
Mechanisms underlying the inhibitory effects of uroguanylin on NHE3 transport activity in renal...
10
CALL FOR PAPERS Renal Solute Cotransporters and Exchangers Mechanisms underlying the inhibitory effects of uroguanylin on NHE3 transport activity in renal proximal tubule Lucília M. A. Lessa, 1,3 Luciene R. Carraro-Lacroix, 1 Renato O. Crajoinas, 2 Camila N. Bezerra, 1 Rafael Dariolli, 2 Adriana C. C. Girardi, 2 Manassés C. Fonteles, 3,4 and Gerhard Malnic 1 1 Department of Physiology and Biophysics, Biomedical Sciences Institute, University of São Paulo, São Paulo/SP, Brazil; 2 Heart Institute (InCor), University of São Paulo Medical School, São Paulo/SP, Brazil; 3 Biomedical Sciences Superior Institute, State University of Ceará, Fortaleza/CE, Brazil; and 4 Mackenzie University, São Paulo, Brazil Submitted 13 July 2011; accepted in final form 3 September 2012 Lessa LM, Carraro-Lacroix LR, Crajoinas RO, Bezerra CN, Dariolli R, Girardi AC, Fonteles MC, Malnic G. Mechanisms underlying the inhibitory effects of uroguanylin on NHE3 transport activity in renal proxi- mal tubule. Am J Physiol Renal Physiol 303: F1399 –F1408, 2012. First published September 5, 2012; doi:10.1152/ajprenal.00385.2011.—We pre- viously demonstrated that uroguanylin (UGN) significantly inhibits Na /H exchanger (NHE)3-mediated bicarbonate reabsorption. In the present study, we aimed to elucidate the molecular mechanisms underlying the action of UGN on NHE3 in rat renal proximal tubules and in a proximal tubule cell line (LLC-PK1 ). The in vivo studies were performed by the stationary microperfusion technique, in which we measured H secretion in rat renal proximal segments, through a H -sensitive microelectrode. UGN (1 M) significantly inhibited the net of proximal bicarbonate reabsorption. The inhibitory effect of UGN was completely abolished by either the protein kinase G (PKG) inhibitor KT5823 or by the protein kinase A (PKA) inhibitor H-89. The effects of UGN in vitro were found to be similar to those obtained by microperfusion. Indeed, we observed that incubation of LLC-PK1 cells with UGN induced an increase in the intracellular levels of cAMP and cGMP, as well as activation of both PKA and PKG. Furthermore, we found that UGN can increase the levels of NHE3 phosphorylation at the PKA consensus sites 552 and 605 in LLC-PK1 cells. Finally, treatment of LLC-PK 1 cells with UGN reduced the amount of NHE3 at the cell surface. Overall, our data suggest that the inhibitory effect of UGN on NHE3 transport activity in proximal tubule is mediated by activation of both cGMP/PKG and cAMP/PKA signaling pathways which in turn leads to NHE3 phosphorylation and reduced NHE3 surface expression. Moreover, this study sheds light on mechanisms by which guanylin peptides are intricately involved in the maintenance of salt and water homeostasis. NHE3; uroguanylin; renal microperfusion; bicarbonate reabsorption; cGMP; cAMP IN THE PAST DECADES, IT HAS been postulated that Sta-like guanylin peptides participate in a control system regulating salt balance in response to oral salt intake, linking intestine and kidney in the process of salt and water homeostasis (8, 19, 22). These peptides are known as endogenous agonists for the Escherichia coli heat- stable toxin receptor, guanylate cyclase C (GC-C), uroguanylin (UGN) being its most potent member (12, 18, 20, 27). UGN is a 16 amino acid peptide (27), expressed throughout the intestinal tract, secreted by enterochromaffin cells as prou- roguanylin that undergoes postsecretory proteolytic cleavage, generating the active form (15, 28, 41). Recently, Qian and colleagues (44) demonstrated that the conversion of prourogua- nylin to UGN occurs within the lumen of renal tubules. Renal effects of UGN include natriuresis, kaliuresis, diure- sis, and increased excretion of cGMP (17, 26). In a previous work using stationary in vivo microperfusion, our group dem- onstrated that UGN perfusion stimulates potassium secretion through Maxi-K channels in connecting and cortical collecting ducts (2). Furthermore, this study demonstrated that UGN perfusion also leads to an inhibition of bicarbonate reabsorp- tion in proximal tubules by reducing the activity of the Na /H exchanger, NHE3 isoform (2). In renal proximal tubule cells, NHE3 is the principal mech- anism of hydrogen secretion, which leads to filtered bicarbon- ate reabsorption (1, 2, 4, 47, 52). NHE3 is also responsible for the isotonic reabsorption of approximately two-thirds of the filtered NaCl and water (3, 43). Accordingly, Schultheis and co-workers (47) demonstrated that NHE3 knockout mice are hypovolemic, hypotensive, and present metabolic acidosis and reduced reabsorption of Na , HCO 3 , and volume. In addition, the mortality of these NHE3 knockout mice is increased when they are submitted to a low-salt diet (33). Considering NHE3 physiological roles, relatively small alterations of its activity may have significant consequences. Indeed, NHE3 is among the most extensively regulated transport proteins of cell mem- branes, being modulated by several physiological and patho- logical conditions (13, 24). The mechanisms underlying UGN renal actions are not completely elucidated, especially regarding the natriuretic ef- fect. High-salt diets increase the concentration of UGN and cGMP in the urine (22, 42), but in GC-C knockout mice, guanylin peptides still cause natriuresis (9). Our previous studies suggested an inhibitory effect of guanylin peptides on NHE3 activity in proximal tubules (2, 35). Thus, the present study was designed to investigate the signaling mechanisms involved in the natriuretic effect of UGN, focusing on the inhibition of NHE3 activity by the peptide. MATERIALS AND METHODS Reagents and antibodies. All chemicals were obtained from Sigma (St. Louis, MO) unless otherwise noted. UGN was purchased from Address for reprint requests and other correspondence: L. M. A. Lessa, Instituto Superior de Ciencias Biomedicas, Universidade Estadual do Ceará (UEC), Fortaleza, Ceara (e-mail: [email protected]). Am J Physiol Renal Physiol 303: F1399–F1408, 2012. First published September 5, 2012; doi:10.1152/ajprenal.00385.2011. 1931-857X/12 Copyright © 2012 the American Physiological Society http://www.ajprenal.org F1399
Transcript of Mechanisms underlying the inhibitory effects of uroguanylin on NHE3 transport activity in renal...

CALL FOR PAPERS Renal Solute Cotransporters and Exchangers
Mechanisms underlying the inhibitory effects of uroguanylin on NHE3transport activity in renal proximal tubule
Lucília M. A. Lessa,1,3 Luciene R. Carraro-Lacroix,1 Renato O. Crajoinas,2 Camila N. Bezerra,1
Rafael Dariolli,2 Adriana C. C. Girardi,2 Manassés C. Fonteles,3,4 and Gerhard Malnic1
1Department of Physiology and Biophysics, Biomedical Sciences Institute, University of São Paulo, São Paulo/SP, Brazil;2Heart Institute (InCor), University of São Paulo Medical School, São Paulo/SP, Brazil; 3Biomedical Sciences SuperiorInstitute, State University of Ceará, Fortaleza/CE, Brazil; and 4Mackenzie University, São Paulo, Brazil
Submitted 13 July 2011; accepted in final form 3 September 2012
Lessa LM, Carraro-Lacroix LR, Crajoinas RO, Bezerra CN, DariolliR, Girardi AC, Fonteles MC, Malnic G. Mechanisms underlying theinhibitory effects of uroguanylin on NHE3 transport activity in renal proxi-mal tubule. Am J Physiol Renal Physiol 303: F1399–F1408, 2012. Firstpublished September 5, 2012; doi:10.1152/ajprenal.00385.2011.—We pre-viously demonstrated that uroguanylin (UGN) significantly inhibitsNa�/H� exchanger (NHE)3-mediated bicarbonate reabsorption. Inthe present study, we aimed to elucidate the molecular mechanismsunderlying the action of UGN on NHE3 in rat renal proximal tubulesand in a proximal tubule cell line (LLC-PK1). The in vivo studies wereperformed by the stationary microperfusion technique, in which wemeasured H� secretion in rat renal proximal segments, through aH�-sensitive microelectrode. UGN (1 �M) significantly inhibited thenet of proximal bicarbonate reabsorption. The inhibitory effect ofUGN was completely abolished by either the protein kinase G (PKG)inhibitor KT5823 or by the protein kinase A (PKA) inhibitor H-89.The effects of UGN in vitro were found to be similar to those obtainedby microperfusion. Indeed, we observed that incubation of LLC-PK1
cells with UGN induced an increase in the intracellular levels ofcAMP and cGMP, as well as activation of both PKA and PKG.Furthermore, we found that UGN can increase the levels of NHE3phosphorylation at the PKA consensus sites 552 and 605 in LLC-PK1
cells. Finally, treatment of LLC-PK1 cells with UGN reduced theamount of NHE3 at the cell surface. Overall, our data suggest that theinhibitory effect of UGN on NHE3 transport activity in proximaltubule is mediated by activation of both cGMP/PKG and cAMP/PKAsignaling pathways which in turn leads to NHE3 phosphorylation andreduced NHE3 surface expression. Moreover, this study sheds light onmechanisms by which guanylin peptides are intricately involved in themaintenance of salt and water homeostasis.
NHE3; uroguanylin; renal microperfusion; bicarbonate reabsorption;cGMP; cAMP
IN THE PAST DECADES, IT HAS been postulated that Sta-like guanylinpeptides participate in a control system regulating salt balance inresponse to oral salt intake, linking intestine and kidney in theprocess of salt and water homeostasis (8, 19, 22). These peptidesare known as endogenous agonists for the Escherichia coli heat-stable toxin receptor, guanylate cyclase C (GC-C), uroguanylin(UGN) being its most potent member (12, 18, 20, 27).
UGN is a 16 amino acid peptide (27), expressed throughoutthe intestinal tract, secreted by enterochromaffin cells as prou-roguanylin that undergoes postsecretory proteolytic cleavage,generating the active form (15, 28, 41). Recently, Qian andcolleagues (44) demonstrated that the conversion of prourogua-nylin to UGN occurs within the lumen of renal tubules.
Renal effects of UGN include natriuresis, kaliuresis, diure-sis, and increased excretion of cGMP (17, 26). In a previouswork using stationary in vivo microperfusion, our group dem-onstrated that UGN perfusion stimulates potassium secretionthrough Maxi-K channels in connecting and cortical collectingducts (2). Furthermore, this study demonstrated that UGNperfusion also leads to an inhibition of bicarbonate reabsorp-tion in proximal tubules by reducing the activity of theNa�/H� exchanger, NHE3 isoform (2).
In renal proximal tubule cells, NHE3 is the principal mech-anism of hydrogen secretion, which leads to filtered bicarbon-ate reabsorption (1, 2, 4, 47, 52). NHE3 is also responsible forthe isotonic reabsorption of approximately two-thirds of thefiltered NaCl and water (3, 43). Accordingly, Schultheis andco-workers (47) demonstrated that NHE3 knockout mice arehypovolemic, hypotensive, and present metabolic acidosis andreduced reabsorption of Na�, HCO3
�, and volume. In addition,the mortality of these NHE3 knockout mice is increased whenthey are submitted to a low-salt diet (33). Considering NHE3physiological roles, relatively small alterations of its activitymay have significant consequences. Indeed, NHE3 is amongthe most extensively regulated transport proteins of cell mem-branes, being modulated by several physiological and patho-logical conditions (13, 24).
The mechanisms underlying UGN renal actions are notcompletely elucidated, especially regarding the natriuretic ef-fect. High-salt diets increase the concentration of UGN andcGMP in the urine (22, 42), but in GC-C knockout mice,guanylin peptides still cause natriuresis (9). Our previousstudies suggested an inhibitory effect of guanylin peptides onNHE3 activity in proximal tubules (2, 35). Thus, the presentstudy was designed to investigate the signaling mechanismsinvolved in the natriuretic effect of UGN, focusing on theinhibition of NHE3 activity by the peptide.
MATERIALS AND METHODS
Reagents and antibodies. All chemicals were obtained from Sigma(St. Louis, MO) unless otherwise noted. UGN was purchased from
Address for reprint requests and other correspondence: L. M. A. Lessa,Instituto Superior de Ciencias Biomedicas, Universidade Estadual do Ceará(UEC), Fortaleza, Ceara (e-mail: [email protected]).
Am J Physiol Renal Physiol 303: F1399–F1408, 2012.First published September 5, 2012; doi:10.1152/ajprenal.00385.2011.
1931-857X/12 Copyright © 2012 the American Physiological Societyhttp://www.ajprenal.org F1399

Bachem (Philadelphia, PA). KT5823, a specific inhibitor of proteinkinase G (PKG), was purchased from Calbiochem (San Diego, CA).S3226 was kindly donated by Dr. Jurgen Punter from Sanofi-AventisDeutschland GmbH (Frankfurt, Germany). EZ-Link Sulfo-NHS-SS-Biotin as well as immunopure immobilized streptavidin were pur-chased from Thermo Fisher Scientific (Rockford, IL). A monoclonalantibody (mAb) raised to the renal brush-border Na�/H� exchangerisoform 3 (NHE3), clone 3H3, was kindly provided by Drs. DanielBiemesderfer and Peter Aronson (Yale University, New Haven, CT).Phosphospecific NHE3 mAbs to NHE3 (phospho-serines 552 and605), anti-PS552, clone 14D5, anti-PS605, and clone 10A8 (32) werepurchased from Santa Cruz Biotechnology (Santa Cruz, CA). Horse-radish peroxidase-conjugated secondary antibodies were purchasedfrom Life Technologies (Carlsbad, CA).
Animals. Animal procedures and protocols were followed in accor-dance with the ethical principles in animal research of the BrazilianCollege of Animal Experimentation and were approved by the insti-tutional animal care and use committee. Experiments were performedusing male Wistar rats (250 to 300 g) housed under standardizedconditions (constant temperature of 22°C, 12:12-h dark-light cycle,and relative humidity of 60%) at the University of São Paulo Bio-medical Institute animal facility. To perform stationary in vivo mi-croperfusion, the animals were anesthetized with intramuscular ket-amine (75 mg/kg) and xylazine (8 mg/kg). The left jugular vein wascannulated for infusion of 3% mannitol in isotonic saline at a rate of0.1 ml/min. The kidney was exposed by a lumbar approach andprepared for “in vivo” micropuncture.
Stationary in vivo microperfusion. The microperfusion procedurewas performed as described previously (23). A proximal tubule waspunctured by means of a double-barreled micropipette, one barrelbeing used to inject FDC-green colored Ringer perfusion solution (inmM: 80 NaCl, 5 KCl, 25 NaHCO3, 1 CaCl2, 1.2 MgSO4, and raffinoseto reach isotonicity, at 0 PCO2), and the other to inject Sudan black-colored castor oil used to block the injected fluid columns in thelumen. A single micropipette containing the same Ringer solution plusthe polypeptide agent was impaled into a neighboring proximal loop.A proximal segment of the same nephron, recognized by the coloredperfusion and by its low transepithelial potential difference (PD), wasimpaled by a double-barreled asymmetric microelectrode to measureintratubular pH, the larger barrel containing at its tip the H� ion-sensitive ion exchange resin (Fluka, Buchs, Switzerland) and thesmaller, 1 M KCl reference solution colored by FDC-green. Propertiesof the microelectrode were described previously (23). The pH micro-electrodes were calibrated before and after every impalement on thekidney’s surface by perfusion with 20 mM phosphate-Ringer buffersolutions containing 130 mM NaCl, at 37°C. The pH values wereadjusted to 6.5, 7.0, and 7.5 with 0.1 N NaOH or HCl. A luminal oilblock was split by perfusions so that the solution was isolated andblocked by oil. The perfusion rate was sufficient to increase luminalpH to values near those of the perfusion fluid, i.e., 25 mM NaHCO3
or pH �8.0. After the luminal solution was blocked with oil, theincrease in luminal H� activities, representing bicarbonate reabsorp-tion, was followed until a stable level was reached.
By this technique, several (�2 to 5 each) control, experimental, andrecovery curves were obtained, the mean of control plus last recoverycurves and experimental curves constituting the pair of values for thistubule. The value of N given for an experimental condition corre-sponds to the number of perfused tubules, �1 to 3 being perfused inone rat, and for each group at least 4 rats were used. Luminal HCO3
�
activity, initially starting with 25 mM, was then progressively reducedto a stationary level (HCO3
�s) by H� secretion. The voltage between
the microelectrode barrels, representing the luminal H� activity, wassampled every 0.5 s by an AD converter (Lynx, São Paulo, Brazil) ina microcomputer. At the same time, the potential difference betweenthe reference barrel and ground (the rat tail) was recorded, giving theevolution of transepithelial PD with time during the perfusion. Lumi-nal bicarbonate was calculated from luminal pH and blood PCO2 was
measured by a Severinghaus electrode. The rate of tubular acidifica-tion was expressed as the half-time of the exponential reduction of theinjected HCO3
� concentration to its stationary level (t1/2). Net HCO3�
reabsorption (JHCO3�) per cm2 of tubule epithelium was calculated
from the equation: JHCO3� � ln 2/t1/2 ([HCO3
�]0 � [HCO3�]s)*r/2,
where t/2 is the half-time of bicarbonate reabsorption, r is the tubuleradius measured by an ocular micrometer, and [HCO3
�]o and [HCO3�]s
are the concentrations of the injected HCO3� and HCO3
� at thestationary level, respectively. The tubules were perfused with controlsolution, 10�6 M UGN, and/or the specific NHE3 inhibitor, 5 �MS3226. In the experiments designed to investigate the signalingmechanisms of UGN action, we used 10�6 M KT5823 (PKG inhib-itor); 10�5 M H89 (PKA inhibitor).
Gene expression of GC-C receptor in the rat kidney and LLC-PK1
cells. RNA was purified from rat kidney and LLC-PK1 cell corticesusing TRIzol Reagent (Invitrogen, Carlsbad, CA) following the man-ufacturer’s protocol. Total RNA (2 �g) was reversed transcribed usingSuper Script III (Invitrogen) and random hexamer primers werefollowed by amplification of cDNA by PCR using Taq DNA Poly-merase (Invitrogen). The specific primers to the rat GC-C receptor(NM_013170.1), forward (5=-ATGACGTCACTCCTGGGCTT-3=)and reverse (5=-GCGCTTTCGCACGATGTCCAGT-3=), were usedfor initial PCR with an expected PCR product of �200 bp plusmultiple bands. Forward primer (5=-GACGTCACTCCTGGGCT-TGGC-3=) and reverse primer (5=-GTGGCACTTCT-GCCTCACCT-3=) were used for nested PCR with an expected PCR product of �90 bp.Thermal cycling for initial rat GC-C analysis included a denaturation stepat 95°C for 5 min followed by 30 cycles of 95°C for 30 s, annealingtemperature of 58°C for initial PCR, and 60°C for nested PCR for 30 sand 72°C for 1 min. LLC-PK1 cell cDNA was subjected to PCR usingtwo sets of oligonucleotide primers [5=-CCCGCTGTTGGCCTTG-GCTT-3= (forward) and 5=-GGTACACGAGGGCCCCAGGA-3= (re-verse)] and [5=-GCCTCCCTG-AAAGCCCAGAGC-3= (forward) and5=-TACACGAGGGCCCCAGGAGG-3= (reverse)]. These two PCRprimer pairs were designed according to the porcine guanylate cyclase 2C(heat-stable enterotoxin receptor) mRNA (NM_214105.1). The expectedsizes of the amplified DNA bands were 367 and 438 bp. PCR productswere resolved by electrophoresis at 100 V through 1% agarose gel andvisualized with ethidium bromide.
Cell culture. LLC-PK1 cells were obtained from American TypeCulture Collection and used from passages 2 to 7. Serial cultures weremaintained in DMEM medium supplemented with 45 mM NaHCO3,25 mM HEPES buffer, 0.1 mM sodium pyruvate, 0.01 mM nones-sential amino acids, 10% vol/vol heat-inactivated fetal bovine serum,100 IU/ml penicillin, and 100 �g/ml streptomycin. Cells were grownat 37°C, 95% humidified air-5% CO2 (pH 7.4) in a CO2 incubator(Lab-Line Instruments, Melrose Park, IL). The cells were subculturedwith trypsin-EGTA (0.02%) and seeded on tissue culture plates withsterile glass coverslips (for pH measurements) to become confluent.For all experiments, cells were placed in serum-free medium 24 hbefore the experiments.
Measurement of Na�/H� exchanger activity by fluorescencemicroscopy. Intracellular pH was examined in cells loaded with thefluorescent pH-sensitive probe BCECF by measuring the fluorescenceintensity at alternating excitation wavelengths of 440 and 495 nmusing a method described previously (7). Briefly, cells were grown toconfluence on glass coverslips and were placed in a thermoregulatedchamber mounted on an inverted epifluorescence microscope (Nikon,TMD) and loaded with 10 �M BCECF-AM (in control solution,containing in mM: 141 NaCl, 5.4 KCl, 1.0 CaCl2, 0.4 KH2PO4, 0.5MgCl2, 0.4 MgSO4, 0.3 Na2HPO4, 10 HEPES, 0.6 glucose) for 5 min.After several washes, the BCECF-loaded cells were prepulsed with 20mM NH4Cl for 2 min for subsequent acid loading and then exposedto control solution in the presence of UGN with or without severalinhibitors, as described later. Measurements of NHE3 activity wereperformed in the presence of 10 �M HOE694 to obviate the contri-bution by NHE1 (7). In all the experiments, we calculated the initial
F1400 MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org
rate of pH recovery (dpHi/dt, pH U/min) from the first 2 min after thestart of the pHi recovery curve by linear regression analysis. Fluores-cence was monitored using alternately 440 or 495 nm as excitationwavelengths using a xenon light source. Emission was measured at530 nm by a photomultiplier-based fluorescence system (GeorgiaInstruments, PMT-4000) at time intervals of 5 s. pHi was calculatedfrom the fluorescence emission ratio of the two excitation wave-lengths using a standard calibration procedure based on the use of 10�M nigericin in high-potassium Ringer at different pHs.
cGMP and cAMP assay. LLC-PK1 cells were cultured to 100%confluence in 96-well plates. Cells were incubated for 10 min with culturemedium containing 1 mM 3-isobutyl-1-methylxanthine (IBMX) orIBMX and 10�6 M UGN or 10�6 M ANP (used as positive control incGMP assay) or 10�4 M forskolin (used as positive control in cAMPassay). cGMP and cAMP were measured by using the Amershamenzyme immunoassay Biotrak (EIA) System (GE Healthcare) accordingto specifications of the manufacturer.
Determination of PKA activity in cell lysates. LLC-PK1 grown toconfluence in 24-well plates were treated or not with UGN andsubsequently solubilized in lysis buffer containing 20 mM MOPS, 50mM �-glycerolphosphate, 50 mM NaF, 1 mM sodium vanadate, 5mM EGTA, 2 mM EDTA, 1% NP-40, 1 mM DTT, 1 mM benzami-dine, 1 mM PMSF, 10 �g/ml aprotinin, and 10 �g/ml leupeptin. PKAactivity was measured in this cell lysate using a nonradioactive PKAKinase Activity Assay (Enzo Life Sciences, Farmingdale, NY) ac-cording to the manufacturer’s instructions.
Determination of PKG activity in cell lysates. LLC-PK1 grown toconfluency in 24-well plates were treated or not with UGN andsubsequently solubilized in extraction buffer containing 20 mM Tris,pH 7.4, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 0.2 mM PMSF,1 �g/ml pepstatin, 0.5 �g/ml leupeptin, 2 mM NaF, 0.2 mM Na3VO4,5 mM �-mercaptoethanol. PKG activity was measured in this celllysate using a single-site and semiquantative CycLex cGK Assay Kit(CycLex, Nagano, Japan) according to the manufacturer’s instruc-tions.
Cell surface biotinylation. The assay was performed as describedpreviously (25). Cells were rinsed twice in ice-cold PBS-Ca-Mg (PBSwith 0.1 mM CaCl2, 1.0 mM MgCl2). Surface membrane proteinswere then biotinylated by incubating the cells twice for 25 min with2 ml of ice-cold biotinylation buffer (150 mM NaCl, 10 mM trieth-anolamine, 2 mM CaCl2, and 2 mg/ml EZ-Link sulfo-NHS-SS-biotin). Cells were then rinsed twice for 20 min with a quenchingbuffer (PBS-Ca-Mg, 100 mM glycine), washed twice with ice-coldPBS-Ca-Mg, and strapped into ice-cold solubilization buffer (50 mMTris, 150 mM NaCl, 1 mM EDTA, 0.5% sodium deoxycholate, 1%Triton X-100, pH 7.4) containing protease inhibitors (0.7 �g/mlpepstatin A, 0.5 �g/ml leupeptin, and 40 �g/ml PMSF). After lysis onice for 60 min, extracts were centrifuged for 10 min at 14,000 g and4°C. The protein concentration of the supernatants was measured (38)and equal protein amounts of cell lysate (500 �g) were equilibratedwith streptavidin-agarose beads at 4°C. The beads were then washedfive times in ice-cold solubilization buffer. Biotinylated proteins werereleased by incubation in Laemmli buffer and subjected to SDS-PAGE and immunoblotting.
SDS-PAGE and immunoblotting. Protein samples were solubilizedin Laemmli sample buffer and separated by SDS-PAGE using 7.5%polyacrylamide gels. For immunoblotting, proteins were transferred toPVDF (Millipore Immobilon-P, Millipore, Bedford, MA) at 350 mAfor 8–10 h at 4°C with a TE 62 transfer electrophoresis unit (GEHealthCare). Sheets of PVDF containing transferred proteins wereincubated first in Blotto (5% nonfat dry milk and 0.1% Tween 20 inPBS, pH 7.4) for 1 h to block nonspecific binding of antibody,followed by overnight incubation in primary antibody. Primary anti-bodies, diluted in Blotto, were used at dilutions ranging from 1:200 to1:50,000. The sheets were then washed in Blotto and incubated for 1h with an appropriate horseradish peroxidase-conjugated secondaryantibody diluted 1:2,000 in Blotto. After being washed 5� in Blotto
and 2� PBS (pH 7.4), an enhanced chemiluminescence detection(ECL) system (GE HealthCare) with Kodak Biomax imaging film(Kodak) was used for visualization of the bound antibodies. Thevisualized bands were digitized using the ImageScanner III (GEHealthCare) and quantified using the Scion Image Software (Scion,Federick, MD). In some experiments, PVDF blots were reprobed withadditional primary antibodies after the first antibody was strippedaway. This was accomplished by incubating the PVDF sheets in RestoreWestern Blot Stripping Buffer (Thermo Scientific, Rockford, IL).
Statistical analysis. The data were analyzed by a Visual Basicprogram in Excel software. Statistical comparisons were made by theunpaired t-test, taking the probability of 0.05 (5%) as the limit ofsignificance. When more than two groups were compared, one-wayANOVA followed by Tukey’s post hoc test, taking 0.05 (5%) as limitof significance, was performed. In microperfusion experiments, aminimum of six tubules was used (n � number of perfused tubules).
RESULTS
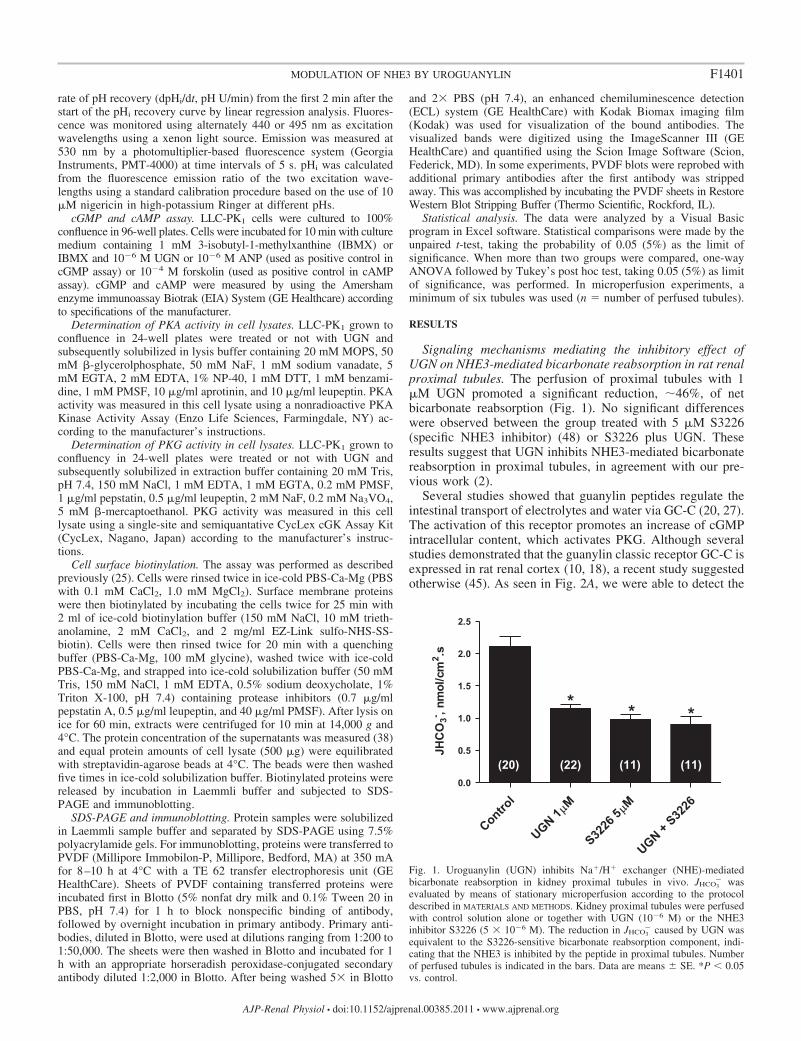
Signaling mechanisms mediating the inhibitory effect ofUGN on NHE3-mediated bicarbonate reabsorption in rat renalproximal tubules. The perfusion of proximal tubules with 1�M UGN promoted a significant reduction, �46%, of netbicarbonate reabsorption (Fig. 1). No significant differenceswere observed between the group treated with 5 �M S3226(specific NHE3 inhibitor) (48) or S3226 plus UGN. Theseresults suggest that UGN inhibits NHE3-mediated bicarbonatereabsorption in proximal tubules, in agreement with our pre-vious work (2).
Several studies showed that guanylin peptides regulate theintestinal transport of electrolytes and water via GC-C (20, 27).The activation of this receptor promotes an increase of cGMPintracellular content, which activates PKG. Although severalstudies demonstrated that the guanylin classic receptor GC-C isexpressed in rat renal cortex (10, 18), a recent study suggestedotherwise (45). As seen in Fig. 2A, we were able to detect the
Control Mµ µ
UGN 1M
S3226
5
UGN + S32
260.0
0.5
1.0
1.5
2.0
2.5
(20) (22)
* *
(11)
*
JHC
O3- , n
mol
/cm
2 .s
(11)
Fig. 1. Uroguanylin (UGN) inhibits Na�/H� exchanger (NHE)-mediatedbicarbonate reabsorption in kidney proximal tubules in vivo. JHCO3
� wasevaluated by means of stationary microperfusion according to the protocoldescribed in MATERIALS AND METHODS. Kidney proximal tubules were perfusedwith control solution alone or together with UGN (10�6 M) or the NHE3inhibitor S3226 (5 � 10�6 M). The reduction in JHCO3
� caused by UGN wasequivalent to the S3226-sensitive bicarbonate reabsorption component, indi-cating that the NHE3 is inhibited by the peptide in proximal tubules. Numberof perfused tubules is indicated in the bars. Data are means � SE. *P 0.05vs. control.
F1401MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org
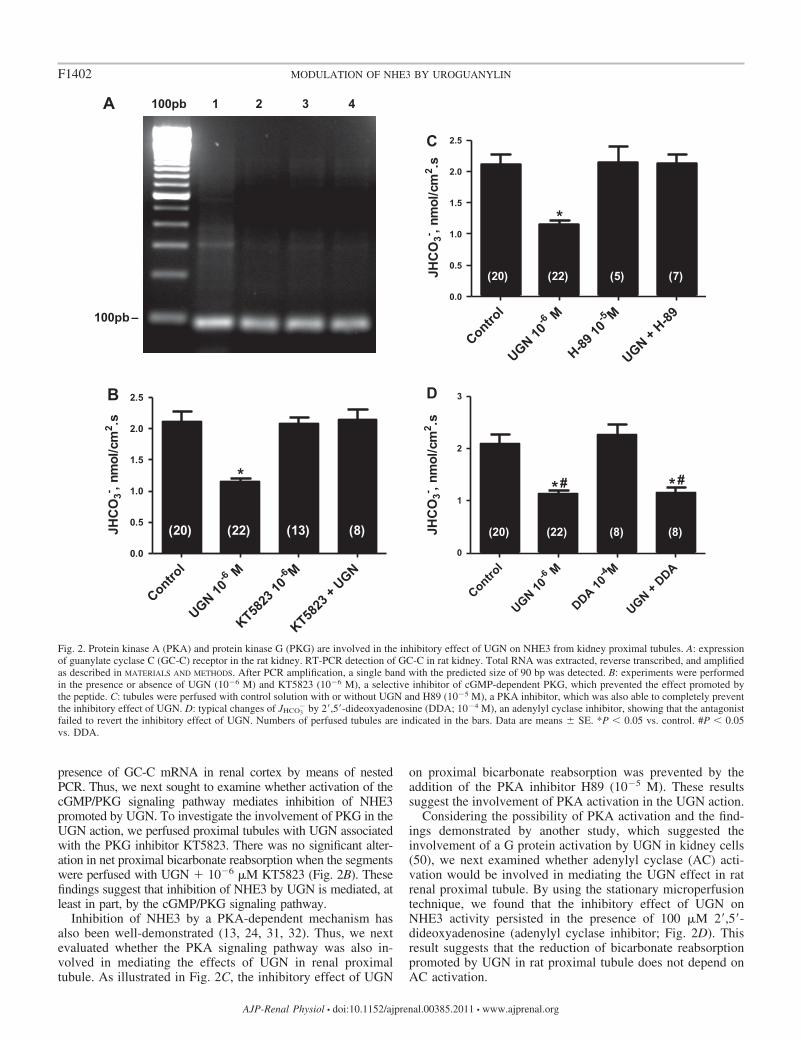
presence of GC-C mRNA in renal cortex by means of nestedPCR. Thus, we next sought to examine whether activation of thecGMP/PKG signaling pathway mediates inhibition of NHE3promoted by UGN. To investigate the involvement of PKG in theUGN action, we perfused proximal tubules with UGN associatedwith the PKG inhibitor KT5823. There was no significant alter-ation in net proximal bicarbonate reabsorption when the segmentswere perfused with UGN � 10�6 �M KT5823 (Fig. 2B). Thesefindings suggest that inhibition of NHE3 by UGN is mediated, atleast in part, by the cGMP/PKG signaling pathway.
Inhibition of NHE3 by a PKA-dependent mechanism hasalso been well-demonstrated (13, 24, 31, 32). Thus, we nextevaluated whether the PKA signaling pathway was also in-volved in mediating the effects of UGN in renal proximaltubule. As illustrated in Fig. 2C, the inhibitory effect of UGN
on proximal bicarbonate reabsorption was prevented by theaddition of the PKA inhibitor H89 (10�5 M). These resultssuggest the involvement of PKA activation in the UGN action.
Considering the possibility of PKA activation and the find-ings demonstrated by another study, which suggested theinvolvement of a G protein activation by UGN in kidney cells(50), we next examined whether adenylyl cyclase (AC) acti-vation would be involved in mediating the UGN effect in ratrenal proximal tubule. By using the stationary microperfusiontechnique, we found that the inhibitory effect of UGN onNHE3 activity persisted in the presence of 100 �M 2=,5=-dideoxyadenosine (adenylyl cyclase inhibitor; Fig. 2D). Thisresult suggests that the reduction of bicarbonate reabsorptionpromoted by UGN in rat proximal tubule does not depend onAC activation.
A
Control M-6
UGN 10M-6
KT5823
10
KT5823
+ UGN
0.0
0.5
1.0
1.5
2.0
2.5
(20) (22) (13) (8)
*
B
JHC
O3- , n
mol
/cm
2 .s
Control M-6
UGN 10M-5
H-89 10
UGN + H-89
0.0
0.5
1.0
1.5
2.0
2.5
(20) (7)(22) (5)
*
C
JHC
O3- , n
mol
/cm
2 .s
Control M-6
UGN 10M-4
DDA 10
UGN + DDA
0
1
2
3D
*#*#
(20) (22) (8) (8)JHC
O3- , n
mol
/cm
2 .s
Fig. 2. Protein kinase A (PKA) and protein kinase G (PKG) are involved in the inhibitory effect of UGN on NHE3 from kidney proximal tubules. A: expressionof guanylate cyclase C (GC-C) receptor in the rat kidney. RT-PCR detection of GC-C in rat kidney. Total RNA was extracted, reverse transcribed, and amplifiedas described in MATERIALS AND METHODS. After PCR amplification, a single band with the predicted size of 90 bp was detected. B: experiments were performedin the presence or absence of UGN (10�6 M) and KT5823 (10�6 M), a selective inhibitor of cGMP-dependent PKG, which prevented the effect promoted bythe peptide. C: tubules were perfused with control solution with or without UGN and H89 (10�5 M), a PKA inhibitor, which was also able to completely preventthe inhibitory effect of UGN. D: typical changes of JHCO3
� by 2=,5=-dideoxyadenosine (DDA; 10�4 M), an adenylyl cyclase inhibitor, showing that the antagonistfailed to revert the inhibitory effect of UGN. Numbers of perfused tubules are indicated in the bars. Data are means � SE. *P 0.05 vs. control. #P 0.05vs. DDA.
F1402 MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org
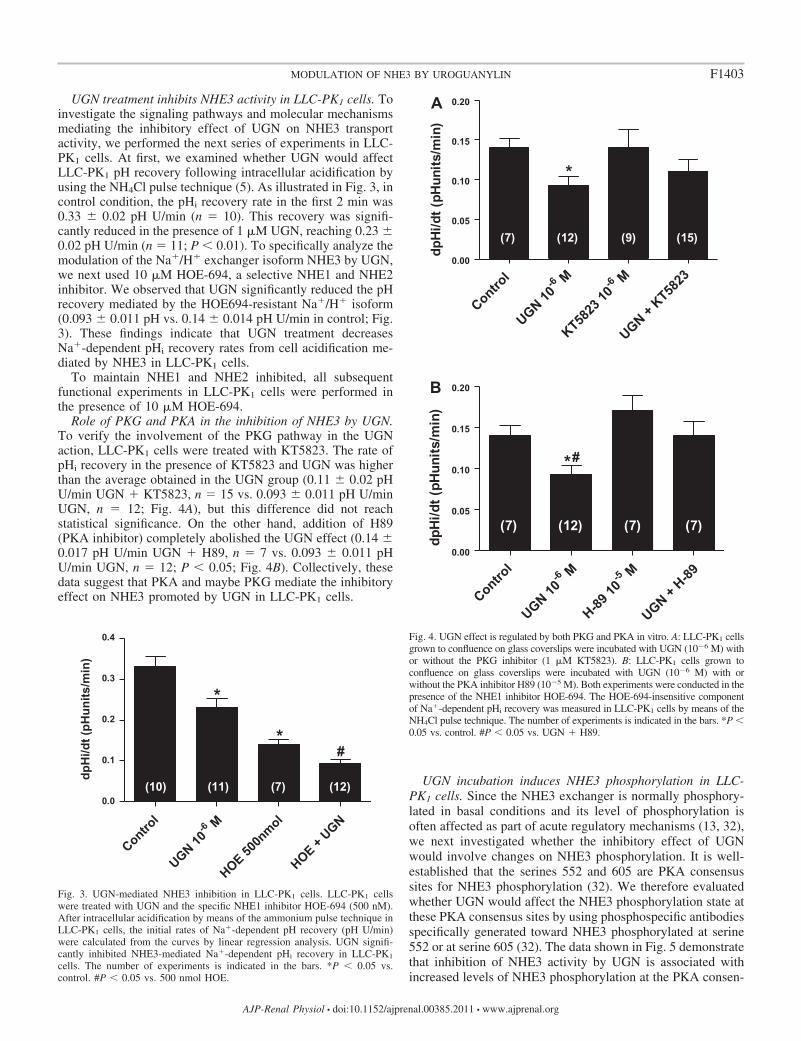
UGN treatment inhibits NHE3 activity in LLC-PK1 cells. Toinvestigate the signaling pathways and molecular mechanismsmediating the inhibitory effect of UGN on NHE3 transportactivity, we performed the next series of experiments in LLC-PK1 cells. At first, we examined whether UGN would affectLLC-PK1 pH recovery following intracellular acidification byusing the NH4Cl pulse technique (5). As illustrated in Fig. 3, incontrol condition, the pHi recovery rate in the first 2 min was0.33 � 0.02 pH U/min (n � 10). This recovery was signifi-cantly reduced in the presence of 1 �M UGN, reaching 0.23 �0.02 pH U/min (n � 11; P 0.01). To specifically analyze themodulation of the Na�/H� exchanger isoform NHE3 by UGN,we next used 10 �M HOE-694, a selective NHE1 and NHE2inhibitor. We observed that UGN significantly reduced the pHrecovery mediated by the HOE694-resistant Na�/H� isoform(0.093 � 0.011 pH vs. 0.14 � 0.014 pH U/min in control; Fig.3). These findings indicate that UGN treatment decreasesNa�-dependent pHi recovery rates from cell acidification me-diated by NHE3 in LLC-PK1 cells.
To maintain NHE1 and NHE2 inhibited, all subsequentfunctional experiments in LLC-PK1 cells were performed inthe presence of 10 �M HOE-694.
Role of PKG and PKA in the inhibition of NHE3 by UGN.To verify the involvement of the PKG pathway in the UGNaction, LLC-PK1 cells were treated with KT5823. The rate ofpHi recovery in the presence of KT5823 and UGN was higherthan the average obtained in the UGN group (0.11 � 0.02 pHU/min UGN � KT5823, n � 15 vs. 0.093 � 0.011 pH U/minUGN, n � 12; Fig. 4A), but this difference did not reachstatistical significance. On the other hand, addition of H89(PKA inhibitor) completely abolished the UGN effect (0.14 �0.017 pH U/min UGN � H89, n � 7 vs. 0.093 � 0.011 pHU/min UGN, n � 12; P 0.05; Fig. 4B). Collectively, thesedata suggest that PKA and maybe PKG mediate the inhibitoryeffect on NHE3 promoted by UGN in LLC-PK1 cells.
UGN incubation induces NHE3 phosphorylation in LLC-PK1 cells. Since the NHE3 exchanger is normally phosphory-lated in basal conditions and its level of phosphorylation isoften affected as part of acute regulatory mechanisms (13, 32),we next investigated whether the inhibitory effect of UGNwould involve changes on NHE3 phosphorylation. It is well-established that the serines 552 and 605 are PKA consensussites for NHE3 phosphorylation (32). We therefore evaluatedwhether UGN would affect the NHE3 phosphorylation state atthese PKA consensus sites by using phosphospecific antibodiesspecifically generated toward NHE3 phosphorylated at serine552 or at serine 605 (32). The data shown in Fig. 5 demonstratethat inhibition of NHE3 activity by UGN is associated withincreased levels of NHE3 phosphorylation at the PKA consen-
Control M-6
UGN 10
HOE 500n
mol
HOE + UGN
0.0
0.1
0.2
0.3
0.4
(10) (12)(7)(11)
*
*#
dpH
i/dt (
pHun
its/m
in)
Fig. 3. UGN-mediated NHE3 inhibition in LLC-PK1 cells. LLC-PK1 cellswere treated with UGN and the specific NHE1 inhibitor HOE-694 (500 nM).After intracellular acidification by means of the ammonium pulse technique inLLC-PK1 cells, the initial rates of Na�-dependent pH recovery (pH U/min)were calculated from the curves by linear regression analysis. UGN signifi-cantly inhibited NHE3-mediated Na�-dependent pHi recovery in LLC-PK1
cells. The number of experiments is indicated in the bars. *P 0.05 vs.control. #P 0.05 vs. 500 nmol HOE.
Control M-6
UGN 10 M-6
KT5823
10
UGN + KT58
230.00
0.05
0.10
0.15
0.20
(7) (15)(9)(12)
*
A
dpH
i/dt (
pHun
its/m
in)
Control M-6
UGN 10 M-5
H-89 10
UGN + H-89
0.00
0.05
0.10
0.15
0.20
(7) (12) (7) (7)
*#
B
dpH
i/dt (
pHun
its/m
in)
Fig. 4. UGN effect is regulated by both PKG and PKA in vitro. A: LLC-PK1 cellsgrown to confluence on glass coverslips were incubated with UGN (10�6 M) withor without the PKG inhibitor (1 �M KT5823). B: LLC-PK1 cells grown toconfluence on glass coverslips were incubated with UGN (10�6 M) with orwithout the PKA inhibitor H89 (10�5 M). Both experiments were conducted in thepresence of the NHE1 inhibitor HOE-694. The HOE-694-insensitive componentof Na�-dependent pHi recovery was measured in LLC-PK1 cells by means of theNH4Cl pulse technique. The number of experiments is indicated in the bars. *P 0.05 vs. control. #P 0.05 vs. UGN � H89.
F1403MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org
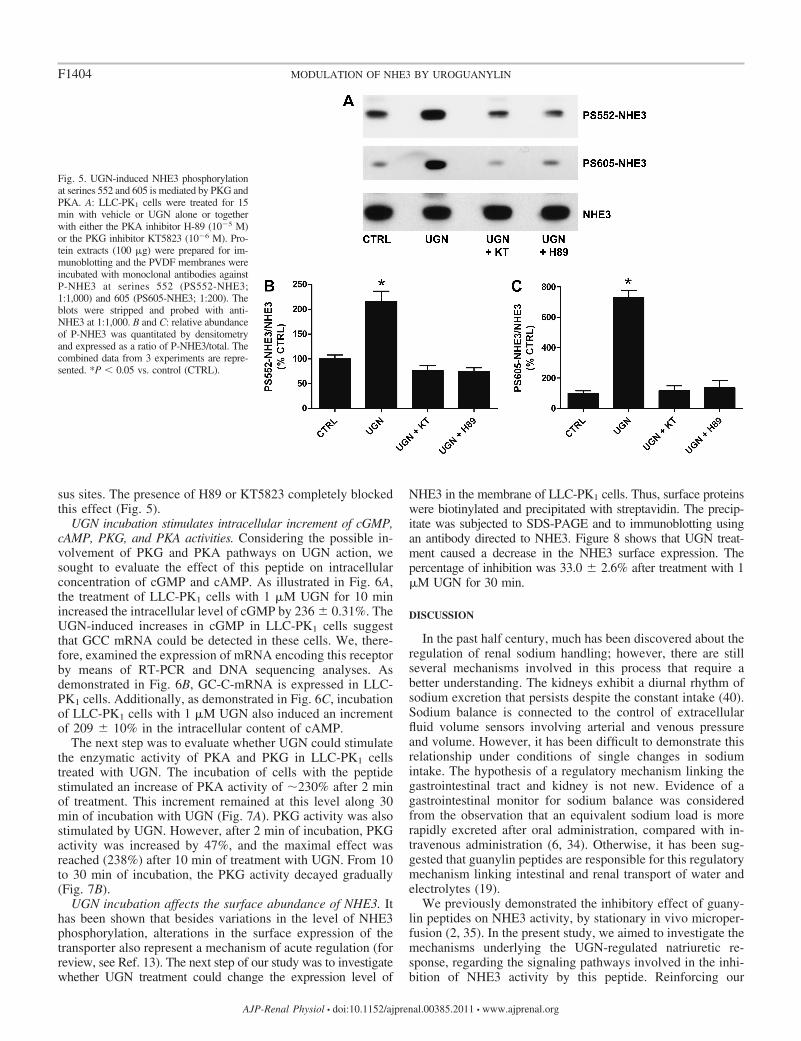
sus sites. The presence of H89 or KT5823 completely blockedthis effect (Fig. 5).
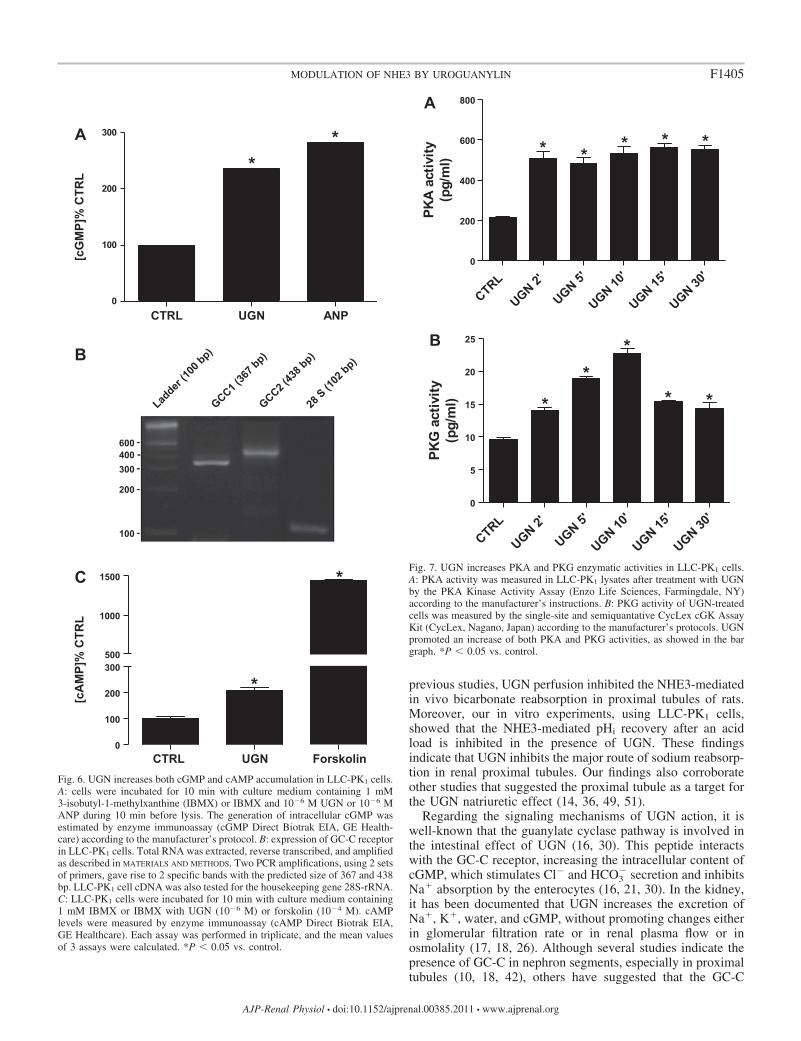
UGN incubation stimulates intracellular increment of cGMP,cAMP, PKG, and PKA activities. Considering the possible in-volvement of PKG and PKA pathways on UGN action, wesought to evaluate the effect of this peptide on intracellularconcentration of cGMP and cAMP. As illustrated in Fig. 6A,the treatment of LLC-PK1 cells with 1 �M UGN for 10 minincreased the intracellular level of cGMP by 236 � 0.31%. TheUGN-induced increases in cGMP in LLC-PK1 cells suggestthat GCC mRNA could be detected in these cells. We, there-fore, examined the expression of mRNA encoding this receptorby means of RT-PCR and DNA sequencing analyses. Asdemonstrated in Fig. 6B, GC-C-mRNA is expressed in LLC-PK1 cells. Additionally, as demonstrated in Fig. 6C, incubationof LLC-PK1 cells with 1 �M UGN also induced an incrementof 209 � 10% in the intracellular content of cAMP.
The next step was to evaluate whether UGN could stimulatethe enzymatic activity of PKA and PKG in LLC-PK1 cellstreated with UGN. The incubation of cells with the peptidestimulated an increase of PKA activity of �230% after 2 minof treatment. This increment remained at this level along 30min of incubation with UGN (Fig. 7A). PKG activity was alsostimulated by UGN. However, after 2 min of incubation, PKGactivity was increased by 47%, and the maximal effect wasreached (238%) after 10 min of treatment with UGN. From 10to 30 min of incubation, the PKG activity decayed gradually(Fig. 7B).
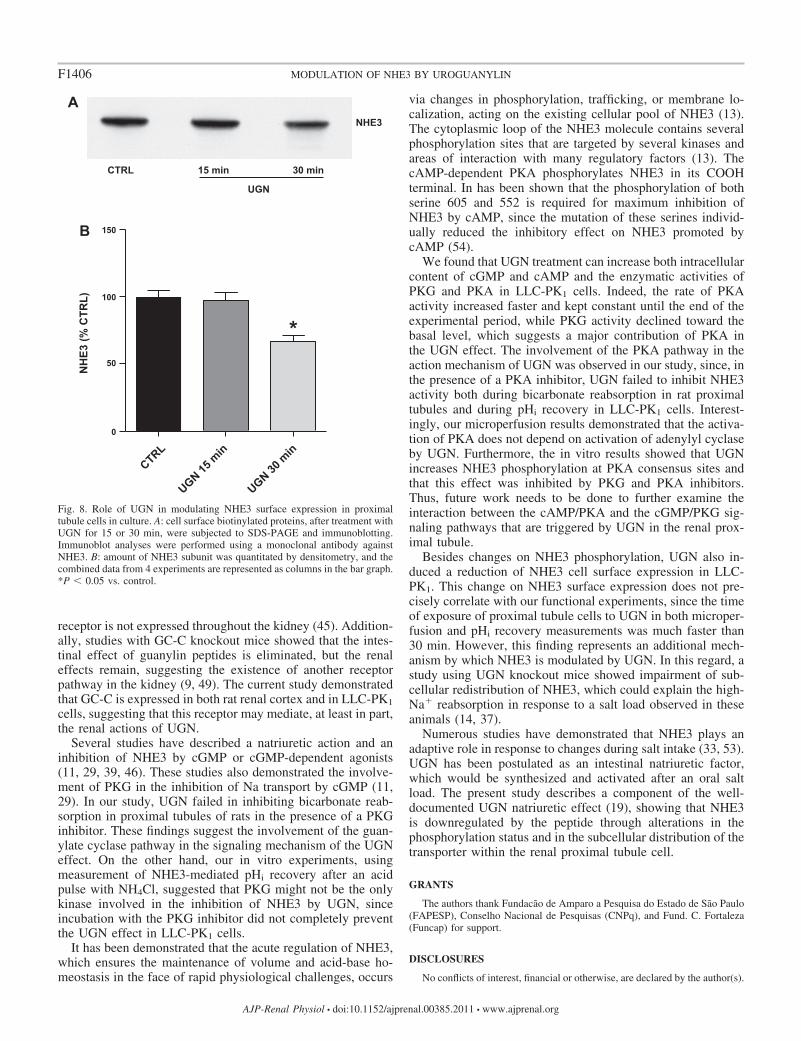
UGN incubation affects the surface abundance of NHE3. Ithas been shown that besides variations in the level of NHE3phosphorylation, alterations in the surface expression of thetransporter also represent a mechanism of acute regulation (forreview, see Ref. 13). The next step of our study was to investigatewhether UGN treatment could change the expression level of
NHE3 in the membrane of LLC-PK1 cells. Thus, surface proteinswere biotinylated and precipitated with streptavidin. The precip-itate was subjected to SDS-PAGE and to immunoblotting usingan antibody directed to NHE3. Figure 8 shows that UGN treat-ment caused a decrease in the NHE3 surface expression. Thepercentage of inhibition was 33.0 � 2.6% after treatment with 1�M UGN for 30 min.
DISCUSSION
In the past half century, much has been discovered about theregulation of renal sodium handling; however, there are stillseveral mechanisms involved in this process that require abetter understanding. The kidneys exhibit a diurnal rhythm ofsodium excretion that persists despite the constant intake (40).Sodium balance is connected to the control of extracellularfluid volume sensors involving arterial and venous pressureand volume. However, it has been difficult to demonstrate thisrelationship under conditions of single changes in sodiumintake. The hypothesis of a regulatory mechanism linking thegastrointestinal tract and kidney is not new. Evidence of agastrointestinal monitor for sodium balance was consideredfrom the observation that an equivalent sodium load is morerapidly excreted after oral administration, compared with in-travenous administration (6, 34). Otherwise, it has been sug-gested that guanylin peptides are responsible for this regulatorymechanism linking intestinal and renal transport of water andelectrolytes (19).
We previously demonstrated the inhibitory effect of guany-lin peptides on NHE3 activity, by stationary in vivo microper-fusion (2, 35). In the present study, we aimed to investigate themechanisms underlying the UGN-regulated natriuretic re-sponse, regarding the signaling pathways involved in the inhi-bition of NHE3 activity by this peptide. Reinforcing our
Fig. 5. UGN-induced NHE3 phosphorylationat serines 552 and 605 is mediated by PKG andPKA. A: LLC-PK1 cells were treated for 15min with vehicle or UGN alone or togetherwith either the PKA inhibitor H-89 (10�5 M)or the PKG inhibitor KT5823 (10�6 M). Pro-tein extracts (100 �g) were prepared for im-munoblotting and the PVDF membranes wereincubated with monoclonal antibodies againstP-NHE3 at serines 552 (PS552-NHE3;1:1,000) and 605 (PS605-NHE3; 1:200). Theblots were stripped and probed with anti-NHE3 at 1:1,000. B and C: relative abundanceof P-NHE3 was quantitated by densitometryand expressed as a ratio of P-NHE3/total. Thecombined data from 3 experiments are repre-sented. *P 0.05 vs. control (CTRL).
F1404 MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org
previous studies, UGN perfusion inhibited the NHE3-mediatedin vivo bicarbonate reabsorption in proximal tubules of rats.Moreover, our in vitro experiments, using LLC-PK1 cells,showed that the NHE3-mediated pHi recovery after an acidload is inhibited in the presence of UGN. These findingsindicate that UGN inhibits the major route of sodium reabsorp-tion in renal proximal tubules. Our findings also corroborateother studies that suggested the proximal tubule as a target forthe UGN natriuretic effect (14, 36, 49, 51).
Regarding the signaling mechanisms of UGN action, it iswell-known that the guanylate cyclase pathway is involved inthe intestinal effect of UGN (16, 30). This peptide interactswith the GC-C receptor, increasing the intracellular content ofcGMP, which stimulates Cl� and HCO3
� secretion and inhibitsNa� absorption by the enterocytes (16, 21, 30). In the kidney,it has been documented that UGN increases the excretion ofNa�, K�, water, and cGMP, without promoting changes eitherin glomerular filtration rate or in renal plasma flow or inosmolality (17, 18, 26). Although several studies indicate thepresence of GC-C in nephron segments, especially in proximaltubules (10, 18, 42), others have suggested that the GC-C
CTRL UGN ANP0
100
200
300
**
[cG
MP]
% C
TRL
CTRL UGN Forskolin0
100
200
300500
1000
1500
*
*
[cA
MP]
% C
TRL
Ladder
(100 b
p)
GCC2 (43
8 bp)
GCC1 (36
7 bp)
28 S (1
02 bp)
600400300
200
100
A
B
C
Fig. 6. UGN increases both cGMP and cAMP accumulation in LLC-PK1 cells.A: cells were incubated for 10 min with culture medium containing 1 mM3-isobutyl-1-methylxanthine (IBMX) or IBMX and 10�6 M UGN or 10�6 MANP during 10 min before lysis. The generation of intracellular cGMP wasestimated by enzyme immunoassay (cGMP Direct Biotrak EIA, GE Health-care) according to the manufacturer’s protocol. B: expression of GC-C receptorin LLC-PK1 cells. Total RNA was extracted, reverse transcribed, and amplifiedas described in MATERIALS AND METHODS. Two PCR amplifications, using 2 setsof primers, gave rise to 2 specific bands with the predicted size of 367 and 438bp. LLC-PK1 cell cDNA was also tested for the housekeeping gene 28S-rRNA.C: LLC-PK1 cells were incubated for 10 min with culture medium containing1 mM IBMX or IBMX with UGN (10�6 M) or forskolin (10�4 M). cAMPlevels were measured by enzyme immunoassay (cAMP Direct Biotrak EIA,GE Healthcare). Each assay was performed in triplicate, and the mean valuesof 3 assays were calculated. *P 0.05 vs. control.
CTRL
UGN 2'
UGN 5'
UGN 10'
UGN 15'
UGN 30'
0
200
400
600
800
* * * * *
A
PKA
activ
ity(p
g/m
l)
CTRL
UGN 2'
UGN 5'
UGN 10'
UGN 15'
UGN 30'
0
5
10
15
20
25B
**
*
* *PK
G a
ctiv
ity(p
g/m
l)
Fig. 7. UGN increases PKA and PKG enzymatic activities in LLC-PK1 cells.A: PKA activity was measured in LLC-PK1 lysates after treatment with UGNby the PKA Kinase Activity Assay (Enzo Life Sciences, Farmingdale, NY)according to the manufacturer’s instructions. B: PKG activity of UGN-treatedcells was measured by the single-site and semiquantative CycLex cGK AssayKit (CycLex, Nagano, Japan) according to the manufacturer’s protocols. UGNpromoted an increase of both PKA and PKG activities, as showed in the bargraph. *P 0.05 vs. control.
F1405MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org
receptor is not expressed throughout the kidney (45). Addition-ally, studies with GC-C knockout mice showed that the intes-tinal effect of guanylin peptides is eliminated, but the renaleffects remain, suggesting the existence of another receptorpathway in the kidney (9, 49). The current study demonstratedthat GC-C is expressed in both rat renal cortex and in LLC-PK1
cells, suggesting that this receptor may mediate, at least in part,the renal actions of UGN.
Several studies have described a natriuretic action and aninhibition of NHE3 by cGMP or cGMP-dependent agonists(11, 29, 39, 46). These studies also demonstrated the involve-ment of PKG in the inhibition of Na transport by cGMP (11,29). In our study, UGN failed in inhibiting bicarbonate reab-sorption in proximal tubules of rats in the presence of a PKGinhibitor. These findings suggest the involvement of the guan-ylate cyclase pathway in the signaling mechanism of the UGNeffect. On the other hand, our in vitro experiments, usingmeasurement of NHE3-mediated pHi recovery after an acidpulse with NH4Cl, suggested that PKG might not be the onlykinase involved in the inhibition of NHE3 by UGN, sinceincubation with the PKG inhibitor did not completely preventthe UGN effect in LLC-PK1 cells.
It has been demonstrated that the acute regulation of NHE3,which ensures the maintenance of volume and acid-base ho-meostasis in the face of rapid physiological challenges, occurs
via changes in phosphorylation, trafficking, or membrane lo-calization, acting on the existing cellular pool of NHE3 (13).The cytoplasmic loop of the NHE3 molecule contains severalphosphorylation sites that are targeted by several kinases andareas of interaction with many regulatory factors (13). ThecAMP-dependent PKA phosphorylates NHE3 in its COOHterminal. In has been shown that the phosphorylation of bothserine 605 and 552 is required for maximum inhibition ofNHE3 by cAMP, since the mutation of these serines individ-ually reduced the inhibitory effect on NHE3 promoted bycAMP (54).
We found that UGN treatment can increase both intracellularcontent of cGMP and cAMP and the enzymatic activities ofPKG and PKA in LLC-PK1 cells. Indeed, the rate of PKAactivity increased faster and kept constant until the end of theexperimental period, while PKG activity declined toward thebasal level, which suggests a major contribution of PKA inthe UGN effect. The involvement of the PKA pathway in theaction mechanism of UGN was observed in our study, since, inthe presence of a PKA inhibitor, UGN failed to inhibit NHE3activity both during bicarbonate reabsorption in rat proximaltubules and during pHi recovery in LLC-PK1 cells. Interest-ingly, our microperfusion results demonstrated that the activa-tion of PKA does not depend on activation of adenylyl cyclaseby UGN. Furthermore, the in vitro results showed that UGNincreases NHE3 phosphorylation at PKA consensus sites andthat this effect was inhibited by PKG and PKA inhibitors.Thus, future work needs to be done to further examine theinteraction between the cAMP/PKA and the cGMP/PKG sig-naling pathways that are triggered by UGN in the renal prox-imal tubule.
Besides changes on NHE3 phosphorylation, UGN also in-duced a reduction of NHE3 cell surface expression in LLC-PK1. This change on NHE3 surface expression does not pre-cisely correlate with our functional experiments, since the timeof exposure of proximal tubule cells to UGN in both microper-fusion and pHi recovery measurements was much faster than30 min. However, this finding represents an additional mech-anism by which NHE3 is modulated by UGN. In this regard, astudy using UGN knockout mice showed impairment of sub-cellular redistribution of NHE3, which could explain the high-Na� reabsorption in response to a salt load observed in theseanimals (14, 37).
Numerous studies have demonstrated that NHE3 plays anadaptive role in response to changes during salt intake (33, 53).UGN has been postulated as an intestinal natriuretic factor,which would be synthesized and activated after an oral saltload. The present study describes a component of the well-documented UGN natriuretic effect (19), showing that NHE3is downregulated by the peptide through alterations in thephosphorylation status and in the subcellular distribution of thetransporter within the renal proximal tubule cell.
GRANTS
The authors thank Fundacão de Amparo a Pesquisa do Estado de São Paulo(FAPESP), Conselho Nacional de Pesquisas (CNPq), and Fund. C. Fortaleza(Funcap) for support.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
CTRL
UGN 15 m
in
UGN 30 m
in0
50
100
150
*
B
CTRL 15 min 30 min
NHE3
UGN
NH
E3 (%
CTR
L)
A
Fig. 8. Role of UGN in modulating NHE3 surface expression in proximaltubule cells in culture. A: cell surface biotinylated proteins, after treatment withUGN for 15 or 30 min, were subjected to SDS-PAGE and immunoblotting.Immunoblot analyses were performed using a monoclonal antibody againstNHE3. B: amount of NHE3 subunit was quantitated by densitometry, and thecombined data from 4 experiments are represented as columns in the bar graph.*P 0.05 vs. control.
F1406 MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org

AUTHOR CONTRIBUTIONS
Author contributions: L.M.A.L., L.R.C.-L., A.C.C.G., M.C.F., and G.M.conception and design of research; L.M.A.L., L.R.C.-L., R.O.C., C.N.A.B.,R.D., A.C.C.G., and G.M. performed experiments; L.M.A.L., L.R.C.-L.,C.N.A.B., R.D., A.C.C.G., M.C.F., and G.M. analyzed data; L.M.A.L., L.R.C.-L., A.C.C.G., M.C.F., and G.M. interpreted results of experiments; L.M.A.L.,L.R.C.-L., C.N.A.B., R.D., and A.C.C.G. prepared figures; L.M.A.L. draftedmanuscript; L.M.A.L., L.R.C.-L., R.O.C., C.N.A.B., R.D., A.C.C.G., M.C.F.,and G.M. edited and revised manuscript; L.M.A.L., L.R.C.-L., R.O.C.,C.N.A.B., R.D., A.C.C.G., M.C.F., and G.M. approved final version ofmanuscript.
REFERENCES
1. Amemiya M, Loffing J, Lotscher M, Kaissling B, Alpern RJ, MoeOW. Expression of NHE-3 in the apical membrane of rat renal proximaltubule and thick ascending limb. Kidney Int 48: 1206–1215, 1995.
2. Amorim JB, Musa-Aziz R, Lessa LM, Malnic G, Fonteles MC. Effectof uroguanylin on potassium and bicarbonate transport in rat renal tubules.Can J Physiol Pharmacol 84: 1003–1010, 2006.
3. Aronson PS. Role of ion exchangers in mediating NaCl transport in theproximal tubule. Kidney Int 49: 1665–1670, 1996.
4. Biemesderfer D, Pizzonia J, Abu-Alfa A, Exner M, Reilly R, IgarashiP, Aronson PS. NHE3: a Na�/H� exchanger isoform of renal brushborder. Am J Physiol Renal Fluid Electrolyte Physiol 265: F736–F742,1993.
5. Boron WF, De Weer P. Intracellular pH transients in squid giant axonscaused by CO2, NH3, and metabolic inhibitors. J Gen Physiol 67: 91–112,1976.
6. Carey RM. Evidence for a splanchnic sodium input monitor regulatingrenal sodium excretion in man. Lack of dependence upon aldosterone.Circ Res 43: 19–23, 1978.
7. Carraro-Lacroix LR, Malnic G, Girardi AC. Regulation of Na�/H�
exchanger NHE3 by glucagon-like peptide 1 receptor agonist exendin-4 inrenal proximal tubule cells. Am J Physiol Renal Physiol 297: F1647–F1655, 2009.
8. Carrithers SL, Jackson BA, Cai WY, Greenberg RN, Ott CE. Site-specific effects of dietary salt intake on guanylin and uroguanylin mRNAexpression in rat intestine. Regul Pept 107: 87–95, 2002.
9. Carrithers SL, Ott CE, Hill MJ, Johnson BR, Cai W, Chang JJ, ShahRG, Sun C, Mann EA, Fonteles MC, Forte LR, Jackson BA, GiannellaRA, Greenberg RN. Guanylin and uroguanylin induce natriuresis in micelacking guanylyl cyclase-C receptor. Kidney Int 65: 40–53, 2004.
10. Carrithers SL, Taylor B, Cai WY, Johnson BR, Ott CE, GreenbergRN, Jackson BA. Guanylyl cyclase-C receptor mRNA distribution alongthe rat nephron. Regul Pept 95: 65–74, 2000.
11. Cha B, Kim JH, Hut H, Hogema BM, Nadarja J, Zizak M, Cavet M,Lee-Kwon W, Lohmann SM, Smolenski A, Tse CM, Yun C, de JongeHR, Donowitz M. cGMP inhibition of Na�/H� antiporter 3 (NHE3)requires PDZ domain adapter NHERF2, a broad specificity protein kinaseG-anchoring protein. J Biol Chem 280: 16642–16650, 2005.
12. Currie MG, Fok KF, Kato J, Moore RJ, Hamra FK, Duffin KL, SmithCE. Guanylin: an endogenous activator of intestinal guanylate cyclase.Proc Natl Acad Sci USA 89: 947–951, 1992.
13. Donowitz M, Li X. Regulatory binding partners and complexes of NHE3.Physiol Rev 87: 825–872, 2007.
14. Elitsur N, Lorenz JN, Hawkins JA, Rudolph JA, Witte D, Yang LE,McDonough AA, Cohen MB. The proximal convoluted tubule is a targetfor the uroguanylin-regulated natriuretic response. J Pediatr GastroenterolNutr 43, Suppl 1: S74–S81, 2006.
15. Fan X, Hamra FK, Freeman RH, Eber SL, Krause WJ, Lim RW, PaceVM, Currie MG, Forte LR. Uroguanylin: cloning of preprouroguanylincDNA, mRNA expression in the intestine and heart and isolation ofuroguanylin and prouroguanylin from plasma. Biochem Biophys ResCommun 219: 457–462, 1996.
16. Fawcus K, Gorton VJ, Lucas ML, McEwan GT. Stimulation of threedistinct guanylate cyclases induces mucosal surface alkalinisation in ratsmall intestine in vitro. Comp Biochem Physiol A Physiol 118: 291–295,1997.
17. Fonteles MC, Greenberg RN, Monteiro HS, Currie MG, Forte LR.Natriuretic and kaliuretic activities of guanylin and uroguanylin in theisolated perfused rat kidney. Am J Physiol Renal Physiol 275: F191–F197,1998.
18. Fonteles MC, Havt A, Prata RB, Prata PH, Monteiro HS, Lima AA,Jorge AR, Santos CF, Greenberg RN, Nascimento NR. High-salt intakeprimes the rat kidney to respond to a subthreshold uroguanylin dose duringex vivo renal perfusion. Regul Pept 158: 6–13, 2009.
19. Forte LR Jr. Uroguanylin: physiological role as a natriuretic hormone. JAm Soc Nephrol 16: 291–292, 2005.
20. Forte LR, London RM, Freeman RH, Krause WJ. Guanylin peptides:renal actions mediated by cyclic GMP. Am J Physiol Renal Physiol 278:F180–F191, 2000.
21. Forte LR, Thorne PK, Eber SL, Krause WJ, Freeman RH, FrancisSH, Corbin JD. Stimulation of intestinal Cl� transport by heat-stableenterotoxin: activation of cAMP-dependent protein kinase by cGMP. AmJ Physiol Cell Physiol 263: C607–C615, 1992.
22. Fukae H, Kinoshita H, Fujimoto S, Kita T, Nakazato M, Eto T.Changes in urinary levels and renal expression of uroguanylin on low orhigh salt diets in rats. Nephron 92: 373–378, 2002.
23. Gil FZ, Malnic G. Effect of amphotericin B on renal tubular acidificationin the rat. Pflügers Arch 413: 280–286, 1989.
24. Girardi AC, Di Sole F. Deciphering the mechanisms of the Na�/H�
exchanger-3 regulation in organ dysfunction. Am J Physiol Cell Physiol302: C1569–C1587, 2012.
25. Girardi AC, Knauf F, Demuth HU, Aronson PS. Role of dipeptidylpeptidase IV in regulating activity of Na�/H� exchanger isoform NHE3 inproximal tubule cells. Am J Physiol Cell Physiol 287: C1238–C1245,2004.
26. Greenberg RN, Hill M, Crytzer J, Krause WJ, Eber SL, Hamra FK,Forte LR. Comparison of effects of uroguanylin, guanylin, and Esche-richia coli heat-stable enterotoxin STa in mouse intestine and kidney:evidence that uroguanylin is an intestinal natriuretic hormone. J InvestigMed 45: 276–282, 1997.
27. Hamra FK, Forte LR, Eber SL, Pidhorodeckyj NV, Krause WJ,Freeman RH, Chin DT, Tompkins JA, Fok KF, Smith CE, Duffin KL,Siegel NR, Currie MG. Uroguanylin: structure and activity of a secondendogenous peptide that stimulates intestinal guanylate cyclase. Proc NatlAcad Sci USA 90: 10464–10468, 1993.
28. Hess R, Kuhn M, Schulz-Knappe P, Raida M, Fuchs M, Klodt J,Adermann K, Kaever V, Cetin Y, Forssmann WG. GCAP-II: isolationand characterization of the circulating form of human uroguanylin. FEBSLett 374: 34–38, 1995.
29. Jin XH, Siragy HM, Carey RM. Renal interstitial cGMP mediatesnatriuresis by direct tubule mechanism. Hypertension 38: 309–316, 2001.
30. Joo NS, London RM, Kim HD, Forte LR, Clarke LL. Regulation ofintestinal Cl� and HCO3
� secretion by uroguanylin. Am J Physiol Gastro-intest Liver Physiol 274: G633–G644, 1998.
31. Kocinsky HS, Dynia DW, Wang T, Aronson PS. NHE3 phosphorylationat serines 552 and 605 does not directly affect NHE3 activity. Am JPhysiol Renal Physiol 293: F212–F218, 2007.
32. Kocinsky HS, Girardi AC, Biemesderfer D, Nguyen T, Mentone S,Orlowski J, Aronson PS. Use of phospho-specific antibodies to deter-mine the phosphorylation of endogenous Na�/H� exchanger NHE3 atPKA consensus sites. Am J Physiol Renal Physiol 289: F249–F258, 2005.
33. Ledoussal C, Lorenz JN, Nieman ML, Soleimani M, Schultheis PJ,Shull GE. Renal salt wasting in mice lacking NHE3 Na�/H� exchangerbut not in mice lacking NHE2. Am J Physiol Renal Physiol 281: F718–F727, 2001.
34. Lennane RJ, Carey RM, Goodwin TJ, Peart WS. A comparison ofnatriuresis after oral and intravenous sodium loading in sodium-depletedman: evidence for a gastrointestinal or portal monitor of sodium intake.Clin Sci Mol Med 49: 437–440, 1975.
35. Lessa LM, Amorim JB, Fonteles MC, Malnic G. Effect of renoguanylinon hydrogen/bicarbonate ion transport in rat renal tubules. Regul Pept 157:37–43, 2009.
36. Lima AA, Monteiro HS, Fonteles MC. The effects of Escherichia coliheat-stable enterotoxin in renal sodium tubular transport. PharmacolToxicol 70: 163–167, 1992.
37. Lorenz JN, Nieman M, Sabo J, Sanford LP, Hawkins JA, Elitsur N,Gawenis LR, Clarke LL, Cohen MB. Uroguanylin knockout mice haveincreased blood pressure and impaired natriuretic response to enteral NaClload. J Clin Invest 112: 1244–1254, 2003.
38. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measure-ment with the Folin phenol reagent. J Biol Chem 193: 265–275, 1951.
39. McKee M, Scavone C, Nathanson JA. Nitric oxide, cGMP, and hormoneregulation of active sodium transport. Proc Natl Acad Sci USA 91:12056–12060, 1994.
F1407MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org

40. Mills JN. Human circadian rhythms. Physiol Rev 46: 128–171, 1966.41. Perkins A, Goy MF, Li Z. Uroguanylin is expressed by enterochromaffin
cells in the rat gastrointestinal tract. Gastroenterology 113: 1007–1014,1997.
42. Potthast R, Ehler E, Scheving LA, Sindic A, Schlatter E, Kuhn M.High salt intake increases uroguanylin expression in mouse kidney.Endocrinology 142: 3087–3097, 2001.
43. Preisig PA, Rector FC Jr. Role of Na�-H� antiport in rat proximaltubule NaCl absorption. Am J Physiol Renal Fluid Electrolyte Physiol 255:F461–F465, 1988.
44. Qian X, Moss NG, Fellner RC, Goy MF. Circulating prouroguanylin isprocessed to its active natriuretic form exclusively within the renaltubules. Endocrinology 149: 4499–4509, 2008.
45. Qian X, Moss NG, Fellner RC, Taylor-Blake B, Goy MF. The ratkidney contains high levels of prouroguanylin (the uroguanylin precursor)but does not express GC-C (the enteric uroguanylin receptor). Am JPhysiol Renal Physiol 300: F561–F573, 2011.
46. Roczniak A, Burns KD. Nitric oxide stimulates guanylate cyclase andregulates sodium transport in rabbit proximal tubule. Am J Physiol RenalFluid Electrolyte Physiol 270: F106–F115, 1996.
47. Schultheis PJ, Clarke LL, Meneton P, Miller ML, Soleimani M,Gawenis LR, Riddle TM, Duffy JJ, Doetschman T, Wang T, GiebischG, Aronson PS, Lorenz JN, Shull GE. Renal and intestinal absorptivedefects in mice lacking the NHE3 Na�/H� exchanger. Nat Genet 19:282–285, 1998.
48. Schwark JR, Jansen HW, Lang HJ, Krick W, Burckhardt G, HropotM. S3226, a novel inhibitor of Na�/H� exchanger subtype 3 in variouscell types. Pflügers Arch 436: 797–800, 1998.
49. Sindic A, Velic A, Basoglu C, Hirsch JR, Edemir B, Kuhn M,Schlatter E. Uroguanylin and guanylin regulate transport of mousecortical collecting duct independent of guanylate cyclase C. Kidney Int 68:1008–1017, 2005.
50. Sindice A, Basoglu C, Cerci A, Hirsch JR, Potthast R, Kuhn M,Ghanekar Y, Visweswariah SS, Schlatter E. Guanylin, uroguanylin, andheat-stable euterotoxin activate guanylate cyclase C and/or a pertussistoxin-sensitive G protein in human proximal tubule cells. J Biol Chem277: 17758–17764, 2002.
51. Wang T, Kawabata M, Haneda M, Takabatake T. Effects of urogua-nylin, an intestinal natriuretic peptide, on tubuloglomerular feedback.Hypertens Res 26: 577–582, 2003.
52. Wang T, Yang CL, Abbiati T, Schultheis PJ, Shull GE, Giebisch G,Aronson PS. Mechanism of proximal tubule bicarbonate absorption inNHE3 null mice. Am J Physiol Renal Physiol 277: F298–F302, 1999.
53. Yang LE, Sandberg MB, Can AD, Pihakaski-Maunsbach K, Mc-Donough AA. Effects of dietary salt on renal Na� transporter subcellulardistribution, abundance, and phosphorylation status. Am J Physiol RenalPhysiol 295: F1003–F1016, 2008.
54. Zhao H, Wiederkehr MR, Fan L, Collazo RL, Crowder LA, Moe OW.Acute inhibition of Na/H exchanger NHE-3 by cAMP. Role of proteinkinase a and NHE-3 phosphoserines 552 and 605. J Biol Chem 274:3978–3987, 1999.
F1408 MODULATION OF NHE3 BY UROGUANYLIN
AJP-Renal Physiol • doi:10.1152/ajprenal.00385.2011 • www.ajprenal.org




















