
Microfingerprinting analysis of human Ia molecules favours a three loci model
11
Molrculur Iww~oloyy Vol. 20, No. 3. pp. 333-343, 1983. Printed in Great Britain. 0161.5890’83/030333-I 1$03.00/O 0 1983Pergamon Press Ltd. MICROFINGERPRINTING ANALYSIS OF HUMAN Ia MOLECULES FAVOURS A THREE LOCI MODEL* ROSA SORRENTINO,t$j GIORGIO CORTE, FRANC0 CALABI,‘l NOBUYUKI TANIGAKIY and ROBERTO TOSIf Wruppo di Microbiologia e Patologia Generale, UniversitB di Roma, Italy; fLaboratorio di Biologia Cellulare, C.N.R.. Roma, Italy; //Istituto di Chimica Biologia, Universita di Genova. Italy; TDepartment of Immunology Research, Roswell Park Memorial Institute. Buffalo, New York, U.S.A. (First receirrd 7 June 1982: accrpted in rc,ri.wd form I1 October 1982) Abstract-Alpha subunits from DC1 la molecules, when compared with DR alpha subunits. are shown to possess distinctive features revealed by differences in microfingerprinting patterns after peptic diges- tion. Alpha chains from BR4X7 molecules differ from DC1 alpha chains and are more similar to DR alpha chains. Since DC1 and BR4X7 beta chains (which carry the HLA-controlled alloantigenic determinants) associate with different alpha subunits, it is considered unlikely that they are controlled by alleles at the same locus. The proposed model implies the existence of three tightly linked HLA loci controlling the beta subunits of DR. DC and BR molecules respectively. INTRODUCTION The study of the polymorphism of Ia-like mol- ecules has been a major task of human immu- nogenetics in the last few years. Through the systematic study of a large number of allo- antisera, mainly performed during the 7th and 8th International Histocompatibility Work- shops, ten allospecificities have been defined, each recognized by a cluster of internally corre- lating alloantisera. These specificities (num- bered from 1 to ~10) have been assigned to a single HLA-linked locus, named HLA-DR locus, on the basis of population and family studies. HLA-DR specificites have been shown to reside on Ia beta chains. In fact, specific alloantisera react with isolated, ‘251-labelled beta chains but not with alpha chains (Tosi et al., 1978; Tanigaki et al., 1980a) and structural differences between chains bearing different allospecificities are much more prominent for beta than for alpha chains (Corte et al., 1981~; Silver & Ferrone, 1979; Shackelford & Strom- inger, 1980). Human DR molecules are prob- ably homologous to murine I-E molecules (Corte et ul., 1981a; Delovitch & Falk, 1979; McMillan et al., 1977). * This work was partially supported by the C.N.R. grant no. 54/80.01525.96 and United States Health Service Grants AI08899, CA17276, and CA16056. $ Correspondence to Rosa Sorrentino, Laboratorio di Biologia Cellulare, Via Romagnosi 18A, 00196 Roma, Italy. Other specificities, recognized by distinct clusters of alloantisera, have been described, which are associated, at the population level, with two or more DR specificities. These have been called ‘supertypic specificities’ to differen- tiate them from the single, or ‘subtypic’ DR specificities. Detailed studies at the molecular level have been performed on two of these supertypic specificities, namely DC1 and BR4X7. The DC1 specificity resides on beta subunit of Ia molecules different from those which carry DR determinants (Tosi et al., 1978). Be- cause of a very high degree of linkage disequili- brium, DC1 is so strongly associated with the DR specificities DRl, DR2 and DRw6 to simu- late cross-reactivity between them. We have previously shown that DC1 molecules differ from DR molecules in papain sensitivity (Tani- gaki et al., 1980b) and in the rate of iodine uptake by the two chains (Tosi et al., 1978). More recently, we have found that not only beta chains, but also alpha chains from DC1 molecules markedly differ from those of DR molecules (Corte et al., 1981b). These findings indicate that beta chains of DC1 molecules are controlled by a second locus, closely linked to HLA-DR, which we name DC locus. The poss- ible homology between human DC molecules and murine I-A molecules has been previously discussed (Corte et al., 1981b). The other supertypic specificity, BR4X7, is possessed by most DR4-positive or DR7-posi- 333
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Microfingerprinting analysis of human Ia molecules favours a three loci model

Molrculur Iww~oloyy Vol. 20, No. 3. pp. 333-343, 1983. Printed in Great Britain.
0161.5890’83/030333-I 1$03.00/O 0 1983 Pergamon Press Ltd.
MICROFINGERPRINTING ANALYSIS OF HUMAN Ia MOLECULES FAVOURS A THREE LOCI MODEL*
ROSA SORRENTINO,t$j GIORGIO CORTE, FRANC0 CALABI,‘l NOBUYUKI TANIGAKIY and ROBERTO TOSIf
Wruppo di Microbiologia e Patologia Generale, UniversitB di Roma, Italy; fLaboratorio di Biologia Cellulare, C.N.R.. Roma, Italy;
//Istituto di Chimica Biologia, Universita di Genova. Italy; TDepartment of Immunology Research, Roswell Park Memorial Institute. Buffalo, New York, U.S.A.
(First receirrd 7 June 1982: accrpted in rc,ri.wd form I1 October 1982)
Abstract-Alpha subunits from DC1 la molecules, when compared with DR alpha subunits. are shown to possess distinctive features revealed by differences in microfingerprinting patterns after peptic diges- tion. Alpha chains from BR4X7 molecules differ from DC1 alpha chains and are more similar to DR alpha chains. Since DC1 and BR4X7 beta chains (which carry the HLA-controlled alloantigenic determinants) associate with different alpha subunits, it is considered unlikely that they are controlled by alleles at the same locus. The proposed model implies the existence of three tightly linked HLA loci controlling the beta subunits of DR. DC and BR molecules respectively.
INTRODUCTION
The study of the polymorphism of Ia-like mol- ecules has been a major task of human immu- nogenetics in the last few years. Through the systematic study of a large number of allo- antisera, mainly performed during the 7th and
8th International Histocompatibility Work- shops, ten allospecificities have been defined, each recognized by a cluster of internally corre- lating alloantisera. These specificities (num- bered from 1 to ~10) have been assigned to a single HLA-linked locus, named HLA-DR locus, on the basis of population and family studies. HLA-DR specificites have been shown to reside on Ia beta chains. In fact, specific alloantisera react with isolated, ‘251-labelled
beta chains but not with alpha chains (Tosi et al., 1978; Tanigaki et al., 1980a) and structural differences between chains bearing different allospecificities are much more prominent for beta than for alpha chains (Corte et al., 1981~; Silver & Ferrone, 1979; Shackelford & Strom- inger, 1980). Human DR molecules are prob- ably homologous to murine I-E molecules (Corte et ul., 1981a; Delovitch & Falk, 1979; McMillan et al., 1977).
* This work was partially supported by the C.N.R. grant no. 54/80.01525.96 and United States Health Service Grants AI08899, CA17276, and CA16056.
$ Correspondence to Rosa Sorrentino, Laboratorio di Biologia Cellulare, Via Romagnosi 18A, 00196 Roma, Italy.
Other specificities, recognized by distinct
clusters of alloantisera, have been described, which are associated, at the population level, with two or more DR specificities. These have been called ‘supertypic specificities’ to differen- tiate them from the single, or ‘subtypic’ DR specificities. Detailed studies at the molecular level have been performed on two of these supertypic specificities, namely DC1 and
BR4X7. The DC1 specificity resides on beta subunit
of Ia molecules different from those which carry DR determinants (Tosi et al., 1978). Be-
cause of a very high degree of linkage disequili- brium, DC1 is so strongly associated with the DR specificities DRl, DR2 and DRw6 to simu- late cross-reactivity between them. We have
previously shown that DC1 molecules differ from DR molecules in papain sensitivity (Tani- gaki et al., 1980b) and in the rate of iodine uptake by the two chains (Tosi et al., 1978). More recently, we have found that not only beta chains, but also alpha chains from DC1 molecules markedly differ from those of DR molecules (Corte et al., 1981b). These findings indicate that beta chains of DC1 molecules are controlled by a second locus, closely linked to HLA-DR, which we name DC locus. The poss- ible homology between human DC molecules and murine I-A molecules has been previously discussed (Corte et al., 1981b).
The other supertypic specificity, BR4X7, is possessed by most DR4-positive or DR7-posi-
333

334 ROSA SORRENTINO <‘I al
tive individuals. It is equivalent to the Hon 7 specificity which, in Japan, is in addition as- sociated with 75 (Katagiri et al., 1979). BR4X7 resides on the beta Ia subunit. Also this determinant is interpreted as a marker of a non-DR locus (Tanigaki et ul., 198Ou), coding for Ia beta chains and in close linkage with the DR locus. The allele controlling BR4X7 has a very high linkage disequilibrium with the DR4, 7 and 75 alleles, thus simulating cross-reactivity between these DR specificities.
The present study is mainly addressed to the following question: are the two non-DR speci- ficities DC1 and BR4X7 allelic at the same locus, or are they controlled by two different loci‘? The approach we followed has been to study the peptide structure of the component chains of these two non-DR specificities. The feature revealed by the microfingerprinting
analysis supports the hypothesis of non-allelism of DC1 with BR4X7, indicating the existence of three HLA-linked loci controlling three differ- ent beta Ia subunits.
MATERIALS AND METHODS
Cell lines
The following human B-type cell lines were used: Chevalier (DR3, 7; BR4X7); U698M (DR2, 4; DCl; BR4X7): Daudi (DRw6; DCl). Typing was performed by cytotoxicity (Darke & Gay, 1974) using 7th Workshop antisera and was confirmed by radio-immunoassay (Tosi et al., 1980a).
Preparation of lahelled antigens
The purification procedure has been de- scribed in detail (Tosi et ul., 1978; Koyama et al., 1977). Briefly, crude cell membranes were isolated by differential centrifugation from about log of packed cultured cells (about 5 x lo9 cells) disrupted by nitrogen decom- pression and solubilized with Renex 30 deter- gent (Ruger Chemical Co., Irvington, NJ) in Tris-HCl buffer (Tris 0.075 M, pH 7.8). The solubilized material was separated by ultracen- trifugation and subjected to affinity chroma- tography on a column of lentil-lectin-coupled Sepharose 4B (LcH*: 4 mg/ml gel, bed volume 15 ml). The retarded fraction was eluted with a
* Abbreviations: LcH: lentil-&tin: SDS: sodium do- decyl sulfate; BSA: bovine serum albumin; CHO: carbo- hydrate; Sac: Stuph~lococcus cweus strain I.
.5’?< mannoside solution. The major portion of the retarded fraction was concentrated by ultrafiltration and fractionated by gel filtration on a column of Biogel A 1.5 M (bed volume 100 ml). The major fractions which had high Ia
activity were pooled. A small amount (0.020 A 280 units) of the pool was labelled with 1251
by the chloramine T method (Greenwood et al., 1977). The labelled preparation was fraction- ated on a column of Bio-Gel A 1.5 M. After absorption with rabbit anti-HLA (A, B, C)- coupled Sepharose (Nakamuro et al., 1975) the
material was again passed through a LcH- Sepharose column. The retarded fractions were pooled and used as antigen. The binding characteristics (high reactivity with rabbit anti- human Ia serum and absence of reactivity with anti-HLA (A, B, C), anti-beta,m and anti-Ig) have been previously described (Tosi et al., 1978; Tanigaki et al., 1980h).
Antisera
Alloantisera used were as follows (their
specificity has been described in the indicated references): DR4: 8~477 (Tosi et al., 1980b) DRw6: Fe88/37 (Tosi et ul., 1978); DR7: 8~770
(Tosi et al., 1980h); DCl: Fe131/6 (Tosi & Tanigaki, 1980~; Tosi & Tanigaki 1980b); BR4X7 in the Chevalier cell line: 8~613 (Tosi, 1980); BR4X7 in the U698M cell line: 8~1075 (Tosi, 1980).
All these antisera have been tested against
purified Ia preparations of different phenotypes in order to define their specificity by the direct binding reaction. Since alloantisera are gener- ally complex mixtures of antibodies directed against products of alleles at different loci in
‘linkage disequilibrium’, a critical point has been to choose the antisera which in a given combination can recognize a single component.
For instance, antiserum 8~613 has been found to contain an anti-DR4 as well as an anti- BR4X7 antibody component. When used with an Ia preparation from the Chevalier cell line (DR3,7; BR4X7) it can only react with BR4X7 molecules. Similar criteria have been applied to the choice of all alloantisera.
Immunoprecipitation
Each specific alloantisera was allowed to react with ‘2sI-labelled antigens for 18 hr at 20°C. Aliquots varying from 5 to 20 ~1 accord- ing to the strength of the antiserum were used for 100~1 of antigen preparation (about 5 x lo6 cpm). The complexes obtained were

Microfingerprinting Analysis of Human la 335
bound to heat-killed and formalin fixed
~taphybcoccus aureus strain I (Sac). Ten ~1 of 10% bacterial suspension were used for each /tl of antiserum. After washing, the bound pro- teins were eluted from SaC by boiling for 2 min with SDS-sample buffer (see below).
SDS polyacrylamide gel electropkoresis
SDS-PAGE was carried out in slab gel in a discontinuous Tris buffer system (Maize1 et al., 1971). The gel was then dried and exposed to Kodak X-ray film for autoradiography. Pro- teins were recovered as described (Corte et al., 1979).
Microjingerprinting analysis
Carrier BSA was added to a final concen- tration of 50&ml to the eluates recovered
from the SDS-PAGE. Complete reduction was effected with dithiothreitol (final concentration: 20mM) and was followed by alkylation with iodoacetamide (final concentration: 60 mM). Proteins were precipitated by cold 15% TCA and the pellet was digested with pepsin (1:50 enzyme to protein ratio, 18 hr at 37°C in 100 ~1
of formic acid/acetic acid/water buffer l/4/45 vol/vol/vol, pH 2.1). Two dimensional peptide maps were obtained by a previously described technique (Corte et al., 1979), based on Feinstein’s modification (Feinstein et al., 1979) of the method of Mole (1975).
Peptide analysis after CNBr digestion
Ia subunits eluted from SDSPAGE were reduced and alkylated as above and then digested with CNBr as described by Ewenstein et al. (1978), with minor modifications (24 hr at room temp. in 70% formic acid, with a 25 fold excess, w/w, of CNBr). The digested material
was lyophilized, resuspended in 30 ~1 of SDS sample buffer and run in a 15/30 SDS-gradient gel electrophoresis. The slab gel was fixed, stained, dried and exposed to Kodak X-ray film for autoradiography.
LcH-afJinity ckromutograpky
Ia subunits eluted from SDS-PAGE, either intact or digested with pepsin as above, were passed through a small column (10 x 10 mm) of LcH-coupled Sepharose, pre-washed with Tris-HCl buffer (Tris 0.075 M, pH 7.8) contain- ing 0.5 M NaCl, 0.2% Renex and 0.02% BSA.
The retarded material was eluted with the same buffer containing 5% alpha-methyl-mannoside.
RESULTS
Microjngerprinting analysis qf pepsin-digested la subunits
Starting from ’ 2 5 I-labelled purified Ia prep- arations, subsets of molecules carrying different determinants were isolated by reacting them with specific alloantisera. The component chains were then separated by SDS-PAGE, eluted and digested with pepsin. The peptic digests (about 30,OOOcpm) were subjected to two-dimensional microfingerprinting analysis. i.e. electrophoresis in the first dimension and chromatography in the second dimension, on silica gel plates. Figures 1 and 2 show the pat- terns obtained with alpha and beta subunits respectively. Each specificity exhibits an indi- vidual beta chain pattern. The BR4X7 beta chains derived from the DR4-positive cell line (U698M) and those derived from the DR7- positive cell line (Chevalier) are very similar. They markedly differ from both DR4 and DR7 beta chains. Alpha chains from the DR specifi- cities examined and from BR4X7 are similar, most of the radioactivity being concentrated in a group of five peptides (poorly resolved in the autoradiograms at the shown exposure time) of slow chromatographic migration. The residual peptides show some differences in their pos- ition and intensity. Also BR4X7 alpha chains derived from a DR7-positive cell line and a DR4-positive cell line, appear similar but not
identical. DC1 alpha chains show a completely different pattern, all the radioactivity being dis- tributed in fast-moving peptides which have no homology with the other specificities.
Luck of labelled glycopeptides in DC1 alpha chains and in beta chains
The cluster of peptides which are common to both DR and BR alpha chains but absent from DC1 alpha chains as well as from all beta chains has the characteristic features (low
chromatographic migration, diffuse spot appearance) of glycopeptides. Possible differ- ences in the distribution of radioactivity between LcH-retarded and LcH-unretarded peptides*was tested. Peptic digests of DC1 and DRw6 isolated subunits were subjected to LcH-affinity chromatography. The elution pro- files are shown in Fig. 3. While DRw6 alpha chain radioactivity is prevalently (about 60%) concentrated in the retarded fraction, the
labelled DC1 alpha chains are mostly (about 80%) unretarded. Also beta chain peptides,
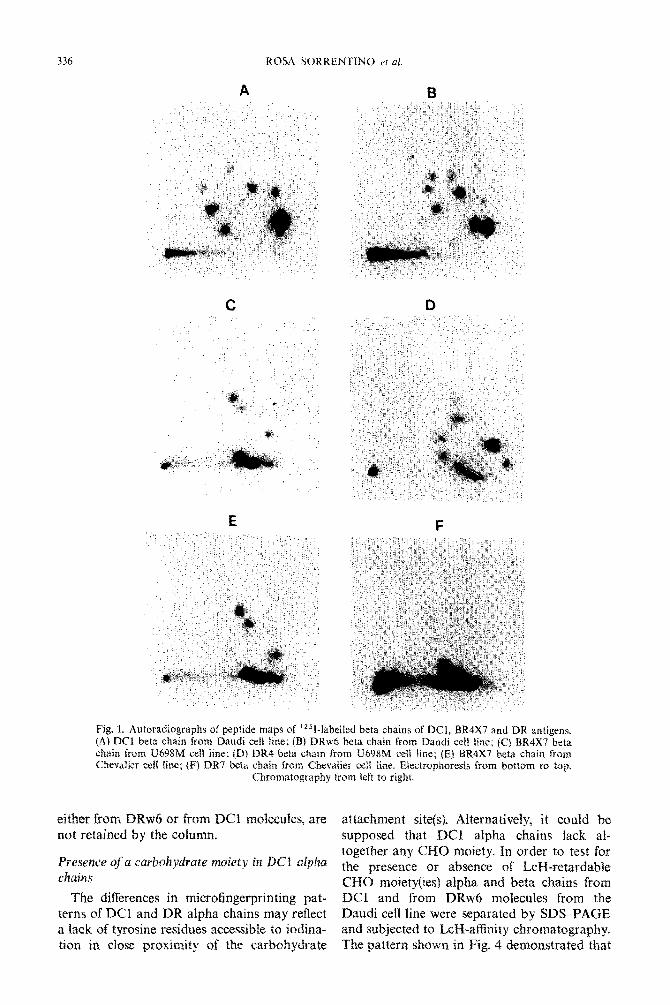
336
E
Fig. 1. Autoradiographs of peptide maps of 1251-labeXled beta chains of DCI, BR4X7 and DR antigens. (Ai DC1 beta chain from Daudi cell line; {S) DRw6 beta chain from Daudi cc11 line; (C} BR4X7 beta chain from US98M cell line; (D) DR4 beta chain from U498M ceiI Ike; jEf BRIX7 beta chain from Chevalier & tine; fF) DR7 beta chain from Chevatier ceii &ne. Etectrophoresis from bottom to top
Chromatog~p~ly from left to rig&.
eitirer from DRw6 or from DC1 molecules, are attachment site(s). Alt~rnatiye~~~ it could be not retained by the column. supposed that DCI alpha chains lack al-
Presexe ofa ~ur~~~~~r~~e ~~~j~~~ in DC1 ~1~~~~ together any CHO moiety. In order to test for
&ains the presence or absence of Lag-retardabIe CIZIO rnoiety~~es~ alpha and beta chains from
The differences in microfingerprinting pat- DCI and from DRw6 molecules from the terns of DC1 and DR alpha chains may reflect Daudi cell line were separated by SDS-PAGE a lack of tyrosine residues accessible to iodina- and subjected to L&I-affinity cbro~natograpb~* tion in cfose proximity of the carbohydrate The pattern shown in Fig. 4 demonstrated that

Microfingerprinting Analysis of Human Ia 337
E F
Fig. 2. Autoradiographs of peptide maps of ‘251-1abe11ed alpha chains of DCl, BR4X7 and DR antigens. (A) DC1 alpha chain from Daudi cell line; (B) DRw6 alpha chain from Daudi cell line; (C) BR4X7 alpha chain from U698M cell line; (D) DR4 alpha chain from U698M cell line; (E) BR4X7 alpha chain from Chevalier cell line; (F) DR7 alpha chain from Chevalier cell line. Electrophoresis from bottom to top.
Chromatography from left to right.
DC1 alpha chains are LcH retarded, thus im- plying the presence of at least one CHO side chain.
Peptide patterns after CNBr cleavage
The lack of iodine incorporation in DC1 alpha glycopeptides suggested some basic structural difference from DR (and BR) alpha chains. Further evidence of this fact was obtained by subjecting the alpha subunits to
CNBr cleavage. The rationale of this approach was that the number and the position of meth- ionine residues is generally a conserved feature in the sequence of homologous proteins. As shown in Fig. 5, the CNBr peptide patterns of DC1 and DRw6 alpha chains in gradient gel electrophoresis are markedly different. DRw6 shows three major bands of 23 K, 17 K and 13.5 K mol. wt, whereas almost all radioac- tivity of DC1 is localized in a diffuse band of
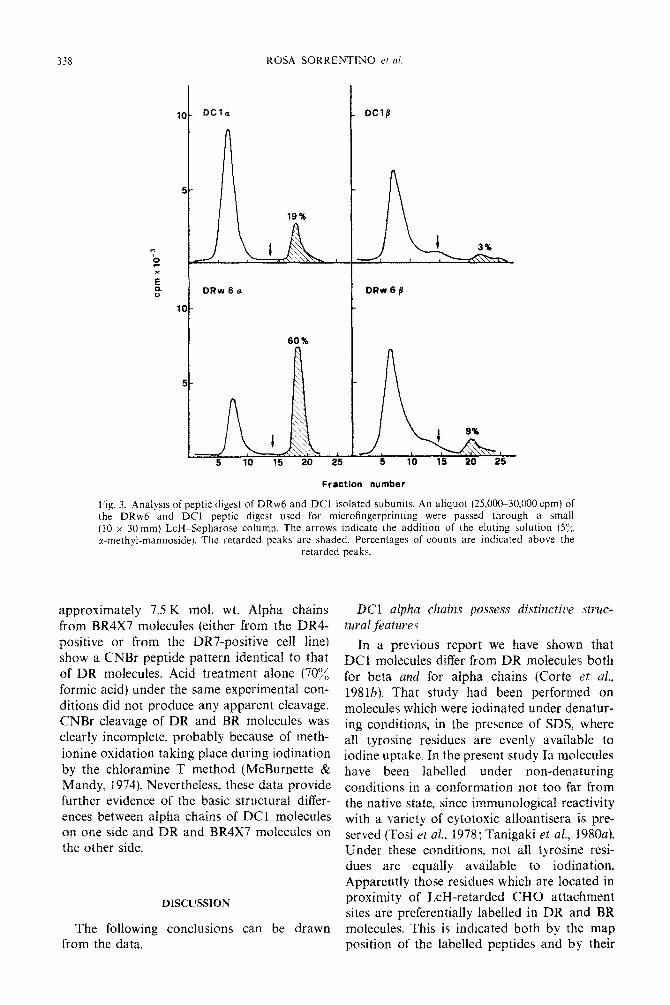
338 ROSA SORRENTINO etai.
DRw6a DRw6/3
60%
5 10 15 20 25 5 10 15 20 25
Fraction number
Fig. 3. Analysis of peptic digest of DRw6 and DC1 isolated subunits. An aliquot (25,00~30.000 cpm) of the DRw6 and DC1 peptic digest used for microfingerprinting were passed through a small (10 x 30 mm) LcH-Sepharose column. The arrows indicate the addition of the eluting solution (5’:;, r-methyl-mannoside). The retarded peaks are shaded. Percentages of counts are indicated above the
retarded peaks.
approxiInately 7.5 K mol. wt. Alpha chains from BR4X7 molecules (either from the DR4- positive or from the DR7-positive cell line) show a CNBr peptide pattern identical to that of DR molecules. Acid treatment alone (70% formic acid) under the same experimental con- ditions did not produce any apparent cleavage. CNBr cleavage of DR and BR molecules was clearly incomplete, probably because of meth- ionine oxidation taking place during iodination by the chloramine T method (McBurnette & Mandy, 1974). Nevertheless, these data provide further evidence of the basic structural differ- ences between alpha chains of DC1 molecules on one side and DR and RR4X7 molecules on the other side.
DISCUSSION
The following conclusions can be drawn from the data.
LX3 ffl~~~a chains possess distinctive struc- tural featu~e~~
In a previous report we have shown that DC1 molecules differ from DR molecules both for beta and for alpha chains (Corte et al., 1981b). That study had been performed on molecules which were iodinated under denatur- ing conditions, in the presence of SDS, where all tyrosine residues are evenly available to iodine uptake. In the present study Ia molecules have been labelled under non-denaturing conditions in a conformation not too far from the native state, since immunological reactivity with a variety of cytotoxic alloantisera is pre- served (Tosi et uE., 1978; Tanigaki et al., 1980a). Under these conditions, not all tyrosine resi- dues are equally available to iodination. Apparently those residues which are located in proximity of LcH-retarded CHO attachment sites are preferentially labelled in DR and BR molecules. This is indicated both by the map position of the labelled peptides and by their
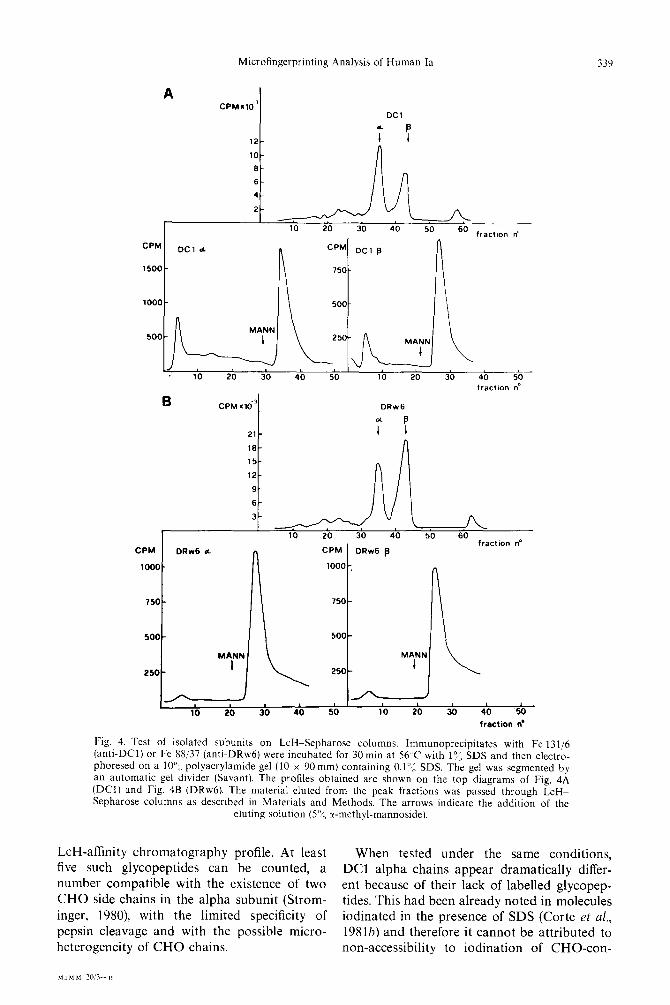
Microfingerprinting Analysis of Human la 339
A CPM.lOP
DC1
12- 7 B
lo- II
B CPMxd
21-
16-
15-
12-
9-
6-
3- A/L _^ _^ ^^ _^ __ I” Z” J” 9” 3” -+ fractKm no
CPM DRw6 d CPM DRw6 p
looO-
9 t
1000
n 750
500 I (I I MANN 1 i
‘“,-a , , , , , , , , , 10 20 30 40 50 10 20 30 40 50
fraction ne
Fig. 4. Test of isolated subunits on LcH-Sepharose columns. Immunoprecipitates with Fe 131/6 (anti-DC11 or Fe 88/37 (anti-DRwh) were incubated for 30 min at 56 C with l”:, SDS and then electro- phoresed on a lo”,, polyacrylamide gel (10 x 90 mm) containing 0.1”; SDS. The gel was segmented by an automatic gel divider (Savant). The profiles obtained are shown on the top diagrams of Fig, 4A (DCI) and Fig. 4B (DRw61. The material eluted from the peak fractions was passed through LcH- Sepharose columns as described in Materials and Methods. The arrows indicate the addition of the
eluting solution (5”); cc-methyl-mannoside).
LcH-affinity chromatography profile. At least When tested under the same conditions, five such glycopeptides can be counted, a DC1 alpha chains appear dramatically differ- number compatible with the existence of two ent because of their lack of labelled glycopep- CHO side chains in the alpha subunit (Strom- tides. This had been already noted in molecules inger, 1980) with the limited specificity of iodinated in the presence of SDS (Corte et al., pepsin cleavage and with the possible micro- 1981b) and therefore it cannot be attributed to heterogeneity of CHO chains. non-accessibility to iodination of CHO-con-
M.I M M. ?0!3 -H
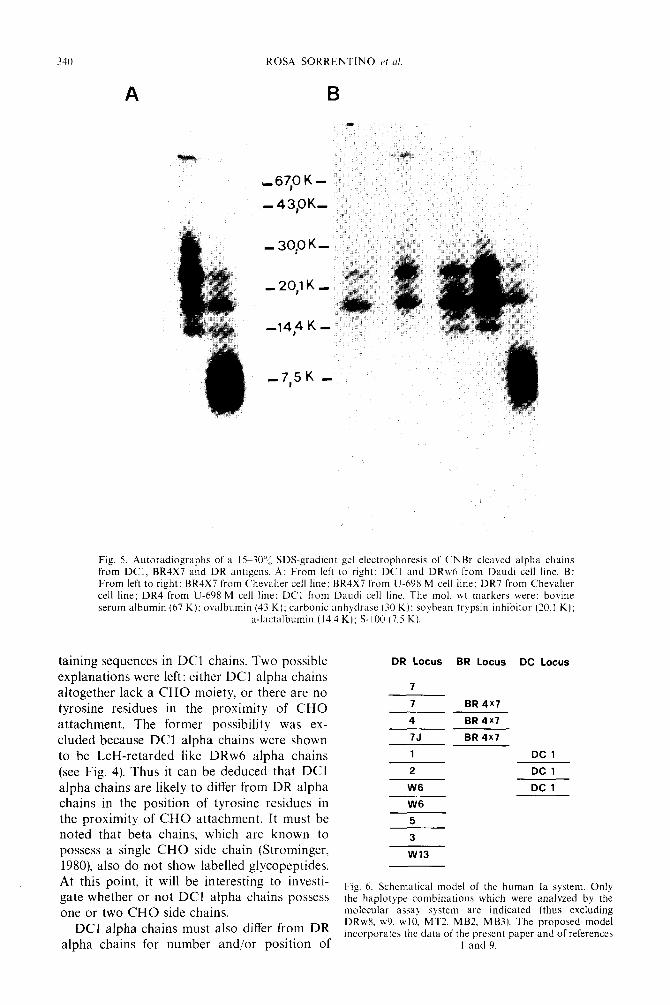
340 ROSA SORRENTINO c’f <I/
A
Fig. 5. Autoradiographs of a 15-?O’?, SDS-gradient gel electrophoresis of CNBr cleaved alpha chains from DCl. BR4X7 and DR antigens. A: From left to right: DCI and DRw6 from Daudi cell line. B: From left to right: BR4X7 from Chevalier cell line: BR4X7 from U-698 M cell line; DR7 from Chevalier cell line; DR4 from U-698 M cell line: DCI from Daudi cell line. The mol. ut markers were: bovine serum albumin (67 K): ovalbumin (43 K); carbonic anhydrasc (30 K): soybean trypsin inhibitor (20.1 K);
a-lactalbumin (14.4 K): S-100 (7.5 K).
taining sequences in DC1 chains. Two possible explanations were left: either DC1 alpha chains altogether lack a CHO moiety, or there are no tyrosine residues in the proximity of CHO attachment. The former possibility was ex- cluded because DC1 alpha chains were shown to be LcH-retarded like DRw6 alpha chains (see Fig. 4). Thus it can be deduced that DC1 alpha chains are likely to differ from DR alpha chains in the position of tyrosine residues in the proximity of CHO attachment. It must be noted that beta chains, which are known to possess a single CHO side chain (Strominger, 1980), also do not show labelled glycopeptides. At this point, it will be interesting to investi- gate whether or not DC1 alpha chains possess one or two CHO side chains.
DC1 alpha chains must also differ from DR alpha chains for number and/or position of
DR Locus BR Locus DC Locus
7
7 BR 4x1
4 BR 4x7
7J BR 4x1
1 DC 1
2 DC 1
W6 DC 1
W6
5
3
w13
Fig. 6. Schematical model of the human Ia system. Only the haplotype combinations which were analyzed by the molecular assay system are indicated (thus excluding DRwX. w9, ~10. MT2. MB2, MB3). The proposed model incorporates the data of the present paper and of references
1 and 9.

Microfingerprinting Analysis of Human la 341
methionine residues. In fact, analysis of CNBr cleavage products shows a distinctive feature in
the pattern of DC1 alpha chains, where most radioactivity is concentrated in a single diffuse band. This band is completely absent from the DR alpha chain pattern, which is composed of 3-5 bands of higher mol. wt. The DC alpha chain pattern can be attributed to an increased number of methionine residues or to their more regular spacing along the sequence, or to a lower susceptibility to oxidation by chlora- mine T, leading to a more complete cleavage. Number and position of methionine residues is generally considered as a conserved feature in homologous proteins. These differences con- firm the low degree of homology observed between DC1 and DR alpha chains peptide
maps.
DC1 and BR4X7 determinants are likely to be
controlled by non-alielic genes
DC1 and BR4X7 are non-DR specificities localized on beta Ia chains. Evidence for these facts has been previously reported (Tosi et al.,
1978: Tanigaki et al., 1980a, Tanigaki et cd., 1981) and has been confirmed by other groups (Katagiri et al., 1979, Shackelford et al., 1981). Evidence that the BR4X7 determinant is local- ized on molecules different from those carrying either the DR4 or the DR7 determinant derives from ‘sequential binding experiments’ (Tani- gaki et al., 1980a). These experiments demon- strated that by removing DR4-carrying mol-
ecules by a specific anti-DR4 alloantiserum, reactivity of anti-BR4X7 sera remained un- affected. The same results were obtained in a
DR7-positive cell line. Here the specific question of whether DC1
and BR4X7 are allelic to each other is con- sidered. Several differences can be seen between DC1 (which are known to carry the alloanti- genie determinants) and BR4X7 beta chains. Unfortunately the phenomenon of complex allelism, i.e. multiple differences between alleles, is a common finding both in human and mouse MHC genes. It is therefore difficult to decide, based on beta chain differences, whether these molecules are allelic products or not. It has been shown in the mouse that the beta chains of the two sets of Ia molecules (AB and EB chains) are associated with alpha chains which possess little or no resemblance to each other, as judged from N-terminal sequences (Cook et al., 1979). ‘A’ allospecificities and ‘E’ allospecificities, even though residing probably
on beta chains, can thus be distinguished also
for the alpha chains to which they associate. Also in man, DC1 and DR carrying beta sub- units are associated with different alpha chains as shown previously (Corte et al., 1981) and confirmed in this paper. It may be hypothe- sized that this phenomenon of differential as- sociation between Ia subunits is a basic prop- erty of the system, linked to some specific func- tions and preserved in evolution.
If we accept this generalization, it can be assumed that subunits controlled by alleles at the same locus associate with the same species of alpha subunits. Since BR4X7 alpha chains do not show any of the features described above as peculiar of DC1 alpha chains, it appears extremely unlikely that DC1 and BR4X7 beta chains are allelic if they associate with alpha chains of such drastically different structures. Even though this is not a formal proof of non-allelism, nevertheless the hypoth- esis of three beta chain loci can be considered as the most likely in the light of the present data. More firm evidence will be gained when further antigenic specificities, allelic to either DC1 or BR4X7, will be discovered. Our efforts are presently conveyed in this direction.
As apparent from Figs 2 and 4, BR4X7 alpha chains are basically similar to DR alpha chains both in their peptic maps and in their CNBr cleavage patterns. A limited degree of struc-
tural polymorphism of DR alpha chains has been previously observed (Corte et al., 1981~; Shackelford & Strominger, 1980) and it is poss- ible that this is amplified in our analysis since small conformational changes which may also depend on the association with a particular beta chain, could affect the iodination pattern. The minor differences which are observed between BR4X7 alpha chains from DR6posit- ive or DR7-positive cell line and the more marked differences between them and DR alpha chains cannot be interpreted with confi- dence. BR4X7 beta chains could associate with a separate species of alpha chains with the same basic features as DR alpha chains. Alternatively, the same alpha chain pool could be shared by DR and BR loci. A third possi- bility is that DR beta chains are associated with two kinds of alpha chains, one of which is shared by BR. Experiments performed more recently by a modified procedure including l 251-labelling under denaturing conditions strongly favour the first of these three possibili- ties (R. Sorrentino, manuscript in preparation).

342 ROSA SORRENTINO er ul.
Multiple linkage disequilibrium
Our present views on the Ia system are sum- marized in the model shown in Fig. 6, which incorporates the proposed assignment of the two supertypic specificities DC1 and BR4X7 to different loci. The model implies multiple link- age disequilibrium by which a single allele at one locus (e.g. DCl) is strongly associated with more than one allele at another locus (e.g. DRl, DR2, DRw6).
However, another interpretation based on single linkage disequilibria is possible and it has been indeed proposed by Shackelford et al. (1981) for the DC1 specificity on the basis of isoelectric point differences. In the present report, beta chains from BR4X7 molecules gave very similar patterns whether they were isolated from a DRCpositive or from a DR7- positive cell line (see Fig. 1). This suggests a multiple linkage disequilibrium relationship between BR4X7 on one side and DR4 and DR7 on the other side. However, microfinger- printing performed under different conditions, e.g. cleavage by different proteolytic enzymes, could reveal other structural variations. Our present opinion is that the multiple linkage disequilibrium hypothesis must be tested case by case with all available analytical tools, with- out a preconceptual commitment to a general model.
Acknor~led~ement-The authors wish to thank MS Dora Centis for technical assistance.
REFERENCES
Calissano P.. Mercanti D. & Levi A. (1976) Ca’*, K+-regulated intramolecular crosslinking of S-100 pro- tein via disulfide bond formation. Eur. .I. Biochrrn. 71, 45-52.
Cook R. G., Siegelman M. H., Capra J. D.. Uhr J. W. & Vitetta E. S. (1979) Structural studies on the murine la alloantigen. IV. NH,-terminal sequence analysis of alle- lit products of the JA and JEjC subregion. J. Imtnun. 122, 2232-2237.
Corte G., Damiani G.. Calabi F., Fabbi M. & Bargellesi A. (1981~) Analysis of HLA-DR polymorphism by two dimensional peptide mapping. Proc. nafn. Acud. Sci. U.S.A. 78, 53&538.
Corte G., Calabi F., Damiani G.. Bargellesi A., Tosi R. & Sorrentino R. (1981h) Human Ia molecules carrying DCI determinants differ in both r and p subunits from la molecules carrving DR determinants. Nature. Lond. 292, 357-360. . -
Corte G.. Tonda P., Cosulich E.. Milstein C. P.. Bargellesi A. & Ferrarini M. (1979) Characterization of IgD. !. Iso- lation of two molecular forms from human serum. Scant/. J. Inl!?lun. 9, 141 149.
Darke C. & Gay E. (1974) An evaluation of ethidium bro- mide microcytotoxicity test. J. Zmmun. Meth. 4, 161-168.
Delovitch T. L. & Falk J. A. (1979) Evidence for structural homology between murine and human la antigens. Im- munagenrtict 8, 405-418.
Ewenstein B. M., Nisizawa T., Uehara H., Nathenson J. G., Coligan J. E. & Kindt T. J. (1978) Primary struc- ture of murine major histocompatibility complex alloan- tigens: isolation. biochemical characterization and pre- Ii&nary alignment of CNBr fragments from the H-2Kb plvconrotein. Proc. rluttl. Acud. Sci. U.S.A. 75, 2909-29 13.
Feinstein A., Richardson N. E. & McIlhinney R. A. J. (1979) Structure of lymphocyte membrane immuno- globulin. In Cells of Imrnur~oglohuli~~ Synthesis. (Edited by Pernis B. & Vogel F.) pp. 165-188.
Greenwood F. C.. Hunter W. M. & Glaver J. S. (1977) The preparation of ‘251-labelled human growth hormone of hiah specific radioactivitv. Biochrm. J. 89. 114123.
Katagiri’M.. lkeda H.. Maruyama N., Mariuchi J., Waki- saka A.. Kimura S., Aizawa M. & Itakura K. (1979) Evidence for two B-cell alloantigen loci in the HLA-D region. Il,ln2llnoyenefi~s 9, 335-352.
Koyama K., Nakamuro K., Tanigaki N. & Pressman D. (1977) Alloantigenic activity and cell line, organ and tissue distribution as determined by radioimmunoassay. Ii?lnzunolog~, 33, 217-230.
McBurnette S. K. & Mandy W. J. (1974) Chemical modifi- cation of a rabbit immunoglobulin allotypic specificity. I/n,nunochrmivtr~, 11, 255-260.
McMillan M., Cecka J. M.. Murphy D. B., Mcdevitt H. 0. & Hood L. (1977) Structure of murine la antigens: par- tial NH,-termmal amino acid sequences of products of the I-E or I-C subregion. Proc. WXH. Acad. Sci. U.S.A. 74,5135-5141.
Maize1 J. V. (1971) Polyacrylamide gel electrophoresis of viral proteins. In Method\ in Virologr. (Edited by Mara- morosch K. & Koprowski H.), Vol. 5. pp. 179-187. Aca- demic Press, New York.
Mole L. E. (1975) A genetic marker in the variable region of rabbit immunoglobulin heavy chain. Biochem. J. 151, 351-357.
Nakamuro K., Tanigaki N. & Pressman D. (1975) Com- mon antigenic structures of HLA antigens. VI. Common antigenic determinants located in the 33,OOOmol. wt alloantigenic fragment portion of papain-solubilized HLA molecules. ~wnuno~yr. 29, 11 19.m 1 126.
Shackelford D. A. & Strominger J. L. (19801 Demon- stration of structural polym&phism among HLA-DR light chains by two dimensional gel electrophoresis. J. exp. Med. 151, 144-165.
Shackclford D. A.. Strominger J. L., Mann D. L.. van Rood J. J. & Ferrara G. B. (1981) Biochemical characterization of the human B all alloantigen DC1 (MTI. LB12) and comparison to the HLA-DR antigen. Proc. nut/l. Acad. Sci. U.S.A. 78, 4566~4571.
Silver J. & Ferrone S. (1979) Structural polymorphism of human DR antigens. Nature Land. 279, 43G437.
Strominger J. L. (1980) Structure of products of the major histocompatibility complex in man and mouse. In Immu- noloyy X0 (Edited by Fougereau M. & Dausset J.). pp. 541-554. Academic Press, New York.
Tanigaki N., Tosi R.. Centis D., Ferrara G. B. & Pressman D. (1981) Further characterization of two DR supertypic specificities: additional evidence that they reside on mol- ecules different from those carrying HLA-DR locus spe- cificities. Human Immuno/og>~ 3, 93-108.
Tanigaki N., Tosi R., Koyama K. & Pressman D. (1980~1) Purification and separation of subsets of human Ia molecules by papain digestion. Im~nu~~oloy~ 3% 615-622.
Tanigaki N.. Tosi R., Pressman D. & Ferrara G. B. (1980h) Molecular identification of human la antigens coded by a gene locus closely linked to HLA-DR locus. Iwiuno- ~ywtic\ 10, 151-167.

Microfingerprinting Analysis of Human Ia 343
Tosi R. (1980) Joint report: MT3. In Mi.stocompatihilitt Testing (Edited by Terasaki P. I.), p. 568. UCLA.
Tosi R. & Tanigaki N. (1980a) 8th Workshop antisera pos- sessing anti-DC1 activity. In Hi~toconzparihilirr Testing (Edited by Terasaki P. I.), p. 917. UCLA.
Tosi R. & Tanigaki N. (1980b) Joint Report: MT1. In Nis- foco~}~~acib~~if~ Testing (Edited by Terasaki P. I.). pp. 558-563. UCLA.
Tosi R., Tanigaki N., Centis D., Ferrara G. B. & Pressman
D. (1978) Immununological dissection of human Ia mol- ecules. J. exp. Med. 148, 1592-1611.
Tosi R., Tanigaki N.. Centis D., Rossi P. L., Alfano G., Ferrara G. B. & Pressman D. (1980~) HLA-DR typing by radioimmunoassay. 7imsphtation 29, 302-305.
Tosi R., Tanigaki N. & Centis D. (1980b) Analysis at mol- ecular level of the specificity of 8th Workshop antisera. In Hisrocompafihiiir?. ‘J’estDzg (Edited by Terasaki P. I.), pp. 9033905. UCLA.










![Colec]ia PLURAL](https://static.fdokumen.com/doc/165x107/63165d23c32ab5e46f0dbd8a/colecia-plural.jpg)









