
Low spatial frequency filtering modulates early brain processing of affective complex pictures
11
Neuropsychologia 45 (2007) 3223–3233 Low spatial frequency filtering modulates early brain processing of affective complex pictures Catalina Alorda a , Ignacio Serrano-Pedraza b , J. Javier Campos-Bueno c , Vicente Sierra-V´ azquez c , Pedro Montoya a,∗ a Department of Psychology and Research Institute on Health Sciences (IUNICS), University of the Balearic Islands, Palma, Spain b Psychology Division, School of Biology and Psychology, Newcastle University, Newcastle upon Tyne, UK c Department of Experimental Psychology, Universidad Complutense, Madrid, Spain Received 24 December 2006; received in revised form 19 June 2007; accepted 20 June 2007 Available online 1 July 2007 Abstract Recent research on affective processing has suggested that low spatial frequency information of fearful faces provide rapid emotional cues to the amygdala, whereas high spatial frequencies convey fine-grained information to the fusiform gyrus, regardless of emotional expression. In the present experiment, we examined the effects of low (LSF, <15 cycles/image width) and high spatial frequency filtering (HSF, >25 cycles/image width) on brain processing of complex pictures depicting pleasant, unpleasant, and neutral scenes. Event-related potentials (ERP), percentage of recognized stimuli and response times were recorded in 19 healthy volunteers. Behavioral results indicated faster reaction times in response to unpleasant LSF than to unpleasant HSF pictures. Unpleasant LSF pictures and pleasant unfiltered pictures also elicited significant enhancements of P1 amplitudes at occipital electrodes as compared to neutral LSF and unfiltered pictures, respectively; whereas no significant effects of affective modulation were found for HSF pictures. Moreover, mean ERP amplitudes in the time between 200 and 500 ms post-stimulus were significantly greater for affective (pleasant and unpleasant) than for neutral unfiltered pictures; whereas no significant affective modulation was found for HSF or LSF pictures at those latencies. The fact that affective LSF pictures elicited an enhancement of brain responses at early, but not at later latencies, suggests the existence of a rapid and preattentive neural mechanism for the processing of motivationally relevant stimuli, which could be driven by LSF cues. Our findings confirm thus previous results showing differences on brain processing of affective LSF and HSF faces, and extend these results to more complex and social affective pictures. © 2007 Elsevier Ltd. All rights reserved. Keywords: Affective pictures; Spatial frequency filtering; Event-related potentials 1. Introduction The processing of stimuli with biological significance requires specialized neural systems, which are able to rapidly detect threat cues and to mobilize the individual for sur- vival. Several EEG and MEG studies have provided convincing evidence that the visual processing of emotionally arousing stimuli elicits specific changes characterized by positive and negative amplitude enhancements of the visual event-related potentials (ERP) starting around 100 ms after stimulus onset ∗ Corresponding author at: Beatriu de Pin ´ os Building, Department of Psychol- ogy, Cra. de Valldemossa km 7.5, 07122 Palma, Spain. Tel.: +34 971 172646; fax: +34 971 172309. E-mail address: [email protected] (P. Montoya). (Batty & Taylor, 2003; Carretie, Hinojosa, Martin-Loeches, Mercado, & Tapia, 2004; Delplanque, Lavoie, Hot, Silvert, & Sequeira, 2004; Eger, Jedynak, Iwaki, & Skrandies, 2003; Keil et al., 2002; Meeren, van Heijnsbergen, & de Gelder, 2005; Montoya & Sitges, 2006; Pizzagalli, Regard, & Lehmann, 1999; Pourtois, Thut, Grave de Peralta, Michel, & Vuilleumier, 2005; Schupp, Junghofer, Weike, & Hamm, 2003; Skrandies, 1998; Smith, Cacioppo, Larsen, & Chartrand, 2003). These ampli- tude enhancements have been related to selective processing mechanisms for emotional stimuli, which could lead to the acti- vation of motivational brain systems (Cuthbert, Schupp, Bradley, Birbaumer, & Lang, 2000; Lang et al., 1998). Studies using neuroimaging techniques (fMRI, PET) and source modeling analyses have further shown that effects of emotional stimuli are strongest in occipital and posterior brain locations (Delplanque et al., 2004; Fredrikson et al., 1993; Keil 0028-3932/$ – see front matter © 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.neuropsychologia.2007.06.017
Transcript of Low spatial frequency filtering modulates early brain processing of affective complex pictures

A
tpwruomgosbr©
K
1
rdvesnp
oT
0d
Neuropsychologia 45 (2007) 3223–3233
Low spatial frequency filtering modulates early brain processingof affective complex pictures
Catalina Alorda a, Ignacio Serrano-Pedraza b, J. Javier Campos-Bueno c,Vicente Sierra-Vazquez c, Pedro Montoya a,∗
a Department of Psychology and Research Institute on Health Sciences (IUNICS), University of the Balearic Islands, Palma, Spainb Psychology Division, School of Biology and Psychology, Newcastle University, Newcastle upon Tyne, UK
c Department of Experimental Psychology, Universidad Complutense, Madrid, Spain
Received 24 December 2006; received in revised form 19 June 2007; accepted 20 June 2007Available online 1 July 2007
bstract
Recent research on affective processing has suggested that low spatial frequency information of fearful faces provide rapid emotional cues tohe amygdala, whereas high spatial frequencies convey fine-grained information to the fusiform gyrus, regardless of emotional expression. In theresent experiment, we examined the effects of low (LSF, <15 cycles/image width) and high spatial frequency filtering (HSF, >25 cycles/imageidth) on brain processing of complex pictures depicting pleasant, unpleasant, and neutral scenes. Event-related potentials (ERP), percentage of
ecognized stimuli and response times were recorded in 19 healthy volunteers. Behavioral results indicated faster reaction times in response tonpleasant LSF than to unpleasant HSF pictures. Unpleasant LSF pictures and pleasant unfiltered pictures also elicited significant enhancementsf P1 amplitudes at occipital electrodes as compared to neutral LSF and unfiltered pictures, respectively; whereas no significant effects of affectiveodulation were found for HSF pictures. Moreover, mean ERP amplitudes in the time between 200 and 500 ms post-stimulus were significantly
reater for affective (pleasant and unpleasant) than for neutral unfiltered pictures; whereas no significant affective modulation was found for HSF
r LSF pictures at those latencies. The fact that affective LSF pictures elicited an enhancement of brain responses at early, but not at later latencies,uggests the existence of a rapid and preattentive neural mechanism for the processing of motivationally relevant stimuli, which could be driveny LSF cues. Our findings confirm thus previous results showing differences on brain processing of affective LSF and HSF faces, and extend theseesults to more complex and social affective pictures.2007 Elsevier Ltd. All rights reserved.
als
(MSeMPS
eywords: Affective pictures; Spatial frequency filtering; Event-related potenti
. Introduction
The processing of stimuli with biological significanceequires specialized neural systems, which are able to rapidlyetect threat cues and to mobilize the individual for sur-ival. Several EEG and MEG studies have provided convincingvidence that the visual processing of emotionally arousing
timuli elicits specific changes characterized by positive andegative amplitude enhancements of the visual event-relatedotentials (ERP) starting around 100 ms after stimulus onset∗ Corresponding author at: Beatriu de Pinos Building, Department of Psychol-gy, Cra. de Valldemossa km 7.5, 07122 Palma, Spain.el.: +34 971 172646; fax: +34 971 172309.
E-mail address: [email protected] (P. Montoya).
StmvB
sel
028-3932/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.oi:10.1016/j.neuropsychologia.2007.06.017
Batty & Taylor, 2003; Carretie, Hinojosa, Martin-Loeches,ercado, & Tapia, 2004; Delplanque, Lavoie, Hot, Silvert, &
equeira, 2004; Eger, Jedynak, Iwaki, & Skrandies, 2003; Keilt al., 2002; Meeren, van Heijnsbergen, & de Gelder, 2005;ontoya & Sitges, 2006; Pizzagalli, Regard, & Lehmann, 1999;
ourtois, Thut, Grave de Peralta, Michel, & Vuilleumier, 2005;chupp, Junghofer, Weike, & Hamm, 2003; Skrandies, 1998;mith, Cacioppo, Larsen, & Chartrand, 2003). These ampli-
ude enhancements have been related to selective processingechanisms for emotional stimuli, which could lead to the acti-
ation of motivational brain systems (Cuthbert, Schupp, Bradley,irbaumer, & Lang, 2000; Lang et al., 1998).
Studies using neuroimaging techniques (fMRI, PET) andource modeling analyses have further shown that effects ofmotional stimuli are strongest in occipital and posterior brainocations (Delplanque et al., 2004; Fredrikson et al., 1993; Keil

3 cholo
eLetit2bffafspdfswhbcTpvrT2
oaihifcat2hostfah
PqntD2aecem
ne2MF
2
2
oitDe
2
t1vp(tNnra
p(oun3bfspost&fT&woBn
2
1daea
224 C. Alorda et al. / Neuropsy
t al., 2002; Lane, Reiman, Ahern, Schwartz, & Davidson, 1997;ane, Reiman, Bradley et al., 1997; Lang et al., 1998; Reimant al., 1997). Recently, fMRI studies have also demonstratedhat high (HSF) and low spatial frequency (LSF) informationnvolved in facial expression of fear might be processed by dis-inct neural channels (Vuilleumier, Armony, Driver, & Dolan,003; Winston, Vuilleumier, & Dolan, 2003). In this sense, it haseen observed that fusiform gyrus was more activated by fearfulaces containing all or only HSF visual information than by fear-ul faces containing only LSF information. By contrast, greatermygdala responses were elicited by LSF than by intact or HSFearful stimuli. Thus, it appeared that LSF information only pre-erves the configuration, particularly important in the emotionalrocessing; whilst HSF provides the visual system with fineetails, which are relevant in individual face identification. Theseunctional data seem to be in agreement with anatomical findingsuggesting the existence of a retinal–collicular–pulvinar path-ay projecting to the amygdala (Jones & Burton, 1976), anduman neuroimaging findings revealing functional interactionsetween the pulvinar nucleus and the amygdala during aversiveonditioning of facial stimuli (Morris, Friston, & Dolan, 1997).here is, therefore, suggestive evidence that pulvinar–amygdalaathways may be important in mediating responses to salientisual stimuli, and form part of an evolutionarily selected neu-al system which responds rapidly to threat or danger (Friston,ononi, Reeke, Sporns, & Edelman, 1994; Vuilleumier et al.,003).
It seems plausible that if such a neural system has been devel-ped as an adaptive mechanism for detecting fear-related stimuli,similar dissociation should also be present during the process-
ng of other emotional stimuli. In this sense, a recent fMRI studyas shown that although differential brain activation is elicitedn response to specific emotional stimuli (happy, sad, anger, andear) compared to neutral stimuli, both facial expressions andomplex pictures activated similar brain structures, including themygdala, posterior hippocampus, ventromedial prefrontal cor-ex and visual cortex (Britton, Taylor, Sudheimer, & Liberzon,006). The present study aimed to investigate the effects ofigh and low spatial frequency information on brain processingf pleasant and unpleasant pictures, depicting complex socialcenes. For this purpose, spatial frequency information for threeypes of affective pictures (pleasant, unpleasant, and neutral)rom the International Affective Picture System (IAPS, Lang etl., 2005) was selected using 2D isotropic Butterworth low- andigh-pass filters (Gonzalez & Wintz, 1977).
According with previous research, we hypothesized that early1 amplitudes of visual ERP will be affected by low spatial fre-uency (LSF) filtering of unpleasant pictures as compared toeutral pictures, reflecting that the automatic processing of emo-ionally aversive information is driven by LSF cues (Pourtois,an, Grandjean, Sander, & Vuilleumier, 2005; Vuilleumier et al.,003). Moreover, we expected that late ERP components (meanmplitudes between 200 and 600 ms post-stimulus) elicited by
ither LSF or HSF pictures would not be modulated by affectiveontent, since these components seem to be more related to thexplicit and cognitive appraisal of salient stimuli and, probably,ight reflect the involvement of more complex neural mecha-p(pefi
gia 45 (2007) 3223–3233
isms during affective processing (Carretie et al., 2004; Cuthbertt al., 2000; Herbert, Kissler, Junghofer, Peyk, & Rockstroh,006; Montoya, Larbig, Pulvermuller, Flor, & Birbaumer, 1996;ontoya & Sitges, 2006; Schupp et al., 2000, 2003; Schupp,
laisch, Stockburger, & Junghofer, 2006).
. Method
.1. Participants
Nineteen healthy undergraduate students (15 women and 4 men; 18–25 yearsld; average age: 21 years) from the University of Balearic Islands participatedn the experiment. All subjects were right-handed and had normal or correctedo normal vision. Participants gave written informed consent according with theeclaration of Helsinki, and received course credit for their participation. The
xperimental procedure was approved by the local institutional ethics committee.
.2. Affective stimuli
Stimuli were 117 standardized pictures selected from the International Affec-ive Picture System (IAPS) (Center for the Study of Emotion and Attention,999) consisting of 39 pleasant, 39 unpleasant, and 39 neutral pictures, based onalence and arousal ratings. Normative valence ratings (S.D. in parenthesis) forleasant, unpleasant, and neutral unfiltered raw pictures were 6.23 (0.77), 2.640.83), and 4.01 (0.93), respectively, on a scale ranging from 0 (very unpleasant)o 9 (very pleasant) (Lang et al., 2005; Spanish version from Molto et al., 1999).ormative arousal ratings (S.D. in parenthesis) for pleasant, unpleasant, andeutral unfiltered raw pictures were 5.34 (0.78), 5.67 (0.95), and 4.69 (0.80),espectively, on a scale ranging from 0 (very calm) to 9 (very excited) (Lang etl., 2005; Spanish version from Molto et al., 1999).
All stimuli were converted to black-and-white (256 grey levels) and encom-assed within a square frame of 512 × 512 pixels (Fig. 1). One set of 39 stimuli13 pleasant, 13 unpleasant, and 13 neutral pictures) was always presented with-ut filtering (broad-band stimuli, BSF). The rest of 78 stimuli (26 pleasant, 26npleasant and 26 neutral pictures) were used for filtering purposes and wereot presented unfiltered. Two different sets of stimuli, each of which contained9 pictures (13 pleasant, 13 unpleasant, and 13 neutral pictures), were obtainedy using a 2D isotropic Butterworth low-pass filter of order two with cut-offrequency of 15 cycles/image width (2.42 cycles/deg) (low spatial frequencytimuli, LSF). Two further sets of stimuli were obtained by filtering the sameictures using a 2D isotropic Butterworth high-pass filter of order two with cut-ff frequency of 25 cycles/image width (8.89 cycles/deg) (high spatial frequencytimuli, HSF). The two classes of spatial (non-causal) isotropic Butterworth fil-ers used have been defined in the 2D spatial-frequency domain [see Gonzalez
Wintz (1977, pp. 170, 181), and Sierra-Vazquez et al. (2006, Appendix A)or depictions of radial profiles of low-pass and high-pass filter Modulationransfer Functions]. Numerical filtering using MATLAB® (Gonzalez, Woods,Eddins, 2004) was applied in the spatial-frequency domain and then the result
as Fourier transformed into the spatial domain. Thus, nine stimulus categoriesf stimuli were used combining affective valence and filtering features: neutralSF, pleasant BSF, unpleasant BSF, neutral HSF, pleasant HSF, unpleasant HSF,eutral LSF, pleasant LSF, and unpleasant LSF.
.3. Procedure
Participants were seated in a dimly lit sound attenuated room in front of a7 in. computer screen with a resolution of 1280 × 1024 pixels, at a viewingistance of 125 cm. The experiment consisted of one practice block (5 trials)nd four blocks of 117 trials each, with a brief pause between blocks. Withinach block, one set of LSF (13 pleasant, 13 unpleasant, and 13 neutral pictures)nd one set of different HSF stimuli (13 pleasant, 13 unpleasant, and 13 neutral
ictures) were presented pseudorandomly together with the set of BSF stimuli13 pleasant, 13 unpleasant, and 13 neutral pictures). Furthermore, a specificicture (either filtered or unfiltered) was presented once within a block. Thus,ach BSF stimulus was repeated four times during the experiment, while eachltered stimulus (either LSF or HSF) was repeated only twice.
C. Alorda et al. / Neuropsychologia 45 (2007) 3223–3233 3225
F) an
pbdprw8
2
M15z(lweaw
2
qos1tf
o(wtmmotP
rbtGw
3
3
re
3
Fig. 1. Sample of broadband (BSF) pictures, as well as high-pass (HS
Each trial began with a fixation cross-followed by the presentation of aicture for a maximum of 5000 ms. Participants were asked to press a YESutton as fast as possible if they recognized the stimulus or a NO button if theyid not. Since subjects were not asked to report the picture content, behavioralarameters of this self-report task were based on the number of YES and NOesponses for each stimulus category. Following the key response, a dark screenith the fixation cross was presented with an intertrial interval varying between00 and 1000 ms. Reaction times were recorded from stimulus onset.
.4. Recording of event-related potentials
Scalp EEG was recorded with a QuickAmp amplifier (Brain Products GmbH,unich, Germany) from 64 electrodes mounted in an electrocap (extended
0–20 system) and using Cz as reference. Electrode impedances were kept belowk�. The amplifier bandpass was 0.5–40 Hz. EEG was sampled with a digiti-ation rate of 1000 Hz and stored for offline averaging. EEG epochs of 1100 ms100 ms pre-, 1000 ms post-stimulus) were obtained for each trial. After a base-ine correction was computed using the pre-stimulus interval (100 ms), epochsith artifacts (a voltage exceeding ±75 �V) were removed, and corrected for
ye blinks using a vertical EOG channel (Gratton & Coles’s algorithm) prior toveraging. Only those trials where the subject explicitly recognized the stimulusere included in the average.
.5. Data analyses
Statistical analyses of brain activity focused on the impact of spatial fre-uency filtering on ERP amplitudes elicited by affective pictures at specific sets
f electrodes. Multivariate analyses of variance (MANOVA) for repeated mea-ures were conducted on P1 amplitudes (positive maximum between 100 and50 ms post-stimulus at Oz electrode as reference), as well as mean ERP ampli-udes in four 100-ms time windows ranging from 200 to 600 ms post-stimulus atrontal (Fz, F3, and F4), central (Cz, C3, and C4), parietal (Pz, P3, and P4), andtsP
d low-pass filtered (LSF) pictures used as stimuli in the experiment.
ccipital electrodes (Oz, O1, and O2). For all these analyses, spatial frequencyBSF, HSF, and LSF) and emotional valence (pleasant, unpleasant, and neutral)ere used as within-subjects factors, and the different brain regions (frontal, cen-
ral, parietal, and occipital) as repeated measures variables. Moreover, repeatedeasures ANOVAs were also conducted on N170 amplitudes (negative maxi-um between 150 and 250 ms post-stimulus at CB1 electrode as reference) at
ccipito-temporal electrodes (CB1 and CB2). Greenhouse–Geisser adjustmentso the degrees of freedom were performed when appropriate, and the corrected
values as well as ε values are reported.Statistical analyses of behavioral performance were conducted using
epeated measures ANOVA on recognition rates and reaction times. Inoth cases, emotional valence (pleasant, unpleasant, and neutral) and spa-ial frequency (BSF, HSF, and LSF) were used as within-subjects factors.reenhouse–Geisser adjustments to the degrees of freedom were performedhen appropriate, and the corrected P values as well as ε values are reported.
. Results
.1. Behavioral performance
Table 1 summarizes the mean and the standard deviation forecognition rates and reaction times to recognized pictures in allxperimental conditions.
.1.1. Recognition rates
A significant emotional valence × spatial frequency interac-ion effect (F[4,72] = 9.6; P < 0.001; ε = 0.615) accompanied byignificant main effects of spatial frequency (F[2,36] = 66.4;< 0.001; ε = 0.868) and emotional valence (F[2,36] = 41.2;
3226 C. Alorda et al. / Neuropsycholo
Table 1Mean (±S.D.) recognition and responses times across experimental conditions(maximum and minimum values are indicated in bold)
Filter Valence Mean ± S.D.
Recognitionrates (%)
Yes responsestimes (ms)
BSF (unfiltered) Neutral 89.0 ± 8.3 654.2 ± 172Unpleasant 90.3 ± 4.5 658.8 ± 188Pleasant 90.7 ± 4.5 673.4 ± 194
HSF Neutral 71.7 ± 13.5 a 792.8 ± 272 bUnpleasant 64.9 ± 12.9 807.0 ± 302Pleasant 85.2 ± 10.5 760.3 ± 280
LSF Neutral 66.6 ± 8.8 a 737.6 ± 231Unpleasant 65.1 ± 9.8 735.2 ± 254Pleasant 73.3 ± 7.4 751.0 ± 225
Notes—a: significant differences between pleasant and neutral stimuli(fa
PyrtPBuSrant(swo
3
i(cε
stpLuStottwurp
3
tfEtwsnasa
3
Pvt(wGfSe(0fgegomtstiooraearH
ftε
a
3
P < 0.001), and between pleasant and unpleasant (P < 0.001); b: significant dif-erences between pleasant and neutral stimuli (P < 0.05), and between pleasantnd unpleasant (P < 0.01).
< 0.001; ε = 0.948) were observed. Follow-up pairwise anal-ses conducted separately for each spatial frequency categoryevealed significant greater recognition rates to pleasant thano unpleasant and to neutral pictures for LSF and HSF (all< 0.001), but no differences due to emotional valence forSF stimuli. No significant differences were observed betweennpleasant and neutral pictures in any spatial frequency category.eparated follow-up pairwise comparisons were conducted onecognition rates to pleasant, unpleasant, and neutral stimuli tossess the effects of spatial frequency. Significant higher recog-ition rates were observed in response to BSF than in responseo LSF or HSF pictures for all three types of affective picturesall P < 0.05). In addition, recognition rates for pleasant HSFtimuli were greater than for pleasant LSF stimuli (P < 0.01);hereas no significant effects of spatial frequency was observedn recognition rates for unpleasant or neutral stimuli.
.1.2. Reaction timesStatistical analyses on reaction times also yielded a signif-
cant emotional valence × spatial frequency interaction effectF[4,64] = 6.9; P < 0.001; ε = 0.717) accompanied by a signifi-ant main effect of spatial frequency (F[2,32] = 17.6; P < 0.001;= 0.677). Pairwise comparisons conducted separately for eachpatial frequency category revealed significant faster reactionimes to pleasant than to unpleasant (P < 0.01) and to neutralictures (P < 0.05) only for HSF stimuli, but not for BSF orSF stimuli. No significant differences were observed betweennpleasant and neutral pictures in any spatial frequency category.eparated pairwise comparisons were conducted on reaction
imes for each affective stimulus category to assess the effectsf spatial frequency. Reaction times were more reduced to BSFhan to LSF and to HSF pictures for all three types of affec-ive pictures (all P < 0.05). Furthermore, faster reaction times
ere observed in response to unpleasant and neutral LSF stim-li than to unpleasant and neutral HSF stimuli (both P < 0.05),espectively; but no significant differences were found betweenleasant LSF and HSF stimuli.asav
gia 45 (2007) 3223–3233
.2. Event-related brain potentials
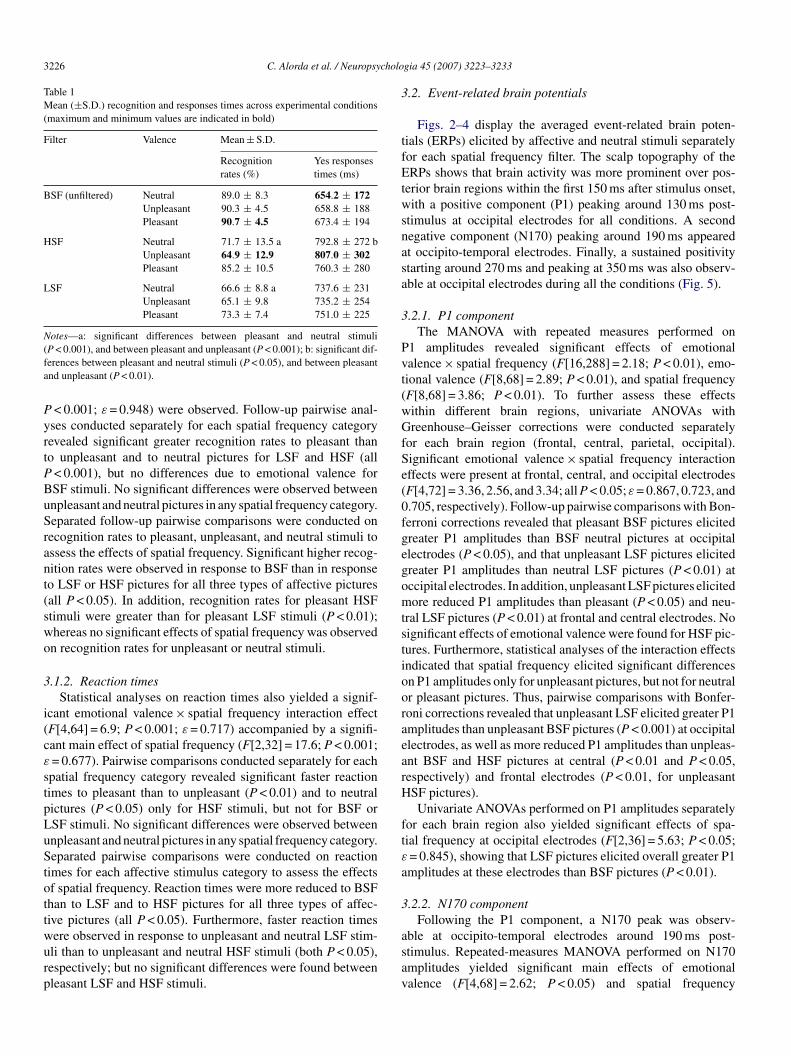
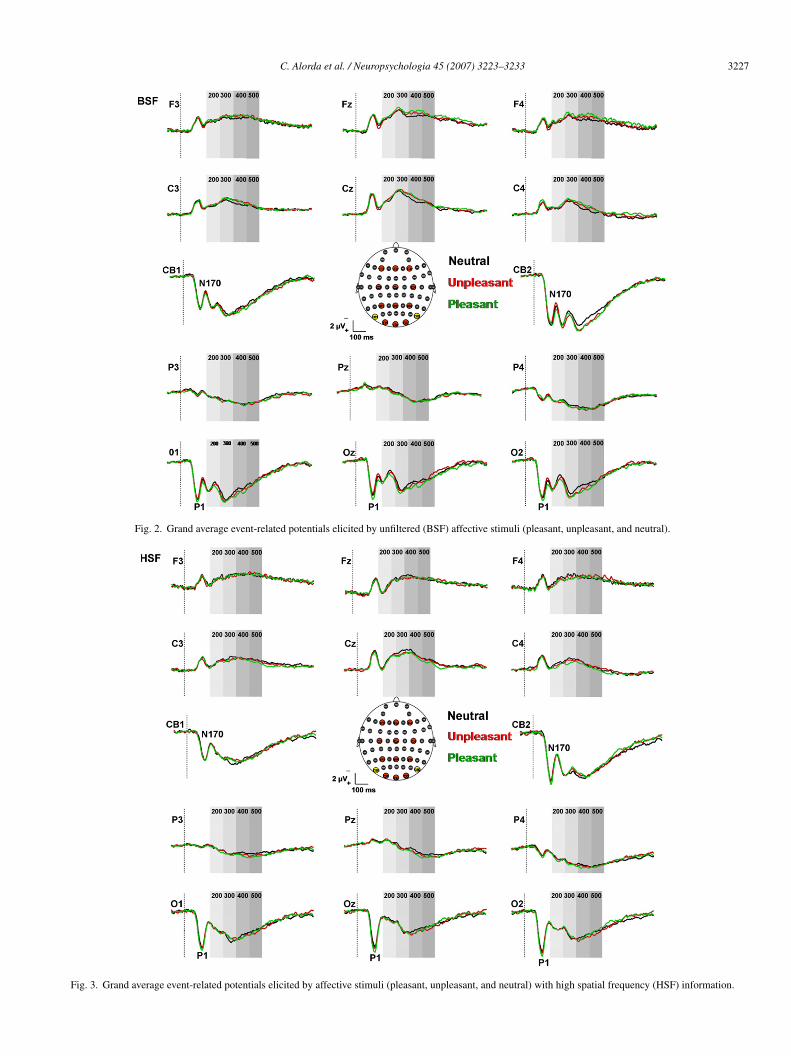
Figs. 2–4 display the averaged event-related brain poten-ials (ERPs) elicited by affective and neutral stimuli separatelyor each spatial frequency filter. The scalp topography of theRPs shows that brain activity was more prominent over pos-
erior brain regions within the first 150 ms after stimulus onset,ith a positive component (P1) peaking around 130 ms post-
timulus at occipital electrodes for all conditions. A secondegative component (N170) peaking around 190 ms appearedt occipito-temporal electrodes. Finally, a sustained positivitytarting around 270 ms and peaking at 350 ms was also observ-ble at occipital electrodes during all the conditions (Fig. 5).
.2.1. P1 componentThe MANOVA with repeated measures performed on
1 amplitudes revealed significant effects of emotionalalence × spatial frequency (F[16,288] = 2.18; P < 0.01), emo-ional valence (F[8,68] = 2.89; P < 0.01), and spatial frequencyF[8,68] = 3.86; P < 0.01). To further assess these effectsithin different brain regions, univariate ANOVAs withreenhouse–Geisser corrections were conducted separately
or each brain region (frontal, central, parietal, occipital).ignificant emotional valence × spatial frequency interactionffects were present at frontal, central, and occipital electrodesF[4,72] = 3.36, 2.56, and 3.34; all P < 0.05; ε = 0.867, 0.723, and.705, respectively). Follow-up pairwise comparisons with Bon-erroni corrections revealed that pleasant BSF pictures elicitedreater P1 amplitudes than BSF neutral pictures at occipitallectrodes (P < 0.05), and that unpleasant LSF pictures elicitedreater P1 amplitudes than neutral LSF pictures (P < 0.01) atccipital electrodes. In addition, unpleasant LSF pictures elicitedore reduced P1 amplitudes than pleasant (P < 0.05) and neu-
ral LSF pictures (P < 0.01) at frontal and central electrodes. Noignificant effects of emotional valence were found for HSF pic-ures. Furthermore, statistical analyses of the interaction effectsndicated that spatial frequency elicited significant differencesn P1 amplitudes only for unpleasant pictures, but not for neutralr pleasant pictures. Thus, pairwise comparisons with Bonfer-oni corrections revealed that unpleasant LSF elicited greater P1mplitudes than unpleasant BSF pictures (P < 0.001) at occipitallectrodes, as well as more reduced P1 amplitudes than unpleas-nt BSF and HSF pictures at central (P < 0.01 and P < 0.05,espectively) and frontal electrodes (P < 0.01, for unpleasantSF pictures).Univariate ANOVAs performed on P1 amplitudes separately
or each brain region also yielded significant effects of spa-ial frequency at occipital electrodes (F[2,36] = 5.63; P < 0.05;= 0.845), showing that LSF pictures elicited overall greater P1mplitudes at these electrodes than BSF pictures (P < 0.01).
.2.2. N170 componentFollowing the P1 component, a N170 peak was observ-
ble at occipito-temporal electrodes around 190 ms post-timulus. Repeated-measures MANOVA performed on N170mplitudes yielded significant main effects of emotionalalence (F[4,68] = 2.62; P < 0.05) and spatial frequency
C. Alorda et al. / Neuropsychologia 45 (2007) 3223–3233 3227
Fig. 2. Grand average event-related potentials elicited by unfiltered (BSF) affective stimuli (pleasant, unpleasant, and neutral).
Fig. 3. Grand average event-related potentials elicited by affective stimuli (pleasant, unpleasant, and neutral) with high spatial frequency (HSF) information.

3228 C. Alorda et al. / Neuropsychologia 45 (2007) 3223–3233
pleasa
(vwrae
c(
Fp
Fig. 4. Grand average event-related potentials elicited by affective stimuli (
F[4,68] = 3.13; P < 0.05); but no interaction effect of emotionalalence × spatial frequency was observed. Univariate ANOVAs
ith Greenhouse–Geisser corrections were conducted sepa-ately for each electrode location (CB1 and CB2), revealingsignificant main effect of emotional valence only for CB2
lectrode (F[2,34] = 4.31; P < 0.05; ε = 0.934), and a signifi-
pppN
ig. 5. Grand average event-related potentials elicited by affective stimuli (pleasant, uanel). Topographic maps of the mean ERP amplitude in the time interval between 2
nt, unpleasant, and neutral) with low spatial frequency (LSF) information.
ant main effect of spatial frequency only for CB1 electrodeF[2,34] = 6.15; P < 0.05; ε = 0.782). Follow-up pair wise com-
arisons with Bonferroni corrections indicated that neutralictures elicited more enhanced N170 amplitudes than pleasantictures (P < 0.05), and that HSF pictures elicited more enhanced170 amplitudes than LSF pictures (P < 0.05).npleasant, and neutral) for each spatial frequency category at Oz electrode (left00 and 400 ms post-stimulus (right panel).

cholo
36
ima0t05m
ctvw(0cupteepwea(
et3043ataficfaa
4
fioGtilau(
oscp(oat(tf
4a
ulaLnpaiicprtHasdcetiu
Evtta(tapSTpfo
C. Alorda et al. / Neuropsy
.2.3. Mean amplitudes in time-windows from 200 to00 ms post-stimulus
The MANOVA with repeated measures yielded signif-cant effects of emotional valence × spatial frequency on
ean ERP amplitudes for time-windows 200–300, 300–400,nd 400–500 ms (F[16,288] = 2.09, 2.41, and 1.82; P < 0.01,.01, and 0.05, respectively), and spatial frequency for allime-windows (F[8,68] = 3.64, 4.88, 2.88, and 2.19; P < 0.01,.001, 0.01, and 0.05, for 200–300, 300–400, 400–500, and00–600 ms, respectively). No significant emotional valenceain effects were found for any time-window.Univariate ANOVAs with Greenhouse–Geisser corrections
onducted separately for each brain region (frontal, cen-ral, parietal, and occipital) revealed significant emotionalalence × spatial frequency interaction effects in the time-indows 200–300 and 300–400 ms at occipital electrodes
F[4,72] = 6.27 and 6.76; P < 0.01 and 0.001; ε = 0.824 and.919, respectively), and in the time-window 400–500 ms atentral electrodes (F[4,72] = 5.28; P < 0.01; ε = 0.795). Follow-p comparisons confirmed that pleasant and unpleasant BSFictures elicited greater mean amplitudes than neutral BSF pic-ures at occipital electrodes (Ps < 0.05) (Fig. 6), and at centrallectrodes (P < 0.05) for those time-windows. No significantffects of emotional valence were found for either HSF or LSFictures at those time-windows. Nevertheless, follow-up pair-ise comparisons also revealed that BSF and LSF pictures
licited more enhanced amplitudes than HSF pictures for pleas-nt and unpleasant pictures at occipital and central electrodesall Ps < 0.05), but not for neutral pictures.
These univariate ANOVAs also yielded significant mainffects of spatial frequency on mean ERP amplitudes in allime-windows at occipital electrodes (F[2,36] = 9.73, 16.35,.52, and 5.84; P < 0.01, 0.001, 0.05, and 0.05; ε = 0.765,,805, 0.911, and 0.816, respectively, for 200–300, 300–400,00–500, and 500–600 ms), and in time-windows 200–300 and00–400 ms at frontal (F[2,36] = 11.22, and 15.06; P < 0.001nd 0.001; ε = 0.946 and 0.885, respectively), and central elec-rodes (F[2,36] = 7.36 and 18.02; P < 0.01 and 0.001; ε = 0.799nd 0.836, respectively). Follow-up pairwise comparisons con-rmed that both LSF and BSF pictures, regardless of affectiveontent, elicited significant greater amplitudes than HSF picturesor time-windows 200–300 and 300–400 ms at frontal, central,nd occipital electrodes (all Ps < 0.5), as well as for 400–500nd 500–600 ms at occipital electrodes (all Ps < 0.5).
. Discussion
The present study examined the influence of spatial frequencyltering on brain processing of affective complex pictures, asbserved previously with emotional fearful faces (Goffaux,authier, & Rossion, 2003; Pourtois, Dan et al., 2005). For
his purpose, event-related potentials (ERPs) were recordedn response to affective pictures depicting pleasant (erotic,
andscapes, babies), unpleasant (mutilations, snakes, roaches,ccidents) or neutral scenes (clothes, mushrooms, neutral faces,mbrellas) from the International Affective Picture SystemIAPS). To manipulate the spatial frequency characteristicsthoa
gia 45 (2007) 3223–3233 3229
f the stimuli, pictures were presented either unfiltered (BSFtimuli), low-pass filtered to retain only low spatial frequencyomponents (<15 cycles/image width) (LSF stimuli), or high-ass filtered to retain only high spatial frequency components>25 cycles/image width) (HSF stimuli). According with previ-us findings using affective facial expressions (Pourtois, Dan etl., 2005), it was hypothesized that the early P1 component ofhe visual ERP would be influenced by LSF filtering of affectivepleasant and unpleasant) as compared to neutral pictures, whilehe late ERP components would not be modulated by the spatialrequency filtering of affective content.
.1. Early brain activity is enhanced by unpleasant picturesnd low spatial frequency filtering
Our data confirmed that early ERP components elicited bynpleasant pictures, but not by pleasant pictures, were modu-ated by LSF filtering. Thus, a significant enhancement of P1mplitude was observed during the processing of unpleasantSF pictures in comparison with neutral LSF pictures, whereaso differences were found during the processing of pleasant LSFictures or affective HSF pictures (either pleasant or unpleas-nt). More interestingly, these interaction effects appeared onlyn the early P1 component over the occipital electrodes, but notn the late ERP components, such as the face-sensitive N170omponent or the positive mean amplitudes starting at 200 msost-stimulus. These findings are in agreement with previousesults, where fearful LSF faces also elicited larger P1 ampli-udes than neutral LSF faces, but no effects were found in theSF condition (Pourtois, Dan et al., 2005). Our results thus
re in line with recent evidence suggesting that low and highpatial frequency information of affective stimuli might provideifferent emotional and perceptual features during the early pro-essing of visual information (Vuilleumier et al., 2003; Winstont al., 2003). Moreover, we extend previous findings by showinghat early visual processing might be driven by LSF cues not onlyn response to affective facial expressions, but also to complexnpleasant stimuli depicting motivationally relevant scenes.
P1 is considered the earliest endogenous component of visualRPs, originating at several extrastriate regions within posteriorisual cortex (Woldorff et al., 2002, 1997). It has been suggestedhat P1 amplitude may be enhanced by both the voluntary alloca-ion of attentional resources (Luck, Woodman, & Vogel, 2000),nd the automatic capture of motivationally relevant stimuliTaylor, 2002). Enhancements of visual P1 amplitude at occipi-al electrodes, together with marked reductions of P1 amplitudest frontal electrodes, have been observed in response to com-lex stimuli depicting affective scenes (Carretie et al., 2004;chupp et al., 2003; Smith et al., 2003), expressive faces (Batty &aylor, 2003; Pizzagalli et al., 1999; Pourtois, Thut et al., 2005),hotographs showing incongruent affective expressions of theace and the body (Meeren et al., 2005), schematic drawingsf emotional facial expressions (Eger et al., 2003), or affec-
ive words (Skrandies, 1998). Previous neuroimaging researchas further shown that the visual cortex, including lingual, mid-ccipital and fusiform gyri might be also differentially activateds a function of valence using stimuli such as affective pictures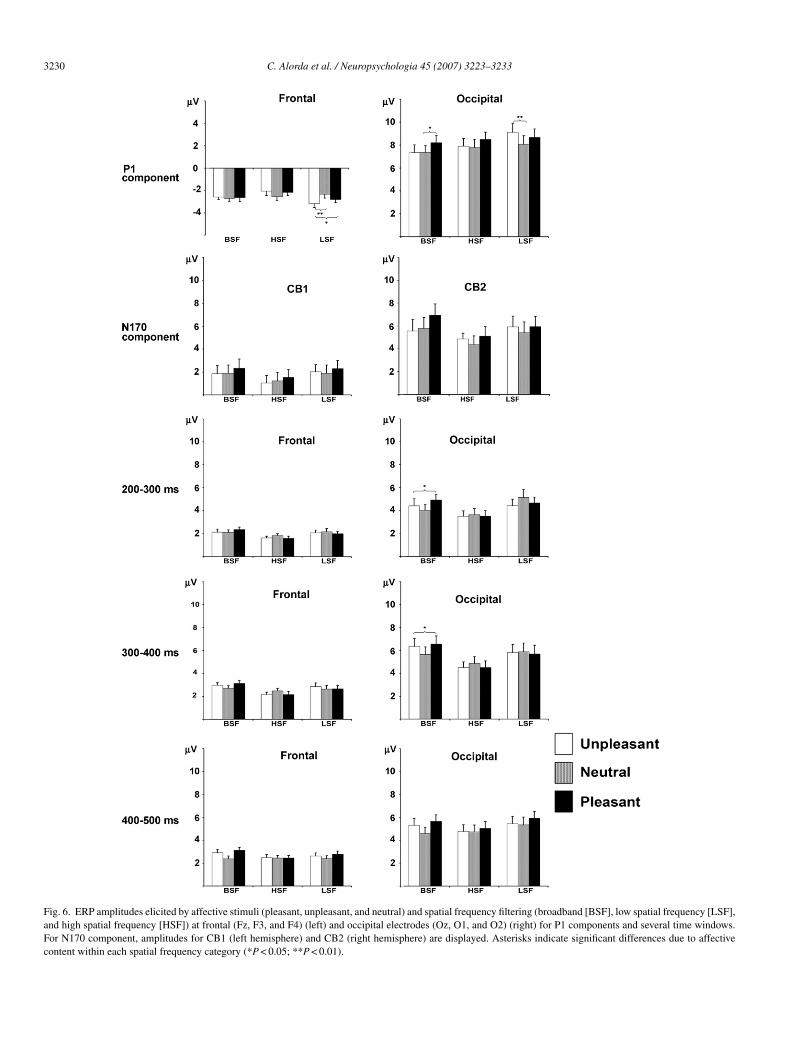
3230 C. Alorda et al. / Neuropsychologia 45 (2007) 3223–3233
Fig. 6. ERP amplitudes elicited by affective stimuli (pleasant, unpleasant, and neutral) and spatial frequency filtering (broadband [BSF], low spatial frequency [LSF],and high spatial frequency [HSF]) at frontal (Fz, F3, and F4) (left) and occipital electrodes (Oz, O1, and O2) (right) for P1 components and several time windows.For N170 component, amplitudes for CB1 (left hemisphere) and CB2 (right hemisphere) are displayed. Asterisks indicate significant differences due to affectivecontent within each spatial frequency category (*P < 0.05; **P < 0.01).

cholo
(&e1msdmMohase
amfvsb(tsttasiteFrsb2ffat
4a
uptf2t1bcmBa
crLGhgCMGCttglNf
4s
tBapEH1alhvAtcs2rBasteDeajmspssi
C. Alorda et al. / Neuropsy
Lang et al., 1998; Sabatinelli, Flaisch, Bradley, Fitzsimmons,Lang, 2004; Taylor et al., 2000), unpleasant films (Reiman
t al., 1997), or imagined traumatic experiences (Rauch et al.,996). Thus, our findings of significant P1 amplitude enhance-ents at occipital electrodes following pleasant and unpleasant
timuli compared to neutral stimuli in the BSF and LSF con-ition, respectively, may reflect an increased attention towardsotivationally relevant stimuli during early brain processing.oreover, the fact that no affective modulation was observed
n P1 amplitudes elicited by HSF stimuli further supports theypothesis that enhanced early processing of affective stimuli,s it has been observed for emotional fearful faces, might beelectively driven by LSF rather than by HSF cues (Vuilleumiert al., 2003).
The P1 amplitude enhancement to unpleasant LSF picturess compared to neutral LSF and to unpleasant HSF picturesight also reflect the effects of modulatory feedback signals
rom amygdala and orbitofrontal cortex to motivationally rele-ant and threat-related stimuli (Pourtois, Dan et al., 2005). In thisense, feedforward and feedback connections among differentrain regions as suggested by recent models of visual processingAdolphs, 2004; Bullier, 2001) would be likely mechanisms forhe observed effects on P1 amplitude in response to affectivetimuli. Thus, a specific retinal–collicular–pulvinar projectiono the amygdala has been proposed as subcortical route forhe processing of masked fear-related stimuli without consciouswareness (Morris, Ohman, & Dolan, 1999). Moreover, a recenttudy using diffusion tensor imaging (DTI) technique has alsodentified that the inferior longitudinal fasciculus might mediatehe rapid transfer of visual signals between the amygdala andarly visual areas in the human brain (Catani, Jones, Donato, &fytche, 2003). Finally, recent fMRI data have shown selectiveesponses to fearful LSF faces in the amygdala, thalamus anduperior colliculus; while fusiform gyrus was more activatedy BSF and HSF faces than by LSF faces (Vuilleumier et al.,003). All these findings strongly suggest that such rapid feed-oward connections from the amygdala might be responsibleor a rapid and preattentive coding of affective stimuli in visualreas, thus leading to an amplified and enhanced response overhe extrastriate cortex during visual processing.
.2. High spatial frequency filtering influenced N170mplitudes regardless of affective content
We also examined whether the N170 component can be mod-lated by affective and spatial frequency information during therocessing of complex stimuli. Source analyses have revealedhat neural generators of this component are located within theusiform gyrus (Iidaka, Matsumoto, Haneda, Okada, & Sadato,006), and are measured over the scalp at lateral occipito-emporal electrodes (Bentin, Allison, Puce, Perez, & McCarthy,996). Traditionally, an enhancement of N170 amplitude haseen observed using face stimuli as compared to hands, houses,
ars, or other common objects, suggesting that this componentay be linked to early stages of facial processing (Eimer, 2000;entin et al., 1996). Nevertheless, neuroimaging research haslso shown that common objects and complex affective scenessacr
gia 45 (2007) 3223–3233 3231
an activate, although to a lesser degree than faces, the poste-ior and middle fusiform gyrus (Britton et al., 2006; Ioannides,iu, Kwapien, Drozdz, & Streit, 2000; Joseph, 2001; Joseph &athers, 2003; Liu, Ioannides, & Streit, 1999). Recent evidenceas further suggested that N170 amplitude might also reflect aeneral perceptual expertise in discriminating objects (Gauthier,urran, Curby, & Collins, 2003; Kolassa, Musial, Kolassa, &iltner, 2006; Rossion, Joyce, Cottrell, & Tarr, 2003; Rossion,authier, Goffaux, Tarr, & Crommelinck, 2002; Tanaka &urran, 2001). In the present study, we found that N170 ampli-
udes were more enhanced in response to HSF as comparedo LSF pictures. These ERP differences were accompanied byreater recognition rates for HSF than for LSF pictures, regard-ess of affective content. Thus, our data may suggest that the170 component is more sensitive to high than to low spatial
requency information.
.3. Enhancement of late ERP amplitudes elicited by lowpatial frequency and unfiltered stimuli
A further finding of the present study was that late ERP ampli-udes were enhanced in response to both pleasant and unpleasantSF compared to neutral BSF stimuli for occipital electrodesnd central electrodes. These findings are in agreement withrevious observations of an affective modulation of middle-lateRP components (Carretie et al., 2004; Cuthbert et al., 2000;erbert et al., 2006; Montoya & Sitges, 2006; Montoya et al.,996; Schupp et al., 2000, 2003, 2006). In those studies, eitherlong-lasting positive shift beginning 200–300 ms after stimu-
us or an enhanced P2 component has been observed followingighly arousing emotional stimuli (e.g., pictures with erotic oriolent content, affective words) as compared to neutral ones.lthough its neural origin is still unknown, it has been suggested
hat activity in orbitofrontal and anterior cingulate cortices mightontribute to the P2 component, reflecting the evaluation of thealience of a given stimulus to the current task (Potts & Tucker,001). Moreover, an enhancement of P2 amplitude has beenelated to conscious processing of affective content (Bernat,unce, & Shevrinm, 2001). According with this assumption,recent study revealed that patients with affective disorders
uch as panic disorder displayed more enhanced P2 amplitudeso panic-related words than to neutral words, whilst no differ-nces appeared in healthy controls (Pauli, Amrhein, Muhlberger,engler, & Wiedemann, 2005). Thus, it appears that amplitude
nhancement around 200 ms post-stimulus might be considereds an attention-related index of stimulus ability to catch sub-ect’s attention and, thus, might reflect the engagement of the
otivational system for more complete and explicit appraisal ofalient stimuli (Cuthbert et al., 2000; Lang et al., 1998). In theresent study, enhanced ERP amplitudes elicited by unpleasanttimuli in the time windows 200–300 and 300–400 ms post-timulus were only present in response to BSF stimuli, but notn response to either LSF or HSF stimuli. Thus, it could be pos-
ible that the lack of affective ERP modulation for LSF stimulit these latencies were related to difficulties for an explicit andonscious appraisal of these stimuli. Behaviorally, the reducedecognition rate and increased reaction times to LSF and HSF
3 cholo
ppectbwfc
sdotwissitedof2itctaa
A
BdFd
R
AB
B
B
B
B
C
C
C
D
E
E
F
F
G
G
G
G
H
H
I
I
J
J
J
K
K
K
L
L
232 C. Alorda et al. / Neuropsy
ictures as compared to BSF pictures seem to support this inter-retation. Nevertheless, the fact that both LSF and BSF pictureslicited higher ERP amplitudes than HSF, regardless of affectiveontent, also suggests that the later enhancement of ERP ampli-udes for LSF pictures could be the result of an arousal differenceetween LSF and HSF pictures. It is possible that future workill uncover reliable arousal differences due to different spatial
requency filtering through, for example, the recording of skinonductance changes.
Summarizing, our results confirm and extend previous studieshowing that low and high spatial frequency information pro-uce differential effects on brain processing of affective stimuliver the primary visual cortex. We observed that unpleasant pic-ures elicited more enhanced P1 amplitudes than neutral pictureshen they were conveying predominantly low spatial frequency
nformation, but not when they were conveying exclusively highpatial frequency information. These findings provide furtherupport for the hypothesis of dissociated neural mechanismsn the brain processing of unpleasant information based onheir sensitivity to different ranges of spatial frequencies. Nev-rtheless, these results must be interpreted with caution, as aisproportionate number of female subjects were included inur sample. In this sense, significant gender effects have beenound in brain responses to unfiltered IAPS pictures (Hofer et al.,006; Kemp, Silberstein, Armstrong, & Nathan, 2004), suggest-ng a neural mechanism underlying gender-related vulnerabilityo some affective disorders, such a depression and anxiety. Tolarify these issues, further research should investigate the puta-ive influence of gender on brain processing of affective LSFnd HSF stimuli in healthy controls, as well as in patients withffective disorders.
cknowledgements
Research was supported by grants SEJ2004-01332 (PM) andSO2002-00192 (VSV & ISP) from the Spanish Ministerioe Ciencia y Tecnologıa and European Regional Developmentunds, as well as by grant PR145/03-12169 from Comunidade Madrid (Spain) to VSV, ISP and JJCB.
eferences
dolphs, R. (2004). Emotional vision. Nature Neuroscience, 7, 1167–1168.atty, M., & Taylor, M. J. (2003). Early processing of the six basic facial
emotional expressions. Cognitive Brain Research, 17, 613–620.entin, S., Allison, T., Puce, A., Perez, E., & McCarthy, G. (1996). Electro-
physiological studies of face perception in humans. Journal of CognitiveNeuroscience, 8, 551–565.
ernat, E., Bunce, S., & Shevrinm, H. (2001). Event-related brain potentialsdifferentiate positive and negative mood adjectives during both supraliminaland subliminal visual processing. International Journal of Psychophysiol-ogy, 42, 11–34.
ritton, J. C., Taylor, S. F., Sudheimer, K. D., & Liberzon, I. (2006). Facialexpressions and complex IAPS pictures: Common and differential networks.Neuroimage, 31, 906–919.
ullier, J. (2001). Integrated model of visual processing. Brain Research BrainResearch Review, 36, 96–107.
arretie, L., Hinojosa, J. A., Martin-Loeches, M., Mercado, F., & Tapia, M.(2004). Automatic attention to emotional stimuli: Neural correlates. HumanBrain Mapping, 22, 290–299.
L
gia 45 (2007) 3223–3233
atani, M., Jones, D. K., Donato, R., & Ffytche, D. H. (2003). Occipito-temporalconnections in the human brain. Brain, 126, 2093–2107.
uthbert, B. N., Schupp, H. T., Bradley, M. M., Birbaumer, N., & Lang, P. J.(2000). Brain potentials in affective picture processing: Covariation withautonomic arousal and affective report. Biological Psychology, 52, 95–111.
elplanque, S., Lavoie, M. E., Hot, P., Silvert, L., & Sequeira, H. (2004).Modulation of cognitive processing by emotional valence studied throughevent-related potentials in humans. Neuroscience Letters, 356, 1–4.
ger, E., Jedynak, A., Iwaki, T., & Skrandies, W. (2003). Rapid extraction ofemotional expression: Evidence from evoked potential fields during briefpresentation of face stimuli. Neuropsychologia, 41, 808–817.
imer, M. (2000). The face-specific N170 component reflects late stages in thestructural encoding of faces. Neuroreport, 11, 2319–2324.
redrikson, M., Wik, G., Greitz, T., Eriksson, L., Stone-Elander, S., Ericson, K.,et al. (1993). Regional cerebral blood flow during experimental phobic fear.Psychophysiology, 30, 126–130.
riston, K. J., Tononi, G., Reeke, G. N., Sporns, O., & Edelman, G. M. (1994).Value-dependent selection in the brain: simulation in a synthetic neuralmodel. Neuroscience, 59, 229–243.
authier, I., Curran, T., Curby, K. M., & Collins, D. (2003). Perceptualinterference supports a non-modular account of face processing. NatureNeuroscience, 6, 428–432.
offaux, V., Gauthier, I., & Rossion, B. (2003). Spatial scale contribution toearly visual differences between face and object processing. Cognitive BrainResearch, 16, 416–424.
onzalez, R. C., & Wintz, P. (1977). Digital Image Processing. Reading, Mass:Addison-Wesley.
onzalez, R. C., Woods, R. E., & Eddins, S. L. (2004). Digital Image ProcessingUsing MATLAB. Upper Saddle River, NJ: Prentice Hall.
erbert, C., Kissler, J., Junghofer, M., Peyk, P., & Rockstroh, B. (2006). Pro-cessing of emotional adjectives: Evidence from startle EMG and ERPs.Psychophysiology, 43, 197–206.
ofer, A., Siedentopf, C. M., Ischebeck, A., Rettenbacher, M. A., Verius, M.,Felber, S., et al. (2006). Gender differences in regional cerebral activityduring the perception of emotion: A functional MRI study. Neuroimage, 32,854–862.
idaka, T., Matsumoto, A., Haneda, K., Okada, T., & Sadato, N. (2006).Hemodynamic and electrophysiological relationship involved in human faceprocessing: Evidence from a combined fMRI–ERP study. Brain Cognition,60, 176–186.
oannides, A. A., Liu, L. C., Kwapien, J., Drozdz, S., & Streit, M. (2000).Coupling of regional activations in a human brain during an object and faceaffect recognition task. Human Brain Mapping, 11, 77–92.
ones, E. G., & Burton, H. (1976). A projection from the medial pulvinar to theamygdala in primates. Brain Research, 104, 142–147.
oseph, J. E. (2001). Functional neuroimaging studies of category specificity inobject recognition: A critical review and meta-analysis. Cognitive, Affectiveand Behavioral Neuroscience, 1, 119–136.
oseph, J. E., & Gathers, A. D. (2003). Natural and manufactured objects activatethe fusiform face area. Neuroreport, 13, 935–938.
eil, A., Bradley, M. M., Hauk, O., Rockstroh, B., Elbert, T., & Lang, P. J.(2002). Large-scale neural correlates of affective picture processing. Psy-chophysiology, 39, 641–649.
emp, A. H., Silberstein, R. B., Armstrong, S. M., & Nathan, P. J. (2004).Gender differences in the cortical electrophysiological processing of visualemotional stimuli. Neuroimage, 21, 632–646.
olassa, I. T., Musial, F., Kolassa, S., & Miltner, W. H. (2006). Event-relatedpotentials when identifying or color-naming threatening schematic stimuliin spider phobic and non-phobic individuals. BMC Psychiatry, 6, 38.
ane, R. D., Reiman, E. M., Ahern, G. L., Schwartz, G. E., & Davidson, R.J. (1997). Neuroanatomical correlates of happiness, sadness, and disgust.American Journal of Psychiatry, 154, 926–933.
ane, R. D., Reiman, E. M., Bradley, M. M., Lang, P. J., Ahern, G. L., Davidson,
R. J., et al. (1997). Neuroanatomical correlates of pleasant and unpleasantemotion. Neuropsychologia, 35, 1437–1444.ang, P. J., Bradley, M. M., Fitzsimmons, J. R., Cuthbert, B. N., Scott, J. D.,Moulder, B., et al. (1998). Emotional arousal and activation of the visualcortex: An fMRI analysis. Psychophysiology, 35, 199–210.

cholo
L
L
L
M
M
M
M
M
M
P
P
P
P
P
R
R
R
R
S
S
S
S
S
S
S
T
T
V
W
W
C. Alorda et al. / Neuropsy
ang, P. J., Bradley, M. M., & Cuthbert, B. N. (2005). International affectivepicture system (IAPS): Affective ratings of pictures and instruction manual.Technical Report A-6. Gainesville, FL: University of Florida.
iu, L., Ioannides, A. A., & Streit, M. (1999). Single trial analysis of neuro-physiological correlates of the recognition of complex objects and facialexpressions of emotion. Brain Topography, 11, 291–303.
uck, S. J., Woodman, G. F., & Vogel, E. K. (2000). Event-related potentialstudies of attention. Trends in Cognitive Science, 4, 432–440.
eeren, H. K., van Heijnsbergen, C. C., & de Gelder, B. (2005). Rapid perceptualintegration of facial expression and emotional body language. Proceedingsof the National Academy of Science of the United States of America, 102,16518–16523.
olto, J., Montanes, S., Poy, R., Segarr, P., Pastor, M. C., & Tormo, M. P.(1999). Un nuevo metodo para el estudio experimental de las emociones: elinternacional affective picture system (hipas), adaptacion espanola. Revistade Psicologıa General y Aplicada, 52, 55–87.
ontoya, P., Larbig, W., Pulvermuller, F., Flor, H., & Birbaumer, N. (1996).Cortical correlates of semantic classical conditioning. Psychophysiology,33, 644–649.
ontoya, P., & Sitges, C. (2006). Affective modulation of somatosensory-evoked potentials elicited by tactile stimulation. Brain Research, 1068,205–212.
orris, J. S., Friston, K. J., & Dolan, R. J. (1997). Neural responses to salientvisual stimuli. Proceedings: Biological sciences, 264, 769–775.
orris, J. S., Ohman, A., & Dolan, R. J. (1999). A subcortical pathway tothe right amygdala mediating “unseen” fear. Proceedings of the NationalAcademy of Science of the United States of America, 96, 1680–1685.
auli, P., Amrhein, C., Muhlberger, A., Dengler, W., & Wiedemann, G. (2005).Electrocortical evidence for an early abnormal processing of panic-relatedwords in panic disorder patients. International Journal of Psychophysiology,57, 33–41.
izzagalli, D., Regard, M., & Lehmann, D. (1999). Rapid emotional face pro-cessing in the human right and left brain hemispheres: An ERP study.Neuroreport, 10, 2691–2698.
otts, G. F., & Tucker, D. M. (2001). Frontal evaluation and posterior repre-sentation in target detection. Brain Research Cognitive Brain Research, 11,147–156.
ourtois, G., Dan, E. S., Grandjean, D., Sander, D., & Vuilleumier, P. (2005).Enhanced extrastriate visual response to bandpass spatial frequency fil-tered fearful faces: Time course and topographic evoked-potentials mapping.Human Brain Mapping, 26, 65–79.
ourtois, G., Thut, G., Grave de Peralta, R., Michel, C., & Vuilleumier, P. (2005).Two electrophysiological stages of spatial orienting towards fearful faces:Early temporo-parietal activation preceding gain control in extrastriate visual
cortex. Neuroimage, 26, 149–163.auch, S. L., van der Kolk, B. A., Fisler, R. E., Alpert, N. M., Orr, S. P., Savage, C.R., et al. (1996). A symptom provocation study of posttraumatic stress disor-der using positron emission tomography and script-driven imagery. Archivesof General Psychiatry, 53, 380–387.
W
gia 45 (2007) 3223–3233 3233
eiman, E. M., Lane, R. D., Ahern, G. L., Schwartz, G. E., Davidson, R. J.,Friston, K. J., et al. (1997). Neuroanatomical correlates of externally andinternally generated human emotion. American Journal of Psychiatry, 154,918–925.
ossion, B., Gauthier, I., Goffaux, V., Tarr, M. J., & Crommelinck, M. (2002).Expertise training with novel objects leads to left-lateralized face-like elec-trophysiological responses. Psychological Science, 13, 250–257.
ossion, B., Joyce, C. A., Cottrell, G. W., & Tarr, M. J. (2003). Early lateral-ization and orientation tuning for face, word, and object processing in thevisual cortex. Neuroimage, 20, 1609–1624.
abatinelli, D., Flaisch, T., Bradley, M. M., Fitzsimmons, J. R., & Lang, P. J.(2004). Affective picture perception: Gender differences in visual cortex?Neuroreport, 15, 1109–1112.
chupp, H. T., Cuthbert, B. N., Bradley, M. M., Cacioppo, J. T., Ito, T., &Lang, P. J. (2000). Affective picture processing: The late positive poten-tial is modulated by motivational relevance. Psychophysiology, 37, 257–261.
chupp, H. T., Flaisch, T., Stockburger, J., & Junghofer, M. (2006). Chapter2 emotion and attention: Event-related brain potential studies. Progress inBrain Research, 156, 31–51.
chupp, H. T., Junghofer, M., Weike, A. I., & Hamm, A. O. (2003). Emotionalfacilitation of sensory processing in the visual cortex. Psychological Science,14, 7–13.
ierra-Vazquez, V., Serrano-Pedraza, I., & Luna, D. (2006). Effect of spatial-frequency filtering on the visual processing of global spatial structure.Perception, 35, 1583–1609.
krandies, W. (1998). Evoked potential correlates of semantic meaning—A brainmapping study. Brain Research Cognitive Brain Research, 6, 173–183.
mith, N. K., Cacioppo, J. T., Larsen, J. T., & Chartrand, T. L. (2003). May Ihave your attention, please: Electrocortical responses to positive and negativestimuli. Neuropsychologia, 41, 171–183.
anaka, J. W., & Curran, T. (2001). A neural basis for expert object recognition.Psychological Science, 12, 43–47.
aylor, M. J. (2002). Non-spatial attentional effects on P1. Clinical Neurophys-iology, 113, 1903–1908.
uilleumier, P., Armony, J., Driver, J., & Dolan, R. J. (2003). Distinct spatial fre-quency sensitivities for processing faces and emotional expressions. NatureNeuroscience, 6, 624–631.
inston, J. S., Vuilleumier, P., & Dolan, R. J. (2003). Effects of low-spatialfrequency components of fearful faces on fusiform cortex activity. CurrentBiology, 13, 1824–1829.
oldorff, M. G., Fox, P. T., Matzke, M., Lancaster, J. L., Veeraswamy, S., Zamar-ripa, F., et al. (1997). Retinotopic organization of early visual spatial attentioneffects as revealed by PET and ERPs. Human Brain Mapping, 5, 280–
286.oldorff, M. G., Liotti, M., Seabolt, M., Busse, L., Lancaster, J. L., & Fox,P. T. (2002). The temporal dynamics of the effects in occipital cortex ofvisual–spatial selective attention. Brain Research Cognitive Brain Research,15, 1–15.




















