
Mathematical modeling of electromechanical function disturbances and recovery in calcium-overloaded...
10
F.B. Sachse and G. Seemann (Eds.): FIMH 2007, LNCS 4466, pp. 383–392, 2007. © Springer-Verlag Berlin Heidelberg 2007 Mathematical Modeling of Electromechanical Function Disturbances and Recovery in Calcium-Overloaded Cardiomyocytes Leonid B. Katsnelson, Tatiana Sulman, Olga Solovyova, and Vladimir S. Markhasin Institute of Immunology and Physiology, Ural Division of the Russian Academy of Sciences Pervomayskaya str. 91, Ekaterinburg 620041, Russia [email protected] Abstract. Rhythm disturbances and mechanical function suppression proper to the acute heart failure in the case of cardiomyocyte calcium overload are simulated in a mathematical model of cardiomyocyte electromechanical activity. Particular attention is paid to the overload caused by diminished activity of the Na + -K + pump. It is shown in the framework of the model that myocardium mechanics may promote arrhythmias in these conditions. In particular, cooperative influence of the attached crossbridges on the calcium- troponin kinetics is shown to contribute to the initiation of spontaneous action potentials. Numerical experiments showed that the recovery of the normal Na + - K + pump activity during the heart failure attack did not always led to the normal electromechanical function recovery in the failed cardiomyocyte. Alternative approaches were suggested in the model and compared to each other for recovery of the myocardium electrical and mechanical performance in the simulated case of the acute heart failure. Keywords: Active myocardium mechanics – Mechano-electric feedback – Heart rhythm – Extrasystole – Arrhythmia. 1 Introduction Rhythm disturbances are prevalent and potentially lethal complications of heart diseases. Given the complexity of intracellular mechanisms underlying arrhyth- mogenesis, detailed mathematical models can help to reveal causal chains of events and to identify possible targets for therapeutic interventions. Calcium overloading of cardiomyocytes is one of the principal factors inducing rhythm disturbances on the cellular level. For example, Ca 2+ accumulation in cardiomyocytes during heart failure is fraught with a risk of triggered activity and fibrillation, as a consequence of spontaneous Ca 2+ releases induced by sarcoplasmic reticulum (SR) overload [1, 2]. It is well known that Na + -K + pumping decrease may cause cardiomyocyte calcium overload [3]. Previous research has focused on mathematical modeling of rhythm disturbances caused by calcium overload [3-5], including the case of reduced Na + -K + pump activity [3]. Nevertheless, those studies dealt with the electrical activity of cardiomyocytes, while not considering simultaneous disturbances of the mechanical function and possible role of the mechano-electric feedback (MEF) in the modelled arrhythmias.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Mathematical modeling of electromechanical function disturbances and recovery in calcium-overloaded...

F.B. Sachse and G. Seemann (Eds.): FIMH 2007, LNCS 4466, pp. 383–392, 2007. © Springer-Verlag Berlin Heidelberg 2007
Mathematical Modeling of Electromechanical Function Disturbances and Recovery in Calcium-Overloaded
Cardiomyocytes
Leonid B. Katsnelson, Tatiana Sulman, Olga Solovyova, and Vladimir S. Markhasin
Institute of Immunology and Physiology, Ural Division of the Russian Academy of Sciences Pervomayskaya str. 91, Ekaterinburg 620041, Russia
Abstract. Rhythm disturbances and mechanical function suppression proper to the acute heart failure in the case of cardiomyocyte calcium overload are simulated in a mathematical model of cardiomyocyte electromechanical activity. Particular attention is paid to the overload caused by diminished activity of the Na+-K+ pump. It is shown in the framework of the model that myocardium mechanics may promote arrhythmias in these conditions. In particular, cooperative influence of the attached crossbridges on the calcium- troponin kinetics is shown to contribute to the initiation of spontaneous action potentials. Numerical experiments showed that the recovery of the normal Na+-K+ pump activity during the heart failure attack did not always led to the normal electromechanical function recovery in the failed cardiomyocyte. Alternative approaches were suggested in the model and compared to each other for recovery of the myocardium electrical and mechanical performance in the simulated case of the acute heart failure.
Keywords: Active myocardium mechanics – Mechano-electric feedback – Heart rhythm – Extrasystole – Arrhythmia.
1 Introduction
Rhythm disturbances are prevalent and potentially lethal complications of heart diseases. Given the complexity of intracellular mechanisms underlying arrhyth-mogenesis, detailed mathematical models can help to reveal causal chains of events and to identify possible targets for therapeutic interventions.
Calcium overloading of cardiomyocytes is one of the principal factors inducing rhythm disturbances on the cellular level. For example, Ca2+ accumulation in cardiomyocytes during heart failure is fraught with a risk of triggered activity and fibrillation, as a consequence of spontaneous Ca2+ releases induced by sarcoplasmic reticulum (SR) overload [1, 2]. It is well known that Na+-K+ pumping decrease may cause cardiomyocyte calcium overload [3]. Previous research has focused on mathematical modeling of rhythm disturbances caused by calcium overload [3-5], including the case of reduced Na+-K+ pump activity [3].
Nevertheless, those studies dealt with the electrical activity of cardiomyocytes, while not considering simultaneous disturbances of the mechanical function and possible role of the mechano-electric feedback (MEF) in the modelled arrhythmias.

384 L.B. Katsnelson et al.
We try to address the lack of such analysis with the help of our ‘Ekaterinburg-Oxford’ mathematical model developed earlier [6] and combining description of both the electrical and mechanical cardiomyocyte activity.
When Na+-K+ pump activity is reduced in the model, calcium overload developed gradually during a series of isometric contractions generated by regular pacing. We found that, if mechanics expelled from the model, only almost complete block of the Na+-K+ pump produced extrasystoles. This observation is fully consistent with the results of the simulation performed by Noble and Varghese [3]. However, effects of the mechanical activity of cardiomyocytes on both their Ca2+ handling and electrical activity proved in the ‘Ekaterinburg-Oxford’ model to increase significantly vulnerability for triggered activity in the case of moderately decreased Na+-K+ pump [7]. Now, using our mathematical model, we focus on the theoretical analysis of possible approaches to the recovery of the electromechanical function of the cardiomyocytes in these pathological conditions.
2 Cardiac MEF in the ‘Ekaterinburg-Oxford’ Mathematical Model
‘Ekaterinburg-Oxford’ mathematical model of mechano-electric interactions in ventricular cardiomyocytes has been developed earlier [6], which inherits the description of the electrical activity from the Noble’98 ventricular cell model [8] and the description of cellular Ca
2+ kinetics and mechanical activity from the Ekaterinburg
model family. Now we use the most recent version of the mechanical model described in detail elsewhere [9]. The combined model realistically simulates mechanical and electrical activity of cardiomyocytes during isometric and afterloaded contractions.
The model includes mechano-dependent cooperativity of the kinetics of Ca2+
- troponin C (CaTnC) binding. This is a principal link between cardiac mechanics and Ca
2+ handling in cardiomyocytes. Three types of experimentally-established
cooperativity of Ca2+
activation [10] are taken into account in the model [11]. Namely, the affinity of TnC for Ca
2+ increases with the increasing of: (i) concentration of
strongly-bound crossbridges (Xb) and (ii) concentration of CaTnC complexes; while, (iii) availability of actin sites for myosin heads increases due to the end-to-end interaction between tropomyosins.
These mechanisms underlie in the model a wide range of experimentally observed phenomena, such as the effects of mechanical load on muscle relaxation and mechano-Ca
2+ coupling (e.g. length- and load-dependence of Ca
2+ transients). In
addition, mechanical modulation of Ca2+
transients (due to mechano-dependent CaTnC kinetics) links through to mechanical and electrical activity, via Ca
2+-
dependent ionic currents. The model reproduces the effects of mechano-electrical coupling as an influence of muscle length on action potential (AP) duration during isometric contractions, the AP duration (APD) dependence on the muscle load during afterloaded contractions, and certain APD changes caused by passive deformations [6]. The cooperativity of the first type (Xb-CaTnC cooperativity) proves to be of

Mathematical Modeling of Electromechanical Function Disturbances and Recovery 385
special importance in this study. It is specified in the model by an exponential function
A(N )π in the differential equation describing CaTnC kinetics [9]:
2on tot i off A A
dA a (A A) [ Ca ] a exp(-k A) (N ) Adt
Here A is CaTnC concentration, AN is an average fraction of attached cross-bridges
per CaTnC complex. A(N )π decreases with the AN increase. In this work Ca2+ overload was reached in the model via attenuation of the Na+-K+
pump activity (other ways of the overload were also simulated and produced similar
results). The pump current pi is described in the model as follows [8].
o ip p
o m ,K i m ,Na
[ K ] [ Na ]ˆi i[ K ] K [ Na ] K
+ +
+ += ⋅ ⋅+ +
Here i[ Na ]+ and o[ K ]+ are concentrations of the intracellular sodium and
extracellular potassium respectively. m ,KK and m ,NaK are corresponding Michaelis
constants of the pump. Reduction of the pump activity was simulated with an increase in m ,NaK . The normal value of this parameter (Km,Na = 24.2 mM) [7] was used as a
reference magnitude in our study.
3 Role of MEF in the Arrhythmias Induced by Decreased Na+-K+ Pump Activity
We have earlier published elsewhere [7] our model analysis of the MEF contribution to the arrhythmias in cardiomyocytes overloaded with Са2+. Here we only summarize those results briefly as a background for describing approaches to the rhythm recovery suggested in Section 4.
Essentially new aspect of that our study [7], in addition to previous works of other authors [3-5, 12] was its focus on the mechanical contribution to inducing arrhythmias in conditions where the calcium overload of cardiomyocytes was not sufficiently large to cause the rhythm disturbances by itself. In particular, Figures 1 and 2 demonstrate an extrasystolic attack followed by the force alternans in a case of such moderate calcium overload.
We showed that the following mechanical factors proved to be potentially arrhythmogenic in cardiomyocytes moderately overloaded with Са2+:
- cross-bridge kinetics during the cardiomyocyte relaxation and effects of the kinetics on the CaTnC dissociation via the Xb-CaTnC cooperativity mechanism;
- cardiomyocyte length during contraction-relaxation cycles; - mechanical loads (as these influence both current lengths and contraction
velocities); - mechanical interaction of cardiomyocytes with different degree of Na+-K+ pump
inhibition.

386 L.B. Katsnelson et al.
We found in the model two mechanisms of the mechanical factor contribution to the arrhythmia in cardiomyocytes moderately overloaded with Са2+. Both mechanisms are analyzed in detail elsewhere [7]. Briefly they are as follows.
First, during any isometric/isotonic twitch following a regular pacing, there are dynamic cooperative interactions between Xb and CaTnC kinetics. The interactions support a high rate of the CaTnC decay during the final relaxation stage of the twitch, and thus provide the cytosol with a small surplus of [Са2+]i at the end of this stage. When cardiomyocytes are moderately overloaded with Са2+, the background diastolic level of the cytosol calcium is also higher than the normal one. Therefore the surplus turns out to provide a critical increment in the diastolic [Са2+]i to trigger spontaneous Са2+ release from the SR. The release, in its turn, activates inward depolarizing current via the forward mode of the Na+-Са2+ exchanger and the following extrasystolic AP. Thus, the described is a mechanism of the Xb-CaTnC-induced spontaneous AP generation.
Second, mechanical conditions (a decrease in the initial length and/or cardiomyocyte dynamic shortening) may intensify gradual (beat-to-beat) Са2+ storage in the SR due to the length dependence of the same Xb-CaTnC-cooperativity [7]. This mechanism may additionally contribute to the cell Са2+ overloading and thus promote overcoming the arrhythmogenic threshold of the SR calcium concentration.
Fig. 1. Contractions of a single virtual cardiomyocyte with moderately reduced activity of the Na+-K+ pump (P-sample; see the text, where the sample is defined). L=0.90 Lmax (Lmax corresponds to the sample length where it develops a maximum active isometric tension); pacing rate: 75 stimuli/m. The panel demonstrates active tension peaks (F – arbitrary units). The magnifying lens indicates the time interval represented in detail in Fig.2. The band under the panel indicates phases of the process where P-sample distinctly responds to the recovery of the normal Na+-K+ pump (see Sec. 4 for details). The data shows that, unlike the stimulation rate of 60 stimuli/min, where no rhythm disturbances observed in the P-sample, a faster pacing equal to 75 stimuli/min (or higher – not shown here) at decreased Na +-K+ pump activity resulted in the following events. A brief increase in the force amplitudes turned into a very gradual decrease that was suddenly disrupted by the drop in force accompanied by extrasystoles (see Fig. 2 for more detail). Then a transient force alternans arose (see also Fig. 2) transferring to steady-state contractions with very low force amplitudes.

Mathematical Modeling of Electromechanical Function Disturbances and Recovery 387
Fig. 2. Time course of the tension development (F), calcium transients ([Ca2+]i) and membrane potential (E) in the P-sample during 137 ÷ 160 s indicated with the magnifying lens in Fig.1. Scores under the time axis show the moments of the regular stimuli (each 0.8 s).
In general, these two mechanisms allow us to consider 3 characteristic ranges of Na+-K+ pump attenuation in respect with the mechanical contribution to the arrhythmogenesis. The ranges are as follows.
(1) Km,Na ≥ 110 mM (while the normal Km,Na is 24.2 mM) – extrasystoles arise independently of the mechanics.
(2) 40 mM ≤ Km,Na < 110 mM - extrasystoles also arise in any mechanical conditions, but only due to the above mechanism of the Xb-CaTnC-induced spontaneous AP generation. Figures 1, 2 demonstrate an example of the rhythm disturbances revealed in the model for Km,Na= 40 mM (where all the other parameter values are the same as in the norm). This virtual cardiomyocyte is named in our work as P-sample (pathological sample).
The behaviour of the P-sample shown in Fig. 1 and 2 reminds sudden force fall, arrhythmia and pulsus alternans observed during the acute heart failure development. Similar disturbances arose in the P-sample not only at the length equal to 0.90 Lmax represented here, but for any lengths ≤ 0.95 Lmax.
(3) 37 mM ≤ Km,Na < 40 mM - extrasystoles arise only due to the direct mechanical impacts (length or load diminution) via the mechanism of the mechanically modulated calcium overloading.
The latter range represents a border zone. When Km,Na lied below this zone, extrasystoles did not arise in the model in any mechanical conditions.

388 L.B. Katsnelson et al.
The first Na+-K+ pump inhibition range (Km,Na≥ 110 mM) has many things in common with findings obtained by Noble and Varghese [3], who studied effects of the almost blocked pump in the Earm-Hilgeman-Noble model of the cardiomyocyte ionic currents [13, 14]. In particular, they observed autorhythmic activity which arose in response to a single stimulus under strongly reduced Na+-K+ pump function. As they showed, the autorhythmic activity in that case resulted from the spontaneous Са2+ release from the SR that in turn activated inward current via the Na+-Са2+ exchanger. The same processes underlie rhythm disturbances in our simulations for Km,Na≥ 110 mM. Reduction of Na+-K+ pump activity in the second (40 mM ≤ Km,Na < 110 mM) and the third (37 mM ≤ Km,Na < 40 mM) ranges in our experiments was much more moderate than they simulated. Respectively, diastolic [Ca2+]i also increased much less and thus could not cause by itself the spontaneous Са2+ release from the SR without assistance of the additional MEF mechanisms we found in the model.
It is important that the predicted mechanism of the Xb-CaTnC-induced spontaneous AP generation may be verified in real experiments with a muscle sample. First, arrhythmia may be induced in the sample via a cardiac glycoside overdose. Then crossbridge attachment may be eliminated in the sample. This may be done using BDM (2,3-butanedione 2-monoxime) that prevents actomyosin formation [15]. The model predicts that spontaneous APs will vanish after BDM application. Moreover, the vanish is predicted to be valid for a significant range of cardiac glycoside concentrations inducing rhythm disturbances (corresponding to the found in the model range (2) of Na+-K+ pump attenuation: 40 mM ≤ Km,Na < 110 mM).
4 Arrhythmia Suppression and Contractile Function Recovery
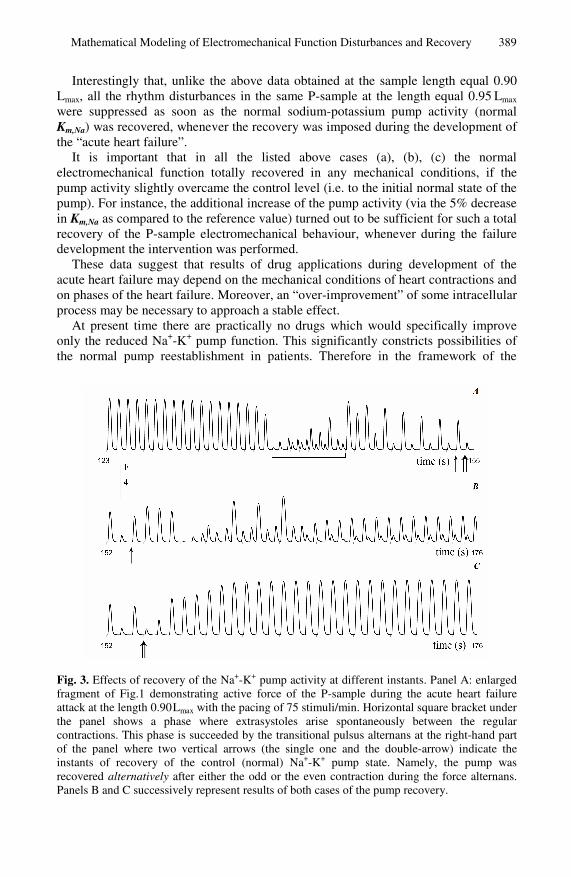
Here we use the model of the acute heart failure associated with the calcium overload to simulate suppression of the rhythm disturbances and restoration of stable contractions with amplitudes close to the normal ones. Figure 3 and Table 1 illustrate results of several “therapeutic” approaches applied to the P-sample.
First, it turned out to be quite unexpectedly that reestablishment of the normal Na+-K+ ATP-ase state did not always provide the recovery of the normal contractions. In this case the result substantially depended on the phases of the “acute heart failure” development, in which the reestablishment (i.e. the normal value of Km,Na) was done.
The band under the time axis in Fig. 1 shows such phases for the P-sample: white parts of the band indicate the phases where reestablishment of the normal Km,Na resulted in the total recovery of both the normal rhythm and normal contraction pattern. However, results of the normal Km,Na reestablishment was not so successful and stable during the shaded part of the band. Particularly, the reestablishment was tested (a) during the extrasystolic sub-phase indicated with the horizontal square bracket under Fig. 3; (b) after each low (odd) contraction during the transitional alternans sub-phase that succeeded extrasystolic one; (c) after each high (even) contraction during the alternans sub-phase. An example of (b) is indicated with the vertical arrow under Fig. 3A, and that of (c) – with the double-arrow. The tests showed that the normal electromechanical activity recovered totally only in the case (c) (see Fig. 3C). In the case (b) the reestablishment of the normal Na+-K+ pump resulted in one more transitional extrasystolic phase that passed into the steady-state alternans (Fig. 3B). The pattern similar to the latter one arose also in the case (a).
Mathematical Modeling of Electromechanical Function Disturbances and Recovery 389
Interestingly that, unlike the above data obtained at the sample length equal 0.90
Lmax, all the rhythm disturbances in the same P-sample at the length equal 0.95 Lmax were suppressed as soon as the normal sodium-potassium pump activity (normal Km,Na) was recovered, whenever the recovery was imposed during the development of the “acute heart failure”.
It is important that in all the listed above cases (a), (b), (c) the normal electromechanical function totally recovered in any mechanical conditions, if the pump activity slightly overcame the control level (i.e. to the initial normal state of the pump). For instance, the additional increase of the pump activity (via the 5% decrease in Km,Na as compared to the reference value) turned out to be sufficient for such a total recovery of the P-sample electromechanical behaviour, whenever during the failure development the intervention was performed.
These data suggest that results of drug applications during development of the acute heart failure may depend on the mechanical conditions of heart contractions and on phases of the heart failure. Moreover, an “over-improvement” of some intracellular process may be necessary to approach a stable effect.
At present time there are practically no drugs which would specifically improve only the reduced Na+-K+ pump function. This significantly constricts possibilities of the normal pump reestablishment in patients. Therefore in the framework of the
Fig. 3. Effects of recovery of the Na+-K+ pump activity at different instants. Panel A: enlarged fragment of Fig.1 demonstrating active force of the P-sample during the acute heart failure attack at the length 0.90 Lmax with the pacing of 75 stimuli/min. Horizontal square bracket under the panel shows a phase where extrasystoles arise spontaneously between the regular contractions. This phase is succeeded by the transitional pulsus alternans at the right-hand part of the panel where two vertical arrows (the single one and the double-arrow) indicate the instants of recovery of the control (normal) Na+-K+ pump state. Namely, the pump was recovered alternatively after either the odd or the even contraction during the force alternans. Panels B and C successively represent results of both cases of the pump recovery.

390 L.B. Katsnelson et al.
model we studied various alternative ways to modulate (amplify or attenuate) other intracellular mechanisms that may lead to some reduction of the cardiomyocyte Ca2+ loading and thus to the recovery of both the normal rhythm and satisfactory mechanical characteristics of the contractions without intensifying Na+-K+ pump activity. Particularly, such alternative interventions applied to the P-sample gave the following results.
A desired anti-arrhythmic effect resulted from a decrease in the rate of calcium uptake by the SR Ca2+ pump, even if the Na+-K+ pump function was not recovered. For example, 13% decrease in the rate of the SR calcium uptake suppressed rhythm disturbances in the P-sample at 0.95 Lmax, while extrasystoles still arose for the smaller cardiomyocyte lengths. Total ‘curing’ of the cardiomyocyte for all the lengths ensued from 18% SR Ca2+ uptake rate decrease. The mechanism of the ‘curing’ is as follows. Redistribution between SR Ca2+ uptake and Ca2+ removal from the cytosol via the Na+-Ca2+ exchange current shifted in favour of the Ca2+ removal from the cell due to the slowing down of the SR uptake. Thanks to this shift, the total level of Ca2+ in both the SR and cytosol remained lower than the arrhythmogenic thresholds.
Alternatively, the arrhythmia was suppressed with a decrease in the L-type Ca2+ current, iCaL, via respective decrease in the Ca2+ permeability of the L-type channels. For example, when it was decreased by 15%, extrasystoles vanished in the P-sample at 0.95 Lmax, but they still arose at the smaller lengths. The total ‘curing’ for all the lengths was observed with the 20% decrease in iCaL.
Antiarrhythmic effects were also obtained at all lengths in the case of the Na+-Ca2+ exchange acceleration (20% increase in its turnover rate as compared to the normal value) or of the 20% decrease in sensitivity of the SR calcium release to [Ca2+]i.
When the “antiarrhythmic treatment” using one of the above parameters was applied after the first extrasystole, the arrhythmia disappeared immediately. However, the time for the recovery of steady-state force amplitudes varied depending on each particular parameter change within the range of 100÷160 s.
Finally, the combined effect of simultaneous SR Ca pump activity attenuation and a decrease in the L-type Ca2+ current was considered. It is a case of a special importance, as in a real heart both effects may result simultaneously from the same drug application (β-blockers). It turned out that 11% simultaneous decrease in both parameters suppressed the rhythm disturbances at all lengths (compared to the ~20% decrease which was necessary in the case of the individual interventions aimed at either SR pumping decrease or iCaL decrease).
Table 1 demonstrates how the SR Ca2+ load ([Ca2+]SR) as well as 2 mechanical characteristics of contraction-relaxation (inotropic one: Fm – the peak active force during the steady-state contractions, and loositropic one: t30 – relaxation time to 30% of Fm) changed in comparison with the control due to the simulated interventions at 0.90 Lmax. The data for the table were obtained when the steady-state contractions set in after the respective intervention.
Summarizing the suggested “therapeutic” approaches, it may be concluded that all of them are aimed at the unloading of the cardiomyocytes overloaded with Ca2+. The interventions considered either promote the beat-to-beat calcium removal from the cell or slow down calcium beat-to-beat influx into the cytosol. All these methods of calcium unloading proved to be palliatives, because Ca2+ level in the SR in all these cases remained essentially higher than in norm. As applied to clinical practice, this

Mathematical Modeling of Electromechanical Function Disturbances and Recovery 391
Table 1. Results of different “treatments” of the P-sample
SR pumprate
18%
L-type Ca2+
current
20%
Na+-Ca2+
current
20%
2+ sensitivityof the SR 2+
release
20%
SR pump rate& L-type Ca2+
current
11% (both)Fm 88.5% 95.7% 99.2% 119.7% 94.4%t30 101.8% 104.1% 108.8% 138.2% 105.3%[Ca2+]SR 122.9% 122.7% 135.1% 134.2% 119.2%
The top row of the table indicates parameter changes as compared to the P-sample. Arrows mean an increase (↑) and a decrease (↓) in the parameter values. [Ca2+]SR, Fm and t30 are given as a percentage of the corresponding values in the normal cardiomyocyte.
might be interpreted as follows. If the prime cause of calcium overload is unavoidable, the drug application is to be used permanently. Drugs in this situation are, in a certain sense, molecular ‘prosthetic tools’ aimed to support impaired native mechanisms of calcium homeostasis. Our modeling confirms that β-blockers are highly effective drugs for the correction of cardiac electromechanical function, when cardiomyocytes are overloaded with calcium.
[Ca2+]SR values in the table reveal that each particular intervention partially relieved SR of Ca2+ as compared with the threshold [Ca2+]SR = 142% (of the norm) in the P-sample achieved just before the beginning of the “acute heart failure” attack.
5 Conclusion
A mathematical model of the acute heart failure development has been created. The model simulates cardiomyocyte electrical and mechanical functions and deals with the heart failure associated with the Ca2+ overload. The model suggests that both cardiomyocyte mechanical properties (crossbridge attachment/detachment kinetics that in turn influence CaTnC kinetics) and mechanical conditions of contractions (length, load) may significantly affect a risk of the acute heart failure attack in cardiomyocytes moderately overloaded with Ca2+.
We analyzed in the model a few alternative “therapeutic” approaches aimed to decrease calcium loading of cardiomyocytes, and thus to prevent from the “heart failure” development. The model predicts that simultaneous decrease in both L-type Ca2+ current and SR Ca2+ pumping seems to be the most relevant ‘therapy’ among the analyzed approaches. Just similar double action is typical for the effect of β-blockers.
The model shows that response of a cardiomyocyte to a prompt change of its state during alternating contractions may differ principally depending on which particular contraction (even or odd) precedes the impact. In the modelled case the impact was an instant recovery of the normal Na+-K+ pump activity. Meantime, there are real experimental data [16] revealing that another impact (a single extrastimulus) led to altering restitution curves depending on even or odd beat preceded the extrastimulus. It might be hypothesized that some intrinsic mechanisms underlie different cases of such altering responses. This hypothesis may be a subject of a future study.

392 L.B. Katsnelson et al.
Acknowledgments. This work is subject of a Wellcome Trust Research Development Award # 074152/Z/04/Z, President Grant for Leading Scientific Schools #4923.2006.4, The Fogarty International Center # 1 RO3 TW006250-01a1, and of a Russian Foundation for Basic Research grant # 05-04-48352.
References
1. Kihara, Y., Morgan, J.P.: Intracellular calcium and ventricular fibrillation. Studies in the aequorin-loaded isovolumic ferret heart. Circ Res. 68.5, 1378–1389 (1991)
2. Lakatta, E.G., Guarnieri, T.: Spontaneous myocardial calcium oscillations: are they linked to ventricular fibrillation? J. Cardiovasc Electrophysiol 4.4, 473–489 (1993)
3. Noble, D., Varghese, A.: Modelling of sodium-overload arrhythmias and their suppression. Can. J. Cardiol 14.1, 97–100 (1998)
4. Luo, C.H., Rudy, Y.: A dynamic model of the cardiac ventricular action potential. II. Afterdepolarizations, triggered activity, and potentiation. Circ Res. 74.6, 1097–1113 (1994)
5. Noble, D.: Modeling the heart–from genes to cells to the whole organ. Science 295.5560, 1678–1682 (2002)
6. Solovyova, O., Vikulova, N., Katsnelson, L.B., Markhasin, V.S., Noble, P.J., Garny, A.F., Kohl, P., Noble, D.: Mechanical interaction of heterogeneous cardiac muscle segments in silico: effects on Ca2+ handling and action potential. International Journal of Bifurcation and Chaos 13.12, 3757–3782 (2003)
7. Katsnelson, L.B., Solovyova, O., Sulman, T., Konovalov, P., Markhasin, V.S.: Modeling cardiomyocyte mechano-electric coupling in norm and pathology. Biophysics 51.6, 917–926 (2006)
8. Noble, D., Varghese, A., Kohl, P., Noble, P.: Improved guinea-pig ventricular cell model incorporating a diadic space, IKr and IKs, and length- and tension-dependent processes. Can. J. Cardiol 1, 123–134 (1998)
9. Katsnelson, L.B., Nikitina, L.V., Chemla, D., Solovyova, O., Coirault, C., Lecarpentier, Y., Markhasin, V.S.: Influence of viscosity on myocardium mechanical activity: a mathematical model. J. Theor Biol 230.3, 385–405 (2004)
10. Gordon, A.M., Regnier, M., Homsher, E.: Skeletal and Cardiac Muscle Contractile Activation: Tropomyosin “Rocks and Rolls”. News Physiol. Sci. 16, 49–55 (2001)
11. Katsnelson, L.B., Markhasin, V.S.: Mathematical modeling of relations between the kinetics of free intracellular calcium and mechanical function of myocardium. J. Mol. Cell Cardiol 28.3, 475–486 (1996)
12. DiFrancesco, D., Noble, D.: A model of cardiac electrical activity incorporating ionic pumps and concentration changes. Philos Trans R. Soc. Lond B. Biol Sci. 307.1133, 353–398 (1985)
13. Earm, Y.E., Noble, D.: A model of the single atrial cell: relation between calcium current and calcium release. Proc R. Soc. Lond B. Biol Sci. 240.1297, 83–96 (1990)
14. Hilgemann, D.W., Noble, D.: Excitation-contraction coupling and extracellular calcium transients in rabbit atrium: reconstruction of basic cellular mechanisms. Proc R. Soc. Lond B. Biol Sci. 230.1259, 163–205 (1987)
15. Mulieri, L.A., Alpert, N.R.: Differential effects of BDM on activation and contraction. Biophys J. 45, 47a (1984)
16. Pastore, J.M., Laurita, K.R., Rosenbaum, D.S.: Importance of spatiotemporal heterogeneity of cellular restitution in mechanism of arrhythmogenic discordant alternans. Heart Rhythm 3.6, 711–719 (2006)




















