
Investigation of immunoglobulins in skin of the Antarctic teleost Trematomus bernacchii
Upload
independentCategory
view
1download
0
BritishJournai of Haernatology, 1980,45,405-416.
Lymphoid Cell Sets and Serum Immunoglobulins in Patients with Thalassaemia Intermedia:
Relationship to Serum Iron and Splenectomy ASHA KAPADIA, MARIA DE SOUSA, ALICEJANE L. MARKENSON,
DENIS R. MILLER, ROBERT A. GOOD AND SUDHIR C U ~ A
Memorial Sloan-Kettering Cancer Center and N e w York Hospital Cornell Medical Center
(Received 2 October 1979; acceptedfor publication 18 December 1979)
SUMMARY. Peripheral blood lymphocyte sets and serum immunoglobulins were examined in 15 patients with thalassaemia intermedia, aged between 5 and 30 years. They were divided in three clinical groups: Group I, non-splenectomized, with serum iron (SI) lower than 200 pg/dl; Group 11, splenectomized with serum iron lower than 200 pg/dl; and Group 111, splenectomized with serum iron levels greater than 200 pg/dl. High absolute lymphocyte counts were observed in the majority of patients, whether splenectomized or not. Following splenectomy, marked increases were observed in absolute number of lymphocytes and percentages of circulating surface immunoglobulin (SIg) bearing B lymphocytes, mouse rosette forming cells (MRFC) and ‘null’ cells. The increases in circulating B lymphocytes following splenectomy were due exclusively to cells bearing SIgM and/or SIgD. No changes were observed in numbers and proportions of circulating SIgA or SIgG bearing cells. Serum IgA levels were high in the majority of patients studied whether splencctomized or not. Following splenectomy incrcases in serum IgA and m u m IgG were observed which seemed to relate to increasing SI levels. No significant changes occurred in serum IgM levels.
Absolute numbers of circulating T lymphocytes were unaffected by splenectomy or SI levels. Changes were observed, however, in distribution of T lymphocyte subsets with decreases in number ofTp cells and concomitant increases in numbers of non-Tp, non-Ty cells. T y cells were the least affected by splencctomy and increasing SI. The results are compatible with the proposition that iron participates in the rcgulation of lymphoid cell migration and in the modulation of expression of cell surface markers. The findings also illustrate the role of the spleen in the control of lymphocyte migration and immunoglobulin production.
Recent studies of lymphoid tissues and immunological functions in clinical and experimental situations of iron deficiency indicate that iron plays a role in the control of thymus lympho- poiesis (Chandra et al, 1977) and is necessary for normal T lymphocyte function (Joynson et al,
U.S.A. Correspondence: Dr Maria de Sousa, Sloan Kettering Institute, 1275 York Avenue, New York, NY 10021,
0007-1048/80/0700-040S $02.00 8 1980 Blackwell Scientific Publications
405

406 Asha Kapadia et a1
1972; MacDougall et al, 1975; Bashkaram & Reddy, 1975) and mucosal IgA synthesis (Chandra e t a l , 1977).
The mechanisms whereby iron deficiency relates to reduced T cell numbers, defective T cell-mediated immunity and mucosal IgA synthesis are not clear. The fact that iron is needed in DNA synthesis for ribonucleotide reductase (Larsson, 1969; Hoarand et al, 1976) has been suggested as a possible cause (Hoffbrand, 1977). One possible additional mechanism relates to the proposed role played by iron in the control of migration and tissue distribution of lymphoid cells (de Sousa, 1978a; de Sousa et al, 1978). According to this proposal, poor lymphocyte migration to peripheral sites would be expected in clinical situations of iron deficiency; in contrast, an increased migration of peripheral lymphocytes would be expected in situations of iron overload, such as thalassaemia.
In the present paper we examine the distribution of the three major populations of human lymphocytes, namely T lymphocytes, B lymphocytes and so-called ‘third population’ cells, and of two sets of T lymphocytes characterized by Fc receptors for IgM (Tp) or IgG (Ty) (Gupta & Good, 1977a, b) in a group of patients with high serum iron and high serum ferritin levels associated with thalassaemia intermedia.
Although the patients with thalassaemia intermedia have clinical problems ofiron overload, increased iron absorption and hypersplenism, similar to those present in thalassaemia major, they were selected for the present study because of a less severe anaemia and only occasional requirement for blood transfusions.
SUBJECTS
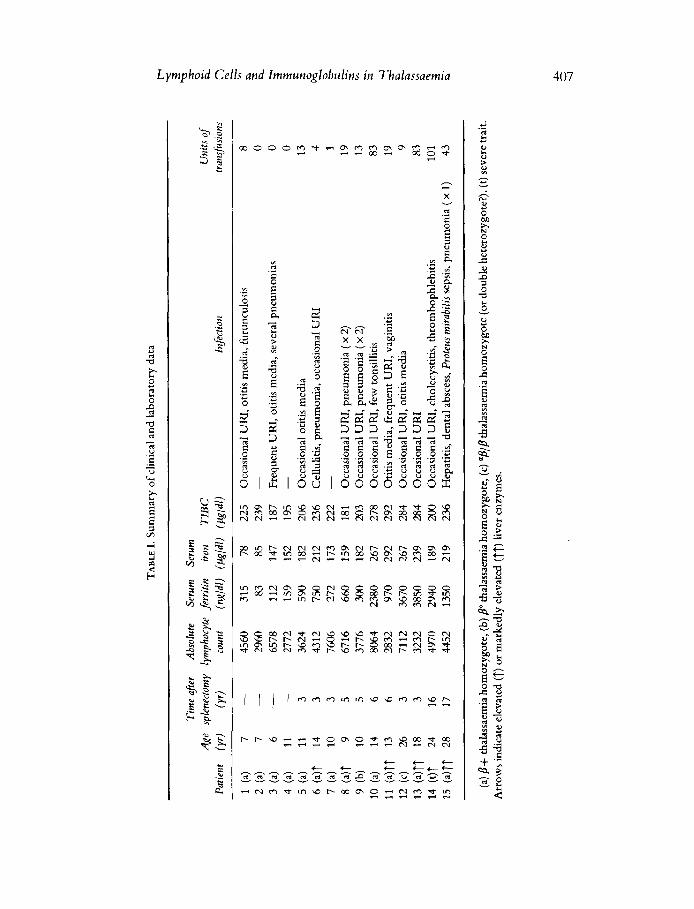
Fifteen patients with thalassaemia intermedia and one patient with thalassaemia major (23 years, female) were studied. Diagnosis was made on the basis of the finding of thalassaemia microcytosis in both parents and hypochromic microcytic anaemia and hepatosplenomegaly in the patient. The patients with thalassaemia intermedia fell into three clinical subgroups: Group 1, included four patients aged 5-15 years who had not undergone splenectomy; Group 2, included seven patients in the same age group who had been splenectomized; Group 3, included four patients aged 15-30 years who had been splenectomized (Table I).
A summary of clinical and laboratory data on these patients is presented in Table I. To discriminate between the effects attributable to splenectomy and those associated with iron overload, the results are grouped according to serum iron levels or splenectomy (Figs 1-4).
Isolation of Mononuclear Cel ls Mononuclear cells were isolated from peripheral blood on Ficoll-Hypaque (FH) gradients,
washed three times in Hanks’ balance salt solution (HBSS) and resuspended at a concentration of 5 x lo6 cellslml in medium containing 20% heat-inactivated fetal calf serum (FCS). Phago- cytic cells were identified by their ability to ingest 0.8 pm polystyrene latex particles (Dow Chemicals, Indianapolis, Ind.). Cells were washed three times and resuspended in HBSS to a final concentration of 5 x 106 cells/ml. Viability of cells tested with trypan blue dye exclusion was regularly greater than 98%.
Sheep Erythrocyte Rosette Forming T Cells (ERFC) T lymphocytes were identified by their capacity to form spontaneous rosettes with sheep red
1 (a
) 7
-
2 (a
) 7
-
3 (a
) 6
-
4 (a
) 11
-
5 (a
) 11
3
6 14
3
7 (a
) 10
3
8 (a
)t
9 5
9 (b
) 10
5
10 (
a)
14
6 11
(.)I
t 13
6
13 (a
)tt
18
3 14
(t)
t 24
16
15
(a)
Tt
28
17
12 (
c)
26
3
4560
29
60
6578
27
72
3624
43
12
7606
67
16
3776
80
64
2832
71
12
3232
49
70
4452
315 83
11
2 15
9 5%
75
0 27
2 66
0 30
0 23
80
970
3670
38
50
2940
13
50
78
85
147
152
182
212
173
159
182
267
292
267
239
189
219
TA
BL
E
I. Su
mm
ary
of c
linic
al a
nd la
bora
tory
dat
a t-r
3
Y
Tim
e uje
r Ab
solu
te
Seru
m
Seru
m
2-
Age
spl
enec
tom
y ly
mph
ocyt
e ferritin
iron
TIB
C
Uni
ts of
8. 2
Infe
ction
tra
nsfu
sions
Pu
atien
t (y
r)
(yr)
cm
nt
(flg
ldll
(Pg
W
(figg
ldl)
T
225
Occ
asio
nal U
RI,
otiti
s m
edia
, fur
uncu
losi
s 8
n
239
187
195
206
236
222
181
203
278
292
284
284
200
236
.- 3
-
0 S
L
3 2 Fr
eque
nt U
RI,
otit
is m
edia
, sev
eral
pne
umon
ias
Occ
asio
nal o
titis
med
ia
13
3
0 0 -
Cel
lulit
is, p
neum
onia
, occ
asio
nal U
RI
4 % 0
-
L
1 13
-
- O
ccas
iona
l UR
I, pn
eum
onia
( x
2)
19
-.
2 O
ccas
iona
l UR
I, f
ew to
nsill
itis
83
Occ
asio
nal U
RI,
otit
is m
edia
9
Occ
asio
nal U
RI,
cho
lecy
stiti
s, th
rom
boph
lebi
tis
101
Hep
atiti
s, d
enta
l abs
cess
, Pro
teus
miru
bilis
seps
is, p
neum
onia
( x
1)
-.
Occ
asio
nal U
RI,
pneu
mon
ia (
x 2
) 3
Otit
is m
edia
, fre
quen
t U
RI,
vag
initi
s 19
2 2. n w
rri - n :
Occ
asio
nal U
RI
83
43
P
(a) fl
+ th
alas
saem
ia h
omoz
ygot
e, (
b) ,9 t
hala
ssae
rnia
hom
ozyg
ote,
(c) "8
18 th
alas
saem
ia h
omoz
ygot
e (o
r dou
ble
hete
rozy
gote
?), (
t) se
vere
trai
t. A
rrow
s in
dica
te el
evat
ed (7
) or
mar
kedl
y el
evat
ed (1
1) liv
er e
nzym
es.
408 Asha Kapadia et al
T. ERFC TY
s - - s - - ‘U 1-
s - - 1 U
s - - ~ 2 0 0 ~ 2 0 0 e200 *200 <200>200 <200 >200
Serum iron concentration (pg/dl)
U U
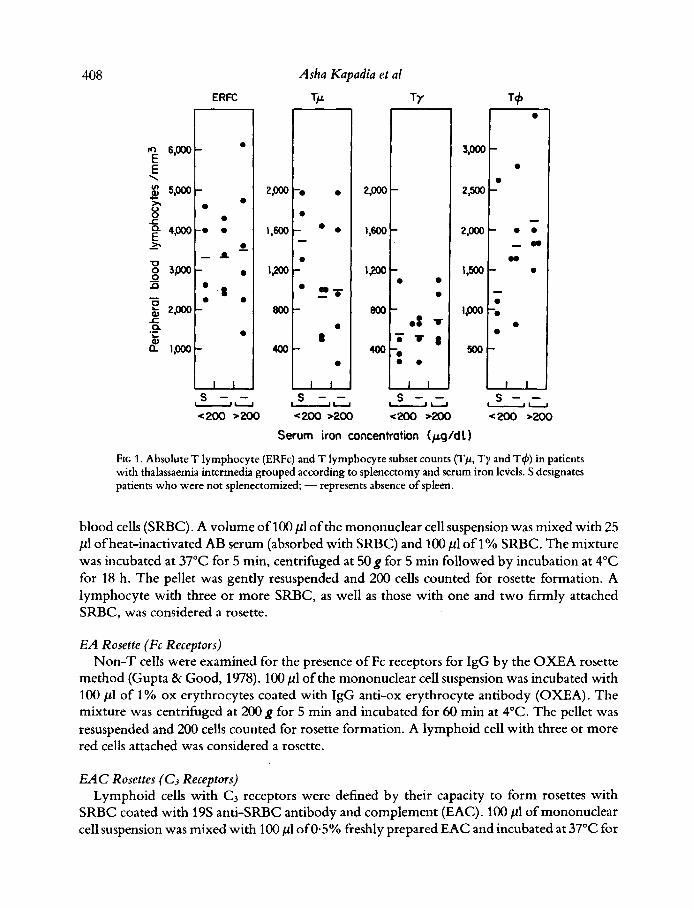
FIG 1. Absolute T lymphocyte (ERFc) and T lymphocyte subset counts (Tp, Ty and T4) in patients with thalassaemia intermedia grouped according to splenectomy and serum iron levels. S designates patients who were not splenectomized; - represents absence of spleen.
blood cells (SRBC). A volume of 100 pl of the mononuclear cell suspension was mixed with 25 p1 of heat-inactivated AB serum (absorbed with SRBC) and 100 pl of 1% SRBC. The mixture was incubated at 37°C for 5 min, centrifuged at 50 g for 5 min followed by incubation a t 4°C for 18 h. The pellet was gently resuspended and 200 cells counted for rosette formation. A lymphocyte with three or more SRBC, as well as those with one and two firmly attached SRBC, was considered a rosette.
E A Rosette (Fc Receptors) Non-T cells were examined for the presence of Fc receptors for IgG by the OXEA rosette
method (Gupta & Good, 1978). 100 pl of the mononuclear cell suspension was incubated with 100 p1 of 1% ox erythrocytes coated with IgG anti-ox erythrocyte antibody (OXEA). The mixture was centrifuged at 200 g for 5 min and incubated for 60 min at 4°C. The pellet was resuspended and 200 cells counted for rosette formation. A lymphoid cell with three or more red cells attached was considered a rosette.
EAC Rosettes (C3 Receptors) Lymphoid cells with C , receptors were defined by their capacity to form rosettes with
SRBC coated with 19s anti-SRBC antibody and complement (EAC). 100 p1 of mononuclear cell suspension was mixed with 100 pl of 0.5% freshly prepared EAC and incubated at 37°C for
Lymphoid Cells and lmmunoglobulins in Thalassaemia 409
Pv M G
300
200
I 0 0
s - - 1 u s - - .U
s - - s - - 'U 1 u
A
<200 s200 <200>200 <200 ~200 <xx) >aM) <aM) w Serum iron cmentmtion ( p g l d l )
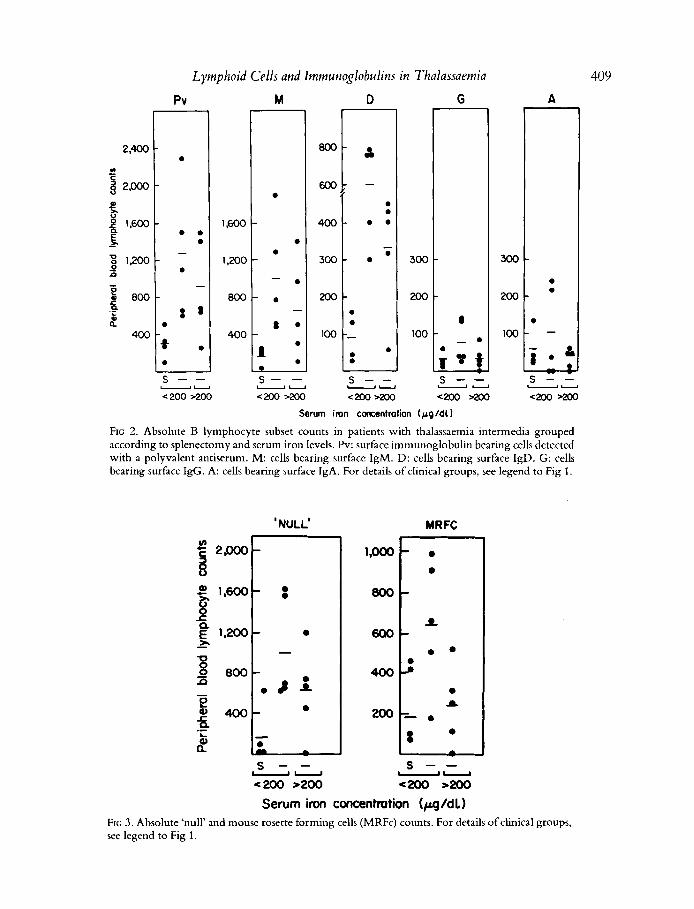
FIG 2. Absolute B lymphocyte subset counts in patients with thalassaemia interrnedia grouped according to splenectomy and serum iron levels. Pv: surface immunoglobulin bearing cells detected with a polyvalent antiserum. M: cells bearing surface IgM. D: cells bearing surface IgD. G: cells bearing surface IgG. A: cells bearing surface IgA. For details of clinical groups, see legend to Fig 1 .
'NULL' MRFC
Serum iron concentration (pg/dl) FIG 3 . Absolute 'null' and mouse rosette forming cells (MRFc) counts. For details of clinical groups, see legend to Fig 1.

410 Asha Kapadia et a1
IgG
3,000
s - - 4200 ~ 2 0 0 .Y
s - - 4200 >Mo - Y
Serum iron concentmtion
FIG 4. Circulating levels of serum immunoglobulins in patients with thalassaemia intermedia. For details of clinical groups see legend to Fig 1. Shaded areas represent normal range (mean 1 SD).
30 min. The pellet was resuspended and 200 cells were counted. A lymphoid cell with three or more RBC attached was considered a rosette.
Mouse Erythrocyte Rosette Forming Cells ( M R F C ) Mouse erythrocyte rosette-forming cells (a subpopulation of B cells) were assayed by the
technique described previously (Gupta & Good, 1978). 100 pl of mononuclear cell suspension was mixed with 25 p1 of heat-inactivated FCS (absorbed with MRBC) and 100 p1 of 1% MRBC. The mixture was centrifuged at 200 g for 5 min followed by incubation in a water bath a t 28°C for 1 h. The pellet was gently resuspended and a drop of trypan blue added. Two hundred cells were counted for rosette formation. A lymphocyte with three or more MRBC attached was considered a rosette.
Surface Immunoglobulins heavy chain determinants of immunoglobulin
conjugated with fluorescein isothiocyanate were used. Mononuclear cells (0.5-1 x 106/tube) were washed with cold 2% bovine serum albumin in phosphate buffered saline (BSA/PBS) containing 0.2% sodium azide and incubated with 25 pl of the antiserum. Staining was carried out at 4°C for 30 min. Cells were washed three times with BSA/PBS and resuspended in a small volume of BSA/PBS. Wet mounts were prepared and 200400 cells were counted under a Leitz orthoplan microscope equipped with Ploem fluorescence epi-illumination.
Antisera monospecific for p, 6 , CL or

Lymphoid Cells and Immunoglobulins in Thalassaemia 41 1
‘Null’ Cells Cells lacking any identifiable surface markers including Fc receptors were designated as
‘null’ cells. Their proportions were enumerated by the formula: lW-[ERFC (%) + EA rosettes (%) 1.
Determination of ‘I‘ Lymphocyte Subpopulations Purlfcation o f T lymphocytes. T lymphocytes were purified from non-T cells by rosetting
with SRBC, as described above, with a minor modification; instead ofan 18 h incubation, cells were incubated for 1 h with 1 % neuraminidase-treated SRBC and separated from non-roset- ting (non-T) cells on a FH gradient. SRBC attached to cells were lysed by using Tris-buffer with 0.83% ammonium chloride. T lymphocytes obtained in this way were more than 98% pure, as determined by rosette formation with SRBC and lack of cells with readily demon- strable surface immunoglobulin. cells were washed three times with PBS and resuspended in RPMI 1640 (Grand Island Biological Co., Grand Island, N.Y.) containing penicillin (100 U/ml), streptomycin (100 pg/ml) and 20% FCS at a final concentration of 4 x 166 cells/ml. These cells were cultured at 37°C in a humidified incubator with 5% COz and 95% air for 20 h. At the end of incubation cells were washed three times in HBSS and resuspended in HBSS to a concentration of 4 x 106/ml. More than 98% of cells were viable as tested by trypan blue die exclusion. Purified T cells were assayed for the proportion of Tp and Ty cells.
Preparation of ox RBC-antibody complexes. Anti-ox RBC antibodies were prepared by the technique previously described (Gupta & Good, 1977a). Ox RBC were washed three times in HBSS and resuspended to a concentration of 2%. Equal volumes of ox RBC and purified IgM anti-ox RBC antibody (1125 dilution) or IgG anti-ox RBC antibody (1/100 dilution) were mixed and incubated at room temperature for 120 min. Following incubation, cells were washed three times in HBSS and resuspended to a final concentration of 1.0%. Ox RBC with IgM (EAm) or IgG (EAg) were prepared fresh each time.
T cells with receptors for ZgM (Tp). 100 p1 of the T lymphocyte suspension were mixed with 100 pl of EAm, centrifuged a t 200g for 5 min and incubated at 4°C for 60 min. The pellet was resuspended and 200 lymphocytes counted for rosette formation. A lymphocyte with three or more red cells attached was considered a rosette.
T cells with receptorsfor ZgC (Ty). 100 pl of the T lymphocyte suspension were mixed with 100 pl of EAg, centrifuged at 200g for 5 min and incubated at 4°C for 30 min. The pellet was resuspended and 200 lymphocytes counted for rosette formation. A rosette was defined as a lymphocyte with three or more red cells attached.
T cells lacking either of the immunoglobulin receptors are designated as T&J. The results of Tp and Ty cells are presented aa a per cent of T lymphocytes.
Serum Ferritin Serum ferritin levels were measured by means of a Ramco-Kit provided by Ramco
Laboratory, Houston, Texas, with serum dilution of 1 : 1000. The statistical analysis was done by unpaired Student’s t test.
RESULTS
The detailed results of analysis of distribution of lymphocyte subsets and immunoglobulins in

41 2 Asha Kupudiu et al
each individual patient are presented in Figs 1-4. To discriminate between the effects related to splenectomy and those associated with iron overload, the results have been grouped according to serum iron levels and whether or not splenectomy had been performed. The value for serum iron of 200 ,ug/dl was chosen arbitrarily as the dividing point between groups. With the exception of patient 14, all patients with serum iron levels lower than 200 yg/dl had serum ferritin levels less than under 660 ng/ml (Table I). Patients with serum iron lower than 200 pg/dl were further subdivided into those who had (patients 5, 7, 8, 9 and 14) or had not (patients 1-4) undergone splenectomy. All patients with serum iron levels higher than 200 pg/dl and serum ferritin levels higher than 600 ng/ml had been splenectomized.
1. Changes Related to Splenectomy (a) Lymphocyte subsets (Figs 1-3). High absolute lymphocyte counts were observed in the
majority of the patients examined (Table I). No significant differences were noted in the lymphocyte counts between splenectomized and nonsplenectomized patients.
Results of analysis of the two major lymphocyte subpopulations (Figs 1 and 2), namely E-rosette forming cells and cells with surface Ig, show that splenectomy had no effect on the absolute T lymphocyte count, regardless of whether the serum iron was lower or higher than 200 pg/dl. The decreased percentage of T cells (not shown) was due to an increase in the absolute number of circulating SIg positive lymphocytes following splenectomy (Fig 2) and was observed in patients with serum iron levels lower or higher than 200.
The numbers of T cells were comparable in both splenectomized and non-splenectomized patients although differences in the absolute number of the subsets of T cells were observed between the two groups. As shown in Fig 1, a moderate though not statistically significant decrease in the number of Tp cells was observed in splenectomized patients. No correlation of these numbers or proportions of T cell subpopulations was observed with the levels of iron. Numbers of T4 cells were moderately but not significantly increased in splenectomized patients.
Analysis of the changes in the circulating B lymphocytes (Fig 2) according to class of surface immunoglobulin showed that the overall increase detected with the polyvalent antiserum in splenectomized patients was due almost entirely to an increase in absolute numbers of SIgM and/or SIgD bearing cells. These changes were observed in both groups of splenectomized patients, but those with higher serum and ferritin levels tended to have lower numbers than those with serum iron under 200 pg/dl. Splenectomy had very little effect on numbers of circulating SIgG and SIgA bearing cells (Fig 2). Total numbers of MRFC (a subset of B lymphocytes) and 'null' cells were increased in splenectomized patients when compared with those of the non-splenectomized group (Fig 3). Significant increases in MRFC (P<0.05) and null cells ( P ~ 0 . 0 2 ) were observed in splenectomized patients with serum iron lower than 200
One patient with thalassaemia major was studied for the proportions of lymphocyte subpopulations in the peripheral blood and spleen; the results are shown in Table 11. Peripheral blood at the time of splenectomy demonstrated a normal distribution of lymphocyte subpopu- lations. No 'null' cells were found. The spleen, however, had higher proportions ofT cells and lower proportions of B cells than have been reported for normal human spleen (Jonsson & Christensen, 1977). Peripheral blood cell surface markers repeated 2 months following
Pg/dl-
Lymphoid Cells and Immunoglobulins in Thalassaemia
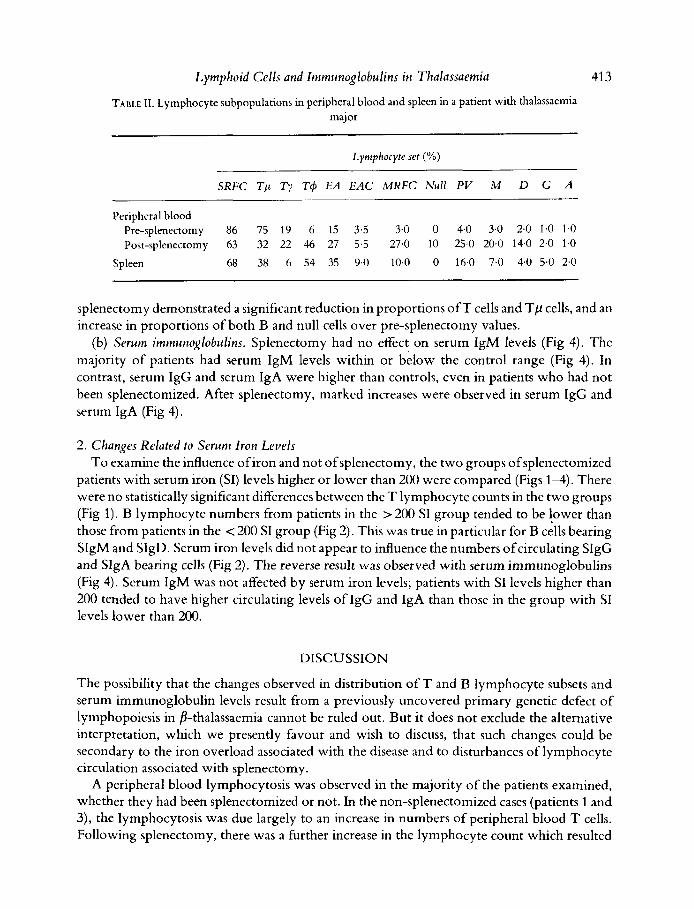
TABLE 11. Lymphocyte subpopulations in peripheral blood and spleen in a patient with thalassaemia major
413
Lymphocyte set (%)
SRFC T p Ty T 4 EA EAC MRFC Null PV M D G A
Peripheral blood Pre-splenectomy 86 75 19 6 15 3.5 3.0 0 4.0 3.0 2.0 1.0 1.0 Post-splenectomy 63 32 22 46 27 5.5 27.0 10 254 20.0 14.0 2.0 1.0
Spleen 68 38 6 54 35 9.0 10.0 0 16.0 7.0 4.0 5.0 2.0
splenectomy demonstrated a significant reduction in proportions o f T cells and Tp cells, and an increase in proportions of both B and null cells over pre-splenectomy values.
(b) Serum immuno~lobulins. Splenectomy had no effect on serum IgM levels (Fig 4). Thc majority of patients had serum IgM levels within or below the control range (Fig 4). In contrast, serum IgG and serum IgA were higher than controls, even in patients who had not been splenectomized. After splenectomy, markcd increases were observed in serum IgG and serum IgA (Fig 4).
2. Changes Related to Serum Iron Levels To examine the influence ofiron and not of splenectomy, the two groups ofsplenectomized
patients with serum iron (SI) levels higher or lower than 200 were comparcd (Figs 1-4). There were no statistically significant differences between the T lymphocyte counts in the two groups (Fig 1). B lyrnphocytc numbcrs from patients in the > 200 SI group tended to be lower than those from patients in the < 200 SI group (Fig 2). This was true in particular for B cells bearing SIgM and SIgD. Serum iron levels did not appear to influence the numbers of circulating SIgG and SIgA bearing cells (Fig 2). The reverse result was observed with serum immunoglobulins (Fig 4). Serum IgM was not affected by serum iron levels; patients with SI levels higher than 200 tended to have higher circulating levels of IgG and IgA than those in the group with SI levels lower than 200.
DISCUSSION
The possibility that the changes observed in distribution of T and B lymphocyte subsets and serum immunoglobulin levels result from a previously uncovered primary genetic defect of lymphopoiesis in 8-thalassaemia cannot be ruled out. But it does not exclude the alternative interpretation, which we presently favour and wish to discuss, that such changes could be secondary to the iron overload associated with the disease and to disturbances of lymphocyte circulation associated with splenectomy.
A peripheral blood lymphocytosis was observed in the majority of the patients examined, whether they had been splenectomized or not. In the non-splenectomized cases (patients 1 and 3), the lymphocytosis was due largely to an increase in numbers of peripheral blood T cells. Following splenectomy, there was a further increase in the lymphocyte count which resulted

41 4 Asha Kapadia et a1 from an increase in the numbers of circulating ‘null’ cells and B lymphocytes (MRFC) while total circulating T cells numbers remained unaffected. Imbalances amongst the T cell subsets did occur, however, which seemed to relate both to splenectomy and to serum iron levels. Thus, following removal of the spleen there was a decrease in the numbers of Tp in the blood with a concomitant increase in number of T4 cells which tended to increase with increasing concentrations of serum iron. Although the increases in T4 were not statistically significant, it is possible that iron plays a regulatory .role in expression of some T cell surface markers in vivo, similarly to its reported effects in vitro (de Sousa & Nishiya, 1978; Nishiya etal , 1979,1980). It is also possible that in the absence of the spleen some T cells, particularly Tp cell subsets, may migrate more readily to other tissues, namely lymph nodes and/or gut. This suggestion is further supported by a recent study showing that Ty cells move well and Ty cells do not move or move very poorly in the presence of the chemoattractant casein (Parrott et al, 1978). Further, lymph nodes contain high proportions of Ty cells and lack Ty cells, demonstrating some affinity of Tp for the lymph node (Gupta & Good, 1978).
The findings of increased serum levels of IgG and IgA, but not IgM, following splenectomy confirms earlier reports of the effect of splenectomy on serum immunoglobulins in p-thalassae- mia (Seitandis et al, 1971; Constantoulakis et al, 1978). It has been suggested that increased IgA may be related to liver damage (Wasi et al, 1971); in this study the finding of high serum IgA levels could not be related to altered liver function or frequency of infections.
The changes in serum immunoglobulins in p thalassaemia are compatible with an increased migration of T helper cells to gut and lymph nodes. Since synthesis of IgG and IgA is a thymus-dependent process (for review see de Sousa & Good, 1978), the probability that iron overload influences this process is further borne out by the fact that serum IgG and IgA levels were already higher than normal in patients who were not splenectomized but were still higher following splenectomy in patients with serum iron levels above 200 &dl. In addition, serum IgG and IgA levels in normal children who suffered splenic rupture do not differ from control values (Schumacher, 1970).
As mentioned, splenectomized patients had higher lymphocyte counts than nonsplenecto- mized patients, and this increase in lymphocyte numbers was due to an increase in ‘null’ cells and B lymphocytes.
The increase in B lymphocytes was mainly due to cells bearing SIgM and SIgD. No significant changes were observed in the populations ofSIgG and SIgA bearing cells, following splenectomy. These differences may relate to the role of the spleen in B cell ecotaxis (de Sousa, 1978b). In other words, the spleen is probably the environment of choice for the lodging of SIgM and SIgD bearing cells and for synthesis of these two immunoglobulin classes. In the absence of the correct ‘lodging site,’ the cells accumulate in the peripheral blood. By contrast, B cell sets with alternative environments for lodging and differentiation, continue to migrate, and function normally in the absence of the spleen.
In conclusion, the present results are compatible with the proposition that iron participates in regulation of lymphoid cell migration and in the modulation of expression of cell surface markers (de Sousa, 1978; Nishiya et al, 1980). They further illustrate the nature ofthe role ofthe spleen in the control of lymphocyte circulation and immunoglobulin synthesis and secretion.

Lymphoid Cells and Zmmunoglobulins in Thalassaemia 41 5
ACKNO W LEDGMENR
We wish to acknowledge the skilled technical assistance ofMs Judith Miller and Joseph Cavallo and help of nurses at the thalassaemia transfusion clinic. This work was aided by grants from the National Institutes of Health CA-08748, CA-17404, CA-19267, AI-11843, NS-11457, AG-O541, Zelda R. Weintraub Cancer Fund, Judith Harrisfelig Foundation, the Fund for Advanced Study of Cancer, Children’s Blood Foundation NIH-RO 1 HL 19898-03, and the H. Margolis (Jamieson) Fund.
REFERENCES
BASHKARAM, C . & REDDY, V. (1975) Cell-mediated immunity in iron and vitamin-deficient children. British MedicalJournal, iii, 522.
CHANDRA, R.K., Au, B., WOODFORD, G. & HYAM, P. (1977) Iron status, immune response and susceptibi- lity to infection. Ciba Foundation Symposium, No. 51, pp. 24P-268.
CONSTANTOULAKIS, M., TRKHOPOULOS, D., Avcous- TAKI, 0. & ECONOMIDOU, J. (1978) Serum im- munoglobulin concentrations before and after splenectomy in patients with homozygous 8-tha- 1assemia.Journal of Clinical Pathology, 31, 546-550.
DE SOUSA, M. (1Y78a) Lymphoid cell positioning: a new proposal for the mechanism of control of lym- phoid cell migration. Symposium on Experimental Biology, 32, 393-409.
DE SOUSA, M. (1978b) Ecotaxis, ecotaxopathy and lymphoid malignancy; terms, facts and predictions. The lmmunopathology of Lymphoreticular Neoplasms (ed. by J. J. Twomey and R. A. Good), pp. 325-359. Plenum Publishing Corporation, New York.
DE SOUSA, M. & GOOD, R.A. (1978) T and B cell populations in gut and gut associated lymphoid organs: arrangement, migration and function. Gas- trointestinal Tract Cancer (ed. by M. Lipkin and R. A. Good), pp. 29-44. Plenum Publishing Corpo- ration, New York.
DE SOUSA, M. & NISHIYA, K. (1978) Inhibition of E-rosette formation by two iron salts. Cellular Im- munology, 38,20>208.
DE SOUSA, M., SMITHYMAN, A.M. 81 TAN, C.T.C. (1978) Suggested models of ecotaxopathy in lym- phoreticular malignancy. AmericanJournal ofPatho-
ENGELHARD, D., CIVIDALLI, G. & RACHMILEWITZ, E.A. (1975) Splencctomy in homozygous beta thalassae- mia: a retrospective study of 30 patients. Britishjournal ofHaematology, 31, 391-403.
GUPTA, S. 81 GOOD, R.A. (1977a) Subpopulations of human T lymphocytes. 11. Effect of thymopoietin, corticosteroids and irradiation. Cellular Immu- nology, 34, 1G-18.
GUFTA, S . & GOOD, R.A. (1977b) Rosette formation
logy, 90,497-520.
with mouse erythrocytes. V. Relationship of mouse erythrocytes rosette forming cells to cells with sur- face immunoglobulin receptors for C3 and IgG Fc in patients with primary immunodeficiency disease. Clinical Immunology and Immunopathology, 8, 52@529.
GUPTA, S. & GOOD, R.A. (1978) Subpopulations of human T lymphocytes. 111. Distribution and quan- titation in peripheral blood, cord blood, tonsils, bone marrow, thymus, lymph nodes, and spleen. Cellular Immunology, 36, 265270.
HOFFBRAND, A.V. (1977) In discussion ofChandra et al (1977) Ciba Foundation Symposium, 51, p. 263.
HOFFBRAND, A.V., GANESHAGURU, K., HOOTEN, J.W.L. & TATTERSALL, M.H.N. (1976) Effect ofiron deficiency and desferrioxamine in DNA synthesis in human cells. British journal of Haematology, 33,
JONSSON. V. & CHRISTENSEN, B.E. (1977) Distribution of B, T and 0 lymphocytes in blood and tissues of normal subjects reflecting a kinetic model. Scan- dinavianJournal of Haematology, 18, 18.51%.
JOYNSON, H.H.M., JACOBS, A,, WALKER, D.M. & DOLBY, A.E. (1972) Defect of cell-mediated im- munity in patients with iron-deficiency anaemia. Lancet, ii, 1058-1059.
LARSSON, A. (1969) Ribonucleotide reductase from regenerating rat liver. EuropeanJournal ofBiochemis- try, 11, 11S121.
MACDOWALL, L.G., ANDERSON, R., McNAB,G.M.& KATZ, J. (1975) Immune response in iron-deficient children; impaired cellular defense mechanism< with altered humoral components. journal of Pedia- trics, 86, 83?443.
NISHIYA, K., GUFTA, S. & DE SOUSA, M. (1979) Differ- ential inhibitory effect of iron on E. EA and EAC rosette formation. Cellular Immunology, 46, 405-408.
NISHIYA, K., DE SOUSA, M., TSOI, E., BOGNACKI, J. & DE HARVEN, E. (1980) Regulation ofcxpression ofa human lymphoid cell surface marker by iron. Crl- lular Immunology, in press.
PARROTT, D.M.V., GOOD, R.A., O’NEILL, G.J. &
517-526.

416 Asha Kapadia et a1 GUPTA, S. (1978) Heterogeneity of locomotion in human T cell subsets. firneedings uf the National Academy $ Sciences $ the United Stutes $America, 75,2392-2395.
SCHUMACHER, M.J. (1970) Serum immunoglobulin and transferrin levels after childhood splenectomy. Archives $Diseases of Children, 45, 114-117.
SERANDIS, B., MIHAS, A. & ANGELOFQULOS, B. (1971) Serum immunoglobulins in /l-thalassemia after splenectomy. Ada Huematologica, 46,267-270.
WASI, C . , WAS, P. & THONGCHAROEN, P. (1971) Serum immunoglobulin levels in thalassaemia and the effects of splenectomy. Lancet, ii, 237-239.
Copyright © 2022 FDOKUMEN




















