
Efficient Bulk Deletes for Multi Dimensionally Clustered Tables in DB2

Journal of Molecular Neuroscience 211 Volume 28, 2006
*Author to whom all correspondence and reprint requests should be addressed. E-mail: [email protected]
BackgroundBasic helix–loop–helix (bHLH) transcription fac-
tors regulate a variety of fundamental biologicalprocesses in organisms ranging from yeast to man(Massari and Murre, 2000). One of the family mem-bers, Math1, is essential for the generation of variouscell types that are missing in Math1 knockout mice.Math1-dependent populations include cerebellargranule neurons (Ben-Arie et al., 1997), D1 spinal cordinterneurons (Bermingham et al., 2001; Helms andJohnson, 1998), inner ear hair cells (Bermingham etal., 1999), and intestinal secretory cells (Yang et al.,2001). Moreover, abnormal levels ofHath1, the human
Journal of Molecular NeuroscienceCopyright © 2006 Humana Press Inc.All rights of any nature whatsoever are reserved.ISSN0895-8696/06/28:211–230/$30.00JMN (Online)ISSN 1559-1166DOI 10.1385/JMN/28:02:211
ORIGINAL ARTICLE
ortholog of Math1 (Ben-Arie et al., 1996) were identi-fied in various diseases such as colon adenocarcino-mas (Leow et al., 2004), Merkel cell cutaneouscarcinoma (Leonard et al., 2002), and medulloblas-toma (Lee et al., 2003; Salsano et al., 2004; Tong et al.,2003). Interestingly, overexpression of Hath1 in theinner ear leads to a generation of new hair cells in thecochlea and organ of corti and improves the hearingin deaf mammals (Izumikawa et al., 2005; Kawamotoet al., 2003; Shou et al., 2003; Zheng and Gao, 2000).Despite its importance in development and relevanceto human disease, direct target genes through whichMath1/Hath1 act are poorly recognized.
Math1 Target Genes Are Enriched With Evolutionarily Conserved Clustered E-box Binding Sites
Valery Krizhanovsky, Lilach Soreq, Vitaly Kliminski, and Nissim Ben-Arie*
Department of Cell and Animal Biology, Institute of Life Sciences, The Hebrew University of Jerusalem, Jerusalem, Israel
Abstract
The basic helix–loop–helix (bHLH) transcription factor Math1 and its orthologs are fundamental for properdevelopment of various neuronal subpopulations, such as cerebellar granule cells, D1 interneurons in the spinalcord, and inner ear hair cells. Although crucial for neurogenesis, the mechanisms by which Math1 specificallyrecognizes its direct targets are not fully understood. To search for direct and indirect target genes and signal-ing pathways controlled by Math1, we analyzed the effect of Math1 knockout on the expression profile of mul-tiple genes in the embryonic cerebellum. Eighteen differentially expressed transcripts were identified and foundto belong to a few developmentally-related functional groups, such as transcriptional regulation, proliferation,organogenesis, signal transduction, and apoptosis. Importantly, genomic analysis of E-box motifs has identi-fied a significant enrichment and clustering of MATH1-binding E-boxes only in a subset of differentially expressedgenes (Nr2f6, Hras1, and Hes5) in both mouse and man. Moreover, Math1 was shown by chromatin immuno-precipitation (ChIP) to bind, and by a luciferase reporter assay to activate transcription, of an upstream genomicfragment of Nr2f6. Taken together, we propose that when putative direct targets of Math1 are being selected fordetailed studies on DNA microarray hybridization, the enrichment and clustering of binding E-boxes in multiple species may be helpful criteria. Our findings may be useful to the study of other bHLHtranscription factors, many of which control the development of the nervous system.
DOI 10.1385/JMN/28:02:211
Index Entries: bHLH transcription factor; Math1; E-box, cerebellum; cerebellar granule cells; target gene; Nr2f6.
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 211

212 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
Neurogenesis is regulated by a complicated net-work of molecular pathways, in which the bHLHtranscription factors are essential and maintain anevolutionarily conserved function (Atchley andFitch, 1997; Bertrand et al., 2002; Massari and Murre,2000). Most of the bHLH proteins underlying neuro-genesis, including MATH1, belong to class II (groupA), whose members are tissue-specific transcrip-tional activators (Atchley and Fitch, 1997; Jones, 2004;Massari and Murre, 2000; Murre et al., 1994). ClassII bHLH factors heterodimerize with a class I memberto bind an E-box core sequence (CANNTG), locatedin the direct target genes. The E-box configurationsCAGCTG and CAGGTG were found to be favoredby class II members, including MATH1 (Akazawaet al., 1995; Atchley and Fitch, 1997; Bertrand et al.,2002; Helms et al., 2000; Jones, 2004; Murre et al.,1994).
One cell population, which is the most affectedby the genomic ablation of Math1, is the cerebellargranule cells (CGCs) at the rhombic lip, which areunable to differentiate and give rise to the cerebel-lar external granule layer (Ben-Arie et al., 1997; Ben-Arie et al., 2000; Gazit et al., 2004). We hypothesizedthat genes acting downstream to Math1 regulate dif-ferentiation of the CGC progenitors and are impor-tant for understanding cerebellar maturation. Thus,expression profiling of early CGC progenitors inembryonic day (E)14.5 wild-type (WT) and Math1mutant animals was combined with large-scaleanalysis of genomic sequences. We aimed at usingbioinformatic tools to identify a shared signature,which may guide the identification of direct Math1target genes. As a first step towards this goal, weutilized a CDNA array with a limited number ofgenes that allows a closer inspection of the data asneeded in order to develop the methodology. Asubset of the 18 differentially expressed genes(DEGs) identified were found to contain clusters ofmultiple binding E-boxes present in both mouse andman. Moreover, one of these genes, Nr2f6, was foundto be expressed in the rhombic lip of wild type, butnot Math1-null mice; and MATH1 bound and acti-vated transcription of an E-box-containing genomicfragment of Nr2f6.
Materials and MethodsAnimals, Genotyping, and Embryonic Tissue
DissectionMath1-null mice were previously described (Ben-
Arie et al., 2000). Experiments were conducted
according to an ethical approval of IACUC of theHebrew University of Jerusalem, Israel.
Cerebella were collected at E14.5. Samples wereimmediately frozen and embryonic tails were col-lected and used for genotyping by Southern analysis(Ben-Arie et al., 1997) or real-time polymerase chainreaction (PCR) (Krizhanovsky et al., 2004). In paral-lel, the embryos were subjected to X-gal staining forconfirmative phenotyping (Ben-Arie et al., 2000).
Array Hybridization and Data AnalysisTotal RNA was isolated from a pool of 10 cere-
bella from each genotype, and 20 µg of total RNAfrom each genotype were treated by DNaseI and[32P]dATP-radiolabeled by reverse transcriptionusing Atlas Pure Total RNA Labeling System (Clon-tech, USA). The resulting probes were hybridized toAtlas 1.2 cDNA arrays (#7853-1, Clontech, USA).Autoradiographs were scanned, and the data ana-lyzed by AtlasImage 1.0 (Clontech, USA), using atwo-fold difference as a cutoff.
Reverse-Transcription PCRConfirmatory semi-quantitative gene-specific
reverse-transcription (RT)-PCR was performedusing commercial Atlas Array Custom Primers(Clontech, USA) according to the manufacturer’sinstructions. Total RNAwas independently extractedfrom WT and Math1-null cerebella by TriReagent(Sigma, USA), treated by DNaseI (Ambion, USA) and4 µg were reverse-transcribed by RevertAid H- FirstStrand Synthesis Kit (MBI Fermentas, Lithuania)using (dT)18 primers. PCR amplification was per-formed using FastStart Taq DNAPolymerase (Roche,Germany). PCR products were gel-separated, quan-tified by densitometry and the linear range of ampli-fication was detected for each gene and primer pair.
Real-time RT-PCR was performed in a Rotor-Genemachine (Corbett Research, Australia) in the pres-ence of SYBR Green I (Molecular Probes, USA). Singleproduct amplification and size were verified by melt-ing curve analysis and gel separation. Data analysiswas carried out by Rotor-Gene 5.0 software (CorbettResearch, Australia).
Identification and Analysis of GenomicSequencesSequences of the putative Math1 target genes and
flanking 5 kb on both sides were identified in mouseand human genome project databases of NationalCenters for Biotechnology Information (NCBI).Murine and human repeat sequences were masked
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 212

E-box Clustering in Math1 Target Genes 213
Journal of Molecular Neuroscience Volume 28, 2006
using RepeatMasker (http://www.repeatmasker.org)before being subjected to E-box analysis. K-meansalgorithm (Hartigan and Wong, 1979) was imple-mented in Matlab code (release 13, version 6.5) andis available upon request. The current Matlab ver-sion (Matlab 7.0.1) includes a native K-means func-tion under the statistics toolbox.
In Situ HybridizationHeads of E14 mouse embryos were fixed in 4%
paraformaldehyde (PFA), cryoprotected by 30%sucrose, and sectioned to 10-µm sagittal sections.Math1 genotype was determined by real-time PCR(Krizhanovsky et al., 2004). Probes were synthesizedusing DIG RNALabeling Kit (Roche, Germany) froma cDNAfragment of Nr2f6 (NM_010150, 584 to 1494).Hybridization and detection were performed asdescribed (Ma et al., 1996), and were followed byfixation and DAPI counterstaining. Pictures weretaken under an Axioskop2 microscope (Zeiss, Germany), using a DP10 digital camera (Olympus,Germany). Images were assembled using NationalInstitutes of Health (NIH) ImageJ software.
Luciferase AssayThe reporter plasmids were constructed by
cloning of mouse Nr2f6 5′-genomic sequences (accession NC_000074) of various lengths into pGL3-Basic luciferase reporter vector (Promega, USA).These plasmids were co-transfected with combina-tions of control plasmids, E47-expressing plasmid,Math1-expressing plasmid, and control Renillaluciferase pRL-null vector (Promega, USA) into HeLacells. The luciferase activity was assayed 36 h fol-lowing transfection by Dual-Luciferase ReporterAssay (Promega, USA). The relative luciferase activ-ity was calculated as the ratio of luciferase to Renillaluciferase activities.
Chromatin ImmunoprecipitationHEK293 cells were cotransfected with 5 µg of
pHM6-Math1 (entire open reading frame [ORF]cloned into pHM6 [Roche, Germany]) and 5 µgpcDNA3.1/Zeo-E47 (mouse ORF cloned intopcDNA3.1/Zeo [Invitrogen, USA], kindly providedby F. Guillemot). Cells were grown for 48 h and thenfixed with 1% PFA. Chromatin immunoprecipita-tion (ChIP) was performed using the ChIP-IT kit(#53001, ActiveMotif, USA) according to the manu-fecturer’s instructions. Briefly, following nuclearextraction, the chromatin was sonicated to approx-imate length of 300–500 bp and precleared.
Chromatin preparations were incubated (4°C, O/N)either with 2 µg of rat anti-HAantibody (3F10, Roche,Germany) or 1 µg of goat IgG (provided with the kit)and precipitated with Protein G-conjugated agarosebeads. The immunoprecipitates were subjected toprotein–DNA cross-link reversal (65°C, O/N), Pro-teinase K treatment (42°C, 1.5 h), and DNA elution.The purified ChIP products were PCR-analyzed uti-lizing the following primers flanking E-box con-taining regions upstream to hNr2f6 gene:
Pair A (-3405 to -3102) L: CACCAGGACTGGCTGAGTCT, R: GTGCTCGCCACCATACTTCT;
Pair B (-2710 to -2403) L: GGGGTAGAGCCTTGATCCAC, R: CACGTCACGTCACTCCACTC;
Pair C (-1777 to -1440) L: GTGCGCAGAAGGCACTGTC, R: GGCTGGACTGCAGATGTCTC;
Pair D (-1425 to -1137) L: CCCTTTCTGCAAAGCACAG, R: TGTGTTTGCTTCCTCTTCCA.
ResultsIdentification and Confirmation of Differentially
Expressed Genes in Cerebella of Math1-Nulland WT MiceCGC precursors are born in the rhombic lip, a tran-
sient embryonic structure at the roof of the fourthventricle of the developing brain (Alder et al., 1996).Following specification and proliferation, CGC pre-cursors migrate and give rise to the cerebellar exter-nal granule/germinal layer (EGL) (Alder et al., 1996).Postnatally, CGCs proliferate further, become post-mitotic, and migrate inward within the cerebellarcortex to give rise to the internal granule layer (IGL).Math1-null mice display a lack of EGL, although theCGC progenitors are properly generated and cor-rectly specified and localized at the rhombic lip (Ben-Arie et al., 1997; Ben-Arie et al., 2000; Gazit et al.,2004). To identify components of the transcriptionalnetwork underlying early CGC development, weperformed a comparison of the expression level ofabout 1200 genes, representing various functionalgroups, using Atlas Mouse 1.2 cDNA arrays. Thesewere probed with radioactively labeled cDNAs syn-thesized from Math1-null embryos and littermateWT cerebellar mRNA from E14.5. This develop-mental stage was selected for analysis because at thistime point, CGC progenitors are present in Math1-null embryos (Gazit et al., 2004).
Out of the 1176 cDNAs tested, 28 genes (2.4%)were found by array hybridization to be differ-entially expressed in WT and null cerebella. To confirm the differential expression of the 28 array-
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 213

214 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
identified genes we performed both semi-quantita-tive RT-PCR as well as quantitative real-time RT-PCR. A few batches of cerebellar RNA preparations,unrelated to the one used for the DNA array analy-sis, were used as templates. The housekeeping genesβ-actin and ornithin decarboxylase (Odc) served ascontrols to ensure that similar levels of cDNA werepresent in the starting material. As shown in Fig. 1and Table 1, out of 28 genes identified by arrayhybridization, 18 (61%) were also found to be dif-ferentially expressed by the two RT-PCR assays. Theexpression of the other nine genes (Vamp1, Bax, RGA,Dbx1, Src, WSB2, Penk1, Dag1, and Hsp84-1) was notsignificantly different between WT and Math1-nullcerebella as determined by RT-PCR. Gnb2, whichwas downregulated according to the arrayhybridization, was also excluded from the list,because it was the only gene tested, which wasrepeatedly downregulated in semi-quantitative RT-PCR but upregulated in real-time RT-PCR. Onlygenes that were found to be differentially expressedby array hybridization and both RT-PCR assays wereconsidered to be confirmed DEGs, and were sub-jected to further analysis.
Identity and Expression of the DifferentiallyExpressed GenesTo obtain a general view of the functional groups
to which the 18 DEGs belong, we employed the GeneOntology Consortium classification (Ashburner etal., 2000). All the identified DEGs belong to a smallnumber of functional categories: proliferation,growth regulation, transcriptional regulation, signaltransduction, and apoptosis (Fig. 2, Table 1). Thesecategories are directly related and relevant to cere-bellar development and to the known defects causedby the lack of Math1. In contrast, other functionalgroups widely represented in the array (e.g., ionchannels, cytoskeleton components, membrane pro-teins, and proteins involved in cell maintenance)were not represented by the DEGs.
The 12 down- and 6 upregulated genes wereunevenly distributed among the functional groups(Table 1). All DEGs classified as transcriptional reg-ulation, proliferation and apoptosis (n = 7) weredownregulated in Math1-null embryonic cerebella.This was not the case for functional categories likecell growth regulators and signal transduction mol-ecules (n = 10), which included similar numbers ofdown- and upregulated genes. These data supportedthe hypothesis that Math1 acts mainly as a tran-scriptional activator during cerebellar development.
Among the DEGs, there are genes already knownto play a major role during development by actingthrough well characterized signaling pathways (Table1): (a) Nr2f6 (v-erbA related proto-oncogene) and hair-less (hr) are regulators of thyroid hormone signaling(Potter et al., 2001; Yoh and Privalsky, 2001); (b) Fibrob-last growth factor (Fgf)15and insulin-like growth factor(Igf)2are growth factors (Holley et al., 2002; McWhirteret al., 1997); (c) hairy and enhancer-of-split (Hes)5 is aNotch signaling effector (Ohtsuka et al., 1999); (d)Refbp1 (Transcriptional coactivator of AML-1 & LEF-1)is an activator and dishevelled homolog (Dvl)3 is an
Fig. 1. Semi-quantitative reverse-transcription (RT)-polymerase chain reaction (PCR) confirmation of array-identified differentially expressed genes (DEGs). cDNAsfrom wild-type (WT) and Math1-null embryonic day (E)14.5cerebella were subjected to RT-PCR and the products quan-tified during the linear phase of amplification. Cytoplasmicβ-actin (Actin) and ornithin decarboxylase (Odc) were usedas control genes. The values (mean and SEM) represent theband intensity ratios in WT/Math1-null. The values origi-nate from at least three different experiments using RNAfrom two separate preparations, unrelated to that used forarray hybridization. The statistical significance of the dif-ferences in gene expression was evaluated by Student’s t-test (*p < 0.05, **p < 0.005).
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 214

Tabl
e 1
Iden
tity
and
Pro
pert
ies
of t
he D
iffe
rent
ially
Exp
ress
ed G
enes
in M
ath1
-Nul
l vs
Wild
-Typ
e C
ereb
ellu
m a
t E
mbr
yoni
c D
ay 1
4.5
WT
/G
ene
NC
BI
Mat
h1-
sym
bol
Nam
ege
ne I
Dnu
llFu
ncti
onE
xpre
ssio
n R
ef.
Tran
scri
pti
onal
reg
ula
tion
Hes
5H
airy
and
enh
ance
r of
sp
lit 5
1520
84.
77N
otch
eff
ecto
r in
V
ario
us
dev
elop
ing
tiss
ues
O
htsu
ka e
t al.,
neu
roge
nesi
sin
clu
din
g d
evel
opin
g ne
uro
ns19
99hr
Hai
rles
s15
460
4.36
Cor
epre
ssor
for
thyr
oid
B
rain
and
ski
nC
acho
n-G
onza
lez
horm
one
rece
pto
ret
al.,
199
9; P
otte
ret
al.,
200
1N
r2f6
Ear
-2; v
-erb
Are
late
d
1386
43.
24R
epre
ssor
of t
hyro
id
Var
iou
s ce
ll ty
pes
, dif
fere
ntia
ting
Yo
h an
d P
riva
lsky
, p
roto
-onc
ogen
eho
rmon
e re
cep
tor
P19
cel
ls20
01M
rg2
Hom
eobo
x m
yelo
id e
cotr
opic
17
537
3.38
Hin
dbr
ain
ante
rio-
pos
teri
or
Em
bryo
nic
hind
brai
n an
d
Dib
ner
et a
l., 2
001
vira
l int
egra
tion
sit
e-re
late
d
pat
tern
ing
spin
al c
ord
, som
e ad
ult
tiss
ues
2 (M
EIS
3)N
ucl
eic
acid
bin
din
g &
rel
ated
Ref
bp1
Tran
scri
pti
onal
coa
ctiv
ator
21
681
2.55
Coa
ctiv
ator
in W
nt p
athw
ayV
ario
us
cell
typ
esB
ruhn
et a
l., 1
997
of A
ML
-1 &
LE
F-1
(ALY
)Sh
fdg1
Split
han
d/
foot
del
eted
gen
e 1
2042
20.
37D
NA
bind
ing
regu
lato
rV
ario
us
tiss
ues
, als
o d
uri
ng th
eir
Cra
ckow
er e
t al.,
d
evel
opm
ent
1996
Pro
life
rati
onH
ras1
H-r
as p
roto
-onc
ogen
e;
1546
12.
11C
ell p
rolif
erat
ion
and
V
ario
us
cell
typ
esK
oera
et a
l., 1
997
tran
sfor
min
g p
rote
in p
21
dif
fere
ntia
tion
(HR
AS)
Cd
k4C
ell d
ivis
ion
pro
tein
kin
ase
412
567
2.67
Cel
l cyc
le p
rogr
essi
on a
t G1
Var
iou
s ce
ll ty
pes
, pro
lifer
atin
g Fe
rgu
son
et a
l.,
neu
rona
l pre
curs
ors
2000
Gro
wth
fac
tors
& o
rgan
ogen
esis
Gd
f1G
row
th/
dif
fere
ntia
tion
14
559
2.85
Rig
ht–l
eft p
atte
rnin
gE
arly
em
bryo
, em
bryo
nic
nerv
ous
Ran
kin
et a
l.,
fact
or 1
syst
em, s
ome
adu
lt ti
ssu
es20
00G
ap43
Neu
rom
odu
lin (a
xona
l 14
432
2.81
Neu
rite
ext
ensi
onD
iffe
rent
iati
ng n
euro
nsB
enow
itz
and
m
embr
ane
prot
ein
GA
P-43
)R
outt
enbe
rg,
1997
(Con
tinu
ed)
215
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 215

Tabl
e 1
(Con
tinu
ed)
WT
/G
ene
NC
BI
Mat
h1-
sym
bol
Nam
ege
ne I
Dnu
llFu
ncti
onE
xpre
ssio
n R
ef.
Nbl
1N
euro
blas
tom
a su
pp
ress
ion
1796
52.
45B
MP
sign
alin
g an
tago
nist
Dev
elop
ing
nerv
ous
syst
em a
nd
Dio
nne
et a
l.,
of tu
mor
igen
icit
y p
rote
in 1
bo
ne, o
ther
tiss
ues
2001
(DA
N)
Fgf1
5Fi
brob
last
gro
wth
fact
or 1
514
170
0.29
Gro
wth
fact
orE
mbr
yoni
c st
em c
ells
and
bra
inM
cWhi
rter
et a
l.,
1997
Igf2
Insu
lin-l
ike
grow
th fa
ctor
II
1600
20.
32G
row
th fa
ctor
, for
mat
ion
of
Ubi
quit
ousl
yH
olle
y et
al.,
200
2(I
GF-
II)
vari
ous
tiss
ues
Sig
nal
tran
sdu
ctio
n &
cel
l com
mu
nic
atio
nFa
bp7
Bra
in li
pid
-bin
din
g p
rote
in
1214
02.
58M
igra
tion
on
rad
ial g
liaR
adia
l glia
cel
ls, C
GC
in E
GL
Feng
et a
l., 1
994
(BL
BP
)P
rkar
1bcA
MP
-dep
end
ent p
rote
in
1908
50.
38cA
MP
dep
end
ent k
inas
eV
ario
us
cell
typ
es, m
ostl
y in
B
rand
on e
t al.,
ki
nase
typ
e I-
βre
gula
tory
ne
uro
ns, i
nclu
din
g C
GC
s 19
95ch
ain
Dvl
3Se
gmen
t pol
arit
y p
rote
in
1354
40.
38W
nt p
athw
ay e
ffec
tor
Wid
ely
exp
ress
ed in
em
bryo
and
Ts
ang
et a
l., 1
996
dis
heve
lled
hom
olog
3so
me
adu
lt ti
ssu
eG
rin1
Glu
tam
ate
(NM
DA
) rec
epto
r 14
810
0.44
NM
DA
rece
pto
rN
ervo
us
syst
em a
nd s
ome
adu
lt
Mas
kos
and
su
buni
t zet
a-1
(NR
1;
tiss
ues
McK
ay, 2
003
NM
D-R
1)A
pop
tosi
sD
ad1
Def
end
er a
gain
st c
ell d
eath
113
135
3.18
Ant
i-ap
opto
tic
agen
t, V
ario
us
cell
typ
es, u
pre
gula
ted
in
Hon
g et
al.,
200
0;
dev
elop
men
tally
ess
enti
ald
iffe
rent
iati
ng n
euro
nsSa
toh
and
K
uro
da,
200
0
BM
P, b
one
mor
pho
gene
tic
pro
tein
; EG
L, e
xter
nal g
ranu
le/
germ
inal
laye
r; C
EC
, cer
ebel
lar
gran
ule
cel
l.
216
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 216

E-box Clustering in Math1 Target Genes 217
Journal of Molecular Neuroscience Volume 28, 2006
effector in the Wnt signaling (Bruhn et al., 1997; Tsanget al., 1996); and (e) Growth/ differentiation factor(Gdf)1 and Nbl1 (Neuroblastoma suppression oftumorigenicity protein 1) are an agonist and an antag-onist,respectively, in the bone morphogenetic protein(BMP) pathway (Dionne et al., 2001; Rankin et al.,2000). Therefore, Math1 may regulate CGC develop-ment, directly or indirectly, not only through modu-lation of various cellular functions, but also by affectingdifferent signaling pathways.
To further relate the identified DEGs to cerebel-lar development, we took advantage of various exist-ing databases in which the DEGs are annotated bytheir function and expression (Table 1). Most of theidentified genes were previously shown to beexpressed in the embryonic nervous system and tocontrol neurodevelopment. Some of them areexpressed in specific neuronal subpopulationsduring development. This provides additional, albeitindirect, evidence for the physiological relevance ofthe results obtained by the array hybridization. Forexample, Hes5, Gdf1, and Mrg2 (myeloid ecotropicviral integration site-related gene 2/Meis3) displaya restricted expression pattern in the developingbrain (Dibner et al., 2001; Ohtsuka et al., 1999; Rankinet al., 2000), whereas Nbl1, Fgf15, and growth asso-
ciated protein (Gap)43/neuromodulin are expressedin a restricted manner in differentiating precursorsof various cells in the nervous system (Benowitz andRouttenberg, 1997; Dionne et al., 2001; McWhirteret al., 1997). Although annotations did not specifythe cerebellum, these DEGs could also be function-ally related to the developing CGC population.
Genomic Analysis of Binding Sites in Mouse and Human DEGsWe expected only some of the DEGs to be primary
targets of Math1, i.e., genes whose transcription isaffected by direct binding of MATH1 to their genomiccontrol elements. To distinguish between direct andindirect targets, genomic DNA sequences flankingthe DEGs were analyzed for the presence of MATH1binding sites. Like most bHLH transcription factors,MATH1 binds an E-box motif (CANNTG). More-over, in vitro assays identified that MATH1, like otherbHLH class II members, has a binding preferencefor CAGCTG and CAGGTG E-boxes (“binding E-boxes”), but not to CAAATG (“nonbinding E-box”) (Akazawa et al., 1995; Helms et al., 2000).Binding sites for bHLH transcription factors may bepresent upstream to the transcription start site (e.g.,in promoters), within the gene itself (e.g., in introns),
Fig. 2. Functional classification of the Math1-regultated differentially expressed genes (DEGs). DEGs were classifiedaccording to the Gene Ontology terms in relation to biological process (A) and molecular function (B). Figure was plot-ted by NetAffx analysis tool (www.affymetrix.com/netaffx), with a probe set count threshold of 5. Note that, based onthe Gene Ontology classification, a gene may be assigned to more than one term. The number of genes assigned to aterm is indicated in parenthesis, as well as the percentage of genes included in the category out of all genes tested.
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 217

218 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
or further downstream (e.g., in enhancers) (Cawleyet al., 2004). Therefore, for each of the DEGs, wetested a genomic window, which included 5 kbupstream of the transcription start site, the entiregene, and 5 kb downstream of the transcription stopsite. Three DEG genes (Grin1, Prkar1b, and Gap43)were not included in the analyses described below,because either their genomic sequences are currentlyunavailable for mouse and/or man or their genomicsequence is around 100 kb. Because the 15 analyzedgenes are of varying size, the E-box cumulative countwas normalized to 10 kb. The analysis strategy anddefinitions are schematically represented in Fig. 3A.
The combined density of the two binding E-boxesin and around the mouse DEGs ranged from 2.6 to23.1/10 kb (Fig. 3B, solid black bars, and Supple-mentary Fig. 1). This compares with 4.9/10 kbexpected from a random distribution model. Nega-tive control genes not regulated by bHLH factorscould not be selected for comparison, because theycould be targets of more than a hundred unrelatedclass II bHLH factors, and therefore contain the samecanonical binding E-boxes. Therefore, we devisedanother strategy for evaluating the statistical sig-nificance of our findings, and used as controls a non-binding E-box (CAAATG) and a scrambled sequenceof a binding E-box (TGGCCA). The density of thecontrol sequences ranged from 3.6 to 8.8/10 kb (Fig.3B, solid gray bars). To further test our findings, thedistribution of two additional control hexamers(CATTTG and GTAAAC) was examined and wasnot significantly different from a random distribu-tion (data not shown).
The density of the binding E-boxes ranged from2.6 to 23.1/10 kb. To distinguish between high andlow occurrences of the tested sequences, one mayuse an arbitrary cutoff value. Instead, we appliedthe rough K-means least-squares clustering algo-rithm (Hartigan and Wong, 1979). The goal of thisnonhierarchical algorithm is to partition a set ofobjects into a prespecified number of K clusters, such
that the within-group variance is minimized. To doso, K objects are selected at random from the popu-lation as initial centers. Then, data points are groupedto their closest center. A very common metric mea-sure of a distance used to assign an object to a clus-ter, which we implemented, is the sum of squaredEuclidean distances of each object from the center.Objects are added to their least distant cluster oneat a time until all components are grouped into clus-ters. Then, the position of the centroid (mean) of eachcluster is calculated. This entire process is repeatedfor as many random starts as the user specifies inorder to minimize the effect of randomly-selectedinitial objects as centers.
In our case, DEGs were classified as “low” or“high” with regard to the density of a sequence (e.g.,binding E-box) per 10 kb, and thus K = 2. To mini-mize the effect of randomly selected initial objects,we used 100 iterations of the entire procedure. TheK-means clustering analysis indicated that six mouseDEGs were more enriched in binding E-boxes (cen-troid = 16.1, range 11.5–23.1/10 kb, Fig. 3B, blackasterisks) than the nine other DEGs (centroid = 6.6,range 2.6–10.1/10 kb). The density of binding E-boxesin the enriched group was also higher than the den-sity of nonbinding hexamers and the one expectedfrom a random distribution model. The increasedappearances of binding E-boxes in only a subset ofthe DEGs and in higher values than nonbinding andother control sequences supports our hypothesis thatthis accumulation is not random in nature.
Binding E-boxes were previously shown to appearin multiple copies and in close proximity to oneanother in target genes of bHLH proteins in mam-mals (Chiaramello et al., 1996; Manohar et al., 2004;Massari and Murre, 2000) and other organisms(Gibert and Simpson, 2003; Roztocil et al., 1998). There-fore, we analyzed not only the density but also thedistribution of binding E-boxes around the DEGs (Fig.3B, striped bars). Indeed, in some of the DEGs (e.g., Dvl3, Nr2f6, and Hras1 [H-ras proto-oncogene;
Fig. 3. (Opposite page) High density and clustering of E-boxes in the genomic loci around the differentially expressed genes(DEGs). (A) Genomic sequences, including each DEG, 5 kb upstream and downstream to the transcription start (curved arrow)and stop (*) sites, respectively, were masked and analyzed for the presence of binding E-boxes (black) and control nonbind-ing and scrambled E-box sequences (gray). Solid bars represent the normalized density of E-boxes, and striped bars the clus-tered E-boxes (less than 500 b apart) in mouse (B) and man (C). The genes from each species were divided into two groups bythe K-Means least-squares clustering algorithm. Asterisks denote clustering into groups with a high level of binding E-boxes(black asterisks) or clustered binding E-boxes (gray asterisks). Nr2f6, Hras1, and Hes5 show cross-species conservation of bothhigh density and high clustering of binding E-boxes. (D) DEGs were assigned by the K-means algorithm into high (H) and low(L) clusters based on the combination of binding and clustering values (per 10 kb). The plot illustrated the two-dimensionalposition of each DEG from mouse (triangles) and man (squares). The names of DEGs that were scored as high in both speciesare shown. Centroids for mouse and human DEGs are indicated in each cluster as X and +, respectively.
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 218

E-box Clustering in Math1 Target Genes 219
Journal of Molecular Neuroscience Volume 28, 2006
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 219

220 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
transforming protein p21]), we identified regions ofclustered binding E-boxes (less than 500 bp apart).K-means analysis was used also here to divide thegenes into clusters containing high (centroid = 13.8,range 9.4–19.5/10 kb, Fig. 3B, black asterisks) or low(centroid = 4.5, range 0.0–8.5/10 kb) numbers of clus-tered binding E-boxes. Clustering level of controlsequences, nonbinding and scrambled E-boxes, waslower (Fig. 3B, striped bars). Notably, there was adirect correlation between high-density of bindingE-boxes (Fig. 3B, black asterisks) and increased clus-tering (Fig. 3B, gray asterisks), because five out ofthe six DEG were grouped into the high-level clus-ters for both density and clustering of binding E-boxes. It should be noted that clustering is not an
obligatory outcome of high density of appearance.In contrast, under a random distribution model,binding E-boxes are expected to be more evenlyspaced, which is not the case for genes such as Hras1(Supplementary Fig. 1).
Math1 and its human ortholog Hath1 probablycontrol overlapping target genes. With this in mind,we examined the evolutionary conservation of thehigh density of binding E-boxes and their tendencyto cluster. The human orthologs of the mouse DEGswere identified and (when available) analyzed. Likein mouse, human binding E-box densities alsovaried among DEGs, and therefore the DEGs weregrouped by K-means analysis, such that the highcentroid was 14.5 (range 11.0–18.8) and the low
Supplementary Fig. 1. Distribution of E-boxes in the genomic loci around differentially expressed genes (DEGs) inmouse. Genomic sequences of mouse DEGs, including 5 kb upstream and downstream to the transcription start andstop sites, are drawn to scale. MATH1-binding (black) and nonbinding (gray) sites are indicated by crossing lines. Thetranscription start and stop sites are indicated by rhombuses. The densities of total and clustered binding and nonbind-ing (black and gray, respectively) sequences per 10 kb are indicated.
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 220

centroid was 6.2 (range 2.1–9.2). As in mouse, thedensity of binding E-boxes in man (2.1–18.8/10 kb)was higher than the density of nonbindingsequences (3.0–9.3/10 kb). Analysis of the clusteredE-boxes split the DEGs such that the group of highlyclustered binding E-boxes was centered at 11.5(range 7.9–15.8) and the low was at 3.6 (range 0.0–6.6;green asterisks in Fig. 3C and Supplementary Fig.2 denote highly clustered DEGs). Because the onlyfunctionally validated direct target for MATH1 bind-ing is Math1 itself (Helms et al., 2000), we searchedfor E-box sequences also in Math1 and Hath1. Asexpected, high binding E-box density (9.9 and 9.0/10kb in mouse and man, respectively) and clusters ofbinding E-boxes were identified in the genomicsequences flanking the two orthologs.
We then looked for DEGs that show both highdensity and tendency to cluster binding E-boxes inboth mouse and man. To do so, we re-analyzed thedata by the K-means algorithm, but this time eachobject (DEG) was described by two parameters: thenumber of total and clustered binding E-boxes per10 kb. The centroids obtained after 100 iterations ofthe entire procedure were: mouse DEGs 17.0, 13.8and 7.1, 4.5, high and low centroids, respectively;and for the human DEGs 14.4, 11.4 and 6.2, 3.5. Tovisualize this, the data values were plotted (Fig. 3D),and the space describing each cluster was markedby a dashed line. Three DEGs (Nr2f6, Hras1, andHes5) were categorized into the “high” cluster inboth mouse and man. One of these genes was Hes5,which had already been proposed as a direct target
E-box Clustering in Math1 Target Genes 221
Journal of Molecular Neuroscience Volume 28, 2006
Supplementary Fig. 2. Distribution of E-boxes in the genomic loci around differentially expressed genes (DEGs) inman. Genomic sequences of mouse DEGs, including 5 kb upstream and downstream to the transcription start and stopsites, are drawn to scale. MATH1-binding (black) and nonbinding (gray) sites are indicated by crossing lines. The tran-scription start and stop sites are indicated by rhombuses. The densities of total and clustered binding and nonbinding(black and gray, respectively) sequences per 10 kb are indicated.
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 221

222 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
of Math1 (Gazit et al., 2004). An identical result wasobtained by examination of the overlap between theDEGs marked by asterisk in Fig. 3 B,C, when eachparameter (density and clustering) was used sepa-rately for clustering. This observation further veri-fied the correlation between density and clusteringthat was noted previously, indicating that when thenumber of E-boxes increased they did not spreadevenly. Taken together, our data revealed new fea-tures that are shared only by a subset of the DEGs,and therefore may be helpful in narrowing down thelist of DEGs taken for experimental validation. Itmight be that direct targets of Math1 display a highdensity of binding E-boxes that tend to cluster in thegenome, as can be detected in more than one species(e.g., a primate and a rodent).
Nr2f6 Expression is Specifically Downregulatedin Math1-Null Rhombic LipTo experimentally validate the notion obtained
by the bioinformatic and expression analysis, wechose as a proof-of-concept to further study Nr2f6.It was selected because it is one of the DEGs con-taining dense and clustered MATH1-binding E-boxes in mouse and man, and therefore could serveas a direct target of MATH1. Therefore, we tested ifexpression of Nr2f6 is specifically altered in CGCprecursors in the developing cerebellum of Math1-null embryos. In situ hybridization was performedon sagittal sections of E14.5 WT and Math1-null
embryos (Fig. 4). In Math1-null mice, Nr2f6 expres-sion decreased in the rhombic lip, but not in otherregions of the developing cerebellum, such as theventricular zone. In WT embryos, Nr2f6 wasexpressed also in the emerging EGL, which is miss-ing in Math1-null embryos. Therefore, althoughexpression of Nr2f6 may be activated also by othergenes, the specific decrease in CGC precursors ofMath1-null embryos implies that it is also regulatedby Math1, which is in agreement with data obtainedby the DNA array and RT-PCR analyses.
Math1 Binds and Activates Transcription of Nr2f6If Nr2f6 is placed directly downstream of Math1
in the signaling pathway, it is expected that its pro-moter will bind MATH1. To test this protein–DNAinteraction, we expressed MATH1-HAtogether withits dimerization partner E47 in HEK293 cells. Thisline was selected because it does not express HATH1as detected by RT-PCR (not shown). At first, we veri-fied that ChIP could identify interactions betweenMATH1-HA and two previously identified targets.MATH1 and HATH1 bind to their own 3′- enhancers,which are shown to be part of a positive autoregu-latory loop (Helms et al., 2000). Additionally, thebHLH factor HES5, which is downregulated inMath1-null cerebellum, was shown by elec-trophoretic mobility shift assay (EMSA) to containa binding E-box recognized by MATH1 (Gazit et al.,
Fig. 4. The expression of Nr2f6 is reduced in the RL of Math1-null embryos. The expression of Nr2f6 in the cerebel-lum (Cb) of wild-type and Math1-null embryonic day (E)14 embryos was evaluated by in situ hybridization. In wild-typeCb, the expression is evident in the external granule layer (EGL), rhombic lip (RL), and choroid plexus of the fourth ven-tricle (CP). In the Math1-null Cb, the EGL is missing and the expression of Nr2f6 is reduced in the RL but not in the CP.Scale bar: 100 µm.
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 222
E-box Clustering in Math1 Target Genes 223
Journal of Molecular Neuroscience Volume 28, 2006
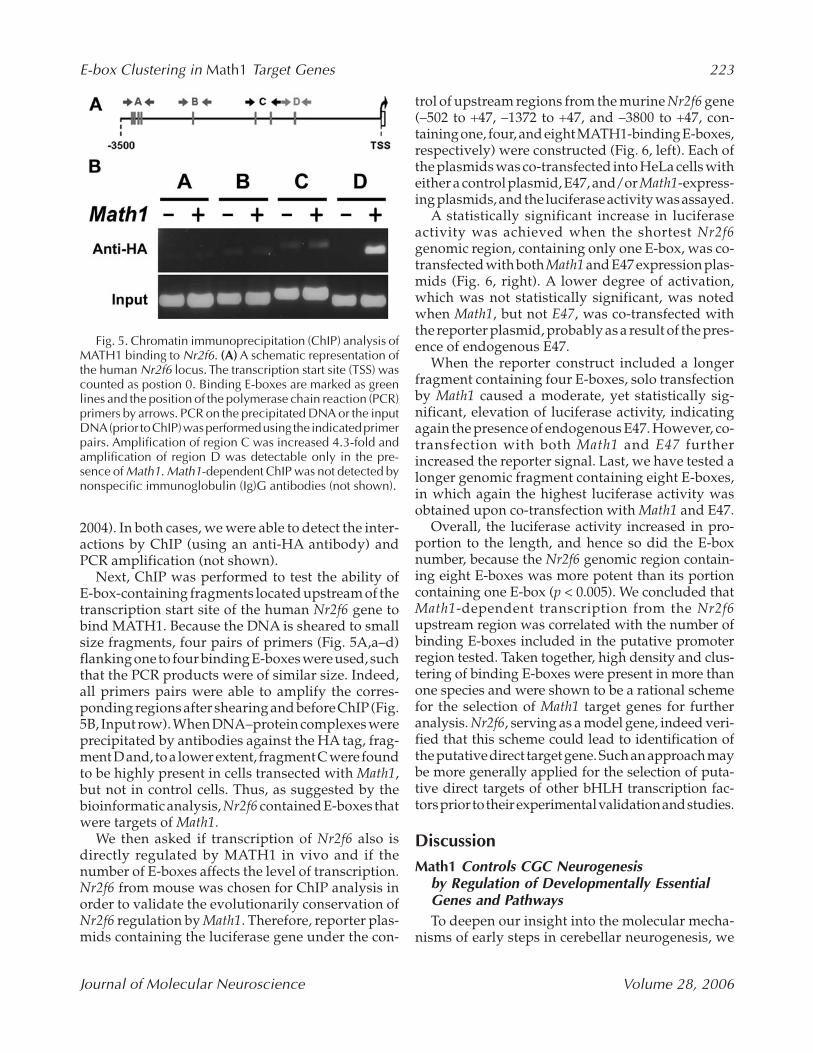
2004). In both cases, we were able to detect the inter-actions by ChIP (using an anti-HA antibody) andPCR amplification (not shown).
Next, ChIP was performed to test the ability of E-box-containing fragments located upstream of thetranscription start site of the human Nr2f6 gene tobind MATH1. Because the DNA is sheared to smallsize fragments, four pairs of primers (Fig. 5A,a–d)flanking one to four binding E-boxes were used, suchthat the PCR products were of similar size. Indeed,all primers pairs were able to amplify the corres-ponding regions after shearing and before ChIP(Fig.5B, Input row). When DNA–protein complexes wereprecipitated by antibodies against the HA tag, frag-ment D and, to a lower extent, fragment C were foundto be highly present in cells transected with Math1,but not in control cells. Thus, as suggested by thebioinformatic analysis, Nr2f6 contained E-boxes thatwere targets of Math1.
We then asked if transcription of Nr2f6 also isdirectly regulated by MATH1 in vivo and if thenumber of E-boxes affects the level of transcription.Nr2f6 from mouse was chosen for ChIP analysis inorder to validate the evolutionarily conservation ofNr2f6 regulation by Math1. Therefore, reporter plas-mids containing the luciferase gene under the con-
trol of upstream regions from the murine Nr2f6 gene(–502 to +47, –1372 to +47, and –3800 to +47, con-taining one, four, and eight MATH1-binding E-boxes,respectively) were constructed (Fig. 6, left). Each ofthe plasmids was co-transfected into HeLa cells witheither a control plasmid, E47, and/or Math1-express-ing plasmids, and the luciferase activity was assayed.
A statistically significant increase in luciferaseactivity was achieved when the shortest Nr2f6genomic region, containing only one E-box, was co-transfected with both Math1 and E47 expression plas-mids (Fig. 6, right). A lower degree of activation,which was not statistically significant, was notedwhen Math1, but not E47, was co-transfected withthe reporter plasmid, probably as a result of the pres-ence of endogenous E47.
When the reporter construct included a longerfragment containing four E-boxes, solo transfectionby Math1 caused a moderate, yet statistically sig-nificant, elevation of luciferase activity, indicatingagain the presence of endogenous E47. However, co-transfection with both Math1 and E47 furtherincreased the reporter signal. Last, we have tested alonger genomic fragment containing eight E-boxes,in which again the highest luciferase activity wasobtained upon co-transfection with Math1 and E47.
Overall, the luciferase activity increased in pro-portion to the length, and hence so did the E-boxnumber, because the Nr2f6 genomic region contain-ing eight E-boxes was more potent than its portioncontaining one E-box (p < 0.005). We concluded thatMath1-dependent transcription from the Nr2f6upstream region was correlated with the number ofbinding E-boxes included in the putative promoterregion tested. Taken together, high density and clus-tering of binding E-boxes were present in more thanone species and were shown to be a rational schemefor the selection of Math1 target genes for furtheranalysis. Nr2f6, serving as a model gene, indeed veri-fied that this scheme could lead to identification ofthe putative direct target gene. Such an approach maybe more generally applied for the selection of puta-tive direct targets of other bHLH transcription fac-tors prior to their experimental validation and studies.
DiscussionMath1 Controls CGC Neurogenesis
by Regulation of Developmentally EssentialGenes and PathwaysTo deepen our insight into the molecular mecha-
nisms of early steps in cerebellar neurogenesis, we
Fig. 5. Chromatin immunoprecipitation (ChIP) analysis ofMATH1 binding to Nr2f6. (A) A schematic representation ofthe human Nr2f6 locus. The transcription start site (TSS) wascounted as postion 0. Binding E-boxes are marked as greenlines and the position of the polymerase chain reaction (PCR)primers by arrows. PCR on the precipitated DNA or the inputDNA (prior to ChIP) was performed using the indicated primerpairs. Amplification of region C was increased 4.3-fold andamplification of region D was detectable only in the pre-sence of Math1. Math1-dependent ChIP was not detected bynonspecific immunoglobulin (Ig)G antibodies (not shown).
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 223

224 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
compared the expression level of 1200 genes innormal vs developmentally arrested CGC precur-sors from Math1-null mice. Eighteen DEGs wereidentified and validated, including 12 downregu-lated and 6 upregulated genes.
The E14.5 cerebellum contains neuronal precur-sors for different neuronal populations. The precur-sors of CGC are present at the rhombic lip, whereasPurkinje cells and neurons of deep cerebellar nucleiare born in a different ventricular zone (Hatten andHeintz, 1995). Math1 is expressed first in the rhom-bic lip, and later in migrated rhombic lip-born cells(Ben-Arie et al., 2000). Moreover, the primary effectof the Math1 knockout (agenesis of the EGL and thepontine nuclei) is cell-autonomous (Ben-Arie et al.,2000; Jensen et al., 2004). To learn whether the DEGsare expressed in CGC precursors, as is expected ofdirect targets of Math1, we searched GenePaint, ahigh-throughput RNAin situ hybridization database(Visel et al., 2004) with the 18 DEG. Images availablefor five genes (Nr2f6, Shfdg1, Igf1, and Grin1) indi-cated that they are expressed in developing cerebellaor adjacent tissues at E14.5 (head images are shownas Supplementary Fig. 3, with permission of Prof. G.Eichele). Moreover, the expression pattern of splithand/foot deleted gene (Shfdg)1 in the rhombic lip
and EGL matched exactly that of Math1, and theexpression domains of other genes (Nr2f6 and Fgf15)overlapped with that of Math1. Nr2f6 is expressed ina wider domain, which extends to the ventricularzone of the fourth ventricle. These data fully supportthe results of in situ hybridization analysis we haveperformed for Nr2f6. The localization of the DEGssupports our hypothesis that they act downstreamto Math1 and participate in cerebellar development.
The expression domain of two other DEGs, Igf2and glutamate receptor, ionotrophic, NMDA1, zeta1 (Grin1), was adjacent to, but did not overlap with,the expression domain of Math1. Grin1 is expressedin the ventricular zone of the fourth ventricle, wherePurkinje cells are born, and in the inner part of deve-loping cerebella. On the other hand, Igf2 is expressedin the meninges, covering the rhombic lip and EGL,which have functional molecular interactions withdeveloping CGCs (Zhu et al., 2004). Those genes areprobably not direct targets of Math1, and were regu-lated as a result of signaling between Math1-positiveCGC precursors and the cells in which they wereexpressed (Math1-negative). Such a mechanism ofindirect regulation is further supported by the obser-vation that, although Math1 is expressed only in CGCs,the Purkinje cell layer is abnormally positioned in
Fig. 6. The number of MATH1-binding E-boxes in Nr2f6 is correlated with promoter activity. Luciferase reporter plas-mids contained a fragment of the murine Nr2f6 promoter, increasing in the size and number of MATH1 binding E-boxes,respectively (as indicated on left side, E-boxes are marked as vertical lines and transcription start sites as empty boxes).Each reporter plasmid was co-transfected with a control plasmid, E47-expressing plasmid, Math1-expressing plasmid,or a combination of Math1- and E47-expressing plasmids. An elevation in luciferase activity correlated with the increasein the number of MATH1-binding E-boxes present in the Nr2f6 luciferase reporter in the presence of Math1 and E47 wasdetected. The values shown are net activity (after substruction of luciferase activity without inducers) and represent themean and SEM of at least four independent transfections. The statistical significance of the differences in gene expres-sion was evaluated by Student’s t-test (*p < 0.05, **p < 0.005).
JMN28_2_211_230_Arie 3/21/06 7:41 PM Page 224

E-box Clustering in Math1 Target Genes 225
Journal of Molecular Neuroscience Volume 28, 2006
Math1-null mice as a result of the lack of an EGL(Ben-Arie et al., 1997). It was recently shown that the EGLresponds to a diffusible attractant secreted by themeninges, which implies a reciprocal interplaybetween the two tissues (Zhu et al., 2004).
Six genes (Cdk4, Mrg2, Fabp7, Hes5, Hras1, andRefbp1) that were downregulated in Math1-nullembryonic cerebella display a time-dependentdecrease during normal postnatal cerebellar deve-lopment, whereas only one (hr) increases its expres-sion postnatally (Kagami and Furuichi, 2001). Hes5,a previously identified putative direct target of Math1(Gazit et al., 2004), was among the downregulatedgenes that showed a decline in expression duringdevelopment. Thus, the expression decline duringnormal postnatal development supported the notionthat some of the genes that were downregulated inMath1-null cerebella at E14.5 are important for normalearly cerebellar development and maturation.
Among all the functional groups represented inthe array, the identified DEGs belong to a few cate-gories: transcriptional regulators, nucleic acid bind-ing proteins, regulators of proliferation, growth andorganogenesis, and mediators of signal transduc-tion, and apoptosis. In contrast, genes regulated bythe bHLH factor MyoD also include other categories,such as metabolism and structural and cytoskeletalproteins (Bergstrom et al., 2002). These functions arephysiologically relevant to the developing cerebel-lum, because both proliferation and differentiationdefects of CGC were previously demonstrated inMath1 mutants (Ben-Arie et al., 1997; Gazit et al.,2004; Jensen et al., 2004).
The fact that the DEGs are members of varioussignaling pathways implies that Math1 controls CGCprecursor development, directly and indirectly, bymodulating multiple molecular pathways involvingthyroid hormone, growth factors, Notch, Wnt, and
Supplementary Fig. 3. Expression pattern of differentially expressed genes in the embryonic day 14.5 cerebellum. Insitu hybridization images of whole embryo sectioned sagittally with the indicated probes were retrieved from the Gene-Paint web site (www.genepaint.org [Visel et al., 2004]). Shown are the head images and enlargement of the cerebellarregions (inserts). Arrow = rhombic lip, arrowhead = external granule/germinal layer.
JMN28_2_211_230_Arie 3/21/06 7:42 PM Page 225

226 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
BMP. All of these mechanisms were identified ear-lier as essential for CGC precursor specification(Wang and Zoghbi, 2001). We found that they func-tion also during differentiation, and Math1 isinvolved in the cross talk between the different path-ways. In conclusion, we propose that Math1 controlsCGC development by modulating multiple molec-ular and cellular mechanisms, and that the devel-opmental arrest of CGC progenitors in Math1-nullembryos is caused by aberrant regulation of thosepathways. Future experiments will reveal thedetailed interplay between the different molecularmechanisms acting during CGC neurogenesis.
Math1 is essential not only for proper develop-ment of the CGC, but also of inner ear hair cells(Bermingham et al., 1999), D1 spinal cord neuronalprecursors (Bermingham et al., 2001), and secretorycells of the gut (Yang et al., 2001). Interestingly, path-ways similar to those affected in the cerebellum ofMath1-null mice were proposed to take part also inthe development of the other Math1-dependent tis-sues. Apoptosis was shown to be the cause of thehair cell loss in Math1-null inner ear (Chen et al.,2002). Notch signaling, which is essential for CGCdifferentiation (Gazit et al., 2004), is involved alsoin normal maturation of the inner ear hair cells (Lan-ford et al., 2000; Zine and de Ribaupierre, 2002) andsecretory cells of the intestine (Schroder and Gossler,2002; Yang et al., 2001). Similarly, Wnt and BMPpath-ways have a prominent role in the development ofthe cerebellum (Wang and Zoghbi, 2001), spinal cordinterneurons (Lee and Jessell, 1999; Muroyama et al.,2002), and intestine (Theodosiou and Tabin, 2003).The fact that some of the signaling pathways areshared among Math1-dependent cells or tissues fur-ther implies that Math1 regulates, at least partially,the same target genes in various tissues.
Frequent Appearance of Clustered Binding E-boxes is a Predictive Tool for the Selectionof bHLH Target GenesThe specificity of class II bHLH proteins is based
on binding to the canonical E-box motif CANNTG,which is further specified by the amino acid sequenceof the basic and helix regions (Bertrand et al., 2002;Nakada et al., 2004). Even when the level of degen-eracy is narrowed down (to CAGc/gTG), these pre-ferred E-box motifs are expected to be present every2048 bases under a random distribution model.Therefore, a single E-box sequence, although essen-tial, is clearly not sufficient by itself to determine thespecificity of DNA–protein interaction. An addi-
tional criterion, by which Math1-regulated genesmust contain dense and clustered binding E-boxes,may be employed. Support for an essential role ofhigh density and clustering of the binding E-boxescomes from the evolutionary conservation of thesefeatures in mouse and man, and from the high inci-dence of binding E-box clusters in Math1 and Hes5,genes shown previously to be recognized by MATH1(Gazit et al., 2004; Helms et al., 2000).
We have shown that MATH1 directly binds a puta-tive promoter region of Nr2f6, and that reporter activ-ity increased in proportion to the E-box numberpresent in upstream genomic region of Nr2f6, one ofthe genes demonstrating high density and cluster-ing of binding E-boxes. Nr2f6 (Ear2) is an orphannuclear receptor of the chicken ovalbumin upstreampromoter-transcription factor (COUP-TF) family(Avram et al., 1999). Nr2f6 is transiently expressedin locus coeruleus (LC) precursors when they stillreside in the rostral rhombic lip. Recently, it wasshown that Nr2f6 knockout mice lack >70% of LCneurons and that Nr2f6 acts downstream to the bHLHfactor Mash1 (Warnecke et al., 2005). Additionally,the expression of Math1 was not changed in E10.5Nr2f6-null embryos, supporting our findings thatNr2f6 acts directly downstream to Math1. Moreover,the dual regulation of Nr2f6, being a direct target ofboth Mash1 and Math1, provides a possible expla-nation for the existence of multiple clustered E-boxesin its control region.
During the last decade, several direct targets ofbHLH transcription factors were found in differentorganisms and their genomic regulatory regionsidentified (Atchley and Fitch, 1997; Bertrand et al.,2002; Gibert and Simpson, 2003; Hu et al., 2004; Massari and Murre, 2000). Also in these cases, E-boxes were found, and more than one E-box per clus-ter was required for correct recognition and controlover transcription activity (Chiaramello et al., 1996;Manohar et al., 2004; Roztocil et al., 1998; Seki et al.,2003). For example, an E-box cluster in Gap43 is rec-ognized by bHLH factors and regulates the expres-sion of a reporter (Chiaramello et al., 1996;Uittenbogaard et al., 2003). Furthermore, the bHLHfactors Cash1, NeuroM, and NeuroD were able to acti-vate expression of a reporter gene under an AchRα1regulatory element containing two CAGCTG E-boxes, but not the AChRβ3 element with only oneE-box (Roztocil et al., 1998). Clustered E-boxes wereidentified also in the NeuroD2 promoter, and two ofthem, E4 and E5, were shown to be functional (Linet al., 2004). Similarly, the bHLH c-Myc recognizes
JMN28_2_211_230_Arie 3/21/06 7:42 PM Page 226

E-box Clustering in Math1 Target Genes 227
Journal of Molecular Neuroscience Volume 28, 2006
several E-boxes in its target genes (Haggerty et al.,2003). It might be that, because the canonical E-boxsequence is short, there is need for clustering toenhance binding. We therefore propose that indeedthe presence of a binding E-box sequence is signifi-cant for target gene recognition and transcriptionregulation by a bHLH protein. In addition, the den-sity of the E-boxes and their clustering around thetarget genes in various species signals a functionalregulatory site. The mechanism of action of the mul-tiple clustered E-boxes is out of the scope of the cur-rent work, but was suggested to increase cell typediversity (Kageyama et al., 2005). Because not all E-boxes that activated transcription were found tobind Math1, various scenarios such as cooperativity,combinatorial binding of different bHLH factors,species-specificity, and cell-context activation arepossible and should be further studied. However,the phenomena of high density and clustering ofbinding E-boxes in multiple species may serve as a“flag” marking regulated genes that are good can-didates to serve as direct targets.
Our data indicated that three genes, Nr2f6, Hras1,and Hes5, fulfill the criteria of transcriptional regu-lation in Math1 knockout and evolutionary conser-vation of high density and clustering of bindingE-boxes, and are therefore putative direct targets ofMath1. These criteria can be applied for the identi-fication of candidate target genes for other bHLHtranscription factors.
AcknowledgmentsWe thank Esther Golenser and Theodora Bar-El
for their valuable assistance. We gratefully acknowl-edge Huda Y. Zoghbi (Houston, TX) for providingus with Math1 knockout mice and Gregor Eichele(Hannover, Germany) for the Nr2f6 probe. We thankH. Soreq, M. Devor, and A. Bar-Sinai for carefullyreading the manuscript. This work was supportedby grants from the Israel Science Foundation(587/02), US-Israel Binational Science Foundation(1998-066), and the European Community (QLG3-CT -2000-00072). N.B.A is a member of the RolandCenter for Neurodegenerative Diseases and the OttoLoewi Minerva Center for Cellular and MolecularNeurobiology, The Hebrew University of Jerusalem.
ReferencesAkazawa C., Ishibashi M., Shimizu C., Nakanishi S., and
Kageyama R. (1995) Amammalian helix-loop-helix factorstructurally related to the product of Drosophilaproneuralgene atonal is a positive transcriptional regulator
expressed in the developing nervous system. J. Biol.Chem. 270, 8730–8738.
Alder J., Cho N., and Hatten M. (1996) Embryonic pre-cursor cells from the rhombic lip are specified to a cere-bellar granule neuron identity. Neuron 17, 389–399.
Ashburner M., Ball C. A., Blake J. A., et al. (2000) Geneontology: tool for the unification of biology. The GeneOntology Consortium. Nat. Genet. 25, 25–29.
Atchley W. R. and Fitch W. M. (1997) A natural classifica-tion of the basic helix-loop-helix class of transcriptionfactors. Proc. Natl. Acad. Sci. U. S. A. 94, 5172–5176.
Avram D., Ishmael J. E., Nevrivy D. J., et al. (1999) Het-erodimeric interactions between chicken ovalbuminupstream promoter-transcription factor family mem-bers ARP1 and ear2. J. Biol. Chem. 274, 14,331–14,336.
Ben-Arie N., Bellen H. J., Armstrong D. L., et al. (1997)Math1 is essential for genesis of cerebellar granule neu-rons. Nature 390, 169–172.
Ben-Arie N., Hassan B. A., Bermingham N. A., et al. (2000)Functional conservation of atonal and Math1 in theCNS and PNS. Development 127, 1039–1048.
Ben-Arie N., McCall A. E., Berkman S., Eichele G., BellenH. J., and Zoghbi H. Y. (1996) Evolutionary conserva-tion of sequence and expression of the bHLH proteinAtonal suggests a conserved role in neurogenesis. Hum.Mol. Genet. 5, 1207–1216.
Benowitz L. I. and Routtenberg A. (1997) GAP-43: an intrin-sic determinant of neuronal development and plasti-city. Trends Neurosci. 20, 84–91.
Bergstrom D. A., Penn B. H., Strand A., Perry R. L., RudnickiM. A., and Tapscott S. J. (2002) Promoter-specific regu-lation of MyoD binding and signal transduction coop-erate to pattern gene expression. Mol. Cell 9, 587–600.
Bermingham N. A., Hassan B. A., Price S. D., et al. (1999)Math1: an essential gene for the generation of inner earhair cells. Science 284, 1837–1841.
Bermingham N. A., Hassan B. A., Wang V. Y., et al. (2001)Proprioceptor pathway development is dependent onMath1. Neuron 30, 411–422.
Bertrand N., Castro D. S., and Guillemot F. (2002) Proneuralgenes and the specification of neural cell types. Nat.Rev. Neurosci. 3, 517–530.
Brandon E. P., Zhuo M., Huang Y. Y., et al. (1995) Hip-pocampal long-term depression and depotentiation aredefective in mice carrying a targeted disruption of thegene encoding the RI beta subunit of cAMP-dependentprotein kinase. Proc. Natl. Acad. Sci. U. S. A. 92, 8851–8855.
Bruhn L., Munnerlyn A., and Grosschedl R. (1997) ALY, acontext-dependent coactivator of LEF-1 and AML-1, isrequired for TCRalpha enhancer function. Genes Dev.11, 640–653.
Cachon-Gonzalez M. B., San-Jose I., Cano A., et al. (1999)The hairless gene of the mouse: relationship of phe-notypic effects with expression profile and genotype.Dev. Dyn. 216, 113–126.
Cawley S., Bekiranov S., Ng H. H., et al. (2004) Unbiasedmapping of transcription factor binding sites alonghuman chromosomes 21 and 22 points to widespreadregulation of noncoding RNAs. Cell 116, 499–509.
JMN28_2_211_230_Arie 3/21/06 7:42 PM Page 227

228 Krizhanovsky et al.
Journal of Molecular Neuroscience Volume 28, 2006
Chen P., Johnson J. E., Zoghbi H. Y., and Segil N. (2002) Therole of Math1 in inner ear development: Uncoupling theestablishment of the sensory primordium from hair cellfate determination. Development 129, 2495–2505.
Chiaramello A., Neuman T., Peavy D. R., and Zuber M. X.(1996) The GAP-43 gene is a direct downstream targetof the basic Helix-Loop-Helix transcription factors. J. Biol. Chem. 271, 22,035–22,043.
Crackower M. A., Scherer S. W., Rommens J. M., et al.(1996) Characterization of the split hand/split foot mal-formation locus SHFM1 at 7q21.3-q22.1 and analysisof a candidate gene for its expression during limb devel-opment. Hum. Mol. Genet. 5, 571–579.
Dibner C., Elias S., and Frank D. (2001) XMeis3 proteinactivity is required for proper hindbrain patterningin Xenopus laevis embryos. Development 128 ,3415–3426.
Dionne M. S., Skarnes W. C., and Harland R. M. (2001)Mutation and analysis of Dan, the founding memberof the Dan family of transforming growth factor betaantagonists. Mol. Cell Biol. 21, 636–643.
Feng L., Hatten M. E., and Heintz N. (1994) Brain lipid-binding protein (BLBP): a novel signaling system in thedeveloping mammalian CNS. Neuron 12, 895–908.
Ferguson K. L., Callaghan S. M., O’Hare M. J., Park D. S.,and Slack R. S. (2000) The Rb-CDK4/6 signaling path-way is critical in neural precursor cell cycle regulation.J. Biol. Chem. 275, 33,593–33,600.
Gazit R., Krizhanovsky V., and Ben-Arie N. (2004) Math1controls cerebellar granule cell differentiation by reg-ulating multiple components of the Notch signalingpathway. Development 131, 903–913.
Gibert J. M. and Simpson P. (2003) Evolution of cis-regu-lation of the proneural genes. Int. J. Dev. Biol. 47, 643–651.
Haggerty T. J., Zeller K. I., Osthus R. C., Wonsey D. R.,and Dang C. V. (2003) A strategy for identifying tran-scription factor binding sites reveals two classes ofgenomic c-Myc target sites. Proc. Natl. Acad. Sci. U S A100, 5313–5318.
Hartigan J. A. and Wong M. A. (1979) A K-Means Clus-tering Algorithm. Applied Statistics 28, 100–108.
Hatten M. E. and Heintz N. (1995) Mechanisms of neuralpatterning and specification in the developing cere-bellum. Ann. Rev. Neurosci. 18, 385–408.
Helms A. W., Abney A. L., Ben-Arie N., Zoghbi H. Y., andJohnson J. E. (2000) Autoregulation and multipleenhancers control Math1 expression in the developingnervous system. Development 127, 1185–1196.
Helms A. W. and Johnson J. E. (1998) Progenitors of dorsalcommissural interneurons are defined by MATH1expression. Development 125, 919–928.
Holley S. J., Hall S. B., and Mellon P. L. (2002) Comple-mentary expression of IGF-II and IGFBP-5 during ante-rior pituitary development. Dev. Biol. 244, 319–328.
Hong N. A., Flannery M., Hsieh S. N., Cado D., PedersenR., and Winoto A. (2000) Mice lacking Dad1, thedefender against apoptotic death-1, express abnormalN-linked glycoproteins and undergo increased embry-onic apoptosis. Dev. Biol. 220, 76–84.
Hu Y., Wang T., Stormo G. D., and Gordon J. I. (2004) RNAinterference of achaete-scute homolog 1 in mouseprostate neuroendocrine cells reveals its gene targetsand DNA binding sites. Proc. Natl. Acad. Sci. USA 101,5559–5564.
Izumikawa M., Minoda R., Kawamoto K., et al. (2005)Auditory hair cell replacement and hearing improve-ment by Atoh1 gene therapy in deaf mammals. Nat.Med. 11, 271–276.
Jensen P., Smeyne R., and Goldowitz D. (2004) Analysisof cerebellar development in math1 null embryos andchimeras. J. Neurosci. 24, 2202–2211.
Jones S. (2004) An overview of the basic helix-loop-helixproteins. Genome Biol. 5, 226.
Kagami Y. and Furuichi T. (2001) Investigation of differ-entially expressed genes during the development ofmouse cerebellum. Gene Expression Patterns 1, 39–59.
Kageyama R., Ohtsuka T., Hatakeyama J., and Ohsawa R.(2005) Roles of bHLH genes in neural stem cell differ-entiation. Exp. Cell Res. 306, 343–348.
Kawamoto K., Ishimoto S., Minoda R., Brough D. E., andRaphael Y. (2003) Math1 gene transfer generates newcochlear hair cells in mature guinea pigs in vivo. J. Neu-rosci. 23, 4395–4400.
Koera K., Nakamura K., Nakao K., et al. (1997) K-ras isessential for the development of the mouse embryo.Oncogene 15, 1151–1159.
Krizhanovsky V., Golenser E., and Ben-Arie N. (2004)Genotype identification of Math1/LacZ knockout micebased on real-time PCR with SYBR Green I dye. J. Neu-rosci. Methods 136, 187–192.
Lanford P. J., Shailam R., Norton C. R., Gridley T., andKelley M. W. (2000) Expression of Math1 and HES5 inthe cochleae of wildtype and Jag2 mutant mice. J. Assoc.Res. Otolaryngol. 1, 161–171.
Lee K. J. and Jessell T. M. (1999) The specification of dorsalcell fates in the vertebrate central nervous system. Ann.Rev. Neurosci. 22, 261–294.
Lee Y., Miller H. L., Jensen P., et al. (2003) A molecular fin-gerprint for medulloblastoma. Cancer Res. 63,5428–5437.
Leonard J. H., Cook A. L., Van Gele M., et al. (2002)Proneural and proneuroendocrine transcription factorexpression in cutaneous mechanoreceptor (Merkel)cells and Merkel cell carcinoma. Int. J. Cancer 101,103–110.
Leow C. C., Romero M. S., Ross S., Polakis P., and Gao W. Q.(2004) Hath1, down-regulated in colon adenocarcino-mas, inhibits proliferation and tumorigenesis of coloncancer cells. Cancer Res. 64, 6050–6057.
Lin C. H., Stoeck J., Ravanpay A. C., Guillemot F., Tap-scott S. J., and Olson J. M. (2004) Regulation of neuroD2expression in mouse brain. Dev. Biol. 265, 234–245.
Ma Q., Kintner C., and Anderson D. J. (1996) Identifica-tion of neurogenin, a vertebrate neuronal determina-tion gene. Cell 87, 43–52.
Manohar C. F., Bray J. A., Salwen H. R., et al. (2004) MYCN-mediated regulation of the MRP1 promoter in humanneuroblastoma. Oncogene 23, 753–762.
JMN28_2_211_230_Arie 3/21/06 7:42 PM Page 228

E-box Clustering in Math1 Target Genes 229
Journal of Molecular Neuroscience Volume 28, 2006
Maskos U. and McKay R. D. (2003) Neural cells withoutfunctional N-Methyl-D-Aspartate (NMDA) receptorscontribute extensively to normal postnatal braindevelopment in efficiently generated chimaeric NMDA R1 −/− <—> +/+ mice. Dev. Biol. 262, 119–136.
Massari M. E. and Murre C. (2000) Helix-loop-helix pro-teins: regulators of transcription in eucaryotic organ-isms. Mol. Cell Biol. 20, 429–440.
McWhirter J. R., Goulding M., Weiner J. A., Chun J., andMurre C. (1997) A novel fibroblast growth factor geneexpressed in the developing nervous system is a down-stream target of the chimeric homeodomain oncopro-tein E2A-Pbx1. Development 124, 3221–3232.
Muroyama Y., Fujihara M., Ikeya M., Kondoh H., andTakada S. (2002) Wnt signaling plays an essential rolein neuronal specification of the dorsal spinal cord. GenesDev. 16, 548–553.
Murre C., Bain G., van Dijk M. A., et al. (1994) Structureand function of helix-loop-helix proteins. Biochem. Bio-phys. Acta 1218, 129–135.
Nakada Y., Hunsaker T. L., Henke R. M., and Johnson J. E.(2004) Distinct domains within Mash1 and Math1 arerequired for function in neuronal differentiation versusneuronal cell-type specification. Development 131,1319–1330.
Ohtsuka T., Ishibashi M., Gradwohl G., Nakanishi S.,Guillemot F., and Kageyama R. (1999) Hes1 and Hes5as notch effectors in mammalian neuronal differentia-tion. EMBO J. 18, 2196–2207.
Potter G. B., Beaudoin G. M., 3rd, DeRenzo C. L., ZarachJ. M., Chen S. H., and Thompson C. C. (2001) The hair-less gene mutated in congenital hair loss disordersencodes a novel nuclear receptor corepressor. GenesDev. 15, 2687–2701.
Rankin C. T., Bunton T., Lawler A. M., and Lee S. J. (2000)Regulation of left-right patterning in mice bygrowth/differentiation factor-1. Nat. Genet. 24,262–265.
Roztocil T., Matter-Sadzinski L., Gomez M., Ballivet M.,and Matter J. M. (1998) Functional properties of theneuronal nicotinic acetylcholine receptor beta3 pro-moter in the developing central nervous system. J. Biol.Chem. 273, 15,131–15,137.
Salsano E., Pollo B., Eoli M., Giordana M. T., and Finocchiaro G. (2004) Expression of MATH1, a markerof cerebellar granule cell progenitors, identifies differ-ent medulloblastoma sub-types. Neurosci. Lett. 370,180–185.
Satoh J. and Kuroda Y. (2000) Differential gene expres-sion between human neurons and neuronal progeni-tor cells in culture: an analysis of arrayed cDNAclones in NTera2 human embryonal carcinoma cell line as a model system. J. Neurosci. Methods 94, 155–164.
Schroder N. and Gossler A. (2002) Expression of Notchpathway components in fetal and adult mouse smallintestine. Gene Expr. Patterns 2, 247–250.
Seki K., Fujimori T., Savagner P., et al. (2003) Mouse Snailfamily transcription repressors regulate chondrocyte,extracellular matrix, type II collagen, and aggrecan. J. Biol. Chem. 278, 41,862–41,870.
Shou J., Zheng J. L., and Gao W. Q. (2003) Robust gener-ation of new hair cells in the mature mammalian innerear by adenoviral expression of Hath1. Mol. Cell Neu-rosci. 23, 169–179.
Theodosiou N. A. and Tabin C. J. (2003) Wnt signalingduring development of the gastrointestinal tract. Dev.Biol. 259, 258–271.
Tong W.-M., Ohgaki H., Huang H., Granier C., KleihuesP., and Wang Z.-Q. (2003) Null Mutation of DNAStrandBreak-Binding Molecule Poly(ADP-ribose) PolymeraseCauses Medulloblastomas in p53-/- Mice. Am. J. Pathol.162, 343–352.
Tsang M., Lijam N., Yang Y., Beier D. R., Wynshaw-BorisA., and Sussman D. J. (1996) Isolation and characteri-zation of mouse dishevelled-3. Dev. Dyn. 207, 253–262.
Uittenbogaard M., Martinka D. L., and Chiaramello A.(2003) The basic helix-loop-helix differentiation factorNex1/MATH-2 functions as a key activator of the GAP-43 gene. J. Neurochem. 84, 678–688.
Visel A., Thaller C., and Eichele G. (2004) GenePaint.org:an atlas of gene expression patterns in the mouseembryo. Nucleic Acids Res. 32, D552–556.
Wang V. Y. and Zoghbi H. Y. (2001) Genetic regulation ofcerebellar development. Nat. Rev. Neurosci. 2, 484–491.
Warnecke M., Oster H., Revelli J. P., Alvarez-Bolado G.,and Eichele G. (2005) Abnormal development of thelocus coeruleus in Ear2 (Nr2f6)-deficient mice impairsthe functionality of the forebrain clock and affects noci-ception. Genes Dev. 19, 614–625.
Yang Q., Bermingham N. A., Finegold M. J., and ZoghbiH. Y. (2001) Requirement of Math1 for secretory celllineage commitment in the mouse intestine. Science 294,2155–2158.
Yoh S. M. and Privalsky M. L. (2001) Transcriptional repres-sion by thyroid hormone receptors. A role for receptorhomodimers in the recruitment of SMRT corepressor.J. Biol. Chem. 276, 16,857–16,867.
Zheng J. L. and Gao W. Q. (2000) Overexpression of Math1induces robust production of extra hair cells in post-natal rat inner ears. Nat Neurosci 3, 580–586.
Zhu Y., Yu T., and Rao Y. (2004) Temporal regulation ofcerebellar EGL migration through a switch in cellularresponsiveness to the meninges. Dev. Biol. 267, 153–164.
Zine A. and de Ribaupierre F. (2002) Notch/Notch lig-ands and Math1 expression patterns in the organ ofCorti of wild-type and Hes1 and Hes5 mutant mice.Hear. Res. 170, 22–31.
JMN28_2_211_230_Arie 3/21/06 7:42 PM Page 229
Copyright © 2022 FDOKUMEN




















