
Catechisms and Confessionarios: Distorting Mirrors of Andean Societies

84
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Kottke I, Suárez JP (2009) Mutualistic, root-inhabiting fungi of orchids identification and functional types.In: Pridgeon A M, Suarez JP (eds) Proceedings of the Second Scientific Conference on Andean Orchids.
Universidad Técnica Particular de Loja, Loja, Ecuador, pp 84-99.
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDSIDENTIFICATION AND FUNCTIONAL TYPES
Ingrid Kottke1 and Juan Pablo Suárez Chacón2
1 Eberhard-Karls-Universität Tübingen, Spezielle Botanik, Mykologie und Botanischer Garten, Auf der Mor-genstelle 1, D-72076 Tübingen, Germany
2 Centro de Biología Celular y Molecular, Universidad Técnica Particular de Loja, San Cayetano Alto s/n C.P. 1101608, Loja, Ecuador
ABSTRACT
Orchids depend on fungi for germination and protocorm development and maintain mycor-rhizas in their adult forms. Mycorrhizal fungi are therefore a driving force in orchid evolution and speciation. However, reliable data on fungal identities are crucial. Information from molecular phylo-genetics and transmission electron microscopy congruently revealed distinct fungal groups as orchid mycobionts. Sebacinales Group B, Tulasnellales, and Ceratobasidiales were found associated with ter-restrial orchids in open grasslands and arbuscular mycorrhizal forests and with epiphytes, whereas Sebacinales Group A, Thelephorales, Russulales, some Euagaricales, and Tuberales form mycorrhizas with terrestrial orchids in ectomycorrhizal forests. Enzyme and isotope analyses revealed that the for-mer obtain carbon from rotten organic material to nourish the protocorm, whereas the latter take car-bon from ectomycorrhizas of woody plants to supply nutrients to protocorms and adult heterotrophic or mixotrophic plants. Mycobionts of terrestrial orchids are of narrower host range than previously expected. The few investigations on mycobionts of epiphytic orchids indicate sharing of hosts. DNA-based fungal identities of mycobionts from tropical terrestrial and epiphytic orchids, host range, and inclusion of data from several ecological parameters are still needed to determine whether association strategies differ between epiphytic and terrestrial orchids or between temperate and tropical habitats.
METHODS FOR IDENTIFICATION OF ORCHID MYCOBIONTS
Orchid mycobionts were previously only known by isolation of the fungi from orchid roots or protocorms, description of hyphal features of the anamorphic state (Currah et al., 1997; Roberts, 1999), and occasional induction of the teleomorphs (Warcup and Talbot, 1966, 1967a,b, 1980; Cur-rah et al., 1987; Warcup, 1988). Re-infection experiments claimed separation of the true mycobiont isolates from endophytes of the velamen and other contaminants (Bernard, 1909; Brundrett, 2006;

85This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Bonnardeaux et al., 2007). Transmission electron microscopical studies of the isolates and in situ hyphae yielded distinctive features to discern and confirm the main systematic groups of orchid my-cobionts (Currah and Sherburne, 1992; Andersen, 1996; Gleason and McGee, 2001; Selosse et al., 2004, Suarez et al., 2006, 2008). Ultrastructural details are also needed to demonstrate that the fungi are forming pelotons (coils) inside vital protocorm and root cortical cells (Peterson and Massicotte, 2004). DNA–sequencing of the mycobionts directly from the mycorrhizas combined with molecular phylogenetics have yielded an unexpected richness of fungal genotypes including fungal groups previ-ously unknown as orchid mycobionts.
ORCHIDS DEPEND ON MYCOBIONTS
Orchids need fungi supplying carbohydrates and soil nutrients to promote growth of the non-photosynthetic protocorm stage (Bernard, 1909; Smith and Read, 1997). Similar mycobionts are ha-bitually harbored in root cortical cells of adult green and non-photosynthetic orchids (Fig. 1a). These fungi colonize not only the velamen and epidermal cells where many other fungi also find shelter (Fig. 1b) but penetrate the root exodermis via living passage cells (Fig. 1d ) and form coils (pelotons) in root cortical cells (Fig. 1c). Hyphae soon collapse and become encased by root cell wall material (Fig. 1c; Peterson et al., 1996). This type of interaction was found in both orchid protocorms and roots independent of orchid and mycobiont species involved (Dörr and Kollmann, 1969; Peterson and Cur-rah, 1990; Rasmussen, 1990; Selosse et al., 2004, Suarez et al., 2006, 2008; Abadie et al., 2006).
Plant access to carbon, nitrogen, and phosphorus via the fungal mycelium was demonstrated (Smith, 1967; Alexander et al., 1984; Alexander and Hadley, 1985; Gebauer and Meyer, 2003; Trudell et al., 2003; Bidartondo et al., 2004; Cameron et al., 2006, 2007), and bidirectional transfer of carbon between the green-leaved orchid Goodyera repens (L.) R.Br. and its mycobiont Ceratobasidium cornige-rum was proven in Petri dish microcosms, supporting the view of a true mutualistic interaction (Cam-eron et al., 2006). With respect to carbon acquisition, two functionally distinct groups of fungi were found associated with the orchids: mycobionts with saprophytic capabilities and mycobionts with ectomycorrhizal associations. Isolates of saprophytic orchid mycobionts displayed enzyme activities for degrading cellulose and other complex organic compounds (Hadley, 1969; Midgley et al., 2006), indicating access to carbon from humus, although this has so far not been demonstrated in nature. Ectomycorrhizal fungi supply carbon obtained from woody plant roots to the achlorophyllous, my-coheterotrophic or green, mixotrophic orchids (McKendrick et al., 2000a; Gebauer and Meyer, 2003; Trudell et al., 2003).
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES

86
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
ORCHID MYCOBIONTS AS IDENTIFIED BY MOLECULAR PHYLOGENYAND ULTRASTRUCTURAL CHARACTERS
For many years, only few and unspecific fungi, so-called Rhizoctonia species, isolated from or-chid mycorrhizas were considered as orchid mycobionts (Hadley, 1970). However, isolation of fungal DNA directly from the mycorrhizas followed by sequencing yielded an unexpected richness of geno-types of not only previously known orchid mycobiont groups but also unexpected ectomycorrhizal fungi. Obviously, many of the mycobionts do not grow on artificial media. Claims for re-infection to prove the mycobiont nature of the fungi can, therefore, not be strictly met. Isolation is also biased by large numbers of fungi, mostly ascomycetes that only colonize the velamen, epidermal cells or root hairs (Williamson and Hadley, 1970; Bayman and Otero, 2006).
Sequencing of mycobionts directly from the mycorrhizas and ultrastructural studies of hy-phae in orchid root cortical cells proved three groups of hymenomycetous heterobasidiomycetes -- the Sebacinales, Tulasnellales, and Ceratobasidiales -- and several groups of hymenomycetous homobasi-diomycetes, especially Thelephorales, Russulales, and Tuberales of Ascomycota as orchid mycobionts (Tables 1-4). Heterobasidiomycetes had already been indicated by teleomorphs induced in cultures by Warcup and Talbot (1966, 1967a,b, 1980) and Warcup (1988). However, molecular phylogenet-ics revealed that Sebacinales fall into two distinct groups, A and B (Weiss et al., 2004). Autotrophic orchids were found associated with Sebacinales group B, related to Sebacina vermifera sensu Warcup and Oberwinkler, whereas mycoheterotrophic or mixotrophic orchids growing with ectomycorrhiza forming trees are associated with Sebacinales group A, related to S. incrustans and S. epigaea (Table 1). Tulasnella calospora and T. violea sensu Warcup came out as polyphyletic, and related genotypes were found with numerous orchids (Table 2). Ceratobasidiales were also found with diverse orchids, but in situ proofs are rare (Gleason and McGee, 2001), and most authors worked on fungal isolates only (Table 3). Taxonomy within the three heterobasidomycete groups is unresolved, and taxon names cannot yet be provided. Although earlier findings pointed to a liaison of orchids with ectomycorrhiza-forming fungi (Warcup 1991; Zelmer and Currah, 1995), DNA-sequencing of orchid mycobionts directly from the mycorrhizas revealed that this is a widespread phenomenon of orchids in ectomycor-rhizal forests (Table 4). These orchids are either heterotrophic or mixotrophic, although the latter was not proven for every species so far (Table. 4).
Ultrastructural hyphal characters allow unambiguous distinctions among the four basidiomy-cete groups and the ascomycetes in root cortical cells and fungal isolates. Sebacinales are characterized by fine hyphae with electron-dense cell walls and dolipores with imperforate, straight or dish-shaped pore caps (Fig. 2a; Andersen 1996; Gleason and McGee, 2001; Setaro et al., 2006; Suarez et al., 2008). Tulasnellales possess dolipores with imperforate, dish-shaped pore caps with recurved ends (Fig. 2b) and slime bodies in the cell walls (Fig. 2c), becoming electron-dense during aging (Fig. 2d; Ander-sen, 1996; Gleason and McGee, 2001; Suarez et al., 2006). Ceratobasidiales are distinct by having dolipores with dome-shaped pore caps that contain large perforations (Fig. 2e; Gleason and McGee, 2001), whereas the homobasidiomycetes have dolipores with dome-shaped but multi-perforate pore caps (Fig. 2f, Abadie et al., 2006). Ascomycete mycobionts possess simple cell wall pori accompanied by Woronin bodies and mostly electron-translucent cell walls (Fig. 2g; Setaro et al., 2006). The only ascomycetes so far proven as mycobionts of orchids in situ are members of the ectomycorrhizal genus Tuber (Selosse et al., 2004). However, further taxonomic resolution cannot be obtained using only ultrastructural characters.

87This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Figure 1. Light micrographs of sections through mycorrhizal roots: a. Patchy distribution of fungal pelotons (p) in root cortical cells (CC) of Elleanthus sp. Outermost root layers comprise the velamen (V). Scale bar = 50 µm; b. Dense fungal colonization (h) of the two-layered velamen (V) and fungal pelotons (p) of Tulasnella sp. occur in the root cor-tical cells (CC), but hyphae are not present in the thick-walled exodermal cells (Ex) of Elleanthus sp. Scale bar = 50 µm; c. Pelotons (p) of fine, sebacinoid fungus and collapsed hyphae (ch) in root cortical cells (CC) of Sobralia rosea Poepp. & Endl. Scale bar = 10 µm; d. Hypha (h) traversing living, thin-walled passage cell (PC) of the exodermis of Elleanthus sp. Shown also are velamen (V) and tilosomes (T), outer wall excrescences of the passage cell. Scale bar = 10 µm.
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES

88
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Sebacinales Group A
Orchid species Geographic area Material and methods References
Cephalanthera damasonium (Mill.) Druceheterotrophic and mixotrophic
France ITS from mycorrhizas, BLAST Julou et al., 2004
Cephalanthera longifolia (L.) Fritsch heterotrophic and mixotrophic
Estonia ITS-RFLPs and sequences from mycorrhizas, BLAST
Abadie et al., 2006
Cypripedium fasciculatum Kellog ex S.Watson mixotrophic
USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas, molec. phylogeny
Shefferson et al., 2005, 2007; Suarez et al., 2008
Epipactis atrorubens (Hoffm.) Bessermixotrophic
Germany ITS, mtLSU from mycorrhizas, BLAST Bidartondo et al., 2004
Epipactis helleborine (L.) Crantzmixotrophic
Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004;Suarez et al., 2008
Epipactis microphylla (Ehrh.) Sw.heterotrophic and mixotrophic
France ITS, nrLSU from mycorrhizas, BLAST Selosse et al., 2004
Hexalectris revoluta CorrellH. spicata (Walter) Barnhartheterotrophic
USA ITS RFLPs, ITS/nrLSU sequences from mycorrhizas, molec. phylogeny
Taylor et al., 2003
Neottia nidus-avis (L.) Rich.heterotrophic
France, Germany, UK ITS, mtLSU sequences from mycorrhizas, molec. phylogeny
McKendrick et al., 2002; Selosse et al., 2002:Bidartondo et al. 2004
Sebacinales Group B
Caladenia carnea R.Br.C. falcata (Nicholls) M.A.Clem.
Australia ITS-RFLPs, nucLSU sequences from isolates, molec. phylogeny
Bougoure et al. 2005:Suarez et al., 2008
Caladenia catenata (Sm.) Druce Australia Warcup’s isolates; nucLSU Warcup 1988, Weiss et al. 2004
Cypripedium californicum A.Gray C. parviflorum Salisb.
USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas, molec. phylogeny
Shefferson et al. 2005, 2007;Suarez et al. 2008
Cyrtostylis reniformis R.Br. Australia Warcup’s isolates; nucLSU, molec. phylogeny Warcup 1988; Weiss et al. 2004
Epipactis palustris (L.) Crantz Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al. 2004: Suarez et al. 2008
Eriochilus scaber Lindl. Australia Warcup’s isolates; nucLSU, molec. phylogeny Warcup 1988; Weiss et al. 2004
Glossodia minor R.Br. Australia Warcup’s isolates; nucLSU, molec. phylogeny Warcup 1988; Weiss et al. 2004
Microtis media R.Br. Australia ITS from isolates, BLAST Bonnardeux et al. 2007
Microtis unifolia (G.Forst.) Rchb.f. Australia Warcup’s isolates, nucLSU, molec. phylogeny Warcup 1988; Weiss et al. 2004
Pleurothallis lilijae Foldats, epiphytic Ecuador nucLSU/ITS from mycorrhizas, molec. phylogeny
Suarez et al. 2008
Stelis concinna Lindl., S. superbiens Lindl.,S. hallii Lindl., epiphytic
Ecuador nucLSU/ITS from mycorrhizas, molec. phylogeny
Suarez et al. 2008
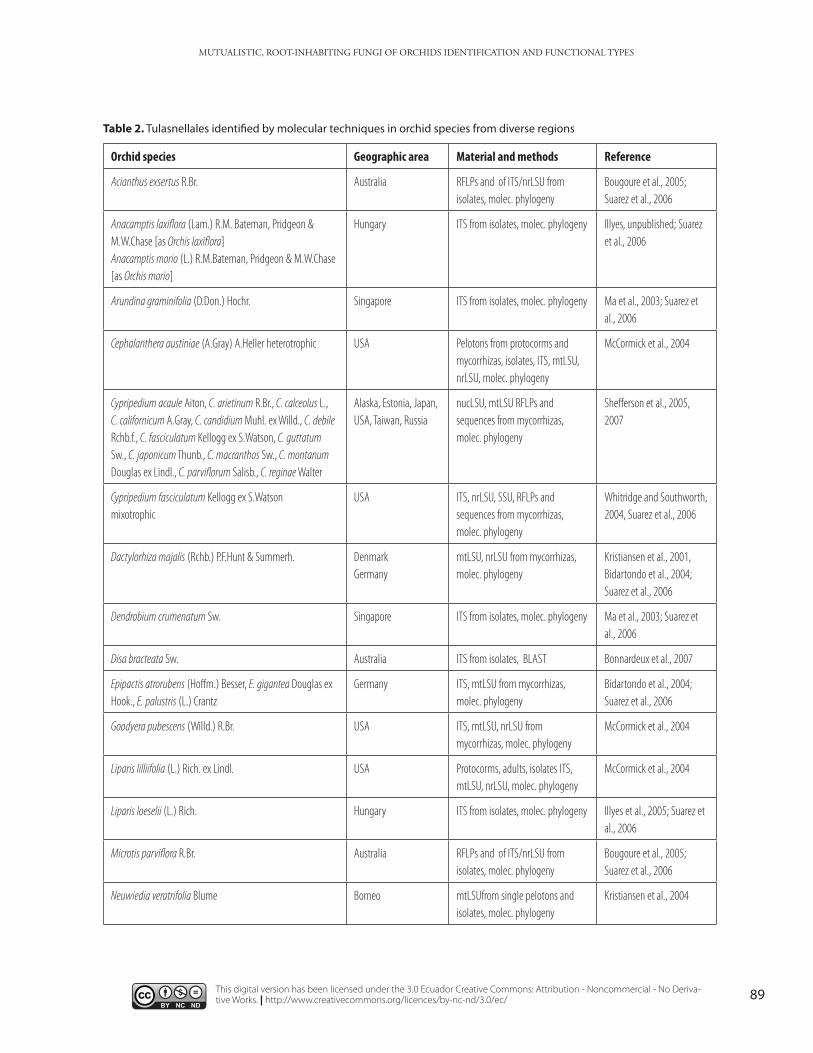
Table 1. Sebacinales Group A and Group B identified by molecular techniques in orchid species from diverse regions.
89This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Orchid species Geographic area Material and methods Reference
Acianthus exsertus R.Br. Australia RFLPs and of ITS/nrLSU from isolates, molec. phylogeny
Bougoure et al., 2005;Suarez et al., 2006
Anacamptis laxiflora (Lam.) R.M. Bateman, Pridgeon & M.W.Chase [as Orchis laxiflora]Anacamptis morio (L.) R.M.Bateman, Pridgeon & M.W.Chase [as Orchis morio]
Hungary ITS from isolates, molec. phylogeny Illyes, unpublished; Suarez et al., 2006
Arundina graminifolia (D.Don.) Hochr. Singapore ITS from isolates, molec. phylogeny Ma et al., 2003; Suarez et al., 2006
Cephalanthera austiniae (A.Gray) A.Heller heterotrophic USA Pelotons from protocorms and mycorrhizas, isolates, ITS, mtLSU, nrLSU, molec. phylogeny
McCormick et al., 2004
Cypripedium acaule Aiton, C. arietinum R.Br., C. calceolus L., C. californicum A.Gray, C. candidium Muhl. ex Willd., C. debile Rchb.f., C. fasciculatum Kellogg ex S.Watson, C. guttatum Sw., C. japonicum Thunb., C. macranthos Sw., C. montanum Douglas ex Lindl., C. parviflorum Salisb., C. reginae Walter
Alaska, Estonia, Japan,USA, Taiwan, Russia
nucLSU, mtLSU RFLPs and sequences from mycorrhizas, molec. phylogeny
Shefferson et al., 2005, 2007
Cypripedium fasciculatum Kellogg ex S.Watsonmixotrophic
USA ITS, nrLSU, SSU, RFLPs and sequences from mycorrhizas, molec. phylogeny
Whitridge and Southworth, 2004, Suarez et al., 2006
Dactylorhiza majalis (Rchb.) P.F.Hunt & Summerh. DenmarkGermany
mtLSU, nrLSU from mycorrhizas, molec. phylogeny
Kristiansen et al., 2001, Bidartondo et al., 2004; Suarez et al., 2006
Dendrobium crumenatum Sw. Singapore ITS from isolates, molec. phylogeny Ma et al., 2003; Suarez et al., 2006
Disa bracteata Sw. Australia ITS from isolates, BLAST Bonnardeux et al., 2007
Epipactis atrorubens (Hoffm.) Besser, E. gigantea Douglas ex Hook., E. palustris (L.) Crantz
Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004;Suarez et al., 2006
Goodyera pubescens (Willd.) R.Br. USA ITS, mtLSU, nrLSU from mycorrhizas, molec. phylogeny
McCormick et al., 2004
Liparis lilliifolia (L.) Rich. ex Lindl. USA Protocorms, adults, isolates ITS, mtLSU, nrLSU, molec. phylogeny
McCormick et al., 2004
Liparis loeselii (L.) Rich. Hungary ITS from isolates, molec. phylogeny Illyes et al., 2005; Suarez et al., 2006
Microtis parviflora R.Br. Australia RFLPs and of ITS/nrLSU from isolates, molec. phylogeny
Bougoure et al., 2005;Suarez et al., 2006
Neuwiedia veratrifolia Blume Borneo mtLSUfrom single pelotons andisolates, molec. phylogeny
Kristiansen et al., 2004
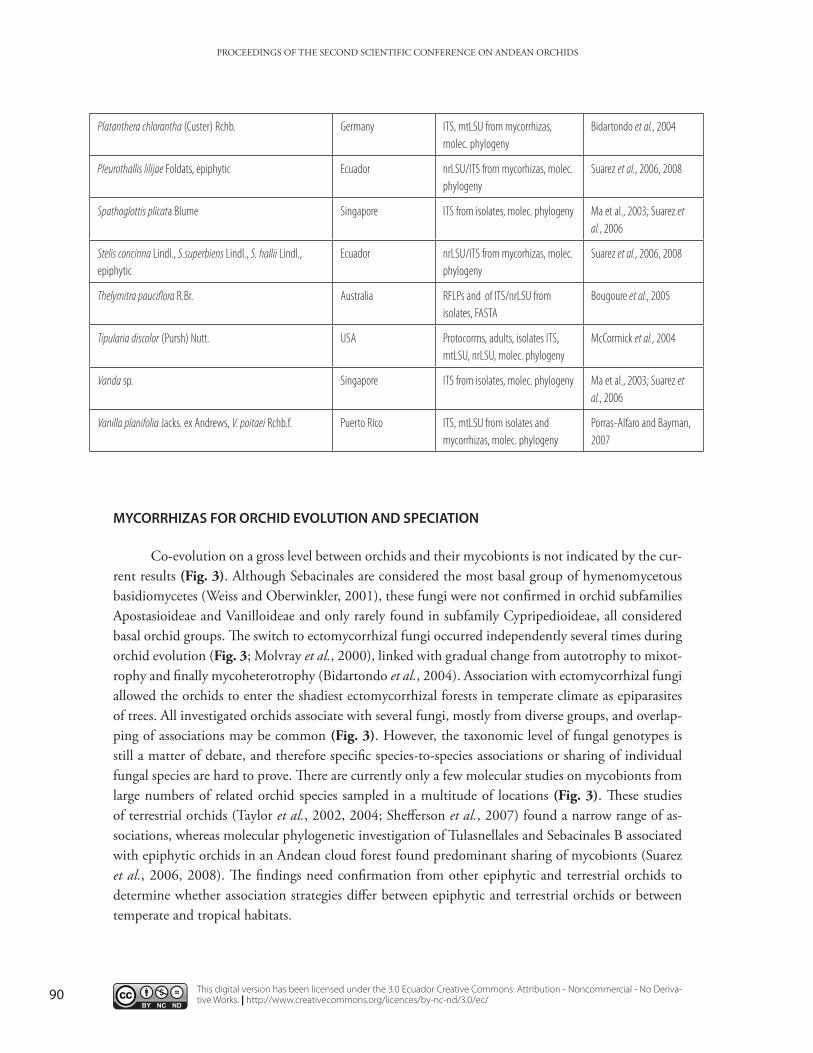
Table 2. Tulasnellales identified by molecular techniques in orchid species from diverse regions
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES
90
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
MYCORRHIZAS FOR ORCHID EVOLUTION AND SPECIATION
Co-evolution on a gross level between orchids and their mycobionts is not indicated by the cur-rent results (Fig. 3). Although Sebacinales are considered the most basal group of hymenomycetous basidiomycetes (Weiss and Oberwinkler, 2001), these fungi were not confirmed in orchid subfamilies Apostasioideae and Vanilloideae and only rarely found in subfamily Cypripedioideae, all considered basal orchid groups. The switch to ectomycorrhizal fungi occurred independently several times during orchid evolution (Fig. 3; Molvray et al., 2000), linked with gradual change from autotrophy to mixot-rophy and finally mycoheterotrophy (Bidartondo et al., 2004). Association with ectomycorrhizal fungi allowed the orchids to enter the shadiest ectomycorrhizal forests in temperate climate as epiparasites of trees. All investigated orchids associate with several fungi, mostly from diverse groups, and overlap-ping of associations may be common (Fig. 3). However, the taxonomic level of fungal genotypes is still a matter of debate, and therefore specific species-to-species associations or sharing of individual fungal species are hard to prove. There are currently only a few molecular studies on mycobionts from large numbers of related orchid species sampled in a multitude of locations (Fig. 3). These studies of terrestrial orchids (Taylor et al., 2002, 2004; Shefferson et al., 2007) found a narrow range of as-sociations, whereas molecular phylogenetic investigation of Tulasnellales and Sebacinales B associated with epiphytic orchids in an Andean cloud forest found predominant sharing of mycobionts (Suarez et al., 2006, 2008). The findings need confirmation from other epiphytic and terrestrial orchids to determine whether association strategies differ between epiphytic and terrestrial orchids or between temperate and tropical habitats.
Platanthera chlorantha (Custer) Rchb. Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004
Pleurothallis lilijae Foldats, epiphytic Ecuador nrLSU/ITS from mycorhizas, molec. phylogeny
Suarez et al., 2006, 2008
Spathoglottis plicata Blume Singapore ITS from isolates, molec. phylogeny Ma et al., 2003; Suarez et al., 2006
Stelis concinna Lindl., S.superbiens Lindl., S. hallii Lindl., epiphytic
Ecuador nrLSU/ITS from mycorhizas, molec. phylogeny
Suarez et al., 2006, 2008
Thelymitra pauciflora R.Br. Australia RFLPs and of ITS/nrLSU from isolates, FASTA
Bougoure et al., 2005
Tipularia discolor (Pursh) Nutt. USA Protocorms, adults, isolates ITS, mtLSU, nrLSU, molec. phylogeny
McCormick et al., 2004
Vanda sp. Singapore ITS from isolates, molec. phylogeny Ma et al., 2003; Suarez et al., 2006
Vanilla planifolia Jacks. ex Andrews, V. poitaei Rchb.f. Puerto Rico ITS, mtLSU from isolates and mycorrhizas, molec. phylogeny
Porras-Alfaro and Bayman, 2007

91This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Figure 2. Transmission electron micrographs of orchid mycobionts. a. Sebacinoid hypha in root cortical cell of So-bralia rosea with imperforate, slightly dish-shaped pore caps (Pa) with straight margins, covering the dolipore. Scale bar = 1 µm; b. Tulasnelloid hypha in root cortical cell of Elleanthus sp. with imperforate pore caps (Pa), dish-shaped with recurved margins. Scale bar =1 µm; c. Square section of tulasnelloid hyphae (h) with slime bodies (Sb) in cell walls (arrowhead) in root cortical cell of Elleanthus sp. Scale bar = 1 µm; d. Tulasnelloid hypha displaying several layers of slime bodies (arrows) in the cell walls. Scale bar = 1 µm; e. Hypha of Ceratobasidiales in root cortical cell of Stelis sp. displaying dome-shaped pore caps (Pa) with several holes. Scale bar = 1 µm; f. Homobasidiomycete in ectomycorrhiza displaying regular perforate pore caps (Pa). Scale bar = 0.5 µm; g. Ascomycete hypha with simple pore and Woronin bodies (Wb). Scale bar = 1 µm.
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES

92
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Figure 3. Distribution of mycobionts in subfamilies of Orchidaceae as determined by molecular techniques. Color of orchid name indicates dominant fungal group; colored lines indicate subordinate fungi associated. Heterotrophic and mixotrophic state of orchids is given when proven by isotope measurements. Homobasidiomycetous ectomy-corrhiza-forming fungi (bECMF).

93This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Orchid species Geographic area Material and methods Reference
Campylocentrum fasciola (Lindl.) Cogn.C. filiforme (Sw.) Cogn.
Puerto Rico ITS from isolates, molec. phylogeny Otero et al., 2002
Cephalanthera damasonium (Mill.) Druceheterotrophic and mixotrophic
France ITS from mycorrhizas, BLAST Julou et al., 2004
Cephalanthera longifolia (L.) Fritschheterotrophic and mixotrophic
Estonia ITS-RFLPs and sequences from mycorrhizas, BLAST
Abadie et al., 2006
Chamaegastrodia shikokiana Makino & Maek. heterotrophic
Japan ITS from isolates, molec. phylogeny Yagame et al., 2008
Cypripedium californicum A.Gray USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas, molec. phylogeny
Shefferson et al., 2005, 2007
Dactylorhiza majalis (Rchb.) P.F.Hunt & Summerh., Epipactis helleborine (L.) Crantz, E. palustris (L.) Crantz, Platanthera chlorantha Custer (Rchb.)
Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004
Hexalectris spicata (Walter) Barnhart, H. spicata var. arizonica (S.Watson) Catling & V.S.Engelmycoheterotroph
USA ITS RFLPs,nrLSU sequences from isolates, molec. phylogeny
Taylor et al., 2003
Ionopsis satyrioides (Sw.) Rchb.f.I. utricularioides (Sw.) Lindl.
Puerto Rico ITS from isolates, molec. phylogeny Otero et al., 2002, 2004, 2007
Neuwiedia veratrifolia Blume Borneo mtLSU, single pelotons from mycorrhizas and isolates, molec. phylogeny
Kristiansen et al., 2004
Oeceoclades maculata (Lindl.) Lindl. Puerto Rico ITS from isolates, molec. phylogeny Otero et al., 2002
Oncidium altissimum (Jacq.) Sw. Puerto Rico ITS from isolates, molec. phylogeny Otero et al., 2002
Psychilis monensis Sauleda Puerto Rico ITS from isolates, molec. phylogeny Otero et al., 2002
Pterostylis nutans R.Br. Australia ITS from isolates, molec. phylogeny Irwin et al., 2007
Pterostylis sanguinea D.L.Jones & M.A.Clem. Australia ITS from isolates, molec. phylogeny Bonnardeux et al., 2007
Pterostylis nutans R.Br.P. obtusa R.Br.
Australia Isolates, ITS-RFLPs, ITS/LSU sequences, FASTA
Bougoure et al., 2005
Sarcochilus falcatus R.Br.S. olivaceus Lindl.
Australia TEM porus of isolates Gleason and McGee, 2001
Tolumnia variegata (Sw.) Braem Puerto Rico ITS from isolates, molec. phylogeny Otero et al., 2002, 2004
Vanilla planifolia Jacks. ex Andrews, V. poitaei Rchb.f., V. aphylla Blume, V. phaeantha Rchb.f.
Puerto Rico, Cuba, Costa Rica
nrITS, mtLSU from isolates and mycorrhizas, molec. phylogeny
Porras-Alfaro & Bayman, 2007
Table 3. Ceratobasidiales identified by molecular techniques or ultrastructure in orchid species from diverse regions.
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES

94
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Fungal group or species
Orchid species Geographic area
Material and methods Reference
Thelephoraceae Cephalanthera austiniae (A.Gray) A.Hellerheterotrophic
USA ITS, mtLSU, nrLSU RFLPs and sequences from isolates and mycorrhizas, molec. phylogeny
Taylor and Bruns, 1997; McCormick et al., 2004
Cortinariaceae, Hymenogaster, Inocybe, Thelephoraceae
Cephalanthera damasonium (Mill.) Druceheterotrophic and mixotrophic
Germany, France
ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004; Julou et al., 2004
Thelephoraceae, Cortniariaceae, Amphinema, Wilcoxina
Cephalanthera longifolia (L.) Fritschheterotrophic and mixotrophic
Estonia ITS-RFLPs and sequences from mycorrhizas, BLAST
Abadie et al., 2006
Tomentella Cephalanthera rubra (L.) Rich.mixotrophic
Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004
Russulaceae Corallorhiza maculata (Raf.) Raf., C. mertensiana Bong., heterotrophic
USA ITS, mtLSU, nrLSU - RFLPs and sequences from mycorrhizas, molec. phylogeny
Taylor and Bruns, 1997, 1999Taylor et al., 2003
Thelephoraceae Corallorhiza trifida Châtelheterotrophic
Scotland ITS from mycorrhizas, BLAST McKendrick et al., 2000b
Thelephoraceae Cypripedium candidium Muhl. ex Willd., C. montanum Douglas ex Lindl.
USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas , molec. phylogeny
Shefferson et al., 2005, 2007
Russula laurocerasii Cypripedium acaule Aiton USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas, molec. phylogeny
Shefferson et al., 2007
Russula, Lactarius, Suillus
Cypripedium fasciculatum Kellogg ex S.Watsonmixotrophic
USA ITS, nrLSU, SSU, RFLPs and sequences from mycorrhizas, molec. phylogeny
Whitridge and Southworth, 2005
Russula Cypripedium parviflorum Salisb. USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas, molec. phylogeny
Shefferson et al., 2005, 2007
Hygrocybe, Cantharellus
Cypripedium reginae Walter USA nucLSU, mtLSU RFLPs and sequences from mycorrhizas , molec. phylogeny
Shefferson et al., 2007
GymnomycesRussula
Dipodium hamiltonianum F.M.Baileyheterotrophic
Australia ITS from mycorrhizas, BLAST Dearnaley and Le Brocque, 2006
Russula Dipodium variegatum M.A.Clem.heterotrophic
Australia ITS, from mycorrhizas, BLAST Bougoure and Dearnaley, 2005
Table 4. Ectomycorrhiza-forming homobasidiomycetes identified by molecular techniques in orchid species from diverse regions.

95This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Literature cited
Abadie, L. C., Puttsepp, Ü., Gebauer, G., Faccio, A., Bonfante, P., and Selosse, M. A. 2006. Cephalan-thera longifolia (Neottieae, Orchidaceae) is mixotrophic: a comparative study between green and nonphotosynthetic individuals. Canadian Journal of Botany 84: 1462-1477.
Alexander, C., Alexander, I. J., and Hadley, G. 1984. Phosphate uptake by Goodyera repens in relation to mycorrhizal infection. New Phytologist 97: 401-411.
Alexander, C. and Hadley, G. 1985. Carbon movement between host and mycorrhizal endophyte dur-ing the development of the orchid Goodyera repens Br. New Phytologist 101: 657-665.
Andersen, T. F. 1996. A comparative study of Rhizoctonia sensu lato employing morphological, ul-trastructural and molecular methods. Mycological Research 100: 1117-1128.
Bayman, P. and Otero, J. T. 2006. Microbial endophytes of orchid roots. In Microbial root endophytes (ed. B. Schulz, C. Boyle, and T. N. Sieber), pp. 153-177. Soil Biology 9. Springer-Verlag, Ber-lin.
Bernard, N. 1909. L’evolution dans la symbiose. Les orchidées et leurs champignons commensaux. Annales des Sciences Naturelles Botanique 9: 1-196.
Bidartondo, M. I., Burghardt, B., Gebauer, G., Bruns, T. D., and Read, D. J. 2004. Changing part-ners in the dark: isotopic and molecular evidence of ectomycorrhizal liaison between forest orchids and trees. Proceedings of the Royal Society London B 271: 1799-1806.
Bonnardeaux, Y., Brundrett, M., Batty, A. L., Dixon, K. W., Koch, J., and Sivasithamparam, K. 2007. Diversity of mycorrhizal fungi of Western Australien terrestrial orchids: compatibility webs, brief encounters, lasting relationships, and alien invasions. Mycological Research 111: 51-61.
Inocybe, Tuber Epipactis atrorubens (Hoffm.) Bessermixotrophic
Germany ITS, mtLSU from mycorrhizas , molec. phylogeny
Bidartondo et al., 2004
Wilcoxina Epipactis distans Arv.-Touv.mixotrophic
Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004
Tuber Epipactis helleborine (L.) Crantzmixotrophic
Germany ITS, mtLSU from mycorrhizas, molec. phylogeny
Bidartondo et al., 2004
Cortinariaceae, Hymenosgastraceae, Thelephoraceae, Russula foetens Fr., Tuber spp.
Epipactis microphylla (Ehrh.) Sw.heterotrophic and mixotrophic
France ITS, nrLSU from mycorrhizas, BLAST
Selosse et al., 2004
Russula delica Fr., R. chloroides Kalchbr., R. brevipes Peck
Limodorum abortivum (L.) Sw., L. brulloi Bartolo & Pulv., L. trabutianum Batt., mixotrophic
Italy ITS-RFLPs and sequences from mycorrhizas, molec. phylogeny
Girlanda et al., 2006
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES

96
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Bougoure, J. J., Bougoure, D. S., Cairney, W. G., and Dearnaley, D. W. 2005. ITS-RFLP and sequence analysis of endophytes from Acianthus, Caladenia and Pterostylis (Orchidaceae) in southeastern Queensland. Mycological Research 109: 452-460.
Bougoure, J. J. and Dearnaley, J. D. 2005. The fungal endophytes of Dipodium variegatum (Or-chidaceae). Australasian Mycologist 24: 15-19.
Brundrett, M. C. 2006. Understanding the roles of multifunctional mycorrhizal and endophytic fun-gi. In Microbial root endophytes (ed. B. Schulz, C. Boyle, and T. N. Sieber), pp. 153-177. Soil Biology 9. Springer-Verlag, Berlin.
Cameron, D. D, Leake, J. R., and Read, D. J. 2006. Mutualistic mycorrhiza in orchids: evidence from plant-fungus carbon and nitrogen transfer in the green-leaved terrestrial orchid Goodyera repens. New Phytologist 171: 405-416.
Cameron, D. D., Johnson, I., Leake, J. R., and Read, D. J. 2007. Mycorrhizal acquisition of inorganic phosphorus by the green-leaved terrestrial orchid Goodyera repens. Annals of Botany 99: 831-834.
Currah, R. S. and Sherburne R. 1992. Septal ultrastructure of some fungal endophytes from boreal orchid mycorrhizas. Mycological Research 96: 583-587.
Currah, R. S., Sigler, L., and Hambleton, S. 1987. New records and taxa of fungi from the mycorrhi-zae of terrestrial orchids of Alberta. Canadian Journal of Botany 65: 2473-2482.
Currah, R. S., Zelmer, C. D., Hambleton, S., and Richardson, K. A. 1997. Fungi from orchid mycor-rhizas. In Orchid biology: reviews and perspectives, VII. (ed. J. Arditti and A. M. Pridgeon), pp. 117-170. Kluwer Academic Publishers, Dordrecht.
Dearnaley, J. D. and Le Brocque, A. F. 2006. Molecular identification of the primary root fungal endophytes of Dipodium hamiltonianum (yellow hyacinth orchid). Australian Journal of Botany 54: 487-491.
Dörr, I. and Kollmann, R. 1969. Fine structure of mycorrhiza in Neottia nidus-avis (L.) L. C. Rich. (Orchidaceae). Planta 89: 372-375.
Gebauer, G. and Meyer, M. 2003. 15N and 13C natural abundance studies of autotrophic and myco-heterotrophic orchids provides insight into nitrogen and carbon gain from fungal associations. New Phytologist 160: 209-223.
Girlanda, M., Selosse, M. A., Cafasso, D., Brilli, F., Delfine, S., Fabbian, R., Ghgignone, S., Pinelli, P., Segreto, R., Loreto, F., Cozzolono, S., and Perotto, S. 2006. Inefficient photosynthesis in the Mediterranean orchid Limodorum abortivum is mirrored by specific associations to ectomycor-rhizal Russulaceae. Molecular Ecology 15: 491-504.
Gleason, F. H. and McGee, P. A. 2001. Septal pore cap ultrastructure of fungi identified as Epulorhiza sp. (sensu Sebacina) isolated from Australian orchids. Australasian Mycologist 20: 12-15
Hadley, G. 1969. Cellulose as a carbon source for orchid mycorrhiza. New Phytologist 68: 933-939.Hadley, G. 1970. Non-specificity of symbiotic infection in orchid mycorrhiza. New Phytologist 69:
1015-1023.Illyes, Z., Rudnoy, S., and Bratek, Z. 2005. Aspects of in situ, in vitro germination and mycorrhizal
partners of Liparis loeselii. Acta Biologica Szegediensis 49: 137-139.

97This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Irwin, M. J., Bougoure, J. J., and Dearnaley, J. D. 2007. Pterostylis nutans (Orchidaceae) has a specific association with two Ceratobasidium root-associated fungi across its range in eastern Australia. Mycoscience 48: 231-239.
Julou, T., Burghardt, B., Gebauer, G., Berveilleir, D., Damesin, C., and Selosse, M. A. 2005. Mixotro-phy in orchids: insight from a comparative study of green individuals and nonphotosynthestic individuals of Cephalanthera damasonium. New Phytologist 166: 639-653.
Kristiansen, K. A., Taylor, D. L., Kjoller, H. N., Rasmussen, N., and Rosendahl, S. 2001. Identifica-tion of mycorrhizal fungi from single pelotons of Dactylorhiza majalis (Orchidaceae) using single-strand conformation polymorphism and mitochondrial ribosomal large subunit DNA sequences. Molecular Ecology 10: 2089-2093.
Kristiansen, K. A., Freudenstein, J. V., Rasmussen, F. N., and Rasmussen, H. N. 2004. Molecular identification of mycorrhizal fungi in Neuwiedia veratrifolia (Orchidaceae). Molecular Phyloge-netics and Evolution 33: 251-258.
Ma, M., Tan, T. K., and Wong, S. M. 2003. Identification and molecular phylogeny of Epulorhiza isolates from tropical orchids. Mycological Research 107: 1041-1049.
McCormick, M. K., Whigham, D. F., and O’Neill, J. 2004. Mycorrhizal diversity in photosynthetic terrestrial orchids. New Phytologist 163: 425-438.
McKendrick, S. L., Leake, J. R., and Read, D. J. 2000a. Symbiotic germination and development of myco-heterotrophic plants in nature: transfer of carbon from ectomycorrhizal Salix repens and Betula pendula to the orchid Corallorhiza trifida through shared hyphal connections. New Phytologist 145: 539-548.
McKendrick, S. L., Leake, J. R., Taylor, D. L., and Read, D. J. 2000b. Symbiotic germination and development of myco-heterotrophic plants in nature: ontogeny of Corallorhiza trifida and char-acterization of its mycorrhizal fungi. New Phytologist 145: 523-537.
McKendrick, S. L., Leake, D. J., Taylor, D. L., and Read, D. J. 2002. Symbiotic germination and development of myco-heterotrophic orchid Neottia nidus-avis in nature and its requirement for locally distributed Sebacina spp. New Phytologist 154: 233-247.
Midgley, D. J., Jordan, L. A., Saleeba, J. A., and McGee, P. A. 2006. Utilisation of carbon substrates by orchid and ericoid mycorrhizal fungi from Australian dry sclerophyll forests. Mycorrhiza 16: 175-182.
Molvray, M., Kores, P. J., and Chase, M. W. 2000. Polyphyly of mycoheterotrophic orchids and func-tional influences on floral and molecular characters. In Monocots: systematics and evolution (ed. K. L. Wilson and D. A. Morrison), pp. 441-448. CSIRO Publishing, Melbourne.
Otero, J. T., Ackerman, J. D., and Bayman, P. 2002. Diversity and host specificity of endophytic Rhi-zoctonia-like fungi from tropical orchids. American Journal of Botany 89: 1852-1858.
Otero J. T., Ackerman, J. D., and Bayman, P. 2004. Differences in mycorrhizal preferences between two tropical orchids. Molecular Ecology 13: 2393-2404.
Otero, J. T., Flanagan, N. S., Herre, E. A., Ackerman, J. D., and Bayman, P. 2007. Widespread mycor-rhizal specificity correlates to mycorrhizal function in the neotropical, epiphytic orchid Ionopsis utricularioides (Orchidaceae). American Journal of Botany 94: 1944-1950
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES

98
PROCEEDINGS OF THE SECOND SCIENTIFIC CONFERENCE ON ANDEAN ORCHIDS
This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Peterson, R. L., Bonfante, P., Faccio, A., and Uetake, Y. 1996. The interface between fungal hyphae and orchid protocorm cells. Canadian Journal of Botany 74: 1861-1870.
Peterson, R. L. and Currah, R. S. 1990. Synthesis of mycorrhizas between protocorms of Goodyera repens (Orchidaceae) and Ceratobasidium cereale. Canadian Journal of Botany 68: 117-125.
Peterson, R. L. and Massicotte, H. B. 2004. Exploring structural definitions of mycorrhizas, with em-phasis on nutrient-exchange interfaces. Canadian Journal of Botany 82: 1074-1088.
Porras-Alfaro, A. and Bayman, P. 2007. Mycorrhizal fungi of Vanilla: diversity, specificity and effects on seed germination and plant growth. Mycologia 99: 510-525.
Rasmussen, H. 1990. Cell differentiation and mycorrhizal infection in Dactylorhiza majalis (Rchb. f.) Hunt and Summerh. (Orchidaceae) during germination in vitro. New Phytologist 116: 137-147.
Roberts, P. 1999. Rhizoctonia-forming fungi: a taxonomic guide. Royal Botanic Gardens, Kew, Rich-mond, Surrey, UK.
Selosse, M.-A., Weiss, M., Jany, J.-L., and Tiller, A. 2002. Communities and populations of sebac-inoid Basidiomycetes associated with the achlorophyllous orchid Neottia nidus-avis (L.) L.C.M. Rich. and neighbouring tree ectomycorrhizae. Molecular Ecology 11: 1831–1844.
Selosse, M.-A., Faccio, A., Scappaticci, G., and Bonfante, P. 2004. Chlorophyllous and achlorophyl-lous specimens of Epipactis microphylla (Neottieae, Orchidaceae) are associated with ectomycor-rhizal septomycetes, including truffles. Microbial Ecology 47: 416-426.
Setaro, S., Oberwinkler, F., and Kottke, I. 2006. Anatomy and ultrastructure of mycorrhizal associa-tions of neotropical Ericaceae. Mycological Progress 5: 243-254.
Shefferson, R. P., Weiss, M., Kull, T., and Taylor, D. L. 2005. High specificity generally characterizes mycorrhizal association in rare lady’s slipper orchids, genus Cypripedium. Molecular Ecology 14: 613-626.
Shefferson, R. P., Taylor, D. L., Weiss, M., Garnica, S., McCormick, M. K., Adams, S., Gray H. M., McFarland, J. W., Kull, T., Tali, K., Yukawa, T., Kawahara, T., Miyoshi, K., and Lee, Y.-L. 2007. The evolutionary history of mycorrhizal specificity among lady’s slipper orchids. Evolu-tion 61: 1380-1390.
Smith, S. E. 1967. Carbohydrate translocation into orchid mycorrhizas. New Phytologist 66: 371-378.
Smith, S. E. and Read, D. J. 1997. Mycorrhizal symbiosis. 2nd ed. Academic Press, San Diego. Suárez, J. P., Weiss, M., Abele, A., Garnica, S., Oberwinkler, F., and Kottke, I. 2006. Diverse tulas-Diverse tulas-
nelloid fungi form mycorrhizas with epiphytic orchids in an Andean cloud forest. Mycological Research 110: 1257-1270.
Suárez, J. P., Weiss, M., Abele, A., Oberwinkler, F., and Kottke, I. 2008. Members of Sebacinales subgroup B form mycorrhizas with epiphytic orchids in a neotropical mountain rain forest. Mycological Progress 7: 75-85.
Taylor, D. L. and Bruns, T. D. 1997. Independent, specialized invasion of ectomycorrhizal mutualism by two non- photosynthetic orchids. Proceedings of the National Academy of Sciences USA 94: 4510-4515.

99This digital version has been licensed under the 3.0 Ecuador Creative Commons: Attribution - Noncommercial - No Deriva-tive Works. | http://www.creativecommons.org/licences/by-nc-nd/3.0/ec/
Taylor, D. L. and Bruns, T. D. 1999. Population, habitat and genetic correlates of mycorrhizal spe-cialization in the “cheating” orchids Corallorhiza maculata and C. mertensiana. Molecular Ecol-ogy 8: 1719-1732.
Taylor, D. L., Bruns, T. D., Leake, J. R., and Read, D. J. 2002. Mycorrhizal specificity and function in myco-heterotrophic plants. In Mycorrhizal ecology. Ecological studies, Vol. 157 (ed. M. G. A. van der Heijden and I. Sanders), pp. 375-413. Springer-Verlag, Berlin.
Taylor, D. L., Bruns, T. D., Szaro, T. M., and Hodges, S. A. 2003. Divergence in mycorrhizal speciali-zation within Hexalectris spicata (Orchidaceae), a non-photosynthetic desert orchid. American Journal of Botany 90: 1168-1179.
Taylor, D. L., Bruns, T. D., and Hodges, S. A. 2004. Evidence for mycorrhizal races in a cheating orchid. Proceedings of the Royal Society London B 271: 35-43.
Trudell, S. A., Rygiewicz, P. T., and Edmonds, R. I. 2003. Nitrogen and carbon stable isotope abun-dance support the myco-heterotrophic nature and host-specificity of certain achlorophyllous plants. New Phytologist 160: 391-401.
Warcup, J. H. 1988. Mycorrhizal associations of isolates of Sebacina vermifera. New Phytologist 110: 227-231.
Warcup, J. H. 1991. The Rhizoctonia endophytes of Rhizanthella gardneri (Orchidaceae). Mycological Research 95: 656-659.
Warcup, J. H. and Talbot, P. H. 1966. Perfect states of some Rhizoctonias. Transaction British Mycologi-cal Society 49: 427-435.
Warcup, J. H. and Talbot, P. H. 1967a. Perfect states of Rhizoctonias associated with orchids. New Phytologist 66: 631-641.
Warcup, J. H. and Talbot, P. H. 1967b. Perfect states of Rhizoctonias associated with orchids. II. New Phytologist 70: 35-40.
Warcup, J. H. and Talbot, P. H. 1980. Perfect states of Rhizoctonias associated with orchids. III. New Phytologist 86: 267-272.
Weiss, M. and Oberwinkler, F. 2001. Phylogenetic relationships in Auriculariales and related groups - hypotheses derived from nuclear ribosomal DNA sequences. Mycological Research 105: 403-415.
Weiss, M., Selosse, M. A., Rexer, K. H., Urban, A., and Oberwinkler, F. 2004. Sebacinales: a hitherto overlooked cosm of heterobasidiomycetes with a broad mycorrhizal potential. Mycological Re-search 108: 1003-1010.
Whitridge, H. and Southworth, D. 2004. Mycorrhizal symbionts of the terrestrial orchid Cypripedium fasciculatum. Selbyana 26: 254-260.
Williamson, B. and Hadley, G. 1970. Penetration and infection of orchid protocorms by Thanatepho-rus cucumeris and other Rhizoctonia isolates. Phytopathology 60: 1092-1096.
Yagame, T., Yamato, M., Suzuki, A., and Iwase, K. 2008. Ceratobasidiaceae mycorrhizal fungi isolated from nonphotosynthetic orchid Chamaegastrodia shikokiana. Mycorrhiza 18: 97-102.
Zelmer, C. D. and Currah, R. S. 1995. Evidence for a fungal liaison between Corallorhiza trifida (Or-chidaceae) and Pinus contorta (Pinaceae). Canadian Journal of Botany 73: 862-866.
MUTUALISTIC, ROOT-INHABITING FUNGI OF ORCHIDS IDENTIFICATION AND FUNCTIONAL TYPES
Copyright © 2022 FDOKUMEN




















