
Medicarpin, a legume phytoalexin, stimulates osteoblast differentiation and promotes peak bone mass...
10
RESEARCH ARTICLE Identification of kaempferol-regulated proteins in rat calvarial osteoblasts during mineralization by proteomics Avinash Kumar 1 , Anand K. Singh 2 , Abnish K. Gautam 1 , Deepak Chandra 3 , Divya Singh 1 , Bendangla Changkija 1 , Mahendra Pratap Singh 2 and Ritu Trivedi 1 1 Division of Endocrinology, Central Drug Research Institute, Chattar Manzil Palace, Lucknow, India 2 Toxicogenomics and Predictive Toxicology Division, Indian Institute of Toxicology Research, Mahatma Gandhi Marg, Lucknow, India 3 Department of Biochemistry, Lucknow University, Lucknow, India Received: September 22, 2009 Revised: January 7, 2010 Accepted: January 20, 2010 Kaempferol, a flavonoid, promotes osteoblast mineralization in vitro and bone formation in vivo; however, its mechanism of action is yet unknown. We adopted proteomic approach to identify the differential effect of kaempferol on rat primary calvarial osteoblasts during mineralization. The primary rat calvarial osteoblasts were treated with kaempferol (5.0 mM) for 9 days under mineralizing condition that resulted in significant increase in alkaline phosphatase activity and mineralization of the cells. Further, 2-D analysis of the kaempferol-treated osteoblast lysates revealed 18 differentially expressed proteins (nine upregulated and nine downregulated) on the basis of 4/o2.0-fold as cut-off (po0.01) that were then identified by MALDI-TOF MS. These included cytoskeletal proteins, intracellular signaling protein, chaperone, extracellular matrix protein, and proteins involved in glycolysis and cell–matrix interactions. Proteomics data were confirmed by Western blotting and quantitative real-time PCR by randomly selecting two upregulated and two downregulated proteins. Western blot analysis confirmed upregulation of HSP-70 and cytokeratin-14 levels, and downregulation of aldose reductase and caldesmon expression. We further demonstrated that kaempferol treatment inhibits aldose reductase activity in osteoblasts indicating an altered cellular metabolism by decelerating polyol pathway that was associated with the kaempferol-induced osteoblast mineralization. In conclusion, this is a first comprehensive study on the differential regulation of proteins by kaempferol in primary osteoblast, which would further help to elucidate the role of the identified proteins in the process of osteoblast mineralization. Keywords: Aldose reductase / Animal proteomics / Caldesmon / Cytokeratin-14 / HSP-70 / Osteoblast 1 Introduction Osteoblast differentiation is the crucial event for bone formation [1]. Primary calvarial cells from neonatal rats or mice can form mineralized matrix in vitro, when induced by L-ascorbate and b-glycerophosphate [2–4]. Robust demand for energy is placed on osteoblasts during the production of mineralized matrix of bone [5, 6]. Studies show that mineralization at the time of bone formation involves tightly regulated expression of several proteins [7–9]. Expression of these proteins follows temporal patterns during successive developmental stages. For example, cell cycle genes, histone H3 and H4, and ribosomal S6 exhibit higher expression levels during proliferative stage, while type I collagen, fibronectin, tumor growth factor-b and osteonectin levels are higher during matrix maturation stage [10]. However, little is known about the molecular mechanism by which Abbreviations: ALP, alkaline phosphatase; AR, aldose reductase; K, kaempferol; PDI, disulfide isomerase; qPCR, quantitative real- time PCR; SDH, sorbitol dehydrogenase Correspondence: Ritu Trivedi, Division of Endocrinology, Central Drug Research Institute, Chattar Manzil Palace, Post Box 173, Lucknow 226 001, India E-mail: [email protected] Fax: 191-522-2623938 & 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com 1730 Proteomics 2010, 10, 1730–1739 DOI 10.1002/pmic.200900666
Transcript of Medicarpin, a legume phytoalexin, stimulates osteoblast differentiation and promotes peak bone mass...

RESEARCH ARTICLE
Identification of kaempferol-regulated proteins in rat
calvarial osteoblasts during mineralization
by proteomics
Avinash Kumar1, Anand K. Singh2, Abnish K. Gautam1, Deepak Chandra3, Divya Singh1,Bendangla Changkija1, Mahendra Pratap Singh2 and Ritu Trivedi1
1 Division of Endocrinology, Central Drug Research Institute, Chattar Manzil Palace, Lucknow, India2 Toxicogenomics and Predictive Toxicology Division, Indian Institute of Toxicology Research,
Mahatma Gandhi Marg, Lucknow, India3 Department of Biochemistry, Lucknow University, Lucknow, India
Received: September 22, 2009
Revised: January 7, 2010
Accepted: January 20, 2010
Kaempferol, a flavonoid, promotes osteoblast mineralization in vitro and bone formation in vivo;
however, its mechanism of action is yet unknown. We adopted proteomic approach to identify
the differential effect of kaempferol on rat primary calvarial osteoblasts during mineralization.
The primary rat calvarial osteoblasts were treated with kaempferol (5.0mM) for 9 days under
mineralizing condition that resulted in significant increase in alkaline phosphatase activity and
mineralization of the cells. Further, 2-D analysis of the kaempferol-treated osteoblast lysates
revealed 18 differentially expressed proteins (nine upregulated and nine downregulated) on the
basis of 4/o2.0-fold as cut-off (po0.01) that were then identified by MALDI-TOF MS. These
included cytoskeletal proteins, intracellular signaling protein, chaperone, extracellular matrix
protein, and proteins involved in glycolysis and cell–matrix interactions. Proteomics data were
confirmed by Western blotting and quantitative real-time PCR by randomly selecting two
upregulated and two downregulated proteins. Western blot analysis confirmed upregulation of
HSP-70 and cytokeratin-14 levels, and downregulation of aldose reductase and caldesmon
expression. We further demonstrated that kaempferol treatment inhibits aldose reductase
activity in osteoblasts indicating an altered cellular metabolism by decelerating polyol pathway
that was associated with the kaempferol-induced osteoblast mineralization. In conclusion, this is
a first comprehensive study on the differential regulation of proteins by kaempferol in primary
osteoblast, which would further help to elucidate the role of the identified proteins in the process
of osteoblast mineralization.
Keywords:
Aldose reductase / Animal proteomics / Caldesmon / Cytokeratin-14 / HSP-70 /
Osteoblast
1 Introduction
Osteoblast differentiation is the crucial event for bone
formation [1]. Primary calvarial cells from neonatal rats or
mice can form mineralized matrix in vitro, when induced by
L-ascorbate and b-glycerophosphate [2–4]. Robust demand
for energy is placed on osteoblasts during the production of
mineralized matrix of bone [5, 6]. Studies show that
mineralization at the time of bone formation involves tightly
regulated expression of several proteins [7–9]. Expression of
these proteins follows temporal patterns during successive
developmental stages. For example, cell cycle genes, histone
H3 and H4, and ribosomal S6 exhibit higher expression
levels during proliferative stage, while type I collagen,
fibronectin, tumor growth factor-b and osteonectin levels are
higher during matrix maturation stage [10]. However, little
is known about the molecular mechanism by which
Abbreviations: ALP, alkaline phosphatase; AR, aldose reductase;
K, kaempferol; PDI, disulfide isomerase; qPCR, quantitative real-
time PCR; SDH, sorbitol dehydrogenase
Correspondence: Ritu Trivedi, Division of Endocrinology, Central
Drug Research Institute, Chattar Manzil Palace, Post Box 173,
Lucknow 226 001, India
E-mail: [email protected]
Fax: 191-522-2623938
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
1730 Proteomics 2010, 10, 1730–1739DOI 10.1002/pmic.200900666

mineralization of bone matrix (osteoid formation) takes
place.
Osteoblast-‘‘like’’ cell lines including MC3T3-E1 [11, 12],
MG-63 [13] and SaOS-2 [14, 15] have been extensively used
to understand osteoblast differentiation; however, these
immortalized lines are phenotypically different from the
primary cultures. Indeed, an in-depth proteomic study using
human mature osteoblasts and SaOS-2 (human osteogenic
sarcoma cell line) revealed significant difference between
them [16]. We, therefore, used primary rat neonatal calvarial
cells to study changes in expression profile of proteins
during osteoblast mineralization as a way to identify the
proteins linked to osteoblast mineralization [17].
Various polyphenolic compounds derived from natural
source have been shown to promote bone health possibly by
stimulating osteoblast functions [18–21]. We have recently
shown that kaempferol (K), a flavonol, promotes differentia-
tion and mineralization of rat calvarial osteoblasts and has
osteogenic effect in vivo [20]. These results encouraged us to
study the differentially regulated proteins during osteoblast
mineralization. Long-term culture of neonatal calvarial osteo-
blasts required for the induction of mineralization in vitroexhibits maturation sequences and developmental stages that
are comparable with those in neonatal bone [22]. We have
applied comparative proteomic approach to identify the
proteins involved in osteoblast mineralization stimulated by K.
The differentially expressed protein spots in osteoblasts, trea-
ted with or without K under mineralizing condition, were
identified by 2-DE. Protein spots were excised from gel, in-gel
digested, and the extracted proteins were further identified by
MALDI-TOF MS. The differential expression of proteins was
validated by Western blotting quantitative real-time PCR
(qPCR) and enzyme assay (in case of aldose reductase, AR).
Identified proteins could provide an insight into the mechan-
ism of osteoblast mineralization.
2 Materials and methods
2.1 Reagents and chemicals
Cell culture media and supplements were purchased from
Invitrogen (Carlsbad, CA). All fine chemicals, including K, were
purchased from Sigma Aldrich (St. Louis, MO). HPLC grade
ACN was obtained from Merck India (Mumbai, India).
2.2 Culture of calvarial osteoblasts
Rat calvarial osteoblasts were isolated following our previously
published protocol of sequential digestion [23]. Briefly, calvaria
from 10–12 1- to 2-day-old Sprague Dawley rats (both sexes)
were pooled. After surgical isolation from the skull and the
removal of sutures and adherent mesenchymal tissues, calvaria
were subjected to five sequential (10–15 min) digestions at 371C
in a solution containing 0.1% dispase and 0.1% collagenase P.
Cells released from the second to fifth digestions were collected,
centrifuged, resuspended and plated in T-25 cm2 flasks in
a-MEM containing 10% FCS and 1% penicillin/streptomycin
(complete growth medium).
2.3 Mineralization of calvarial osteoblasts
Calvarial osteoblasts cultured until 80% confluence were
trypsinized and plated in the differentiation medium (25 000
cells/well in 12-well plate), consisting of complete growth
medium with ascorbic acid (50 mg/mL) and b-glyceropho-
sphate (10 mM). The medium was changed every alternate
day up to 9 days. A 10 mM stock solution of K was made in
DMSO. Treatment with K at 5.0 mM was given in such a
manner that the final concentration of DMSO did not
exceed 0.01%. The treatment group contained osteoblast
differentiation medium with K (5.0 mM). At the end of the
experiment, cells were washed with PBS and fixed with 4%
paraformaldehyde in PBS for 15 min. The fixed cells were
stained with 40 mM (pH 4.5) Alizarin Red S for 30 min
followed by washing with distilled water [24].
2.4 Measurement of osteoblast alkaline
phosphatase activity
Calvarial osteoblasts (1200 cells/96well plate) were plated in
osteoblast differentiation medium and cultured in the
presence or absence of K for 2 days. The cells were then
freeze thawed by first keeping them at �701C for an hour
and then bringing them to room temperature to determine
the ALP activity. The assay is based on colorimetric esti-
mation of p-nitrophenol formed after breakdown of
p-nitrophenol-phosphate by alkaline phosphatase (ALP) [25].
Absorbance was read at 405 nm using a pre-programmed
semi-automatic photometer (model 5010, Boehringer
Mannheim, Germany).
2.5 2-D PAGE
2.5.1 Sample preparation
The cell pellet, obtained following harvesting and centrifu-
gation, was suspended in lysis buffer (urea: 7 M; thiourea:
2 M; Tris-HCl: 20 mM; CHAPS: 4% w/v; DTT: 10 mM;
EDTA: 1 mM; pPMSF: 1 mM and protease inhibitor cock-
tail). The mixed content was centrifuged and protein was
quantified [26].
2.5.2 IEF
The IPG strips (11 cm) were rehydrated with proteins
(0.5 mg), mixed in rehydration buffer (8 M urea, 2% w/v
Proteomics 2010, 10, 1730–1739 1731
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

CHAPS, 130 mM DTT, 0.002% bromophenol blue) and IPG
buffer (10mL) in a total volume of 150mL before subjecting
to IEF [27]. The rehydrated strips (pH 4.0–7.0) were focused
in a multiphor-II electrophoresis unit up to 14 000 V h at
201C (500 V for 30 min linear gradient, 1000 V for 10 min,
8000 V for 3 h, 8000 V for 6 h and 8000 V for 7 h).
2.5.3 SDS-PAGE and visualization of proteins
After IEF, the strips were kept in equilibration buffer
(50 mM Tris-HCl pH 8.8, 6 M urea, 30% v/v glycerol, 2%
SDS, 65 mM DTT, 2.5% iodoacetamide and 0.002%
bromophenol blue) for 20 min prior to SDS-PAGE. The pre-
focused and equilibrated IPG strips were placed over the
resolving gels, covered with sealing gel (0.5% agarose in
SDS electrophoresis buffer) and SDS-PAGE was carried out.
Markers were run in parallel to the strips during second
dimension to calculate the approximate molecular weight of
the separated proteins. SDS-PAGE was performed at
constant current (10 mA/gel) using running buffer (25 mM
Tris-HCl, 192 mM glycine and 0.1% SDS). The proteins
were visualized by silver staining, as reported previously
[27]. Equal amount of proteins was loaded in each experi-
ment and similar procedure for staining was used to mini-
mize the variations arising due to sample loading, staining
procedures and run-to-run variability.
2.5.4 2-DE gel image analysis
The 2-DE gel images were analyzed using Image
Master 2D-Platinum software (Gene Bio and Amersham
Biosciences). The gels were aligned properly and the inte-
grated intensity was normalized. The differentially displayed
protein spots were determined on the basis of spot
volume and spot density. Each spot was quantified using
total spot volume normalization method multiplied by the
total area of all the spots. The differentially displayed
proteins that showed significant alteration were selected for
MS.
2.5.5 Sample preparation for MS
The sample for MS was prepared using standard procedure
[26] with slight modifications. In brief, differentially
expressed silver-stained spots were excised, cut into pieces
and washed with 500mL water with agitation on a vortex
mixer. To remove silver stain, the excised spots were incu-
bated with 250 mL of 50 mM sodium thiosulfate and 15 mM
potassium ferricyanide for 5 min and washed twice with
500 mL water to remove reducing agents. In a tube, excised
spot and 500 mL of 100 mM NH4HCO3 and 45 mM DTT was
added and incubated at 601C for 30 min. On cooling, 500mL
of 100 mM iodoacetamide was added and further incubated
for 30 min in dark at room temperature. Excised spots were
sliced and equilibrated with 500mL of 50 mM ammonium
bicarbonate in 50% ACN, dehydrated with 500mL of 100%
ACN for 20 min and air dried [26]. Further the spots were
rehydrated in 30 mL of a solution containing 0.02 mg/mL
trypsin and 50 mM ammonium bicarbonate at 41C for
60 min. The supernatant was discarded and 30–50 mL of
50 mM ammonium bicarbonate was added in excised spots,
incubated at 371C for 16–18 h and supernatant was taken.
To the pellet, 25–50mL of 1% TFA in 60% ACN was added
and sonicated for 10 min. The supernatant was collected
following centrifugation at 12 000� g for 30 s. Both the
supernatants were mixed, freeze dried and concentrated by
centrifugal evaporation to dryness.
2.5.6 MALDI-TOF and LC-MS
Mass spectrometric analysis of samples was carried out at
The Centre for Genomic Applications, New Delhi. In brief,
trypsinized peptide samples were dissolved and mixed with
matrix, namely CHCA. Following drying, the peptides were
spotted on ground steel plate and subjected to Bruker
Ultraflex MALDI-TOF/TOF and 2D Nano LC-ESI-Trap
(Agilent) for mass spectrometric identification. The MALDI-
TOF/TOF instrument was equipped with a pulsed nitrogen
laser. Peptide fragment peaks produced by auto-digestion
of trypsin were used as an internal standard for every
peptide sample to ensure high mass accuracy and to control
possible variations arose due to protein extraction, trypsini-
zation, reconstitution and suppression of ionization by
highly abundant species and incomplete/non-homogeneous
crystallization of proteins and peptides during matrix
preparation.
2.5.7 Analysis of peptide sequences
The analyses of peptide sequences were performed as
reported elsewhere [26, 27]. In brief, trypsinized peptides
were used to generate peptide mass fingerprints. The
MASCOT search engine based on NCBI and Swiss-Prot
protein database were used to identify peptide mass
fingerprints. Probability based MOWSE scores were esti-
mated as ion scores �10� log10 (P), where P is the prob-
ability. The MASCOT search engine was used to identify the
observed peptide mass fingerprints.
2.6 Western blotting
Cells were grown to 60–70% confluence following, which
they were treated with K for 9 days. The cells were lysed in
Mammalian Cell Lysis buffer (Sigma). Lysate were centri-
fuged at 12 000� g for 10 min and supernatant collected.
Protein concentration was determined by the Bradford
1732 A. Kumar et al. Proteomics 2010, 10, 1730–1739
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
assay. Fifty micrograms of total protein was loaded on 10%
SDS-PAGE gel. After electrophoresis, proteins were trans-
ferred onto PVDF membranes (Immobilon-P, Millipore).
The membranes were probed with AR, cytokeratin, HSP-70
and caldesmon (Santa Cruz Biotechnology) and then incu-
bated with anti-rabbit antibodies conjugated with HRP (Cell
Signaling); and visualized using an enhanced chemilumi-
nescence kit (Amersham Pharmacia Biotech).
2.7 Determination of AR activity
At the end of treatment of K at days 3, 6 and 9 for 9 days,
osteoblasts were harvested by trypsinization and resus-
pended in ice-cold buffer (100 mM phosphate buffer, pH
7.4). Cells were sonicated and after centrifugation super-
natants were kept on ice and assayed in a reaction mixture
containing 0.15 mM NADPH, 10 mM glyceraldehyde,
10 mM HEPES buffer (pH 7.4) and enzyme (protein lysate)
in a total volume of 1 mL. The reaction was initiated by the
addition of substrate (glyceraldehydes) and was
followed at 340 nm for 0, 60, 120 and 180 s at room
temperature with a UV spectrophotometer. As a control,
absorbance at 340 nm was monitored in the presence of
treatment.
2.8 qPCR
PCR was performed using a light cycler (Roche@480). SYBR
Green PCR kit (Roche) and cDNA synthesized from 1.0 mg
total RNA in a 20mL reactions were subjected to PCR
according to the manufacture’s instructions.
2.9 Statistics
Data were presented as the mean7SD of results from three
or four cultures and the significance of differences was
analyzed by Student’s t-test. Groups were analyzed via t-tests
0d 3d 6d 9d0d 3d 6d
0.03
0.04
0.05
. [40
5]n
m
**
0
0.01
0.02
5C μM K
mea
n O
.D.
C 5μM K
A
B
C
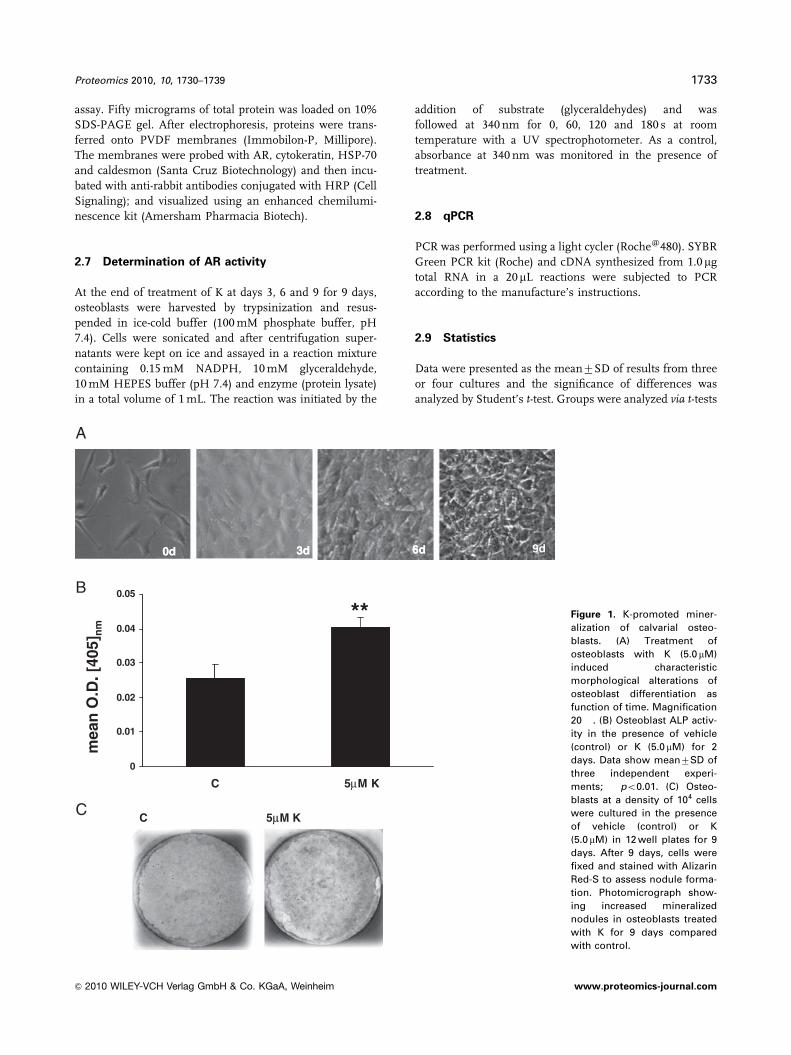
Figure 1. K-promoted miner-
alization of calvarial osteo-
blasts. (A) Treatment of
osteoblasts with K (5.0 mM)
induced characteristic
morphological alterations of
osteoblast differentiation as
function of time. Magnification
20� . (B) Osteoblast ALP activ-
ity in the presence of vehicle
(control) or K (5.0 mM) for 2
days. Data show mean7SD of
three independent experi-
ments; ��po0.01. (C) Osteo-
blasts at a density of 104 cells
were cultured in the presence
of vehicle (control) or K
(5.0 mM) in 12 well plates for 9
days. After 9 days, cells were
fixed and stained with Alizarin
Red-S to assess nodule forma-
tion. Photomicrograph show-
ing increased mineralized
nodules in osteoblasts treated
with K for 9 days compared
with control.
Proteomics 2010, 10, 1730–1739 1733
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

(two-sided). A p-value of o0.05 was considered to be
statistically significant.
3 Results
3.1 K increased ALP activity and mineralization in
primary osteoblasts
Cells extracted from neonatal calvariae are predominantly
preosteoblasts with proliferating ability [28]. When these
preosteoblasts were treated with K (5.0mM) in the presence of
osteoblast differentiation medium, cell morphology became
more cuboidal with the passage of time that was characteristic
of osteoblast differentiation (Fig. 1A). By day 9, cells displayed
more developed intercellular networks (Fig. 1A). Determina-
tion of ALP activity (measure of osteoblast differentiation) on
day 2 demonstrated significantly higher ALP levels (Fig. 1B). K
treatment continued for 9 days, resulted in increased forma-
tion of mineralized nodules compared with vehicle-treated
control osteoblasts (Fig. 1C).
3.2 Proteomic analysis of osteoblasts treated with K
Based on the data described above, osteoblasts treated with
or without K for 9 days were subjected to 2-DE analysis.
Three independent experiments were performed under
similar conditions and only those spots having altered
expressions in all the three experiments were selected
(Fig. 2). Cut-off criterion for changes in the expression levels
was 2.0-folds. By applying this criterion, a total of 18 spots
were identified as differentially expressed proteins by K
treatment. Nine of these proteins were overexpressed and as
many proteins were underexpressed (Table 1).
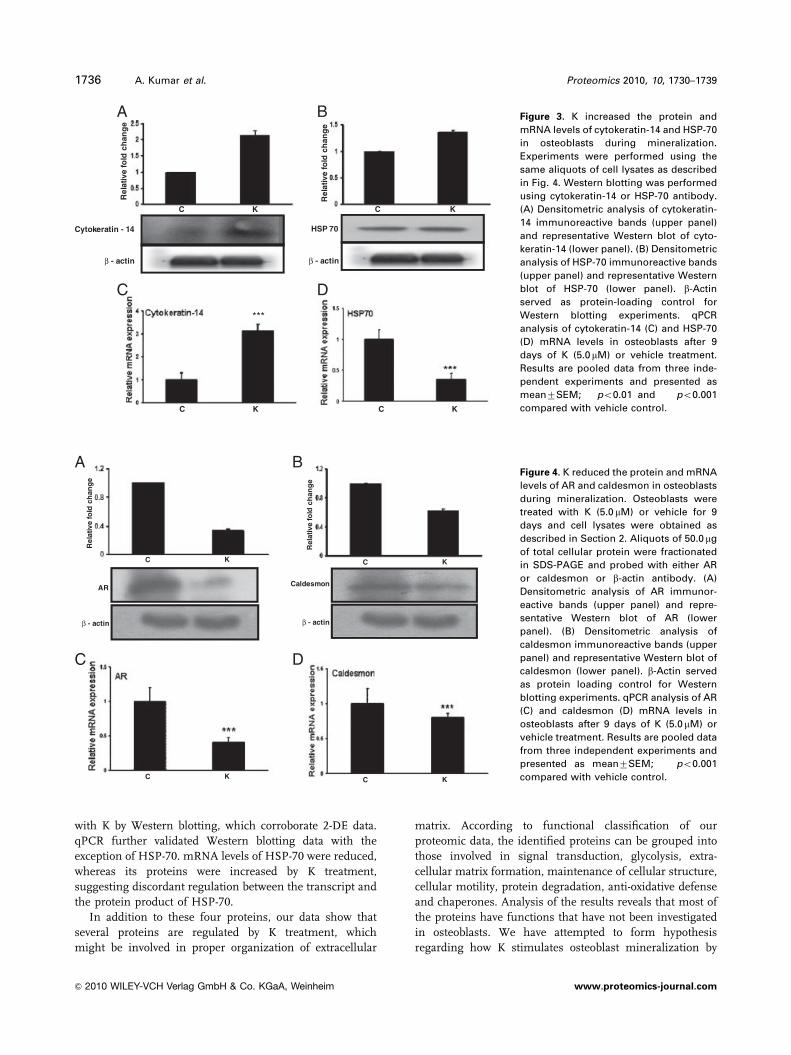
3.3 Confirmation of proteomic data
We randomly selected caldesmon, AR, HSP-70 and cytokera-
tin-14 from the proteomic data and performed western blotting
to confirm changes in their protein levels following K treat-
ment of osteoblasts. Our 2-DE data revealed marked increases
in cytokeratin-14 and HSP-70 spots in K-treated osteoblasts.
Western blotting data show that K treatment significantly
increased levels of both of these proteins in osteoblasts
(Figs. 3A and B). Cytokeratin-14 levels were increased more
robustly by K treatment compared with HSP-70 levels. In
addition, K treatment reduced levels of AR and caldesmon, as
assessed by Western blotting (Figs. 4A and B).
In addition, we confirmed 2-DE data by qPCR analysis of
mRNA levels of caldesmon, AR, cytokeratin-14 and HSP-70. K
treatment increased cytokeratin-14 mRNA levels in osteoblasts
(Fig. 3C), confirming Western blotting data. In contrast, HSP-
70 mRNA levels were decreased despite increased protein
levels by K treatment of osteoblasts (Fig. 3D), thereby
suggesting discordant regulation of mRNA and protein levels
of HSP-70. mRNA levels of AR and caldesmon were both
decreased by K treatment of osteoblasts (Figs. 4C and D),
further corroborating western blotting data.
200 kDa
C 5 μM K
6868 18 18
2
7
13
412
17
1
3
7
2
12
16
17
4
10
11
14
15
16
910
11
14
15
9
5
13
5
pH 4.0 pH 7.0
20kDa
13
Figure 2. Differential protein expression in
calvarial osteoblasts during mineralization
induced by K. Osteoblasts were plated at a
density of 104 cells in T-75 cm2 flasks, and
treated with 5.0 mM of K for 9 days. On ninth
day, cells were lysed in the protein lysis
buffer as described in Section 2. Only the
spots that showed altered expression (with a
minimum of 2.0-fold change in either direc-
tion) in three independent experiments
under similar conditions were defined as
differentially expressed proteins. The spots
from 2-D silver-stained gels were cut out and
digested, and the resulting peptides were
extracted, purified and identified by MALDI-
TOF. List of differentially expressed proteins
are given in Table 1.
1734 A. Kumar et al. Proteomics 2010, 10, 1730–1739
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

3.4 K modulates the AR activity in osteoblasts
Having observed that K inhibits AR production in osteo-
blasts, we next investigated the effect of K on AR activity. AR
activity was studied at various time points during the course
of osteoblast mineralization (Fig. 5). Data show that induc-
tion of mineralization by ascorbate and b-glycerophosphate
had no effect on AR activity (Fig. 5). K treatment resulted in
modest but significant increase in AR activity at day 3 but
was significantly inhibited at day 9 of the treatment (Fig. 5).
Inhibition of AR activity at day 9 of K treatment demon-
strated that K not only inhibits AR production but also its
activity in osteoblasts.
4 Discussion
Acquisition of bone matrix and mineralization is a complex
phenomenon and is a hallmark of osteoblast function.
Mechanism of osteoblast mineralization and matrix acqui-
sition remains poorly understood. Based on the studies
using a murine calvarial osteoblast cell line, MC3T3-E1,
collagen, osteonectin and osteocalcin represent proteins
associated with bone matrix accumulation and mineraliza-
tion [10]. Given a variety of MC3T3-E1 clones being used by
the researchers and that these clones are phenotypically
different from each other [29, 30], studies on
osteoblast mineralization in MC3T3-E1 do not truly repre-
sent the process of osteoblast mineralization in primary
osteoblasts.
We undertook proteomic study, using K as mineraliza-
tion stimulus in rat primary osteoblasts. Our data indicate
that the expression patterns of proteins, which are differ-
entially regulated by K treatment of osteoblasts during
mineralization, are unique to primary culture system
compared with cell lines [31]. Authenticity of proteomic data
was confirmed independently by western blotting and qPCR
of randomly selected proteins. We observed that protein
levels of HSP-70 and cytokeratin-14 were increased and that
of caldesmon and AR were reduced in osteoblasts treated
Table 1. Identification of differentially expressed proteins in osteoblasts whose expression is modulated by K treatment
S.no.
Proteins Totalscore
Matchedpeptides
Accession no. Description K treatment
Downregulated Upregulated
1. Ubiquitin ligase Nedd4-rat 80 3 S70642 Protein rafficking,signal transduction
4.18
2. RATER60PNID 276 21 BAA09695 2.23. Ribophorin I 38 1 Q6P7A7_RAT Uncharacterized protein 2.644. Chaperonin containing TCP1,
subunit 2 (b)142 7 Q5XIM9_RAT Chaperone 2.13
5. Cytokeratin-14 67 3 K1C14_RAT Cytoskeletal protein 3.316. Caldesmon 60 9 A55887 Intracellular cell-
signaling protein2.0
7. Stress 70 protein 48 3 GRP75_RAT Cell stress and actinorganization
2.0
8. AR 43 4 ALDR_HUMAN Glycolysis 2.139. Procollagen, type XI, a-2-rat 30 4 Q6MGB2_RAT Cell–matrix interaction 2.13
10. Prohibitin 91 6 A39682 Intracellular proteinblocks DNAsynthesis
2.01
11. Vimentin rat 284 17 VIME_RAT Cytoskeletal protein 2.7312. Reticulocalbin 3, EF hand calcium
binding domain137 10 Q5XIG3_RAT Intracellular cell-
signaling protein2.24
13. Translation initiation factor eIF-5Ahuman
65 9 FIHUA 4.64
14. K1 intermediate/smallconductance Ca21- activatedchannel
20 2 Q3S914_RAT Membrane protein 2.02
15. Mitochondrial import stimulationfactor S1-chain rat
159 12 JC2502 3.33
16. Fibronectin precursor rat 29 2 S14428 Extracellular matrixglycoprotein
2.01
17. PDI 54 8 A28807 Oxidoreductase 4.3818. Rho GDP dissociation inihibitor
GDI-a68 3 Q5X173 2.13
Proteomics 2010, 10, 1730–1739 1735
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
with K by Western blotting, which corroborate 2-DE data.
qPCR further validated Western blotting data with the
exception of HSP-70. mRNA levels of HSP-70 were reduced,
whereas its proteins were increased by K treatment,
suggesting discordant regulation between the transcript and
the protein product of HSP-70.
In addition to these four proteins, our data show that
several proteins are regulated by K treatment, which
might be involved in proper organization of extracellular
matrix. According to functional classification of our
proteomic data, the identified proteins can be grouped into
those involved in signal transduction, glycolysis, extra-
cellular matrix formation, maintenance of cellular structure,
cellular motility, protein degradation, anti-oxidative defense
and chaperones. Analysis of the results reveals that most of
the proteins have functions that have not been investigated
in osteoblasts. We have attempted to form hypothesis
regarding how K stimulates osteoblast mineralization by
Cytokeratin - 14
β - actin β - actin
***
Rel
ativ
e fo
ld c
han
ge
Rel
ativ
e fo
ld c
han
ge
HSP 70
KC KC
C KC KC K
A B
DC
Figure 3. K increased the protein and
mRNA levels of cytokeratin-14 and HSP-70
in osteoblasts during mineralization.
Experiments were performed using the
same aliquots of cell lysates as described
in Fig. 4. Western blotting was performed
using cytokeratin-14 or HSP-70 antibody.
(A) Densitometric analysis of cytokeratin-
14 immunoreactive bands (upper panel)
and representative Western blot of cyto-
keratin-14 (lower panel). (B) Densitometric
analysis of HSP-70 immunoreactive bands
(upper panel) and representative Western
blot of HSP-70 (lower panel). b-Actin
served as protein-loading control for
Western blotting experiments. qPCR
analysis of cytokeratin-14 (C) and HSP-70
(D) mRNA levels in osteoblasts after 9
days of K (5.0mM) or vehicle treatment.
Results are pooled data from three inde-
pendent experiments and presented as
mean7SEM; ��po0.01 and ���po0.001
compared with vehicle control.
AR
β - actin β - actin
Caldesmon
C K
C K
C K
C K
Rel
ativ
e fo
ld c
han
ge
Rel
ativ
e fo
ld c
han
ge
A B
C D
Figure 4. K reduced the protein and mRNA
levels of AR and caldesmon in osteoblasts
during mineralization. Osteoblasts were
treated with K (5.0 mM) or vehicle for 9
days and cell lysates were obtained as
described in Section 2. Aliquots of 50.0 mg
of total cellular protein were fractionated
in SDS-PAGE and probed with either AR
or caldesmon or b-actin antibody. (A)
Densitometric analysis of AR immunor-
eactive bands (upper panel) and repre-
sentative Western blot of AR (lower
panel). (B) Densitometric analysis of
caldesmon immunoreactive bands (upper
panel) and representative Western blot of
caldesmon (lower panel). b-Actin served
as protein loading control for Western
blotting experiments. qPCR analysis of AR
(C) and caldesmon (D) mRNA levels in
osteoblasts after 9 days of K (5.0mM) or
vehicle treatment. Results are pooled data
from three independent experiments and
presented as mean7SEM; ���po0.001
compared with vehicle control.
1736 A. Kumar et al. Proteomics 2010, 10, 1730–1739
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

reviewing the functions of the K-regulated proteins in
osteoblast mineralization with their known functions in
other cell types.
For example, we observed that K treatment during
osteoblast mineralization upregulated prohibitin, whose
function in osteoblast has never been studied. Prohibitin has
been shown to block DNA synthesis in normal fibroblasts
and HeLa cells [32]. It is surmised that by increasing the
levels of prohibitin in osteoblasts, K signals termination of
cellular proliferation, which could be crucial for the
commencement of osteoblast differentiation leading to its
mineralization. On the contrary, caldesmon, which is
downregulated by K treatment in osteoblast, is known to
inhibit mitosis in non-muscle cells [33]. As end of mitotic
cell growth signals, induction of osteoblast differentiation
caldesmon may have important role in osteoblast differ-
entiation.
Mineralization is characterized by deposition of both
collagenous and non-collagenous matrix proteins. We
observed that K increases fibronectin protein levels in
osteoblasts. Fibronectin is maximally produced in rat
calvarial osteoblasts during the matrix maturation stage
[34, 35]. It would be interesting to understand the effect
of fibronectin in osteoblast mineralization in general and
K-stimulated mineralization in particular. Cellular archi-
tecture during mineralization is required to be maintained
for optimum osteoblast function, particularly during the
transformation of osteoblasts to osteocytes. Our data show
that cytokeratin-14 levels are increased by K treatment. Like
in other cells [36], increased expression of cytokeratin-14 in
osteoblasts could result in alterations in cellular morphol-
ogy, adhesion and motility. Regarding cytoskeletal protein
levels being altered by K treatment, our 2-DE data also show
that vimentin levels are reduced in osteoblasts. Intermittent
PTH treatment to rats, which is known to exert osteogenic
effect, has been shown to decrease vimentin levels in bone
marrow cells [37, 38]. As K too has osteogenic effect in vivo,
decrease in vimentin levels by K treatment of osteoblasts
may favor osteoblast mineralization.
Mineralization involves deposition of nascent calcium in
the form of hydroxyapatite crystals. Intracellular calcium
homeostasis is expected to play a vital role in the process of
mineralization by osteoblasts, although nothing is known
about intracellular calcium homeostasis in mineralized
osteoblasts. Our data show that reticulocalbin 3-EF, a
calcium-binding protein [39] is upregulated in K-treated
osteoblasts. Reticulocalbin has no significant sequence
homology with any other proteins except the EF hand
motifs; however, the overall structure of reticulocalbin is
similar to that of calbindin D28K and calretinin [40, 41].
Although there are reports on the structure and localization
of reticulocalbin [42, 43], exact function of this protein
remains unknown. Separate studies are required to learn
whether or not reticulocalbin regulates K-stimulated osteo-
blast mineralization. Endoplasmic reticulum plays an
essential role in intracellular calcium homeostasis by being
an intracellular calcium store. The lumen of endoplasmic
reticulum contains a number of soluble proteins including
protein disulfide isomerase (PDI) [44]. Our 2-DE data show
that PDI levels are significantly increased upon K treatment
of osteoblasts. PDI is involved in the initial stages of
maturation of newly synthesized secretary proteins through
folding and formation of disulfide bonds [45, 46]. In addition
to HSP-70, K treatment increased TCP1 subunit beta, a
molecular chaperone [47], in osteoblasts. HSP-70 has high
affinity for non-native proteins and interacts with hydro-
phobic residues exposed to incompletely folded nascent
polypeptides. Therefore, it appears that increased levels of
reticulocalbin, PDI and TCP1 by K treatment of osteoblasts
could result in nascent polypeptide modification of proteins
that may be important for osteoblast mineralization, where
maturation of matrix protein is required.
There is considerable energy demand during the process
of osteoblast differentiation and mineralization that is met
not only by increasing activities of enzymes involved in
energy metabolic process such as glycolysis but also by their
increased production [48, 49]. AR catalyzes glucose to
sorbitol conversion, which is known as sorbitol/polyol
pathway. Sorbitol is the intermediate step of glucose to
fructose conversion that is catalyzed by sorbitol dehy-
drogenase (SDH). Therefore, by catalyzing glucose to
sorbitol conversion, AR bypasses the control points of
hexokinase and phosphofructokinase in glucose metabo-
lism. It is now well recognized that diabetes is a disease of
impaired glycolysis due to increased glucose flux via AR. As
a result, AR inhibitor is considered as therapeutic candidate
for diabetes and other metabolic disorders [50, 51]. Role of
polyol pathway in osteoblast mineralization is hardly
0.75C
T
0.6 **
**
0.3
0.45
OD
(34
0nm
)
9630
Days post treatment
Figure 5. K inhibits AR activity in osteoblasts. Effect of K (5.0 mM)
on AR activity in osteoblasts was studied as described in Section
2. Cells were cultured in 6-well plates in the presence or absence
of K at days 0 (control), 3, 6 and 9. Lysates containing equal
amounts of protein were sonicated and AR activity was assayed
in the reaction mixture (containing NADPH, glyceraldehydes and
HEPES buffer). The reaction was initiated by adding glycer-
aldehydes and was followed at 340 nm at various time points till
180 s. Results are pooled from six independent experiments with
similar results. Values are expressed as mean7SD. ��po0.01
versus their respective controls in the corresponding groups.
Proteomics 2010, 10, 1730–1739 1737
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

understood. Osteoblast differentiation and formation of
mineralized nodules have been correlated with increase in
osteoblast glycolysis [48]. In addition, intracellular sorbitol
accumulation is thought to be involved in the development
of osteoblast dysfunction [51, 52]. Furthermore, AR inhibitor
has been shown to attenuate development of galactose-
induced diabetic osteopenia [50]. Our data show that treating
osteoblasts with K resulted in reduced mRNA and protein
levels of AR. SDH mRNA levels were also reduced by K
treatment (data not shown). As K promotes osteoblast
mineralization, inhibition of AR and SDH expressions by K
suggest that activation of polyol pathway negatively regu-
lates osteoblast function.
In conclusion, our study has led to the identification of
proteins not hitherto known for their roles in promoting
osteoblast mineralization. Future studies are required to
demonstrate their specific roles either in elevated or reduced
levels as observed with K treatment in the formation of
mineralized matrix in osteoblasts derived from calvaria
as well as bone marrow. It would also be interesting to
determine whether these proteins are specifically linked
to K-stimulated osteoblast mineralization or other differ-
entiation promoting agents such as bone morphogenetic
proteins.
The authors thank Dr. Sanghamitra Bandhopadhyay forcritical reading of the manuscript. Grant support from Ministryof Health and Family Welfare is acknowledged.
The authors have declared no conflict of interest.
5 References
[1] Harada, S., Rodan, G. A., Control of osteoblast function and
regulation of bone mass. Nature 2003, 423, 349–355.
[2] Beck, G. R., Jr., Sullivan, E. C., Moran, E., Zerler, B.,
Relationship between alkaline phosphatase levels,
osteopontin expression, and mineralization in differentiat-
ing MC3T3-E1 osteoblasts. J. Cell. Biochem. 1998, 68,
269–280.
[3] Aubin, J. E., Regulation of osteoblast formation and func-
tion. Rev. Endocr. Metab. Disord. 2001, 2, 81–94.
[4] BeckJr., G. R., Zerler, B., Moran, E., Gene array analysis of
osteoblast differentiation. Cell. Growth Differ. 2001, 12,
61–83.
[5] Malaval, L., Modrowski, D., Gupta, A. K., Aubin, J. E.,
Cellular expression of bone-related proteins during in vitro
osteogenesis in rat bone marrow stromal cell cultures.
J. Cell. Physiol. 1994, 158, 555–572.
[6] Owen, T. A., Aronow, M., Calhoun, V., Baronet, L. M. et al.,
Progressive development of the rat osteoblast phenotype
in vitro: reciprocal relationships in expression of genes
associated with osteoblast proliferation and differentiation
during formation of the bone extracellular matrix. J. Cell.
Physiol. 1990, 143, 420–430.
[7] Yoon, K., Buenaga, R., Rodan, G. A., Tissue specificity and
developmental expression of rat osteopontin. Biochem.
Biophys. Res. Commun. 1987, 148, 1129–1136.
[8] Weinreb, M., Shinar, D., Rodan, G. A., Different pattern of
alkaline phosphatase, osteopontin, and osteocalcin
expression in developing rat bone visualized by in situ
hybridization. J. Bone Miner. Res. 1990, 5, 831–842.
[9] Sandberg, M., Autio-Harmainen, H., Vuorio, E., Localization
of the expression of types I, III, and IV collagen, TGF-beta 1
and c-fos genes in developing human calvarial bones. Dev.
Biol. 1988, 130, 324–334.
[10] Choi, J. Y., Lee, B. H., Song, K. B., Park, R. W. et al.,
Expression patterns of bone-related proteins during osteo-
blastic differentiation in MC3T3-E1 cells. J. Cell. Biochem.
1996, 61, 609–618.
[11] Kumegawa, M., Hiramatsu, M., Hatakeyama, K., Yajima, T.
et al., Effects of epidermal growth factor on osteoblastic
cells in vitro. Calcif. Tissue Int. 1983, 35, 542–548.
[12] Sudo, H., Kodama, H. A., Amagai, Y., Yamamoto, S., Kasai,
S., In vitro differentiation and calcification in a new clonal
osteogenic cell line derived from newborn mouse calvaria.
J. Cell. Biol. 1983, 96, 191–198.
[13] Prouillet, C., Maziere, J. C., Maziere, C., Wattel, A. et al.,
Stimulatory effect of naturally occurring flavonols
quercetin and kaempferol on alkaline phosphatase
activity in MG-63 human osteoblasts through ERK and
estrogen receptor pathway. Biochem. Pharmacol. 2004, 67,
1307–1313.
[14] Myung, J. K., Afjehi-Sadat, L., Felizardo-Cabatic, M., Slavc,
I., Lubec, G., Expressional patterns of chaperones in ten
human tumor cell lines. Proteome Sci. 2004, 2, 8.
[15] Im, G. I., Qureshi, S. A., Kenney, J., Rubash, H. E., Shanb-
hag, A. S., Osteoblast proliferation and maturation by
bisphosphonates. Biomaterials 2004, 25, 4105–4115.
[16] Spreafico, A., Frediani, B., Capperucci, C., Chellini, F. et al.,
A proteomic study on human osteoblastic cells proliferation
and differentiation. Proteomics 2006, 6, 3520–3532.
[17] Bellows, C. G., Aubin, J. E., Heersche, J. N., Antosz, M. E.,
Mineralized bone nodules formed in vitro from enzymati-
cally released rat calvaria cell populations. Calcif. Tissue Int.
1986, 38, 143–154.
[18] Hsu, Y. L., Chang, J. K., Tsai, C. H., Chien, T. T., Kuo, P. L.,
Myricetin induces human osteoblast differentiation through
bone morphogenetic protein-2/p38 mitogen-activated
protein kinase pathway. Biochem. Pharmacol. 2007, 73,
504–514.
[19] Trivedi, R., Kumar, A., Gupta, V., Kumar, S. et al., Effects of
Egb 761 on bone mineral density, bone microstructure, and
osteoblast function Possible roles of quercetin and
kaempferol. Mol. Cell. Endocrinol. 2009, 302, 86–91.
[20] Trivedi, R., Kumar, S., Kumar, A., Siddiqui, J. A. et al.,
Kaempferol has osteogenic effect in ovariectomized adult
Sprague-Dawley rats. Mol. Cell. Endocrinol. 2008, 289,
85–93.
[21] Trivedi, R., Mithal, A., Chattopadhyay, N., Anabolics in
osteoporosis: the emerging therapeutic tool. Curr. Mol.
Med. 2010, 10, 14–28.
1738 A. Kumar et al. Proteomics 2010, 10, 1730–1739
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

[22] Stein, G. S., Lian, J. B., Owen, T. A., Relationship of cell
growth to the regulation of tissue-specific gene expression
during osteoblast differentiation. FASEB J. 1990, 4, 3111–3123.
[23] Wong, G. L., Cohn, D. V., Target cells in bone for para-
thormone and calcitonin are different: enrichment for each
cell type by sequential digestion of mouse calvaria and
selective adhesion to polymeric surfaces. Proc. Natl. Acad.
Sci. USA 1975, 72, 3167–3171.
[24] Prabhakar, U., James, I. E., Dodds, R. A., Lee-Rykaczewski,
E. et al. A novel human bone marrow stroma-derived cell
line TF274 is highly osteogenic in vitro and in vivo. Calcif.
Tissue Int. 1998, 63, 214–220.
[25] Akcakaya, H., Aroymak, A., Gokce, S., A quantitative
colorimetric method of measuring alkaline phosphatase
activity in eukaryotic cell membranes. Cell Biol. Int. 2007,
31, 186–190.
[26] Patel, S., Sinha, A., Singh, M. P., Identification of differen-
tially expressed proteins in striatum of maneb-and para-
quat-induced Parkinson’s disease phenotype in mouse.
Neurotoxicol. Teratol. 2007, 29, 578–585.
[27] Sinha, A., Srivastava, N., Singh, S., Singh, A. K. et al.,
Identification of differentially displayed proteins in cere-
brospinal fluid of Parkinson’s disease patients: a proteomic
approach. Clin. Chim. Acta 2009, 400, 14–20.
[28] Chattopadhyay, N., Yano, S., Tfelt-Hansen, J., Rooney, P. et al.,
Mitogenic action of calcium-sensing receptor on rat calvarial
osteoblasts. Endocrinology 2004, 145, 3451–3462.
[29] Wenstrup, R. J., Fowlkes, J. L., Witte, D. P., Florer, J. B.,
Discordant expression of osteoblast markers in MC3T3-E1
cells that synthesize a high turnover matrix. J. Biol. Chem.
1996, 271, 10271–10276.
[30] Bellows, C. G., Sodek, J., Yao, K. L., Aubin, J. E., Phenotypic
differences in subclones and long-term cultures of clonally
derived rat bone cell lines. J. Cell. Biochem. 1986, 31,
153–169.
[31] Miyake, M., Arai, N., Ushio, S., Iwaki, K. et al., Promoting
effect of kaempferol on the differentiation and mineraliza-
tion of murine pre-osteoblastic cell line MC3T3-E1. J. Biosci.
Biotechnol. Biochem. 2003, 67, 1199–1205.
[32] Nuell, M. J., Stewart, D. A., Walker, L., Friedman, V. et al.,
Prohibitin, an evolutionarily conserved intracellular protein
that blocks DNA synthesis in normal fibroblasts and HeLa
cells. Mol. Cell. Biol. 1991, 11, 1372–1381.
[33] Yamashiro, S., Yamakita, Y., Yoshida, K., Takiguchi, K.,
Matsumura, F., Characterization of the COOH terminus of
non-muscle caldesmon mutants lacking mitosis-
specific phosphorylation sites. J. Biol. Chem. 1995, 270,
4023–4030.
[34] Moursi, A. M., Damsky, C. H., Lull, J., Zimmerman, D. et al.,
Fibronectin regulates calvarial osteoblast differentiation.
J. Cell. Sci. 1996, 109, 1369–1380.
[35] Globus, R. K., Doty, S. B., Lull, J. C., Holmuhamedov, E.
et al., Fibronectin is a survival factor for differentiated
osteoblasts. J. Cell. Sci. 1998, 111, 1385–1393.
[36] Lee, C. H., Coulombe, P. A., Self-organization of keratin
intermediate filaments into cross-linked networks. J. Cell.
Biol. 2009, 186, 409–421.
[37] Kim, S. H., Jun, S., Jang, H. S., Lim, S. K., Identification of
parathyroid hormone-regulated proteins in mouse bone
marrow cells by proteomics. Biochem. Biophys. Res.
Commun. 2005, 330, 423–429.
[38] Steinert, P. M., Roop, D. R., Molecular and cellular biology of
intermediate filaments. Annu. Rev. Biochem. 1988, 57, 593–625.
[39] Ozawa, M., Muramatsu, T., Reticulocalbin, a novel endo-
plasmic reticulum resident Ca(21)-binding protein with
multiple EF-hand motifs and a carboxyl-terminal HDEL
sequence. J. Biol. Chem. 1993, 268, 699–705.
[40] Rogers, J. H., Immunoreactivity for calretinin and other
calcium-binding proteins in cerebellum. Neuroscience
1989, 31, 711–721.
[41] Tiwari, S., Gupta, S. K., Kumar, K., Trivedi, R., Godbole,
M. M., Simultaneous exposure of excess fluoride and
calcium deficiency alters VDR, CaR, and calbindin D 9 k
mRNA levels in rat duodenal mucosa. Calcif. Tissue Int.
2004, 75, 313–320.
[42] Booth, C., Koch, G. L., Perturbation of cellular calcium indu-
ces secretion of luminal ER proteins. Cell 1989, 59, 729–737.
[43] Kelly, R. B., Cell biology Tracking an elusive receptor.
Nature 1990, 345, 480–481.
[44] Freedman, R. B., Protein disulfide isomerase: multiple roles
in the modification of nascent secretory proteins. Cell 1989,
57, 1069–1072.
[45] Ferrari, D. M., Soling, H. D., The protein disulphide-
isomerase family: unravelling a string of folds. Biochem.
J. 1999, 339, 1–10.
[46] Kemmink, J., Darby, N. J., Dijkstra, K., Nilges, M.,
Creighton, T. E., The folding catalyst protein disulfide
isomerase is constructed of active and inactive thioredoxin
modules. Curr. Biol. 1997, 7, 239–245.
[47] Brackley, K. I., Grantham, J., Activities of the chaperonin
containing TCP-1 (CCT): implications for cell cycle
progression and cytoskeletal organization. Cell Stress
Chaperones 2009, 14, 23–31.
[48] Komarova, S. V., Ataullakhanov, F. I., Globus, R. K., Bio-
energetics and mitochondrial transmembrane potential
during differentiation of cultured osteoblasts. Am. J.
Physiol. Cell. Physiol. 2000, 279, C1220–C1229.
[49] Schirrmacher, K., Lauterbach, S., Bingmann, D., Oxygen
consumption of calvarial bone cells in vitro. J. Orthop. Res.
1997, 15, 558–562.
[50] Inaba, M., Terada, M., Nishizawa, Y., Shioi, A. et al.,
Protective effect of an aldose reductase inhibitor against
bone loss in galactose-fed rats: possible involvement of the
polyol pathway in bone metabolism. Metabolism 1999, 48,
904–909.
[51] Inaba, M., Effect of aldose reductase on the abnormality of
calcium metabolism in diabetic patients. Clin. Calcium 2006,
16, 1360–1365.
[52] Takizawa, M., Suzuki, K., Matsubayashi, T., Kikuyama, M. et al.,
Increased bone resorption may play a crucial role in the
occurrence of osteopenia in patients with type 2 diabetes:
possible involvement of accelerated polyol pathway in its
pathogenesis. Diabetes Res. Clin. Pract. 2008, 82 119–126.
Proteomics 2010, 10, 1730–1739 1739
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com




















