
Modelling the dynamics of intramammary E. coli infections in dairy cows: understanding mechanisms...
15
Original article Modelling the dynamics of intramammary E. coli infections in dairy cows: understanding mechanisms that distinguish transient from persistent infections Lisa J. WHITE 1 * , Ynte H. SCHUKKEN 2 , Belgin DOGAN 2 , Laura GREEN 3 , Do ¨rte DO ¨ PFER 4 , Mike J. CHAPPELL 5 , Graham F. MEDLEY 3 1 Mahidol Oxford Research Unit, Faculty of Tropical Medicine, Mahidol University, Bangkok, Thailand 2 Departments of Population Medicine and Diagnostic Sciences, and Clinical Sciences, Cornell University, Ithaca, NY, 14853, USA 3 Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, United Kingdom 4 Department of Medical Sciences, University of Madison, Wisconsin-Madison, Madison, WI, 53706, USA 5 School of Engineering, University of Warwick, Coventry, CV4 7AL, United Kingdom (Received 6 March 2009; accepted 15 October 2009) Abstract – The majority of intramammary infections with Escherichia coli in dairy cows result in transient infections with duration of about 10 days or less, although more persistent infections (2 months or longer) have been identified. We apply a mathematical model to explore the role of an intracellular mammary epithelial cell reservoir in the dynamics of infection. We included biological knowledge of the bovine immune response and known characteristics of the bacterial population in both transient and persistent infections. The results indicate that varying the survival duration of the intracellular reservoir reproduces the data for both transient and persistent infections. Survival in an intracellular reservoir is the most likely mechanism that ensures persistence of E. coli infections in mammary glands. Knowledge of the pathogenesis of persistent infections is essential to develop preventive and treatment programmes for these important infections in dairy cows. mathematical model / mastitis / Escherichia coli / persistent infection 1. INTRODUCTION Mastitis is one of the most important produc- tion and welfare diseases affecting dairy cows. In recent decades, coliform infections have been identified in an ever increasing proportion of mastitis cases [22]. Among the coliform bacteria Escherichia coli is the dominant pathogenic spe- cies in dairy cow mastitis [5, 26]. The typical infection pattern for E. coli intramammary infection includes a clinically severe inflamma- tory response by the host with outcomes of either elimination of coliforms within 96 h of the initial infection [20, 23] or a deleterious outcome for the host including shock, sepsis and often death [8, 9]. In addition to severe disease several authors have reported persistent and clinically less severe E. coli infection [11, 12, 15]. Persistent E. coli infections are not uncom- mon in other organs or species. In particular, persistent infections of the human female uri- nary tract are often observed, and have inspired * Corresponding author: [email protected] Vet. Res. (2010) 41:13 DOI: 10.1051/vetres/2009061 Ó INRA, EDP Sciences, 2009 www.vetres.org This is an Open Access article distributed under the terms of the Creative Commons Attribution-Noncommercial License (http://creativecommons.org/licenses/by-nc/3.0/), which permits unrestricted use, distribution, and reproduction in any noncommercial medium, provided the original work is properly cited. Article published by EDP Sciences
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Modelling the dynamics of intramammary E. coli infections in dairy cows: understanding mechanisms...

Original article
Modelling the dynamics of intramammaryE. coli infections in dairy cows:
understanding mechanisms that distinguish transientfrom persistent infections
Lisa J. WHITE1*, Ynte H. SCHUKKEN2, Belgin DOGAN
2, Laura GREEN3,
Dorte DOPFER4, Mike J. CHAPPELL
5, Graham F. MEDLEY3
1 Mahidol Oxford Research Unit, Faculty of Tropical Medicine, Mahidol University, Bangkok, Thailand2 Departments of Population Medicine and Diagnostic Sciences, and Clinical Sciences,
Cornell University, Ithaca, NY, 14853, USA3 Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, United Kingdom
4 Department of Medical Sciences, University of Madison, Wisconsin-Madison, Madison, WI, 53706, USA5 School of Engineering, University of Warwick, Coventry, CV4 7AL, United Kingdom
(Received 6 March 2009; accepted 15 October 2009)
Abstract – The majority of intramammary infections with Escherichia coli in dairy cows result in transientinfections with duration of about 10 days or less, although more persistent infections (2 months or longer)have been identified. We apply a mathematical model to explore the role of an intracellular mammaryepithelial cell reservoir in the dynamics of infection. We included biological knowledge of the bovineimmune response and known characteristics of the bacterial population in both transient and persistentinfections. The results indicate that varying the survival duration of the intracellular reservoir reproduces thedata for both transient and persistent infections. Survival in an intracellular reservoir is the most likelymechanism that ensures persistence of E. coli infections in mammary glands. Knowledge of thepathogenesis of persistent infections is essential to develop preventive and treatment programmes for theseimportant infections in dairy cows.
mathematical model / mastitis / Escherichia coli / persistent infection
1. INTRODUCTION
Mastitis is one of the most important produc-tion and welfare diseases affecting dairy cows.In recent decades, coliform infections have beenidentified in an ever increasing proportion ofmastitis cases [22]. Among the coliform bacteriaEscherichia coli is the dominant pathogenic spe-cies in dairy cow mastitis [5, 26]. The typicalinfection pattern for E. coli intramammary
infection includes a clinically severe inflamma-tory response by the host with outcomes of eitherelimination of coliformswithin 96 h of the initialinfection [20, 23] or a deleterious outcome forthe host including shock, sepsis and often death[8, 9]. In addition to severe disease severalauthors have reported persistent and clinicallyless severe E. coli infection [11, 12, 15].
Persistent E. coli infections are not uncom-mon in other organs or species. In particular,persistent infections of the human female uri-nary tract are often observed, and have inspired* Corresponding author: [email protected]
Vet. Res. (2010) 41:13DOI: 10.1051/vetres/2009061
� INRA, EDP Sciences, 2009
www.vetres.org
This is an Open Access article distributed under the terms of the Creative Commons Attribution-Noncommercial License(http://creativecommons.org/licenses/by-nc/3.0/), which permits unrestricted use, distribution, and reproduction in anynoncommercial medium, provided the original work is properly cited.
Article published by EDP Sciences

research into the pathogenesis of persistentE. coli infections [1, 6, 7, 14, 16, 21]. Thereis still debate about the precise mechanismsinvolved with persistent urinary tract infections,but it is clear that fimbriae that enable adhesionto bladder epithelial cells [6] and subsequentinvasion of bladder epithelial cells, togetherwith prolonged intracellular survival of the bac-teria play an important role in persistence ofinfection [1]. A number of similarities betweenpersistent urinary tract and persistent intramam-mary E. coli infections have been noted [11].
Recent studies on bovine infection patternsand immune response mechanisms have identi-fied complex patterns of innate and adaptiveimmune mechanisms [2, 3] after intramammarychallenge with E. coli and other mastitis – caus-ing pathogens. Lipopolysacharides (LPS) in thebacterial cell wall are recognized by host cellreceptors (such as Toll-like receptor-4) on mac-rophages that circulate in milk. The recognitionof LPS by the host cell receptors leads to acascade of events and eventually to anincreased production of pro-inflammatory cyto-kines [2, 3]. These cytokines have, amongstother characteristics, a chemotactic effect onpolymorphonuclear cells (PMN) in the blood-stream, which are, consequently, attracted intothe mammary gland towards the site of infec-tion where they attempt to phagocytise and killthe bacterial invaders [9, 23]. The outcome ofthe bacterial invasion into the mammary glandis eventually determined by the host responseand the ability of the bacterial pathogen to cir-cumvent or evade this host response [9, 20].One of the mechanisms that bacterial pathogensemploy to persist in the host is the creation ofan intracellular niche [6], the importance of thisin E. coli mastitis has not yet been established.
Mathematical modelling may be valuable inproviding insight into the mechanisms of persis-tent infections, both at the population level [28]and the individual animal level [10, 17]. Themodel described by Detilleux et al. [10] modelstransient infections and includes the compart-ments: milk, blood leukocytes and bacteria inmilk. This simple model was fitted to transientinfection data, but it does not adequatelydescribe the complex pathogenesis of bacterial
infections that is essential to reproduce thedynamics of both transient and persistent intra-mammary infections.
In this study, we aim to improve understand-ing of the pathogenesis of transient and persis-tent intramammary E. coli infections. Inparticular, we are interested in understandingthe processes that underlie the divergence inE. coli infection outcome (elimination or persis-tence): why do a majority of intramammaryE. coli infections results in transient infectionsand how do other E. coli intramammary infec-tions result in persistent infections?
2. MATERIALS AND METHODS
2.1. The model
We model the interaction between the bovineimmune system and numbers of E. coli in milk andin intracellular reservoirs in a quarter (1 of the 4 com-partments of the bovine mammary gland). We assumethat the total number of colony forming units (cfu) ofE. coli inmilk (E) grows exponentially, but the bacteriaare phagocytised and killed bymacrophages and PMN[18]. Bacteria can move between milk and an intracel-lular mammary epithelial cell reservoir (I) [11]. Fromnumerous challenge infections it is known that produc-tion of pro-inflammatory cytokines (C+) is stimulatedby contact between bacteria and immune cells, eithermacrophages (M) or PMN (P) [4]. The production ofpro-inflammatory cytokines is inhibited by the pres-ence of anti-inflammatory or inhibitory cytokines(C�), which increase at a rate dependent on the con-centration of pro-inflammatory cytokines, thus thereis a negative feedback loop [2–4]. The macrophagespresent in the milk are assumed to remain at a constantconcentration, whereas the concentration of PMNincreases at a rate dependent on the concentration ofpro-inflammatory cytokines present in the milk [23].
An additional process that determines the bacte-rial dynamics is the twice daily process of milkingwhich rapidly depletes the volume of milk in themammary gland, and so changes the population sizeof the bacterial cells. We assume a time dependentfunction, V, that defines the linearly changing volumeof milk in the quarter (Fig. 1).
This simplified description of inflammatorydynamics leads to the following series of differentialequations for the concentrations of cytokines (C+
and C�), and the numbers of PMN (P) and E. coli
Vet. Res. (2010) 41:13 L.J. White et al.
Page 2 of 15 (page number not for citation purpose)

cfu (E) in milk (which decreases due to milking,see Appendix for derivation) and intracellularE. coli (I):
dCþdt¼
gcp
ð1þ icpC�ÞEV 2 M þ Pð Þ � dcCþ
dC�dt¼ gcaCþ � dcC�
dEdt¼ geE � kE
M þ Pð ÞV
� aE þ bI þ EV
dVdt
dPdt¼ gpCþV � dpP þ
PV
dVdt
dIdt¼ aE � bI � d iI
v ¼1�pmilk
0:5�Dmilk
modð2t; 1Þ2
h iþ pmilk
for mod 2t; 1ð Þ � 1� 2Dmilk
pmilk�1Dmilk
modð2t; 1Þ2� 0:5
h iþ pmilk
for mod 2t; 1ð Þ > 1� 2Dmilk
9>>>>>>>=>>>>>>>;
V ¼ V max v
dVdt¼
0pmilk � 1
Dmilk
for mod 2t; 1ð Þ � 1� 2Dmilk
for mod 2t; 1ð Þ � 1� 2Dmilk
9=;ð1Þ
In all equations, t represents time in days. The ini-tial values of these variables were set to 0, exceptfor E, which had a low initial infection level of1 cfu/mL (or 250 cfu in total immediately aftermilking) representing initial invasion. The baselinelevel for M was set at 5 000 and P was set at 25%of 50 000 (see below). The parameters are definedin Table I and discussed in more detail below.
To explore fadeout of the bacterial population,the model equations for the bacterial populations(E and I) were discretised using the Euler approxima-tion with an interval, dt, of 1 s. The equations for theconcentrations of immune cells and cytokinesremained deterministic and continuous. Thus
producing a stochastic version of the bacterial popu-lation section of the model as follows:
Etþdt ¼ max
�H
�Et þ
�geEt � kEt M þ Pð Þ
� aEt þ bI t þdVdt
Et
V
��dt�; 0
�; ð2Þ
I tþdt ¼ max H I t þ aEt � bI t � d iI tð Þ�dt½ �; 0f gwhere H[x] is sampled from a Poisson distributionof mean x. The Poisson distribution is a discretedistribution and expresses the probability of a num-ber of events (of the type that occur with a knownaverage rate and independently of the time since theprevious event) occurring in a fixed period of time.An event in this case is defined as a change in thepopulation size due to growth, killing or milking.This distribution therefore appears to be the mostappropriate for this system.
2.2. Biological data
2.2.1. Differential cell counts in milk
Basic milk somatic cell counts (SCC) at the time ofinitial infectionwere estimated using data fromhealthynon-infected quarters [13]. Differential milk leukocytecounts for healthy cows (n = 21) with low SCC milk(SCC < 200 000 cells/mL, [25]) in early lactation(22 ± 4 d after calving), mid lactation (185 ± 6 d aftercalving) and late lactation (245 ± 12 d after calving)were used [12]. Approximately 10% of all somaticcells in a healthy quarter are macrophages and 25%PMN. If we assume 50 000 cells/mL in a healthy quar-ter [24], then M
V = 5 000 cells/mL and approximately25% would be PMN.
Washout of cells and bacteria due tomilking is esti-mated at about 90% (see also [10]). With a milk yieldof 2.5 kg per quarter (i.e. Vmax = 2 500 mL) andacceptablemilkwithhold of 0.25 kg at the end ofmilk-ing, it is reasonable to assume that 90% of cells andbacteria are removed at each milking (i.e. pmilk = 0.1).
2.2.2. Killing capacity of PMN
No direct data on bacterial killing were availablefor bovine intramammary PMN or macrophages.To obtain a quantitative estimate of killing capacitywe utilized data from a mouse model of E. coli infec-tion published by Iwahi and Imada [16], in whichthe susceptibility of E. coli to killing by murine per-itoneal exudate PMN was evaluated. A 0.05-mLvolume of bacterial suspension (2 · 107 to
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 3 of 15

8 · 107 cfu/mL) was mixed with 0.25 mL of PMNsuspension (6 · 106 to 8 · 106 cells/mL). A killingrate of approximately 1 log per hour was observed,with a decreasing rate of killing over time. Thisestimate was very similar to an estimate used byDetilleux et al. [10]. This corresponds to an estimateof k between 4.8 · 10�6 and 3.6 · 10�6 cell/day (SeeAppendix for the derivation of these values).
2.2.3. Growth of E. coli in vitro and in vivo
Data on growth of E. coli both in vitro and in vivowere taken from experiments by Kornalijnslijperet al. [18, 19], in which 200 lL of bacterial suspen-sion (aiming at an inoculum of 100 cfu) was added to10 mL whole milk. Bacterial counts in whole milkafter 6 h of incubation varied up to a 100-foldbetween individual cows (3.8–6.0 log10 cfu/mL)[18]. This corresponds to a value of ge between25.8 and 46.1 day�1 (See Appendix for the deriva-tion of these values).
2.2.4. Cytokine dynamics
Data on cytokine dynamics during E. coli infec-tion were taken from Bannerman et al. [2, 4]. In this
study, cows were challenged with approximately72 cfu of E. coli. Sterile milk samples were collectedfrom all quarters at 0, 8, 16, 24, 32, 40 and 48 h afterchallenge and evaluated for cytokine profiles, cfu andSCC. An ELISA assay was used to measure the con-centrations of IL-8 and TNF-a in milk obtained fromquarters infused with E. coli, and saline controls. Asexpected, pro-inflammatory cytokines increased rap-idly after intramammary challenge [2–4]. An increasewas observed at 8 h after challenge and the concen-tration peaked at 16 h after challenge. Pro-inflamma-tory cytokine concentrations then dropped andreached baseline values between 32 and 48 h afterchallenge. Inhibitory cytokines (IL-10) showed arapid increase at 16 h post-challenge (at the peak ofpro-inflammatory cytokines) and peaked at 24 hpost-challenge. Thereafter the concentration droppedand fell back to baseline at 72 h after challenge [2].No data were available in peer-reviewed publicationsfor cytokine profiles in persistent E. coli infections.However, a similar study using a Klebsiella challengeprovides some insight in cytokine dynamics in morepersistent gram-negative infections [3]. The modeloutput was considered with reference to this generalbehavior and timescale. Cytokines have a half-life(T) of between 1 and 24 h [4]. Thus the rate of loss,dc is given by dc ¼ Ln ð2Þ
T : Then dc is between 0.69
21.510.50
1
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
V
Linear increase in volume of milk after milking period
Linear decrease in volume of milk during milking period
Proportion of maximum volume remaining after milking period
Milking period
Time (days)
Figure 1. Annotated functional form (not to scale during milking periods) of the milk volume before,during and after milking.
Vet. Res. (2010) 41:13 L.J. White et al.
Page 4 of 15 (page number not for citation purpose)

Tab
leI.Mod
elparameter
definition
s,initialvalues
andmod
elestimates.
Sym
bol
Definition
Initialvalue
Estim
ated
value
Units
Param
eterswiththesameva
lueforallcows
g cp
Baselinegrow
thrate
ofpro-inflam
matorycytokinesin
absenceof
anti-infl
ammatorycytokines.Stimulated
bycontactbetweenE.coliandmacroph
ages
Unk
nown
0.00
24day�
1
i cp
Inhibitory
effect
ofanti-infl
ammatorycytokineson
grow
thof
pro-inflam
matorycytokines
Unk
nown
0.03
5no
ne
g ca
Rateof
grow
thof
anti-infl
ammatorycytokines(dependent
onconcentrationof
pro-inflam
matorycytokines)
Unk
nown
9.8e-5
g eGrowth
rate
ofE.coliin
milk
25.8–4
6.1
34day�
1
d pRateof
loss
ofPMN
from
milk
0.1–
1.8
0.10
day�
1
d cRateof
loss
ofcytokines
0.7–
1710
day�
1
MBaselinenu
mberof
macroph
ages
inmilk
5000
(fixed)
Cells/m
LVmax
Maxim
umvo
lumeof
milkin
aqu
arter
2.5(fixed)
Litre
Dmilk
Durationof
milking
5/(60
·24
)(fixed)
Day
p milk
Propo
rtionof
milkremaining
aftermilking
0.1(fixed)
Non
ek
Killing
rate
bymacroph
ages
(MandP)
3.6e-6–4
.8e-6
1.0e-5
Cell/day
r EMeanstandard
deviationof
themod
elfitfrom
thedata
forlogE.coliconcentration
Unk
nown
0.54
logcells/mL
r SMeanstandard
deviationof
themod
elfitfrom
thedata
forlogsomatic
cellconcentration
Unk
nown
0.75
logcells/mL
Param
etersthat
hav
eadifferentva
lueforeach
typeof
infection(transient,persistent)
aRateof
mov
ementof
E.colifrom
milkto
intra-cellular
reservoirs
Unk
nown
0.51
(transient)
5.1(persistent)
day�
1
bRateof
mov
ementof
E.colifrom
intra-cellular
reservoirs
tomilk
Unk
nown
0.55
(transient)
7.5e-3
(persistent)
day�
1
Param
etersthat
haveadifferentvalueforeach
individual
cow
d iDeath
rate
ofE.coliin
intra-cellular
reservoirs
�1(transient)
�0.1(persistent)
2.1,
0.9,
1.4,
1.2,
1.4,
1.8,
1.9(transient)
0.1(persistent)
day�
1
g pRateof
influx
ofPMN
into
themilkin
presence
ofpro-inflam
matorycytokines
Unk
nown
0.02
,0.04
,0.02
,0.03
,0.03
,0.50
,2.0,
0.8,
6.0,
2.0,
0.3
day�
1
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 5 of 15

and 17, which are the upper and lower limits used forestimation of this parameter.
2.2.5. Intracellular E. coli
Data on intracellular survival of E. coli were takenfrom an in vitro study [11] in which E. coli strainsfrom transient and persistent field infections werecompared in adhesion and invasion assays. MAC-Tcells were infected at a multiplicity of infection of10 (10 bacteria per epithelial cell) for 1 h. TheE. coli strains associated with persistent infectionswere 10 times more invasive than strains associatedwith transient infections. This translates to the con-straint of the value for the parameter, a, for chronicinfections being 10 times its value for acuteinfections.
Intracellular survival of E. coli strains in MAC-Tcells was studied by gentamycin protection assays.MAC-T cells were infected with E. coli strains for1 h, after which extracellular bacteria were killedby exposure to gentamycin (100 lg/mL), an extracel-lular antibiotic, for 2 h. MAC-T cells were incubatedfor a further 2, 24 and 48 h. The number of intracel-lular bacteria surviving was determined by disruptingthe cell wall and plating and counting cfu. Strains ofE. coli from transient infections showed a decrease ofthe number of viable bacteria to approximately 80%of the initial concentration after 48 h. In contrast, thenumber of cfu for persistent strains had almost dou-bled during the same time period (180% of baseline).These findings are in concordance with similar obser-vations in mouse urinary tract epithelial cells [1, 21].
We use an initial value of di = 0.1 day�1 for chronicinfections. This is equivalent to intracellular survivalfor an average of 10 days [21]. To be conservativewith regard to the importance of the intracellular res-ervoir we initially assumed no growth in the intracel-lular reservoir.
2.2.6. E. coli infection data
E. coli intramammary challenge longitudinalinfection data used in this modelling study were gath-ered from several published [11, 12, 19, 27] andunpublished studies1. We identified a total of 7transient infections (cow numbers: 218, 4 003,4 310, 7 036, 9 658, 2 058 and 2 109, all fromKornalijnslijper et al. [18] and VanWerven et al.[27]) and 4 persistent infections (cow numbers:171 103 (from Dogan et al. [11]), 3a, 3b and 3c(from Dopfer et al. [12]). All data used for simu-lations are summarized in Tables II and III.
2.3. Parameter estimation
The model predictions for the logarithms of theE. coli and somatic cell concentrations generatedusing (1) were fitted to the data. The parameters wereestimated within Berkeley-Madonna [28], by maxi-mising likelihood assuming a normal distribution oferror. The initial values for the parameter estimation
Table II. Data for seven transient infections. Cows 218, 4 003, 4 310, 7 036, 9 658, 2 058, and 2 109 arefrom Van Werven et al. [27]. Time is in days, lg(E) is log10 E. coli cfu/mL, lg(S) is log10 SCC/mL.
Time(days)
Cow number
218 4 003 4 310 7 036 9 658 2 058 2 109
Lg(E) Lg(S) Lg(E) Lg(S) Lg(E) Lg(S) Lg(E) Lg(S) Lg(E) Lg(S) Lg(E) Lg(S) Lg(E) Lg(S)
0 0 5.0 0 4.4 0 4.0 0 4.0 0 4.4 0 4.0 0 5.41 9.0 7.0 5.9 7.0 7.7 6.9 8.7 5.6 8.6 6.7 5.3 7.0 3.7 6.32 5.2 7.0 6.5 7.0 6.2 7.0 6.4 7.0 4.9 7.0 2.4 7.0 0 7.03 4.1 7.0 6.0 7.0 5.0 7.0 5.4 7.0 4.9 7.0 2.4 7.0 0 6.94 3.6 7.0 5.7 5.5 5.1 6.9 5.5 6.8 4.5 6.5 2.2 6.8 0 6.45 2.9 7.0 5.0 6.8 4.5 7.0 4.5 7.0 4.2 6.8 0 7.0 0 6.36 3.2 7.0 4.9 6.4 3.3 7.0 3.9 7.0 3.7 6.8 0 7.1 0 7.07 0 7.0 4.0 6.2 3.0 7.0 2.3 7.0 3.1 7.0 0 6.6 0 6.38 0 7.0 3.4 7.0 2.4 7.0 2.3 7.0 3.4 7.0 0 6.4 0 5.89 0 7.0 3.1 7.0 2.3 7.0 1.7 7.0 2.9 7.0 0 6.2 0 6.010 0 0 0 0 0 0 0
1 Dopfer D., Ph.D. Thesis, Utrecht University,2000.
Vet. Res. (2010) 41:13 L.J. White et al.
Page 6 of 15 (page number not for citation purpose)

Tab
leIII.Dataforfour
persistent
infections.C
ow17
110
3isfrom
Dog
anetal.[11]andcows3a,3band3c
arefrom
Dop
feretal.[12
].Tim
eisin
days,
lg(E)is10
logE.colicfu/mL,lg(S)is10
logSCC/m
L.
Cow
number
Tim
e(days)
17110
3Tim
e(days)
3aTim
e(days)
3bTim
e(days)
3c
lg(E)lg(S)days
lg(E)lg(S)
lg(E)lg(S)days
lg(E)lg(S)
lg(E)lg(S)days
lg(E)lg(S)
lg(E)lg(S)days
lg(E)lg(S)
04.3
413.9
7.1
03.5
6.7
413.6
6.4
02.0
6.7
413.6
6.9
03.0
6.8
412.0
5.7
15.5
422.0
7.1
13.8
6.6
423.4
6.2
12.8
6.6
422.6
6.8
12.0
6.6
422.0
6.5
23.5
431.9
6.7
24.2
6.5
433.8
6.1
22.3
6.8
433.2
6.6
20
6.4
430
6.1
33.0
442.8
7.2
33.5
6.2
443.1
6.9
32.5
6.8
443.4
6.8
30
6.1
442.3
5.8
43.1
453.2
7.0
44.1
6.8
453.3
7.0
42.5
6.6
453.3
6.7
40
5.6
452.0
5.4
55.8
463.3
7.3
52.9
6.8
463.6
6.6
53.5
6.9
462.4
6.8
52.0
5.5
462.3
5.5
64.3
473.7
7.1
62.9
6.7
473.6
6.5
63.3
7.0
472.5
6.8
62.5
5.5
473.1
7.0
74.7
484.8
7.3
72.0
6.5
484.5
6.5
73.5
6.8
482.6
6.6
70
6.3
482.3
6.7
83.9
494.3
5.2
82.0
6.7
493.3
6.5
83.2
6.8
492.7
6.4
82.0
6.2
492.3
6.6
93.9
504.4
5.5
92.0
6.3
503.3
6.3
92.4
6.7
506.3
92.0
6.4
502.3
6.5
104.4
514.3
7.4
103.1
6.7
513.4
6.4
102.3
6.7
513.4
6.4
100
6.1
513.3
6.2
114.5
524.7
7.4
112.3
6.7
523.3
6.4
112.2
6.7
523.3
6.4
112.0
5.8
520
6.4
124.2
535.2
7.4
122.0
6.7
534.0
6.1
122.2
6.5
534.0
6.1
122.3
6.2
532.0
5.9
134.7
545.4
7.0
133.3
6.5
543.8
6.1
132.1
6.5
543.8
6.1
130
5.9
540
5.7
144.0
555.4
7.0
143.2
6.7
553.3
6.9
142.1
6.3
553.3
6.9
142.3
5.7
552.6
6.9
153.9
5.2
565.5
7.0
152.8
6.7
564.0
6.8
152.0
6.0
564.0
6.8
150
6.2
560.0
6.6
164.1
7.3
573.2
7.2
162.5
6.6
573.4
6.6
162.0
6.1
573.4
6.6
162.0
5.3
572.3
6.3
173.0
7.0
582.3
6.6
174.3
6.4
583.7
6.1
172.5
6.7
583.7
6.1
170
5.4
582.0
6.4
183.0
5.9
592.6
6.7
182.8
6.7
594.0
6.2
184.2
6.9
594.0
6.2
182.3
5.8
590
6.0
193.6
7.4
602.9
6.9
192.0
6.7
604.1
6.3
194.4
6.8
604.1
6.3
190
6.4
600
5.9
203.7
7.4
611.8
7.2
202.8
6.5
613.5
6.5
204.7
6.8
613.5
6.5
202.0
6.1
610
5.5
214.1
7.5
623.1
7.4
213.2
6.7
624.1
6.6
213.9
7.0
624.1
6.6
212.0
5.9
622.7
5.5
222.4
7.2
633.5
7.2
223.6
6.5
634.0
6.4
223.6
7.0
634.0
6.4
222.3
6.5
632.0
6.7
231.9
6.9
642.1
7.0
233.5
6.3
644.4
6.1
234.1
6.7
644.4
6.1
232.0
6.3
642.0
6.3
242.3
7.1
652.0
6.4
243.4
6.8
654.5
6.5
244.4
7.0
654.5
6.5
242.5
6.4
652.0
6.0
251.3
7.1
663.5
7.3
252.6
7.0
663.9
6.5
253.5
6.7
663.9
6.5
250
6.3
662.5
6.5
262.4
6.6
672.6
7.2
263.7
6.9
673.8
6.4
263.7
6.9
673.8
6.4
260
6.0
670
5.7
270.0
7.3
682.3
6.6
273.4
6.4
683.5
6.5
274.2
7.0
683.5
6.5
272.3
5.8
686.8
continuedon
next
page
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 7 of 15

Tab
leIII.Con
tinu
ed.
Cow
number
Tim
e(days)
17110
3Tim
e(days)
3aTim
e(days)
3bTim
e(days)
3c
lg(E)lg(S)days
lg(E)lg(S)
lg(E)lg(S)days
lg(E)lg(S)
lg(E)lg(S)days
lg(E)lg(S)
lg(E)lg(S)days
lg(E)lg(S)
280.0
6.9
693.1
7.2
283.7
7.0
693.4
6.4
284.2
6.8
693.4
6.4
282.3
5.9
693.4
6.6
292.5
7.0
702.4
6.7
293.1
7.0
703.3
6.2
293.1
6.8
703.3
6.2
292.0
5.7
703.7
6.1
302.7
6.2
712.4
7.1
302.3
6.6
713.9
6.1
302.0
6.7
713.9
6.1
302.9
5.8
714.0
6.2
312.0
4.1
722.5
6.9
312.9
6.5
724.1
5.9
312.5
6.7
724.1
5.9
312.0
6.5
724.1
6.3
322.5
7.1
731.3
6.6
323.6
6.1
322.9
6.4
322.0
6.1
733.5
6.5
333.3
7.4
743.2
7.4
333.5
6.1
333.3
6.7
332.0
6.2
744.1
6.6
343.1
7.2
752.5
7.2
343.4
6.7
342.0
6.6
342.7
6.0
754.0
6.4
352.7
4.9
353.9
6.5
352.0
6.4
352.0
6.6
764.4
6.1
363.6
6.4
363.8
6.4
363.2
6.6
362.0
6.1
774.5
6.5
373.4
7.0
374.0
6.3
373.5
6.9
372.3
6.6
783.9
6.5
383.9
7.0
384.0
6.6
383.5
6.9
382.5
6.4
793.8
6.4
393.3
7.0
393.0
6.2
393.3
6.8
392.0
6.4
803.5
6.5
403.5
7.1
403.4
6.5
403.6
6.8
402.0
6.1
813.4
6.4
Vet. Res. (2010) 41:13 L.J. White et al.
Page 8 of 15 (page number not for citation purpose)

process are given in Table I. We assume that if thedataset includes a zero value and the model predictsa value less than or equal to zero (that is less than orequal to 1 cell/mL), then this is the best fit for thispoint due to limits of detection. We also assume thatthe persistent infections had reached equilibriumwhen they were sampled.
3. RESULTS
The model reproduced the data with the esti-mated parameter values as given in Table I. Theoptimised negative log-likelihood was 650.35.The model also reproduced both transient andpersistent infection behaviour (see Figs. 2 and 3).
10 L
og E
. col
i/mL
or 1
0 Lo
g S
CC
/mL
0
2
4
6
8
10
12
E. coli: 10 log cfu/mLSCC: 10 log SCC/mL
Cow 218
0
2
4
6
8
10
12
0
2
4
6
8
10
12
0 1 2 3 4 5 6 7 8 9 10
Days0 1 2 3 4 5 6 7 8 9 10
0
2
4
6
8
10
12
Cow 4003
Cow 4310 Cow 7036
Cow 9658
Cow 2109
Cow 2058
Days
Figure 2. Observed (points) and model predicted (lines) bacteria count and SCC data for seven cows withtransient infections. Cow numbers are shown on the top-right of each graph. The model prediction is asingle realisation of the stochastic version of the model including variation in measurement based on thestandard deviations of the model fit to the data (rE and rS).
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 9 of 15

For all cows, themodel consistently predicteda 3 h differencebetween the timing of thepeakofpro- and anti-inflammatory cytokine concentra-tions. The model also predicted that the pro-inflammatory cytokines peak within the rangeof 13 to 22 h after infection for all cows and thatthe anti-inflammatory cytokines show a rapidincrease that started just before the peak in pro-inflammatory cytokines. Figure 4 shows anexample of this predicted behaviour for cownumber 218. The sampling frequency of theexperiment by Bannerman et al. [4] was every8 h. Therefore from the data we can concludethat the delay between peaks should be between4 and 12 h. The model predicts an approximate3 h difference which is somewhat lower thanthe range predicted by experimental data. Thus,the model is qualitatively but not quantitativelycapturing the cytokine dynamics and a morecomplex model of the mechanism would berequired for a more precise quantitative predic-tion of this phenomenon.
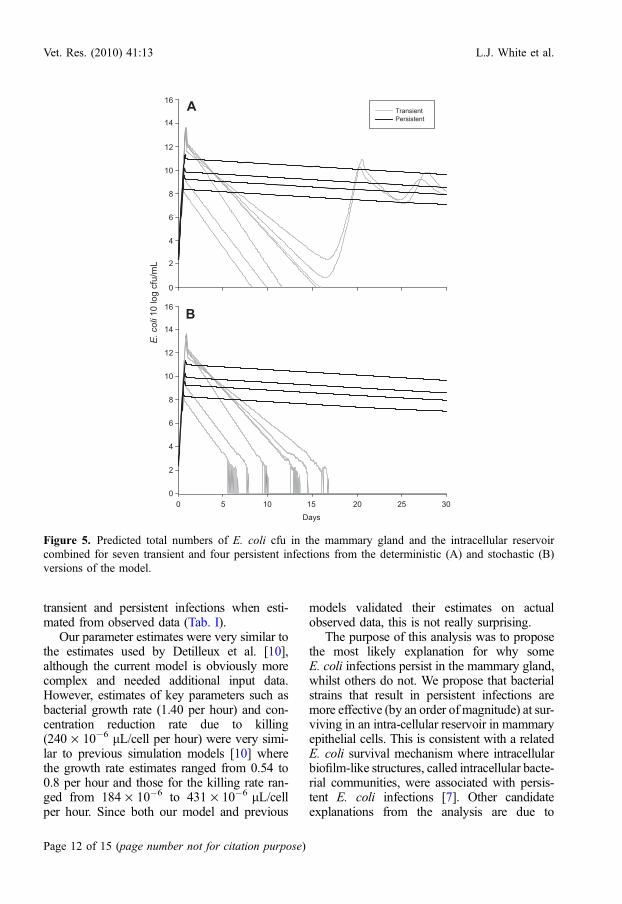
The key result from our model is the transi-tion from transient to steady state dynamics forthe two distinct types of infection. The outputfrom the stochastic and deterministic versionsof the model in Figure 5 is the model predictionsfor the total bacterial population in the milk andthe intracellular reservoir combined. We use thisoutput to demonstrate the potential for the ani-mal to clear the infection due to very low num-bers after the first 5 to 12 days. This wouldcorrespond with an observed clearance in 2 to10 days since only the concentration in the milkwas being observed in the experiment and sothis value would drop below the detection limit(of about 1 cfu/mL) 2 or 3 days before trueclearance by the animal. For transient infections,the number of bacteria drop below 10 cfu (notper mL but in the whole cow), whereas for thepersistent infections the number of bacterianever drop below 106.5 cfu in the wholecow: with approximately 2 500 mL of milk inthe gland (quarter) this would equate to
0 20 40 60 80
E. coli 10 log cfu/mL SCC 10 log SCC/mL
Days0 20 40 60 80
0
2
4
6
8
100
2
4
6
8
10 Cow 3a10
Log
E. c
oli/m
L or
10
Log
SC
C/m
LCow 3b
Cow 3c Cow 171103
Figure 3. Observed (points) and model predicted (lines) bacteria count and SCC data for four cows with apersistent infection. The model prediction is a single realisation of the stochastic version of the modelincluding variation in measurement based on the standard deviations of the model fit to the data (rE and rS).
Vet. Res. (2010) 41:13 L.J. White et al.
Page 10 of 15 (page number not for citation purpose)

approximately 4 · 103 cfu/mL for a persistentlyinfected quarter (see Fig. 3).
This value of the nadir in E. coli bacterialnumbers was highly sensitive to the assumedduration of survival of bacteria in the intracellu-lar reservoir. This duration of survival has athreshold value for simulations of the stochasticmodel of about 1 day where below the thresh-old fadeout was most likely and above thethreshold persistence was more likely. For thesesets of models we assumed this duration to beapproximately 10 days (an alternative estimateof 2 days was also considered with similarresults) for persistent infections and estimated(by fitting) the duration to be about 0.8 daysfor transient infections.
The discretized stochastic model was run1 000 times and fadeout in the transientlyinfected cows and no fadeout in the persistentlyinfected cows was predicted for 996 out of1 000 runs. In four runs one or more of themodelled transient infections did not fadeout.In 1 000 runs the modelled persistent infectionsdid not fadeout. This result confirms the conclu-sion from the deterministic model that the nadirin E. coli bacterial numbers for transient infec-tions after the initial acute phase will generallyresult in fadeout of the bacterial population andthus clearance of infection.
4. DISCUSSION
The mathematical model presented repro-duces the observed dynamic behaviour of bothtransient and persistent infections. The pre-dicted time-dependent behaviour of the pro-and anti-inflammatory cytokine concentrationsis also consistent with our current biologicalknowledge.
We show that for parameter values suitablefor transient infections, the model predicts thatthe total number of bacteria in the mammarygland drops to values at which spontaneousclearance of the infection is highly likely dueto stochastic effects. The predicted minimumlevel of bacteria present is sensitive to changesin the duration of survival in the intracellularreservoir. Long durations of survival in theintracellular reservoir correspond with high bac-teria counts in the nadir of cfu following theacute phase, indicating a failure to spontane-ously clear the infection and thereby leadingto a persistent infection. Hence, the key param-eters distinguishing transient from persistentinfections were the parameters a, b and di(respectively the rates of movement of bacteriabetween the milk and the intra-cellular reservoirand the intracellular bacterial death rate). Thesethree parameters were indeed different between
Time (hours)010 12 16 20 24 28 32 36 40 44 48
Pro
-infla
mm
ator
y cy
toki
ne (u
nits
/mL)
0.0
2.0e+7
4.0e+7
6.0e+7
8.0e+7
1.0e+8
1.2e+8
1.4e+8
Ant
i-inf
lam
mat
ory
cyto
kine
(uni
ts/m
L)
0
2e+3
4e+3
6e+3
8e+3
1e+4Pro-inflammatoryAnti-inflammatory
Figure 4. Dynamics of the concentrations of pro- and anti-inflammatory cytokines predicted by the modelfor cow number 218.
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 11 of 15
transient and persistent infections when esti-mated from observed data (Tab. I).
Our parameter estimates were very similar tothe estimates used by Detilleux et al. [10],although the current model is obviously morecomplex and needed additional input data.However, estimates of key parameters such asbacterial growth rate (1.40 per hour) and con-centration reduction rate due to killing(240 · 10�6 lL/cell per hour) were very simi-lar to previous simulation models [10] wherethe growth rate estimates ranged from 0.54 to0.8 per hour and those for the killing rate ran-ged from 184 · 10�6 to 431 · 10�6 lL/cellper hour. Since both our model and previous
models validated their estimates on actualobserved data, this is not really surprising.
The purpose of this analysis was to proposethe most likely explanation for why someE. coli infections persist in the mammary gland,whilst others do not. We propose that bacterialstrains that result in persistent infections aremore effective (by an order of magnitude) at sur-viving in an intra-cellular reservoir in mammaryepithelial cells. This is consistent with a relatedE. coli survival mechanism where intracellularbiofilm-like structures, called intracellular bacte-rial communities, were associated with persis-tent E. coli infections [7]. Other candidateexplanations from the analysis are due to
Days
0 5 10 15 20 25 300
2
4
6
8
10
12
14
16
E. c
oli 1
0 lo
g cf
u/m
L
0
2
4
6
8
10
12
14
16TransientPersistent
A
B
Figure 5. Predicted total numbers of E. coli cfu in the mammary gland and the intracellular reservoircombined for seven transient and four persistent infections from the deterministic (A) and stochastic (B)versions of the model.
Vet. Res. (2010) 41:13 L.J. White et al.
Page 12 of 15 (page number not for citation purpose)

differences at the host response rather than thebacterial strain level [8, 23]. Namely that(a) persistent infections occur when there is alower or delayed influx of PMN in the presenceof pro-inflammatory cytokines [20, 27], or (b)persistent infections result from a short half-lifeand decreased functionality of PMN such asimpaired phagocytosis and/or killing capacity[9, 23]. However, these alternative explanationsare unlikely to be a general mechanism respon-sible for creating the observed patterns. In thecase of low or delayed PMN influx, the modelpredicts unrealistically high numbers of E. coliduring the transient infection dynamics. In thecase of a short life expectancy of PMN, ourmodel would require an unrealistically shorthalf-life to be able to fit data for cows with a per-sistent intramammary infection. Also, there isgood in vitro experimental evidence to suggestthat E. coli strains present in persistent infectionshave different attributes from those present intransient infections [11], thereby suggesting thatthe difference in pathogenesis between transientand persistent infections is due to the type ofbacteria rather than the type of responsemounted by individual cows.
Although macrophages, PMN and lympho-cytes will probably undergo some homingtowards the supramammary lymphnodes [24],we did not include this behaviour in the model.We also ignored the potential impact of mem-ory dependent immunity, since it is generallyassumed that antibody mediated immuneresponse mostly reduces clinical severity andhas relatively little impact on the incidence ofinfection or the distinction between transientand persistent infection [29]. In future genera-tions of our infection model, more componentsof the immune response may be added and eval-uated for their importance. The hypothesis of alonger survival time in an intracellular epithelialreservoir, resulting in persistent rather than tran-sient infections, appeared to be supported by thedata and our current knowledge of the biologyof these infections.
Acknowledgements. The Wellcome Trust-MahidolUniversity-Oxford Tropical Medicine Research Pro-gramme is funded by the Wellcome Trust of Great
Britain. Y.H. Schukken and G.F. Medley acknowledgethe support of the BBSRC (BBS/S/04854).
REFERENCES
[1] Anderson G.G., Dodson K.W., Hooton T.M.,Hultgren S.J., Intracellular bacterial communities ofuropathogenic Escherichia coli in urinary tract pathogen-esis, Trends Microbiol. (2004) 12:424–430.
[2] Bannerman D.D., Paape M.J., Lee J.W., Zhao X.,Hope J.C., Rainard P., Escherichia coli and Staphylococ-cus aureus elicit differential innate immune responsesfollowing intramammary infection, Clin. Diagn. Lab.Immunol. (2004) 11:463–472.
[3] Bannerman D.D., Paape M.J., Hare W.R., Hope J.C.,Characterization of the bovine innate immune response tointramammary infection with Klebsiella pneumoniae,J. Dairy Sci. (2004) 87:2420–2432.
[4] Bannerman D.D., Pathogen-dependent induction ofcytokines and other soluble inflammatory mediatorsduring intramammary infection of dairy cows, J. Anim.Sci. (2008) 87:10–25.
[5] Barkema H.W., Schukken Y.H., Lam T.J.G.M.,Beiboer M.L., Wilmink H., Benedictus G., Brand A.,Incidence of clinical mastitis in dairy herds grouped inthree categories by bulk milk somatic cell counts, J. DairySci. (1998) 81:411–419.
[6] Bergsten G., Wullt B., Svanborg C., Escherichia coli,fimbriae, bacterial persistence and host response inductionin the human urinary tract, Int. J. Med. Microbiol. (2005)295:487–502.
[7] Berry R.E., Klumpp D.J., Schaeffer A.J., Urothelialcultures support intracellular bacterial community forma-tion by uropathogenic Escherichia coli, Infect. Immun.(2009) 77:2762–2772.
[8] Burvenich C., Van Merris V., Mehrzad J., Diez-FraileA., Duchateau L., Severity of E. coli mastitis is mainlydetermined by cow factors, Vet. Res. (2003) 34:521–564.
[9] Burvenich C., Monfardini E., Mehrzad J., CapucoA.V., Paape M.J., Role of neutrophil polymorphonuclearleukocytes during bovine coliform mastitis: physiology orpathology?, Verh. K. Acad. Geneeskd. Belg. (2004)66:97–150.
[10] Detilleux J., Vangroenweghe F., Burvenich C.,Mathematical model of the acute inflammatory responseto Escherichia coli in intramammary challenge, J. DairySci. (2006) 89:3455–3465.
[11] Dogan B., Klaessig S., Rishniw M., Almeida R.,Oliver S.P., Simpson K.W., Schukken Y.H., Adherent andInvasive Escherichia coli are associated with persistentbovine mastitis, Vet. Microbiol. (2006) 116:270–282.
[12] Dopfer D., Barkema H.W., Lam T.J.G.M., SchukkenY.H., Gaastra W., Recurrent clinical mastitis caused byEscherichia coli in dairy cows, J. Dairy Sci. (1999)82:80–85.
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 13 of 15

[13] Dosogne H., Vangroenweghe F., Mehrzad J.,Massart-Leen A.M., Burvenich C., Differential leukocytecount method for bovine low somatic cell count milk,J. Dairy Sci. (2003) 86:828–834.
[14] Franco A.V., Recurrent urinary tract infections, BestPract. Res. Clin. Obstet. Gynaecol. (2005) 19:861–873.
[15] Green M.J., Green L.E., Medley G.F., SchukkenY.H., Bradley A.J., Influence of dry period bacterialintramammary infection on clinical mastitis in dairy cows,J. Dairy Sci. (2002) 85:2589–2599.
[16] Iwahi T., Imada A., Interaction of Escherichia coliwith polymorphonuclear leukocytes in pathogenesis ofurinary tract infection in mice, Infect. Immun. (1988)56:947–953.
[17] Kirschner D., Marino S., Mycobacterium tubercu-losis as viewed through a computer, Trends Microbiol.(2005) 13:206–211.
[18] Kornalijnslijper J.E., van Werven T., Daemen A.J.,van den Broek J., Niewold T.A., Rutten V.P., Noordhu-izen-Stassen E.N., In vitro growth of mastitis-inducingEscherichia coli in milk and milk fractions of dairy cows,Vet. Microbiol. (2003) 91:125–134.
[19] Kornalijnslijper J.E., Daemen A.J., van Werven T.,Niewold T.A., Rutten V.P., Noordhuizen-Stassen E.N.,Bacterial growth during the early phase of infectiondetermines the severity of experimental Escherichia colimastitis in dairy cows,Vet.Microbiol. (2004) 101:177–186.
[20] Lohuis J.A., Schukken Y.H., Henricks P.A.,Heyneman R., Burvenich C., Verheijden J.H., et al.,Preinfection functions of blood polymorphonuclearleukocytes and the outcome of experimental Escherichiacoli mastitis in the cow, J. Dairy Sci. (1990) 73:342–350.
[21] Mulvey M.A., Schilling J.D., Hultgren S.J., Estab-lishment of a persistent Escherichia coli reservoir duringthe acute phase of a bladder infection, Infect. Immun.(2001) 69:4572–4579.
[22] Olde Riekerink R.G., Barkema H.W., Kelton D.F.,Scholl D.T., Incidence rate of clinical mastitis onCanadian dairy farms, J. Dairy Sci. (2008) 91:1366–1377.
[23] Paape M., Mehrzad J., Zhao X., Detilleux J.,Burvenich C., Defense of the bovine mammary gland bypolymorphonuclear neutrophil leukocytes, J. MammaryGland. Biol. Neoplasia (2002) 7:109–121.
[24] Saad A.M., Ostensson K., Flow cytofluorometricstudies on the alteration of leukocyte populations in bloodand milk during endotoxin-induced mastitis in cows, Am.J. Vet. Res. (1990) 51:1603–1607.
[25] Schepers A.J., Lam T.J., Schukken Y.H.,Wilmink J.B., Hanekamp W.J., Estimation of variancecomponents for somatic cell counts to determine thresh-olds for uninfected quarters, J. Dairy Sci. (1997) 80:1833–1840.
[26] Schukken Y.H., Grommers F.J., Vandegeer D.,Brand A., Incidence of clinical mastitis on farms withlow somatic-cell counts in bulk milk, Vet. Rec. (1989)125:60–63.
[27] VanWerven T., NoordhuizenStassen E.N.,Daemen A.J., Schukken Y.H., Brand A., Burvenich C.,
Preinfection in vitro chemotaxis, phagocytosis, oxidativeburst, and expression of CD11/CD18 receptors and theirpredictive capacity on the outcome of mastitis induced indairy cows with Escherichia coli, J. Dairy Sci. (1997)80:67–74.
[28] Wearing H.J., Rohani P., Keeling M.J., Appropriatemodels for the management of infectious diseases, PLoSMed. (2005) 2:e174.
[29] Wilson D.J., Grohn Y.T., Bennett G.J., GonzalezR.N., Schukken Y.H., Spatz J., Comparison of J5vaccinates and controls for incidence, etiologic agent,clinical severity, and survival in the herd followingnaturally occurring cases of clinical mastitis, J. DairySci. (2007) 90:4282–4288.
APPENDIX
Derivation of equation for E. coli
The variables are expressed as concentrationsapart from bacterial load and PMN which isexpressed in absolute numbers. We assume thatmilking does not alter the concentrations of anyof these quantities and thus there is no loss termin the concentration formulae. However, milk-ing will affect numbers of cells in the milk thusthe following analysis was performed in order topredict total bacterial load. Let concentration ofE. coli be X. Then the differential equation forconcentration of E. coli is given by:
dXdt¼ geX � kX M þ Pð Þ � aX þ b
1
V
Then the total number, E, is given by:
E ¼ VX
Since the volume, V, also varies in time, thedifferential of E is derived using the productrule:
dEdt¼ V
dXdtþ X
dVdt
Thus combining the two previous equationsgives the following formula for the rate ofchange of E. coli cfu in the milk:
dEdt¼ V geX � kX M þ Pð Þ � aX Þð
þbI þ XdVdt
Vet. Res. (2010) 41:13 L.J. White et al.
Page 14 of 15 (page number not for citation purpose)

then
dEdt¼ geE � kE M þ Pð Þ � aE
þ bI þ EV
dVdt
where dVdtis the rate of change of the volume of
milk.The differential equation for the numbers of
PMN in the milk is derived similarly.
Killing rate, k:
Given 0.25 mL of p (6 to 8) · 106 cells/mLPMN added to 0.05 mL of E. coli. solution.Then the number of PMN cells would be0.25 p · 106 and the volume would be0.25 + 0.05 = 0.3 mL. Thus the concentration,P, of the resulting solution is 0:25�P : 106
0:3¼ P�107
12.
Then, given a killing rate of k cell day�1, theconcentration of E. coli over time, y, is givenby the following:
dydt¼ �kPy
y ¼ y0 expð�kPtÞ
ln y ¼ ln y0 � kPt
k ¼ � ln y � ln y0Pt
Since a reduction of one log per hour is
observed: t = 1/24, P¼ p�10712
, In y0 � ln y = 1then k¼ 12�24
p�107, so for p = 6, k = 4.8 · 10�6
and for p = 8, k = 3.6 · 10�6.
Growth rate of bacteria in milk, ge:
Let x(t) = x0 exp(get) where x is the concen-tration of E. coli over time, x0 is the initial con-centration and t is time, then
In x ¼ In x0 þ get
Iog10 xIog10 e
¼ Iog10 x0Iog10 e
þ get
ge ¼Iog10 x� Iog10 x0
t Iog10 e
Then, the upper limit of ge is (6�1)/(0.25 log10 e) or equal to 46.1, and the lowerlimit of ge is (3.8�1)/(0.25 log10 e) or equal to25.8.
Mathematical model of intramammary E. coli Vet. Res. (2010) 41:13
(page number not for citation purpose) Page 15 of 15




















