
Modern pollen assemblages from cultivated rice fields and rice pollen morphology: Application to a...
12
The Holocene 22(12) 1393–1404 © The Author(s) 2012 Reprints and permission: sagepub.co.uk/journalsPermissions.nav DOI: 10.1177/0959683612449761 hol.sagepub.com Introduction Pollen analysis is commonly considered as one of the important archives providing information on vegetation and climate changes as well as human impacts during the Holocene. In recent years, the quantitative analysis of Quaternary pollen records for evidence of early anthropogenically induced land cover changes and agricul- tural development has attracted increasing attention from the sci- entific community. For more than a decade, fossil pollen data have been investigated as a proxy employed for human impact on the landscape and agriculture in China (Atahan et al., 2008; Behre, 1986; Davies and Tipping, 2004; Dearing et al., 2006; Edwards and Hirons, 1984; Li et al., 2008; Yasuda et al., 2004; Zong et al., 2007). Some studies further developed pollen analysis of sedi- ments from archaeological sites as a proxy method for ecological condition surrounding the relics and related human activity (e.g. Behre, 1986; Dimbleby, 1985; Kong and Du, 1981; Mo et al., 2003; Yang et al., 2006; Zong et al., 2012a). For the Holocene in particular, pollen assemblages reveal valuable insight into the beginning of cereal cultivation and the transition from hunting and gathering by Neolithic foragers to a food-producing economy (Innes et al., 2009; Lu, 1999; Zheng et al., 2004). Previous studies show that early rice farming was widespread in China, especially in the middle and lower reaches of the Yangtze River, as well as throughout south China (Gong et al., 2007; Itzstein-Davey et al., 2007; Zong et al., 2012b). Zheng et al. (2004) argued that sudden increases in the frequency of heliophilous plants in the upper part of the Holocene pollen diagrams for south China may reflect human disturbance of forests linked to the development of agricul- ture. However, pollen evidence for human activity is not always unambiguous, and interpretation of Holocene pollen spectra and methods for the quantitative estimate of human-induced land cover change in China is still problematic. We address this topic in relation to the investigation of rice, which is significant as the most important tropical-subtropical crop. The historical development of rice agriculture played a major role in regional environmental 449761HOL 0010.1177/09596 83612449761Yang et al.The Holocene 2012 1 Sun Yat-Sen University, China 2 The University of Hong Kong, SAR China 3 Hebei Normal University, China 4 University of Hawaii, USA Corresponding author: Zheng Zhuo, Department of Earth Sciences, Sun Yat-Sen University, Guangzhou 510275, China. Email: [email protected] Modern pollen assemblages from cultivated rice fields and rice pollen morphology: Application to a study of ancient land use and agriculture in the Pearl River Delta, China Shixiong Yang, 1 Zhuo Zheng, 1 Kangyou Huang, 1 Yongqiang Zong, 2 Jianhua Wang, 1 Qinghai Xu, 3 Barry V Rolett 4 and Jie Li 1 Abstract Pollen from a series of surface soil samples collected along a transect spanning southeast China was investigated to better understand palynological signals of ancient agriculture and other human activity. The transect surface samples consist of pairs taken inside and outside rice paddy fields. Pollen assemblages from these samples are valuable as modern analogs of human-altered environments and rice agriculture. Our measurements of Poaceae pollen grains from inside the modern rice fields discovered that 34–40 µm is the statistically significant size range for identifying domesticated rice in fossil pollen samples. This conclusion is also based on a size comparison of raw and chemically treated modern pollen grains from the plants. Pollen measurements for local wild grasses show that most native weeds have pollen grains less than 30 µm in size. The modern analogs and our study of the influence of chemical treatment on pollen grain size made it possible to examine a sediment core from the Pearl River delta for evidence of anthropogenic influence, including rice farming. Pollen assemblages from around 2200 cal. yr BP are highly similar to those of our modern analogs representing disturbed landscapes outside modern rice fields. The pollen spectra reveal abrupt increases in Poaceae, Dicranopteris, Artemisia and Pinus indicative of rice farming and forest clearance, at around 2200 cal. yr BP. Major factors associated with this abrupt transition were the rapid formation of the deltaic flood plain and massive increases in the Pearl River delta area population during the Qin Dynasty. Keywords Holocene, human activity, Oryza sativa, pollen analysis, rice paddy field, southeastern China Received 10 November 2011; revised manuscript accepted 12 March 2012 Research paper
-
Upload
manoa-hawaii -
Category
Documents
-
view
3 -
download
0
Transcript of Modern pollen assemblages from cultivated rice fields and rice pollen morphology: Application to a...

The Holocene22(12) 1393 –1404© The Author(s) 2012Reprints and permission:sagepub.co.uk/journalsPermissions.navDOI: 10.1177/0959683612449761hol.sagepub.com
IntroductionPollen analysis is commonly considered as one of the important archives providing information on vegetation and climate changes as well as human impacts during the Holocene. In recent years, the quantitative analysis of Quaternary pollen records for evidence of early anthropogenically induced land cover changes and agricul-tural development has attracted increasing attention from the sci-entific community. For more than a decade, fossil pollen data have been investigated as a proxy employed for human impact on the landscape and agriculture in China (Atahan et al., 2008; Behre, 1986; Davies and Tipping, 2004; Dearing et al., 2006; Edwards and Hirons, 1984; Li et al., 2008; Yasuda et al., 2004; Zong et al., 2007). Some studies further developed pollen analysis of sedi-ments from archaeological sites as a proxy method for ecological condition surrounding the relics and related human activity (e.g. Behre, 1986; Dimbleby, 1985; Kong and Du, 1981; Mo et al., 2003; Yang et al., 2006; Zong et al., 2012a). For the Holocene in particular, pollen assemblages reveal valuable insight into the beginning of cereal cultivation and the transition from hunting and gathering by Neolithic foragers to a food-producing economy (Innes et al., 2009; Lu, 1999; Zheng et al., 2004). Previous studies show that early rice farming was widespread in China, especially in the middle and lower reaches of the Yangtze River, as well as
throughout south China (Gong et al., 2007; Itzstein-Davey et al., 2007; Zong et al., 2012b). Zheng et al. (2004) argued that sudden increases in the frequency of heliophilous plants in the upper part of the Holocene pollen diagrams for south China may reflect human disturbance of forests linked to the development of agricul-ture. However, pollen evidence for human activity is not always unambiguous, and interpretation of Holocene pollen spectra and methods for the quantitative estimate of human-induced land cover change in China is still problematic. We address this topic in relation to the investigation of rice, which is significant as the most important tropical-subtropical crop. The historical development of rice agriculture played a major role in regional environmental
449761 HOL0010.1177/0959683612449761Yang et al.The Holocene2012
1Sun Yat-Sen University, China2The University of Hong Kong, SAR China3Hebei Normal University, China4University of Hawaii, USA
Corresponding author:Zheng Zhuo, Department of Earth Sciences, Sun Yat-Sen University, Guangzhou 510275, China. Email: [email protected]
Modern pollen assemblages from cultivated rice fields and rice pollen morphology: Application to a study of ancient land use and agriculture in the Pearl River Delta, China
Shixiong Yang,1 Zhuo Zheng,1 Kangyou Huang,1 Yongqiang Zong,2
Jianhua Wang,1 Qinghai Xu,3 Barry V Rolett4 and Jie Li1
AbstractPollen from a series of surface soil samples collected along a transect spanning southeast China was investigated to better understand palynological signals of ancient agriculture and other human activity. The transect surface samples consist of pairs taken inside and outside rice paddy fields. Pollen assemblages from these samples are valuable as modern analogs of human-altered environments and rice agriculture. Our measurements of Poaceae pollen grains from inside the modern rice fields discovered that 34–40 µm is the statistically significant size range for identifying domesticated rice in fossil pollen samples. This conclusion is also based on a size comparison of raw and chemically treated modern pollen grains from the plants. Pollen measurements for local wild grasses show that most native weeds have pollen grains less than 30 µm in size. The modern analogs and our study of the influence of chemical treatment on pollen grain size made it possible to examine a sediment core from the Pearl River delta for evidence of anthropogenic influence, including rice farming. Pollen assemblages from around 2200 cal. yr BP are highly similar to those of our modern analogs representing disturbed landscapes outside modern rice fields. The pollen spectra reveal abrupt increases in Poaceae, Dicranopteris, Artemisia and Pinus indicative of rice farming and forest clearance, at around 2200 cal. yr BP. Major factors associated with this abrupt transition were the rapid formation of the deltaic flood plain and massive increases in the Pearl River delta area population during the Qin Dynasty.
KeywordsHolocene, human activity, Oryza sativa, pollen analysis, rice paddy field, southeastern China
Received 10 November 2011; revised manuscript accepted 12 March 2012
Research paper

1394 The Holocene 22(12)
changes throughout much of Asia. In order to fully understand the signature of rice agriculture in fossil pollen assemblages, it is use-ful to study the modern pollen rain in agricultural areas. We apply this approach in order to develop methods for distinguishing rice pollen from that of other Poaceae in both modern and fossil pollen assemblages. Refining this distinction is an important step towards using pollen as a proxy to study not only the origins and spread of rice farming but also the intensification of cultivation systems.
Some scientists (Andersen and Bertelsen, 1972; Köhler and Lange, 1979) have used scanning electron microscopy to distin-guish rice pollen from that of other Poaceae. Another method is the use of a size threshold to separate the cereal grain from grass pollen on the basis of Poaceae pollen grain size. In this case, some researchers (e.g. Atahan et al., 2008; Innes et al., 2009; Zong et al., 2007) identify Poaceae pollen greater than 40 µm in length as rice, while others (Zheng et al., 2009) use a size threshold of greater than 38 µm. With the goal of refining the pollen grain size method for differentiating rice from other Poaceae, we examined modern pollen samples from southern China, a region where rice agriculture has been common since ancient times.
This paper aims to reveal the characteristic pollen spectra linked with rice agriculture and human vegetation disturbance in rice cultivation areas, which helps better differentiate the human disturbance on the vegetation from climate change. Our study
compares modern pollen assemblages from inside and outside of rice paddy fields in areas of rice cultivation across southern China. In addition to the Poaceae pollen, we also examine the associated pollen taxa from our modern soil samples. The modern samples were collected along a latitudinal transect in the subtropi-cal double-cropping rice agriculture area of southern China. Finally, we use our modern data to interpret fossil pollen assem-blages from a sediment core obtained from the Pearl River delta plain, with the goal of identifying evidence for the beginning of intensive rice agriculture in this region.
Study areaRegional setting of surface soil sample collection areasWe collected surface sediment samples along a latitudinal tran-sect in the double-cropping rice production region of southern China (Figure 1), extending through the provinces of Guangdong, Hunan and Hubei. The spacing interval between samples is about 30~50 km, and ground altitudes of the sampling locations vary between 30 and 800 m above sea level (Table 1). Two soil sam-ples were collected at each location, one from inside a rice field and a second from about 10 m outside the cultivated field. Sam-ples collected inside the fields consist of surface sediments from
Figure 1. Locations of the surface pollen sampling sites in the subtropical double-cropping rice region of southern China and a Holocene borehole from the center of the Pearl River delta plain.

Yang et al. 1395
the top ~5 cm of the rice paddies. Samples taken from outside the fields consist of a mixture of surface sediment collected at several random points of uncultivated land surrounding the rice field. All samples were collected in September 2008 before the flowering period of the second rice crop.
For each sampling site, we also recorded data documenting the vegetation on land surrounding the rice fields investigated. Weeds are plentiful on the uncropped land near the paddy fields. The most common species are Poaceae, including Echinochloa crus-galli, Arundinella hirta, Digitaria sanguinalis, Digitaria ciliaris, Imperata cylindrical, Alopecurus aequalis, Setaria palmifolia, Setaria plicata and Setaria viridis (Table 2). Other frequent herbs are Asteraceae, Polygonaceae, Cyperaceae, Amaranthaceae, Scrophulariaceae, Papillionaceae, Phytolaccaceae, Com-melinaceae, Verbenaceae and Ranunculaceae. The aquatic plants Potamogeton (Potamogeton distinctus) and Onagraceae are com-mon in nearby wetlands.
On hill slopes in upland areas near the rice fields, the most common trees are Pinus massoniana and Cunninghamia lanceo-lata. Roadside trees near rice fields include Eucalyptus spp., Cin-namomum spp., Liquidambar formosana, Sapium sebiferum, Aleurites moluccana, and Populus sp. Secondary forests in our study area, which are not located in close proximity to the sam-pling sites, comprise mainly shrubs and small trees. The major secondary forest species are Rhus chinensis Mill, Loropetalum chinensis and Mallotus apelta in the southern areas, and Castanea mollissima, Pinus tabulaeformis, Quercus acutissima, Ilex
purpurea, Juglans cathayensis in the northern zone of the sam-pling transect.
Location and chronology of the Pearl River Delta sediment coreThe plain of Pearl River Delta is one of the largest flat regions in southeastern China. It is a highly productive area for agriculture, and has been throughout modern and extending back into ancient times. The climate is characterized by monsoon circulation, with annual precipitation as high as 1900 mm and a mean annual tem-perature of about 22°C. The local vegetation has been seriously disturbed by human activity and the modern vegetational land-scape consists mainly of agriculture, secondary shrubs, and planted forest. A few natural forests can be found in the nearby mountains. One example is the Dinghu Mountain area, where the vegetation is characteristic of monsoon broadleaved evergreen forests, dominated by Castanopsis, Cyclobalanopsis, Moraceae, Magnoliaceae, Proteaceae, and Palmae.
The Quaternary sediments of the delta are up to 60 m or more thick for the time period from the Late Pleistocene to Holocene (Zong et al., 2009a, 2012c). The sediment core (GZ-2) for our study was collected in Wanqingsha, Panyu County in the south-eastern part of the Pearl River Delta plain. The GIS location is 22°42.339'N, 113°30.831'E and 0.68 m altitude. The Holocene sediment sequence of this core is contained within the top 14 m, and its chronology is based on four AMS dates (Table 3) (Wang
Table 1. Location of surface sample sites of the present study.
Site no. Latitude (°N) Longitude (°E) Altitude (m) Site no. Latitude (°N) Longitude (°E) Altitude (m)
1 22.84 115.17 114 14 29.29 113.32 56 2 23.00 115.63 97 15 29.63 113.72 44 3 23.15 114.58 133 16 29.73 113.83 37 4 23.86 113.54 89 17 30.24 114.26 31 5 24.16 113.66 143 18 31.13 113.74 49 6 24.83 113.19 232 19 31.53 113.39 106 7 25.09 113.11 763 20 31.92 112.93 119 8 26.00 113.07 119 21 32.13 112.46 109 9 26.46 112.87 95 22 32.22 112.02 15510 26.94 112.83 77 23 32.22 110.72 59911 27.53 113.03 82 24 32.29 111.73 9112 28.44 113.18 59 25 32.51 111.11 18013 28.79 113.27 53
Table 2. The most frequent herbs commonly found from land surface around rice fields.
Family species Family species
Poaceae Echinochloa crusgalli Cyperaceae C. compressus Arundinella hirta C. glomeratus Digitaria sanguinalis Scirpus spp. Digitaria ciliaris Amaranthaceae Alternanthera spp. Imperata cylindrica, Scrophulariaceae Scopia dulcis Alopecurus aequalis Papillionaceae Sesbania cannabina Setaria viridis Phytolaccaceae Phytolacca acinosa Setaria palmifolia Commelinaceae Commelina communis Setaria plicata Verbenaceae Vitex negundoAsteraceae Bidens pilosa Ranunculaceae Anemone vitifolia Ageratum conyzoides Polygonaceae Fagopyrum sp Artemisia princeps Polygonum orientale Erigeron acer P. lapathifolium Xanthium sibiricum P. hydropiperCyperaceae Juncellus serotinus P. orientale, Cyperus iria

1396 The Holocene 22(12)
et al., 2009). All dates are calibrated into calendar ages by means of CalPal software of Cologne Radiocarbon Calibration & Pal-aeoclimate Research Package (Weninger and Jöris, 2002) and lin-ear interpolation was performed into each sample depth.
MethodsThe soil samples were prepared for palynological analysis using hydrochloric (10%) and hydrofluoric acids (40%) to remove the carbonates and silicates, followed by KOH treatment to remove the organic matter. The fossil pollen grains and spores were con-centrated using standard heavy liquid flotation techniques grains (Du and Chen, 1990). Pollen identification and counting were carried out with a Nikon E200 stereomicroscope at 400× and 1000× (oil immersion) magnifications. A minimum sum of 300 pollen grains was counted for each sample. Pollen percentages were calculated based on the pollen sum excluding fern spores.
We collected pollen grains from flowering rice (Oryza sativa L) plants and wild grasses to compare their morphology and to determine methods for distinguishing rice from other Poaceae. The raw pollen was treated using two procedures. The Erdtman acetic anhydride method uses an acetolysis solution made by mix-ing 10 ml of sulphuric acid with 90 ml of acetic anhydride. The
fossil pollen preparation procedure is based on the alkaline-acidic and heavy liquid method. Treated pollen grains were compared with untreated grains to determine the influence of treatment on grain size.
Analysis of the modern pollen assemblagesTaxonomic compositionA total of 23,571 pollen grains representing 77 pollen taxa have been identified and counted for this study. In addition to grasses (Poaceae), which are the dominant taxa, other abundant pollen taxa are Pinus, Asteraceae, Brassicaceae, Chenopodiaceae, Polygonaceae, Cyperaceae, Quercus, Castanopsis, Euphorbia-ceae, Apiaceae, Caryophyllaceae, Ranunculaceae, Artemisia and Papillionaceae. Dicranopteris is the most abundant taxon among the fern spores. Other common spores are Pteris, Lygodium and Cibotium barometz.
Poaceae accounts for up to 36–88% (average 68.8%) of the total pollen among pollen assemblages from samples collected inside the rice fields (Figure 2). The pollen assemblages from samples collected outside rice fields are also dominated by
Figure 2. Pollen diagram of the surface soil samples inside rice fields northwards (upward). The regional divisions of the three Provinces are indicated.
Table 3. AMS 14C ages of core GZ-2.
Depth (m) AMS lab no.a Material Dating method AMS 14C age/yr BP Cal. yr BP
2.4 GZ2088 Shell 14C AMS 1175±33 1110±606.70-6.90 GZ2089 Shell 14C AMS 1817±29 1770±4010.53 GZ2090 Shell 14C AMS 2452±40 2450±12012.8 GZ2091 Charcoal 14C AMS 3698±28 4040±50
aDating samples were prepared in the laboratory of Guangzhou Institute of Geochemistry, Chinese Academy of Sciences and the AMS 14C was measured by the State Key Laboratory of nuclear physics and nuclear technology, Beijing University.

Yang et al. 1397
Poaceae pollen, which accounts for an average 57% of the pollen in these samples (Figure 3). The relative abundance of Poaceae is lower, at about 40%, among the Hubei Province samples, where other herbs and tree pollen are comparatively abundant.
The composition of non-Poaceae pollen types is similar among samples from both inside and outside the rice fields. Some of the herb pollen in our assemblages may derive from vegetables farmed nearby, including cabbage and brassica (Brassicaceae), carrots (Apiaceae), beans and peas (Papillionaceae), leeks and daylily (Liliaceae). Other less abundant herbs are from the Astera-ceae, Apiaceae, Caryophyllaceae, Ranunculaceae, and Chenopo-diaceae families. The tree pollen is mainly Pinus, with smaller quantities of Quercus (deciduous type), Castanopsis, Euphorbia-ceae, Juglans and so on. Pinus pollen is particularly abundant (41.5%) at sample site 25, which is located near a pine forest. However, even at other locations comparatively distant from pine forests, the relative abundance of Pinus pollen is still as high as 10%, because it is easily transported by wind over long distances. The pollen frequencies also vary by latitude. Poaceae is compara-tively abundant in the south (especially among samples collected outside rice fields), together with higher numbers of fern spores (mainly Dicranopteris), while the frequencies of other herb and tree pollen is higher in the north.
Statistical analysisA statistical analysis was carried out to correlate pollen taxa. We performed principal component analysis of the main 21 pollen taxa from 25 rice field sample sites using MVSP software pack-age Version 3.1 (Kovach, 1999). The results show that the first axis is significant, with its eigenvalue as high as 47.85%, and a cumulative percentage of 80.71%. The scores loading on the first factor display two groups characterized by positive and negative values on the axis (Figure 4). The taxa with positive loadings are Poaceae (mostly rice), Pinus, Brassicaceae, Dicranopteris, Asteraceae, Ranunculaceae and Artemisia, reflecting rice farming
and severe deforestation in the region. On the negative side of the axis are Hamamelidadceae, Papillionaceae, Myrica, Taxodiaceae, Juglans and Quercus, which represent other major taxa in the region. The combination of Poaceae, Pinus and Dicranopteris is a clear indicator of agriculture and forest clearance.
The relative frequency of Poaceae pollen is highest among the rice fields of Hunan Province, an area famous for rice production, where it averages 76.2% (Table 4). In all three provinces, how-ever, the total relative abundance of Poaceae is significantly higher inside the rice fields than outside. Beyond this general trend, there are notable differences among results for the three provinces. Hunan and Guangdong have the highest frequencies of Poaceae outside the fields, with averages ranging from 65% to 69%. By contrast, in Hubei Poaceae represents only 40% of the
Figure 3. Pollen diagram of principal taxa from samples outside of rice fields. The regional divisions of the three Provinces are indicated.
Figure 4. Loadings of the principal pollen taxa on the first factor axis for the samples inside rice fields, showing that Pinus, Brassicaceae and Dicranopteris Asteraceae, Ranunculaceae and Artemisia are positively associated with Poaceae (mostly rice pollen).

1398 The Holocene 22(12)
pollen assemblage collected in areas surrounding the fields. These results imply that, in fossil pollen assemblages evidence for rice cultivation may be obscured by two factors: (1) ancient rice fields contained greater numbers of weeds than modern fields, and (2) most fossil pollen assemblages will represent areas outside, rather than inside former rice fields.
Certain pollen taxa, including Dicranopteris, herbs such as Artemisia and some secondary trees (Pinus and Taxodiaceae) are indirectly but closely associated with agriculture. For example, Brassicaceae pollen is common in the surface soil of cultivated land and it is possibly related to vegetable production. Among these taxa indicative of highly anthropogenic landscapes, the most highly represented in our study are Dicranopteris (15%~30%), herbs such as Artemisia and some secondary trees (Pinus and Taxodiaceae). Dicranopteris is a heliophilous fern of tropical-subtropical hilly areas where it thrives in areas disturbed by forest clearance and which have been repeatedly burned. Dicranopteris is also closely linked with land clearing for agricul-ture and human disturbance in general. It is well adapted to acidic soils such as those common in Guangdong, and it often grows in association with planted Pinus massonia and other pioneer vege-tation. In Hunan and Hubei, where the soils are generally less acidic than in Guangdong, Dicranopteris spores occur at lower frequencies.
Artemisia becomes increasingly significant in the northern reaches of our sampling transect, particularly in Hubei (Figure 3). Previous studies link dramatic increases in Artemisia pollen with intensive human activity, especially in the temperate floodplains (Zheng et al., 2007). Artemisia is the most abundant NAP in the dust flux samples, with peak values recorded at latitudes 44°–48°N and 35°–37°N, corresponding to the Northeast and North China Plain regions (Zheng et al., 2007). Levels of Artemisia decrease gradually from north to south, before falling off rapidly to 0.5% at 20°N latitude in eastern China. The ratio of Poaceae to Artemisia pollen is also a useful latitudinal indicator (Zheng et al., 2007). Our study shows Artemisia present at frequencies of 5–10% in rice fields in the north of our transect, but falling off to lower levels the south.
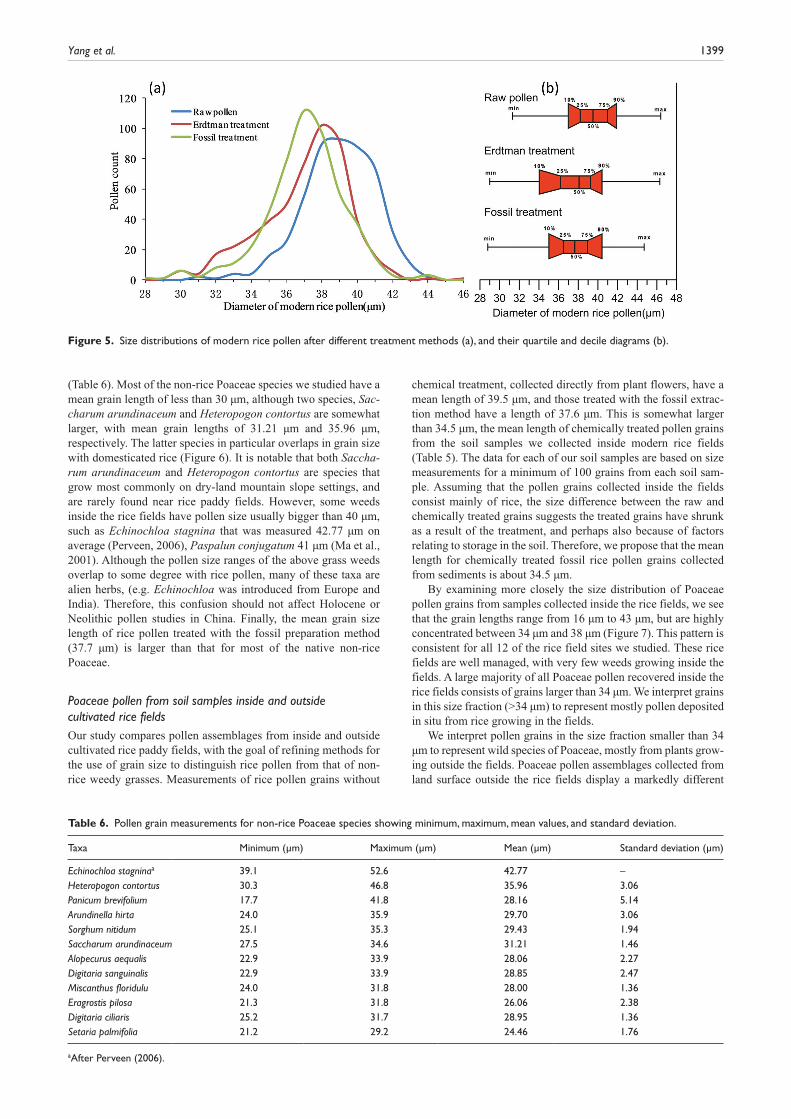
Rice pollen morphologyPollen morphology of rice (Oryza sativa L)Rice (Oryza sativa L) pollen is ovoid or nearly spherical in shape. The grains are monoporate, circumscribed by an annulus with a thickening belt about 2–3 µm in bandwidth. Microscopy shows that the outline of rice pollen grains is smooth, with a fuzzy sculpture. Previous studies indicate that chemical treatment of pollen affects the overall size of the grains, as well as the size of aperture (Andersen, 1960; Meltsov et al., 2008; Reitsma, 1969; Sluyter, 1997). The influence of chemical treatment and storage on pollen grain size differs, however, by treatment method (Schüler, 2011). For exam-ple, while acetolysis swells the grains, hydrofluoric acid causes them to shrink (Martin, 2000). For this study, we compared the size of rice pollen grains before and after two chemical treatment meth-ods. Grain size length for the raw, untreated grains varies from 31.3 to 46.4 µm, with an average of 39.5 µm, while pore size ranges from 3.53 to 6.86 µm with an average of 5.01 µm (Table 5). We compared the effects of the Erdtman acetolysis and fossil chemical treatments on pollen grain size. As shown in Table 5 and Figure 5, both methods cause the pollen grains and their apertures to shrink. Thus studies involving the identification of fossil rice pollen must be based on measurements that account for the effects of chemical treatment on grain size. Rice pollen treated with the standard fossil extraction method shrinks to a mean size of 37.6 µm, and aperture size was also reduced. Although the size range for pollen treated with the fossil extraction method is from 28.8 µm to 44.7 µm, 80% of the grains measure between 35 µm to 40.4 µm, with an average of 37.6 µm.
Pollen morphology of wild grassesFor comparison with rice, we also collected and studied pollen from eleven other species of Poaceae, including the most com-mon wild grasses in our study area. We collected the pollen grains directly from the plants and prepared pollen slides using the stan-dard chemical treatment method for fossil grains. For each spe-cies, 200 grains were measured to obtain grain size length data
Table 5. Pollen grain length and aperture diameter size for rice (Oryza sativa L) pollen before and after chemical treatment, compared with size of all Poaceae pollen grains from soil samples collected inside rice fields.
Treatment methods Sample size Minimum Decile (10%) Quartile (25%) Median Mean Quartile (75%) Decile (90%) Maximum
Pollen size of rice (µm)Untreatment 500 31.3 37 38.2 39.5 39.5 41 41.95 46.4Erdtman treatment 500 29 34 36.2 38.1 37.65 39.3 40.3 46.3Fossil treatment 500 28.8 35 36.5 37.7 37.6 38.9 40.4 44.7Size of aperture (µm)Untreatment 200 3.53 – – 4.98 5.01 – – 6.86Erdtman treatment 200 3.52 – – 4.54 4.57 – – 5.93Fossil treatment 200 3.37 – – 4.4 4.45 – – 5.86Pollen size of all Poaceae from soil samples inside rice fieldsFossil treatment 1200 16.8 26.63 33.09 35.58 34.5 37.48 39 46.82
Table 4. Comparison of the average pollen content among the samples inside and outside the rice cultivated land.
Region Sampling location Arboreal (%) Non-arboreal (%) Fern (%) Poaceae (%)
Hubei Prov. Inside rice fields 17.85 16.74 1.97 65.41 Outside rice fields 28.95 30.69 1.71 40.36Hunan Prov. Inside rice fields 11.20 13.60 3.23 76.20 Outside rice fields 19.16 11.40 3.54 69.44Guangdong Prov. Inside rice fields 13.63 19.60 17.6 66.67 Outside rice fields 12.24 22.04 11.28 65.72
Yang et al. 1399
(Table 6). Most of the non-rice Poaceae species we studied have a mean grain length of less than 30 µm, although two species, Sac-charum arundinaceum and Heteropogon contortus are somewhat larger, with mean grain lengths of 31.21 µm and 35.96 µm, respectively. The latter species in particular overlaps in grain size with domesticated rice (Figure 6). It is notable that both Saccha-rum arundinaceum and Heteropogon contortus are species that grow most commonly on dry-land mountain slope settings, and are rarely found near rice paddy fields. However, some weeds inside the rice fields have pollen size usually bigger than 40 µm, such as Echinochloa stagnina that was measured 42.77 µm on average (Perveen, 2006), Paspalun conjugatum 41 µm (Ma et al., 2001). Although the pollen size ranges of the above grass weeds overlap to some degree with rice pollen, many of these taxa are alien herbs, (e.g. Echinochloa was introduced from Europe and India). Therefore, this confusion should not affect Holocene or Neolithic pollen studies in China. Finally, the mean grain size length of rice pollen treated with the fossil preparation method (37.7 µm) is larger than that for most of the native non-rice Poaceae.
Poaceae pollen from soil samples inside and outside cultivated rice fieldsOur study compares pollen assemblages from inside and outside cultivated rice paddy fields, with the goal of refining methods for the use of grain size to distinguish rice pollen from that of non-rice weedy grasses. Measurements of rice pollen grains without
chemical treatment, collected directly from plant flowers, have a mean length of 39.5 µm, and those treated with the fossil extrac-tion method have a length of 37.6 µm. This is somewhat larger than 34.5 µm, the mean length of chemically treated pollen grains from the soil samples we collected inside modern rice fields (Table 5). The data for each of our soil samples are based on size measurements for a minimum of 100 grains from each soil sam-ple. Assuming that the pollen grains collected inside the fields consist mainly of rice, the size difference between the raw and chemically treated grains suggests the treated grains have shrunk as a result of the treatment, and perhaps also because of factors relating to storage in the soil. Therefore, we propose that the mean length for chemically treated fossil rice pollen grains collected from sediments is about 34.5 µm.
By examining more closely the size distribution of Poaceae pollen grains from samples collected inside the rice fields, we see that the grain lengths range from 16 µm to 43 µm, but are highly concentrated between 34 µm and 38 µm (Figure 7). This pattern is consistent for all 12 of the rice field sites we studied. These rice fields are well managed, with very few weeds growing inside the fields. A large majority of all Poaceae pollen recovered inside the rice fields consists of grains larger than 34 µm. We interpret grains in this size fraction (>34 µm) to represent mostly pollen deposited in situ from rice growing in the fields.
We interpret pollen grains in the size fraction smaller than 34 µm to represent wild species of Poaceae, mostly from plants grow-ing outside the fields. Poaceae pollen assemblages collected from land surface outside the rice fields display a markedly different
Figure 5. Size distributions of modern rice pollen after different treatment methods (a), and their quartile and decile diagrams (b).
Table 6. Pollen grain measurements for non-rice Poaceae species showing minimum, maximum, mean values, and standard deviation.
Taxa Minimum (µm) Maximum (µm) Mean (µm) Standard deviation (µm)
Echinochloa stagninaa 39.1 52.6 42.77 –Heteropogon contortus 30.3 46.8 35.96 3.06Panicum brevifolium 17.7 41.8 28.16 5.14Arundinella hirta 24.0 35.9 29.70 3.06Sorghum nitidum 25.1 35.3 29.43 1.94Saccharum arundinaceum 27.5 34.6 31.21 1.46Alopecurus aequalis 22.9 33.9 28.06 2.27Digitaria sanguinalis 22.9 33.9 28.85 2.47Miscanthus floridulu 24.0 31.8 28.00 1.36Eragrostis pilosa 21.3 31.8 26.06 2.38Digitaria ciliaris 25.2 31.7 28.95 1.36Setaria palmifolia 21.2 29.2 24.46 1.76
aAfter Perveen (2006).

1400 The Holocene 22(12)
size distribution pattern characterized by a broad, unimodal distri-bution pattern with a broad peak value span around 25–36 µm (Figure 7). This pattern suggests that pollen from soil samples col-lected outside the rice fields contains highly mixed assemblages of both rice and wild non-rice Poaceae. Thus the contrasting size dis-tribution patterns are valuable for establishing a grain size thresh-old for distinguishing rice pollen from that of other Poaceae species. Our results indicate that based on grain length, the size threshold for differentiating rice Poaceae from wild non-rice Poa-ceae lies at ~34 µm and the pollen grains measuring between 34 and 40 µm consist mainly of rice.
A Holocene pollen record from the Pearl River DeltaArchaeologists and palaeoecologists have long recognized that changes over time in Poaceae pollen can be a valuable proxy for anthropogenically induced land cover change. However, in areas where rice appears during the Neolithic era as a major economic crop, the quantitative analysis of Poaceae pollen from sediment cores has further significance. In southern China, as well as throughout much of East Asia, changing frequencies of Poaceae pollen are important for identifying the beginning of
Figure 7. Size distributions of Poaceae pollen from inside and outside of the rice cultivated fields. Graphs (a) and (c) show the diameter ranges of Poaceae pollen counted from different sites inside (a) and outside (c) of rice fields. (b) and (d) 12 sites inside (b) and outside (d) of the rice fields.
Figure 6. Pollen diameter(µm) ranges of the principal local wild grasses. Minimum, maximum and mean values are shown for each species, and the decile values (10 and 90) for Oryza sativa are indicated.

Yang et al. 1401
rice cultivation and agriculture (e.g. Atahan et al., 2008; Innes et al., 2009; Zheng et al., 2009). Our study advances these efforts by refining the size criteria for distinguishing rice from non-rice Poaceae pollen. We illustrate the application of our approach by applying it to the study of a sediment core from the deltaic plain of the Pearl River.
The pollen results from this core can be divided into five pol-len zones spanning a continuous time sequence from around 5500 to 500 cal. yr BP (Figure 8). Zones I and II are dominated by arboreal pollen, consisting mainly of Castanopsis, Pinus, Quer-cus and Altingia. By contrast, zones III, IV and V are character-ized by high percentages of Poaceae, Dicranopteris, and Pinus, together with increasing frequencies of Artemisia, Chenopodia-ceae, and Cyperaceae. The most remarkable feature of the pollen diagram is an abrupt rise in Poaceae, at about 2200 cal. yr BP, from less than 10% in the zones I–II to over 45% in zone III. At its peak in zone III, Poaceae reaches a maximum value of 68.5%. Notably, the other taxa associated with Poaceae in zones III–V, including Dicranopteris, Pinus, Artemisia and Chenopodiaceae, are ones typical of modern surface soil pollen assemblages of southern China. High frequencies of Dicranopteris, in particular, are indicative of forest clearance, as shown by the Guangdong Province surface transect data (Figure 2).
The high levels of Poaceae pollen, in combination with sig-nificant frequencies of pioneer plants such as Dicranopteris, Arte-misia, Chenopodiaceae and Pinus, suggest the intensification of agriculture as part of the development of a strongly anthropo-genic, cultural landscape by human since 2200 cal. yr BP. The timing of this dramatic transition coincides with rapid formation of the Pearl River delta plain (Zong et al., 2009b).
The grain size distribution pattern of pollen from zone III is markedly different from that for the Poaceae modern samples col-lected inside rice fields, but more similar to the pattern for sam-ples from outside rice fields (Figure 9). Paleogeographic reconstruction of the Pearl River delta is useful for understanding the depositional setting for the zone III sediments. As shown by a recent study documenting the evolution of the Pearl River delta (Zong et al., 2009b), at around 2200 cal. yr BP the location marked by core GZ-2 was a subaqueous delta front environment receiving fluvial sediments. Thus the zone III sediments for core GZ-2 do not represent in situ rice agriculture; rather they
represent rice agricultural activity in the apex area of the Pearl River delta, recorded by rice pollen transported by the river to the core site. This helps to explain why the zone III sediments contain highly mixed assemblages of both rice and wild non-rice Poaceae. Furthermore, the ancient Chinese rice farming systems of 2000 years ago were less intensive than their modern analogs (Chao, 1986; Ruddiman and Erle, 2009), and the comparatively primitive irrigation techniques may have resulted in large numbers of weeds (wild Poaceae) growing with rice in the paddy fields.
Modern analog analysis is valuable in allowing a quantitative comparison of the fossil pollen assemblages from core GZ-2 with modern assemblages. We used pollen assemblages from our lati-tudinal transect study as the modern analogs. The analog method is based on the assumption that if a fossil pollen assemblage closely matches a modern pollen assemblage, then the vegeta-tional and environmental setting of the modern assemblage is similar to that which yielded the fossil assemblage (Jackson and Williams, 2004). We used Euclidean Distance as a statistical method to quantify the dissimilarity between fossil and modern pollen samples (e.g. Schweitzer, 1994). A score of 0 indicates exact similarity and higher values indicate increasing dissimilar-ity. The results show that the best matches between fossil and modern pollen samples are for fossil samples in zones III and IV, which represent a time period of intense anthropogenically induced land cover change marked by sharply increases in Poa-ceae and Dicranopteris. The pattern of dissimilarity scores sug-gests a phase of rapid development of rice agriculture in the deltaic plain (Figure 9). The best matches with modern analogs are during the time period marked by sharp increases in Poaceae pollen and Dicranopteris spores.
Evidence for an abrupt transition in the vegetational landscape is significant when considered in the historical context of south-ern China. This transition took place around 2200 years ago, coin-ciding with the end of the Late Warring States Period and the beginning of Qin Dynasty. The population in the Pearl River delta increased rapidly during this period as a result of the inflow of Han immigrants from northern China (He, 2006; Museum of Guangdong Province, 1981). Successive waves of migration stimulated cultural change while also transforming the genetic makeup of southern populations (Wen et al., 2004). Previous studies show that sea water inundated the delta area by 7000 cal.
Figure 8. Pollen percentage diagram of Core GZ-2. All percentages were exaggerated (shadow) by five times (modified from Wang et al., 2010).

1402 The Holocene 22(12)
yr BP (Yuan et al., 1986; Zong, 2004), after which the delta plain advanced progressively (Zheng et al., 2004; Zong et al., 2009b). Archaeological work shows that the Neolithic economy of this region extended through the Spring and Autumn (771–476 bc) and Warring States (475–221 bc) periods. People lived in villages with pottery and other aspects of Neolithic technology, supported mainly by hunting, fishing, and heavy exploitation of shellfish. Agriculture and food production was apparently limited until around 2200 cal. yr BP when the upper part of the delta plain emerged, creating fertile wetlands well suited for agriculture. Our results support the findings of Zong et al. (2010) who suggest that a complex form of agriculture developed about 2000 years ago, involving intensive cultivation of multiple crops, land reclama-tion techniques, and the use of metal tools. Our findings show that intensive rice agriculture did not develop until the Qin Dynasty, more than 4000 years after the emergence of intensive rice culti-vation in the lower Yangtze River area and the northern plain of China.
ConclusionsWe studied pollen assemblages from top soil surface samples col-lected inside rice paddy fields, as well as in neighboring unculti-vated grasslands from a subtropical region of China. Double-cropping rice agriculture is the mainstay of the economy in our study area. Pollen assemblages from the rice paddy fields are characterized by high percentages of Poaceae, with Poaceae accounting for an average of 68.8% of the total assemblage. Other pollen and spores are also indicative of agriculture. The abun-dance of fern spores (Dicranopteris) and weed pollen (small pol-len grain Poaceae and Artemisia) and the low quantity of natural arboreal taxa indicate vegetation disturbance, particularly defor-estation. Statistical analysis helps explain the relationship among
pollen taxa, revealing two distinctive groups. The first group is composed of Poaceae (mostly rice), Pinus, Brassicaceae, Dicranopteris, Asteraceae, Ranunculaceae and Artemisia. Taxa in this group are related to human activity including forest clearance and rice agriculture. The second group of taxa, which includes Hamamelidaceae, Myrica, Taxodiaceae, Juglans and Quercus, represents distant forests and dry land herbs.
Our investigation of rice pollen morphology indicates that the average grain length of rice pollen is 39.5 µm and 37.6 µm, respectively, before and after the standard chemical treatment used for fossil extraction. Rice pollen grains shrank about 1.8 µm in grain size as a result of the treatment and aperture size also was altered. After treatment, 80% of the rice pollen grains measure between 35 µm to 40.4 µm. In comparison, most native species of non-rice Poaceae are smaller in grain size. The main exception, which displays the greatest overlap in grain size with rice, is Het-eropogon, a genus of historic era introductions that usually grows on dry-land hillside and mountain slopes.
The grain size distribution of Poaceae pollen from soil sam-ples collected inside modern rice fields is highly concentrated between 34 µm and 38 µm. This is consistent with the observation that 80% of the raw rice pollen grains measured between 35 µm to 40.4 µm after chemical treatment. Therefore, considering the complex chemical and climatic conditions that affect the size of pollen grains in cultivated soil, it is suggested that 34 µm to 40 µm is a good estimate for the size distribution range of fossil rice pol-len grains. These results are crucial for scientists investigating ancient evidence for anthropogenic influence, especially early rice agriculture, in the double cropping rice area of southern China. As a case study for identifying rice pollen through grain size analysis, we investigated pollen assemblages from a sedi-ment core collected in the Pearl River delta plain. The results show that the high levels of Poaceae pollen, in combination with
Figure 9. Poaceae size distribution for the selected samples of core GZ-2 and dissimilarity scores of the fossil pollen from core GZ-2 with surface sample data from inside rice fields. The black curves in the right figures show the Poaceae size distribution in the selected samples at ages between 2226 to 1692 cal. yr BP; the grey curves show the average frequency of Poaceae grain sizes from samples outside rice fields (Figure 3). The shaded area shows the suggested grain size range for rice pollen.

Yang et al. 1403
significant frequencies of pioneer plants such as Dicranopteris, Artemisia, Chenopodiaceae and Pinus, suggest the intensification of agriculture as part of the development of a strongly anthropo-genic, cultural landscape. The beginning of intensive rice agricul-ture coincides with a phase of rapid development in plain formation which created wetlands suitable for paddy fields. The transition from a broadleaf evergreen forest ecosystem to a herb-dominated environment occurred at about 2200 cal. yr BP. The timing of the transition coincides with a period of human migra-tion from the north and the rapid development of rice cultivation during the Qin Dynasty. These developments are associated with a phase of deforestation indicated by decreases in broadleaved tree pollen and increases in the pioneer plants, particularly Dicranopteris. However, the proportion of rice-type Poaceae as a percentage of the total Poaceae pollen assemblage is low. This can be explained in part by the fact that Qin Dynasty agricultural tech-niques were primitive in comparison with modern cultivation sys-tems, and there were many weeds growing in the fields. It is also significant that the land suitable for irrigated rice was limited and the core sediments do not represent paddy field deposits. During the Qin Dynasty the coring site was situated in estuarine condi-tions, near the delta front but not on the deltaic plain. The fossil pollen assemblage likely reflects a high level of rice agricultural activity upstream on the emerging deltaic plain, in a highly anthropogenic setting characterized by forest clearance and other consequences of human activity.
FundingThis research was funded by the National Science Foundation of China (41072128, 40772113), the Fundamental Research Funds for the Central Universities (11lgjc13, 10lgzd08), the Hong Kong University Seed Funds (2009-02159004) and the General Research Fund of Hong Kong (HKU707109P). The modern pol-len data was collected and analyzed under the frame of National projects (Grants: 40730103 and 40331011). We thank the drilling work that was partly supported by the grant of Guangzhou Qua-ternary Environment Investigation.
ReferencesAndersen ST (1960) Silicon oil as a mounting medium for pollen grains. Dan-
murks geologiske Undersøgelse Series IV 4(1): 24–24.Andersen ST and Bertelsen F (1972) Scanning electron microscope studied of
pollen of cereals and other grsses. Grana 12: 79–86.Atahan P, Davey FI, Taylor D et al. (2008) Holocene-aged sedimentary records
of environmental changes and early agriculture in the lower Yangtze, China. Quaternary Science Reviews 27: 556–570.
Behre KE (1986) Anthropogenic Indicators in Pollen Diagram. Rotterdam: A A Balkema.
Chao K (1986) Man and Land in Chinese History: An Economic Analysis. Stanford: Stanford University Press.
Davies AL and Tipping R (2004) Sensing small-scale human activity in the palaeoecological record: Fine spatial resolution pollen analyses from Glen Affric, northern Scotland. The Holocene 14: 233–245.
Dearing JA, Battarbee RW, Dikau R et al. (2006) Human–environment inter-actions: Learning from the past. Regional Environmental Change 6: 115–123.
Dimbleby GW (1985) The Palynology of Archaeological Sites. London: Aca-demic Press.
Du NQ and Chen YS (1990) Effects of heavy liquid floating on the calculation of pollen concentration. Acta Botanica Sinica 32: 794–798.
Edwards KJ and Hirons KR (1984) Cereal pollen grains in pre-elm decline deposits: Implications for the earliest agriculture in Britain and Ireland. Journal of Archaeological Science 11: 71–80.
Gong ZT, Chen HZ, Yuan DG et al. (2007) The temporal and spatial distribu-tion of ancient rice in China and its implications. Chinese Science Bulletin 52: 1071–1079.
He HL (2006) From uncivilized state to civilized society: The transportation and Yue nationality of Lingnan in the Qin and Han Dynasties. Guizhou Ethnic Studies 26: 201–206.
Innes JB, Zong Y, Chen Z et al. (2009) Environmental history, palaeoecol-ogy and human activity at the early Neolithic forager/cultivator site at
Kuahuqiao, Hangzhou, eastern China. Quaternary Science Reviews 28: 2277–2294.
Itzstein-Davey F, Atahan P, Dodson JR et al. (2007) Environmental and cul-tural changes during the terminal Neolithic: Qingpu, Yangtze delta, east-ern China. The Holocene 17: 875–887.
Jackson ST and Williams JW (2004) Modern analogs in Quaternary paleoecol-ogy: Here today, gone yesterday, gone tomorrow? Annual Review of Earth and Planetary Sciences 32: 495–537.
Köhler E and Lange E (1979) A contribution to distinguishing cereal from wild grass pollen grains by LM and SEM. Grana 18: 133–140.
Kong ZC and Du NQ (1981) Sporo-pollen analysis of some archaeologi-cal sites and preliminary discussion on the past flora and climate of Nei-Monggol Zizhiqu. Acta Phytoecologia et Geobotanica Sinica 5: 193–202.
Kovach WL (1999) MVSP-A Multivariate Statistical Package for Windows,Version 3.1. Pentraeth: Kovach Computing Services, 133 pp.
Li YY, Zhou LP and Cui HT (2008) Pollen indicators of human activity. Chi-nese Sciences Bulletin 53: 1281–1293.
Lu TLD (1999) The Transition from Foraging to Farming and the Origin of Agriculture in China. British Archaeological Reports International Series 774, Oxford: John & Erica Hedges.
Ma GH, Zhao NX, Hu XY et al. (2001) Pollen morphology and poly-aperture in Papalum. Journal of Tropical and Subtropical Botany 9: 201–204 (in Chinese with English abstract).
Martin HA (2000) Re-assignment of the affinities of the fossil pollen type Tri-colpites trioblatus Mildenhall and Pocknall to Wilsonia (Convolvulaceae) and a reassessment of the ecological interpretations. Review of Palaeo-botany and Palynology 111: 237–251.
Meltsov V, Poska A and Saar M (2008) Pollen size in Carex: The effect of different chemical treatments and mounting media. Grana 47: 220–233.
Mo DW, Wang H and Li SC (2003) Effects of Holocene environmental changes on the development of archaeological cultures in different regions of North China. Quaternary Sciences 23: 200–210.
Museum of Guangdong Province (1981) Archaeological achievement in Guangdong, opening a new chapter in the history of Lingnan. Relic’s Edi-torial Committee edit. Thirty Years of Archaeological Works (1949–1979). Beijing: Cultural Relics Publishing House, pp. 328–331.
Perveen A (2006) A contribution to the pollen morphology of family Gra-mineae. World Applied Sciences Journal 1: 60–65.
Reitsma T (1969) Size modification of recent pollen grains under different treatments. Review of Palaeobotany and Palynology 9: 175–202.
Ruddiman WF and Erle CE (2009) Effect of per-capita land use changes on Holocene forest clearance and CO2 emissions. Quaternary Science Reviews 28: 3011–3015.
Schüler LHB (2011) Poaceae pollen grain size as a tool to distinguish past grasslands in South America: A new methodological approach. Vegetation History and Archaeobotany 20: 83–96.
Schweitzer PN (1994) ANALOG: A program for estimating paleoclimate parameters using the method of modern analogs. U.S. Geological Survey Open-file Report, pp. 594–645.
Sluyter A (1997) Analysis of maize (Zea mays subsp. mays) pollen: Normal-izing the effects of microscope-slide mountingmedia on diameter determi-nations. Palynology 21: 35–39.
Wang JH, Wang XJ, Cao LL et al. (2009) The Holocene sporopollen charac-teristics and their paleoenvironmental significance of Core GZ - 2 in Pearl River Delta. Journal of Palaeography 11: 661–669.
Wang XJ, Wang JH, Cao LL et al. (2010) Late Quaternary pollen records and climate significance in Guangzhou. Acta Scientiarum Naturalium Univer-sitatis Sunyatsen I 49: 113–121.
Wen B, Li H, Lu D et al. (2004) Genetic evidence supports demic diffusion of Han culture. Nature 431: 302–305.
Weninger B and Jöris O (2002) Glacial radiocarbon calibration: The CalPal program. In: Higham T, Ramsey CB and Owen C (eds) Radiocarbon and Archaeology. Oxford: Oxford University, pp. 9–15.
Yang YZ, Li FC, Jin ZD et al. (2006) New evidence for the presence of middle Holocene Paddy soil in the Chuodun archaeological site. Quaternary Sci-ences 26: 864–871.
Yasuda Y, Fujiki T, Nasu H et al. (2004) Environmental archaeology at the Chengtoushan site, Hunan Province, China, and implications for envi-ronmental change and the rise and fall of the Yangtze River civilization. Quaternary International 123–125: 149–158.
Yuan JY, Liang GX, Chen MH et al. (1986) The limits of Holocene Transgres-sion in Guangzhou-Huaxian Plain. Acta Scientiarum Naturalium Univer-sitatis Sunyatseni 3: 63–68.
Zheng Y, Sun G, Qin L et al. (2009) Rice fields and modes of rice cultivation between 5000 and 2500 BC in east China. Journal of Archaeological Sci-ence 36: 2609–2616.

1404 The Holocene 22(12)
Zheng Z, Cour P, Huang CX et al. (2007) Dust pollen distribution on a conti-nental scale and its relation to present-day vegetation along north–south transects in east China. Science of China Series D-Earth Science 250: 236–246.
Zheng Z, Deng Y, Zhang H et al. (2004) Holocene environmental changes in the tropical and subtropical areas of the south China and the relation to human activities. Quaternary Sciences 244: 387–393.
Zong Y ( 2004) Mid-Holocene sea-level highstand along southeast coast of China. Quaternary International 117: 55–67.
Zong Y, Chen Z, Innes JB et al. (2007) Fire and flood management of coastal swamp enabled first rice paddy cultivation in east China. Nature 449: 459–463.
Zong Y, Wang Z, Innes JB et al. (2012a) Holocene environmental change and Neolithic rice agriculture in lower Yangtze region of China: A review, China. The Holocene doi:10.1177/0959683611409775.
Zong Y, Innes JB, Wang Z et al. (2012b) Environmental change and Neolithic settlement movement in the lower Yangtze wetlands of China. The Holo-cene doi:10.1177/0959683611414933.
Zong Y, Huang K, Yu F et al. (2012c) The role of sea-level rise, monsoonal dis-charge and the palaeo-landscape in the evolution of the Pearl River delta, southern China. Quaternary Science Reviews in press.
Zong Y, Yim WW-S, Yu F, Huang G (2009a) Late Quaternary environmental changes in the Pearl River mouth region, China. Quaternary International 206: 35-45.
Zong Y, Huang G, Switzer AD et al. (2009b) An evolutionary model for the Holocene formation of the Pearl River delta, China. The Holocene 19: 129–142.
Zong Y, Yu F, Huang G et al. (2010) Sedimentary evidence of Late Holocene human activity in the Pearl River delta, China. Earth Surface Processes and Landforms 35: 1095–1102.




















