series of genetic variants over the last 20 years, someof which persist and replace the previous circulating variants(Siebenga et al., 2010). Norovirus epidemic patterns in human pop-ulations include epidemic outbreaks of disease every 2–3 years,punctuated by the emergence of an antigenically distinct GII.4strain that appears to escape human herd immunity to the previ-ous circulating strains (Debbink et al., 2012; Lindesmith et al.,

66 A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73
2013). The continuous evolution of the P2 sub-domain of the sur-face exposed major capsid protein (VP1) has been proposed as akey mechanism by which new antigenic variants are generated.
Norovirus recognizes histo-blood group antigen (HBGAs), whichplay an important role in host susceptibility to norovirus infection(Tan and Jiang, 2011). HBGAs are a diverse family of carbohydratesexpressed on red blood cells, mucosal epithelia, saliva, milk andother body fluids, which are highly polymorphic and are relatedto the ABO, secretor and Lewis families. They have been hypothe-sized to be receptors or coreceptors that allow norovirus to attachand enter permissive cells (Debbink et al., 2012). The HBGA bind-ing interfaces are located at the top of the P dimer, correspondingto the outermost surface of the capsid. The carbohydrate bindingpockets involve several scattered amino acid residues in the P do-main that form extensive hydrogen bond network with individualsaccharides, and thus stabilizing the binding of HBGAs to the cap-sid protein (Zhang et al., 2013).
The increased detections of norovirus GII.4 variants have beenobserved globally. Since the mid-90s, norovirus GII.4 variants havebeen shown to be associated with at least six pandemics of acutegastroenteritis and continue to cause millions of infections acrossthe globe annually. The norovirus GII.4 variants reported previ-ously include the US 1995/96 in 1996 (Noel et al., 1999; Whiteet al., 2002), Farmington_Hills_2002 in 2002 (Lopman et al.,2004; Widdowson et al., 2004), Hunter_2004 in 2004 (Bull et al.,2006), Minerva_2006b and Den_Haag_2006b virus in 2006–2008(Eden et al., 2010), New_Orleans_2009 in 2009–2012 (Yen et al.,2011), and most recently Sydney_2012 (Bennett et al., 2013;Fonager et al., 2013; Mai et al., 2013; van Beek et al., 2013).
The periodic increase of norovirus outbreaks have been shownto be related to the emergence of new variant strains, probably ow-ing to evasion of population immunity (Zheng et al., 2010). Theemergence of such variants has been shown to be associated withsubstantial increases in cases worldwide (Pang et al., 2010). Sincelate 2012, surveillance systems in many countries showed theincrease of norovirus activity levels compared to the previousseasons, and associated with the emergence of a new variantGII.4 named Sydney_2012 (Bennett et al., 2013; Fonager et al.,2013; Mai et al., 2013; van Beek et al., 2013). Therefore, rapid anti-genic variations are important evolutionary forces that shape epi-demiological success and persistence of the GII.4 viruses in thepopulation. The objectives of this study were: (1) to describe asurveillance of norovirus infections that are associated with gastro-enteritis in Japanese pediatric patients; and (2) to investigate P2sub-domain of the new variant Sydney_2012 which may affectantigenic property.
2. Materials and methods
2.1. Patients and samples
A total of 2381 fecal specimens were collected from non-hospi-talized children with acute gastroenteritis in pediatric clinics in sixprefectures from northern to southern Japan including Hokkaido,Tokyo, Shizuoka, Kyoto, Osaka, and Saga from July 2009 to June2013. Among these, 515, 536, 592, and 738 samples were obtainedduring the period of 2009–2010, 2010–2011, 2011–2012, and2012–2013, respectively. In this study, the annual observation per-iod for norovirus gastroenteritis began in July and ended in June ofthe following year. The spectrum of clinical signs exhibited by pa-tients with acute gastroenteritis included acute watery diarrhea,vomiting, abdominal cramps, and fever of various severity and invarious combinations. The age of the patients ranged from neonateto 15 years (0–178 months; mean age 24.4 months). For subse-quent data analysis, seven different age-groups were established:
<6 months (213 samples), 6–11 months (673 samples), 12–23 months (829 samples), 24–35 months (334 samples), 36–47 months (113 samples), 48–59 months (63 samples), and>60 months (156 samples). The specimens were stored at �20 �Cuntil use.
2.2. Viral RNA extraction
The fecal specimens were prepared as 10% fecal suspensions indistilled water. Viral genome was extracted from the supernatantof 10% suspension using the QIAamp Viral RNA MiniKit, accordingto the manufacturer’s instructions (QIAgen, Hilden, Germany). Theextracted viral RNAs were stored at �80 �C until use.
2.3. Detection of norovirus
The reverse transcription (RT) was performed with RNA tem-plate using SuperScriptIII reverse transcriptase (Invitrogen, Carls-bad, CA, USA). The presence of norovirus was detected by RT-PCRusing the protocol described previously (Yan et al., 2003). Thespecimens were tested for the presence of norovirus GI and GIIby amplification of partial gene. For norovirus GI, a combinationof forward primer G1-SKF (nt 5342–5361) 50-CTGCCCGAATTYG-TAAATGA-30 and reverse primer G1SKR (nt 5653–5671) 50-CCAACCCARCCATTRTACA-30 were used. For norovirus GII, a forwardprimer COG2F (nt 5003–5028; 50-CARGARBCNATGTTYAGRTGGAT-GAG-30) was used in combination with the reverse primer G2SKR(nt 5367–5389; 50-CCRCCNGCATRHCCRTTRTACAT-30). Additionallyfor norovirus GII, semi-nested PCR was performed using G2SKF (nt5389–5367; 50-CNTGGGAGGGCGATCGCAA-30) and G2SKR primersfor the samples that were negative by the first set of primer fornorovirus GII.
2.4. Amplification of polymerase and capsid genes of norovirus
The partial polymerase and full-length capsid region of a newvariant was amplified using two pairs of primers, GV21 (50-GTBGGNGGYCARATGGGNATG-30)/G2SKR (50-CCRCCNGCATRHCCRTTRTACAT-30) and COG2F (50-CARGARBCNATGTTYAGRTGGATGAG-30)/TX30SXN (50-GACTAGTTCTAGATCGCGAGCGGCCGCCC(T)30-30).Long-range PCR was amplified with High Fidelity DNA polymeraseKOD-Plus (Toyobo, Osaka, Japan). The PCR was carried out with2 ll of cDNA using TX30SXN primer, in the reagent mixture con-taining 2.5 ll of 10� buffer for KOD-Plus, 1 ll of 2 mM dNTPs,1 ll of 25 mM MgSO4, 0.75 ll of each specific primer (20 lM)and 0.5 ll of KOD Plus enzyme. MilliQ water was added to makeup a total volume of 25 ll. The PCR was performed at 94 �C for2 min, followed by 35 cycles of 98 �C 20 s, 50 �C or 55 �C 30 s,68 �C 2 min, and a final extension at 68 �C for 10 min, and then heldat 4 �C.
2.5. Sequence and Phylogenetic Analyses
The genotypes of the norovirus were characterized by directnucleotide sequencing of partial capsid gene and then performingphylogenetic analysis as described previously by Kageyama et al.(2004). The PCR products were purified using QIAquick Gel Extrac-tion Kit (Qiagen, Hilden, Germany) and sequenced by using theBigDye Terminator Cycle Sequencing kit (Perkin Elmer-AppliedBiosystems, Inc., Foster City, CA) on an automated DNA sequencer(ABI 3100; Perkin Elmer-Applied Biosystems, Inc., Foster City, CA).The nucleotide sequences were analysed in comparison with thoseof norovirus strains deposited in GenBank database. Phylogeneticrelationship of norovirus was examined by aligning sequenceswith the ClustalX program. A phylogenetic tree was constructed

A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73 67
according to the maximum likelihood (ML) method using MEGAversion 5 (Tamura et al., 2011).
2.6. Nucleotide sequence accession numbers
The partial nucleotide sequences of the capsid gene of norovirusidentified in this study were deposited in GenBank under accessionnumbers KF658275–KF658339. Accession number for partialpolymerase and full-length capsid sequences of norovirus GII.4Sydney_2012 variant used in this study is KF145149.
2.7. Three-dimensional structure
The three-dimensional structures of norovirus genomes of vari-ants were constructed by homology modeling technique usingGeno3D web server (Combet et al., 2002), the structures werechecked the quality of geometry by PROCHECK (Laskowski et al.,1993). For the model structures, all sequences were based on thestructure of a norovirus GII.4 2004 norovirus using PDB code 3sej(Shanker et al., 2011).
3. Results
3.1. Detection and genotypic distribution of norovirus
A high proportion (39.3%; 936/2381) of norovirus were detectedin the pediatric patients during 2009–2013, comprising 0.3% of GI(6/2381), 39.0% of GII (928/2381), and 0.1% mixed GI/GII (2/2381).Most of norovirus belonged to norovirus GII genogroup (39.1%;930/2381). As shown in Table 1, the detection rate of norovirusGII were 40.8% (210/515) in 2009–2010, 40.1% (215/536) in2010–2011, 31.8% (188/592) in 2011–2012, and 43.0% (317/738)in 2012–2013.
Based on the analysis of partial sequence of capsid gene of noro-virus GII, nine distinct genotypes were identified. Of these, GII.4was the most predominant genotype (71.4%; 664/930), followedby GII.3 (13.1%; 122/930), GII.2 (6.5%; 60/930), GII.14 (4.4%; 41/
Table 1Genotype distributions of noroviruses detected in Japanese pediatric patients with diarrhe
Year No. ofsamplestested
No. ofnoroviruspositive (%)
No. ofgenogrouppositive (%)
No. of ge
GI GII GII.1 G
2004–2005 752 139 (18.5) 2(0.3)
137(18.2)
1 0
2005–2006 603 51 (8.5) 0 (0) 51(8.5)
1 1
2006–2007 628 97 (15.4) 2(0.3)
95(15.1)
0 2
2007–2008 477 125 (26.2) 7(1.5)
118(24.7)
0 0
2008–2009 477 129 (27.0) 0 (0) 129(27.0)
1 4
2009–2010 515 210 (40.8) 0 (0) 210(40.8)
0 2
2010–2011 536 215 (40.1) 1(0.2)
215(40.1)
0 1
2011–2012 592 188 (31.8) 0 (0) 188(31.8)
0 7
2012–2013 738 323 (43.8) 7(0.9)
317(43.0)
0 1
930), GII.7 (1.4%; 13/930), GII.6 (1.3%; 12/930), GII.13 (1.1%; 10/930), GII.12 (0.8%; 7/930), and GII.17 (0.1%; 1/930) (Table 1). Thenorovirus GI was detected at 0.3% (8/2381), comprising GI.1(0.04%), GI.4 (0.04%), and GI.6 (0.25%).
Out of total samples, 28.1% of the patients were infected bynorovirus GII alone, while 11.0% were co-circulated with other en-teric viruses. Double-infection and triple-infection were found at10.5% (250/2381) and 0.5% (11/2381), respectively. Mix-infectionbetween norovirus GII and group A rotavirus was the most pre-dominant (4.5%; 106/2381), followed by human parechovirus(2.3%), adenovirus (1.9%), enterovirus (1.4%), sapovirus (0.3%),astrovirus (0.1%), and norovirus GI (0.1%).
3.2. Identification of epidemic norovirus GII.4 variants
Phylogenetic tree of nucleotide sequences of the partial capsidregion of norovirus GII isolates detected in the present study wasconstructed in comparison with those of the reference strainsavailable in GenBank database (Fig. 1). The norovirus GII.4 con-sisted of three major variants strains detected in pediatric patientsin this study (Fig. 2). The Den_Haag_2006b strain that appearedduring the study period comprised 287 of 664 norovirus GII.4strains (43.2%). The sequences of these norovirus strains exhibited98.0–100% nucleotide identity and 98.7–100% amino acid identitywith Hu/GII.4/DenHaag89/2006/NL (GenBank accession numberEF126965). The second variant, New_Orleans_2009 strain, was de-tected in 17.8% (118/664) and displayed 96.0–100% nucleotide se-quence identity and 97.5–100% amino acid identity with the capsidregion of Hu/GII.4/New Orleans1805/2009/USA (GU445325). Inter-estingly, the new Sydney_2012 variant was detected at 36.4% (242/664) and clustered within the same cluster of the Sydney/NSW0514/2012/AU (JX459908) and Hong Kong/CUHK3655/2012/CHN (JX629456) reference strains and showed 98.7–100% nucleo-tide sequence identity and 100% amino acid sequence identity withreference strains. However, a number of additional GII.4 variantshave also been identified, including Hunter 2004, Yerseke_2006a,Apeldoorn_2007, and Hokkaido1_2008. These variants were lesscommon in this study.
a from 2004 to 2013.
notype cases Reference
II.2 GII.3 GII.4 GII.6 GII.7 GII.12 GII.13 GII.14 GII.17
22 108 6 0 0 0 0 0 Phanet al.(2006)
27 19 2 1 0 0 0 0 Phanet al.(2007)
6 84 0 3 0 0 0 0 Dey et al.(2012)
6 92 6 0 0 0 14 0 Chan-itet al.(2011)
7 92 18 1 3 0 3 0 Chan-itet al.(2011)
4 30 135 1 6 5 1 7 1 Thisstudy
7 60 118 2 1 1 2 14 0 Thisstudy
30 132 3 0 0 5 11 0 Thisstudy
2 2 279 6 6 1 2 9 0 Thisstudy

Fig. 1. Phylogenetic analysis of the partial capsid sequence of norovirus GII.4 strains. The phylogenetic tree constructed using the maximum likelihood method.Representative strains detected in this study are presented in boldface.
68 A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73

Fig. 2. Monthly distribution of norovirus GII.4 norovirus infection in Japan pediatric patients during the epidemic seasons of 2009/2010–2012/2013. (A) norovirus genotypes.(B) Norovirus GII.4 variants.
A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73 69
3.3. Seasonal pattern and age distribution of norovirus infection
The seasonal pattern of norovirus infection is shown in Fig. 2.Norovirus was detected all year round with a highest peak inDecember. From this study, it was clearly observed that manynoroviruses genotypes were common in winter season in Japan(Fig. 2A). The highest prevalence of norovirus was found in Decem-ber (23.6%; 221/936), followed by January (19.6%; 183/936),November (12.3%; 115/936), and February (9.8%; 92/936),respectively.
Focusing on norovirus GII.4 variants, most of norovirus GII.4 oc-curred between November and January, with a highest peak inDecember (25.9%). The Den_Haag_2006b variant was detected asthe most predominant variant until March 2012 in co-circulationwith New_Orleans_2009 variant. Subsequently, they were replacedby variant Sydney_2012 which was described in June 2012(Fig. 2B). However, the Sydney_2012 was first detected in a Japa-nese patient in Tokyo since November 2011 before spreadingthroughout Japan from mid-2012.
Of the patients with acute gastroenteritis, norovirus infectionwas detected in all age groups. The mean age of norovirus infectedpatients was 22.4 months. The highest prevalence of norovirus wasfound in infants and children aged 12–23 months (42.4%, n = 397),
followed by 6–11 months (25.6%, n = 240), 24–36 months (14.9%,n = 139), less than 6 months (6.3%, n = 59), 36–47 months (4.5%,n = 42), more than 60 months (3.5%, n = 33), and the lowest in chil-dren aged 48–59 months (1.1%, n = 26).
3.4. Analysis of polymerase and capsid of norovirus GII.4 Sydney_2012
To determine antigenic differences between GII.4 Sydney_2012and the recent circulating ancestral strains Den_Haag_2006b andNew_Orleans_2009 strains. We analysed VP1 and P2 sub-domainespecially in blockade epitopes of Sydney_2012 and compared withother variant strains. The analysis of partial VP1 capsid of GII.4 var-iant 2012 representative strains revealed a high diversity of 6 posi-tions of amino acid substitutions compared to Den_Haag_2006band New_Orleans_2009 variants; positions 294(A2006b ? P2009 ?T2012), 359(T2006b ? S2009 ? A2012), 368(S2006b ? A2009 ? E2012),377(T2006b ? T2009 ? A2012), 393(S2006b ? S2009 ? G2012), and413(V2006b ? I2009 ? T2012), as shown in Fig. 2A and B. In the anal-ysis of P2 sub-domain, the Japanese norovirus GII.4 variant 2012strains showed P98.9% nucleotide and P99.2% amino acid se-quence identities with the Sydney/NSW0514/2012 (JX459908).Moreover, analysis of polymerase region of the Sydney_2012 vari-ant was found to be similar to Apeldoorm_2007 but distinct from

70 A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73
other variants and accounted for three amino acid substitutions inpolymerase region.
3.5. Effect of HBGA binding site
In order to investigate the HBGA binding site, the superimposi-tion between the crystal structure of Hunter_2004 strain (pdb code3sej) and other variants were studied as shown in Fig. 4A. Thestructure of Hunter_2004 strain revealed the hydrogen bond inter-action between Lewisb hexasaccharide and some amino acid resi-dues in both dimers (i.e. Chain A: Gln390, Asp391, Gly392,Ser393, Thr394, Thr395, Ser442, and Gly443, Chain C: Ser343,Thr344, Arg345, and Asp374), as shown in Fig. 4B. For the strainsof Sydney_2012 (JP41), the amino acid residue position 393 wasglycine which was different from other variant strains. From thecrystal structure, the hydroxyl group of Ser393 formed the hydro-gen bond interaction to Lewis fucose (LeFuc) while Gly393 in Syd-ney_2012 (JP41) strain could not form the hydrogen bondinteraction to the LeFuc (Fig. 4A).
4. Discussion
Viral gastroenteritis is still a health burden and one of the mostfrequently encountered problems in developed and developingcountries. Global outbreaks of acute gastroenteritis by norovirushave frequently reported. Our group also previously reported theepidemic occurrence of norovirus in a Japanese population (Phanet al., 2006; Chan-it et al., 2011; Dey et al., 2011). The current studyextended these findings by investigating different epidemic pat-terns. A high prevalence (39.3%) of norovirus infection was foundin these Japanese pediatric patients with acute gastroenteritis inJapan during July 2009 to June 2013; 99.4% of all norovirus caseswere infected by norovirus GII. In contrast, the prevalences of rota-virus, human parechovirus, enterovirus, adenovirus, sapovirus,astrovirus, and Aichi virus were 20.1%, 6.6%, 6.1%, 5.6%, 4.8%,2.3%, and 0.1%, respectively (unpublished data). The prevalence ofnorovirus is higher than that found in the preceding studies con-ducted in the same geographical area during 2004–2009 whichshowed the prevalence rates of norovirus ranged from 8.5% to27.0% (Phan et al., 2006; Chan-it et al., 2011; Dey et al., 2011). Ta-ken together, results in the present and preceding studies con-firmed that the detection rate increased year by year andnorovirus is the most predominant agent causing acute gastroen-teritis in Japan (Kageyama et al., 2004; Phan et al., 2006; Chan-itet al., 2011; Dey et al., 2011). This study documents 11.0% mixedinfection between norovirus and other enteric viruses, in which4.5% mixed infection between norovirus and group A rotaviruswas most predominant. Differences in rotavirus and norovirus sea-sonality may relate to co-infection rate in this study. Moreover,mixed infection was not detected more frequently in any particularage group.
Regarding norovirus seasonality, the seasonal distribution ofnorovirus infection exhibits a peak in winter seasons, a highestpeak of incidence in December (23.6%). It is clearly observed thatnorovirus infection, especially norovirus GII.4 is commonly de-tected in cold months in Japan; 65.3% of confirmed norovirus infec-tion cases occurred between December and February. Our data arein good agreement with the other surveillance on pediatric caseswhich demonstrated the main peak of norovirus during the wintermonths in Japan (Iritani et al., 2010; Dey et al., 2010). The temper-ature and humidity had highly consistent effects on driving noro-virus epidemic patterns (Lopman et al., 2009). A high number ofnorovirus cases detected in every winter season in this study mightbe associated with cold and dry temperature.
Several studies reported that norovirus infection occurredfrequently in young children under 2 years-old (Sai et al., 2013;Yoneda et al., 2014). In agreement with this, our results showeda marked trend toward higher rates of infection in children un-der-2-years of age (74.4%, 696/936), particularly children between12 and 23 months are the most frequently affected group. The find-ings show that norovirus infections usually occur in early child-hood. It is possible that the children younger than 2 years oldmight lack of maternal antibody protection.
Based on antigenic analyses and more recent extensive se-quence analyses, the circulation of norovirus in nature have beenshown to be highly variable. In this study, genetic analysis amongnorovirus GII demonstrated that GII.4 genotype is the most pre-dominant genotype accounting for 71.4%, followed by GII.3, GII.2,GII.14, GII.7, GII.6, GII.13, GII.12, and GII.17. The highest prevalenceof GII.4 is consistent with those obtained from earlier studies con-ducted in the same settings as well as other studies conductedelsewhere in Japan (Phan et al., 2006; Dey et al., 2011; Chan-itet al., 2011; Hoa Tran et al., 2013; Yoneda et al., 2014). NorovirusGII.4 strains evolved rapidly and spread globally with the emergingvariants. On the other hand, the increase of norovirus GII.3 hasbeen identified and this strain is the second leading genotype in Ja-pan after dropping during 2006–2009. A sharp decrease in thenumber of patients infected by GII.3 was observed when the newemerging variant was spread in 2012–2013 (14.3% in 2009–2010;27.9% in 2010–2011; 16.0% in 2012–2013; 0.6% in 2012–2013).However, norovirus GI was low detectable in this surveillancestudy.
Focusing on norovirus GII.4 variants, analysis of noroviruses cir-culating in the past 4 years in Japanese pediatric patients revealeda change of the predominant variant of norovirus GII.4 in each epi-demic season over time. Among norovirus GII.4 identified in thisstudy, three separate norovirus GII.4 variants were mainly identi-fied as the etiologic agents of norovirus-associated acute gastroen-teritis, including Den_Haag_2006b (43.2%), New_Orleans_2009(17.8%), and Sydney_2012 (36.4%) variants.
The Sydney_2012 new variant was shown to emerge since late2011 and this emergence has been shown to be associated with theincrease of norovirus infections in Japanese pediatric patients fromMay 2012. Interestingly, the previous dominant Den_Haag_2006band New_Orleans_2009 variants were displaced by new emer-gence variant Sydney_2012 (Fig. 2B). The Sydney_2012 varianthas been shown to be responsible for the majority of norovirusinfection in 2012–2013, accounting for 85.7% of all norovirusGII.4 detected in 2012–2013. Even though many studies reportedthe detection of Sydney_2012 variant from late 2012 (Bennettet al., 2013; Fonager et al., 2013; van Beek et al., 2013), the presentstudy described the detection of Sydney_2012 variant among pedi-atric patients in Tokyo since November 2011 before spreadingthroughout Japan from mid-2012. This study clearly indicated thatnorovirus variant Sydney_2012 was the predominant group caus-ing acute gastroenteritis among Japanese pediatric populationssince mid-2012. Interestingly, the Sydney_2012 strains detectedin the current study were clustered in the same lineage andshowed a high nucleotide sequence (P98.7%) and amino acid se-quence (100%) similar to norovirus GII found previously Australia,the United States, Europe, Canada, and China (Bennett et al., 2013;Fonager et al., 2013; Mai et al., 2013; van Beek et al., 2013). Then,the Sydney_2012 had emerged as the etiologic agent in acutegastroenteritis across the United States, Europe and Asia, includingJapan in the same period.
Although the Sydney_2012 variant has common ancestor withthe dominant New_Orleans_2009 variants, analysis of partial VP1capsid of GII.4 variant 2012 representative strains revealed a highdiversity with 6 positions of amino acid substitutions, positions294, 359, 368, 377, 393, and 413, compared to Den_Haag_2006b
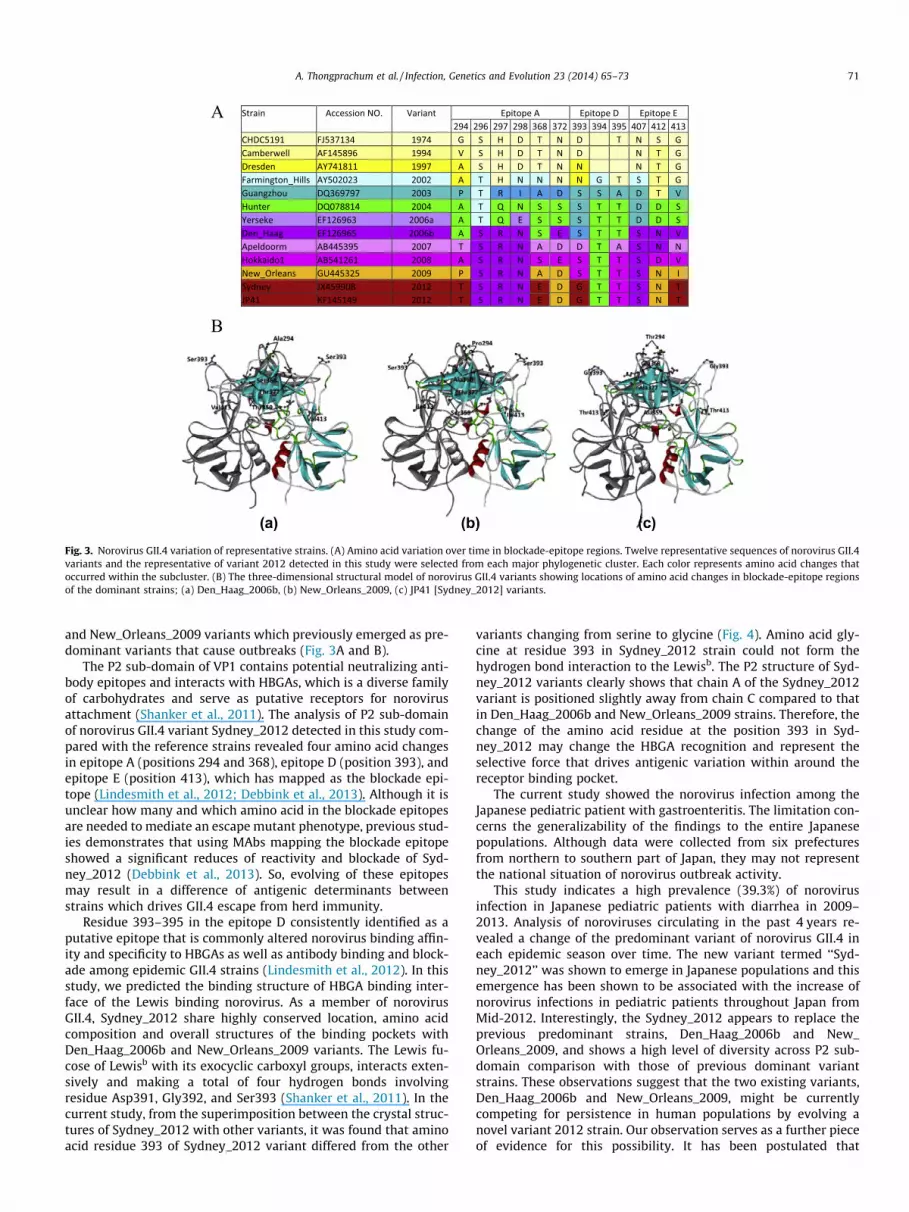
Strain Accession NO. Variant Epitope A Epitope D Epitope E294 296 297 298 368 372 393 394 395 407 412 413
CHDC5191 FJ537134 1974 G S H D T N D T N S G Camberwell AF145896 1994 V S H D T N D N T G Dresden AY741811 1997 A S H D T N N N T G Farmington_Hills AY502023 2002 A T H N N N N G T S T G Guangzhou DQ369797 2003 P T R I A D S S A D T V Hunter DQ078814 2004 A T Q N S S S T T D D S Yerseke EF126963 2006a A T Q E S S S T T D D S Den_Haag EF126965 2006b A S R N S E S T T S N V Apeldoorm AB445395 2007 T S R N A D D T A S N N Hokkaido1 AB541261 2008 A S R N S E S T T S D V New_Orleans GU445325 2009 P S R N A D S T T S N I Sydney JX459908 2012 T S R N E D G T T S N T JP41 KF145149 2012 T S R N E D G T T S N T
A
B
Fig. 3. Norovirus GII.4 variation of representative strains. (A) Amino acid variation over time in blockade-epitope regions. Twelve representative sequences of norovirus GII.4variants and the representative of variant 2012 detected in this study were selected from each major phylogenetic cluster. Each color represents amino acid changes thatoccurred within the subcluster. (B) The three-dimensional structural model of norovirus GII.4 variants showing locations of amino acid changes in blockade-epitope regionsof the dominant strains; (a) Den_Haag_2006b, (b) New_Orleans_2009, (c) JP41 [Sydney_2012] variants.
A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73 71
and New_Orleans_2009 variants which previously emerged as pre-dominant variants that cause outbreaks (Fig. 3A and B).
The P2 sub-domain of VP1 contains potential neutralizing anti-body epitopes and interacts with HBGAs, which is a diverse familyof carbohydrates and serve as putative receptors for norovirusattachment (Shanker et al., 2011). The analysis of P2 sub-domainof norovirus GII.4 variant Sydney_2012 detected in this study com-pared with the reference strains revealed four amino acid changesin epitope A (positions 294 and 368), epitope D (position 393), andepitope E (position 413), which has mapped as the blockade epi-tope (Lindesmith et al., 2012; Debbink et al., 2013). Although it isunclear how many and which amino acid in the blockade epitopesare needed to mediate an escape mutant phenotype, previous stud-ies demonstrates that using MAbs mapping the blockade epitopeshowed a significant reduces of reactivity and blockade of Syd-ney_2012 (Debbink et al., 2013). So, evolving of these epitopesmay result in a difference of antigenic determinants betweenstrains which drives GII.4 escape from herd immunity.
Residue 393–395 in the epitope D consistently identified as aputative epitope that is commonly altered norovirus binding affin-ity and specificity to HBGAs as well as antibody binding and block-ade among epidemic GII.4 strains (Lindesmith et al., 2012). In thisstudy, we predicted the binding structure of HBGA binding inter-face of the Lewis binding norovirus. As a member of norovirusGII.4, Sydney_2012 share highly conserved location, amino acidcomposition and overall structures of the binding pockets withDen_Haag_2006b and New_Orleans_2009 variants. The Lewis fu-cose of Lewisb with its exocyclic carboxyl groups, interacts exten-sively and making a total of four hydrogen bonds involvingresidue Asp391, Gly392, and Ser393 (Shanker et al., 2011). In thecurrent study, from the superimposition between the crystal struc-tures of Sydney_2012 with other variants, it was found that aminoacid residue 393 of Sydney_2012 variant differed from the other
variants changing from serine to glycine (Fig. 4). Amino acid gly-cine at residue 393 in Sydney_2012 strain could not form thehydrogen bond interaction to the Lewisb. The P2 structure of Syd-ney_2012 variants clearly shows that chain A of the Sydney_2012variant is positioned slightly away from chain C compared to thatin Den_Haag_2006b and New_Orleans_2009 strains. Therefore, thechange of the amino acid residue at the position 393 in Syd-ney_2012 may change the HBGA recognition and represent theselective force that drives antigenic variation within around thereceptor binding pocket.
The current study showed the norovirus infection among theJapanese pediatric patient with gastroenteritis. The limitation con-cerns the generalizability of the findings to the entire Japanesepopulations. Although data were collected from six prefecturesfrom northern to southern part of Japan, they may not representthe national situation of norovirus outbreak activity.
This study indicates a high prevalence (39.3%) of norovirusinfection in Japanese pediatric patients with diarrhea in 2009–2013. Analysis of noroviruses circulating in the past 4 years re-vealed a change of the predominant variant of norovirus GII.4 ineach epidemic season over time. The new variant termed ‘‘Syd-ney_2012’’ was shown to emerge in Japanese populations and thisemergence has been shown to be associated with the increase ofnorovirus infections in pediatric patients throughout Japan fromMid-2012. Interestingly, the Sydney_2012 appears to replace theprevious predominant strains, Den_Haag_2006b and New_Orleans_2009, and shows a high level of diversity across P2 sub-domain comparison with those of previous dominant variantstrains. These observations suggest that the two existing variants,Den_Haag_2006b and New_Orleans_2009, might be currentlycompeting for persistence in human populations by evolving anovel variant 2012 strain. Our observation serves as a further pieceof evidence for this possibility. It has been postulated that

Strain Variant Chain A Chain C
390 391 392 393 442 443 343 344 345 374
Hunter 2004 Q D G S S G S T R D
Den_Haag 2006b Q D G S S G S T R D
New_Orleans 2009 Q D G S S G S T R D
Sydney 2012 Q D G G S G S T R D
JP41 2012 Q D G G S G S T R D
A
B
Fig. 4. HBGA binding of norovirus GII. 4 variant representative strains. (A)Hydrogen-bond interaction (red dash line) in HBGA binding site between Lewisb
hexasaccharide and Hunter_2004 strain (pdb code 3sej) (Grey). Structures ofDen_Haag_2006 (Blue), New Orleans_2009 (Green), Sydney_2012 (Pink) and JP41(Orange) are also shown. ⁄SeFuc (secretor fucose), LeFuc (Lewis fucose), Gal(galactose), GlcNAc (N-acetylglucosamine), and Glc (glucose). (B) Amino acidresidues interacting with Secretor Lewis HBGA (Lewisb). (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version ofthis article.)
72 A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73
diversification of the P2 sub-domain through accumulated muta-tions in antigenic blockade epitopes and HBGAs binding pocketin Sydney_2012 may affect HBGA binding and has been linked toantigenic escape from host immune responses directed to previousinfection that permits the emergence of widespread a new variantSydney_2012.
Acknowledgements
We are grateful to Dr. Shuichi Nishimura, Dr. Hideaki Kikuta, Dr.Atsuko Yamamoto, Dr. Kumiko Sugita, Dr. Masaaki Kobayashi, andDr. Tsuneyoshi Baba for specimen collections. This study wassupported by Japan Society for the Promotion of Science (JSPS)Grants-in-Aid for Scientific Research (Grant Number; 23406036and 24390266).
References
Bennett, S., MacLean, A., Miller, R.S., Aitken, C., Gunson, R.N., 2013. Increasednorovirus activity in Scotland in 2012 is associated with emergence of a newnorovirus GII.4 variant. Euro. Surveill. 18, 20349.
Bull, R.A., Tu, E.T., Mciver, C.J., Rawlinson, W.D., White, P.A., 2006. Emergence of anew norovirus genotype II.4 variant associated with global outbreaks ofgastroenteritis emergence of a new norovirus genotype II.4 variant associatedwith global outbreaks of gastroenteritis. J. Clin. Microbiol. 44, 327–333.
Chan-It, W., Thongprachum, A., Okitsu, S., Nishimura, S., Kikuta, H., Baba, T.,Yamamoto, A., Sugita, K., Hashira, S., Tajima, T., Mizuguchi, M., Ushijima, H.,2011. Detection and genetic characterization of norovirus infections in childrenwith acute gastroenteritis in Japan, 2007–2009. Clin. lab. 57, 213–220.
Combet, C., Jambon, M., Deléage, G., Geourjon, C., 2002. Geno3D: automaticcomparative molecular modeling of protein. Bioinformatics 18, 213–214.
Debbink, K., Donaldson, E.F., Lindesmith, L.C., Baric, R.S., 2012. Genetic mapping of ahighly variable norovirus GII.4 blockade epitope: potential role in escape fromhuman herd immunity. J. Virol. 86, 1214–1226.
Debbink, K., Lindesmith, L.C., Donaldson, E.F., Costantini, V., Beltramello, M., Corti,D., Swanstrom, J., Lanzavecchia, A., Vinjé, J., Baric, R.S., 2013. Emergence of newpandemic GII.4 Sydney norovirus strain correlates with escape from herdimmunity. J. Infect. Dis. 27 [Epub ahead of print].
Dey, S.K., Phathammavong, O., Okitsu, S., Mizuguchi, M., Ohta, Y., Ushijima, H., 2010.Seasonal pattern and genotype distribution of norovirus infection in Japan.Pediatr. Infect. Dis. J. 29, 32–34.
Dey, S.K., Phan, T.G., Mizuguchi, M., Okitsu, S., Ushijima, H., 2011. Genetic diversityand emergence of norovoruis GII/4-2006b in Japan during 2006–2007. Clin. Lab.57, 193–199.
Eden, J.S., Bull, R.A., Tu, E., McIver, C.J., Lyon, M.J., Marshall, J.A., Smith, D.W., Musto,J., Rawlinson, W.D., White, P.A., 2010. Norovirus GII.4 variant 2006b causedepidemics of acute gastroenteritis in Australia during 2007 and 2008. J. Clin.Virol. 49, 265–271.
Fonager, J., Hindbæk, L.S., Fischer, T.K., 2013. Rapid emergence and antigenicdiversification of the norovirus 2012 Sydney variant in Denmark, October–December, 2012. Euro. Surveill. 18, 10–13.
Hoa Tran, T.N., Trainor, E., Nakagomi, T., Cunliffe, N.A., Nakagomi, O., 2013.Molecular epidemiology of noroviruses associated with acute sporadicgastroenteritis in children: global distribution of genogroups, genotypes andGII.4 variants. J. Clin. Virol. 56, 185–193.
Iritani, N., Kaida, A., Kubo, H., Abe, N., Goto, K., Ogura, H., 2010. MolecularEpidemiology of noroviruses detected in seasonal outbreaks of acutenonbacterial gastroenteritis in Osaka city, Japan, from 1996–1997 to 2008–2009. J. Med. Virol. 2105, 2097–2105.
Kageyama, T., Shinohara, M., Uchida, K., Fukushi, S., Hoshino, F.B., Kojima, S., Takai,R., Oka, T., Takeda, N., Katayama, K., Kageyama, T., Shinohara, M., Uchida, K.,Fukushi, S., Takeda, N., Katayama, K., 2004. Coexistence of multiple genotypes,including newly identified genotypes, in outbreaks of gastroenteritis due tonorovirus in Japan. J. Clin. Microbiol. 42, 2988–2995.
Kroneman, A., Vega, E., Vennema, H., Vinje, J., White, P.A., Hansman, G., Green, K.,Martella, V., Katayama, M., Koopmans, M., 2013. Proposal for unified norovirusnomenclature and genotyping. Arch. Virol. 158, 2059–2068.
Laskowski, R.A., MacArthur, M.W., Moss, D.S., Thornton, J.M., 1993. PROCHECK: aprogram to check the stereochemical quality of protein structures. J. Appl.Crystallogr. 26, 283–291.
Lindesmith, L.C., Beltramello, M., Donaldson, E.F., Corti, D., Swanstrom, J., Debbink,K., Lanzavecchia, A., Baric, R.S., 2012. Immunogenetic mechanisms drivingnorovirus GII.4 antigenic variation. PLoS Pathog. 8, e1002705.
Lindesmith, L.C., Costantini, V., Swanstrom, J., Debbink, K., Donaldson, E.F., Vinjé, J.,Baric, R.S., 2013. Emergence of a norovirus GII.4 strain correlates with changesin evolving blockade epitopes. J. Virol. 87, 2803–2813.
Lopman, B., Vennema, H., Kohli, E., Pothier, P., Sanchez, A., Negredo, A., Buesa, J.,Schreier, E., Reacher, M., Brown, D., Gray, J., Iturriza, M., Gallimore, C., Bottiger,B., Hedlund, K.O., Torvén, M., von Bonsdorff, C.H., Maunula, L., Poljsak-Prijatelj,M., Zimsek, J., Reuter, G., Szücs, G., Melegh, B., Svennson, L., van Duijnhoven, Y.,Koopmans, M., 2004. Increase in viral gastroenteritis outbreaks in Europe andepidemic spread of new norovirus variant. Lancet 363, 682–688.
Lopman, B., Armstrong, B., Atchison, C., Gray, J.J., 2009. Host, weather and virologicalfactors drive norovirus epidemiology: time-series analysis of laboratorysurveillance data in England and Wales. PLoS ONE 4, e6671.
Mai, H., Jin, M., Guo, X., Liu, J., Liu, N., Cong, X., Gao, Y., Wei, L., 2013. Clinical andepidemiologic characteristics of norovirus GII.4 Sydney during winter 2012–13in Beijing, China following its global emergence. PLoS ONE 8, e71483.
Noel, J.S., Fankhauser, R.L., Ando, T., Monroe, S.S., Glass, R.I., 1999. Identification of adistinct common strain of ‘‘Norwalk-like viruses’’ having a global distribution. J.Infect. Dis. 179, 1334–1344.
Pang, X.L., Preiksaitis, J.K., Wong, S., Li, V., Lee, B.E., 2010. Influence of novelnorovirus GII.4 variants on gastroenteritis outbreak dynamics in Alberta and theNorthern Territories, Canada between 2000 and 2008. PLoS ONE 5, e11599.
Phan, T.G., Takanashi, S., Kaneshi, K., Ueda, Y., Nakaya, S., Nishimura, S., Sugita, K.,Nishimura, T., Yamamoto, A., Yagyu, F., Okitsu, S., Maneekarn, N., Ushijima, H.,2006. Detection and genetic characterization of norovirus strains circulatingamong infants and children with acute gastroenteritis in Japan during 2004–2005. Clin. Lab. 52, 519–525.
Phan, T.G., Nishimura, S., Sugita, K., Nishimura, T., Okitsu, S., Ushijima, H., 2007.Multiple recombinant noroviruses in Japan. Clin. Lab. 53, 567–570.
Prasad, B.V., Hardy, M.E., Dokland, T., Bella, J., Rossmann, M.G., Estes, M.K., 1999. X-ray crystallographic structure of the norwalk virus capsid. Science 286, 287–290.
Sai, L., Sun, J., Shao, L., Chen, S., Liu, H., Ma, L., 2013. Epidemiology and clinicalfeatures of rotavirus and norovirus infection among children in Ji’nan. ChinaVirol. J. 10, 302.
Shanker, S., Choi, J.M., Sankaran, B., Atmar, R.L., Estes, M.K., Prasad, B.V., 2011.Structural analysis of histo-blood group antigen binding specificity in anorovirus GII.4 epidemic variant: implications for epochal evolution. J. Virol.85, 8635–8645.
Siebenga, J.J., Lemey, P., Kosakovsky Pond, S.L., Rambaut, A., Vennema, H.,Koopmans, M., 2010. Phylodynamic reconstruction reveals norovirus GII.4epidemic expansions and their molecular determinants. PLoS pathog. 6, 8–84.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28,2731–2739.
Tan, M., Jiang, X., 2011. Norovirus-host interaction: multi-selections by humanhisto-blood group antigens. Trends Microbiol. 19, 382–388.

A. Thongprachum et al. / Infection, Genetics and Evolution 23 (2014) 65–73 73
van Beek, J., Ambert-Balay, K., Botteldoorn, N., Eden, J.S., Fonager, J., Hewitt, J.,Iritani, N., Kroneman, A., Vennema, H., Vinjé, J., White, P.A., Koopmans, M., 2013.Indications for worldwide increased norovirus activity associated withemergence of a new variant of genotype II.4, late 2012. Euro. Surveill. 18, 8–9.
White, P., Hansman, G.S., Li, A., Dable, J., Isaacs, M., Ferson, M., McIver, C.J.,Rawlinson, W.D., 2002. Norwalk-like virus 95/96-US strain is a major cause ofgastroenteritis outbreaks in Australia. J. Med. Virol. 68, 113–118.
Widdowson, M.A., Cramer, E.H., Hadley, L., Bresee, J.S., Beard, R.S., Bulens, S.N.,Charles, M., Chege, W., Isakbaeva, E., Wright, J.G., Mintz, E., Forney, D., Massey, J.,Glass, R.I., Monroe, S.S., 2004. Outbreaks of acute gastroenteritis on cruise shipsand on land: identification of a predominant circulating strain of norovirus–United States, 2002. J. Infect. Dis. 190, 27–36.
Yan, H., Yagyu, F., Okitsu, S., Nishio, O., Ushijima, H., 2003. Detection of norovirus(GI, GII), Sapovirus and astrovirus in fecal samples using reverse transcriptionsingle-round multiplex PCR. J. Virol. Methods 114, 37–44.
Yen, C., Wikswo, M.E., Lopman, B.A., Vinje, J., Parashar, U.D., Hall, A.J., 2011. Impactof an emergent norovirus variant in 2009 on norovirus outbreak activity in theUnited States. Clin. Infect. Dis. 53, 568–571.
Yoneda, M., Okayama, A., Kitahori, Y., 2014. Epidemiological characteristics ofnorovirus associated with sporadic gastroenteritis among children from the2006/2007–2011/2012 season in Nara prefecture. Jpn. Intervirol. 57, 31–35.
Zhang, X.F., Tan, M., Chhabra, M., Dai, Y.C., Meller, J., Jiang, X., 2013. Inhibition ofhisto-blood group antigen binding as a novel strategy to block norovirusinfections. PLoS ONE 8, e69379.
Zheng, D.P., Widdowson, M.A., Glass, R.I., Vinjé, J., 2010. Molecular epidemiology ofgenogroup II-genotype 4 noroviruses in the United States between 1994 and2006. J. Clin. Microbiol. 48, 168–177.




















