
Mutants of Escherichia coli - NCBI
11
JOURNAL OF BACrERIOLOGY, Mar. 1974, p. 1082-1092 Copyright i 1974 American Society for Microbiology Vol. 117. No. 3 Printed in U.S.A. Genetic Analysis of Cold-Sensitive Ribosome Maturation Mutants of Escherichia coli ROBERT E. BRYANT AND PAUL S. SYPHERD Department of Medical Microbiology, College of Medicine, University of California, Irvine, California 92664 Received for publication 3 December 1973 Four cold-sensitive mutants of Escherichia coli, which have defects in the maturation of the 50S ribosomal subunit, were isolated. Each of the mutations was shown to map at a different locus. The loci were assigned the name rim (ribosome maturation) and were shown to map as follows: rimA is co-transduced with ilvD and with pyrE; rimB is co-transduced with aroD; conjugation experiments limited rimD to a region between ilv and malB, and conjugation experiments limited rimC to the 22 to 30 min region of the chromosome. In merodiploids heterozygous for rimA, rimB, or rimD, the wild-type allele was shown to be dominant to the mutant allele. The observation that the rim loci lie outside the strA region and separate from each other, as well as the recessive character of the rim loci, suggests that the mutants may be defective in ribosome maturation factors rather than being defective in ribosomal structural proteins. The study of ribosome assembly in bacteria has been pursued by a variety of techniques, such as pulse labeling of cells, treatment of cells with chloramphenicol, production of "relaxed" particles, and mutant analysis (for recent re- views, see references 8, 19, 27). The genetic approach to investigation of ribosome matura- tion was limited until the development of a means of isolating a large number of ribosome assembly mutants. Guthrie, Nashimoto, and Nomura (12) and Tai, Kessler, and Ingraham (33) found that a large percentage of cold-sensi- tive mutants have conditional blocks in ribo- somal subunit assembly. Many cold-sensitive mutations reported thus far are closely linked to strA and may involve ribosomal proteins (12, 33, 38). Our interest in such conditional mutants was the possibility that nonstructural components might be involved in ribosome biosynthesis and that these components might be revealed by certain mutations. We report here the mapping of four cold-sensitive, ribosome assembly mu- tants of Escherichia coli. These four loci are shown to lie outside the strA region and sepa- rate from each other. The existence of these loci suggests that the mutants may be defective in ribosome maturation factors rather than defec- tive in ribosomal structural proteins. (This work was submitted to the Department of Microbiol- ogy, University of Illinois, by R. E. B. in partial fulfillment of the requirements of the Ph.D. degree.) MATERIALS AND METHODS Strains. Information pertaining to the strains used is shown in Table 1. In describing new strains, the recommendations of Demerec et al. (9) are followed. The symbols and map positions for genetic loci are those compiled by Taylor and Trotter (35). Pheno- types are indicated by nonitalicized symbols begin- ning with a capital letter. The terms Cld-S and Cld-R refer to the phenotypic growth responses of cold sensitivity and cold resistance (wild type), respec- tively. The term rim (ribosome maturation) denotes a genotypic determinant for ribosome maturation. Media. Stock cultures, except strains bearing epi- somes, were maintained frozen at -20 C in L broth containing 20% glycerol. For episomal strains, selec- tive liquid medium was substituted for L broth. "Selective" medium consisted of EC basal salts (31) supplemented with only those nutrients required for growth of a given strain, with 0.5% D-glucose (wt/vol) as the carbon source. Amino acids were used in the L-form at a concentration of 25 ug/ml. Nucleosides were also used at 25 tsg/ml. Vitamins and cofactors were used at 10 ,g/ml. Amino acids, nucleosides, and vitamins were obtained from Sigma or from Nutrional Biochemicals. Difco special agar-Noble was used for selective medium at a concentration of 1.5% (wt/vol). Dihydrostreptomycin sulfate (Squibb), when present, was at a concentration of 200 Ag/ml. The medium used for mutagenesis and isolation of mutants was ECTNV, which is composed of EC basal salts, tryptone (Difco) at 1% (wt/vol), nucleosides, and a vitamin mixture. The nucleosides were guano- sine, adenosine, uridine, thymidine, and cytidine at concentrations of 25 gg/ml. The stock vitamin mix- ture had the following composition per 100 ml of solution: biotin, 0.2 mg; folic acid, 0.2 mg; pyridoxine 1082
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Mutants of Escherichia coli - NCBI

JOURNAL OF BACrERIOLOGY, Mar. 1974, p. 1082-1092Copyright i 1974 American Society for Microbiology
Vol. 117. No. 3Printed in U.S.A.
Genetic Analysis of Cold-Sensitive Ribosome MaturationMutants of Escherichia coliROBERT E. BRYANT AND PAUL S. SYPHERD
Department of Medical Microbiology, College of Medicine, University of California, Irvine, California 92664
Received for publication 3 December 1973
Four cold-sensitive mutants of Escherichia coli, which have defects in thematuration of the 50S ribosomal subunit, were isolated. Each of the mutationswas shown to map at a different locus. The loci were assigned the name rim(ribosome maturation) and were shown to map as follows: rimA is co-transducedwith ilvD and with pyrE; rimB is co-transduced with aroD; conjugationexperiments limited rimD to a region between ilv and malB, and conjugationexperiments limited rimC to the 22 to 30 min region of the chromosome. Inmerodiploids heterozygous for rimA, rimB, or rimD, the wild-type allele was
shown to be dominant to the mutant allele. The observation that the rim loci lieoutside the strA region and separate from each other, as well as the recessivecharacter of the rim loci, suggests that the mutants may be defective in ribosomematuration factors rather than being defective in ribosomal structural proteins.
The study of ribosome assembly in bacteriahas been pursued by a variety of techniques,such as pulse labeling of cells, treatment of cellswith chloramphenicol, production of "relaxed"particles, and mutant analysis (for recent re-views, see references 8, 19, 27). The geneticapproach to investigation of ribosome matura-tion was limited until the development of ameans of isolating a large number of ribosomeassembly mutants. Guthrie, Nashimoto, andNomura (12) and Tai, Kessler, and Ingraham(33) found that a large percentage of cold-sensi-tive mutants have conditional blocks in ribo-somal subunit assembly. Many cold-sensitivemutations reported thus far are closely linked tostrA and may involve ribosomal proteins (12,33, 38).Our interest in such conditional mutants was
the possibility that nonstructural componentsmight be involved in ribosome biosynthesis andthat these components might be revealed bycertain mutations. We report here the mappingof four cold-sensitive, ribosome assembly mu-tants of Escherichia coli. These four loci areshown to lie outside the strA region and sepa-rate from each other. The existence of these locisuggests that the mutants may be defective inribosome maturation factors rather than defec-tive in ribosomal structural proteins. (This workwas submitted to the Department of Microbiol-ogy, University of Illinois, by R. E. B. in partialfulfillment of the requirements of the Ph.D.degree.)
MATERIALS AND METHODSStrains. Information pertaining to the strains used
is shown in Table 1. In describing new strains, therecommendations of Demerec et al. (9) are followed.The symbols and map positions for genetic loci arethose compiled by Taylor and Trotter (35). Pheno-types are indicated by nonitalicized symbols begin-ning with a capital letter. The terms Cld-S and Cld-Rrefer to the phenotypic growth responses of coldsensitivity and cold resistance (wild type), respec-tively. The term rim (ribosome maturation) denotes agenotypic determinant for ribosome maturation.
Media. Stock cultures, except strains bearing epi-somes, were maintained frozen at -20 C in L brothcontaining 20% glycerol. For episomal strains, selec-tive liquid medium was substituted for L broth."Selective" medium consisted of EC basal salts (31)supplemented with only those nutrients required forgrowth of a given strain, with 0.5% D-glucose (wt/vol)as the carbon source. Amino acids were used in theL-form at a concentration of 25 ug/ml. Nucleosideswere also used at 25 tsg/ml. Vitamins and cofactorswere used at 10 ,g/ml. Amino acids, nucleosides, andvitamins were obtained from Sigma or from NutrionalBiochemicals. Difco special agar-Noble was used forselective medium at a concentration of 1.5% (wt/vol).Dihydrostreptomycin sulfate (Squibb), when present,was at a concentration of 200 Ag/ml.The medium used for mutagenesis and isolation of
mutants was ECTNV, which is composed of EC basalsalts, tryptone (Difco) at 1% (wt/vol), nucleosides,and a vitamin mixture. The nucleosides were guano-sine, adenosine, uridine, thymidine, and cytidine atconcentrations of 25 gg/ml. The stock vitamin mix-ture had the following composition per 100 ml ofsolution: biotin, 0.2 mg; folic acid, 0.2 mg; pyridoxine
1082

GENETICS OF RIBOSOME MUTANTS
TABLE 1. Bacterial strains
Strain no. J Genotype Source f Strain no. ] Genotype ] Source
F- strainsAT 722
AB 1472
AB 744
X 461
JC 1553
x 474
JC 355
W 2961
AB 2834AB 1360
AB 470
AT 3143
AB 3294
AB 2463
Shigella 16
argG pyrE thi str malArbs
ilvD metB argH thi spcgal malA lacY X-/Ar
lac xvl thi argC his ilvproB str
ara leu lacZ proA purEtrp lys str xyl mtlmetE thi T, ,rT,r
argG his metB str' leulac malA xyl mtlrecA
Same as X462 exceptproB
leu lac gal his malAargG metB str
thr leu proB lacY gaixyl mtl thi str
aroE mal thipro lac gal his arg thiaroD str
thi his purB proA mtlxyl galK lacY rel?
pyrC ilv met his purEproC pdxC cyc xyllac Y str tsx rel?
thi ilvC argE proA hispabB lacY mtl xyl strtsx fer sup-37?
thi argE his proA mtixyl ara galK tsx strsup-37 recA
Unknown
A. L. Taylor
S. Kaplan
D. Apirion
R. Curtiss
B. Low
R. Curtiss
41 A. J. Clark
E. A. Adelberg
CGSCaA. J. Clark
CGSC
CGSC
CGSC
S. Kaplan
H. Moyed
Hfr strainsAB 3311
A 19
JC 5088
X 879
X 895
AT 3141
KL 25
KL 209
F' strainsKLF11/JC 1553
F' 148/AB2463
met thi streO ... ilv ...leu
xylmet rnsA rel A+O ... pil ... lac
purAilu thr thi recAO ... lysA ... hisserA
Prototrophic Stra T.8O... purE... .leu ...
galPrototrophic Str" T,6O ... his ... lacpurF
thi argD proA thr leustrO ... argG .ilv ...
lysAsupEO ... ilv ... leu ...
xylmalB sup-53 Xr/X-O ... malB ...ilv ...
leu
pyrE+ ilv+ metB+argH+ malB+: argGhis metB leu lacmalA xyl mtl recA str
PhenotypeHis-Leu-Arg-
his+ aroD+: thi argE hisproA mtl xyl aragalK tsx str sup-37recA
Phenotype ThiArg- Pro-
a Coli Genetic Stock Center, Yale University, Barbara Bachmann, Curator.
hydrochloride, 1.0 mg; thiamine hydrochloride, 0.5mg; riboflavine, 0.5 mg; niacin, 0.5 mg; calciumpantothenate, 0.5 mg; cyanocobalamine, 0.33 mg;
p-aminobenzoic acid, 0.5 mg. This stock solution was
diluted 1:100 into growth medium.A medium containing morpholinopropansulfonic
acid (MOPS) was used for experiments where 32Pincorporation was involved. MOPS was obtained fromCalbiochem. MOPS medium was constituted as fol-lows (per 2.5 liters of solution): MOPS, 50 g; KCL, 5g; NH4Cl, 5 g; Na2SO4, 0.125 g; Na2HPO4, 0.10 g;
MgCl2, 1.25 g; adjusted to pH 7.2 with 10 M NaOH.MOPS medium was supplemented with 0.5% (wt/vol)casein enzymatic hydrolysate (Difco), 10 gg of thia-mine per ml when required, and 0.5% glucose (wt/vol).
Isolation of Cld-S mnutants. Cold-sensitive mu-
tants were isolated from AB 1472 by mutagenesis witheither N-methyl-N'-nitro-N-nitrosoguanidine (NTG)or by ethylmethane sulfonate (EMS), followed bypenicillin counterselection at 20 C. Procedures havebeen described for mutagenesis with NTG (1), EMS(22), and for penicillin counterselection (7). NTG was
purchased from Aldrich Chemical Co., and EMS was
purchased from Eastman Kodak Co. Penicillin was
obtained from Squibb. Penicillin counterselection was
not used in the isolation of strain PS 1583. After thepenicillin treatment, washed cells were plated atvarious dilutions on ECTNV agar plates and incu-bated at 37 C. After colonies appeared, replica plateswere made for incubation at 20 C. Those coloniesfailing to grow after 2 to 3 days at 20 C were thenretested for cold sensitivity by inoculating duplicateECTNV plates for incubation at 37 C and 20 C,respectively.
Strain PS 1518 was found to have acquired Pro-and His- mutations in addition to Cld-S. Strains PS1537 and PS 1500 acquired Pro- and Cld-S mutations.Strain PS 1583 did not acquire any auxotrophy inaddition to that of the parental strain.
Test for ribosomal maturation defect. A strain tobe tested for defective ribosome maturation was
grown at 37 C with aeration in 10 ml of supplementedMOPS medium in the presence of 0.1 to 0.5 MCi of[5-3H]uracil per ml (obtained from Schwarz-Mann;18 Ci/mmol) with unlabeled uracil carrier at 0.5
VOL. 117, 1974 1083
A. J. Clark
Gesteland
A. J. Clark
R. Curtiss
R. Curtiss
CGSC
B. Low via
D. WulffB. Low viaD. Wulff
CGSC
S. Kaplan

BRYANT AND SYPHERD
jg/ml. The culture was grown for two to threegenerations to a cell density of 4 x 10' to 6 x 10'cells/ml. The culture was then shifted to 20 C. Ten totwenty minutes after the shift from 37 C to 20 C,H332PO4 (International Chemical and Nuclear Corp.)was added to make 0.2 to 1.0 MCi/ml. When Klett-Summerson colorimeter readings had increased from25 to 50%, the culture was harvested.Uptake of [3H]uracil was complete within one
generation. Uptake of 32p continued throughout 20 Cgrowth. Uptake of isotopes was measured by precipi-tation of samples from a culture in 5% trhchloroaceticacid. The precipitated cells were filtered ontoSchleicher and Schuell B-6 membrane filters. Theradioactivity on the dried filters was measured with aNuclear-Chicago Mark II liquid scintillation counter.Settings were made so that less than 1% crossover of32P into the 3H channel was observed. No correctionfor this minimal crossover was made.
Cultures were harvested by pouring them over anequal volume of crushed ice. The ice was composed of100 Ag of lysozyme per ml (obtained from Calbio-chem), 20% sucrose (wt/vol), 10 mM tris(hydroxy-methyl)aminomethane (Tris) (adjusted to pH 7.6 atroom temperature with HCI), and 0.1 mM MgCl2.The cells were centrifuged at low speed, the superna-tant was decanted, and the pellets were frozen. Cellextracts were prepared by thawing and resuspendingthe cells in 10 mM Tris, 0.1 mM MgCl2 (TM4) buffercontaining 10 ig of pancreatic deoxyribonuclease perml (electrophoretically pure, obtained from Worth-ington) and 0.2 to 0.5% LOC. LOC is a nonionicdetergent produced by Amway Corp., Ada, Mich.Cells lyse upon resuspension. This procedure of har-vesting cells and preparing extracts is a modificationof a procedure described by Anderson (Ph.D. Thesis,University of Illinois, Urbana, 1969).
Extracts, after being centrifuged at 10,000 x g for10 min to remove cell debris, were layered on a 5 to20% linear sucrose gradient (wt/vol) in TM4 bufferand centrifuged using the Beckman SW27 rotor withSW27.1 buckets or with the SW50.1 rotor. Thegradients were fractionated by collecting drops fromthe bottom of the tube. To each gradient fraction wasadded 0.15 mg of bovine serum albumin, Fraction Vpowder (Miles Laboratories), and the fractions wereprecipitated in 5% trichloroacetic acid in a finalvolume of 2 ml. The fractions were filtered usingReeve Angel glass fiber filters (grade 934 AH) anddried. Radioactivity was measured as describedabove.
Conjugation methods. Matings of Hfr and F-strains were carried out as described by Curtiss et al.(6), except that Hfr strains were shaken at 200 rpm toprovide aeration until the time of mating. Matingmixtures were diluted into a flask containing L brothand rotated at 30 to 80 rpm at 37 C. The ratio of Hfr toF - cells was 1: 10 to 1:20 with a total cell density of 2x 10' to 4 x 108/ml. Where matings were interrupted,interruption was accomplished by shaking a 1:10dilution of the mating mixture in 2.0 ml of EC saltsmedium in an 18-mm test tube containing 10 glassbeads, using a Vortex shaker for 30 s. Episomaltransfers were carried out in the same manner as forHfr matings except that the F' to F- ratio was 10:1 as
suggested by Low (23). Volumes of mating mixturevaried from 10 to 100 ml. Times of mating forepisomal strains varied from 1.5 to 5 h. All conjuga-tion experiments included control plates of donor andrecipient cells, respectively.The rapid mapping method of Low (23) was used
for the preliminary mapping of two Cld-S mutants.Mating plates were sprayed with streptomycin after10 min of incubation at 37 C so that only Hfr strainshaving points of origin proximal to the Cld-S muta-tion gave rise to recombinants. Selection was made forCld-R by incubation of L-agar mating plates at 2' Cfor 1 to 2 days.
Transduction methods. For the preparation andtitration of P1 lysates, the methods of Lennox (21)were used with the exceptions noted below. The P1transducing phage was the strain Plbt described inHashimoto (14) and was a gift of H. Moyed. Shigella16 was used as an indicator strain for titrations of P1lysates. Transductions were carried out by suspendingrecipient cells in log phase in phage buffer (4) and P1lysate to give a final suspension of 2 x 109 cells and1010 phage particles in a volume of 1 ml with a CaCl,concentration of 5 mM. This suspension was incu-bated for 20 min at 37 C and then plated on selectivemedia. Control plates of the P1 lysate and of therecipient were always included.
Scoring for Cld-S and Cld-R recombinants. Acolony to be tested for Cld-S was first inoculated into1 ml of liquid selective medium in a 13-mm culturetube, incubated at 37 C overnight, and then inocu-lated with a sterile toothpick onto a nutrient agarplate for 20 C incubation and onto a nutrient agar orselective agar plate for 37 C incubation as a control.This procedure serves to dilute the inoculum and toinsure that approximately the same amount of inocu-lum is used when many colonies are to be tested. Coldsensitivity is defined as the failure to grow on nutrientagar after two days of incubation at 20 C, providedgrowth at 37 C occurs, when tested by this procedure.Controls consisting of known Cld-S and known Cld-Rstrains were always included in any Cld-S test ofisolates.
Preparation of recA strains. To carry out domi-nance tests for cold-sensitive loci, the recA locus wasintroduced into Cld-S mutants. The first step was toisolate a thyA derivative of a given Cld-S strain bytrimethoprim enrichment (25, 28). Strain JC 5088,which is an Hfr strain bearing recA, was used totransfer recA to the Cld-S mutant. Selection wasmade for thyA+. Recombinants were then scored forsensitivity to ultraviolet (UV) radiation using a GEgermicidal lamp under conditions where recA + strainssuffer a 50% loss of viability but recA strains areapproximately 99.9% killed (3). Scoring was simpli-fied by replica-plating a nutrient agar master-plate ofcolonies to be tested to a second plate. The secondplate was UV irradiated. Colonies which failed to growafter 2 days of incubation at 37 C were retested for UVsensitivity by constructing a kill curve for them, withthe parental Cld-S (recA+) strain and strain PS 1062(recA) as controls.
Preparation of rnsA strains. Cld-S strains weremade ribonuclease (RNase) I deficient by mating withstrain A 19, which is an Hfr strain bearing rnsA.
1084 J. BACTERIOL.
GENETICS OF RIBOSOME MUTANTS
Selection was made for Pro+, Gal+. RNase I-deficientrecombinants were detected by testing for the inabil-ity to degrade yeast ribonucleic acid (RNA) in an agarplate.
RESULTS
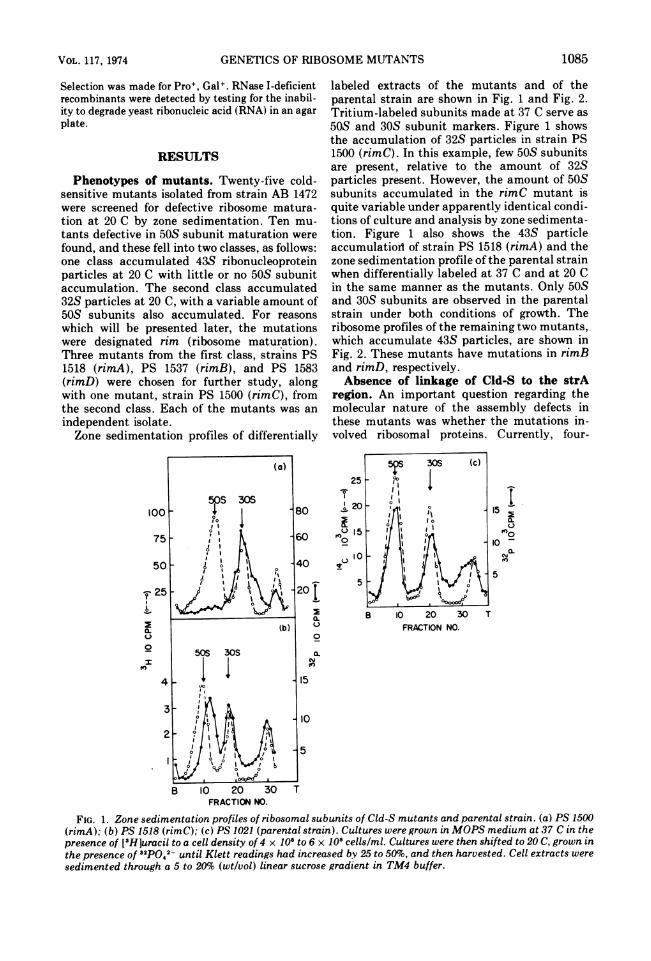
Phenotypes of mutants. Twenty-five cold-sensitive mutants isolated from strain AB 1472were screened for defective ribosome matura-tion at 20 C by zone sedimentation. Ten mu-tants defective in 50S subunit maturation werefound, and these fell into two classes, as follows:one class accumulated 43S ribonucleoproteinparticles at 20 C with little or no 50S subunitaccumulation. The second class accumulated32S particles at 20 C, with a variable amount of50S subunits also accumulated. For reasonswhich will be presented later, the mutationswere designated rim (ribosome maturation).Three mutants from the first class, strains PS1518 (rimA), PS 1537 (rimB), and PS 1583(rimD) were chosen for further study, alongwith one mutant, strain PS 1500 (rimC), fromthe second class. Each of the mutants was anindependent isolate.Zone sedimentation profiles of differentially
100
75
50
-F 25
0.0.0
zfEO
4
labeled extracts of the mutants and of theparental strain are shown in Fig. 1 and Fig. 2.Tritium-labeled subunits made at 37 C serve as50S and 30S subunit markers. Figure 1 showsthe accumulation of 32S particles in strain PS1500 (rimC). In this example, few 50S subunitsare present, relative to the amount of 32Sparticles present. However, the amount of 50Ssubunits accumulated in the rimC mutant isquite variable under apparently identical condi-tions of culture and analysis by zone sedimenta-tion. Figure 1 also shows the 43S particleaccumulatiod of strain PS 1518 (rimA) and thezone sedimentation profile of the parental strainwhen differentially labeled at 37 C and at 20 Cin the same manner as the mutants. Only 50Sand 30S subunits are observed in the parentalstrain under both conditions of growth. Theribosome profiles of the remaining two mutants,which accumulate 43S particles, are shown inFig. 2. These mutants have mutations in rimBand rimD, respectively.Absence of linkage of Cld-S to the strA
region. An important question regarding themolecular nature of the assembly defects inthese mutants was whether the mutations in-volved ribosomal proteins. Currently, four-
150.~80too
10 -
a-
10 20 30FRACTION NO.
FIG. 1. Zone sedimentation profiles of ribosomal subunits of Cld-S mutants and parental strain. (a) PS 1500(rimA); (b) PS 1518 (rimC); (c) PS 1021 (parental strain). Cultures were grown in MOPS medium at 37 C in thepresence of [3HJuracil to a cell density of 4 x 108 to 6 x 108 cells/ml. Cultures were then shifted to 20 C, grown inthe presence of 32pQ42- until Klett readings had increased by 25 to 50%o, and then harvested. Cell extracts weresedimented through a 5 to 20%o (wt/vol) linear sucrose gradient in TM4 buffer.
1085VOL. 117, 1974

BRYANT AND SYPHERD
0
a-
0
K)0
B 10 20 30 TFRACTION NO.
FIG. 2. Zone sedimentation profiles of ribosomalsubunits of cold-sensitive mutants. (a) PS 1662 (anrnsA derivative of PS 1537, rimB); (b) PS 1583(rimD). Conditions of culture growth and zone sedi-mentation analysis are the same as for Fig. 1.
tenths of the 55 ribosomal proteins cataloguedby Kaltschmidt and Wittmann (18) have beenshown to be specified by loci near strA, atminute 64 on the E. coli chromosome (34). Theone exception is S18, which maps between 76and 88 min (2, 17). The demonstration that a
ribosome mutation is at strA constitutes stronggenetic evidence that the mutation resides in agene coding for a ribosomal protein (34). Ac-cordingly, three of our mutants were tested forco-transduction of Cld-S with aroE, whichserves as a nutritional marker for the strAregion. Transducing phage lysates were pre-pared on mutants defective in rimA, rimB, and
rimC. Selection was made for aroE+ usingstrain AB 2834 as the recipient strain followedby scoring for Cld-S, which serves as a markerfor the maturation defect. No transduction ofCld-S with aroE was obs-,. --d1 for any of themutations. All aroE recombinants tested werewild type with respect to subunit maturation.The absence of linkage of the mutations causingmaturation defects to the strA region suggeststhat the mutations do not involve ribosomalproteins.Mapping of rimA. The genetic analysis was
continued with the object of determiningwhether the mutations occupied one or severalgenetic loci. Map locations were narrowed downto small regions of the chromosome in conjuga-tion experiments using Hfr strains with differ-ent points of origin. Then co-transduction ex-periments served to pinpoint the maturationdefective or Cld-S loci.
Preliminary mapping experiments for rimAinvolved the mating of a leu derivative of strainPS 1518 with the Hfr strains X884, x895, andx879. Distal markers were selected as follows:for strain X884, His+; for strain x895, Leu+; andfor strain x879, Ilv+. Donor strains were coun-terselected with streptomycin. Matings weremechanically interrupted after 50 min. Recom-binants were scored for linkage with Cld-R.The results of these experiments indfcate that
the Cld-S locus is located between 64 and 88min. Six markers within this chromosomalregion were tested for co-transduction withCld-S. The results of these experiments aregiven in Table 2. These data indicate that Cld-Sof strain PS 1518 co-transduces with pyrE andilvD. The lack of co-transduction of Cld-S withmetE, together with the 3-min separation ofpyrE and ilvD, shows that Cld-S lies betweenthese two markers, probably near min 74.
Tests for defective subunit assembly weremade for the following recombinants: an ilvD+Cld-S recombinant, an ilvD+ Cld-R recombi-nant, a pyrE+ Cld-S recombinant, and a pyrE+Cld-R recombinant. The Cld-S recombinantsaccumulated 43S particles at 20 C. The Cld-Rrecombinants were wild type with respect tosubunit maturation at 20 C. The subunit matu-ration defect and Cld-S are thus shown toco-transduce together when tested by selectionof two different nutritional markers. Defectivesubunit maturation and Cld-S in strain PS 1518are probably the same mutation; if not, themutations are very closely linked. In any case,demonstration of co-transduction of the matu-ration defect with a nutritional marker unequiv-ocally maps the locus giving rise to the matura-tion defect.Mapping of rimB. The rapid mapping proce-
1086 J. BACTERIOL.
GENETICS OF RIBOSOME MUTANTS
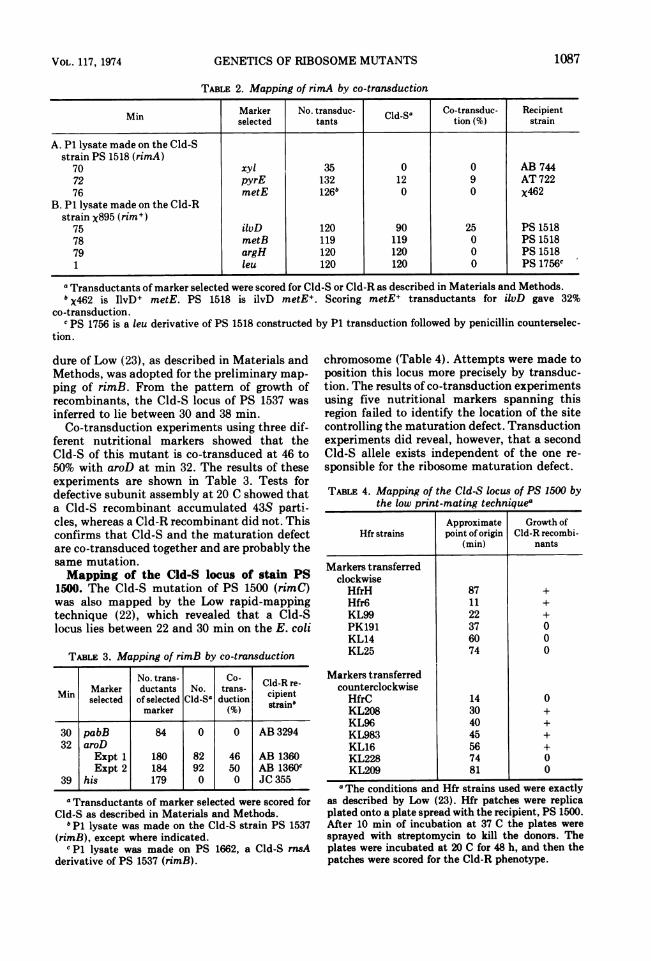
TABLE 2. Mapping of rimA by co-transduction
Min Marker No. transduc- Cld-Sa Co-transduc- Recipientselected tants tion (%) strain
A. P1 lysate made on the Cld-Sstrain PS 1518 (rimA)
70 xyl 35 0 0 AB 74472 pyrE 132 12 9 AT 72276 metE 126b 0 0 X462
B. P1 lysate made on the Cld-Rstrain x895 (rim+)
75 ilvD 120 90 25 PS 151878 metB 119 119 0 PS 151879 argH 120 120 0 PS 15181 leu 120 120 0 PS 1756C
a Transductants of marker selected were scored for Cld-S or Cld-R as described in Materials and Methods.b X462 is IlvD+ metE. PS 1518 is ilvD metE+. Scoring metE+ transductants for ilvD gave 32%
co-transduction.c PS 1756 is a leu derivative of PS 1518 constructed by P1 transduction followed by penicillin counterselec-
tion.
dure of Low (23), as described in Materials andMethods, was adopted for the preliminary map-ping of rimB. From the pattern of growth ofrecombinants, the Cld-S locus of PS 1537 was
inferred to lie between 30 and 38 min.Co-transduction experiments using three dif-
ferent nutritional markers showed that theCld-S of this mutant is co-transduced at 46 to50% with aroD at min 32. The results of theseexperiments are shown in Table 3. Tests fordefective subunit assembly at 20 C showed thata Cld-S recombinant accumulated 43S parti-cles, whereas a Cld-R recombinant did not. Thisconfirms that Cld-S and the maturation defectare co-transduced together and are probably thesame mutation.Mapping of the Cld-S locus of stain PS
1500. The Cld-S mutation of PS 1500 (rimC)was also mapped by the Low rapid-mappingtechnique (22), which revealed that a Cld-Slocus lies between 22 and 30 min on the E. coli
TABLE 3. Mapping of rimB by co-transduction
No. trans- Co- Cld-R re-
Min Marker ductants No. trans- cipientselected of selected Cld-S0 duction strain"marker (%)
30 pabB 84 0 0 AB 329432 aroD
Expt 1 180 82 46 AB 1360Expt 2 184 92 50 AB 1360c
39 his 179 0 0 JC 355
a Transductants of marker selected were scored forCld-S as described in Materials and Methods.
b P1 lysate was made on the Cld-S strain PS 1537(rimB), except where indicated.cP1 lysate was made on PS 1662, a Cld-S mnsA
derivative of PS 1537 (rimB).
chromosome (Table 4). Attempts were made toposition this locus more precisely by transduc-tion. The results of co-transduction experimentsusing five nutritional markers spanning thisregion failed to identify the location of the sitecontrolling the maturation defect. Transductionexperiments did reveal, however, that a secondCld-S allele exists independent of the one re-sponsible for the ribosome maturation defect.
TABLE 4. Mapping of the Cld-S locus of PS 1500 bythe low print-mating techniquea
Approximate Growth ofHfr strains point of origin Cld-R recombi-
(min) nants
Markers transferredclockwiseHfrH 87 +Hfr6 11 +KL99 22 +PK191 37 0KL14 60 0KL25 74 0
Markers transferredcounterclockwiseHfrC 14 0KL208 30 +KL96 40 +KL983 45 +KL16 56 +KL228 74 0KL209 81 0
a The conditions and Hfr strains used were exactlyas described by Low (23). Hfr patches were replicaplated onto a plate spread with the recipient, PS 1500.After 10 min of incubation at 37 C the plates weresprayed with streptomycin to kill the donors. Theplates were incubated at 20 C for 48 h, and then thepatches were scored for the Cld-R phenotype.
1087VOL. 117, 1974

BRYANT AND SYPHERD
Mapping of rimD. The Cld-S locus control-ling ribosome maturation in strain PS 1583(rimD) was mapped by conjugation. Figure 3shows the Hfr strains and their points of origin.Selection was made for Ilv+ in matings of strainPS 1583 (rimD) with Hfr strains X879 and AT3141 followed by scoring for Cld-R. Matingswere carried out for 45 min and were notmechanically interrupted. Donor strains werecounterselected by streptomycin (x879) or byomission of essential nutrient (AT 3141). Theresults of these matings (Table 5) revealed thatCld-S was located between 54 min clockwise asthe chromosome map is usually drawn to 12 minand probably near ilv. Two additional matingswere performed using the Hfr strains KL 25(selecting for argH+) and KL 209 (selecting forilvD+). Matings were for 30 min and were notmechanically interrupted. Donor strains werecounterselected by streptomycin. Linkage ofCld-R to each of these markers was 99%, indi-cating that the Cld-S locus lies between thepoints of origin of these two Hfr strains, whichare 74 min (KL 25) and 81 min (KL 209),respectively. Tests were made for defective
,_ _s~~~~~~4
"AT 3141
SI r,, 64
\ o,Q6 59
38
79
e /75
I\sr^ 64
KL209
FIG. 3. Direction and origin of transfer of chromo-some by Hfr strains used in locating the rimD locus.The dotted lines indicated the portion of the chromo-some distal to the selected marker. Selected markerswere Ilv for the matings with PS 1583 (rimD) andHfrs AT3141, X879, and KL209, and Arg+ for matingswith PS1583 (rimD) and KL25.
TABLE 5. Mapping of the Cld-S locus of strain PS1583 (rimD) by conjugationa
No. of re- Linkage ofstrain Selected combinants No. Cld-R andHfr stranmarker inheriting Cld-Rb selected
selected marker (%)marker
X879 ilv 117 105 90AT 3141 ilv 117 110 94KL 25 arg 141 139 99KL 209 ilv 142 141 99
a Points of origin and polarity of chromosomaltransfer of Hfr strains are shown in Fig. 3. Matingswith X879 and AT 3141 were for 45 min. Matings withKL 25 and KL 209 were for 30 min. At the end of themating period, cells were plated on selective mediawithout mechanical interruption of the mating. Donorcells were counterselected with streptomycin exceptfor AT 3141, which was counterselected by starvationfor pro, thr, and leu.
b Recombinants inheriting the selected marker werescored for Cld-R as described in Materials andMethods.
subunit assembly at 20 C for Ilv+ and Arg+recombinants. Cld-R recombinants were wildtype with respect to subunit assembly. Cld-Srecombinants accumulated 43S particles at20 C, confirming the association of Cld-S andthe maturation defect.
Strain PS 1583 is not efficiently infected byP1 phage, giving rise to lysate titers and tofrequencies of transduction, when used as arecipient, of values of 10- to 100-fold less thanthe titers or transduction frequencies observedfor other E. coli K-12 strains. In addition,recombinants of PS 1583 frequently segregatecolonies with a variety of growth rates, therebyobscuring definitive scoring for Cld-R andCld-S. For these reasons, a more precise maplocation for rimD has not yet been established.Preliminary co-transduction experiments indi-cate that rimD does not map near ilv. Thematuration-defective loci rimD and rimA are,therefore, probably not allelic.Of some interest is the observation that a
recombinant obtained from a mating of strainsKL 25 and PS 1583, where selection was madefor Ilv+, is Cld-S but not maturation defective.This Cld-S locus co-transduces with metB at afrequency of 84%. Therefore, there are at leasttwo Cld-S loci in strain PS 1583, one of thembeing unrelated to the maturation defect. Thefinding of more than one Cld-S locus is notsurprising since mutagenesis was by NTG. TheCld-S locus controlling ribosome maturationquite clearly lies in the 74- to 78-min region.Dominance tests. Dominance tests consisted
of testing a stable partial diploid strain (Cld-R/
1088 J. BACTERIOL.

GENETICS OF RIBOSOME MUTANTS
Cld-S) for defective ribosomal subunit matura-tion at 20 C. Partial diploids were formed bymating an appropriate F' strain with a recAderivative of the mutant to be tested. The recAderivatives were prepared as described in Mate-rials and Methods. Strain KLF11/JC1553 wasused to transfer the episome KLF 11 to a rimA,recA strain and to a rimD, recA strain. Selectionwas made for Ilv+ Met+ Arg+ Thi+ in bothcases. In a mating of strain F' 148/JC1553 witha rimB, recA strain, selection was made forHis+. In all matings, the episomal donor strainwas counterselected by omission of an essen-tial nutrient.The partial diploids were tested for the matu-
ration defect at 20 C by differentially labelingthe cells in nonselective medium, followed byzone sedimentation of cell extracts. To test thecultures for their diploid state at the time thecultures were harvested, samples of the cultureswere plated on nutrient agar. Colonies werepicked and tested for their donor ability byinoculating log-phase L broth cultures of eachcolony onto selective agar medium containinga lawn of recipient F- cells. For strain KLF-11/rimA recA, 100 colonies were tested for theability to transfer the episome to strain AT 722,selecting for pyrE+. All 100 colonies gave rise topyrE+ recombinants, demonstrating that all ofthe strain KLF11/rimA recA cells were genuinepartial diploids. In addition, the appearance ofpyrE+ recombinants confirms that the episomeKLF11 extends from ilvD to pyrE, therebyrendering the rimA, recA strain diploid at theCld-S locus. In the case of KLF11/rimD recA,which is spc+, 77 putative diploid colonies weretested for the ability to transfer the episome to aspc derivative of AB 1472. Selection was madefor Ilv+ Met+ Arg+ Thi+. The rimD, recA, spc+strain was counterselected by spectinomycin.Donor ability was demonstrated for 68 coloniesor 88% of the population at the time the culturewas harvested. In an independent experiment,44 out of 44 colonies tested had donor ability. In
SO 0S SOS
20 - 4 20
T ,, l it', I t \
the case of strain F' 148/rimB, recA, 373 colo-nies were tested for the inability to grow onmedium lacking histidine, which would revealepisomeless segregants. Three such colonieswere found, or 0.8% of the population. Tests fordonor ability were made on these three segre-gants along with 22 His+ colonies, using a Pro+derivative of strain AB 1360 as the recipientstrain and selecting for aroD+ his+. The 22 His+colonies gave rise to Aro+ His+ recombinants;the three His- colonies did not. This testdemonstrates that the episome F' 148 conferreddiploidy for both his and aroD, which is closelylinked to rimB.
Figure 4 shows the zone sedimentation pro-files of extracts of the differentially labeledpartial diploids. Substantial amounts of 50Ssubunits were accumulated in all three diploids,indicating that the wild-type allele is dominant.However, the dominance is incomplete becausea significant amount of material still accumu-lates in the 43S region at 20 C.
DISCUSSIONThe mutants studied here were isolated in
much the same way as those described byGuthrie et al. (12), and which they called sad(for subunit assembly defective). It is possiblethat rim and sad mutants constituted the sameheterogeneous group. However, since rim mu-tants are shown here to lie in regions far awayfrom the strA gene and the genetics reported forsad mutations place them close to strA, it wouldbe premature to regard the two genotypes asequivalent. This distinction seems especiallycompelling in view of the finding that severalsad mutants have alterations in the gene for the30S ribosomal protein S5 (26).There are three types of mutations that
conceivably could give rise to a block in ribo-somal subunit maturation. These are (i) a mu-tated component of the ribosome (i.e., an alter-ation in the primary structure of ribosomalRNA [rRNA ] or a ribosomal protein) that
25
204
6 - 6
a l 20 30 TFRAC:TION NO.
FIG. 4. Dominance tests for the rim mutations: zone sedimentation profiles of ribosomes isolated frommerodiploids heterozygous for the wild-type allele and (a) rimA, (b) rimB, and (c) rimD. Merodiploids wereprepared by transferring an episome into F- recA strains bearing the respective rim loci, as described in thetext. Conditions of culture growth and zone sedimentation analysis are the same as for Fig. 1.
VOL. 117, 1974 1089

BRYANT AND SYPHERD
prevents binding of all ribosomal components,blocks modification of components, or preventsfolding of the ribosome into the mature configu-ration; (ii) an altered enzyme that fails tocatalyze a modification reaction that is neces-sary for proper assembly; (iii) a factor thatpromotes assembly in the wild type, but, whenaltered, prevents proper assembly and is notretained in the mature subunit. Many of thecold-sensitive, ribosome assembly-defectivemutants isolated by other investigators (12, 13,26, 32, 33, 38) apparently b)elong to the firstclass since they are linked to strA. However,some mutants of the class referred to as sadapparently are not linked to strA (Nomura,personal communication). With regard to thesecond class, modification reactions, such asrRNA methylation, removal of oligonucleotidesfrom precursor rRNA, and acetylation andmethylation of the 50S protein L7, are known tooccur (8), Whether any of these modifications isnecessary for proper assembly is not known. Atleast one 16S rRNA methylation reaction is notnecessary for assembly (15). With regard to thethird class of mutation, Traub and Nomura (36)have shown that both the 50S and 30S subunitsare capable of self assembly in vitro into ma-ture, functional subunits. There are two notabledifferences between in vitro reconstitution andin vivo maturation. In vitro reconstitution of30S ribosomes employs mature, fully methyl-ated RNA and has an optimum temperature of50 C (10). In vivo maturation begins with sub-methylated precursor RNA and takes place attemperatures down to 8 to 15 C, the lowergrowth limit of E. coli (16). That in vivomaturation proceeds by a mechanism otherthan self assembly seems quite possible. Thereis precedence for "assembly factors or enzymes"in other maturation systems; for example, in T4head and tail assembly (20, 39). Mutations inT4 head assembly factors have also been re-ported (5).We have found four classes of ribosome matu-
ration mutations based on map positions. Onlyin the case of rimA and rimD are the locipossibly allelic. Our tentative conclusion is thatthese two mutations constitute separate loci. InFig. 5 the map positions of the four loci aresummarized. Each of the four mutationalclasses is represented by a single mutant. It isquite probable that additional non-strA-linkedloci exist. Turnock (24, 37) showed that amutation responsible for accumulation of 43Sparticles in a non-Cld-S mutant lies near xyl.That mutation might be similar to those re-ported here.The fact that all four mutations lie outside
rID,u*`-Aat4 rim c JPur aproteinsstr A
rIG. 5. Genetic map of E. coli indicating the mappositions of rim loc&. The positions of rimC and rimDare based on conjugation data, and are approximate.The positions of rimA and rimB were determined byco-transduction with ilv and aroD, respectively.
the s_,rA region suggests that the loci do not codefor ribosomal proteins. Additional evidence thatthe mutations do not involve ribosomal proteinsis provided by an analysis of proteins isolatedfrom the 50S and 30S subunits. Two-dimen-sional polyacrylamide gel electrophoresis hasrevealed that the ribosomal proteins from themutants are identical to the parental strain inelectrophoretic mobility (unpublished data).We consider the possibilty that these mutationsinvolve the primary structure of rRNA to beunlikely because of the redundancy of rRNAgenes.The recessive character of the ribosome mat-
uration defects in rimA, rimB, and rimD mu-tants is most simply explained as a defectivecatalytic activity. The wild-type alleles are in-completely dominant as evidenced by the oc-currence of detectable amounts of 43S particlesin the merodiploids. This phenomenon is notunderstood at present, but could be explained ifthe catalytic factors were transiently involvedin stoichiometric quantities. For example, if afactor were bound to the developing particle fora significant time, and then released to be usedagain, there could be some limitation of thefactor in the merodiploids. At any rate, the few43S particles seen in the merodiploids do de-velop into mature particles since they "chase"into the 50S region. One would expect thatwild-type modification enzymes or assemblyfactors used in catalytic amounts would restoreribosome maturation in the merodiploids. Spar-ling et al. (30) have presented evidence that, instrains diploid for ribosomal protein loci, ribo-somes are assembled from a common poolcontaining proteins encoded by both episomeand chromosome. A mutation in a ribosomal
1090 J. BACTERIOL.

GENETICS OF RIBOSOME MUTANTS
structural protein that causes a block in subunitassembly should be partially dominant to wildtype since ribosomal proteins are used stoichio-metrically. The strA-linked or spcA-linkedCld-S mutations that have been tested aredominant or partially dominant (13, 38).
It is possible that the mutations examinedhere, and which we regard as ribosome matura-tion mutations, constitute a very heterogeneousgroup of mutations and are similar to thosereferred to as sad (subunit assembly defective)by Guthrie et al. (12). In one case (sad-19), thesad character occurred at 64 min on the chromo-some. However, other sad mutations showrather complicated genetics and several areunlinked strA, but lie in the general region. Someof the cold-sensitive mutants of Salmonellatyphimurium also lie outside the strA region(38).The genetic data reported here suggest that
the ribosome assembly defects which we callrim involve either modification enzymes orassembly factors. Recent biochemical analysesof the ribosome maturation defects of strain PS1518 and strain PS 1537 performed in ourlaboratory support this hypothesis. These ex-periments have shown that the 43S particlesaccumulated in these two mutants are con-verted to 50S subunits when treated with ex-tracts from wild-type cells. This conversion isnot due merely to addition of ribosomal pro-teins. In addition, a partial purificatiQn of theconverting activity by diethylaminoethyl col-umn chromatography has been achieved.
ACKNOWLEDGMENTSThis investigation was supported by grant GM 18293/3
from the National Institute of Health and grant GB 25529from the National Science Foundation. R. E. B. was sup-ported by a stipend from a training grant awarded to theDepartment of Microbiology, University of Ilinois, from theNational Institute of Health. We wish to thank Mary JoSypherd and Kathleen Dimmitt for their capable assistancein the search and screening for mutants.
LITERATURE CITED
1. Adelberg, E. A., M. Mandel, and G. C. C. Chen. 1965.Optimal conditions for mutagenesis by N-meth-yl-N'-N-nitroso-guanidine in Escherichia coli K12.Biochem. Biophys. Res. Commun. 18:788-795.
2. Bollen, A., M. Faelen, J. P. Lecocq, A. Herzog, J. Zengel,L. Kahan, and M. Nomura. 1973. The structural genefor the ribosomal protein S18 in Escherichia coli. I.Genetic studies on a mutant having an alteration in theprotein S18. J. Mol. Biol. 76:463-472.
3. Clark, A. J., and A. D. Margulies. 1965. Isolation andcharacterization of recombination-deficient mutants ofEscherichia coli K12. Proc. Nat. Acad. Sci. U.S.A.53:451-459.
4. Clowes, R. C., and W. Hayes. 1968. Experiments inmicrobial genetics, p. 188. John Wiley and Sons, Inc.,New York.
5. Coppo, A., A. Manzi, J. F. Pulitzer, and H. Takahashi.
1973. Abortive bacteriophage T4 head assembly inmutants of Escherichia coli. J. Mol. Biol. 76:61-87.
6, Curtiss, R., III, L. G. Caro, D. P. Allison, and D. R.Stallions. 1969. Early stages of conjugation in Esche-richia coli. J. Bacteriol. 100:1091-1104.
7. Curtiss, R., III, L. J. Charamella, C. M. Berg, and P. E.Harris. 1965. Kinetic and genetic analyses of D-cyclo-serine inhibition and resistance in Escherichia coli. J.Bacteriol. 90:1238-1250.
8. Davies, J., and M. Nomura. 1972. The genetics ofbacterial ribosomes. Annu. Rev. Genet. 6:203-234.
9. Demerec, H., E. A. Adelberg, A. J. Clark, and P. E.Hartman. 1966. A proposal of uniform nomenclature inbacterial genetics. Genetics 54:61-76.
10. Egberts, E., P. Traub, P. Herrlich, and M. Schweiger.1972. Functional integrity of Escherichia coli 30Sribosomes reconstituted from RNA and protein: invitro synthesis of S-adenosylmethionine cleaving en-zyme. Biochim. Biophys. Acta 277:681-684.
11. Gesteland, R. F. 1966. Isolation and characterization ofribonuclease I mutants of Escherichia coli. J. Mol. Biol.16:67-84.
12. Guthrie, C., H. Nashimoto, and M. Nomura. 1969.Structure and function of Escherichia coli ribosomes.VIII. Cold-sensitive mutants defective in ribosomeassembly. Proc. Nat. Acad. Sci. U.S.A. 63:384-391.
13. Guthrie, C., H. Hashimoto, and M. Nomura. 1969.Studies on the assembly of ribosomes in vivo. ColdSpring Harbor Symp. Quant. Biol. 34:69-75.
14. Hashimoto, K. 1960. Streptomycin resistance in Esche-richia coli analyzed by transduction. Genetics45:49-62.
15. Helser, T. L., J. E. Davies, and J. E. Dahlberg. 1971.Mechanism of kasugamycin resistance in Escherichiacoli. Nature N. Biol. 235:6-9.
16. Ingraham, J. L. 1958. Growth of psychrophilic bacteria. J.Bacteriol. 76:75-80.
17. Kahan, L., J. Zengel, M. Nomura, A. Bollen, and A.Herzog. 1973. The structural gene for the ribosomalprotein S18 in Escherichia coli, II. Chemical studies onthe protein S18 having an altered electrophoreticmobility. J. Mol. Biol. 76:473-483.
18. Kaltschmidt, E., and H. G. Wittmann. 1970. Ribosomalproteins. XII. Number of proteins in small and largesubunits of Escherichia coli as determined by two-dimensional gel electrophoresis. Proc. Nat. Acad. Sci.U.S.A. 67:1276-1282.
19. Kurland, C. G. 1972. Structure and function of thebacterial ribosome. Annu. Rev. Biochem. 41:377-408.
20. Laemmli, U. K. 1970. Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature (London) 227:680-685.
21. Lennox, E. S. 1955. Transduction of linked geneticcharacters of the host by bacteriophage P1. Virology1:190-206.
22. Lin, E. C. C., S. A. Lerner, and S. E. Jorgensen. 1962. Amethod for isolating constitutive mutants for carbohy-drate-catabolizing enzymes. Biochim. Biophys. Acta60:422-429.
23. Low, B. 1973. Rapid mapping of conditional and auxo-trophic mutations in Escherichia coli K12. J. Bacteriol.113:798-812.
24. MacDonald, R. E., G. Tumock, and J. Forchhammer.1967. The synthesis and function of ribosomes in a newmutant of Escherichia coli. Proc. Nat. Acad. Sci.U.S.A. 57:141-147.
25. Miller, J. H. 1972. Experiments in molecular genetics, p.218-220. Cold Spring Harbor Laboratory, Cold SpringHarbor, New York.
26. Nashimoto, H., W. Held, E. Kaltschmidt, and M. Nomu-ra. 1971. Structure and function of bacterial ribosomes.XII. Accumulation of 21S particles by some cold-sensi-
VOL. 117, 1974 1091

BRYANT AND SYPHERD
tive mutants of Escherichia coli. J. Mol. Biol.62:121-138.
27. Nomura, M. 1970. Bacterial ribosome. Bacteriol. Rev.34:228-277.
28. Okada, T., K. Yanagisawa, and F. J. Ryan. 1961. Amethod for securing thymineless mutants from strainsof E. coli Z. Verebungslehre 92:403-412.
29. Pace, B., and N. R. Pace. 1971. Gene dosage for 5Sribosomal ribonucleic acid in Escherichia coli andBacillus megaterium. J. Bacteriol. 105:142-149.
30. Sparling, P. F., J. Modolell, I. Takeda, and B. D. Davis.1968. Ribosomes from Escherichia coli merodiploidsheterozygous for resistance to streptomycin and specti-nomycin. J. Mol. Biol. 37:407-421.
31. Sypherd, P. S., and N. Strauss. 1962. Chloramphenicol-promoted repression of 0-galactosidase synthesis inEscherichia coli. Proc. Nat. Acad. Sci. U.S.A.49:400-407.
32. Tai, P. C., and J. L. Ingraham. 1971. A ribosome-likeparticle accumulated at low temperature by a cold-sen-sitive mutant of Salmonella typhimurium. Biochim.Biophys. Acta 232:151-166.
33. Tai, P. C., D. P. Kessler, and J. Ingraham. 1969.Cold-sensitive mutations in Salmonella typhimuriumwhich affect ribosome synthesis. J. Bacteriol.97:1298-1304.
34. Takata, R. 1972. Genetic studies of the ribosomal pro-teins in Escherichia coli. VII. Mapping of ribosomalprotein components by intergeneric mating experi-ments between Serratia marcescens and Escherichiacoli. Mol. Gen. Genet. 118:363-371.
35. Taylor, A. L., and C. D. Trotter. 1972. Linkage map ofEscherichia coli strain K12. Bacteriol. Rev. 36:504-524.
36. Traub, P., and M. Nomura. 1968. Structure and functionof Escherichia coli ribosomes. V. Reconstitution offuntionally active 30S ribosomal particles from RNAand proteins. Proc. Nat. Acad. Sci. U.S.A. 59:777-784.
37. Turnock, G. 1969. A genetic analysis of a mutant ofEscherichia coli with a defect in the assembly ofribosomes. Mol. Gen. Genet. 104:295-312.
38. Tyler, B., and J. L. Ingraham. 1973. Studies on ribosomalmutants of Salmonella typhimurium LT-2. Mol. Gen.Genet. 112:197-214.
39. Wood, W. B., and M. Henninger. 1969. Attachment oftail fibers in bacteriophage T4 assembly: some proper-ties of the reaction in vitro and its genetic control. J.Mol. Biol. 39:603-618.
40. Yankofsky, S. A., and S. Spiegelman. 1962. The identifi-cation of the ribosomal RNA cistron by sequencecomplementarity. II. Saturation of and competitiveinteraction at the RNA cistron. Proc. Nat. Acad. Sci.U.S.A. 48:1466-1472.
1092 J. BACTERIOL.




















