
Modulation of gene expression through chromosomal positioning in Escherichia coli
8
Microbio/ogy (1 997), 143, 207 1-2078 Printed in Great Britain Modulation of gene expression through chromosomal positioning in Escherichia coli Carolina Sousa,t Victor de Lorenzo and Angel Cebollat Author for correspondence: Victor de Lorenzo. Tel: +34 1 5854536. Fax: +34 1 5854506. e-mail: VDLORENZO(wSAMBA.CNB.UAM.ES Centro Nacional de B iotecno log ia, CSI C, Campus de Cantoblanco, 28049 Madrid, Spain Variations in expression of the nah genes of the NAH7 (naphthalene biodegradation) plasmid of Pseudomonas putida when placed in different chromosomal locations in Escherichia coli have been studied by employing a collection of hybrid mini-Tn5 transposons bearing lac2 fusions to the Psal promoter, along with the cognate regulatory gene nahR. Insertions of Psal-lacZ reporters in the proximity of the chromosomal origin of replication, oriC, increased accumulation of jl-galactosidase in vivo. Position-dependent changes in expression of the reporter product could not be associated with local variations of the supercoiling in the DNA region, as revealed by probing the chromosome with mobile gyrB-lacZ elements. Such variations in p- galactosidase activity (and, therefore, the expression of catabolic genes) seemed, instead, to be linked to the increase in gene dosage associated with regions close to oriC, and not to local variations in chromosome structure. The tolerance of strains to the selection markers borne by the transposons also varied in parallel with the changes in Lac2 levels. The role of chromosomal positioning as a mechanism for the outcome of adaptation phenotypes is discussed. ~ Keywords : biodegradation, catabolic operons, positional effects, NahR, gyrB ~ INTRODUCTION Our laboratory is interested in the mechanisms by which G r a m - n ega t i v e b acter i a, in part ic u 1 a r Pseudo m o nas , evolve the ability to employ recalcitrant hydrocarbons as carbon sources (de Lorenzo & Perez-Martin, 1996; van der Meer et al., 1992). A critical step in the success of a new pathway is the acquisition of an adequate expression control system that is responsive to pathway substrates or related metabolic intermediates (de Lorenzo & Perez-Martin, 1996). Generally, promoters are controlled by specific regulators that trigger tran- scription initiation when cells face a certain chemical or physical signal. However, the effective level of ex- pression of a particular gene or operon is subject to the general physiological status of the cells. Such status selects a window of expression for the sake of an adequate energetic return. General mechanisms that influence gene expression in viva include histone- like proteins (Nash, 1996), superhelical density (Liu & t Present address: Centre National de la Recherche Scientifique, lnstitut des Sciences VQcAtales, Avenue de la Terrasse, 91 198 Gif-sur-Yvette, France. Abbreviation : p-Gal, p-galactosidase. Wang, 1987; Menzel & Gellert, 1983) and DNA methylation (Plasterk et al., 1983). These may differ depending on the physiological status or even, as has been suggested, on the chromosomal location (Schmid & Roth, 1987). This last possibility is particularly appealing, since catabolic operons appear frequently within transposons (Wyndham et al., 1994) and could, therefore, employ differential positioning within a replicon as a mechanism for fine-tuning an optimal level of expression. During the course of previous studies (de Lorenzo et al., 1993), we had noticed that the activity of three different catabolic systems of Pseudomonas putida varied signifi- cantly depending on their position in the bacterial chromosome, as revealed through lacZ fusions to relevant promoters. These were: (i) the Pu promoter of the TOL (toluene biodegradation) plasmid pWW0, (ii) the Pm promoter of the TOL plasmid and (iii) the Psal promoter of the NAH7 (naphthalene biodegradation) plasmid. Since very minor variations in the expression of adaptation genes have a major influence on the long- term outcome of a particular strain (Elena et al., 1996; Negri et at., 1994), we were interested in exploring chromosomal positioning as a factor influencing gene 0002-1356 0 1997 SGM 207 1
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Modulation of gene expression through chromosomal positioning in Escherichia coli

Microbio/ogy (1 997), 143, 207 1-2078 Printed in Great Britain
Modulation of gene expression through chromosomal positioning in Escherichia coli
Carolina Sousa,t Victor de Lorenzo and Angel Cebollat
Author for correspondence: Victor de Lorenzo. Tel : +34 1 5854536. Fax: +34 1 5854506. e-mail: VDLORENZO(wSAMBA.CNB.UAM.ES
Centro Nacional de B iotecno log ia, CSI C, Campus de Cantoblanco, 28049 Madrid, Spain
Variations in expression of the nah genes of the NAH7 (naphthalene biodegradation) plasmid of Pseudomonas putida when placed in different chromosomal locations in Escherichia coli have been studied by employing a collection of hybrid mini-Tn5 transposons bearing lac2 fusions to the Psal promoter, along with the cognate regulatory gene nahR. Insertions of Psal-lacZ reporters in the proximity of the chromosomal origin of replication, oriC, increased accumulation of jl-galactosidase in vivo. Position-dependent changes in expression of the reporter product could not be associated with local variations of the supercoiling in the DNA region, as revealed by probing the chromosome with mobile gyrB-lacZ elements. Such variations in p- galactosidase activity (and, therefore, the expression of catabolic genes) seemed, instead, to be linked to the increase in gene dosage associated with regions close to oriC, and not to local variations in chromosome structure. The tolerance of strains to the selection markers borne by the transposons also varied in parallel with the changes in Lac2 levels. The role of chromosomal positioning as a mechanism for the outcome of adaptation phenotypes is discussed.
~ Keywords : biodegradation, catabolic operons, positional effects, NahR, gyrB ~
INTRODUCTION
Our laboratory is interested in the mechanisms by which G r a m - n eg a t i v e b act e r i a, in part i c u 1 a r Pseudo m o nas , evolve the ability to employ recalcitrant hydrocarbons as carbon sources (de Lorenzo & Perez-Martin, 1996; van der Meer et al., 1992). A critical step in the success of a new pathway is the acquisition of an adequate expression control system that is responsive to pathway substrates or related metabolic intermediates (de Lorenzo & Perez-Martin, 1996). Generally, promoters are controlled by specific regulators that trigger tran- scription initiation when cells face a certain chemical or physical signal. However, the effective level of ex- pression of a particular gene or operon is subject to the general physiological status of the cells. Such status selects a window of expression for the sake of an adequate energetic return. General mechanisms that influence gene expression i n viva include histone- like proteins (Nash, 1996), superhelical density (Liu &
t Present address: Centre National de la Recherche Scientifique, lnstitut des Sciences VQcAtales, Avenue de la Terrasse, 91 198 Gif-sur-Yvette, France.
Abbreviation : p-Gal, p-galactosidase.
Wang, 1987; Menzel & Gellert, 1983) and DNA methylation (Plasterk et al., 1983). These may differ depending on the physiological status or even, as has been suggested, on the chromosomal location (Schmid & Roth, 1987). This last possibility is particularly appealing, since catabolic operons appear frequently within transposons (Wyndham et al., 1994) and could, therefore, employ differential positioning within a replicon as a mechanism for fine-tuning an optimal level of expression.
During the course of previous studies (de Lorenzo et al., 1993), we had noticed that the activity of three different catabolic systems of Pseudomonas putida varied signifi- cantly depending on their position in the bacterial chromosome, as revealed through lacZ fusions to relevant promoters. These were: (i) the Pu promoter of the TOL (toluene biodegradation) plasmid pWW0, (i i ) the Pm promoter of the TOL plasmid and (iii) the Psal promoter of the NAH7 (naphthalene biodegradation) plasmid. Since very minor variations in the expression of adaptation genes have a major influence on the long- term outcome of a particular strain (Elena et al., 1996; Negri et at., 1994), we were interested in exploring chromosomal positioning as a factor influencing gene
0002-1356 0 1997 SGM 207 1

C. S O U S A a n d OTHERS
expression. In this report, we present genetic evidence that location modulates operative (but riot intrinsic) catabolic gene expression in vivo by virtue of the distance from the chromosomal replication origin, oriC. Our data suggest that such an influence is predominantly due to the operative increase in gene dosage associated with regions close to oriC, and not becaluse of local variations in DNA supercoiling. Finally, we show that the chromosomal location affects the survival of clonal populations under antibiotic selective pressure, de- pending on the position of the resistance genes.
METHODS
Strains, media and general procedures. Bacteria and plasmids used in this work are listed in Table 1. Cells were grown in Luria broth (LB) or M9-glucose (0.2%) minimal medium (Maniatis et al., 1982) supplemented, where appropriate, with 50 pg rifampicin (Rif) ml-', 50 pg kanamycin (Km) ml-', 150 pg ampicillin (Ap) ml-', 25-50 pg streptomycin (Sm) ml-I or 50 pg X-Gal m1-I. DNA manipulations, including Southern blotting of agarose gels, were carried out according to published protocols (Maniatis e t al., 1982; Sarnbrook et al., 1989). B-Galactosidase (@-Gal) accumulation was determined according to Miller (1972). Determination of thLe MIC of Km and Sm for the Escherichia coli strains indicated was done using the dilution method of Washington & Sutter (1980).
Table 1. Bacteria and plasmids
Design of a hybrid transposon bearing a PgyrB-laczfusion. A genetic probe responsive to differential supercoiling was constructed as follows. Plasmid pIC549 (Table l ) , bearing an Sm selection marker and a transcriptional fusion between the gyrB promoter of E. coli (PgyrB) and the reporter hybrid gene ga1T'-'galK, was used as the source of a 3.9 kb EcoRI-BamHI restriction fragment containing the promoter and part of the adjacent downstream sequence, which was cloned at the same sites of the lacZ vector pUJ9 (de Lorenzo et al., 1990). This gave rise to plasmid pUgyrB, which bears a DNA segment with the sequence PgyrB: :galT'-'ga1K'-'lacZ (in essence, a transcriptional PgyrB-lacZ fusion) as a 7.8 kb NotI fragment. This segment was cloned into the transposon delivery plasmid pUT/mini-TnSKm2 (de Lorenzo et al., 1990) to generate pTgyrB. This plasmid contains a hybrid mini-transposon in which the PgyrB-lacZ fusion is flanked by Km and Sm interposons that shield the reporter system from any read- through transcription from nearby promoters after chromo- somal integration. The organization of the resulting supercoiling-responsive probe is shown in Fig. 1.
Mobilization and transposition. Insertion of hybrid mini- transposons into the chromosome of the K-12 E. coli strain W3110 was done following the procedure described in detail by de Lorenzo & Timmis (1994). Matings between the donor strain E. coli S17-l~pir , transformed with the delivery plasmid indicated in each case, and the recipient strain were run at 30 "C for 2-4 h on the surface of a filter placed on an LB plate supplemented with 1% sodium citrate. The cells were then
S traidplasmid Relevant genotype/phenotype/characteristics Reference/origin
F . coli K12 CC118
CC118ipir S 17-lipir
pCNB4-lacZ
pGP704
FUT
pUTlmini-Tn5 Km2
pICS48 PUP0
A(ara-leu) araD AlacX 74 galE galK phoA thi-1 Herrero et al. (1990)
CC118 lysogenized with l p i r phage Herrero et al. (1990) Tp' Smr, recA thi hsdRA4+ RP4: : 2- de Lorenzo & Timmis
hyA deoC 3,- F- INVLrrnD-rrnE] Alac Daniels (1990) Ap', trp'-'lac2 promoter probe vector de Lorenzo et al. (1990) Ap', lac2 promoter probe vector de Lorenzo et al. (1990) Ap' Km', oriR6K mobRP4, delivery vector for de Lorenzo et al. (1993)
Same as pCNB4, but inserted with a de Lorenzo et al. (1993)
vpsE rpoB argE (Am) rtvA1
Tc : : Mu : : Km Tn7, l p i r phage lysogen (1994)
mini-Tn5 nahR/Psal
promoterless trp'-'lac2 reporter downstream of the Psal promoter
Ap', oriR6K mo6RP4 M13 tg131 polylinker Miller & Mekalanos
Ap', tnp gene of Tn.5-ISSO, inserted in SalI site Herrero et al. (1990)
Ap' Km', RbKoriV RP4oriT mini-Tn5 Km2
Ap' Sm', PgyrB : : galK '-'lacZ Ap', aadA promoter inserted as a 1.1 kb
BQWZHI-BC~I insert in pUJ8 Ap' Sm', pUJS derivative bearing a
PgyrB: :galK'-'lad-Sm' cassette as a NotI insert
vector for mini-Tn5 PgyrB-lacZ
(1988)
of pCP704
transposon-vector delivery plasmid de Lorenzo et a!. (1990)
Macian et al. (1994) This work
This work
Ap' Km' Sm", R6KoriV l<P4oriT, delivery This work
2072

Modulation of gene expression
I
upper operon lower uperon 0
........ 0
AvrIf 'Not1 Not1
Fig. 1. The catabolic NAH system and organization of Psal-lacZ and Pgyr-lacZ transposons. (a) Regulatory features of the naphthalene-degrading pathway of the NAH7 plasmid of f . putida. Upon exposure of the cells t o naphthalene vapour, the basal expression of the upper operon is enough to generate a certain amount of intracellular salicylate. This intermediate binds the product of the regulatory gene nahR which, in turn, further activates the Pnah promoter of the upper operon and the Psal divergent promoter of the lower pathway. The regulatory elements of the pathway included in the mini-Tn5 nahRIfsal-lacZ transposon represented below the scheme are indicated. Note that the segment of the NAH operon included in the mini-transposon maintains the same orientation as the control elements and that nahR i s transcribed through i t s original promoter, f r . The scheme also shows (not to scale) the arrangement of other segments, i.e. a Km-resistance gene, the transcriptional terminator a t the end of the lacZ reporter (symbolized as a stem-loop a t the end of the box) and the I and 0 termini of Tn5. (b) Structure of the mini-Tn5 fgyrf3-lacZ transposon. This mobile element includes a lacZ transcriptional fusion to the PgyrB promoter flanked upstream and downstream by Sm and Km R elements. The Not1 and Awl1 sites within the transposons are indicated.
resuspended in 1% NaCl and plated on M9-glucose plates containing the appropriate antibiotics for selection of chromo- somal transposon insertions and counterselection of the donor strain.
Induction conditions. The Psal promoter in cells bearing insertions of mini-Tn5 nahR/Psnl-lacZ was induced by growing the corresponding cultures in LB medium at 30 "C up to ODGoo 0-4, after which 2 m M salicylate was added. Incubation was then continued for 3 h and the accumulation of P-Gal measured. In order to examine the effects of the gyrase inhibitor coumermycin A1 on expression of the reporter l a c 2 fusions, E . coli cells were grown in LB medium to OD600 0 . 3 4 4 , after which the antibiotic (pre-dissolved in DMSO) was added to a final concentration of 10 pg ml-'. Control cultures received only the corresponding amount of DMSO. Incubation was continued for a further 2 h and the amount of P-galactosidase accumulated in the cells was measured. Similarly, the effect of increased osmolarity was examined by raising the salt concentration of cell cultures grown to OD,,, in LB (85 mM NaCl) to 500 m M NaCl for 2 h.
Pulsed-field gel electrophoresis (PFGE). T o prepare intact DNA in agarose plugs, cells were grown in LB to approxi-
mately 1.5 x 10' c.f.u. ml-'. Cultures were then treated for 1 h with 180 pg chloramphenicol ml-' (Ohki & Smith, 1989), pelleted and resuspended in 1 vol. 1 M NaCI, 10 mM Tris/HCl pH 8. The cells were warmed to 3 0 4 0 "C and 0.5 ml of the suspension was mixed with an equal volume of 1 '/o low-melting-point agarose in sterile water, poured into plug moulds (Bio-Rad) and allowed to cool. Each set of four solidified plugs was incubated overnight at 37 "C in 3 ml EC lysis solution (6 mM Tris/HCl pH 8, 1 M NaC1, 100 mM EDTA pH 8,0.5 '/o Brij 58,0.2 '/o deoxycholate, 0.5 '/o sarkosyl, 1 mg lysozyme ml-' and 20 pg RNase ml-'). The EC solution was then replaced with an equal volume of ESP buffer (0-5 M EDTA pH 9-5, 1 '/o sarkosyl, 1 mg proteinase K ml-'), and the incubation was continued overnight at 50 "C, after which the solution was replaced by fresh ESP buffer and the incubation continued for 1 d. Finally, the plugs were placed for 1 h in TE buffer (10 mM Tris/HCl pH 7.5, 1 m M EDTA pH 7.5) at 50 "C with 40 pg ml-' of the protease inhibitor phenyl- methylsulfonyl fluoride. The resulting samples were stored at 4 "C in 0.5 M EDTA pH 8. For restriction analysis, the plugs were washed thoroughly, first with TE buffer and then with the appropriate enzyme buffer at 4 "C. Digestions were set by placing each plug in 750 pl of the digestion buffer and adding 10-20 units of the required enzyme. Reactions were run for 2-4 h at 37 "C. After complete digestion, the buffer was removed and the plugs were transferred to cold TE buffer (4 "C) for 30 min. Samples were subjected to PFGE using a CHEF-DRII system (Bio-Rad) based on the procedure desc- ribed by Chu et al. (1986). Seaplaque GTG agarose (Bio-Rad) gels were run for 20 h at 200 V in TBE buffer (45 mM Tris/HCl, 45 mM boric acid, 1 mM EDTA pH 7.6) at 9 "C. Fragments of 20-500 kb were separated in 1 '/o agarose gels with a pulse time ramp of 5-40 s. DNA fragments of 100-1200 kb were examined in 0.8% gels with a pulse time ramp of 20-120 s. Chromosomes of Sacharonzyces cereuisiae were used as size markers.
RESULTS AND DISCUSSION
Chromosomal location modulates expression of nah genes of the NAH7 plasmid
This study was initiated by our earlier observation that independent insertions of hybrid mini-transposons bear- ing lac2 fusions to the Pu, Pm and Psal promoters of catabolic plasmids TOL and NAH of P. putida gave rise to different levels of accumulation of P-Gal in response to the pathway inducers m-xylene, m-toluate and salicylate, respectively, when randomly inserted into the chromosome of P. putida strain KT2442 (de Lorenzo et al., 1993). This behaviour could not be attributed to readthrough transcription from nearby external pro- moters, since design of the lacZ-bearing mini-trans- posons employed included strong terminators flanking the reporter system. We therefore examined the basis of such differential expression. For this, we used the salicylate-responsive Psal promoter of the NAH7 plasmid and transferred the regulatory elements into E. coli strain W3110 in order to have a more defined background for genetic analyses.
We started by reproducing the phenomenon in E . coli W3110. For this, we mutagenized this strain with the transposon mini-Tn5 nahR/Psal-lac2 using plasmid pCNB4-lac2 (Table 1) as suicide delivery vector. As
2073
C. S O U S A a n d O T H E R S
Nano. 1 4 8 11 18 20
(a) ' = Non-induced +Salicylate
24 25
Avrll C B/E G A D C B B
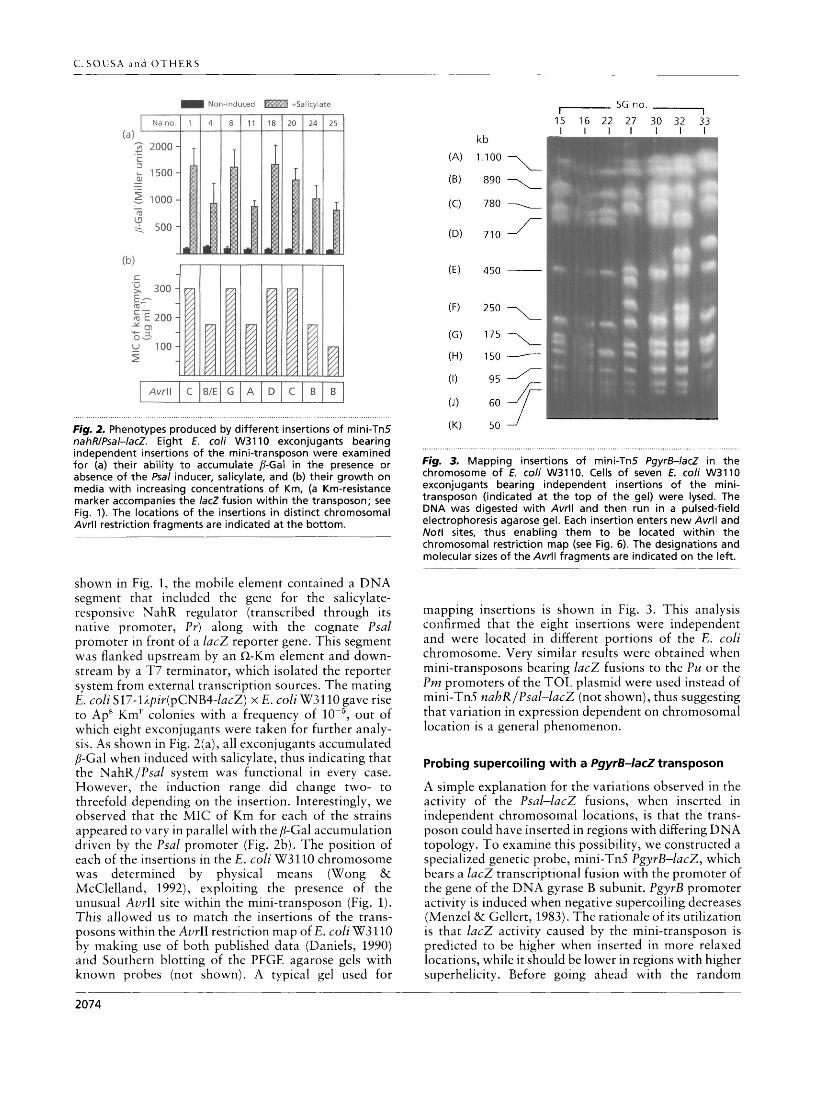
Fig, 2. Phenotypes produced by different insertions of mini-Tn5 nahRIPsal-lacZ. Eight E. coli W3110 exconjugants bearing independent insertions of the mini-transposon were examined for (a) their ability to accumulate P-Gal in the presence or absence of the Psal inducer, salicylate, and (b) their growth on media with increasing concentrations of Km, (a Km-resistance marker accompanies the lac2 fusion within the transposon; see Fig. 1). The locations of the insertions in distinct chromosomal AvrlI restriction fragments are indicated a t the bottom.
kb
1.100
890
780
710
450
250
175
150
95
60
50
I I SG no.
15 16 22 27 30 32 33 I I 1 I l I I
Mappinq insertions of mini-Tn5 Pqyr5-lacZ in the chromosome 'of 2. coli W3110. Cells of seven E. coli W3110 exconjugants bearing independent insertions of the mini- transposon (indicated a t the top of the gel) were lysed. The DNA was digested with Awl1 and then run in a pulsed-field electrophoresis agarose gel. Each insertion enters new Avrll and Not1 sites, thus enabling them to be located within the chromosomal restriction map (see Fig. 6). The designations and molecular sizes of the Avrll fragments are indicated on the left.
shown in Fig. 1, the mobile element contained a DNA segment that included the gene for the salicylate- responsive NahR regulator (transcribed through its native promoter, Pr) along with the cognate Psal promoter in front of a lac2 reporter gene. This segment was flanked upstream by an n-Km element and down- stream by a T7 terminator, which isolated the reporter system from external transcription sources. The mating E . coli S17-lipir(pCNB4-facZ) x E. coli W3110 gave rise to ApS Km" colonies with a frequency of out of which eight exconjugants were taken for further analy- sis. As shown in Fig. 2(a), all exconjugants accumulated /?-Gal when induced with salicylate, thus indicating that the NahRIPsal system was functional in every case. However, the induction range did change two- to threefold depending on the insertion. Interestingly, we observed that the MIC of Km for each of the strains appeared to vary in parallel with the /?-Gal accumulation driven by the Psal promoter (Fig. 2b). The position of each of the insertions in the E . coli W3110 chromosome was determined by physical means (Wong & McClelland, 1992), exploiting the presence of the unusual AvrII site within the mini-transposon (Fig. 1). This allowed us to match the insertions of the trans- posons within the A d 1 restriction map of 12, coli W3110 by making use of both published data (Dlaniels, 1990) arid Southern blotting of the PFGE agarose gels with known probes (not shown). A typical gel used for
mapping insertions is shown in Fig. 3. This analysis confirmed that the eight insertions were independent and were located in different portions of the E. coli chromosome. Very similar results were obtained when mini-transposons bearing lacZ fusions to the Pu or the Pm promoters of the TOL plasmid were used instead of mini-Tn5 nahR/Psal-lac2 (not shown), thus suggesting that variation in expression dependent on chromosomal location is a general phenomenon.
Pro b i ng su pe rco i I i ng with a PgyrB-lacZ trans poso n
A simple explanation for the variations observed in the activity of the Psal-lacZ fusions, when inserted in independent chromosomal locations, is that the trans- poson could have inserted in regions with differing DNA topology. To examine this possibility, we constructed a specialized genetic probe, mini-Tn.5 PgyrB-lad, which bears a lac2 transcriptional fusion with the promoter of the gene of the DNA gyrase B subunit. PgyrB promoter activity is induced when negative supercoiling decreases (Menzel & Gellert, 1983). The rationale of its utilization is that lacZ activity caused by the mini-transposon is predicted to be higher when inserted in more relaxed locations, while it should be lower in regions with higher superhelicity. Before going ahead with the random
2074
Modulation of gene expression
0 Nil +Courn. m + N a C I
2o 1,1 30 300
'5 250
20 .- 2 - 150 2 -
v 10
&
z
h
w h
$-' .- C
aJ L 5 - 200
m g 100
- - .- m
W
X 10 50 m
0 0
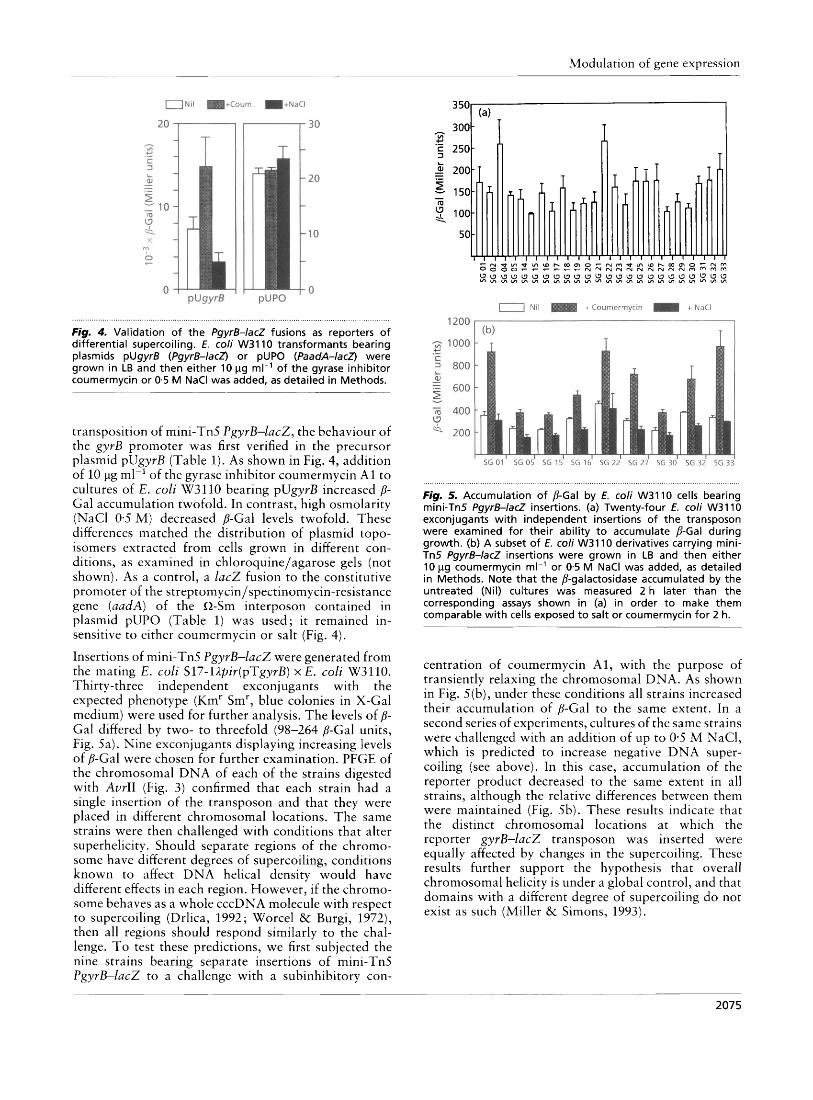
Fig, 4. Validation of the Pgyrf3-lacZ fusions as reporters of differential supercoiling. E. coli W3110 transformants bearing plasmids pUgyrB (PgyrB-lacZ) or pUPO (PaadA-lacZ) were grown in LB and then either 10 pg ml-' of the gyrase inhibitor coumermycin or 0.5 M NaCl was added, as detailed in Methods.
transposition of mini-Tn5 PgyrB-lac2, the behaviour of the gyrB promoter was first verified in the precursor plasmid pUgyrB (Table 1). As shown in Fig. 4, addition of 10 pg ml-' of the gyrase inhibitor coumermycin A1 to cultures of E. coli W3110 bearing pUgyrB increased P- Gal accumulation twofold. In contrast, high osmolarity (NaCl 0.5 M ) decreased P-Gal levels twofold. These differences matched the distribution of plasmid topo- isomers extracted from cells grown in different con- ditions, as examined in chloroquine/agarose gels (not shown). As a control, a lac2 fusion to the constitutive promoter of the streptomycin/spectinomycin-resistance gene (aadA) of the 0-Sm interposon contained in plasmid pUPO (Table 1) was used; it remained in- sensitive to either coumermycin or salt (Fig. 4).
Insertions of mini-Tn5 PgyrB-lac2 were generated from the mating E . coli S17-l@ir(pTgyrB) x E. coli W3110. Thirty-three independent exconjugants with the expected phenotype (Kmr Smr, blue colonies in X-Gal medium) were used for further analysis. The levels of p- Gal differed by two- to threefold (98-264 P-Gal units, Fig. Sa). Nine exconjugants displaying increasing levels of P-Gal were chosen for further examination. PFGE of the chromosomal DNA of each of the strains digested with AvrII (Fig. 3) confirmed that each strain had a single insertion of the transposon and that they were placed in different chromosomal locations. The same strains were then challenged with conditions that alter superhelicity. Should separate regions of the chromo- some have different degrees of supercoiling, conditions known to affect DNA helical density would have different effects in each region. However, if the chromo- some behaves as a whole cccDNA molecule with respect to supercoiling (Drlica, 1992; Worcel & Burgi, 1972), then all regions should respond similarly to the chal- lenge. To test these predictions, we first subjected the nine strains bearing separate insertions of mini-Tn5 PgyrB-lacZ to a challenge with a subinhibitory con-
T I Nil - +Cournerrnycin + NaCl
1200 T
';;; 10001- T T 1 + S
aJ
.- 3 800
600 2 ii 400 13
- v
@k 200
' S G 0 1 ' S G O j SG l j SG 161 S G 2 i S G 2 j S G 3 d S G 3 i S G 3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . , . , . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . , .
Fig. 5. Accumulation of P-Gal by E. coli W3110 cells bearing mini-Tn5 PgyrB-lacZ insertions. (a) Twenty-four E. coli W3110 exconjugants with independent insertions of the transposon were examined for their ability t o accumulate P-Gal during growth. (b) A subset o f E. coli W3110 derivatives carrying mini- Tn5 PgyrB-lacZ insertions were grown in LB and then either 10 pg coumermycin ml-' or 0.5 M NaCl was added, as detailed in Methods. Note that the P-galactosidase accumulated by the untreated (Nil) cultures was measured 2 h later than the corresponding assays shown in (a) in order t o make them comparable with cells exposed to salt or coumermycin for 2 h.
centration of coumermycin Al , with the purpose of transiently relaxing the chromosomal DNA. As shown in Fig. 5(b), under these conditions all strains increased their accumulation of P-Gal to the same extent. In a second series of experiments, cultures of the same strains were challenged with an addition of up to 0-5 M NaC1, which is predicted to increase negative DNA super- coiling (see above). In this case, accumulation of the reporter product decreased to the same extent in all strains, although the relative differences between them were maintained (Fig. 5b). These results indicate that the distinct chromosomal locations at which the rep or t er gy rB-lac2 t r an spo son was inserted were equally affected by changes in the supercoiling. These results further support the hypothesis that overall chromosomal helicity is under a global control, and that domains with a different degree of supercoiling do not exist as such (Miller & Simons, 1993).
2075
C . S O U S A a n d OTHERS
PI J 200
2 150
'z 100 E ," 5 0 -
9 -
C
0
of the E. coli W3110 chromosome is shown lined up with the genetic map. The positions
- of various gene probes (Beck & Bremer, 1980; Boulain e t a/., 1986; Braun-Brenton & - Hofnung, 1986; Daniels, 1990; de Lorenzo et a/., 1990; Hayward eta/ . , 1991; Nakamura & Inouye, 1979; Wee e t a/., 1988; Williams e t
-
Variations in promoter activity depend orr the distance from the chromosomal replication origin
f u r ompA trpBA
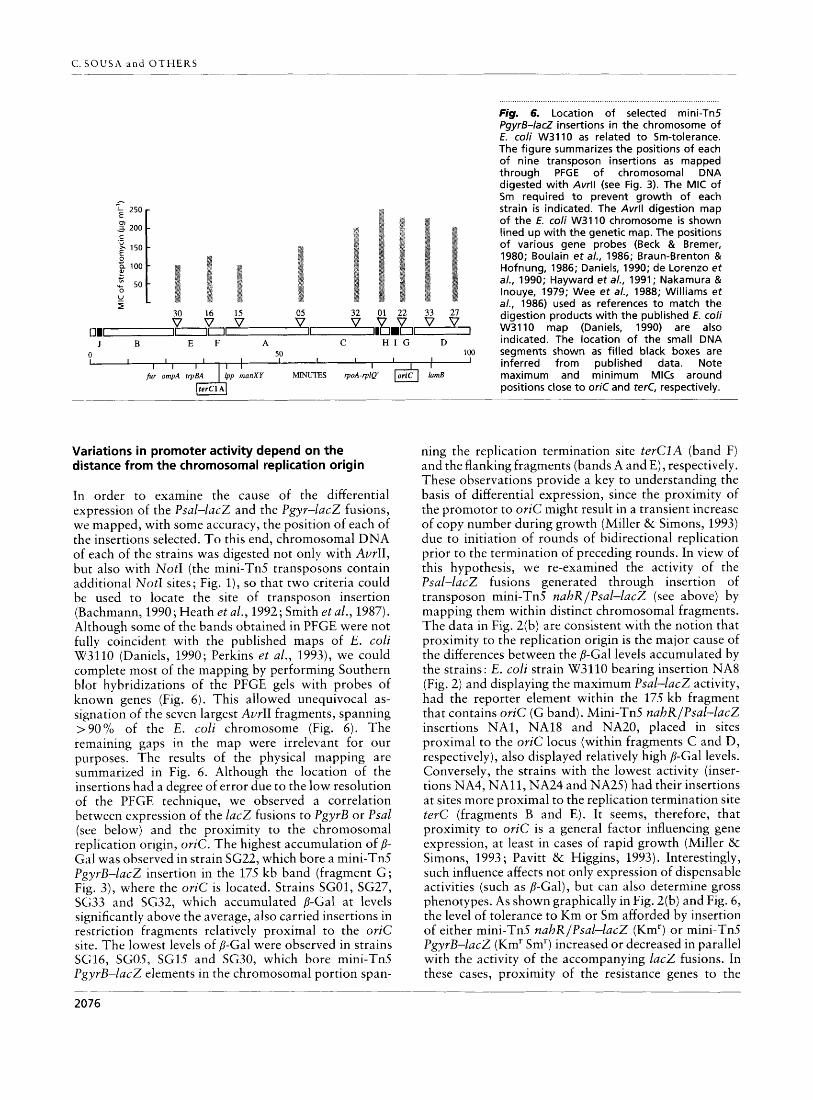
In order to examine the cause of the differential expression of the Psal-lac2 and the Pgyr-lac2 fusions, we mapped, with some accuracy, the position of each of the insertions selected. T o this end, chromosomal DNA of each of the strains was digested not only with AvrII, but also with NotI (the mini-Tn.5 transposons contain additional NotI sites; Fig. 1), so that two criteria could be used to locate the site of transposon insertion (Bachmann, 1990 ; Heath et al., 1992 ; Smith et al., 1987). Although some of the bands obtained in PFGE were not fully coincident with the published maps of E. coli W3110 (Daniels, 1990; Perkins et al., 1993), we could complete most of the mapping by performing Southern blot hybridizations of the PFGE gels with probes of known genes (Fig. 6). This allowed unequivocal as- signation of the seven largest AvrII fragments, spanning >90% of the E. coli chromosome (Fig. 6 ) . The remaining gaps in the map were irrelevant for our purposes. The results of the physical rnapping are summarized in Fig. 6. Although the loc,ation of the insertions had a degree of error due to the lo'w resolution of the PFGE technique, we observed a correlation between expression of the l a c 2 fusions to I'gyrB or Psal (see below) and the proximity to the chromosomal replication origin, oriC. The highest accumulation of p- Gal was observed in strain SG22, which bore a mini-Tn5 PgyrB-lac2 insertion in the 175 kb band (fragment G ; Fig. 3), where the oriC is located. Strains SGO1, SG27, SG33 and SG32, which accumulated D-Gal at levels significantly above the average, also carried insertions in restriction fragments relatively proximal to the oriC site. The lowest levels of P-Gal were observed in strains SG16, SGO.5, SG1.5 and SG30, which bore mini-Tn.5 PgyrB-lac2 elements in the chromosomal portion span-
Ipp manXY MWU'IES rpoA-rplQ' otiC l a d maximum and minimum MlCs around
ning the replication termination site terCZA (band F) and the flanking fragments (bands A and E), respectively. These observations provide a key to understanding the basis of differential expression, since the proximity of the promotor to oriC might result in a transient increase of copy number during growth (Miller & Simons, 1993) due to initiation of rounds of bidirectional replication prior to the termination of preceding rounds. In view of this hypothesis, we re-examined the activity of the Psal-lacZ fusions generated through insertion of transposon mini-Tn5 nahR/Psal-lac2 (see above) by mapping them within distinct chromosomal fragments. The data in Fig. 2(b) are consistent with the notion that proximity to the replication origin is the major cause of the differences between the p-Gal levels accumulated by the strains: E. coli strain W3110 bearing insertion NA8 (Fig. 2) and displaying the maximum Psal-lac2 activity, had the reporter element within the 175 kb fragment that contains oriC (G band). Mini-Tn.5 nahR/Psal-lac2 insertions NA1, NA18 and NA20, placed in sites proximal to the oriC locus (within fragments C and D, respectively), also displayed relatively high p-Gal levels. Conversely, the strains with the lowest activity (inser- tions NA4, NA11, NA24 and NA25) had their insertions at sites more proximal to the replication termination site terC (fragments B and E). I t seems, therefore, that proximity to oriC is a general factor influencing gene expression, at least in cases of rapid growth (Miller & Simons, 1993 ; Pavitt & Higgins, 1993). Interestingly, such influence affects not only expression of dispensable activities (such as P-Gal), but can also determine gross phenotypes. As shown graphically in Fig. 2(b) and Fig. 6, the level of tolerance to Km or Sm afforded by insertion of either mini-Tn5 nahR/Psal-lacZ (Kmr) or mini-Tn.5 PgyrB-lacZ (Kmr Smr) increased or decreased in parallel with the activity of the accompanying lacZ fusions. In these cases, proximity of the resistance genes to the
2076
. . . . . . . . . . . . . . . . . . . . . . , . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . . Fig. 6. Location of selected mini-Tn5 PgyrS-lacZ insertions in the chromosome of E. coli W3110 as related t o Sm-tolerance. The figure summarizes the positions of each of nine transposon insertions as mapped through PFGE of chromosomal DNA digested with Avrll (see Fig. 3). The MIC of Sm required t o prevent growth of each strain is indicated. The Avrll digestion map of the E. coli W3110 chromosome is shown lined up with the genetic map. The positions of various gene probes (Beck & Bremer, 1980; Boulain e t a/., 1986; Braun-Brenton & Hofnung, 1986; Daniels, 1990; de Lorenzo et a/., 1990; Hayward e t a / . , 1991; Nakamura & Inouye, 1979; Wee e t a/., 1988; Williams e t a/., 1986) used as references t o match the digestion products with the published E. coii W3110 map (Daniels, 1990) are also indicated. The location of the small DNA segments shown as filled black boxes are inferred from published data. Note maximum and minimum MlCs around positions close t o oriC and terC, respectively.

Modulation of gene expression
replication origin determined production of a phenotype required for s u rv i v a 1.
Modulation of gene expression through chromosomal positioning ?
There is increasing evidence that building of a higher- order chromosomal structure is basically a stochastic process (Higgins et al., 1996). Although barriers to supercoil diffusion exist (Worcel & Burgi, 1972), they seem to vary in position from cell t o cell or within one cell over time (Higgins et al., 1996). O n this basis, it seems that variable expression of differently positioned genes is unrelated to the responsiveness of the promoters to supercoiling. In fact, it seems that both strong and weak promoters are stochastically distributed through- out the entire chromosome (Chuang et al., 1993), which argues against the existence of defined silent or active chromosomal domains in bacteria. However, genes placed in the proximity of the replication origin are systematically expressed at higher levels (Miller & Simons, 1993 ; Pavitt & Higgins, 1993 ; this work). This may happen because of a transient increase in copy number during growth. Although the differences ob- served are relatively small, they may become significant for organisms adapting to a harsh environment (e.g. antibiotic resistance or growth on unusual carbon sources). At least in the case of antibiotics, it seems that minimal variations in MICs determine the outcome of resistant strains in the presence of subinhibitory con- centrations of the selective agent (Negri et al., 1994). By the same token, it is possible that small variations in the expression of catabolic operons contribute to the success of a certain pathway in non-extreme selection con- ditions. In this context, our results suggest that chromo- somal positioning might be one more mechanism of adaptation to changing environments by modulating the degree of gene expression to an optimal window of activity.
ACKNOWLEDGEMENTS
This work was funded by the ENV4-CT95-0141 (ENVIRON- MENT) contract of the European Union and by grant BI095- 0788 of the Comision Interministerial de Ciencia y Tecnologia, Spain. T h e authors are indebted t o Ernest0 Garcia (Centro de Investigaciones Biologicas del CSIC, Madrid) for his help with the pulsed-field electrophoresis procedures.
REFERENCES
Bachmann, B. (1990). Linkage map of Escherichra colt K-12, edition 8. Microbiol Reu 54, 130-197. Beck, E. & Bremer, E. (1980). Nucleotide sequence of the gene ompA for the outer membrane protein I1 of Escherichra coli. Nuclerc Acids Res 8, 301 1-3027. Boulain, 1. C., Charbit, A. & Hofnung, M. (1986). Mutagenesis by random linker insertion into the IamB gene of Eschertchra colr K-12. Mol Geir Genet 205, 339-348. Braun-Brenton, C. & Hofnung, M. (1986). In urvo and zn i’itro functional alterations of the bacteriophage lambda receptor in lamB missense mutants of Escherrchta coli K12. J Bacteriol 148, 845-852.
Chu, G., Vollrath, D. & Davis, R. W. (1986). Separation of large DNA molecules by contour-clamped homogeneous electric fields. Science 234, 1582-1585. Chuang, S., Daniels, D. L. & Blattner, F. R. (1993). Global regulation of gene expression in Escherichia coli. Bacteriol 175,
Daniels, D. L. (1990). The complete AvrII restriction map of the E . coli genome and comparisons of several laboratory strains. Nucfeic Acids Res 18, 2649-2651. Drlica, K. (1992). Control of bacterial supercoiling. Mol Microbiol 6,425433. Elena, S., Cooper, V. 5. & Lenski, R. (1996). Punctuated evolution caused by selection of rare beneficial mutations. Science 272,
Hayward, R. S., Igarashi, K. & Ishihama, A. (1991). Functional specialization within the a-subunit of Escherichia coli RNA polymerase. J Mol Biol 221, 23-29. Heath, J. D., Perkins, J. D., Sharma, B. & Weinstock, G. M. (1992). Not1 genomic cleavage map of Escherichia coli K-12 strain MG1655. J Bacteriol 174, 558-567. Herrero, M., de Lorenzo, V. & Timmis. K. T. (1990). Transposon vectors containing non-antibiotic selection markers for cloning and stable chromosomal insertion of foreign DNA in Gram- negative bacteria. J Bacteriol 172, 6557-6567. Higgins, N. P., Yang, X., Fu, Q. & Roth, J. (1996). Surveying a supercoil domain by using the yb resolution system in Salmonella typhimurium. ] Bacteriol 178, 2825-2835. Liu, L. & Wang, J. C. (1987). Supercoiling of the DNA template during transcription. Proc Natf Acad Sci U S A 84, 7024-7027. de Lorenzo, V. & P&ez-Martln, J. (1996). Regulatory noise in prokaryotic promoters: how bacteria learn to respond to novel environmental signals. Mol Microbiof 19, 1177-1 184. de Lorenzo, V. & Timmis, K. N. (1994). Analysis and construction of stable phenotypes in Gram-negative bacteria with TnS- and TnlO-derived minitransposons. Methods Enzymol235, 386405. de Lorenzo, V., Herrero, M., Jakubzik, U. & Timmis, K. N. (1990). Mini-TnS transposon derivatives for insertion mutagenesis, promoter probing, and chromosomal insertion of cloned DNA in Gram negative eubacteria. J Bacteriol 172, 65684572. de Lorenzo, V., Ferndndez, S., Herrero, M., Jakubzik, U. & Timmis, K. N. (1 993). Engineering of alkyl- and haloaromatic-responsive gene expression with mini-transposons containing regulated promoters of biodegradative pathways of Pseudomoms. Gene 130, 4146. MaciAn, F., Perez-Roger, 1. & Arrnengod, M. E. (1994). An improved vector system for constructing lacZ fusions : analysis of regulation of the dnaA, dnaN, recF and gyrB genes of Escherichia coli. Gene 145, 17-24. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982). Molecular Cloning: a Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. van der Meer, 1. R., de Vos, W., Harayama, 5. & Zehnder, A. (1992). Molecular mechanisms of bacterial adaptation to xeno- biotic compounds. Microbiol Rev 56, 677-694. Menzel, R. & Gellert, M. (1983). Regulation of the genes for E. coli DNA gyrase : homeostatic control of DNA supercoiling. Cell 34,
Miller, J. H. (1972). Experiments in Molecular Genetics. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. Miller, V. L. & Mekalanos, J. (1988). A novel suicide vector and its use in construction of insertion mutations : osmoregulation of
2026-2036.
1802-1804.
105-1 13.
2077

C. S O U S A a n d OTHERS
outer membrane proteins and virulence determinants in Vibrio cholerae requires toxR. ] Bacteriol 170, 2575-2583. Miller, W. G. & Simons, R. (1993). Chromosomal supercoiling in Escherichia coli. Mol Microbiol 10, 675-684. Nakamura, K. & Inouye, M. (1979). DNA sequence of the gene for the outer membrane lipoprotein of E . coli: an extremely AT-rich promoter. Cell 18, 1109-1 117. Nash, H. A. (1996). The E. coli HU and IHF proteins: accessory factors for complex protein-DNA assemblies. In Regulation of Gene Expression in Escherichia coli, pp. 149-179. Edited by E. C. C. Lin & A. S. Lynch. Bethesda, M D : RG Landes Co. Negri, M. C., Morosini, M. I., Loza, E. & Baquero, F. (1994). In vitro selective antibiotic concentrations of p-lactams for penicillin resistant Streptococcus pneumoniae populations. Antimicrob Agents Chemother 38, 122-125. Ohki, M. & Smith, C. L. (1989). Tracking bacterial replication forks in vivo by pulsed field gel electrophoresis. Nucleic Acids Res
Pavitt, G. D. & Higgins, C. F. (1993). Chromosomal domains of supercoiling in Salmonella typhimurium. Mol Microbiol 10, 68.5-696. Plasterk. R. H., Vrieling, H. & Van de Putte, P. (1983). Trans- cription initiation of Mu mom depends on niethylation of the promoter region and a phage-coded transactivator. Nature 301, 344-347. Perkins, 1. D., Heath, J., Sharma, B. R. & Weinstock, G. M. (1993). XbaI and Blnl genomic cleavage maps of Escherichia cofi K-12 strain MG165.5 and comparative analysis of other strains. J Mol Biol232, 419445. Sambrook, 1.. Fritsch, E. F. & Maniatis, T. (1989). Molecular
17, 3479-3490.
Cloning: a Laboratory Manual, 2nd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. Schmid, M. & Roth, 1. (1987). Gene location affects expression level in Salmonella typhimurium. ] Bacteriol 169, 2872-2875. Smith, C. L., Econome, J., Schutt,A., Klco, S. & Cantor, C. R. (1987). A physical map of the Escherichia coli K-12 genome. Science 236,
Washington, J. A. & Sutter, V. (1980). Dilution susceptibility test: agar and macro-broth dilution procedures. In Manual of Clinical Microbiology, 3rd edn, pp. 453458. Edited by R. H. Lennette, A. Balows, W. J. Hausler, Jr & J. P. Truant. Washington, DC: American Society for Microbiology. Wee, S., Neilands, J., Bittner, M., Hemming, B., Haymore, B. & Seetharam, R. (1 988). Expression, isolation and properties of Fur (ferric uptake regulation) protein of Escherichia coli K-12. Biol Metals 1, 62-68. Williams, N., Fox, D., Shea, C. & Roseman, 5. (1986). Pel, the protein that permits DNA penetration of Escherichia coli, is encoded by a gene in ptsM and is required for mannose utilization by the phosphotransferase system. Proc Natl Acad Sci USA 83, 8934-893 8. Wong, K. K. & McClelland, M. (1992). Dissection of the Salmonella typhimurium genome by use of a Tn5 derivative carrying rare restriction sites. J Bacteriol 174, 3807-381 1. Worcel, A. & Burgi, E. (1972). O n the structure of the folded chromosome of Escherichia coli.
Wyndham, R. C., Cashore, A. E., Nakatsu, C. H. & Peel, M. (1994). Catabolic transposons. Biodegradation 5, 323-342.
1448-1453.
Mol Biol71, 127-147.
Received 14 October 1996; revised 3 January 1997; accepted 6 February 1997.
2078




















