
Uptake and incorporation of sialic acid by the tick Ixodes ricinus

Soldati-FavreStephen Matthews, Ten Feizi and Dominique Saouros, Makoto Kiso, Michael J. Blackman,Angelina S. Palma, Ester Leon, Savvas Nikolas Friedrich, Joana M. Santos, Yan Liu, Apicomplexan Parasitesduring Host Cell Invasion by Domains Act as Sialic Acid-binding LectinsContaining Microneme Adhesive Repeat Members of a Novel Protein FamilyGlycobiology and Extracellular Matrices:
doi: 10.1074/jbc.M109.060988 originally published online November 9, 20092010, 285:2064-2076.J. Biol. Chem.
10.1074/jbc.M109.060988Access the most updated version of this article at doi:
.JBC Affinity SitesFind articles, minireviews, Reflections and Classics on similar topics on the
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2009/11/09/M109.060988.DC1.html
http://www.jbc.org/content/285/3/2064.full.html#ref-list-1
This article cites 48 references, 19 of which can be accessed free at
at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

Members of a Novel Protein Family Containing MicronemeAdhesive Repeat Domains Act as Sialic Acid-binding Lectinsduring Host Cell Invasion by Apicomplexan Parasites*□S
Received for publication, August 29, 2009, and in revised form, November 5, 2009 Published, JBC Papers in Press, November 9, 2009, DOI 10.1074/jbc.M109.060988
Nikolas Friedrich‡, Joana M. Santos‡1,2, Yan Liu§1, Angelina S. Palma§3, Ester Leon¶, Savvas Saouros¶, Makoto Kiso�,Michael J. Blackman**, Stephen Matthews¶, Ten Feizi§4, and Dominique Soldati-Favre‡5
From the ‡Department of Microbiology and Molecular Medicine, Faculty of Medicine, University of Geneva Centre MedicalUniversitaire, 1 Rue Michel-Servet, 1211 Geneva 4, Switzerland, §Glycosciences Laboratory, Division of Medicine, Imperial CollegeLondon, Northwick Park Campus, Harrow HA1 3UJ, United Kingdom, ¶Division of Molecular Biosciences, Imperial College London,South Kensington Campus, London SW7 2AZ, United Kingdom, �Department of Applied Bioorganic Chemistry, Gifu University,Gifu 501-11, Japan, and **Division of Parasitology, Medical Research Council National Institute for Medical Research, Mill Hill,London NW7 1AA, United Kingdom
Numerous intracellular pathogens exploit cell surface glyco-conjugates for host cell recognition and entry. Unlike bacteriaand viruses, Toxoplasma gondii and other parasites of the phy-lum Apicomplexa actively invade host cells, and this processcritically depends on adhesins (microneme proteins) releasedonto the parasite surface from intracellular organelles calledmicronemes (MIC). The microneme adhesive repeat (MAR)domain ofT. gondiiMIC1 (TgMIC1) recognizes sialic acid (Sia),a key determinant on the host cell surface for invasion by thispathogen. By complementation and invasion assays, we demon-strate that TgMIC1 is one important player in Sia-dependentinvasion and that another novel Sia-binding lectin, designatedTgMIC13, is also involved. Using BLAST searches, we identify afamily of MAR-containing proteins in enteroparasitic coccid-ians, a subclass of apicomplexans, including T. gondii, suggest-ing that all these parasites exploit sialylated glycoconjugates onhost cells as determinants for enteric invasion. Furthermore,this protein family might provide a basis for the broad host cellrange observed for coccidians that form tissue cysts duringchronic infection. Carbohydrate microarray analyses, cor-roborated by structural considerations, show that TgMIC13,TgMIC1, and its homologue Neospora caninumMIC1 (NcMIC1)
share a preference for �2–3- over �2–6-linked sialyl-N-acetyllac-tosamine sequences. However, the three lectins also display differ-ences in binding preferences. Intense binding of TgMIC13 to�2–9-linkeddisialyl sequencereportedonembryonal cells andrel-atively strong binding to 4-O-acetylated-Sia found on gut epithe-lium and binding of NcMIC1 to 6�sulfo-sialyl Lewisx might haveimplications for tissue tropism.
Sialic acids (Sias)6 occur abundantly in glycoproteins and gly-colipids on the cell surface and are exploited by many virusesand bacteria for attachment and host cell entry. Recognition ofcarbohydrates and in particular sialylated glycoconjugates isimportant also for host cell invasion by the Apicomplexa (1–4),a phylum that includes several thousand species of obligateintracellular parasites, among them the Plasmodium spp. caus-ing malaria. Enteroparasitic coccidians are a subclass of Api-complexa comprising Eimeria spp. responsible for coccidiosisin poultry, Neospora spp. causing neosporosis in cattle, andToxoplasma, the causative agent of toxoplasmosis in warm-blooded animals and humans.The host range and cell type targeted by these parasites vary
widely across the phylum. Whereas Plasmodium falciparummerozoites exclusively invade erythrocytes of humans andgreat apes (5), Toxoplasma gondii tachyzoites (the form of theparasite associated with acute infection) invade an extremelybroad range of cell types in humans and virtually all warm-blooded animals, enabling rapid establishment of infection inthe host and dissemination into deep tissues (6). Information isemerging on the involvement of carbohydrate-protein interac-tions in this broad host cell recognition (1).Many intracellular pathogens have evolved tomanipulate the
phagocytic pathways of host cells during invasion. This con-trasts with invasion by apicomplexans, which express their own
* This work was supported in part by the Swiss National Foundation (to D. S.),the UK Medical Research Council, and UK Research Council Basic Technol-ogy Grant GR/S79268 and Translational Grant EP/G037604/1 (to T. F.). Thiswork is part of the activities of the BioMalPar European Network of Excel-lence supported by a European Grant LSHP-CT-2004-503578 from the Pri-ority 1 “Life Sciences, Genomics, and Biotechnology for Health” in the 6thFramework Programme.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Materials and Methods, Figs. S1–S7, and Tables I and II.
1 Both authors contributed equally to this work.2 Recipient of the European Union-funded Marie Curie Action MalPar Training
Grant MEST-CT-2005-020492 (The Challenge of Malaria in the Post-genomic Era).
3 Fellow of the Fundacao para a Ciencia e Tecnologia and supported by GrantSFRH/BPD/26515/2006, Portugal. Present address: REQUIMTE, Centro deQuímica Fina e Biotecnologia, Dept. de Química, Faculdade de Ciencias eTecnologia, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal.
4 To whom correspondence may be addressed. Tel.: 44-20-8869-3460; E-mail:[email protected].
5 International Howard Hughes Medical Institute scholar. To whom corre-spondence may be addressed. Tel.: 41-22-379-5672; Fax: 41-22-379-5702;E-mail: [email protected].
6 The abbreviations used are: Sia, sialic acid; EGF, epidermal growth factor;ESA, excreted secreted antigens fraction; HFF, human foreskin fibroblast;MAR, microneme adhesive repeat; MCP, MAR-containing protein; MIC,microneme protein; NANA, N-acetylneuraminic acid; 3�SiaLacNAc, 3�sialyl-N-acetyllactosamine; PBS, phosphate-buffered saline; CHO, Chinese ham-ster ovary; IFA, immunofluorescence assay; WT, wild type; EST, expressedsequence tag; SP, signal peptide.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 3, pp. 2064 –2076, January 15, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
2064 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

machinery for active host cell entry. Invasion is a multistepprocess requiring the tightly regulated discharge of parasiteorganelles called micronemes and rhoptries (7). Micronemesrelease adhesins (MICs) onto the parasite surface, which formmultiprotein complexes with nonoverlapping roles in motility,host cell attachment, secretion of rhoptry organelles, and cellpenetration (8). After attachment and reorientation of the par-asite, invasion induces the formation of a nonfusogenic parasi-tophorous vacuole derived in large part from host cell plasmamembranes (9). The MICs share a limited number of adhesivedomains arranged in various combinations and numbers (10).These domains are implicated in host cell recognition andattachment and are believed to contribute to host cell typespecificity and hence disease pathology.T. gondii microneme protein 1 (TgMIC1) forms a complex
with TgMIC4 and TgMIC6 (11, 12) and binds to sialylated gly-coconjugates on the host cell surface (1). Previous studies basedon gene disruption have established a critical role for the com-plex in host cell invasion in tissue culture and its contribution tovirulence in vivo (13). The N-terminal region of TgMIC1 inter-acts with TgMIC4, a protein comprising six “apple” domainsthat has been shown to bind to host cells in the presence ofTgMIC1 (11). TgMIC6 contains three epidermal growth factor(EGF)-like domains and is a type I membrane protein, whichserves as an escorter and anchors the TgMIC1-MIC4-MIC6(TgMIC1-4-6) complex to the parasite surface during invasion(12). The first EGF-like domain (TgMIC6-EGF1) is cleaved offduring secretory transport of the complex, probably in a post-Golgi compartment (14). Each of the remaining two EGF-likedomains is able to recruit one molecule of TgMIC1 via interac-tion with its C-terminal galectin-like domain (for a schematicsee Fig. 1A) (12, 15, 16). Correct trafficking of the complex tothe micronemes depends not only on a sorting determinant inthe C-terminal tail of TgMIC6 but also on the interactionbetween the galectin-like domain of TgMIC1 (TgMIC1-GLD)with the third membrane-proximal EGF-like domain ofTgMIC6 (TgMIC6-EGF3). This interaction is crucial for trans-port of the entire complex through the early secretory pathwayas it assists proper folding of TgMIC6-EGF3, providing a qual-ity control checkpoint (12, 15, 17).Several studies have shown that recognition of carbohydrate
structures on the host cell surface is critical for efficient inva-sion by T. gondii (18–20). We have recently demonstrated thatthe N-terminal region of TgMIC1 contains two copies of anovel MAR domain and that this region termed TgMIC1-MARR binds specifically to sialylated oligosaccharides asshown by cell binding assays and carbohydrate microarrayanalyses (1). Also, we observed a 90% reduction of invasionefficiency when N-acetylneuraminic acid (NANA) was used asa competitor or when host cells were treated with neuramini-dase (1). This suggested that Sia is a major determinant of hostcell invasion by T. gondii and that the effect observed in theinvasion assays may be attributed, at least in part, to inhibitionof the interaction betweenTgMIC1 and its host cell receptor(s).Here, we compare the invasion efficiency of several T. gondii
knock-out strains and complemented mutants and demon-strate that the TgMIC1-Sia interaction is indeed important forefficient host cell invasion. We also show that the TgMIC1-4-6
complex is not the only molecular player involved in Sia-de-pendent invasion by T. gondii tachyzoites. By BLAST searches,we identify a family of MAR domain-containing proteins(MCPs) in T. gondii and related apicomplexan parasites Sarco-cystis neurona,Neospora caninum, andEimeria tenella. Amongthese we characterize TgMIC13, and we compare the bindingspecificities of recombinant TgMIC13 with TgMIC1 and itshomologue from the closely related organism N. caninum,NcMIC1, by carbohydrate microarray and cell binding assays.Our results indicate that the MAR domain is unique to andconserved among coccidians and that it acts as an importantdeterminant in host cell recognition by these parasites throughselective binding to sialylated glycoconjugates.
EXPERIMENTAL PROCEDURES
Generation of T. gondii Mutant Parasite Strains—All T. gon-dii strains were grown in human foreskin fibroblasts (HFF) orVero cells in Dulbecco’s modified Eagle’s medium (Invitrogen),10% fetal calf serum, 2 mM glutamine, 25 �g/ml gentamicin.The RH strain is referred to as “wild type.”The mic1ko parasite strain, generated previously (12),
was stably complemented with linearized plasmidspM2MIC1myc, pM2MIC1T220Amyc, pM2MIC1T126A,T220Amyc,and pROP1mycMIC1-GLD coding for the expression ofTgMIC1myc, TgMIC1T220Amyc, TgMIC1TTAAmyc, andmycTgMIC1-GLD (encompassing amino acids 299–456 ofTgMIC1), respectively, using a standard electroporationtransfection protocol with restriction enzyme-mediatedinsertion. For selection, plasmids were cotransfected withp2854_DHFR or pTUB5-CAT (carrying the pyrimethamineand the chloramphenicol resistance marker, respectively)at a 5:1 ratio. pM2MIC1T220Amyc was generated frompM2MIC1myc using the QuikChange kit (Stratagene) andprimers TgMIC1-20_1731 and TgMIC1-21_1732.pM2MIC1T126A,T220Amyc was obtained by replacing thefragment between restriction sites NdeI and EcoNI inpM2MIC1myc with the equivalent region in plasmidpPICZ�-TgMIC1NTT126A,T220A (see Ref. 1). To obtain aclonal line, parasites were cloned at least twice by limitingdilution from the drug-resistant pool of parasites obtainedfrom transfection and selection.Cloning of T. gondii MCPs—Restriction enzymes were pur-
chased from New England Biolabs. All primers are listed insupplemental Table 2. Accession numbers are listed in Table 1.TgMIC13, TgMCP3, and TgMCP4 were amplified by PCRfrom aT. gondii tachyzoite cDNApool. TgMIC13with or with-out the signal peptide was amplified using primer pairs MIC1/2–5/1047 andMIC1/2–6/1048 or MIC1/2–1/1147 andMIC1/2–2/1148, respectively. TgMCP3 was cloned using primersMIC1/3-7_1981 and MIC1/3-7_1982. To obtain the codingsequence for TgMCP4 with or without the signal peptide,primer pairs TgMIC1/4-7_1983 and TgMIC1/4-7_1984 as wellas TgMIC1/4-3_1847 and TgMIC1/4-4_1848 were used foramplification, respectively. The fragments were cloned intopGEM and sequenced. The sequence encoding full-lengthTgMIC13 was digested with EcoRI and SbfI and subcloned intopTUB8TgMLCTy_HX between EcoRI and NsiI restrictionsites.
Family of MAR Domain-containing Proteins in Apicomplexa
JANUARY 15, 2010 • VOLUME 285 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 2065 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from
pTUB8TgMLCTy_HXwas cutwith EcoRI, blunt-endedwithendonuclease, and cut again with NsiI prior to insertion of full-length TgMCP3 coding sequence cut from pGEM-TgMCP3with EcoRV and PstI. TgMCP3 without signal peptide (SP) wascloned into pROP1mycMIC1-GLD in between NsiI and PacI(blunt end cloning) resulting in pROP1mycTgMCP3(3430).
Full-length TgMCP4was cloned into pTUB8TgMLCTy_HXusing EcoRI and NsiI restriction sites. The sequence corre-sponding to TgMCP4 without SP was subcloned intopROP1mycMIC1-GLD using NsiI and PacI restriction sites.Constructs were used in transient transfection experiments
in either RH�Hx, mic1ko, or mic6ko parasites. Stable parasite
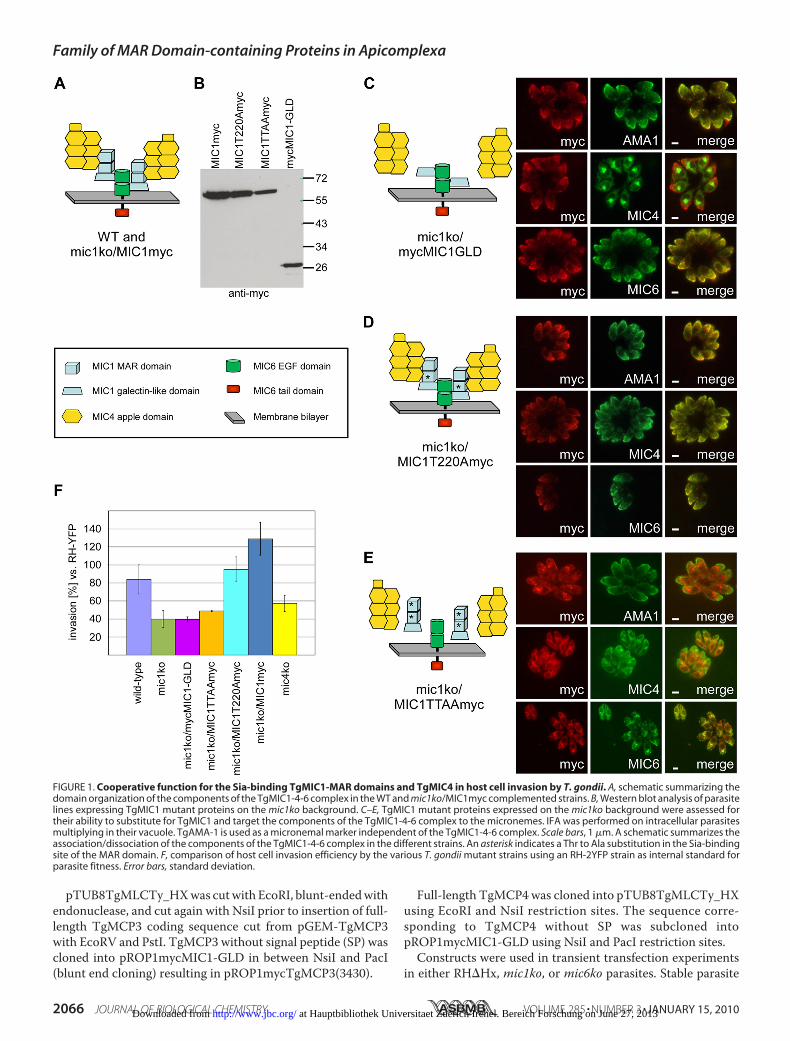
FIGURE 1. Cooperative function for the Sia-binding TgMIC1-MAR domains and TgMIC4 in host cell invasion by T. gondii. A, schematic summarizing thedomain organization of the components of the TgMIC1-4-6 complex in the WT and mic1ko/MIC1myc complemented strains. B, Western blot analysis of parasitelines expressing TgMIC1 mutant proteins on the mic1ko background. C–E, TgMIC1 mutant proteins expressed on the mic1ko background were assessed fortheir ability to substitute for TgMIC1 and target the components of the TgMIC1-4-6 complex to the micronemes. IFA was performed on intracellular parasitesmultiplying in their vacuole. TgAMA-1 is used as a micronemal marker independent of the TgMIC1-4-6 complex. Scale bars, 1 �m. A schematic summarizes theassociation/dissociation of the components of the TgMIC1-4-6 complex in the different strains. An asterisk indicates a Thr to Ala substitution in the Sia-bindingsite of the MAR domain. F, comparison of host cell invasion efficiency by the various T. gondii mutant strains using an RH-2YFP strain as internal standard forparasite fitness. Error bars, standard deviation.
Family of MAR Domain-containing Proteins in Apicomplexa
2066 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

lines were derived from transfection of the pTUB8TgMLCTy_HX-based vectors carrying the mycophenolic acid/xanthineresistance marker into the RH�Hx strain.Expression and Purification of MCPs in P. pastoris—Pichia
pastoris transformation and expression were performed usingpPICZ�-based plasmids according to the Pichia expression kitprotocols (Invitrogen). Transformation of the supplied hoststrain GS115 was performed by electroporation following lin-earization of the vector with PmeI, SacI, or DraI. To inhibitglycosylation, expression was occasionally carried out in thepresence of 10 �g/ml tunicamycin. Recombinant TgMIC13(amino acids 23–468) was either used directly in the form ofculture supernatant or purified on nickel-nitrilotriacetic acid(Qiagen) and concentrated into 20 mM NaH2PO4/Na2HPO4,pH 7.3. TgMCP3 (amino acids 54–565) and TgMCP4 (aminoacids 526–1016) were not secreted, but soluble �-factor fusionprotein was obtained by lysis of cells in 50 mM NaH2PO4/Na2HPO4, pH 7.4, 5% glycerol, 1 mM phenylmethylsulfonylfluoride.Preparation of Excreted Secreted Antigen (ESA) Fraction—T.
gondii tachyzoites freshly lysed from their host cells were har-vested by centrifugation at 240� g for 10min andwashed twicein IM (Dulbecco’s modified Eagle’s medium, 3% fetal bovineserum, 10 mM HEPES) prewarmed to 37 °C. A pellet of 2.0–4.88 � 108 parasites was resuspended in 1 ml of IM, and analiquot of 50 �l was taken as reference standard to allow esti-mation of the degree of secretion, and microneme secretionwas stimulated by adding 10 �l of 100% EtOH to the remaining950 �l. The sample was incubated for 10 min at room temper-ature, then 40min at 37 °C, and finally cooled down to 0 °C in anice-water bath for 5 min. Parasites were pelleted at 1000 � g,4 °C for 5 min. The supernatant was transferred to a new tubeand centrifuged again for 5 min, 4 °C at 2000 � g. The superna-tant (ESA) was collected and stored at �80 °C.Cell Binding Assays—These were performed as described
previously (1). Briefly, confluent monolayers of HFF cells wereblocked for 1 h at 4 °C with 1% bovine serum albumin in coldPBS, 1 mM CaCl2, 0.5 mM MgCl2 (CM-PBS). Excess bovineserum albumin was removed by two 5-min washes with ice-cold CM-PBS. The proteins to be assayed were then added inthe form of P. pastoris culture supernatant (�0.25�g in 250�l)or parasite ESA, together or not with different concentrationsof competitors (N-acetylneuraminic acid and glucuronic acidwere purchased from Sigma; stock solutions in PBS wereadjusted to pH 7.0), and diluted in cold CM-PBS to a total vol-ume of 500 �l. After incubation at 4 °C for 1 h, the supernatantwas removed, and the cells were washed four times for 5 minwith ice-cold CM-PBS. The cell-bound fraction was collectedby the direct addition of 50 �l of 1� SDS-PAGE loading bufferwith 0.1 M dithiothreitol. In some cases, prior to blocking, HFFcells were pretreated with 66 milliunits/ml �2–3, -6, -8 Vibriocholerae neuraminidase (RocheApplied Science) in RPMI 1640medium, 25 mM HEPES, L-glutamine (Invitrogen) for 1 h at37 °C in a total reaction volume of 1 ml.CHO-lec2 cells were purchased from the ATCC. All CHO
cells were cultured according to ATCC indications. C6 rat gli-oma cells were propagated in RPMI 1640medium (Invitrogen),5% fetal calf serum, 2 mM L-glutamine, 25 �g/ml gentamicin.
Invasion Assays—Comparison of different T. gondii strainsfor invasion efficiency was done using an RH-2YFP strain (21)as internal standard for parasite fitness. The details weredescribed previously (16). Total number of parasite vacuolesand RH-2YFP parasite vacuoles were counted on 20 micro-scopic fields on each IFA slide with a minimum of 440 vacuolesin total per slide.Only vacuoles containing at least two parasiteswere counted to be sure not to count extracellular parasites thatresisted washing. Each experiment was repeated at least threetimes. Statistical analysis was performed using PRISM.MicroarrayAnalyses of RecombinantMICProteins—The sia-
lyloligosaccharide microarrays were generated with 88 lipid-linked oligosaccharide probes (supplemental Table 1), whichwere arrayed in duplicate on nitrocellulose-coated glass slidesat 2 and 5 fmol per spot using a noncontact instrument (22).Analysis of carbohydrate binding of the recombinant His-taggedMICproteinswas performed essentially as described (1).In brief, each His-tagged MIC protein was precomplexed withmouse monoclonal anti-polyhistidine and biotinylated goatanti-mouse IgG antibodies (Sigma) in a ratio of 1:2.5:2.5 (byweight) and overlaid onto the arrays at 40 �g/ml for TgMIC1-MARR andTgMIC13 and 20�g/ml for NcMIC1-MARR. Bind-ing was detected using Alexa 647 fluor-conjugated streptavidin(Molecular Probes). Microarray data analysis and presentationwere carried out using dedicated software.7 The binding tooligosaccharide probes was dose-related, and results of 5 fmolper spot are shown.
RESULTS
TgMIC1 Is an Important Player in Sia-dependent Host CellInvasion by T. gondii—A previously generated TgMIC1 knock-out strain (mic1ko) showed a 50% reduction in invasion effi-ciency compared with the wild-type strain (13). As theTgMIC1-4-6 complex is disrupted in this strain, and the trans-port of TgMIC4 and TgMIC6 to the micronemes is ablatedresulting in their retention in the early secretory pathway (12),the contribution of the individual components to host cellinvasion has remained unresolved (for a schematic of the com-plex see Fig. 1A). To address this question, we complementedthe mic1ko strain with three different TgMIC1 mutant con-structs and then examined whether expression of these pro-teins is able to improve or restore invasion efficiency. A firstparasite mutant line named mic1ko/mycMIC1-GLDexpresses the TgMIC1 galectin-like domain (TgMIC1-GLD)on the mic1ko background. A second parasite line calledmic1ko/MIC1TTAAmyc expresses full-length TgMIC1 carryingThr to Ala substitutions at two positions (126 and 220) in thebinding site of each of the twoMARdomains, previously shownto be critical for the host cell binding activity of the TgMIC1-MAR region (TgMIC1-MARR) (1). A third complemented linecalled mic1ko/MIC1T220Amyc expresses TgMIC1 containingjust a single T220A substitution that also abolishes the host cellbinding activity of the protein (1). As a control, the mic1kostrain was complemented with full-length wild-type TgMIC1carrying a Myc tag epitope at the C terminus (parasite linemic1ko/MIC1myc). Expression of the respective mutant pro-
7 M. S. Stoll, unpublished data.
Family of MAR Domain-containing Proteins in Apicomplexa
JANUARY 15, 2010 • VOLUME 285 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 2067 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

teins was confirmed by Western blot (Fig. 1B). In addition, thepreviously generatedmic4ko line (12) was included in the anal-yses. Fig. 1F shows a comparison of invasion efficiency of theparental wild-type and the mic1ko lines and confirms the 50%reduction in invasion phenotype previously reported for themic1ko (13).Given that TgMIC4 and TgMIC6 are retained in the early
secretory pathway in themic1ko strain, assessment of the sub-cellular localization of the TgMIC1 mutant proteins, as well asof TgMIC4 andTgMIC6, in the complementedmic1ko parasitelines was important for interpretation of the invasion assayresults. Expression of TgMIC1myc in themic1ko strain rescuedthe targeting of TgMIC4 and TgMIC6 to the micronemes sup-plemental Fig. S1 (12). Invasion efficiency in this strain wasrestored to a level rather higher than the parental wild-type line(Fig. 1F), showing that a complex composed of endogenousTgMIC4 and TgMIC6 together with the epitope-taggedTgMIC1myc is functional in invasion. In agreement with pre-vious studies (15), expression of TgMIC1-GLD in the mic1kostrain brought TgMIC6 to the micronemes but TgMIC4remained in the endoplasmic reticulum (Fig. 1C). In the inva-sion assay (Fig. 1F), when compared with mic1ko, mic1ko/mycMIC1-GLD showed no rescue of phenotype, demonstrat-ing that the TgMIC6-TgMIC1-GLD mutant complex does notcontribute to invasion, likely due to the absence of adhesivedomains.In the mic1ko/MIC1T220Amyc line, TgMIC1T220A was tar-
geted to the micronemes, restoring proper trafficking ofTgMIC4 and TgMIC6 (Fig. 1D). This mutant invaded signifi-cantly better thanmic1ko/mycMIC1-GLD (Fig. 1F) despite thefact that the mutation in TgMIC1T220A abrogates the adhesiveproperties of the protein (1). This suggested that the invasionenhancement observed upon expression of this protein mightbe solely a result of its capacity to recruit TgMIC4 to the mem-brane-bound complex and/or residual weak binding of the firstTgMIC1 MAR domain not carrying a Thr to Ala substitution.Invasion efficiency of this line was similar to the wild type butlower than the control line mic1ko/MIC1myc, underlining theimportance of the adhesive properties of TgMIC1. In themic1ko/MIC1TTAAmyc line, TgMIC1TTAAwas foundmainly inthe early secretory pathway (Fig. 1E) probably due to incorrectfolding of this mutant protein, precluding corroboration of theabove results with the single T220A mutant. Therefore, it wasnot surprising that there was no improvement of invasion effi-ciency in this line compared with themic1ko (Fig. 1F).
In the mic4ko strain, the partial TgMIC1-6 complex wasdelivered to the micronemes (12). This strain invaded moreefficiently thanmic1ko andmic1ko/mycMIC1-GLD (t tests,p�0.05, respectively), confirming the critical role of TgMIC1-MARR in invasion (Fig. 1F). In addition the mic4ko strain didnot invade as well as the wild type (t test, p � 0.05), suggestinga role for TgMIC4 in invasion in agreement with our observa-tionswith themic1ko/MIC1T220Amyc line or, alternatively, thatthe presence of TgMIC4 impacts on the proper function of theMAR domains.Collectively, the characterization of thesemutants suggests a
cooperative function for TgMIC1-MARR and TgMIC4 inreceptor binding and confirms the absence of any adhesive
function for TgMIC6 and TgMIC1-GLD. The invasion experi-ments establish that TgMIC1 function and its contribution toefficient invasion resides in TgMIC1-MARR binding to sialy-lated glycoconjugates. Furthermore, our data support the ideathat TgMIC4 is an adhesin with an important function in hostcell invasion.T. gondii Possesses More Than One Sia-binding Factor
Involved in Host Cell Invasion—To address the question ofwhether other parasite lectins bind to host sialylated glycocon-jugates during invasion, we tested the effect of free NANA oninvasion by the mic1ko strain. Interestingly, this assay showedthat host cell invasion by the mic1ko strain was considerablyimpaired in the presence of free NANA (Fig. 2A), indicatingthat TgMIC1 is not the only Sia-binding parasite lectin contrib-uting to invasion by T. gondii tachyzoites. This result was cor-roborated by performing an invasion assay with neuramini-dase-treated host cells that resulted in a substantial reduction ofinvasion by themic1ko strain (Fig. 2B).Identification of a Novel Family of MCPs in Coccidia—In the
light of the above observations, we hypothesized that T. gondiihas at least one additional Sia-binding parasite lectin. A surveyof the T. gondii genome sequence revealed that it encodes afamily of four MCPs, herein named TgMIC1, TgMCP2,TgMCP3, andTgMCP4 (see Table 1, Fig. 3A, and supplementalFig. S2). The presence in all cases of a predicted N-terminal SPsuggests that these proteins are delivered to the secretorypathway. In contrast to TgMIC1, which includes two MARdomains (TgMIC1-MARR) followed by a galectin-likedomain (TgMIC1-GLD), the other three sequences containfour consecutive MAR domains but lack a galectin-likedomain. In addition, TgMCP4 possesses a novel N-terminalrepeat region (16 repeats, 17–22 amino acids per repeat),with the number of repeats varying between different strainsof T. gondii. Examination of the EST data set indicate that allthese MCP genes are transcribed both in tachyzoite and bra-dyzoite stages, and a proteomic study (23) showed theexpression of TgMCP2 in tachyzoites.
FIGURE 2. Cell invasion assays using the T. gondii mic1ko strain demon-strate the existence of at least one more Sia-specific parasite lectin.A, invasion by mic1ko parasites in the absence and presence of 10 and 20 mM
free NANA or galactose. B, assay comparing invasion of mic1ko parasites intohost cells (HFFs) pretreated or not with neuraminidase. Error bars, standarddeviation. Note that, compared with the parental strain, the mic1ko shows a50% reduced invasion phenotype (Fig. 1F), but its invasion efficiency was setto 100% here.
Family of MAR Domain-containing Proteins in Apicomplexa
2068 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from
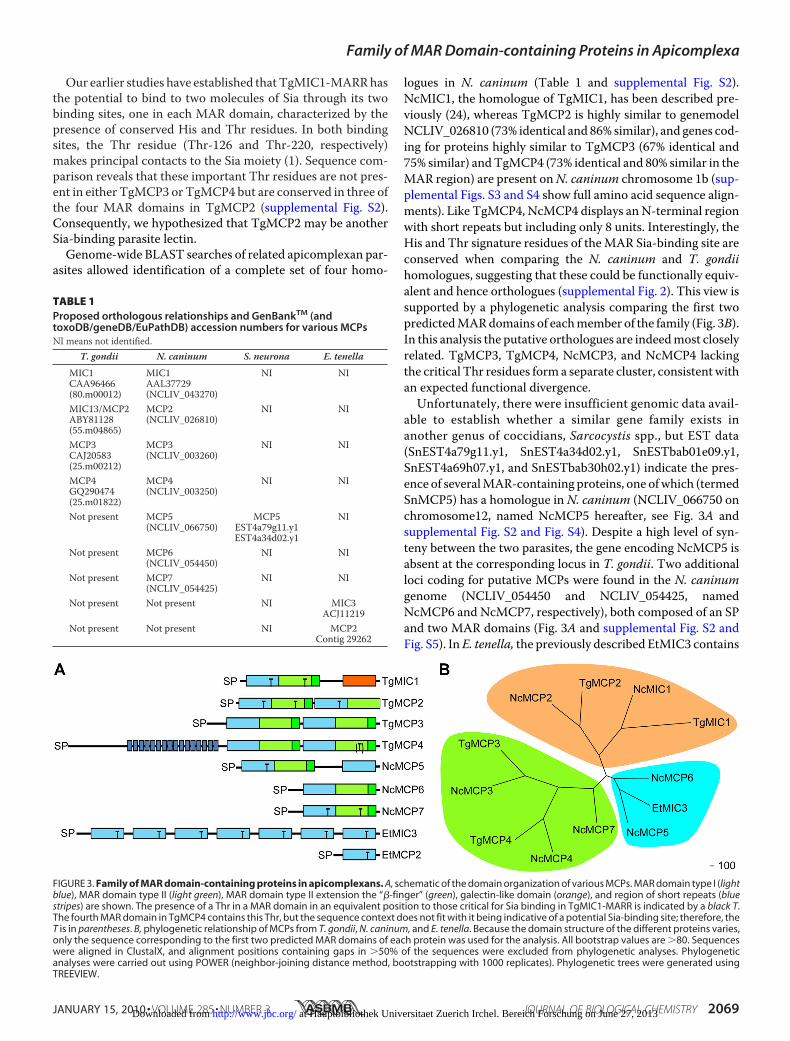
Our earlier studies have established thatTgMIC1-MARRhasthe potential to bind to two molecules of Sia through its twobinding sites, one in each MAR domain, characterized by thepresence of conserved His and Thr residues. In both bindingsites, the Thr residue (Thr-126 and Thr-220, respectively)makes principal contacts to the Sia moiety (1). Sequence com-parison reveals that these important Thr residues are not pres-ent in either TgMCP3 or TgMCP4 but are conserved in three ofthe four MAR domains in TgMCP2 (supplemental Fig. S2).Consequently, we hypothesized that TgMCP2 may be anotherSia-binding parasite lectin.Genome-wide BLAST searches of related apicomplexan par-
asites allowed identification of a complete set of four homo-
logues in N. caninum (Table 1 and supplemental Fig. S2).NcMIC1, the homologue of TgMIC1, has been described pre-viously (24), whereas TgMCP2 is highly similar to genemodelNCLIV_026810 (73% identical and 86% similar), and genes cod-ing for proteins highly similar to TgMCP3 (67% identical and75% similar) andTgMCP4 (73% identical and 80% similar in theMAR region) are present onN. caninum chromosome 1b (sup-plemental Figs. S3 and S4 show full amino acid sequence align-ments). Like TgMCP4, NcMCP4 displays anN-terminal regionwith short repeats but including only 8 units. Interestingly, theHis and Thr signature residues of theMAR Sia-binding site areconserved when comparing the N. caninum and T. gondiihomologues, suggesting that these could be functionally equiv-alent and hence orthologues (supplemental Fig. 2). This view issupported by a phylogenetic analysis comparing the first twopredictedMARdomains of eachmember of the family (Fig. 3B).In this analysis the putative orthologues are indeedmost closelyrelated. TgMCP3, TgMCP4, NcMCP3, and NcMCP4 lackingthe critical Thr residues form a separate cluster, consistentwithan expected functional divergence.Unfortunately, there were insufficient genomic data avail-
able to establish whether a similar gene family exists inanother genus of coccidians, Sarcocystis spp., but EST data(SnEST4a79g11.y1, SnEST4a34d02.y1, SnESTbab01e09.y1,SnEST4a69h07.y1, and SnESTbab30h02.y1) indicate the pres-ence of severalMAR-containing proteins, one ofwhich (termedSnMCP5) has a homologue in N. caninum (NCLIV_066750 onchromosome12, named NcMCP5 hereafter, see Fig. 3A andsupplemental Fig. S2 and Fig. S4). Despite a high level of syn-teny between the two parasites, the gene encoding NcMCP5 isabsent at the corresponding locus in T. gondii. Two additionalloci coding for putative MCPs were found in the N. caninumgenome (NCLIV_054450 and NCLIV_054425, namedNcMCP6 and NcMCP7, respectively), both composed of an SPand two MAR domains (Fig. 3A and supplemental Fig. S2 andFig. S5). In E. tenella, the previously described EtMIC3 contains
FIGURE 3. Family of MAR domain-containing proteins in apicomplexans. A, schematic of the domain organization of various MCPs. MAR domain type I (lightblue), MAR domain type II (light green), MAR domain type II extension the “�-finger” (green), galectin-like domain (orange), and region of short repeats (bluestripes) are shown. The presence of a Thr in a MAR domain in an equivalent position to those critical for Sia binding in TgMIC1-MARR is indicated by a black T.The fourth MAR domain in TgMCP4 contains this Thr, but the sequence context does not fit with it being indicative of a potential Sia-binding site; therefore, theT is in parentheses. B, phylogenetic relationship of MCPs from T. gondii, N. caninum, and E. tenella. Because the domain structure of the different proteins varies,only the sequence corresponding to the first two predicted MAR domains of each protein was used for the analysis. All bootstrap values are �80. Sequenceswere aligned in ClustalX, and alignment positions containing gaps in �50% of the sequences were excluded from phylogenetic analyses. Phylogeneticanalyses were carried out using POWER (neighbor-joining distance method, bootstrapping with 1000 replicates). Phylogenetic trees were generated usingTREEVIEW.
TABLE 1Proposed orthologous relationships and GenBankTM (andtoxoDB/geneDB/EuPathDB) accession numbers for various MCPsNI means not identified.
T. gondii N. caninum S. neurona E. tenella
MIC1 MIC1 NI NICAA96466 AAL37729(80.m00012) (NCLIV_043270)MIC13/MCP2 MCP2 NI NIABY81128 (NCLIV_026810)(55.m04865)MCP3 MCP3 NI NICAJ20583 (NCLIV_003260)(25.m00212)MCP4 MCP4 NI NIGQ290474 (NCLIV_003250)(25.m01822)Not present MCP5 MCP5 NI
(NCLIV_066750) EST4a79g11.y1EST4a34d02.y1
Not present MCP6 NI NI(NCLIV_054450)
Not present MCP7 NI NI(NCLIV_054425)
Not present Not present NI MIC3ACJ11219
Not present Not present NI MCP2Contig 29262
Family of MAR Domain-containing Proteins in Apicomplexa
JANUARY 15, 2010 • VOLUME 285 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 2069 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

seven MAR domains, in which repeats 3–5 are identical (25).EtMCP2, lying downstream of EtMIC3 on contig 29262, wasannotated in silico by comparison with EST data from sporu-lated oocyst and sporozoite stages. EtMCP2 is composed of anSP and a singleMAR domain (Fig. 3A and supplemental Fig. S2and Fig. S5). In addition, this parasite possesses a number ofhypothetical MCPs (two gene models, respectively, on contigs29652 and 14843). EST data support expression of at least oneof them in first generation merozoites. Our BLAST searchesfailed to identify anyMCPs in Plasmodium, Theileria, or Cryp-tosporidium. Accession numbers and proposed orthologousrelationships for all MCPs are depicted in Table 1.A comparison of MAR domains from Toxoplasma, Neos-
pora, and Sarcocystis highlights differences in disulfide bondpatterns between two tandemly arranged MAR domains in agiven MCP (supplemental Fig. S2). As in the TgMIC1-MARRprototype, the beginning of the second domain (referred to astype II) lacks a stretch of amino acids containing two cysteineresidues shown to participate in the formation of a disulfidebond in the TgMIC1-MARR crystal structure, but it possessesan additional C-terminal extension (with the notable exceptionof the second MAR domain in TgMCP2 and NcMCP2) that
adopts a �-hairpin (1). Curiously, inthis regard all the MAR domains inEtMIC3 and in all other predictedMCPs in Eimeria resemble the firstof the two tandemly arranged MARdomains, referred to as type I (Fig.3A and supplemental Fig. S2). Thissuggests that these proteins haveevolved differently in Eimeria com-paredwith theMCPs found inToxo-plasma, Neospora, and Sarcocystis.Therefore, despite the fact thatsome of the MCPs in Eimeria dis-play the binding site His-Thr or asimilar motif, these proteins mightpossess different properties andfunctions.TgMIC13, a Novel Sia-binding
Lectin—A parasite line expressinga C-terminal ty-epitope-taggedcopy of TgMCP2 was generated.TgMCP2ty was co-localized withTgMIC4 to the micronemes by IFAandwas renamedTgMIC13 accord-ing to the current nomenclaturestatus. IFA on wild-type parasitesusing antibodies raised againstrecombinant TgMIC13 expressedin P. pastoris confirmed localizationto the micronemes, although onlypartial co-localization with TgMIC3was observed (Fig. 4A). Analysis oflysates from wild type andWT/TgMIC13ty parasites byWest-ern blot indicated that TgMIC13migrates on SDS-PAGE as a single
protein species with an apparent molecular mass (56 kDa)somewhat higher than expected (49.1 kDa calculated from theamino acid sequence) (Fig. 4B). TgMIC13ty migrates slightlyfaster than endogenous TgMIC13. A similar discrepancybetween apparent molecular mass on SDS-PAGE and theexpected mass was observed for recombinant TgMIC13expressed in P. pastoris (Fig. 5A, 59 kDa on SDS-PAGE versus51.9 kDa expected). No glycosylation sites are predicted forTgMIC13, and mass spectrometric analysis of recombinantTgMIC13 confirmed that its truemass is close to that predicted(supplemental Fig. S7). Therefore, we conclude that the aber-rant migration behavior is related to intrinsic properties of theprotein. The solubility profile of TgMIC13 is identical toTgMIC4 and is consistent with it being a soluble protein withinthe parasite organelles (Fig. 4C,upper panel). To assesswhetherTgMIC13 displays cell binding activity, ESA prepared fromWT/TgMIC13ty parasites was tested in a cell binding assay; theproteinwas found to bind to the cell surface (Fig. 4C, lower panel).Furthermore, transient expression of TgMIC13ty in mic1ko andmic6ko recipient strains indicated that TgMIC13 traffics to themicronemes independently of theTgMIC1-4-6 complex anddoesnotassociatewith its components (Fig. 4D).These results establish
FIGURE 4. Characterization of TgMIC13 (TgMCP2) in T. gondii. A, IFA shows co-localization of TgMIC13 withmicroneme proteins in wild-type (wt) (top, confocal images) and in WT/TgMIC13ty parasites (bottom). B, West-ern blot analysis of endogenous and epitope-tagged TgMIC13 in wild-type and WT/TgMIC13ty parasites. Anextract of HFF cells was also loaded as a control. C, assessment of TgMIC13 solubility by fractionation (top). Cellbinding assays using ESA from WT/TgMIC13ty parasites (bottom). I, input; W, last of four washes; CB, cell-boundfraction. D, transient transfection of TgMIC13ty into the mic1ko and the mic6ko show correct localization ofTgMIC13ty to the micronemes. Scale bars, 1 �m.
Family of MAR Domain-containing Proteins in Apicomplexa
2070 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from
that expression of TgMIC13 cannot functionally rescue themic1ko, strongly suggesting that TgMIC13 belongs to a distinctcomplex.The cell binding activity of TgMIC13 was reproduced with
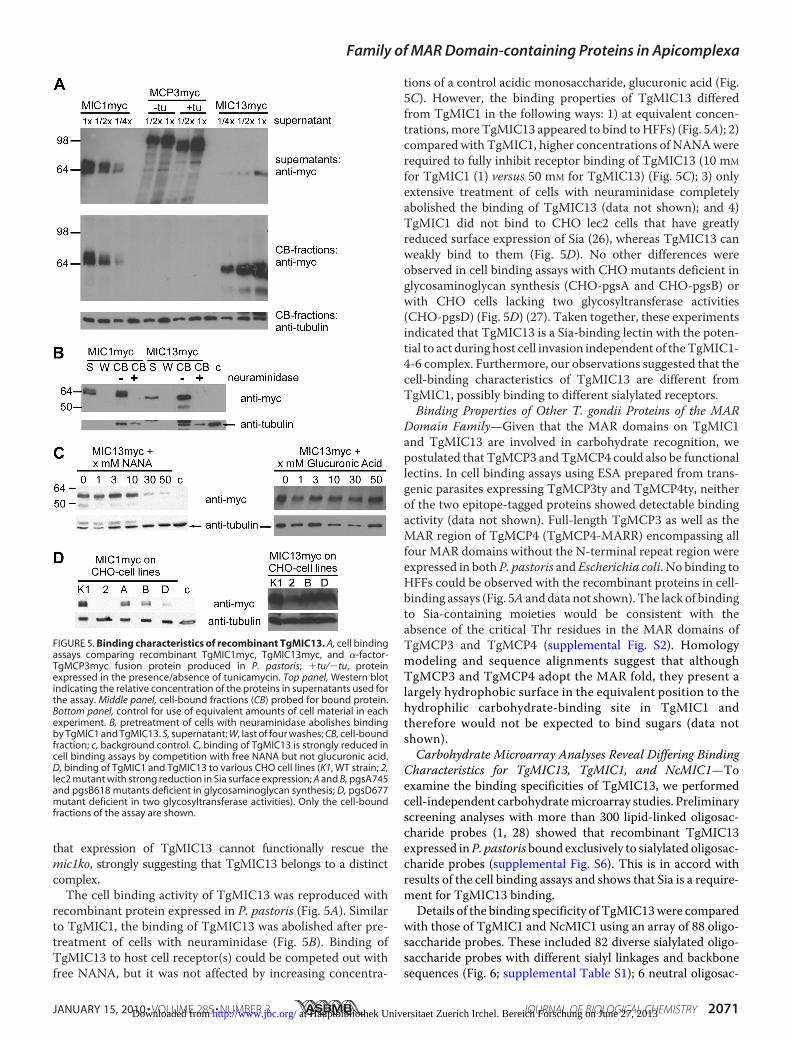
recombinant protein expressed in P. pastoris (Fig. 5A). Similarto TgMIC1, the binding of TgMIC13 was abolished after pre-treatment of cells with neuraminidase (Fig. 5B). Binding ofTgMIC13 to host cell receptor(s) could be competed out withfree NANA, but it was not affected by increasing concentra-
tions of a control acidic monosaccharide, glucuronic acid (Fig.5C). However, the binding properties of TgMIC13 differedfrom TgMIC1 in the following ways: 1) at equivalent concen-trations,more TgMIC13 appeared to bind toHFFs) (Fig. 5A); 2)compared with TgMIC1, higher concentrations of NANAwererequired to fully inhibit receptor binding of TgMIC13 (10 mM
for TgMIC1 (1) versus 50 mM for TgMIC13) (Fig. 5C); 3) onlyextensive treatment of cells with neuraminidase completelyabolished the binding of TgMIC13 (data not shown); and 4)TgMIC1 did not bind to CHO lec2 cells that have greatlyreduced surface expression of Sia (26), whereas TgMIC13 canweakly bind to them (Fig. 5D). No other differences wereobserved in cell binding assays with CHO mutants deficient inglycosaminoglycan synthesis (CHO-pgsA and CHO-pgsB) orwith CHO cells lacking two glycosyltransferase activities(CHO-pgsD) (Fig. 5D) (27). Taken together, these experimentsindicated that TgMIC13 is a Sia-binding lectin with the poten-tial to act during host cell invasion independent of theTgMIC1-4-6 complex. Furthermore, our observations suggested that thecell-binding characteristics of TgMIC13 are different fromTgMIC1, possibly binding to different sialylated receptors.Binding Properties of Other T. gondii Proteins of the MAR
Domain Family—Given that the MAR domains on TgMIC1and TgMIC13 are involved in carbohydrate recognition, wepostulated that TgMCP3 andTgMCP4 could also be functionallectins. In cell binding assays using ESA prepared from trans-genic parasites expressing TgMCP3ty and TgMCP4ty, neitherof the two epitope-tagged proteins showed detectable bindingactivity (data not shown). Full-length TgMCP3 as well as theMAR region of TgMCP4 (TgMCP4-MARR) encompassing allfour MAR domains without the N-terminal repeat region wereexpressed in bothP. pastoris andEscherichia coli. No binding toHFFs could be observed with the recombinant proteins in cell-binding assays (Fig. 5A anddata not shown). The lack of bindingto Sia-containing moieties would be consistent with theabsence of the critical Thr residues in the MAR domains ofTgMCP3 and TgMCP4 (supplemental Fig. S2). Homologymodeling and sequence alignments suggest that althoughTgMCP3 and TgMCP4 adopt the MAR fold, they present alargely hydrophobic surface in the equivalent position to thehydrophilic carbohydrate-binding site in TgMIC1 andtherefore would not be expected to bind sugars (data notshown).Carbohydrate Microarray Analyses Reveal Differing Binding
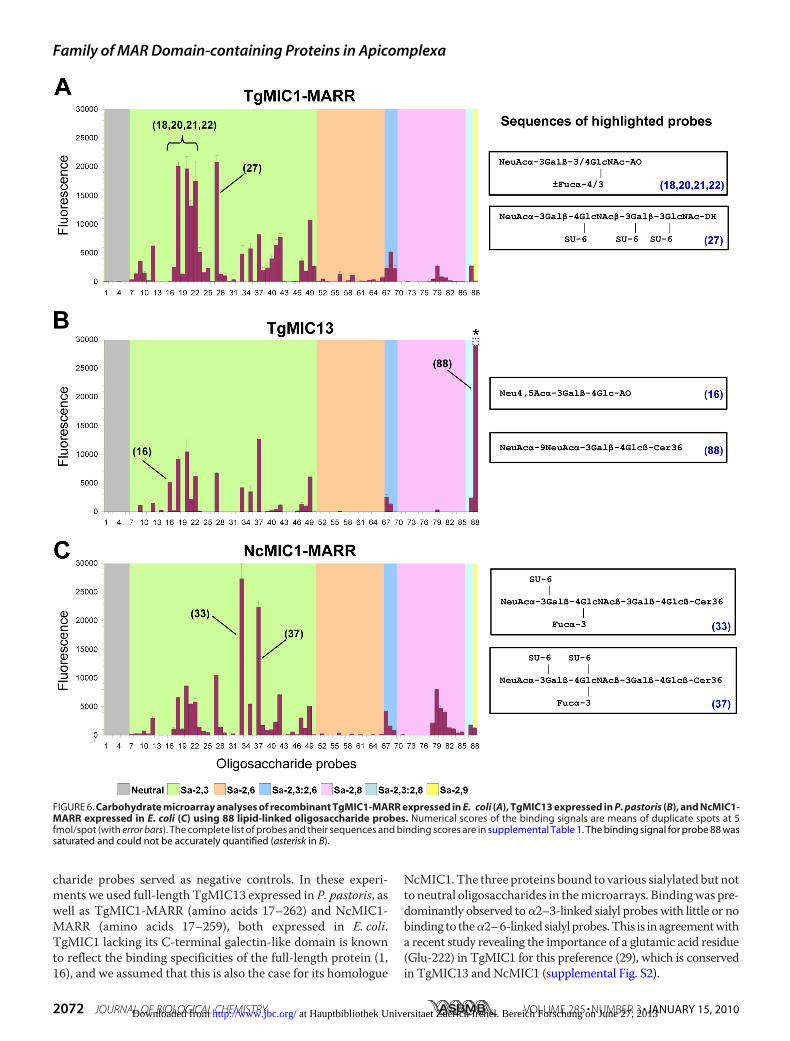
Characteristics for TgMIC13, TgMIC1, and NcMIC1—Toexamine the binding specificities of TgMIC13, we performedcell-independent carbohydratemicroarray studies. Preliminaryscreening analyses with more than 300 lipid-linked oligosac-charide probes (1, 28) showed that recombinant TgMIC13expressed inP. pastoris bound exclusively to sialylated oligosac-charide probes (supplemental Fig. S6). This is in accord withresults of the cell binding assays and shows that Sia is a require-ment for TgMIC13 binding.Details of the binding specificity of TgMIC13were compared
with those of TgMIC1 and NcMIC1 using an array of 88 oligo-saccharide probes. These included 82 diverse sialylated oligo-saccharide probes with different sialyl linkages and backbonesequences (Fig. 6; supplemental Table S1); 6 neutral oligosac-
FIGURE 5. Binding characteristics of recombinant TgMIC13. A, cell bindingassays comparing recombinant TgMIC1myc, TgMIC13myc, and �-factor-TgMCP3myc fusion protein produced in P. pastoris; �tu/�tu, proteinexpressed in the presence/absence of tunicamycin. Top panel, Western blotindicating the relative concentration of the proteins in supernatants used forthe assay. Middle panel, cell-bound fractions (CB) probed for bound protein.Bottom panel, control for use of equivalent amounts of cell material in eachexperiment. B, pretreatment of cells with neuraminidase abolishes bindingby TgMIC1 and TgMIC13. S, supernatant; W, last of four washes; CB, cell-boundfraction; c, background control. C, binding of TgMIC13 is strongly reduced incell binding assays by competition with free NANA but not glucuronic acid.D, binding of TgMIC1 and TgMIC13 to various CHO cell lines (K1, WT strain; 2,lec2 mutant with strong reduction in Sia surface expression; A and B, pgsA745and pgsB618 mutants deficient in glycosaminoglycan synthesis; D, pgsD677mutant deficient in two glycosyltransferase activities). Only the cell-boundfractions of the assay are shown.
Family of MAR Domain-containing Proteins in Apicomplexa
JANUARY 15, 2010 • VOLUME 285 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 2071 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from
charide probes served as negative controls. In these experi-ments we used full-length TgMIC13 expressed in P. pastoris, aswell as TgMIC1-MARR (amino acids 17–262) and NcMIC1-MARR (amino acids 17–259), both expressed in E. coli.TgMIC1 lacking its C-terminal galectin-like domain is knownto reflect the binding specificities of the full-length protein (1,16), and we assumed that this is also the case for its homologue
NcMIC1.The three proteins bound to various sialylated but notto neutral oligosaccharides in themicroarrays. Bindingwas pre-dominantly observed to�2–3-linked sialyl probes with little or nobinding to the�2–6-linkedsialyl probes.This is inagreementwitha recent study revealing the importance of a glutamic acid residue(Glu-222) in TgMIC1 for this preference (29), which is conservedin TgMIC13 and NcMIC1 (supplemental Fig. S2).
FIGURE 6. Carbohydrate microarray analyses of recombinant TgMIC1-MARR expressed in E. coli (A), TgMIC13 expressed in P. pastoris (B), and NcMIC1-MARR expressed in E. coli (C) using 88 lipid-linked oligosaccharide probes. Numerical scores of the binding signals are means of duplicate spots at 5fmol/spot (with error bars). The complete list of probes and their sequences and binding scores are in supplemental Table 1. The binding signal for probe 88 wassaturated and could not be accurately quantified (asterisk in B).
Family of MAR Domain-containing Proteins in Apicomplexa
2072 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

Interestingly, for each protein, a distinct binding profile wasobserved. Among the broad range of sialylated probes bound byTgMIC1-MARR, the preferred ligands are �2–3-linked sialyl-N-acetyllactosamine (3�SiaLacNAc) sequences with or withoutfucosylation/sulfation on the N-acetylglucosamine residue(highlighted probes in Fig. 6A). This is in overall agreementwith previous findings (1). Although the oligosaccharidesequences of probes 18, 20, 21, and 22 are short tri- or tetrasac-charides, their recognition demonstrates the effective presen-tation of these as oxime-linked neoglycolipids on the array sur-face (30).TgMIC13 bound to a more restricted spectrum of probes
compared with TgMIC1-MARR (Fig. 6B). Several 3�SiaLacNAc-based probes were bound, but there was little or no binding tosialyl sequences having long chain polyLacNAc or N-glycanbackbones (probes 38–41) and to �2–8-polysialyl sequences(probes 76–85). Strikingly, the strongest binding of TgMIC13was to a disialyllactose probe having �2–9 linkage between thetwo Sia residues (probe 88) (31). The binding signal for thisprobe was extremely high (saturated) at the protein concentra-tion tested. In addition, TgMIC13, but not TgMIC1, bound to alittle studied 4-O-acetylated 3�sialyllactose sequence as inprobe 16. It is interesting that the binding to probe 16 wasstronger than that to the non-O-acetylated analogue (probe 12).For NcMIC1-MARR, there was strikingly high binding to twosialyl Lex-related probes (probes 33 and 37, Fig. 6C), which havein common a sulfate group at the 6-position of the galactoseresidue. In contrast, if the sulfation is on the N-acetylglucos-amine residue as in probe 35, binding of NcMIC1-MARR wasmuch weaker. NcMIC1-MARR gave stronger binding signalsthan TgMIC1 and TgMIC13 with polysialyl sequences.TgMIC1-MARR Crystal Structure Provides Explanations for
the Findings inMicroarrayAnalyses—Tobetter understand thebasis of the binding preference revealed by themicroarray stud-ies, we compared the crystal structure of TgMIC1-MARR incomplex with 3�SiaLacNAc1–3 (Protein Data Bank code 3F5A)(29) to the TgMIC13 MAR domains. Although the first threeMAR domains of TgMIC13 possess the Sia-binding signature,we have assumed that the type II domains (MAR2) make thedominant contribution to Sia recognition as observed forTgMIC1-MARR (1); therefore, MAR2 of TgMIC1 andTgMIC13, in particular, was considered. In TgMIC1-MAR2,Lys-216 and Phe-169 from loops �1-�2 and �4-�5 are posi-tioned in proximity to the hydroxyl group on C4 of the Sia ring(Fig. 7A). In the equivalent MAR domain from TgMIC13, thebulky side chains of Lys-216 and Phe-169 are replaced bysmaller Ser and Thr residues, respectively, which together withamovement in backbone atoms could enable TgMIC13-MAR2to accommodate the additional acetyl group at the C4 of Sia.In a 2–9-linked disialyloligosaccharide, the two Sia moieties
are separated by three additional bonds comparedwith the sep-aration between Sia and galactose in 3�SiaLacNAc resulting inincreased flexibility and making it difficult to predict whichamino acids will interact. Assuming a similar mode of recogni-tion for the outermost Sia moiety, one possibility is that thelonger linkage would shift the second Sia to a position directlyover the end of helix II in TgMIC1-MAR2 (Fig. 7B). In TgMIC1,three consecutive Glu residues (Glu-221–223) cap this helix,
which might induce electrostatic repulsion of the negativelycharged carboxylate group and hinder any preference for 2–9-disialyl sugars. In TgMIC13, Glu-221 and -223 are replaced bypositively charged Arg and Lys residues, which would not onlyreduce any electrostatic repulsion but provide opportunities forthe formation of specific charge-charge interaction with thesecond Sia.The preference of NcMIC1-MARR for the two sialyl Lex-
related probes sulfated in the 6-position of galactose could beexplained by the presence of a positively charged Lys (Lys-176)inNcMIC1-MAR2,which is replaced by anAla inTgMIC1 (Fig.7C). By analogy to the complex of TgMIC1-MARR with3�SiaLacNAc1–3, Lys-176 of NcMIC1 would be spatially closelypositioned to the 6-position of galactose and could form a spe-cific charge-charge interaction with the negatively charged sul-fate. Lys-176 in NcMIC1 is not conserved in TgMIC13 but ispresent in NcMCP2.
DISCUSSION
Sia is abundant on glycoconjugates decorating the surface ofall vertebrate cells and is widely exploited by viruses and bacte-ria for host cell entry. In this study, we have characterized keyparasite molecular players involved in Sia-dependent host cellinvasion by T. gondii. The salient findings are as follows: first,the characterization of TgMIC1-4-6 complex as one importantplayer in Sia-dependent host cell invasion; second, the relianceof the parasite on an additional Sia-binding lectin for invasion,identified as TgMIC13; and third, the determination of distinctSia-dependent binding specificities for three members of anovel family of MCPs conserved in coccidians. In T. gondii, allthe fourMCPs are expressed in tachyzoites. Among them, onlyTgMIC1 and TgMIC13 possess the sequence characteristics ofa Sia-binding MAR domain, and only these specifically recog-nize sialyloligosaccharides.Role of TgMIC13 in Host Cell Invasion by T. gondii—To con-
tribute to host cell invasion, TgMIC13 needs to be targetedaccurately to the micronemes and then delivered in a timelyfashion to become firmly anchored on the parasite surface.Because TgMIC13 does not contain a predicted transmem-brane domain or glycosylphosphatidylinositol-anchor motif, itlikely belongs to a yet undefined complex. Expression ofTgMIC13 in themic1ko andmic6ko strains ruled out any inter-action with the TgMIC1-4-6 complex. TgMIC13 does not con-tain a galectin-like domain like TgMIC1, which precludes asimilar architecture for the putative TgMIC13-containingcomplex.Unraveling themechanismofTgMIC13 sorting to themicronemes must await the identification of its escorter pro-tein, as described for other solublemicronemeproteins (12, 14).To assess precisely the role of TgMIC13 in invasion, we
repeatedly tried to knock-out the gene in wild-type (RH strain)and inmic1ko parasites without success using a knock-out cas-sette with 5-kb flanking elements homologous to the TgMIC135�- and 3�-UTRs.8 In addition, knock-out attempts8 failed in a�-ku80 strain efficiently amenable to gene disruption by dra-matically enhanced frequency of homologous recombination
8 N. Friedrich, J. Santos, and D. Soldati-Favre, unpublished data.
Family of MAR Domain-containing Proteins in Apicomplexa
JANUARY 15, 2010 • VOLUME 285 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 2073 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

(32, 33). These results suggest that this gene might play animportant role for parasite propagation.Distinct Binding Specificities Might Determine Tissue Tro-
pism of Parasitic Infections—With carbohydrate microarraytechnology, we examined details of carbohydrate-binding spec-ificities of TgMIC13 in comparison with MAR regions ofTgMIC1 andNcMIC1. The preponderant binding activity of allthree proteins was to �2–3-linked sialyl probes. However, therelative binding intensities to specific sialyl probes were quitedifferent for the three proteins. TgMIC13 bound to two sialyl-oligosaccharide probes, namely the 4-O-acetylated sialyllactoseand the �2–9-linked disiayl sequences, which were not recog-nized by TgMIC1 or NcMIC1. The 4-O-acetyl-substituted Siahas been described in various tissues in mice, especially in thegut; its presence has been documented in a number of otheranimal species and in trace amounts in humans (34, 35). Theresistance of 4-O-acetyl-substituted Sia toward neuramini-dases from V. cholerae and Clostridium perfringens has previ-ously been described (36, 37), but the susceptibility of this Siaform to neuraminidases requires further detailed investigation.
There is so far limited information on the occurrence of 2–9-linked Sia in animals. It has been reported as a component on amurine neuroblastoma cell line (38) and a human embryonalcarcinoma cell line (39). The distribution of �2–9-linked disia-lyllactose in HFFs, C6 rat glioma, and CHO cells has not beendescribed; this and its susceptibility to neuraminidases will bethe subject of future investigation.A special feature of NcMIC1 as revealed by carbohydrate
microarray analyses is the strong binding to the two sulfatedsialyl Lex-related probes. Sulfation was previously reported toplay a role in cell surface binding byNcMIC1 (24); in that study,glycosaminoglycans were proposed to be targeted by NcMIC1;however, the influence of Sia was not investigated. The distinctbinding specificities of the three MIC proteins might haveimplications for tissue tropism, and the cellular targets for theseproteins warrant further investigation.In general, TgMIC1 and TgMIC13 bind with high affinity to
3�SiaLacNAc sequences. Considering the potential spectrumofsialyloligosaccharides that can be bound by these two lectins,and the wide distribution of Sia in vertebrate tissues, it is likely
FIGURE 7. Three orientations of the crystal structure of TgMIC1-MARR in complex with 3�SiaLacNAc1–3 (Protein Data Bank code 3F5A) (29);3�SiaLacNAc1–3 is shown in an orange stick representation. Key side chains are drawn as stick representations minus attached protons and labeled withsequence position. A and B, amino acid differences between TgMIC1 and TgMIC13 relevant to their binding specificities are labeled in cyan and violet,respectively. C, amino acid differences between TgMIC1 and NcMIC1 relevant to their binding specificities are labeled in cyan and violet, respectively.
Family of MAR Domain-containing Proteins in Apicomplexa
2074 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

that the two proteins will bind to most cell types. This is inmarked contrast to the P. falciparum erythrocyte-binding anti-gen 175 (PfEBA175), which binds selectively to a unique sialy-lated glycoprotein, glycophorin A (40, 41). Our results suggestcooperativity between TgMIC1 and TgMIC4 in receptor bind-ing by the TgMIC1-4-6 complex. It will be interesting to inves-tigate if receptor recognition of TgMIC1 is influenced by itsassociation with TgMIC4.Is There a Molecular Basis for Invasion of a Broad Range of
Cells?—The presence of the MAR domain-containing proteinsin Toxoplasma, Neospora, Sarcocystis, and Eimeria parasitesand its absence in Plasmodium, Theileria, and Cryptospo-ridium suggest that this domain is exclusively conserved amongthe Coccidia. We hypothesize that sialylated receptors may beexploited as a common target by a subset of MCPs carrying theappropriate binding motif in this group of enteroparasites. Assuggested in our study, not all MCPs would bind to Sia or evento host cells, which raises questions regarding their alternativefunctions. We have identified a complete set of homologues ofT. gondii MCPs in N. caninum, a parasite causing significanteconomic loss by infecting cattle. T. gondii and N. caninumbelong to a group of tissue-cyst-forming coccidians that alsoinclude the genera Hammonida, Besnoita, Sarcocystis, andFrenkelia. Similar to T. gondii tachyzoites, N. caninumtachyzoites and S. neuronamerozoites are known to be promis-cuous pathogens that are able to infect a large variety of cells(42, 43). This is probably related to their common behavior ofspreading through tissues during acute infection prior to cystformation. Considering the fact that the MAR family isrestricted to enteroparasites, these adhesins might also be ofimportance in the context of the natural route of infection viathe intestine. To our knowledge, no study has so far addressedthe importance of receptor-ligand interactions for invasionof the midgut epithelium.It is tempting to speculate that the MAR family of lectins is
instrumental to the infection of such a broad range of cells.Several lines of evidence support the involvement of TgMIC1and TgMIC13 and more in general MCPs. First, their bindingcharacteristics are suited for engagement with a variety of cellsurface sialyl-glycoconjugates, which are widely distributed invertebrate tissues. Second, a strain deficient in TgMIC1 andTgMIC3 displays decreased virulence in vivo (13). Importantly,infection of mice with this attenuated strain results in reducedparasite load in a variety of tissues compared with the parentalstrain (44). Third, as discussed above, theMARdomain appearsto be restricted to coccidians. Of note, E. tenella is a coccidianthat does not spread through tissues and does not form tissue-cysts but nevertheless also expresses at least oneMCP, EtMIC3(25). Furthermore, EST and genomic data indicate that moreMCPs might be expressed. However, our sequence and phylo-genetic analyses indicate thatEimeriaMCPs are divergent fromthose present in the cyst-forming coccidians. Therefore, theymight serve a different function.In addition to TgMIC1 and TgMIC13, numerous proteins
are known to act on the surface of T. gondii and might as wellcontribute to the phenomenon of promiscuous host cell inva-sion. In particular, proteins containing coccidian-specificdomains other than MAR should be considered. Interestingly,
the apple domain, present in TgMIC4, is specific to coccidians(45), although a divergent fold belonging to the plasminogenapple nematode superfamily is present in apical membraneantigen-1 homologues from all Apicomplexans (46, 47). Fur-thermore, our BLAST searches indicate that the chitin binding-like domain is restricted to coccidians. The chitin binding-likedomain is present in TgMIC3 and TgMIC8, which formanother major complex involved in invasion. Dimerization ofthese chitin binding-like domains allows host cell binding andhas been associated with parasite virulence (13, 48). Intrigu-ingly, a synergy effect has been demonstrated between thiscomplex and TgMIC1-4-6 (13). However, for most proteincomplexes functioning in invasion, the timing and coordina-tion of action are only beginning to be understood. In summaryour work provides detailed insights into recognition of a broadhost cell range by cyst-forming enteroparasites during earlystages of invasion and raises the possibility that glycomimeticdrugs might be useful to reduce parasite burden in tissues dur-ing acute infection.
Acknowledgments—We thank C. Cherbuliez and N. Klages for excel-lent technical assistance; N. Jemmely for the generation of the anti-TgAMA-1 antibodies; W. Chai and Y. Zhang for mass spectrometricanalyses of oligosaccharide probes; R. Childs for the microarrays; andM. P. Stoll for the microarray analyses software. We thank Dr. C.Rodriguez (University of Oviedo), Dr. F. Tomley (Institute for AnimalHealth, Compton), andDr. A.Hemphill (University of Bern) for kindlyproviding us with the C6 rat glioma cell line, CHO cells, and NcMIC1cDNA, respectively.We also thankM.Messer andT.Urashima for the4-O-acetylated sialyllactose used in themicroarray analyses.We alsoacknowledge Dr. A. Pain and Dr. A. Reid (Sanger Institute, Hinxton,UK) for integrating our gene models into public data bases.
REFERENCES1. Blumenschein, T. M., Friedrich, N., Childs, R. A., Saouros, S., Carpenter,
E. P., Campanero-Rhodes, M. A., Simpson, P., Chai,W., Koutroukides, T.,Blackman, M. J., Feizi, T., Soldati-Favre, D., and Matthews, S. (2007)EMBO J. 26, 2808–2820
2. Persson, K. E., McCallum, F. J., Reiling, L., Lister, N. A., Stubbs, J., Cow-man, A. F.,Marsh, K., and Beeson, J. G. (2008) J. Clin. Invest. 118, 342–351
3. Sim, B. K., Orlandi, P. A., Haynes, J. D., Klotz, F. W., Carter, J. M., Camus,D., Zegans, M. E., and Chulay, J. D. (1990) J. Cell Biol. 111, 1877–1884
4. Takabatake, N., Okamura, M., Yokoyama, N., Ikehara, Y., Akimitsu, N.,Arimitsu, N., Hamamoto, H., Sekimizu, K., Suzuki, H., and Igarashi, I.(2007) Vet. Parasitol. 148, 93–101
5. Martin, M. J., Rayner, J. C., Gagneux, P., Barnwell, J. W., and Varki, A.(2005) Proc. Natl. Acad. Sci. U.S.A. 102, 12819–12824
6. Montoya, J. G., and Liesenfeld, O. (2004) Lancet 363, 1965–19767. Carruthers, V. B., and Sibley, L. D. (1997) Eur. J. Cell Biol. 73, 114–1238. Soldati-Favre, D. (2008) Parasite 15, 197–2059. Carruthers, V., and Boothroyd, J. C. (2007) Curr. Opin. Microbiol. 10,
83–8910. Carruthers, V. B., and Tomley, F. M. (2008) Subcell. Biochem. 47, 33–4511. Brecht, S., Carruthers, V. B., Ferguson, D. J., Giddings, O. K., Wang, G.,
Jakle, U., Harper, J. M., Sibley, L. D., and Soldati, D. (2001) J. Biol. Chem.276, 4119–4127
12. Reiss, M., Viebig, N., Brecht, S., Fourmaux, M. N., Soete, M., Di Cristina,M., Dubremetz, J. F., and Soldati, D. (2001) J. Cell Biol. 152, 563–578
13. Cerede, O., Dubremetz, J. F., Soete, M., Deslee, D., Vial, H., Bout, D., andLebrun, M. (2005) J. Exp. Med. 201, 453–463
14. Meissner,M., Reiss,M., Viebig,N., Carruthers, V. B., Toursel, C., Tomavo,S., Ajioka, J. W., and Soldati, D. (2002) J. Cell Sci. 115, 563–574
Family of MAR Domain-containing Proteins in Apicomplexa
JANUARY 15, 2010 • VOLUME 285 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 2075 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

15. Saouros, S., Edwards-Jones, B., Reiss,M., Sawmynaden, K., Cota, E., Simp-son, P., Dowse, T. J., Jakle, U., Ramboarina, S., Shivarattan, T., Matthews,S., and Soldati-Favre, D. (2005) J. Biol. Chem. 280, 38583–38591
16. Sawmynaden, K., Saouros, S., Friedrich, N., Marchant, J., Simpson, P.,Bleijlevens, B., Blackman,M. J., Soldati-Favre, D., andMatthews, S. (2008)EMBO Rep. 9, 1149–1155
17. Huynh,M.H., Opitz, C., Kwok, L. Y., Tomley, F.M., Carruthers, V. B., andSoldati, D. (2004) Cell. Microbiol. 6, 771–782
18. Carruthers, V. B., Håkansson, S., Giddings, O. K., and Sibley, L. D. (2000)Infect. Immun. 68, 4005–4011
19. Monteiro, V. G., Soares, C. P., and de Souza, W. (1998) FEMS Microbiol.Lett. 164, 323–327
20. Ortega-Barria, E., and Boothroyd, J. C. (1999) J. Biol. Chem. 274,1267–1276
21. Gubbels, M. J., Li, C., and Striepen, B. (2003) Antimicrob. Agents Che-mother. 47, 309–316
22. Palma, A. S., Feizi, T., Zhang, Y., Stoll, M. S., Lawson, A. M., Díaz-Rodríguez, E., Campanero-Rhodes, M. A., Costa, J., Gordon, S., Brown,G. D., and Chai, W. G. (2006) J. Biol. Chem. 281, 5771–5779
23. Xia, D., Sanderson, S. J., Jones, A. R., Prieto, J. H., Yates, J. R., Bromley, E.,Tomley, F.M., Lal, K., Sinden, R. E., Brunk, B. P., Roos, D. S., andWastling,J. M. (2008) Genome Biol. 9, R116
24. Keller, N., Naguleswaran, A., Cannas, A., Vonlaufen, N., Bienz, M., Bjork-man, C., Bohne, W., and Hemphill, A. (2002) Infect. Immun. 70,3187–3198
25. Labbe, M., de Venevelles, P., Girard-Misguich, F., Bourdieu, C., Guil-laume, A., and Pery, P. (2005)Mol. Biochem. Parasitol. 140, 43–53
26. Deutscher, S. L., Nuwayhid, N., Stanley, P., Briles, E. I., and Hirschberg,C. B. (1984) Cell 39, 295–299
27. Esko, J. D., Rostand, K. S., andWeinke, J. L. (1988) Science 241, 1092–109628. Feizi, T., and Chai, W. (2004) Nat. Rev. Mol. Cell Biol. 5, 582–58829. Garnett, J. A., Liu, Y., Leon, E., Allman, S. A., Friedrich, N., Saouros, S.,
Curry, S., Soldati-Favre, D., Davis, B. G., Feizi, T., andMatthews, S. (2009)Protein Sci. 18, 1935–1947
30. Liu, Y., Feizi, T., Campanero-Rhodes, M. A., Childs, R. A., Zhang, Y.,Mulloy, B., Evans, P. G., Osborn, H. M., Otto, D., Crocker, P. R., and Chai,
W. (2007) Chem. Biol. 14, 847–85931. Hasegawa, A., Ogawa, H., and Kiso, M. (1992) Carbohydr. Res. 224,
185–19232. Fox, B. A., Ristuccia, J. G., Gigley, J. P., and Bzik, D. J. (2009) Eukaryot. Cell
8, 520–52933. Huynh, M. H., and Carruthers, V. B. (2009) Eukaryot. Cell 8, 530–53934. Iwersen, M., Dora, H., Kohla, G., Gasa, S., and Schauer, R. (2003) Biol.
Chem. 384, 1035–104735. Rinninger, A., Richet, C., Pons, A., Kohla, G., Schauer, R., Bauer, H. C.,
Zanetta, J. P., and Vlasak, R. (2006) Glycoconj. J. 23, 73–8436. Matrosovich, M. N., Gambaryan, A. S., and Chumakov, M. P. (1992) Vi-
rology 188, 854–85837. Messer, M. (1974) Biochem. J. 139, 415–42038. Inoue, S., Poongodi, G. L., Suresh, N., Jennings, H. J., and Inoue, Y. (2003)
J. Biol. Chem. 278, 8541–854639. Fukuda, M. N., Dell, A., Oates, J. E., and Fukuda, M. (1985) J. Biol. Chem.
260, 6623–663140. Orlandi, P. A., Klotz, F. W., and Haynes, J. D. (1992) J. Cell Biol. 116,
901–90941. Tolia, N. H., Enemark, E. J., Sim, B. K., and Joshua-Tor, L. (2005)Cell 122,
183–19342. Ellison, S. P., Greiner, E., andDame, J. B. (2001)Vet. Parasitol. 95, 251–26143. Hemphill, A., Vonlaufen, N., and Naguleswaran, A. (2006) Parasitology
133, 261–27844. Moire, N., Dion, S., Lebrun, M., Dubremetz, J. F., and Dimier-Poisson, I.
(2009) Exp. Parasitol. 123, 111–11745. Chen, Z., Harb, O. S., and Roos, D. S. (2008) PLoS One 3, e361146. Chesne-Seck, M. L., Pizarro, J. C., Vulliez-Le Normand, B., Collins, C. R.,
Blackman, M. J., Faber, B. W., Remarque, E. J., Kocken, C. H., Thomas,A. W., and Bentley, G. A. (2005)Mol. Biochem. Parasitol. 144, 55–67
47. Pizarro, J. C., Vulliez-Le Normand, B., Chesne-Seck, M. L., Collins, C. R.,Withers-Martinez, C., Hackett, F., Blackman, M. J., Faber, B. W., Re-marque, E. J., Kocken, C. H., Thomas, A. W., and Bentley, G. A. (2005)Science 308, 408–411
48. Cerede, O., Dubremetz, J. F., Bout, D., and Lebrun,M. (2002) EMBO J. 21,2526–2536
Family of MAR Domain-containing Proteins in Apicomplexa
2076 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 3 • JANUARY 15, 2010 at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from

Citations http://www.jbc.org/content/285/3/2064#otherarticles
This article has been cited by 5 HighWire-hosted articles:
at Hauptbibliothek Universitaet Zuerich Irchel. Bereich Forschung on June 27, 2013http://www.jbc.org/Downloaded from
Copyright © 2022 FDOKUMEN




















