
Thinking About Morality With Orchids
22
Jonathan G. Wald Ethics, Agency, and Aesthetics in the Anthropocene Berkeley, California April 17, 2015 Thinking About Morality With Orchids Introduction The only appropriate way to begin my argument is with an orchid, or at the very least the image of one (slide 2). This is angraceum sesquipedale, otherwise known as “Darwin’s Orchid.” Darwin was fascinated by orchids to such an extent that he dedicated an entire book to their subject, On The Various Contrivances by Which British and Foreign Orchids are Fertilised by Insects, published in 1862. Yet of all the orchids that drew Darwin’s attention, only this one has the honor of being remembered as “Darwin’s” orchid. This flower earned that distinction by playing a unique role in the development of evolutionary theory. Darwin’s orchid has a long spur that can sometimes extend over a foot long. The spur contains a small basin of aromatic nectar, yet its remarkable length prevents most insects from reaching it. When Darwin received a sample of the orchids from James Bateman, he was perplexed by this feature. The model of coevolution he had considered in the evolution of plants required that some insect would be drawn to the nectar, thus carrying pollen from the orchid to other plants and vice versa, facilitating pollination. However, at the time, naturalists were not aware of any insect with a proboscis thin and small enough to find visiting Darwin’s orchid worthwhile. Despite this mystery, Darwin theorized that such an insect must exist in order to maintain the coherence of his model. This gap in knowledge remained until 1903, when naturalists discovered xanthopan morganii (slide 3), a hawk moth with a foot-long proboscis, that lived in Madagascar with the orchids and, in accordance with Darwin’s prediction, fed on its nectar (Arditti et al. 2012). I have started with this story to demonstrate the form of the argument I will make today; namely that the form of a plant can provide clues to the existence of entities beyond itself. While
Transcript of Thinking About Morality With Orchids

Jonathan G. Wald Ethics, Agency, and Aesthetics in the Anthropocene Berkeley, California April 17, 2015
Thinking About Morality With Orchids
Introduction
The only appropriate way to begin my argument is with an orchid, or at the very least the
image of one (slide 2). This is angraceum sesquipedale, otherwise known as “Darwin’s Orchid.”
Darwin was fascinated by orchids to such an extent that he dedicated an entire book to their
subject, On The Various Contrivances by Which British and Foreign Orchids are Fertilised by
Insects, published in 1862. Yet of all the orchids that drew Darwin’s attention, only this one has
the honor of being remembered as “Darwin’s” orchid.
This flower earned that distinction by playing a unique role in the development of
evolutionary theory. Darwin’s orchid has a long spur that can sometimes extend over a foot long.
The spur contains a small basin of aromatic nectar, yet its remarkable length prevents most
insects from reaching it. When Darwin received a sample of the orchids from James Bateman, he
was perplexed by this feature. The model of coevolution he had considered in the evolution of
plants required that some insect would be drawn to the nectar, thus carrying pollen from the
orchid to other plants and vice versa, facilitating pollination. However, at the time, naturalists
were not aware of any insect with a proboscis thin and small enough to find visiting Darwin’s
orchid worthwhile. Despite this mystery, Darwin theorized that such an insect must exist in order
to maintain the coherence of his model. This gap in knowledge remained until 1903, when
naturalists discovered xanthopan morganii (slide 3), a hawk moth with a foot-long proboscis,
that lived in Madagascar with the orchids and, in accordance with Darwin’s prediction, fed on its
nectar (Arditti et al. 2012).
I have started with this story to demonstrate the form of the argument I will make today;
namely that the form of a plant can provide clues to the existence of entities beyond itself. While

Wald 2
Darwin used this fact to predict the existence of xanthopan morganii, I will argue that the forms
of contemporary orchids and corn provide clues about the moral values that shaped their
evolution. Consequently, identifying the biosemiotic impact of morals suggests that moral values
themselves may have importantly material elements. I will begin by laying out the basic
principles of biosemiotics as they may relate to normativity before proceeding to an analysis of
the two plants. In concluding, I will comment on the implications of this approach to the future
empirical study of morality.
Concepts
Biosemiotics is the study of biological processes and organisms through the concepts of
signification, information, and reference. Upon this understanding, all living processes
fundamentally interpret elements in their surroundings. This approach can include the study of
animal perception, as in Uexküll’s foundational elaboration on the concept of umwelt (Uexküll
2010), or examples like Darwin’s “reading” of orchids to understand something about their
ecosystem. Semiotics can elaborate species-level phenomenon like the evolution of an anteater’s
snout to fit the particular form of an anthill to the microscopic analysis of how hemoglobin’s
structure perfectly fits an oxygen molecule (Deacon 2013:9).
All of these features share what Terrence Deacon calls a “constitutive absence.” In other
words, “Such concepts as information, function, purpose, meaning, intention, significance,
consciousness, and value are intrinsically defined by their fundamental incompleteness. They
exist only in relation to something they are not.” (Deacon 2013:23) An anteater does not contain
an anthill. However, in order to make sense of the shape of the anteater’s snout, one must realize
that it is the negative image of the long, narrow passage of the ant’s tunnel (Kohn 2013:74).

Wald 3
Biosemiotics thus arrives at a limited form of teleology. By reinscribing function into
biology, i.e. studying hemoglobin in order to understand oxygen transfer, biological entities can
be said to succeed or fail at a particular goal. For example, in the case of Darwin’s orchid, we
may realize that xanthopan morganii’s long proboscis serves the organism by allowing it to
access the orchid’s nectar. If a particular moth’s proboscis were too short or too wide, it would
be reasonable to say that the proboscis “failed” at its purpose. Over the course of prolonged
natural selection, “functional” or “successful” forms will be selected, leading to the inscription of
this ends-based teleology into the form of the organism.
Tantalizingly, a group of biosemioticians led by Kalevi Kull has labeled this process
“minimal normativity,” by which they refer to the way in which a biological organism can
succeed or fail to functionally represent their surroundings (Kull et al. 2009:171). Taking
Eduardo Kohn’s challenge to “situate morality ontologically,” (Kohn 2013:133), I will attempt to
see how far we can extend the concept of “minimal normativity” into the domain of moral
systems. If biological systems represent the world around them, what happens when human
interests, desires, and motivations constitute their world? In other words, how does morality
shape life itself?
Let me clarify what I mean by “morality,” as although this term has received increased
attention in anthropology, the discipline has not yet settled on a shared meaning. Like a number
of anthropologists of morality, particularly James Faubion and James Laidlaw (Faubion 2011;
Laidlaw 2014), I draw on the Foucauldian tradition in this regard, and thus implicitly on
Nietzsche as well. In this framework, “morality,” refers to “a set of values and rules of action
that are recommended to individuals through the intermediary of various prescriptive agencies
such as the family (in one of its roles), educational institutions, churches, and so forth.”

Wald 4
(Foucault 1988:25). This understanding of morality is importantly distinct from “ethics,” which
refers to the manners in which a subject forms themselves in relation to moral injunctions.
Within the anthropology of morality, this ethical portion of self-formation has garnered a greater
degree of attention than the impersonal abstraction of morality.
In order to make the connection between “minimal normativity” and “morality,” it is
important to note that the Manichean concepts of “Good” and “Evil” are entirely secondary to
the more foundational sense of “morality.” Rather, as Nietzsche argues, the primordial binary is
between “good” and “bad” (Nietzsche 1989:40). If we take morality to refer to the diverse modes
by which individuals are prescribed sets of “good” and “bad” behavior, the concept of “minimal
normativity” appears to be a simple translation of morality into the generalized domain of life via
the modes by which an organism succeeds or fails to adapt to its surroundings. In cases where
these surroundings are primarily anthropogenic, this distinction fades.
Grains
To this end, few plants provide as detailed a test case as corn, the focal plant for much of
the development of modern agriculture. As Michael Pollan notes, “Few plants can manufacture
quite as much organic matter (and calories) from the same quantities of sunlight and water and
basic elements as corn.” (Pollan 2007:22). This peculiar quality has made corn a staple of
alimentary production as well as contemporary biotechnical research. However, the corn that we
all know so well is quite distinct from its pre-human iterations. As this comparison shows (slide
4), corn has been selectively bred far away from its close relative teosinte. Paleoethnobotanists
have long been aware of the ways in which corn has evolved as a result of human pressures in
order to maximize the production of sugars and other materials (Cummings 2014). The
importance of corn to contemporary biotechnology is due to a combination of these human

Wald 5
interventions, but these alone would not explain why, of all plants, corn has risen to such
dominance in agricultural production.
In order to make sense of corn’s unnatural history, it is helpful to note the particular place
that agricultural grains have held in modern history. While biopolitics may have reached its end
with the gutting of social welfare programs, the end of the “social” as a domain of intervention in
favor of the “global” or the “individual,” and the development of new, neoliberal technologies of
governance (Rose 1996; Rabinow and Bennett 2012:157), there is little question that the forms
of biopolitics played an enormous role in the formation of contemporary modes of economic and
political governance. While many in medical anthropology have noted the strong links between
the genealogy of biopower, public health and sexuality, I would like to turn our attention towards
Foucault’s first example for the concepts of biopolitics: agricultural scarcity. In the second half
of the eighteenth century, “scarcity” appeared as a natural, artificial, and moral threat brought
about by environmental factors as well as human failures (Foucault 2009:30–31). Security, that
realist technology of power, initially arose as a technology in order to reduce the economic
uncertainty introduced by scarcity and was later expanded to include the apparatuses of the
police.
Obviously, Foucault was not an agricultural historian, and hence his brief foray into the
field quickly diverts into an analysis of the state. For my purposes, however, the securitization of
agriculture, understood as the realist strategies by which grain prices were regulated,
unmistakably appears in the legacy of corn evolution, both in the form of the plant itself and its
economic importance. In essence, corn has been intentionally made into a relatively easily
regulated plant, and the relative ease with which this project was accomplished accounts for its
widespread consumption (Cronon 1992:99).

Wald 6
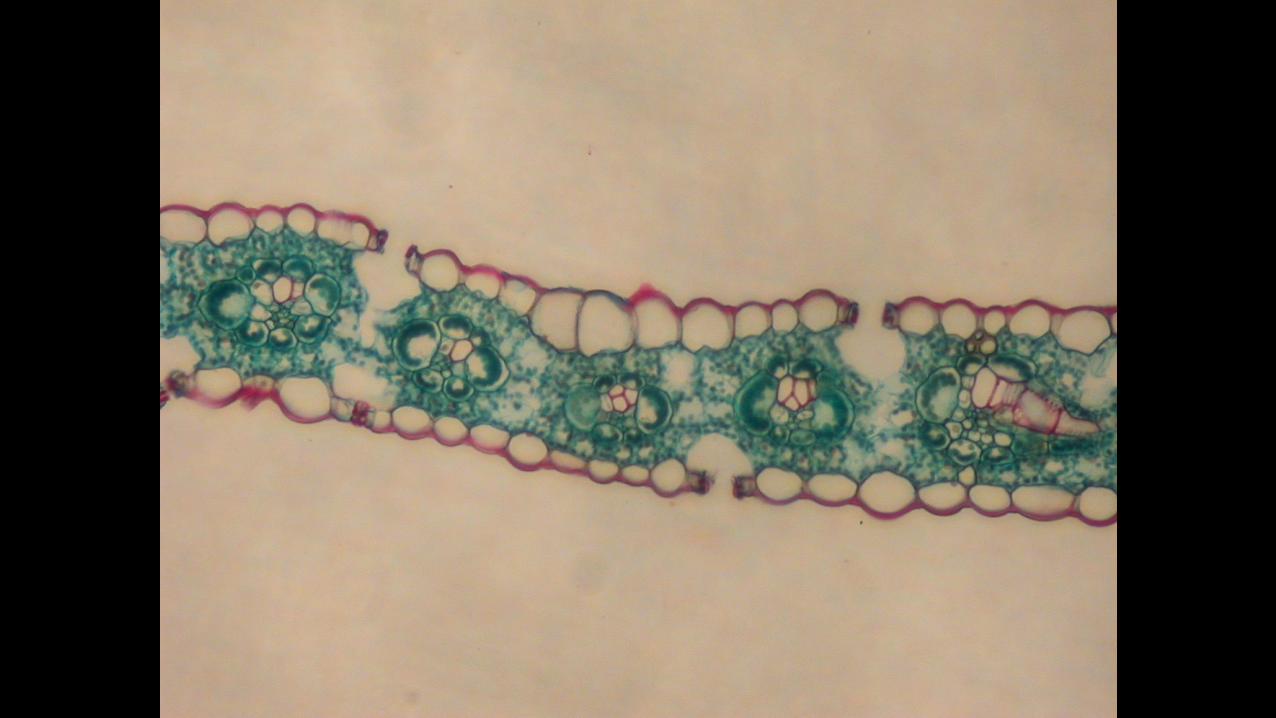
Consider corn’s metabolic pathway, C4 carbon fixation. In order to facilitate more
efficient photosynthesis, C4 plants uses an alternate enzyme to transport C02 through “bundle
sheath” arrangements in the leaf structure (slide 5). This process achieves the effective
production of sugars through a spatial rearranging of the cell but remains temporally static. So
long as light, air, and nutrients are available, corn is able to produce glucose. This trait makes
corn a quick grower and easily predictable in time, both traits that make it an exemplary crop for
the demands of regulated, securitized production.
In the contemporary period of biotechnological modification, engineers have been
working to extend these traits in corn. Contrary to the worries of “frankenplants” running
rampant with unpredictable modifications, one of the key tasks for genetic engineers has been to
reduce the pre-existing rate of genetic mutation in crops. In essence, the goal has been to make
plants more, not less predictable in order to regulate crop traits and farming methods (Murphy
2011). This process of regulating crop diversity corresponds with the trend towards the
“deskilling” of agricultural labor, a process begun in the last century that links with the
movement of knowledge of corn varieties from the tacit knowledge of the farmer to the
specialized knowledge of the biologist (Fitzgerald 1993). In short, corn is now an ideal crop for
economic concerns; it grows relatively reliably, efficiently, and with minimal specialized input.
Orchids
Orchids, on the other hand, could hardly be more different from the image of corn. While
grains have figured centrally in the formation of modern economics and have evolved into their
counterpart, orchids have long been something of agriculture’s “other.” Personally, I began this
research out of a sense of surprise. I had gone to Brazil hoping to understand the motivations
behind agricultural biotechnology, and in my first month there, I found myself surrounded by

Wald 7
orchids. While trying to find people working in biotechnology, I more or less stumbled across
three different centers for orchid cultivation (slides 6-8). The first image is from the greenhouse
for the Sociedade Orquidófila de Belo Horizonte, housed in a public park and open to anyone
interested in cultivating orchids. The second image is the orchid veranda at Inhotim, an art
museum and botanical garden, funded by the mining company Vale. Finally, I had a chance to
speak with biologists studying orchids and other plants at a research greenhouse at the
Universidade Federal do Minas Gerais.
It is immediately apparent how orchids have garnered so much attention over the years.
Even the highly trained and professional biologist I spoke with at UFMG would stop speaking
occasionally just to admire the aesthetic beauty of the flowers. Without much difficulty, I can
imagine Darwin acting in the same manner, nor is it difficult to see why author Susan Orlean, in
her bestselling book The Orchid Thief, notes that orchids “seem to drive people crazy. Those
who love them love them madly.” (Orlean 2000:50) The orchids themselves, through their
beauty, draw people into the study of botany, thus securing their position as one of the early
plants to attract biological interest.
As I tried to make sense of this attraction, or why I kept finding orchids when I was
looking for biotechnology, the more I realized that diversity appears as a pattern throughout the
biology of orchids. While famous for their beauty, not all orchids are much to look at. Some
contain only tiny, barely visible flowers. Some grow on the sides of trees like vines, with their
roots dangling in the air, while others stay low and close to the ground, easily mistaken for a
weed. Some have thick, trunk like features, while others spread out like moss. To my untrained
eye, nothing about orchid morphology appeared regular.

Wald 8
Orchids are also notoriously difficult to cultivate. Some species can take up to seven
years to flower, while many who have tried to cultivate them have been frustrated to find that a
slight change in light, temperature, or humidity can spell the difference between an orchid
growing or dying. Many species of orchid only grow symbiotically in complex ecosystems
within highly specific biomes, down to the species of bacteria that live within their petals.
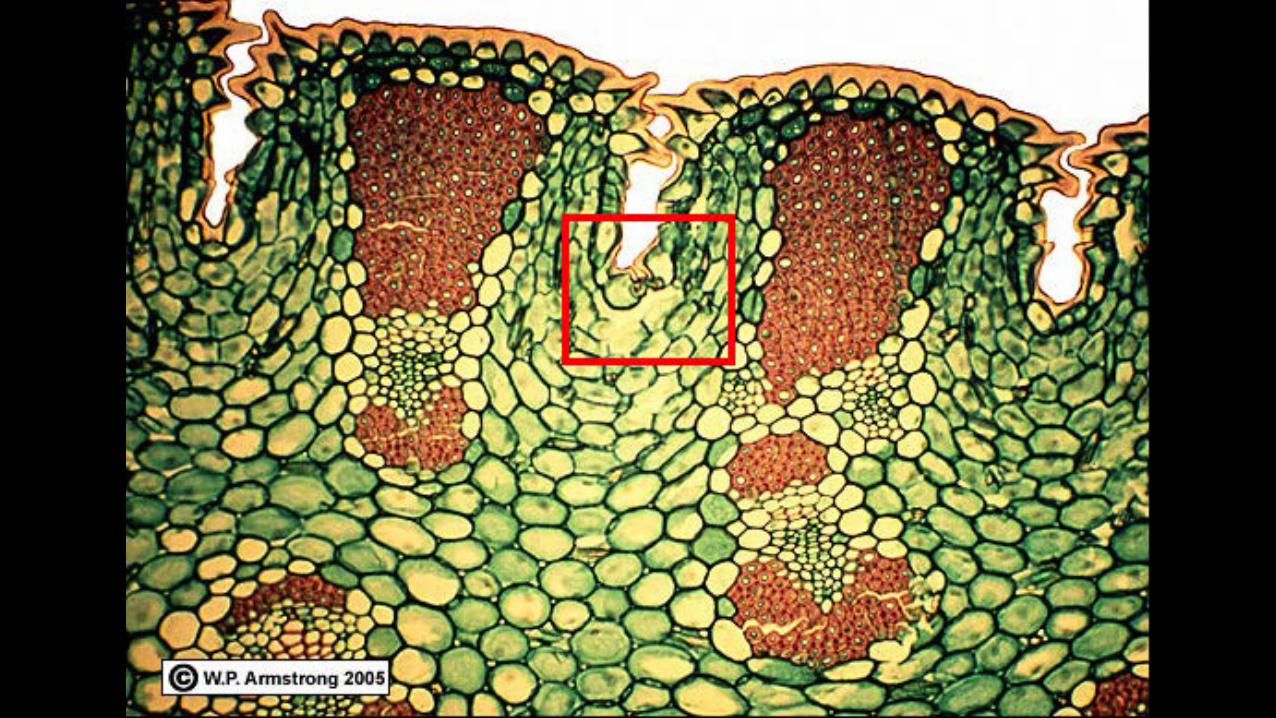
On the molecular level, orchids are also quite unusual (slide 9). Unlike most plants that
use C3 or C4 pathways, orchids are one of the rare plants to use CAM photosynthesis. This
process conserves water in arid regions by differentiating the plant structure not only spatially, as
in the C4 pathway, but also temporally. Orchids engage in different portions of the
photosynthetic pathway at night than they do during the day.
The intense diversity of orchids has resulted in them playing two primary roles in human
interactions. First, they are an ornamental and aesthetic plant. Botanists frequently hybridize
orchids in a style of art. For example, all of the orchids at Inhotim, the botanical garden, are
hybrids selected for maximum brilliance.
Second, orchid diversity opens up a number of possibilities for biologists attempting to
understand the diversity of botany. From their molecular structure to their phenotypical variation,
orchids have been a wealth of information for modern biology. It is no accident that Darwin, the
founder of modern biology, used orchids as one of his first test cases. In more recent years,
genomic technology has seized on orchids for their genetic diversity, especially hoping to
determine their rare procedure for CAM photosynthesis (Cai et al. 2015).
As a biologist was guiding me around UFMG’s orchid greenhouse (slide 10), she
explained that unlike the public gardens, the research centers avoided hybridization in order to
maintain genetic distinctions between the strains and to maximize the possibilities for research.

Wald 9
To this end, I noticed as she swept the dust-like pollen from a flower through the cracks in the
worn metal grate. While this kept the invading pollen away from the tagged and marked
specimens, I could not help but notice that an unruly green carpet littered the floor (slide 11),
remnants of past experiments. It struck me that this verdant laboratory stood in a chain with
sanitized workbenches in more typical laboratory settings.
While the aesthetic and scientific values of orchids may be conceptually distinct, there is
little doubt that the current orchid research owes much to a past of aesthetic valuation.
Throughout the history of colonialism, naturalists scouted conquered landscapes and reported
back about the rare and exotic plants they found (Grove 1996). The enormous capital
investments required to maintain museums like Inhotim or a public orchid greenhouse are only
the most contemporary legacy of orchid’s beauty garnering them a place before our gaze.
Furthermore, their delicate preservation and documentation, due to the valuation of their
appearance, has resulted in orchids maintaining a high degree of genetic and morphological
diversity in contrast with the biopolitical standardization of grains (slide 10).
Conclusion: Morality
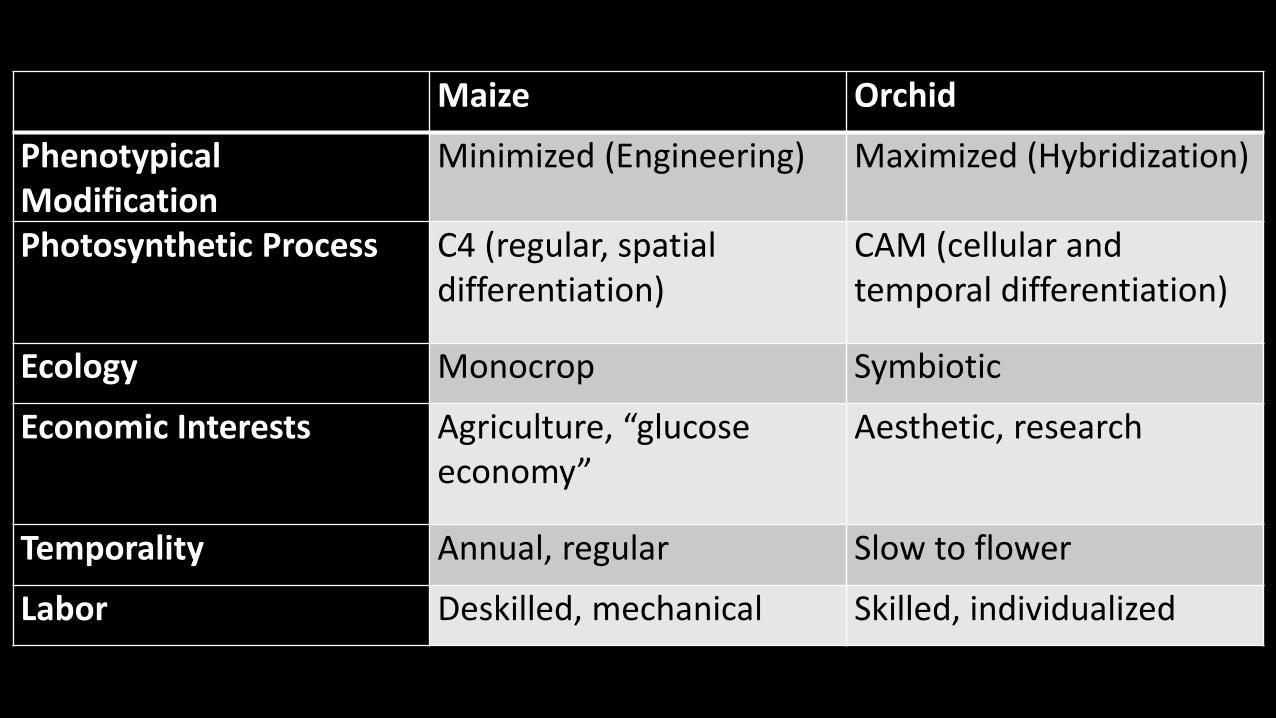
Two very different plants united by the shared fact that they have simultaneously drawn
in human interests and bear the stamps of those interactions; corn in its high regularity and
simplified growing process, orchids in their fragile ornateness and diversity. If biological
systems really do symbolize their world, it is clear that these two organisms exist in worlds
populated not only by environmental conditions and genetic heredity, but also “sets of values and
rules of action” pertaining to their cultivation, preservation, and modification. In other words,
they are in part the products of moral systems.

Wald 10
Ultimately, my argument is not for botanists attempting to understand plants, but for
anthropologists, philosophers, and other attempting to understand the nature of morality. If traces
of morality can be found in non-human, non-symbolic biological systems, then anthropologists
of morality must broaden their focus away from linguistic instances of evaluation or
anthropocentric conceptions of virtue and instead observe the ways in which morality itself
shapes a variety of different worlds populated by a plenitude of different beings. In other words,
my goal with this argument is to begin the task of opening our tools of moral thinking beyond the
confines of purely humanist anthropology. While moral values indisputably rely on human
agency to gain effect, I have shown that they may take on a material, enlivened existence as a
trace of their former instantiations and that the biological systems themselves may offer
possibilities and resistances to human moral actions. Biosemiotics provides only one conceptual
tool for the task of moral anthropology in that it opens the conceptual view to the ways in which
life itself adapts to the polyvalent world around it. Our task now is to determine the form and
content of those worlds.
Works Cited Arditti, Joseph, John Elliott, Ian J. Kitching, and Lutz T. Wasserthal 2012 “Good Heavens What Insect Can Suck It”– Charles Darwin, Angraecum Sesquipedale and Xanthopan Morganii Praedicta. Botanical Journal of the Linnean Society 169(3): 403–432. Cai, Jing, Xin Liu, Kevin Vanneste, et al. 2015 The Genome Sequence of the Orchid Phalaenopsis Equestris. Nature Genetics 47(1): 65–72. Cronon, William 1992 Nature’s Metropolis: Chicago and the Great West. Reprint edition. New York: W. W. Norton & Company. Cummings, Linda Scott 2014 Corn Cob Phytoliths as Indicators of Genetics and Environmental Conditions. In Ancient Plants and People. Marco Madella, Carla Lancelotti, and Manon Savard, eds. Pp. 292–300. Tuscon: University of Arizona Press.

Wald 11
Deacon, Terrence W. 2013 Incomplete Nature: How Mind Emerged from Matter. 1 edition. W. W. Norton & Company. Faubion, James D. 2011 An Anthropology of Ethics. New Departures in Anthropology. Cambridge ; New York: Cambridge University Press. Fitzgerald, Deborah 1993 Farmers Deskilled: Hybrid Corn and Farmers’ Work. Technology and Culture 34(2): 324–343. Foucault, Michel 1988 The Use of Pleasure. Robert Hurley, tran. 1st Vintage Books ed. New York: Vintage Books. 2009 Security, Territory, Population: Lectures at the Collège de France 1977--1978. Michel Senellart, François Ewald, Alessandro Fontana, and Arnold I. Davidson, eds. Graham Burchell, tran. New York: Picador. Grove, Richard H. 1996 Green Imperialism: Colonial Expansion, Tropical Island Edens and the Origins of Environmentalism, 1600-1860. Cambridge; New York: Cambridge University Press. Kohn, Eduardo 2013 How Forests Think: Toward an Anthropology Beyond the Human. Berkeley: University of California Press. Kull, Kalevi, Terrence Deacon, Claus Emmeche, Jesper Hoffmeyer, and Frederik Stjernfelt 2009 Theses on Biosemiotics: Prolegomena to a Theoretical Biology. Biological Theory 4(2): 167–173. Laidlaw, James 2014 The Subject of Virtue. Cambridge: Cambridge University Press. Murphy, Denis J 2011 Plants, Biotechnology and Agriculture. Wallingford: CABI. Nietzsche, Friedrich Wilhelm 1989 On the Genealogy of Morals. Walter Arnold Kaufmann and R. J Hollingdale, trans. New York: Vintage Books. Orlean, Susan 2000 The Orchid Thief: A True Story of Beauty and Obsession. New York: Ballantine Books.

Wald 12
Pollan, Michael 2007 The Omnivore’s Dilemma: A Natural History of Four Meals. New York: Penguin. Rabinow, Paul, and Gaymon Bennett 2012 Designing Human Practices: An Experiment with Synthetic Biology. Chicago ; London: The University of Chicago Press. Rose, Nikolas 1996 The Death of the Social? Re-Figuring the Territory of Government. Economy and Society 25(3): 327–356. Uexküll, Jakob von 2010 A Foray into the Worlds of Animals and Humans: With A Theory of Meaning. Joseph D. O’Neil, tran. Minneapolis: University of Minnesota Press.






Maize Orchid
Phenotypical Modification
Minimized (Engineering) Maximized (Hybridization)
Photosynthetic Process C4 (regular, spatial differentiation)
CAM (cellular and temporal differentiation)
Ecology Monocrop Symbiotic
Economic Interests Agriculture, “glucose economy”
Aesthetic, research
Temporality Annual, regular Slow to flower
Labor Deskilled, mechanical Skilled, individualized

Image Sources• Slide 2: Orchid Corner. 2009. Ursula’s Page. http://www.kammlott.net/Jan2009.html.
(Image)Slide 3: Xanthopan Morganii Praedicta• Slide 3: N.d. Encyclopedia Britannica.
http://www.britannica.com/EBchecked/topic/1315526/Xanthopan-morganii-praedicta, accessed March 31, 2015. (Image)
• Slide 4: 2015 Zea (Genus). Wikipedia. http://www.faculty.umb.edu/yvonne_vaillancourt/Biology/C3%20C4%20images.htm
• Slide 5: Vaillancourt, Yvonne. 2014 C3 and C4 Images. University of Massachusetts Boston. http://www.faculty.umb.edu/yvonne_vaillancourt/Biology/C3%20C4%20images.htm.
• Slide 7: A., Marcio. 2014. Vandário – Foto de Inhotim, Brumadinho - TripAdvisor. http://www.tripadvisor.com.br/LocationPhotoDirectLink-g1747395-d1743976-i94993263-Inhotim-Brumadinho_State_of_Minas_Gerais.html, accessed April 4, 2015.
• Slide 9: Armstrong, Wayne P. 2005. Ecological Principles #2. Wayne’s Word. http://waynesword.palomar.edu/lmexe10a.htm.




















