
M.E. (CIVIL ENGG) Specialization: Transportation Engineering
Upload
independentCategory
view
5download
0
IG
b
tlSp
Neuroscience 194 (2011) 112–123
MITOCHONDRIAL PROTEINS, LEARNING AND MEMORY:
BIOCHEMICAL SPECIALIZATION OF A MEMORY SYSTEMR. O. SOLOMONIA,a N. KUNELAURI,a E. MIKAUTADZE,a
D. APKHAZAVA,a B. J. MCCABEb AND G. HORNb*aInstitute of Chemical Biology, Ilia State University and I. Beritashvilinstitute of Physiology, 14 L. Gotua Street, Tbilisi 0160, Republic ofeorgia
bSub-Department of Animal Behaviour, Department of Zoology, Cam-ridge University, Madingley, Cambridge CB23 8AA, UK
Abstract—The enzyme cytochrome c oxidase is a mitochon-drial protein complex that plays a crucial role in oxidativemetabolism. In the present study we show that amounts oftwo of its protein subunits (cytochrome c oxidase subunit I[CO-I] and II [CO-II]) are influenced by both learning-indepen-dent and learning-dependent factors. Converging evidencehas consistently implicated the left intermediate medialmesopallium (IMM) in the chick brain as a memory store forthe learning process of visual imprinting. This form of learn-ing proceeds very shortly after chicks have been hatched. Inthe left IMM, but not in three other brain regions studied,amounts of CO-I and CO-II co-varied: the correlation betweenthem was highly significant. This relationship did not dependon learning. However, learning influenced the amounts ofboth proteins, but did so only in the left IMM. In this region,amounts of each protein increased with the strength of learn-ing. These findings raise the possibility that the molecularmechanisms involved in the coordinated assembly of cyto-chrome c oxidase are precociously developed in the left IMMcompared to the other regions studied. This precocious de-velopment may enable the region to respond efficiently to theoxidative demands made by the changes in synaptic connec-tivity that underlie memory formation and would allow the leftIMM to function as a storage site within hours after hatching.© 2011 IBRO. Published by Elsevier Ltd. All rights reserved.
Key words: behavioral imprinting, cytochrome c oxidase,intermediate medial mesopallium, IMM/IMHV.
A substantial body of evidence implicates a restricted partof the avian forebrain, the medial mesopallium (MM), as amemory storage site. The intermediate extent of the medialmesopallium, the intermediate medial mesopallium (IMM)
*Corresponding author. Tel: �44-1223-741801; fax: �44-1223-741802.E-mail address: [email protected] (G. Horn).Abbreviations: APP, amyloid precursor protein; BLAST, Basic LocalAlignment Search Tool; cDNA, complementary DNA; CMM, caudalmedial mesopallium; CO-I, cytochrome c oxidase subunit I; CO-II,cytochrome c oxidase subunit II; df, degrees of freedom; F, varianceratio; HSP90, heat shock protein 90; IMHV, intermediate medial hy-perstriatum ventrale; IMM, intermediate medial mesopallium;MARCKS, myristoylated alanine-rich C kinase substrate; MM, medialmesopallium; NMDA, N-methyl-D-aspartic acid; PPN, posterior pole ofhe nidopallium; PSD, postsynaptic density; r, product-moment corre-ation coefficient; SE, standard error; SEM, standard error of the mean;
SH, suppressive subtractive hybridization; t, Student’s t; w/v, weighter unit volume.0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All righdoi:10.1016/j.neuroscience.2011.07.053
112
(formerly known as intermediate medial hyperstriatum ven-trale, IMHV), is such a site for the memory underlyingvisual imprinting (Horn, 1985, 2004; Johnston et al., 1993)and passive avoidance learning (Rose, 2000; Gibbs andSummers, 2002) in domestic chicks. The anterior MM hasbeen implicated in auditory imprinting in domestic chicks(Bredenkötter and Braun, 1997) and the caudal part, cau-dal medial mesopallium (CMM), in storing informationabout the tutor’s song in songbirds (Bolhuis and Gahr,2006; Gobes et al., 2010).
Through the learning process of imprinting, young vi-sually naïve domestic chicks come to recognize a visualobject by being exposed to it and, subsequently, to ap-proach that object in preference to others (see e.g. Bol-huis, 1991). Lesions of IMM impair both the acquisition andretention of the visual preference (McCabe et al., 1981,1982). Imprinting leads to an increase in the number ofIMM neurons responding selectively to the visual trainingstimulus (Brown and Horn, 1994; Nicol et al., 1995; Horn etal., 2001; Jackson et al., 2008) and to an increase in lengthof the postsynaptic density of dendritic spine synapses(Bradley et al., 1981; Horn et al., 1985). Biochemicalchanges also occur in the IMM, following training (seeHorn, 1985, 2004). These changes, which are closely re-lated to the strength of learning, include increases in theexpression of a number of proteins, all of which are knownto play an important role in neuronal function (Solomonia etal., 1997, 1998, 2003, 2005); the changes in proteinamounts occur consistently in the left IMM.
As the number of such proteins increases, it becomesimpracticable to inquire whether each of them plays a rolein information storage. Therefore we undertook a broaderanalysis of genes whose expression in the IMM may bechanged by learning, using subtractive cDNA cloningmethodology (Solomonia et al., 2003). In that study twopooled IMM samples were compared: one from a stronglyand one from a weakly imprinted group of chicks (seeMcCabe and Horn, 1994). The samples were removed24 h after the end of training. Suppressive subtractivehybridization (SSH) followed by differential dot-blot hybrid-ization disclosed the existence of �50 partial cDNA clonescorresponding to genes up- or downregulated during theimprinting procedure. These clones were sequenced andDNA sequence data were searched for homology throughGenBank analysis using Basic Local Alignment SearchTool (BLAST) software.
Because the SSH analysis was conducted on only twoIMM samples, some of the changes in gene expressionobserved might reflect random variation in differences be-
tween samples. Furthermore, a change in the expressionts reserved.
aBdmtFusb
aopCi
o1sc�p
otoHmioid
w(ncmipm
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123 113
of a gene does not necessarily result in a change in theamount of protein that it encodes. We subsequently in-quired whether the amounts of the protein products of twocandidate genes—encoding amyloid precursor protein(APP) and myristoylated, alanine-rich C-kinase substrate(MARCKS)—were changed by imprinting (Solomonia etal., 2003). We found that the levels of each of theseproteins in the IMM increased significantly with thestrength of learning. There were no significant effects oflearning in two control brain regions (the left and rightposterior poles of the nidopallium [PPN]).
In the present study we have further explored theresults obtained by the SSH experiments. Brain sampleswere removed 24 h after training and the protein productsof the following candidate genes were assayed: subunits Iand II of cytochrome c oxidase (CO-I and CO-II, respec-tively), �-fodrin and heat-shock protein 90 (HSP90). In theSSH experiments the mRNA levels of CO-I, CO-II andHSP90 were upregulated in the left IMM of chicks thatshowed a strong preference for the imprinted object,whereas the mRNA level for �-fodrin was downregulated.These gene products were selected for the following rea-sons: (i) CO-I and CO-II are catalytic subunits of the cyto-chrome c oxidase enzyme complex and are expressed inmitochondria. This enzyme is central to energy metabolism(Gennis and Ferguson-Miller, 1995), and mitochondriaplay a role in many aspects of neuronal function including,it has been suggested, the synaptic changes that underlielearning and memory (Horn, 1952; Li et al., 2004; Mattsonet al., 2008). (ii) �-fodrin, or brain spectrin, is a membrane-cytoskeletal linker protein (Malchiodi-Albedi et al., 1993)nd has been implicated in synaptic plasticity (Lynch andaudry, 1984; Lynch et al., 2007). (iii) HSP90 is widelyistributed in neurons (D’Souza and Brown, 1998) anday be involved in synaptic transmission through its role in
he recycling of synaptic vesicles (Sakisaka et al., 2002).or none of these proteins have correlative studies beenndertaken to relate changes in their amounts to thetrength of learning in a region of vertebrate brain known toe a memory store.
EXPERIMENTAL PROCEDURES
Ethical considerations
Chicks experienced no detectable suffering and all experimentswere licensed under the United Kingdom Animals (Scientific Pro-cedures) Act 1986.
Behavior and tissue collection
Chicks (Gallus gallus domesticus) were hatched, reared andtrained as described previously (Solomonia et al., 2003). Eightbatches of chicks (Ross 308) were hatched and reared in dark-ness. When 22- to 28-h-old, chicks were placed individually inrunning wheels and trained by exposure for 1 h to a visual im-printing (training) stimulus (a rotating, internally illuminated redbox; for details see Bolhuis et al. (2000)). As a chick attempted topproach the training stimulus, it rotated the running wheel; rev-lutions of the wheel (circumference, 94 cm) were counted torovide a measure of approach activity (“training approach”).hicks that have become imprinted have learned the character-
stics of the training stimulus and subsequently prefer it to an
alternative visual stimulus. Ten minutes after training, each chickwas given a preference test. In this test the chick was shown,sequentially, either the visual training stimulus or an alternativestimulus—a rotating, internally illuminated blue cylinder (Bolhuiset al., 2000). A preference score (McCabe et al., 1981) was thencalculated to provide a measure of each chick’s preference andhence of the strength of learning. The preference score was thechick’s approach to the training stimulus expressed as a percent-age of approach to the training stimulus plus approach to thealternative stimulus. If both stimuli were approached equally, thepreference score would be 50 (no choice, indicative of no learn-ing). If the chick approached only the training stimulus, the pref-erence score would be 100, indicating strong learning. There areindividual differences in chicks’ preference scores after a fixedperiod of training. This variation was used to relate changes inCO-I, CO-II, �-fodrin and HSP90 protein amounts to the strengthf learning (cf. McCabe and Horn, 1988; Horn and Johnson,989). To ensure that the samples from trained chicks repre-ented the range of preference scores between 40 and 100, onehick in each batch was selected with a preference score �40 and60; one with a preference score �60 and �80; and one withreferences score �80.
Chicks were killed 24 h after the end of training. Four piecesf tissue were removed from the brain of each selected chick: fromhe left and right IMM and from the left and right PPN. For detailsf the methods of removal of tissue from the IMM and PPN seeorn (1991) and Solomonia et al. (1998), respectively. After re-oval, each piece of tissue (a sample) was immediately covered
n dry ice. Thus in each batch there were four samples from eachf four chicks (one untrained, three trained), yielding 16 samples
n all. Samples were coded after collection and all further proce-ures were conducted blind.
Electrophoresis and electroblotting
The tissue samples were rapidly homogenized in 20 mM Tris–HCl(pH 7.4), 0.32 M sucrose, 1 mM ethylenediaminetetraacetic acid,0.5 mM ethylene glycol-bis (2-aminoethylether)-N,N,N’,-N=-tet-raacetic acid and a cocktail of protease inhibitors (Sigma-Aldrich,St. Louis, MO, USA), and centrifuged at 1000 g for 10 min. Thesupernatant will be referred to as nucleus-free homogenate. Aconcentrated solution of sodium dodecyl sulfate (SDS) was addedto this supernatant to give a final concentration of 5% w/v. Proteinconcentration was determined in quadruplicate using a microbicinchoninic acid protein assay kit (Thermo Fisher Scientific,Cramlington, UK). Aliquots containing 30 �g of protein in 30 �l
ere subjected to SDS gel electrophoresis and Western blottingSolomonia et al., 2003). After protein had been transferred ontoitrocellulose membranes, the membranes were stained with Pon-eau S solution to confirm transfer and uniform gel loading. Theembranes were then washed with phosphate-buffered sal-
ne � 0.05% Tween 20 and cut into three parts according to theositions of molecular weight standards. The upper part of theembrane was stained with a monoclonal antibody against �-fo-
drin (Biomol International, Hamburg, Germany), the middle partwith a monoclonal antibody against HSP90 (Sigma-Aldrich, St.Louis, MO, USA) and the third part with a monoclonal antibodyagainst subunit I of cytochrome c oxidase (CO-I; Molecular ProbesA6403, Carlsbad, CA, USA). After staining with the anti–CO-Iantibody the third part of the filter was stripped and equal proteinloading was verified by total actin staining of the membrane (an-tibody from Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA).For CO-II staining, separate electrophoresis and Western immu-noblotting was carried out as described above. The CO-II–immu-noreactive band was stained with anti–CO-II antibody (Molecularprobes A6404, Carlsbad, CA, USA), the filter stripped and equalprotein loading verified by total actin staining. Data from experi-
mental stained bands were not normalized with respect to actin
t(
Pevb
tdcm
rarpap
tac
wbiHwbiptldsf
P
Fti
IelTo
to
tt
t1wpsersf
cw
I
wAi2tsu
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123114
staining because such normalization has been found to be anunreliable loading control (Dittmer and Dittmer, 2006).
Standard immunochemical procedures were carried out usingperoxidase-labeled secondary antibodies and a SuperSignal WestPico chemiluminescent substrate (Thermo Fisher Scientific,Cramlington, UK). The blots were then exposed with intensifyingscreens to X-ray films pre-flashed with Sensitize (GE HealthcareLife Sciences, Amersham Place, UK). The optical densities ofprotein bands were measured using LabWorks 4.0 (UVP) soft-ware. The autoradiographs were calibrated using standardamounts of protein obtained from the nucleus-free homogenatefraction of the IMMs of a group of untrained chicks. Four standards(15, 30, 45 and 60 �g total protein) were applied to each gel. Forhese standards the optical densities of the immunostained bandsfor CO-I, CO-II, �-fodrin or HSP90) were plotted against the
amounts of protein; in all these standards, least squares regres-sion showed a significant fit to a straight line. To obtain the datagiven in Figs. 2–6, the optical density of each band from eachexperimental sample was divided by the optical density which,from the calibration of the same autoradiograph, corresponded to30 �g of total protein in the standard (Solomonia et al., 2003).
rotein amounts from the blots were divided by the batch mean toliminate unavoidable variation between batches (e.g. behavioralariation; see Bateson (1974)). The data expressed in this way wille referred to as “standardized relative amount” of CO-I, CO-II,
�-fodrin and HSP90. Samples were obtained from a total of 24rained chicks and eight untrained chicks in eight batches; someata, identified as artefactual under blind conditions, were dis-arded. The number of animals used was estimated to be theinimum required for adequate statistical analysis.
Statistical analysis
Our previous work, including the subtractive hybridization exper-iment that prompted the present experiment (Solomonia et al.,2003), had implicated the IMM in learning and memory. The PPNhad not been implicated in learning and memory. For this reason,in the present study, data from the IMM and PPN were analyzedseparately. Standardized relative amount of protein was subjectedto analysis of variance (ANOVA) with the factor Training Condition(untrained chicks, trained chicks) and the means of subgroups ofthe data compared using t-tests. The data were also subjected toegression analysis with preference score as the predictor vari-ble. In cases where this regression was significant, a furtheregression analysis was conducted with training approach as theredictor variable in order to determine whether protein level wasssociated with preference score alone, or also with training ap-roach. Relationships between levels of CO-I, CO-II, �-fodrin and
HSP90 were investigated by correlation analysis and partial cor-relation analysis (Snedecor and Cochran, 1989), correcting theserelationships for any association between protein amount andpreference score. When predicting the amount of protein corre-sponding to the “no-preference” score of 50 (see e.g. McCabe andHorn, 1994), least squares regression was used; when describingthe association between the amounts of two proteins, the standardmajor axis was fitted in order to correct for regression towards themean (cf. Warton et al., 2006). Correlation coefficients were com-pared using Fisher’s z transformation and �2 as the test statistic(Snedecor and Cochran, 1989). All tests were two tailed unlessotherwise stated and a probability of 0.05 was taken as statisticallysignificant. Probabilities have been reported for all significant re-sults.
RESULTS
Behavior and immunostaining patterns
The mean preference score of trained chicks was
75.73�3.54 SEM, a value significantly greater than the eno-preference value of 50 (F1,21�52.83, P�0.001). Meanraining approach was 227.88�33.89 m. Preference scorend approach activity during training were not significantlyorrelated with each other (r20��0.37, P�0.090).
Anti–CO-I antibodies bound to a band of moleculareight 57 kDa, anti–CO-II antibodies stained a proteinand with molecular weight 30 kDa, anti–�-fodrin antibod-
es stained a band of molecular weight 240 kDa and anti-SP90 antibodies reacted with a protein of moleculareight 86 kDa (Fig. 1A–D). Antibodies against total actinound to a band of 43 kDa (Fig. 1E). This staining pattern
s in accordance with well-documented features of theroteins that were studied. Staining with antibodies againstotal actin verified that equal amounts of protein wereoaded for each sample. Furthermore, the existence ofifferential effects of training on different proteins within aample (see below) excludes the possibility that such ef-ects could be attributable to artefacts of loading.
rotein amounts, brain regions and behavior
or each brain region and training condition (untrained,rained), the results of t-tests, means and SEMs are givenn Table 1.
We compared the mean level of each protein in the leftMM with that in the right. This comparison was made forach training condition (e.g. mean amount of CO-I in the
eft IMM vs. mean amount in right IMM of untrained chicks).he same comparisons were made between mean levelsf proteins in the left and right PPN. With the exception of
�-fodrin in the PPN, the mean amounts of each protein inhe left and right sides did not differ significantly from eachther (P�0.05). For this reason, with the exception of
�-fodrin, data for left and right sides combined were usedo calculate the mean amount of a given protein for a givenraining condition.
Not all chicks subjected to the training procedure learnhe features of the training object (McCabe and Horn,988, 1994; Solomonia et al., 2003). Correlation analysesere therefore performed to enquire whether the level of arotein was related to the measure of learning (preferencecore). Such analyses have, in the past, disclosed differ-nces between left and right IMM in the strength of thiselationship (Solomonia et al., 1998, 2003). For this reasoneparate correlation analyses were performed on datarom left and right brain regions (IMM and PPN).
For all proteins and for all regions except the PPN, theorrelation between training approach and protein amountas not significant (see Table 2 and below).
MM
CO-I. The mean amount of CO-I in trained chicksas 38% higher than in untrained chicks (Fig. 2A).mongst trained chicks, amount of the protein in the IMM
ncreased significantly as preference score increased (Fig.B). Furthermore, the interpolated amount of CO-I inrained chicks that corresponded to the no-preferencecore of 50 did not differ significantly from the mean inntrained chicks (Fig. 2B). This preference score would be
xpected in chicks that had been subjected to the training
tafs
his papel.
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123 115
procedure but had failed to learn the features of the visualimprinting stimulus. Taken together, these results implythat the amount of CO-I increases as the strength of learn-ing increases.
To determine whether this implication also applies toCO-I amounts in left and right IMM, respectively, separatecorrelograms were plotted for each region. For the left IMM,the correlation between CO-I level and preference scoreremained significant, and the mean amount interpolated to apreference score of 50 was not significantly different from themean in untrained chicks (Fig. 2C). For the right IMM thecorrelation was not significant (see legend, Fig. 2). Thesefindings suggest that the left IMM is the major contributor tothe correlation found in the combined data.
CO-II. The mean level of CO-II in trained chicks was34% higher than that in untrained chicks (Fig. 3A, Table 1). Intrained chicks the amount of CO-II (left and right IMM com-bined) was not significantly correlated with preference score(Table 2). However, the slopes of the regression lines ofCO-II amount in left and right IMM, respectively, on prefer-ence score differed significantly from each other (see Fig. 3B,C). For levels of the subunit in the right IMM the regressionslope was negative but not significant on either a two-tailed or
C α-Fodrin
B CO-II
A CO-I
C α-Fodrin
E Actin
D HSP90
Fig. 1. (A–E) Western blot autoradiographs for CO-I, CO-II, �-fodrin,he four chicks from the same batch. Lanes 1–4 are from chicks with pnd �80; lanes 9–12 are from chicks with preference scores �40 an
our consecutive lanes, the samples were arranged in the order: left IMtandard sample of tissue from the IMM of untrained chicks containing r
after training were conducted “blind.” Note that the effects reported in tand are not necessarily apparent from visual inspection of a single ge
one-tailed probability test (Fig. 3C). For the left IMM, the
regression was significant only on a one-tailed test (Fig. 3B);however, there were separate grounds for predicting thedirection of the association, namely that high amounts ofCO-II are associated with high levels of CO-I in this brainregion (see below). For both left and right IMM, respectively,the interpolated levels of CO-II corresponding to a preferencescore of 50 did not differ significantly from the mean amountsin untrained chicks (Fig. 3B, C).
Relationship between CO-I and CO-II. There was asignificant correlation between the levels of CO-I andCO-II in the left IMM of trained chicks (r16�0.67,P�0.0024; Fig. 4, closed circles and continuous line).That is, across chicks, high levels of CO-I were associ-ated with high levels of CO-II, while low amounts of CO-Iwere associated with low amounts of CO-II. Such acorrelation would be expected because the levels ofCO-I and CO-II both increase with the strength of learn-ing (see Figs. 2C and 3B). However, the correlationbetween the two proteins remained significant even aftercorrecting for preference score (partial correlation coef-ficient, rxy,z;15�0.58, P�0.015; see Fig. 4, open circles,broken line). The slope of the line corrected for prefer-ence score (“corrected data”) is very similar to the slope
nd actin. Each lane corresponds to one sample. Lanes 1–16 are frome scores �80; lanes 5–8 are from chicks with preference scores �60d lanes 13–16 are from untrained chicks. In each of these blocks of
IMM, left PPN, right PPN. Lanes 17–20 (17–19 in C) are from a singlely 15, 30, 45 and 60 �g protein (15, 30 and 45 �g in C). All procedures
r and illustrated in Figs. 2–6 were obtained from the complete dataset
HSP90 areferencd �60 anM, rightespective
of the line fitted to the uncorrected data (see legend to
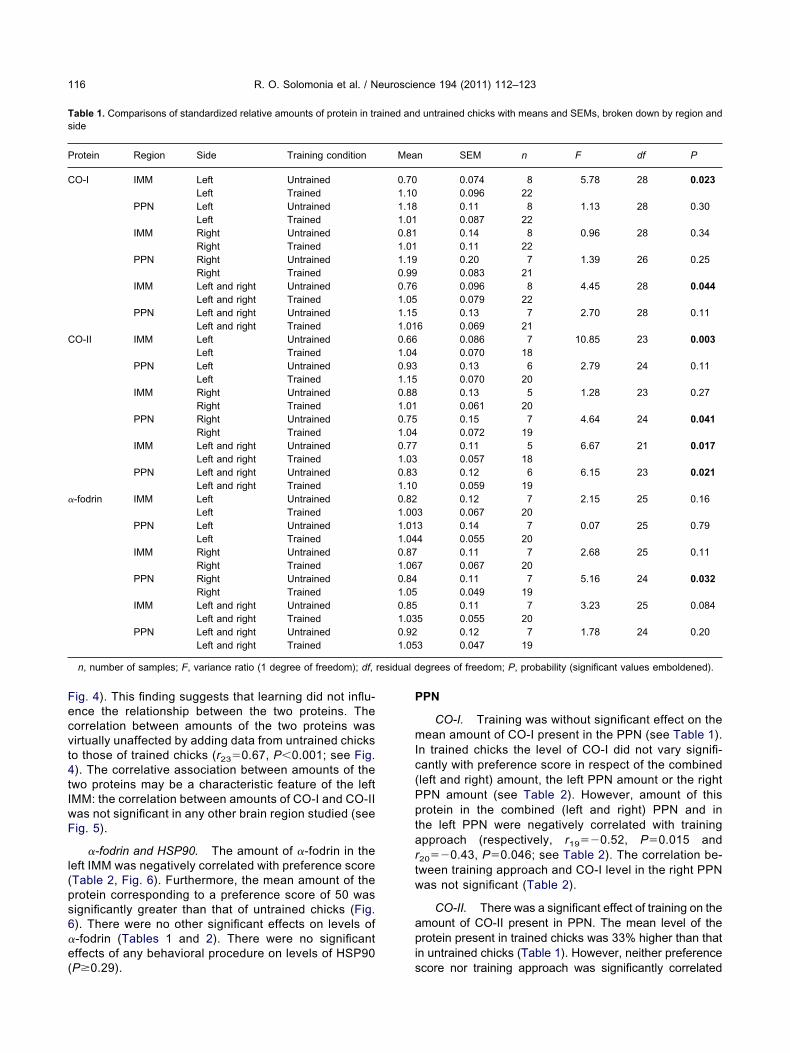
l(ps6
e(
P
mIc(Ppta
tw
api
residual d
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123116
Fig. 4). This finding suggests that learning did not influ-ence the relationship between the two proteins. Thecorrelation between amounts of the two proteins wasvirtually unaffected by adding data from untrained chicksto those of trained chicks (r23�0.67, P�0.001; see Fig.4). The correlative association between amounts of thetwo proteins may be a characteristic feature of the leftIMM: the correlation between amounts of CO-I and CO-IIwas not significant in any other brain region studied (seeFig. 5).
�-fodrin and HSP90. The amount of �-fodrin in theeft IMM was negatively correlated with preference scoreTable 2, Fig. 6). Furthermore, the mean amount of therotein corresponding to a preference score of 50 wasignificantly greater than that of untrained chicks (Fig.). There were no other significant effects on levels of
�-fodrin (Tables 1 and 2). There were no significantffects of any behavioral procedure on levels of HSP90
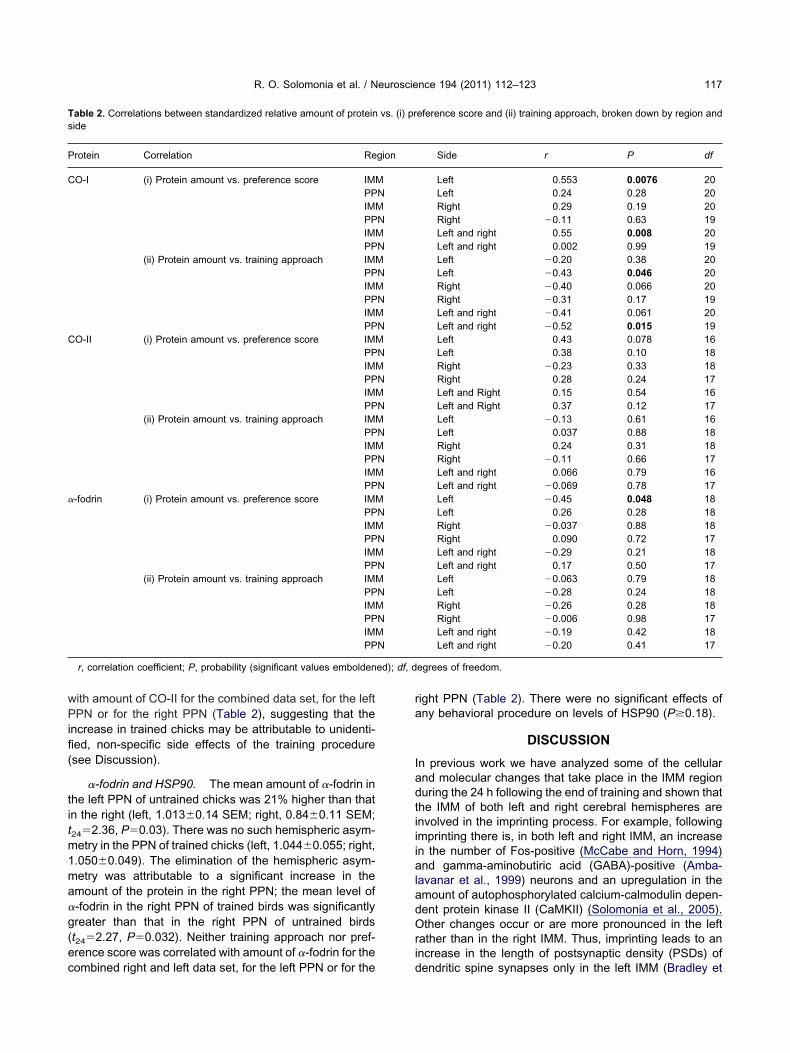
Table 1. Comparisons of standardized relative amounts of protein in trside
Protein Region Side Training condition
CO-I IMM Left UntrainedLeft Trained
PPN Left UntrainedLeft Trained
IMM Right UntrainedRight Trained
PPN Right UntrainedRight Trained
IMM Left and right UntrainedLeft and right Trained
PPN Left and right UntrainedLeft and right Trained
CO-II IMM Left UntrainedLeft Trained
PPN Left UntrainedLeft Trained
IMM Right UntrainedRight Trained
PPN Right UntrainedRight Trained
IMM Left and right UntrainedLeft and right Trained
PPN Left and right UntrainedLeft and right Trained
�-fodrin IMM Left UntrainedLeft Trained
PPN Left UntrainedLeft Trained
IMM Right UntrainedRight Trained
PPN Right UntrainedRight Trained
IMM Left and right UntrainedLeft and right Trained
PPN Left and right UntrainedLeft and right Trained
n, number of samples; F, variance ratio (1 degree of freedom); df,
P�0.29). s
PN
CO-I. Training was without significant effect on theean amount of CO-I present in the PPN (see Table 1).
n trained chicks the level of CO-I did not vary signifi-antly with preference score in respect of the combinedleft and right) amount, the left PPN amount or the rightPN amount (see Table 2). However, amount of thisrotein in the combined (left and right) PPN and in
he left PPN were negatively correlated with trainingpproach (respectively, r19��0.52, P�0.015 and
r20��0.43, P�0.046; see Table 2). The correlation be-ween training approach and CO-I level in the right PPNas not significant (Table 2).
CO-II. There was a significant effect of training on themount of CO-II present in PPN. The mean level of therotein present in trained chicks was 33% higher than that
n untrained chicks (Table 1). However, neither preference
untrained chicks with means and SEMs, broken down by region and
n SEM n F df P
0.074 8 5.78 28 0.0230.096 220.11 8 1.13 28 0.300.087 220.14 8 0.96 28 0.340.11 220.20 7 1.39 26 0.250.083 210.096 8 4.45 28 0.0440.079 220.13 7 2.70 28 0.11
6 0.069 210.086 7 10.85 23 0.0030.070 180.13 6 2.79 24 0.110.070 200.13 5 1.28 23 0.270.061 200.15 7 4.64 24 0.0410.072 190.11 5 6.67 21 0.0170.057 180.12 6 6.15 23 0.0210.059 190.12 7 2.15 25 0.16
3 0.067 203 0.14 7 0.07 25 0.794 0.055 20
0.11 7 2.68 25 0.117 0.067 20
0.11 7 5.16 24 0.0320.049 190.11 7 3.23 25 0.084
5 0.055 200.12 7 1.78 24 0.20
3 0.047 19
egrees of freedom; P, probability (significant values emboldened).
ained and
Mea
0.701.101.181.010.811.011.190.990.761.051.151.010.661.040.931.150.881.010.751.040.771.030.831.100.821.001.011.040.871.060.841.050.851.030.921.05
core nor training approach was significantly correlated
titm1ma
g(
c
ra
ladOri
s
ed); df, d
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123 117
with amount of CO-II for the combined data set, for the leftPPN or for the right PPN (Table 2), suggesting that theincrease in trained chicks may be attributable to unidenti-fied, non-specific side effects of the training procedure(see Discussion).
�-fodrin and HSP90. The mean amount of �-fodrin inhe left PPN of untrained chicks was 21% higher than thatn the right (left, 1.013�0.14 SEM; right, 0.84�0.11 SEM;
24�2.36, P�0.03). There was no such hemispheric asym-etry in the PPN of trained chicks (left, 1.044�0.055; right,.050�0.049). The elimination of the hemispheric asym-etry was attributable to a significant increase in themount of the protein in the right PPN; the mean level of
�-fodrin in the right PPN of trained birds was significantlyreater than that in the right PPN of untrained birdst24�2.27, P�0.032). Neither training approach nor pref-
erence score was correlated with amount of �-fodrin for the
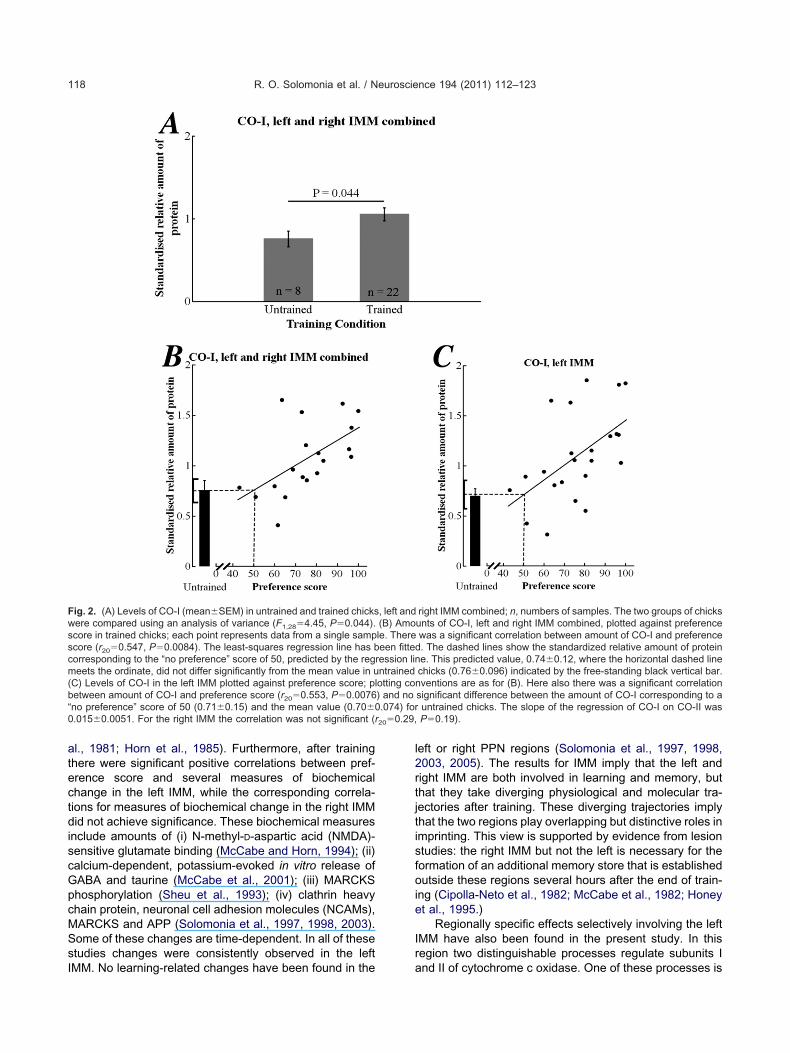
Table 2. Correlations between standardized relative amount of proteinide
Protein Correlation Re
CO-I (i) Protein amount vs. preference score IMPPIMPPIMPP
(ii) Protein amount vs. training approach IMPPIMPPIMPP
CO-II (i) Protein amount vs. preference score IMPPIMPPIMPP
(ii) Protein amount vs. training approach IMPPIMPPIMPP
�-fodrin (i) Protein amount vs. preference score IMPPIMPPIMPP
(ii) Protein amount vs. training approach IMPPIMPPIMPP
r, correlation coefficient; P, probability (significant values embolden
ombined right and left data set, for the left PPN or for the d
ight PPN (Table 2). There were no significant effects ofny behavioral procedure on levels of HSP90 (P�0.18).
DISCUSSION
In previous work we have analyzed some of the cellularand molecular changes that take place in the IMM regionduring the 24 h following the end of training and shown thatthe IMM of both left and right cerebral hemispheres areinvolved in the imprinting process. For example, followingimprinting there is, in both left and right IMM, an increasein the number of Fos-positive (McCabe and Horn, 1994)and gamma-aminobutiric acid (GABA)-positive (Amba-avanar et al., 1999) neurons and an upregulation in themount of autophosphorylated calcium-calmodulin depen-ent protein kinase II (CaMKII) (Solomonia et al., 2005).ther changes occur or are more pronounced in the left
ather than in the right IMM. Thus, imprinting leads to anncrease in the length of postsynaptic density (PSDs) of
eference score and (ii) training approach, broken down by region and
Side r P df
Left 0.553 0.0076 20Left 0.24 0.28 20Right 0.29 0.19 20Right �0.11 0.63 19Left and right 0.55 0.008 20Left and right 0.002 0.99 19Left �0.20 0.38 20Left �0.43 0.046 20Right �0.40 0.066 20Right �0.31 0.17 19Left and right �0.41 0.061 20Left and right �0.52 0.015 19Left 0.43 0.078 16Left 0.38 0.10 18Right �0.23 0.33 18Right 0.28 0.24 17Left and Right 0.15 0.54 16Left and Right 0.37 0.12 17Left �0.13 0.61 16Left 0.037 0.88 18Right 0.24 0.31 18Right �0.11 0.66 17Left and right 0.066 0.79 16Left and right �0.069 0.78 17Left �0.45 0.048 18Left 0.26 0.28 18Right �0.037 0.88 18Right 0.090 0.72 17Left and right �0.29 0.21 18Left and right 0.17 0.50 17Left �0.063 0.79 18Left �0.28 0.24 18Right �0.26 0.28 18Right �0.006 0.98 17Left and right �0.19 0.42 18Left and right �0.20 0.41 17
egrees of freedom.
vs. (i) pr
gion
MNMNMNMNMNMNMNMNMNMNMNMNMNMNMNMNMNMN
endritic spine synapses only in the left IMM (Bradley et
ss
“ .074) for0 20�0.29,
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123118
al., 1981; Horn et al., 1985). Furthermore, after trainingthere were significant positive correlations between pref-erence score and several measures of biochemicalchange in the left IMM, while the corresponding correla-tions for measures of biochemical change in the right IMMdid not achieve significance. These biochemical measuresinclude amounts of (i) N-methyl-D-aspartic acid (NMDA)-sensitive glutamate binding (McCabe and Horn, 1994); (ii)calcium-dependent, potassium-evoked in vitro release ofGABA and taurine (McCabe et al., 2001); (iii) MARCKSphosphorylation (Sheu et al., 1993); (iv) clathrin heavychain protein, neuronal cell adhesion molecules (NCAMs),MARCKS and APP (Solomonia et al., 1997, 1998, 2003).Some of these changes are time-dependent. In all of thesestudies changes were consistently observed in the left
Fig. 2. (A) Levels of CO-I (mean�SEM) in untrained and trained chickswere compared using an analysis of variance (F1,28�4.45, P�0.044).core in trained chicks; each point represents data from a single sampcore (r20�0.547, P�0.0084). The least-squares regression line has b
corresponding to the “no preference” score of 50, predicted by the regmeets the ordinate, did not differ significantly from the mean value in u(C) Levels of CO-I in the left IMM plotted against preference score; plbetween amount of CO-I and preference score (r20�0.553, P�0.0076no preference” score of 50 (0.71�0.15) and the mean value (0.70�0.015�0.0051. For the right IMM the correlation was not significant (r
IMM. No learning-related changes have been found in the
left or right PPN regions (Solomonia et al., 1997, 1998,2003, 2005). The results for IMM imply that the left andright IMM are both involved in learning and memory, butthat they take diverging physiological and molecular tra-jectories after training. These diverging trajectories implythat the two regions play overlapping but distinctive roles inimprinting. This view is supported by evidence from lesionstudies: the right IMM but not the left is necessary for theformation of an additional memory store that is establishedoutside these regions several hours after the end of train-ing (Cipolla-Neto et al., 1982; McCabe et al., 1982; Honeyet al., 1995.)
Regionally specific effects selectively involving the leftIMM have also been found in the present study. In thisregion two distinguishable processes regulate subunits I
right IMM combined; n, numbers of samples. The two groups of chicksunts of CO-I, left and right IMM combined, plotted against preferencewas a significant correlation between amount of CO-I and preference
d. The dashed lines show the standardized relative amount of proteine. This predicted value, 0.74�0.12, where the horizontal dashed linechicks (0.76�0.096) indicated by the free-standing black vertical bar.nventions are as for (B). Here also there was a significant correlationsignificant difference between the amount of CO-I corresponding to auntrained chicks. The slope of the regression of CO-I on CO-II wasP�0.19).
, left and(B) Amole. Thereeen fitte
ression linntrainedotting co) and no
and II of cytochrome c oxidase. One of these processes is
c
of
d e mean acantly dif
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123 119
learning independent and concerns the relationship be-tween the amounts of the subunits; the other process islearning-dependent and influences the amounts of the twosubunits.
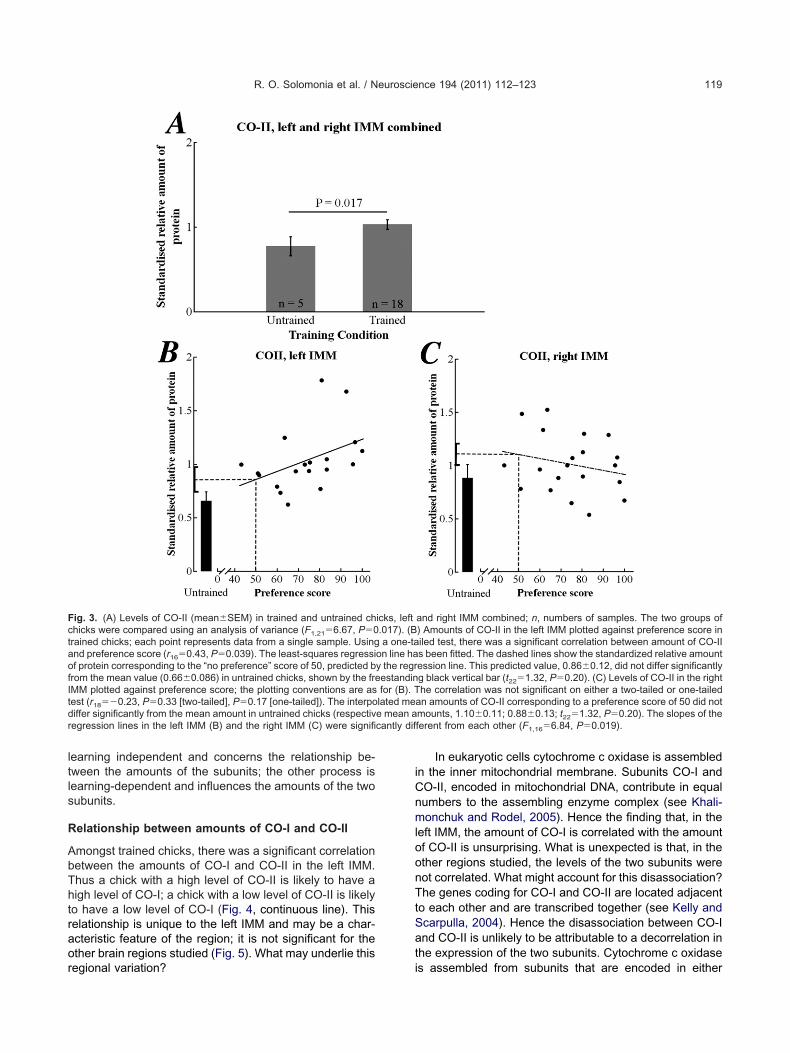
Relationship between amounts of CO-I and CO-II
Amongst trained chicks, there was a significant correlationbetween the amounts of CO-I and CO-II in the left IMM.Thus a chick with a high level of CO-II is likely to have ahigh level of CO-I; a chick with a low level of CO-II is likelyto have a low level of CO-I (Fig. 4, continuous line). Thisrelationship is unique to the left IMM and may be a char-acteristic feature of the region; it is not significant for theother brain regions studied (Fig. 5). What may underlie this
Fig. 3. (A) Levels of CO-II (mean�SEM) in trained and untrained chhicks were compared using an analysis of variance (F1,21�6.67, P�0
trained chicks; each point represents data from a single sample. Usingand preference score (r16�0.43, P�0.039). The least-squares regressif protein corresponding to the “no preference” score of 50, predicted brom the mean value (0.66�0.086) in untrained chicks, shown by the freIMM plotted against preference score; the plotting conventions are astest (r18��0.23, P�0.33 [two-tailed], P�0.17 [one-tailed]). The interpoiffer significantly from the mean amount in untrained chicks (respectiv
regression lines in the left IMM (B) and the right IMM (C) were signifi
regional variation?
In eukaryotic cells cytochrome c oxidase is assembledin the inner mitochondrial membrane. Subunits CO-I andCO-II, encoded in mitochondrial DNA, contribute in equalnumbers to the assembling enzyme complex (see Khali-monchuk and Rodel, 2005). Hence the finding that, in theleft IMM, the amount of CO-I is correlated with the amountof CO-II is unsurprising. What is unexpected is that, in theother regions studied, the levels of the two subunits werenot correlated. What might account for this disassociation?The genes coding for CO-I and CO-II are located adjacentto each other and are transcribed together (see Kelly andScarpulla, 2004). Hence the disassociation between CO-Iand CO-II is unlikely to be attributable to a decorrelation inthe expression of the two subunits. Cytochrome c oxidase
and right IMM combined; n, numbers of samples. The two groups of) Amounts of CO-II in the left IMM plotted against preference score iniled test, there was a significant correlation between amount of CO-IIs been fitted. The dashed lines show the standardized relative amountession line. This predicted value, 0.86�0.12, did not differ significantlyg black vertical bar (t22�1.32, P�0.20). (C) Levels of CO-II in the righthe correlation was not significant on either a two-tailed or one-tailed
an amounts of CO-II corresponding to a preference score of 50 did notmounts, 1.10�0.11; 0.88�0.13; t22�1.32, P�0.20). The slopes of theferent from each other (F1,16�6.84, P�0.019).
icks, left.017). (Ba one-ta
on line hay the regrestandinfor (B). Tlated me
is assembled from subunits that are encoded in either
0P(b
as(firoswuA
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123120
mitochondrial or nuclear DNA. Many factors are involved inthe assembly of the subunits and in the insertion of cofac-tors and metal ions into the maturing enzyme complex (seeFontanesi et al., 2006; Mick et al., 2011). In addition a lessunderstood group of proteins is involved in stabilizing im-mature cytochrome c oxidase pre-complexes (Khalimon-chuk and Rodel, 2005). It is possible that the left IMMcontains all the factors necessary for assembling the en-zyme but that they are less abundant in the other regionsstudied, so that in these regions incorporation of the sub-
Fig. 4. Left IMM. Amount of CO-I plotted against amount of CO-II,trained chicks. Closed symbols, uncorrected for preference score;open symbols, corrected for preference score by partial correlation.Standard major axes have been fitted respectively to the uncorrecteddata (solid line) and corrected data (broken line). Uncorrected data:correlation coefficient, r16�0.67, P�0.0024; slope of fitted line, 1.57 �.29 SE. Corrected data: partial correlation coefficient, rxy,z;15�0.58,�0.015; slope of fitted line, 1.44�0.30 SE. The correlation coefficient
r) and slope of the fitted line for trained and untrained chicks com-ined (r23�0.67, P�0.001; slope�1.37�0.21) were not significantly
different from the corresponding values (whether corrected or uncor-rected) for the trained chicks alone. There was significantly more CO-Iand CO-II in trained than in untrained chicks (CO-I, F1,28�5.78,P�0.023; CO-II, F1,23�10.85, P�0.0032).
Fig. 5. Pearson product moment correlation coefficients betweenamount of CO-I and amount of CO-II in the brain regions studied. Onlyin the left IMM was the correlation significant (in all other regionsP�0.6). The correlation coefficients in the four regions are significantlydifferent from each other (�2�9.12, 3 df, P�0.028). The right IMM, leftPPN and right PPN do not differ significantly from one another�2
( �0.095, 2 df, P�0.95), and the left IMM differs significantly from theother three regions combined (�2�9.21, 1 df, P�0.0024).
units into the assembling complex becomes less coordi-nated. As a result, one or other or both of the unassembledsubunits degrade (see Fontanesi et al., 2006). A differencein the rates of degradation of the two subunits wouldpredict the observed disassociation between amounts ofCO-I and CO-II.
The positive correlation between the amounts of CO-Iand CO-II in the left IMM and the absence of a significantcorrelation in the other brain regions raise the possibilitythat the left IMM contains a greater proportion of combinedto uncombined subunits and more of the assembled en-zyme complex than the other regions. This possibility is ofinterest in light of the specific role the region plays inlearning and memory in the young chick. The domesticchick belongs to a group of animals (precocial species)that exhibit relatively mature forms of behavior soon afterhatching or birth. The group includes some birds and somemammals (especially ungulates and guinea pigs; seeSluckin (1972)). The domestic chick is hatched with itseyes open and within a few hours is highly mobile and ableto learn the features of its mother and of other visuallyconspicuous objects. The neural processes that are en-gaged in the formation of the memory trace and that areparticularly clear in the left IMM may place a heavy de-mand on oxidative metabolism (Gibbs et al., 2006) andhence on cytochrome c oxidase (see below). It is possiblethat the molecular mechanisms involved in the coordinatedassembly of this enzyme complex are precociously devel-oped in the left IMM compared with the other brain regionsenabling it to function efficiently as a memory store within
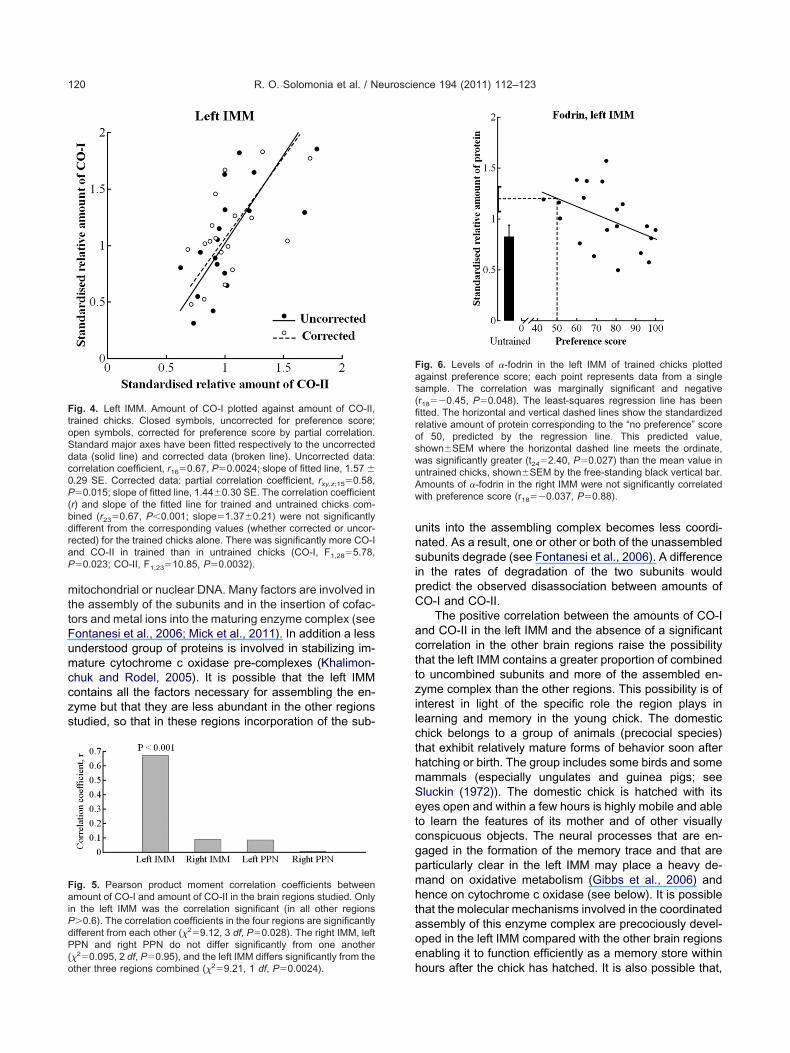
Fig. 6. Levels of �-fodrin in the left IMM of trained chicks plottedgainst preference score; each point represents data from a singleample. The correlation was marginally significant and negativer18��0.45, P�0.048). The least-squares regression line has beentted. The horizontal and vertical dashed lines show the standardizedelative amount of protein corresponding to the “no preference” scoref 50, predicted by the regression line. This predicted value,hown�SEM where the horizontal dashed line meets the ordinate,as significantly greater (t24�2.40, P�0.027) than the mean value inntrained chicks, shown�SEM by the free-standing black vertical bar.mounts of �-fodrin in the right IMM were not significantly correlated
with preference score (r18��0.037, P�0.88).
hours after the chick has hatched. It is also possible that,

sC
msi
IstiTtb
cttf
rwTitcf
Is2sattcmn
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123 121
for forms of early learning that do not involve IMM (seeJohnson and Horn, 1986), the brain regions storing theacquired information have a similar profile to that of the leftIMM in respect of amounts of CO-I and CO-II.
The impact of learning on amounts of CO-I and CO-II
We infer that the change in amount of CO-I in the left IMMafter training (Fig. 2C) was a function of learning for thefollowing reasons. (i) There was a significant positive cor-relation between preference score and the amount of CO-I:the more chicks had learned, the higher the level of CO-I.(ii) The mean level of CO-I in untrained controls was al-most identical to the amount corresponding to a “no-choice” preference score of 50 (Fig. 2C); subjecting chicksto the training conditions alone did not give rise to achange in the amount of CO-I unless there was evidenceof learning. (iii) The amount of CO-I was not correlated withapproach activity during training, indicating that locomotoractivity during training per se was without significant effecton the level of CO-I. Side effects of training such as sen-sory stimulation, movement, arousal or motivational statecannot therefore account for the results obtained. (iv) Theresults for CO-I were regionally specific and contrast withthe findings for the left PPN where the amount of CO-I wasnot correlated with preference score but was negativelycorrelated with approach activity during training.
The results for CO-II followed a similar pattern to thosefor CO-I in the left IMM, and were also regionally specific.There was a positive correlation between the amount ofCO-II in the region and preference score (Fig. 3B), and theamount of the protein was not significantly correlated withtraining approach. Furthermore the interpolated level of theprotein corresponding to a preference score of 50 did notdiffer significantly from the mean amount in untrainedchicks.
The learning-related biochemical changes that occur inthe left IMM after training (see above) (Solomonia et al.,1997, 1998, 2003) would require an adequate energy sup-ply; the increases in CO-I and CO-II in this brain regionmay reflect the mitochondrial response to these demands(see Attwell and Laughlin, 2001).
There are other links between mitochondrial functionand some of the neural changes that follow imprinting. Thislearning process leads to both presynaptic and postsyn-aptic changes in the left IMM and so may be expected toengage mitochondria. The mean length of the PSD ofdendritic spine synapses in the region is increased inimprinted chicks compared with the length in untrainedcontrols (Horn et al., 1985). The PSD of spine synapsesanchors glutamate receptors, including calcium-permeableNMDA receptors (for review see Sheng and Hoogenraad,2007). Synaptic activation of dendritic spines modified byimprinting is likely to lead to enhanced Ca2� entry into thepines through the larger PSD. This enhanced influx ofa2� may trigger downstream signaling cascades, includ-
ing the activation of protein kinases (for review seeDerkach et al., 2007) and hence increase the energy re-quirements of the modified spines. The increased influx of
Ca2� may lead to the translocation of a mitochondrion tothe base of a modified spine and so meet this enhancedenergy need (Li et al., 2004; Mattson et al., 2008;MacAskill and Kittler, 2010). In respect of the presynapticside of a synapse, it has been suggested that mitochondriaare involved in the recycling of synaptic vesicles (Rowlandet al., 2000; Perkins et al., 2010). Imprinting influences akey element in this process, clathrin heavy chain protein,the amount of which increases in the left IMM with thestrength of learning (Solomonia et al., 1997). Mitochondriaalso buffer cytoplasmic Ca2� and so may influence trans-
ission in some presynaptic terminals (Billups and For-ythe, 2002, for review see MacAskill et al., 2010), as doesmprinting (Meredith et al., 2004).
�-fodrin and HSP90
�-fodrin levels varied with preference score only in the leftMM; in this region protein level decreased as preferencecore increased (Fig. 6). Furthermore, the amount of pro-ein corresponding to a preference score of 50 was signif-cantly greater than the mean value for untrained chicks.his finding raises the possibility that some side effect of
raining, such as a high level of arousal elicited for exampley handling, led to an initial increase in the amount of
�-fodrin in the left IMM. It is also possible that, as thehicks became familiar with and learned the features ofhe imprinting stimulus, the level of arousal declined, as didhe level of �-fodrin. This pattern of change was differentrom that found for �-fodrin in the right PPN; training sig-nificantly increased the mean amount of �-fodrin in theight PPN (Table 1), but the amount was not correlatedith preference score (r�0.090; P�0.72; see Table 2).hese findings suggest that for the right PPN (i) the train-
ng-related increase was attributable to some side effect ofhe training procedure, possibly arousal, and (ii) this in-rease was stable, being uninfluenced by learning or byurther changes in levels of arousal.
Although the expression of HSP90 mRNA in the leftMM was found by subtractive hybridization to be greater introngly imprinted chicks than in weakly imprinted chicks4 h after training (Solomonia et al., 2003), in the presenttudy no significant changes were found for this protein inny of the brain regions investigated. The difference be-ween left IMM samples detected by subtractive hybridiza-ion may, in the absence of replication, have been due tohance. However, it is possible that the increased HSP90RNA observed after training (Solomonia et al., 2003) wasot translated.
CONCLUSION
The amounts of CO-I and of CO-II in the left IMM arestrongly correlated; chicks with higher levels of CO-II arealso likely to have higher levels of CO-I. This relationshipdoes not depend on learning and it is regionally specific;the levels of the two proteins are not significantly corre-lated in other brain regions. These findings raise the pos-sibility that the molecular mechanisms involved in the co-ordinated assembly of cytochrome c oxidase are preco-
ciously developed in the left IMM compared to the other
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123122
regions studied. This precocious development may enablethe region to function as a memory store within hours afterthe chick has hatched. Our results also provide strongevidence that mitochondria are involved in learning andmemory; the amounts of the two proteins increase with thestrength of prior learning. Taken together these findingsare consistent with a model involving (i) a factor or factorsconstitutively present in the left IMM but absent or lessabundant in the other brain regions studied, that coordi-nates the incorporation of the two subunits into the assem-bling enzyme complex; and (ii) additional factors, related tolearning, that enhance the pre-existing coordinated ex-pression of the two subunits.
Acknowledgments—We are grateful to Professor Sir John Walkerand Dr. Antonella Spinazolla for helpful discussions. This workwas supported by award N235 from the Georgian National Sci-ence Foundation.
REFERENCES
Ambalavanar R, McCabe BJ, Potter KN, Horn G (1999) Learning-related Fos-like immunoreactivity in the chick brain: time courseand co-localisation with GABA and parvalbumin. Neuroscience93:1515–1524.
Attwell D, Laughlin SB (2001) An energy budget for signaling in thegrey matter of the brain. J Cereb Blood Flow Metab 21:1133–1145.
Bateson PPG (1974) Atmospheric-pressure during incubation andposthatch behavior in chicks. Nature 284:805–807.
Billups B, Forsythe ID (2002) Presynaptic mitochondrial calcium se-questration influences transmission at mammalian central syn-apses. J Neurosci 22:5840–5847.
Bolhuis JJ (1991) Mechanisms of avian imprinting: a review. Biol Rev66:303–345.
Bolhuis JJ, Cook S, Horn G (2000) Getting better all the time: improv-ing preference scores reflect increases in the strength of filialimprinting. Anim Behav 59:1153–1159.
Bolhuis JJ, Gahr M (2006) Neural mechanisms of birdsong memory.Nat Rev Neurosci 7:347–357.
Bradley P, Horn G, Bateson P (1981) Imprinting: an electron micro-scopic study of chick hyperstriatum ventrale. Exp Brain Res41:115–120.
Bredenkötter M, Braun K (1997) Changes of neuronal responsivenessin the mediorostral neostriatum/hyperstriatum after auditory filialimprinting in the domestic chick. Neuroscience 76:355–365.
Brown MW, Horn G (1994) Learning-related alterations in the visualresponsiveness of neurons in a memory system of the chick brain.Eur J Neurosci 6:1479–1490.
Cipolla-Neto J, Horn G, McCabe BJ (1982) Hemispheric asymmetryand imprinting: the effect of sequential lesions to the hyperstriatumventrale. Exp Brain Res 48:22–27.
D’Souza SM, Brown IR (1998) Constitutive expression of heat shockproteins HSP90, HSC70, HSP70 and HSP60 in neural and non-neural tissues of the rat during postnatal development. Cell StressChaperones 3:188–199.
Derkach VA, Oh MC, Guire ES, Soderling TR (2007) Regulatorymechanisms of AMPA receptors in synaptic plasticity. Nat RevNeurosci 8:101–113.
Dittmer A, Dittmer J (2006) beta-actin is not a reliable loading controlin Western blot analysis. Electrophoresis 27:2844–2845.
Fontanesi F, Soto IC, Horn D, Barrientos A (2006) Assembly of mito-chondrial cytochrome c-oxidase, a complicated and highly regu-lated cellular process. Am J Physiol 291:C1129–C1147.
Gennis R, Ferguson-Miller S (1995) Structure of cytochrome c oxi-
dase, energy generator of aerobic life. Science 269:1063–1064.Gibbs ME, O’Dowd BS, Hertz E, Hertz L (2006) Astrocytic energymetabolism consolidates memory in young chicks. Neuroscience141:9–13.
Gibbs ME, Summers RJ (2002) Role of adrenoceptor subtypes inmemory consolidation. Prog Neurobiol 67:345–391.
Gobes SM, Zandbergen MA, Bolhuis JJ (2010) Memory in the making:localised brain activation to song learning in young songbirds. ProcR Soc B 277:3343–3351.
Honey RC, Horn G, Bateson P, Walpole M (1995) Functionally distinctmemories for imprinting stimuli—behavioral and neural dissocia-tions. Behav Neurosci 109:689–698.
Horn G (1952) The neurological basis of thought. Mermaid 18:17–25([email protected]_20100401_091829.pdf).
Horn G (1985) Memory, imprinting and the brain. Oxford: OxfordUniversity Press.
Horn G (1991) Technique for removing IMHV from the chick brain. In:Neural and behavioural plasticity: the use of the domestic chick asa model (Andrew RJ, ed), pp 44–48. Oxford: Oxford UniversityPress.
Horn G (2004) Pathways of the past: the imprint of memory. Nat RevNeurosci 5:108–120.
Horn G, Bradley P, McCabe BJ (1985) Changes in the structure ofsynapses associated with learning. J Neurosci 5:3161–3168.
Horn G, Johnson MH (1989) Memory systems in the chick: dissocia-tions and neuronal analysis. Neuropsychologia 27:1–22.
Horn G, Nicol AU, Brown MW (2001) Tracking memory’s trace. ProcNatl Acad Sci U S A 98:5282–5287.
Jackson C, McCabe BJ, Nicol AU, Grout AS, Brown MW, Horn G(2008) Dynamics of a memory trace: effects of sleep on consoli-dation. Curr Biol 18:393–400.
Johnson MH, Horn G (1986) Dissociation of recognition memory andassociative learning by a restricted lesion of the chick forebrain.Neuropsychologia 24:329–340.
Johnston AN, Rogers LJ, Johnston GAR (1993) Glutamate and im-printing memory—the role of glutamate receptors in the encodingof imprinting memory. Behav Brain Res 54:137–143.
Kelly DP, Scarpulla RC (2004) Transcriptional regulatory circuits con-trolling mitochondrial biogenesis and function. Genes Dev18:357–368.
Khalimonchuk E, Rodel G (2005) Biogenesis of cytochrome c oxidase.Mitrochondrion 5:363–388.
Li Z, Okamoto K-I, Hayashi Y, Sheng M (2004) The importance ofdendritic mitochondria in the morphogenesis and plasticity ofspines and synapses. Cell 119:873–887.
Lynch G, Baudry M (1984) The biochemistry of memory: a new andspecific hypothesis. Science 224:1057–1063.
Lynch G, Rex CS, Gall CM (2007) LTP consolidation: substrates,explanatory power, and functional significance. Neuropharmacol-ogy 52:12–23.
MacAskill AF, Kittler JT (2010) Control of mitochondrial transport andlocalization in neurons. Trends Cell Biol 20:102–112.
MacAskill AF, Atkin TA, Kittler JT (2010) Mitochondrial trafficking andthe provision of energy and calcium buffering at excitatory syn-apses. Eur J Neurosci 32:231–240.
Malchiodi-Albedi F, Ceccarini M, Winkelmann JC, Morrow JS, PetrucciTC (1993) The 270-kDa splice variant of erythrocyte beta-spectrin(beta-I-sigma-2) segregates in vivo and in vitro to specific domainsof cerebellar neurons. J Cell Sci 106:67–78.
Mattson MP, Gleichmann M, Cheng A (2008) Mitochondria in neuro-plasticity and neurological disorders. Neuron 60:748–766.
McCabe BJ, Cipolla-Neto J, Horn G, Bateson P (1982) Amnesiceffects of bilateral lesions placed in the hyperstriatum ventrale ofthe chick after imprinting. Exp Brain Res 48:13–21.
McCabe BJ, Horn G (1988) Learning and memory: regional changes inN-methyl-D-aspartate receptors in the chick brain after imprinting.
Proc Natl Acad Sci U S A 85:2849–2853.
S
S
S
W
R. O. Solomonia et al. / Neuroscience 194 (2011) 112–123 123
McCabe BJ, Horn G (1994) Learning-related changes in Fos-likeimmunoreactivity in the chick forebrain after imprinting. Proc NatlAcad Sci U S A 91:11417–11421.
McCabe BJ, Horn G, Bateson PPG (1981) Effects of restricted lesionsof the chick forebrain on the acquisition of filial preferences duringimprinting. Brain Res 205:29–37.
McCabe BJ, Kendrick KM, Horn G (2001) �-aminobutyric acid, taurineand learning: release of amino acids from slices of chick brainfollowing filial imprinting. Neuroscience 105:317–324.
Meredith RM, McCabe BJ, Kendrick KM, Horn G (2004) Amino acidneurotransmitter release and learning: a study of visual imprinting.Neuroscience 126:249–256.
Mick DU, Fox TD, Rehling P (2011) Inventory control: cytochrome coxidase assembly regulates mitochondrial translation. Nat Rev MolCell Biol 12:14–20.
Nicol AU, Brown MW, Horn G (1995) Neurophysiological investiga-tions of a recognition memory system for imprinting in the domesticchick. Eur J Neurosci 7:766–776.
Perkins GA, Tjong J, Brown JM, Poquiz PH, Scott RT, Kolson DR,Ellisman MH, Spirou GA (2010) The micro-architecture of mito-chondria at active zones: electron tomography reveals novel an-choring scaffolds and cristae structured for high-rate metabolism.J Neurosci 30:1015–1026.
Rose SPR (2000) God’s organism? The chick as a model system formemory studies. Learn Mem 7:1–17.
Rowland KC, Irby NK, Spirou GA (2000) Specialized synapse-associ-
ated structures within the Calyx of Held. J Neurosci 20:9135–9144.Sakisaka T, Meerlo T, Matteson J, Plutner H, Balch WE (2002) Rab-alphaGDI activity is regulated by a HSP90 chaperone complex.EMBO J 21:6125–6135.
Sheng M, Hoogenraad C (2007) The postynaptic architecture of ex-citatory synapses: a more quantitative view. Annu Review Biochem76:823–847.
Sheu FS, McCabe BJ, Horn G, Routtenberg A (1993) Learning selec-tively increases protein kinase C substrate phosphorylation in spe-cific regions of the chick brain. Proc Natl Acad Sci U S A90:2705–2709.
Sluckin W (1972) Imprinting and early learning. London: Methuen.Snedecor GW, Cochran WG (1989) Statistical methods. Ames: Iowa
State University Press.Solomonia RO, Kotorashvili A, Kiguradze T, McCabe BJ, Horn G
(2005) Ca2�/calmodulin protein kinase II and memory: learning-related changes in a localized region of the domestic chick brain.J Physiol 569:643–653.
olomonia RO, McCabe BJ, Horn G (1998) Neural cell adhesionmolecules, learning and memory in the domestic chick. BehavNeurosci 112:646–655.
olomonia RO, McCabe BJ, Jackson AP, Horn G (1997) Clathrinproteins and recognition memory. Neuroscience 80:59–67.
olomonia RO, Morgan K, Kotorashvili A, McCabe BJ, Jackson AP,Horn G (2003) Analysis of differential gene expression supports arole for amyloid precursor protein and a protein kinase C substrate(MARCKS) in long-term memory. Eur J Neurosci 17:1073–1081.
arton DI, Wright IJ, Falster DS, Westoby M (2006) Bivariate line-
fitting methods for allometry. Biol Rev 81:259–291.(Accepted 22 July 2011)(Available online 1 August 2011)
Copyright © 2022 FDOKUMEN




















