
Zinc Finger Protein 467 Is a Novel Regulator of Osteoblast and Adipocyte Commitment
Upload
independentCategory
view
0download
0
Pharmacological Research 52 (2005) 475–484
Methylamine but not mafenide mimics insulin-like activity of thesemicarbazide-sensitive amine oxidase-substrate benzylamine
on glucose tolerance and on human adipocyte metabolism
Maria Carmen Iglesias-Osmaa, Sandy Bourb, Maria Jose Garcia-Barradoa, Virgile Visentinb,Maria Francisca Pastora, Xavier Testarc, Luc Martic, Gemma Enrique-Taranconc,
Philippe Valetb, Julio Moratinosa, Christian Carpeneb,∗a Departamento de Fisiologıa y Farmacologıa, Facultad de Medicina, Universidad de Salamanca, 37007 Salamanca, Spain
b INSERM (Institut National de la Sante et de la Recherche Medicale), U586 Universite Paul Sabatier, Unite de recherches sur les obesites IFR31,Centre Hospitalier Universitaire de Toulouse (CHU Rangueil), F-31432 Toulouse, France
c Facultat de Biologia, Departament de Bioquimica i Biologia Molecular, Universitat de Barcelona, Avda. Diagonal 645, 08028 Barcelona, Spain
Accepted 29 July 2005
use, rabbit,diabetic rats.icarbazide-l antidiabetictly reportedubstrate. Allt mafenide,lin-releasingas not aould not bey contrast,
vel SSAO
ito-theasesevenine is
itant
enicentlyion
Abstract
It has been reported that benzylamine reduces blood glucose in rabbits, stimulates hexose uptake, and inhibits lipolysis in moand human adipocytes. In the presence of vanadate, benzylamine is also able to improve glucose disposal in normoglycaemic andSuch insulin-mimicking properties are the consequence of hydrogen peroxide production during benzylamine oxidation by semsensitive amine oxidase (SSAO). The aim of the study was to determine whether other SSAO-substrates could share such potentiaproperties. Thus, mafenide, a synthetic antimicrobial sulfonamide structurally related to benzylamine, and which has been recento interact with SSAO, was tested in the above mentioned models, in parallel with methylamine, a proposed endogenous SSAO-stested amines stimulated glucose uptake and inhibited lipolysis in rat and mouse fat cells. Methylamine and benzylamine, but noreduced the hyperglycaemic response during a glucose tolerance test in rabbits while the three amines tested were devoid of insuactivity under both in vivo and in vitro conditions. In human adipocytes, mafenide did not stimulate glucose transport since it whigh-affinity substrate for SSAO and generated less hydrogen peroxide than benzylamine or methylamine. Therefore, mafenide cconsidered as an antidiabetic drug despite being oxidized and exhibiting insulin-mimicking effects in rat and mouse adipocytes. Bthe endogenous substrate methylamine improved glucose utilization in all in vitro and in vivo models, leading to consider nosubstrates as drugs with potential anti-hyperglycaemic properties.© 2005 Published by Elsevier Ltd.
Keywords: Glucose homeostasis; Insulin-like activity; Lipolysis; Semicarbazide-sensitive amine oxidase
1. Introduction
Besides its involvement in lipid storage, white adipose tis-sue (WAT) is now recognized as an endocrine organ whichis involved in the regulation of glucose homeostasis[1,2].WAT pathophysiology is related to obesity, diabetes melli-tus, metabolic syndrome, and associated complications[3,4].Another function of WAT remains poorly known: its capacity
∗ Corresponding author. Tel.: +33 5 62 17 29 55; fax: +33 5 61 33 17 21.E-mail address: [email protected] (C. Carpene).
to oxidize biogenic or exogenous amines. In fact, the mchondrial monoamine oxidases (MAO, EC 1.4.3.4.) andcopper-containing semicarbazide-sensitive amine oxid(SSAO, EC 1.4.3.6.) are highly expressed in adipocytes,in human beings[5–7]. Both amine oxidases catalyze,the presence of oxygen, a reaction by which an aminconverted into its corresponding aldehyde with concomproduction of hydrogen peroxide and ammonia[8]. AlthoughSSAO shares the classical scavenger function of biogamines with MAO, this ecto-enzyme possesses other recreported functions[9]. SSAO, also called vascular adhes
1043-6618/$ – see front matter © 2005 Published by Elsevier Ltd.doi:10.1016/j.phrs.2005.07.008

476 M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484
Fig. 1. Structure of the amines tested in in vitro and in vivo experiments.
protein-1 (VAP-1), is present in endothelial cells, particu-larly of lymphatic tissues where it contributes to lymphocyteadhesion to vascular walls[9]. In vascular smooth musclecells, SSAO also interferes with extracellular matrix devel-opment[10]. More interestingly, the abundant SSAO activitypresent in adipocytes[5,6] interacts with the regulation ofglucose metabolism since benzylamine, a SSAO substrateof reference, has been reported to activate glucose uptakeinto fat cells [11]. Generation of hydrogen peroxide dur-ing SSAO-catalyzed amine oxidation is a key element inthe benzylamine-induced stimulation of glucose utilization[11,12]. The combination of benzylamine plus vanadate pro-motes translocation of glucose transporters to the cell sur-face and stimulates glucose transport in rat fat cells up to80% of the maximal insulin effect[11–13]. In vivo, theassociation benzylamine-vanadate leads to an enhanced glucose tolerance and a reduced hyperglycaemia in diabetic rats[14,15]. Moreover, it has been reported that benzylamineon its own was able to stimulate hexose uptake in human[16], mouse or rabbit adipocytes[17]. Benzylamine alonealso exerted in vivo insulin mimicry: its infusion in rab-bits clearly limited the rise in blood glucose provoked dur-ing an intravenous glucose tolerance test (IVGTT) withoutmodifying the insulin secretory pattern[17]. These obser-vations together with the capacity of benzylamine to inhibitlipolysis in human and rodent adipocytes[16–18] led us tos ates.I pha-a ei ppli-c fore,t fullyr ne,a oralt
rma-c sub-s tess en-z ide;F branep d our
interest in this drug, also endowed with a sulfonamide struc-ture [20], and thus with a theoretical potential as an insulinsecretagogue. We have also explored the insulin-like effectsof methylamine (Fig. 1), an endogenous amine derived fromseveral metabolic processes which is smaller than benzy-lamine but which has been defined as a physiological sub-strate of SSAO[21].
In this work, we first verified whether benzylamine,mafenide and methylamine could act in vitro as insulin-mimicking agents – with and without vanadate – by acti-vating hexose uptake or by inhibiting lipolysis in rodentadipocytes. Then, the compounds were compared in vivo onIVGTT in conscious rabbits. We also evaluated whether thethree amines were oxidized and produced hydrogen peroxidewhen incubated with WAT homogenates. Finally, we inves-tigated their capacity to activate glucose transport in humanadipocytes and to undergo SSAO-dependent oxidation.
The following results describe the in vitro and in vivoinsulin-mimicking effects of the tested amines. In rabbits,benzylamine and methylamine, but not mafenide, exhib-ited an antihyperglycaemic profile without increasing insulinsecretion. Although mafenide was readily oxidized andexhibited promising in vitro insulin-mimicking effects inrodent adipocytes, it was unable to stimulate hexose uptakein human adipocytes, probably as a consequence of its lowaffinity for human amine oxidases.
2
2
c ces( mi-c fate,s vines dase,a icalsw ises rs fort cheD
2
Isle,F .5 kg( ed;m erea ow ina irec-t andb inc werep s and
tudy the potential therapeutic interest of SSAO substrndeed, benzylamine is a relatively simple molecule (almino-toluene;Fig. 1) which is better known for its us
n chemical organic synthesis than for therapeutic aations. The objective of the present work was, thereo evaluate whether other primary amines could useeproduce the insulin-mimicking activity of benzylamind could exhibit improved properties for antidiabetic
reatment.Recently, efforts have been made to identify the pha
ophoric characteristics that modulate the recognition oftrates by SSAO[19]. Among a group of potential substracreened on the basis of a structural similarity with bylamine, mafenide (4-aminomethyl-benzenesulfonamig. 1) was found to behave as a SSAO substrate in memreparations from adipose tissue. Therefore, we focuse
-. Materials and methods
.1. Drugs and reagents
2-[1,2-3H]Deoxyglucose (2-DG, 26 Ci mmol−1) was pur-hased from Perkin-Elmer Life and Analytical ScienBoston, MA). Benzylamine, methylamine, mafenide, searbazide (all hydrochloride salts), phenelzine sulodium orthovanadate, cytochalasin B, fatty acid-free boerum albumin, hydrogen peroxide, horseradish peroxintiproteases cocktail, and most commonly used chemere from Sigma–Aldrich (St. Louis, MO), unless otherwpecified. Liberase, collagenase, enzymes and cofactohe determination of glycerol and glucose were from Roiagnostics (Mannheim, Germany).
.2. Animals, subjects and tissue sampling
Male Wistar rats of 300–400 g (Janvier, Le Genest St.rance) and male New Zealand White rabbits of 2.8–3Charles River, Madrid, Spain) were individually housice from the C57BL/6J strain (25–30 g) of both sexes wlso used. All the animals had free access to water and chccordance with the European Communities Council D
ives for experimental animal care. Drug administrationlood sampling necessary for IVGTT were performedonscious fasted rabbits, whereas in vitro experimentserformed with tissues removed from euthanized animal

M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484 477
immediately used for amine oxidase activity or hydrogen per-oxide determination, or even for fat cell or pancreatic isletisolation and subsequent determinations of lipolysis, glucosetransport, or insulin secretion.
Samples of human subcutaneous adipose tissue wereobtained from 24 non-obese women undergoing abdom-inal lipectomy in the Department of Plastic Surgery ofRangueil Hospital (mean body mass index was 25.4± 0.7and age ranged from 28 to 45 years) under approval by theEthical Committee of Toulouse University Hospital. Sam-ples of human adipose tissue were transferred in less than15 min from surgery to the lab in which they were sub-jected to digestion for adipocyte isolation or stored at−80◦Cuntil preparation of homogenates for amine oxidase activityassays.
2.3. Adipocyte isolation and phosphatase activitydetermination
Epididymal and perirenal white adipose tissues fromrats or mice and human subcutaneous WAT samples wereminced in Krebs-Ringer containing 15 mM sodium bicar-bonate, 10 mM HEPES, 6 mM glucose and bovine serumalbumin (35 mg ml−1) (KRBH buffer, pH 7.4). Adipose tis-sue was digested for 35–45 min at 37◦C with 40–60�g ml−1
Liberase blendzyme 3 (Roche Diagnostics, Mannheim, Ger-m imest -tv e dis-t trate6 n afi s-pr hekT ularP
2m
ol-u edi itht wasag -v cells prese ionso2 ationo1 erec late( tes
from the buffer and to count the radioactive intracellular 2-DG as described previously[22].
2.5. Intravenous glucose tolerance tests (IVGTT)
The experimental design for IVGTT was carried out inconscious rabbits after 24 h fasting as already described[17].Briefly, arterial blood was sampled by means of an indwellingcannula placed in the central artery of one ear. Two con-trol samples were taken before drug infusion. Drug solutionswere infused at a constant rate (0.15 ml min−1) for 30 minthrough an indwelling cannula in the marginal vein of thecontralateral ear. Plasma glucose was estimated by means ofthe glucose oxidase procedure while immunoreactive insulin(IRI) was determined by using a radioimmunoassay kit (CISRadioquimica-Schering, Madrid, Spain).
2.6. In vitro insulin release
Islets were isolated by collagenase digestion of rabbitor mouse pancreas. After isolation, islets were first pre-incubated for 1 h at 37◦C in a bicarbonate buffer medium,pH 7.4, containing 15 mM glucose and supplemented with1 mg ml−1 bovine serum albumin. They were then incubatedfor 60 min in 1 ml of medium containing appropriate con-centrations of glucose and tested substances as detailed[23].A wasw ove.
2h
tricm -x im-M lyd ar-a ed ina Lab-s siona , thefl pt d atd erep T in1v ase-d d outihp gy-l Ooch s-c d at
any). Adipocyte preparations were washed three then distributed and incubated at 37◦C for 30 min in plasic tubes containing the tested compounds in 400�l finalolume. Then, the cells were sonicated and aliquots werributed into 96-well plates containing the synthetic subs,8-difluoro-4-methylumbelliferyl phosphate (DiFMUP) inal volume of 100�l of appropriate reaction buffer. Phohatase activity was assessed after 60 min at 37◦C by fluo-ometric measurement following the RediPlate 96 EnzCyrosine Phosphate Assay Kit recommendations (Molecrobes/Interchim).
.4. Lipolysis and deoxyglucose transporteasurements in isolated adipocytes
Isolated fat cells were diluted in around 10-fold their vme of KRBH, and 500�l of cell suspension was distribut
nto plastic incubation vials. After a 90-min incubation whe tested drugs, the glycerol released into the mediumssayed and lipolytic activity was expressed as�moles oflycerol released 100 mg−1 cellular lipid 90 min−1, as preiously described[18]. For hexose uptake assays, fatuspensions were prepared as described above in thence of 2 mM pyruvate instead of 6 mM glucose. Incubatf the tested drugs lasted 45 min at 37◦C. Then, 0.4�Ci of-deoxyglucose (2-DG) were added at a final concentrf 0.1 mM for 10 min. Assays were stopped with 100�l of00�M cytochalasin B, and aliquots of cell suspension wentrifuged in microtubes containing di-isononyl phthadensity 0.97 g ml−1), which allowed to separate adipocy
-
t the end of the incubation, a portion of the mediumithdrawn and diluted for insulin assay as described ab
.7. Generation of hydrogen peroxide by adipose tissueomogenates and amine oxidase activity
Hydrogen peroxide was detected with a fluoromeethod using the probe,N-acetyl-3,7,-dihydroxypheno
azine, commercially available as Amplex Red (Intercholecular Probes, Montluc¸on, France), as previousescribed[9]. Amplex Red oxidation resulted in the appence of the fluorescent dye resorufin which was detectfluorescence microplate reader (Fluoroskan Ascent,
ystems, Helsinki, Finland) set for excitation and emist 544 and 590 nm, respectively. Under these conditionsuorescence increased linearly with H2O2 concentration uo 10�M, then reached a plateau and was even inhibiteoses higher than 100�M. Adipose tissue homogenates wrepared by mixing 100 mg of rat, mouse or human WAml of phosphate buffer 200 mM (Na2HPO4, NaH2PO4, 4/1 v, pH 7.4) with antiprotease cocktail. The amine oxidependent generation of hydrogen peroxide was carrie
n 96-well black microplates, by adding 50�l of tissueomogenate to 50�l of the tested substrates and 50�l ofhosphate buffer with or without enzyme inhibitor (par
ine 0.5 mM or semicarbazide 1 mM, for inhibiting MAr SSAO, respectively). After 10 min at 37◦C, 50�l of thehromogenic mixture containing Amplex Red (100�M) plusorseradish peroxidase (1 U ml−1) was added and fluoreence was continuously monitored on a 30-min perio

478 M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484
37◦C. After protein determination using Bio-Rad DC Pro-tein Assay (CA, USA), results were expressed in nanomolesof hydrogen peroxide produced min−1 mg−1 protein.
Measurement of amine oxidase activity was also con-ducted on WAT homogenates with the radiochemical methodusing [14C]-benzylamine (57 mCi mmol−1) from AmershamBiosciences (Buckinghamshire, UK) as previously reported[16]. Oxidation lasted 15 min at 37◦C when the homogenateswere added to isotopic dilution of benzylamine at 0.5 mMtogether with the indicated final concentration of the testedcompounds.
2.8. Statistical analysis
Results are given as mean± S.E.M. Statistical signifi-cance was assessed by use of Student’st-test. For in vivoexperiments, the one-way analysis of variance (ANOVA) cor-rected by the Newman–Keuls test was used.
3. Results
3.1. Hydrogen peroxide-dependent effects ofbenzylamine in adipocytes
It was first verified whether inhibition of protein-tyrosinep e 3-k oseu mana ep inb amed hos-p timu-l d byw ntp rs ofi ibi-t pro-d bya anda lead-i ake.S ulina beend ru e andm sis.
3a
tedg date,w edg syn-
Table 1Tyrosine phosphatase activity and hexose transport in benzylamine-treatedrat and human fat cells
Addition to medium Tyrosine-phosphatase activity(percentage of basal activity)
Rat fat cells Human fat cells
Insulin, 100 nM 99± 25 79± 10Hydrogen peroxide, 1 mM 61± 6** 54 ± 6***
Benzylamine, 1 mM 72± 9* 61 ± 4***
Vanadate, 1 mM 12± 4*** 2 ± 2***
Addition to benzylamine,0.1 mM
Hexose transport (percentageof benzylamine-stimulated)
Rat fat cells Human fat cells
Wortmannin, 10 nM 77± 5* 66 ± 18Wortmannin, 100 nM 20± 3** 47 ± 9**
Wortmannin, 1�M 17 ± 4** 10 ± 15**
Phosphatase activity is expressed as percentage of basal which wasequivalent to 480± 45 nmol and 81± 14 nmol of phosphate released per100 mg lipid during 60 min for rat and human preparations, respectively.Hexose transport is expressed as percentage of 0.1 mM benzylamine-dependent stimulation, which increased from 1.07± 0.19 nmol (unstim-ulated) to 5.66± 0.38 nmol of 2-DG 10 min−1 100 mg lipid−1 in rat fatcells (in the presence of 0.1 mM vanadate,n = 3, p < 0.001). Benzylamine0.1 mM also increased 2-DG uptake from 0.79± 0.10 nmol (basal) to1.57± 0.31 nmol 10 min−1 100 mg lipid−1 in human adipocytes (n = 6,p < 0.01). Means± S.E.M. different from control set at 100%.
* p < 0.05.** p < 0.01.
*** p < 0.001.
ergism with vanadate but not the two other end-products ofbenzylamine oxidation, benzaldehyde and ammonia, evenwhen tested at 1 mM (Fig. 2A). Methylamine and mafenideweakly activated glucose uptake when tested alone: at 1 mM,their action represented less than 20% of the maximal effectof insulin (not shown). However, when tested at 1 mM in thepresence of vanadate, the amines stimulated uptake as highas 100 nM insulin (Fig. 2B). When mafenide was tested at0.1 mM in combination with vanadate, it hardly reproduced20% of the insulin stimulation, while 0.1 mM methylamineor benzylamine plus vanadate mimicked more than 80% ofthe insulin activation of hexose uptake. The three aminesalso partially reproduced the antilipolytic effect of insulin:at 0.1–1 mM, they inhibited the lipolytic response evoked byisoproterenol, a�-adrenergic receptor agonist, even withoutvanadate (Fig. 2C).
3.3. Effects on hexose uptake and lipolysis in mouse fatcells
The effects of the three amines on hexose transport inmurine adipocytes are shown inFig. 3A. As recently reported,benzylamine was able by its own to activate glucose uptake inthis model, without any addition of vanadate[17]. Mafenide1 mM was less efficient than benzylamine or methylamine.The stimulatory action of the amines was enhanced by0 was
hosphatase activity and activation of phosphoinositidinase activity were involved in the stimulation of hexptake by benzylamine previously reported in rat and hudipocytes[11,12,16]. Table 1shows that 1 mM benzylaminartially inhibited protein-tyrosine phosphatase activityoth species, as did 1 mM hydrogen peroxide. At the sose, sodium orthovanadate completely abolished phatase activity. Moreover, benzylamine-dependent s
ation of hexose uptake was dose-dependently inhibiteortmaninn in both species (Table 1). Since thiol-dependerotein-tyrosine phosphatases are negative regulato
nsulin signalling, these observations indicated that inhion of phosphatase activity by the hydrogen peroxideuced during benzylamine oxidation was likely followedn increased ratio of tyrosine-phosphorylated proteinssubsequent activation of phosphoinositide 3-kinase,
ng to the observed insulin-like stimulation of hexose uptince the mechanisms by which benzylamine mimics insction and recruits glucose transporters have alreadyescribed in rat adipocytes[11,12], this model was furthesed to compare the capacity of benzylamine, mafenidethylamine to activate hexose uptake or to inhibit lipoly
.2. Effects on glucose uptake and lipolysis in ratdipocytes
Fig. 2A shows that 0.1 mM benzylamine clearly activalucose uptake only in the presence of 0.1 mM vanahich by itself did not modify basal or insulin-stimulatlucose uptake. Hydrogen peroxide exhibited a similar
.1 mM vanadate as in rat fat cells. Again, mafenide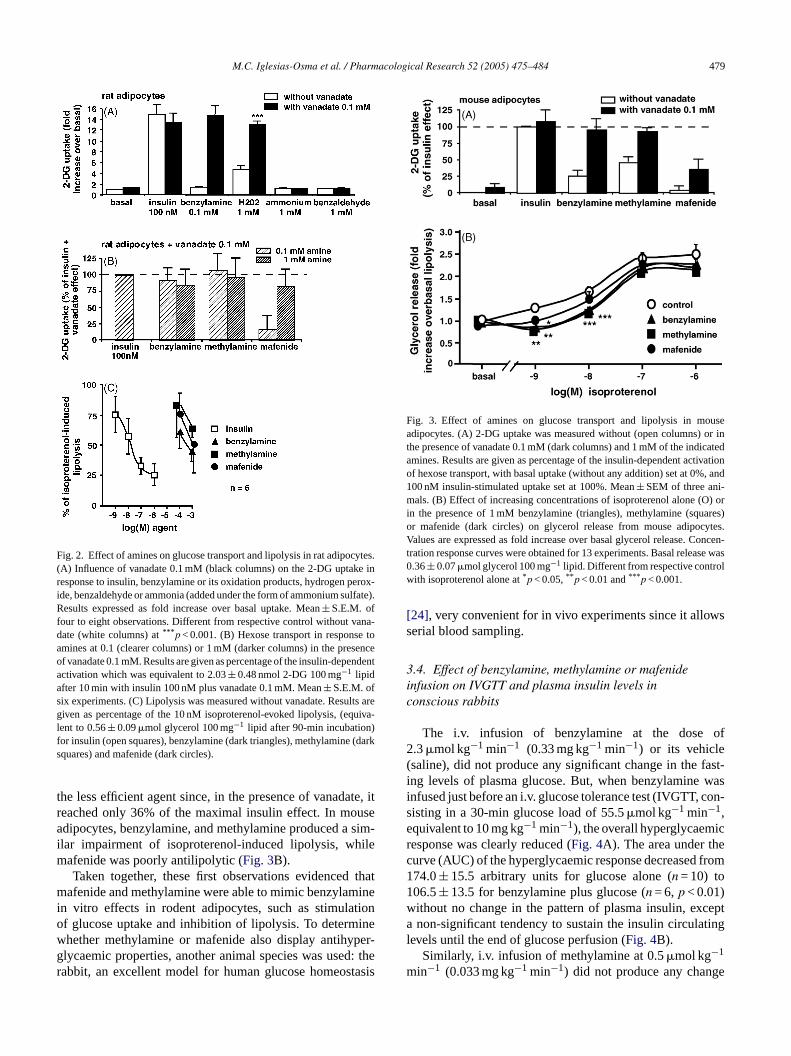
M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484 479
Fig. 2. Effect of amines on glucose transport and lipolysis in rat adipocytes.(A) Influence of vanadate 0.1 mM (black columns) on the 2-DG uptake inresponse to insulin, benzylamine or its oxidation products, hydrogen perox-ide, benzaldehyde or ammonia (added under the form of ammonium sulfate).Results expressed as fold increase over basal uptake. Mean± S.E.M. offour to eight observations. Different from respective control without vana-date (white columns) at*** p < 0.001. (B) Hexose transport in response toamines at 0.1 (clearer columns) or 1 mM (darker columns) in the presenceof vanadate 0.1 mM. Results are given as percentage of the insulin-dependentactivation which was equivalent to 2.03± 0.48 nmol 2-DG 100 mg−1 lipidafter 10 min with insulin 100 nM plus vanadate 0.1 mM. Mean± S.E.M. ofsix experiments. (C) Lipolysis was measured without vanadate. Results aregiven as percentage of the 10 nM isoproterenol-evoked lipolysis, (equiva-lent to 0.56± 0.09�mol glycerol 100 mg−1 lipid after 90-min incubation)for insulin (open squares), benzylamine (dark triangles), methylamine (darksquares) and mafenide (dark circles).
the less efficient agent since, in the presence of vanadate, itreached only 36% of the maximal insulin effect. In mouseadipocytes, benzylamine, and methylamine produced a sim-ilar impairment of isoproterenol-induced lipolysis, whilemafenide was poorly antilipolytic (Fig. 3B).
Taken together, these first observations evidenced thatmafenide and methylamine were able to mimic benzylaminein vitro effects in rodent adipocytes, such as stimulationof glucose uptake and inhibition of lipolysis. To determinewhether methylamine or mafenide also display antihyper-glycaemic properties, another animal species was used: therabbit, an excellent model for human glucose homeostasis
Fig. 3. Effect of amines on glucose transport and lipolysis in mouseadipocytes. (A) 2-DG uptake was measured without (open columns) or inthe presence of vanadate 0.1 mM (dark columns) and 1 mM of the indicatedamines. Results are given as percentage of the insulin-dependent activationof hexose transport, with basal uptake (without any addition) set at 0%, and100 nM insulin-stimulated uptake set at 100%. Mean± SEM of three ani-mals. (B) Effect of increasing concentrations of isoproterenol alone (O) orin the presence of 1 mM benzylamine (triangles), methylamine (squares)or mafenide (dark circles) on glycerol release from mouse adipocytes.Values are expressed as fold increase over basal glycerol release. Concen-tration response curves were obtained for 13 experiments. Basal release was0.36± 0.07�mol glycerol 100 mg−1 lipid. Different from respective controlwith isoproterenol alone at* p < 0.05,** p < 0.01 and*** p < 0.001.
[24], very convenient for in vivo experiments since it allowsserial blood sampling.
3.4. Effect of benzylamine, methylamine or mafenideinfusion on IVGTT and plasma insulin levels inconscious rabbits
The i.v. infusion of benzylamine at the dose of2.3�mol kg−1 min−1 (0.33 mg kg−1 min−1) or its vehicle(saline), did not produce any significant change in the fast-ing levels of plasma glucose. But, when benzylamine wasinfused just before an i.v. glucose tolerance test (IVGTT, con-sisting in a 30-min glucose load of 55.5�mol kg−1 min−1,equivalent to 10 mg kg−1 min−1), the overall hyperglycaemicresponse was clearly reduced (Fig. 4A). The area under thecurve (AUC) of the hyperglycaemic response decreased from174.0± 15.5 arbitrary units for glucose alone (n = 10) to106.5± 13.5 for benzylamine plus glucose (n = 6, p < 0.01)without no change in the pattern of plasma insulin, excepta non-significant tendency to sustain the insulin circulatinglevels until the end of glucose perfusion (Fig. 4B).
Similarly, i.v. infusion of methylamine at 0.5�mol kg−1
min−1 (0.033 mg kg−1 min−1) did not produce any change

480 M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484
Fig. 4. Effect of benzylamine infusion (2.3�mol kg−1 min−1) on IVGTTin conscious fasted rabbits. Changes in plasma glucose levels (A)and in immunoreactive insulin (IRI) levels (B) were measured duringand after i.v. administration of saline alone (S), glucose alone (Glc,55.5�mol kg−1 min−1), or benzylamine followed by glucose (Bza + Glc)in conscious fasted rabbits. Saline or benzylamine were administered for30 min (white horizontal bar) previously to the 30-min infusion of the glu-cose load (black horizontal bar). Variations from control values are given inmM for plasma glucose and in percentage of control for plasma IRI. Eachpoint is the mean± S.E.M. of 6 (Bza + Glc, triangles), 7 (S, circles), or 10(Glc, diamonds) rabbits. Significant difference between glucose alone andglucose plus benzylamine at:*** p < 0.001. Significant increase above the cor-responding condition without glucose (dotted line) at:+p < 0.05,++p < 0.01,+++p < 0.001.
in fasting levels of plasma glucose (Fig. 5A) or insulin(Fig. 5B). However, when methylamine was administeredjust before the glucose load, it clearly reduced the hypergly-caemic response. The increase in glycaemia at the end ofglucose infusion declined from 3.55± 0.31 mM in controlto 1.77± 0.54 mM in methylamine-treated rabbits (n = 11,p < 0.01), the mean AUC values of which was, in arbi-trary units, 84.6± 20.8 (i.e. significantly lower than con-trol at p < 0.01; Fig. 5A). No significant effect of methy-lamine was observed on plasma insulin since the maxi-mum increase reached 15 min after glucose infusion was208± 62% versus 314± 69% of fasting control, in theabsence or presence of methylamine, respectively (n = 11,NS) (Fig. 5B). The i.v. infusion of mafenide did not pro-duce any significant change in fasting levels of plasma glu-cose, even at the maximal dose tested: 1.12�mol kg−1 min−1
(0.25 mg kg−1 min−1; Fig. 6A). Moreover, when adminis-tered at this dose just before IVGTT, mafenide did not modifythe glycaemic response (Fig. 6A) or the pattern of circulatinginsulin (Fig. 6B).
Fig. 5. Changes in plasma glucose (A) and in immunoreactive insulin (IRI)levels (B) in conscious fasted rabbits, measured after the i.v. infusion ofphysiological saline (S, circles), glucose alone (Glc, 10 mg kg−1 min−1, dia-monds), methylamine (Mta, 0.5�mol kg−1 min−1, squares), and of methy-lamine just before the hexose (Mta + Glc, triangles); for the latter, the SSAOsubstrate was infused for 30 min (open bar) just before a 30-min hexose infu-sion (black horizontal bar). Ordinate scales,� mM plasma glucose refersto the variations from control values.� IRI levels are expressed as per-centage changes from the control level (control = 100%). Each point of anygiven curve represents the mean± S.E.M. from 10 to 11 rabbits.** p < 0.01,values significantly different between Glc and Mta + Glc. Different fromthe corresponding condition without glucose at+p < 0.05,++p < 0.01, and+++p < 0.001.
Despite the lack of hypoglycaemic effect of mafenide,this sulfonamide derivative was tested on insulin secretionin islets of Langerhans isolated from rabbit pancreas. In thepresence of 8 mM glucose, the islets did not increase insulinsecretion in response to 60-min incubation with 20�M or1 mM mafenide (110± 25% and 100± 9% of control insulinrelease, set at 100%, NS,n = 21). Islets did not respond to20�M or 1 mM methylamine (87± 15% and 116± 12%,NS,n = 22), or to benzylamine as already reported[17], whilethey responded to 200�M of the sulfonylurea secretagogueagent tolbutamide, or to 1�M forskolin by 195± 10% and250± 36% increase over basal insulin release, respectively(not shown). Similar results were obtained with murine pan-creatic islets (not shown).
3.5. Effects of benzylamine, methylamine and mafenideon hexose uptake in human adipocytes
Finally, to evaluate the relevance of the insulin-mimickingproperties of benzylamine, methylamine and mafenide, their

M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484 481
Fig. 6. Changes in plasma glucose (A) and in immunoreactive insulin (IRI)levels (B) induced in conscious fasted rabbits by the infusion of mafenide(Maf, 1.12�mol kg−1 min−1, squares) alone or just before a 30-min per-fusion of 55.5�mol kg−1 min−1 glucose (Maf + Glc, triangles). Differentfrom the corresponding condition without glucose (circles) at+p < 0.05,++p < 0.01, and+++p < 0.001. For more details see legend forFig. 5.
capacity to activate glucose uptake was compared in humanadipocytes. The response to benzylamine or methylaminereached a plateau at 0.1–1 mM which was equivalent to onethird of the maximal insulin stimulation (Fig. 7), identicalto that reached with hydrogen peroxide[16]. Involvementof SSAO activity in this response has been already reported,
Fig. 7. Effect of amines on glucose transport in human adipocytes obtainedfrom subcutaneous WAT in 16 women undergoing abdominal lipectomy(mean age: 42± 2, BMI: 25.4± 0.7). Effect of 0.1 (clearer columns) or1 mM amine (darker columns) different from basal uptake (white column)at ** p < 0.01 and*** p < 0.001.
since the effect of benzylamine was prevented by semicar-bazide[16]. Again, mafenide was the poorest activator, sinceit hardly reproduced less than 20% of insulin activation. Theeffects of these amines were not further increased by 0.1 mMvanadate while methylamine and benzylamine effects werecompletely suppressed by 1 mM glutathione (not shown).Mafenide did not alter insulin stimulation of glucose trans-port since response to 1 mM mafenide plus 100 nM insulinwas equivalent to 117± 21% of insulin alone (n = 3).
3.6. Amine oxidation and hydrogen peroxide generationin human, rat and mouse adipose tissues
To better understand why mafenide, although beingstructurally closer to benzylamine than methylamine, wasunable to activate glucose uptake, its capacity to behave as aSSAO substrate was tested in rat, mouse and human adiposetissues. When hydrogen peroxide production was measuredon homogenates of subcutaneous white adipose tissues(SCWAT), interspecies differences appeared regardingthe spontaneous release of hydrogen peroxide, with ratshowing the highest production. Mafenide was less ablethan benzylamine or methylamine to increase hydrogenperoxide production, especially in human SCWAT whereit was without effect (Table 2). Both methylamine- andb xidew thes
tratef ities
TH umana
NBBMMMM
H 6-wellpp ithout( e forhr np nits ofAe
tiona
tiona* tiona
enzylamine-induced production of hydrogen peroere totally inhibited by 1 mM semicarbazide, whateverpecies (not shown).
Since mafenide apparently did not work as a subsor human SSAO, an additional comparison of the capac
able 2ydrogen peroxide production by homogenates of rat, mouse and hdipose tissues
Hydrogen peroxide productionnmol−1 mg protein min−1
Rat Mouse Human
o addition 0.61± 0.04 0.38± 0.09 0.17± 0.03enzylamine, 0.1 mM 0.85± 0.07** 0.91± 0.13** 0.79± 0.20**
enzylamine, 1 mM ND 0.78± 0.12* 1.77± 0.21***
ethylamine, 0.1 mM 0.73± 0.06 0.49± 0.10 0.49± 0.07**
ethylamine, 1 mM 0.88± 0.07** 0.78± 0.13* 1.80± 0.19***
afenide, 0.1 mM 0.66± 0.04 0.48± 0.11 0.20± 0.04afenide, 1 mM ND 0.70± 0.13* 0.27± 0.06
omogenates of subcutaneous adipose tissues were distributed in 9lates in a 50�l volume (containing 75± 6�g, 110± 10�g and 43± 2�grotein for rat, mouse and human tissues, respectively) and incubated wno addition) or with the indicated amines and the fluorescent reactivydrogen peroxide detection in a final volume of 200�l for 30 min prioreading fluorescence as detailed in Section2. A standard curve of hydrogeeroxide was made in parallel and served to convert fluorescence umplex red into nmol of hydrogen peroxide produced. Mean± S.E.M. ofight (rat, mouse) or five (human) experiments. ND: not determined.* Significantly different from corresponding control without any addit p < 0.05.
** Significantly different from corresponding control without any addit p < 0.01.
** Significantly different from corresponding control without any addit p < 0.001.

482 M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484
Fig. 8. Inhibition of [14C]-benzylamine oxidation by phenelzine and amines in rat and human adipose tissue homogenates. In each species, oxidation ofan isotopic dilution of 0.5 mM [14C]-benzylamine was set at 100% reference for the expression of oxidation remaining in the presence of the indicatedconcentrations of mafenide (circles), methylamine (black squares) or benzylamine (triangles). Dose-dependent curve for phenelzine (white squares) is shownas a representative inhibition by irreversible amine oxidase inhibitor. Mean± S.E.M. ofn cases.
of methylamine and mafenide to inhibit [14C]-benzylamineoxidation was undertaken. When incubated 15 min with0.5 mM benzylamine (a dose activating glucose uptake),human WAT homogenates oxidized 59± 5 nmol of ben-zylamine min−1 g−1 tissue (n = 18), while homogenates ofmouse and rat WAT oxidized 46± 7 nmol min−1 g−1 and13± 2 nmol min−1 g−1, respectively (n = 3). Fig. 8 showsthat, in all species, [14C]-benzylamine oxidation was totallyabolished with the irreversible inhibitor phenelzine. It alsoillustrates that both methylamine and mafenide inhibited[14C]-benzylamine oxidation in mouse and rat WAT, whilemafenide was practically unable to inhibit benzylamine oxi-dation in human WAT. Analysis of the dose-dependent curvesobtained in man gave an IC50 of 606 and 2178�M for benzy-lamine and methylamine, whereas the value was >10,000�Mfor mafenide. These data definitively indicated that mafenidedoes not readily interact with human SSAO neither as a highaffinity substrate nor as an inhibitor, while it was an efficientcompetitor for benzylamine oxidation by rodent SSAO.
4. Discussion
Benzylamine is known to exhibit interesting insulin-mimicking effects on in vitro models[11] and on glucosed m-p anneb p-e ions( gro-c e tod thep ichs d/orS ereda sul-
fonamide topically used for its antimicrobial properties[20],was recently shown to interact with benzylamine oxidationin a pharmacological screening of novel SSAO substrates[19], and appeared as a good candidate for benzylamine-like drug development. Methylamine was also included inthe study since it is currently considered as a physiologicalSSAO substrate[21]. Thus, we tested the capacity of theseamines to activate glucose transport and to inhibit lipolysis inadipocytes, or to improve glucose disposal in models alreadyused to study the insulin-like actions of benzylamine[17].
Hydrogen peroxide generated during SSAO substrate oxi-dation appeared to be responsible for the amine-induced acti-vation of hexose transport, as previously reported[11,18]. Infact, hydrogen peroxide exhibited the same synergism withvanadate than the amines used to mimic the insulin stimu-lation of glucose transport in rodent adipocytes. Moreover,it was clearly demonstrated that neither benzaldehyde norammonia, i.e. other end-products of amine oxidation, wereable to reproduce the effect of benzylamine on glucose uptakeinto rat adipocytes. The insulin-like effects of the SSAOsubstrates, benzylamine and methylamine, observed in thepresent study are, therefore, in perfect agreement with pre-vious findings, such as: activation of glucose transport in thepresence of vanadate in rodent adipocytes[11,26], or in itsabsence in human fat cells[16], adipogenesis in 3T3 celllines [27,28] and antilipolysis[18]. Besides this confirma-t inea w.
thy-l d toe di-a ess-m coset hasn s orw on-s ion
isposal[13,15,17], while several benzylamine-related coounds have been reported to act as potassium chlockers[25]. However, in spite of its pharmacological prorties, this compound largely used for industrial applicatmanufacture of dyestuffs, pigments, textile auxiliaries, ahemicals) is devoid of any clinical and therapeutic usate. The aim of this work was, therefore, to investigateotential insulin-mimicking action of primary amines whhare with benzylamine a similar chemical structure anSAO substrate properties and which could be consids possible antidiabetic drugs. In this view, mafenide, a
lory aspect, the original findings obtained with methylamnd mafenide in the present work will be discussed belo
Indeed, our results indicate for the first time that meamine is able to improve in vivo glucose disposal anxhibit an antihyperglycaemic action which is not meted by a clear stimulation of insulin secretion. This assent agrees with the fact that methylamine activates glu
ransport and inhibits lipolysis in adipocytes, while ito effect on insulin release either from isolated islethen measuring circulating insulin after infusion in ccious rabbits. Unfortunately, this insulin-mimicking act

M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484 483
of methylamine could be counterbalanced by its deaminatedmetabolite, formaldehyde, reported to exert deleterious vas-cular effects in diabetes mellitus[21,29]. In keeping with this,the plasma and urine levels of methylamine are known to varyduring diabetes mellitus, and this biogenic amine has mainlybeen considered as a generator of formaldehyde and cardio-vascular complications[21,30,31], rather than an activator ofglucose uptake, a notion we initiated few years ago[12] andwhich is actually shared by other research groups[26,28].Likewise, a recent report also describes a beneficial effect ofa chronic treatment with methylamine on glucose utilizationin transgenic mice[32]. Our present data clearly showing thatmethylamine is able to increase glucose uptake or reduce lipidmobilization via its oxidation by SSAO in diverse speciesshould therefore be taken into account before pursuing thebasic and clinical investigations that recommend to limit vas-cular complications of diabetes by inhibiting SSAO activity[29]. Obviously, methylamine on its own cannot be consid-ered as a drug, but it makes useful the concept of exploringthe antihyperglycaemic properties of chemically synthetizedamine oxidase substrates that should become potential drugsfor metabolic diseases.
Regarding to its amine moiety, mafenide was supposedto be an SSAO substrate and to exert insulin-like actionsvia the formation of hydrogen peroxide resulting from itsputative deaminative oxidation. Considering that moleculeswr sub-sa cre-t ablet ivoc oods ot inm ple-t ide( e duet mod-es er-tl anS cesi nota rtiesf vari-a ewn -m M[ ter-a ided thanb genp -d port
observed in rat and mouse adipocytes with mafenide plusvanadate did not exceed benzylamine effect. Our compara-tive approach indicated that mafenide could not be consideredas an antihyperglycaemic drug, keeping in mind its pooreffect on glucose transport at least in human fat cells, andits inability to increase insulin release or to lower blood glu-cose in rabbits. However, our rationale for the investigationof drugs that could be more efficient than benzylamine can-not be discarded. A special attention to the animal modelsused must be taken for future pharmacological screening ofSSAO substrates with potential antidiabetic properties andeven if several of them reproduce the pathology of type2 diabetes[36], their interest will be limited by the inter-species differences in amine oxidase substrate selectivity.Any “benzylamine-related” drug, detected via modelling ofstructure/activity catalytic domain of the SSAO/VAP-1[19],or by other pharmacological survey, must be checked for itsactivity towards human SSAO and should be tested for glu-cose uptake in human adipocytes, before going into furtherstudies.
5. Conclusion
Taken together, our observations led to conclude that thea anti-h vivoi sub-s con-n emica torso AOi dia-b find-i andm s foro casef hs
A
deI stillay ed by“ sP
tech-n
R
rends
ith a sulfonamide structure reduce blood glucose[33], as aesult of ATP-sensitive potassium channels closure andequent insulin release from islets of Langerhans[34], it waslso hypothesized that mafenide could improve insulin se
ion. The present work indicates that mafenide was uno activate insulin secretion under both in vitro and in vonditions. In addition, this diamine appeared to be a gubstrate for SSAO activity in rodents (rat, mouse) but nan. A similar observation was obtained during the com
ion of this work when 4-aminomethyl-benzenesulfonamidentical to mafenide) was tested as a SSAO substrato its resemblance to a pharmacophore revealed bylling of the catalytic domain of SSAO/VAP-1[19]. In thistructure-activity study, mafenide exhibited kinetic propies that corresponded to a very highKm (poor affinity) andow Vmax (poor capacity to be oxidized) towards humSAO/VAP-1 [19]. Therefore, species-specific differen
n the substrate selectivity of SSAO unfortunately didllowed to extrapolate to human its promising prope
ound in rodent models. However, such species-specifictions are well admitted[35] since, for instance, tyraminhich can be oxidized by SSAO in rat adipocytes[18,22]doesot behave as SSAO substrate in human fat cells[7]. Suppleentary experiments on inhibition of the oxidation of 0.5 m
14C]-tyramine evidenced that mafenide did not readily inct either with murine or human MAO. Noteworthy, mafenid not exhibit better pharmacological characteristicsenzylamine in rats or mice, when considering hydroeroxide production, inhibition of [14C]-benzylamine oxiation. Even the satisfactory activation of glucose trans
ntimicrobial sulfonamide mafenide does not work as anyperglycaemic drug. Nevertheless, the in vitro and in
nsulin-mimicking properties of the endogenous SSAOtrate methylamine add support to our initial hypothesisecting SSAO substrate properties and anti-hyperglycactivity. They also indicate that the impact of SSAO inhibin glycaemic control should be considered when SS
nhibitors are used to prevent vascular complications ofetes. Finally, although somewhat negative, the present
ngs leave open the search for similar – though saferore efficient – SSAO-interacting drugs as candidateral treatment of diabetes mellitus, as it is currently the
or novel PPAR exogenous ligands[37], the design of whictill results in an increased benefit/risk ratio.
cknowledgments
This work was partly supported by grants from Fondonvestigaciones Sanitarias (FIS) and from Junta de CaLeon. Spanish–French exchanges were partly financ
Accords INSERM-CSIC” and “Communaute de Travail deyrenees”.The authors express gratitude to the “Service de Zoo
ie de l’IFR31” (Toulouse, France) for animal care.
eferences
[1] Ahima RS, Flier JS. Adipose tissue as an endocrine organ. TEndocrinol Metab 2000;11:327–32.

484 M.C. Iglesias-Osma et al. / Pharmacological Research 52 (2005) 475–484
[2] Fruhbeck G, Gomez-Ambrosi J, Muruzabal FJ, Burrell MA. Theadipocyte: a model for integration of endocrine and metabolic sig-naling in energy metabolism regulation. Am J Physiol EndocrinolMetab 2001;280:E827–47.
[3] Abel ED, Peroni O, Kim JK, Kim YB, Boss O, Hadro E, et al.Adipose-selective targeting of the GLUT4 gene impairs insulin actionin muscle and liver. Nature 2001;409:729–33.
[4] Rajala MW, Scherer PE. The adipocyte at the crossroads ofenergy homeostasis, inflammation, and atherosclerosis. Endocrinol-ogy 2003;144:3765–73.
[5] Raimondi L, Pirisino R, Ignesti G, Capecchi S, Banchelli G, BuffoniF. Semicarbazide-sensitive amine oxidase activity (SSAO) of rat epi-didymal white adipose tissue. Biochem Pharmacol 1991;41:467–9.
[6] Morris NJ, Ducret A, Aebersold R, Ross SA, Keller SR, Lien-hard GE. Membrane amine oxidase cloning and identification asa major protein in the adipocyte plasma membrane. J Biol Chem1997;272:9388–92.
[7] Pizzinat N, Marti L, Remaury A, Leger F, Langin D, Lafontan M, etal. High expression of monoamine oxidases in human white adiposetissue: evidence for their involvement in noradrenaline clearance.Biochem Pharmacol 1999;58:1735–42.
[8] Buffoni F. Semicarbazide-sensitive amine oxidases: some bio-chemical properties and general considerations. Progr Brain Res1995;106:323–31.
[9] Salmi M, Yegutkin GG, Lehvonen R, Koskinen K, Salminen T, Jalka-nen S. A cell surface amine oxidase directly controls lymphocytemigration. Immunity 2001;14:265–76.
[10] Langford SD, Trent MB, Boor PJ. Semicarbazide-sensitive amineoxidase and extracellular matrix deposition by smooth-muscle cells.Cardiovasc Toxicol 2002;2:141–50.
[11] Enrique-Tarancon G, Marti L, Morin N, Lizcano JM, Unzeta M,e onadi-
[ pscoop-IRSslo-
[ A.minehem
[ nos ofmia in
.[ mis
esionrats.
[ al.lucosel Exp
[ sillactsbbits
[ llmine
[ M,ide-
sensitive amine oxidase/VAP-1: identification of novel substrateswith insulin-like activity. J Med Chem 2004;47:4865–74.
[20] Eagon RG, McManus AT. The effect of mafenide on dihydropteroatesynthase. J Antimicrob Chemother 1990;25:25–9.
[21] Yu PH, Wright S, Fan EH, Lun ZR, Gubisne-Harberle D. Physiolog-ical and pathological implications of semicarbazide-sensitive amineoxidase. Biochim Biophys Acta 2003;1647:193–9.
[22] Marti L, Morin N, Enrique-Tarancon G, Prevot D, Lafontan M, Tes-tar X, et al. Tyramine and vanadate synergistically stimulate glucosetransport in rat adipocytes by amine oxidase-dependent generationof hydrogen peroxide. J Pharmacol Exp Ther 1998;285:342–9.
[23] Garcia-Barrado MJ, Sancho C, Iglesias-Osma MC, MoratinosJ. Effects of verapamil and elgodipine on isoprenaline-inducedmetabolic responses in rabbits. Eur J Pharmacol 2001;415:105–15.
[24] Moratinos J, Carpene C, De Pablos I, Reverte M. Role of alpha1-and alpha2-adrenoceptors in catecholamine induced hyperglycaemia,lipolysis and insulin secretion in conscious fasted rabbits. Br J Phar-macol 1988;24:299–310.
[25] Banchelli G, Raimondi L, Ghelardini C, Pirisino R, Bertini V, DeMunno A, et al. Benzylamine-related compounds stimulate rat vasdeferens neurotransmission and potentiate memory in the mouse act-ing as potassium channel blockers. Pharmacol Res 2000;41:151–62.
[26] Yu PH, Wang M, Fan H, Gubisne-Haberle D. Involvement SSAO-mediated deamination in adipose glucose transport and weight gainin obese diabetic KKAy mice. Am J Physiol Endocrinol Metab2004;286:E634–41.
[27] Fontana E, Boucher J, Marti L, Lizcano JM, Testar X, ZorzanoA, et al. Amine oxidase substrates mimic several of the insulineffects on adipocyte differentiation in 3T3 F442A cells. Biochem
[ ive3-L1
[ tingon in
[ bymagebetes
[ ofation
[ t al.al con-SEB
[ ho-ogia
[ dlogia
[ zide-toxi-
[ ina-t: Amacol
[ cal
Sevilla L, et al. Role of semicarbazide-sensitive amine oxidasglucose transport and GLUT4 recruitment to the cell surface inpose cells. J Biol Chem 1998;273:8025–32.
12] Enrique-Tarancon G, Castan I, Morin N, Marti L, Abella A, CamM, et al. Substrates of semicarbazide-sensitive amine oxidaseerate with vanadate to stimulate tyrosine phosphorylation ofproteins, phosphatidylinositol 3-kinase activity and GLUT4 trancation in adipose cells. Biochem J 2000;350:171–80.
13] Abella A, Marti L, Carpene C, Palacin M, Testar X, ZorzanoStimulation of glucose transport by semicarbazide-sensitive aoxidase activity in adipocytes from diabetic rats. J Physiol Bioc2003;59:153–60.
14] Marti L, Abella A, Carpene C, Palacin M, Testar X, ZorzaA. Combined treatment with benzylamine and low dosagevanadate enhances glucose tolerance and reduces hyperglycestreptozotocin-induced diabetic rats. Diabetes 2001;50:2061–8
15] Abella A, Marti L, Camps M, Claret M, Fernandez-Alvarez J, GoR, et al. Semicarbazide-sensitive amine oxidase/vascular adhprotein-1 activity exerts an antidiabetic action in Goto-KakizakiDiabetes 2003;52:1004–13.
16] Morin N, Lizcano JM, Fontana E, Marti L, Smih F, Rouet P, etSemicarbazide-sensitive amine oxidase substrates stimulate gtransport and inhibit lipolysis in human adipocytes. J PharmacoTher 2001;297:563–72.
17] Iglesias-Osma MC, Garcia-Barrado MJ, Visentin V, Pastor-ManMF, Bour S, Prevot D, et al. Benzylamine exhibits insulin-like effeon glucose disposal, glucose transport and fat cell lipolysis in raand diabetic mice. J Pharmacol Exp Ther 2004;309:1020–8.
18] Visentin V, Prevot D, Marti L, Carpene C. Inhibition of rat fat celipolysis by monoamine oxidase and semicarbazide-sensitive aoxidase substrates. Eur J Pharmacol 2003;466:235–43.
19] Marti L, Abella A, De la Cruz X, Garcia-Vicente S, UnzetaCarpene C, et al. Exploring the binding mode of semicarbaz
J 2001;356:769–77.28] Mercier N, Moldes M, El Hadri K, Feve B. Semicarbazide-sensit
amine oxidase activation promotes adipose conversion of 3Tcells. Biochem J 2001;358:335–42.
29] Ekblom J. Potential therapeutic value of drugs inhibisemicarbazide-sensitive amine oxidase: vascular cytoprotectidiabetes mellitus. Pharmacol Res 1998;37:87–92.
30] Yu PH, Zuo DM. Oxidative deamination of methylaminesemicarbazide-sensitive amine oxidase leads to cytotoxic dain endothelial cells. Possible consequences for diabetes. Dia1993;42:594–603.
31] Yu PH. Deamination of methylamine and angiopathy; toxicityformaldehyde, oxidative stress and relevance to protein glycoxidin diabetes. J Neural Transm Suppl 1998;105:201–16.
32] Stolen C, Madanat R, Marti L, Kari S, Yegutkin GG, Sariola H, eSemicarbazide sensitive amine oxidase overexpression has dusequences: insulin mimicry and diabetes-like complications. FAJ 2004;18:702–4.
33] Henquin JC. The fiftieth anniversary of hypoglycaemic sulpnamides. How did the mother compound work? Diabetol1992;35:907–12.
34] Ashcroft FM, Gribble FM. KATP-sensitive K+ channels aninsulin secretion: their role in health and disease. Diabeto1999;42:903–19.
35] Lyles GA. Mammalian plasma and tissue-bound semicarbasensitive amine oxidases: biochemical, pharmacological andcological aspects. Int J Biochem Cell Biol 1996;28:259–74.
36] Srinivasan K, Viswanad B, Asrat L, Kaul CL, Ramarao P. Combtion of high-fat diet-fed and low-dose streptozotocin-treated ramodel for type 2 diabetes and pharmacological screening. PharRes 2005;52:313–20.
37] Kota BP, Huang TH-W, Roufogalis BD. An overview of biologimechanisms of PPARs. Pharmacol Res 2005;51:85–94.
Copyright © 2022 FDOKUMEN




















