
Mitochondrial diversity of Western spadefoot toads, Pelobates cultripes, in northwestern Spain
6
Mitochondrial diversity of Western spadefoot toads, Pelobates cultripes, in northwestern Spain Angelica Crottini 1,2,* , Pedro Galán 3 , Miguel Vences 1 Abstract. We analysed the DNA sequences of the mitochondrial cytochrome b gene for 49 specimens of the Western spadefoot toad, Pelobates cultripes, from Spain, and one sample from France, with a focus on populations from Galicia in northwestern Spain. Our results point to a generally low mitochondrial differentiation of the species across its distributional range although some phylogeographic structure was detected. Galician populations from coastal localities, from the A Limia region, and from Monforte appeared to be closely related, whereas the only available sample from the relict Galician population from near O Barco de Valdeorras has the same haplotype as the samples from Burgos, in central Spain. Keywords: Amphibia, cytochrome b, Galicia, genetic diversity, Iberian Peninsula, Pelobates cultripes, Pelobatidae. The family Pelobatidae consists of one ex- tant genus, Pelobates Wagler, 1830, contain- ing four species of Afro-European spadefoot toads (García-París, Buchholz and Parra-Olea, 2003). This genus includes predominantly bur- rowing species, generally not considered to be very mobile over long distances (Eggert, 2002). Spadefoot toads have narrow habitat require- ments (Shpun et al., 1993; Martínez-Rica, 1997; Nöllert, 1997), and environmental conditions strongly influence their population dynamics. Therefore paleoenvironmental conditions have played a key role in structuring current popula- tions (Eggert et al., 2006; Crottini et al., 2007). The Western spadefoot toad, Pelobates cultripes Cuvier, 1829, is distributed across most of the Iberian Peninsula (except for the north), along the French Mediterranean littoral, and in iso- lated populations at the Western Atlantic French coast (Lizana, 1997a, 1997b; ACEMAV, Duguet 1 - Zoological Institute, Technical University of Braun- schweig, Spielmannstr. 8, 38106 Braunschweig, Ger- many 2 - Sezione di Zoologia e Citologia, Dipartimento di Biolo- gia, Universitá degli Studi di Milano, Via Celoria 26, 20133 Milano, Italy 3 - Departamento de Bioloxía Animal, Bioloxía Vexetal e Ecoloxía, Facultade de Ciencias, Universidade da Coruña, 15071 A Coruña, Spain; e-mail: [email protected] * Corresponding author; e-mail: [email protected] and Melki, 2003; Malkmus, 2004). Pelobates cultripes is a widespread Iberian species with, however, a relatively small area of occupancy within its range. In the Iberian Peninsula it is mostly present in the Mediterranean region and it is more abundant in the south, but is ab- sent from the Cantabrian area and the Pyre- nees (Tejedo and Reques, 2002; Cruz, 2008). It mostly inhabits siliceous sandy soils, where its populations are more uniformly distributed, while on calcareous substrates its populations are scarce (García-París, Montori and Herrero, 2004). The species is typically found in exposed habitats with low vegetation cover and tempo- rary lentic water bodies. In the region of Galicia, northwest of Spain, Pelobates cultripes is the amphibian with the most restricted distribution, only occurring pat- chily in the south. Historically it was present in Galician inland zones (Monforte de Lemos, in Lugo; O Barco de Valdeorras, Verín, and A Limia in Ourense; Gándaras de Budiño in Pontevedra) and on the southern coast, princi- pally in the Rías Baixas (Pontevedra) and the southern littoral of A Coruña province (Galán and Fernández, 1993; Balado, Bas and Galán, 1995; Tejedo and Reques, 2002). The species is generally not threatened, except for some pe- ripheral populations. However, during the last three decades several Galician populations have © Koninklijke Brill NV, Leiden, 2010. Amphibia-Reptilia 31 (2010): 443-448 Also available online - www.brill.nl/amre
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Mitochondrial diversity of Western spadefoot toads, Pelobates cultripes, in northwestern Spain

Mitochondrial diversity of Western spadefoot toads,Pelobates cultripes, in northwestern Spain
Angelica Crottini1,2,*, Pedro Galán3, Miguel Vences1
Abstract. We analysed the DNA sequences of the mitochondrial cytochrome b gene for 49 specimens of the Westernspadefoot toad, Pelobates cultripes, from Spain, and one sample from France, with a focus on populations from Galicia innorthwestern Spain. Our results point to a generally low mitochondrial differentiation of the species across its distributionalrange although some phylogeographic structure was detected. Galician populations from coastal localities, from the ALimia region, and from Monforte appeared to be closely related, whereas the only available sample from the relict Galicianpopulation from near O Barco de Valdeorras has the same haplotype as the samples from Burgos, in central Spain.
Keywords: Amphibia, cytochrome b, Galicia, genetic diversity, Iberian Peninsula, Pelobates cultripes, Pelobatidae.
The family Pelobatidae consists of one ex-tant genus, Pelobates Wagler, 1830, contain-ing four species of Afro-European spadefoottoads (García-París, Buchholz and Parra-Olea,2003). This genus includes predominantly bur-rowing species, generally not considered to bevery mobile over long distances (Eggert, 2002).Spadefoot toads have narrow habitat require-ments (Shpun et al., 1993; Martínez-Rica, 1997;Nöllert, 1997), and environmental conditionsstrongly influence their population dynamics.Therefore paleoenvironmental conditions haveplayed a key role in structuring current popula-tions (Eggert et al., 2006; Crottini et al., 2007).The Western spadefoot toad, Pelobates cultripesCuvier, 1829, is distributed across most of theIberian Peninsula (except for the north), alongthe French Mediterranean littoral, and in iso-lated populations at the Western Atlantic Frenchcoast (Lizana, 1997a, 1997b; ACEMAV, Duguet
1 - Zoological Institute, Technical University of Braun-schweig, Spielmannstr. 8, 38106 Braunschweig, Ger-many
2 - Sezione di Zoologia e Citologia, Dipartimento di Biolo-gia, Universitá degli Studi di Milano, Via Celoria 26,20133 Milano, Italy
3 - Departamento de Bioloxía Animal, Bioloxía Vexetale Ecoloxía, Facultade de Ciencias, Universidade daCoruña, 15071 A Coruña, Spain; e-mail: [email protected]*Corresponding author; e-mail: [email protected]
and Melki, 2003; Malkmus, 2004). Pelobatescultripes is a widespread Iberian species with,however, a relatively small area of occupancywithin its range. In the Iberian Peninsula it ismostly present in the Mediterranean region andit is more abundant in the south, but is ab-sent from the Cantabrian area and the Pyre-nees (Tejedo and Reques, 2002; Cruz, 2008).It mostly inhabits siliceous sandy soils, whereits populations are more uniformly distributed,while on calcareous substrates its populationsare scarce (García-París, Montori and Herrero,2004). The species is typically found in exposedhabitats with low vegetation cover and tempo-rary lentic water bodies.
In the region of Galicia, northwest of Spain,Pelobates cultripes is the amphibian with themost restricted distribution, only occurring pat-chily in the south. Historically it was presentin Galician inland zones (Monforte de Lemos,in Lugo; O Barco de Valdeorras, Verín, andA Limia in Ourense; Gándaras de Budiño inPontevedra) and on the southern coast, princi-pally in the Rías Baixas (Pontevedra) and thesouthern littoral of A Coruña province (Galánand Fernández, 1993; Balado, Bas and Galán,1995; Tejedo and Reques, 2002). The speciesis generally not threatened, except for some pe-ripheral populations. However, during the lastthree decades several Galician populations have
© Koninklijke Brill NV, Leiden, 2010. Amphibia-Reptilia 31 (2010): 443-448Also available online - www.brill.nl/amre

444 Short Note
become extinct, e.g., in Gándaras de Budiño inthe inland, in Carnota-Louro, and in the RíasBaixas area, mostly due to the alteration of wet-lands and the introduction of invasive species(Galán, 2008).
Pelobates cultripes is the sister species of theNorth African P. varaldii (García-París, Buch-holz and Parra-Olea, 2003; Veith et al., 2006).Besides general assessments of the phyloge-netic position of Pelobates cultripes amongpelobatoid frogs on the basis of partial or com-plete mitochondrial sequences (García-París,Buchholz and Parra-Olea, 2003; Gissi et al.,2006; Veith et al., 2006), no studies havethus far addressed the molecular differentia-tion of this species. The aim of this study isto assess the molecular differentiation of thenorthwesternmost populations of Pelobates cul-tripes which currently occur in two major geo-graphic clusters of populations in Galicia: oneof these in coastal areas of southwestern Gali-cia, mainly in sandy dune areas, a second onein the inland close to the area of the nowdesiccated Antela lake around A Limia. Iso-lated relict populations also occur near Mon-forte de Lemos (in Lugo province) and O Barcode Valdeorras (in Ourense province). We com-pare mitochondrial DNA sequences of indi-viduals from these populations with represen-tative samples from other sites in Spain anda sample from the French Mediterranean lit-toral.
We extracted total genomic DNA from finclips of tadpoles preserved in 99% ethanol usinga salt extraction protocol (Bruford et al., 1992).We amplified fragments of 689 bp of the mi-tochondrial cytochrome b gene via polymerasechain reaction using the primers L15162 (Taber-let, Meyer and Bouvet, 1992) and H15915 (Ir-win, Kocher and Wilson, 1991) following Crot-tini et al. (2007). A minimum-spanning net-work was constructed using the TCS softwarepackage (Clement, Posada and Crandall, 2000),which employs the statistical parsimony methodof Templeton, Crandall and Sing (1992). Carewas taken to use only one tadpole per pond
(sometimes two tadpoles from large ponds) toavoid sampling full siblings. Altogether, DNAsequences of 28 specimens from eight Galicianlocalities and 20 specimens of 11 additionalpopulations across Spain (fig. 1 and table 1)were determined and deposited in Genbank (ac-cession numbers: GU983048-GU983095). TwoCyt b sequences of P. cultripes were retrievedfrom GenBank (Accession Numbers: AJ871086and DQ333373).
The resulting haplotype network (fig. 1) con-sists of 12 haplotypes separated by a maximumof six mutational steps. However, because ofthe restricted number of individuals studied weassume that some additional genetic variationexists within this species, especially consider-ing that only one sample from France was usedand we did not include samples from Portu-gal.
The central haplotype H2 is predominant inall Galician populations from the southern coast(populations: 1-4), from A Limia (Ponteliñares,7 and Porqueira, 8), and from Monforte deLemos (5). Haplotype H2 was not found inthe population from O Barco de Valdeorras (6)where, however, only one individual was se-quenced. In addition, we observed one singletonhaplotype each in one coastal population (H1; atA Lanzada, population 4) and at A Limia (Pon-teliñares, population 7) (H3). The predominantGalician haplotype H2 was shared with popula-tions from Madrid (19), Ávila (10) and Toledo(11-11b) in central Spain. In contrast, the Gali-cian population from O Barco de Valdeorras (6)shared its haplotype (H4) with the populationfrom Burgos (9). For distribution of haplotypesin other localities, see fig. 1 and table 1. Inter-estingly, in Spain the haplotype H8 has beenfound along the Mediterranean littoral in Mur-cia (16) and Granada (17), and also in France(20).
Our data point to a general low mitochondr-ial differentiation of the Western spadefood toadacross most of its range. In Galicia, the pres-ence of altogether four haplotypes in 28 sam-ples clearly suggests a lower variability than
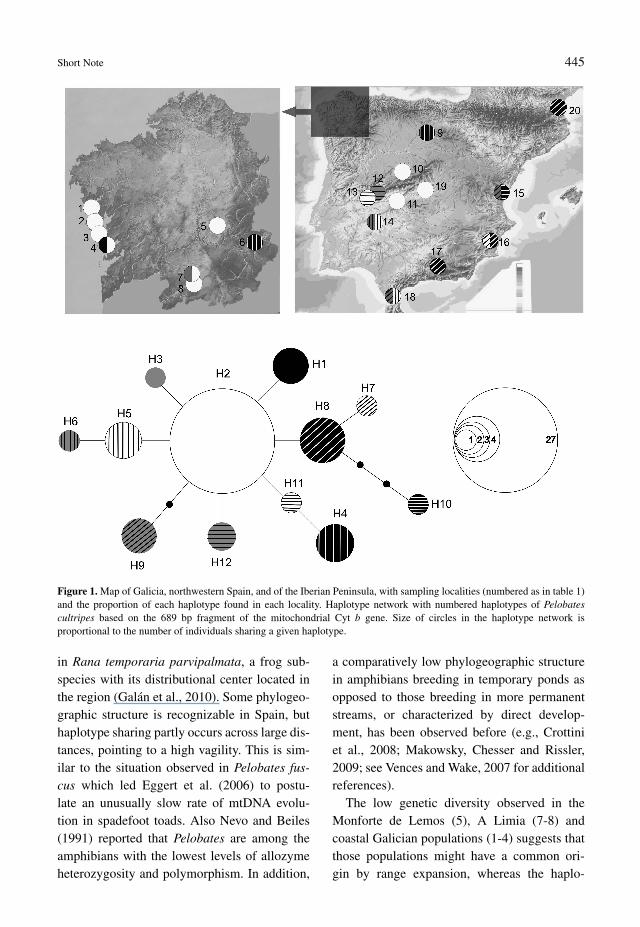
Short Note 445
Figure 1. Map of Galicia, northwestern Spain, and of the Iberian Peninsula, with sampling localities (numbered as in table 1)and the proportion of each haplotype found in each locality. Haplotype network with numbered haplotypes of Pelobatescultripes based on the 689 bp fragment of the mitochondrial Cyt b gene. Size of circles in the haplotype network isproportional to the number of individuals sharing a given haplotype.
in Rana temporaria parvipalmata, a frog sub-species with its distributional center located inthe region (Galán et al., 2010). Some phylogeo-graphic structure is recognizable in Spain, buthaplotype sharing partly occurs across large dis-tances, pointing to a high vagility. This is sim-ilar to the situation observed in Pelobates fus-cus which led Eggert et al. (2006) to postu-late an unusually slow rate of mtDNA evolu-tion in spadefoot toads. Also Nevo and Beiles(1991) reported that Pelobates are among theamphibians with the lowest levels of allozymeheterozygosity and polymorphism. In addition,
a comparatively low phylogeographic structurein amphibians breeding in temporary ponds asopposed to those breeding in more permanentstreams, or characterized by direct develop-ment, has been observed before (e.g., Crottiniet al., 2008; Makowsky, Chesser and Rissler,2009; see Vences and Wake, 2007 for additionalreferences).
The low genetic diversity observed in theMonforte de Lemos (5), A Limia (7-8) andcoastal Galician populations (1-4) suggests thatthose populations might have a common ori-gin by range expansion, whereas the haplo-

446 Short Note
Table 1. Locality number (as in map in fig. 1), localities sampled, province, number of individuals sampled per locality (N ),and haplotypes encountered at each locality (haplotype number as in fig. 1, with numbers of individuals per haplotype inparentheses).
Locality number Locality Province N Haplotypes
1 Lagoas de Xuño and Muro, Porto do Son A Coruña 6 H2 (6)2 Corrubedo, Ribeira A Coruña 6 H2 (6)3 Siradella, O Grove Pontevedra 1 H2 (1)4 A Lanzada, O Grove Pontevedra 6 H1 (3), H2 (3)5 Monforte de Lemos Lugo 3 H2 (3)6 O Barco de Valdeorras Ourense 1 H4 (1)7 Ponteliñares, Rairiz de Veiga, A Limia Ourense 2 H2 (1), H3 (1)8 Porqueira, A Limia Ourense 3 H2 (3)
9 Cubillo del Campo Burgos 2 H4 (2)10 El Hoyo de Pinares Ávila 1 H2 (1)11 Fuentes Toledo 1 H2 (1)
11b Pozo del Cordel de Merinos Toledo 1 H2 (1)12 Near Monfragüe Cáceres 2 H12 (2)13 Los Barruecos Cáceres 1 H11 (1)14 Mérida Badajoz 2 H5 (1), H6 (1)15 Pla dels Vorrals Valencia 2 H9 (1), H10 (1)16 Gaitán Murcia 2 H7 (1), H8 (1)17 Granada Granada 2 H8 (2)18 Tarifa Cádiz 4 H5 (2), H9 (2)19 Las Rozas; AJ871086 Madrid 1 H2 (1)20 Argeles-Sur-Mer, France; DQ333373 Languedoc-Roussillon 1 H8 (1)
type sharing between the sample from the relictpopulation of O Barco de Valdeorras and thesamples from Burgos suggests a closer connec-tions to the eastern central Spanish populations.Further samples from O Barco de Valdeorrasneed to be investigated to verify this hypoth-esized close relationship. For more conclusivephylogeographic analyses it will be crucial toinclude several populations from adjacent Por-tugal and France and to sensibly increase thepopulation sampling sizes of the non Galicianlocalities. Besides ensuring a more comprehen-sive geographic coverage, additional studies areneeded to reduce the bias of small samplingsizes in order to ensure a better representationof the genetic diversity hosted by each popula-tion, and include additional (nuclear) markers toobtain a more complete assessment of geneticdiversity in Pelobates cultripes.
Acknowledgements. This study was performed with au-thorization from the regional government of Galicia (Direc-ción Xeral de Conservación da Natureza, Xunta de Galicia).Field data and tissue samples from Galicia were obtained as
part of the project “Elaboración das bases do plan de con-servación de Pelobates cultripes en Galicia”. Martiño Ca-bana Otero and Ricardo Ferreiro Sanjurjo contributed sev-eral samples. Manuel Arzúa Piñeiro provided informationon the population of O Barco de Valdeorras. Comparativetissue samples from localities across Spain were obtainedfrom the tissue collection of the Museo Nacional de Cien-cias Naturales in Madrid; we are particularly grateful to Is-abel Rey Fraile for her efficient and fast processing of ourtissue loan request, and to Mario García-París for his openexchange of information. AC was supported by a fellowshipof the Universitá degli Studi di Milano.
References
ACEMAV coll., Duguet, R., Melki, F. (Eds) (2003): LesAmphibiens de France, Belgique et Luxembourg. Col-lection Parthénope, Éditions Biotope, Mèze.
Balado, R., Bas, S., Galán, P. (1995): Anfibios e réptiles. In:Atlas de Vertebrados de Galicia. Aproximación a dis-tribución dos vertebrados terrestres de Galicia durante oquinquenio 1980-85. Tomo 1: Peixes, Anfibios, Réptilese Mamíferos, p. 65-170. Consello da Cultura Galega& Sociedade Galega de Historia Natural, Eds, AgenciaGráfica, Santiago de Compostela.
Bruford, M.W., Hanotte, O., Brookfield, J.F.Y., Burke, T.(1992): Single-locus and multilocus DNA fingerprint-ing. In: Molecular Genetic Analysis of Populations:A Practical Approach, p. 225-270. Hoelzel, A.R., Ed.,IRL Press, Oxford.

Short Note 447
Clement, X., Posada, D., Crandall, K. (2000): TCS: a com-puter program to estimate gene genealogies. Mol. Ecol.9: 1657-1659.
Crottini, A., Andreone, F., Kosuch, J., Borkin, L.,Litvinchuk, S.N., Eggert, C., Veith, M. (2007): Fosso-rial but widespread: the phylogeography of the commonspadefoot toad (Pelobates fuscus), and the role of thePo Valley as a major source of genetic variability. Mol.Ecol. 16: 2734-2754.
Crottini, A., Chiari, Y., Mercurio, V., Meyer, A., Vences,M., Andreone, F. (2008): Into the canyons: the phylo-geography of the Malagasy frogs Mantella expectata andScaphiophryne gottlebei in the arid Isalo Massif, andits significance for conservation (Amphibia: Mantellidaeand Microhylidae). Org. Divers. Evol. 8: 368-377.
Cruz, M.J. (2008): Pelobates cultripes (Cuvier, 1829). In:Atlas dos Anfíbios e Répteis de Portugal, p. 110-111.Loureiro, A., Ferrand de Almeida, N., Carretero, M.A.,Paulo, O.S., Eds, Instituto de Conservação da Naturezae da Biodiversidade, Lisboa.
Eggert, C. (2002): Use of fluorescent pigments and im-plantable transmitters to track a fossorial toad (Pelobatesfuscus). Herpetol. J. 12: 69-74.
Eggert, C., Cogalniceanu, D., Veith, M., Dzukic, G., Taber-let, P. (2006): The declining spadefoot toad, Pelobatesfuscus (Pelobatidae): paleo and recent environmentalchanges as a major influence on current population struc-ture and status. Conserv. Gen. 7: 185-195.
Galán, P. (2008): Asistencia técnica para la elaboraciónde las bases del plan de conservación de Pelobatescultripes en Galicia. I Parte: diagnóstico. Situación dela especie. Unpublished document, Dirección Xeral deConservación da Natureza, Xunta de Galicia, Santiagode Compostela.
Galán, P., Fernández, G. (1993): Anfibios e réptiles deGalicia. Edicions Xerais, Vigo.
Galán, P., Ludewig, A.-K., Kmiec, J., Hauswaldt, S., Ca-bana, M., Ferreiro, R., Vences, M. (2010): Low mi-tochondrial divergence of rediscovered southern relictpopulations of Rana temporaria parvipalmata in Spain.Amphibia-Reptilia 31: 144-148.
García-París, M., Buchholz, D.R., Parra-Olea, G. (2003):Phylogenetic relationships of Pelobatoidea re-examinedusing mtDNA. Mol. Phylogenet. Evol. 28: 12-23.
García-París, M., Montori, A., Herrero, P. (2004): Am-phibia, Lissamphibia. In: Fauna Ibérica, Vol. 24, p. 1-640. Ramos, M.A. et al., Eds, Museo Nacional de Cien-cias Naturales, CSIC, Madrid.
Gissi, C., San Mauro, D., Pesole, G., Zardoya, R. (2006):Mitochondrial phylogeny of Anura (Amphibia): a casestudy of congruent phylogenetic reconstruction usingamino acid and nucleotide characters. Gene 366: 228-237.
Irwin, D.M., Kocher, T.D., Wilson, A.C. (1991): Evolutionof the cytochrome b gene of mammals. J. Mol. Evol. 32:128-144.
Lizana, M. (1997a): Pelobates cultripes (Cuvier, 1829).Sapo de espuelas, Sapo-de-unha-negra, Gripaud’esperons (C), Sapo de esporóns (G), Apo ezproidunarrunta (V). In: Distribución y biogeografía de los
Anfibios y Reptiles en España y Portugal. Monografíasde Herpetología, n◦ 3, p. 140-142. Pleguezuelos, J.M.,Ed., Editorial Universidad de Granada & AsociaciónHerpetológica Española, Granada.
Lizana, M. (1997b): Pelobates cultripes (Cuvier, 1829). In:Atlas of Amphibians and Reptiles in Europe, p. 108-109. Gasc, J.-P., Cabela, A., Crnobrnja-Isailovic, J.,Dolmen, D., Grossenbacher, K., Haffner, P., Lescure, J.,Martens, H., Martínez Rica, J.P., Maurin, H., Oliveira,M.E., Sofianidou, T.S., Veith, M., Zuiderwijk, A., Eds,Societas Europaea Herpetologica & Muséum Nationald’Histoire Naturelle (IEGB/SPN), Paris.
Makowsky, R., Chesser, J., Rissler, L.J. (2009): A strikinglack of genetic diversity across the wide-ranging am-phibian Gastrophryne carolinensis (Anura: Microhyli-dae). Genetica 135: 169-183.
Malkmus, R. (2004): Amphibians and Reptiles of Portugal,Madeira and the Azores-Archipelago. Distribution andNatural History Notes. A.R.G. Gantner Verlag, Ruggell.
Martínez-Rica, J.P. (1997): General features of Europeanclimate and vegetation consequences on the distributionof herpetofauna. In: Atlas of Amphibians and Reptilesin Europe, p. 17-21. Gasc, J.-P., Cabela, A., Crnobrnja-Isailovic, J., Dolmen, D., Grossenbacher, K., Haffner, P.,Lescure, J., Martens, H., Martínez Rica, J.P., Maurin, H.,Oliveira, M.E., Sofianidou, T.S., Veith, M., Zuiderwijk,A., Eds, Societas Europaea Herpetologica & MuséumNational d’Histoire Naturelle (IEGB/SPN), Paris.
Nevo, E., Beiles, A. (1991): Genetic diversity and ecologi-cal heterogeneity in amphibian evolution. Copeia 1991:565-592.
Nöllert, A. (1997): Pelobates fuscus (Laurenti, 1768). In:Atlas of Amphibians and Reptiles in Europe, p. 110-111. Gasc, J.-P., Cabela, A., Crnobrnja-Isailovic, J.,Dolmen, D., Grossenbacher, K., Haffner, P., Lescure, J.,Martens, H., Martínez Rica, J.P., Maurin, H., Oliveira,M.E., Sofianidou, T.S., Veith, M., Zuiderwijk, A., Eds,Societas Europaea Herpetologica & Muséum Nationald’Histoire Naturelle (IEGB/SPN), Paris.
Shpun, S., Hoffman, E., Nevo, E., Katz, U. (1993): Is thedistribution of Pelobates syriacus related to its lim-ited osmoregulatory capacity? Comp. Biochem. Physiol.105A: 135-139.
Taberlet, P., Meyer, A., Bouvet, J. (1992): Unusual mito-chondrial DNA polymorphism in two local populationsof blue tit Parus caeruleus. Mol. Ecol. 1: 27-36.
Tejedo, M., Reques, R. (2002): Pelobates cultripes (Cuvier,1829). Sapo de espuelas. In: Atlas y libro rojo de losanfibios y reptiles de España, p. 94-96. Pleguezuelos,J.M., Márquez, R., Lizana, M., Eds, Dirección Generalde Conservación de la Naturaleza & Asociación Her-petológica Española, Madrid.
Templeton, A.R., Crandall, K.A., Sing, C.F. (1992):A cladistic analysis of phenotypic associations with hap-lotypes inferred from restriction endonuclease mappingand DNA sequence data. III. Cladogram estimation. Ge-netics 132: 619-633.

448 Short Note
Veith, M., Fromhage, L., Kosuch, J., Vences, M. (2006):Historical biogeography of Western Palearctic pelobatidand pelodytid frogs: a molecular phylogenetic perspec-tive. Contrib. Zool. 75: 109-120.
Vences, M., Wake, D.B. (2007): Speciation, species bound-aries and phylogeography of amphibians. In: AmphibianBiology, Vol. 6, Systematics, p. 2613-2669. Heatwole,
H.H., Tyler, M., Eds, Surrey Beatty & Sons, ChippingNorton, Australia.
Received: January 24, 2010. Accepted: March 15, 2010.




















