
Members of Sebacinales subgroup B form mycorrhizae with epiphytic orchids in a neotropical mountain...
11
ORIGINAL ARTICLE Members of Sebacinales subgroup B form mycorrhizae with epiphytic orchids in a neotropical mountain rain forest Juan Pablo Suárez & Michael Weiß & Andrea Abele & Franz Oberwinkler & Ingrid Kottke Received: 25 June 2007 / Revised: 12 February 2008 / Accepted: 26 February 2008 / Published online: 16 April 2008 # German Mycological Society and Springer 2008 Abstract Previous investigations revealed that epiphytic orchids in a mountain rain forest in southern Ecuador formed mycorrhizae with diverse members of Tulasnellales. Using specific primers, we now show that the same orchids are also associated with Sebacinales. Ultrastructural obser- vations confirmed the Sebacinales mycobionts in situ. Mycorrhizae of flowering individuals of Stelis hallii, S. superbiens, S. concinna and Pleurothallis lilijae were sampled in different forest types of the mountain rain forest of southern Ecuador along an altitudinal gradient between 1,850 and 2,100 m a.s.l. Phylogenetic analysis of fungal nuclear rDNA sequences coding for the ribosomal large subunit (nucLSU) showed the presence of eight sequence types based on proportional differences of <1% bp. All sequence types clustered in the Sebacinales subgroup B which also contained sequences of mycobionts from ericads and terrestrial orchids. Sequences of the nuclear rDNA 5.8S subunit, including parts of the internal transcribed spacers ITS1 and ITS2 (5.8-ITS) from the mycobionts of the epiphytic orchids, were distinct from published sequences of sebacinoid mycobionts of green terrestrial orchids and ericads. Sebacinales sequences from different epiphytic orchid species differed at least by 1% bp as was previously found for Tulasnella sequences. Sebacinales occurred less frequently and with a lower number of sequence types than Tulasnellales, but distribution along the altitudinal gradient was similar. Introduction Recent investigations have shown that epiphytic orchids constantly form mycorrhizae in roots that are in close contact with the substrate (Otero et al. 2002, 2004; Pereira et al. 2003, 2005; Ma et al. 2003; Suárez et al. 2006) suggesting that mycorrhizal fungi are important for adult, green, epiphytic plants. Our previous work revealed by molecular identification Tulasnellales as frequently associ- ated mycobionts of epiphytic Stelis and Pleurothallis (Pleurothallidinae) in an Andean cloud forest of southern Ecuador (Suárez et al. 2006). Sequence types from seven clades of Tulasnellales, new to science and distinct from published sequences from terrestrial orchid fungi, were identified in the roots of these closely related epiphytic orchids (Suárez et al. 2006). In the tropical mountain rain forest, relative humidity is close to saturation most of the time, but periods of very low humidity (20%) can occur day and night in the drier months (Bendix et al. 2008). During the periods of water shortage, the mycorrhiza-forming fungi may improve water and nutrient uptake of the orchids (Yoder et al. 2000; Zotz and Hietz 2001). We hypothesized that other fungi may also be present as mycobionts in these epiphytic orchids. Beside Tulasnel- lales, Sebacinales and Ceratobasidiales are common orchid mycorrhiza-forming fungi (Currah et al. 1997). In a first attempt, using specific primers, we obtained partial rDNA sequences of the nuclear gene coding for the ribosomal large subunit (nucLSU) of Sebacinales and found that their Mycol Progress (2008) 7:75–85 DOI 10.1007/s11557-008-0554-4 J. P. Suárez (*) Centro de Biología Celular y Molecular, Universidad Técnica Particular de Loja, San Cayetano Alto s/n C.P., 11 01 608 Loja, Ecuador e-mail: [email protected] J. P. Suárez : M. Weiß : A. Abele : F. Oberwinkler : I. Kottke Spezielle Botanik und Mykologie, Eberhard-Karls-Universität Tübingen, Botanisches Institut, Auf der Morgenstelle 1, 72076 Tübingen, Germany
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Members of Sebacinales subgroup B form mycorrhizae with epiphytic orchids in a neotropical mountain...

ORIGINAL ARTICLE
Members of Sebacinales subgroup B form mycorrhizaewith epiphytic orchids in a neotropicalmountain rain forest
Juan Pablo Suárez & Michael Weiß & Andrea Abele &
Franz Oberwinkler & Ingrid Kottke
Received: 25 June 2007 /Revised: 12 February 2008 /Accepted: 26 February 2008 /Published online: 16 April 2008# German Mycological Society and Springer 2008
Abstract Previous investigations revealed that epiphyticorchids in a mountain rain forest in southern Ecuadorformed mycorrhizae with diverse members of Tulasnellales.Using specific primers, we now show that the same orchidsare also associated with Sebacinales. Ultrastructural obser-vations confirmed the Sebacinales mycobionts in situ.Mycorrhizae of flowering individuals of Stelis hallii, S.superbiens, S. concinna and Pleurothallis lilijae weresampled in different forest types of the mountain rain forestof southern Ecuador along an altitudinal gradient between1,850 and 2,100 m a.s.l. Phylogenetic analysis of fungalnuclear rDNA sequences coding for the ribosomal largesubunit (nucLSU) showed the presence of eight sequencetypes based on proportional differences of <1% bp. Allsequence types clustered in the Sebacinales subgroup Bwhich also contained sequences of mycobionts from ericadsand terrestrial orchids. Sequences of the nuclear rDNA 5.8Ssubunit, including parts of the internal transcribed spacersITS1 and ITS2 (5.8-ITS) from the mycobionts of theepiphytic orchids, were distinct from published sequencesof sebacinoid mycobionts of green terrestrial orchids andericads. Sebacinales sequences from different epiphyticorchid species differed at least by 1% bp as was previously
found for Tulasnella sequences. Sebacinales occurred lessfrequently and with a lower number of sequence types thanTulasnellales, but distribution along the altitudinal gradientwas similar.
Introduction
Recent investigations have shown that epiphytic orchidsconstantly form mycorrhizae in roots that are in closecontact with the substrate (Otero et al. 2002, 2004; Pereiraet al. 2003, 2005; Ma et al. 2003; Suárez et al. 2006)suggesting that mycorrhizal fungi are important for adult,green, epiphytic plants. Our previous work revealed bymolecular identification Tulasnellales as frequently associ-ated mycobionts of epiphytic Stelis and Pleurothallis(Pleurothallidinae) in an Andean cloud forest of southernEcuador (Suárez et al. 2006). Sequence types from sevenclades of Tulasnellales, new to science and distinct frompublished sequences from terrestrial orchid fungi, wereidentified in the roots of these closely related epiphyticorchids (Suárez et al. 2006). In the tropical mountain rainforest, relative humidity is close to saturation most of thetime, but periods of very low humidity (20%) can occur dayand night in the drier months (Bendix et al. 2008). Duringthe periods of water shortage, the mycorrhiza-forming fungimay improve water and nutrient uptake of the orchids(Yoder et al. 2000; Zotz and Hietz 2001).
We hypothesized that other fungi may also be present asmycobionts in these epiphytic orchids. Beside Tulasnel-lales, Sebacinales and Ceratobasidiales are common orchidmycorrhiza-forming fungi (Currah et al. 1997). In a firstattempt, using specific primers, we obtained partial rDNAsequences of the nuclear gene coding for the ribosomallarge subunit (nucLSU) of Sebacinales and found that their
Mycol Progress (2008) 7:75–85DOI 10.1007/s11557-008-0554-4
J. P. Suárez (*)Centro de Biología Celular y Molecular,Universidad Técnica Particular de Loja,San Cayetano Alto s/n C.P.,11 01 608 Loja, Ecuadore-mail: [email protected]
J. P. Suárez :M. Weiß :A. Abele : F. Oberwinkler : I. KottkeSpezielle Botanik und Mykologie,Eberhard-Karls-Universität Tübingen,Botanisches Institut,Auf der Morgenstelle 1,72076 Tübingen, Germany

sequence types were distinct from those detected in Andeanericad mycorrhizae sampled in the same forest (Setaro et al.2006b; Kottke et al. 2008). To further define the myco-bionts of the epiphytic orchids and clarify their phyloge-netic position within the Sebacinales, additional sequencingwas carried out including the nuclear rDNA 5.8S subunitand parts of the internal transcribed spacers ITS1 and ITS2(5.8S-ITS). Molecular phylogenetic analysis was combinedwith ultrastructural observations to verify the presence ofSebacinales in situ. Finally, we compared fungal richnessand distribution of the Sebacinales and Tulasnellalesmycobionts of the epiphytic orchids from different foresttypes along an altitudinal gradient in the tropical mountainrain forest.
Materials and methods
Study site
The study site, the Reserva Biológica San Francisco(RBSF), is located on the eastern slope of the CordilleraEl Consuelo in the Northern Andes of southern Ecuadorhalf way between Loja and Zamora, Zamora-Chinchipeprovince (3°58!S, 79°04!W). The forest area where sam-pling was carried out belongs to tierra templada (averageannual temperature 15.5°C) and is classified as an ever-green lower and upper montane rain forest (Ceja Andina)especially characterized by a richness of vascular andcryptogamic epiphytes (Beck et al. 2008). The climate inthe forest area is mainly influenced by strong easterliesthroughout the year, which bring up the clouds from theAmazonian valley. Cloudiness, sheltering the ground fromdirect irradiance, has its maximum at altitudes above1,800 m a.s.l. which is also the zone of intense precipitation(2,200 mm average per year) (Bendix et al. 2006). Relativehumidity is close to saturation most of the time, but periodsof very low humidity (20%) can occur day and night in thedrier months (Bendix et al. 2008).
The forest covering the ridges and steep slopes (35–45°inclination) is extraordinarily rich in tree species, gaining aheight of 15–20 m, and mostly difficult to identify in a non-flowering state. During sampling of orchid mycorrhizae,tree species were therefore not recorded. The altitudinalgradient between 1,850 and 2,100 m a.s.l. comprises threetypes of forests as differentiated on a floristic and astructural basis (Homeier et al. 2008; Paulsch et al. 2008).Stem height and diameter, stand and crown density declinewith increasing altitude resulting in higher irradiance andwind exposure in the upper parts of the forest (Homeier2004). Microclimate, therefore, cannot be expected to bethe same for all sampled orchids, but long-term measure-ments at a microscale level were not available.
Sampling
Sampling of Stelis hallii Lindl., Stelis superbiens Lindl.,Stelis concinna Lind. and Pleurothallis lilijae Foldats wascarried out along the mountain ridge between 1,850 and2,100 m a.s.l. as described in Suárez et al. (2006). Stelishallii was found in the lower part of the forest (1,850–1900 m a.s.l., forest type Ia according to Homeier 2004,megaphyll ridge forest according to Paulsch et al. 2008), S.superbiens and P. lilijae were sampled between 1,900 and2,100 m a.s.l. (forest type II of Homeier 2004, macrophyllridge forest of Paulsch et al. 2008). Stelis concinna wasrestricted to the upper part of forest type II at the lower endof forest type III (Homeier 2004; microphyll ridge forest ofPaulsch et al. 2008). Root samples were collected continu-ously during 3 years from 2003 to 2005 from a total of 83flowering individuals, 22 of S. hallii, 17 of S. superbiens, 13of P. lilijae, and 31 of S. concinna; 6 samples of S. concinnain addition to our previous work (Suárez et al. 2006).
All selected plants were epiphytes on trunks or branchesof standing trees. One epiphytic orchid individual wascollected per tree. Only roots that were in direct contactwith the substrate were collected. One to four roots werecollected per plant, packed in aluminium foil to preventdesiccation and transported to the laboratory the same day.Vouchers of the orchid specimens were deposited in theHerbarium of UTPL, Loja, Ecuador, including flowersfixed in ethanol. Root samples were processed the same dayof collection as pre-investigation had revealed a rapid lossof vitality of the symbiotic fungi. Light microscopicexamination was used to select material with fungal coils.
Transmission electron microscopy
Anatomy of mycorrhizae and ultrastructure of hyphae wasstudied as described in Suárez et al. (2006). Shortly, rootpieces of samples displaying high frequency of vitallooking hyphal coils were fixed in 2.5% glutaraldehyde-formaldehyde in Sørensen buffer (Karnovsky 1965), post-fixed in 1% osmium tetroxide for 1 h, dehydrated in anacetone series and flat-embedded in Spurr’s resin lowviscosity, longer pot-life formulation (Spurr 1969). Twentysamples with apparently vital hyphae according to semi-thin section observation were selected for ultra-thin cutting.Sections were mounted on Formvar-coated copper gridsand stained with 1% uranyl acetate (40 min) and lead citrate(12 min) and studied using a TEM Zeiss 109 or Zeiss 902.
DNA extraction, PCR and sequencing
DNA was extracted from fresh or dried mycorrhizal planttissue, from which the velamen was removed, using a PlantMini Kit (Qiagen, Hilden, Germany) according to the
76 Mycol Progress (2008) 7:75–85

manufacturer’s instructions. Nested PCR was conducted tospecifically amplify DNA from sebacinoid fungi. The firstamplification was carried out with the primer combinationITS1F (5!-CTTGGTCATTTAGAGGAAGTAA-3!; Gardesand Bruns 1993) and TW14 (5!-GCTATCCTGAGGGAAACTTC-3!; Cullings 1994) or ITS1 (5!-TCCGTAGGTGAACCTGCGG-3!; White et al. 1990) and TW14, and thesecond with the primer combinations ITS3Seb (5!-TGAGTGTCATTGTAATCTCAC-3!; M. Berbee; Setaro etal. 2006b) and NL4 (5!-GGTCCGTGTTTCAAGACGG-3!;White et al. 1990) or ITS3Seb and LR5 (5!-TCCTGAGGGAAACTTCG-3!; Vilgalys and Hester 1990) for theinternal transcribed spacers ITS2 and part of the ribosomallarge subunit (nucLSU). The primer combination ITS1 andITS3SebR (5!-GTGAGATTACAATGACACTCA-3!; re-verse complement of ITS3Seb) was used to amplify theinternal transcribed spacer ITS1, the 5.8S region and part ofITS2 of the nuclear rDNA (5.8S-ITS). For the nested PCR,the product obtained in the first PCR was diluted 1:10,1:100, or 1:1,000.
PCR conditions were as detailed in Suárez et al. (2006).Success of the PCR amplifications was tested in 0.7%agarose, stained in a solution of ethidium bromide 0.5 μgml!1. Negative controls without template DNA were usedin every PCR. PCR products were purified using theQIAquick protocol (Qiagen). Cycle sequencing was con-ducted using BigDye version 3.1 chemistry, and sequencingwas done on an ABI 3100 Genetic Analyzer (AppliedBiosystems, Foster City, Calif.). Both strands of DNA weresequenced. Sequence editing was performed usingSequencher version 4.5 (Gene Codes, Ann Arbor, Mich.).
Phylogenetic analyses
A collection of representative nucLSU Sebacinales sequen-ces for phylogenetic analysis was downloaded fromGenBank (http://www.ncbi.nlm.nih.gov). We also usedGenBank BLAST (Altschul et al. 1997) to detect publishedsequences with high similarity to the nucLSU sequences ofthe Sebacinales mycobionts detected in the four Ecuadorianepiphytic orchids. The obtained sequences were alignedusing the G-INS-i strategy implemented in MAFFT v.5.667(Katoh et al. 2005). A neighbor-joining (NJ) analysis of aninitial data set was performed in PAUP* (Swofford 2002)using the BIONJ modification of the NJ algorithm (Gascuel1997) with a general time-reversible model of nucleotidesubstitution, additionally assuming a proportion of invariantsites and gamma-distributed substitution rates at theremaining sites (GTR+I+G; see, e.g., Swofford et al.1996). The final dataset, covering the D1/D2 region of thenucLSU, was the result of pruning the initial tree to removerepeated sequences from the same host without closerelationship to our sequences, and to select a representative
set of Sebacinales subgroup A (Weiß et al. 2004) that weused as outgroup sequences. The resulting dataset wasanalyzed using heuristic maximum likelihood (ML) asimplemented in the PHYML software, v.2.4.4 (Guindonand Gascuel 2003), starting from a BIONJ tree and usingthe GTR+I+G DNA substitution model. The gammadistribution was approximated with four discrete ratecategories. All model parameters were estimated usingML. Branch support was inferred from 1000 replicates ofnon-parametric bootstrapping (Felsenstein 1985). In addi-tion, we applied a Bayesian approach based on Markovchain Monte Carlo (MCMC) as implemented in MrBayes,v.3.1 (Ronquist and Huelsenbeck 2003), again using theGTR+I+G model, with all parameters sampled duringMCMC. We ran two independent MCMC analyses, eachinvolving four incrementally heated Markov chains over5 million generations and using random starting trees. Treeswere sampled every 100 generations resulting in a total of50,000 trees in each run from which the last 30,000 wereused to compute a pooled majority rule consensus tree.
For 5.8S-ITS analysis, we used BLAST to search theGenBank nucleotide database for Sebacinales sequencesrelated to those detected in our orchid root samples. Some5.8S-ITS reference sequences used in this study are derivedfrom Sebacina vermifera strains kindly provided by theNational Institute of Agrobiological Sciences (NIAS),Japan, that were previously isolated from Australianorchids and determined by J. H. Warcup (Warcup 1988).The obtained dataset was aligned using the L-INS-i strategyimplemented in MAFFT v.5.667 (Katoh et al. 2005). Due tothe heterogeneity of the Sebacinales sequences we had toexclude considerable portions of the 5.8S-ITS sequencesfor phylogenetic analysis. The resulting dataset wasanalyzed using heuristic maximum likelihood (ML) and aBayesian approach based on Markov chain Monte Carlo(MCMC) as mentioned before.
Sequence types were defined based on <1% bp propor-tional differences between nucLSU D1/D2 sequences. Wecalculated a sample-based rarefaction accumulation curvewith 95% confidence intervals and estimated the totalsequence type richness with Chao2 and Jackknife2 usingthe software EstimateS, v.7.5.0 (Colwell 2004).
Results
Sebacinales in the root cortical tissue of the epiphyticorchids
The ultrastructural studies were carried out in order toverify whether sebacinoid hyphae colonized the corticaltissue of the orchids as mutualistic symbionts and not onlythe velamen or surface of the roots as contaminants.
Mycol Progress (2008) 7:75–85 77
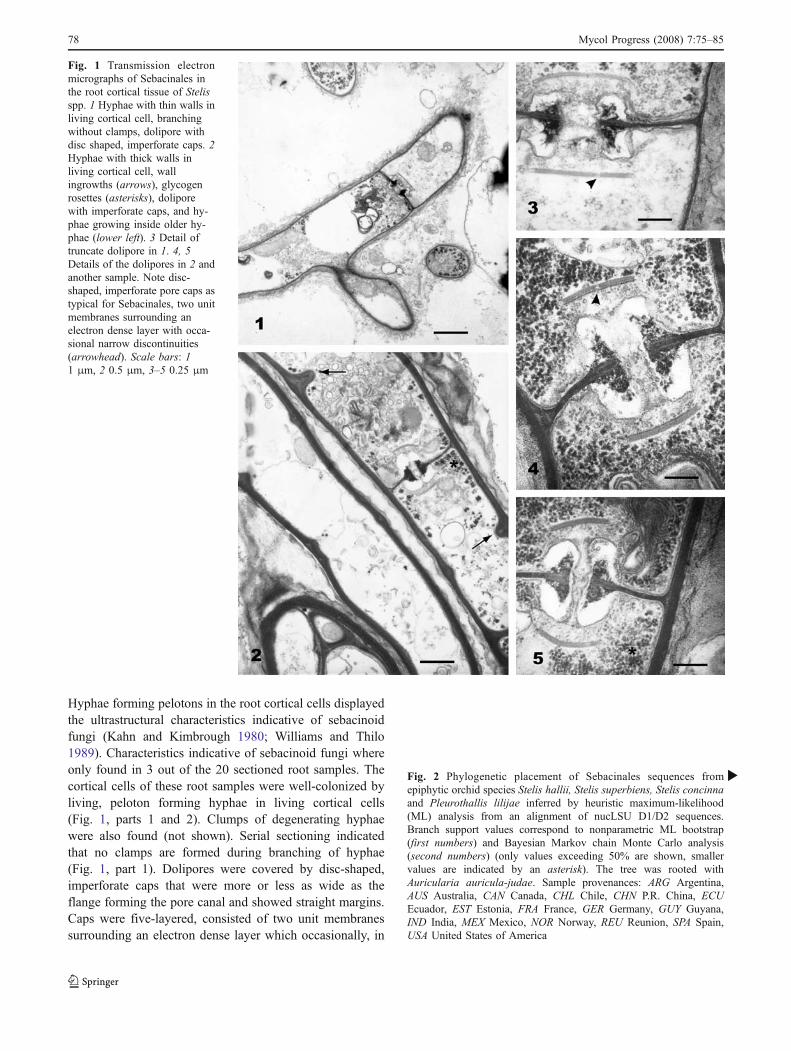
Hyphae forming pelotons in the root cortical cells displayedthe ultrastructural characteristics indicative of sebacinoidfungi (Kahn and Kimbrough 1980; Williams and Thilo1989). Characteristics indicative of sebacinoid fungi whereonly found in 3 out of the 20 sectioned root samples. Thecortical cells of these root samples were well-colonized byliving, peloton forming hyphae in living cortical cells(Fig. 1, parts 1 and 2). Clumps of degenerating hyphaewere also found (not shown). Serial sectioning indicatedthat no clamps are formed during branching of hyphae(Fig. 1, part 1). Dolipores were covered by disc-shaped,imperforate caps that were more or less as wide as theflange forming the pore canal and showed straight margins.Caps were five-layered, consisted of two unit membranessurrounding an electron dense layer which occasionally, in
Fig. 1 Transmission electronmicrographs of Sebacinales inthe root cortical tissue of Stelisspp. 1 Hyphae with thin walls inliving cortical cell, branchingwithout clamps, dolipore withdisc shaped, imperforate caps. 2Hyphae with thick walls inliving cortical cell, wallingrowths (arrows), glycogenrosettes (asterisks), doliporewith imperforate caps, and hy-phae growing inside older hy-phae (lower left). 3 Detail oftruncate dolipore in 1. 4, 5Details of the dolipores in 2 andanother sample. Note disc-shaped, imperforate pore caps astypical for Sebacinales, two unitmembranes surrounding anelectron dense layer with occa-sional narrow discontinuities(arrowhead). Scale bars: 11 μm, 2 0.5 μm, 3–5 0.25 μm
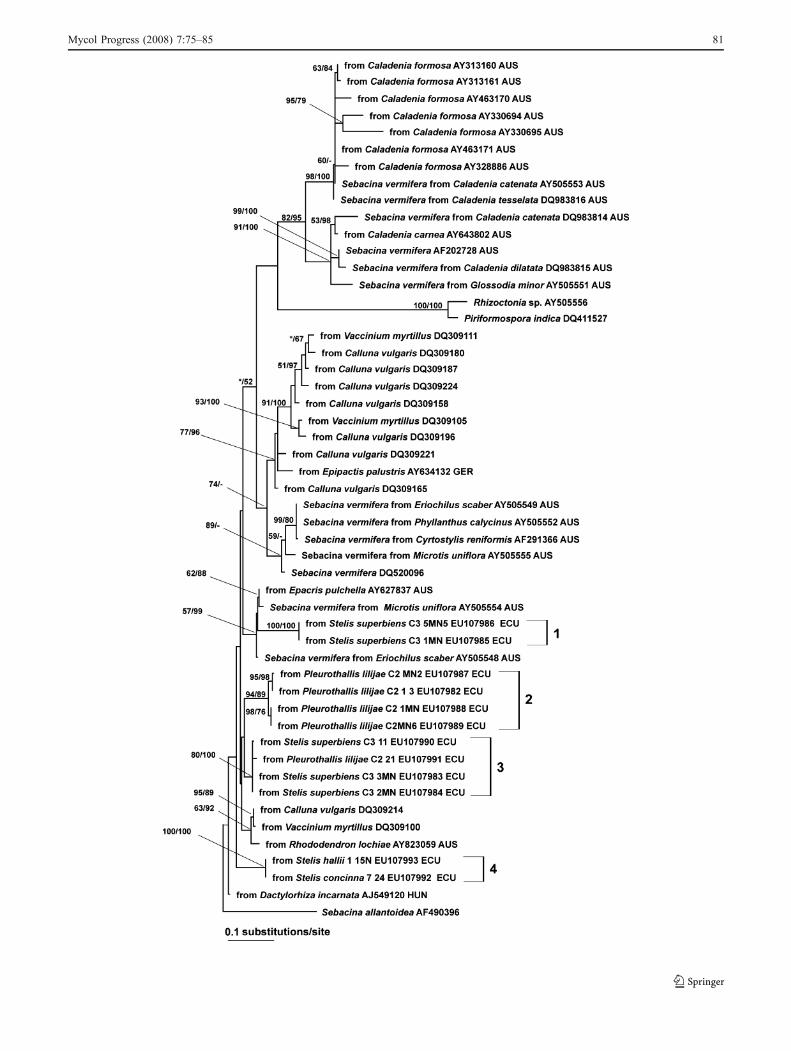
Fig. 2 Phylogenetic placement of Sebacinales sequences fromepiphytic orchid species Stelis hallii, Stelis superbiens, Stelis concinnaand Pleurothallis lilijae inferred by heuristic maximum-likelihood(ML) analysis from an alignment of nucLSU D1/D2 sequences.Branch support values correspond to nonparametric ML bootstrap(first numbers) and Bayesian Markov chain Monte Carlo analysis(second numbers) (only values exceeding 50% are shown, smallervalues are indicated by an asterisk). The tree was rooted withAuricularia auricula-judae. Sample provenances: ARG Argentina,AUS Australia, CAN Canada, CHL Chile, CHN P.R. China, ECUEcuador, EST Estonia, FRA France, GER Germany, GUY Guyana,IND India, MEX Mexico, NOR Norway, REU Reunion, SPA Spain,USA United States of America
b
78 Mycol Progress (2008) 7:75–85

Mycol Progress (2008) 7:75–85 79

median sections, showed a narrow discontinuity (Fig. 1,parts 3 and 4). In one mycorrhiza, the intracellular hyphaehad thin walls (Fig. 1, part 1) and the dolipore was largeand truncate (Fig. 1, part 3). In the two other mycorrhizalroots, the hyphae had thicker walls (0.1 μm), with initials ofnew septa formation, and large amounts of glycogen rosettes(Fig. 1, parts 2, 4 and 5). All the other studied root samplescontained hyphae of Tulasnella spp. as indicated by wall-associated slime bodies and dome-shaped, imperforatedolipore caps with recurved ends (Suárez et al. 2006). NoAscomycetes were observed in cortical tissue, and Cerato-basidiales were only indicated once by the dolipore withdome-shaped caps with large perforations (not shown).
Phylogenetic analysis of Sebacinales associatedwith the epiphytic orchids
PCR products of Sebacinales were obtained by use ofspecific primers in 27% of the 83 orchid individualsinvestigated. The heuristic ML analysis and the twoindependent MCMC runs involving the nucLSU consis-tently yielded eight distinct sequence types (Fig. 2). Differ-ences between heuristic ML analysis and the twoindependent MCMC were mostly in the branch supportvalues, but some differences also occurred in the treetopologies. All sequence types appeared in Sebacinalessubgroup B (Fig. 2) (Weiß et al. 2004). Within subgroup B,all but sequence type 8 belong to a well-supported clade,which also includes sequences obtained from ericads, andSebacina vermifera sequences from Australian orchids(Fig. 2). The high heterogeneity made it difficult to obtaina reliable alignment from the ITS sequences for phyloge-netic analysis (data not shown). The 5.8S-ITS sequencesretrieved from the orchid roots for this study formed fourdistinct clades, which are separate from published ITSsequences from terrestrial orchids and available sequencesobtained from ericads (Fig. 3).
Comparison of Sebacinales and Tulasnellales mycobiontsof the same epiphytic orchids
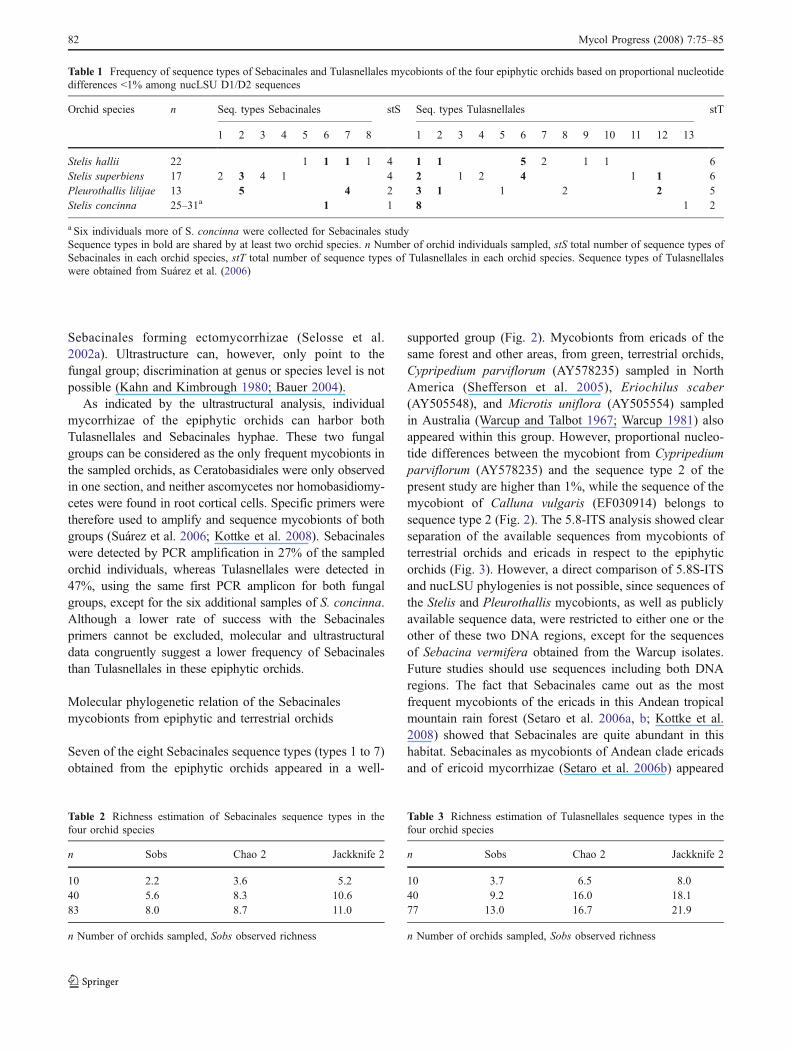
Four sequence types of Sebacinales were associated with S.hallii and S. superbiens, respectively, two sequence typeswere found in P. lilijae, and one in S. concinna mycorrhizae(Table 1). Six Tulasnella sequence types were previouslyfound as associated with S. hallii and S. superbiens, fivewith P. lilijae, and only two with S. concinna (Table 1).Sebacinales sequence types 1, 2 and 4 were shared amongat least two orchid species, similar situation was observedfor some sequence types of the Tulasnellales in ourprevious work (Table 1).
The sample-based rarefied accumulation curve with 95%confidence intervals did not reach its asymptote (not
shown). The observed richness of sequence types was8 in the case of the Sebacinales and 13 in the case of theTulasnellales (Tables 2 and 3). The difference between theobserved sequence type richness and the expected totalrichness was also higher in Tulasnellales than in Sebacinales(Chao2 and Jackknife2; Tables 2 and 3). In total, observedand expected diversity of Sebacinales and Tulasnellalessequence types of the four closely related orchid speciesappeared to be surprisingly high.
Stelis concinna was associated with only three sequencetypes of the Sebacinales and the Tulasnellales (Table 1), buttwo of these showed a wide altitudinal distribution range.Sebacinales sequence type 1, the only Sebacinales typedetected in S. concinna, was also found associated with S.hallii, occurring at the lowest elevation in forest type I, andTulasnellales sequence type 1 was associated with orchidsfrom all the altitudinal ranges (Table 1).
Discussion
Sebacinales are mycobionts of epiphytic Stelisand Pleurothallis species
The results obtained by a combination of ultrastructuralresearch and DNA sequencing revealed that Sebacinales aresymbiotically associated with the epiphytic Stelis andPleurothallis species. The ultrastructural features enableddiscrimination between Sebacinales and Tulasnellales in thecortical tissue of the individual mycorrhizae and confirmedthe mutualistic interaction as indicative of orchid mycor-rhizas (Peterson and Massicotte 2004) for Sebacinales, aswas previously shown for Tulasnellales in the same orchidmycorrhizae (Suárez et al. 2006). Sebacinoid hyphae were,however, only found in low numbers of the sectionedmycorrhizae (3 out of 20), while Tulasnellales wereobserved in all 20 mycorrhizae (Suárez et al. 2006),indicating a low frequency of Sebacinales in these epiphyticorchids. Similar pore caps as described here were found inSebacinales mycorrhizae of Andean ericads sampled in thesame habitat (Setaro et al. 2006a, b), in ericads from aworldwide distribution study (Selosse et al. 2007), and in
Fig. 3 Phylogenetic placement of Sebacinales sequences fromepiphytic orchid species Stelis hallii, Stelis superbiens, Stelis concinnaand Pleurothallis lilijae inferred by heuristic maximum-likelihood(ML) analysis from an alignment of 5.8S-ITS. Note that geneticdistances cannot be directly correlated to branch lengths in the tree,since highly diverse alignment regions were excluded for treeconstruction. Branch support values correspond to nonparametricML bootstrap (first numbers) and Bayesian Markov chain MonteCarlo analysis (second numbers) (only values exceeding 50% areshown, smaller values are indicated by an asterisk). The tree wasrooted with Sebacina allantoidea. Sample provenances: AUS Aus-tralia, ECU Ecuador, GER Germany, HUN Hungary
b
80 Mycol Progress (2008) 7:75–85
Mycol Progress (2008) 7:75–85 81
Sebacinales forming ectomycorrhizae (Selosse et al.2002a). Ultrastructure can, however, only point to thefungal group; discrimination at genus or species level is notpossible (Kahn and Kimbrough 1980; Bauer 2004).
As indicated by the ultrastructural analysis, individualmycorrhizae of the epiphytic orchids can harbor bothTulasnellales and Sebacinales hyphae. These two fungalgroups can be considered as the only frequent mycobionts inthe sampled orchids, as Ceratobasidiales were only observedin one section, and neither ascomycetes nor homobasidiomy-cetes were found in root cortical cells. Specific primers weretherefore used to amplify and sequence mycobionts of bothgroups (Suárez et al. 2006; Kottke et al. 2008). Sebacinaleswere detected by PCR amplification in 27% of the sampledorchid individuals, whereas Tulasnellales were detected in47%, using the same first PCR amplicon for both fungalgroups, except for the six additional samples of S. concinna.Although a lower rate of success with the Sebacinalesprimers cannot be excluded, molecular and ultrastructuraldata congruently suggest a lower frequency of Sebacinalesthan Tulasnellales in these epiphytic orchids.
Molecular phylogenetic relation of the Sebacinalesmycobionts from epiphytic and terrestrial orchids
Seven of the eight Sebacinales sequence types (types 1 to 7)obtained from the epiphytic orchids appeared in a well-
supported group (Fig. 2). Mycobionts from ericads of thesame forest and other areas, from green, terrestrial orchids,Cypripedium parviflorum (AY578235) sampled in NorthAmerica (Shefferson et al. 2005), Eriochilus scaber(AY505548), and Microtis uniflora (AY505554) sampledin Australia (Warcup and Talbot 1967; Warcup 1981) alsoappeared within this group. However, proportional nucleo-tide differences between the mycobiont from Cypripediumparviflorum (AY578235) and the sequence type 2 of thepresent study are higher than 1%, while the sequence of themycobiont of Calluna vulgaris (EF030914) belongs tosequence type 2 (Fig. 2). The 5.8-ITS analysis showed clearseparation of the available sequences from mycobionts ofterrestrial orchids and ericads in respect to the epiphyticorchids (Fig. 3). However, a direct comparison of 5.8S-ITSand nucLSU phylogenies is not possible, since sequences ofthe Stelis and Pleurothallis mycobionts, as well as publiclyavailable sequence data, were restricted to either one or theother of these two DNA regions, except for the sequencesof Sebacina vermifera obtained from the Warcup isolates.Future studies should use sequences including both DNAregions. The fact that Sebacinales came out as the mostfrequent mycobionts of the ericads in this Andean tropicalmountain rain forest (Setaro et al. 2006a, b; Kottke et al.2008) showed that Sebacinales are quite abundant in thishabitat. Sebacinales as mycobionts of Andean clade ericadsand of ericoid mycorrhizae (Setaro et al. 2006b) appeared
Table 1 Frequency of sequence types of Sebacinales and Tulasnellales mycobionts of the four epiphytic orchids based on proportional nucleotidedifferences <1% among nucLSU D1/D2 sequences
Orchid species n Seq. types Sebacinales stS Seq. types Tulasnellales stT
1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8 9 10 11 12 13
Stelis hallii 22 1 1 1 1 4 1 1 5 2 1 1 6Stelis superbiens 17 2 3 4 1 4 2 1 2 4 1 1 6Pleurothallis lilijae 13 5 4 2 3 1 1 2 2 5Stelis concinna 25–31a 1 1 8 1 2
a Six individuals more of S. concinna were collected for Sebacinales studySequence types in bold are shared by at least two orchid species. n Number of orchid individuals sampled, stS total number of sequence types ofSebacinales in each orchid species, stT total number of sequence types of Tulasnellales in each orchid species. Sequence types of Tulasnellaleswere obtained from Suárez et al. (2006)
Table 2 Richness estimation of Sebacinales sequence types in thefour orchid species
n Sobs Chao 2 Jackknife 2
10 2.2 3.6 5.240 5.6 8.3 10.683 8.0 8.7 11.0
n Number of orchids sampled, Sobs observed richness
Table 3 Richness estimation of Tulasnellales sequence types in thefour orchid species
n Sobs Chao 2 Jackknife 2
10 3.7 6.5 8.040 9.2 16.0 18.177 13.0 16.7 21.9
n Number of orchids sampled, Sobs observed richness
82 Mycol Progress (2008) 7:75–85

in the well-supported group that included the epiphyticorchid mycobionts (Fig. 2). A switch of the mycobiontsbetween ericads and orchids cannot be strictly excluded,although currently these two host groups do not share thesame sequence types (Kottke et al. 2008).
One sequence from S. hallii, referred to as sequence type8 (Fig. 2), fell in an uncertain position closest tomycobionts from liverworts Lophozia sudetica and Calyp-ogeia muelleriana from Europe (Kottke et al. 2003), and toericoid mycorrhizal fungi from several locations (Allen etal. 2003; Selosse et al. 2007). Mycobionts of the greenterrestrial orchids Eriochilus scaber, Cyrtostylis reniformis,Phyllanthus calycinus, Microtis uniflora, Caladenia spp.and Glossodia minor from Australia (Warcup 1981;Bougoure et al. 2005), Cypripedium californicum fromNorth America, and Epipactis palustris from Europe(Shefferson et al. 2005; Bidartondo et al. 2004) alsoappeared in the Sebacinales subgroup B. At the currentstage, phylogeny of Sebacinales subgroup B is not wellenough resolved to draw detailed conclusions on theirphylogenetic relationships.
Sebacinales subgroup A (Weiß et al. 2004) appears asthe sister group to subgroup B in our nucLSU tree.Subgroup A forms ectomycorrhizae with diverse treespecies (Richard et al. 2005; Tedersoo et al. 2006; Selosseet al. 2002a; Avis et al. 2003; Kennedy et al. 2003; Walkerand Parrent 2004; Glen et al. 2002) and establishes tripartiteassociations with achlorophyllous and mixotrophic orchids(Warcup 1988; Selosse et al. 2002b; Taylor et al. 2003;Selosse et al. 2004; Urban et al. 2003; McKendrick et al.2002; Julou et al. 2005). Sebacinales appearing in subgroupA or in subgroup B probably correlate with their mycor-rhizal interaction types. Our nucLSU tree shows that evenorchid species from the same genus are specifically linkedto fungi that presumably represent different mycorrhizaltype. The Sebacinales mycobionts of Epipactis palustris(AY634132) and of Cypripedium californicum (AY578234)appear in Sebacinales subgroup B, whereas the Sebacinalesmycobiont of Epipactis helleborine (AY452674) andCypripedium fasciculatum (AY578232), both commonforest understorey species, are in the Sebacinales subgroupA. Epipactis helleborine is a mixotrophic orchid associatedwith ectomycorrhiza forming fungi (Bidartondo et al. 2004;Julou et al. 2005).
Functional comparison of Sebacinales and Tulasnellalesassociated with the epiphytic orchids
The mountain rain forest in the study site shows highrelative humidity offering extraordinarily good conditionsfor epiphytes (Beck et al. 2008). Sebacinales and Tulasnel-lales, considering their saprotrophic capabilities (Roberts1999), are expected to grow on and in the tree bark if
humidity is more or less permanently sufficient. During thedry months, the epiphytic orchids may suffer from watershortage. Mycobionts should then be able to survive andimprove water uptake and nutrient supply (Yoder et al.2000). The fungi, on the other side, may profit from thewater retention capacity of the velamen covering the rootsof epiphytic orchids. The particular conditions in theepiphytic habitat possibly forced adaptation of specificgroups of Sebacinales and Tulasnellales. The observedsharing of mycobionts between the three Stelis and onePleurothallis species could then be partly explained by theirclose phylogenetic relationships (Pridgeon and Chase2001). Sharing of mycobionts will improve the rate ofsuccessful germination of orchid seeds and also promoteco-existence of closely related species as observed in thetropical mountain rain forest.
Species definition based on sequence variation iscurrently not possible in Sebacinales and Tulasnellales.Recently, McCormick et al. (2006) suggested sequencedifferences around 2% in the 5.8S-ITS region to separatepure culture isolates of Tulasnella, which in our case wouldhave yielded a very high number of Tulasnellales andSebacinales sequence types. rDNA substitution rates areconsidered higher in the Tulasnellales than in the Sebaci-nales (Bidartondo et al. 2003; Moncalvo et al. 2007), but itis not yet possible to define a precise percentage ofnucleotide differences suitable to define meaningful se-quence types for each fungal group. We pragmaticallydecided to use proportional nucleotide differences of <1%between nucLSU D1/D2 sequences to define sequencetypes for a tentative comparison of “species” richness ofSebacinales and Tulasnellales mycobionts of the orchids.
Our previous investigation (Suárez et al. 2006) revealedsome differences among the four orchid species in thenumber of Tulasnellales sequence types. We found a similarsituation for the Sebacinales mycobionts. The lowernumber of sequence types associated with P. lilijae maybe explained by the lower number of samples collected ofthis species, but in the case of S. concinna the highestnumber of plant individuals was investigated and still thelowest number of fungi detected. Most likely, fewer fungiare adapted to the more exposed conditions of S. concinnaoccurring in the upper part of the altitudinal range (2,100 ma.s.l., forest type III, microphyll forest), which is moreexposed to wind and changes in air humidity (Bendix et al.2008).
A larger dataset including mycobionts of terrestrialorchids from the same area will be required to test thehypothesis that Sebacinales and Tulasnellales radiated inthe Andean tropical mountain rain forest together with theepiphytic pleurothallid orchids. Switches from terrestrial toepiphytic habitat or back were found to be major drivingforces in radiation and speciation of orchids (Cameron
Mycol Progress (2008) 7:75–85 83

2005). Parallel radiation could occur for the mycobionts,but no arguments can be given in favor of this presumptionas long as information on mycobionts of terrestrial orchidsin the tropical mountain rain forest of Ecuador is missing.
Acknowledgements This research was funded by the DeutscheForschungsgemeinschaft (DFG project FOR 402). We thank theFundación Científica San Francisco for providing research facilities,Lorena Endara for help in orchid identification and Paulo Herrera forhis help in laboratory work.
References
Allen TR, Millar T, Berch SM, Berbee ML (2003) Culturing anddirect DNA extraction find different fungi from the same ericoidmycorrhizal roots. New Phytol 160:255–272
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-Blast: a newgeneration of protein database search programs. Nucleic AcidsRes 25:3389–3402
Avis PG, McLaughlin DJ, Dentinger BC, Reich PB (2003) Long-termincrease in nitrogen supply alters above- and below-groundectomycorrhizal communities and increases the dominance ofRussula spp. in a temperate oak savanna. New Phytol 160:239–253
Bauer R (2004) Basidiomycetous interfungal cellular interactions - asynopsis. In: Agerer R, Piepenbring M, Blanz P (eds) Frontiers inbasidiomycote mycology. IHW, Eching, Germany, pp 325–337
Beck E, Makeschin F, Haubrich F, Richter M, Bendix J, Valarezo C(2008) The ecosystem (Reserva Biológica San Francisco). In: BeckE, Bendix J, Kottke I, Makeschin F, Mosandl R (eds) Gradients ina tropical mountain ecosystem of Ecuador. Ecological Studies,no.198. Springer, Berlin Heidelberg New York, pp 1–14
Bendix J, Rollenbeck R, Reudenbach C (2006) Diurnal patterns ofrainfall in a tropical Andean valley of southern Ecuador as seenby a vertically pointing K-band Doppler Radar. Int J Climatol26:829–847
Bendix J, Rollenbeck R, Richter M, Fabian P, Emck P (2008) Climate. In:Beck E, Bendix J, Kottke I, Makeschin F, Mosandl R (eds)Gradients in a tropical mountain ecosystem of Ecuador. EcologicalStudies, no.198. Springer, Berlin Heidelberg New York, pp 63–74
Bidartondo MI, Bruns TD, Weiß M, Sergio S, Read DJ (2003)Specialized cheating of the ectomycorrhizal symbiosis by anepiparasitic liverwort. Proc R Soc London B 270:835–842
Bidartondo MI, Burghardt B, Gebauer G, Bruns TD, Read DJ (2004)Changing partners in the dark: isotopic and molecular evidenceof ectomycorrhizal liaisons between forest orchids and trees. ProcR Soc Lond B 271:1799–1806
Bougoure JJ, Bougoure DS, Cairney JW, Dearnaley JD (2005) ITS-RFLP and sequence analysis of endophytes from Acianthus,Caladenia and Pterostylis (Orchidaceae) in southeastern Queens-land. Mycol Res 109:452–460
Cameron KM (2005) Leave it to the leaves: a molecular phylogeneticstudy of Malaxideae (Epidendroideae, Orchidaceae). Am J Bot96:1025–1032
Colwell RK (2004) EstimateS: statistical estimate of species richnessand shared species from samples. Version 7. Persistent URL:purl.oclc.org/estimates
Cullings KW (1994) Molecular phylogeny of the Monotropoideae(Ericaceae) with a note on the placement of the Pyroloideae. JEvol Biol 7:501–516
Currah RS, Zelmer CD, Hambleton S, Richardson KA (1997) Fungifrom orchid mycorrhizae. In: Arditti J, Pridgeon AM (eds)Orchid biology: reviews and perspectives, VII. Kluwer,Dordrecht, Netherlands, pp 117–170
Felsenstein J (1985) Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 39:783–791
Gardes M, Bruns D (1993) ITS primers with enhanced specifity forbasidiomycetes–application to the identification of mycorrhizaeand rusts. Mol Ecol 2:113–118
Gascuel O (1997) BIONJ: An improved version of the NJ algorithm basedon a simple model of sequence data. Mol Biol Evol 14:685–695
Glen M, Tommerup IC, Bougher NL, O, Brien PA (2002) AreSebacinaceae common and widespread ectomycorrhizal associatesof Eucalyptus species in Australian forests? Mycorrhiza 12:243–247
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithmto estimate large phylogenies by maximum likelihood. Syst Biol52:696–704
Homeier J (2004) Baumdiversität, Waldstruktur und Wachstumsdynamikzweier tropischer Bergregenwälder in Ecuador und Costa Rica.Dissertationes Botanicae, vol. 391, Cramer, Berlin
Homeier J, Werner FA, Breckle S-W, Gradstein SR, Richter M (2008)Potential vegetation and floristic composition of Andean forestsin South Ecuador, with a focus on the RBSF. In: Beck E, BendixJ, Kottke I, Makeschin F, Mosandl R (eds) Gradients in a tropicalmountain ecosystem of ecuador. Ecological Studies, no.198.Springer, Berlin Heidelberg New York, pp 87–100
Julou T, Burghardt B, Gebauer G, Berveilleir D, Damesin C, SelosseM-A (2005) Mixotrophy in orchids: insight from a comparativestudy of green individuals and non-photosynthestic individuals ofCephalanthera damasonium. New Phytol 166:639–653
Kahn SR, Kimbrough JW (1980) Septal ultrastructure in some generaof the Tremellaceae. Can J Bot 58:55–60
Karnovsky MJ (1965) A formaldehyde glutaraldehyde fixation of highosmolarity for use in electron microscopy. J Cell Biol 27:137–138
Katoh K, Kuma K, Toh H, Miyata T (2005) MAFFT version 5:improvement in accuracy of multiple sequence alignment.Nucleic Acids Res 33:511–518
Kennedy PG, Izzo AD, Bruns TD (2003) There is high potential forthe formation of common mycorrhizal networks between under-storey and canopy trees in a mixed evergreen forest. J Ecol91:1071–1080
Kottke I, Beiter A, Weiss M, Haug I, Oberwinkler F, Nebel M (2003)Heterobasidiomycetes form symbiotic associations with hepatics:Jungermanniales have sebacinoid mycobionts while Aneurapinguis (Metzgeriales) is associated with a Tulasnella species.Mycol Res 107:957–968
Kottke I, Haug I, Setaro S, Suárez JP, Weiß M, Preußing M, Nebel M,Oberwinkler F (2008) Guilds of mycorrhizal fungi and theirrelation to trees, ericads, orchids and liverworts in a neotropicalmountain rain forest. Basic Appl Ecol 9:13–23
Ma M, Tan TK, Wong SM (2003) Identification and molecularphylogeny of Epulorhiza isolates from tropical orchids. MycolRes 107:1041–1049
McCormick M, Whigham DF, Sloan D, O, Malley K, Hodkinson B(2006) Orchid-fungus fidelity: a marriage meant to last? Ecology87:903–911
McKendrick SL, Leake JR, Taylor DL, Read DJ (2002) Symbioticgermination and development of the myco-heterotrophic orchidNeottia nidus-avis in nature and its requirement for locallydistributed Sebacina spp. New Phytol 154:233–247
Moncalvo J-M, Nilsson RH, Koster B, Dunham SM, Bernauer T,Matheny PB, McLenon T, Margaritescu S, Weiß M, Garnica S,Danell E, Langer G, Langer E, Larsson E, Larsson K-H, VilgalysR (2007) [“2006”] The cantharelloid clade: dealing with
84 Mycol Progress (2008) 7:75–85

incongruent gene trees and phylogenetic reconstruction methods.Mycologia 98:937–948
Otero JT, Ackerman JD, Bayman P (2002) Diversity and hostspecificity of endophytic Rhizoctonia-like fungi from tropicalorchids. Am J Bot 89:1852–1858
Otero JT, Ackerman JD, Bayman P (2004) Differences in mycorrhizalpreferences between two tropical orchids. Mol Ecol 13:2393–2404
Paulsch A, Piechowski D, Müller-Hohenstein K (2008) Forestvegetation structure along an altitudinal gradient in southernEcuador. In: Beck E, Bendix J, Kottke I, Makeschin F, MosandlR (eds) Gradients in a tropical mountain ecosystem of Ecuador.Ecological Studies, no.198. Springer, Berlin Heidelberg NewYork, pp 113–122
Pereira OL, Rollemberg CL, Borges AC, Matsuoka K, Kasuya MCM(2003) Epulorhiza epiphytica sp. nov. isolated from mycorrhizalroots of epiphytic orchids in Brazil. Mycoscience 44:153–155
Pereira OL, Kasuya MCM, Borges AC, Fernandes de Araújo E (2005)Morphological and molecular characterization of mycorrhizalfungi isolated from neotropical orchids in Brazil. Can J Bot83:54–65
Peterson RL, Massicotte HB (2004) Exploring structural definitions ofmycorrhizas, with emphasis on nutrient-exchange interfaces. CanJ Bot 82:1074–1088
Pridgeon AM, Chase MW (2001) A phylogenetic reclassification ofPleurothallidinae (Orchidaceae). Lindleyana 16:235–271
Richard F, Millot S, Gardes M, Selosse M-A (2005) Diversity andspecificity of ectomycorrhizal fungi retrieved from an old-growthMediterranean forest dominated by Quercus ilex. New Phytol166:1011–1023
Roberts P (1999) Rhizoctonia-forming fungi: a taxonomic guide.Royal Botanic Gardens, Kew
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phyloge-netic inference under mixed models. Bioinformatics 19:1572–1574
Selosse M-A, Bauer R, Moyersoen B (2002a) Basal Hymenomycetesbelonging to the Sebacinaceae are ectomycorrhizal on temperatedeciduous trees in silva: microscopical and molecular evidence.New Phytol 155:183–195
Selosse M-A, Weiß M, Jany JL, Tillier A (2002b) Communities andpopulations of sebacinoid basidiomycetes associated with theachlorophyllous orchid Neottia nidus-avis (L.) L.C.M. Rich. andneighbouring tree ectomycorrhizae. Mol Ecol 11:1831–1844
Selosse M-A, Faccio A, Scappaticci G, Bonfante P (2004) Chlorophyl-lous and achlorophyllous specimens of Epipactis microphylla(Neottieae, Orchidaceae) are associated with ectomycorrhizalseptomycetes, including truffles. Microb Ecol 47:416–426
Selosse M-A, Setaro S, Glatard F, Richard F, Urcelay C, Weiß M(2007) Sebacinales are common mycorrhizal associates ofEricaceae. New Phytol 174:864–878
Setaro S, Oberwinkler F, Kottke I (2006a) Anatomy and ultrastructureof mycorrhizal associations of neotropical Ericaceae. MycolProgr 5:243–254
Setaro S, Weiß M, Oberwinkler F, Kottke I (2006b) Sebacinales formectendomycorrhizae with Cavendishia nobilis, a member of the
Andean clade of Ericaceae, in the mountain rain forest ofsouthern Ecuador. New Phytol 169:355–365
Shefferson RP, Weiß M, Kull T, Taylor L (2005) High specificitygenerally characterizes mycorrhizal association in rare lady’sslipper orchids, genus Cypripedium. Mol Ecol 14:613–626
Spurr AR (1969) A low viscosity epoxy resin embedding medium forelectron microscopy. J Ultrastruct Res 26:31–43
Suárez JP, Weiß M, Abele A, Garnica S, Oberwinkler F, Kottke I(2006) Diverse tulasnelloid fungi form mycorrhizae with epi-phytic orchids in an Andean cloud forest. Mycol Res 110:1257–1270
Swofford DL (2002) PAUP*4.0 Phylogenetic analysis using parsimo-ny (*and other methods). Sinauer, Sunderland, Mass., USA
Swofford DL, Olsen GJ, Waddell PJ, Hillis DM (1996) PhylogeneticInference. In: Hillis DM, Moritz C, Mable BK (eds) Molecularsystematics. Sinauer, Sunderland, Mass., USA, pp 407–514
Taylor DL, Bruns TD, Szaro TM, Hodges SA (2003) Divergence inmycorrhizal specialization with Hexalectris spicata (Orchid-aceae), a nonphotosynthetic desert orchid. Am J Bot 90:1168–1179
Tedersoo L, Suvi T, Larsson E, Kõljalg U (2006) Diversity andcommunity structure of ectomycorrhizal fungi in a woodedmeadow. Mycol Res 110:734–748
Urban A, Weiß M, Bauer R (2003) Ectomycorrhizae involvingsebacinoid mycobionts. Mycol Res 107:3–14
Vilgalys R, Hester M (1990) Rapid genetic identification and mappingof enzymatically amplified ribosomal DNA from several Cryp-tococcus species. J Bacteriol 172:4238–4246
Walker JF, Parrent JL (2004) Molecular phylogenetic evidence for themycorrhizal status of Tremellodendron (Sebacinaceae). Mem NY Bot Gard 89:291–296
Warcup JH (1981) The mycorrhizal relationships of Australianorchids. New Phytol 87:371–381
Warcup JH (1988) Mycorrhizal associations of isolates of Sebacinavermifera. New Phytol 110:227–231
Warcup JH, Talbot PHB (1967) Perfect states of rhizoctoniasassociated with orchids I. New Phytol 66:631–641
Weiß M, Selosse M-A, Rexer K, Urban A, Oberwinkler F (2004)Sebacinales: a hitherto overlooked cosm of heterobasidiomyceteswith a broad mycorhizal potential. Mycol Res 108:1003–1010
White TJ, Bruns TD, Lee SB, Taylor JW (1990) Analysis ofphylogenetic relationships by amplification and direct sequencingof ribosomal RNA genes. In: Innis MA, Gelfand DH, SninskyJN, White TJ (eds) PCR protocols: a guide to methods andapplications. Academic, New York, pp 315–322
Williams PG, Thilo E (1989) Ultrastructural evidence for the identityof some multinucleate rhizoctonias. New Phytol 112:513–518
Yoder JA, Zettler LW, Stewart SL (2000) Water requirements ofterrestrial and epiphytic orchid seeds and seedlings, and evidencefor water uptake by means of mycotrophy. Plant Sci 156:145–150
Zotz G, Hietz P (2001) The physiological ecology of vascularepiphytes: current knowledge, open questions. J Exp Bot52:2067–2078
Mycol Progress (2008) 7:75–85 85




















