
EVIDENCE OF ADAPTATION FROM ANCESTRAL VARIATION IN YOUNG POPULATIONS OF BEACH MICE
Upload
personal-psuCategory
view
3download
0
Major diversification of voltage-gated K+ channelsoccurred in ancestral parahoxozoansXiaofan Lia,b, Hansi Liua, Jose Chu Luoa, Sarah A. Rhodesa, Liana M. Trigga, Damian B. van Rossuma,b, Andriy Anishkinc,Fortunay H. Diattaa, Jessica K. Sassica, David K. Simmonsd, Bishoy Kamela, Monica Medinaa, Mark Q. Martindaled,and Timothy Jeglaa,b,1
aDepartment of Biology and bHuck Institutes of the Life Sciences, Penn State University, University Park, PA 16802; cDepartment of Biology, University ofMaryland, College Park, MD 20742; and dThe Whitney Marine Laboratory for Marine Bioscience, University of Florida, St. Augustine, FL 32080
Edited by Richard W. Aldrich, The University of Texas at Austin, Austin, TX, and approved January 22, 2015 (received for review December 1, 2014)
We examined the origins and functional evolution of the Shakerand KCNQ families of voltage-gated K+ channels to better under-stand how neuronal excitability evolved. In bilaterians, the Shakerfamily consists of four functionally distinct gene families (Shaker,Shab, Shal, and Shaw) that share a subunit structure consisting ofa voltage-gated K+ channel motif coupled to a cytoplasmic domainthat mediates subfamily-exclusive assembly (T1). We traced the or-igin of this unique Shaker subunit structure to a common ancestorof ctenophores and parahoxozoans (cnidarians, bilaterians, and pla-cozoans). Thus, the Shaker family is metazoan specific but is likelyto have evolved in a basal metazoan. Phylogenetic analysis sug-gested that the Shaker subfamily could predate the divergence ofctenophores and parahoxozoans, but that the Shab, Shal, and Shawsubfamilies are parahoxozoan specific. In support of this, putativectenophore Shaker subfamily channel subunits coassembled withcnidarian and mouse Shaker subunits, but not with cnidarian Shab,Shal, or Shaw subunits. The KCNQ family, which has a distinct sub-unit structure, also appears solely within the parahoxozoan lineage.Functional analysis indicated that the characteristic properties ofShaker, Shab, Shal, Shaw, and KCNQ currents evolved before thedivergence of cnidarians and bilaterians. These results show thata major diversification of voltage-gated K+ channels occurred inancestral parahoxozoans and imply that many fundamental mech-anisms for the regulation of action potential propagation evolvedat this time. Our results further suggest that there are likely tobe substantial differences in the regulation of neuronal excitabilitybetween ctenophores and parahoxozoans.
Shaker | KCNQ | Nematostella | ctenophore | Mnemiopsis
Voltage-gated K+ channels are highly conserved among bilat-erian metazoans and play a central role in the regulation of
excitation in neurons and muscle. Understanding the functionalevolution of these channels may therefore provide importantinsights into how neuromuscular excitation evolved within theMetazoa. Three major gene families, Shaker, KCNQ, and Ether-a-go-go (EAG) encode all voltage-gated K+ channels in bilat-erians (1, 2). In this study, we examine the functional evolutionand origins of the Shaker and KCNQ gene families. Shaker familychannels can be definitively identified by a unique subunitstructure that includes both a voltage-gated K+ channel core anda family-specific cytoplasmic domain within the N terminus knownas the T1 domain. T1 mediates assembly of Shaker family sub-units into functional tetrameric channels (3, 4). KCNQ channelsare also tetrameric but lack a T1 domain and use a distinct coiled-coil assembly domain in the C terminus (5, 6). KCNQ channelscan be identified by the presence of this family-specific assemblymotif and high amino acid conservation within the K+ channelcore. Both channel families are found in cnidarians (1, 7) and thuspredate the divergence of cnidarians and bilaterians, but theirultimate evolutionary origins have not yet been defined.Shaker family K+ channels serve diverse roles in the regulation
of neuronal firing and can be divided into four gene subfamiliesbased on function and sequence homology: Shaker, Shab, Shal,
and Shaw (8, 9). The T1 assembly domain is only compatiblebetween subunits from the same gene subfamily (4, 10) and thusserves to keep the subfamilies functionally segregated. Shakersubfamily channels activate rapidly near action potential thresh-old and range from rapidly inactivating to noninactivating. Mul-tiple roles for Shaker channels in neurons and muscles have beendescribed, but their most unique and fundamental role may bethat of axonal action potential repolarization. Shaker channelsare clustered to the axon initial segment and nodes of Ranvier invertebrate neurons (11–13) and underlie the delayed rectifier insquid giant axons (14). The Shaker subfamily is diverse in cni-darians (15, 16), and the starlet sea anemone Nematostella vec-tensis has functional orthologs of most identified Shaker currenttypes observed in bilaterians (16).The Shab and Shal gene subfamilies encode somatodendritic
delayed rectifiers and A currents, respectively (17–20). Shabchannels are important for maintaining sustained firing (21, 22),whereas the Kv4-based A current modulates spike threshold andfrequency (17). Shab and Shal channels are present in cnidarians,but cnidarian Shab channels have not been functionally charac-terized, and the only cnidarian Shal channels expressed to datedisplay atypical voltage dependence and kinetics compared withbilaterian channels (23). Shaw channels are rapid, high-thresholdchannels specialized for sustaining fast firing in vertebrates (24, 25)but have a low activation threshold and may contribute to resting
Significance
We examined the origin and evolution of two major families ofvoltage-gated K+ channels, Shaker and KCNQ, which regulateaction potential repolarization, patterning, and threshold. Shakerfamily channels evolved in a basal metazoan ancestor of cteno-phores and parahoxozoans (including cnidarians and bilaterians),but functional diversification of the Shaker family and the emer-gence of the KCNQ family occurred specifically within theparahoxozoan lineage. Our results suggest that many majorinnovations in the regulation of cellular excitability by voltage-gated K+ channels are unique to parahoxozoans and that theseinnovations occurred before the divergence of cnidarians andbilaterians. Ctenophores and sponges separated prior to this burstof innovation and thus either lack major mechanisms for actionpotential regulation or evolved such mechanisms independently.
Author contributions: T.J. designed research; X.L., H.L., J.C.L., S.A.R., L.M.T., D.B.v.R., A.A.,F.H.D., J.K.S., and T.J. performed research; D.K.S., B.K., M.M., and M.Q.M. contributednew reagents/analytic tools; X.L., H.L., J.C.L., D.B.v.R., A.A., F.H.D., and T.J. analyzed data;and X.L., D.B.v.R., A.A., and T.J. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The sequences reported in this paper have been deposited in the Gen-Bank database (accession nos. KP219389–KP219399).1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1422941112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1422941112 PNAS Early Edition | 1 of 10
NEU
ROSC
IENCE
PNASPL
US

potential in Drosophila (19, 26, 27). A Caenorhabditis elegansShaw has slow kinetics but a high activation threshold (28), anda single expressed cnidarian Shaw channel has the opposite:a low activation threshold but relatively fast kinetics (29). Thus,the ancestral properties and function of Shaw channels is not yetunderstood. Further functional characterization of cnidarianShab, Shal, and Shaw channels would provide a better un-derstanding of the evolutionary status of the Shaker family inearly parahoxozoans.KCNQ family channels underlie the M current in vertebrate
neurons (30) that regulates subthreshold excitability (31). The Mcurrent provides a fundamental mechanism for regulation offiring threshold through the Gq G-protein pathway becauseKCNQ channels require phosphatidylinositol 4,5-bisphosphate(PIP2) for activation (32, 33). PIP2 hydrolysis and subsequentKCNQ channel closure initiated by Gq-coupled receptors pro-duces slow excitatory postsynaptic potentials, during which theprobability of firing is greatly increased (32, 33). The key func-tional adaptations of KCNQ channels for this physiological rolethat can be observed in vitro are (i) a requirement for PIP2 tocouple voltage-sensor activation to pore opening (34, 35), and(ii) a hyperpolarized voltage–activation curve that allows chan-nels to open below typical action potential thresholds. Both keyfeatures are found in vertebrate (30, 34, 36–38), Drosophila (39),and C. elegans (40) KCNQ channels, suggesting they may havebeen present in KCNQ channels in a bilaterian ancestor. Evo-lution of the M current likely represented a major advance in theability to modulate the activity of neuronal circuits, but it is notyet clear when PIP2-dependent KCNQ channels first evolved.Here, we examine the origins and functional evolution of the
Shaker and KCNQ gene families. If we assume the evolution ofneuronal signaling provided a major selective pressure for thefunctional diversification of voltage-gated K+ channels, then wecan hypothesize that the appearance of these gene families mightaccompany the emergence of the first nervous systems or a majorevent in nervous system evolution. Recent phylogenies that placethe divergence of ctenophores near the root of the metazoan treesuggest that the first nervous systems, or at least the capacity tomake neurons, may have been present in a basal metazoan an-cestor (41–43) (Fig. S1). One hypothesis then is that much of thediversity of metazoan voltage-gated channels should be sharedbetween ctenophores and parahoxozoans [cnidarians, bilaterians,and placozoans (44)]. However, genome analysis indicates thatmany “typical” neuronal genes are missing in ctenophores andthe sponges lack a nervous system, leading to the suggestion thatextant nervous systems may have evolved independently in cteno-phores and parahoxozoans (42, 45). Thus, a second hypothesis isthat important steps in voltage-gated K+ channel evolutionmight have occurred separately in ctenophores and parahoxozoans.We tested these hypotheses by carefully examining the phyloge-netic distribution and functional evolution of Shaker and KCNQfamily K+ channels. Our results support a model in which majorinnovations in neuromuscular excitability occurred specificallywithin the parahoxozoan lineage.
ResultsTo determine the evolutionary origin of the Shaker family, weused a BLAST (46) search strategy to identify potential Shakerfamily genes in choanoflagellates, ctenophores, and sponge withbilaterian and cnidarian Shaker, Shab, Shal, and Shaw proteinsas queries. We defined Shaker family channels as (i) includinga T1 domain, and (ii) having best matches to known Shakerfamily channels within the voltage-gated K+ channel core motifin reciprocal BLAST searches against bilaterian and cnidariansequences. Shaker family channels were not present in genomedrafts and gene predictions from two choanoflagellates, Sal-pingoeca rosetta and Monosiga brevicollis (47, 48). Becausechoanoflagellates are believed to represent the closest extant
relatives of metazoans, this is a strong indication that the Shakerfamily is metazoan specific. In support of this view, voltage-gatedK+ channels cloned from prokaryotes (49), plants (50, 51), andfungi (52) lack the T1 domain and Shaker-specific homology.We next searched for Shaker family channels in sponges and
ctenophores because current views of metazoan phylogeny placectenophores or sponges as the most basally branching extantphyla (41–43). We were unable to find Shaker channels in thegenome of the sponge Amphimedon queenslandica (53), or inseven of eight sponge species transcriptomes (54). However,three ESTs from the Corticium candelabrum transcriptome couldbe assembled into two nonoverlapping fragments with specifi-cally high homology to the Shaker subfamily, one covering T1–S1and one covering S3 to the C terminus (Fig. S2). In contrast, weidentified 49 Shaker family genes in the draft genome andtranscriptome of the ctenophore Mnemiopsis leidyi (41). Aminoacid predictions for channel proteins encoded by these genes areincluded in Dataset S1. Multiple Shaker family channels werealso present in the genome draft of a second ctenophore, Pleu-robachia bachei, and four ctenophore transcriptomes (Pleuro-bachia plus three additional species) (42). However, we did notassemble Shaker channel sets from these species because manyPleurobachia gene predictions and transcriptome sequences werefragmentary. As a whole, these results indicate that the Shakerfamily originated in basal metazoans. The reason for the ap-parent absence of Shaker family channels in many sponges iscurrently unclear but could point to loss of the family withinvarious sponge lineages.To determine the evolutionary relationship between cteno-
phore and parahoxozoan Shaker family channels, we constructeda Bayesian inference phylogeny (Fig. 1) based on alignment ofthe T1 and voltage-gated K+ channel core motifs. We excludedseven ctenophore channels and the Corticium candelabrumfragments because of large sequence gaps in these motifs. Eachof the seven excluded ctenophore channels had highest homol-ogy to other Mnemiopsis sequences in BLAST comparisons. Thephylogeny splits Mnemiopsis Shaker family channels into twoclades, one separate from the parahoxozoan channels and onewithin the parahoxozoan Shaker subfamily (Fig. 1). The phy-logeny therefore supports a model in which the Shaker subfamilyevolved before the divergence of ctenophores and parahoxozoans,and that Shab, Shal, and Shaw later evolved from Shaker spe-cifically within the parahoxozoan lineage. All top hits identifiedin BLAST searches of Pleurobrachia bachei (42) using Shab,Shal, and Shaw channels best matched identified MnemiopsisShaker family sequences when used as queries in reciprocalBLAST searches. Therefore, both ctenophores species appearto lack Shab, Shal, and Shaw channels. We rebuilt and tested thephylogeny using both minimum-evolution and maximum-likeli-hood methods to further assess these findings. Minimum evo-lution supports the same topology as Bayesian inference (Fig.S3). Maximum likelihood supports the late evolution of Shab,Shal, and Shaw and a close relationship between one cteno-phore channel clade and the Shaker subfamily, but is unable toresolve the branching pattern (Fig. S3) between them. Sequencesimilarity network (SSN) analysis also agrees with the Bayesianinference topology (Fig. S4). These sequence analyses thereforeconverge on a model in which the Shaker subfamily evolvedbefore the Shab, Shal, and Shaw subfamilies and possibly beforethe ctenophore/parahoxozoan divergence.Nematostella Shaker channels have been extensively charac-
terized and demonstrate functional conservation between hydro-zoans, anthozoans, and bilaterians (15, 16, 55). To better understandthe functional evolution of the Shaker family, we first func-tionally expressed Nematostella whole Shab, Shaw, and Shalchannels in Xenopus oocytes. Cnidarian Shab channels have notpreviously been expressed. Nematostella Shab (NvShab) currentsare compared with the mouse Shab channel Kv2.1 in Fig. 2A. They
2 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1422941112 Li et al.

T.adh_2482H.sap_Kv2.2
N.vec_114311
M.lei_50420
0.97
0.960.82
0.88
0.93
0.79
0.73
0.71
0.580.61
0.79
0.87
0.86
0.92
0.92
0.81
0.62
0.86
0.68
0.94
0.89
0.68
0.68
0.97
0.65
0.71
0.69
0.77 0.96
0.69
0.94
0.72
0.61
0.70
0.81
0.53
0.51
0.85
0.79
0.53
0.61
0.68
0.71
0.52
0.75
0.80
0.55
0.940.96
0.2
H.sap_Kv2.1D.mel_ShabS.pur_00856 H.sap_Kv8.2
H.sap_Kv9.2H.sap_Kv9.3
H.sap_Kv9.1H.sap_Kv8.1 H.sap_Kv6.4H.sap_Kv6.3
H.sap_Kv6.1H.sap_Kv6.2H.sap_Kv5.1
N.vec_NvShabT.adh_3098 C.ele_kvs-3
C.ele_kvs-2C.ele_exp2C.ele_kvs-5
C.ele_kvs-1C.ele_kvs-4H.sap_Kv3.4H.sap_Kv3.2
H.sap_Kv3.3H.sap_Kv3.1 D.mel_Shaw2
D.mel_ShawC.ele_shw-3
C.ele_egl2C.ele_shw-1S.pur_06470
N.vec_208071N.vec_104830
N.vec_36427N.vec_228740 N.vec_NvShawR2N.vec_84381
N.vec_NvShawR1N.vec_40487
N.vec_NvShaw1N.vec_109242
T.adh_1922H.sap_Kv4.3
H.sap_Kv4.2H.sap_Kv4.1
D.mel_ShalC.ele_shl-1S.pur_12871
N.vec_NvShal1
N.vec_85827
N.vec_90372
N.vec_NvShalR3N.vec_NvShalR2
N.vec_517238N.vec_NvShalR1N.vec_33143
N.vec_1001
N.vec_218758N.vec_86018
N.vec_137731
H.sap_Kv1.8H.sap_Kv1.4
H.sap_Kv1.1H.sap_Kv1.2
H.sap_Kv1.3H.sap_Kv1.6
H.sap_Kv1.5H.sap_Kv1.7D.mel_Shaker
S.pur_26003N.vec_NvShak4 C.ele_shk-1
N.vec_NvShakR8N.vec_NvShakR1N.vec_NvShak3
N.vec_NvShak5N.vec_NvShak6 N.vec_NvShakR11
N.vec_NvShakR9N.vec_NvShakR10
N.vec_NvShakR14N.vec_NvShakR12N.vec_NvShakR6
N.vec_NvShakR7 N.vec_NvShakR5N.vec_NvShakR4N.vec_NvShak1
N.vec_NvShak2 N.vec_NvShakR2
M.lei_MlShak2
M.lei_MlShak1
M.lei_131129 M.lei_022314M.lei_375927
M.lei_1381M.Lei_06263 M.lei_070821M.lei_14971
M.lei_149210 M.lei_050920M.lei_07015
M.lei_35172 M.lei_08827M.lei_101015
M.lei_04675
M.lei_070211M.lei_025912
M.lei_11462 M.lei_351735M.lei_351734
M.lei_258237M.lei_01366
M.lei_10392M.lei_07807
M.lei_141713M.lei_05482M.lei_02524
M.lei_40863 M.lei_204441M.lei_050918
M.lei_32225M.lei_21868
M.lei_00641M.lei_030524
M.lei_1117M.lei_35174
M.lei_17471M.lei_00483
M.lei_004421M.lei_18152
Shab
Sub
fam
ilySh
aker
Sub
fam
ily
CtenophoreCnidarianPlacozoanProtostomeDeuterostome
Shaw
Sub
fam
ilySh
al S
ubfa
mily
Fig. 1. Bayesian inference phylogeny of the metazoan Shaker K+ channel family. Major metazoan clades are identified by color according to the legend atthe Upper Left; the Shaker, Shab, Shal, and Shaw subfamilies are marked at the right margin, and protein names are given at branch termini. Species prefixesin the protein names in alphabetical order are as follows: C.ele (Caenorhabditis elegans, nematode), D.mel (Drosophila melanogaster, fruit fly), H.sap (Homosapiens, human), M.lei (Mnemiopsis leidyi, ctenophore), N.vec (Nematostella vectensis, sea anemone), S.pur (Stronglyocentrotus purpuratus, sea urchin), andT.adh (Trichoplax adhaerens, placozoan). Green asterisks mark channels functionally expressed in this study. Sequences used in phylogeny construction areprovided in Dataset S1. The scale bar indicates number of substitutions per site, and posterior probabilities are given at branch nodes. Unlabeled nodes hadposterior probabilities ≥0.97.
Li et al. PNAS Early Edition | 3 of 10
NEU
ROSC
IENCE
PNASPL
US
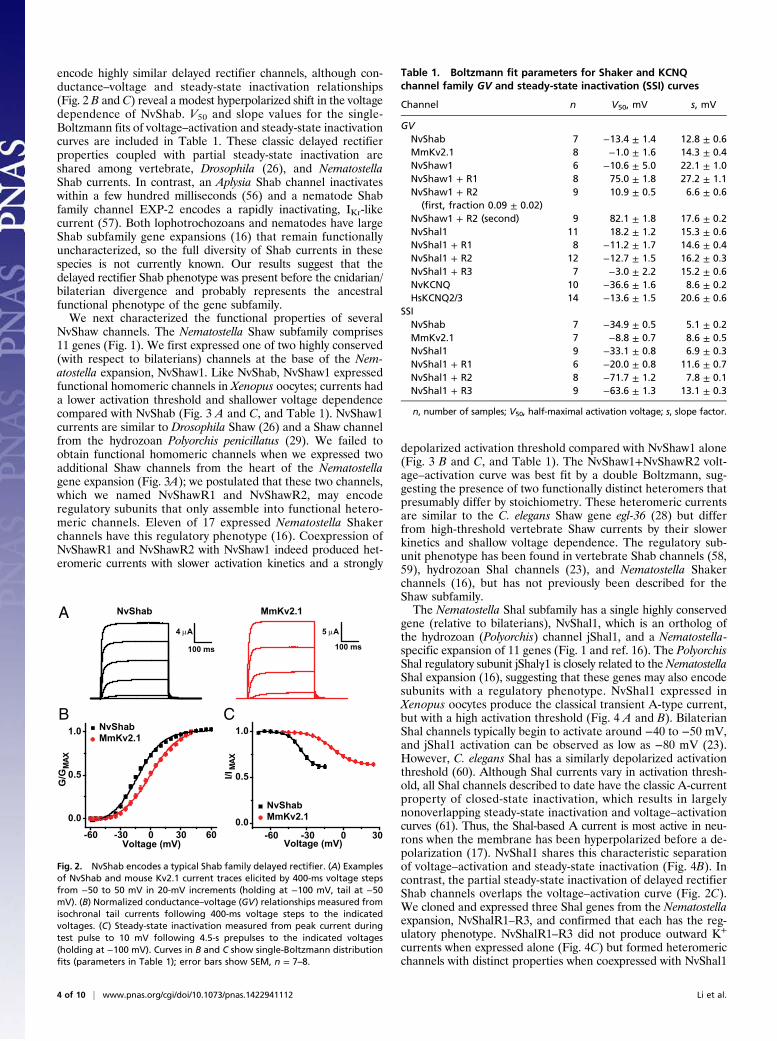
encode highly similar delayed rectifier channels, although con-ductance–voltage and steady-state inactivation relationships(Fig. 2 B and C) reveal a modest hyperpolarized shift in the voltagedependence of NvShab. V50 and slope values for the single-Boltzmann fits of voltage–activation and steady-state inactivationcurves are included in Table 1. These classic delayed rectifierproperties coupled with partial steady-state inactivation areshared among vertebrate, Drosophila (26), and NematostellaShab currents. In contrast, an Aplysia Shab channel inactivateswithin a few hundred milliseconds (56) and a nematode Shabfamily channel EXP-2 encodes a rapidly inactivating, IKr-likecurrent (57). Both lophotrochozoans and nematodes have largeShab subfamily gene expansions (16) that remain functionallyuncharacterized, so the full diversity of Shab currents in thesespecies is not currently known. Our results suggest that thedelayed rectifier Shab phenotype was present before the cnidarian/bilaterian divergence and probably represents the ancestralfunctional phenotype of the gene subfamily.We next characterized the functional properties of several
NvShaw channels. The Nematostella Shaw subfamily comprises11 genes (Fig. 1). We first expressed one of two highly conserved(with respect to bilaterians) channels at the base of the Nem-atostella expansion, NvShaw1. Like NvShab, NvShaw1 expressedfunctional homomeric channels in Xenopus oocytes; currents hada lower activation threshold and shallower voltage dependencecompared with NvShab (Fig. 3 A and C, and Table 1). NvShaw1currents are similar to Drosophila Shaw (26) and a Shaw channelfrom the hydrozoan Polyorchis penicillatus (29). We failed toobtain functional homomeric channels when we expressed twoadditional Shaw channels from the heart of the Nematostellagene expansion (Fig. 3A); we postulated that these two channels,which we named NvShawR1 and NvShawR2, may encoderegulatory subunits that only assemble into functional hetero-meric channels. Eleven of 17 expressed Nematostella Shakerchannels have this regulatory phenotype (16). Coexpression ofNvShawR1 and NvShawR2 with NvShaw1 indeed produced het-eromeric currents with slower activation kinetics and a strongly
depolarized activation threshold compared with NvShaw1 alone(Fig. 3 B and C, and Table 1). The NvShaw1+NvShawR2 volt-age–activation curve was best fit by a double Boltzmann, sug-gesting the presence of two functionally distinct heteromers thatpresumably differ by stoichiometry. These heteromeric currentsare similar to the C. elegans Shaw gene egl-36 (28) but differfrom high-threshold vertebrate Shaw currents by their slowerkinetics and shallow voltage dependence. The regulatory sub-unit phenotype has been found in vertebrate Shab channels (58,59), hydrozoan Shal channels (23), and Nematostella Shakerchannels (16), but has not previously been described for theShaw subfamily.The Nematostella Shal subfamily has a single highly conserved
gene (relative to bilaterians), NvShal1, which is an ortholog ofthe hydrozoan (Polyorchis) channel jShal1, and a Nematostella-specific expansion of 11 genes (Fig. 1 and ref. 16). The PolyorchisShal regulatory subunit jShalγ1 is closely related to theNematostellaShal expansion (16), suggesting that these genes may also encodesubunits with a regulatory phenotype. NvShal1 expressed inXenopus oocytes produce the classical transient A-type current,but with a high activation threshold (Fig. 4 A and B). BilaterianShal channels typically begin to activate around −40 to −50 mV,and jShal1 activation can be observed as low as −80 mV (23).However, C. elegans Shal has a similarly depolarized activationthreshold (60). Although Shal currents vary in activation thresh-old, all Shal channels described to date have the classic A-currentproperty of closed-state inactivation, which results in largelynonoverlapping steady-state inactivation and voltage–activationcurves (61). Thus, the Shal-based A current is most active in neu-rons when the membrane has been hyperpolarized before a de-polarization (17). NvShal1 shares this characteristic separationof voltage–activation and steady-state inactivation (Fig. 4B). Incontrast, the partial steady-state inactivation of delayed rectifierShab channels overlaps the voltage–activation curve (Fig. 2C).We cloned and expressed three Shal genes from the Nematostellaexpansion, NvShalR1–R3, and confirmed that each has the reg-ulatory phenotype. NvShalR1–R3 did not produce outward K+
currents when expressed alone (Fig. 4C) but formed heteromericchannels with distinct properties when coexpressed with NvShal1
-60 -30 0 30 60
0.0
0.5
1.0 NvShabMmKv2.1
G/G
MAX
Voltage (mV)
B C
NvShab MmKv2.1
-60 -30 0 300.0
0.5
1.0
Voltage (mV)
NvShabMmKv2.1
I/IM
AX
A4 μA
100 ms
5 μA
100 ms
Fig. 2. NvShab encodes a typical Shab family delayed rectifier. (A) Examplesof NvShab and mouse Kv2.1 current traces elicited by 400-ms voltage stepsfrom −50 to 50 mV in 20-mV increments (holding at −100 mV, tail at −50mV). (B) Normalized conductance–voltage (GV) relationships measured fromisochronal tail currents following 400-ms voltage steps to the indicatedvoltages. (C) Steady-state inactivation measured from peak current duringtest pulse to 10 mV following 4.5-s prepulses to the indicated voltages(holding at −100 mV). Curves in B and C show single-Boltzmann distributionfits (parameters in Table 1); error bars show SEM, n = 7–8.
Table 1. Boltzmann fit parameters for Shaker and KCNQchannel family GV and steady-state inactivation (SSI) curves
Channel n V50, mV s, mV
GVNvShab 7 −13.4 ± 1.4 12.8 ± 0.6MmKv2.1 8 −1.0 ± 1.6 14.3 ± 0.4NvShaw1 6 −10.6 ± 5.0 22.1 ± 1.0NvShaw1 + R1 8 75.0 ± 1.8 27.2 ± 1.1NvShaw1 + R2
(first, fraction 0.09 ± 0.02)9 10.9 ± 0.5 6.6 ± 0.6
NvShaw1 + R2 (second) 9 82.1 ± 1.8 17.6 ± 0.2NvShal1 11 18.2 ± 1.2 15.3 ± 0.6NvShal1 + R1 8 −11.2 ± 1.7 14.6 ± 0.4NvShal1 + R2 12 −12.7 ± 1.5 16.2 ± 0.3NvShal1 + R3 7 −3.0 ± 2.2 15.2 ± 0.6NvKCNQ 10 −36.6 ± 1.6 8.6 ± 0.2HsKCNQ2/3 14 −13.6 ± 1.5 20.6 ± 0.6
SSINvShab 7 −34.9 ± 0.5 5.1 ± 0.2MmKv2.1 7 −8.8 ± 0.7 8.6 ± 0.5NvShal1 9 −33.1 ± 0.8 6.9 ± 0.3NvShal1 + R1 6 −20.0 ± 0.8 11.6 ± 0.7NvShal1 + R2 8 −71.7 ± 1.2 7.8 ± 0.1NvShal1 + R3 9 −63.6 ± 1.3 13.1 ± 0.3
n, number of samples; V50, half-maximal activation voltage; s, slope factor.
4 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1422941112 Li et al.

(Fig. 4 D–F). NvShalR1 hyperpolarized the GV curve and con-verted the phenotype of NvShal1 to that of a Shab-like delayedrectifier: inactivation was greatly reduced during short steps (Fig.4D) and partial steady-state inactivation shifted to the Shab patternof overlap with the voltage–activation curve (Fig. 4E and Table 1).This is the first example to our knowledge of a Shal channelthat lacks an A-current phenotype. In contrast, NvShalR2 andNvShalR3 altered the inactivation rate observed in current traces(Fig. 4D) and hyperpolarized both the GV and steady-state in-activation curves (Fig. 4E and Table 1), but preserved the classicA-current phenotype. NvShalR2 and NvShalR3 also slow therate of recovery from inactivation (Fig. 4F). These results in-dicate that the large Nematostella Shal subfamily encodes a varietyof A currents and that at least one heteromeric Shal current(NvShal1+NvShalR1) instead contributes to delayed rectifierdiversity, typically the domain of the Shab subfamily.We next cloned two of the potential Mnemiopsis Shaker sub-
family channels, MlShak1 and MlShak2, to examine their func-tional phenotypes in vitro. However, neither of the two channelsproduced functional voltage-gated potassium currents whenexpressed alone or in combination in Xenopus oocytes (Fig. 5A).We reasoned that the channels we chose to express could beregulatory subunits, but identifying viable heteromeric combi-nations of subunits among 49 Mnemiopsis Shaker family geneswith no a priori knowledge of mixing rules was a daunting prop-osition. We therefore focused on functionally testing their puta-tive Shaker subfamily identity suggested by phylogenetic analysis.If MlShak1 and MlShak2 are Shaker subfamily genes, then theyshould specifically coassemble with parahoxozoan Shaker sub-family channels. We therefore coexpressed them with Nematostella(NvShak3) and mouse (MmKv1.2) Shaker channels. RNAs weremixed at a 10:1 ratio in favor of Mnemiopsis subunits to biasformation of heteromers. Both ctenophore channels introduceda slowly activating component into NvShak3 currents andslowed channel deactivation (Fig. 5B), indicating that functional
heteromers were formed. Currents were also significantly re-duced by MlShak1 coexpression (Fig. 5D), suggesting eithera smaller single-channel conductance for heteromers or that notall assembled channel stoichiometries were functional. Whenthe ctenophore channels were coexpressed with mouse Kv1.2,this dominant-negative suppression was the major phenotype;currents were reduced by ∼90% (Fig. 5 C and E). We did notclosely examine the small residual currents for evidence offunctional heteromer formation; dominant-negative suppressionis commonly used as evidence for coassembly of K+ channelsubunits (62–64).We further tested whether coassembly with MlShak1 and
MlShak2 was specific to the Shaker subfamily by coexpressingthe channels with NvShab, NvShal1, and NvShaw1. We did notobserve novel components in the currents when these channelswere mixed with MlShak1 and MlShak2 at RNA concentra-tions that specifically altered NvShak3 and MmKv1.2 currents
A
B CNvShawR1 NvShawR2
-100 -50 0 50 1000.0
0.5
1.0
G/G
MA
X
Voltage (mV)
NvShaw1
4 μA100 ms
1 μA150 ms
1 μA150 ms
300 ms 300 ms
NvShaw1 + R1 NvShaw1 + R2
NvShaw1+ R1+ R2
Fig. 3. Functional properties of homomeric and heteromeric NematostellaShaw channels. (A) Current traces elicited from oocytes injected with NvShaw1,NvShawR1, and NvShawR2 by 400-ms voltage steps from −60 to 60 mV in20-mV increments (holding at −100 mV; tail at −50 mV). (B) Superimposedcurrent traces from oocytes injected with NvShaw1 alone (gray), orNvShaw1+NvShawR1 (Left) and NvShaw1+NvShawR2 (Right). Traces werenormalized to peak current at 60 mV for comparison. (C) Normalized GVcurves of homomeric and heteromeric Nematostella Shaw channels. Datawere taken from isochronal tail currents following 400-ms voltage steps forNvShaw1, and from peak current adjusted for driving force (assuming a−100-mV reversal potential) for heteromeric currents. Curves show single(NvShaw1 and NvShaw1+R1)- or double (NvShaw1+R2)-Boltzmann fits(parameters reported in Table 1; error bars show SEM, n = 6–9). NvShaw1was injected at a 1:10 RNA ratio relative to NvShawR1 and NvShawR2 toproduce a predominantly heteromeric current for comparison.
C
NvShal1
NvShal1+R1 NvShal1+R2 NvShal1+R3
A B
D
NvShal R1
NvShal R2
NvShal R3-60 -30 0 30 60
0.0
0.5
1.0
0.0
0.5
1.0
G/G
Voltage (mV)
I/I
E F
-90 -60 -30 0 30 60
0.0
0.5
1.0
Voltage (mV)
I/I
0 200 400 600 800 1000
0.0
0.5
1.0
Interpulse Interval (ms)
I/I
4 μA
300 ms
300 ms1 μA
4 μA
300 ms 0.4 μA
300 ms
2 μA
300 ms
MA
X
0.0
0.5
1.0
G/G
MA
X
MA
X
NvShal1+R1NvShal1+R2NvShal1+R3
NvShal1NvShal1+R2NvShal1+R3
MA
X
MA
X
Fig. 4. Functional properties of homomeric and heteromeric NematostellaShal channels. (A) Current traces from an oocyte expressing NvShal1 elicitedby 1-s voltage steps (−60 to 60 mV in 20-mV increments; holding at −100 mVand tail at −50 mV). (B) Normalized GV (black) and steady-state inactivation(gray) relationships for NvShal1. GV was measured from peak current duringdepolarizing steps and adjusted for driving force (assuming −100-mV re-versal). Steady-state inactivation was measured from peak current at +40 mVfollowing 4.5-s steps to the indicated voltages. (C) No K+ currents could beelicited from oocytes injected with NvShalR1-3; voltage protocols are thesame as in A. (D) Current traces elicited by the same voltage protocols fromoocytes injected with NvShal1+NvShalR1–R3 (1:10 RNA ratio). (E) Normal-ized GV (black) and steady-state inactivation (gray) relationships for het-eromeric Nematostella Shal currents, measured and fit as in B. The dashedlines are Boltzmann fits of data from homomeric NvShal1 currents shownin B. Data in B and E show average values, and error bars indicate SEM (n =6–12); curves in B and E show single-Boltzmann fits (V50 and slope valuesreported in Table 1). (F) Time course of recovery from inactivation forNvShal1, NvShal1+NvShalR2, and NvShal1+NvShalR3. Recovery was de-termined using a double 1-s, +40-mV pulse protocol: data show fraction ofcurrent recovered in a second depolarizing pulse after a repolarizing step to−100 mV of the indicated duration. Curves show a double-exponential fit ofthe data. Time constants (τ) for NvShal1 were 7.8 ± 1.2 (0.68 ± 0.03 fractionalamplitude) and 36.5 ± 6.3 (n = 10; ±SEM). Time constants for NvShal1+NvShalR2 were 74.4 ± 4.5 (0.60 ± 0.02 fractional amplitude) and 350.5 ± 30.5(n = 14), and time constants for NvShal1+NvShalR3 were 98.0 ± 9.4 (0.25 ±0.01 fractional amplitude) and 1,030.9 ± 119.2 (n = 8).
Li et al. PNAS Early Edition | 5 of 10
NEU
ROSC
IENCE
PNASPL
US

(Fig. 6 A, C, and E). Furthermore, we did not observe dominant-negative suppression of currents during these coexpression experi-ments (Fig. 6 B, D, and F). These results therefore indicate thatfunctional and dominant-negative assembly with MlShak1 andMlShak2 is limited to Shaker subfamily and thus support the hy-pothesis that MlShak1 andMlShak2 are Shaker subfamily channels.We next examined the origins and functional evolution of the
KCNQ gene family, starting with a similar BLAST search strategy.We defined KCNQ genes as containing the C-terminal coiled-coil assembly domains and having greater homology to KCNQchannels in voltage-gated K+ channel core in reciprocal BLASTsearches against bilaterian sequences. We did not find KCNQfamily genes outside the parahoxozoan lineage: There were noKCNQ family genes in two choanoflagellate genomes (47, 48),two ctenophore genomes and five ctenophore transcriptomes(41, 42), or one sponge genome and eight sponge transcriptomes(53, 54). Furthermore, we were unable to find KCNQ channels
in the placozoan Trichoplax adhaerens (65), a parahoxozoan.However, KCNQ channels can be found in all cnidarian andbilaterian genomes we have examined. Anthozoans Nematostellavectensis, Acropora digitifera, and Orbicella faveolata had a singleKCNQ channel, whereas the hydrozoan Hydra magnipapillata(66) had six KCNQ channels. Phylogenetic analysis indicatedthat two distinct clades of KCNQ channels are present inbilaterians and that all cnidarian KCNQ channels fall outsidethese clades (Fig. 7). The phylogeny supports a model in whicha cnidarian/bilaterian ancestor had a single KCNQ channel, whichwas then duplicated in an ancestral bilaterian after divergencefrom cnidarians. Members of both bilaterian KCNQ clades dis-play the characteristic PIP2 dependence of the channel family(34), suggesting that this defining feature evolved before the du-plication of KCNQ in bilaterians.We functionally expressed Nematostella KCNQ (NvKCNQ)
and compared it to human KCNQ2/KCNQ3 heteromers, a majorM channel found in vivo in vertebrate nervous systems (30).NvKCNQ activates more slowly than HsKCNQ2/3 and hasa flatter conductance–voltage curve (Fig. 8 A and B), but retainsthe low-threshold activation, which is believed to allow KCNQchannels to control excitation threshold. NvKCNQ reaches half-maximal activation (V50) around −13.6 ± 1.5 mV (n = 14), sig-nificantly more positive than HsKCNQ2/3 channels (V50 = −36.6 ±1.6 mV; n = 10), but activation of both channels can be ob-served at voltages below −40 mV (Fig. 8 A and B). We coex-pressed a voltage-sensitive phosphoinositide phosphatase from
A
B
C
MmKv1.2
0
5
10
15
20
******
0
2
4
6
8
*
n.s.p = 0.08
NvShak3
D E
NvShak3/MlShak1 NvShak3/MlShak2
MlShak1 MlShak2 MlShak1/2
MmKv1.2/MlShak1 MmKv1.2/MlShak2
1 μA
300 ms
2 μA150 ms
5 μA150 ms
Cur
rent
at 6
0 m
V (μ
A)
+ MlShak1
NvShak3
+ MlShak2Control
n =
14
n =
7 n =
8
Cur
rent
at 6
0 m
V (μ
A)
MmKv1.2
n =
7
n =
7 n =
8
+ MlShak1 + MlShak2Control
Fig. 5. Ctenophore Shaker family subunits coassemble with Nematostellaand mouse Shaker subfamily subunits. (A) No outward K+ currents weredetected in oocytes expressing the Mnemiopsis leidyi Shaker family channelsMlShak1 and MlShak2 either alone or in combination (1:1 RNA ratio) (1-svoltage steps from −60 to 60 mV in 20-mV increments, holding at −100 mV).(B and C) Current traces showing the Nematostella Shaker subfamily channelNvShak3 (B) and the mouse Shaker subfamily channel Kv1.2 (MmKv1.2; C)expressed alone or together with either MlShak1 or MlShak2 (1:10 RNAratio). Currents were elicited by 400-ms voltage steps from −60 to 60 mV in20-mV increments (holding at −100 mV; tail at 0 mV for NvShak3 and −50mV for MmKv1.2). Gray traces in B show NvShak3 current at 60 mV nor-malized to the heteromeric channel peak current at 60 mV to illustratechanges in current shape; note the pronounced slowing of deactivation(arrows). (D and E) Comparison of average peak outward current amplitudesrecorded at 60 mV for NvShak3 (D) and MmKv1.2 (E) alone and in combi-nation with MlShak1 and MlShak2. Error bars show SEM, and n is indicatedon the graphs. Asterisks indicate significant difference in amplitude fromNvShak3 or MmKv1.2: *P < 0.05; ***P < 0.001 (two-tailed t test).
C
NvShaw1/MlShak1 NvShaw1/MlShak2
NvShab/MlShak1 NvShab/MlShak2
NvShal1/MlShak1 NvShal1/MlShak2
E
0
5
10
15n.s.
n.s.
n = 9 n = 8 n = 9
0
5
10
15
20
n = 9 n = 8 n = 6
n.s. n.s.D
0
4
8
12
n.s.n.s.
n = 8 n = 9 n = 10
F
A B
NvShab
NvShab
/
MlShak1
NvShab
/
MlShak2
NvShaw
1
NvShaw
1
/MlShak
1
NvShaw
1
/MlShak
2
NvShal1
NvShal1
/
MlShak1
NvShaw
1/
MlShak2
Cur
rent
at 6
0 m
V (μ
A)
Cur
rent
at 6
0 m
V (μ
A)
Cur
rent
at 6
0 m
V (μ
A)
200 ms
200 ms
200 ms
Fig. 6. Ctenophore Shaker family subunits do not coassemble with Nem-atostella Shab, Shal, or Shaw channels. (A) Current traces are compared forNvShab (gray) and NvShab+MlShak1 (Left, black) and NvShab+MlShak2(Right, black). Currents were elicited by 1-s steps ranging from −60 to 60 mVin 20-mV increments and normalized to the amplitude at 60 mV. No dif-ferences in current shape were observed. (B) Peak outward current ampli-tudes at 60 mV are shown for NvShab, NvShab+MlShak1, and NvShab+MlShak2 (average of the indicated number of measurements; error barsshow SEM). No significant differences in current amplitude were observed intwo-tailed t test comparisons (n.s.). (C–F) Identical analyses are shown forNvShaw1 and NvShal1. NvShab, NvShaw1, and NvShal1 were coexpressedwith MlShak1 and MlShak2 at a 1:10 RNA ratio.
6 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1422941112 Li et al.

Ciona intestinalis, VSP, with NvKCNQ to test PIP2 dependence.VSP dephosphorylates PIP2 into PI4P when activated bymembrane depolarization, and can reduce PIP2 levels in theoocyte membrane (67). If NvKCNQ requires PIP2 for activa-tion, a decrease in current should be observed during repeateddepolarization in the presence of VSP. NvKCNQ currents re-duced in magnitude during repeated steps to 0 mV from a hold-ing potential of −100 mV when coexpressed with VSP, but not incontrols (Fig. 8 C and D). HsKCNQ2/3 currents were similarlyreduced in repeated 2-s steps to +40 mV specifically whencoexpressed with VSP (Fig. 8 E and F). We did not observea significant effect of VSP on HsKCNQ2/3 at 0 mV (where lessVSP activation occurs), and a single sweep to +40 mV was suf-ficient to eliminate NvKCNQ currents. Both results suggestNvKCNQ has a lower affinity for PIP2, and a smaller decreasein PIP2 abundance may be sufficient to eliminate activation.Coexpression of NvKCNQ with PI(4)P5-kinase (PIP5K), whichcatalyzes PIP2 production from PI4P (68), increased peak NvKCNQcurrent size during 2-s voltage step to 40 mV (1 d postinjection)from 0.43 ± 0.09 μA (n = 8) in controls to 0.98 ± 0.16 μA (n = 10).This indicates that NvKCNQ is not PIP2 saturated at restingXenopus oocyte PIP2 concentration. The VSP and PIP5K coex-pression experiments together demonstrate PIP2 dependencefor NvKCNQ. Thus, PIP2-dependent gating of KCNQ channelsevolved before the cnidarian/bilaterian divergence.The structural basis of PIP2 sensitivity in KCNQ channels has
been studied using mammalian channels. Two separate bindingsites for PIP2 have been proposed: one that couples the voltagesensor to the activation gate and includes positively chargedresidues in the intracellular linkers of the voltage sensor and nearthe S6 activation gate of the pore (35), and one that implicates
positively charged residues in the distal C terminus (69). Se-quence alignments of 13 KCNQ sequences from eight species,including NvKCNQ, show high conservation of putative PIP2-binding residues of the voltage sensor/activation gate site (Fig.S5). In contrast, we observed no significant alignment of theproposed C terminus binding site between KCNQ channels fromdistant species. We therefore suggest that the major evolution-arily conserved site for PIP2 modulation of KCNQ channelsresides at the voltage-sensor/activation gate interface. It is pos-sible that the proposed C terminus binding site found in somevertebrate KCNQ channels (69) may contribute to their highPIP2 affinity.
DiscussionOur results combined with previous studies (15, 16, 23, 29, 55)show that the functional and molecular diversification of Shakerand KCNQ families of voltage-gated K+ channels was largelycomplete before the divergence of cnidarians and bilaterians.Furthermore, Erg K+ channels, which constitute one of threebilaterian Ether-a-go-go gene subfamilies, are also highly con-served on functional level between cnidarians and vertebrates(70). The other two Ether-a-go-go subfamilies, Elk and Eag, havebeen identified in Nematostella (1). Thus, eight major classes ofvoltage-gated K+ channel are conserved between cnidarians andbilaterians, and the characteristic functional properties of six(Shaker, Shab, Shal, Shaw, KCNQ, and Erg) have now been shownto have evolved before the cnidarian/bilaterian divergence. Fur-ther support for an early diversification of voltage-gated K+
channels within parahoxozoans is the fact that Shaker, Shab,Shaw, and Erg channels can also be found in the placozoanTrichoplax adhaerens (65). Bilaterians are unique only in sharingtwo distinct KCNQ lineages; both KCNQ channel types are PIP2dependent (34, 40), and lineage-specific functional propertieshave not yet been described. Thus, ancestral parahoxozoans hada set of functionally diverse voltage-gated K+ channels that wassufficiently adaptable to regulate excitability in the highly diversenervous systems of extant cnidarians and bilaterians. The evolu-tion of voltage-gated Ca2+ channels tells a similar story; para-hoxozoans share L-type, T-type, and N/P/Q/R-type channels,but ctenophores only have an N/P/Q/R-type channel (71).Nematostella has single Shab and KCNQ genes, but contains
large and functionally diverse expansions of the Shaker, Shal,and Shaw subfamilies. Interestingly, many of the genes in theseexpansions encode regulatory subunits that only function inheteromeric channels. The large Shab family expansion in ver-tebrates also contains regulatory subunits (58, 59). The regulatorysubunit phenotype has therefore evolved independently in allfour gene subfamilies of the Shaker family during metazoan evo-lution. These gene expansions provide increased functional di-versity to specific subfamilies within various metazoan phylogeneticgroups. Nevertheless, each of the voltage-gated K+ channel typeswe have examined has characteristic functional properties thathave been highly conserved throughout parahoxozoan evolu-tion. These include the delayed rectifier phenotype of Shabchannels, the A-current phenotype of Shal channels, the rapidactivation and steep voltage dependence of Shaker channels (16),the low activation threshold and PIP2 dependence of KCNQchannels, and the IKr phenotype of Erg channels (59). The an-cestral properties of Shaw channels are less clear, but our studiesshow a high degree of similarity between ecdysozoan and cni-darian Shaw currents. The steeply voltage-dependent and rap-idly activating high-threshold Shaw currents of vertebratesmay therefore represent a recent adaption specialized for rapidfiring (24, 25).Our results also demonstrate that many of the voltage-gated
K+ channels that regulate excitability in bilaterian nervous sys-tems are absent in the basal metazoan ancestor of ctenophores,sponges, and parahoxozoans. Shab, Shal, Shaw, and KCNQ
0.60
0.86
0.850.81
M.mus_KCNQ1
M.mus_KCNQ2M.mus_KCNQ3
M.mus_KCNQ5M.mus_KCNQ4
C.int_143587S.pur_3726072
C.ele_KLQ1D.mel_KCNQ
A.gam_4228C.tel_183094
C.gig_1608L.gig_91828
C.int_148713S.pur_3729658
C.ele_KQT3A.gam_8384
C.gig_1282C.tel_111874
C.gig_473L.gig_81703
N.vec_KCNQO.fav_KCNQ
A.dig_KCNQH.mag_1235562
H.mag_1235969H.mag_36426
H.mag_32182H.mag_39101H.mag_37287
0.2
* * *
BilaterianKCNQ1-like
BilaterianKCNQ2-like
CnidarianKCNQs
CnidarianPlacozoanProtostomeDeuterostome
Fig. 7. Bayesian inference phylogeny of the metazoan KCNQ voltage-gatedK+ channel family. Color indicates major phylogenetic groups of metazoans,and dashed gray lines separate bilaterian KCNQ1-like, bilaterian KCNQ2-like,and cnidarian KCNQ sequences. Channel names include the following speciesprefixes: A.dig (Acropora digitifera, coral), A.gam (Anopheles gambiae,mosquito), C.ele (Caenorhabditis elegans, nematode), C.tel (Capitella teleta,polychaete), C.int (Ciona intestinalis, tunicate), C.gig (Crassostrea gigas, oyster),D.mel (Drosophila melanogaster, fruit fly), H.mag (Hydra magnipapillata,hydra), L.gig (Lottia gigantea, limpet), M.mus (Mus musculus, mouse), N.vec(Nematostella vectensis, sea anemone), O.fav (Orbicella faveolata, coral),and S.pur (Stronglyocentrotus purpuratus, sea urchin). Green asterisks markchannels we functionally compared. Sequences used in phylogeny con-struction are provided in Dataset S2. A scale bar indicates substitutions persite, and posterior probabilities <0.97 are given at branch nodes.
Li et al. PNAS Early Edition | 7 of 10
NEU
ROSC
IENCE
PNASPL
US

channels are all found exclusively within the parahoxozoan lin-eage. Shaker subfamily channels, in contrast, are older and mayhave been present in a common ancestor of ctenophores andparahoxozoans, as well as at least one sponge lineage based ontranscriptome sequences. We show evidence that twoMnemiopsisShaker family channels from a clade that is closely associated withthe parahoxozoan Shaker subfamily in phylogenies coassemblewith Shaker subfamily channels from Nematostella and mouse.The ability for subunits to coassemble is considered a definingproperty of gene subfamilies (72) within the Shaker family. Ourresults therefore suggest that the Shaker subfamily emergedbefore the ctenophore/parahoxozoan divergence. Functional char-acterization of pure Mnemiopsis Shaker channels, which we wereunable to obtain in this study, would resolve whether there isclose conservation of biophysical properties between the cteno-phore and parahoxozoan Shaker subfamilies.The high conservation of functional phenotype within K+ gene
families over long evolutionary distances suggests that at leastsome important aspects of the parahoxozoan voltage-gated K+
current diversity will be missing in ctenophores. However, ourresults do not necessarily mean that the complexity of electricalsignaling is “simple” in the ctenophore nervous systems. Mne-miopsis leidyi has 49 Shaker family genes, more than we haveobserved in any other species, and these fall into two distinctclades. One possibility is that the clades represent functionallydistinct gene subfamilies. However, because we have so far beenunable to express ctenophore channels, we do not yet know theextent of their functional diversity. It is entirely possible that
some of the diversity provided by parahoxozoan-specific Shaker K+
channel subfamilies could be provided by functional diversificationwithin the large Mnemiopsis gene expansions. Future cross-speciescomparisons of ctenophore Shaker family channels will be neededto determine how the diversity we observed inMnemiopsis evolvedwithin the ctenophore lineage. What is clear from our findings isthat if ctenophore voltage-gated K+ channels are functionally di-verse, that diversity evolved separately within ctenophores.The selective pressure that drove evolutionary diversification
of voltage-gated K+ channels in ancestral parahoxozoans has notyet been determined. If the hypothesis that the nervous systemevolved separately in ctenophores and parahoxozoans is true (42,45), then it may have been the de novo evolution of neurons andmuscle within the parahoxozoan lineage that provided a selectivepressure for the diversification of electrical signaling mechanisms.However, if nervous systems were present in the ctenophore/parahoxozoan ancestor, what might have driven the evolution ofso many new channel types in parahoxozoans? We can only spec-ulate because the evolution of nervous system structure and func-tion is still poorly understood. Parahoxozoan-specific channels(Shab, Shal, Shaw, and KCNQ) specifically regulate firing thresh-old or complex action potential patterning, whereas Shakerchannels, which our study suggests are shared by ctenophoresand parahoxozoans, play a critical role in axonal action potentialpropagation. It is therefore tempting to speculate that voltage-gated K+ channel evolution in basal metazoans was driven bythe evolution of action potential propagation, whereas para-hoxozoan-specific K+ channel diversification accompanied more
C
-120 -80 -40 0 40 80
0.0
0.5
1.0
G/G
Voltage (mV)
NvKCNQHsKCNQ2/3
NvKCNQ HsKCNQ2/3
A B
Dsweep 1
F
NvKCNQ NvKCNQ + VSP
sweep 1
HsKCNQ2/3 HsKCNQ2/3 + VSP
E
sweep 3 sweep 100.0
0.5
1.0NvKCNQ + VSP
***
Nor
mal
ized
peak
curr
ent NvKCNQ
***
sweep 3 sweep 100.0
0.5
1.0
******
HsKCNQ2/3 + VSP
Nor
mal
ized
peak
curr
ent HsKCNQ2/3
0.2 μA
500 ms 500 ms1 μA
0.2 μA
500 ms0.2 μA
500 ms
500 ms5 μA
500 ms5 μA
MA
X
Fig. 8. Nematostella KCNQ encodes a subthreshold PIP2-sensitive voltage-gated K+ channel. (A) Current traces from oocytes expressing Nematostella(NvKCNQ, Left) or human (HsKCNQ2/3, Right) KCNQ family channels. Currents were elicited by 2-s voltage steps from −120 to 60 mV in 20-mV increments(holding at −100 mV; tails at −50 mV). HsKCNQ2 and HsKCNQ3 were injected at a 1:1 ratio. (B) Normalized GV curves for NvKCNQ and HsKCNQ2/3 measuredfrom isochronal tail currents at −50 mV following 2-s voltage steps to the indicated voltages. Curves show fits to a single-Boltzmann distribution; V50 andslope values are reported in Table 1. (C) Example NvKCNQ current traces recorded in response to a series of 15 2-s voltage sweeps to 0 mV for NvKCNQexpressed alone (Left) or coexpressed with the Ciona VSP to progressively deplete PIP2 (Right). (D) Peak current sizes observed in sweep 3 and sweep10 relative to the peak current sizes of the first sweep. Error bars show SEM; n = 9 (control), 11 (VSP). The triple asterisk (***) denotes significance at P < 0.001(t test). (E and F) Experiments were repeated for HsKCNQ2/3; voltage steps were taken to 40 mV, n = 10 (control) and 15 (VSP).
8 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1422941112 Li et al.

complex patterning of signaling within networks. However, fur-ther characterization of cnidarian and ctenophore channels, neu-rons, and circuits on functional level will be needed to betterunderstand how the evolution of voltage-gated K+ channels fitsinto the broader picture of the evolution of neuronal function.
MethodsGene Identification, Cloning, and Phylogenetic Analysis. Cnidarian and Mne-miopsis Shaker and KCNQ family K+ channel genes described in this studywere identified and compiled through comprehensive BLAST (46) searchesof genome drafts, transcriptomes, and gene predictions ofMnemiopsis leidyi(41) and Nematostella vectensis (73), and for KCNQ only, Hydra magnipa-pillata (66), and Acropora digitifera (74) and Orbicella faveolata. Multiplebilaterian members of each channel type were used as query sequences, andreciprocal BLAST searches of identified sequences against bilaterian data-bases were used to classify the sequences before phylogenetic analysis. Mostqueries identified all voltage-gated K+, Na+, and Ca2+ channels, but re-ciprocal searches sorted target sequences by gene family and were used torefine gene predictions when necessary.
Alignments for phylogenetic analysis were produced using the MuscleAlgorithm as implemented in MEGA 6 (75), and regions of low lengthconservation were trimmed before phylogenetic analysis. Shaker familyalignments included the N-terminal T1 domain and S1–S6 voltage-gated K+
channel core. KCNQ alignments contained the channel core and KCNQ-specific coiled-coil region of the C terminus. Sequences used for phyloge-netic analysis are provided in Dataset S1 (Shaker) and Dataset S2 (KCNQ).Bayesian inference phylogenies of channel proteins were constructed usingMrBayes (76). Analyses were run under a mixed model for 1 million gen-erations with two runs of four chains. The first 25% of the analysis wasdiscarded as a burn in phase, and the displayed phylogenies are the con-sensus. The convergence diagnostic potential scale reduction factor was1.000 for Shaker and 0.999 for KCNQ, indicating appropriate convergenceof independent runs. Minimum-evolution, maximum-likelihood, and SSNanalyses are described in SI Methods.
Nematostella and Mnemiopsis channel genes were cloned using standardRT-PCR techniques from total RNA samples from whole adult animals.Briefly, 500 ng to 2 μg of total RNA was reverse transcribed using oligo-dT,and gene-specific primers were used to amplify coding regions. When nec-essary, overlap extension PCR was used to assemble full-length codingregions from overlapping fragments. Coding regions were cloned into thepOX vector (23) for expression in Xenopus oocytes and sequence confirmed.Consensus DNA coding sequences are reported in Dataset S3, and onlyclones matching these sequences were used for expression analysis.
Electrophysiology. Xenopus laevis ovaries were obtained from Xenopus I.Mature oocytes were enzymatically defolliculated with type II collagenase
(Sigma-Aldrich) at 1 mg/mL in Ca2+-free ND98 solution (98 mM NaCl, 2 mMKCl, 1 mM MgCl2, 5 mM Hepes, pH 7.2). Following digestion, oocytes weremaintained in ND98 culture solution (98 mM NaCl, 2 mM KCl, 1.8 mM CaCl2,1 mM MgCl2, 2.5 mM Na-pyruvate, 100 U/mL penicillin, 100 μg/mL strepto-mycin, and 5 mM Hepes, pH 7.2). Capped, polyadenylated cRNA transcriptswere generated from linearized expression plasmids using T3 mMessagemMachine kit and Poly-A tailing kit (Life Technologies). LiCl precipitationwas used to purify transcripts, and pellets were resuspended in nuclease-freewater supplemented with RNase inhibitor. Transcripts were injected in 50-nLvolumes into mature oocytes, which were incubated at 18 °C in ND98 culturesolution for 1–3 d before recording.
Two-electrode voltage-clamp recordings were carried out at room tem-perature (22–24 °C) using a Dagan CA-1B amplifier, and data were collectedand analyzed with the pClamp 10 acquisition suite (Molecular Devices).Recordings were performed under constant perfusion (98 mM NaOH, 2 mMKCl, 1 mMCaCl2, 1 mMMgCl2, and 5 mMHepes, pH 7.0). Electrodes (0.4–1MΩ)were filled with 3 M KCl and bath-clamp circuitry was isolated with a 1 MNaCl/agarose bridge. Currents were sampled at 10 kHz and filtered at 2 kHz.Hardware capacitance and leak compensation was used in some recordings.
Data were fit and plotted in Origin 8.1 (OriginLab). Single-Boltzmanndistributions were fit using the equation f(V) = A2 + (A1 − A2)/(1+e
(V−V50)/s),where V50 is the half-maximal activation voltage, s is the slope factor, andA1 and A2 are the lower and upper asymptotes, respectively. Double-Boltzmanndistributions were fit using the equation f(V ) = y0 + A[p/(1 + e(v−x01)/s1) +(1 − p)/(1 + e(v−x02)/s2)], where y0 is the current offset, A is the amplitude span,and p is the fraction of current component in the more hyperpolarizedvoltage range. Data points show averages of normalized values for individualcells. Arithmetic means of V50 and s values from individual measurementswere used to generate the Boltzmann fits shown in figures. Recovery timecourse for NvShal channels was fit with two exponentials using the equationf(t) = A1*exp(−x/t1) + A2*exp(−x/t2) + y0 (x, time interval of recovery; t1 and t2,time constants of recovery; A1 and A2, amplitudes of the recovering com-ponents; y0, current offset). Peak current amplitudes measured from testpulses were normalized to the peak current amplitude of the first pulse. Datapoints are averages of measurements from individual cells, and fits shownwere generated with arithmetic means of fit parameters.
Statistical comparisons between datasets were carried out using two-tailed t test.
ACKNOWLEDGMENTS. J.C.L. and L.M.T. were supported by Penn StateDiscovery Grants, F.H.D. received undergraduate support from the EberlyCollege of Science, and T.J. received support from the Departmentof Biology, Huck Institute of Life Sciences, and NIH Grant R01 NS069842.D.B.v.R. was supported by a grant from the Pennsylvania Department ofHealth using Tobacco Settlement Funds.
1. Jegla TJ, Zmasek CM, Batalov S, Nayak SK (2009) Evolution of the human ion channelset. Comb Chem High Throughput Screen 12(1):2–23.
2. Yu FH, Catterall WA (2004) The VGL-chanome: A protein superfamily specialized forelectrical signaling and ionic homeostasis. Sci STKE 2004(253):re15.
3. Kreusch A, Pfaffinger PJ, Stevens CF, Choe S (1998) Crystal structure of the tetrame-rization domain of the Shaker potassium channel. Nature 392(6679):945–948.
4. Shen NV, Pfaffinger PJ (1995) Molecular recognition and assembly sequences involvedin the subfamily-specific assembly of voltage-gated K+ channel subunit proteins.Neuron 14(3):625–633.
5. Howard RJ, Clark KA, Holton JM, Minor DL, Jr (2007) Structural insight into KCNQ(Kv7) channel assembly and channelopathy. Neuron 53(5):663–675.
6. Schwake M, et al. (2006) Structural determinants of M-type KCNQ (Kv7) K+ channelassembly. J Neurosci 26(14):3757–3766.
7. Jegla T, Salkoff L (1994) Molecular evolution of K+ channels in primitive eukaryotes.Soc Gen Physiol Ser 49:213–222.
8. Wei A, Jegla T, Salkoff L (1996) Eight potassium channel families revealed by theC. elegans genome project. Neuropharmacology 35(7):805–829.
9. Salkoff L, et al. (1992) An essential “set” of K+ channels conserved in flies, mice andhumans. Trends Neurosci 15(5):161–166.
10. Xu J, Yu W, Jan YN, Jan LY, Li M (1995) Assembly of voltage-gated potassium chan-nels. Conserved hydrophilic motifs determine subfamily-specific interactions betweenthe alpha-subunits. J Biol Chem 270(42):24761–24768.
11. Ogawa Y, et al. (2008) Postsynaptic density-93 clusters Kv1 channels at axon initialsegments independently of Caspr2. J Neurosci 28(22):5731–5739.
12. Ogawa Y, et al. (2010) ADAM22, a Kv1 channel-interacting protein, recruits membrane-associated guanylate kinases to juxtaparanodes of myelinated axons. J Neurosci 30(3):1038–1048.
13. Wang H, Kunkel DD, Martin TM, Schwartzkroin PA, Tempel BL (1993) Hetero-multimeric K+ channels in terminal and juxtaparanodal regions of neurons. Nature365(6441):75–79.
14. Rosenthal JJ, Vickery RG, Gilly WF (1996) Molecular identification of SqKv1A. A can-didate for the delayed rectifier K channel in squid giant axon. J Gen Physiol 108(3):207–219.
15. Jegla T, Grigoriev N, Gallin WJ, Salkoff L, Spencer AN (1995) Multiple Shaker potas-sium channels in a primitive metazoan. J Neurosci 15(12):7989–7999.
16. Jegla T, et al. (2012) Expanded functional diversity of shaker K+ channels in cnidariansis driven by gene expansion. PLoS One 7(12):e51366.
17. Carrasquillo Y, Nerbonne JM (2014) IA channels: Diverse regulatory mechanisms.Neuroscientist 20(2):104–111.
18. Tsunoda S, Salkoff L (1995) The major delayed rectifier in both Drosophila neuronsand muscle is encoded by Shab. J Neurosci 15(7 Pt 2):5209–5221.
19. Tsunoda S, Salkoff L (1995) Genetic analysis of Drosophila neurons: Shal, Shaw, andShab encode most embryonic potassium currents. J Neurosci 15(3 Pt 1):1741–1754.
20. Misonou H, Mohapatra DP, Trimmer JS (2005) Kv2.1: A voltage-gated K+ channelcritical to dynamic control of neuronal excitability. Neurotoxicology 26(5):743–752.
21. Du J, Haak LL, Phillips-Tansey E, Russell JT, McBain CJ (2000) Frequency-dependentregulation of rat hippocampal somato-dendritic excitability by the K+ channel sub-unit Kv2.1. J Physiol 522(Pt 1):19–31.
22. Guan D, Armstrong WE, Foehring RC (2013) Kv2 channels regulate firing rate in py-ramidal neurons from rat sensorimotor cortex. J Physiol 591(Pt 19):4807–4825.
23. Jegla T, Salkoff L (1997) A novel subunit for shal K+ channels radically alters activationand inactivation. J Neurosci 17(1):32–44.
24. Lien CC, Jonas P (2003) Kv3 potassium conductance is necessary and kinetically opti-mized for high-frequency action potential generation in hippocampal interneurons.J Neurosci 23(6):2058–2068.
25. Wang LY, Gan L, Forsythe ID, Kaczmarek LK (1998) Contribution of the Kv3.1 po-tassium channel to high-frequency firing in mouse auditory neurones. J Physiol509(Pt 1):183–194.
26. Wei A, et al. (1990) K+ current diversity is produced by an extended gene familyconserved in Drosophila and mouse. Science 248(4955):599–603.
Li et al. PNAS Early Edition | 9 of 10
NEU
ROSC
IENCE
PNASPL
US

27. Hodge JJ, Choi JC, O’Kane CJ, Griffith LC (2005) Shaw potassium channel genes inDrosophila. J Neurobiol 63(3):235–254.
28. Johnstone DB, Wei A, Butler A, Salkoff L, Thomas JH (1997) Behavioral defects inC. elegans egl-36 mutants result from potassium channels shifted in voltage-dependence of activation. Neuron 19(1):151–164.
29. Sand RM, Atherton DM, Spencer AN, Gallin WJ (2011) jShaw1, a low-threshold,fast-activating Kv3 from the hydrozoan jellyfish Polyorchis penicillatus. J Exp Biol214(Pt 18):3124–3137.
30. Wang HS, et al. (1998) KCNQ2 and KCNQ3 potassium channel subunits: Molecularcorrelates of the M-channel. Science 282(5395):1890–1893.
31. Peters HC, Hu H, Pongs O, Storm JF, Isbrandt D (2005) Conditional transgenic sup-pression of M channels in mouse brain reveals functions in neuronal excitability,resonance and behavior. Nat Neurosci 8(1):51–60.
32. Brown DA, Passmore GM (2009) Neural KCNQ (Kv7) channels. Br J Pharmacol 156(8):1185–1195.
33. Delmas P, Brown DA (2005) Pathways modulating neural KCNQ/M (Kv7) potassiumchannels. Nat Rev Neurosci 6(11):850–862.
34. Zaydman MA, Cui J (2014) PIP2 regulation of KCNQ channels: Biophysical and mo-lecular mechanisms for lipid modulation of voltage-dependent gating. Front Physiol5:195.
35. ZaydmanMA, et al. (2013) Kv7.1 ion channels require a lipid to couple voltage sensingto pore opening. Proc Natl Acad Sci USA 110(32):13180–13185.
36. Kubisch C, et al. (1999) KCNQ4, a novel potassium channel expressed in sensory outerhair cells, is mutated in dominant deafness. Cell 96(3):437–446.
37. Wickenden AD, Zou A, Wagoner PK, Jegla T (2001) Characterization of KCNQ5/Q3potassium channels expressed in mammalian cells. Br J Pharmacol 132(2):381–384.
38. Zhang H, et al. (2003) PIP2 activates KCNQ channels, and its hydrolysis underliesreceptor-mediated inhibition of M currents. Neuron 37(6):963–975.
39. Wen H, et al. (2005) A Drosophila KCNQ channel essential for early embryonic de-velopment. J Neurosci 25(44):10147–10156.
40. Wei AD, Butler A, Salkoff L (2005) KCNQ-like potassium channels in Caenorhabditiselegans. Conserved properties and modulation. J Biol Chem 280(22):21337–21345.
41. Ryan JF, et al.; NISC Comparative Sequencing Program (2013) The genome of thectenophore Mnemiopsis leidyi and its implications for cell type evolution. Science342(6164):1242592.
42. Moroz LL, et al. (2014) The ctenophore genome and the evolutionary origins ofneural systems. Nature 510(7503):109–114.
43. Dunn CW, et al. (2008) Broad phylogenomic sampling improves resolution of theanimal tree of life. Nature 452(7188):745–749.
44. Ryan JF, Pang K, Mullikin JC, Martindale MQ, Baxevanis AD; NISC ComparativeSequencing Program (2010) The homeodomain complement of the ctenophoreMnemiopsis leidyi suggests that Ctenophora and Porifera diverged prior to theParaHoxozoa. Evodevo 1(1):9.
45. Ryan JF (2014) Did the ctenophore nervous system evolve independently? Zoology(Jena) 117(4):225–226.
46. Altschul SF, et al. (1997) Gapped BLAST and PSI-BLAST: A new generation of proteindatabase search programs. Nucleic Acids Res 25(17):3389–3402.
47. Fairclough SR, et al. (2013) Premetazoan genome evolution and the regulation of celldifferentiation in the choanoflagellate Salpingoeca rosetta. Genome Biol 14(2):R15.
48. King N, et al. (2008) The genome of the choanoflagellate Monosiga brevicollis andthe origin of metazoans. Nature 451(7180):783–788.
49. Lee SY, Lee A, Chen J, MacKinnon R (2005) Structure of the KvAP voltage-dependentK+ channel and its dependence on the lipid membrane. Proc Natl Acad Sci USA102(43):15441–15446.
50. Sentenac H, et al. (1992) Cloning and expression in yeast of a plant potassium iontransport system. Science 256(5057):663–665.
51. Schachtman DP, Schroeder JI, Lucas WJ, Anderson JA, Gaber RF (1992) Expression ofan inward-rectifying potassium channel by the Arabidopsis KAT1 cDNA. Science258(5088):1654–1658.
52. Ketchum KA, Joiner WJ, Sellers AJ, Kaczmarek LK, Goldstein SA (1995) A new familyof outwardly rectifying potassium channel proteins with two pore domains in tan-dem. Nature 376(6542):690–695.
53. Srivastava M, et al. (2010) The Amphimedon queenslandica genome and the evolu-tion of animal complexity. Nature 466(7307):720–726.
54. Riesgo A, Farrar N, Windsor PJ, Giribet G, Leys SP (2014) The analysis of eight tran-scriptomes from all poriferan classes reveals surprising genetic complexity in sponges.Mol Biol Evol 31(5):1102–1120.
55. Bouchard C, et al. (2006) Cloning and functional expression of voltage-gated ionchannel subunits from cnidocytes of the Portuguese Man O’War Physalia physalis.J Exp Biol 209(Pt 15):2979–2989.
56. Quattrocki EA, Marshall J, Kaczmarek LK (1994) A Shab potassium channel contrib-utes to action potential broadening in peptidergic neurons. Neuron 12(1):73–86.
57. Davis MW, Fleischhauer R, Dent JA, Joho RH, Avery L (1999) A mutation in theC. elegans EXP-2 potassium channel that alters feeding behavior. Science 286(5449):2501–2504.
58. Post MA, Kirsch GE, Brown AM (1996) Kv2.1 and electrically silent Kv6.1 potassiumchannel subunits combine and express a novel current. FEBS Lett 399(1-2):177–182.
59. Ottschytsch N, Raes A, Van Hoorick D, Snyders DJ (2002) Obligatory hetero-tetramerization of three previously uncharacterized Kv channel alpha-subunitsidentified in the human genome. Proc Natl Acad Sci USA 99(12):7986–7991.
60. Fawcett GL, et al. (2006) Mutant analysis of the Shal (Kv4) voltage-gated fast transientK+ channel in Caenorhabditis elegans. J Biol Chem 281(41):30725–30735.
61. Bähring R, Covarrubias M (2011) Mechanisms of closed-state inactivation in voltage-gated ion channels. J Physiol 589(Pt 3):461–479.
62. Tang CY, Schulteis CT, Jiménez RM, Papazian DM (1998) Shaker and ether-à-go-go K+
channel subunits fail to coassemble in Xenopus oocytes. Biophys J 75(3):1263–1270.63. Zou A, et al. (2003) Distribution and functional properties of human KCNH8 (Elk1)
potassium channels. Am J Physiol Cell Physiol 285(6):C1356–C1366.64. Wimmers S, Wulfsen I, Bauer CK, Schwarz JR (2001) Erg1, erg2 and erg3 K channel
subunits are able to form heteromultimers. Pflugers Arch 441(4):450–455.65. Srivastava M, et al. (2008) The Trichoplax genome and the nature of placozoans.
Nature 454(7207):955–960.66. Chapman JA, et al. (2010) The dynamic genome of Hydra. Nature 464(7288):592–596.67. Murata Y, Okamura Y (2007) Depolarization activates the phosphoinositide phos-
phatase Ci-VSP, as detected in Xenopus oocytes coexpressing sensors of PIP2. J Physiol583(Pt 3):875–889.
68. Loijens JC, Boronenkov IV, Parker GJ, Anderson RA (1996) The phosphatidylinositol4-phosphate 5-kinase family. Adv Enzyme Regul 36:115–140.
69. Hernandez CC, Zaika O, Shapiro MS (2008) A carboxy-terminal inter-helix linker as thesite of phosphatidylinositol 4,5-bisphosphate action on Kv7 (M-type) K+ channels.J Gen Physiol 132(3):361–381.
70. Martinson AS, et al. (2014) Functional evolution of Erg potassium channel gatingreveals an ancient origin for IKr. Proc Natl Acad Sci USA 111(15):5712–5717.
71. Moran Y, Zakon HH (2014) The evolution of the four subunits of voltage-gatedcalcium channels: Ancient roots, increasing complexity, and multiple losses. GenomeBiol Evol 6(9):2210–2217.
72. Covarrubias M, Wei AA, Salkoff L (1991) Shaker, Shal, Shab, and Shaw express in-dependent K+ current systems. Neuron 7(5):763–773.
73. Putnam NH, et al. (2007) Sea anemone genome reveals ancestral eumetazoan generepertoire and genomic organization. Science 317(5834):86–94.
74. Shinzato C, et al. (2011) Using the Acropora digitifera genome to understand coralresponses to environmental change. Nature 476(7360):320–323.
75. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular Evo-lutionary Genetics Analysis version 6.0. Mol Biol Evol 30(12):2725–2729.
76. Ronquist F, et al. (2012) MrBayes 3.2: Efficient Bayesian phylogenetic inference andmodel choice across a large model space. Syst Biol 61(3):539–542.
10 of 10 | www.pnas.org/cgi/doi/10.1073/pnas.1422941112 Li et al.
Copyright © 2022 FDOKUMEN




















